Embed Size (px)
Citation preview

IntroductionCA125 is a tumor antigen that was defined by the monoclonalantibody (mAb) OC125 more than 20 years ago (Bast et al.,1981). On the basis of an immunological serum assay (Bast etal., 1983; Bon et al., 1996), the CA125 antigen has beendefined as a marker primarily for ovarian carcinoma (Davis etal., 1986; Lloyd and Yin, 2001). Although CA125 is expressedboth by normal and tumor cells (O’Brien et al., 1986; Zurawskiet al., 1988; Hardardottir et al., 1990; Nap et al., 1996), cell-surface expression and release of soluble proteolytic fragmentsof CA125 into the extracellular space (Lloyd and Yin, 2001)appear to be associated with the conversion from benign tocancer cells (Meyer and Rustin, 2000). Consistently, CA125has been shown to accumulate in the serum of cancer patientsbearing ovarian as well as other carcinomas (Bast et al., 1983;Bon et al., 1996). However, the biological function of CA125remains to be established.
Only recently was the primary structure of CA125elucidated, demonstrating that CA125 represents a giantmucin-like glycoprotein (O’Brien et al., 2001; Yin and Lloyd,2001). On this basis, CA125 has been termed Muc16 to reflectthe nature of CA125 as a new member of the protein family ofmucins (Yin and Lloyd, 2001). Full-length CA125 containsmore than 11,000 amino acids that form the proteinaceous core
structure (O’Brien et al., 2001; Yin and Lloyd, 2001). CA125is both N- and O-glycosylated in its N-terminal extracellulardomain (Zurawski et al., 1988; Nagata et al., 1991; Lloyd etal., 1997; Lloyd and Yin, 2001), which is composed of a stalkdomain next to the transmembrane span, more than 60 repeatstructures (each of which consists of 156 amino acids) and anN-terminal extension (O’Brien et al., 2001; Yin and Lloyd,2001). Towards the C-terminus, CA125 contains a putativetransmembrane span and a short cytoplasmic tail (O’Brien etal., 2001). The release of soluble fragments of CA125 into theextracellular space appears to be triggered by serine/threonin-and/or tyrosine-dependent phosphorylation within thecytoplasmic domain (Fendrick et al., 1997; Lloyd and Yin,2001).
Tumor-specific cell-surface expression concomitant with therelease of extracellular fragments suggests a role for CA125 inthe regulation of cell proliferation and/or tumor progression.Owing to its nature as a glycoprotein, potential ligands ofCA125 include lectins of the extracellular matrix (ECM) suchas the β-galactoside-specific family of galectins (Perillo et al.,1998; Rabinovich et al., 2002). Intriguingly, galectin-1 andgalectin-3 expression is upregulated in various cancer-derivedcell lines compared with benign tissue (Iurisci et al., 2000;Lahm et al., 2001; Lloyd, 2001; Armstrong et al., 2002).
1305
CA125 is an ovarian cancer antigen whose recentlyelucidated primary structure suggests that CA125 is a giantmucin-like glycoprotein present on the cell surface oftumor cells. Here, we establish a functional link betweenCA125 and β-galactoside-binding, cell-surface lectins,which are components of the extracellular matriximplicated in the regulation of cell adhesion, apoptosis, cellproliferation and tumor progression. On the basis of massspectrometry and immunological analyses, we find thatCA125 is a counter receptor for galectin-1, as both solubleand membrane-associated fragments of CA125 derivedfrom HeLa cell lysates are shown to bind specifically tohuman galectin-1 with high efficiency. This interaction isdemonstrated (1) to depend on β-galactose-terminated,O-linked oligosaccharide chains of CA125, (2) to bepreferential for galectin-1 versus galectin-3 and (3) to beregulated by the cellular background in which CA125 isexpressed. Despite lacking a conventional signal peptide,a CA125 C-terminal fragment of 1148 amino acids,
representing less than 10% of the full-length protein,retains the ability to integrate into secretory membranessuch as the endoplasmic reticulum (ER) and the Golgi, andis targeted to the plasma membrane by conventionalsecretory transport. As demonstrated by a novel assay thatreconstitutes non-conventional secretion of galectin-1 basedon fluorescence-activated cell sorting (FACS), we find thattumor-derived HeLa cells expressing endogenous CA125present more than ten times as much galectin-1 on theirsurface compared with non-tumor-derived, CA125-deficient CHO cells. Intriguingly, both the galectin-1expression level and the cell-surface binding capacity forgalectin-1 are shown to be similar in CHO and HeLa cells,suggesting that CA125 might be a factor involved in theregulation of galectin-1 export to the cell surface.
Key words: CA125, Galectin, Counter receptor, Nonclassical export,Cell-surface expression, Tumor antigen
Summary
The cancer antigen CA125 represents a novel counterreceptor for galectin-1Claudia Seelenmeyer*, Sabine Wegehingel*, Johannes Lechner and Walter Nickel ‡
Biochemie-Zentrum Heidelberg (BZH), Im Neuenheimer Feld 328, 69120 Heidelberg, Germany*These authors contributed equally to this work‡Author for correspondence (e-mail: [email protected])
Accepted 9 December 2002Journal of Cell Science 116, 1305-1318 © 2003 The Company of Biologists Ltddoi:10.1242/jcs.00312
Research Article

1306
Moreover, one member of this family, galectin-3, has beenreported to represent a ligand of a colon cancer mucin(Bresalier et al., 1996).
At the molecular level, the best-characterized members ofthis protein family are galectin-1 and -3 (Hughes, 1997;Hughes, 1999). Owing to its dimeric character, cell-surfacerecruitment of galectin-1 is thought to affect the conformationand oligomeric status of glycosylated protein domains byforming intra- or intermolecular bridges that, in turn, mightexert a cellular response (Perillo et al., 1998). Biologicalcounter receptors for galectin-1 include laminin, fibronectin,lamp 1 and 2, GM1 glycolipid (reviewed by Perillo et al., 1998)as well as cell-type-specific molecules such as the T-cellglycoproteins CD43 and CD45 (Perillo et al., 1995; Nguyen etal., 2001). The pattern of oligosaccharide chains presented onthe cell surface of individual cells is likely to influence the waygalectin-1 interacts with their surface. Therefore, the way aparticular cell type responds to galectin-1 might also beregulated by variations in the activity of glycosyl transferasesand/or glycosidases (Perillo et al., 1998). Known cellularresponses to the cell-surface recruitment of galectin-1 includea change in proliferation activity, regulation of cell survival andregulation of cell adhesion. Interestingly, depending both onthe cellular context and its local concentration, galectin-1exerts both inhibitory and stimulatory effects on theseprocesses (Perillo et al., 1998).
Galectin export from mammalian cells has been shown tooccur in a nonclassical manner independent of the function ofthe endoplasmic reticulum (ER) and the Golgi (Cooper andBarondes, 1990; Cleves et al., 1996). Consistently, galectinslack a conventional signal peptide for translocation into theER (Cleves, 1997; Hughes, 1999). The balance betweencytoplasmic and extracellular populations appears to be tightlyregulated (Hughes, 1999). For example, galectin-1 export frommuscle cells is developmentally regulated as increased exportis observed upon differentiation from myoblasts to myotubes(Cooper and Barondes, 1990). Moreover, it has been reportedthat galectin-1 externalization can be triggered upondifferentiation of K562 leukemia-derived cells induced byerythropoietin (Lutomski et al., 1997).
In the current study, we identify CA125 as a novel counterreceptor for galectin-1. Using affinity chromatography andmass spectrometry, as well as immunological analyses,galectin-1 is shown to bind specifically to CA125 in a directmanner. A comparison with the second most-abundant familymember, galectin-3, indicates that CA125 exhibits specificitytowards galectin-1. A C-terminal fragment of CA125,CA125-C-TERM (defined by NCBI clone AK024365) retainsthe ability to integrate into secretory membranes and, likefull-length CA125, is shown to be transported to the cellsurface. Cell-surface delivery of CA125-C-TERM isdemonstrated to occur by ER/Golgi-dependent vesiculartransport. CA125-C-TERM is found to bind to galectin-1with twofold higher efficiency compared with galectin-3when expressed in HeLa cells, and with sevenfold higherefficiency compared with galectin-3 when expressed in CHOcells. These results demonstrate that CA125 represents anovel counter receptor for galectins and has bindingcharacteristics that can be regulated by the cellularbackground in which it is expressed.
In order to investigate the functional significance of the
interaction reported, we compared tumor-derived, CA125-expressing HeLa cells with non-tumor-derived, CA125-deficient CHO cells with regard to various galectin-1parameters. Although we find that galectin-1 expression andcell-surface binding capacity for galectin-1 is similar in HeLaand CHO cells, we demonstrate that HeLa cells contain morethan ten times as much cell-surface galectin-1 compared withCHO cells. Our results suggest that CA125 might allow tumorcells to interact differentially with the ECM in a galectin-1-dependent manner.
Materials and MethodsRecombinant proteins and antibodiesGST–galectin-1 and GST–galectin-3 fusion constructs were clonedusing the vector pGEX-2T (Amersham). For this purpose, appropriatePCR products were generated using the IMAGE clones 2666528 and2419761 as a source for the ORFs (ORF) of human galectin-1 and-3, respectively. The corresponding proteins were expressed inEscherichia coliBL21(DE3) cells (Novagen). Protein purification wasachieved by affinity chromatography using GSH-sepharose(Amersham) according to standard procedures. An N-terminalfragment of CA125-C-TERM (defined by the NCBI cloneAK024365) that corresponds to amino acids 1-356 (CA125-C-TERM1-356) was cloned into the vector pIVEX 2.4b Nde (Roche)in order to express a His6-tagged version of the protein in vitroemploying the rapid translation system (Roche). Homogenouspreparations of GST–galectin-1 and His6-CA125-C-TERM1-356,respectively, were used to generate polyclonal antisera in rabbits.Anti-galectin-1 antibodies were affinity-purified from thecorresponding rabbit serum in two steps: the serum was first incubatedwith GST beads to remove anti-GST antibodies, followed by affinitypurification of anti-galectin-1 antibodies on GST–galectin-1 beads.Anti-galectin-1 antibodies were eluted from the GST–galectin-1affinity matrix under acidic conditions according to standardprocedures. Affinity-purified anti-GST antibodies were eluted underacidic conditions from GST beads obtained in step 1.
The mAb anti-CA125 antibody OC125 was purchased fromZymed. Secondary antibodies used for western blotting werefrom Bio-Rad, those used for FACS sorting and indirectimmunofluorescence confocal microscopy were from MolecularProbes.
Galectin-1 affinity matrix and binding experiments employingsubcellular fractions from HeLa cellsTo conduct affinity purification of galectin-1-interacting proteins,GST–galectin-1 and GST–galectin-3 fusion proteins, as well as GSTas a control, were expressed in E. coli BL21(DE3) cells. Cells wereresuspended in PBS containing 1 mM DTT, 0.1% (w/v) Triton X-100,10% (w/v) glycerol and protease inhibitor tabs (Roche), followed byhomogenization using a cell disruptor (Avestin). A 100,000 gavsupernatant was obtained and incubated with glutathione beads for 2hours at 4°C on a rotating wheel. Following extensive washing usinghomogenization buffer (see above), 250 µl of beads containing 250µg of coupled protein were used per binding experiment.
