Embed Size (px)
Citation preview

278
THE BIOLOGY AND BEHAVIOUR OF PTINUSTECTUS BOIE. (COLEOPTERA, PTINIDAE),
A PEST OF STORED PRODUCTSII. THE AMOUNT OF LOCOMOTORY ACTIVITY IN RELATION
TO EXPERIMENTAL AND TO PREVIOUS TEMPERATURES
BY D. L. GUNN AND H. S. HOPF
From the Zoology Department, University of Birmingham
(Received 10 August 1941)
(With Six Text-figures)
I. INTRODUCTION
IT has long been known that temperature has a great effect on the speed of bio-logical processes. For this reason, when the speed of such a process is beingmeasured, the experimental temperature has to be controlled. It has only recentlybeen realized, however, that the temperature at which the biological material hasbeen kept before the experiment may also be important, even in cases whendamaging temperatures are not in question. For example, Thompson (1937)recorded the rate of heart beat of embryos of certain grasshoppers; for late embryosof Melanoplus femur-rubrum reared and tested at 20° C. this rate was considerablyhigher than for similar embryos tested at the same temperature but reared at 350 C.In fact, the rate for embryos reared and tested at 200 C. was about the same asthat for embryos reared at 35° C. and tested at 300 C. Mellanby (1940) found thatthe rates of heart beat of the crested newt {Triton cristatus) were much higher atboth o and 10° C. if the animals had been kept at io° C. merely for the previousday than if they had been kept at 300 C. for the same time. Again, Hopkins (1937)found that the rate of locomotion of an amoeba, Flabellula ndra, depended ontemperature in the usual way only if some hours had been allowed for temperatureacclimatization; during that period, the rate fluctuated widely before becomingsteady. On the other hand, Mellanby (1939) concluded that the speed of walkingof bedbugs {Cimex lectularius) depended only on the experimental temperature andnot at all on the previous temperature, provided the previous temperature had notbeen low enough to cause, chill coma nor high enough to be damaging, and pro-vided the insects were allowed half a minute to acclimatize to the new temperature.
There is some other information about the effects of previous temperatures onprocesses such as these, but it is scanty; our work on the subject was to have beena comprehensive study of Ptinus tectus, but it was interrupted at an early stage byinjuries sustained by one of us (H.S.H.) during an air raid. The incomplete results

The biology and behaviour of Ptinus tectus 279
on locomotory activity are presented now because they include new and usefulinformation and because there is no immediate prospect of extending them.
Locomotory activity is important in the life of this beetle not only because it ispartly responsible for spreading the species to neighbouring places, but also becausethe effectiveness of certain lethal agents depends on how much the insects aremoving about.
In order to find the total amount of movement in a given time, it is necessary toknow two things: how much of the time is spent in movement, and the speed ofthe movement while it is taking place. An estimate of the intensity of locomotoryactivity could, of course, be made by means of a suitable combination of measure-ments of oxygen consumption; our work, however, deals only with the amount oftime spent on locomotion. It will be seen that this does depend on the previoustemperature as well as on the temperature during observation.
Up to the present, three kinds of observation have been made on the effects oftemperature on the locomotory activity of invertebrates. First, Crozier (1924)measured the rate of walking of a millipede, Parajulus peimsylvanicus, as a functionof temperature, the object being to find the 'critical thermal increment'. Second,Chapman et al. (1926) raised the temperature of a vessel containing various insectsat a rate of 210 C. per hour, and recorded qualitatively the kind of activity—firstmovement of a limb, first crawling, first sign of heat paralysis, etc.—correspondingto each temperature. Third, Nicholson (1934) estimated the proportion moving ina batch of blowflies under given temperature conditions; since this is essentiallythe measurement we have adopted, it will be dealt with more fully.
In assessing the activity of Ptinus, the kind of activity was practically ignored.Locomotion was recorded separately from mere movements of the legs, or antennaewhich did not result in the insect changing its location ('virtual inactivity'), but nospecial attention was paid to the speed of locomotion. Ptinus tectus does not fly.When separate curves were drawn, showing locomotory activity alone and activityplus 'virtual inactivity', the latter were somewhat the higher, but there was noessential difference in shape. The general conclusions to be reached are not affectedby discussing or including the records of 'virtual inactivity', so they are omittedfor simplicity. We are concerned simply with the number of animals in locomotionunder given conditions.
