Embed Size (px)
Citation preview

The breakdown and decomposition of allochthonous and autochthonous plant litter in anoligotrophic lake (Llyn Frongoch)
R. David G. HanlonDepartment of Zoology, University College of Wales, Aberystwyth, Dyfed SY23 3DA, Wales
Keywords: decomposition, plant litter, oligotrophic lake
Abstract
The breakdown and decomposition of two species of deciduous leaf litter, Fagus sylvatica L. and Salixviminalis L. and two species of aquatic macrophyte Isoetes lacustris L . and Potamogeton perfoliatus L . wereexamined in an oligotrophic lake . In all cases plant litter in coarse mesh litter bags lost significantly morematerial than the fine mesh after 1 years submergence in the lake. This however was considered to be the resultof physical environmental factors and microbial activity rather than animal processing. The litter was rankedin order of fastest to slowest rates of decay as follows - Isoetes, Potamogeton, Salix and Fagus .Decomposition processes proceeded at a relatively slow rate as a result of low temperatures and lowphosphate and mineral ion concentration . The results suggested that there was an accumulation of organicmaterial in the lake .
Introduction
The flow of energy and the cycling of nutrientsare two of the major processes taking place withinecosystems, and the quantity of energy derivedfrom primary production and the rate of nutrientturnover are generally the prime factors deter-mining the productivity of organisms at successivetrophic levels (Egglishaw 1968) . In many lakes alarge proportion of the energy entering the systemoriginates from phytoplankton productivity . Insmall oligotrophic lakes, however, phytoplanktonproduction is often very low and aquatic macro-phytes and airborne terrestrial plant litter maymake a larger contribution to the energy budget incomparison with eutrophic lakes . The rate of plantlitter decomposition, particularly in oligotrophicsystems, may therefore provide an indication of therate of nutrient cycling and energy flow within theecosystem .
Much information has previously been collectedon the decomposition of deciduous leaf litter in
Hydrobiologia 88,281-288(1982) . 0018-8158/82/0883-0281/$01 .60.9 Dr W. Junk Publishers, The Hague . Printed in the Netherlands .
lakes (Szczepanski 1965 ; Pieczynska 1972 ; Gasith& Lawacz 1976; Reed 1979) and streams (Petersen& Cummins 1974 ; Iversen 1975 ; Benfield et al. 1977,1979; and many others), but much less informationis available on the decomposition of aquatic ma-crophyte litter (for reviews see Danell & Sjoberg1979 and Howard-Williams & Davies 1979) . Therate of plant litter breakdown and decomposition isdetermined by several factors, including the chem-ical and structural composition of the litter, tempe-rature, nutrient availability and invertebrate com-munity structure (Gasith & Lawacz 1976) . The roleof animal populations in the breakdown of plantlitter is however uncertain . Mathews & Kowal-czewski (1969) found no significant differences inweight loss from litter bags in exclusion experi-ments; nevertheless, Petersen & Cummins (1974)suggested that litter breakdown by invertebrateswas important for certain leaf species . The im-portance of animals in litter fragmentation will bedependent on the number and species of inverte-brates in the community which will, naturally, vary
282
between different freshwater habitats .The aim of this study is to examine the rate of
allochthonous and autochthonous plant litterbreakdown and decomposition in association withinvertebrate activity . This study is part of a largerproject designed to evaluate the energy budget foran oligotrophic lake (Llyn Frongoch) in order todetermine future management policies aimed atimproving secondary production .