S-HeLa cells (ATCC CCL-2.2) were cultured in spinner flasksaccording to standard procedures. Typically, cultures were grown toa density of about 6-7×105 cells per ml. Cells were collected bycentrifugation and resuspended in HeLa homogenization buffer (25mM Tris, pH 7.5; 130 mM KCl; protease inhibitor tabs) at 1 g cellsper ml. Following cell breakage using a Balch homogenizer (Balchand Rothman, 1985), the homogenate was sequentially centrifugedtwice at 1000 gav and twice at 3500 gav. The resulting supernatant wassubjected to centrifugation at 100,000 gav. Following separation ofsupernatant and sediment, the soluble fraction was diluted with PBS
Journal of Cell Science 116 (7)

1307CA125 is a galectin-1 receptor
(supplemented with 1 mM DTT and protease inhibitor tabs) to give afinal protein concentration of about 0.25 mg/ml. Typically, whenstarting with 5 g of cells, the soluble fraction was adjusted to a finalvolume of 50 ml. The corresponding sediment was then resuspendedin 50 ml of PBS supplemented with 1 mM DTT, protease inhibitortabs and 1% (w/v) NP-40 (Roche). Per experimental condition, 25 mlof the soluble or the membrane fraction, respectively, were incubatedwith 250 µl of GSH beads containing about 250 µg GST–galectin-1,GST–galectin-3 or GST, respectively. Bound proteins were elutedsequentially with 100 mM lactose and 25 mM glutathione,respectively. Further details are given in the corresponding figurelegends.
Protein identification employing MALDI-TOF massspectrometryIn order to identify individual proteins eluted from the galectin-1affinity matrix, the eluates were separated on 10% Novex Bis-Tris gels(Invitrogen) followed by protein staining using the SilverQuest system(Invitrogen). After excision of gel pieces containing individualproteins, in-gel trypsin digestion allowed extraction of trypticpeptides. Proteins were identified based on the masses of the peptidesobtained in this way by employing MALDI-TOF mass spectrometry(Wilm et al., 1996).
Retroviral transduction of MCAT-expressing HeLa and CHOcellsIn order to transduce target cells with the cDNA of CA125-C-TERM,the ORF was cloned into the vector pFB (Stratagene). This vectorpromotes constitutive expression of the cDNA in question. Retroviralparticles were generated by conventional transfection of HEK293Tcells employing the VPack vector system (Stratagene). In this context,an envelope protein with an ecotropic host range was used, encodedby the vector pVPack-eco (Stratagene). Retroviral particles harvestedfrom the medium of triple-transfected HEK293T cells were added tothe medium of the target cells. As target cells, HeLa and CHO cellswere used that stably express the murine cation amino acid transporterMCAT-1 (Albritton et al., 1989; Davey et al., 1997) and a doxicycline-dependent transactivator (Urlinger et al., 2000). These cell lineswere designated HeLaMCAT-TAM2 and CHOMCAT-TAM2, respectively(Engling et al., 2002).
FACSHeLaMCAT-TAM2 and CHOMCAT-TAM2 were cultured according tostandard procedures. Further details are given in the correspondingfigure legends. In order to detach the cells from the culture plateswithout using protease-based protocols, cell-dissociation buffer(Life Technologies) was used to generate a cell suspension devoidof cell aggregates. Cell-surface antigens were detected with theprimary antibodies indicated, followed by decoration withsecondary antibodies coupled to either Alexa-488 orallophycocyanine (APC; Molecular Probes), respectively. Antibodyincubations were performed on a rotating wheel for 1 hour at 4°C(primary antibody) and 30 minutes at 4°C (secondary antibody),respectively. Wash procedures were carried out by sedimenting thecells at 200 g for 3 minutes at 4°C. Where indicated, propidiumiodide (1 µg/ml) was added prior to the FACS analysis in order todetect damaged cells.
Flow cytometric measurements were performed using a BectonDickinson FACSCalibur system. Autofluorescence was determined bymeasuring trypsinized cells that were otherwise treated identicallycompared with the positive controls. Alexa-488-derived and APC-derived fluorescence can be measured simultaneously on aFACSCalibur two-laser system without the need of channelcompensation.
Confocal microscopyCells were grown on glass cover slips to about 75% confluency.Following a wash with PBS, the cells were further processed byparaformaldehyde fixation (3% w/v, 20 minutes at 4°C), with orwithout permeabilization, employing 0.5% (w/v) Triton X-100.Antibody processing was achieved as indicated in the correspondingfigure legends. For double-staining procedures, Alexa-488- andAlexa-546-coupled secondary antibodies (Molecular Probes) wereused in all experiments. The specimens were mounted in FluoromountG (Southern Biotechnology Associates) and viewed with a Zeiss LSM510 confocal microscope.
ResultsIdentification of CA125 as a galectin-1 counter receptorTo search for ligands of human galectin-1, we attached arecombinant GST–galectin-1 fusion protein to glutathionesepharose beads in order to prepare an affinity matrix. As asource for galectin-1-binding proteins, HeLa cells werefractionated into a soluble and a membrane fraction (seeabove). Following dilution to about 0.25 mg/ml of total protein,these fractions were incubated with the galectin-1 affinitymatrix. Bound proteins were eluted sequentially using lactose(Fig. 1A, lanes 1-4) and glutathione (Fig. 1A, lanes 5-8). Thisprocedure allowed us to collect proteins that interact with thegalectin-1 matrix in a galactose-dependent manner, followedby the elution of proteins bound to the matrix by a galactose-independent mechanism. As shown in Fig. 1A, severalspecifically bound proteins (some of which have been labeledwith an asterisk) could be identified in lanes 2 (GST–galectin-1 matrix; soluble HeLa fraction; elution with lactose), 4(GST–galectin-1 matrix; HeLa membrane fraction; elutionwith lactose), 6 (GST–galectin-1 matrix, soluble HeLafraction, elution with glutathione) and 8 (GST–galectin-1matrix, HeLa membrane fraction, elution with glutathione)when compared with the corresponding GST control matrices(lanes 1, 3, 5 and 7). GST (j; lanes 5 and 7) and GST–galectin-1 (m; lanes 6 and 8) were co-eluted upon treatment withglutathione. Using mass spectrometry, protein band #2 wasidentified as a chondroitin sulfate proteoglycan, band #3 wasidentified as the cell adhesion molecule L1-CAM, and bands 4and 5 were identified as galectin-1. These results suggest thatrecombinant galectin-1 and HeLa-derived galectin-1 form adimer that is disassembled in the presence of lactose.
In addition to these known interaction partners of galectin-1, we identified 16 tryptic peptides from protein band #1whose masses were consistent with corresponding trypticfragments of a potential ORF defined by cDNA cloneAK024365 (NCBI database; Fig. 1D, boxed sequencesindicate peptides identified by mass spectrometry). In orderto verify whether band #1 is the product of a genecorresponding to AK024365, we generated a polyclonalantiserum against an artificial recombinant proteincorresponding to the N-terminal part (amino acids 1-356;Mr: ~39 kDa) of AK024365. As shown in Fig. 1B,immunoreactive material with a broad high-molecular-weightmigration behavior was detected in lanes 2, 4, 6 and 8, whichcorrespond to the various eluates (see above) of theGST–galectin-1 affinity matrix. No signal could be observedunder control conditions. Binding of immunoreactivematerial to galectin-1 appeared to be mediated by agalactose–lectin interaction as more than 90% (determined by

1308
a quantitative analysis using Bio-Rad® QuantityOne®
Software) eluted upon treatment of the affinity matrix withlactose. About 80% of the total immunoreactive material wasrecovered from the soluble fraction, with the remainingpopulation derived from the membrane fraction.
More recently, the AK024365 ORF was found to representa C-terminal fragment of 1148 amino acids in length of agiant mucin-like glycoprotein (O’Brien et al., 2001; Yin andLloyd, 2001). Molecular cloning of the corresponding generevealed that this new mucin is identical to the ovarian cancerantigen CA125, a putative integral membrane protein presenton the cell surface of tumor cells that has originally beendefined by the mAb OC125 (Bast et al., 1981). Therefore, weanalyzed the various eluates from the galectin-1 affinity
matrix with regard to immunoreactivity based on OC125. Asshown in Fig. 1C, the pattern of immunoreactive bandsdetected with OC125 is strikingly similar to the patterndetected with the polyclonal anti-AK024365 antiserum (fromnow on referred to as anti-CA125-C-TERM1-356 antiserum)described above (Fig. 1B). Since CA125 was reported torepresent an integral membrane protein with a singlemembrane span that is cleaved in the extracellular domain inorder to release soluble fragments, we conclude that thepattern of immunoreactive bands eluted from the galectinaffinity matrix represents both soluble and membrane-anchored fragments of CA125. From now on, the 1148 aminoacid, C-terminal part of CA125 (defined by cDNA cloneAK024365) will be termed CA125-C-TERM.
Journal of Cell Science 116 (7)
D MPLFKNTSVSSLYSGCRLTLLRPEKDGAATRVDAVCTHRPDPKSPGLDRERLYWKLSQLTHGI TELGPYTLDRHSLYVNGFTHQSSMTTTRTPDTSTMHLATSRTPASLSGPTTASPLLVLFTI NFTI TNLRYEENMHHPGSRKFNTTERVLQGLLRPVFKNTSVGPLYSGCRLTLLRPKKDGAATKVDAI CTYRPDPKSPGLDREQLYWELSQLTHSI TELGPYTLDRDSLYVNGFTQRSSVPTTSI PGTPTVDLGTSGTPVSKPGPSAASPLLVLFTLNFTI TNLRYEENMQHPGSRKFNTTERVLQGLLRSLFKSTSVGPLYSGCRLTLLRPEKDGTATGVDAI CTHHPDPKSPRLDREQLYWELSQLTHNI TELGHYALDNDSLFVNGFTHRSSVSTTSTPGTPTVYLGASKTPASI FGPSAASHLL I LFTLNFTI TNLRYEENMWPGSRKFNTTERVLQGLLRPLFKNTSVGPLYSGSRLTLLRPEKDGEATGVDAI CTHRPDPTGPGLDREQLYLELSQLTHSI TELGPYTLDRDSLYVNGFTHRSSVPTTSTGVVSEEPFTLNFTI NNLRYMADMGQPGSLKFNI TDNVMKHLLSPLFQRSSLGARYTGCRVI ALRSVKNGAETRVDLLCTYLQPLSGPGLPI KQVFHELSQQTHGI TRLGPYSLDKDSLYLNGYNEPGLDEPPTTPKPATTFLPPLSEATTAMGYHLKTLTLNFTI SNLQYSPDMGKGSATFNSTEGVLQHLLRPLFQKSSMGPFYLGCQLI SLRPEKDGAATGVDTTCTYHPDPVGPGLDI QQLYWELSQLTHGVTQLGFYVLDRDSLFI NGYAPQNLSI RGEYQI NFHI VNWNLSNPDPTSSEYI TLLRDI QDKVTTLYKGSQLHDTFRFCLVTNLTMDSVLVTVKALFSSNLDPSLVEQVFLDKTLNASFHWLGSTYQLVDI HVTEMESSVYQPTSSSSTQHFYPNFTI TNLPYSQDKAQPGTTNYQRNKRNI EDALNQLFRNSSI KSYFSDCQVSTFRSVPNRHHTGVDSLCNFSPLARRVDRVAI YEEFLRMTRNGTQLQNFTLDRSSVLVDGYSPNRNEPLTGNSDLPFWAVI FI GLAGLLGLI TCLI CGVLVTTRRRKKEGEYNVQQQCPGYYQSHLDLEDLQ
Fig. 1. Identification ofCA125 as a counter receptorof galectin-1. (A) Affinitypurification of galectin-1-interacting proteins. Bothsoluble (lanes 1, 2, 5, 6) andmembrane (lanes 3, 4, 7, 8)fractions of HeLa cells wereincubated with eitherGST–galectin-1 beads(lanes 2, 4, 6, 8) or GSTbeads as a control (lanes 1,3, 5, 7). Bound proteinswere eluted sequentiallywith lactose (lanes 1-4) andglutathione (lanes 5-8),followed by separation onNovex NuPage 10% Bis-Tris gels. Protein bandswere visualized usingSilverQuest.(B) Immunoblot analysis ofthe proteins eluted from theGST–galectin-1 and GSTmatrices, respectively. Thefractions were loaded in thesame order as shown inpanel A. The anti-CA125antibody OC125 was usedas primary antibodyfollowed by detection byelectrochemiluminescence.(C) Immunoblot analysis asshown in panel B employingan anti-CA125-C-TERM1-
356 antiserum for thedetection of CA125-derivedfragments. (D) Amino acidsequence of CA125-C-TERM. Boxed sequencesindicate tryptic peptidesderived from band 1 (panelA, lane 2) as identified bymass spectrometry.