The observations of activity were made by the ' cross-section method' (Bentleyet al. 1941). The aim is thus to count the animals which are moving at one instant;if an individual observation were to take an appreciable time, then the number ofanimals counted as moving would increase as the duration of this time increased,for some individuals would start to walk during the count. By this cross-sectionmethod, one might, for example, find that 60 % of the animals were moving at aninstant; with a reasonably constant level of activity over a period, it can be inferredfrom this that on the average each animal walked about for 60 % of the period inquestion. If individual animals were to be considered, it would of course be foundthat the time spent in activity varied from one individual to another.
Some of the factors, other than temperature, which affect the activity of Ptinus18-2

280 D. L. GUNN and H. S. H O P F
were easily eliminated: thus the normal diurnal rhythm of activity was abolishedby keeping the insects long enough at constant temperature and in unvarying light(Bentley et al. 1941); observations were usually not started for several hours afterthe animals had been placed in the observation chamber, so that the activity due tothe mechanical stimulation of handling had had time to subside; as far as possible,the light intensity in the observations was the same as for the cultures. With theaid of these precautions, reasonably self-consistent results were obtained, but verysmooth curves are not to be expected, because the activity of Ptinus appears to besensitive to many conditions.
In considering the effects of temperature both before and during the observationperiod, there appears to be only one set of conditions in which no arbitrary choiceof temperature or time relations has to be made; that is, the set of conditions inwhich a number of batches of animals are bred at different temperatures and theactivity of each batch is assessed at its temperature of breeding. Such a procedureallows only a limited comparison to be made between different batches. If animalsare to be tested at a temperature other than the temperature of breeding, then thespeed at which their temperature is changed and the time they are left at the newtemperature before testing»have to be chosen arbitrarily. Had sufficient data beenaccumulated, it might have been possible to reduce the a/bitrariness of the choiceof conditions. The conditions chosen are mentioned under the appropriate headings.
II. MATERIAL AND METHODS
The experimental animals were normally bred and kept at 25° C, except whenthe effects of a different culture temperature were to be investigated. They werebred in z\ 1. jars (Breffitts) containing about 600 g. of wholemeal flour and 30 g. ofdried yeast; the animals could drink water from a tube filled with wet cotton wool;some pads of dry cotton wool lay on the flour, and animals for experiments wereusually picked off these pads. The light was on constantly, day and night, and itsintensity was about 25 m.c. around the cultures. The animals used were of mixedand unknown ages: during the observations of activity they were neither fed norallowed to drink; none of them was used more than once.
The observations were carried out with the animals in glass dishes immersedin water baths. The type of dish was cylindrical, 16 cm. in diameter and 85 cm.high, with the rim ground flat to make a good joint with a plate glass lid. This lidwas held on by a pair of joiner's clamps, which also served to sink the dish, and thejoint was made air tight with vaseline and, at temperatures over 30° C , an outsidelayer of paraffin wax. There were two holes in the lid, one for a thermometer and theother carrying a piece of 6 mm. glass tubing through which the animals were drop-ped into the dish. This tubing was not stoppered, but a piece of capillary tubing wasfixed to the open end with rubber tubing, thus preventing gross exchanges of airbetween the dish and the outside but allowing pressure adjustments. The beetlescould walk about freely on a perforated zinc platform, but were prevented fromgetting around the edges and below the platform by a ring of electric flex, freed

The biology and behaviour of Ptinus tectus 281
from its insulating cloth and rubber, squeezed into the small gaps between theplatform and the glass wall. Below the platform there were glass pots of potassiumhydroxide or sulphuric acid solution of appropriate concentration to control thehumidity. Observations were almost always made concurrently at three differenthumidities (approx. 34, 60 and 95 % R.H.) ; the effect of humidity on activity wassmall and uncertain compared with the effects of temperature and it will not bedealt with here. Five animals were used in each dish in each experiment.
