Embed Size (px)
Citation preview

The Central Dogmaof molecular biology
RNA
DNA
transcription
replication
translation
Protein
study in detailthe molecular machines – the central dogma in action…
RNA polymerase; (creates mRNA transcript)
Ribosomes (synthesize proteins from mRNA)
Helicase;DNA polymerase(cell division)
(Show movie)

DNA unzipping and motor proteins: effect of the genetic code
• Statistical mechanicsof unzipping long dsDNA at constant force (Fc=15 pN)�Strong first order transition � delocalization of the unzipping fork
• Sequence heterogeneitydominates within ~5pN of the transition� Energy barriers ~ kBT√M, M = genome size! (kBT ≈ 0.6 kcal/mole)
� Anomalous dynamicsof the unzipping fork for F > Fc� Experimental evidence for jumps and pauses while unzipping λ-phage DNA
• Anomalous dynamics of molecular motors near the stall force� Dynamics of molecular motors on DNA substrates (driven by NTP’s…)
� Polymerases, exonucleases and translocation via SSB proteins
TheoristsDavid Lubensky
Yariv KafriJulius Lucks
Buddhapriya Chakrabarti
ExperimentersClaudia Danilowicz
Vincent ColjeeCedric Bouzigues
Mara Prentiss

DNA Stretching Experiments
Smith et. al. Science 271, 785 (1996)
21 1/ (1 )
2 4
x xFa kT
L L= − − +
Bustamante, Marko, Siggia and Smith, Science 265, 1599(1994)
�Single molecule DNA stretching experiments,made possible by laser tweezers and highlyspecific biological linking agents, provideddirect experimental evidence for the Kratky-Porod(or “worm-like chain”) theory of entropic polymer elasticity; circa 1949.
�Excellent fits to force-extension curves of dsDNAcharacterized only by a “contour length” L and an effective monomer size a.
Entropic spring

Random walks and entropic elasticity
7 random walks, all starting at the origin
�The total number of random walks terminating at r is a Gaussian….
2 2/2 3/ 2
1( )
( )N r Nar z e
Nlπ−≈r
�
N ~ 10,000 step random walk
rr
L = contour lengtha=monomer sizeN = L/a = number of
independent units
21( ) . ( )
2G r const kr F r F kr= + − =
ur rr r��Add a force F to obtain an “entropic spring”:
�The free energy G(r) associated with end-to-end distance r is purely entropic:
[ ]2
( ) ln ( )
1. ; 2 /
2
B
B
G r TS T k r
const kr k k T L a
= − = −
= + =
�

Single Molecule Biophysics Experiments
Bustamante et. al., Nature 421,423 (2003); see also van Oijenet. al. Science 30129 (2003)
Reversible action of DNA polymerase/exonuclease…
Strick et. al., Nature 404, 901 (2000)
Relaxation of supercoiledDNA by topoisomerase II …
chiral specificity revealed;no “inactive” enzymes

Force-induced unfolding of RNA hairpins
Liphardt et. al., Science 292, 733 (2001)

Single Molecule DNA Unzipping Transitions
Bockelmann et. al., Biophys. J. 82, 1537 (2002)
�Previous experiments and simulationsperformed in a “constant extension” ensemble
�However, exact results possible in a more
tractable (and biologically relevant?) “constant
force” ensemble ….
�Unzipping for F < Fc(T) dominated by pauses and jumps determined by the base pair sequence
� Precise predictions available even for single molecule experiments
� Anomalous dynamics of the DNA unzipping fork for F > Fc(T); barriers scale as square root of genome size!
David K. Lubensky and drn:

ThermalDenaturation of DNA (poly A:T or G:C)
for homopolymers like poly(A:T)or poly(G:C)…
2
20
( ))
( ) . ( )
bm
m
N T
a T const T T
1 ∼ (Τ − Τ
∼ − − {
( )2 2
( ,0; ) / ( ,0; ) ( ) ( ,0; )2N
TT Z r N F T Z r N U r Z r N
g
−∂ = ∇ − +r r ur ur r r r r r
( ,0; )Z r N ∝r r probability that N base pairs of
DNA are separated by r at top
rr
applied force at top…

Force-induced denaturation of poly(A:T)
a0(T)
= 2a1(F)
�First order “unzipping” transition at Fc(T)
�Two “phases” coexist at the unzipping fork
unzipping fork
If m base pairs are unzipped, the unzipping bias f = 2a1(F) – a0
controls the energy landscape ε(m)
0 1
0
( ) ( ) 2m N m a ma
a N fm
ε = − + = +{