1309CA125 is a galectin-1 receptor
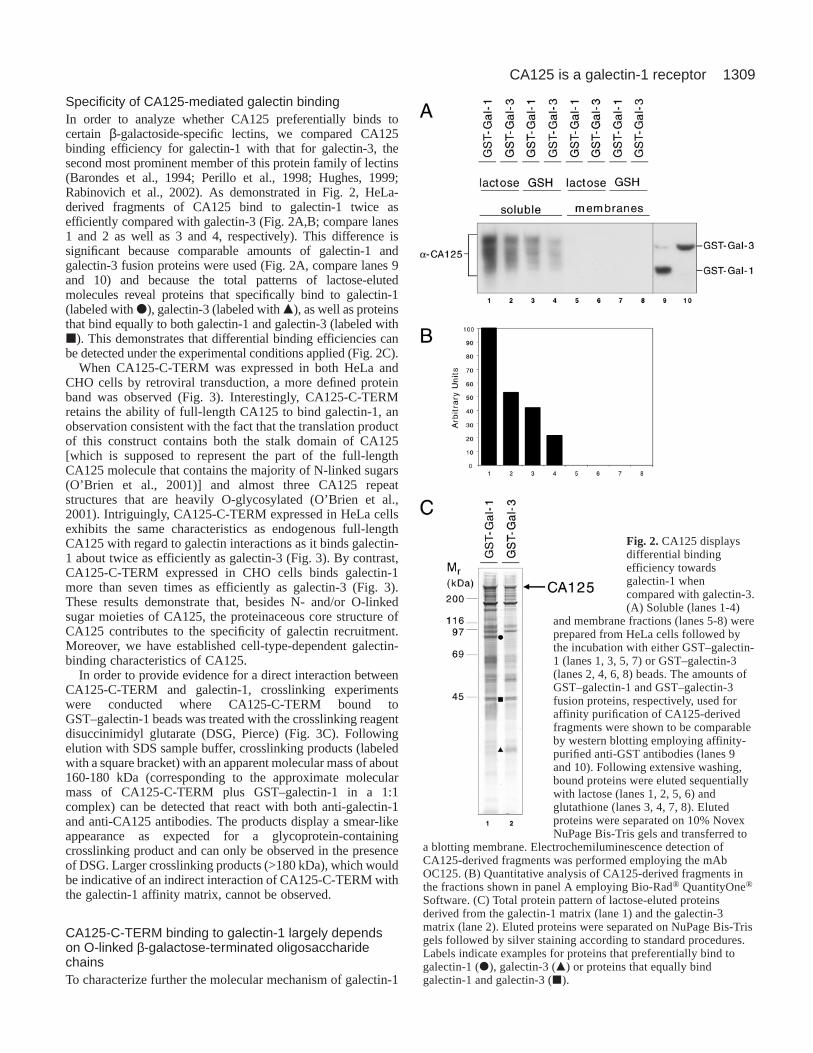
Specificity of CA125-mediated galectin bindingIn order to analyze whether CA125 preferentially binds tocertain β-galactoside-specific lectins, we compared CA125binding efficiency for galectin-1 with that for galectin-3, thesecond most prominent member of this protein family of lectins(Barondes et al., 1994; Perillo et al., 1998; Hughes, 1999;Rabinovich et al., 2002). As demonstrated in Fig. 2, HeLa-derived fragments of CA125 bind to galectin-1 twice asefficiently compared with galectin-3 (Fig. 2A,B; compare lanes1 and 2 as well as 3 and 4, respectively). This difference issignificant because comparable amounts of galectin-1 andgalectin-3 fusion proteins were used (Fig. 2A, compare lanes 9and 10) and because the total patterns of lactose-elutedmolecules reveal proteins that specifically bind to galectin-1(labeled with d), galectin-3 (labeled with m), as well as proteinsthat bind equally to both galectin-1 and galectin-3 (labeled withj). This demonstrates that differential binding efficiencies canbe detected under the experimental conditions applied (Fig. 2C).
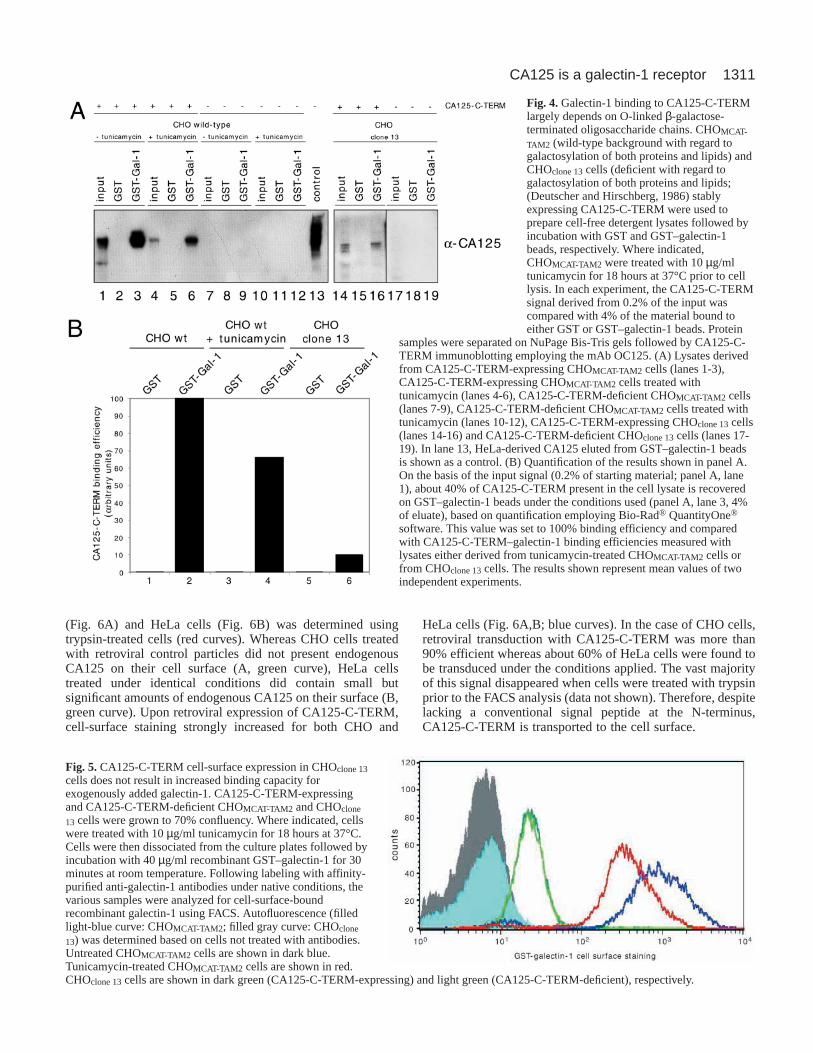
When CA125-C-TERM was expressed in both HeLa andCHO cells by retroviral transduction, a more defined proteinband was observed (Fig. 3). Interestingly, CA125-C-TERMretains the ability of full-length CA125 to bind galectin-1, anobservation consistent with the fact that the translation productof this construct contains both the stalk domain of CA125[which is supposed to represent the part of the full-lengthCA125 molecule that contains the majority of N-linked sugars(O’Brien et al., 2001)] and almost three CA125 repeatstructures that are heavily O-glycosylated (O’Brien et al.,2001). Intriguingly, CA125-C-TERM expressed in HeLa cellsexhibits the same characteristics as endogenous full-lengthCA125 with regard to galectin interactions as it binds galectin-1 about twice as efficiently as galectin-3 (Fig. 3). By contrast,CA125-C-TERM expressed in CHO cells binds galectin-1more than seven times as efficiently as galectin-3 (Fig. 3).These results demonstrate that, besides N- and/or O-linkedsugar moieties of CA125, the proteinaceous core structure ofCA125 contributes to the specificity of galectin recruitment.Moreover, we have established cell-type-dependent galectin-binding characteristics of CA125.
In order to provide evidence for a direct interaction betweenCA125-C-TERM and galectin-1, crosslinking experimentswere conducted where CA125-C-TERM bound toGST–galectin-1 beads was treated with the crosslinking reagentdisuccinimidyl glutarate (DSG, Pierce) (Fig. 3C). Followingelution with SDS sample buffer, crosslinking products (labeledwith a square bracket) with an apparent molecular mass of about160-180 kDa (corresponding to the approximate molecularmass of CA125-C-TERM plus GST–galectin-1 in a 1:1complex) can be detected that react with both anti-galectin-1and anti-CA125 antibodies. The products display a smear-likeappearance as expected for a glycoprotein-containingcrosslinking product and can only be observed in the presenceof DSG. Larger crosslinking products (>180 kDa), which wouldbe indicative of an indirect interaction of CA125-C-TERM withthe galectin-1 affinity matrix, cannot be observed.
CA125-C-TERM binding to galectin-1 largely dependson O-linked β-galactose-terminated oligosaccharidechainsTo characterize further the molecular mechanism of galectin-1
Fig. 2.CA125 displaysdifferential bindingefficiency towardsgalectin-1 whencompared with galectin-3.(A) Soluble (lanes 1-4)
and membrane fractions (lanes 5-8) wereprepared from HeLa cells followed bythe incubation with either GST–galectin-1 (lanes 1, 3, 5, 7) or GST–galectin-3(lanes 2, 4, 6, 8) beads. The amounts ofGST–galectin-1 and GST–galectin-3fusion proteins, respectively, used foraffinity purification of CA125-derivedfragments were shown to be comparableby western blotting employing affinity-purified anti-GST antibodies (lanes 9and 10). Following extensive washing,bound proteins were eluted sequentiallywith lactose (lanes 1, 2, 5, 6) andglutathione (lanes 3, 4, 7, 8). Elutedproteins were separated on 10% NovexNuPage Bis-Tris gels and transferred to
a blotting membrane. Electrochemiluminescence detection ofCA125-derived fragments was performed employing the mAbOC125. (B) Quantitative analysis of CA125-derived fragments inthe fractions shown in panel A employing Bio-Rad® QuantityOne®
Software. (C) Total protein pattern of lactose-eluted proteinsderived from the galectin-1 matrix (lane 1) and the galectin-3matrix (lane 2). Eluted proteins were separated on NuPage Bis-Trisgels followed by silver staining according to standard procedures.Labels indicate examples for proteins that preferentially bind togalectin-1 (d), galectin-3 (m) or proteins that equally bindgalectin-1 and galectin-3 (j).