Study area
Llyn Frongoch (National grid reference S .N. 721753) is a small (7 .2 ha), shallow (4 m deep)oligotrophic lake situated at an altitude of 300 m inthe highland plateau of Mid-Wales . The lake wasoriginally constructed as a reservoir and lies in ashallow basin with a catchment area of 1 .15 km 2.The lake is surrounded by rough pastureland andhas several tree species growing around its margin,beech (Fagus silvatica L.) and the common osier(Salix viminalis L.) being the two most commonspecies . The surrounding deciduous trees make anannual contribution of approximately 100 kg (0.625g C M-2 y-1 ) . of airborne plant litter to the organicmatter input of the lake (Hanlon 1981) . There arealso several species of aquatic macrophyte growingin and around the lake margins . Two of the majorspecies contributing to macrophyte primary pro-duction are Potamogeton perfoliatus L . and Isoeteslacustris L . which yield 720.0 kg (1 .68 g C m-2 Y 1)
and 460 kg (2 .187 g C m-2 y -1 ) respectively from atotal annual macrophyte production of 2400 kg (15g C m-2 y -1 ) . Hanlon (in prep . a) . The phytoplank-
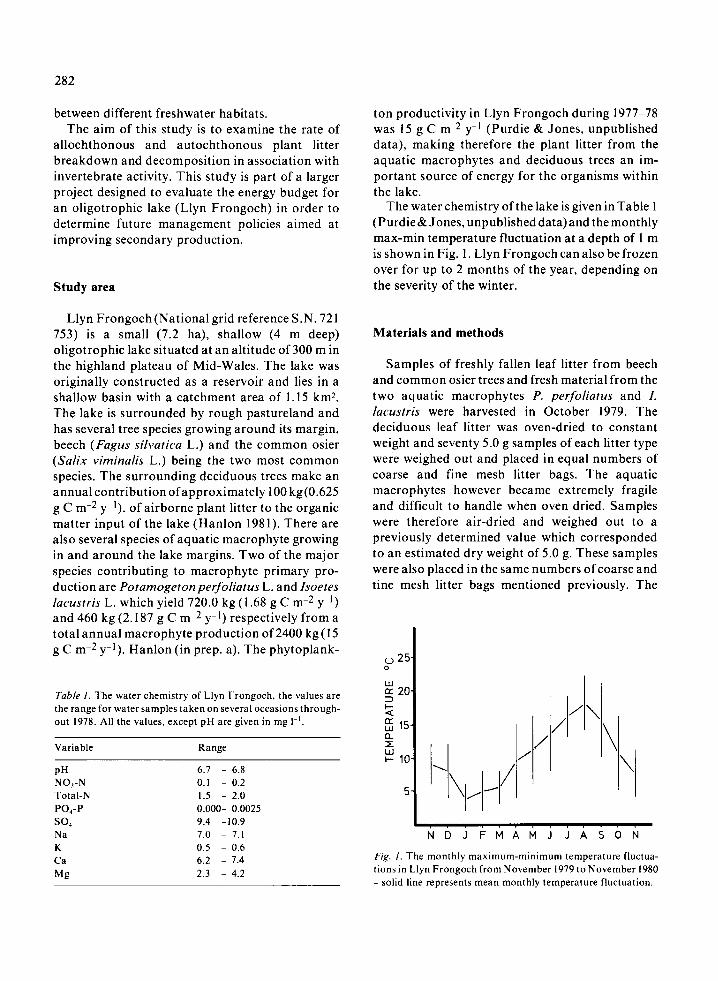
Table 1 . The water chemistry of Llyn Frongoch, the values arethe range for water samples taken on several occasions through-out 1978 . All the values, except pH are given in mg 1 -1 .
ton productivity in Llyn Frongoch during 1977-78was 15 g C m-2 Y-1 (Purdie & Jones, unpublisheddata), making therefore the plant litter from theaquatic macrophytes and deciduous trees an im-portant source of energy for the organisms withinthe lake .
The water chemistry of the lake is given in Table 1(Purdie& Jones, unpublished data) and the monthlymax-min temperature fluctuation at a depth of 1 mis shown in Fig . 1 . Llyn Frongoch can also be frozenover for up to 2 months of the year, depending onthe severity of the winter .
Materials and methods
Samples of freshly fallen leaf litter from beechand common osier trees and fresh material from thetwo aquatic macrophytes P. perfoliatus and I.lacustris were harvested in October 1979. Thedeciduous leaf litter was oven-dried to constantweight and seventy 5 .0 g samples of each litter typewere weighed out and placed in equal numbers ofcoarse and fine mesh litter bags . The aquaticmacrophytes however became extremely fragileand difficult to handle when oven dried . Sampleswere therefore air-dried and weighed out to apreviously determined value which correspondedto an estimated dry weight of 5 .0 g. These sampleswere also placed in the same numbers of coarse andtine mesh litter bags mentioned previously . The
u 250
w 20FQ
w 15a -Xw1- 10-
5- i
Z
N D J F M A M J J A S O N
Fig. 1 . The monthly maximum-minimum temperature fluctua-tions in Llyn Frongoch from November 1979 to November 1980- solid line represents mean monthly temperature fluctuation .
Variable Range
pH 6 .7 - 6 .8NO,-N 0 .1 - 0.2Total-N 1 .5 - 2.0P0,-P 0.000- 0.0025so, 9 .4 -10.9Na 7 .0 - 7 .1K 0 .5 - 0 .6Ca 6 .2 - 7 .4Mg 2 .3 - 4 .2
litter bags (0.15 X 0.2 m) were made from coarse (4X 4 mm) and fine (0 .1 X 0.1 mm) nylon mesh toenable the effects of the presence and absence ofbenthic invertebrates on litter breakdown to beexamined . The bags were heat-sealed and thenmoistened in the laboratory by the method ofHodkinson (1975) to prevent loss of material fromthe bags during handling and transportation .
