Embed Size (px)
Citation preview

THE AUSTRALIAN NATIONAL UNIVERSITY _________________________________________________________________________________
Research School of Biology Division of Evolution, Ecology & Genetics CANBERRA ACT 0200 AUSTRALIA TELEPHONE: +61 26 125 2873 FACSIMILE: +61 26 125 5573 EMAIL: [email protected] 7 February 2014
To whom it concerns, Dr Gabor Neumann has asked me to provide documentation on the identity of dead, preserved lac insects (order Hemiptera, superfamily Coccoidea, family Kerriidae; formerly Tachardiidae) that were sent to me for taxonomic identification. All specimens from Australia (Christmas Island) and Malaysia (Klang in Selangor and Sarawak in Borneo) have morphology consistent with that of Tachardina aurantiaca (Cockerell), which is a common species in Southeast Asia. This species was named and described in 1903 from specimens collected on citrus in Java, Indonesia. Subsequent publications record this species from Australia (Christmas Island), Malaysia, the Maldives, Singapore and Thailand. All other species of the genus Tachardina occur in Africa and thus it is relatively easy to distinguish T. aurantiaca from other lac insects present in Southeast Asia. The only published taxonomic descriptions and illustrations of T. aurantiaca that are useful for morphological identification are by Chamberlin (1923) and Lit (2002). The latter is an unpublished chapter in a PhD thesis of one of my former students. I identified the adult female specimens from Christmas Island and Malaysia by examination of the insect cuticle (“skin”) under a compound microscope. This required the body contents of the specimens to be cleared (removed) by soaking for 24 hours in 10% potassium hydroxide, and then the cleaned cuticles were stained and dehydrated before being mounted in a resin on a microscope slide. The features useful for identification can only be seen under high magnification on the compound microscope. Tachardina aurantiaca is distinguished from other lac insects by a combination of morphological features, including the four-sectored anal ring, the presence of two secondary depressions on the brachial plate and the shape and composition of the marginal duct clusters. There had been no recent collection and study of T. aurantiaca until the work of Dr Neumann and his colleagues and associates. Regards,
Dr P.J. Gullan Emeritus Professor Literature cited Chamberlin, J.C. (1923) A systematic monograph of the Tachardiinae or lac insects (Coccidae).
Bulletin of Entomological Research 14: 147–212. Lit, I.L., Jr (2002) Morphology, Systematics and Phylogeny of the Lac Insects (Kerriidae,
Coccoidea, Hemiptera). Doctor of Philosophy thesis, University of the Philippines Los Baños, Philippines. 286 pp.

1
RESEARCH AND DEVELOPMENT OF BIOLOGICAL CONTROL FOR
SCALE INSECTS: INDIRECT CONTROL OF THE YELLOW CRAZY
ANT ON CHRISTMAS ISLAND, 2009‐2013
Peter T. Green, Dennis J. O’Dowd, Gabor Neumann, and Sarah Wittman
Department of Botany, La Trobe University, Bundoora, Vic 3086
Final Report to the Director of National Parks
3 July 2013

2
Executive Summary
Research Project 1. YCA dependence on honeydew
Project 1a. Honeydew use by YCA. Successful biological control of the yellow lac scale Tachardina
aurantiaca could remove a large fraction (an average of 70% to an average of 87%, depending on
assumptions) of honeydew available to the yellow crazy ant. However, there is considerable site‐to‐
site variation in the likely contribution of T. aurantiaca to the total honeydew economy, and it is not
certain that targeting this species alone would provide adequate indirect control for the YCA in all
supercolonies. The most prudent course of action is to target the entire assemblage of honeydew‐
producing scale insects through the introduction of a biological control agent for T. aurantiaca,
complemented by the use parasitoids already present on Christmas Island to target the coccoid soft
scales.
Project 1b. Stable Isotope Analysis. Stable isotope analyses of YCA workers, plants, herbivores and
predators collected from four declining supercolonies in 2010‐2011 indicate that at supercolony
densities, a substantial fraction of YCA dietary intake is plant‐derived. This is consistent with the idea
that YCA supercolonies depend heavily on honeydew derived from scale insects for a large fraction
of colony food and energy requirements, and provides support for the idea that indirect control over
YCA supercolonies could be achieved by targeting honeydew‐producing scale insects for biological
control.
Project 1c. Scale Insect Removal Experiment. The exclusion of YCA from access to scale insects at a
large experimental field site caused YCA activity on the ground to decline 5‐fold within 4 weeks,
compared to pre‐treatment levels. This large field experiment validates the key concept of indirect
biological control for YCA on Christmas Island; exclusion of honeydew‐producing scale insects from
YCA caused a significant and substantial reduction in YCA abundance on the ground.
Project 1d. Carbohydrate supply and YCA growth and behaviour. Dynamics and behaviour of YCA in
laboratory colonies depended on carbohydrate supply. When sugar supply was elevated,
reproductive output by queens increased, death rates of workers decreased, foraging tempo
quickened, and interspecific aggression intensified. These results suggest that sugar supply, through
honeydew supplied from scale insects, plays an important role in YCA supercolony dynamics,
foraging efficiency, and interspecific aggression.
Research Project 2. Scale insects and natural enemies
Projects 2a and 2c. Natural enemies survey, parasitization rates of lac scale insects (Tachardina
aurantiaca and Paratachardina pseudolobata) and parasitoid behaviour in area of origin
(Southeast Asia) and on Christmas Island. Few natural enemies of the key honeydew‐producing
scale insect Tachardina aurantiaca occur on Christmas Island, and they do not regulate its population
size. No parasitoids of female T. aurantiaca were found. Conversely, within its native range in
Malaysia, T. aurantiaca is rare and patchily distributed, associated with diverse natural enemies,
including at least five species of primary parasitoids, and it suffers high parasitization rates – all
attributes consistent with population regulation by natural enemies. Importantly, high parasitism
rates in Malaysia occurred in the presence of honeydew‐collecting ants, including the yellow crazy
ant. Of the primary parasitoids known to attack yellow lac scale from our studies in Malaysia,

3
Tachardiaephagus somervillei (Encyrtidae) is the most promising agent for introduction and release
on Christmas Island. All Tachardiaephagus species have a narrow host range and appear to be family
specialists, known only to attack the Kerriidae, the family to which the yellow lac scale belongs (Table
3). Our initial studies indicate that (a) T. somervillei attacks T. aurantiaca across 1900‐km range in
Peninsular Malaysia and Malaysian Borneo, (b) is the most abundant natural enemy of T. aurantiaca,
(c) has a short life cycle compared to its host, (d) exhibits superparasitism (i.e., where multiple
progeny emergence from a single host individual); (e) causes high rates of parasitism on T.
aurantiaca in the presence of tending ants, including the yellow crazy ant, and (f) can be reared
under laboratory conditions. Coccophagus ceroplastae and Encyrtus infelix, both parasitoids of a
wide variety of coccid scale insects, are already present on Christmas Island. The three parasitoids T.
somervillei. C. ceroplastae and E. infelix could be deployed in combination against the entire
assemblage of honeydew‐producing scale insects in YCA supercolonies.
Project 2b. Scale insect survey on Christmas Island (biology, host range, habitats, natural history,
identification). No native or endemic scale insect species have been discovered in intensive and
extensive searches for scale insect species on Christmas Island. However, 400 hours of search over
two years did yield five additional exotic scale insect species previously unknown to the island.
Assuming that any prospective biological control agent for Tachardina aurantiaca on Christmas
Island would be a narrow family specialist (Kerriidae) and all known scale insect species are non‐
native and invasive, the probability of any direct non‐target effects is negligible.
Project 2d. Genetic and morphological matching between Tachardina aurantiaca and Paratachardina
pseudolobata on Christmas Island and Southeast Asia. The single population of T. aurantiaca on
Christmas Island matches morphologically and genetically those populations examined so far within
its native region in Southeast Asia. This establishes the identity of Tachardina aurantiaca on
Christmas Island, making the successful establishment on Christmas Island of a biological control
agent that attacks T. aurantiaca in its native range more likely. Further collections and analyses of T.
aurantiaca in Eastern Malaysia (Sabah) and exploration in southern Thailand should establish if any
morphological or genetic variation occurs across its native distribution in Southeast Asia.
Project 2e. Scale population dynamics, especially the ecology of lac scale, Tachardina aurantiaca. The
life cycle of Tachardina aurantiaca is relatively long for a scale insect but several overlapping
generations occur in a single year. Female size is significantly correlated with fecundity, indicating
that conditions that affect female size are likely to have a strong influence on population growth
rates and density. The role of males in reproduction is not yet clear although it appears that some
females can reproduce parthenogenetically. All of these attributes will influence host‐parasitoid
dynamics on Christmas Island.
Research Project 3. Strategic planning for biocontrol introduction
Project 3a. Planning research in step with legislative and regulatory requirements for biological
control introductions. Two regulatory frameworks govern the importation and release of an exotic
biological control agent on Christmas Island. To gain approval to import and release a biological
control agent on Christmas Island, we will need to prepare and submit two sets of documents: an
Approvals Package under the DAFF protocol and an Environmental Referral to EACD, DSEWPaC. The
key government contacts involved in the two frameworks are described and currently consulting

4
each other.
Project 3b. Identification and agreement with co‐operators. Collaborations have been established
with researchers with expertise in the systematics, biology, and molecular genetics of scale insects,
taxonomy of microhymenopteran parasitoids, and biological control of scale insects. Cooperative
agreements have been established with the Forestry Research Institute of Malaysia (FRIM) and
Sarawak Forestry that provide benchspace and technical assistance. Dr Neumann is serving as an
advisor of a Masters student (Universiti Sains Malaysia, Penang) who is conducting research on this
project at FRIM. Research and collecting permits have been obtained from the States of Sarawak
and Sabah, as well as a national collecting permit.
Project 3c. Safety and sanitary protocols anticipating a pre‐release environmental assessment of
introduction. An agreed, peer‐reviewed host list and host specificity protocol has been prepared for
evaluating the risk of introducing T. somervillei to Christmas Island to control the invasive yellow lac
scale, Tachardina aurantiaca. It is recommended that host specificity testing be conducted in the
area of origin of Tachardina aurantiaca. A protocol for ensuring that hyperparasitoids are not
accidentally co‐introduced with importation of T. somervillei to Christmas Island has also been
prepared.
Project 3d. Rearing, release and monitoring protocols. The requirements to establish a founder
population of Tachardiaephagus in Malaysia free of pathogens and hyperparasites, to import this
population under permit to Christmas Island, to establish and expand this population in a mass‐
rearing facility on Christmas Island have all been identified. An agreed protocol for the rearing and
release of biological control agents, and pre‐ and post‐release monitoring for their efficacy in
controlling target scale insects and reducing densities and impacts of the invasive yellow crazy ant
Anoplolepis gracilipes has also been prepared.

5
Introduction
Background to the Program
Biology and Impacts of the Yellow Crazy Ant on Christmas Island. Anoplolepis gracilipes (the yellow
crazy ant, YCA) is a ‘tramp’ species that has become invasive throughout the tropics (Wetterer 2005).
The YCA is listed by the IUCN as one of the world’s 100 worst invasive species (Lowe et al. 2000), and
was accidentally introduced to Christmas Island between 1915 and 1934 (O’Dowd et al. 1999). Because
of its negative impacts on species, interactions and ecosystem processes, YCA is recognised as the most
significant and pervasive threatening process affecting biodiversity on Christmas Island, reflected by the
listing of the Loss of biodiversity and ecosystem integrity following invasion by the Yellow Crazy Ant on
Christmas Island, Indian Ocean as a Key Threatening Process under the EPBC Act 1999, and as identified
in Threat abatement plan to reduce the impacts of tramp ants on biodiversity in Australia and its
territories (Commonwealth of Australia 2006). The control of this ant features prominently in many
Recovery Plans for EPBC‐Listed species on Christmas Island, and has been the focus of natural resource
management activities on the island for more than a decade at the cost of millions of dollars.
The attribute that makes the YCA so problematic on Christmas Island and elsewhere is its capacity to
form high‐density, expansive ‘supercolonies’. Many tramp ant species form supercolonies, often
defined using a combination of criteria including genetic relatedness, intraspecific behavioural
interactions, and ant abundance. For example, the invasive argentine ant Linepithema humile is
genetically uniform across most of its European range and is regarded as a single supercolony (Giraud et
al. 2002), while intercontinental behavioural assays and analyses of cuticular hydrocarbons suggest this
species may form a single supercolony spanning Australia, Europe, North America and Japan (Sunamura
et al. 2009; Suhr et al. 2010). Although two distinct genotypes of YCA occur on Christmas Island, these
co‐occur at very small spatial scales (Thomas et al. 2010) and behavioural assays pairing individual ants
from opposite ends of the island suggest that the population on Christmas Island behaves as a single
supercolony (Abbott 2005). Nevertheless, YCA supercolonies on Christmas Island have always been
defined in terms of very high ant densities. YCA occurred at many locations across the island in very low
abundance with no obvious impact on biodiversity, but in 1989, and then again in late 1997, YCA was
discovered in several locations at extremely high densities sufficient to extirpate local populations of the
abundant red land crab Gecarcoidea natalis (O’Dowd et al. 1999, 2003). The red crab is a keystone
species in rainforest on the island that controls patterns seedling recruitment and rates of litter
breakdown and nutrient cycling (Green et al. 1997, 1999, 2008). Since then, the density at which YCA
kills land crabs leading to understorey transformation has become the operational definition of
‘supercolony’ on Christmas Island. On the ground, the density of YCA in supercolonies can be
extraordinary – up to several thousand per square meter (Abbott 2005). A systematic, island‐wide
survey in 2001 found multiple supercolonies ranging in area from tens to hundreds of hectares, totalling
c. 2500 ha, or 25% of rainforest on the island (Green et al. 2004; Green and O’Dowd 2009).
Supercolonies have continued to form and reform; upwards of 5000 ha of rainforest have been treated
with toxic bait to 2012.
The key trait that has allowed YCA to form high‐density supercolonies is its ability to form mutualistic
associations with honeydew‐producing hemipterans, principally scale insects (O’Dowd et al. 2003;
Abbott and Green 2007). The scale insects suck sap from trees and secrete carbohydrate‐rich
honeydew on which the ants feed. The ants provide sanitation services for the scale insects, removing
honeydew that might otherwise build up and kill them either through asphyxiation or the growth of

6
sooty moulds, but the ants may also provide limited protection for the scale insects from generalist
natural enemies. Supercolony‐level densities of YCA and outbreak‐densities of several species of scale
insects invariably co‐occur (Abbott 2004), and in supercolonies high densities of ants can typically be
seen ascending the boles of most trees to tend scale insects in the canopy. The gasters of descending
ants are swollen with carbohydrate‐rich honeydew that they take back to the nest to share with non‐
forager conspecifics. Site‐scale, manipulative experiments on Christmas Island have demonstrated a
causal link between co‐occurring high densities and ants and scale; the exclusion of ants using toxic bait
leads to a dramatic decline in scale abundance (Abbott and Green 2007). Part of the current project
was to determine if this link is bi‐directional, that is, if the exclusion of scale insects from the ants leads
to a dramatic decline in ant density (see Project 1c, below). On Christmas Island YCA interacts with
several species of honeydew‐secreting scale insects including Tachardina aurantiaca, the principal
target of the biocontrol program (see below).
The mutualism between YCA and scale insects has manifold negative impacts on species abundances,
interactions among species, and forest structure. At the core of these impacts is the devastating effect
of YCA on land crabs, especially red crabs. YCA sprays formic acid as a weapon both to subdue prey and
in self‐defence, and although the amount sprayed by individual ants is tiny, at supercolony densities the
overall effect is overwhelming. YCA supercolonies reduce formerly high densities of red crabs
(averaging c. 0.5 – 1.0 crabs m‐2) to nil, deregulating seedling recruitment and litter dynamics and
resulting in a thick, diverse understorey of seedlings and saplings with an almost permanent layer of leaf
litter (O’Dowd et al. 2003). In forest dominated by red crabs, the understorey is sparse and dominated
by a few crab‐resistant species, and the forest floor is almost devoid of litter for much of the year
(Green et al. 1997, 1999, 2008). These impacts are widespread. Based on the spatial extent of
supercolony formation over the last 12 years, it is likely that YCA has extirpated at least 20 million red
land crabs, or about 30% of the total population in areas where they have formed supercolonies.
YCA has also caused declines in the density of red land crabs at sites where high‐density supercolonies
have never formed. About half of the red crab population migrates to the coast each year to complete
breeding activities, and many YCA supercolonies have formed across traditional crab migration routes.
Thus, significant numbers of migrating red crabs have been killed en route to the coast over many years,
never to return to their former home ranges. As a result, some areas of rainforest are practically devoid
of red crabs even though YCA supercolonies have never occurred there, and the same processes of
understorey transformation are in train there too. It is hard to gauge the severity and extent of this
“ghosting” effect because pre‐YCA invasion data on red crab densities are sparse. It is likely to be
significant; Green et al. (2011) estimated that around 25% of rainforest may have been ghosted at some
time in the last decade. The direct and indirect (ghosting) impacts of YCA supercolony formation have
been so widespread since the late 1990s that just 28% of rainforest could still be considered as “intact”
(i.e. no YCA supercolony formation and unaffected by ghosting) by 2007 (Green et al. 2011).
In YCA supercolonies, scale insects themselves can have large negative impacts on their host plants.
Especially vulnerable is the Tahitian Chestnut Inocarpus fagifer, a widespread canopy tree that hosts
very high densities of Tachardina on its outer twigs and leaves. In supercolonies, seedlings, saplings and
small trees all suffer extremely high mortality, and the canopies of large trees are much reduced
through the dieback of fine twigs and branches (Green et al. 2001, O’Dowd et al. 2003, P. Green and D.
O’Dowd, unpublished results). There is also evidence that fruit production is reduced in supercolonies.
Excess honeydew that YCA does not harvest, or which the scale insects deliberately flick off, settles on

7
leaves of all plant species and is colonised by sooty moulds, which probably interferes with
photosynthesis and growth.
YCA may affect many of the island’s bird species through direct interference and through altered
resource availability and habitat structure (Davis et al. 2008). The Christmas Island Emerald Dove
Chalcophaps indica natalis is 9‐14 times less abundant in ant‐invaded forest, and because it forages on
the forest floor, is probably vulnerable to direct predation by YCA. The nesting success and density of
juvenile Christmas Island Thrushes Turdus poliocephalus erythropleurus is lower in supercolonies, where
they also show altered foraging and reproductive behaviours. Furthermore, these birds alter their
choice of tree species in which to build nests, with lower frequency on tree species that typically
experience high densities of scale insects and ants. The density of foraging Christmas Island white‐eye
Zosterops natalis is higher in supercolonies, perhaps because scale insects (as prey) are more common
there. It is possible that impacts of YCA on thrushes and white‐eyes affect frugivory and seed dispersal
on the island; assays with both real and model fruit showed handling rates to be more than two‐fold
lower in YCA supercolonies, and manipulative experiments showed this to be a direct consequence of
the presence of ants (Davis et al. 2009). There is no evidence that YCA supercolony formation
significantly affects the density of nesting success of Abbott’s Booby Papasula abbotti (P. Green,
unpublished data), while the impact of YCA on other seabirds and on other land birds such as the
goshawk and owl are unknown.
Several endemic vertebrate species, including the pipistrelle bat Pipistrellus murrayi and endemic
reptiles have experienced precipitous declines over recent years, but the causes of the declines are
enigmatic. In the case of the pipistrelle it is possible that YCA supercolony formation has contributed to
the decline, either directly through predation of bats at roost sites or indirectly by eliminating red crabs
and facilitating the expansion of predators such as giant centipedes Scolopendra subspinipes, wolf
snakes Lycodon aulicus, cats and rats (Schulz and Lumsden 2004, Lumsden et al. 2007, Beeton et al.
2010). However, the decline of the pipistrelle was well in train before the rise of YCA supercolonies in
the late 1990s. The endemic reptiles were similarly in decline long before YCA supercolonies became
common, and the role of YCA in their decline is also uncertain (Smith et al. 2012).
In addition to impacts on species of concern, supercolony formation by YCA has also led to an altered
web of species interactions that facilitates the entry, spread, and abundance of other invasive species.
The best example of this is the entry and spread of the giant African landsnail Achatina fulica (GALS) in
rainforest on the island. GALS was introduced to the island in the 1940s, and despite being a
notoriously invasive species (Lowe et al. 2000), its distribution was for many decades limited to settled
areas, abandoned mining fields and roadsides. Experiments showed that predation by red crabs
excluded this invader from establishing in rainforest (Lake and O’Dowd 1991), but the extirpation of red
crabs in YCA supercolonies, coupled with the ability of GALS to coexist alongside YCA in supercolonies,
has permitted this snail to establish high densities in rainforest at many locations across the island
(Green et al. 2011). The facilitation of GALS by YCA could be due to the creation of enemy‐free space,
augmented understorey resources, or both.
The rise of YCA supercolonies and the extirpation of red crabs have also affected the invasion dynamics
of other non‐native organisms. These effects encompass both inhibition and facilitation for a range of
non‐native ant and snail species (O’Dowd and Green 2010; P. Green and L. O’Loughlin, unpublished
results), while invasion by several weeds including Capsicum frutescens, Carica papaya, Cordia
curassavica, and Muntingia calabura appears to be facilitated in areas affected by YCA supercolony
formation (P. Green and D. O’Dowd, personal observations).

8
Management. Given all of the above, supercolony formation by YCA is considered a major and on‐
going threat to biodiversity values on Christmas Island. To date, the management of YCA has depended
on surveillance, monitoring, and control using toxic bait (Green and O’Dowd 2009, Boland et al. 2011).
New supercolonies continue to form, and there is concern for the sustainability of this program in terms
of its expense, non‐target impacts, and the resources it diverts from other conservation programs
(Beeton et al. 2010). Further, this program can only ever be reactive, and it has not been able to find an
effective solution to the difficult issue of the management of incipient supercolonies, where YCA density
is high enough for them to monopolize the toxic bait, making the risk of non‐target impacts too high to
justify the treatment of these areas. There is widespread agreement that the development of a more
cost‐effective, sustainable alternative to the use of toxic bait is needed to manage the YCA invasion on
Christmas Island (Beeton et al. 2010).
Ten years ago, O’Dowd and Green (2003) suggested that long‐term, sustainable suppression of YCA
supercolonies could be achieved using biological control agents. Classical biological control works on
the principle that in their area of origin, populations of native species are kept in check by their natural
enemies (predators, parasites or pathogens). There is a large body of literature demonstrating that in
many cases, species introduced outside of their native ranges become invasive because they have
effectively left their enemies behind. This is known as the “Enemies Release Hypothesis”. The principle
of classical biological control then is the converse: to re‐establish population control over invasive
species by first identifying and then importing a natural enemy – a biological control agent – from within
the native range of the target organism. In all instances, this now involves selection and testing of
biological control agents to verify narrow host ranges that minimize the risk of non‐target impacts in the
area of introduction.
Indirect Biocontrol for YCA on Christmas Island. Despite the diversity and significant ecological
and economic impacts of invasive ants worldwide, they have proved to be an especially difficult target
group for biological control. Although a program for the biocontrol of the Red Imported Fire Ant
(Solenopsis invicta) using a parasitic fly and a protozoan disease as agents is currently under
development in the southeastern United States, no species of ants have yet been successfully controlled
in the field using biological control agents and principles. Instead, O’Dowd and Green (2003) proposed a
novel solution for managing the YCA invasion; rather than targeting the ant itself, biological control
could target the key mutualist species (honeydew‐producing scale insects) that are likely to play a
significant role in sustaining YCA supercolonies at very high and ecologically damaging densities. Long‐
term, sustainable suppression of YCA supercolonies could be achieved through the introduction of a
biological control agent that would indirectly affect YCA by reducing carbohydrate supply provided by
honeydew‐producing scale insects, a key resource implicated in supercolony dynamics.
The Biocontrol Project 2009 – 2013. Funding for a four‐year program of research and development
to progress the concept and feasibility of indirect biocontrol was allocated in the Australian Federal
Budget 2007/08, as part of a broader, 10‐year plan for the control of YCA on Christmas Island. We
proposed a research project with three overarching questions:
1. Do YCA depend on honeydew‐producing scale insects? 2. Which species of honeydew‐secreting scale insects occur on Christmas Island, and are appropriate
natural enemies for scale insects available for introduction?
3. What regulatory frameworks govern the implementation of a biological control program on
Christmas Island, an external territory of Australia?

9
Two Postdoctoral Fellows were employed to perform most of the research under Questions 1 & 2. Dr.
Sarah Wittman was recruited from the University of Vermont in the United States, with a project brief
to establish the dependence of YCA on honeydew‐producing scale insects. She arrived on Christmas
Island in November 2009, and finished in December 2012. She is currently back in the United States
working on the preparation of manuscripts for publication arising from her work. Dr. Gabor Neumann
was recruited from the USDA‐ARS in Hilo, Hawaii as a biocontrol practitioner, with a project brief to
investigate the scale insects of Christmas Island, to describe their local suite of natural enemies, and to
search for suitable biocontrol agents in Southeast Asia. His tenure on this contract expires in July 2013.
The biocontrol project was overseen by an independent steering committee, the Christmas Island Crazy
Ant Scientific Advisory Panel (CASAP), which comprises scientists, managers, and policymakers. CASAP
provided expert advice and reviewed progress through panel meetings and evaluation of interim and
annual reports.
Structure of the Final Report. The broad research questions (above) were refined into three
“Research Projects” and 13 subprojects, that were listed as Services in the Schedule to the Contract
between the Director of National Parks and La Trobe University, signed on 29 January 2009. Below, we
report against these subprojects as they are laid out in the Contract Schedule, and provide an
introductory rationale for each using text (in italics) copied from the original Schedule.

10
Research Project 1. YCA dependence on honeydew
Numerical dominance and impacts of invasive ants may be sustained by their association with
honeydew‐secreting Hemiptera. For example, ~1/2 of the energy supply of the red imported fire
ant is obtained from honeydew secreted by Hemiptera and ecologically dominant ants in the
rainforest canopy are highly dependent on honeydew. How dependent is the YCA on honeydew
obtained from scale insects and is honeydew critical to population size? Research outlined
below is directed at estimating the use and sources of honeydew by the YCA on Christmas
Island, the importance of carbohydrates in its diet, and the role of honeydew in sustaining high
populations.
Project 1a. Honeydew use by YCA
Mutualisms between invasive ants and Hemiptera may drive positive population growth leading to
numerical dominance of ants. This project will give a quantitative picture of honeydew use by the YCA
and focus on the relative importance of the lac scale Tachardina aurantiaca and soft scale Coccus spp. as
sources of honeydew. Knowledge of the importance of honeydew derived from lac scale is critical since
in the first instance they are the proposed target of biological control.
Research Findings 2009‐2013
The original intention of this project to combine data on per capita honeydew production by the most
important (common) honeydew‐producing scale insects, analyses of the quality of honeydew (sugars and
their concentrations), and estimates of scale abundance and YCA visitation rates to estimate empirically
the ‘honeydew economy’ of YCA supercolonies, and the relative contribution of Tachardina to that
economy. However, collecting sufficient honeydew for analyses using glass microcapillary tubes proved
technically very challenging, there were also issues (packaging, quarantine) of transporting fresh samples
to our preferred analytical laboratory in Melbourne. The logistics of collecting spatially and temporally
replicated data on YCA visitation rates to scale insects using video recordings was also extremely
challenging.
Instead, we devised an indirect measure of the honeydew economy, called the Site Honeydew Index
(SHI). The SHI is a relative estimate of the total volume of honeydew that could be produced at a site. It
is a compound measure that considers the capacity of different plant species to host either Tachardina
aurantiaca, or coccoid honeydew‐producing species of soft scale insects, together with tree size and tree
abundance. The basis of the SHI is canopy surface area, generated for each tree using published
allometric equations that predict crown dimensions from stem diameter (Poorter and Bongers 2006).
Presumably, large trees have the capacity to host more scale insects, and canopy surface area is
preferable to canopy volume because scale insects live on leaves and thin twigs of the outer canopy. This
estimate is then multiplied by the average abundance of scale insects per unit length of stem, using host
species‐specific data for both Tachardina and soft scales (data from Table 4.3 in Abbott 2004). The SHI is
the sum of these products, divided by 10000 for convenience. Thus, the SHI is the sum of the
contributions from individual trees for both Tachardina and soft scales, weighted by tree size (canopy
surface area) and species identity. SHIs were calculated for 10 50 m x 50 m plots in YCA supercolonies,
on which all trees ≥ 5 cm dbh were enumerated in June‐October 2000 (P. Green, unpublished data). The
SHI varied 1.6‐fold (from 78 to 125) suggesting that the capacity of forest stands to support YCA
supercolonies varies considerably. Tachardina is estimated to contribute a large fraction of the

11
honeydew economy at forest sites with YCA supercolonies (mean = 70%, range 46‐86%; blue symbols in
Fig. 1). This assumes per capita parity in the quantity and quality of honeydew produced by Tachardina
and the coccoid soft scales species. However, adult female Tachardina are much larger than the adult
females of the coccoid species, and its reasonable to suppose that per capita honeydew production is
much higher in Tachardina. Assuming a three‐fold higher rate of honeydew production, Tachardina may
contribute a mean of 87% to the SHI (range 72 – 95%; red symbols in Fig. 1). Successful biological control
of the yellow lac scale Tachardina aurantiaca could remove a large fraction of honeydew available to the
yellow crazy ant. One caveat to this conclusion is that the balance between the contributions made by
Tachardina and the coccoid soft scales to the total honeydew economy may have declined over time at
some sites where supercolonies have reformed after baiting. Inocarpus fagifer is a key host species for
Tachardina in YCA supercolonies; just this single species contributed an average of 50% to the Tachardina
component of the SHI in 2000. The species is so heavily utilised by Tachardina and YCA that significant
tree mortality occurred in supercolonies between 2000 and 2002 on the 10 50 x 50 m tree census plots
(P. Green, unpublished data). Although these supercolonies were destroyed by the aerial baiting
campaign of 2002 (Green and O’Dowd 2009) and YCA supercolonies have not reformed on these sites
since (D. Maple, unpublished data), a targeted recensus of Inocarpus on 8 plots in 2012 showed some
further dramatic declines, possibly caused by Tachardina tending by the ant Camponotus sp. in the
decade after baiting. The death of many mid‐ to large‐sized Inocarpus trees has resulted in a decline of
that species’ contribution to the SHI by an average of 50% (range 11 – 91%; Fig. 2). The implication of this
finding is that where contemporary YCA supercolonies have reappeared at former but baited YCA
supercolony sites, the new supercolonies may now be supported to a greater extent by soft scales, and
because of that, the introduction of a parasitoid against Tachardina may be less effective in those
locations. The extent of this issue could be estimated from the Island‐Wide Survey data by determining
the proportion of supercolonies that reform, and the proportion of time supercolonies occupied each
site.
Key Research Outcome: Successful biological control of the yellow lac scale Tachardina aurantiaca could
remove a large fraction (an average of 70% to an average of 87%, depending on assumptions) of
honeydew available to the yellow crazy ant. However, there is considerable site‐to‐site variation in the
likely contribution of T. aurantiaca to the total honeydew economy, and it is not certain that targeting
this species alone would provide adequate indirect control for the YCA in all supercolonies. The most
prudent course of action is to target the entire assemblage of honeydew‐producing scale insects through
the introduction of a biological control agent for T. aurantiaca, complemented by the use of parasitoids
already present on Christmas Island to target the coccoid soft scales.

12
Project 1b. Stable Isotope Analysis
Analyses of Nitrogen (N) isotope ratios show that ecologically dominant ants and many invasive
ants obtain little N through predation and scavenging, but feed primarily as herbivores, deriving
carbohydrate through exudates produced by Hemiptera, primarily scale insects. This project
will use this widely accepted technique to determine N and C stable isotope ratios of the YCA,
other ant species, and known primary producers, herbivores, and predators to characterize how
much of the dietary requirements of the YCA is derived from plant‐derived carbohydrates and
how this changes with invasion dynamics.
Research Findings 2009‐2013
Honeydew from scale insects is essentially plant sap that has passed through the bodies of scale insects,
so when feeding on honeydew YCA are essentially acting as herbivores. If YCA at supercolony densities
derive most of their dietary intake from scale insects, then stable isotope analysis of their body parts
(δ15N) should indicate a mainly herbivorous diet, while YCA at non‐supercolony densities should show a
less herbivorous diet.
We estimated the change in trophic position of YCA at four sites (Island Wide Survey points) as the
abundance of ants declined from supercolony to non‐supercolony densities. YCA workers were sampled
6 to 8 times over a 16‐month period (April 2010 – September 2011). Trophic position was calculated
from mixing models that considered site‐specific stable isotope ratios of known plants, herbivores (stick
insects), and carnivores (spiders). All samples were analysed by a commercial company, Natural
Isotopes, in Perth, Western Australia.
Stable isotope analyses showed that trophic position increased (i.e. YCA became more carnivorous) as
population densities decreased over 16 months (Site 206, R2 = 0.75, P = 0.025; Site 318, R2 = 0.67, P =
0.025; Site 403, R2 = 0.58, P = 0.046, Site 582, R2 = 0.72, P = 0.033)(Fig. 3). This result suggests that a
waning carbohydrate supply, as indicated by an upward shift in the trophic position of YCA, is associated
with supercolony decline. A series of YCA collected over two years from 10 supercolonies in 2000‐2002
were also analyzed for δ15N after storage in ethanol for over a decade. Unlike the four sites above, δ15N
values and YCA abundance were not negatively correlated at three supercolonies that declined to zero
over the period. However, as expected, δ15N values for the other seven supercolonies with
consistently high YCA densities did not vary over time.
Key Research Outcome: Stable isotope analyses of YCA workers, plants, herbivores and predators
collected from four declining supercolonies in 2010‐2011 indicate that at supercolony densities, a
substantial fraction of YCA dietary intake is plant‐derived. This is consistent with the idea that YCA
supercolonies depend heavily on honeydew derived from scale insects for a large fraction of colony
food and energy requirements, and provides support for the idea that indirect control over YCA
supercolonies could be achieved by targeting honeydew‐producing scale insects for biological control.