1310
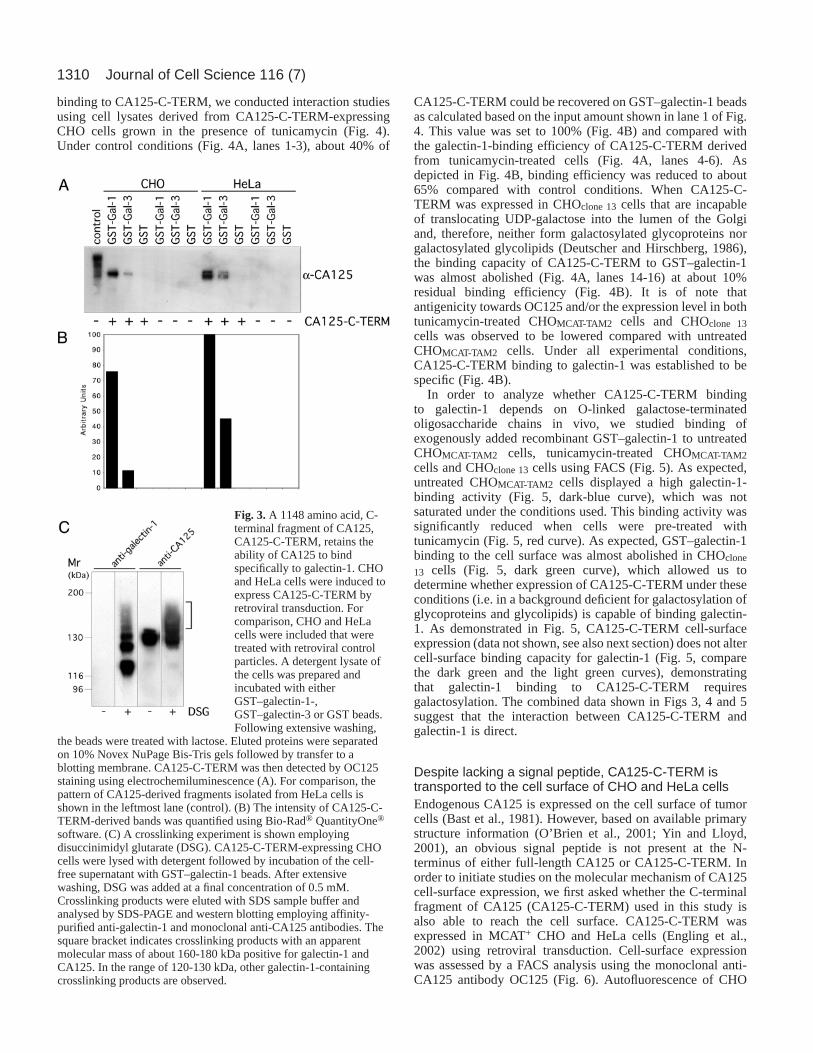
binding to CA125-C-TERM, we conducted interaction studiesusing cell lysates derived from CA125-C-TERM-expressingCHO cells grown in the presence of tunicamycin (Fig. 4).Under control conditions (Fig. 4A, lanes 1-3), about 40% of
CA125-C-TERM could be recovered on GST–galectin-1 beadsas calculated based on the input amount shown in lane 1 of Fig.4. This value was set to 100% (Fig. 4B) and compared withthe galectin-1-binding efficiency of CA125-C-TERM derivedfrom tunicamycin-treated cells (Fig. 4A, lanes 4-6). Asdepicted in Fig. 4B, binding efficiency was reduced to about65% compared with control conditions. When CA125-C-TERM was expressed in CHOclone 13cells that are incapableof translocating UDP-galactose into the lumen of the Golgiand, therefore, neither form galactosylated glycoproteins norgalactosylated glycolipids (Deutscher and Hirschberg, 1986),the binding capacity of CA125-C-TERM to GST–galectin-1was almost abolished (Fig. 4A, lanes 14-16) at about 10%residual binding efficiency (Fig. 4B). It is of note thatantigenicity towards OC125 and/or the expression level in bothtunicamycin-treated CHOMCAT-TAM2 cells and CHOclone 13cells was observed to be lowered compared with untreatedCHOMCAT-TAM2 cells. Under all experimental conditions,CA125-C-TERM binding to galectin-1 was established to bespecific (Fig. 4B).
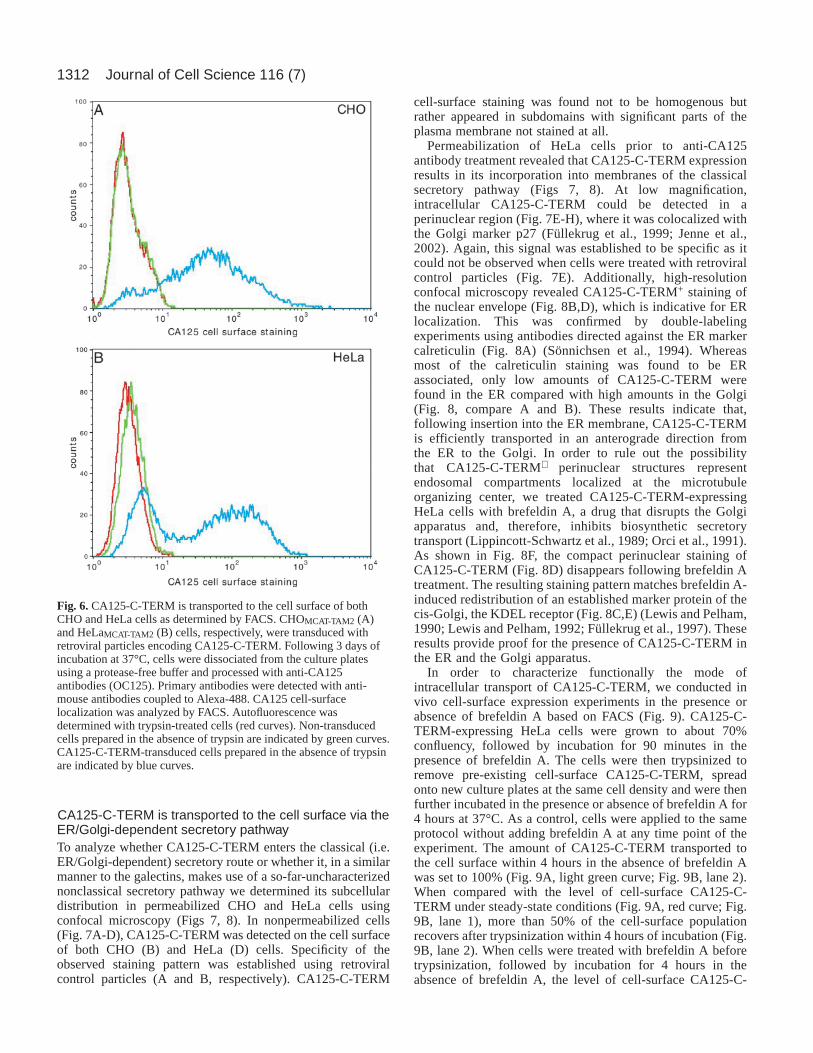
In order to analyze whether CA125-C-TERM bindingto galectin-1 depends on O-linked galactose-terminatedoligosaccharide chains in vivo, we studied binding ofexogenously added recombinant GST–galectin-1 to untreatedCHOMCAT-TAM2 cells, tunicamycin-treated CHOMCAT-TAM2cells and CHOclone 13cells using FACS (Fig. 5). As expected,untreated CHOMCAT-TAM2 cells displayed a high galectin-1-binding activity (Fig. 5, dark-blue curve), which was notsaturated under the conditions used. This binding activity wassignificantly reduced when cells were pre-treated withtunicamycin (Fig. 5, red curve). As expected, GST–galectin-1binding to the cell surface was almost abolished in CHOclone
13 cells (Fig. 5, dark green curve), which allowed us todetermine whether expression of CA125-C-TERM under theseconditions (i.e. in a background deficient for galactosylation ofglycoproteins and glycolipids) is capable of binding galectin-1. As demonstrated in Fig. 5, CA125-C-TERM cell-surfaceexpression (data not shown, see also next section) does not altercell-surface binding capacity for galectin-1 (Fig. 5, comparethe dark green and the light green curves), demonstratingthat galectin-1 binding to CA125-C-TERM requiresgalactosylation. The combined data shown in Figs 3, 4 and 5suggest that the interaction between CA125-C-TERM andgalectin-1 is direct.
Despite lacking a signal peptide, CA125-C-TERM istransported to the cell surface of CHO and HeLa cellsEndogenous CA125 is expressed on the cell surface of tumorcells (Bast et al., 1981). However, based on available primarystructure information (O’Brien et al., 2001; Yin and Lloyd,2001), an obvious signal peptide is not present at the N-terminus of either full-length CA125 or CA125-C-TERM. Inorder to initiate studies on the molecular mechanism of CA125cell-surface expression, we first asked whether the C-terminalfragment of CA125 (CA125-C-TERM) used in this study isalso able to reach the cell surface. CA125-C-TERM wasexpressed in MCAT+ CHO and HeLa cells (Engling et al.,2002) using retroviral transduction. Cell-surface expressionwas assessed by a FACS analysis using the monoclonal anti-CA125 antibody OC125 (Fig. 6). Autofluorescence of CHO
Journal of Cell Science 116 (7)
Fig. 3.A 1148 amino acid, C-terminal fragment of CA125,CA125-C-TERM, retains theability of CA125 to bindspecifically to galectin-1. CHOand HeLa cells were induced toexpress CA125-C-TERM byretroviral transduction. Forcomparison, CHO and HeLacells were included that weretreated with retroviral controlparticles. A detergent lysate ofthe cells was prepared andincubated with eitherGST–galectin-1-,GST–galectin-3 or GST beads.Following extensive washing,
the beads were treated with lactose. Eluted proteins were separatedon 10% Novex NuPage Bis-Tris gels followed by transfer to ablotting membrane. CA125-C-TERM was then detected by OC125staining using electrochemiluminescence (A). For comparison, thepattern of CA125-derived fragments isolated from HeLa cells isshown in the leftmost lane (control). (B) The intensity of CA125-C-TERM-derived bands was quantified using Bio-Rad® QuantityOne®
software. (C) A crosslinking experiment is shown employingdisuccinimidyl glutarate (DSG). CA125-C-TERM-expressing CHOcells were lysed with detergent followed by incubation of the cell-free supernatant with GST–galectin-1 beads. After extensivewashing, DSG was added at a final concentration of 0.5 mM.Crosslinking products were eluted with SDS sample buffer andanalysed by SDS-PAGE and western blotting employing affinity-purified anti-galectin-1 and monoclonal anti-CA125 antibodies. Thesquare bracket indicates crosslinking products with an apparentmolecular mass of about 160-180 kDa positive for galectin-1 andCA125. In the range of 120-130 kDa, other galectin-1-containingcrosslinking products are observed.
1311CA125 is a galectin-1 receptor
(Fig. 6A) and HeLa cells (Fig. 6B) was determined usingtrypsin-treated cells (red curves). Whereas CHO cells treatedwith retroviral control particles did not present endogenousCA125 on their cell surface (A, green curve), HeLa cellstreated under identical conditions did contain small butsignificant amounts of endogenous CA125 on their surface (B,green curve). Upon retroviral expression of CA125-C-TERM,cell-surface staining strongly increased for both CHO and
HeLa cells (Fig. 6A,B; blue curves). In the case of CHO cells,retroviral transduction with CA125-C-TERM was more than90% efficient whereas about 60% of HeLa cells were found tobe transduced under the conditions applied. The vast majorityof this signal disappeared when cells were treated with trypsinprior to the FACS analysis (data not shown). Therefore, despitelacking a conventional signal peptide at the N-terminus,CA125-C-TERM is transported to the cell surface.
Fig. 4.Galectin-1 binding to CA125-C-TERMlargely depends on O-linked β-galactose-terminated oligosaccharide chains. CHOMCAT-
TAM2 (wild-type background with regard togalactosylation of both proteins and lipids) andCHOclone 13cells (deficient with regard togalactosylation of both proteins and lipids;(Deutscher and Hirschberg, 1986) stablyexpressing CA125-C-TERM were used toprepare cell-free detergent lysates followed byincubation with GST and GST–galectin-1beads, respectively. Where indicated,CHOMCAT-TAM2 were treated with 10 µg/mltunicamycin for 18 hours at 37°C prior to celllysis. In each experiment, the CA125-C-TERMsignal derived from 0.2% of the input wascompared with 4% of the material bound toeither GST or GST–galectin-1 beads. Protein
samples were separated on NuPage Bis-Tris gels followed by CA125-C-TERM immunoblotting employing the mAb OC125. (A) Lysates derivedfrom CA125-C-TERM-expressing CHOMCAT-TAM2 cells (lanes 1-3),CA125-C-TERM-expressing CHOMCAT-TAM2 cells treated withtunicamycin (lanes 4-6), CA125-C-TERM-deficient CHOMCAT-TAM2 cells(lanes 7-9), CA125-C-TERM-deficient CHOMCAT-TAM2 cells treated withtunicamycin (lanes 10-12), CA125-C-TERM-expressing CHOclone 13cells(lanes 14-16) and CA125-C-TERM-deficient CHOclone 13cells (lanes 17-19). In lane 13, HeLa-derived CA125 eluted from GST–galectin-1 beadsis shown as a control. (B) Quantification of the results shown in panel A.On the basis of the input signal (0.2% of starting material; panel A, lane1), about 40% of CA125-C-TERM present in the cell lysate is recoveredon GST–galectin-1 beads under the conditions used (panel A, lane 3, 4%of eluate), based on quantification employing Bio-Rad® QuantityOne®
software. This value was set to 100% binding efficiency and comparedwith CA125-C-TERM–galectin-1 binding efficiencies measured withlysates either derived from tunicamycin-treated CHOMCAT-TAM2 cells orfrom CHOclone 13cells. The results shown represent mean values of twoindependent experiments.