One of the two water baths used for temperature control was a refrigeratedthermostat designed and constructed by Mr R. J. Whitney. There was artificialillumination of about 25 m.c. around the dishes, the tank being completely enclosedin wood. This tank was used for experiments at constant temperatures between1-5 and 150 C. The other water bath could be used as a thermostat with the normalarrangement of a 400 W. heater, mercury-toluene regulator and Sunvic vacuumswitch. For experiments between room temperature and tap-water temperature,eooling was carried out with tap water passing through metal tubing. This tank wassurrounded by light-proof curtains and, once more, the light intensity was about25 m.c. This second tank could be heated rapidly with bunsen burners and cooledwith ice or a stream of water.
III. OBSERVATIONS AT CONSTANT TEMPERATURE
(a) Animals bred and kept at 250 C.
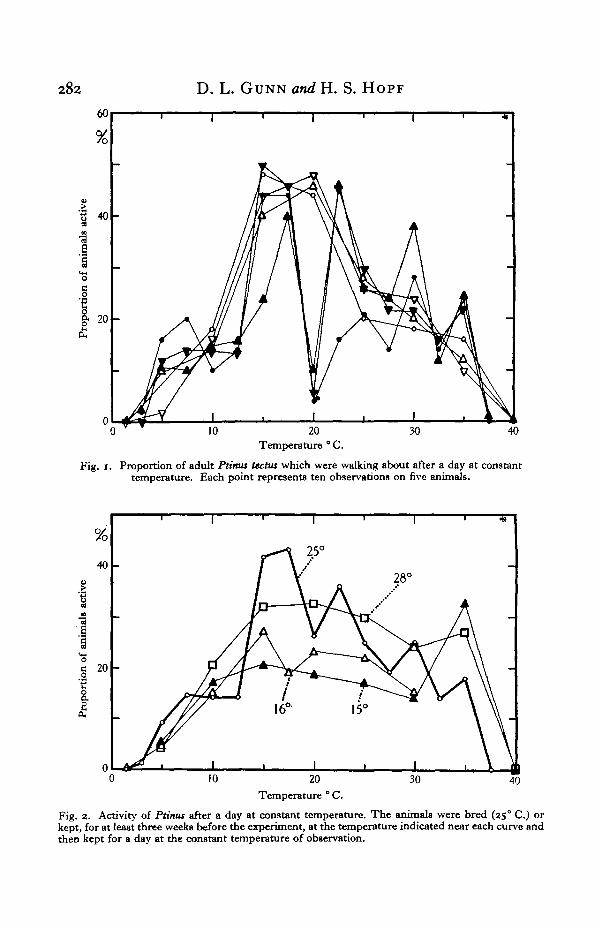
The first observations were made on the activity of animals bred and kept at250 C.; they were transferred to the experimental dishes, which had already beenallowed half an hour to equilibrate in the thermostat, and then left at the constanttemperature of the particular experiment for over 24 hr. The first reading wastaken after 23 hr., and nine further readings were made at 10 min. intervals. Foreach of the three dishes with five animals in each, the number of records of activityfrom one experiment was thus fifty. Each point on Fig. 1 shows what percentageof these fifty records were records of animals moving from place to place. .Thustwo series of experiments each with three dishes show that at 10° C. averages ofbetween 10 and 18 % of the animals were walking about at any instant.
In Fig. 1 the seventy-one points represent observations on that number of diffe-rent batches of five animals. Generally speaking, this graph shows a surprising self-consistency. The only notable inconsistency was at 20° C; the three batches inthe second series at this temperature were all rather inactive, and no explanationhas been found for this.
In Fig. 2 the six sets of observations of Fig. 1 are shown combined in the curvemarked 250 C.
(b) Animals previously kept at temperatures other than 25° C.
A preliminary series of experiments was done with animals which had been keptcooler than 250 C. A culture of young adults (less than a month old) was simplyimmersed in a tank which was kept cool with tap water. Between the beginning of
282 D. L. GUNN and H. S. HOPF
10 20Temperature ° C.
30 40
Fig. i. Proportion of adult Ptinus tectus which were walking about after a day at constanttemperature. Each point represents ten observations on five animals.