13
Project 1c. Scale Insect Removal Experiment
Although Projects 1a and 1b above will indicate the importance of honeydew in the diet of YCA
and the relative contribution by scale insect type, large‐scale field experiments are needed to
establish the influence of scale insects on population dynamics of the YCA. This experiment uses
the systemic insecticide imidacloprid to remove scale insects in YCA supercolonies and follows
ant population dynamics.
Research Findings 2009‐2013
The original intention of the is experiment was to use the systemic insecticide imidacloprid to remove
scale insects from an experimental plot, and to monitor ant activity before and after to determine the
dependency of YCA at supercolony densities on scale insects. This was essentially a proof‐of‐concept
experiment for indirect biocontrol. The intent was sow the treatment plot with tablets containing
imidacloprid, which would be taken up by the root systems of trees and transmitted to scale insects,
removing them from the canopy. This would mimic the effect achieved through the introduction of a
biological control agent for scale insects on the honeydew supply to YCA.
The experiment was not carried out as planned. Although imidacloprid is routinely used in single
species Eucalyptus plantations to control herbivorous insect pests, it has not been used in natural
forests where plant species diversity, and a range of size classes of trees from sapling to canopy giants,
poses significant challenges for achieving consistent uptake of the toxin across all individual trees. In
addition, analyses of honeydew would have been mandatory to test the possibility that any decline in
YCA density on the treatment plot was more likely due to scale insect death and a reduction in the
honeydew supply, rather than a toxic effect due to the presence of imidacloprid or it metabolites in
honeydew. As pointed out above, collecting and preparing honeydew for analysis was challenging in
itself. Instead, we used a physical barrier applied to tree trunks to prevent YCA from accessing their
honeydew supply in tree canopies.
The experiment was a Before‐After‐Control‐Impact (BACI) design on two forest plots in a YCA
supercolony abutting the Winifred Beach Track. There were two plots, each 40 m x 40 m, and one was
designated as the control plot, the other as the treatment plot. On the treatment plot, tree bands were
made by winding Gladwrap™ around the boles of all trees >2 cm DBH. Bands blocked YCA traffic flow to
and from the forest floor, resulting in “log jams” of downward moving ants, most replete with
honeydew, above the bands (Fig. 4a). Throughout the experiment these stranded ants were returned to
the forest floor by brushing them gently off each bole, so that any decline in YCA abundance on the
ground could not be attributed to the retention of these ants on trees. Repeated application of Mr
Sheen™ spray‐on furniture polish to each band over the experiment greatly increased the effectiveness
of the barriers. No bands were applied to trees on the control plot.
The measured response variables were YCA trunk traffic and YCA ground activity, both measured in the
inner 20 x 20 m core of each plot. YCA trunk traffic was estimated by counts per 30 s in a 10 x 10 cm
quadrat on each of 10 trees, while ground activity was monitored using ant counts per 30 s on one
quadrant on each of twelve 20 x 20 cm cards. (Fig. 5a). Both variables were monitored at 3‐4 day
intervals, 9 times before and after applying tree bands, during the dry season in 2012. Results were
analyzed as a one‐way repeated measures ANOVA, using ant exclusion from the forest canopy as the
main factor and time as the repeated measures factor. In this design, the time x treatment interaction is
the key term, with a significant treatment difference after, but not before treatment application.

14
Bands effectively excluded YCA from the canopy, resulting in a precipitous decline and the virtual
elimination of YCA traffic on tree boles 4 weeks after the tree bands were in place (Fig. 4b; Treatment x
Time interaction, F1,32 = 90.198, P = 0.000). YCA abundance on the ground fell and diverged markedly
from the control plot two weeks after tree bands were placed (Fig. 5b; Treatment x Time interaction,
F1,32 = 37.604, P = 0.000). YCA abundance declined ~3‐fold compared to average pre‐exclusion values,
and was ~5‐fold lower than on the control plot 4 weeks. If card counts are converted to YCA densities
(using the regression y = 15.694x – 21.612; Abbott 2005), YCA densities on the forest floor fell from ~400
m‐2 before to ~140 m‐2 after exclusion from the canopy, and when the experiment was terminated were
~600 m‐2 on the control plot compared to ~100 m
‐2 on the exclusion plot. We attribute the decline in
YCA density on the ground to their exclusion from honeydew resources from scale insects in the canopy.
Two alternative explanations are not likely. First, plot disturbance when setting up the tree bands on
the treatment plot could have affected YCA activity on the ground but would be unlikely to explain the
magnitude of change in YCA abundance after bands were in place. Second, this experiment excluded
YCA from all canopy resources, not just honeydew, and its possible that it was exclusion from other food
sources, such as invertebrate prey, that caused the observed decline in density on the ground. In
anticipation of this, video records were made of samples of ants descending tree trunks to determine
the proportion with distended shiny gasters (indicative of honeydew foraging), versus the percentage of
workers carrying prey items. These records are still being processed, but observations indicate most
workers were foraging for honeydew, and < 1% were carrying prey.
Key Research Outcome: The exclusion of YCA from access to scale insects at a large experimental field
site caused YCA activity on the ground to decline 5‐fold within 4 weeks, compared to pre‐treatment
levels. This large field experiment validates the key concept of indirect biological control for YCA on
Christmas Island; exclusion of honeydew‐producing scale insects from YCA caused a significant and
substantial reduction in YCA abundance on the ground.

15
Project 1d. Carbohydrate supply and YCA growth and behaviour
The importance of simple carbohydrates in the population growth and behaviour of the yellow
crazy ant (YCA) will be assessed by varying carbohydrate supply to laboratory colonies and
measuring brood production, worker survival, and aggression.
Research Findings 2009‐2013
This experiment consisted of nine treatments that varied the absolute amount, but not the
concentration, of sugars available to YCA colonies. Eighteen YCA nests were set up in plastic containers
sitting in water traps to contain the ants. Each nest initially contained 2 queens and 200 workers, and
they had unlimited access to water and protein (thawed crickets) for the duration of the experiment.
Two nests were assigned to each of nine sugar treatments, 0, 10, 20, 40, 80, 160, 320, 640, and 1280 µl,
delivered as 13% honey water every 3‐4 days for two months. In the last week, novel objects (a small
frame of bamboo skewers) were added to track YCA exploratory behaviour, and behavioural assays
against the big‐headed ant Pheidole megacephala were conducted to assess the impact of sugar supply
on interspecific aggression of YCA. In these aggression assays, three YCAs and three Pheidole were
placed in a 6 cm diameter vial with fluon‐coated sides for 10 minutes. The time to the first spray of
formic acid by each YCA, the total number of sprays by each YCA, and the total number of P.
megacephala dead after 10 minutes was recorded.
Performance and foraging behaviour of yellow crazy ants depend on sugar supply. Per capita
recruitment to sugar indicated that a smaller fraction of colony workers was needed to collect sugar
with increasing sugar supply (Fig. 6a). Colony performance, as measured by production of workers and
males (Fig. 6b) increased with sugar availability (P = 0.034) whereas per capita death rate decreased
with increasing sugar availability (Fig. 6c). Per capita encounter rate of YCA with novel, non‐food objects
placed in the nest, a measure of foraging tempo, also increased with sugar supply (Fig. 6d).
Aggressive behaviours in the yellow crazy ant increased with sugar supply. YCA with access to more
sugar sprayed P. megacephala with formic acid sooner and more often (Fig. 7a; P = 0.006), and killed
more P. megacephala in 3:3 interaction trials (Fig. 7b).
Key Research Outcome: Dynamics and behaviour of YCA in laboratory colonies depended on
carbohydrate supply. When sugar supply was elevated, reproductive output by queens increased,
death rates of workers decreased, foraging tempo quickened, and interspecific aggression intensified.
These results suggest that sugar supply, through honeydew supplied from scale insects, plays an
important role in YCA supercolony dynamics, foraging efficiency, and interspecific aggression.

16
Research Project 2. Scale insects and natural enemies
For indirect biological control of the YCA to be feasible, appropriate natural enemies must be
available. The most likely source of specialist natural enemies is in association with the target
scale insect in its area of origin. Our co‐operator (Dr. R. Pemberton, USDA‐ARS Ft Lauderdale
FL) is already well advanced in development of biological control of the lac scale Paratachardina
sp. nov. and recently determined SEA as its area of origin. Further, he agreed to search for and
collect Tachardina aurantiaca in Thailand and Malaysia for us from which they have now reared
several species of hymenopteran parasitoids that are currently being described. Research
outlined below is directed at building on these preliminary results to locate and evaluate
potential biological control agents for T. aurantiaca in SEA, defining the scale insect fauna of
Christmas Island, determining any natural enemies of scale insects already on Christmas Island,
and establishing a pre‐release monitoring program for T. aurantiaca.
Projects 2a and 2c. Natural enemies survey, parasitization rates of lac scale
insects (Tachardina aurantiaca and Paratachardina pseudolobata) and
parasitoid behaviour in area of origin (Southeast Asia) and on Christmas Island
2a Collection of scale insect material to rear enemies and identification of candidate parasitoids
for consideration in biological control. Species descriptions, biology and behaviours of suitable
candidates as biological control agents. Separation of parasitoids into wanted primary
parasitoids and undesirable hyperparasitoids.
2c Identification of existing natural enemies (if any), and predation and parasitism rates on key
scale insects on Christmas Island. This will identify if significant natural enemies of scale insects
are already present on the island.
Research Findings 2009‐2013
This part of the research very quickly focused on Tachardina aurantiaca as the key Kerriidae honeydew
producer on Christmas Island. Although Paratachardina pseudolobata produces honeydew, females
flick droplets away from their bodies (Howard et al. 2010) making it unavailable to foraging YCA. Below,
we present information on the target T. aurantiaca including its geographic range in Southeast Asia, its
life cycle, its natural enemies and incidence of parasitism in Southeast Asia and on Christmas Island, and
a description of the biology of its candidate biological control agent, Tachardiaephagus somervillei. We
conclude this section with a short description of the suite of parasitoids of soft scales found on
Christmas Island.
Distribution of T. aurantiaca in Southeast Asia. We located populations of T. aurantiaca across a 1900‐
km east‐west distribution in Sundaland, its putative area of origin (Fig. 8). This region is the part of
Southeast Asian continental shelf that was exposed during the last ice age, encompassing Peninsular
Malaysia, Borneo, Java, and Sumatra). Much of the search for T. aurantiaca in Malaysia was targeted,
using a highly suitable and widespread host plant, Milletia pinnata, as a focus. Search effort totalled 27
days In Malaysian Borneo, and 5 days were spent searching in Singapore. Live aggregates of yellow lac
scale (yellow circles in Fig. 8) were located on Penang Island, and in Klang (Selangor) and Kepong

17
(Selangor) in Peninsular Malaysia. In Singapore, T. aurantiaca was found on the campus of the National
University of Singapore. In Sarawak, live aggregates were discovered around Santubong and Kuching
(two sites ‐ Kampung Istana and Kampung Boyan). In Sabah, live aggregates were found in the Sandakan
area and Sepilok. Dead aggregates (not shown above) were found in Melaka, peninsular Malaysia and
in Temon, 190 km south of Kota Kinabalu, Sabah. Ants tended T. aurantiaca at all sites and collected
honeydew (Penang – Crematogaster sp.; Klang – Oecophylla smaragdina; Kepong – Dolichoderus sp.;
Kampung Istana – Anoplolepis gracilipes; Kampung Boyan – Oecophylla smaragdina; Singapore,
Sandakan and Sepilok – Anoplolepis gracilipes).
Assemblage of natural enemies of T. aurantiaca in Southeast Asia and on Christmas Island. On
Christmas Island, search focused on seven areas (see Figure 12) and examined over 11,000 females and
2000 males of the yellow lac scale. In Malaysia, targeted search for T. aurantiaca on known host plants
(e.g., Milletia pinnata, Acacia mangium x A. auriculiformis) was frequently used to locate T. aurantiaca.
Because the yellow lac scale is so rare across Malaysia, many fewer individuals were inspected for
parasitization. To determine natural enemies, host plant twigs with aggregates of T. aurantiaca were
first inspected, and then isolated so that emerging parasitoids could be collected for later identification.
Female T. aurantiaca, either individually or in aggregates, were inspected for parasitoid exit holes to
estimate parasitization rates. Individual females were also isolated to collect emergent parasitoids and
dissected to determine overall rates of parasitization.
The assemblage of natural enemies of T. aurantiaca is much more diverse in the area of origin that in its
introduced range on Christmas Island (Table 1). Using a combination of historical records (Noyes 2012)
and direct field surveys, there are six primary parasitoids that use T. aurantiaca as a host in Malaysia,
but only one on Christmas Island. Furthermore, the one parasitoid on Christmas Island, Marietta
leopardina, can only successfully attack males of T. aurantiaca. Females of this species attempt to lay
eggs inside females of Tachardina, but the test is seemingly too tough for the ovipositor to penetrate.
In addition to parasitoids, two lepidopteran predators of female T. aurantiaca were also been found in
both the native and introduced ranges, but on Christmas Island these are extremely rare. Two
hyperparasitoids, Promuscidea unfasciativentris and Aprostocetus purpureus, were isolated from T.
aurantiaca in Malaysia, but not on Christmas Island (Table 1).
Parasitism of T. aurantiaca in its native and introduced ranges. In Southeast Asia, T. aurantiaca is rare
and evidence of female parasitization was frequently seen in its native range in SE Asia (Fig. 9). In
contrast, in its introduced range on Christmas Island, T. aurantiaca is superabundant and there was no
evidence of female parasitization (Table 2). In Southeast Asia, the rate of parasitism was high, varying
from a mean of 29% to 81% at different sites. The rates in Table 2, based on the incidence of
emergence holes, almost certainly underestimate actual parasitization rates because parasitized
females from which parasitoids have not yet emerged would not be included. To investigate this,
parasitisation rates of 5 additional aggregates in Sarawak were assessed by counting emergence holes,
and then by dissecting the females to count unemerged parasitoid larvae. Parasitization was estimated
as 44.6 ± 9.6% (mean ± SE) by emergence holes, but as 61.4 ± 7.7% when females with unemerged
larvae were added. As a further example, a small aggregate near Kuching that showed no visible sign of
parasitization (i.e. no exit holes) was found to be 100% parasitised after dissections.
Tachardiaephagus somervillei parasitized T. aurantiaca across all sites examined (Fig. 9a).
Superparasitism, where more than one progeny is produced per host individual, is frequent. The mean
number of T. somervillei emerging from each parasitized female T. aurantiaca was 2.1 (range 1‐ 4, N =
30) and up to 5 emergences occurred from each female at sites in Peninsular Malaysia. Coccophagus

18
tschirchii and C. euxanthodes were also isolated from T. aurantiaca (Table 1), but these species were
uncommon and found only in Peninsular Malaysia (Fig. 9b).
In contrast to the widespread parasitism of T. aurantiaca in Southeast Asia, parasitism of females was
never observed on Christmas Island. More than 11,000 females from multiple sites (Table 2) were
collected and inspected under magnification over two years to determine the presence of any
parasitoid emergence holes. Single exit holes were observed in some male T. aurantiaca at a few sites.
In the laboratory, the primary parasite Marietta leopardina emerged from these males. Parasitization
rates were low, ranging from 0 ‐ 10% (N= 558, 751, and 696 males examined on three trees at Hugh’s
Dale, Anderson’s Dale, and Sydney’s Dale), but M. leopardina is clearly not an effective biological control
agent of female T. aurantiaca on the island. High densities of intact females occured where M.
leopardina was present and, on Christmas Island, T. aurantiaca may be parthenogenetic (G. Neumann,
unpublished results).
Biology, ecology and life cycle of the parasitoid microhymenopteran wasp, Tachardiaepagus
somervillei. T. somervillei was first described by Mahdihassan in 1923 as Lissencytus somervilli but in
1928 Ferriere transferred L. somervillei Mahdihassan to Tachardiaephagus and also later downgraded
the species somervillei to a variety of T. tachardiae Howard (Hayat et al. 2010). However, Hayat et al.
(2010) regarded T. somervillei as a valid species. T. somervillei is known to occur in India, Thailand and
Malaysia (Hayat et al. 2010 and references within). During our exploration for natural enemies of
Tachardina aurantiaca, T. somervillei was found and its identity was confirmed by Dr. Mohammad Hayat
(Table 7) from sites in Selangor, West Malaysia and in southwestern Sarawak, East Malaysia. We also
observed this parasitoid in eastern Sabah, East Malaysia but its identity has not yet been confirmed.
With its large geographical range (Fig. 8), T. somervillei is a very widespread and frequent natural enemy
of T. aurantiaca.
Host records for T. somervillei indicate that this parasitoid attacks four species in the genus Kerria
(Noyes 2012) and T. aurantiaca (Table 3). The host records sometimes do not specify the species of
Tachardina parasitized but the only known species in the genus Tachardina within the recorded
geographical range of T. somervillei is T. aurantiaca. The apparently restricted host range of T.
somervillei based on historical records is not surprising: all known parasitoids within the genus
Tachardiaephagus have been recorded only from lac scales (Kerriidae), with the single exception of a
coccid (Ceroplastes eucleae) recorded as a host of T. similis in Africa. This latter host record, however,
has been discounted by Prinsloo (1977). While host ranges of the species in the genus
Tachardiaephagus have not been assessed in detail, Sharma et al. (2006) mentions T. somervillei (along
with T. tachardiae) as “exclusive to lac ecosystem”.
Besides taxonomic work, very little research has been done on the biology of Tachardiaephagus spp.
even though the species attacks economically important lac scales, such as Kerria lacca (Kerr) and are
considered pests in lac‐producing regions (Sharma et al. 2006). Our initial observations in peninsular
Malaysia and Sarawak provide basic information on the life cycle and biology of T. somervillei in relation
to T. aurantiaca. The basic biology and life cycle is similar to that of other encyrtids that attack scale
insects. T. somervillei attacks mature female hosts, but never attacks males. Parasitoid eggs are
deposited inside the host’s test (the test is a secretion that covers the scale insect’s body), probably
through the anal pore, and the hatching parasitoid larvae consume the host’s body inside the test. The
larva pupates after larval development is completed. The adult wasp emerges from the pupa and
through a round, smooth‐edged whole which it drills through the test and exits the dead host.
Development time from oviposition to adult emergence can vary, but it is usually about 3 weeks.

19
More than one parasitoid may emerge from a single host (superparasitism), and the number of
emerging progeny is correlated with host size. In order to investigate the relationship between the
number of emerging parasitoid progeny and the size of the female T. aurantiaca host in field conditions
(Kuching, Sarawak), a random sample of 50, parasitized female T. aurantiaca were measured (greatest
horizontal diameter of test) and the emerging adult parasitoids were counted from each host. There
was a significant positive correlation between host size and parasitoid progeny produced (R2 = 0.47, p <
0.001) (Fig. 11). This relationship is probably best explained by the simple fact that larger hosts provide
more food resources for more parasitoid larvae to complete development. It is not clear whether this
superparasitism is self‐superparasitism (a single female parasitoid ovipositing multiple eggs in a single
host) or if it is superparasitism in terms of intraspecific competition (different individual parasitoids
ovipositing in the same single host). This positive relationship between host size and production of T.
somervillei progeny will be important in captive rearing, release site selection, and ensuring
establishment during inoculative releases.
The longevity of adult parasitoids and their fecundity can vary greatly depending on diet and the source
of nutrients such as sugar from honeydew or nectar (Lee et al. 2004). It is possible that T. somervillei
feeds on honeydew secreted by its host. Further, some parasitoids stab their hosts with their ovipositor
but do not lay eggs, instead feeding on the hemolymph that leaks out. Such "host feeding" can also be
very important in longevity and fecundity (Wheeler 1996). Host feeding, although not yet observed in
the field, would be advantageous because it can result in the death of the host without it being
parasitized.
Natural enemies of soft scales on Christmas Island. Parasitization of soft scale insects (Coccidae) on
Christmas Island was very rare during the searches between 2010 and 2012. Signs of parasitoid
presence were initially found only in the settled area in the northeast of the Island. The reason for the
apparently limited distribution of soft scale parasitoids on Christmas Island is not clear. The 2010 dry
season was unusually wet and scale insect populations were very low in general throughout the island.
It is possible that parasitoid populations went locally extinct in areas outside of the settled areas and the
recolonization of soft scale aggregates simply did not occur at detectable levels in 2011 and 2012.
Alternatively, despite their presence in the settled areas, these parasitoids are naturally absent from
large parts of the rainforest on Christmas Island due to dispersal limitation.
Parasitization was observed in Coccus sp. and Pulvinaria urbicola, a newly recorded invasive soft scale
on the island (Neumann et al. 2011). Two parasitoids, Coccophagus ceroplastae (Aphelinidae) and Encyrtus infelix (Encyrtidae) were recovered from Coccus sp., both first recorded in the settled areas.
Additionally, two Metaphycus spp. (Encyrtidae) and Coccophagus nr. bivittatus (tentatively
identification) were recovered from immature soft scales (most likely P. urbicola) near the 'Boulder'
(South Point) on the host plant Pisonia grandis.
Coccophagus ceroplastae and E. infelix are both known to attack a variety of soft scale species, including
three that are important in YCA supercolonies (Table 4: Coccus hesperidium, Saissetia oleae, S. coffeae).
It is possible that these two parasitoid species could be used to control all, or almost all, soft scales at
acceptable levels on Christmas Island. For example, C. ceroplastae can control the damaging scale
insect P. urbicola under laboratory conditions. Potted Pisonia umbellifera plants were infested with P.
urbicola and the scale insects were allowed to reproduce. When the scale insects of the first generation
were estimated to be in the second instar of their development, 60 field‐collected C. coccophagus were
liberated on the plants in two batches one week apart. Parasitism was observed approximately three
weeks later. Parasitization rates were low and not quantified, but the suitability of the host was

20
demonstrated. Approximately 11 weeks after the first parasitoid introduction, two of the host plants
had no remaining live scale insects. The other two plants died before the eradication of the scale insect
infestation but parasitism rates were high (G. Neumann, unpublished data). The potential of C. nr.
bivittatus and the two Metaphycus species is not clear. Study of the potential of these parasitoids will
commence in Phase 2 of the biological control project as well as the mass rearing, redistribution, and
impact survey of the identified soft scale parasitoids, C. ceroplastae and E. infelix.
Key Research Outcomes: Few natural enemies of the key honeydew‐producing scale insect Tachardina
aurantiaca occur on Christmas Island, and they do not regulate its population size. No parasitoids of
female T. aurantiaca were found. Conversely, within its native range in Malaysia, T. aurantiaca is rare
and patchily distributed, associated with diverse natural enemies, including at least five species of
primary parasitoids, and it suffers high parasitization rates – all attributes consistent with population
regulation by natural enemies. Importantly, high parasitism rates in Malaysia occurred in the presence
of honeydew‐collecting ants, including the yellow crazy ant. Of the primary parasitoids known to attack
yellow lac scale from our studies in Malaysia, Tachardiaephagus somervillei (Encyrtidae) is the most
promising agent for introduction and release on Christmas Island. All Tachardiaephagus species have a
narrow host range and appear to be family specialists, known only to attack the Kerriidae, the family to
which the yellow lac scale belongs (Table 3). Our initial studies indicate that (a) T. somervillei attacks T.
aurantiaca across 1900‐km range in Peninsular Malaysia and Malaysian Borneo, (b) is the most
abundant natural enemy of T. aurantiaca, (c) has a short life cycle compared to its host, (d) exhibits
superparasitism (i.e., where multiple progeny emergence from a single host individual); (e) causes high
rates of parasitism on T. aurantiaca in the presence of tending ants, including the yellow crazy ant, and
(f) can be reared under laboratory conditions. Coccophagus ceroplastae and Encyrtus infelix, both
parasitoids of a wide variety of coccid scale insects, are already present on Christmas Island. The three
parasitoids T. somervillei, C. ceroplastae and E. infelix could be deployed in combination against the
entire assemblage of honeydew‐producing scale insects in YCA supercolonies.

21
Project 2b. Scale insect survey on Christmas Island (biology, host range,
habitats, natural history, identification)
Comprehensive knowledge of the scale insect species present in rainforest and the built
environment on Christmas Island. This information will indicate habitats and host plants of T.
aurantiaca and other scale insects on Christmas Island. It will provide information on nativeness
of the scale insect fauna which will aid determination of a host specificity test list, testing
protocols, and other regulatory matters for biological control agents.
Research Findings 2009‐2013
Extensive searches for scale insects were carried out over three years 2010‐2013, using five methods.
First, surveys were conducted using Timed Searches, in which 30 min was spent looking within a 50‐m
radius at each of 151 waypoints from the CINP island‐wide survey (Fig. 12). Searches were replicated
three times in each dry season in 2010‐2012. Second, searches were conducted along the entire length
of five tracks, a total of 21 km, twice in each of the dry seasons of 2011 and 2012 (Northwest Point
Track, 4.7 km; Boulder Track, 6 km; Blowholes Road, 3.4 km; Martin Point Lookout to CINP boundary,
1.9 km; and, Dolly Beach Boardwalk, 1.8 km). Search effort totalled ~405 hours that did not include
travel between sites. Third, endemic plant species were a focal point of further search assuming that
endemic scale insects would be more likely to be associated with them (Neumann et al. 2007). Fifteen
of the 18 species of endemic plant species were located and examined between 2010‐2012. Ten rare
and endemic plants species from the CINP database were searched for scale insects at 125 locations. For
each common endemic plant species, 30 haphazardly selected individuals were examined each year
between 2010‐2012. Fourth, opportunistic searches were made of exotic plants at many locations
around the settled areas in the ‘Dog’s Head’ area of the island. Fifth, CINP personnel also aided the
search by being aware of scale insects during the island‐wide survey in 2011.
Twenty‐one species of scale insects in six families have been found on Christmas Island (Table 4). It is
highly probable that these species, all with broad host plant ranges and geographic distributions, were
introduced following human settlement on Christmas Island and are non‐native. No endemic scale insects
were found.
Key Research Outcomes: No native or endemic scale insect species have been discovered in intensive
and extensive searches for scale insect species on Christmas Island. However, 400 hours of search over
two years did yield five additional exotic scale insect species previously unknown to the island.
Assuming that any prospective biological control agent for Tachardina aurantiaca on Christmas Island
would be a narrow family specialist (Kerriidae) and all known scale insect species are non‐native and
invasive, the probability of any direct non‐target effects is negligible.

22
Project 2d. Genetic and morphological matching between Tachardina
aurantiaca and Paratachardina pseudolobata on Christmas Island and
Southeast Asia.
Determination of species identity and likely area of origin of T. aurantiaca and P. pseudolobata.
This is critical for further targeted search for biocontrol candidates. Specimens from Christmas
Island and Southeast Asia (Indonesia, Thailand, West Malaysia, and East Malaysia) will be
compared genetically and morphologically.
Research Findings 2009‐2013
T. aurantiaca collected from 5 different sites and host plant species across Christmas Island are
morphologically and genetically identical, based on cuticular morphology and mitochondrial cytochrome
c oxidase subunit 1 (COI) and 28S ribosomal RNA sequences (P. Gullan and L. Cook, personal
communications, 2012). Initial morphological and genetic studies of T. aurantiaca collected from sites in
Sarawak indicate that they are morphologically and genetically identical to T. aurantiaca on Christmas
Island (P. Gullan, personal communication, 2012).
Further study of Paratachardina pseudolobata was not pursued because, as pointed out above, it is not
a source of useable honeydew for YCA on Christmas Island.
Key Research Outcome: The single population of T. aurantiaca on Christmas Island matches
morphologically and genetically those populations examined so far within its native region in Southeast
Asia. This establishes the identity of Tachardina aurantiaca on Christmas Island, making the successful
establishment on Christmas Island of a biological control agent that attacks T. aurantiaca in its native
range more likely. Further collections and analyses of T. aurantiaca in Eastern Malaysia (Sabah) and
exploration in southern Thailand should establish if any morphological or genetic variation occurs across
its native distribution in Southeast Asia.

23
Project 2e. Scale population dynamics, especially the ecology of lac scale,
Tachardina aurantiaca
Pre‐release monitoring of T. aurantiaca and P. pseudolobata prior to any introduction and
release of natural enemies so that the efficacy of natural enemies in controlling scale insect
numbers and reducing yellow crazy ant densities can be determined.
Research Findings 2009‐2013
The life cycle of Tachardina aurantiaca is relatively long, with one complete generation from F1 crawler
settlement, through female maturation, to F2 crawler production and settlement completed between
88 ‐ 100 days, depending on host plant species (Table 5). Males emerge about 6 weeks after crawler
settlement, consistent across host plant species (Table 5). On Christmas Island, there are overlapping
generations such that all stages, including crawlers, female nymphs, and mature females can all be
found together on twigs and along the midribs of leaves. This suggests that suitable life stages of T.
aurantiaca susceptible to attack by Tachardiaephagus somervillei are available most of each year.
Female size and offspring production vary considerably and is significantly correlated at the time of
crawler release both in field and laboratory populations (R2 = 0.295, p < 0.001 in the laboratory and R2 =
0.207, p = 0.006 in the field) (Fig. 13). Large females can produce over 600 crawlers, indicating that
conditions that affect female growth before crawler production could have a very strong effect on
population growth. Conditions affecting grown are not well understood but could relate to settlement
density of crawlers, position of crawler settlement on the host, host plant identity, and time of year.
The role of males in reproduction is not clear, because it appears that some females are capable of
producing offspring parthenogenetically (Ong and Neumann, unpublished data). The approximately 6‐
week development time of males suggests that females are mature at that time. Females apparently
continue to live and grow well beyond the completion of development and offspring production occurs
long after the female becomes mature (mating might not occur at all). Further experiments are under
way to clarify the role of males in reproduction and the effect (if any) of parthenogenetic reproduction
on life span and fecundity of T. aurantiaca.
Key Research Outcomes: The life cycle of Tachardina aurantiaca is relatively long for a scale insect but
several overlapping generations occur in a single year. Female size is significantly correlated with
fecundity, indicating that conditions that affect female size are likely to have a strong influence on
population growth rates and density. The role of males in reproduction is not yet clear although it
appears that some females can reproduce parthenogenetically. All of these attributes will influence
host‐parasitoid dynamics on Christmas Island.

24
Research Project 3. Strategic planning for biocontrol introduction
Australia is well advanced in the regulation of import and release of exotic invertebrate
biological control agents. Legislation stipulates import conditions, risk assessment, host‐
specificity test lists, release protocols, and pre‐release studies prior to approval of biological
control introductions. Consultation with regulators early in the project is essential in the
planning, conduct, and evaluation of research. Furthermore, early identification and agreement
with co‐operators is essential for completion of research components key to proof of concept of
indirect biological control of the YCA. Consultation outlined below will put project planning
within the regulatory frameworks for biological control introductions in Australia and determine
key co‐operators necessary to advance the project.
Project 3a. Planning research in step with legislative and regulatory
requirements for biological control introductions
Smooth preparation and transparency in planning for introduction of biological control agents.
Coordination of research findings in step with regulations (i.e. DAFF, DSEWPaC). Written report
anticipating the regulatory steps and hurdles for advancement of the project.
Research Findings 2009‐2013
The importation and release of an exotic biological control agent (BCA) on Christmas Island, an external
territory of Australia, is regulated under Australian Government legislation by two independent, but
parallel frameworks. The first framework falls under the Quarantine Act 1908, and subordinate
legislation in the Quarantine (Christmas Island) Proclamation 2004. The process to seek approval to
import and release an exotic biological control agent on Christmas Island is outlined by Biosecurity DAFF
(Department of Agriculture, Fisheries and Forestry) in a document entitled Revised Biosecurity
Guidelines for the introduction of exotic biological control agents for the control of weeds and plant
pests
(http://www.daff.gov.au/ba/reviews/biological_control_agents/protocol_for_biological_control_agents
/guidelines‐introduction‐exotic‐bcas‐weed‐and‐plants
Second, because the proposed action could have a significant impact on the environment, an
Environmental Referral is required under legislation in the EPBC Act (1999). This framework is outlined
by the Department of Sustainability, Environment, Water, Populations and Communities at
http://www.environment.gov.au/epbc/assessments/index.html
The criteria for evaluating importation and release of a BCA under the two frameworks differ
somewhat. Under the Biosecurity DAFF framework, import risk analysis for a BCA differs from a
standard commodity import analysis that assesses the probability of entry, establishment and spread.
Because BCAs intended for release are deliberately introduced, risk analysis focuses on off‐target effects
alone. Under the EPBC Act, DSEWPaC also has a linked approval process for the import and release of
BCAs which uses the final risk analysis report produced by Biosecurity DAFF. This may be used by the
DSEWPaC minister to include the BCA on the live import list.
Under separate legislation under the EPBC Act, a proposed action is referred if it is likely to have a
significant impact on a matter of national environmental significance (e.g. wetlands of international

25
significance, threatened species and ecological communities, migratory species). The process for
submission and evaluation of an Environmental Referral is given at
http://www.environment.gov.au/epbc/assessments/pubs/flow‐chart.pdf
Harmonization of these two frameworks is critical as we move through these regulatory processes. Key
issues and Australian government contacts for progressing issues in each framework are given in Table
6. In discussions with Biosecurity DAFF, it was clear that they were unfamiliar with the Environmental
Referral process (EACD) in DSEWPaC. It was equally clear that the Environmental Assessment and
Compliance Division in DSEWPaC had not previously encountered Environmental Referrals that
addressed importation and release of BCAs. A search of the 100 most recent environmental referrals
indicated that 42% related to mineral or energy projects, 31% to infrastructure development, 20% to
residential development, 5% to biodiversity management, and 2% to other. None of the Environmental
Referrals since July 2000 on the referrals list page
(http://www.environment.gov.au/cgi‐
bin/epbc/epbc_ap.pl?name=current_referrals&limit=999999&text_search=)
addressed importation and release of a BCA. However, Biosecurity DAFF and the EACD DSEWPaC are
now exchanging information on the CI project.
Key Research Outcomes: Two regulatory frameworks govern the importation and release of an exotic
biological control agent on Christmas Island. To gain approval to import and release a biological control
agent on Christmas Island, we will need to prepare and submit two sets of documents: an Approvals
Package under the DAFF protocol and an Environmental Referral to EACD, DSEWPaC. The key
government contacts involved in the two frameworks are described and currently consulting each
other.