The bags were placed in Llyn Frongoch in earlyNovember 1979 at a depth of 1 m and at a distanceof 5 m from the shoreline . The bags were tied ingroups of five and anchored in place with heavystones obtained from the lake . Five coarse and finemesh litter bags for each of the different litter typeswere removed from the lake after 1, 2, 3, 4, 6, 9 and12 months. Once removed, the samples were gentlywashed to remove sediment and animals, thenoven-dried (105 °C) to constant weight. The ben-thic invertebrates colonising the coarse mesh litterbags were collected and preserved in ethanol for
later identification and enumeration . No macro-invertebrates were found in the fine mesh bags .The breakdown and decomposition of the plant
litter was measured as a function of weight lossfrom the litter bags . The model most commonlyused to express weight loss from litter bags in bothterrestrial and aquatic systems is :
Wt = Wo e-Kt
where W t is the weight at time t, and Wo is the initialweight at t = 0, and K is the decay coefficient for thedifferent litter types . The decay coefficients for thefour litter types in both coarse and fine mesh litterbags wer calculated by fitting a linear regression toIn (W t/W) by the least squares method and thecoefficients of determination (r2) were calculatedfor each regression . The results were statisticallyanalysed by a Student t-test, analysis of varianceand the Student Newman-Keuls multiple com-parison test at P = 0 .05 (Soakl & Rolhf 1969) .
100
co 80-zzawcr
MwI-
J
wU
zwU(YwIL
60-
40
20
D J F M A M J J A S O N
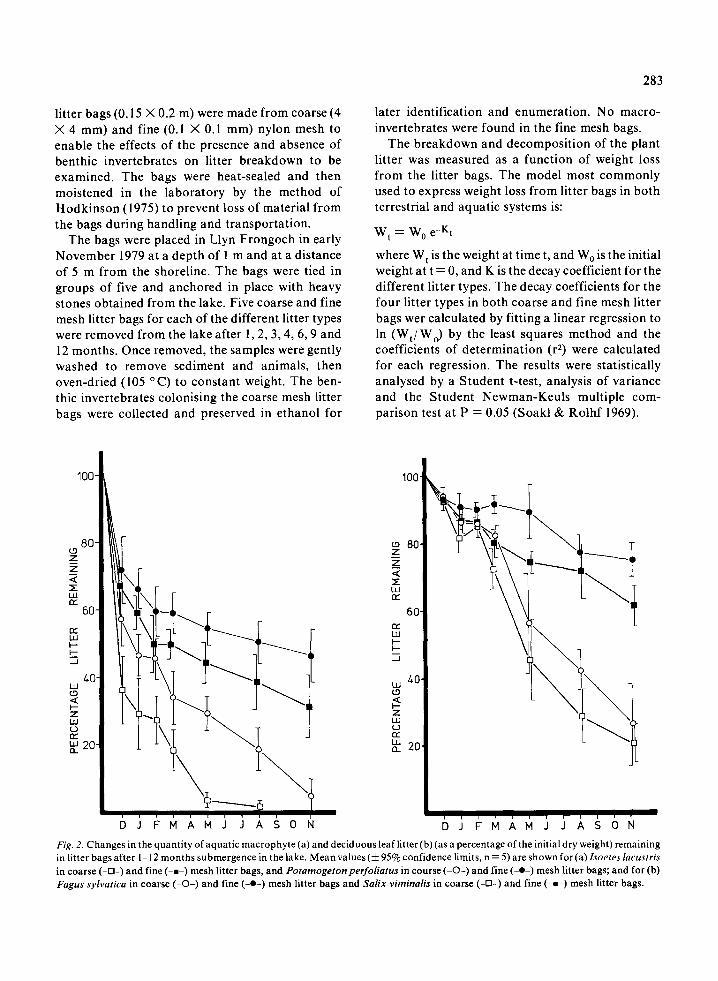
Fig . 2 . Changes in the quantity of aquatic macrophyte (a) and deciduous leaf litter (b) (as a percentage of the initial dry weight) remainingin litter bags after 1-12 months submergence in the lake . Mean values (±95% confidence limits, n=5) are shown for (a) Isoetes lacustrisin coarse(-[]-) and fine (-m-) mesh litter bags, and Potamogeton perfoliatus in course (-0-) and fine(-*-) mesh litter bags ; and for (b)Fagus sylvatica in coarse (-0-) and fine (-•-) mesh litter bags and Salix viminalis in coarse (-0-) and fine (-a-) mesh litter bags .