Fig. 5.CA125-C-TERM cell-surface expression in CHOclone 13cells does not result in increased binding capacity forexogenously added galectin-1. CA125-C-TERM-expressingand CA125-C-TERM-deficient CHOMCAT-TAM2 and CHOclone
13 cells were grown to 70% confluency. Where indicated, cellswere treated with 10 µg/ml tunicamycin for 18 hours at 37°C.Cells were then dissociated from the culture plates followed byincubation with 40 µg/ml recombinant GST–galectin-1 for 30minutes at room temperature. Following labeling with affinity-purified anti-galectin-1 antibodies under native conditions, thevarious samples were analyzed for cell-surface-boundrecombinant galectin-1 using FACS. Autofluorescence (filledlight-blue curve: CHOMCAT-TAM2; filled gray curve: CHOclone
13) was determined based on cells not treated with antibodies.Untreated CHOMCAT-TAM2 cells are shown in dark blue.Tunicamycin-treated CHOMCAT-TAM2 cells are shown in red.CHOclone 13cells are shown in dark green (CA125-C-TERM-expressing) and light green (CA125-C-TERM-deficient), respectively.
1312
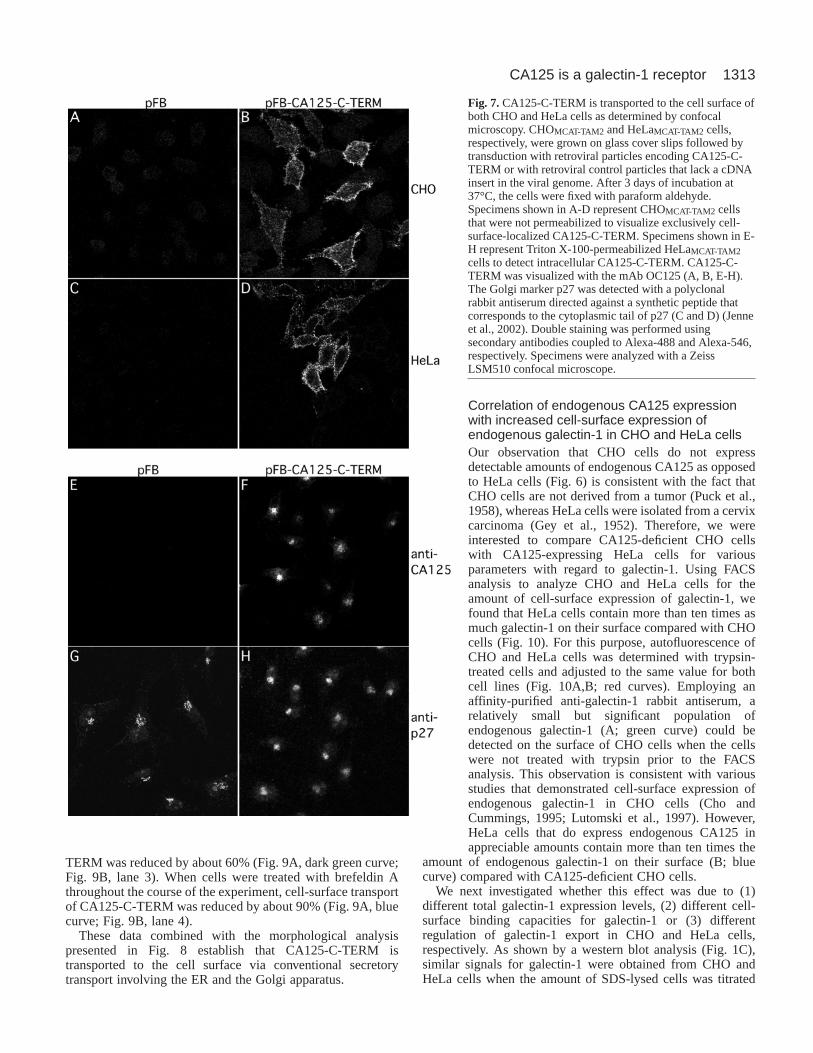
CA125-C-TERM is transported to the cell surface via theER/Golgi-dependent secretory pathwayTo analyze whether CA125-C-TERM enters the classical (i.e.ER/Golgi-dependent) secretory route or whether it, in a similarmanner to the galectins, makes use of a so-far-uncharacterizednonclassical secretory pathway we determined its subcellulardistribution in permeabilized CHO and HeLa cells usingconfocal microscopy (Figs 7, 8). In nonpermeabilized cells(Fig. 7A-D), CA125-C-TERM was detected on the cell surfaceof both CHO (B) and HeLa (D) cells. Specificity of theobserved staining pattern was established using retroviralcontrol particles (A and B, respectively). CA125-C-TERM
cell-surface staining was found not to be homogenous butrather appeared in subdomains with significant parts of theplasma membrane not stained at all.
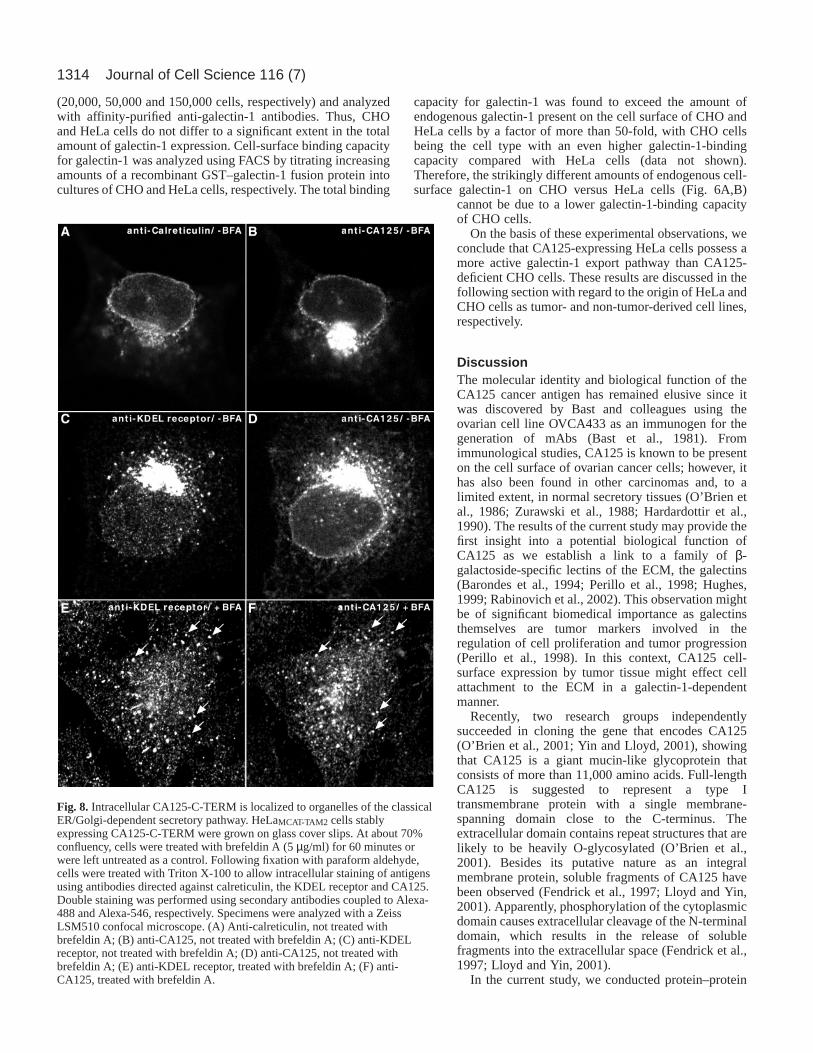
Permeabilization of HeLa cells prior to anti-CA125antibody treatment revealed that CA125-C-TERM expressionresults in its incorporation into membranes of the classicalsecretory pathway (Figs 7, 8). At low magnification,intracellular CA125-C-TERM could be detected in aperinuclear region (Fig. 7E-H), where it was colocalized withthe Golgi marker p27 (Füllekrug et al., 1999; Jenne et al.,2002). Again, this signal was established to be specific as itcould not be observed when cells were treated with retroviralcontrol particles (Fig. 7E). Additionally, high-resolutionconfocal microscopy revealed CA125-C-TERM+ staining ofthe nuclear envelope (Fig. 8B,D), which is indicative for ERlocalization. This was confirmed by double-labelingexperiments using antibodies directed against the ER markercalreticulin (Fig. 8A) (Sönnichsen et al., 1994). Whereasmost of the calreticulin staining was found to be ERassociated, only low amounts of CA125-C-TERM werefound in the ER compared with high amounts in the Golgi(Fig. 8, compare A and B). These results indicate that,following insertion into the ER membrane, CA125-C-TERMis efficiently transported in an anterograde direction fromthe ER to the Golgi. In order to rule out the possibilitythat CA125-C-TERM+ perinuclear structures representendosomal compartments localized at the microtubuleorganizing center, we treated CA125-C-TERM-expressingHeLa cells with brefeldin A, a drug that disrupts the Golgiapparatus and, therefore, inhibits biosynthetic secretorytransport (Lippincott-Schwartz et al., 1989; Orci et al., 1991).As shown in Fig. 8F, the compact perinuclear staining ofCA125-C-TERM (Fig. 8D) disappears following brefeldin Atreatment. The resulting staining pattern matches brefeldin A-induced redistribution of an established marker protein of thecis-Golgi, the KDEL receptor (Fig. 8C,E) (Lewis and Pelham,1990; Lewis and Pelham, 1992; Füllekrug et al., 1997). Theseresults provide proof for the presence of CA125-C-TERM inthe ER and the Golgi apparatus.