26
Project 3b. Identification and agreement with co‐operators
Identification of key co‐operators and agreements to advance the project. Cooperation will be
necessary between Australian universities (e.g., project coordination, research staff mentoring
and supervision); Australian federal agencies (e.g., assistance with regulatory issues in
biocontrol introductions; taxonomic expertise on scale insects, ants, and parasitoids; Overseas
universities (e.g., morphological and genetic studies of scale insects); Overseas government
agencies (e.g., linked biocontrol program with foreign exploration and testing of natural
enemies, biocontrol regulatory expertise ‐ USDA‐ARS, Invasive Plant Research Laboratory, FL;
offshore facilities for safe host specificity testing – e.g., Malaysian Department of Agriculture)
Research Findings 2009‐2013
We have established relationships with many researchers in Australia, India, Malaysia, Singapore and
the United States, all of whom bring different expertise and capacity to this project (Table 7). Prof.
Gullan and Dr. Cook provide taxonomic expertise on scale insects as well as morphological and genetic
analyses for Tachardina aurantiaca on Christmas Island and in Southeast Asia. Dr. Hayat provides
taxonomic expertise in the identification of encyrtid and aphelinid parasitoids associated with T.
aurantiaca. A cooperative research agreement with Forestry Research Institute Malaysia provides
benchspace and facilities for rearing T. aurantiaca and its natural enemies in Kepong, Selangor,
Peninsular Malaysia. Ms Ong, a Masters student under supervision of Dr. Neumann, is conducting her
research on the biology of T. aurantiaca and developing rearing techniques for it and one of its
important natural enemies, Tachardiaephagus somervillei. Sarawak Forestry and the National
University of Singapore provide laboratory space and field assistance. Dr. Pemberton, who has worked
on the biological control of the invasive lobate lac scale in Florida, provides advice and was instrumental
in establishing some of the cooperators listed in Table 7.
The complex permitting system in Malaysia has been negotiated successfully and permits to conduct
research and collect specimens in West Malaysia, Sarawak, and Sabah have been granted.
An ad hoc advisory group of expert biocontrol practitioners has been assembled to review and advise on
development of a host test list and a host‐specificity testing protocol (Table 8). Expertise in this group
covers key aspects of in the development of biocontrol relevant to Christmas Island (e.g safety and risk
assessment in biological control, host specificity of parasitoid natural enemies, biocontrol in natural
areas, foreign exploration, scale insects as targets in biological control).
Key Research Outcomes: Collaborations have been established with researchers with expertise in the
systematics, biology, and molecular genetics of scale insects, taxonomy of microhymenopteran
parasitoids, and biological control of scale insects. Cooperative agreements have been established with
FRIM and Sarawak Forestry that provide benchspace and technical assistance. Dr Neumann is serving as
an advisor of a Masters student (Universiti Sain, Penang) who is conducting research on this project at
FRIM. Research and collecting permits have been obtained from the States of Sarawak and Sabah, as
well as a national collecting permit.

27
Project 3c. Safety and sanitary protocols anticipating a pre‐release
environmental assessment of introduction
An agreed list and protocol for host specificity testing of candidate bio‐control agents and
protocols to eliminate and prevent the accidental introduction of hyperparasitoids to the island.
Research Findings 2009‐2013
To focus host‐specificity testing for Tachardiaephagus somervillei (Hymenoptera: Encyrtidae), a
candidate biological control agent for the invasive introduced yellow lac scale (Tachardina aurantiaca)
(Hemiptera: Kerriidae), we used four approaches to assess the risk of importation and release of the
candidate biological control agent T. somervillei to Christmas Island. Risk was defined as off‐target
impacts, including those that could affect endemic species or threatened and endangered species.
These risk evaluations were based on host records of Tachardiaephagus from the scientific literature,
the diversity and attributes of other scale insects (superfamily Coccoidea) on the island, the occurrence
of endemic hemipterans known from Christmas Island, and the host ranges of encyrtid parasitoids
known to attack scale insects. All lines of evidence indicate that there is a very low likelihood that
importation and release of T. somervillei would harm species of concern (i.e. those that occur on
Christmas Island).
We then describe a protocol for host‐specificity testing recommending that it be conducted within the
area of origin (Malaysia) of Tachardina aurantiaca. Identification of an agreed host test list and protocol
for host range testing of a candidate biological control agent are necessary requisites under the
Biosecurity DAFF guidelines for the approval of importation of exotic biological control agents for the
control of weeds and plant pests (see Project 3a). Lastly, in their native range, many primary parasitoids
have their own insect natural enemies, including those known as hyperparasitoids (i.e. parasites of
parasites). This is so for T. somervillei (Project 2a, Table 1). We describe standard rearing and sanitary
techniques to ensure that the founding population from Malaysia is free of hyperparasitoids before
importation to Christmas Island
Known host range of Tachardiaephagus somervillei. We evaluated known host species records for all
species of Tachardiaephagus using the Universal Chalcidoid Database (Noyes 2012) so that host range
could be estimated and risk of release of T. somervillei assessed for species of concern on Christmas
Island (see Sands and Van Driesche 2004). The Universal Chalcidoidea Databases (Noyes 2012) is the
most comprehensive database for chalcidoid parasitoids, with over 120,000 host/associate records
(including associations with food plants of the hosts) and > 140,000 distribution records of the
parasitoids in the superfamily Chalcidoidea. It is very well developed, regularly updated and extremely
well referenced. Nevertheless, large databases can contain errors affecting reliability, such as
erroneous published host records and outdated parasitoid taxonomy (Kuhlmann et al. 2006). The most
important source of error in this database is actual published erroneous records. Since all records are
referenced, doubtful records can be investigated and if necessary, filtered out.
Seven described species of Tachardiaephagus are distributed in Southeast and South Asia, and sub‐
Saharan Africa (Noyes 2012). Host records of Tachardiaephagus are restricted to species within three
genera (Kerria, Tachardina, and Paratachardina) of lac insects (Kerriidae) (Project 2a, Table 3).
Furthermore, all host species records for Tachardiaephagus somervillei are within two genera: Kerria,
comprising the lac insects of commerce, and Tachardina. These records suggest that Tachardiaephagus
somervillei is highly likely to have a narrow host range restricted to kerriid host species.

Status o
that cou
(Superfa
were co
search o
A total o
2b, Tabl
are non‐
were dis
Phyloge
Island sh
somervil
2006) an
Christma
aurantia
three pl
Aucheno
Thus, an
a host ra
Urban 2
host tax
Host ran
endemic
family to
for the b
(Noyes a
in 2 subf
Tetracne
associat
endopar
hatching
mostly p
are egg
We used
the hem
primary
of other scale
uld conceivab
amily Coccoid
mpiled from
over two yea
of 24 species
e 4). All scal
‐natives on C
scovered.
eny of endem
hould be con
llei. We obta
nd narrowed
as Island in t
aca belongs.
anthoppers
orrhyncha or
ny potential f
ange that bri
012), as wel
a in the Kerr
nges of ency
c species on
o which T. so
biological con
and Hayat 19
families. The
eminae inclu
ed with scale
rasitoids mea
g larva comp
parasitize im
predators (N
d the Univers
mipteran fam
hosts of enc
e insects on
bly constitut
dea, the sam
m the literatu
rs for endem
s of scale inse
le insects tha
Christmas Isl
mic hemipter
nsidered to e
ained a list o
d considerati
the Order He
These spec
(Table 9). A
r Heteropter
for non‐targe
idges this ve
l as distinctiv
riidae.
yrtid parasito
Christmas Is
omervillei be
ntrol of harm
994, Noyes 2
e subfamily E
udes 107 gen
e insects (He
aning that th
pletes develo
mature life s
Noyes 2012).
sal Chalcidoi
ilies represe
cyrtid specie
Christmas Is
e non‐target
me superfam
re and then
mic scale inse
ects in six fam
at could be c
and and non
rans. The clo
evaluate the
of known end
on of taxa o
emiptera, the
ies comprise
ll of these en
ra) than the y
et impacts b
ry substantia
ve morpholo
oids known t
sland. The E
longs, is one
mful insects,
2012). The E
Encyrtinae in
nera and 815
emiptera: Co
he parasitoid
opment feed
stages (or, ra
idea Databas
ented by end
s associated
sland. We d
t species. Re
ily to which
supplement
ects (see Pro
milies have b
considered a
ne are known
osest endem
risk of becom
demic insect
n this list to
e same orde
e one true bu
ndemic speci
yellow lac sc
y T. somervi
al phylogene
ogies, life‐his
to attack me
Encyrtidae (H
e of the most
including a v
ncyrtidae cu
ncludes 353
5 species. Ap
occoidea)(No
d egg is laid d
ing on the ho
arely, adults)
se to analyze
emic species
with these t
etermined in
ecords for oc
Tachardina a
ted by condu
oject 2b).
been recorde
s the most li
n to be bene
mic relatives o
ming non‐tar
t species on C
the closest e
r to which th
ug, a cicada,
ies occur in d
cale (suborde
llei on these
etic distance
stories and e
embers of th
Hymenoptera
t important p
variety of sca
urrently com
genera and 2
proximately
oyes 2012).
directly insid
ost’s tissue,
), but some s
e all records
s on Christm
taxa were de
nsect species
ccurrence of
aurantiaca b
ucting >400 h
ed on Christm
kely potenti
ficial. No en
of the yellow
rget hosts of
Christmas Isl
endemic rela
he known tar
a leafhoppe
different sub
er Sternorrhy
endemic He
(see above a
cological att
he hemiptera
a, Superfami
parasitic was
ale insects in
prises 460 ge
2920 species
half of all en
Encyrtids are
e the host’s
ultimately ki
species in on
of encyrtid p
as Island. N
etermined. C
s on Christm
scale insect
belongs) on t
hours of stru
mas Island (s
al non‐targe
ndemic scale
w lac scale on
f Tachardiaep
land (James
atives of lac s
rget species
r, a spittlebu
borders (eith
yncha).
emiptera wo
and Fig. 2 in
ributes to its
an families w
ily: Chalcidoi
sp (parasitoid
nfesting woo
enera and 37
s, while the
ncyrtid speci
e generally
body where
illing the hos
ne genus (Mi
parasitoid as
ext, all recor
Consequentl
28
as Island
species
he island
ctured
see Project
et organisms
insects
n Christmas
phagus
and Milly
scales on
Tachardina
ug, and
er suborder
uld require
Cryan and
s known
with
idea), the
d) families
dy plants
735 species
es are
the
st. Encyrtids
icroterys)
sociates of
rded
y, any
8
s

29
encyrtid species that share (as hosts) both coccoid species (scale insects) and any family represented by
endemic species on Christmas Island could be found.
There are no known records in the Encyrtidae of any parasitoid species that attacks both scale insects
(Coccoidea) and host species in hemipteran families that have endemic representatives on Christmas
Island (Table 9). All known encyrtid primary parasitoids known to attack species in families which have
endemic representatives on Christmas Island have primary hosts only within suborder
Auchenorrhyncha. While species in these hemipteran families are attacked by many species of
chalcidoid parasitoids, many fewer or no encyrtid species are known to use them as hosts, and not one
encyrtid species is reported with a host range so extremely broad that it encompasses both scale insects
and any of these hemipteran families.
Host specificity testing protocol. Actual records for host range of Tachardiaephagus and known
patterns of parasitism in the Encyrtidae indicate that it is highly improbable that T. somervillei would
exhibit an extremely broad host range (one that would be unprecedented in any single species of
Encyrtidae known to attack scale insects) and utilize any of the endemic hemipteran species on
Christmas Island. Thus, when the host specificity testing of the biological control agent is initiated using
the centrifugal phylogenetic approach, the outer boundaries of the sequential testing should be set
initially at a significantly closer distance from the target organism than the taxa represented on
Christmas Island by endemics.
A ‘test species’ is an insect species that is tested as a potential non‐target insect. Since no non‐target
scale insects were identified on Christmas Island, the list of test species will be determined using the
centrifugal approach (Kuhlmann et al. 2006, Neumann et al. 2010). In this approach, species other than
the known target host of the biological control agent are tested with the most closely related species
(least phylogenetic distance, and with close similarities in biology and ecology) tested first then less
similar species thereafter (“centrifugal principle” ‐ Wapshere 1974).
A test species will be considered a ‘suitable host’ if parasitoid emergence is observed during any of the
exposure tests. To be a suitable host, the parasitoid must (a) accept the test species when exposed, (b)
must oviposit, (c) the eggs must hatch into larvae, (d) the larvae must complete development using the
test species as the resource, (e) the larvae must pupate and, finally, (f) the emerging adult parasitoid
must be able to exit (emerge from) the test species. If parasitoid emergence from a test species is
observed, the test species will be considered a suitable host without assessing the viability, fecundity,
sex ratio and other characteristics of the emerging parasitoid generation.
A test species will be considered ‘susceptible’ if the parasitoid causes significant mortality due to
probing, host feeding (when the parasitoid stabs the insect with its ovipositor and then feeds on the
insect’s hemolymph), oviposition, or oviposition and larval development. Note that even if some
mortality is observed due to the above, the test species will only be considered susceptible if the
mortality resulting to the exposure to the parasitoid is significantly higher than in control groups
(negative controls) of the test species where the test individuals are not exposed to the parasitoid.
The ‘parasitoid’ in this case is T. somervillei (Hymenoptera: Encyrtidae), a widespread parasitoid native
to Southeast Asia. T. somervillei is an important parasitoid of Tachardina aurantiaca (Project 2a, Table
1; Project 2c, Table 2). It is known to parasitize some scale insects in the family Kerriidae (lac scales)
including the ‘target host’, Tachardina aurantiaca (Table 3).
Selecting test species. Host specificity testing can be a long and difficult process, depending on how
problematic it is to establish and manage colonies of the parasitoid, the target host, the host plant of

30
the target host, and the test species. Furthermore, resources and time are not unlimited and permitting
issues to collect insects and run field experiments (when needed) in Malaysia can add to the time
required to complete host testing. Therefore, a manageable number of species should be used (10 to
15, but fewer are acceptable if high specificity is found initially). The species used will be largely
determined by availability, so determining the actual test species at species or genus level is not
possible at this point. We can, however, predict to some degree the families of the test species that will
be selected. We will focus on neococcid taxa including the Kerriidae, the family to which the target host
belongs (see Fig. 14, also Gullan and Cook 2007, Ross et al. 2012). Considering the phylogenetic
relationships of scale insect families (Fig. 14), we will aim to test one species in the Kerriidae,
Paratachardina pseudolobata, which is also invasive on Christmas Island (see Project 2b, Table 4), and
more than one species from the Coccidae. Species in the Diaspididae will also be considered as a less
closely related group of scale insects. Early in the host testing process, an ‘out‐group’ (test species
phylogenetically more distant) will also be used, most likely selected from the Pseudococcidae (Fig. 14).
As mentioned above, availability will determine species selection to some degree but in some of the
groups (e.g. Coccidae) there may be a number of species available. Species that can be reared in
laboratory conditions would be preferred but carrying out tests in field conditions should also be
acceptable as long as the tests can be well controlled.
No‐choice tests. In no‐choice tests, only the non‐target test species will be provided to the parasitoid in
the experimental replicates. This is the most stringent test method that places the parasitoid under
high “oviposition pressure”.
For each test species, trials will be replicated 10 times. Each replicate will consist of 1) one
experimental cage with the test species (50 individuals in three different age groups/instars; 150
individuals in total) exposed to 10 female and 10 male parasitoids, 2) one positive control cage
consisting of 50 mature female T. aurantiaca exposed to 5 female and 5 male parasitoids in order to
confirm that the parasitoids used in the experiment are of good quality and capable of parasitism in the
given exposure time, and 3) one negative control cage which will be similar to the experimental cage
except the test species will not be exposed to parasitoids. The negative controls will be used to
determine any effect of parasitoid exposure on insect mortality other than successful parasitization
such as probing, host feeding and oviposition without successful parasitoid development.
The cages will consist of fine mesh bags (sleeves) fixed on the branches of plants that serve as host
plants for the scale insects. Parasitoids will be collected from a lab colony and introduced to the cages <
24 h after emergence. Exposure time to parasitoids will be determined during preliminary studies to
determine the optimal time frame for mating, egg maturation and oviposition by the parasitoid.
Evaluation:
1) If the positive control does not yield any parasitoid emergence, the replicate will not be evaluated
(failed replicate).
2) If the positive control yields parasitoid emergence the replicate will be evaluated. The parasitization
rate in the positive will be noted but will not be compared to parasitization rates in the test replicate.
3) Parasitoid emergence in the test replicate will be noted along with the approximate age and
developmental stage of the individuals yielding parasitoid emergence. These individuals will not be
used to compare mortality rates due to reasons other than successful parasitism.
4) Mortality of test species in test replicates and negative controls will be noted.

31
Analysis:
1) If any test replicates yielded any number of parasitoid emergences, the test species will be
considered a suitable host. The level of suitability of the test species will not be compared to the
suitability of the target host at this stage of the project.
2) Mortality (except mortality due to successful parasitism) rates of test insects and insects in the
negative control will be compared using 2‐sample t‐tests. If the mortality rate is significantly higher
(α = 0.05), the test species will be considered susceptible and the reasons for mortality will be
investigated.
If a test species is susceptible (but not suitable), behavioural observations will be made to determine
the cause of the effect of parasitoid exposure. The observational arena will be a ‘window box’ arena.
This arena is constructed by constructing a 10 cm W x 10 cm L x 5 cm H box with one side being glass.
The box is constructed so that it can be placed on the stem of a potted host plant on which the test
insects feed. Female parasitoids are then liberated inside the box and the box closed. The potted plant
then can be placed flat on its side and the box positioned under a dissecting microscope. Video footage
is then captured using a high definition video camera with a microscope adaptor. The footage (4‐6 h per
observation) can be analyzed later for the cause(s) of mortality (i.e. probing, host feeding, or oviposition
without full parasitoid development). If oviposition is suspected, dissections will also be made to
investigate parasitoid larval development.
Other testing methods. We considered using choice tests (where the parasitoid is presented with the
test species and the target host simultaneously) and sequential no‐choice tests (where the parasitoid is
presented with the target host first then it is transferred on to the test species). However, in our case
these tests may have limited value for reasons given below.
A choice test would show whether a test species that is a suitable host in no‐choice tests would be
consistently attacked when the target host is also available. A physiologically suitable host may not be
preferred over the target host and the assumption could be made that parasitization would not occur
(or at very low level) in field conditions. This test would be used in cases where the suitable host (found
suitable in a no‐choice test) is of concern, e.g. a beneficial or endemic non‐target species that coexists
with the target host. However, this situation does not arise on Christmas Island where all known scale
insects are non‐native and none of them are beneficial. This test is further limited by the assumption
that the test species and the target host occupy the same space hence providing the parasitoid with a
choice. This would also not be the case on Christmas Island.
A sequential no‐choice test would further show the unsuitability of a test species. The parasitoid may
be ‘motivated’ to attack the test species even if it did not attack it in a simple no‐choice test if the
parasitoid is first allowed to initiate oviposition behaviour and to gain experience. This test may be even
more stringent than the simple no‐choice test and one would consider using it in the case of a test
species that is of great concern to further show the unsuitability of the test species. This would again
require a beneficial or endemic non‐target species neither of which exists on Christmas Island.
Furthermore, similar to the choice test, this would assume either the coexistence or very close
proximity of the test species and the target host.
Options for the location for host‐specificity testing. There are three options for the location where
host specificity testing could be conducted: 1) in quarantine containment at the release location
(Christmas Island); 2) in quarantine containment on mainland Australia; and 3) in the native
geographical range of the biological control agent where no containment would be necessary.

32
Option 1. Testing in containment at the release location (Christmas Island). This option is the least
attractive and involves increased risks and expenses. There is no quarantine containment facility on
Christmas Island, a remote oceanic island. The expense of constructing such a facility to Quarantine
Approved Premises Criteria (Quarantine Insectary Level 2) for host‐specificity testing of a single agent
would be prohibitive. Even if there were such a facility, in case of escape, the biological control agent
would find an environment very similar to its native range with its natural host T. aurantiaca in
abundance. The benefits of this option would include easy access to the natural host for the parasitoid
colony and no further travel if and when a release permit is obtained.
Option 2. Testing in containment on mainland Australia. While this option is more attractive than
Option 1 above, it is still somewhat risky, and labour and cost intensive. It brings few benefits. The
biological control agent (parasitoid) would have to be brought into containment along with its natural
host (Tachardina aurantiaca). This would pose the risk of not only a parasitoid escaping but also a
potentially invasive, host‐generalist scale insect (on Christmas Island, for example, at least 15
horticultural species are attacked by T. aurantiaca, including three species of Citrus, Macadamia, Guava,
Pomegranate, Chili, Eggplant, Star fruit, and Soursop)(R.W. Pemberton and D.J. O’Dowd, unpublished
results). Using a containment facility in the temperate region of Australia could decrease these risks. In
case of quality control issues of the biological control agent due to ‘lab selection,’ additional travel,
collection, and importation of the biological control agent would be necessary, further increasing costs.
Natural host colony loss (which can easily happen due to overexposure to the parasitoid agent, fungal
infections, etc.) would necessitate multiple importations of the natural host, which would once again
significantly increase costs and risk.
Option 3. Testing in the native geographical range of the biological control agent with no containment
necessary. In our case, this option is the most attractive, most cost‐effective, and least risky. There are
many benefits to study biological control agents and carry out host specificity studies in the native
geographical range of the agent. In our case, both the parasitoid and its natural host is readily available
from the wild in Malaysia which ensures good quality parasitoid and host cultures. The risk factor
would be zero as there would be no importation of any organisms. Although obviously desirable, such
studies in the native range are not common, in part because of the lack of local research facilities, lack
of skilled and reliable cooperators, or sufficient time in what are sometimes difficult locations (Van
Driesche et al. 2008). However, scientific work is cost‐effective in Malaysia compared to Options 1 and 2
and our network of collaborators and cooperators is well established (Project 3b, Table 7). Options 1
(test in containment on Christmas Island) and 2 (test in containment on the Australian mainland) offer
no benefits over this option. We therefore suggest that host specificity testing is carried out in the
native range of the biological control agent. This option carries the least risk and is consistent with
Recommendation 2 in a review of biosecurity risks in biological control commissioned by the Australian
Government (Ferrar et al. 2004) that biological control practitioners undertake host specificity testing in
the native region before any importation.
Protocol for insuring that hyperparasitoids associated with T. somervillei are not imported to
Christmas Island. The unintended importation of a hyperparasite of T. somervillei is inimical to the
successful suppression of YCA supercolonies, because it could compromise the capacity of
Tachardiaephagus to build up population densities sufficient to control Tachardina. For this reason,
great care will be taken to implement standard agent rearing and sanitary techniques to ensure that the
founding population of Tachardiaephagus from Malaysia is free of hyperparasitoids and pathogens.

33
Free‐living adults are the safest to import because this would ensure that hyperparasitoids would not be
co‐introduced.
Tachardiaephagus somervillei is frequently attacked by Promuscidea unfasciativentris (Aphelinidae) (see
Project 2a; Table 1). This hyperparasitoid was abundant near Kuching, Sarawak and present in Selangor,
West Malaysia, although rare. The impact of hyperparasitism is not known; nevertheless, the exclusion
of any hyperparasitoids from captive colonies in Malaysia is critical before adult T. somervillei are
imported to Christmas Island as the founding population.
The exclusion of hyperparasitoids from captive populations at locations where the hyperparasitoids are
native can be difficult in the long run unless care is taken. Basic containment is usually sufficient but
standard practices to keep hyperparasitoids out of captive populations must be followed. These include:
Field collected, parasitized scale insects will never be exposed to laboratory scale insects directly;
twigs with parasitized scale insects will be placed into emergence cages; emerging parasitoids will
be examined individually before being removed from emergence cages.
If hyperparasitoids (or any organism other than T. somervillei) are found in the emergence cages,
they will be destroyed.
Emergence cages will be kept in a dedicated room (the emergence room) in a separate building
from that where the dedicated room with captive scale/parasitoid populations (the rearing
facility) is located
T. somervillei will be moved to the rearing facility and placed on host plants with suitable scale
insect hosts in a mesh bag; the mesh bag ensures that parasitoids remain on the plant and the
scale insects are protected.
Mesh bags will not be removed (only shortly for monitoring purposes) from the plants until
parasitoid emergence.
When parasitoid emergence is expected, host plants with parasitized scales in their mesh bags
will be moved to the emergence room.
Personnel conducting field collections or any other field activity will not be allowed in the rearing
facility on the same day.
It is critical that the captive population is monitored continuously for hyperparasitoids even if best
practices are followed closely. In addition to individual examinations of parasitoids, yellow sticky cards
will be used in both the emergence room and the rearing facility to monitor (and also trap) any
hyperparasitoids or other insects. Controlled exposures inside mesh bags will protect T. somervillei.
The elimination of hyperparasitoids before importation to Christmas Island will involve the following
steps. First, the sanitary practices followed during rearing (see above) will ensure a "clean" captive
population. Second, emerged, adult parasitoids will be placed in airtight, glass vials. All individual
parasitoids will be inspected during the packing and hand‐carried from Kuala Lumpur to Christmas
Island Airport. All individuals will be examined again at the point of entry on Christmas Island while still
in the glass vials (training of quarantine personnel on Christmas Island will be provided in advance); if in
any doubt, the vial containing the questionable organism(s) will be destroyed. Third, an individual
examination will be conducted in the rearing facility on Christmas Island before the parasitoids are
removed from the glass vials.
In practice, the above procedures will ensure that hyperparasitoids are not transported to Christmas
Island from Malaysia.

34
Initial studies suggest that pathogens do not play a significant role in T. somervillei populations. In
Malaysia, not a single T. somervillei, either field collected or lab reared, showed any signs of infection by
entomopathogenic fungi or entomopathogenic nematodes.
Key Research Outcomes: an agreed, peer‐reviewed host list and host specificity protocol for evaluating
risk of introduction of T. somervillei to Christmas Island to control the invasive yellow lac scale,
Tachardina aurantiaca, with a recommendation that host specificity testing be conducted in the area of
origin of Tachardina aurantiaca. A protocol for ensuring that hyperparasitoids are not accidentally co‐
introduced with importation of T. somervillei to Christmas Island.

35
Project 3d. Rearing, release and monitoring protocols
An agreed protocol for rearing and release of biological control agents, and pre‐ and post‐
release monitoring for their efficacy in controlling target scale insects.
Research Findings 2009‐2013
Protocol for mass rearing of Tachardiaephagus in Malaysia (FRIM) and import this population into a
mass rearing facility on Christmas Island. The transfer of the founder population from Malaysia to
Christmas Island could be as simple as packaging freshly emerged adult T. somervillei inside a travel
cooler to buffer them against extremes of temperature in transit. The parasitoids should survive well
without access to food or water for 48 hours. There are direct flights between Kuala Lumpur and
Christmas Island, but changes in scheduling may mean the parasitoids could be transported via Perth.
This is the less desirable option because the probability of mortality is greater, and arrangements may
have to be made with DAFF Biosecurity Australia to hold the agent in quarantine if there is a delay
between flights.
On Christmas Island, a production facility will have to be built to mass‐rear Tachardiaephagus for field
release. As in Malaysia, the maintenance of captive parasitoid populations depends on the production
and maintenance of optimal host life stages of Tachardina on suitable host plants. The best locally
available host plant species for Tachardina are Inocarpus fagifer and Milletia pinnata.
The major infrastructure item for this project will be a dedicated glasshouse/screenhouse (preferably
closed, but solid‐roof screen house would probably be sufficient) for rearing of the biological control
agent. Suitable host plants will be transferred to the glasshouse/screenhouse. If chemical control of
pests and diseases on host plants is needed at the nursery, host plants must remain uninfested in the
glasshouse/screenhouse until insecticidal residues are no longer active. Host plants can be inoculated
with Tachardina within the same facility as long as plants in "waiting" and infested plants are separated
with a barrier that prevents Tachardina crawler movement (e.g., a water moat system). Availability of a
second, smaller glasshouse/screenhouse, to serve as a backup should the colony of Tachardiaephagus
fail in the primary glasshouse, would be optimal, although not absolutely necessary.
Tachardina and Tachardiaephagus may be difficult to mass‐rear due to the relatively long life cycle of
Tachardina (Project 2e, Table 5 ‐ from crawler stage to reproductive female is 80‐100 days, but
generations overlap) and the need for fresh host plants (that may or may not be reused) on a regular
basis. Depending on the difficulty of rearing Tachardiaephagus, their availability for releases maybe
limited at any given time. As mass‐rearing methods improve and production increases on Christmas
Island, the goal will be to provide the biological control agent for releases in all areas as needed.
The population of Tachardiaephagus will require careful monitoring and maintenance to minimize any
selection of laboratory‐adapted insects that perform poorly under field conditions. It may be necessary
to replenish the genetic diversity of the captive population through the subsequent introduction of
more insects from the founder population in Malaysia, or, more likely, from the field on Christmas
Island once the initial releases have been conducted. Population renewal will also counter the inherent
susceptibility of microhymenoptera to the loss of population heterozygosity. Sex determination in
microhymenoptera is usually haplodiploid – males are haploid, females diploid, and heterozygosity at a
multi‐allelic sex‐determining locus is required for femaleness. Inbreeding can lead to a preponderance
of homozygous diploids that will either be sterile males, or experience a very high rate of mortality.

36
Field release of Tachardiaephagus. The goal in releasing biological control agents is to generate
sufficient ‘propagule pressure’ (e.g., the size of each release, the frequency of releases, and the number
and spatial arrangement of release sites) to enable their successful establishment. Increasing propagule
pressure can enhance the likelihood of establishment by diminishing the role of chance (i.e., both
demographic and environmental stochasticity), and potentially increase the rate of spread from release
sites (Simberloff 2009). Initial timing, number of individuals released, and the frequency of releases of
both agents will depend on (a) the capacity and sustainability of mass rearing, (b) knowledge of the
biology of Tachardiaephagus, especially in relation to its host, Tachardina; and, (c) the attributes of
release sites.
Criteria for choosing suitable primary release sites will include: (a) positive evidence of host scale
infestation; (b) relatively high percentage of host plants of Tachardina or soft scale in the overstorey
and understorey; (c) occurrence of a high‐density YCA supercolony or at least the presence of YCA; and,
(d) site not subjected to current pesticide exposure (contact or systemic) or residues that could
compromise establishment of the agents. Some criteria (e.g., c and d) can be gleaned from the biennial
Islandwide Survey (Green and O'Dowd 2009) and followed up by more detailed site assessments to
determine (a) and (b). Each release at a site is likely to involve a specified number of adult insects;
specific methodology of release will largely depend on knowledge currently being gathered about
Tachardiaephagus in Malaysia and parasitoids of soft scale insects (e.g. Coccophagus) already present
on Christmas Island.
Fipronil, the intoxicant in Antoff®, the bait currently used to control YCA supercolonies on Christmas
Island, is known to affect the longevity, fecundity, and behaviour of some parasitoids (e.g., fipronil used
in vineyards to control ants can have acute toxic effects on Anagyrus sp. nr pseudococci and
Coccidoxenoides perminutus, two microhymenopteran parasitoids of mealybugs – Mgocheki and
Addison 2009). Thus, exposure of the biological control agents at release sites to baiting (especially
aerial baiting where a fine dust is produced and a fraction of the bait is retained in the canopy) must be
avoided. Coordination between field release and monitoring of biological control agents with National
Parks staff involved in chemical control of YCA supercolonies will be critical during this phase of this
project.
Training on release methodology and criteria will be provided to National Park personnel. Some
additions to the Island Wide Survey (e.g., determination of host tree species composition, inspection of
understorey for Tachardina and soft scale insects) could facilitate selection of release sites.
Interrogation of the survey database to identify the baiting history at waypoints will be an essential
precursor to release of the biological control agents. The National Parks field crew will receive training
in identifying and collecting scale insects and parasitoids.
Monitor the establishment, spread, and impact of Tachardiaephagus and Coccophagus. The absence
of effective, quantitative monitoring for the establishment, spread and impact of most introduced
biological control agents has been the Achilles’ heel of many biological control programs (McEvoy 1996).
Estimation of the success or failure of many past biological control programs has relied on subjective
measures, often post hoc expert opinion alone (e.g., DeBach et al. 1971, Greathead 1989, Griffiths and
Julien 1998). For biological control on Christmas Island, protocols to quantify the establishment,
population status, spread, and impact of biological control agents are essential.
Two approaches will be used. First, a field experiment will be conducted using a Before‐After‐Control‐
Impact design to determine the establishment and population dynamics of the agents, and the effect of

37
their release on host scale densities (counts per length of stem or per leaf) and parasitization rates, both
in the canopy (random sampling of host plant material collected using a shotgun) and in the
understorey (from saplings of known host trees), and abundance of YCA (using counts on tree trunks
and on the forest floor) at release and control sites before and after release of biological control agents.
Sites (each 2‐4 hectares) would be sampled 4 times before release of the agents and 4 times afterwards
at two monthly intervals. Results will be analyzed as a one‐way repeated measures ANOVA, using
release of the biological control agents as the main factor, and comparing response variables before and
after release. In this design, the time x treatment interaction is the key term, with a significant
difference in response variables after, but not before release. Thus, this experiment at the forest plot
scale would establish both the outcome of the release and the mechanism(s) driving any change in YCA
abundance.
Second, at the much broader, island‐wide scale the outcome of agents releases on YCA supercolonies
will be determined by comparing changes in YCA trunk traffic and ground activity (using card counts) at
four‐month intervals at replicated release and control sites across the island. The number of control
sites will be determined based on the release sites and area availability. Ideally, control sites should be
distant enough from release sites so that the chances for biological control agent dispersal are low for a
reasonable period of time. It will be necessary to determine how many of the selected release sites will
be actually available for releases and the available areas for control sites where no other YCA
management practices (i.e., application of toxic ant baits) will be applied.
Spread of the biological control agents beyond release sites will be determined by placing potted
‘sentinel’ host plants, infested with Tachardina or coccoid scales, at set distances (probably at a
logarithmic scale) from replicated release points, followed by their later collection to determine
parasitization rates with distance from each release point. It may also be feasible to use the biennial
Island Wide Survey to document spread of the biological control agent, at least onto understorey
seedlings and saplings, at waypoints surrounding release sites.
Key Research Outcomes: Identification of the requirements for the establishment of a founder
population of Tachardiaephagus in Malaysia free of pathogens and hyperparasites, importation of this
population under permit to Christmas Island, establishment and expansion of this population in a mass‐
rearing facility on Christmas Island, an agreed protocol for rearing and release of biological control
agents, and pre‐ and post‐release monitoring for their efficacy in controlling target scale insects and
reducing densities and impacts of the invasive yellow crazy ant Anoplolepis gracilipes.