0, '( ) ( ')
0,
m mm m
otherwiseη η
∆ = ≈
average alongsequence
For most coding DNA…
0' 0
( ) ( ')m
m
m a N fm mη=
ε = + +∑
sequenceinformation!
Generic energy landscape for DNA unzipping (f/kBT= 0.01)
0 1000 2000 3000 4000 5000
m
-40
-20
0
20
40
60
ε(m
)
Energy landscape for bacteriophage X174; (f/kBT = 0.01; Santa Lucia et.al.)
Unzipping of heterogeneousDNA

Statistical mechanics of the unzipping fork( ) /
0
' 0
( ) , ( 1)
( ) ( ')
( )
N m TB
m
m
c
Z m e dm k
m fm m
f F T F
η
−∆ε
=
= =
∆ε = +
∝ −
∫
∑
�For homopolymers [η(m) = ∆0 = 0], <m> ≈ T/(Fc-F)
�However, sequence heterogeneity [∆0 ≠ 0] leads toplateaus, jumps and astronger divergence:
20
2
/
1/( )c
m f
F F
< > ≈ ∆
∝ −
/( )cm T F F< > ∝ −

The dynamics of the unzipping fork is controlled by a random forceenergy landscape
0 0( ) | | ( ') '
mm a N f m m dmηε = − − + ∫
0 1000 2000 3000 4000 5000
m
-80
-60
-40
-20
0
20
ε(m
)
Energy landscape for bacteriophage X174
f/kT = -0.01
Langevin equation for position of the unzipping fork m(t):
( ) ( ))
| | ( ) )
dm t d mt
dt dmf m t
ξ
η ξ
ε= − Γ + (
= Γ −Γ + (
Physics same as particle in a random force field: Sinai, Derrida, ~1982-83; Long time dynamics depends only onµ = 2kBT|f|/∆0
22 2
00
| | 0,
( )[ ( ) (0)] ln ( / )
0 1,
( ) ~
B
if f then
k Tm t m t
if then
m t tµ
µ
τ
µ
= =
< − > =∆
< <
< >
(logarithmic localization)
(sub-ballistic drift)

DNA unzipped under a constant force exhibits multiple metastable intermediates
C. Danilowicz, M. Prentiss, et. al. PNAS 100, 1694 (2003)
�Several dozen identical λ-phage DNA’s attached to magnetic beads and unzipped in parallel in a magnetic fieldgradient.
�Positions and pauses duringunzipping process provide a sequence-dependent molecularfingerprint.
�Unzipping is very slowat piconewton forces; several hoursrequired to unzip the 48,502base pairs of phage lambda.
Fully zipped
Fully unzipped
(top view)

Unzipping histories and energy landscapes for F = 15pN
�Experiments consistent with largeenergy barriers and sequence specificpause points.
�Positions and local jumps betweentwo-level systems concide with thosepredicted for phage lambda
3 different DNA’s unzpped @ 15 pN
4 different DNA’s unzipped @20pN
}

Simulation of unzipping dynamics for bacteriophage X174
Simulations and genomelandscapes courtesy ofJulius Lucks….
See movie…..
f =-0.02


0, '( ) ( ')
0,
m mm m
otherwiseη η
∆ = ≈
average alongsequence
For most coding DNA…
0' 0
( ) ( ')m
m
m a N fm mη=
ε = + +∑
sequenceinformation!
Helicase����Unzipping of heterogeneous DNA
0 1000 2000 3000 4000 5000
m
-80
-60
-40
-20
0
20
ε(m
)
Energy landscape for bacteriophage X174
f/kT = -0.01
Langevin equation for position of the unzipping fork m(t):
( ) ( ))
dm t d mt
dt dmξε= − Γ + (
Physics same as particle in a random force field: Sinai, Derrida, ~1982-83; Long time dynamics depends only on µ = 2kBT|f|/∆0
0 1,
( ) ~
if then
m t tµ
µ < <
< >(sub-ballistic drift)

Overview for DNA unzipping: energy barriers scale like √M!
0 1000 2000 3000 4000 5000
m
-40
-20
0
20
40
ε(m
)
73 kT
Bacteriophage φX174 M = 5386, √M ≈ 73
RNA hairpinM = 25, √M ≈ 5
Danilowicz et. al. Phys. Rev. Lett. 93, 078101 (2004)
5 kT
Liphardt et. al
�
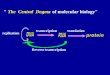
Are energy landscapes for viral genomes really ‘random walks’?
E. coli phage T7 (dsDNA, linear)M =39,936, √M ≈ 200
Fluctuations scale like √√√√M in many viruses, but …..
ε (m)
m
kT√M
E. coli phage T7
M = 39,936
E. coli phage X174 (ssDNA, circular)M = 5386, √M ≈ 73
kT√M
E. coli phage X174
M = 5386
ε (m)
m
kT√M
Roseobacter phage
SIO1, M = 39898
ε (m)
m
m
E. coli phage K
M = 6089
ε (m) kT√M