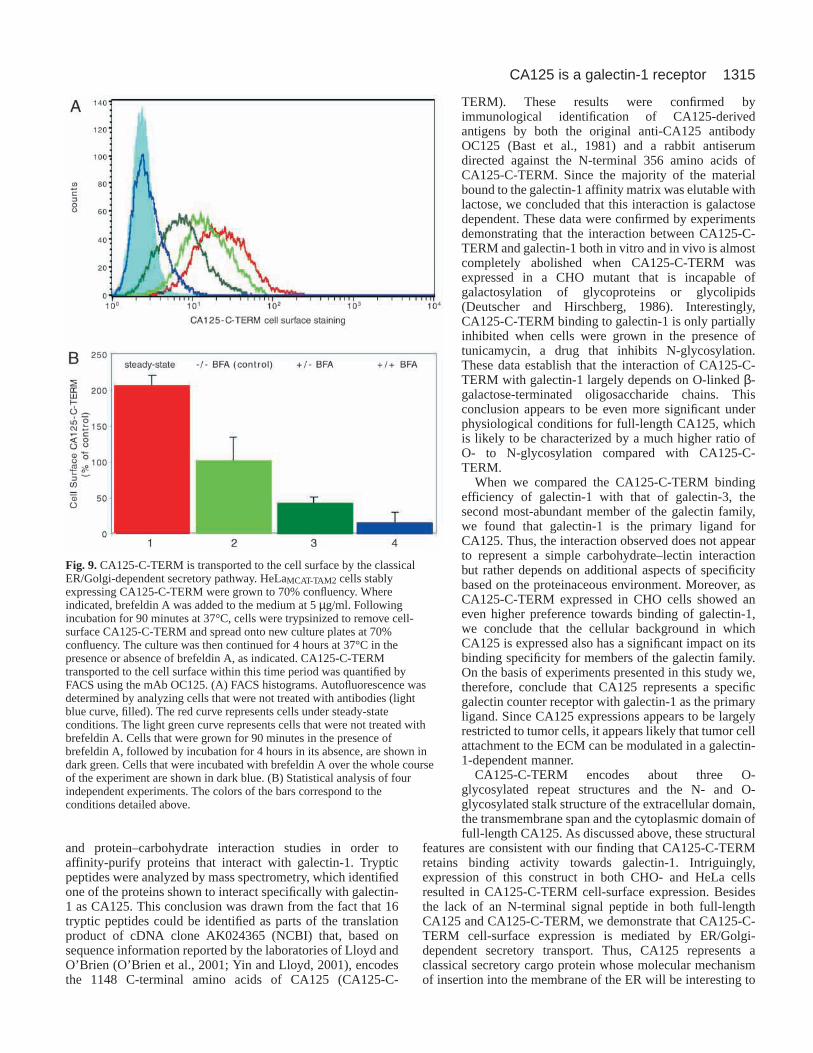
In order to characterize functionally the mode ofintracellular transport of CA125-C-TERM, we conducted invivo cell-surface expression experiments in the presence orabsence of brefeldin A based on FACS (Fig. 9). CA125-C-TERM-expressing HeLa cells were grown to about 70%confluency, followed by incubation for 90 minutes in thepresence of brefeldin A. The cells were then trypsinized toremove pre-existing cell-surface CA125-C-TERM, spreadonto new culture plates at the same cell density and were thenfurther incubated in the presence or absence of brefeldin A for4 hours at 37°C. As a control, cells were applied to the sameprotocol without adding brefeldin A at any time point of theexperiment. The amount of CA125-C-TERM transported tothe cell surface within 4 hours in the absence of brefeldin Awas set to 100% (Fig. 9A, light green curve; Fig. 9B, lane 2).When compared with the level of cell-surface CA125-C-TERM under steady-state conditions (Fig. 9A, red curve; Fig.9B, lane 1), more than 50% of the cell-surface populationrecovers after trypsinization within 4 hours of incubation (Fig.9B, lane 2). When cells were treated with brefeldin A beforetrypsinization, followed by incubation for 4 hours in theabsence of brefeldin A, the level of cell-surface CA125-C-
Journal of Cell Science 116 (7)
Fig. 6.CA125-C-TERM is transported to the cell surface of bothCHO and HeLa cells as determined by FACS. CHOMCAT-TAM2 (A)and HeLaMCAT-TAM2 (B) cells, respectively, were transduced withretroviral particles encoding CA125-C-TERM. Following 3 days ofincubation at 37°C, cells were dissociated from the culture platesusing a protease-free buffer and processed with anti-CA125antibodies (OC125). Primary antibodies were detected with anti-mouse antibodies coupled to Alexa-488. CA125 cell-surfacelocalization was analyzed by FACS. Autofluorescence wasdetermined with trypsin-treated cells (red curves). Non-transducedcells prepared in the absence of trypsin are indicated by green curves.CA125-C-TERM-transduced cells prepared in the absence of trypsinare indicated by blue curves.
1313CA125 is a galectin-1 receptor
TERM was reduced by about 60% (Fig. 9A, dark green curve;Fig. 9B, lane 3). When cells were treated with brefeldin Athroughout the course of the experiment, cell-surface transportof CA125-C-TERM was reduced by about 90% (Fig. 9A, bluecurve; Fig. 9B, lane 4).
These data combined with the morphological analysispresented in Fig. 8 establish that CA125-C-TERM istransported to the cell surface via conventional secretorytransport involving the ER and the Golgi apparatus.
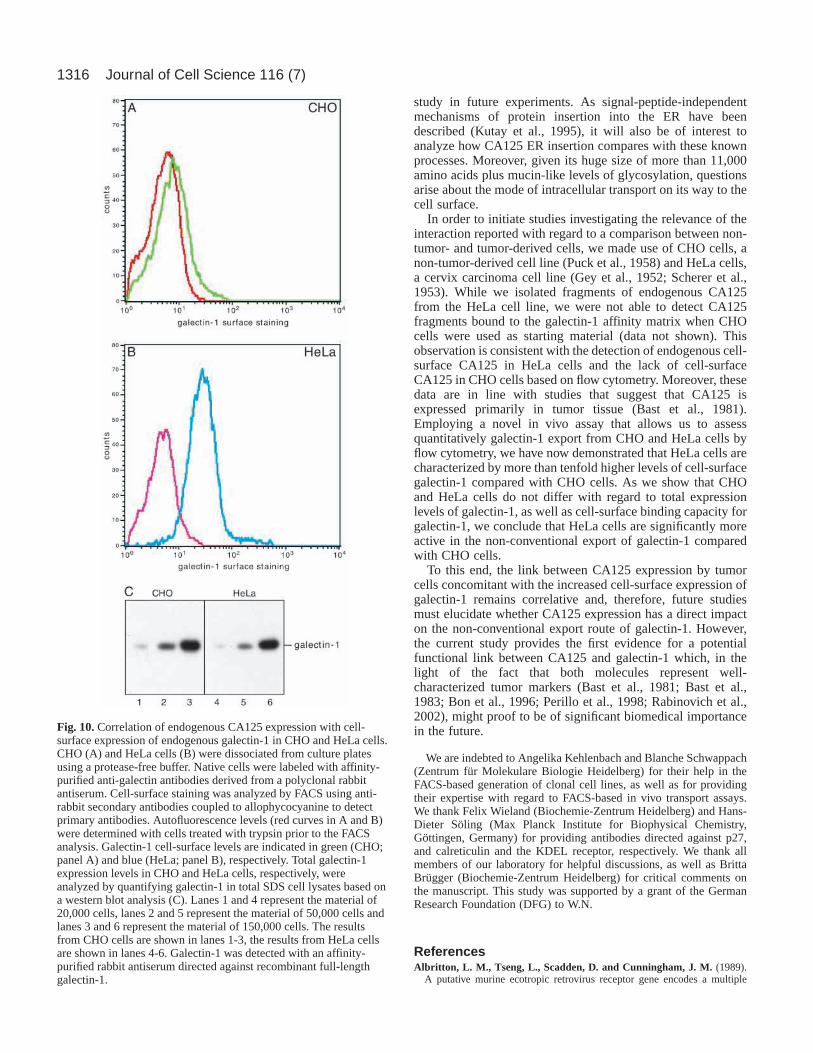
Correlation of endogenous CA125 expressionwith increased cell-surface expression ofendogenous galectin-1 in CHO and HeLa cellsOur observation that CHO cells do not expressdetectable amounts of endogenous CA125 as opposedto HeLa cells (Fig. 6) is consistent with the fact thatCHO cells are not derived from a tumor (Puck et al.,1958), whereas HeLa cells were isolated from a cervixcarcinoma (Gey et al., 1952). Therefore, we wereinterested to compare CA125-deficient CHO cellswith CA125-expressing HeLa cells for variousparameters with regard to galectin-1. Using FACSanalysis to analyze CHO and HeLa cells for theamount of cell-surface expression of galectin-1, wefound that HeLa cells contain more than ten times asmuch galectin-1 on their surface compared with CHOcells (Fig. 10). For this purpose, autofluorescence ofCHO and HeLa cells was determined with trypsin-treated cells and adjusted to the same value for bothcell lines (Fig. 10A,B; red curves). Employing anaffinity-purified anti-galectin-1 rabbit antiserum, arelatively small but significant population ofendogenous galectin-1 (A; green curve) could bedetected on the surface of CHO cells when the cellswere not treated with trypsin prior to the FACSanalysis. This observation is consistent with variousstudies that demonstrated cell-surface expression ofendogenous galectin-1 in CHO cells (Cho andCummings, 1995; Lutomski et al., 1997). However,HeLa cells that do express endogenous CA125 inappreciable amounts contain more than ten times the
amount of endogenous galectin-1 on their surface (B; bluecurve) compared with CA125-deficient CHO cells.
We next investigated whether this effect was due to (1)different total galectin-1 expression levels, (2) different cell-surface binding capacities for galectin-1 or (3) differentregulation of galectin-1 export in CHO and HeLa cells,respectively. As shown by a western blot analysis (Fig. 1C),similar signals for galectin-1 were obtained from CHO andHeLa cells when the amount of SDS-lysed cells was titrated
Fig. 7.CA125-C-TERM is transported to the cell surface ofboth CHO and HeLa cells as determined by confocalmicroscopy. CHOMCAT-TAM2 and HeLaMCAT-TAM2 cells,respectively, were grown on glass cover slips followed bytransduction with retroviral particles encoding CA125-C-TERM or with retroviral control particles that lack a cDNAinsert in the viral genome. After 3 days of incubation at37°C, the cells were fixed with paraform aldehyde.Specimens shown in A-D represent CHOMCAT-TAM2 cellsthat were not permeabilized to visualize exclusively cell-surface-localized CA125-C-TERM. Specimens shown in E-H represent Triton X-100-permeabilized HeLaMCAT-TAM2cells to detect intracellular CA125-C-TERM. CA125-C-TERM was visualized with the mAb OC125 (A, B, E-H).The Golgi marker p27 was detected with a polyclonalrabbit antiserum directed against a synthetic peptide thatcorresponds to the cytoplasmic tail of p27 (C and D) (Jenneet al., 2002). Double staining was performed usingsecondary antibodies coupled to Alexa-488 and Alexa-546,respectively. Specimens were analyzed with a ZeissLSM510 confocal microscope.
1314
(20,000, 50,000 and 150,000 cells, respectively) and analyzedwith affinity-purified anti-galectin-1 antibodies. Thus, CHOand HeLa cells do not differ to a significant extent in the totalamount of galectin-1 expression. Cell-surface binding capacityfor galectin-1 was analyzed using FACS by titrating increasingamounts of a recombinant GST–galectin-1 fusion protein intocultures of CHO and HeLa cells, respectively. The total binding
capacity for galectin-1 was found to exceed the amount ofendogenous galectin-1 present on the cell surface of CHO andHeLa cells by a factor of more than 50-fold, with CHO cellsbeing the cell type with an even higher galectin-1-bindingcapacity compared with HeLa cells (data not shown).Therefore, the strikingly different amounts of endogenous cell-surface galectin-1 on CHO versus HeLa cells (Fig. 6A,B)
cannot be due to a lower galectin-1-binding capacityof CHO cells.
On the basis of these experimental observations, weconclude that CA125-expressing HeLa cells possess amore active galectin-1 export pathway than CA125-deficient CHO cells. These results are discussed in thefollowing section with regard to the origin of HeLa andCHO cells as tumor- and non-tumor-derived cell lines,respectively.
DiscussionThe molecular identity and biological function of theCA125 cancer antigen has remained elusive since itwas discovered by Bast and colleagues using theovarian cell line OVCA433 as an immunogen for thegeneration of mAbs (Bast et al., 1981). Fromimmunological studies, CA125 is known to be presenton the cell surface of ovarian cancer cells; however, ithas also been found in other carcinomas and, to alimited extent, in normal secretory tissues (O’Brien etal., 1986; Zurawski et al., 1988; Hardardottir et al.,1990). The results of the current study may provide thefirst insight into a potential biological function ofCA125 as we establish a link to a family of β-galactoside-specific lectins of the ECM, the galectins(Barondes et al., 1994; Perillo et al., 1998; Hughes,1999; Rabinovich et al., 2002). This observation mightbe of significant biomedical importance as galectinsthemselves are tumor markers involved in theregulation of cell proliferation and tumor progression(Perillo et al., 1998). In this context, CA125 cell-surface expression by tumor tissue might effect cellattachment to the ECM in a galectin-1-dependentmanner.