38
References
Abbott, K.L. 2004. Alien ant invasion on Christmas Island, Indian Ocean: The role of ant‐scale
associations in the dynamics of supercolonies of the yellow crazy ant, Anoplolepis gracilipes. Ph.D
Thesis, Monash University, Melbourne, Australia.
http://arrow.monash.edu.au/vital/access/manager/Repository/monash:6496 (accessed 21 January
2013)
Abbott, K.L. 2005. Supercolonies of the invasive yellow crazy ant, Anoplolepis gracilipes, on an oceanic
island: forager activity patterns, density and biomass. Insectes Sociaux 52: 266–273.
Abbott, K.L. and P.T. Green. 2007. Collapse of an ant‐scale mutualism in a rainforest on Christmas Island.
Oikos 116: 1238‐1246.
Beeton, B., A. Burbidge, G. Grigg, R. How, N. McKenzie, and J. Woinarski. 2010. Final Report of the
Christmas Island Expert Working Group to Minister for the Environment, Heritage and the Arts.
Canberra, A.C.T., Australia. http://www.environment.gov.au/parks/publications/christmas/final‐
report.html (accessed 21 January 2013)
Bellis, G.A., J.F. Donaldson, M. Carver, D.L. Hancock, and M.J. Fletcher. 2004. Records of insect pests on
Christmas Island and the Cocos (Keeling) Islands, Indian Ocean. Australian Entomologist 31: 93‐102.
Ben‐Dov, Y. 2012. Scalenet (http://www.sel.barc.usda. gov/scalenet/scalenet.htm).
Boland, C.R.J., M.J. Smith, D.J. Maple, B. Tiernan, R. Barr, R. Reeves, and F. Napier. 2011. Heli‐baiting
using low concentration fipronil to control invasive yellow crazy ant supercolonies on Christmas Island,
Indian Ocean. Pp. 152‐156 in Veitch, C.R., Clout, M.N., and Towns, D.R. (eds.). Island invasives:
eradication and management. Proceedings of the International Conference on Island Invasives. Gland,
Switzerland: IUCN and Auckland, New Zealand.
http://www.issg.org/pdf/publications/island_invasives/pdfhqprint/2boland.pdf (accessed 21 January
2013)
Butcher, R. and J. Hale. 2010. Ecological character description for The Dales Ramsar site. Unpublished
report to the Department of the Environment, Water, Heritage and the Arts, Canberra, A.C.T., Australia.
http://www.environment.gov.au/water/publications/environmental/wetlands/61‐ecd.html (accessed
22 January 2013)
Campbell, T.G. 1968. Entomological survey of Christmas Island (Indian Ocean) with special reference to
the insects of medical, veterinary, agricultural and forestry significance. Unpublished Report, CSIRO
Division of Entomology, Canberra, A.C.T., Australia. 48 pp.
Centre for Environment Stress and Adaptation. 2011. Monitoring of the 2009 aerial baiting of yellow
crazy ants (Anoplolepis gracilipes) on non‐target invertebrate fauna on Christmas Island. Unpublished
Report for the Director of National Parks, Canberra, A.C.T., Australia.
http://www.environment.gov.au/parks/publications/christmas/fipronil‐report.html (accessed 22
January 2013)
Claussen, J. 2005. Native plants of Christmas Island. Flora of Australia, Supplementary Series 22.
Australian Government Department of the Environment and Heritage, Australian Biological Resources
Study. Canberra, A.C.T., Australia.

39
Commonwealth of Australia. 2006. Threat Abatement Plan to Reduce the Impacts of Tramp Ants on
Biodiversity in Australia and its Territories. Department of the Environment and Heritage, Canberra,
A.C.T., Australia.
http://www.environment.gov.au/biodiversity/threatened/publications/tap/trampants.html (accessed
21 January 2013)
Cryan, R.C. and J.M. Urban. 2012. Higher‐level phylogeny of the insect order Hemiptera: is
Auchenorrhyncha really paraphyletic? Systematic Entomology 37: 7‐21.
CSIRO. 1990. CSIRO Entomological survey of Christmas Island. Phase 2. Unpublished Report. Australian
National Parks and Wildlife Service Consultancy Agreement. 67 pp.
Davis, N.E., O’Dowd, D.J., Green, P.T., and Mac Nally, R. 2008. Effects of alien ant invasion on
abundance, behaviour, and reproductive success of endemic island birds. Conservation Biology 22:
1165‐1176.
Davis, N.E., D.J. O'Dowd, R. Mac Nally, and P.T. Green. 2010. Invasive ants disrupt frugivory by endemic
island birds. Biology Letters 6: 85‐88.
DeBach, P., D. Rosen, and C.E. Kennett. 1971. Biological control of coccids by introduced natural
enemies. Pp. 165 ‐194 in Huffaker, C.B. (ed.) Biological Control. Plenum Press, New York.
Detto, T. and B. Tiernan. 2011. The 2011 island‐wide survey report on Yellow Crazy Ants and Red Crabs.
Unpublished report to Christmas Island National Park. Christmas Island.
Drew, M.M., S. Harzsch, M. Stensmyr, S. Erland, and B.S. Hansson. 2010. A review of the biology and
ecology of the Robber Crab, Birgus latro (Linnaeus, 1767) (Anomura:Coenobitidae). Zoologischer
Anzeiger 249: 45‐67.
Ferrar, P., I.W. Forno, and A.L. Yen. 2004. Report of the Review of the Management of Biosecurity Risks
Associated with the Importation and Release of Biological Control Agents. Australian Government
Department of Agriculture, Fisheries and Forestry, Canberra, Australia. 27 pp.
Giraud, T., J.S. Pedersen, and L. Keller. 2002. Evolution of supercolonies: the Argentine ants of southern
Europe. Proceedings of the National Academy of Science USA 99: 6075–6079.
Göllner‐Scheiding, U. 1980. Revision der afrikanischen Arten sowie Bemerkungen zu weiteren Arten der
Gattungen Leptocoris Hahn, 1833, und Boisea Kirkaldy, 1910. Deutsche Entomologische Zeitschrift, N.F.
27: 103‐148.
Greathead, D.J. 1989. Biological control as an introduction phenomenon: a preliminary examination of
programmes against Homoptera. The Entomologist 108: 28‐37.
Green, P.T. and D.J. O’Dowd. 2009. Management of invasive invertebrates: lessons from the
management of an invasive alien ant. Pp. 153‐172 in Clout, M.N. and P.A. Williams (eds.). Invasive
Species Management: A Handbook of Principles and Techniques. Oxford University Press, Oxford.
Green, P.T., S. Comport, and D. Slip. 2004. The management and control of the invasive alien crazy ant
(Anoplolepis gracilipes) on Christmas Island, Indian Ocean: the aerial baiting campaign, September 2002.
Unpublished final report to Environment Australia and the Crazy Ant Steering Committee.
Green P.T., P.S. Lake, and D.J. O’Dowd. 1999. Monopolization of litter processing by a dominant land
crab on a tropical oceanic island. Oecologia 119: 435‐444.

40
Green, P.T., D.J. O’Dowd, and P.S. Lake. 2008. Recruitment dynamics in a rainforest seedling community:
context‐independent impact of a keystone consumer. Oecologia 156: 373‐385.
Green, P.T., D.J. O’Dowd, K.L. Abbott, M. Jeffery, K. Retallick, and R. Mac Nally. 2011. Invasional
meltdown: invader‐invader mutualism facilitates a secondary invasion. Ecology 92: 1758‐1768.
Green, P.T., D.J. O’Dowd, and P.S. Lake. 1997. Control of seedling recruitment by land crabs in rain
forest on a remote oceanic island. Ecology 78: 2474‐2486.
Green, P.T., D.J. O’Dowd, and P.S. Lake. 2001. From resistance to meltdown: secondary invasion of an
island rain forest. Pp. 451‐455 in Ganeshaiah, K.N., R. Uma Shankar, and K.S. Bawa (eds.). Tropical
Ecosystems: Structure, Diversity and Human Welfare. Proceedings of the International Conference on
Tropical Ecosystems, Bangalore, India. Oxford‐IBH, New Delhi.
Griffiths, M.W. and M.H. Julien. 1998. Biological control of weeds: a world catalogue of agents and their
target weeds. CABI, Oxford.
Gullan, P. J. and L. G. Cook. 2007. Phylogeny and higher classification of the scale insects (Hemiptera:
Sternorrhyncha: Coccoidea). Zootaxa 1668: 413‐425.
Hale, J. and R. Butcher. 2010. Ecological Character Description for Hosnie’s Spring Ramsar Site.
Unpublished report to the Department of the Environment, Water, Heritage and the Arts, Canberra,
A.C.T., Australia. http://www.environment.gov.au/water/publications/environmental/wetlands/40‐
ecd.html (accessed 21 January 2013)
Hayat, M., S. Schroer, and R.W. Pemberton. 2010. On some Encyrtidae (Hymenoptera: Chalcidoidea) on
lac insects (Hemiptera: Kerriidae) from Indonesia, Malaysia and Thailand. Oriental Insects 44: 23‐33.
Holmes, J. and G. Holmes. 2002. Conservation status of the flora of Christmas Island, Indian Ocean.
Unpublished report to Environment Australia/Parks Australia North. Glenn Holmes and Associates,
Atherton, Queensland.
Howard, F.W., R. Pemberton, S. Schroer, and G. Hodges. 2010. Paratachardina pseudolobata
(Coccoidea: Kerriidae): Bionomics in Florida. Florida Entomologist 93: 1‐7.
James, D. and N. Milly. 2006. A biodiversity inventory database for Christmas Island National Park. A
report for the Department of Finance & Administration and Department of Environment & Heritage.
Director of National Parks, Australian Government, Canberra. 49 pages.
Kuhlmann, U., U. Schafner, and P.G. Mason. 2006. Selection of non‐target species for host specificity
testing. Pp. 15‐37 in Bigler, F., D. Babendreier, and U. Kuhlmann (eds.). Environmental Impact of
Invertebrates for Biological Control of Arthropods: Methods and Risk Assessment. CABI Publishing,
Delemont, Switzerland.
Lake, P.S. and D.J. O’Dowd. 1991. Red crabs in rainforest, Christmas Island: biotic resistance to invasion
by an exotic snail. Oikos 62: 25‐29.
Lee, J.C., G.E. Heimpel, and L. Leibee. 2004. Comparing floral nectar and aphid honeydew diets on the
longevity and nutrient levels of a parasitoid wasp. Entomologia Experimentalis et Applicata 111: 189‐
199.
Linnavuori, R. 1975. Homoptera: Cicadellidae, Supplement. Insects of Micronesia 6. No. 9. pp. 611‐632

41
Lowe, S., M. Browne, and S. Boudjelas. 2000. 100 of the world’s worst invasive alien species. Aliens 12:
S1‐S12. http://www.issg.org/database/species/search.asp?st=100ss (accessed 21 January 2013)
Lumsden, L., M. Schulz, R. Ashton, and D. Middleton. 2007. Investigation of the threats to the Christmas
Island Pipistrelle. Unpublished report to the Department of Environment and Water Resources,
Canberra, A.C.T., Australia. http::// http://ausbats.org.au/#/resources/4553704436 (accessed 21
January 2013)
Marr, R.M., D.J. O’Dowd, and P.T. Green. 2003. Assessment of non‐target impacts of Presto® 01 ant bait
on litter invertebrates in Christmas Island National Park, Indian Ocean. Unpublished report to Parks
Australia North, Darwin, NT.
McEvoy, P.B. 1996. Host specificity and biological pest control. BioScience 46: 401‐405.
Mgocheki, N. and P. Addison. 2009. Effect of contact pesticides on vine mealybug parasitoids, Anagyrus
sp. near pseudococci (Girault) and Coccidoxenoides perminutus (Timberlake) (Hymenoptera: Encyrtidae).
South African Journal of Entomology and Viticulture 30: 110 – 116.
Narayanan, E.S. 1962. Pests of lac in India. Pp. 90‐113 in Mukhopadhyay, B. and Muthana, M.S. (eds.). A
Monograph on Lac. Indian Lac Research Institute. Nancum, Ranchi, India.
Neumann, G., P.T. Green, and D.J. O’Dowd. 2011. First record of Pulvinaria urbicola (Hemiptera:
Coccidae), a potentially damaging scale insect, on Christmas Island, Indian Ocean. Unpublished report
to Parks Australia. http://www.environment.gov.au/parks/publications/christmas/pubs/pulvinaria‐
report‐2011.docx (accessed 21 January 2013)
Neumann, G., R.G. Hollingsworth, and P.A. Follett. 2007. First record of the Hawaiian endemic scale,
Colobopyga pritchardiae (Hemiptera: Holimococcidae), on the Island of Hawaii. Proceedings of the
Hawaiian Entomological Society 39: 39‐41.
Neumann, G., P. A. Follett, R. G. Hollingsworth, and J. de León. 2010. High host specificity in Encarsia
diaspidicola (Hymenoptera: Aphelinidae), a biological control candidate against the white peach scale in
Hawaii. Biological Control 54: 107‐113.
Noyes, J.S. 2012. Universal Chalcidoidea Database. World Wide Web electronic publication.
http://www.nhm.ac.uk/chalcidoids
Noyes, J.S. and M. Hayat. 1994. Oriental mealybug parasitoids of the Anagyrini (Hymenoptera:
Encyrtidae). CAB International, Wallingford, UK.
O’Dowd, D.J. and P.T. Green. 2000. Design and feasibility of an island‐wide survey of the invasive alien
ant Anoplolepis gracilipes and its impact on Christmas Island, Indian Ocean. Unpublished report to Parks
Australia North, Darwin, NT.
O’Dowd, D.J. and P.T. Green. 2010. Invasional meltdown: do invasive ants facilitate secondary
invasions? Pp. 271‐272 in Lach, L., C. Parr, and K. Abbott (eds.) Ant Ecology. Oxford University Press,
Oxford.
O’Dowd, D.J. and P.T. Green. 2003. Potential for indirect biological control of the yellow crazy ant
(Anoplolepis gracilipes) on Christmas Island, Indian Ocean. Report to Parks Australia North, Darwin, NT.
O’Dowd, D.J., P.T. Green, and P.S. Lake. 2003. Invasional ‘meltdown’ on an oceanic island. Ecology
Letters 6: 812‐817.

42
O’Dowd, D.J., P.T. Green, and P.S. Lake. 1999. Status, impact, and recommendations for research and
management of exotic invasive ants in Christmas Island National Park. Unpublished report to
Environment Australia, Canberra, A.C.T., Australia
http://www.issg.org/database/species/reference_files/Christmas_Island_Report.pdf (accessed 21
January 2013).
O’Dowd, D.J., P.T. Green, G. Neumann, and S. Wittman. 2012. Executive summary: research and
development for indirect biological control of the yellow crazy ant (Anoplolepis gracilipes) on Christmas
Island, Indian Ocean. Unpublished report to the Director of National Parks and the Crazy Ant Scientific
Advisory Panel, 27 pp.
Palmer, W.A., T.A. Heard, and A.W. Sheppard. 2010. A review of Australian classical biological control of
weeds programs and research activities over the past 12 years. Biological Control 52: 271‐287.
Poorter, L. and F. Bongers. 2006. Leaf traits are good predictors of plant performance across 53 rain
forest species. Ecology 87: 1289‐1301
Prinsloo, G.L. 1977. On the encyrtid parasitoids (Hymenoptera: Chalcidoidea) of lac insects (Hemiptera:
Lacciferidae) from southern Africa. Journal of the Entomological Society of South Africa 40: 47‐72.
Prinsloo, G.L. 1983. A parasitoid‐host Index of Afrotropical Encyrtidae (Hymenoptera: Chalcidoidea).
Entomological Memoir of the Department of Agriculture Republic of South Africa no. 60. 35 pages.
Ross, L., N.B. Hardy, A. Okuso, and B.B. Normark. 2012. Large population size predicts the distribution of
asexuality in scale insects. Evolution 67: 196‐206.
Sands, D.P.A. and R.G. Van Driesche. 2004. Using the scientific literature to estimate the host range of a
biological control agent. Pages 16‐23 in Van Driesche, R.G. and R. Reardon (eds.). Assessing host ranges
for parasitoids and predators used for classical biological control: a guide to pest practice. University
States Department of Agriculture Forest Health Technology Enterprise Team, Morgantown, West
Virginia USA. FHTET‐2004‐3.
Schultz, M. 2004. National Recovery Plan for the Christmas Island Shrew Crocidura attenuata trichura.
Commonwealth of Australia, Canberra.
http://www.environment.gov.au/biodiversity/threatened/publications/recovery/c‐attenuata‐
trichura/index.html (accessed 21 January 2013)
Schulz, M. and C. Barker. 2008. A Terrestrial Reptile Survey of Christmas Island, May‐June 2008.
Unpublished report for Parks Australia North, Darwin, NT.
Schulz, M. and L.F. Lumsden. 2004. National Recovery Plan for the Christmas Island Pipistrelle
Pipistrellus murrayi. Commonwealth of Australia, Canberra.
http://www.environment.gov.au/biodiversity/threatened/publications/recovery/p‐murrayi/pubs/p‐
murrayi.pdf (accessed 21 January 2013)
Secord, D. and P. Karieva. 1996. Perils and pitfalls in the host specificity paradigm. BioScience 46: 448‐
453.
Sharma, K.K., A.K. Jaiswal, and K.K. Kumar. 2006. Role of lac culture in biodiversity conservation: issues
at stake and conservation strategy. Current Science 91: 894‐898.
Simberloff, D. 2009. The role of propagule pressure in biological invasions. Annual Review of Ecology,
Evolution and Systematics 40: 81‐102.

43
Smith. M.J., H. Cogger, B. Tiernan, D. Maple, C. Boland, F. Napier, T. Detto, and P. Smith. 2012. An
oceanic island reptile community under threat: the decline of reptiles on Christmas Island, Indian
Ocean. Herpetological Conservation and Biology 7: 206‐218.
Suhr, E.L., D.J. O’Dowd, S.W. McKechnie, and D.A. Mackay. 2011. Genetic structure, behaviour and
invasion history of the Argentine ant in Australia. Evolutionary Applications 4: 471–484.
Sunamura, E., X. Espadaler, H. Sakamoto, S. Suzuki, M. Terayama, and S. Tatsuki. 2009. Intercontinental
union of Argentine ants: behavioral relationships among introduced populations in Europe, North
America, and Asia. Insectes Sociaux 56: 143–147.
Thomas, M.L., K. Becker, K. Abbott, and H. Feldhaar. 2010. Supercolony mosaics: two different invasions
by the yellow crazy ant, Anoplolepis gracilipes, on Christmas Island, Indian Ocean. Biological Invasions
12: 677‐687.
Van Driesche, R., T. Center, M. Hoddle, and N. Mills. 2008. Can efficacy of new biological control agents
be predicted before their release? Pp. 1‐13 in Mason, P.G., D.R. Gillespie, and C. Vincent (eds.).
Proceedings of the Third International Symposium on Biological Control of Arthropods. Christchurch,
New Zealand, 8‐13 February 2008. United States Department of Agriculture, Forest Service,
Morgantown, WV, FHTET‐2008‐06, December, 2008, 636 p.
Wapshere, A.J. 1974. A strategy for evaluating the safety of organisms for biological weed control.
Annals of Applied Biology 77: 201‐211.
Wetterer, J.K. 2005. Worldwide distribution and potential spread of the long‐legged ant, Anoplolepis
gracilipes (Hymenoptera: Formicidae). Sociobiology 45: 77‐97.
Wheeler, D.E. 1996. The role of nourishment in oogenesis. Annual Review of Entomology 41: 407‐431.
Woods, B. and E. Steiner. 2012. Christmas Island fruit fly and scale survey. Report to Department of
Agriculture and Food, Government of Western Australia. 22 pp.
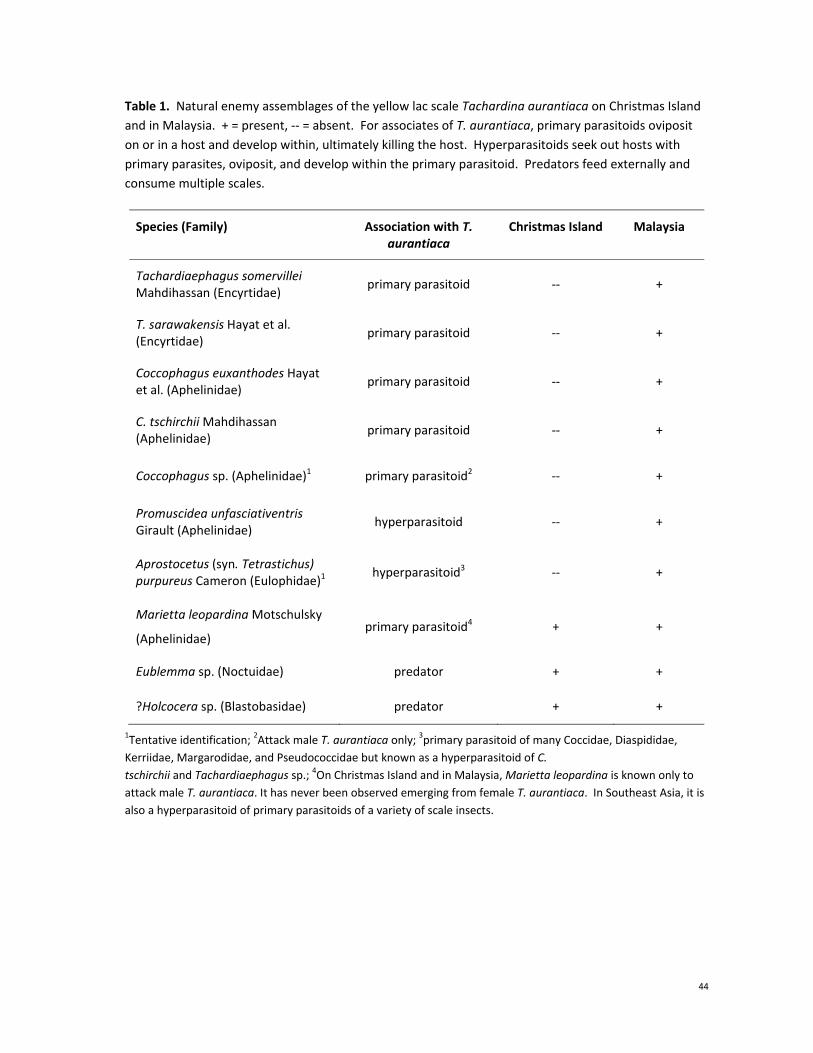
44
Table 1. Natural enemy assemblages of the yellow lac scale Tachardina aurantiaca on Christmas Island
and in Malaysia. + = present, ‐‐ = absent. For associates of T. aurantiaca, primary parasitoids oviposit
on or in a host and develop within, ultimately killing the host. Hyperparasitoids seek out hosts with
primary parasites, oviposit, and develop within the primary parasitoid. Predators feed externally and
consume multiple scales.
Species (Family) Association with T. aurantiaca
Christmas Island Malaysia
Tachardiaephagus somervillei Mahdihassan (Encyrtidae)
primary parasitoid ‐‐ +
T. sarawakensis Hayat et al. (Encyrtidae)
primary parasitoid ‐‐ +
Coccophagus euxanthodes Hayat et al. (Aphelinidae)
primary parasitoid ‐‐ +
C. tschirchii Mahdihassan (Aphelinidae)
primary parasitoid ‐‐ +
Coccophagus sp. (Aphelinidae)1 primary parasitoid2 ‐‐ +
Promuscidea unfasciativentris Girault (Aphelinidae)
hyperparasitoid ‐‐ +
Aprostocetus (syn. Tetrastichus) purpureus Cameron (Eulophidae)1
hyperparasitoid3 ‐‐ +
Marietta leopardina Motschulsky
(Aphelinidae) primary parasitoid4 + +
Eublemma sp. (Noctuidae) predator + +
?Holcocera sp. (Blastobasidae) predator + +
1Tentative identification;
2Attack male T. aurantiaca only;
3primary parasitoid of many Coccidae, Diaspididae,
Kerriidae, Margarodidae, and Pseudococcidae but known as a hyperparasitoid of C.
tschirchii and Tachardiaephagus sp.; 4On Christmas Island and in Malaysia, Marietta leopardina is known only to
attack male T. aurantiaca. It has never been observed emerging from female T. aurantiaca. In Southeast Asia, it is
also a hyperparasitoid of primary parasitoids of a variety of scale insects.

45
Table 2. Parasitization rates on mature females of Tachardina aurantiaca in the native range (Southeast
Asia) and in the introduced range (Christmas Island). Parasitization rates were calculated as the
proportion of mature female scale insects with one or more visible parasitoid emergence hole, either in
isolated aggregates (N = 5) at sites in Southeast Asia or pooled within sites on Christmas Island (N is in
brackets). This gives rates of parasitization at each site, but not the identity of the parasitoids. However,
only T. somervillei was collected at sites in Kuching and Singapore. All other locations in Southeast Asia
had a parasitoid assemblage of more than one species. For Christmas Island, number in parentheses
after site name indicates the total number of mature females examined.
Location Rate of Parasitization
(%, mean ± SE)
Native range (Southeast Asia)
Klang (Selangor, West Malaysia) 38 ± 17
Taman Ehsan (Selangor, West Malaysia) 46 ± 21
Singapore (National University Singapore campus) 73 ± 12
Kampung Istana, Kuching (Sarawak) 42 ± 23
Kampung Boyan, Kuching (Sarawak) 81 ± 6
Sandakan (Sabah) 76 ± 8
Sepilok (Sabah) 29 ± 13
Introduced Range (Christmas Island)
The Dales (Hugh’s – Sydney’s) (4000) 0 ± 0
Martin Point to CINP Boundary (1500) 0 ± 0
Dolly Beach Track (1000) 0 ± 0
North West Point Track (1500) 0 ± 0
Circuit Road (2000) 0 ± 0

46
Table 3. Records of known host families and genera for the primary parasitoid Tachardiaephagus
(Encyrtidae). Taxonomy follows Prinsloo (1977). The biological control agent under investigation,
Tachardiaephagus somervillei, is in bold. As a genus, Tachardiaephagus has an extremely broad
geographic range. With the exception of one probably erroneous host record in Africa (Prinsloo 1983),
all Tachardiaephagus species appear to be family specialists and restricted to the Kerriidae. For host
genera, number of species recorded as hosts is in parentheses. Based on Noyes (2012, Universal
Chalcidoidea Database, http://www.nhm.ac.uk/research‐
curation/research/projects/chalcidoids/database/), except for records for T. somervillei and T.
sarawakensis (Hayat et al. 2010; R.W. Pemberton, pers. comm.).
Parasitoid species Distribution Recorded hosts (all Kerriidae)
Tachardiaephagus somervillei
India, Malaysia, Thailand Kerria spp. (4)1
Tachardina aurantiaca
Tachardina sp.2
T. sarawakensis Sarawak (East Malaysia) Tachardina aurantiaca
T. tachardiae Brunei, China, India, Indonesia, Malaysia, Sri Lanka, Taiwan, Vietnam, Azerbaijan
Kerria spp. (8)
Paratachardina lobata
T. similis Afrotropical, South Africa Tachardina sp. (1)
T. absonus Afrotropical, South Africa Tachardina spp. (2)
T. communis Afrotropical, South Africa Tachardina spp. (5)
T. gracilis Afrotropical, South Africa Tachardina sp. (1)
1In Noyes (2012) both Kerria and Laccifer species are listed as hosts. However, Scalenet (Ben‐Dov et al. 2012) indicates that Laccifer is a synonym for Kerria, so we have synonymized these records with Kerria species.
2 Probably T. aurantiaca, since it is the only known Tachardina species in Asia.

47
Table 4. Scale insects of Christmas Island. It is highly probable that all of these species, with broad host
plant ranges and geographic distributions, are exotic to Christmas Island and introduced following
human settlement. The target species, Tachardina aurantiaca, for biological control is in bold.
Honeydew‐producing scale insects in bold occur commonly tended by YCA in supercolonies. Families are
arranged in increasing phylogenetic distance from the Kerriidae based on Gullan and Cook (2007) and
Ross et al. (2012). All scale insect taxa are 'neococcids' except for Icerya purchasi ('archeococcid').
Taxonomy and distributions from Ben‐Dov et al. (2012), http://www.sel.barc.usda.
gov/scalenet/scalenet.htm).
Family and Species1
Common Name Distribution Honeydew Producer
Kerriidae (lac scales) Paratachardina pseudolobata (Kondo & Gullan) Tachardina aurantiaca (Cockerell)
False lobate lac scale Yellow lac scale
Oriental, Nearctic, Neotropical Oriental
yes2
yes
Coccidae (soft scales) Ceroplastes ceriferus (Fabricius) C. destructor Newstead Coccus celatus De Lotto C. hesperidium Linnaeus Milviscutulus mangiferae (Green) Parasaissetia nigra (Nietner) Pulvinaria urbicola Cockerell P. psidii Maskell3 Saissetia coffeae (Walker) S. oleae (Olivier)
Indian wax scale White wax scale Green coffee scale Brown soft scale Mango shield scale Nigra scale Urbicola soft scale Green shield scale Black olive scale Hemispherical scale
Cosmopolitan Afrotropical, Australasia, Oriental Afrotropical, Australasia, Oriental Cosmopolitan Cosmopolitan Cosmopolitan Pantropical Cosmopolitan Pantropical Cosmopolitan
yes yes
yes
yes yes yes yes yes yes yes
Diaspididae (armoured scales) Aspidiotus destructor (Signoret) Hemiberlesia palmae (Cockerell) Ischnaspis longirostris (Signoret) 3 Lindingaspis sp. Pseudaulacaspis pentagona (Targioni Tozzetti) Unaspis citri (Comstock)
Coconut scale Tropical palm scale Black thread scale ‐‐ White peach scale White louse scale
Cosmopolitan Cosmopolitan Cosmopolitan ‐‐ Cosmopolitan Cosmopolitan
no no no ‐‐ no
no
Cerococcidae (ornate pit scales) Cerococcus indicus (Maskell)
Spiny brown coccid Cosmopolitan yes?
Pseudococcidae (mealybugs) Dysmicoccus finitimus Williams Asian coconut mealybug Australasia, Oriental yes
Ferrisia virgata (Cockerell) Nipaecoccus viridis (Newstead) Pseudococcus longispinus (Targioni Tozzetti)
Monophlebidae (giant scales) Icerya purchasi (Maskell)
Striped mealybug Spherical mealy bug Long‐tailed mealy bug
Cottony cushion scale
Cosmopolitan Cosmopolitan Cosmopolitan
Cosmopolitan
yes yes yes
yes
1Records from Campbell (1968), CSIRO (1999), O'Dowd et al. (2003), Bellis et al. (2004), Abbott (2004), Woods and Steiner (2012) and Neumann et al. (unpubl. results); 2Paratachardina pseudolobata produces honeydew but eject it instead of producing droplets that can be collected by ants (Howard et al. 2010) 3Tentative identifications

48
Table 5. Time (days ± SE) from crawler stage to emergence of either (a) adult males or (b) the
production of the next generation of crawlers from adult females in Tachardina aurantiaca on
seedlings of Acacia mangium x A. auriculiformis (n = 6), Milletia sp. (n = 7) and Inocarpus fagifer (n =
6) under laboratory conditions. All host plants were potted plants less than 1 m tall. Observations
on hosts A. mangium x A. auriculiformis and Milletia sp. were conducted in at the Forest Research
Institute of Malaysia in West Malaysia (Ong and Neumann, unpublished results). Observations on I.
fagifer were conducted on Christmas Island.
Acacia mangium x A.
auriculiformis Milletia sp. Inocarpus fagifer
(a) Days to male
emergence
40.1 ± 0.2 41.71 ± 0.13 42.5 ± 1.3
(b) Days to female
crawler production
87.6 ± 0.2 98.3 ± 0.5 99.7 ± 1.2
49
Table 6. Key Australian government contacts for progressing the importation and release of an exotic biological control agent on Christmas Island, Indian Ocean. Framework refers either to the DAFF protocol for biological control agents or the Environmental assessment procedure under the EPBC Act (EACD, DSEWPaC)
Framework Issue (Key URL) Section Key contact Addresses
DAFF
Import risk analysis
(http://www.daff.gov.au/ba/reviews/biological_control_agents/risk_analyses)
Grains & Forestry, DAFF
Tara Dempsey Assistant Manager, Processed Products, Biologicals & Pacific Plant Biosecurity, DAFF
Nomination of a target species for a biological control agent (http://www.daff.gov.au/ba/reviews/biological_control_agents/protocol_for_biological_control_agents/guidelines‐introduction‐exotic‐bcas‐weed‐and‐plants)
Approvals package to import and release an exotic biological control agent (URL as for nomination)
Office of the Plant Protection Officer, Plant Health Committee
Susie Collins Secretariat, Plant Health Committee, OCPPO
Amendment to live imports list (http://www.environment.gov.au/biodiversity/wildlife‐trade/lists/import/index. html)
Wildlife Trade Regulation, DSEWPaC
Michelle van der VoortWildlife Trade Regulation Section, DSEWPaC
[email protected] [email protected]
EACD Environmental referral and assessment (http://www.environment.gov.au/epbc/assessments/index.html)
Environmental Assessment and Compliance Division, DSEWPaC
Felicity McLean Assistant Secretary, Northwest Section, EACD, DSEWPaC
[email protected] [email protected]

50
Table 7. Established collaborators and cooperators for the biological control project.
Location Collaborator Expertise
Australia
ANU, Biological Sciences
University of Queensland, Biological Sciences
Prof. Penny Gullan
Dr. Lyn Cook
Biology and phylogeny of scale insects
Phylogeny and genetics of scale insects
India
Aligarh Muslim University, Aligarh, Uttar Pradesh
Dr. Mohammed Hayat
Microhymenopteran taxonomy
Peninsular Malaysia
Forestry Research Institute Malaysia (FRIM)
Universiti Sains Malaysia
Dr. Laurence Kirton
Ms Ong Su Ping
Entomology
Entomology (M.Sc student)
Sarawak
Sarawak Forestry, Botanical Research Centre, Semenggoh
Ms Lucy Chong
Mr Het Bin Kaliang
Entomology
Entomology
Sabah
Sabah Forestry Department, Forest Research Centre, Sepilok
Dr. A. Chung Yaw Chyang
Dr. Chey Vun Khen
Entomology
Entomology
Singapore
National University of Singapore (NUS), Biological Sciences
Dr. Rudolf Meier
Dr. Hugh Tan Tiang Wah
Entomology
Entomology
United States
USDA‐ARS, Invasive Plant Research Laboratory, Ft. Lauderdale FL
Dr. Robert Pemberton
Biological control of insects and weeds in natural areas, biosafety

51
Table 8. Expert ad hoc reviewers for the host test list and host‐specificity testing protocol (Project
3c). All of these colleagues have agreed to continue to advise us on this and other aspects of the
project.
Reviewer Expertise
Australia
Dr Don Sands OAM, CSIRO Ecosystem Sciences, Brisbane QLD
Biological control of scale insects
New Zealand
Dr Barbara Barratt, Principal Scientist, AgResearch Invermay, Biocontrol, Biosecurity and Bioprocessing, Mosgiel, New Zealand
Biosafety of biological control agents, risk assessment, biocontrol in natural areas, developed the BIREA (Biocontrol Information Resource for EPA Applicants) for New Zealand
Switzerland
Dr. Tim Haye, Commonwealth Agricultural Bureaux International, Geneva
Host range assessment and impacts of parasitoids.
United States
Dr. Robert Pemberton, USDA‐ARS Invasive Plant Research Laboratory, Ft. Lauderdale FL
Dr. Matthew Purcell, Team Leader, USDA‐ARS, Australian Biocontrol Laboratory, Brisbane QLD
Biological control of insects and weeds in natural areas, biosafety, biocontrol of the lobate lac scale
Biological control of insects and weeds