28 3
284
Results
Figures 2a and b show the percentage weightremaining of the four different plant litter types,enclosed in coarse and fine mesh litter bags . Weightloss from the fine mesh bags was initially rapid,most probably the result of leaching, followed by amuch slower rate of weight loss over the remainingmonths. Weight loss from the coarse mesh bagsfollowed a similar pattern, though it proceeded at amuch faster rate . In all cases the litter in the coarsemesh bags showed a significantly greater weightloss (0 .01 > P < 0.001) after 1 year than the litter inthe fine mesh bags, when analysed by a t-test. Acomparison between the quantity of material re-maining of each litter type, in both coarse (after 9months) and fine mesh (after 12 months) litter bags,was made using a multiple comparison test . Thecomparison between the quantity of the different
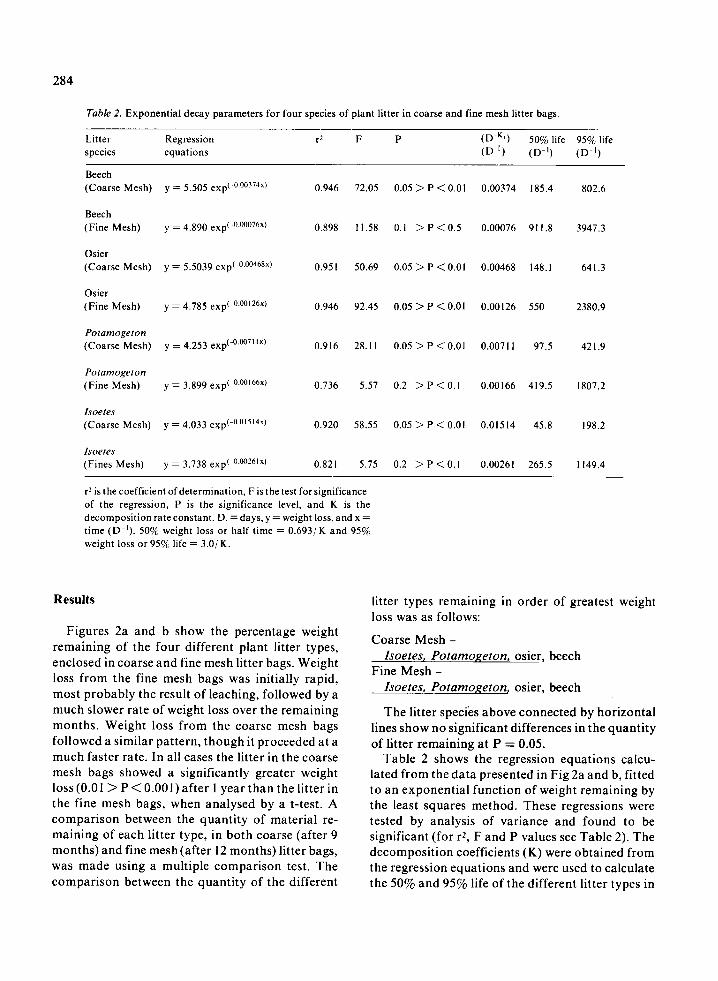
Table 2 . Exponential decay parameters for four species of plant litter in coarse and fine mesh litter bags .
Litter
Regression
r2species
equations
Beech(Coarse Mesh) y = 5 .505 exp( 0 .00374x)
Beech(Fine Mesh)
y = 4.890 exp) 0 .00076x)
Osier(Coarse Mesh) y = 5 .5039 exp(-0 .00468x)
Osier(Fine Mesh)
y = 4.785 exp( -0 .00 ' 26x)
Potamogeton(Coarse Mesh) y = 4 .253 exp(-0.00711 x)
Potamogeton(Fine Mesh) y = 3 .899 exp(-0°°' 66x)
0.736
0.946
0.898
0 .951
0 .946
0.916
Isoetes(Coarse Mesh) y = 4 .033 exp(-0 .01514x)
0.920
Isoetes(Fines Mesh)
y = 3 .738 exp ( 000261 x)
0 .821
r2 is the coefficient of determination, F is the test for significanceof the regression, P is the significance level, and K is thedecomposition rate constant . D. = days, y = weight loss, and x =time (D -1 ) . 50% weight loss or half time = 0 .693/ K and 95%weight loss or 95% life = 3 .0/ K .