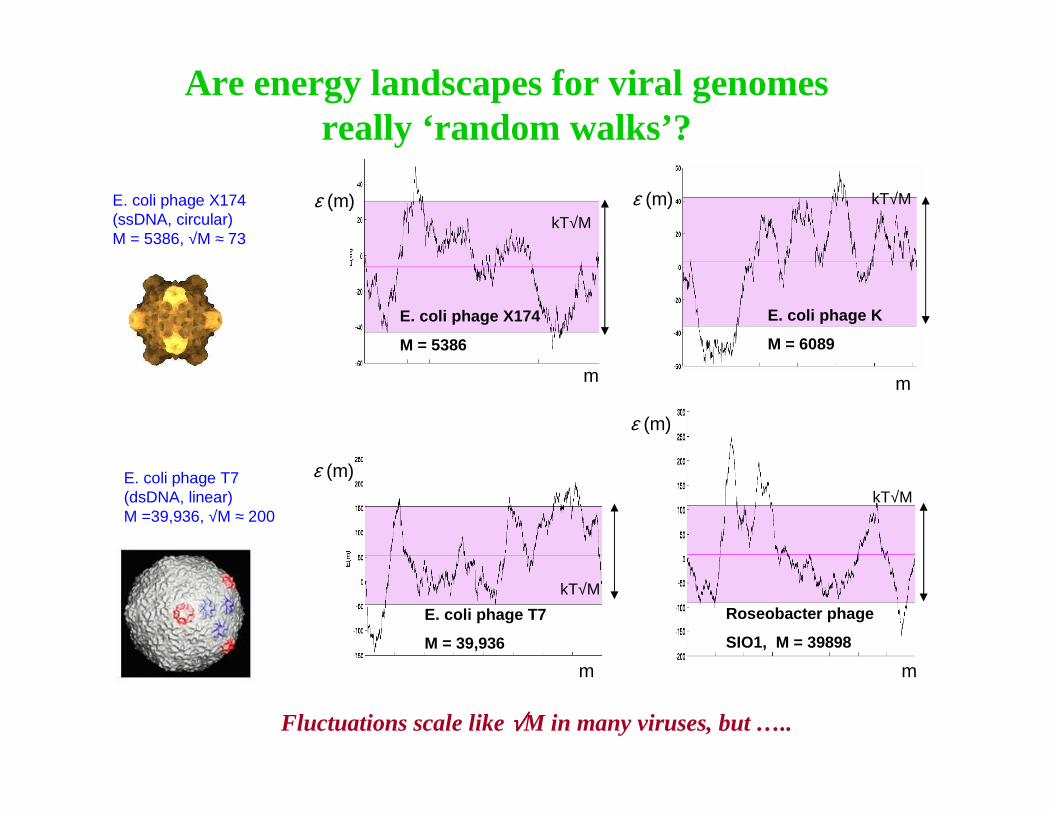
phage lambda is exceptional…
E. coli phage lambda (dsDNA, linear) M=48502, √M ≈ 220
�front end of λ-phage is GC-rich and back end is AT-rich
�is this the result of an ancient splicing of genomes infecting thermophillic and thermophobic organisms?
�mechanical explanation from packingconstraints?
ε (m)
m
kT√M

Anomalous landscapes also appear in other temperate phages (share lysis/lysogeny switch)
upward slopes � head/tail genes….
Phage P2Phage HK022
�P2 landscape hard to reconcile with “mechanical packing explanation” ….
�Can we determine what part of landscape is due to the amino acid sequences in the proteins and what part is due to codon usage?

Synonymous mutations in temperate phages
Phage P2
●Residual features are due to conserving the protein sequences….
● Does codon bias preserve the original landscape over evolutionary time scales?
Phage Lambda
(assume mutation rate ~ 1 x 10-6/bp-replication)

R. W. Hendrix & S. Casjens , “Bacteriophage λλλλ and Its Genetic Neighborhood”, The Bacteriophages ( Oxford, 2004)
0 10000 20000 30000 40000 50000
m
0
500
1000
1500
2000
2500
3000
ε(m)
m
kT√M
ε (m)
Hypothesis: GC content of phage lambda is related to lysogenic/lytic life cycle and/or codon bias of E. coli host
phage tail ≈ 0.15 µ \