52
Table 9. Endemic hemipteran species known from Christmas Island and primary parasitoids
(superfamily Chalcidoidea: family Encyrtidae) associated with the families represented by the
endemic species. The data were extracted from the Universal Chalcidoidea Database (Noyes 2012).
The families Nogodonidae and Rhopalidae have no associated chalcidoid primary parasitoids and
therefore endemic species in these families on Christmas Island can most likely be excluded from all
further consideration. Cicadellidae and Delphacidae have the highest diversity of chalcidoid primary
parasitoids but have magnitudes lower diversity of encyrtid primary parasitoids. These data suggest
that the encyrtid primary parasitoids of families with endemic species on Christmas Island appear to
not have host range overlap with taxa where the target lac scale is included and the host range
separation is at the suborder level suggesting very distant phylogenetic separation. During the
database analysis, only records with species‐level chalcidoid identification were used. N/A indicates
not applicable
1 Xestocephalus izzardi is also reported from Palau in the western Pacific Ocean (Linnavuori 1975). Its status as an endemic on Christmas Island is questionable. 2 Leptocoris subrufescens on Christmas Island has been classified to subspecies status (L. subrufescens subrufescens,). Another subspecies (L. s. flava) is described from Yap, western Pacific Ocean (Göllner‐Scheiding 1980). More research is needed to resolve the taxonomic status of these two subspecies of L. subrufescens.
Endemic species Family No. chalcidoid associates of family
No. encyrtid 1o parasitoid species
of family
Suborder/Family host range of encyrtids parasitizing family
Xestocephalus
Izzardi1
Cicadellidae 627 6 Auchenorrhyncha
(Cicadellidae)
Oxypleura
calypso
Cicadidae 35 0 N/A
Clovia eximia Cercopidae 71 4 Auchenorrhyncha
(Cercopidae‐
Aphrophoridae)
Ugyops aristella Delphacidae 248 5 Auchenorrhyncha
(Delphacidae‐
Cicadellidae)
Varcia
flavicostalis
Salona oceanica
Nogodinidae 0 N/A N/A
Leptocoris
subrufescens2
Rhopalidae 0 N/A N/A

53
Figure 1. The yellow lac scale Tachardina aurantiaca is estimated to contribute a
large fraction of the honeydew economy at forest sites with YCA supercolonies.
At each site the estimated range in % contribution to SHI by T. aurantiaca is given,
based on per capita parity in honeydew production by T. aurantiaca and coccoid
scale insects (blue circles, mean = 70%, range 46‐86%) or a 3 times greater per
capita honeydew production by T. aurantiaca (red circles, mean 86.5%, range 72 –
95%).
0
10
20
30
40
50
60
70
80
90
100
70 80 90 100 110 120 130
% Contribution by Tachardina in
2000
Site Honeydew Index in 2000

54
Figure 2. Estimated change in the Site Honeydew Index specifically for Tachardina
aurantiaca on Inocarpus fagifer from 2000‐2012, at eight 0.25 ha sites in former
YCA supercolonies. Numbers on the y‐axis are an index of Tachardina abundance
and follow the methodology for calculation of the SHI, but applied only to
Inocarpus.
0
10
20
30
40
50
60
70
80
SHI o
f Tachardinaon In
ocarpus
2002 2012

55
Figure 3. Stable isotope analyses (δ15N) of yellow crazy ants from four declining
supercolonies showed that trophic position increased (i.e. YCA became more
carnivorous) as population densities decreased over 16 months in 2010‐2011. In
this figure, a trophic position > 3 indicates stronger carnivory while < 3 indicates
increasing herbivory. The datapoints are connected in temporal order for each
site, beginning at the right‐hand side.
2.4
2.6
2.8
3.0
3.2
3.4
3.6
1 10 100 1000
Trophic Position
YCA abundance (no. ants 30 s‐1)
Site 206
Site 318
Site 403
Site 582

56
Figure 4. Banding trees effectively excluded YCA from honeydew resources in the
forest canopy (A), resulting in a precipitous decline and the virtual elimination of
YCA traffic on tree boles 4 weeks after the tree bands were in place (B). Red
symbols indicate control plot (unbanded trees), blue symbols indicate treatment
plot (banded trees). Solid black line indicates when the bands were applied.
0
5
10
15
20
25
30
35
40
45
50
YCA trunk counts (no. ants 30 s‐1
B
A

57
Figure 5. Mean YCA abundance was estimated from replicate counts of the number of ants crossing
one quadrant of a 20 cm x 20 cm card in 30 s (A). Exclusion of the yellow crazy ant from the forest
canopy resulted in a significant and rapid decline in YCA abundance on the forest floor; YCA abundance
fell and diverged markedly from the control plot two weeks after tree bands were placed (B). Red
symbols indicate control plot (unbanded trees), blue symbols indicate treatment plot (banded trees).
Solid black line indicates when the bands were applied.
A
0
5
10
15
20
25
30
35
40
45
YCA ground counts (n0. card‐130 s‐1)
B
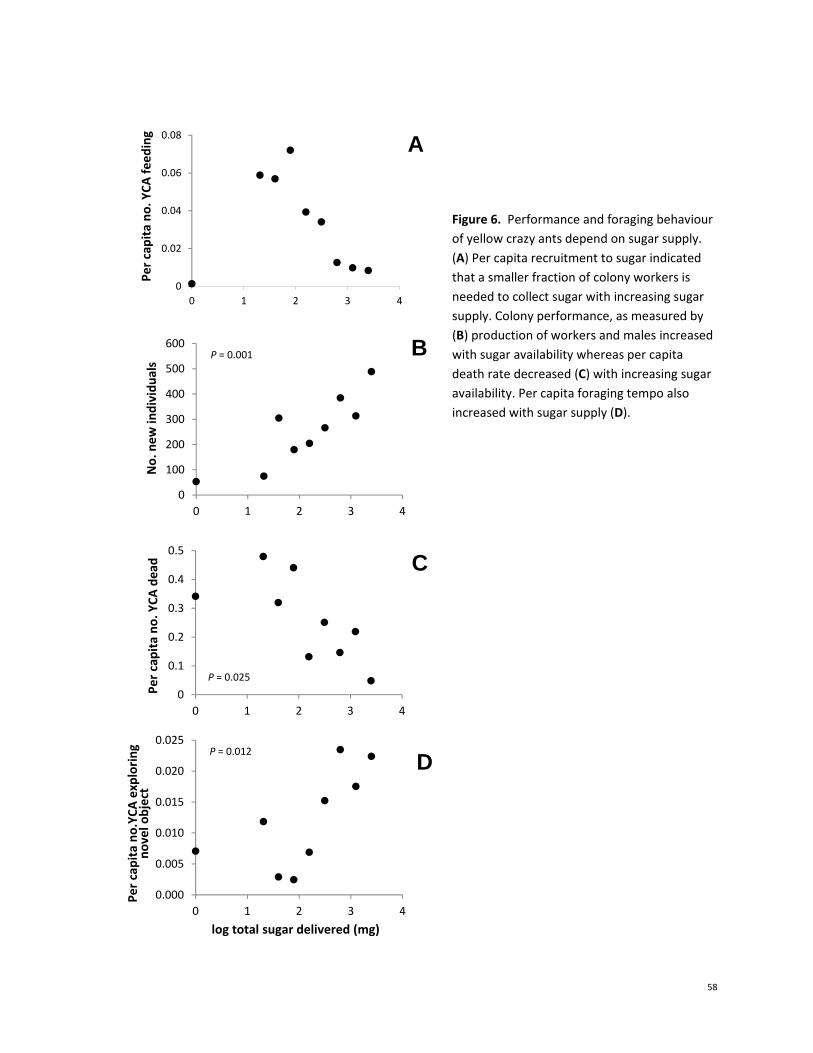
58
Figure 6. Performance and foraging behaviour
of yellow crazy ants depend on sugar supply.
(A) Per capita recruitment to sugar indicated
that a smaller fraction of colony workers is
needed to collect sugar with increasing sugar
supply. Colony performance, as measured by
(B) production of workers and males increased
with sugar availability whereas per capita
death rate decreased (C) with increasing sugar
availability. Per capita foraging tempo also
increased with sugar supply (D).
0
0.02
0.04
0.06
0.08
0 1 2 3 4
Per capita no. Y
CA feeding
0
100
200
300
400
500
600
0 1 2 3 4
No. n
ew in
dividuals
P = 0.001
0
0.1
0.2
0.3
0.4
0.5
0 1 2 3 4
Per capita no. Y
CA dead
P = 0.025
A
B
0.000
0.005
0.010
0.015
0.020
0.025
0 1 2 3 4
Per cap
ita no.YCA exploring
novel o
bject
log total sugar delivered (mg)
P = 0.012
D
C

59
Figure 7. Aggressive behaviours in the yellow crazy ant increased with sugar supply.
YCA with access to more sugar sprayed P. megacephala with formic acid sooner more
often (A), and killed more P. megacephala in 3:3 interaction trials (B).
0
1
2
3
4
0 1 2 3 4 5
No. tim
es YCA spray
P < 0.001
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0 1 2 3 4 5
No. d
ead
P. m
egacephala
log total sugar delivered (mg)
P = 0.077
A
B

60
Figure 8. Christmas Island (red circle) is the only known area of introduction and
invasion of the yellow lac scale Tachardina aurantiaca (Kerriidae). Yellow symbols
indicate sites in Peninsular Malaysia (Penang Island, Klang, Selangor, Singapore) and
Malaysian Borneo (Sarawak ‐ Kuching; Sabah ‐ Sandakan, Sepilok) where live
aggregates of Tachardina were found.
Christmas Island

61
Figure 9. Parasitism of the yellow lac scale Tachardina aurantiaca in Malaysia. A.
Parasitoid emergence holes of Tachardiaephagus sp. (Encyrtidae) in tests of an
aggregate of old adult females of T. aurantiaca on Milletia pinnata near Sandakan,
Sabah, Malaysian Borneo. Yellow crazy ants (Anoplolepis gracilipes) tended T.
aurantiaca at this site. B. Emergence holes of Coccophagus euxanthodes
(Aphelinidae) in the test of a T. aurantiaca on Acacia auriculiformis at Klang,
Selangor, Peninsular Malaysia. The smaller opening in the centre of the test is the
anal pore through which honeydew is produced. Weaver ants (Oecophylla
smaragdina) tended T. aurantiaca at Klang.
A B

62
Figure 10. Tachardiaephagus somervillei, a primary parasite of the yellow lac scale
Tachardina aurantiaca in Malaysia and Singapore (drawing from Narayanan 1962).
In initial studies in 2011‐2013, T. somervillei attacked T. aurantiaca across
Peninsular Malaysia and Malaysian Borneo, is the most abundant natural enemy of
T. aurantiaca, exhibited superparasitism (i.e., where multiple progeny emergence
from a single host individual), and heavily parasitized T. aurantiaca in the presence
of tending ants, including the yellow crazy ant. T. somervillei can be reared under
laboratory conditions (Ong and Neumann, unpublished results).
1 mm

63
Figure 11. The positive and significant relationship between female size of the
yellow lac scale Tachardina aurantiaca and the number of progeny of the parasitoid
Tachardiaephagus somervillei emerging from each host scale under field conditions
in Malaysia (R2 = 0.471, N = 50 females, p < 0.01).
0
1
2
3
4
5
1.0 1.5 2.0 2.5 3.0 3.5 4.0
No. o
f progeny of T. somervillei
Female T. aurantiaca size (mm)

64
Figure 12. Search methodologies for detecting native or endemic scale insects and
natural enemies of the yellow lac scale T. aurantiaca on Christmas Island.
Northwest Point is enlarged in the upper left‐hand side of the figure.

65
Figure 13. Size‐dependent fecundity of female Tachardina aurantiaca under
laboratory and field conditions, across a range of plant hosts. Female size was
determined by measuring the test at the greatest horizontal diameter with callipers.
The number of crawlers was determined by dissecting the females shortly before
crawler release. There is a significant correlation between female size and number
offspring produced (laboratory, open circles: R2 = 0.295, p < 0.001, n = 50; field solid
circles: R2 = 0.207, p = 0.006, n = 50)(Ong and Neumann, unpublished results).
0
100
200
300
400
500
600
700
2.0 2.5 3.0 3.5 4.0 4.5 5.0
No. crawlers / female
Female size (mm)

66
Figure 14. Portion of a phylogram of the scale insects based on nucleotide
sequencing of nuclear 18S rRNA for 72 species of scale insects and 10 outgroup taxa
(see Gullan and Cook [2007] for details). Only the neococcoids (red) are shown here.
For all families given here (except the Kermesidae), Bayesian posterior probabilities
(numerical values above branches) are > 90%. For a host test list, we will select
species from those families given in bold. The lac scales Kerriidae (bold and italics)
is a sister family of the soft scales (Coccidae), more distant to the armoured scales
(Diaspididae), and even further removed from the mealybugs (Pseudococcidae).
Pseudococcidae
Coccidae
Kerriidae
Kermesidae
Diaspididae
Gondwanan clade
BSE clade
Asterolecaniidae
Acanthococcid group
Dactylopiidae

Tropical Ecosystems Research Centre
PMB 44 Winnellie, NT 0822, Australia
T (03) 8944 8400 • ABN 41 687 119 230
CSIRO ECOSYSTEMS SCIENCES
15 January 2013 Mr Mike Misso Manager, Christmas Island National Park [email protected] Dear Mike, During CASAP’s meeting on 12 December 2012 we considered the executive summary of the report ‘Research and development for indirect biological control of the yellow crazy ant (Anoplolepis gracilipes) on Christmas Island, Indian Ocean’. Drs Green and O’Dowd were excused from this consideration due to their significant roles in the writing of the report. The report summarises results of research related to the potential use of biocontrol of the scale insect Tachardina aurantiaca as a means of controlling crazy ant infestations on Christmas Island. The research shows that: (1) The supply of liquid carbohydrate plays a key role in the dynamics of crazy ant colonies on CI; (2) T. aurantiaca is a key provider of liquid carbohydrate on CI; (3) In contrast to CI, within its native range in Malaysia T. aurantiaca has a diverse range of natural enemies, and is rare and patchily distributed; (4) In Malaysia, T. aurantiaca has high rates of parasitism by the encyrtid Tachardiaephagus somervillei , which appears to specialise on the family Kerriidae (not native to CI) and can be reared under laboratory conditions. The report concludes that prospects of successful biocontrol are good, and therefore the biocontrol program should proceed. CASAP members were unanimously of the view that the research had very high scientific merit, and effectively demonstrates the potential for successful biocontrol. It was considered that the risks associated with introducing a bio-control agent were low, particularly given existing bio-security risks on CI. CASAP endorses the conclusion of the report that the biocontrol program should proceed, and recommends that it covers soft scale as well as T. aurantiaca, since it is highly feasible and requires minimal extra resources to do so. Yours sincerely,
Prof Alan N. Andersen Chair, Christmas Island Crazy Ant Scientific Advisory Panel [email protected] (08) 8944 8431

1
Host specificity testing of Tachardiaephagus somervillei (Hymenoptera: 1
Encyrtidae), a biological control agent for the yellow lac scale 2
Tachardina aurantiaca (Hemiptera: Kerriidae) 3
G. Neumann1, D.J. O'Dowd2 and P.T. Green1 4
1Department of Botany, La Trobe University, Bundoora, Victoria 3086 Australia and 2School of 5
Biological Sciences, Monash University, Melbourne, Victoria 3800 Australia 6
7
Preamble 8
The yellow lac scale, Tachardina aurantiaca (Hemiptera: Kerriidae) is a damaging, invasive 9
pest on Christmas Island (Indian Ocean) and implicated in the formation of widespread, high-10
density supercolonies of the invasive yellow crazy ant (Anoplolepis gracilipes) (O'Dowd et al. 11
2003, Abbott and Green 2007). Suppression and control of crazy ant supercolonies may be 12
afforded by the importation and release of Tachardiaephagus somervillei (Hymenoptera: 13
Encyrtidae), a widespread and abundant parasitoid of Tachardina that is known to parasitize 14
lac scales in its native range in Southeast Asia (Hayat et al. 2010, Green et al. 2013). Below we 15
provide a ‘desktop’ evaluation (Sands and Van Driesche 2004) of risk of importation and 16
release of T. somervillei on Christmas Island and then describe a protocol for host-specificity 17
testing recommending that it be conducted in the area of origin of Tachardina aurantiaca. 18
19
1. Evaluation of risk based on host records of Tachardiaephagus from the 20
scientific literature, hemipteran diversity on Christmas Island, and host 21
records of encyrtid parasitoids 22
To focus host-specificity testing for Tachardiaephagus somervillei, we used four approaches to 23
assess the risk of importation and release of the exotic biological control agent T. somervillei 24
to Christmas Island. All four lines of evidence indicate that there is a very low likelihood that 25
importation and release of T. somervillei would harm species of concern on Christmas Island. 26
27
1.1 Known host range of Tachardiaephagus somervillei 28
We evaluated known host species records for all species of Tachardiaephagus using the 29
Universal Chalcidoid Database (Noyes 2012) so that host range could be estimated and risk of 30
release of T. somervillei assessed for species of concern on Christmas Island. The Universal 31
Chalcidoidea Databases (Noyes 2012) is the most comprehensive database for chalcidoid 32
parasitoids, with over 120,000 host/associate records (including associations with food plants 33
of the hosts) and > 140,000 distribution records of the parasitoids in the superfamily 34
Chalcidoidea. It is very well developed, regularly updated and extremely well referenced. 35
Nevertheless, large databases can contain errors affecting reliability, such as erroneous 36

2
published host records and outdated parasitoid taxonomy (Kuhlmann et al. 2006). The most 37
important source of error in this database is actual published erroneous records. Since all 38
records are referenced, doubtful records can be investigated and if necessary, filtered out. 39
Seven described species of Tachardiaephagus are distributed in Southeast and South Asia, 40
and sub-Saharan Africa (Noyes 2012). Host records of Tachardiaephagus are restricted to 41
species within three genera (Kerria, Tachardina, and Paratachardina) of lac insects (Kerriidae) 42
(Table 1). Furthermore, all host species records for Tachardiaephagus somervillei are within 43
two genera: Kerria, comprising the lac insects of commerce, and Tachardina. These records 44
suggest that Tachardiaephagus somervillei is highly likely to have a narrow host range 45
restricted to kerriid host species. 46
47
1.2 Status of other scale insects on Christmas Island 48
We determined insect species on Christmas Island that could conceivably constitute non-49
target species. Records for occurrence of scale insect species (Superfamily Coccoidea, the 50
same superfamily to which Tachardina aurantiaca belongs) on the island were compiled from 51
the literature and then supplemented by conducting >400 hours of structured search over 52
two years for endemic scale insects (Green et al. 2013). For specimens from the structured 53
search, all identifications were verified by Prof Penny Gullan (Australian National University), 54
a scale insect systematist. 55
A total of 24 species of scale insects in six families have been recorded on Christmas Island 56
(Table 2). All scale insects that could be considered as the most likely potential non-target 57
organisms are non-natives on Christmas Island and none are beneficial (Table 2). 58
59
1.3 Phylogeny of endemic hemipterans 60
The closest endemic relatives of the yellow lac scale on Christmas Island should be considered 61
to evaluate the risk of becoming non-target hosts of Tachardiaephagus somervillei. We 62
obtained a list of known endemic insect species on Christmas Island (James and Milly 2006) 63
and narrowed consideration of taxa on this list to the closest endemic relatives of lac scales 64
on Christmas Island in the Order Hemiptera, the same order to which the known target 65
species Tachardina aurantiaca belongs. These species comprise one true bug, a cicada, a 66
leafhopper, a spittlebug, and three planthoppers (Table 3). All of these endemic species occur 67
in different suborders (either suborder Auchenorrhyncha or Heteroptera) than the yellow lac 68
scale (suborder Sternorrhyncha). 69
70
71
72

3
Thus, any potential for non-target impacts by T. somervillei on these endemic Hemiptera 73
would require a host range that bridges this very substantial phylogenetic distance (see above 74
and Fig. 2 in Cryan and Urban 2012), as well as distinctive morphologies, life-histories and 75
ecological attributes to its known host taxa in the Kerriidae. 76
77
1.4 Host ranges of encyrtid parasitoids known to attack members of the hemipteran 78
families with endemic species on Christmas Island. 79
The Encyrtidae (Hymenoptera, Superfamily: Chalcidoidea), the family to which T. somervillei 80
belongs, is one of the most important parasitic wasp (parasitoid) families for the biological 81
control of harmful insects, including a variety of scale insects infesting woody plants (Noyes 82
and Hayat 1994, Noyes 2012). The Encyrtidae currently comprises 460 genera and 3735 83
species in 2 subfamilies. The subfamily Encyrtinae includes 353 genera and 2920 species, 84
while the Tetracneminae includes 107 genera and 815 species. Approximately half of all 85
encyrtid species are associated with scale insects (Hemiptera: Coccoidea)(Noyes 2012). 86
Encyrtids are generally endoparasitoids meaning that the parasitoid egg is laid directly inside 87
the host’s body where the hatching larva completes development feeding on the host’s 88
tissue, ultimately killing the host. Encyrtids mostly parasitize immature life stages (or, rarely, 89
adults), but some species in one genus (Microterys) are egg predators (Noyes 2012). 90
We used the Universal Chalcidoidea Database to analyze all records of encyrtid parasitoid 91
associates of the hemipteran families represented by endemic species on Christmas Island. 92
Next, all recorded primary hosts of encyrtid species associated with these taxa were 93
determined. Consequently, any encyrtid species that share (as hosts) both coccoid species 94
(scale insects) and any family represented by endemic species on Christmas Island could be 95
found. 96
There are no known records in the Encyrtidae of any parasitoid species that attacks both scale 97
insects (Coccoidea) and host species in hemipteran families that have endemic 98
representatives on Christmas Island (Table 3). All known encyrtid primary parasitoids known 99
to attack species in families which have endemic representatives on Christmas Island have 100
primary hosts only within suborder Auchenorrhyncha. While species in these hemipteran 101
families are attacked by many species of chalcidoid parasitoids, many fewer or no encyrtid 102
species are known to use them as hosts, and not one encyrtid species is reported with a host 103
range so extremely broad that it encompasses both scale insects and any of these hemipteran 104
families. 105
106
2. Host specificity testing protocol 107
Actual records for host range of Tachardiaephagus and known patterns of parasitism in the 108
Encyrtidae indicate that it is highly improbable that T. somervillei would exhibit an extremely 109
broad host range (one that would be unprecedented in any single species of Encyrtidae 110

4
known to attack scale insects) and utilize any of the endemic hemipteran species on 111
Christmas Island. 112
2.1 Terminology 113
A ‘test species’ is an insect species that is tested as a potential non-target insect. Since no 114
non-target scale insects were identified on Christmas Island, the list of test species will be 115
determined using the centrifugal approach (Kuhlmann et al. 2006, Neumann et al. 2010). In 116
this approach, species other than the known target host of the biological control agent are 117
tested with the most closely related species (least phylogenetic distance, and with close 118
similarities in biology and ecology) tested first then less similar species thereafter (“centrifugal 119
principle” - Wapshere 1974). 120
A test species will be considered a ‘suitable host’ if parasitoid emergence is observed during 121
any of the exposure tests. To be a suitable host, the parasitoid must (a) accept the test 122
species when exposed, (b) must oviposit, (c) the eggs must hatch into larvae, (d) the larvae 123
must complete development using the test species as the resource, (e) the larvae must 124
pupate and, finally, (f) the emerging adult parasitoid must be able to exit (emerge from) the 125
test species. If parasitoid emergence from a test species is observed, the test species will be 126
considered a suitable host without assessing the viability, fecundity, sex ratio and other 127
characteristics of the emerging parasitoid generation. 128
A test species will be considered ‘susceptible’ if the parasitoid causes significant mortality due 129
to probing, host feeding (when the parasitoid stabs the insect with its ovipositor and then 130
feeds on the insect’s hemolymph), oviposition, or oviposition and larval development. Note 131
that even if some mortality is observed due to the above, the test species will only be 132
considered susceptible if the mortality resulting to the exposure to the parasitoid is 133
significantly higher than in control groups (negative controls) of the test species where the 134
test individuals are not exposed to the parasitoid. 135
The ‘parasitoid’ in this case is T. somervillei (Hymenoptera: Encyrtidae), a widespread 136
parasitoid native to Southeast Asia. It is known to parasitize some scale insects in the family 137
Kerriidae (lac scales) including the ‘target host’, Tachardina aurantiaca (O’Dowd et al. 2012). 138
2.2 Selecting test species 139
Host specificity testing can be a long and difficult process, depending on how problematic it is 140
to establish and manage colonies of the parasitoid, the target host, the host plant of the 141
target host, and the test species. Furthermore, resources and time are not unlimited and 142
permitting issues to collect insects and run field experiments (when needed) in Malaysia can 143
add to the time required to complete host testing. Therefore, a manageable number of 144
species should be used (10 to 15, but fewer are acceptable if high specificity is found initially). 145
The species used will be largely determined by availability, so determining the actual test 146
species at species or genus level is not possible at this point. We can, however, predict to 147

5
some degree the families of the test species that will be selected. We will focus on neococcid 148
taxa including the Kerriidae, the family to which the target host belongs (see Fig. 1 and Gullan 149
and Cook 2007). Considering the phylogenetic relationships of scale insect families (Fig. 1), 150
we will aim to test one species in the Kerriidae, Paratachardina pseudolobata, which is also 151
invasive on Christmas Island (Table 2), and more than one species from the Coccidae. Species 152
in the Diaspididae will also be considered as a less closely related group of scale insects. Early 153
in the host testing process, an ‘out-group’ (test species phylogenetically more distant) will 154
also be used, most likely selected from the Pseudococcidae (Fig. 1). 155
As mentioned above, availability will determine species selection to some degree but in some 156
of the groups (e.g. Coccidae) there may be a number of species available. Species that can be 157
reared in laboratory conditions would be preferred but carrying out tests in field conditions 158
should also be acceptable as long as the tests can be well controlled. 159
2.3 No-choice tests 160
In no-choice tests, only the non-target test species will be provided to the parasitoid in the 161
experimental replicates. This is the most stringent test method that places the parasitoid 162
under high “oviposition pressure”. 163
For each test species, trials will be replicated 10 times. Each replicate will consist of 1) one 164
experimental cage with the test species (50 individuals in three different age groups/instars; 165
150 individuals in total) exposed to 10 female and 10 male parasitoids, 2) one positive control 166
cage consisting of 50 mature female T. aurantiaca exposed to 5 female and 5 male parasitoids 167
in order to confirm that the parasitoids used in the experiment are of good quality and 168
capable of parasitism in the given exposure time, and 3) one negative control cage which will 169
be similar to the experimental cage except the test species will not be exposed to parasitoids. 170
The negative controls will be used to determine any effect of parasitoid exposure on insect 171
mortality other than successful parasitization such as probing, host feeding and oviposition 172
without successful parasitoid development. 173
The cages will consist of fine mesh bags (sleeves) fixed on the branches of plants that serve as 174
host plants for the scale insects. Parasitoids will be collected from a lab colony and 175
introduced to the cages < 24 h after emergence. Exposure time to parasitoids will be 176
determined during preliminary studies to determine the optimal time frame for mating, egg 177
maturation and oviposition by the parasitoid. 178
Evaluation: 179
1) If the positive control does not yield any parasitoid emergence, the replicate will not be 180
evaluated (failed replicate). 181
2) If the positive control yields parasitoid emergence the replicate will be evaluated. The 182
parasitization rate in the positive will be noted but will not be compared to parasitization 183
rates in the test replicate. 184

6
3) Parasitoid emergence in the test replicate will be noted along with the approximate age 185
and developmental stage of the individuals yielding parasitoid emergence. These 186
individuals will not be used to compare mortality rates due to reasons other than 187
successful parasitism. 188
4) Mortality of test species in test replicates and negative controls will be noted. 189
Analysis: 190
1) If any test replicates yielded any number of parasitoid emergences, the test species will 191
be considered a suitable host. The level of suitability of the test species will not be 192
compared to the suitability of the target host at this stage of the project. 193
2) Mortality (except mortality due to successful parasitism) rates of test insects and insects 194
in the negative control will be compared using 2-sample t-tests. If the mortality rate is 195
significantly higher (α = 0.05), the test species will be considered susceptible and the 196
reasons for mortality will be investigated. 197
If a test species is susceptible (but not suitable), behavioural observations will be made to 198
determine the cause of the effect of parasitoid exposure. The observational arena will be a 199
‘window box’ arena. This arena is constructed by constructing a 10-cm W x 10-cm L x 5-cm H 200
box with one side being glass. The box is constructed so that it can be placed on the stem of a 201
potted host plant on which the test insects feed. Female parasitoids are then liberated inside 202
the box and the box closed. The potted plant then can be placed flat on its side and the box 203
positioned under a dissecting microscope. Video footage is then captured using a high 204
definition video camera with a microscope adaptor. The footage (4-6 h per observation) can 205
be analyzed later for the cause(s) of mortality (i.e. probing, host feeding, or oviposition 206
without full parasitoid development). If oviposition is suspected, dissections will also be 207
made to investigate parasitoid larval development. 208
2.4 Other testing methods 209
We considered using choice tests (where the parasitoid is presented with the test species and 210
the target host simultaneously) and sequential no-choice tests (where the parasitoid is 211
presented with the target host first then it is transferred on to the test species). However, in 212
our case these tests may have limited value for reasons given below. 213
A choice test would show whether a test species that is a suitable host in no-choice tests 214
would be consistently attacked when the target host is also available. A physiologically 215
suitable host may not be preferred over the target host and the assumption could be made 216
that parasitization would not occur (or at very low level) in field conditions. This test would 217
be used in cases where the suitable host (found suitable in a no-choice test) is of concern, e.g. 218
a beneficial or endemic non-target species that coexists with the target host. However, this 219
situation does not arise on Christmas Island where all known scale insects are non-native and 220
none of them are beneficial. This test is further limited by the assumption that the test 221

7
species and the target host occupy the same space hence providing the parasitoid with a 222
choice. This would also not be the case on Christmas Island. 223
A sequential no-choice test would further show the unsuitability of a test species. The 224
parasitoid may be ‘motivated’ to attack the test species even if it did not attack it in a simple 225
no-choice test if the parasitoid is first allowed to initiate oviposition behavior and to gain 226
experience. This test may be even more stringent than the simple no-choice test and one 227
would consider using it in the case of a test species that is of great concern to further show 228
the unsuitability of the test species. This would again require a beneficial or endemic non-229
target species neither of which exists on Christmas Island. Furthermore, similar to the choice 230
test, this would assume either the coexistence or very close proximity of the test species and 231
the target host. 232
233
3. Options for the location for host-specificity testing 234
There are three options for the location where host specificity testing could be conducted: 1) 235
in quarantine containment at the release location (Christmas Island); 2) in quarantine 236
containment on mainland Australia; and 3) in the native geographical range of the biological 237
control agent where no containment would be necessary. 238
Option 1. Testing in containment at the release location (Christmas Island) 239
This option is the least attractive and involves increased risks and expenses. There is no 240
quarantine containment facility on Christmas Island, a remote oceanic island. The expense of 241
constructing such a facility to Quarantine Approved Premises Criteria (Quarantine Insectary 242
Level 2) for host-specificity testing of a single agent would be prohibitive. Even if there were 243
such a facility, in case of escape, the biological control agent would find an environment very 244
similar to its native range with its natural host T. aurantiaca in abundance. The benefits of 245
this option would include easy access to the natural host for the parasitoid colony and no 246
further travel if and when a release permit is obtained. 247
Option 2. Testing in containment on mainland Australia 248
While this option is more attractive than Option 1 above, it is still somewhat risky, and labour 249
and cost intensive. It brings few benefits. The biological control agent (parasitoid) would have 250
to be brought into containment along with its natural host (Tachardina aurantiaca). This 251
would pose the risk of not only a parasitoid escaping but also a potentially invasive, host-252
generalist scale insect (on Christmas Island, for example, at least 15 horticultural species are 253
attacked by T. aurantiaca, including three species of Citrus, Macadamia, Guava, Pomegranate, 254
Chili, Eggplant, Star fruit, and Soursop)(R.W. Pemberton and D.J. O’Dowd, unpublished 255
results). Using a containment facility in the temperate region of Australia could decrease 256
these risks. In case of quality control issues of the biological control agent due to ‘lab 257
selection,’ additional travel, collection, and importation of the biological control agent would 258

8
be necessary, further increasing costs. Natural host colony loss (which can easily happen due 259
to overexposure to the parasitoid agent, fungal infections, etc.) would necessitate multiple 260
importations of the natural host, which would once again significantly increase costs and risk. 261
Option 3. Testing in the native geographical range of the biological control agent with no 262
containment necessary 263
In our case, this option is the most attractive, most cost-effective, and least risky. There are 264
many benefits to study biological control agents and carry out host specificity studies in the 265
native geographical range of the agent. In our case, both the parasitoid and its natural host is 266
readily available from the wild in Malaysia which ensures good quality parasitoid and host 267
cultures. The risk factor would be zero as there would be no importation of any organisms. 268
Although obviously desirable, such studies in the native range are not common, in part 269
because of the lack of local research facilities, lack of skilled and reliable cooperators, or 270
sufficient time in what are sometimes difficult locations (Van Driesche et al. 2008). However, 271
scientific work is cost-effective in Malaysia compared to Options 1 and 2 and our network of 272
collaborators is well established (see Table B1, O’Dowd et al. 2012). Options 1 (test in 273
containment on Christmas Island) and 2 (test in containment on the Australian mainland) 274
offer no benefits over this option. We therefore suggest that host specificity testing is carried 275
out in the native range of the biological control agent. This option carries the least risk and is 276
consistent with Recommendation 2 in a review of biosecurity risks in biological control 277
commissioned by the Australian Government (Ferrar et al. 2004) that biological control 278
practitioners undertake host specificity testing in the native region before any importation. 279
280
References 281
Abbott, K.L. 2004. Alien ant invasion on Christmas Island, Indian Ocean: the role of ant-scale 282
associations in the dynamics of supercolonies of the yellow crazy ant, Anoplolepis gracilipes. 283
PhD Thesis, Monash University, Melbourne, Australia. 284
Abbott, K. and P.T. Green. 2007. Collapse of an ant-scale mutualism in a rainforest on 285
Christmas Island. Oikos 116: 1238-1246. 286
Bellis, G.A., J.F. Donaldson, M. Carver, D.L. Hancock, and M.J. Fletcher. 2004. Records of insect 287
pests on Christmas Island and the Cocos (Keeling) Islands, Indian Ocean. Australian 288
Entomologist 31: 93-102. 289
Ben-Dov, Y. 2012. Scalenet (http://www.sel.barc.usda. gov/scalenet/scalenet.htm). 290
Campbell, T.G. 1968. Entomological survey of Christmas Island (Indian Ocean) with special 291
reference to the insects of medical, veterinary, agricultural and forestry significance. 292
Unpublished Report, CSIRO Division of Entomology. 48 pp. 293