10 20Temperature ° C.
30 40
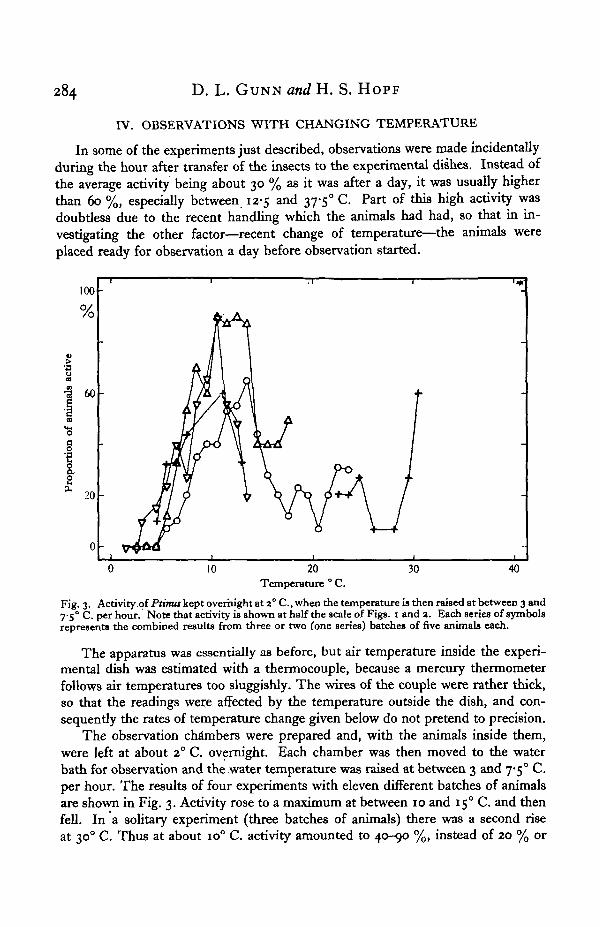
Fig. 2. Activity of Ptinus after a day at constant temperature. The animals were bred (25° C.) orkept, for at least three weeks before the experiment, at the temperature indicated near each curve andthen kept for a day at the constant temperature of observation.

The biology and behaviour of Ptinus tectus 283
the experiments in May and the end in July, the temperature rose from 13-5 to18° C. The observations began after 3 weeks and the culture temperature wasabout 16° C. when most of the experiments were done. In other respects theexperiments were like the previous ones.
The results are shown in the curve marked 160 in Fig. 2, and they were generallysimilar to those for a previous temperature of 250 C, but the activity recorded wasless. Further experiments were therefore done with carefully controlled previoustemperatures of 15 and 280 C. It was not then known that 280 C. is too high atemperature for the development of Ptinus (Ewer & Ewer, 1941), but it does notseem to have been too high for the purpose of these experiments.
These two sets of observations were like each other, but different from the twoprevious sets in t£iat the cultures had to be kept in constant darkness. At each tem-perature and humidity two batches of five animals were studied simultaneously,one from 15 and one from 28° C. Since all the animals had originally been takenfrom one culture, the two sets of experiments are comparable.
It will be seen from Fig. 2 that when observed at any temperature between 10 and300 C. the activity of animals previously kept at 280 C. was always higher than thatof animals previously kept at 15° C, the increase being about 60 % on the average.
Leaving aside for the moment the differences between the four curves of Fig: 2and considering only their general shape, it will be seen that they are mutuallyconfirmatory. They have been combined in the lowest curve of Fig. 4; in this case,a number of the set of experiments for a previous temperature of 250 C. (e.g. at7*5, 12-5, i7-5° C, etc.) have been omitted, so as to avoid unduly weighting thecurve with this set.