Codon bias in E. Coli
http://www.evolvingcode.net/codon/cai/cais.php
Leucine is spelled “CUG”with frequency ~ 65%
�Codon bias pattern of a gene correlates with the levels with which it is translated into protein and indeed the accuracy of the translation process
�Codon bias tables are typically constructed from abundantly produced ribosomal proteins (~12,000 ribosomes per cell), elongation factors, etc.
But…11/19 (58%) of the most highly preferred codons have GC in the 3rd position
�tRNA abundances and translational machinery of E. coli (and other bacteria) are biased towards rapid transcription of certain codons.
In E. coli 8/19 (42%) of the most highly preferred codons have AT in the 3rd position
Phage Lambda
GC Landscape
GC3 Landscape
Cumulative CAI Landscape

kinesin –periodic substrate; small, simple
RNAp –is sequenceheterogeneityImportant?
Anomalous dynamics for motor proteins?[Y. Kafri, D. Lubensky, drn; Biophys. J. 86: 3373 (2004)]
(fit due to Fisher and Kolomeisky PNAS 2001 98: 7748-7753; see alsoJulicher, Prost and Ajdari)) K. Vissher, M. J. Schnitzer, S. M. Block
Nature 400, 184 (1999)
F
velocity
M. Wang et al, Science282, 902 (1998)

Cis side trans side
MEMBRANE
m monomers translocated
− +
Dynamics with heterogeneity: one possibility is a random energy landscape:
Electric field drives hetero polymer like ssDNA through a pore…)
( )mε
m
�Local interaction of nucleotides with pore leads to tilted “random energy” landscape with bounded fluctuations….
�Resulting dynamics is “diffusion with drift”, with diffusion constant and drift velocity renormalized by the randomness….
( ) v 2R Rm t t D t≅ ±
ssDNA translocation through a pore

Motors on periodic tracks: kinesinwalking along a microtubule
Kinesin-microtubule binding potential (note asymmetry)
Binding potential with ATP driving force
Motor moves from the plus end to the minus end, powered by ATP hydrolysis and opposed by an applied force F
-
F
• Describe motion of chemically driven motors by a simple two-sublattice model
• Lack of inversion symmetry means
, b b a aw w w w→ ← → ←≠ ≠
0 2 4 6
51 3 7
8
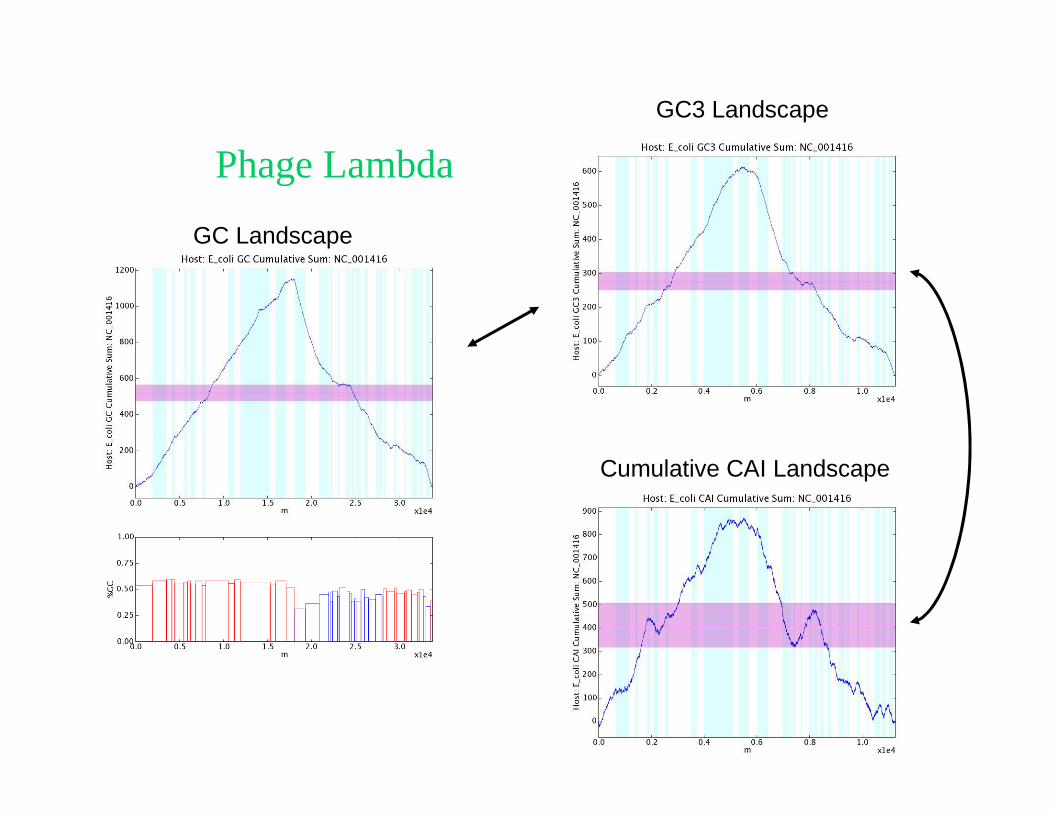
Lattice Model of Motor Proteins
/ / / 2
/ 2
/ / / 2
/ 2
( )
( )
( )
( )
T T f Ta
f Tb
T T f Ta
f Tb
w e e
w e
w e e
w e
µ ε
µ ε
α ωα ωα ωα ω
→ ∆ −∆ −
←
← ∆ −∆ +
→ −
= +
= +′ ′= +′ ′= +
1 1
1 1
( )( ) ( ) ( ) ( )
(odd sites)
( )( ) ( ) ( ) ( )
(even sites)
na n a n b b n
nb n b n a a n
dP tw P t w P t w w P t
dt
dP tw P t w P t w w P t
dt
→ ← → ←− +
→ ← → ←− +
= + − +
= + − +
, , a a b bw w w and w→ ← → ←
Motor dynamics can be captured by a simplified two-state modelwith two sublattices a and b and rate constants
� Two parallel channels, α and ω,
both biased by an applied force f
� ω-channel describes thermal transitions over a barrier ∆ε, unassisted by ATP
[ ][ ][ ]
[ ][ ] [ ]
ATPATPln ln
ADP ADPeq
i ieq eq
TP P
µ ∆ = −
� α-channel powered by ATP hydrolysis
� Directional motion requires α ≠ α’, ω ≠ ω’,
(due to lack of inversion symmetry)
On a periodic substrate, dynamics is again diffusion with drift….