9
Cryan, R.C. and J.M. Urban. 2012. Higher-level phylogeny of the insect order Hemiptera: is 294
Auchenorrhyncha really paraphyletic? Systematic Entomology 37: 7-21. 295
CSIRO. 1990. CSIRO Entomological survey of Christmas Island. Phase 2. Unpublished Report. 296
Australian National Parks and Wildlife Service Consultancy Agreement. 67 pp. 297
Ferrar, P., I.W. Forno, and A.L. Yen. 2004. Report of the Review of the Management of 298
Biosecurity Risks Associated with the Importation and Release of Biological Control Agents. 299
Australian Government Department of Agriculture, Fisheries and Forestry, Canberra, 300
Australia. 27 pp. 301
Green, P.T., D.J. O’Dowd, G. Neumann, and S. Wittman. 2013. Research and development of 302
biological for scale insects: indirect control of the yellow crazy ant (Anoplolepis gracilipes) on 303
Christmas Island, 2009-2013. Final report to the Director of National Parks, Parks Australia, 304
Canberra, A.C.T. 66 pp. 305
Gullan, P. J. and L. G. Cook. 2007. Phylogeny and higher classification of the scale insects 306
(Hemiptera: Sternorrhyncha: Coccoidea). Zootaxa 1668: 413-425. 307
Göllner-Scheiding, U. 1980. Revision der afrikanischen Arten sowie Bemerkungen zu weiteren 308
Arten der Gattungen Leptocoris Hahn, 1833, und Boisea Kirkaldy, 1910. Deutsche 309
Entomologische Zeitschrift, N.F. 27: 103-148. 310
Hayat, M., S. Schroer, and R.W. Pemberton. 2010. On some Encyrtidae (Hymenoptera: 311
Chalcidoidea) on lac insects (Hemiptera: Kerriidae) from Indonesia, Malaysia and Thailand. 312
Oriental Insects 44: 23-33. 313
James, D. and N. Milly. 2006. A biodiversity inventory database for Christmas Island National 314
Park. A report for the Department of Finance & Administration and Department of 315
Environment & Heritage. Director of National Parks, Australian Government, Canberra. 49 316
pages. 317
Kuhlmann, U., U. Schafner, and P. G. Mason. 2006. Selection of non-target species for host 318
specificity testing. Pages 15-37 in Bigler, F., D. Babendreier and U. Kuhlmann (eds) 319
Environmental Impact of Invertebrates for Biological Control of Arthropods: Methods and Risk 320
Assessment. CAB International, Oxford. 321
Linnavuori, R. 1975. Homoptera: Cicadellidae, Supplement. Insects of Micronesia 6. No. 9. 322
pp. 611-632 323
Neumann, G., P. A. Follett, R. G. Hollingsworth, and J. de León. 2010. High host specificity in 324
Encarsia diaspidicola (Hymenoptera: Aphelinidae), a biological control candidate against the 325
white peach scale in Hawaii. Biological Control 54: 107-113. 326
Noyes, J.S. and M. Hayat. 1994. Oriental mealybug parasitoids of the Anagyrini (Hymenoptera: 327
Encyrtidae) viii+554pp. CAB International, Wallingford, UK. 328

10
Noyes, J.S. 2012. Universal Chalcidoidea Database. World Wide Web electronic publication. 329
http://www.nhm.ac.uk/chalcidoids 330
O’Dowd, D.J., P.T. Green, and P.S. Lake. 2003. Invasional ‘meltdown’ on an oceanic island. 331
Ecology Letters 6: 812-817. 332
Prinsloo, G.L. 1977. On the encyrtid parasitoids (Hymenoptera: Chalcidoidea) of lac insects 333
(Hemiptera: Lacciferidae) from southern Africa. Journal of the Entomological Society of South 334
Africa 40: 47-72. 335
Prinsloo, G.L. 1983. A parasitoid-host Index of Afrotropcial Encyrtidae (Hymenoptera: 336
Chalcidoidea). Entomological Memoir of the Department of Agriculture Republic of South 337
Africa no. 60. 35 pages. 338
Sands, D.P.A. and R.G. Van Driesche. 2004. Using the scientific literature to estimate the host 339
range of a biological control agent. Pages 16-23 in Van Driesche R.G. and R. Reardon (eds.). 340
Assessing host ranges for parasitoids and predators used for classical biological control: a 341
guide to pest practice. University States Department of Agriculture Forest Health Technology 342
Enterprise Team, Morgantown, West Virginia USA. FHTET-2004-3. 343
Van Driesche, R., T. Center, M. Hoddle, and N. Mills. 2008. Can efficacy of new biological 344
control agents be predicted before their release? Pp. 1-13 in Mason, P.G., D.R. Gillespie, and 345
C. Vincent (eds.). Proceedings of the Third International Symposium on Biological Control of 346
Arthropods. Christchurch, New Zealand, 8-13 February 2008. United States Department of 347
Agriculture, Forest Service, Morgantown, WV, FHTET-2008-06, December, 2008, 636 p. 348
Wapshere, A.J. 1974. A strategy for evaluating the safety of organisms for biological weed 349
control. Annals of Applied Biology 77: 201-211. 350
Woods, B. and E. Steiner. 2012. Christmas Island fruit fly and scale survey. Report to 351
Department of Agriculture and Food, Government of Western Australia. 22 pp. 352
353

11
Table 1. Records of known host families and genera for the primary parasitoid 354
Tachardiaephagus (Encyrtidae). Taxonomy follows Prinsloo (1977). The biological control 355
agent under investigation, Tachardiaephagus somervillei, is in bold. As a genus, 356
Tachardiaephagus has an extremely broad geographic range. With the exception of one 357
probably erroneous host record in Africa (Prinsloo 1983), all Tachardiaephagus species appear 358
to be family specialists and restricted to the Kerriidae. For host genera, number of species 359
recorded as hosts is in parentheses. Based on Noyes (2012, Universal Chalcidoidea Database, 360
http://www.nhm.ac.uk/research-curation/research/projects/chalcidoids/database/), except 361
for records for T. somervillei and T. sarawakensis (Hayat et al. 2010; Green et al. 2012; R.W. 362
Pemberton, pers. comm.) 363
Parasitoid species Distribution Recorded hosts (all Kerriidae)
Tachardiaephagus
somervillei
India, Malaysia, Thailand Kerria spp. (4)1
Tachardina aurantiaca
Tachardina sp.2
T. sarawakensis Sarawak (East Malaysia) Tachardina aurantiaca
T. tachardiae Brunei, China, India, Indonesia,
Malaysia, Sri Lanka, Taiwan,
Vietnam, Azerbaijan
Kerria spp. (8)
Paratachardina lobata
T. similis Afrotropical, South Africa Tachardina sp. (1)
T. absonus Afrotropical, South Africa Tachardina spp. (2)
T. communis Afrotropical, South Africa Tachardina spp. (5)
T. gracilis Afrotropical, South Africa Tachardina sp. (1)
1In Noyes (2012) both Kerria and Laccifer species are listed as hosts. However, Scalenet (Ben-364
Dov et al. 2012) indicates that Laccifer is a synonym for Kerria, so we have synonymized these 365
records with Kerria species. 366
2 Probably T. aurantiaca, since it is the only known Tachardina species in Asia. 367

12
Table 2. Scale insects of Christmas Island. It is highly probable that all of these species, with broad host
plant ranges and geographic distributions, are non-native to Christmas Island and introduced following
human settlement. The target species, Tachardina aurantiaca, for biological control is in bold. Families
are arranged in increasing phylogenetic distance from the Kerriidae based on Gullan and Cook (2007).
All scale insect taxa are 'neococcids' except for Icerya purchasi ('archeococcid'). Taxonomy and
distributions from Ben-Dov et al. (2012), http://www.sel.barc.usda. gov/scalenet/scalenet.htm).
Family and Species1
Common Name Distribution
Kerriidae (lac scales) Paratachardina pseudolobata (Kondo & Gullan) Tachardina aurantiaca (Cockerell)
False lobate lac scale Yellow lac scale
Oriental, Nearctic, Neotropical Oriental
Coccidae (soft scales) Ceroplastes ceriferus (Fabricius) C. destructor Newstead Coccus celatus De Lotto C. hesperidium Linnaeus Milviscutulus mangiferae (Green) Parasaissetia nigra (Nietner)
Pulvinaria urbicola Cockerell P. psidii Maskell2 S. coffeae (Walker) Saissetia oleae (Olivier)
Indian wax scale White wax scale Green coffee scale Brown soft scale Mango shield scale Nigra scale Urbicola soft scale Green shield scale Black olive scale Hemispherical scale
Cosmopolitan Afrotropical, Australasia, Oriental Afrotropical, Australasia, Oriental Cosmopolitan Cosmopolitan Cosmopolitan Pantropical Cosmopolitan Pantropical Cosmopolitan
Diaspididae (armoured scales) Aspidiotus destructor (Signoret) Hemiberlesia palmae (Cockerell) Ischnaspis longirostris (Signoret) 2
Lindingaspis sp. Pseudaulacaspis pentagona (Targioni Tozzetti) Unaspis citri (Comstock)
Coconut scale Tropical palm scale Black thread scale -- White peach scale White louse scale
Cosmopolitan Cosmopolitan Cosmopolitan -- Cosmopolitan Cosmopolitan
Cerococcidae (ornate pit scales) Cerococcus indicus (Maskell)
Spiny brown coccid
Cosmopolitan
Pseudococcidae (mealybugs) Dysmicoccus finitimus Williams
Asian coconut mealybug
Australasia, Oriental
Ferrisia virgata (Cockerell) Nipaecoccus viridis (Newstead) Pseudococcus longispinus (Targioni Tozzetti)
Monophlebidae (giant scales) Icerya purchasi (Maskell)
Striped mealybug Spherical mealy bug Long-tailed mealy bug
Cottony cushion scale
Cosmopolitan Cosmopolitan Cosmopolitan
Cosmopolitan
1Records from Campbell (1968), CSIRO (1999), O'Dowd et al. (2003), Bellis et al. (2004), Abbott (2004), Woods and
Steiner (2012) and Neumann et al. (unpubl. results); 2Tentative identifications

13
Table 3. Endemic hemipteran species known from Christmas Island and primary parasitoids
(superfamily Chalcidoidea: family Encyrtidae) associated with the families represented by the
endemic species. The data were extracted from the Universal Chalcidoidea Database (Noyes
2012). The families Nogodonidae and Rhopalidae have no associated chalcidoid primary
parasitoids and therefore endemic species in these families on Christmas Island can most
likely be excluded from all further consideration. Cicadellidae and Delphacidae have the
highest diversity of chalcidoid primary parasitoids but have magnitudes lower diversity of
encyrtid primary parasitoids. These data suggest that the encyrtid primary parasitoids of
families with endemic species on Christmas Island appear to not have host range overlap with
taxa where the target lac scale is included and the host range separation is at the suborder
level suggesting very distant phylogenetic separation. During the database analysis, only
records with species-level chalcidoid identification were used. N/A indicates not applicable.
1 Xestocephalus izzardi is also reported from Palau in the western Pacific Ocean (Linnavuori 1975). Its
status as an endemic on Christmas Island is questionable. 2 Leptocoris subrufescens on Christmas Island has been classified to subspecies status (L. subrufescens
subrufescens,). Another subspecies (L. s. flava) is described from Yap, western Pacific Ocean (Göllner-
Scheiding 1980). More research is needed to resolve the taxonomic status of these two subspecies of
L. subrufescens.
Endemic species Family
No. chalcidoid associates of family
No. encyrtid 1o parasitoid species of
family
Suborder/Family host range of encyrtids parasitizing family
Xestocephalus Izzardi1
Cicadellidae 627 6 Auchenorrhyncha (Cicadellidae)
Oxypleura calypso
Cicadidae 35 0 N/A
Clovia eximia Cercopidae 71 4 Auchenorrhyncha (Cercopidae- Aphrophoridae)
Ugyops aristella Delphacidae 248 5 Auchenorrhyncha (Delphacidae- Cicadellidae)
Varcia flavicostalis
Salona oceanica
Nogodinidae 0 N/A N/A
Leptocoris subrufescens2
Rhopalidae 0 N/A N/A
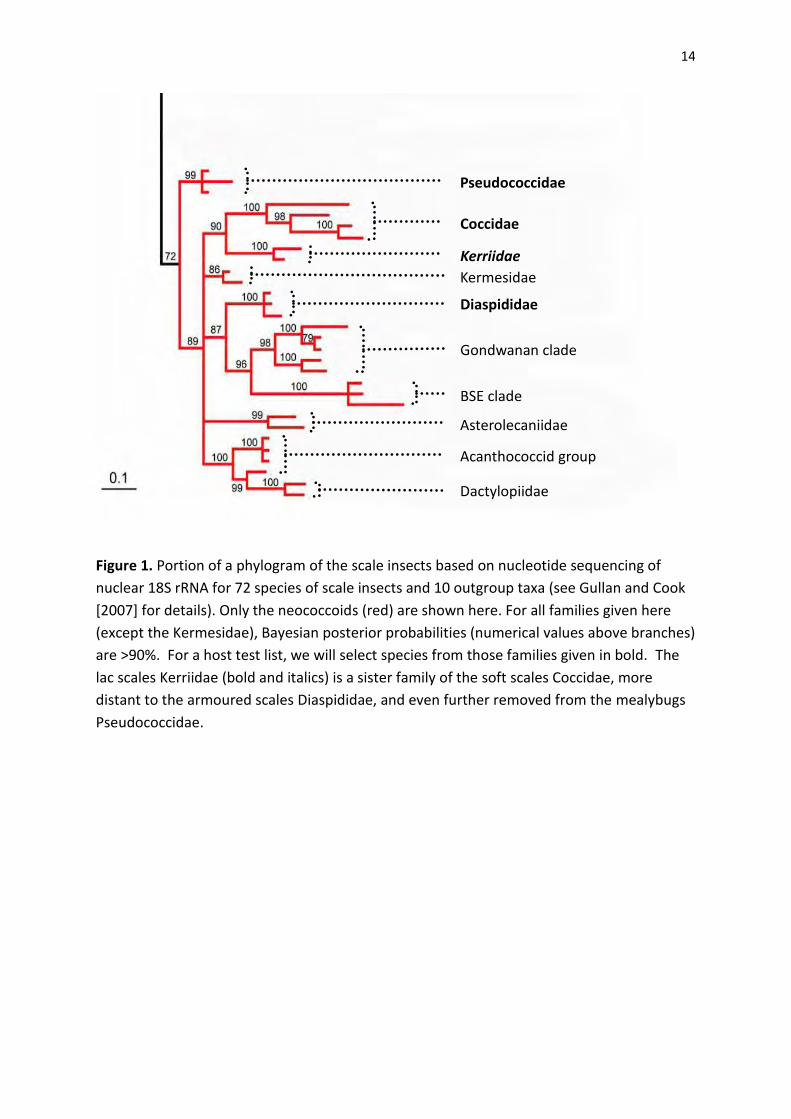
14
Figure 1. Portion of a phylogram of the scale insects based on nucleotide sequencing of
nuclear 18S rRNA for 72 species of scale insects and 10 outgroup taxa (see Gullan and Cook
[2007] for details). Only the neococcoids (red) are shown here. For all families given here
(except the Kermesidae), Bayesian posterior probabilities (numerical values above branches)
are >90%. For a host test list, we will select species from those families given in bold. The
lac scales Kerriidae (bold and italics) is a sister family of the soft scales Coccidae, more
distant to the armoured scales Diaspididae, and even further removed from the mealybugs
Pseudococcidae.
Pseudococcidae
Coccidae
Kerriidae
Kermesidae
Diaspididae
Gondwanan clade
BSE clade
Asterolecaniidae
Acanthococcid group
Dactylopiidae

Comments and suggestions for host specificity testing protocol (Neumann et al. 2013) from 5 ad hoc reviewers
Dr. Bob Pemberton (USDA-ARS, Invasive Plant Research Laboratory, Ft. Lauderdale, Florida)
Leading proponent of safety in classical biological control of weeds and insects; development of biological control for insect pests and weeds, including the lobate lac scale, foreign exploration for natural enemies) You all have done an excellent job on your testing proposal! I have attached the reviewed doc which has only a few comments.
1. More information on T. somervillei. 2. Say that it attacks only attached females not the smaller attached males. 3. Some reviewers will be unfamiliar with the basic biologies of scales and parasitoids
so it would be helpful to describe the life history of your lac scale. Then what stages the parasitoid has been reared from, and if known, what stages are attacked.
4. If it is known what host plants that Tachardina aurantiaca was on when parasitized by T. somervillei and list some.
5. Say something about the phenologies of both the scale and parasitoid to indicate that they overlap or that both appear to have continuous populations because of the moist tropical conditions. This would suggest that there should by synchrony and climatic suitability on Christmas Island.
6. Indicate that there is no parasitism of the scale on Christmas Island but that it is common or usual in its native Malaysia. Give some parasitism rates found. More needs to be said about the potential of the parasitoid and probable establishment based on similarities of the native area and Christmas Island. This information helps justify the host range testing.
7. For the use of "suitable host" some people say "suitable laboratory host" or "potential host." Could use "suitable physiological host" (note: same comment made by Barbara Barratt).
8. Although you say containment would not be needed in Malaysia, a containment level that excludes hyperparasitoids is needed.

Dr. Barbara Barratt (Principal Scientist, AgResearch Invermay, Biocontrol, Biosecurity and Bioprocessing, Mosgiel, New Zealand) Leading researcher in host specificity testing of parasitoids, risk assessment for classical biological control, biocontrol in natural areas, and developed the BIREA for New Zealand Basically looks pretty good to me but I have posed one or two questions.
1. Line 11. What damage is caused by T. aurantiaca? 2. Line 42. Clarify "actual published erroneous records." 3. Line 64. Revise subheading so as to not prejudge phylogenetic status 4. Line 71. "They would seem to be a real long-shot!" 5. Line 132. Add Wapshere 1974 as reference for centrifugal testing. 6. Line 140. Clarify - this determines physiological host range as distinct from
ecological host range. 7. Line 154. Add "and the host plant of the target" to the issue of establishment and
maintenance of colonies of the parasitoid. 8. Line 198. Rationale for not comparing parasitization rates in the control to the test
replicate. 9. Line 206. Same issue as at Line 198. 10. Line 208. Clarify reasoning here. 11. Line 292. Are coccid and diaspidid non-target species you want going to be
available in Malaysia?

Dr Matthew Purcell (Director, USDA-ARS Australian Biological Control Laboratory, Brisbane) Leading researcher in classical biological control of weeds. I’ve gone through the document a few times and it seems bullet proof to me. Given the remote location of Christmas Island to Australia, and to anywhere else for that matter, the risks are very low. Given that you have looked thoroughly into the insect fauna of the Island and that no endemic scales exist, your host testing protocol is more than adequate and fully justified.
1. It may be appropriate to mention in the Preamble that host-specificity testing is recommended to be conducted in Malaysia.
2. In section 2, I think you should make it implicit that only scales are being tested. 3. In section 2.2, it may be best to explain why it is not possible to determine the
species of scale insects to be tested at this point in time. 4. In section 2.3, 2nd paragraph, mature adult females are obviously the preferred stage
for the parasitoid to attack. It may be worth discussing here or elsewhere a little about the biology of the host and its parasite, especially since most Encyrtidae attack immatures. This would be particularly useful information when evaluating the experimental procedure.
5. In section 2.3, Evaluation 1. It would be beneficial to explain why parasitization rate in the positive control will not be compared to parasitization in rates in the test replicate. Ditto for Evaluation 3.

Dr. Don Sands OAM (retired, CSIRO, Ecosystems Sciences, Brisbane, QLD) Leading researcher in the biological control of scale insects and biological control of weeds I could add very little to your thorough testing protocol - you covered it very well. I have attached a few thoughts and comments that might be included or considered.
1. A quarantine facility on Christmas Island should be adequate for needed studies of specificity. Adequate lighting allowing sunlight to enter through double-glass will minimise artefacts in parasitoid behaviour.
2. Light and cage materials may be important when host testing and breeding parasitoids in cages. Encyrtidae often use light, certain spectra, intensity & absorption as ovipositional stimuli and for host recognition. I recommend black cages be used, as now used for rearing parasitoids by USDA and elsewhere, to avoid false positives and negatives.
3. Host specificity testing of parasitoids. Based on indigenous Hemiptera species and parasitoid host range only a small precautionary list of non-targets species would appear to require testing. As exotic scales on Christmas Island would appear to be the closest relatives to the target lac, these could be easily included in preliminary tests.
4. Many encyrtid parasitoids behave differently to their arthropod hosts feeding on different plant hosts. Tachardiaephagus could be tested with scales attached to a small range of plants, particularly native species.
5. Retain vouchers from each original shipment of wasps and also second generation of lab. – reared wasps, and have permanent preserved samples of insects released on the island.
6. Choice tests (target and non-target in same cage) can give misleading information and incorrect results from the diffusing pheromones derived from host affecting non-target host identity.
7. Nursery & mass rearing parasitoids for distribution. Once approvals for release minimise the numbers of parasitoids and number of their generations, in the confinement of quarantine, to avoid lab-adaptation or bottle-necking (can happen in 4 generations).
8. Mass rearing post approval is best done in cages outside of quarantine and with hosts + parasitoids exposed to diurnal temperature fluctuations, to enhance climate pre-adaptation and maximise numbers of parasitoids for release.
9. Release of parasitised scales on live plants (or bouquets) is more effective than the release of adult parasitoids.
10. There may be unrecognised hyper-parasitoids on Christmas Island likely to attack the Tachardiaephagus after it is introduced – worth predicting this possibility in case it influences parasitoid efficiency.
11. Release methods and localities, post release and establishment monitoring might best be outlined and methods included (& for what period) in the protocol?

Dr Tim Haye (Research Scientist, Centre for Agricultural Bioscience International [CABI], Rue des Grillons 1, CH-2800, Delemont, Switzerland) Leading researcher on the biology and use of parasitoids in classical biological control of insect pests I think the approach you suggest is well done. For us here at CABI it is a common practise to test the parasitoids in their native range. I do this for all the European insects that got introduced to North America. As lab tests often overestimate the host range of parasitioids, you may consider to look at the ecological host range (field host range) of your biocontrol agent in Malaysia. I would just take some samples from sites where maybe your target and non-targets co-occur. This would back up your lab data in case you may get non-target parasitism. The chalcidoid data base is a good tool, but the host records are often not complete, so any additional field data would make a later petition for releases of the biocontrol agent much stronger. The only thing I would hesitate about in your test design is that you are planning to test groups of parasitoids instead of individual wasps. Is there a specific reason for this? When testing groups you may have competition of parasitoids which may negatively influence your test outcome. Is there much known on the biology of the wasps (fecundity, pre-oviposition period, longevity)? All these factors may influence the test outcome, but I assume most of this is not known?

Environment Protection and Biodiversity Conservation Act 1999
Page 1 of 44
Referral of proposed action
Project title: Importation, rearing and release of Tachardiaephagus somervillei (Hymenoptera: Encyrtidae) as a biological control agent for the yellow lac scale Tachardina aurantiaca (Hemiptera: Kerriidae) on Christmas Island, Indian Ocean
1 Summary of proposed action NOTE: You must also attach a map/plan(s) showing the location and approximate boundaries of the area in which the project is to occur. Maps in A4 size are preferred. You must also attach a map(s)/plan(s) showing the location and boundaries of the project area in respect to any features identified in 3.1 & 3.2, as well as the extent of any freehold, leasehold or other tenure identified in 3.3(i). 1.1 Short description
Use 2 or 3 sentences to uniquely identify the proposed action and its location.
Supercolonies of the invasive yellow crazy ant Anoplolepis gracilipes (YCA) are a major and on-going threat to biodiversity values on Christmas Island, especially to red land crabs Gecarcoidea natalis and robber crabs Birgus latro. To date, the management of YCA supercolonies has depended on surveillance, monitoring and control using toxic bait (mostly fipronil), particularly through aerial baiting programs in 2002, 2009 and 2012. While this program has been very effective in suppressing YCA supercolonies and there are encouraging signs of recovery in many treated areas, new supercolonies continue to form. There is widespread concern for the sustainability of this program in terms of its expense, non-target impacts, and the resources it diverts from other conservation programs. Recent research conducted by La Trobe University, funded by the Director of National Parks and endorsed by the Christmas Island Crazy Ant Scientific Advisory Panel, indicates that long-term, sustainable suppression of YCA supercolonies could be achieved through the introduction of a host-specific biological control agent that would indirectly affect YCA by reducing carbohydrate supply provided by scale insects, a key resource implicated in supercolony dynamics. The yellow lac scale Tarchardina aurantiaca (Hemiptera, Kerriidae; hereafter Tachardina) is likely to be the single biggest contributor to the honeydew economy of YCA supercolonies across the island. This species, like all other scale insect species on the island, is not native to Christmas Island.
The proposed action is to import, rear and release on Christmas Island a key natural enemy of Tachardina, the parasitoid microhymenopteran wasp Tachardiaephagus somervillei (Hymenoptera, Encyrtidae; hereafter Tachardiaephagus) as a biological control agent for this scale insect. The expectation is that Tachardiaephagus should indirectly suppress supercolony formation by YCA by controlling its key scale insect mutualist.
1.2 Latitude and longitude Latitude and longitude details are used to accurately map the boundary of the proposed action. If these coordinates are inaccurate or insufficient it may delay the processing of your referral.
Latitude Longitude location point degrees minutes seconds degrees minutes seconds NE extreme -10 24 40 105 42 14 NW extreme -10 26 14 105 32 59 SW extreme -10 30 54 105 32 1.6 S extreme -10 34 41 105 39 53 E extreme -10 28 4 105 42 49

Environment Protection and Biodiversity Conservation Act 1999
Page 2 of 44
1.3 Locality and property description Provide a brief physical description of the property on which the proposed action will take place and the project location (eg. proximity to major towns, or for off-shore projects, shortest distance to mainland).
The project will be carried out on Christmas Island, an Australian External Territory located in the northeastern Indian Ocean at 10° 25’S and 105° 40’E (Fig. 1a). The island is approximately 2600 km northwest of Perth and 360 km south of the western end of Java, Indonesia. The island covers 135 km2, of which approximately 85 km2 (64%) is National Park (Fig. 1b). The remaining area is unallocated Commonwealth Crown Land (18%); mine lease (15%) (leased by Phosphate Resources Limited from the Commonwealth) and committed land, including private dwellings (3%). The Park contains two Ramsar wetlands at The Dales and Hosnie’s Spring (Fig. 1b).
Christmas Island rises from the sea in a series of cliffs and terraces to a maximum elevation of 361 meters. The terraces are separated by slopes covered by loose and jagged limestone boulders, or by sheer cliffs (Fig. 1c). The interior of the island is a slightly undulating plateau, from 160-361 metres above sea level (Fig. 1d). About 75% of the island is covered by vegetation, mostly rain forest.
Figure 1. (a) Geographic location of Christmas Island in the north eastern Indian Ocean, (b) Christmas Island showing the National Park (green), unallocated crown land (yellow), Ramsar sites (Blue hatching) and phosphate mining leases (green stippling). The National Park and Unallocated Crown Land together show the extent of remaining rainforest on Christmas Island, (c) Christmas Island rises in a series of cliff and terraces to a maximum elevation of 361 m (d) the interior of the Island is a slightly undulating plateau, from 160-361 m above sea level.
(a) (b)
(c) (d)

Environment Protection and Biodiversity Conservation Act 1999
Page 3 of 44
1.4 Size of the development footprint or work area (hectares)
YCA supercolonies have formed patchily in rainforest across the entire Island (Fig. 2). The intention is for the biological control agent to both control existing supercolonies and suppress the formation of new ones, by eventually colonising and establishing self-sustaining populations across the entire forested area from an initially small number of release points. Thus, the proposed action is forecast to occur over the entire forested area of the Christmas Island National Park and adjacent unallocated Crown Land, totalling c. 10,000 ha.
Figure 2. Composite map showing YCA Supercolonies treated with toxic bait between 2000-2012. This map effectively shows the extent of supercolony formation over the last twelve years.
1.5 Street address of the site
Christmas Island National Park, PO Box 867, Drumsite, Christmas Island, Indian Ocean, 6798.
Unallocated Crown Land, managed by the Commonwealth Department of Department of Regional Australia, Local Government and Sports (DRALGAS).
1.6 Lot description Describe the lot numbers and title description, if known.
N/A
1.7 Local Government Area and Council contact (if known) If the project is subject to local government planning approval, provide the name of the relevant council contact officer.
Although local government approval is not required for the proposed action, consultation will still occur. The relevant local government is the Shire of Christmas Island – Western Australia Local Government Association. CEO Kelvin Mathews.

Environment Protection and Biodiversity Conservation Act 1999
Page 4 of 44
1.8 Time frame Specify the time frame in which the action will be taken including the estimated start date of construction/operation. Foreign exploration in Southeast Asia has already located Tachardina at sites across 1900 km in Malaysia, within its native range in Southeast Asia. Research on the biology of its main parasitoid Tachardiaephagus is already underway in Southeast Asia, principally at the Forest Research Institute of Malaysia in Kuala Lumpur and at Sarawak Forestry in Semenggoh, Sarawak (see O’Dowd et al. 2012, and Section 2.1 below). A preliminary draft for host specificity testing has been produced (see Attachment 1) and host specificity testing could be completed in Malaysia by June 2013. Coincident with these activities, rearing facilities for Tachardiaephagus will be constructed on Christmas Island (see Attachment 2). Tachardiaephagus will be imported to Christmas Island when both tasks are completed, and a population will be established under controlled laboratory conditions in the rearing facility. The first field releases are planned for mid-2014, and there will be multiple releases thereafter through 2015 and 2016.
1.9 Alternatives to proposed action Were any feasible alternatives to taking the proposed action (including not taking the action) considered but are not proposed?
No
Yes, you must also complete section 2.2
1.10 Alternative time frames etc Does the proposed action include alternative time frames, locations or activities?
No
Yes, you must also complete Section 2.3. For each alternative, location, time frame, or activity identified, you must also complete details in Sections 1.2-1.9, 2.4-2.7 and 3.3 (where relevant).
1.11 State assessment Is the action subject to a state or territory environmental impact assessment?
No
Yes, you must also complete Section 2.5 NOTE: As described in Section 2.4, the regulatory framework under which the introduction of a biological control agent to the external Australian territory of Christmas Island would be considered cannot yet be clearly identified. Therefore, the requirement for a formal state or territory environmental impact assessment is also unclear. The matters of greatest relevance in the assessment of this Referral are the host specificity of the proposed biological control agent, and the likelihood of off-target impacts on other scale insects or species of concern. These issues are addressed in Section 2.5 in lieu of a formal environmental impact assessment.
1.12 Component of larger action Is the proposed action a component of a larger action?
No
Yes, you must also complete Section 2.7
1.13 Related actions/proposals Is the proposed action related to other actions or proposals in the region (if known)?
No
Yes, provide details:

Environment Protection and Biodiversity Conservation Act 1999
Page 5 of 44
1.14 Australian Government funding Has the person proposing to take the action received any Australian Government grant funding to undertake this project?
No
Yes, provide details: The research supporting the proposed action is being funded by DSEWPaC though the Director of National Parks to La Trobe University. One postdoctoral fellow has just finished a three-year project establishing the importance of liquid carbohydrate resources provided by scale insects for YCA supercolony formation (November 2009 – November 2012), while a second postdoctoral fellow has been working on research and survey for prospective biological control agents of Tachardina (June 2010 – June 2013). This postdoctoral researcher has spent considerable time in Southeast Asia. It is intended that DSEWPaC continue to fund La Trobe University through the host-specificity testing phase in Kuala Lumpur, and on to the importation, rearing and release phases on Christmas Island (July 2013 – July 2016).
1.15 Great Barrier Reef Marine Park Is the proposed action inside the Great Barrier Reef Marine Park?
No Yes, you must also complete Section 3.1 (h), 3.2 (e)
2 Detailed description of proposed action NOTE: It is important that the description is complete and includes all components and activities associated with the action. If certain related components are not intended to be included within the scope of the referral, this should be clearly explained in section 2.7. 2.1 Description of proposed action This should be a detailed description outlining all activities and aspects of the proposed action and should reference figures and/or attachments, as appropriate.
Background – Biology and Impacts. Anoplolepis gracilipes (the yellow crazy ant, YCA) is a ‘tramp’ species that has become invasive throughout the tropics (Wetterer 2005). The YCA is listed by the IUCN as one of the world’s 100 worst invasive species (Lowe et al. 2000), and was accidentally introduced to Christmas Island between 1915 and 1934 (O’Dowd et al. 1999). Because of its negative impacts on species, interactions and ecosystem processes, YCA is recognised as the most significant and pervasive threatening process affecting biodiversity on Christmas Island, reflected by the listing of the Loss of biodiversity and ecosystem integrity following invasion by the Yellow Crazy
Ant on Christmas Island, Indian Ocean as a Key Threatening Process under the EPBC Act 1999, and as identified in Threat abatement plan to reduce the impacts of tramp ants on biodiversity in Australia
and its territories (Commonwealth of Australia 2006). The control of this ant features prominently in many Recovery Plans for EPBC-Listed species on Christmas Island, and has been the focus of natural resource management activities on the island for more than a decade at the cost of millions of dollars.
The attribute that makes the YCA so problematic on Christmas Island and elsewhere is its capacity to form high-density, expansive ‘supercolonies’. Many tramp ant species form supercolonies, often defined using a combination of criteria including genetic relatedness,intraspecific behavioural interactions, and ant abundance. For example, the invasive argentine ant Linepithema humile is genetically uniform across most of its European range and is regarded as a single supercolony (Giraud et al. 2002), while intercontinental behavioural assays and analyses of cuticular hydrocarbons suggest this species may form a single supercolony spanning Australia, Europe, North

Environment Protection and Biodiversity Conservation Act 1999
Page 6 of 44
America and Japan (Sunamura et al. 2009; Suhr et al. 2010). Although two distinct genotypes of YCA occur on Christmas Island, these co-occur at very small spatial scales (Thomas et al. 2010) and behavioural assays pairing individual ants from opposite ends of the island suggest that the population on Christmas Island behaves as a single supercolony (Abbott 2005). Nevertheless, YCA supercolonies on Christmas Island have always been defined in terms of very high ant densities. YCA occurs at many locations across the island in very low abundance with no obvious impact on biodiversity, but in 1989, and then again in late 1997, YCA was discovered in several locations at extremely high densities sufficient to extirpate local populations of the abundant red land crab Gecarcoidea natalis (O’Dowd et al. 1999, 2003). The red crab is a keystone species in rainforest on the island that controls patterns seedling recruitment and rates of litter breakdown and nutrient cycling (Green et al. 1997, 2008). Since then, the density at which YCA kills land crabs leading to understorey transformation has become the operational definition of ‘supercolony’ on Christmas Island. On the ground, the density of YCA in supercolonies can be extraordinary – up to several thousand per square meter (Abbott 2005). A systematic, island-wide survey in 2001 found multiple supercolonies ranging in area from tens to hundreds of hectares, totalling c. 2500 ha, or 25% of rainforest on the island (Green et al. 2004; Green and O’Dowd 2009). Since 2001, supercolonies have continued to form and/or reform and upwards of 5000 ha of rainforest have been treated with toxic bait to 2012.
The key trait that has allowed YCA to form high-density supercolonies is its ability to form mutualistic associations with honeydew-producing hemipterans, principally scale insects (O’Dowd et al. 2003; Abbott and Green 2007). The scale insects suck sap from trees and secrete carbohydrate-rich honeydew on which the ants feed. The ants provide sanitation services for the scale insects, removing honeydew that might otherwise build up and kill them either through asphyxiation or the growth of sooty moulds, but the ants may also provide limited protection for the scale insects from generalist natural enemies. Supercolony-level densities of YCA and outbreak-densities of several species of scale insects invariably co-occur, and in supercolonies high densities of ants can typically be seen ascending the boles of most trees to visit scale insects in the canopy. The gasters of descending ants are swollen with carbohydrate-rich honeydew that they will take back to the nest to be shared with non-forager conspecifics. Site-scale, manipulative experiments on Christmas Island have demonstrated a bi-directional, causal link between co-occuring high densities and ants and scale – the exclusion of ants using toxic bait leads to a dramatic decline in scale abundance (Abbott and Green 2007), while the prevention of access by ants to scale insects using tree bands leads to a dramatic decline in ant density (O’Dowd et al. 2012). The tree banding experiment especially is important as “proof of concept” that removing a key food resource for ants could lead to supercolony suppression. On Christmas Island YCA interacts with several species of honeydew-secreting scale insects including Tachardina aurantiaca, the principal target of the biocontrol program (see below).
The mutualism between YCA and scale insects has manifold negative impacts on species abundances, interactions among species, and forest structure. At the core of these impacts is the devastating effect of YCA on land crabs, especially red crabs. YCA sprays formic acid as a weapon both to subdue prey and in self-defence, and although the amount sprayed by individual ants is tiny, at supercolony densities the overall effect is overwhelming. YCA supercolonies reduce formerly high densities of red crabs (averaging c. 0.5 – 1.0 crabs m-2) to nil, deregulating seedling recruitment and litter dynamics and resulting in a thick, diverse understorey of seedlings and saplings with an almost permanent layer of leaf litter (O’Dowd et al. 2003). In forest dominated by red crabs, the understorey is sparse and dominated by a few crab-resistant species, and the forest floor is almost devoid of litter for much of the year (Green et al. 1997, 2008). These impacts are widespread. Based on the spatial extent of supercolony formation over the last 12 years, it is likely that YCA has extirpated at least 20 million red land crabs, or about 30% of the total population in areas where they have formed supercolonies.