Up to an experimental temperature of 150 C. the activity after a day at constanttemperature is higher at each successively higher temperature. This is in line withwell-known physiological effects of temperature. But between 15 and 250 C. theactivity is relatively constant. It seems very unlikely that activity in this broadtemperature region reaches a maximum because of harmful effects, of temperature.For example, Deal (1939) has shown that Ptinus has a preferred temperature zonearound 23° C. and this has been confirmed by Dr Gunn and Miss Walshe in thislaboratory. Ewer & Ewer (1941) have shown that the life history of Ptinus is shortestat 23-250 C. and they did not find any harmful effects of temperature on adults at250 C. or below. We have had generation after generation of Ptinus in cultures keptat 250 C. Indeed, Fig. 2 itself shows that after a day at 35° C. Ptinus walks about tomuch the same extent as it does after a day at 150 C, and this activity was notnotably abnormal in character. We therefore consider that the shape of the curvesbetween 15 and 25° C. in Fig. 2 represents the relation between temperature—previously constant for a day—and the frequency of normal non-pathologicallocomotory activity. This relation is quite unlike that approximately expressed bythe Q10 rule. Indeed, if one does not attach much value to the somewhat higheraverage activity at 150 C, one may say that the frequency of locomotory activityof Ptinus (not the velocity of locomotion) does not vary much between 15 and350 C , provided the temperature has been constant for a day.
284 D. L. GUNN andH. S. H O P F
IV. OBSERVATIONS WITH CHANGING TEMPERATURE
In some of the experiments just described, observations were made incidentallyduring the hour after transfer of the insects to the experimental dishes. Instead ofthe average activity being about 30 % as it was after a day, it was usually higherthan 60 %, especially between 12-5 and 37-5° C. Part of this high activity wasdoubtless due to the recent handling which the animals had had, so that in in-vestigating the other factor—recent change of temperature—the animals wereplaced ready for observation a day before observation started.
20 30 40Temperature '
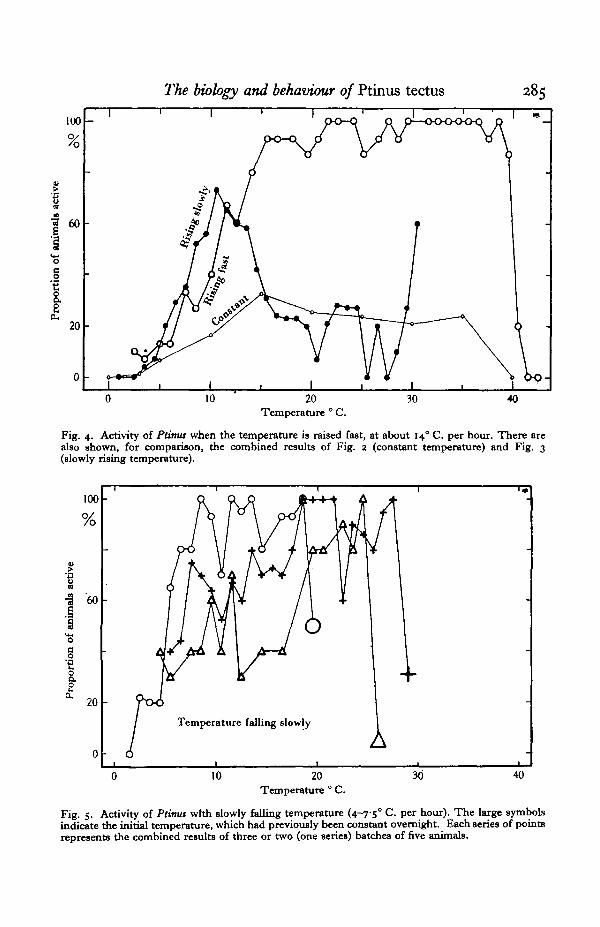
Fig. 3. Activity.of Ptima kept overnight at 2° C , when the temperature is then raised at between 3 and7-5° C. per hour. Note that activity is shown at half the scale of Figs. 1 and a. Each series of symbolsrepresents the combined results from three or two Cone series) batches of five animals each.
The apparatus was essentially as before, but air temperature inside the experi-mental dish was estimated with a thermocouple, because a mercury thermometerfollows air temperatures too sluggishly. The wires of the couple were rather thick,so that the readings were affected by the temperature outside the dish, and con-sequently the rates of temperature change given below do not pretend to precision.