litter types remaining in order of greatest weightloss was as follows :
Coarse Mesh - Isoetes, Potamogeton,osier, beech
Fine Mesh - Isoetes, Potamogeton, osier, beech
The litter species above connected by horizontallines show no significant differences in the quantityof litter remaining at P = 0 .05 .Table 2 shows the regression equations calcu-
lated from the data presented in Fig 2a and b, fittedto an exponential function of weight remaining bythe least squares method . These regressions weretested by analysis of variance and found to besignificant (for r 2 , F and P values see Table 2) . Thedecomposition coefficients (K) were obtained fromthe regression equations and were used to calculatethe 50% and 95% life of the different litter types in
F P (D K ')(D')
50% life(D - ')
95% life(D ')
72 .05 0.05 > P < 0 .01 0 .00374 185 .4 802 .6
11 .58 0.1 >P<0.5 0 .00076 911 .8 3947 .3
50 .69 0.05 > P < 0 .01 0.00468 148 .1 641 .3
92 .45 0 .05 > P < 0 .01 0.00126 550 2380 .9
28 .11 0 .05 > P < 0 .01 0 .00711 97 .5 421 .9
5 .57 0 .2 >P<0.1 0.00166 419 .5 1807 .2
58 .55 0 .05 > P < 0 .01 0.01514 45 .8 198 .2
5 .75 0 .2 >P<0.1 0.00261 265 .5 1149 .4
1
1
V)J 3Q
Z 20
< I
LL0 7
6
rrL11 5[E7
4
Z3
2
1
DEC
JAN FEB MAR MAY
DEC JAN FEB MAR MAY
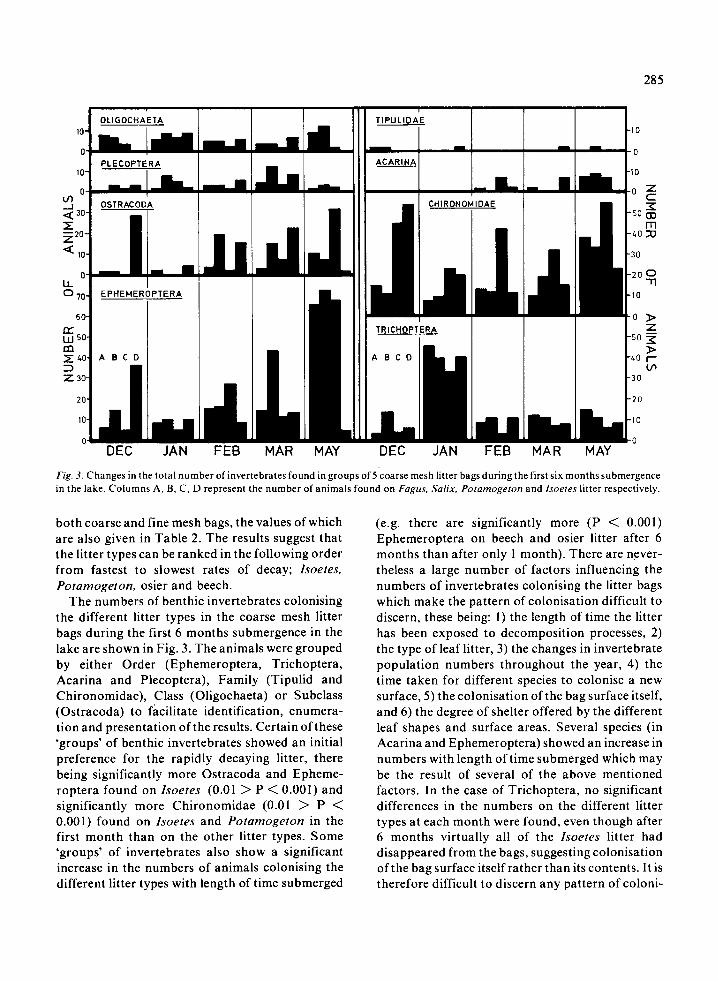
Fig. 3 . Changes in the total number of invertebrates found in groups of 5 coarse mesh litter bags during the first six months submergencein the lake . Columns A, B, C, D represent the number of animals found on Fagus, Salix, Potamogeton and Isoetes litter respectively .
both coarse and fine mesh bags, the values of whichare also given in Table 2 . The results suggest thatthe litter types can be ranked in the following orderfrom fastest to slowest rates of decay ; Isoetes,Potamogeton, osier and beech .