Environment Protection and Biodiversity Conservation Act 1999
Page 7 of 44
YCA has also caused declines in the density of red land crabs at sites where high-density supercolonies have never formed. About half of the red crab population migrates to the coast each year to complete breeding activities, and many YCA supercolonies have formed across the crabs’ traditional migration routes. Thus, significant numbers of migrating red crabs have been killed en
route to the coast over many years, never to return to the former home ranges. As a result, some areas of rainforest are practically devoid of red crabs even though YCA supercolonies have never occurred there, and the same processes of understorey transformation are in train there too. It is hard to gauge the severity and extent of this “ghosting” effect because pre-YCA invasion data on red crab densities are sparse, but is likely to be significant; Green et al. (2011) estimated that around 25% of rainforest may have been ghosted at some time in the last decade. The direct and indirect (ghosting) impacts of YCA supercolony formation have been so widespread since the late 1990s that just 28% of rainforest could still be considered as “intact” (no YCA supercolony formation, and unaffected by ghosting) by 2007 (Green et al. 2011).
In YCA supercolonies, scale insects themselves can have large negative impacts on their host plants. Especially vulnerable is the Tahitian Chestnut Inocarpus fagifer, a widespread canopy tree that hosts very high densities of Tachardina on its outer twigs and leaves. In supercolonies, seedlings, saplings and small trees all suffer extremely high mortality, and the canopies of large trees are much reduced through the dieback of fine twigs and branches (Green et al. 2001, O’Dowd et al. 2003, P. Green and D. O’Dowd, unpublished results). There is also evidence that fruit production is reduced in supercolonies. Excess honeydew that YCA does not harvest settles on leaves of all plant species and is colonised by sooty moulds, which probably interferes with photosynthesis and growth.
YCA may affect many of the island’s bird species through direct interference and through altered resource availability and habitat structure (Davis et al. 2008, Davis et al. 2009). The Christmas Island Emerald Dove Chalcophaps indica natalis is 9-14 times less abundant in ant-invaded forest, and because it forages on the forest floor, is probably vulnerable to direct predation by YCA. The nesting success and density of juvenile Christmas Island Thrushes Turdus poliocephalus erythropleurus is lower in supercolonies, where they also show altered foraging and reproductive behaviours. Furthermore, these birds alter their choice of tree species in which to build nests, with lower frequency on tree species that typically experience high densities of scale insects and ants. The density of foraging Christmas Island white-eye Zosterops natalis is higher in supercolonies, perhaps because scale insects (as prey) are more common there. It is possible that impacts of YCA on thrushes and white-eyes affect frugivory and seed dispersal on the island; assays with both real and model fruit showed handling rates to be more than two-fold lower in YCA supercolonies, and manipulative experiments showed this to be a direct consequence of the presence of ants. There is no evidence that YCA supercolony formation significantly affects the density of nesting success of Abbott’s Booby Papasula abbotti (P. Green, unpublished data), while the impact of YCA on other seabirds and on other land birds such as the goshawk and owl are unknown.
Several endemic vertebrate species, including the pipistrelle bat Pipistrellus murrayi and endemic reptiles have experienced precipitous declines over recent years, but the causes of the declines are opaque. In the case of the pipistrelle it is possible that YCA supercolony formation has contributed to the decline, either directly through predation of bats at roost sites or indirectly by eliminating red crabs and facilitating the expansion of predators such as giant centipedes Scolopendra subspinipes, wolf snakes Lycodon aulicus, cats and rats (Schulz and Lumsden 2004, Lumsden et al. 2007, Beeton et al. 2010). However, the decline of the pipistrelle was well in train before the rise of YCA supercolonies in the late 1990s. The endemic reptiles were similarly in decline long before YCA supercolonies became common, and the role of YCA in their decline is also uncertain (Smith et al. 2012).

Environment Protection and Biodiversity Conservation Act 1999
Page 8 of 44
In addition to impacts on species of concern, supercolony formation by YCA has also led to an altered web of species interactions that facilitates the entry and spread of other invasive species. The best example of this is the entry and spread of the giant African landsnail Achatina fulica (GALS) in rainforest on the island. GALS was introduced to the island in the 1940s, and despite being a notoriously invasive species (Lowe et al. 2000), its distribution was for many decades limited to settled areas, abandoned mining fields and roadsides. Experiments showed that predation by red crabs excluded this invader from establishing in rainforest (Lake and O’Dowd 1991), but the extirpation of red crabs in YCA supercolonies, coupled with the ability of GALS to coexist alongside YCA in supercolonies, has permitted this snail to establish high densities in rainforest at many locations across the island (Green et al. 2011). The facilitation of GALS by YCA could be due to the creation of enemy-free space, augmented understorey resources, or both.
The rise of YCA supercolonies and the extirpation of red crabs have also affected the invasion dynamics of other non-native organisms. These effects encompass both inhibition and facilitation for a range of non-native ant and snail species (O’Dowd and Green 2010; P. Green and L. O’Loughlin, unpublished results), while invasion by several weeds including Capsicum frutescens, Carica
papaya, Cordia curassavica, and Muntingia calabura appears to be facilitated in areas affected by YCA supercolony formation (P. Green and D. O’Dowd, personal observations).
Background – Management. Given all the above, supercolony formation by YCA is considered a major and on-going threat to biodiversity values on Christmas Island. To date the management of YCA has depended on surveillance, monitoring, and control using toxic bait (Green and O’Dowd 2009, Boland et al. 2011). New supercolonies continue to form, and there is concern for the sustainability of this program in terms of its expense, non-target impacts, and the resources it diverts from other conservation programs. Further, this program can only ever be reactive, and it has not been able to find an effective solution to the difficult issue of the management of incipient supercolonies at an islandwide spatial scale (see section 2.2, below). There is widespread agreement that the development of a more cost-effective, sustainable alternative to the use of toxic bait is needed to manage the YCA invasion on Christmas Island.
For the last three years, La Trobe University researchers have been investigating the potential for supercolony suppression by the biocontrol of scale insects. These researchers presented a summary of their findings to the Crazy Ant Scientific Advisory Panel (CASAP), and independent group of scientists who provide advice to the Director of National Parks and Parks Australia on matters pertaining to the management of the YCA invasion. CASAP advice was formally sought on the veracity of the research findings, and on the proposal to import, rear and release a biological control agent(s) as an option for controlling YCA supercolony formation and spread. At a meeting in December 2012, CASAP assessed the scientific merit of the research conducted by La Trobe University as well as the feasibility and risks of introducing a biological agent to Christmas Island. CASAP members were convinced that the option of biological control was a viable and feasible option for controlling YCA, the risks to the island’s biodiversity were very low, and the risk of doing nothing outweighed the risk posed by the importation and release of a biological control agent. CASAP advised the DNP to proceed with the implementation phase of the biological control research (see Attachment 3).
Biological Control. Classical biological control works on the principle that in their area of origin, populations of native species are kept in check by their natural enemies (predators, parasites or pathogens). There is a large body of literature demonstrating that in many cases, species introduced outside of their native ranges become invasive because they have effectively left their enemies behind. This is known as the “Enemies Release Hypothesis”. The principle of classical biological control then is the converse: to re-establish population control over invasive species by first identifying and then importing a natural enemy – a biological control agent – from within the native

Environment Protection and Biodiversity Conservation Act 1999
Page 9 of 44
range of the target organism. In all instances, this now involves selection and testing of biological control agents to verify narrow host ranges that minimize the risk of non-target impacts in the area of introduction.
Indirect Biocontrol for YCA on Christmas Island. Despite the diversity and significant ecological and economic impacts of invasive ants worldwide, they have proved to be an especially difficult target group for biological control. A program for the biocontrol of the Red Imported Fire Ant (Solenopsis invicta) using a parasitic fly and a protozoan disease as agents is currently under development in the southeastern United States, but no species of ants have yet been controlled in the field using biological control agents and principles. Instead, a novel solution for managing the YCA invasion is proposed; rather than targeting the ant itself, a key mutualist species that plays a significant role in sustaining YCA supercolonies at very high and ecologically damaging densities will be targeted instead. Long-term, sustainable suppression of YCA supercolonies could be achieved through the introduction of a biological control agent that would indirectly affect YCA by reducing carbohydrate supply provided by honeydew-producing scale insects, a key resource implicated in supercolony dynamics.
The Proposed Action. While several species of honeydew-producing scale insects are common in YCA supercolonies, the yellow lac scale Tarchardina aurantiaca (Hempitera, Kerriidae) is almost certainly the single biggest contributor to the honeydew economy of YCA supercolonies across the island (O’Dowd et al. 2012). This referral is a proposal to import, rear and release on Christmas Island a key natural enemy of this scale insect, the parasitoid microhymenopteran wasp Tachardiaephagus somervillei (Hymenoptera: Encyrtidae - hereafter Tachardiaephagus) as a biological control agent for Tachardina. The expectation is that Tachardiaephagus will indirectly suppress existing YCA supercolonies and arrest formation of new supercolonies by controlling its key scale insect mutualist.
The Proposed Biological Control Agent. Tachardiaephagus somervillei (Fig. 3) is the most promising agent for introduction and release on Christmas Island. It is one of several primary parasitoids that have been found to attack yellow lac scale during studies conducted within it native distribution in Malaysia (Table 1). This parasitoid
attacks Tachardina across a 1900-km range in Peninsular Malaysia and Malaysian Borneo is the most abundant natural enemy of Tachardina has a short life cycle compared to its host exhibits superparasitism (i.e., where multiple progeny emergence from a single host
individual) causes high rates of parasitism on Tachardina in the presence of tending ants, including the
yellow crazy ant (Fig. 4) can be reared under laboratory conditions.

Environment Protection and Biodiversity Conservation Act 1999
Page 10 of 44
Figure 3. Tachardiaephagus somervillei, a primary parasite of the yellow lac scale Tachardina aurantiaca in Malaysia and Singapore (drawing from Narayanan 1962. Pests of lac in India. Pp. 90-113 in Mukhopadhyay, B and Muthana, MS (eds) A Monograph on Lac. Indian Lac Res. Institut. Nancum, Ranchi, India). A Master’s student at Universiti Sains in Malaysia has also successfully reared T. somervillei under laboratory conditions.
Figure 4. Parasitism of the yellow lac scale Tachardina aurantiaca in Malaysia. The image shows emergence holes of the parasitoid Tachardiaephagus sp. (Encyrtidae) in tests of an aggregate of old adult females of Tarchardina near Sandakan, Sabah, Malaysian Borneo. Yellow crazy ants (Anoplolepis gracilipes) tended Tachardina at this site. Superparasitism (i.e., where more than one progeny is produced per host individual) is frequent. The mean number of Tachardiaephagus somervillei emerging from each parasitized female Tachardina was 2.1 (range 1- 4, N = 30) and up to 5 emergences occurred from each female in Peninsular Malaysia. In contrast, parasitism of Tachardina on Christmas Island was nil; emergence holes were never seen in examination of over 11,000 females over two years (or over the past 15 years of observations of Tachardina on the island).
1 mm
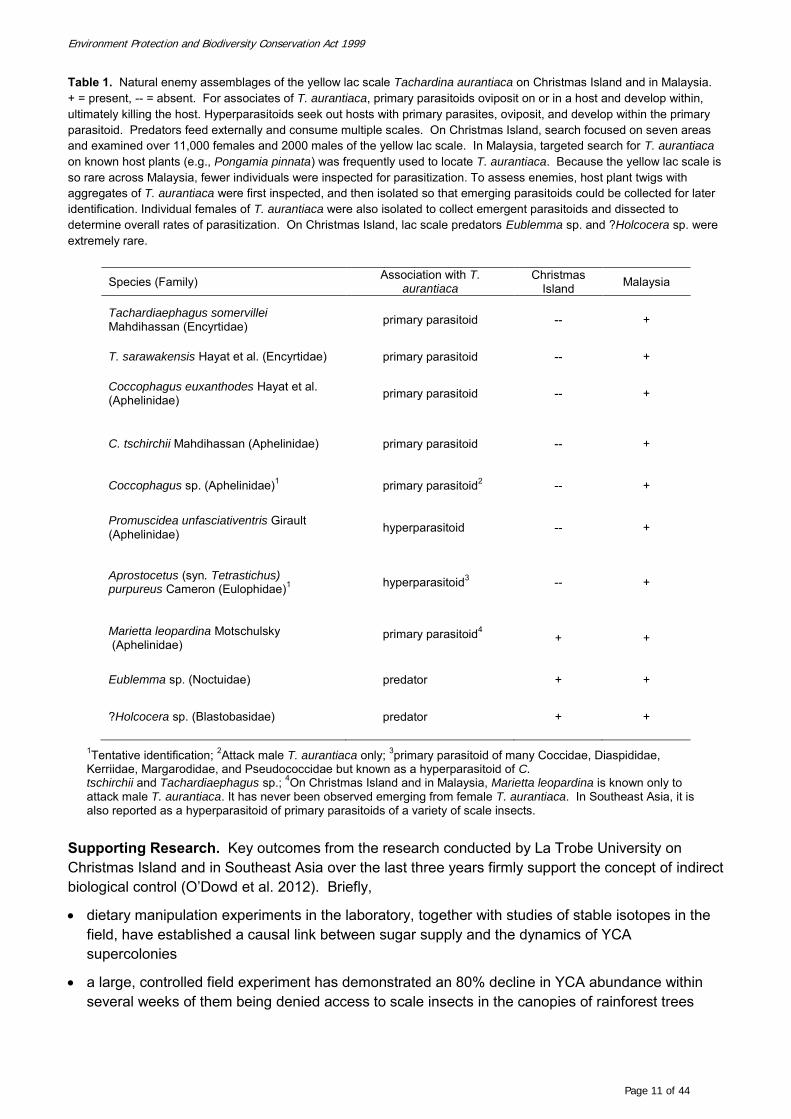
Environment Protection and Biodiversity Conservation Act 1999
Page 11 of 44
Table 1. Natural enemy assemblages of the yellow lac scale Tachardina aurantiaca on Christmas Island and in Malaysia. + = present, -- = absent. For associates of T. aurantiaca, primary parasitoids oviposit on or in a host and develop within, ultimately killing the host. Hyperparasitoids seek out hosts with primary parasites, oviposit, and develop within the primary parasitoid. Predators feed externally and consume multiple scales. On Christmas Island, search focused on seven areas and examined over 11,000 females and 2000 males of the yellow lac scale. In Malaysia, targeted search for T. aurantiaca on known host plants (e.g., Pongamia pinnata) was frequently used to locate T. aurantiaca. Because the yellow lac scale is so rare across Malaysia, fewer individuals were inspected for parasitization. To assess enemies, host plant twigs with aggregates of T. aurantiaca were first inspected, and then isolated so that emerging parasitoids could be collected for later identification. Individual females of T. aurantiaca were also isolated to collect emergent parasitoids and dissected to determine overall rates of parasitization. On Christmas Island, lac scale predators Eublemma sp. and ?Holcocera sp. were extremely rare.
Species (Family) Association with T. aurantiaca
Christmas Island Malaysia
Tachardiaephagus somervillei Mahdihassan (Encyrtidae) primary parasitoid -- +
T. sarawakensis Hayat et al. (Encyrtidae) primary parasitoid -- +
Coccophagus euxanthodes Hayat et al. (Aphelinidae) primary parasitoid -- +
C. tschirchii Mahdihassan (Aphelinidae) primary parasitoid -- +
Coccophagus sp. (Aphelinidae)1 primary parasitoid2 -- +
Promuscidea unfasciativentris Girault (Aphelinidae) hyperparasitoid -- +
Aprostocetus (syn. Tetrastichus) purpureus Cameron (Eulophidae)1 hyperparasitoid3 -- +
Marietta leopardina Motschulsky (Aphelinidae)
primary parasitoid4 + +
Eublemma sp. (Noctuidae) predator + +
?Holcocera sp. (Blastobasidae) predator + +
1Tentative identification; 2Attack male T. aurantiaca only; 3primary parasitoid of many Coccidae, Diaspididae, Kerriidae, Margarodidae, and Pseudococcidae but known as a hyperparasitoid of C. tschirchii and Tachardiaephagus sp.; 4On Christmas Island and in Malaysia, Marietta leopardina is known only to attack male T. aurantiaca. It has never been observed emerging from female T. aurantiaca. In Southeast Asia, it is also reported as a hyperparasitoid of primary parasitoids of a variety of scale insects.
Supporting Research. Key outcomes from the research conducted by La Trobe University on Christmas Island and in Southeast Asia over the last three years firmly support the concept of indirect biological control (O’Dowd et al. 2012). Briefly,
dietary manipulation experiments in the laboratory, together with studies of stable isotopes in the field, have established a causal link between sugar supply and the dynamics of YCA supercolonies
a large, controlled field experiment has demonstrated an 80% decline in YCA abundance within several weeks of them being denied access to scale insects in the canopies of rainforest trees

Environment Protection and Biodiversity Conservation Act 1999
Page 12 of 44
while variable, the average contribution made by Tachardina to the honeydew economy of YCA supercolonies is estimated to be 70%
within its native range in Malaysia, Tachardina is rare and patchily distributed, associated with diverse natural enemies including at least five species of primary parasitoids, and suffers high parasitization rates – all attributes consistent with population regulation by natural enemies
host records of Tachardiaephagus somervillei, the most abundant parasitoid of Tachardina across its native range in Malaysia suggest that it is a specialist within the lac scale family, Kerriidae. There are no native lac scale insects on Christmas Island
Tachardiaephagus does not occur on Christmas Island and Tachardina is not currently under effective biological control from the few existing natural enemies.
there is a single ecotype of female Tachardina on Christmas Island, which has been morphologically and genetically matched to populations in Southeast Asia that show evidence of being under control from their primary parasitoids
Approval, Importation & Rearing, and Release & Monitoring of Tachardiaephagus Approval: Currently, there is some uncertainty about the appropriate regulatory framework under which a biological control agent might be considered for introduction to Christmas Island (see Section 2.4, below). However, the framework will almost certainly include host-specificity testing as a necessary precursor to release (Palmer et al. 2010). The purpose of these trials is to test the proposition that Tachardiaephagus is as narrowly specific as the host records suggest, and that the probability of non-target impacts on species of concern is extremely low.
Australia has had a long history of conducting host-specificity tests in the area of agent origin, and it has only been relatively recently that these tests have been conducted within Australia by bringing agents into quarantine facilities (Palmer et al. 2010). However, testing within the native range is still desirable because it is safe (no need to import the agent), and it also allows for the possibility of conducting field tests which are seen as more realistic than laboratory trials (Secord and Karieva 1996). In any case, there is no suitable quarantine facility on Christmas Island in which to test Tarchardiaephagus against potential local non-target species, and the cost of constructing one would be prohibitive. Alternatively, Tachardiaephagus could be imported into containment in Australia, but this poses some risk to native kerriid species in Australia, and would present formidable regulatory issues. Instead, host-specificity tests will be conducted within the native distribution of Tachardina aurantiaca at the Forest Research Institute of Malaysia, where La Trobe University has established a research laboratory with the assistance of local co-operators. It is proposed to use “centrifugal host range testing” as the guiding principle to assess the host range of the Tachardiaephagus (Kuhlmann et al 2006, Neumann et al. 2010). In this approach, species other than the known host of the biological control agent are tested with the most closely related species (least phylogenetic distance, and with close similarities in biology and ecology) tested first then less similar species thereafter (“centrifugal principle”). This is outlined in more detail in Attachment 1.
Importation & Rearing: Many parasitoids have their own insect natural enemies, known as hyperparasitoids (i.e. parasites of parasites). The importation of a hyperparasite of Tachardieaphagus is inimical to the successful suppression of YCA supercolonies, because it could compromise seriously the capacity of Tachardieaphagus to build up population densities sufficient to control Tachardina. For this reason, great care will be taken to implement standard agent rearing and sanitation techniques to ensure that the founding population of Tachardiaephagus from Malaysia is free of hyperparasitoids and pathogens before importation to Christmas Island.

Environment Protection and Biodiversity Conservation Act 1999
Page 13 of 44
Importation of Tachardiaephagus to the island should conservatively be regarded as the point of release. A production facility will be built on Christmas Island to receive the founder population, and to mass-rear these insects for field-release. This facility will be designed to contain the parasitoids through the rearing phase. The facility will produce Tachardina host plants using existing nursery facilities, and their maintenance in a dedicated glasshouse/greenhouse for the mass rearing of agents. This population will require careful monitoring and maintenance to minimize the selection of laboratory-adapted insects, and to counter the possible susceptibility of Tachardiaephagus to the loss of population heterozygosity. Sex determination in microhymenoptera is usually haplodiploid – males are haploid, females diploid, and heterozygosity at a multi-allelic sex-determining locus is required for femaleness. Inbreeding can lead to a preponderance of homozygous diploids that will either be sterile males, or experience a very high rate of mortality. The infrastructure, personnel and logistic support required for this phase of the biological control program have been outlined in Attachment 2.
Release & Monitoring: Protocols will be developed to estimate the impact of Tachardiaephagus on Tachardina and on YCA densities, and criteria will be established to select the number and location of primary release sites and control points, establish the most appropriate timing of the releases according to local site variation in Tachardina phenology, and to determine the rate of spread of Tachardiaephagus from release points. It is premature to give precise details on most aspects of the release and monitoring phase because they will largely depend on the number of insects that can be reared in the production facility, and on the extent and location of YCA supercolonies at the time. 2.2 Alternatives to taking the proposed action
This should be a detailed description outlining any feasible alternatives to taking the proposed action (including not taking the action) that were considered but are not proposed (note, this is distinct from any proposed alternatives relating to location, time frames, or activities – see section 2.3).
There are two alternatives to taking the proposed action:
1. Do not take the proposed action. Not taking the proposed action will ensure the management of YCA supercolonies and the amelioration of their impacts will continue to rely on chemical control, mainly the toxicant fipronil. However, the use of fipronil is perpetually reactive, because it relies on continual surveillance, monitoring and control using toxic bait and considerable human resources to keep abreast of supercolony formation. Further, it has not been able to deal with incipient supercolonies where YCA densities are increasing but still not high enough for YCA to monopolise the bait, and thereby minimise non-target impacts. Despite no evidence for the bioaccumulation of fipronil or its metabolites in baited areas (CESAR Consultants 2011), there is still disquiet among all the stakeholders over its continued use but a grudging acceptance of the lack of alternatives (Beeton et al. 2010). For all of these reasons, business-as-usual baiting with fipronil is seen as unsustainable in the long term and is now being considered as a transitional measure until biocontrol can be developed as an alternative (EPBC Referral, Helicopter baiting of
the exotic yellow crazy Anoplolepis gracilipes supercolonies on Christmas Island, Indian Ocean, 2012).
2. Develop a biocontrol program for several species of soft scales common in YCA supercolonies, using parasitoids already present on Christmas Island. Coccophagus
ceroplastae (Hymenoptera: Aphelinidae) and Encyrtus infelix (Hymenoptera: Encyrtidae), both parasitoids of a variety of coccid scale insects, have been discovered on Christmas Island as part of the supporting research leading up to this referral (O’Dowd et al. 2012). Both were almost certainly introduced inadvertently to the island in the past with importation of plant material with host scale insects. Both species attack a wide variety of soft scales, including four that are important in YCA supercolonies: Coccus hesperidium, C. celatus, Saissetia oleae and S. coffeae.

Environment Protection and Biodiversity Conservation Act 1999
Page 14 of 44
The difficult issues of foreign exploration, host-specificity testing and navigating regulatory frameworks have been obviated by the presence of these parasitoids on the Island. Experience with the efficacy of these agents in dealing with the outbreak of Pulvinaria urbicola on Christmas Island (Neumann et al. 2011) suggests that the current lack of control of soft scales in YCA supercolonies is a result of dispersal limitation of their parasitoids. Dispersal limitation could presumably be overcome by releasing these parasitoids at multiple sites and at multiple times across the island. However, soft scales are estimated to contribute on average just 30% to the honeydew economy of YCA supercolonies, so targeting them alone is less likely to achieve suppression of YCA supercolonies than targeting Tachardina. For this reason, the development of biological control for soft scales using parasitoids already present on the island should be considered as being complementary, rather than as an alternative, to biological control for Tachardina.
The goal of the proposed action is to achieve ecologically-effective, cost-effective, and self-sustaining suppression of YCA supercolonies through the importation of a biological control agent specifically for Tachardina. Using these criteria, Alternative 1 is not a viable option because it is not self-sustaining, and Alternative 2 is not a viable option by itself because it is unlikely to achieve supercolony suppression.
2.3 Alternative locations, time frames or activities that form part of the referred action If you have identified that the proposed action includes alternative time frames, locations or activities (in section 1.10) you must complete this section. Describe any alternatives related to the physical location of the action, time frames within which the action is to be taken and alternative methods or activities for undertaking the action. For each alternative location, time frame or activity identified, you must also complete (where relevant) the details in sections 1.2-1.9, 2.4-2.7, 3.3 and 4. Please note, if the action that you propose to take is determined to be a controlled action, any alternative locations, time frames or activities that are identified here may be subject to environmental assessment and a decision on whether to approve the alternative.
N/A 2.4 Context, planning framework and state/local government requirements Explain the context in which the action is proposed, including any relevant planning framework at the state and/or local government level (e.g. within scope of a management plan, planning initiative or policy framework). Describe any Commonwealth or state legislation or policies under which approvals are required or will be considered against.
Ecological Context. The Expert Working Group’s Final Report (Beeton et al. 2010) provides an ecosystem-wide context for the proposed action. The EWG’s terms of reference included an examination of all threats to Christmas Island’s ecology, biodiversity management and any other issues relating to conservation management of the island and its surrounds. The principal finding of the EWG was that the extremely high biodiversity values of Christmas Island are in a “parlous state” (p. 9), to the extent that Christmas Island and its surrounding seas should be considered for listing as a threatened ecological community under the EPBC Act 1999. Invasion and supercolony formation by YCA was identified as a major contributing factor, and Loss of biodiversity and ecosystem integrity
following invasion by the Yellow Crazy Ant (Anoplolepis gracilipes on Christmas Island, Indian Ocean has been listed as a Key Threatening Process under the EPBC Act since 2005. Numerous scientific publications and reports over more than a decade have documented the direct and indirect negative impacts of YCA supercolony formation on various aspects of the island’s ecology (O’Dowd et al. 1999; Green et al. 2001; Marr et al. 2003; O’Dowd et al. 2003; Davis et al. 2008, 2010; Green and O’Dowd 2009; O’Dowd and Green 2010; Green et al. 2011, Smith et al., unpublished results).
Planning Framework – The Yellow Crazy Ant Research & Control Program. This 10-year plan was developed by DNP/CINP and CASAP to map out the management of Yellow Crazy Ants from 2007 to 2017. The major objectives of the program include significantly reducing the impact of Yellow Crazy Ants on the biodiversity values of Christmas Island by removing all existing, and halting future, development of supercolonies through:

Environment Protection and Biodiversity Conservation Act 1999
Page 15 of 44
a) development of more target-specific baits which can be used on a more regular and widespread basis.
b) moving the baiting control methodology to a predominantly aerial approach.
c) research and development of biological control agents to reduce the densities of scale insect mutualists, potentially reducing the main cause of YCA supercolony formation.
Part C of this ten-year plan was fully endorsed by two recommendations in the EWG Final Report (Beeton et al. 2010):
Recommendation 9: (High priority) The initial steps taken already to explore biological control of the introduced scale insects be accelerated and biological control trials be started as soon as possible.
Recommendation 29: (High priority) Fundamental investigations continue and be augmented by adaptive management and aspects of Integrated Pest Control experimental work to develop cost-effective methods to break the scale insect - Yellow Crazy Ant mutualistic dependence.
Regulatory Framework – Director of National Parks and the Christmas Island Management Plan. The EPBC Act 1999 outlines the responsibilities of the Director of National Parks (DNP) to protect, conserve and manage biological diversity in Commonwealth reserves. The EPBC Act Regulations provide for the conservation of biological diversity in Commonwealth areas.
The EPBC Act requires the Director of National Parks to prepare a management plan for the Park at all times after the first plan for managing the park comes into effect. The third management plan for Christmas Island National Park (2002 - 2009) ceased to be in operation in March 2009 and the fourth management plan is currently in draft (2012) but is likely to be in force in early to mid 2013, before any importation of a biological control agent would occur. Until the fourth Plan comes into operation the Park will be managed in accordance with Section 357 of the EPBC Act. Section 357(1) states:
While a management plan is not in operation for a Commonwealth Reserve, the Director must exercise the Director's powers and perform the Director's functions in relation to the reserve or to a zone of the reserve so as to manage the reserve in accordance with:
a) the Australian IUCN reserve management principles for the IUCN category to which the reserve or zone has most recently been assigned by:
i. a Proclamation made under Subdivision B; or ii. a management plan that was in operation for the reserve (but is no longer); and
b) if the Director holds land or seabed included in the reserve under lease - the Directors obligations under the lease.
The specific objectives of the draft Christmas Island Management Plan relevant to this referral are:
Landscape
4.1.1 Impacts on the values and integrity of the park’s landscape, including visitor experience values will be avoided or minimised.
4.1.7 If the landscape changes in ways that threaten landscape or ecosystem values or otherwise beyond levels of acceptable change, the Director, in consultation with relevant stakeholders, will identify further monitoring requirements and will decide whether protection, rehabilitation or adaptation measures can and will be implemented.

Environment Protection and Biodiversity Conservation Act 1999
Page 16 of 44
Terrestrial vegetation 4.2.3 Subject to risk and required approvals including from the Australian Quarantine and Inspection Service, the Director may introduce, or issue a permit for the introduction of, non-native species for conservation purposes such as the biological control of invasive species
4.2.4 So far as practicable, implement multi-species recovery plans, and relevant threat abatement plans for listed threatened species.
Terrestrial animals
4.3.1 Determine priorities for actions to conserve native plant and animal species based on:
a) a species’ conservation or biodiversity value, with high priority placed on endemic, keystone and EPBC Act listed and threatened species, and the risks of taking no action
b) likelihood that proposed actions will have ecosystem or multiple species benefits
c) likelihood that proposed actions will achieve their conservation aims
d) cost benefits and effectiveness of implementing proposed actions.
4.3.2 If EPBC Act listed, endemic, keystone or otherwise threatened or significant species are in decline to a level that may threaten their conservation status, the Director will:
a) assess the likelihood of mitigating known threats and, if feasible, implement threat mitigation strategies
b) if threats are not known, seek to determine (so far as possible) the threats and appropriate mitigation measures
c) if threats are not known or not likely to be mitigated for some time, assess the feasibility and effectiveness of implementing interventionist programs, such as captive breeding, that have the long-term aim of conserving the species in their natural environment.
4.3.3 The Director may take actions concerning animal species, including species listed under Part 13 of the EPBC Act, that are otherwise prohibited by the EPBC Act or Regulations where they are necessary to implement this plan, or where they are otherwise necessary for preserving or protecting the park, protecting or conserving biodiversity, or protecting persons or property in the park.
4.3.4 So far as practicable implement, multi-species recovery plans, and relevant threat abatement plans for listed threatened species.
4.3.5 Implement and contribute to on-park and off-park actions for conserving ecosystems and native species, particularly EPBC Act listed and other threatened species including red crabs and other keystone species.
4.3.7 Assess and monitor threats to native species. This will include the risk of invasive species which are currently considered a low threat becoming a greater threat.
4.3.9 Continue to implement the crazy ant control program including monitoring the impact of crazy ants on red crabs and other native species.
Ramsar wetlands and other freshwater wetlands
4.4.1 Applicable policies and actions from this plan will be applied to managing The Dales and Hosnies Spring.
4.4.4 Monitor and maintain the ecological character of the Ramsar listed wetlands known as Hosnies Spring and The Dales. This may include mitigating the impacts of potential threats, such as invasive species and unmanaged visitor use.