The observation chambers were prepared and, with the animals inside them,were left at about 2° C. overnight. Each chamber was then moved to the waterbath for observation and the water temperature was raised at between 3 and 7-5° C.per hour. The results of four experiments with eleven different batches of animalsare shown in Fig. 3. Activity rose to a maximum at between 10 and 150 C. and thenfell. In a solitary experiment (three batches of animals) there was a second riseat 300 C. Thus at about io° C. activity amounted to 40-90 %, instead of 20 % or
100O/
60
20
0
-
-
The biology and behaviour of Ptinus tectus
p
fv /• i . i i i i
285
10 20Temperature ° C.
30 40
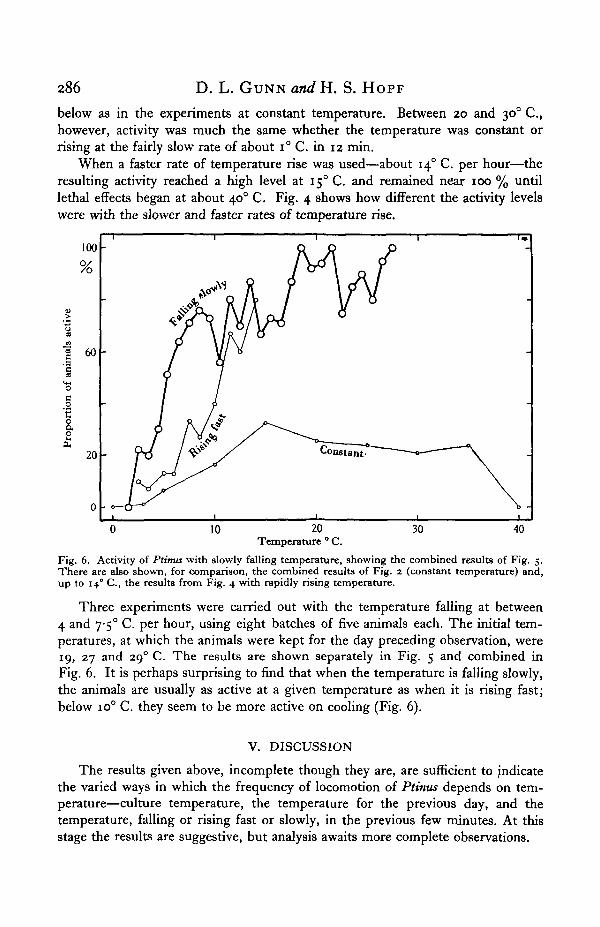
Fig. 4. Activity of Ptinus when the temperature is raised fast, at about 140 C. per hour. There arealso shown, for comparison, the combined results of Fig. 2 (constant temperature) and Fig. 3(slowly rising temperature).
100
%
I60 -
20 -
I Temperature falling slowly
1 1 1
•
• •
• •
• '
•
10 20Temperature c C.
30 40
Fig. 5. Activity of Ptimu wtth slowly falling temperature (4-7-5° C. per hour). The large symbolsindicate the initial temperature, which had previously been constant overnight. Each series of pointBrepresents the combined results of three or two (one series) batches of five animals.
286 D. L. GuNNarufH. S. H O P F
below as in the experiments at constant temperature. Between 20 and 300 C ,however, activity was much the same whether the temperature was constant orrising at the fairly slow rate of about i° C. in 12 min.
When a faster rate of temperature rise was used—about 140 C. per hour—theresulting activity reached a high level at 150 C. and remained near 100 % untillethal effects began at about 400 C. Fig. 4 shows how different the activity levelswere with the slower and faster rates of temperature rise.
20 30 40Temperature '
Fig. 6. Activity of Ptinus with slowly falling temperature, showing the combined results of Fig. 5.There are also shown, for comparison, the combined results of Fig. 2 (constant temperature) and,up to 140 C, the results from Fig. 4 with rapidly rising temperature.
Three experiments were carried out with the temperature falling at between4 and 7-5° C. per hour, using eight batches of five animals each. The initial tem-peratures, at which the animals were kept for the day preceding observation, were19, 27 and 290 C. The results are shown separately in Fig. 5 and combined inFig. 6. It is perhaps surprising to find that when the temperature is falling slowly,the animals are usually as active at a given temperature as when it is rising fast;below io° C. they seem to be more active on cooling (Fig. 6).