Recently, two research groups independentlysucceeded in cloning the gene that encodes CA125(O’Brien et al., 2001; Yin and Lloyd, 2001), showingthat CA125 is a giant mucin-like glycoprotein thatconsists of more than 11,000 amino acids. Full-lengthCA125 is suggested to represent a type Itransmembrane protein with a single membrane-spanning domain close to the C-terminus. Theextracellular domain contains repeat structures that arelikely to be heavily O-glycosylated (O’Brien et al.,2001). Besides its putative nature as an integralmembrane protein, soluble fragments of CA125 havebeen observed (Fendrick et al., 1997; Lloyd and Yin,2001). Apparently, phosphorylation of the cytoplasmicdomain causes extracellular cleavage of the N-terminaldomain, which results in the release of solublefragments into the extracellular space (Fendrick et al.,1997; Lloyd and Yin, 2001).
In the current study, we conducted protein–protein
Journal of Cell Science 116 (7)
Fig. 8. Intracellular CA125-C-TERM is localized to organelles of the classicalER/Golgi-dependent secretory pathway. HeLaMCAT-TAM2 cells stablyexpressing CA125-C-TERM were grown on glass cover slips. At about 70%confluency, cells were treated with brefeldin A (5 µg/ml) for 60 minutes orwere left untreated as a control. Following fixation with paraform aldehyde,cells were treated with Triton X-100 to allow intracellular staining of antigensusing antibodies directed against calreticulin, the KDEL receptor and CA125.Double staining was performed using secondary antibodies coupled to Alexa-488 and Alexa-546, respectively. Specimens were analyzed with a ZeissLSM510 confocal microscope. (A) Anti-calreticulin, not treated withbrefeldin A; (B) anti-CA125, not treated with brefeldin A; (C) anti-KDELreceptor, not treated with brefeldin A; (D) anti-CA125, not treated withbrefeldin A; (E) anti-KDEL receptor, treated with brefeldin A; (F) anti-CA125, treated with brefeldin A.
1315CA125 is a galectin-1 receptor
and protein–carbohydrate interaction studies in order toaffinity-purify proteins that interact with galectin-1. Trypticpeptides were analyzed by mass spectrometry, which identifiedone of the proteins shown to interact specifically with galectin-1 as CA125. This conclusion was drawn from the fact that 16tryptic peptides could be identified as parts of the translationproduct of cDNA clone AK024365 (NCBI) that, based onsequence information reported by the laboratories of Lloyd andO’Brien (O’Brien et al., 2001; Yin and Lloyd, 2001), encodesthe 1148 C-terminal amino acids of CA125 (CA125-C-
TERM). These results were confirmed byimmunological identification of CA125-derivedantigens by both the original anti-CA125 antibodyOC125 (Bast et al., 1981) and a rabbit antiserumdirected against the N-terminal 356 amino acids ofCA125-C-TERM. Since the majority of the materialbound to the galectin-1 affinity matrix was elutable withlactose, we concluded that this interaction is galactosedependent. These data were confirmed by experimentsdemonstrating that the interaction between CA125-C-TERM and galectin-1 both in vitro and in vivo is almostcompletely abolished when CA125-C-TERM wasexpressed in a CHO mutant that is incapable ofgalactosylation of glycoproteins or glycolipids(Deutscher and Hirschberg, 1986). Interestingly,CA125-C-TERM binding to galectin-1 is only partiallyinhibited when cells were grown in the presence oftunicamycin, a drug that inhibits N-glycosylation.These data establish that the interaction of CA125-C-TERM with galectin-1 largely depends on O-linked β-galactose-terminated oligosaccharide chains. Thisconclusion appears to be even more significant underphysiological conditions for full-length CA125, whichis likely to be characterized by a much higher ratio ofO- to N-glycosylation compared with CA125-C-TERM.
When we compared the CA125-C-TERM bindingefficiency of galectin-1 with that of galectin-3, thesecond most-abundant member of the galectin family,we found that galectin-1 is the primary ligand forCA125. Thus, the interaction observed does not appearto represent a simple carbohydrate–lectin interactionbut rather depends on additional aspects of specificitybased on the proteinaceous environment. Moreover, asCA125-C-TERM expressed in CHO cells showed aneven higher preference towards binding of galectin-1,we conclude that the cellular background in whichCA125 is expressed also has a significant impact on itsbinding specificity for members of the galectin family.On the basis of experiments presented in this study we,therefore, conclude that CA125 represents a specificgalectin counter receptor with galectin-1 as the primaryligand. Since CA125 expressions appears to be largelyrestricted to tumor cells, it appears likely that tumor cellattachment to the ECM can be modulated in a galectin-1-dependent manner.
CA125-C-TERM encodes about three O-glycosylated repeat structures and the N- and O-glycosylated stalk structure of the extracellular domain,the transmembrane span and the cytoplasmic domain offull-length CA125. As discussed above, these structural
features are consistent with our finding that CA125-C-TERMretains binding activity towards galectin-1. Intriguingly,expression of this construct in both CHO- and HeLa cellsresulted in CA125-C-TERM cell-surface expression. Besidesthe lack of an N-terminal signal peptide in both full-lengthCA125 and CA125-C-TERM, we demonstrate that CA125-C-TERM cell-surface expression is mediated by ER/Golgi-dependent secretory transport. Thus, CA125 represents aclassical secretory cargo protein whose molecular mechanismof insertion into the membrane of the ER will be interesting to
Fig. 9.CA125-C-TERM is transported to the cell surface by the classicalER/Golgi-dependent secretory pathway. HeLaMCAT-TAM2 cells stablyexpressing CA125-C-TERM were grown to 70% confluency. Whereindicated, brefeldin A was added to the medium at 5 µg/ml. Followingincubation for 90 minutes at 37°C, cells were trypsinized to remove cell-surface CA125-C-TERM and spread onto new culture plates at 70%confluency. The culture was then continued for 4 hours at 37°C in thepresence or absence of brefeldin A, as indicated. CA125-C-TERMtransported to the cell surface within this time period was quantified byFACS using the mAb OC125. (A) FACS histograms. Autofluorescence wasdetermined by analyzing cells that were not treated with antibodies (lightblue curve, filled). The red curve represents cells under steady-stateconditions. The light green curve represents cells that were not treated withbrefeldin A. Cells that were grown for 90 minutes in the presence ofbrefeldin A, followed by incubation for 4 hours in its absence, are shown indark green. Cells that were incubated with brefeldin A over the whole courseof the experiment are shown in dark blue. (B) Statistical analysis of fourindependent experiments. The colors of the bars correspond to theconditions detailed above.
1316
study in future experiments. As signal-peptide-independentmechanisms of protein insertion into the ER have beendescribed (Kutay et al., 1995), it will also be of interest toanalyze how CA125 ER insertion compares with these knownprocesses. Moreover, given its huge size of more than 11,000amino acids plus mucin-like levels of glycosylation, questionsarise about the mode of intracellular transport on its way to thecell surface.
In order to initiate studies investigating the relevance of theinteraction reported with regard to a comparison between non-tumor- and tumor-derived cells, we made use of CHO cells, anon-tumor-derived cell line (Puck et al., 1958) and HeLa cells,a cervix carcinoma cell line (Gey et al., 1952; Scherer et al.,1953). While we isolated fragments of endogenous CA125from the HeLa cell line, we were not able to detect CA125fragments bound to the galectin-1 affinity matrix when CHOcells were used as starting material (data not shown). Thisobservation is consistent with the detection of endogenous cell-surface CA125 in HeLa cells and the lack of cell-surfaceCA125 in CHO cells based on flow cytometry. Moreover, thesedata are in line with studies that suggest that CA125 isexpressed primarily in tumor tissue (Bast et al., 1981).Employing a novel in vivo assay that allows us to assessquantitatively galectin-1 export from CHO and HeLa cells byflow cytometry, we have now demonstrated that HeLa cells arecharacterized by more than tenfold higher levels of cell-surfacegalectin-1 compared with CHO cells. As we show that CHOand HeLa cells do not differ with regard to total expressionlevels of galectin-1, as well as cell-surface binding capacity forgalectin-1, we conclude that HeLa cells are significantly moreactive in the non-conventional export of galectin-1 comparedwith CHO cells.
To this end, the link between CA125 expression by tumorcells concomitant with the increased cell-surface expression ofgalectin-1 remains correlative and, therefore, future studiesmust elucidate whether CA125 expression has a direct impacton the non-conventional export route of galectin-1. However,the current study provides the first evidence for a potentialfunctional link between CA125 and galectin-1 which, in thelight of the fact that both molecules represent well-characterized tumor markers (Bast et al., 1981; Bast et al.,1983; Bon et al., 1996; Perillo et al., 1998; Rabinovich et al.,2002), might proof to be of significant biomedical importancein the future.
We are indebted to Angelika Kehlenbach and Blanche Schwappach(Zentrum für Molekulare Biologie Heidelberg) for their help in theFACS-based generation of clonal cell lines, as well as for providingtheir expertise with regard to FACS-based in vivo transport assays.We thank Felix Wieland (Biochemie-Zentrum Heidelberg) and Hans-Dieter Söling (Max Planck Institute for Biophysical Chemistry,Göttingen, Germany) for providing antibodies directed against p27,and calreticulin and the KDEL receptor, respectively. We thank allmembers of our laboratory for helpful discussions, as well as BrittaBrügger (Biochemie-Zentrum Heidelberg) for critical comments onthe manuscript. This study was supported by a grant of the GermanResearch Foundation (DFG) to W.N.
ReferencesAlbritton, L. M., Tseng, L., Scadden, D. and Cunningham, J. M. (1989).
A putative murine ecotropic retrovirus receptor gene encodes a multiple
Journal of Cell Science 116 (7)
Fig. 10.Correlation of endogenous CA125 expression with cell-surface expression of endogenous galectin-1 in CHO and HeLa cells.CHO (A) and HeLa cells (B) were dissociated from culture platesusing a protease-free buffer. Native cells were labeled with affinity-purified anti-galectin antibodies derived from a polyclonal rabbitantiserum. Cell-surface staining was analyzed by FACS using anti-rabbit secondary antibodies coupled to allophycocyanine to detectprimary antibodies. Autofluorescence levels (red curves in A and B)were determined with cells treated with trypsin prior to the FACSanalysis. Galectin-1 cell-surface levels are indicated in green (CHO;panel A) and blue (HeLa; panel B), respectively. Total galectin-1expression levels in CHO and HeLa cells, respectively, wereanalyzed by quantifying galectin-1 in total SDS cell lysates based ona western blot analysis (C). Lanes 1 and 4 represent the material of20,000 cells, lanes 2 and 5 represent the material of 50,000 cells andlanes 3 and 6 represent the material of 150,000 cells. The resultsfrom CHO cells are shown in lanes 1-3, the results from HeLa cellsare shown in lanes 4-6. Galectin-1 was detected with an affinity-purified rabbit antiserum directed against recombinant full-lengthgalectin-1.

1317CA125 is a galectin-1 receptor
membrane-spanning protein and confers susceptibility to virus infection.Cell 57, 659-666.
Armstrong, S. A., Staunton, J. E., Silverman, L. B., Pieters, R., den Boer,M. L., Minden, M. D., Sallan, S. E., Lander, E. S., Golub, T. R. andKorsmeyer, S. J. (2002). MLL translocations specify a distinct geneexpression profile that distinguishes a unique leukemia. Nat. Genet. 30, 41-47.