The numbers of benthic invertebrates colonisingthe different litter types in the coarse mesh litterbags during the first 6 months submergence in thelake are shown in Fig . 3 . The animals were groupedby either Order (Ephemeroptera, Trichoptera,Acarina and Plecoptera), Family (Tipulid andChironomidae), Class (Oligochaeta) or Subclass(Ostracoda) to facilitate identification, enumera-tion and presentation of the results . Certain of these`groups' of benthic invertebrates showed an initialpreference for the rapidly decaying litter, therebeing significantly more Ostracoda and Epheme-roptera found on Isoetes (0 .01 > P < 0.001) andsignificantly more Chironomidae (0 .01 > P <0.001) found on Isoetes and Potamogeton in thefirst month than on the other litter types. Some`groups' of invertebrates also show a significantincrease in the numbers of animals colonising thedifferent litter types with length of time submerged
0
0
0
0 ZC
50 00
40 70
0
200
0
285
0>Z
so 3>
0 r-tn
30
20
10
0
(e.g . there are significantly more (P < 0 .001)Ephemeroptera on beech and osier litter after 6months than after only I month) . There are never-theless a large number of factors influencing thenumbers of invertebrates colonising the litter bagswhich make the pattern of colonisation difficult todiscern, these being: 1) the length of time the litterhas been exposed to decomposition processes, 2)the type of leaf litter, 3) the changes in invertebratepopulation numbers throughout the year, 4) thetime taken for different species to colonise a newsurface, 5) the colonisation of the bag surface itself,and 6) the degree of shelter offered by the differentleaf shapes and surface areas . Several species (inAcarina and Ephemeroptera) showed an increase innumbers with length of time submerged which maybe the result of several of the above mentionedfactors. In the case of Trichoptera, no significantdifferences in the numbers on the different littertypes at each month were found, even though after6 months virtually all of the Isoetes litter haddisappeared from the bags, suggesting colonisationof the bag surface itself rather than its contents . It istherefore difficult to discern any pattern of coloni-
OLIGOCHAETA TIPULIDAE
ACARINAPLECOPTERA
iCHIRONOMIDAEOSTRACODA
I^ -
-I
I . EPHEMEROPTERA
)-TRICHOPTERA
-
) • A BCD A BCD -
l- -
1- -
)- -

28 6
sation; However, there was no difference in thenumber of invertebrate species found in the litterbags .
Discussion
In previous studies in both terrestrial and aquaticsystems, litter breakdown has often been confusedwith actual decomposition . Many researchersequated weight loss in exclusion experiments withdecay rates ; but weight loss in the presence ofanimal processing is principally a measure ofbreakdown and not total decomposition (Anderson1973), litter breakdown being the weight loss re-sulting from physical fragmentation of the litter byanimal feeding, environmental factors and micro-bial activity and decomposition being the degrada-tion of the plant materials into their constituentelements by microbial activity and digestion byanimals .
The decay coefficient (K) derived from theformula Wt = W O e-Kt is a convenient model fordescribing the initial stages of plant litter decom-position, but it is not strictly a true measure of theoverall decay rates . Plant litter consists of a varietyof compounds (i .e . sugars, hemicelluloses, cellu-lose, lignin and waxes) which all decay at differentrates. The rate of litter decomposition is the sum-mation of all the exponential decay curves for thedifferent litter constituents, the result of which isnot an exponential function (Minderman 1968) .The reason for the close fit of the regressions inTable 2 to the exponential model is the result of therelatively short time period (1 year) over which themeasurements were made . The decay curves ini-tially tend to follow the exponential decay func-tions of the more readily degradable fractionswhich account for a large proportion of the weightof plant litter, particularly aquatic macrophytes,and therefore the majority of the weight loss in thefirst year. However as the readily degradable frac-tions disappear, the decay curves then follow theexponential function represented by the more de-cay resistant materials, making the calculated valuefor the 95% life an underestimate . This change inthe decay rates over the longer term will be moreapparent for deciduous leaf litter which has a largerproportion of decay resistant materials present asstructural elements than aquatic macrophytes . The
rate of litter loss from the coarse mesh bagsnevertheless does not follow an exponential func-tion but for reasons mentioned previously thesevalues are a measure of litter breakdown and notdecomposition .The results of the experiments reported here
suggest that weight loss from coarse mesh litterbags was significantly greater than weight loss fromfine mesh bags and that the different litter typesdecayed at different rates . In comparison with thedecomposition coefficients (K) of previous studieson litter decay (Boiling et al. 1974 ; Sedall et a!. 1975 ;Gasith & Lawacz 1976; Hodkinson 1975; Danell &Sjoberg 1979 ; Howard-Williams & Davies 1979),the K-values for litter in coarse mesh bags showedsimilar values, though the results for the fine meshbags showed much lower K-values . However ameaningful comparison of litter decay rates fromdifferent sites is difficult owing to differences inexperimental design and environmental influences(Gasith & Lawacz 1976) . Nevertheless the resultsreported here suggest that litter decompositionproceeds at a relatively slow rate and an examina-tion of the lake sediments found a high proportionof plant material present, suggesting an accumula-tion of organic material in the lake (Hanlon, inprep . a) . The deciduous leaf litter decayed at a muchslower rate than the aquatic macrophytes whichmay be the result of differences in the chemical andstructural composition of the two litter types,terrestrial plant litter requiring more structuralcomponents consisting of the more decay resistantplant materials than the aquatic plants .