V. DISCUSSION
The results given above, incomplete though they are, are sufficient to jndicatethe varied ways in which the frequency of locomotion of Ptinus depends on tem-perature—culture temperature, the temperature for the previous day, and thetemperature, falling or rising fast or slowly, in the previous few minutes. At thisstage the results are suggestive, but analysis awaits more complete observations.

The biology and behaviour of Ptinus tectus 287
It is well known that the rate of frequency of the most varied biological pro-cesses, such as oxygen consumption, mitosis, growth and even walking, increaseswith temperature in a way which can be approximately expressed in the Q10 ruleor, what comes to much the same thing, the Arrhenius formula (Belehradek, 1935;Heilbrunn, 1938). The value of Qw is often about 2 or 3, as long as the temperatureregion considered is not too close to or above the optimum. Examination of ourcurves shows no resemblance whatever to the usual type of Q10 curve, unless theoptimum is considered to be at or below 15° C, for above this temperature activityrises little in any of the temperature conditions we have dealt with. It would bereducing the already difficult conception of optimum to the absurd to regard 150 C.as optimal in any way for Ptinus; this temperature lies at or below the middle ofthe temperature range in which Ptinus can develop and carry on its normal activities(Ewer & Ewer, 1941). It is not particularly surprising that the frequency of loco-motion of Ptinus does not follow the Q10 rule; whatjs surprising, even with selecteddata, is that the velocity of walking of certain animals does roughly follow this rule.
In Nicholson's (1934) experiments on LuciUa cuprina, activity after 12 hr. atconstant temperature was highest at 300 C , which is about the preferred tempera-ture, and fell away rapidly above and below that temperature; in temperaturerising at about 70 C. per hour, activity was actually less at medium temperaturesand there appeared to be a stimulating effect of changing temperature only outsidethe preferred zone, below 15 and above 370 C. This corresponds roughly with ourresults for slowly rising temperature (3-7-5° C), as far as they go. But with slowlyfalling temperature the activity of Ptinus is much greater, and so it is with quicklyrising temperature (14° C. per hour). These results demonstrate an effect of tem-perature change as distinct from temperature level. Such an effect has already beenshown by Kennedy (1939) in experiments on the locust, Schistocerca gregaria, inthe Sudan. A sudden change of temperature eventually resulted in the locustsreaching a new activity level corresponding to the new temperature, but the im-mediate result of the change was a rise above the appropriate constant level.Indeed, when the temperature fell there was an absolute rise in activity which wasmore prolonged than the temporary rise due to a rise in temperature. In this workof Kennedy's, the change of temperature was very rapid, being initially i° C. in10—15 s e c -
Our work reveals a weakness of the observations made by a number of authorson the effect of rising temperature on insect activity. They raised the temperatureat rates of about 70 (Nicholson, 1934), io° (Bodenheimer et al. 1929), n ° (Hussein,1937), 200 (Nieschulz, 1933, 1935), and 210 C. (Chapman et al. 1926) per hour.But the temperature at which most of the animals are first active in our experimentsis about io° C. when the temperature is raised at 3-7-5° C. per hour and about150 C. when it is raised at about 140 C. per hour. In fact, over the range of rateswhich have been used by the various authors, the rate itself is likely to be as im-portant as the temperature in determining the point of first maximum activity, theother standard points may be affected in a similar way.
Nicholson's work (1934) on activity in blowflies seemed to indicate a relation

288 D. L. GuNNaniH. S. HOPF
between activity, as measured by the method we have adopted from him, and tem-perature preference (Fraenkel & Gunn, 1940, pp. 206-11). In the temperaturegradient apparatus in use in this laboratory (Gunn, 1934, 1935), however, Ptinuscan walk along the gradient at a rate equivalent to io° C. per minute or 6oo° C. perhour! It would therefore be unwise to attempt to draw from our experiments anyconclusion whatever about the mechanism of preferred temperature in this par-ticular gradient.