For ∆µ >> ∆ε can again integrate out even sites to obtain, e. g.,
Decimation for Motors on Heterogeneous Tracks
Rates are now location-dependent!
13 3 1
13
If (13) = 0, then E E - E 2 "random energy" landscape
Otherwise, is a random function of position along the track and a
"random force" landscape is generated....
fµ
η
∆ ∆ = +

Sequence heterogeneity leads toa random force landscape for RNAp…..
functions of location along track
for this setup is not a function of position
Random force landscape with energyfluctuations which grow as
1( ) 2 ( )
m
lE m fm lη
== +∑
Simulation with η(l) nonzero
YK10

Slide 31
YK10 DNA or RNAYariv Kafri; 09/12/2003

Other sources of random forcing
RNA polymerase
produces RNAusing NTP energy
Both random chemical energy & random chemical potential for each nucleotide play a role….
effective energy landscape
explicit random forcing due to nucleotide chemical potentials
1 1( ) 2 ( ) ( )
m m
l lE m fm f l lµ η
= == + +∑ ∑

convexvelocity force curve…
Experimental tests of predications
window-dependent effective velocity…
(MCS)
Close to the stall force, the observed “velocity” will depend on the experimental time scale tE!
1
( ) ,
( )lim 0
( ) 1!
t
m t t so
m tv t
tif f
µ
µ
µ
−
→∞
= →
<
�
�
Fs
stall force

Summary: anomalous dynamics near the stall forceof molecular motors…
�Sequence heterogeneity matters near the stall force Fc of RNAp.
�Effective velocity vanishes at very long times in a window around Fs…
stall force
M. D. Wang et. al., Science 282, 902 (1998)
RNAp ?
Lambda exonuclease? [see, e.g., van Oijen et. al. Science 301 29 (2003)]
Simulations, courtesy of Yariv Kafri
Velocity dependson the time scale!

2. Rec A - or SSB protein – inducedtranslocation of DNA through ananopore � random force landscape
cis
trans
Cis side trans side
MEMBRANE
m monomers translocated
− +1. ssDNA translocation through a nanopore � random energylandscape
Does sequence heterogeneity affect the dynamics of even simplermodels of molecular “motors”?

Pauses and jumps in molecular machines…
Pauses were actually ‘edited out’of single molecule experimentson RNA polymerase in an attemptto obtain a well-defined velocity!!
Is the velocity even well defined??
T. T. Perkins et.al., Science 301, 1914 (2003)In later experiments on lambda exonuclease, the velocity clearly depends on time scale!!!
Note scale-dependentvelocities…
M. D. Wang et. al., Science 282, 902 (1998)
Low load
high load