The relatively slower rate of litter decompositionin Llyn Frongoch may be the result of severalenvironmental factors. Figure 1 shows that the laketemperature is low for a relatively large proportionof the year, and low temperatures during the wintercan have an important influence on decompositionrates in freshwater habitats (Suberkropp et a!1975) . The variation in temperature throughout theyear may influence the form of the decay curves inFigs 2a and b and may account for a plateau in therate of weight loss from fine mesh bags during thecolder months of January - March, followed by aslight increase in the decay rates during the summermonths. The nitrogen and phosphate levels infreshwater lakes and streams can also have animportant influence on decay rates (Hynes &Kaushik 1969) and Llyn Frongoch has a relatively

low phosphate concentration but a much highernitrate level (see Table 1) . Iversen (1975) howeversuggested that a high nitrate concentration alonedid not appear to enhance decomposition processesand both high phosphate and nitrate levels arerequired to increase decay rates . The inorganic ionconcentration of the lake water is also low whichmay have some influence on decay rates as well asaffect the benthic invertebrate population numberswhich may influence the breakdown of plant litter.
Although the litter in the coarse mesh bagsdisappeared more rapidly than in the fine meshbags, the actual rate of decomposition of the litterfragments may be similar to the rates in the finemesh bags as the fragments lost from the coarsemesh bags have not been fully decomposed . Thelitter in the coarse mesh bags showed little sign ofinvertebrate feeding, and the majority of the ani-mals colonising the bags may have been feeding onfine sediments, epiphytic algae and micro-orga-nisms on the surface of the leaves . Trichoptera werethe major litter `shredders' collected from the bags,and as well as feeding on the litter the animals usedleaf fragments in case building . Llyn Frongochnevertheless has a poorly developed macro-inver-tebrate fauna (Hanlon, in prep . b) lacking potential`shredders' such as Gammarus and Asellus and hasonly a sparse population of litter feeding aquaticgastropods, oligochaetes and dipteran larvae. Thelack of evidence of invertebrate feeding suggeststhat the litter breakdown was mainly caused bymicrobial activity and physical environmental fac-tors, such water movement causing abrasion be-tween leaves . The leaves in the fine mesh bags alsoshowed signs of fragmentation though no materialwas lost from the bags .
Mathews & Kowalczewski (1969) suggested thatanimal activity played an insignificant role in litterbreakdown in the River Thames, and the experi-ments reported here reached similar conclusions asa result of the poorly developed invertebrate com-munity in Llyn Frongoch . In contrast, however,with the findings of Mathews & Kowalczewski(1969), the litter in the coarse mesh bags here brokedown more rapidly than that in the fine mesh bagsas the result of microbial activity and abrasionbetween leaves. The results also suggest that therates of nutrient turnover and energy flow in LlynFrongoch are low, resulting in a poorly developedpopulation at secondary trophic levels . Plant litter
287
forms an important energy source in the lakeaccounting for more than 100% of that representedby phytoplankton productivity. The slow rate oflitter decomposition and the accumulation of or-ganic matter in the lake sediments suggest that thepotential energy entering the system is not beingfully utilised. This may be the result of severalfactors including low temperatures and low phos-phate and mineral element concentrations, factorswhich must be taken into consideration whenplanning future management policies aimed atimproving secondary production .
Acknowledgements
I would like to thank D. Purdie and A . Jones fortheir data concerning phytoplankton productionand water chemistry of Llyn Frongoch. This re-search was supported by the Davies Trust .
References
Anderson, J . M ., 1973 . The breakdown and decomposition ofsweet chestnut (Castanea sativa) and beech (Fagus sylvatica)leaf litter in two deciduous woodland soils . Oecologia 12 :251-274 .
Benfield, E . F ., Jones, D . S . & Patterson, M . F., 1977 . Leaf packprocessing in a pastureland stream . Oikos 29 : 99-103 .
Benfield, E . F ., Paul, R . W. & Webster, J . R., 1979 . Theinfluence of exposure technique on leaf breakdown rates instreams . Oikos 33 : 386-391 .
Boiling, R . H., Goodman, E . D ., Sickle, J . A ., Zimmer, J . 0 .,Cummins, K . W., Petersen, R. C. & Reice, S . R ., 1975 .Toward a model of detritus processing in a woodland stream .Ecology 56: 141-151 .