VI. SUMMARY
The amount of walking about shown by adult Ptinus tectus is not a simplefunction of temperature. It depends on culture temperature, on whether thetemperature is constant or changing and, with changing temperatures, on the speedand direction of change. Thus, at 20° C , about 25 % of animals walk about if thetemperature is constant or rising slowly, while about 90 % are active if it is risingfast or falling slowly. None of the temperature-activity curves bear any resemblanceto the usual kind of Qw curve.
For experiments in which the temperature has been constant for a day thetemperature-activity curve of Ptinus rises from 3 to 150 C., then is steady or fallingsomewhat to 30 or 35° C, and then falls steeply to 400 C. The activity level dependson the temperature at which the animals have been kept during the previous weeks.When the temperature is raised slowly (3-7*5° C. per hour), activity rises steeplyup to about io° C , then falls to the level appropriate to constant temperatures and,at about 30° C, rises steeply again. When the temperature is raised more quickly(140 C. per hour) the activity rise is at first similar to that for slowly rising tempera-ture, but it continues until 90-100 % activity is reached at about 150 C.; activityremains at this high level until damaging changes occur at about 40° C. When thetemperature is slowly lowered, the activity curve is similar to that for quickly risingtemperature. There is thus a stimulating effect of change of temperature—whetherrising or falling—the magnitude of which depends on the speed and direction ofthe change.
The results obtained have a considerable bearing on the testing and use ofinsecticides whose effectiveness depends on the activity of the insects.
REFERENCES
BELEHRADEK, J. (1935). Temperature and Living Matter. 277 pp., 70 figs. Berlin.BKNTLEY, E. W., GUNN, D. L. & EWER, D. W. (1941). J. exp. Biol. 18, 182-95.BoDKNHBiMBR, F. S., FRAENKEL, G., REICH, K. & SEGAL, N. (1929). Z. angexa. Ent. 15, 435-557.CHAPMAN, R. N., MICKEL, C. E., PARKER, J. R., MILLER, G. E. & KELLY, E. G. (1926). Ecology,
7, 416^-26.CROZIER, W. J. (1924). J. gen. Pkysiol. 7, 123-36.DEAL, J. H. (1939). Tropic Reactions of Insects. Ph.D. Thesis, Univ. Lond. (Fac. Sci.).EWER, D. W. & EWER, R. F. (1941). J. exp. Biol. 18, 290-305.FRAKNKEL, G. S. & GUNN, D . L. (1940). The Orientation of Animals: JCineses, Taxes, and Compost
Reactions. 352 pp., 135 figs. Oxford.GUNN, D. L. (1934). Z. vergl. Pkysiol. 20, 617-25.
(i935)- J- e*P- Biol. 12, 185-90.

The biology and behaviour of Ptinus tectus 289HEILBRUNN, L. V. (1938). An Outline of General Physiology, pp. 340-5. 603 pp., 122 figs. Phila-
delphia and London.HOPKINS, D. L. (1937). Protoplatma, 28, 161-74.HUSSEIN, M. (1937). Bull. Minitt. •Agric. Egypt, No. 184, 1-55.KENNEDY, J. S. (1939). Trans. R. ent. Soc. Land. 89, 385-542.MHLLANBY, K. (1939). Proc. roy. Soc. B, 127, 473-87.
(1940). Nature, Lond., 146, 165.NICHOLSON, A. J. (1934). Bull. ent. Res. 25, 85-99.NIEBCHULZ, O. (1933). Z. Parasitenk. 6, 220-42.
(1935). Zool. Ansc. 110, 225-33.THOMPSON, V. (1937). Physiol. ZoSl. 10, 21-30.









![1909] EASTON COLEOPTERA...1909] EASTON COLEOPTERA 49 A LIST OF COLEOPTERA COLLECTED WITHIN TEN MILES OF FALL RIVER, MASSACHUSETTS. BY NORMAN S. EASTON FALL …](https://img.pdfslide.us/doc/110x75/611535a3861718272235983f/1909-easton-coleoptera-1909-easton-coleoptera-49-a-list-of-coleoptera-collected.jpg)



