Balch, W. E. and Rothman, J. E. (1985). Characterization of protein transportbetween successive compartments of the Golgi apparatus: asymmetricproperties of donor and acceptor activities in a cell-free system. Arch.Biochem. Biophys. 240, 413-425.
Barondes, S. H., Cooper, D. N., Gitt, M. A. and Leffler, H. (1994). Galectins.Structure and function of a large family of animal lectins. J. Biol. Chem.269, 20807-20810.
Bast, R. C., Jr, Feeney, M., Lazarus, H., Nadler, L. M., Colvin, R. B. andKnapp, R. C. (1981). Reactivity of a monoclonal antibody with humanovarian carcinoma. J. Clin. Invest. 68, 1331-1337.
Bast, R. C., Jr, Klug, T. L., St John, E., Jenison, E., Niloff, J. M., Lazarus,H., Berkowitz, R. S., Leavitt, T., Griffiths, C. T., Parker, L. et al. (1983).A radioimmunoassay using a monoclonal antibody to monitor the course ofepithelial ovarian cancer. N. Engl. J. Med. 309, 883-887.
Bon, G. G., Kenemans, P., Verstraeten, R., van Kamp, G. J. andHilgers, J. (1996). Serum tumor marker immunoassays in gynecologiconcology: establishment of reference values. Am. J. Obstet. Gynecol. 174,107-114.
Bresalier, R. S., Byrd, J. C., Wang, L. and Raz, A. (1996). Colon cancermucin: a new ligand for the beta-galactoside-binding protein galectin-3.Cancer Res. 56, 4354-4357.
Cho, M. and Cummings, R. D. (1995). Galectin-1, a β-galactoside-bindinglectin in Chinese hamster ovary cells. II. Localization and biosynthesis. J.Biol. Chem. 270, 5207-5212.
Cleves, A. E. (1997). Protein transports: the nonclassical ins and outs. Curr.Biol. 7, R318-320.
Cleves, A. E., Cooper, D. N., Barondes, S. H. and Kelly, R. B. (1996). Anew pathway for protein export in Saccharomyces cerevisiae. J. Cell Biol.133, 1017-1026.
Cooper, D. N. and Barondes, S. H. (1990). Evidence for export of a musclelectin from cytosol to extracellular matrix and for a novel secretorymechanism. J. Cell Biol. 110, 1681-1691.
Davey, R. A., Hamson, C. A., Healey, J. J. and Cunningham, J. M. (1997).In vitro binding of purified murine ecotropic retrovirus envelope surfaceprotein to its receptor, MCAT-1. J. Virol. 71, 8096-8102.
Davis, H. M., Zurawski, V. R., Jr, Bast, R. C., Jr and Klug, T. L. (1986).Characterization of the CA 125 antigen associated with human epithelialovarian carcinomas. Cancer Res. 46, 6143-6148.
Deutscher, S. L. and Hirschberg, C. B. (1986). Mechanism of galactosylationin the Golgi apparatus. A Chinese hamster ovary cell mutant deficient intranslocation of UDP-galactose across Golgi vesicle membranes. J. Biol.Chem. 261, 96-100.
Engling, A., Backhaus, R., Stegmayer, C., Zehe, C., Seelenmeyer, C.,Kehlenbach, A., Schwappach, B., Wegehingel, S. and Nickel, W. (2002).Biosynthetic FGF-2 is targeted to non-lipid raft microdomains followingtranslocation to the extracellular surface of CHO cells. J. Cell Sci.115,3619-3631.
Fendrick, J. L., Konishi, I., Geary, S. M., Parmley, T. H., Quirk, J. G., Jrand O’Brien, T. J. (1997). CA125 phosphorylation is associated with itssecretion from the WISH human amnion cell line. Tumour Biol. 18, 278-289.
Füllekrug, J., Sönnichsen, B., Schafer, U., Nguyen Van, P., Söling, H. D.and Mieskes, G. (1997). Characterization of brefeldin A induced vesicularstructures containing cycling proteins of the intermediate compartment/cis-Golgi network. FEBS Lett. 404, 75-81.
Füllekrug, J., Suganuma, T., Tang, B. L., Hong, W., Storrie, B. andNilsson, T. (1999). Localization and recycling of gp27 (hp24gamma3):complex formation with other p24 family members. Mol. Biol. Cell 10,1939-1955.
Gey, G. O., Coffman, W. D. and Kubicek, M. T. (1952). Tissue culturestudies of the prolieferative capacity of cervical carcinoma and normalepithelium. Cancer Res. 12, 264-265.
Hardardottir, H., Parmley, T., Quirk, J., Sanders, M., Miller, F. andO’Brien, T. (1990). Distribution of CA125 in embryonic tissue and adultderivatives of the fetal periderm. Am. J. Obstet. Gynecol. 163, 1925-1931.
Hughes, R. C. (1997). The galectin family of mammalian carbohydrate-binding molecules. Biochem. Soc. Trans. 25, 1194-1198.
Hughes, R. C. (1999). Secretion of the galectin family of mammaliancarbohydrate-binding proteins. Biochim. Biophys. Acta1473, 172-185.
Iurisci, I., Tinari, N., Natoli, C., Angelucci, D., Cianchetti, E. and Iacobelli,S. (2000). Concentrations of galectin-3 in the sera of normal controls andcancer patients. Clin. Cancer Res. 6, 1389-1393.
Jenne, N., Frey, K., Brugger, B. and Wieland, F. T. (2002). Oligomeric stateand stoichiometry of p24-proteins in the early secretory pathway. J. Biol.Chem. 277, 46504-46511.
Kutay, U., Ahnert-Hilger, G., Hartmann, E., Wiedenmann, B. andRapoport, T. A. (1995). Transport route for synaptobrevin via a novelpathway of insertion into the endoplasmic reticulum membrane. EMBO J.14, 217-223.
Lahm, H., Andre, S., Hoeflich, A., Fischer, J. R., Sordat, B., Kaltner, H.,Wolf, E. and Gabius, H. J. (2001). Comprehensive galectin fingerprintingin a panel of 61 human tumor cell lines by RT-PCR and its implications fordiagnostic and therapeutic procedures. J. Cancer Res. Clin. Oncol. 127, 375-386.
Lewis, M. J. and Pelham, H. R. (1990). A human homologue of the yeastHDEL receptor. Nature348, 162-163.
Lewis, M. J. and Pelham, H. R. (1992). Ligand-induced redistribution of ahuman KDEL receptor from the Golgi complex to the endoplasmicreticulum. Cell 68, 353-364.
Lippincott-Schwartz, J., Yuan, L. C., Bonifacino, J. S. and Klausner, R.D. (1989). Rapid redistribution of Golgi proteins into the ER in cells treatedwith brefeldin A: evidence for membrane cycling from Golgi to ER. Cell56, 801-813.
Lloyd, K. O. and Yin, B. W. (2001). Synthesis and secretion of the ovariancancer antigen CA 125 by the human cancer cell line NIH:OVCAR-3.Tumour Biol. 22, 77-82.
Lloyd, K. O., Yin, B. W. and Kudryashov, V. (1997). Isolation andcharacterization of ovarian cancer antigen CA 125 using a new monoclonalantibody (VK-8): identification as a mucin-type molecule. Int. J. Cancer71,842-850.
Lloyd, R. V. (2001). Distinguishing benign from malignant thyroid lesions:galectin 3 as the latest candidate. Endocr. Pathol. 12, 255-257.
Lutomski, D., Fouillit, M., Bourin, P., Mellottee, D., Denize, N., Pontet, M.,Bladier, D., Caron, M. and Joubert-Caron, R. (1997). Externalization andbinding of galectin-1 on cell surface of K562 cells upon erythroiddifferentiation. Glycobiology7, 1193-1199.
Meyer, T. and Rustin, G. J. (2000). Role of tumour markers in monitoringepithelial ovarian cancer. Br. J. Cancer82, 1535-1538.
Nagata, A., Hirota, N., Sakai, T., Fujimoto, M. and Komoda, T. (1991).Molecular nature and possible presence of a membranous glycan-phosphatidylinositol anchor of CA125 antigen. Tumour Biol. 12, 279-286.
Nap, M., Vitali, A., Nustad, K., Bast, R. C., Jr, O’Brien, T. J., Nilsson, O.,Seguin, P., Suresh, M. R., Bormer, O. P., Saga, T. et al. (1996).Immunohistochemical characterization of 22 monoclonal antibodies againstthe CA125 antigen: 2nd report from the ISOBM TD-1 Workshop. TumourBiol. 17, 325-331.
Nguyen, J. T., Evans, D. P., Galvan, M., Pace, K. E., Leitenberg, D., Bui,T. N. and Baum, L. G. (2001). CD45 modulates galectin-1-induced T celldeath: regulation by expression of core 2 O-glycans. J. Immunol. 167, 5697-5707.
O’Brien, T., Hardin, J., Bannon, G., Norris, J. and Quirk, J. (1986). CA125antigen in human amniotic fluid and fetal membranes. Am. J. Obstet.Gynecol. 155, 50-55.
O’Brien, T. J., Beard, J. B., Underwood, L. J., Dennis, R. A., Santin, A.D. and York, L. (2001). The CA 125 gene: an extracellular superstructuredominated by repeat sequences. Tumour Biol. 22, 348-366.
Orci, L., Tagaya, M., Amherdt, M., Perrelet, A., Donaldson, J. G.,Lippincott-Schwartz, J., Klausner, R. D. and Rothman, J. E. (1991).Brefeldin A, a drug that blocks secretion, prevents the assembly of non-clathrin-coated buds on Golgi cisternae. Cell 64, 1183-1195.
Perillo, N. L., Pace, K. E., Seilhamer, J. J. and Baum, L. G. (1995).Apoptosis of T cells mediated by galectin-1. Nature378, 736-739.
Perillo, N. L., Marcus, M. E. and Baum, L. G. (1998). Galectins: versatilemodulators of cell adhesion, cell proliferation, and cell death. J. Mol. Med.76, 402-412.
Puck, T. T., Cieciura, S. J. and Robinson, A. (1958). Genetics of somaticmammalian cells. J. Mol. Med. 108, 945-955.
Rabinovich, G. A., Baum, L. G., Tinari, N., Paganelli, R., Natoli, C., Liu,F. T. and Iacobelli, S. (2002). Galectins and their ligands: amplifiers,silencers or tuners of the inflammatory response? Trends Immunol. 23, 313-320.

1318
Scherer, W. F., Syverton, J. T. and Gey, G. O. (1953). Studies on thepropagation in vitro of poliomyelitis viruses. J. Exp. Med. 97, 695-710.
Sönnichsen, B., Füllekrug, J., Nguyen Van, P., Diekmann, W., Robinson,D. G. and Mieskes, G. (1994). Retention and retrieval: both mechanismscooperate to maintain calreticulin in the endoplasmic reticulum. J. Cell Sci.107, 2705-2717.
Urlinger, S., Baron, U., Thellmann, M., Hasan, M. T., Bujard, H. andHillen, W. (2000). Exploring the sequence space for tetracycline-dependenttranscriptional activators: novel mutations yield expanded range andsensitivity. Proc. Natl. Acad. Sci. USA 97, 7963-7968.
Wilm, M., Shevchenko, A., Houthaeve, T., Breit, S., Schweigerer, L., Fotsis,T. and Mann, M. (1996). Femtomole sequencing of proteins frompolyacrylamide gels by nano-electrospray mass spectrometry. Nature379,466-469.
Yin, B. W. and Lloyd, K. O. (2001). Molecular cloning of the CA125 ovariancancer antigen: identification as a new mucin, MUC16. J. Biol. Chem. 276,27371-27375.
Zurawski, V., Davis, H., Finkler, N., Harrison, C., Bast, R. and Knapp, R.(1988). Tissue distribution and characteristics of the CA125 antigen. CancerRev. 11-12, 102-118.
Journal of Cell Science 116 (7)





