Danell, K . & Sjoberg, K ., 1979 . Decomposition of Carex andEquisetum in a northern Swedish lake : dry weight loss andcolonisation by macro-invertebrates . J . Ecol . 67 : 191-200 .
Egglishaw, H . J ., 1968 . The quantitative relationship betweenbottom fauna and plant detritus in streams of differentcalcium concentrations . J . appl . Ecol . 5 : 731-740 .
Fenchel, T. & Harrison, P . G., 1976 . The significance ofbacterial grazing and mineral cycling for the decompositionof particulate detritus . In : Anderson J . M. & Macfaden, A .(eds) The Role of Terrestrial and Aquatic Organisms inDecomposition Processes, pp . 285-299 . Blackwell, Oxford .
Gasith, A. & Lawacz, W ., 1976 . The breakdown of leaf litter inthe littoral zone of a eutrophic lake . Ekol, Pol . 24 : 421-430 .
Hanlon, R. D . G ., 1981 . Allochthonous plant litter as a source oforganic material in an oligotrophic lake (Llyn Frongoch) .Hydrobiologia 80: 257-261 .
Hanlon, R . D. G . (in prep . a) . An organic carbon budget for anoligotrophic lake (Llyn Frongoch) .

288
Hanlon, R . D . G . (in prep . b) . The productivity of the benthicinvertebrates in an oligotrophic lake (Llyn Frongoch) .
Hargrave, B . T ., 1970 . The effect of a deposit-feeding amphipodon the metabolism of benthic microflora. Limnol. Oceanogr .15 :21-30.
Hargrave, B . T ., 1976 . The central role of invertebrate faeces insediment decomposition . In : Anderson J . M . & Macfadyen,A. (eds) The Role of Terrestrial and Aquatic Organisms inDecomposition Processes. Blackwell, Oxford .
Hodkinson, 1. D., 1975 . Dry weight loss and chemical changes invascular plant litter of terrestrial origin, occurring in a beaverpond system . J . Ecol . 63 : 131-142 .
Howard-Williams, C. & Davies, B . R ., 1979 . The rates of drymatter and nutrient loss from decomposing Potamogetonpectinatus in a brackish south-temperature coastal lake .Freshwat . Biol. 9 : 13-21 .
Hynes, H . B . N . & Kaushik, N . K ., 1969 . The relationshipbetween dissolved nutrient salts and protein production insubmerged autumnal leaves . Verh . int . Ver . Limnol . 17 :95-103 .
Iversen, T . N ., 1975 . Disappearance of autumn shed beech leavesplaced in bays in small streams . Verh . int . Ver . Limnol. 19:1687-1692 .
Lopez, G . R ., Levington, J . R . & Slobodkin, L. B ., 1977 . Theeffect of grazing by the detritivore Orchestia grillus onSpartina litter and its associated community . Oecologia 30:111-127 .
Mathews, C . P . & Kowalczewski, A ., 1969. The disappearanceof leaf litter and its contribution to production in the RiverThames . J . Ecol . 57 : 543-552 .
Minderman, G ., 1968 . Addition, decomposition and accumula-tion of organic matter in forests . J . Ecol . 56 : 355-362 .
Petersen, R . C . & Cummins, K . W ., 1974 . Leaf processing in awoodland stream . Freshwater Biol . 4 : 343-368 .
Pieczynska, E ., 1972 . Role of allochthonous matter in lakes .Wiad . Ekol . 18 : 131-140 .
Reed, F . C ., 1979 . Decomposition of Acer rubrum leaves atthree depths in an eutrophic Ohio lake. Hydrobiologia 64 :195-197 .
Satchell, J . E ., 1974 . Introduction : litter interface of animate/inanimate matter . In : Dickenson, C . H . & Pugh, G . J . F .(eds) . Biology of Plant Litter Decomposition . AcademicPress, London.
Sedell, J . R ., Triska, F . J . & Triska, N . S ., 1975 . The processingof conifer and hardwood leaves in two coniferous foreststreams . I . Weight loss and associated invertebrates . Verh .int . Ver . Limnol . 19: 1617-1627 .
Sokal, R. R. & Rohlf, F . J ., 1969 . Biometry . Freeman, SanFrancisco .
Suberkropp, K ., Klug, M . J. & Cummins, K . W., 1975 .Community processing of leaf litter in woodland streams .Verh . int . Ver . Limnol . 19 : 1653-1658 .
Szczepanski, A ., 1965 . Deciduous leaves as a source of organicmatter in lakes . Bull . Acad . pol. Sci ., Cl . 11 13 : 215-217 .
Received 2 March 1981 .





