Environment Protection and Biodiversity Conservation Act 1999
Page 17 of 44
Research and monitoring
8.7.1 Research and monitoring will focus on conserving, managing or restoring the park’s and island’s natural heritage values.
8.7.2 The Director or permitted organisations or individuals may carry out research and monitoring actions concerning native species, including species listed under Part 13 of the EPBC Act.
8.7.3 Permits authorising research and monitoring may be issued for conservation focused research consistent with this plan and with the park’s IUCN category (see Section 3.1).
8.7.4 Persons carrying out research and monitoring in the park will be required to provide the Director with reports on their work in ways requested by the Director.
8.7.6 Where needed, seek independent scientific and other specialist advice, for example through relevant advisory groups, for conserving and managing natural heritage.
8.7.8 Conduct and/or support or encourage conservation management research, monitoring and studies that assist with conserving and managing the park’s and island’s natural heritage, particularly studies on:
a) landscape and ecosystem changes and threats including monitoring and management of biodiversity condition
b) restoration, recovery and adaptive management of ecosystems and their species including rehabilitated minesites and red crab migration routes/habitat
c) populations of threatened and otherwise significant species, including but not limited to seabirds and land crabs (particularly red and robber crabs) and, as needed, invasive species
d) identification, quantification and, as needed, management of threatening processes impacting on ecosystems and threatened species. Give priority to invasive species, including the indirect biological control of crazy ants and control of cats and rats
8.7.9 Where possible, provide support for suitably qualified research organisations or individuals conducting natural heritage research consistent with Section 8.7.8.
Regulatory Framework – Commonwealth or state legislation or policies under which approvals are required or will be considered against
Any regulatory framework under which the introduction of a biological control agent to Christmas Island would be conducted is currently unclear. However, the Biological Control Act 1984, indicates in Section 4(1) that the responsible Minister, by giving notice in the Gazette, can extend the Act to the external territory of Christmas Island (http://www.comlaw.gov.au/Details/C2008C00315). In the absence of this extension, there is no obvious regulatory framework applicable to these circumstances. For example, the advisory committees (Plant Health Committee [PHC] and the National Biosecurity Committee [NBC]) recommend approvals to the Primary Industry Standing Committee (PISC). A clear approvals process exists for nomination of a biological control target (the yellow lac scale Tachardina aurantiaca [Kerriidae], in this case), but as an external territory Christmas Island appears to be outside of the jurisdiction of both the PHC and NBC. Currently no member on the PHC represents this external island territory; and, even if there were, the island is not within the jurisdiction of the NBC, which approves recommendations made by the PHC.
Nevertheless, initial steps have been followed in the Biosecurity Guidelines for the Introduction of Exotic Biological Control Agents for the Control of Weeds and Plant Pests

Environment Protection and Biodiversity Conservation Act 1999
Page 18 of 44
(http://www.daff.gov.au/ba/reviews/biological_control_agents/protocol_for_biological_control_agents).A nomination of the target species as a candidate for biological control has been produced (see Attachment 4); offshore research in Malaysia on prospective biological control agents is underway; and a rationale for host-specificity testing has been produced (Appendix 1). Approval would be sought to undertake host-specificity testing within the natural range of the biological control agent (Malaysia) rather than in containment on the Australian mainland.
It is intended that in the absence of a regulatory framework for introduction of an exotic biological control agent that is applicable to Christmas Island, this referral will act as a catalyst and help provide a basis for the relevant government agencies to identify, in a timely manner, a workable framework under which to consider import approval. 2.5 Environmental impact assessments under Commonwealth, state or territory legislation If you have identified that the proposed action will be or has been subject to a state or territory environmental impact statement (in section 1.11) you must complete this section. Describe any environmental assessment of the relevant impacts of the project that has been, is being, or will be carried out under state or territory legislation. Specify the type and nature of the assessment, the relevant legislation and the current status of any assessments or approvals. Where possible, provide contact details for the state/territory assessment contact officer. Describe or summarise any public consultation undertaken, or to be undertaken, during the assessment. Attach copies of relevant assessment documentation and outcomes of public consultations (if available). As described above, the regulatory framework under which the introduction of a biological control agent to Christmas Island would be considered cannot be clearly identified. Therefore, the requirement for a formal state or territory environmental impact assessment is also unclear but is considered unlikely, given that the import of exotic species (e.g. plants for propagation) may be permitted (and occurs) under existing biosecurity import arrangements. In lieu of a formal environmental impact assessment, the matters of greatest relevance in the assessment of this Referral are addressed; the host specificity of the proposed biological control agent, and the likelihood of off-target impacts on other scale insects or species of concern.
Host Specificity of Tachardiaephagus somervillei. Based on known host records, all Tachardiaephagus species have narrow host ranges and appear to be family specialists, known only to attack scale insects in the Kerriidae, the family to which the yellow lac scale belongs (Table 2; see also O’Dowd et al. 2012). Table 2. Records of known host families and genera for the primary parasitoid Tachardiaephagus (Encyrtidae). As a genus, Tachardiaephagus has an extremely broad geographic range. With the exception of one probably erroneous host record in Africa, all Tachardiaephagus species appear to be family specialists and restricted to the Kerriidae. For host genera, number of species recorded as hosts is in parentheses. Based on Noyes (2012, Universal Chalcidoidea Database, http://www.nhm.ac.uk/research-curation/research/projects/chalcidoids/database/), except for records for T. somervillei and T. sarawakensis (Hayat et al. 2010. Oriental Insects 44, 23; R.W. Pemberton, pers. comm.; this study)
Species Distribution Recorded hosts
Tachardiaephagus somervillei India, Malaysia, Thailand Kerriidae
Kerria spp. (4)
Tachardina aurantiaca
Tachardina sp. (1)
T. sarawakensis Sarawak
Kerriidae
Tachardina aurantiaca
T. tachardiae Brunei, China, India, Indonesia, Malaysia,
Sri Lanka, Taiwan, Vietnam, Azerbaijan
Kerriidae
Kerria spp. (6)

Environment Protection and Biodiversity Conservation Act 1999
Page 19 of 44
Laccifer spp. (3)
Paratachardina lobata
T. similis Afrotropical,
South Africa
Kerriidae
Tachardina sp. (1)
Coccidae
?Ceroplastes eucleae1
T. absonus Afrotropical,
South Africa
Kerriidae
Tachardina spp. (2)
T. communis Afrotropical,
South Africa
Kerriidae
Tachardina spp. (5)
T. gracilis Afrotropical,
South Africa
Kerriidae
Tachardina sp. (1)
1This record is likely to be erroneous (see Prinsloo1983. Entomol. Mem. Dept. Agric. Rep. S. Afr. 60, 26). Scale insect diversity on Christmas Island. No native or endemic scale insect species have been discovered in 400 hours of search over two years of intensive and extensive searches for scale insect species on Christmas Island (O’Dowd et al. 2012). However, this effort did yield five additional exotic scale insect species previously unknown to the island (Table 3). Table 3. Scale insects of Christmas Island. It is highly probably that all of these species, with broad host plant ranges and geographic distributions, are exotic to Christmas Island and introduced following human settlement. The yellow lac scale, Tachardina aurantiaca (Kerriidae) and Coccidae (soft scales) are the primary honeydew suppliers to the yellow crazy ant. Scale insects in bold occur commonly in YCA supercolonies. Taxonomy and distribution from Ben-Dov et al. (2012, http://www.sel.barc.usda. gov/scalenet/scalenet.htm).
Family and Species
Common Name Distribution Honeydew Producer
Kerriidae (lac scales) Tachardina aurantiaca (Cockerell) *
Paratachardina pseudolobata (Kondo & Gullan)
Yellow lac scale False lobate lac scale
Oriental Oriental, Nearctic, Neotropical
Yes Yes2
Coccidae (soft scales) Coccus celatus De Lotto *C. hesperidium Linnaeus *Milviscutulus mangiferae (Green) *Ceroplastes ceriferus (Fabricius) *C. destructor Newstead Saissetia oleae (Olivier) *S. coffeae (Walker) **Parasaissetia nigra
1
**Pulvinaria urbicola Cockerell
**P. psidii
1
Green coffee scale Brown soft scale Mango shield scale Indian wax scale White wax scale Black olive scale Hemispherical scale Nigra scale Urbicola soft scale Green shield scale
Afrotropical, Australasia, Oriental Cosmopolitan Cosmopolitan Cosmopolitan Afrotropical, Australasia, Oriental Pantropical Cosmopolitan Cosmopolitan Pantropical Cosmopolitan
Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes
Monophlebidae (giant scales) Icerya purchasi (Maskell)
Cottony cushion scale
Cosmopolitan
Yes
Cerococcidae (ornate pit scales) **Cerococcus indicus (Maskell)
Spiny brown coccid
Cosmopolitan
Yes
Pseudococcidae (mealybugs) **Dysmicoccus finitimus Williams
**Ferrisia virgata (Cockerell)1
Asian coconut mealybug Striped mealybug
Australasia, Oriental Cosmopolitan
Yes Yes
Diaspididae (armoured scales) Aspidiotus destructor (Signoret) Pseudaulacaspis pentagona
Coconut scale White peach scale
Cosmopolitan Cosmopolitan
No No

Environment Protection and Biodiversity Conservation Act 1999
Page 20 of 44
(Targioni Tozzetti) *Hemiberlesia palmae (Cockerell) *Lindingaspis sp. **Lepidosaphes sp.1
Tropical palm scale -- Oystershell scale
Cosmopolitan -- --
No No No
*Record added by Abbott (2004, PhD Thesis, Monash University); **Record added by Neumann et al. (unpublished results); 1Tentative identification; 2P. pseudolobata produces honeydew but ejects it instead of producing droplets that can be collected by ants (Howard 2010, Fl. Entomol. 93, 1). Likelihood of off-target impacts. All known scale insect species on the island are exotic/invasive, and the proposed biological control agent for Tachardina on Christmas Island appears to be a narrow family specialist (Kerriidae). After intensive survey, no scale insect species of concern have been found on the island, and the probability of any direct non-target effects on any scale insects is negligible. Further, given the co-evolved nature of host-parasitoid interactions, it is extremely unlikely that Tachardiaephagus, as a primary parasitoid wasp, will attack any native Christmas Island insects. Risk of off-target impacts will be addressed by conducting host-specificity tests within the native range of Tachardina in Malaysia where it is safest to conduct these tests. 2.6 Public consultation (including with Indigenous stakeholders) Your referral must include a description of any public consultation that has been, or is being, undertaken. Where Indigenous stakeholders are likely to be affected by your proposed action, your referral should describe any consultations undertaken with Indigenous stakeholders. Identify the relevant stakeholders and the status of consultations at the time of the referral. Where appropriate include copies of documents recording the outcomes of any consultations.
Christmas Island has never been settled by Indigenous peoples prior to European settlement.
The local community on Christmas Island is very knowledgeable about YCA and its ecological impacts because of public outreach initiatives (community meetings, information brochures and stakeholder meetings) carried out by the DNP and La Trobe University over a number of years The local community is also very aware of the need to suppress supercolonies using toxic bait, especially because of the high community profile of the issue generated by three helicopter baiting campaigns, and the recent (September 2012) ground baiting of YCA supercolonies behind the Kampong, a major residential area. The concept of a biological control program for the suppression of YCA supercolonies was first introduced to the community through public presentations and meetings in 2009. University researchers Green and O’Dowd gave repeat presentations at the Malay Club, Sports and Recreation Centre, the Chinese Literary Association, the Christmas Island District High School and to Phosphates Resources Limited, in an effort to inform a wide cross section of the community of the proposal. The proposal was welcomed insofar as it presents an alternative means of suppressing supercolonies. All community groups appreciated the potential risks of introducing another non-native species to Christmas Island, but were reassured that risk assessment and strict regulatory oversight would occur prior to any importation and release of any biological control agent. Further public meetings will be held closer to the time of importation and release.
In addition, several key island based stakeholders, especially the Department of Regional Australia, Local Government, Arts and Sport (DRALGAS) are aware of and support (in-principle) the biological control research. This is reflected in the Australian Government’s response (http://www.environment.gov.au/parks/publications/christmas/ewg-response.html) to the Expert Working Groups final report (Beeton et al. 2010), particularly recommendation nine that: “the initial steps taken already to explore biological control of the introduced scale insects be accelerated and biological control trials be started as soon as possible”.
2.7 A staged development or component of a larger project If you have identified that the proposed action is a component of a larger action (in section 1.12) you must complete this section. Provide information about the larger action and details of any interdependency between the stages/components and the larger action. You may also provide justification as to why you believe it is reasonable for the referred action to be

Environment Protection and Biodiversity Conservation Act 1999
Page 21 of 44
considered separately from the larger proposal (eg. the referred action is ‘stand-alone’ and viable in its own right, there are separate responsibilities for component actions or approvals have been split in a similar way at the state or local government levels).
The proposed action is part of a larger program of integrated pest management designed to ameliorate the impacts of YCA invasion by suppressing supercolony formation (see section 2.4 above, Planning Framework). The current backbone of the program is surveillance and monitoring to detect and map supercolonies, followed by the use of toxic bait, distributed by hand on the ground or by helicopter, to suppress them. The bait is AntOff®, with the active ingredient Fipronil at 0.01 g kg-1 in a fishmeal protein matrix, broadcast at 4 kg ha-1. Several alternative chemical approaches, including different neurotoxins and insect growth regulators (IGRs), have also been trialled but so far without success. New trials using IGRs were commenced during the aerial baiting campaign of September 2012 but the outcome of those trials will not be known until early to mid-2013.
3 Description of environment & likely impacts
3.1 Matters of national environmental significance Describe the affected area and the likely impacts of the proposal, emphasising the relevant matters protected by the EPBC Act. Refer to relevant maps as appropriate. The interactive map tool can help determine whether matters of national environmental significance or other matters protected by the EPBC Act are likely to occur in your area of interest. Your assessment of likely impacts should refer to the following resources (available from the Department’s web site): specific values of individual World Heritage properties and National Heritage places and the ecological character of
Ramsar wetlands; profiles of relevant species/communities (where available), that will assist in the identification of whether there is likely
to be a significant impact on them if the proposal proceeds; Significant Impact Guidelines 1.1 – Matters of National Environmental Significance; and associated sectoral and species policy statements available on the web site, as relevant. Your assessment of likely impacts should consider whether a bioregional plan is relevant to your proposal. The Minister has prepared four marine bioregional plans (MBP) in accordance with section 176. It is likely that the MBP’s will be more commonly relevant where listed threatened species, listed migratory species or a Commonwealth marine area is considered. Note that even if your proposal will not be taken in a World Heritage area, Ramsar wetland, Commonwealth marine area, the Great Barrier Reef Marine Park or on Commonwealth land, it could still impact upon these areas (for example, through downstream impacts). Consideration of likely impacts should include both direct and indirect impacts.
3.1 (a) World Heritage Properties
Description
N/A. There are no World Heritage Properties on Christmas Island
Nature and extent of likely impact
Address any impacts on the World Heritage values of any World Heritage property.
N/A. There are no World Heritage Properties on Christmas Island

Environment Protection and Biodiversity Conservation Act 1999
Page 22 of 44
3.1 (b) National Heritage Places
Description (from Draft Christmas Island National Park Management Plan, Commonwealth of Australia 2012).
Christmas Island is not listed as a National Heritage Place but is listed on the Commonwealth Heritage register for both natural and cultural reasons (Commonwealth of Australia 2012).
Christmas Island has a number of unique ecological characteristics, as well as other features common to other oceanic islands. Characteristics of Christmas Island’s ecosystems include: fewer species than comparable areas of land on continents or islands near continents but with a relatively higher proportion of endemic terrestrial species; evolutionary isolation for thousands or millions of years until recent human arrival; native species that have evolved with few competitors; and many species with small population sizes. These characteristics make the island’s ecosystems and species of high conservation value but also highly vulnerable to environmental change, particularly from invasive species, climate change, and habitat clearing, fragmentation and degradation.
Christmas Island also has nine cultural sites on the register. Eight of the nine sites are located within settled areas of the island, and the other site (South Point Settlement remains) is well away from settled areas. Nature and extent of likely impact
Address any impacts on the National Heritage values of any National Heritage place.
The likelihood of negative impacts on species of concern from the introduction of a biological control agent is rated as negligible (see Section 2.5); therefore, the likelihood of negative impacts on Commonwealth Heritage Places is also rated as negligible. The likelihood of negative impacts on cultural sites is nil.
3.1 (c) Wetlands of International Importance (declared Ramsar wetlands)
Description (extracted from the Draft Christmas Island National Park Management Plan, Commonwealth of Australia 2012)
The Dales (RS1225). The Dales Ramsar site includes a series of seven dales, three of which support permanent springs and four support intermittent streams. The Dales are surrounded predominantly by semi-deciduous forest. On the seaward side at the edge of the shore terrace there is a line of coastal shrubland which merges with sea cliffs and rocky marine shores. The site extends seaward 50 metres and includes part of a narrow, shallow, sloping reef. Mixed amongst the terrestrial and marine environments is a range of karst features, highly representative of the environment of Christmas Island. The combination of this variety of habitats and the presence of permanent surface water supports a wide diversity of endemic and threatened species.
The site hosts part of the annual red crab migration and provides critical habitat for blue crabs as well as other land crabs. The Dales supports a diverse community of tree species and epiphytes. At Hugh’s Dale, and in parts of Anderson Dale and Sydney’s Dale, there are mono-specific stands of Tahitian Chestnut, Inocarpus fagifer, and the rare epiphytic Ribbon Fern, Ophioglossum pendulum. The endemic Arenga Palm, Arenga listeri, and endemic Ridley’s Orchid, Brachypeza archytas, are common in The Dales. Terminalia catappa grows to an unusual size on Christmas Island and several large specimens occur in The Dales. A number of endemic fauna occur within The Dales including the Abbott’s Booby and several land bird species. The endemic freshwater fish, the Brown Gudgeon, Eleotris fusca, has also been recently sighted within The Dales.
Hosnies Spring (RS512). The Hosnies Spring Ramsar site is an area of permanent, shallow freshwater wetland, fed by a natural spring system located approximately 30 metres above sea level

Environment Protection and Biodiversity Conservation Act 1999
Page 23 of 44
and 120 metres inland of the seaward cliff. The wetland is covered by a stand of mangroves including Bruguiera gymnorhiza and B. sexangula estimated to be 120,000 years old. The margins of the wetland are well defined, with limestone cliffs to the north and west and a sharp transition to hibiscus and pandanus to the south. The area that surrounds the wetland site is predominantly rainforest, characterised by a 20 to 30 metre tall canopy of evergreen and deciduous tree species such as Pisonia
grandis and Barringtonia racemosa and a conspicuous lack of herb and shrub layers. There is a narrow band of coastal scrub with hardy species such as Scaevola taccada at the seaward margin of the shore terrace, with an unvegetated area of limestone pinnacles at the top of the sea cliff. The cliff descends some 17 metres almost vertically to the rocky marine shore below. The site extends 50 metres seaward of the low water mark and includes areas of shallow coral reef.
Hosnies Spring is remarkable for a number of reasons. First, it is one the few permanent freshwater areas on Christmas Island. Second, the mangroves occur at an elevation not recorded anywhere else in the world. Third, the age of the mangrove stand is extraordinary and finally, the individual trees are very large. The site also supports endemic and other significant fauna species including land crabs (in particular red, robber and blue crabs), sea and land birds and the Christmas Island flying fox.
In 2010, Ecological Character Descriptions for The Dales and Hosnies Spring were prepared under the National Framework and Guidance for Describing the Ecological Character of Australia’s Ramsar Wetlands (Hale and Butcher 2010). These descriptions form a baseline reference to help monitor and maintain the ecological character of The Dales and Hosnies Spring.
Nature and extent of likely impact
Address any impacts on the ecological character of any Ramsar wetlands.
Because the biological control agent is a host-specific, parasitoid insect, there are no foreseeable negative impacts associated with its introduction on the conservation values of either Ramsar Wetland. It is anticipated that there will be positive impacts on Ramsar sites, because of the likely reduction of YCA densities at these sites.

Environment Protection and Biodiversity Conservation Act 1999
Page 24 of 44
3.1 (d) Listed threatened species and ecological communities
Description
EPBC Act - Listed threatened species occurring in Christmas Island National Park
Common name Scientific name Status
MAMMALS
Christmas Island shrew Crocidura attenuata trichura EN
Humpback whale Megaptera novaeangliae VU
Christmas Island pipistrelle Pipistrellus murrayi CR
Maclear's rat Rattus macleari EX
Bulldog rat Rattus nativitatis EX
BIRDS
Abbott's booby Papasula abbotti EN
Christmas Island emerald dove Chalcophaps indica natalis EN
Christmas Island frigatebird Fregata andrewsi VU
Christmas Island goshawk Accipiter fasciatus natalis EN
Christmas Island hawk-owl Ninox natalis VU
Christmas Island thrush Turdus poliocephalus erythropleurus EN
REPTILES
Green turtle Chelonia mydas VU
Hawksbill turtle Eretmochelys imbricata VU
Tree gecko Lepidodactylus listeri VU
Pink blind snake Typhlops exocoeti VU
FISH
Whale shark Rhincodon typus VU
VASCULAR PLANTS
Christmas Island spleenwort Asplenium listeri CR
A fern Pneumatopteris truncata CR
A fern Tectaria devexa var. minor EN
EN: endangered; EX: extinct; CR: critically endangered; VU: vulnerable
Nature and extent of likely impact
Address any impacts on the members of any listened threatened species (except a conservation dependent species) or any threatened ecological community, or their habitat.
Because the proposed biological agent is a primary parasitoid of one particular family of introduced scale insects, there are no foreseeable negative impacts of its introduction on any EPBC-Listed species. It is anticipated that there will be positive impacts on any EPBC-Listed (terrestrial) species, because of the likely reduction of YCA densities in their habitats.

Environment Protection and Biodiversity Conservation Act 1999
Page 25 of 44
3.1 (e) Listed migratory species
Description
There are 68 bird species classed as migratory (or vagrant) detected on Christmas Island and five marine species including reptiles, mammals and one fish (see table in section 3.1d) that are recognised by EPBC, JAMBA and/or CAMBA and the Bonn Convention.
Common name Scientific name
EPBC Act marine
EPBC Act migratory JAMBA CAMBA ROKAMBA BONN
MAMMALS
Humpback whale Megaptera novaeangliae
Long-snouted spinner dolphin
Stenella longirostris (E. tropical Pacific/SE Asian pops)
BIRDS
Abbott’s booby Papasula abbotti = Sula abbotti
Antarctic prion Pachyptila desolata
Australian pelican Pelecanus conspicillatus
Australian pratincole Stiltia isabella
Arctic jaeger Stercorarius parasiticus
Baillon’s crake Porzana pusilla
Barn swallow Hirundo rustica
Bar-tailed godwit Limosa lapponica
Black-winged stilt Himanotopus himanotopus
Brown booby Sula leucogaster
Brown goshawk Accipiter fasciatus
Bulwer’s petrel Bulweria bulwerii
Cattle egret Ardea ibis = Bubulcus ibis
Christmas Island frigatebird Fregata andrewsi
Common greenshank, greenshank Tringa nebularia
Common koel Eudynamys scolopacea
Common noddy Anous stolidus
Common sandpiper Actitis hypoleucos =Tringa hypoleucos
Common tern Sterna hirundo
Crested tern Sterna bergii
Curlew sandpiper Calidris ferruginea
Dollarbird Eurystomus orientalis
Eastern reef egret Egretta sacra
Fork-tailed swift Apus pacificus
Garganey Anas querquedula
Glossy ibis Plegadis falcinellus
Great egret Ardea alba = Egretta alba
Great frigatebird Fregata minor
Great knot Calidris tenuirostris
Great skua Catharacta skua

Environment Protection and Biodiversity Conservation Act 1999
Page 26 of 44
Greater sand plover Charadrius leschenaultii
Grey phalarope Phalaropus fulicarius
Grey plover Pluvialis squaterola
Grey wagtail Motacilla cinerea
Grey-tailed tattler Heteroscelus brevipes
Least frigatebird Fregata ariel
Little curlew Numenius minutus
Little egret Egretta garzetta
Little ringed plover Charadrius dubius
Little tern Sterna albifrons
Malay night heron Gorsachius melanolophus
Marsh sandpiper, little greenshank Tringa stagnatilis
Mongolian plover = lesser sand plover Charadrius mongolus
Nankeen kestrel Falco cenchroides
Nankeen night heron Nycticorax caledonicus
Oriental cuckoo Cuculus saturatus
Oriental plover Charadrius veredus = C. asiaticus veredus
Oriental pratincole Glareola maldivarum
Oriental reed-warbler Acrocephalus orientalis Pacific (= lesser) golden plover Pluvialis fulva
Pallid cuckoo Cuculus pallidus Pied imperial-pigeon Ducula bicolor Pin-tailed snipe Gallinago stenura Red-footed booby Sula sula Red-necked phalarope Phalaropus lobatus Red-necked stint Calidris ruficollis Red-rumped swallow Hirundo daurica Red-tailed tropicbird Phaethon rubricauda
Richard’s pipit Anthus novaeseelandiae
Ruddy turnstone Arenaria interpres Sacred kingfisher Todiramphus sanctus
Sanderling Calidris alba = Crocethia alba
Sharp-tailed sandpiper Calidris acuminata Sooty tern Sterna fuscatai Swinhoe’s snipe Gallinago megala Terek sandpiper Xenus cinereus Tree martin Hirundo nigricans Wedge-tailed shearwater Puffinus pacificus Whimbrel Numenius phaeopus Whiskered tern Chlidonias hybridus White tern Gygis alba White-tailed tropicbird Phaethon lepturus White wagtail Motacilla alba White-bellied sea-eagle Haliaeetus leucogaster
White-throated needletail = Spine-tailed swift
Hirundapus caudacutus

Environment Protection and Biodiversity Conservation Act 1999
Page 27 of 44
White-winged black tern Chlidonias leucopterus Wood sandpiper Tringa glareola Yellow wagtail Motacilla flava REPTILES Green turtle Chelonia mydas
Hawksbill turtle Eretmochelys imbricata
Yellow-bellied sea snake Pelamis platurus FISH Bluestripe pipefish Doryramphus excisus
Fijian pipefish Corythoichthys amplexus
Ornate ghost pipefish Solenostomus paradoxus
Redstripe pipefish Doryramphus baldwini
Reef-top pipefish Corythoichthys haematopterus
Robust ghost pipefish Solenostomus cyanopterus
Roughridge pipefish Cosmocampus banneri Schultz’s pipefish Corythoichthys schultzi Sculptured pipefish Choeroichthys sculptus
Short-bodied pipefish Choeroichthys brachysoma
Thorn-tailed pipefish = pygmy pipefish
Micrognathus brevirostris pygmaeus
Whale shark Rhincodon typus
Yellow-banded pipefish Corythoichthys flavofasciatus
JAMBA – Japan-Australia Migratory Bird Agreement; CAMBA – China-Australia Migratory Bird Agreement; ROKAMBA – Korea-Australia Migratory Bird Agreement; Bonn – Bonn Convention
Nature and extent of likely impact
Address any impacts on the members of any listed migratory species, or their habitat.
Because the proposed biological agent is a primary parasitoid with hosts within one particular family of introduced scale insects, there are no foreseeable negative impacts of its introduction on any listed migratory species. It is anticipated that there will be positive impacts on EPBC-Listed migratory (terrestrial) species, because of the likely reduction of YCA densities in their habitats.
3.1 (f) Commonwealth marine area (If the action is in the Commonwealth marine area, complete 3.2(c) instead. This section is for actions taken outside the Commonwealth marine area that may have impacts on that area.) Description
N/A. The proposed action will not be taken in a marine area that lies outside the Commonwealth marine area
Nature and extent of likely impact
Address any impacts on any part of the environment in the Commonwealth marine area.
N/A. The proposed action will not be taken in a marine area that lies outside the Commonwealth marine area

Environment Protection and Biodiversity Conservation Act 1999
Page 28 of 44
3.1 (g) Commonwealth land (If the action is on Commonwealth land, complete 3.2(d) instead. This section is for actions taken outside Commonwealth land that may have impacts on that land.) Description If the action will affect Commonwealth land also describe the more general environment. The Policy Statement titled Significant Impact Guidelines 1.2 - Actions on, or impacting upon, Commonwealth land, and actions by Commonwealth agencies provides further details on the type of information needed. If applicable, identify any potential impacts from actions taken outside the Australian jurisdiction on the environment in a Commonwealth Heritage Place overseas.
N/A. The proposed action will not be taken in a terrestrial area that lies outside Commonwealth land
Nature and extent of likely impact
Address any impacts on any part of the environment in the Commonwealth land. Your assessment of impacts should refer to the Significant Impact Guidelines 1.2 - Actions on, or impacting upon, Commonwealth land, and actions by Commonwealth agencies and specifically address impacts on: ecosystems and their constituent parts, including people and communities; natural and physical resources; the qualities and characteristics of locations, places and areas; the heritage values of places; and the social, economic and cultural aspects of the above things.
N/A. The proposed action will not be taken in a terrestrial area that lies outside Commonwealth land
3.1 (h) The Great Barrier Reef Marine Park
Description
N/A. The proposed action will take place on Christmas Island, several thousand kilometres away from the GBRMP
Nature and extent of likely impact
Address any impacts on any part of the environment of the Great Barrier Reef Marine Park.
Note: If your action occurs in the Great Barrier Reef Marine Park you may also require permission under the Great Barrier Reef Marine Park Act 1975 (GBRMP Act). If so, section 37AB of the GBRMP Act provides that your referral under the EPBC Act is deemed to be an application under the GBRMP Act and Regulations for necessary permissions and a single integrated process will generally apply. Further information is available at www.gbrmpa.gov.au
N/A. The proposed action will take place on Christmas Island, several thousand kilometres away from the GBRMP
3.2 Nuclear actions, actions taken by the Commonwealth (or Commonwealth agency), actions taken in a Commonwealth marine area, actions taken on Commonwealth land, or actions taken in the Great Barrier Reef Marine Park
You must describe the nature and extent of likely impacts (both direct & indirect) on the whole environment if your project: is a nuclear action; will be taken by the Commonwealth or a Commonwealth agency; will be taken in a Commonwealth marine area; will be taken on Commonwealth land; or will be taken in the Great Barrier Reef marine Park. Your assessment of impacts should refer to the Significant Impact Guidelines 1.2 - Actions on, or impacting upon, Commonwealth land, and actions by Commonwealth agencies and specifically address impacts on: ecosystems and their constituent parts, including people and communities; natural and physical resources; the qualities and characteristics of locations, places and areas; the heritage values of places; and the social, economic and cultural aspects of the above things.

Environment Protection and Biodiversity Conservation Act 1999
Page 29 of 44
3.2 (a) Is the proposed action a nuclear action?
No
Yes (provide details below)
If yes, nature & extent of likely impact on the whole environment
3.2 (b) Is the proposed action to be taken by the
Commonwealth or a Commonwealth agency?
No
Yes (provide details below)
If yes, nature & extent of likely impact on the whole environment
Researchers from La Trobe University will undertake the proposed action under contract to the Director of National Parks. As outlined in detail in Sections 2.5, 3.1(b-e) and Section 5, there are no foreseeable negative impacts on the environment.
3.2 (c) Is the proposed action to be taken in a
Commonwealth marine area?
No
Yes (provide details below)
If yes, nature & extent of likely impact on the whole environment (in addition to 3.1(f))
3.2 (d) Is the proposed action to be taken on Commonwealth land?
No
Yes (provide details below)
If yes, nature & extent of likely impact on the whole environment (in addition to 3.1(g))
As outlined in detail in Sections 2.5, 3.1(b-e) and Section 5, there are no foreseeable negative impacts on the environment.
3.2 (e) Is the proposed action to be taken in the
Great Barrier Reef Marine Park?
No
Yes (provide details below)
If yes, nature & extent of likely impact on the whole environment (in addition to 3.1(h))

Environment Protection and Biodiversity Conservation Act 1999
Page 30 of 44
3.3 Other important features of the environment Provide a description of the project area and the affected area, including information about the following features (where relevant to the project area and/or affected area, and to the extent not otherwise addressed above). If at Section 2.3 you identified any alternative locations, time frames or activities for your proposed action, you must complete each of the details below (where relevant) for each alternative identified. 3.3 (a) Flora and fauna
Christmas Island’s isolation, climate and the influence of land crabs have resulted in the development of distinct tropical rainforest vegetation, including some species that have evolved to be taller and larger than examples of the same species found elsewhere.
There are approximately 213 native plant species on Christmas Island, of which 17 are endemic (Claussen 2005). Holmes and Holmes (2002) list six endangered species, three vulnerable species, thirty three rare species and eleven as poorly known. A number of these species are listed as threatened under the EPBC Act.
The island’s terrestrial fauna is dominated by ecologically important and diverse land crabs. Red crabs (Gecarcoidea natalis) are the most numerous, and they act as keystone species because by regulating seedling recruitment, they influence structure and species composition of the island’s rainforest vegetation (Green et al. 1997, 2008). Red crabs also regulate the entry and spread of some invasive species in rainforest on the island (Lake and O’Dowd 1991, Green et al. 2011). The island also supports the world’s largest remaining population of the Robber Crab, Birgus latro (Drew et al. 2010).
Christmas Island is classified an Endemic Bird Area by BirdLife International, with nine nesting varieties /species of seabirds and around 80,000 resident birds. More than 100 seabird species have been recorded, including two endemic seabird species and one endemic subspecies that breed on the island. The Abbott's Booby Papasula abbotti (Listed as Endangered), Christmas Island Frigate Bird Fregata andrewsi, and the Golden Bosun Phaethon lepturus fulvus only nest on Christmas Island. There are seven endemic land birds, including the threatened Christmas Island Hawk-owl Ninox natalis, Christmas Island Thrush Turdus poliocaphalus erythropleurus, Christmas Island Goshawk Accipiter fasciatus natalis, and Christmas Island Emerald Dove Chalcophaps indica natalis.
The island is also important for other terrestrial fauna species. Five of the six recorded native reptile species are endemic; the Blue-tailed Skink Cryptoblepharus egeriae, Lister’s Gecko Lepidodactylus
listeri, the Forest Skink Emoia nativitatis, the Giant Gecko Cyrtodactylus sadlieri, the Coastal Skink Emoia atrocostata, and the Christmas Island Blind Snake Typhlops exocoeti. Lister’s gecko and the Christmas Island Blind Snake are listed under the EPBC Act as vulnerable. The range and abundance of Blue-tailed Skinks, Forest Skinks, Costal Skinks and Lister’s Geckos have dramatically contracted over the past decade and all are currently on the brink of extinction in the wild (Smith et al. unpublished results).
Five native endemic mammals have been recorded. The Bulldog Rat Rattus nativitatus, and Maclear’s Rat Rattus macleari, are extinct and the Christmas Island Shrew, Crocidura attenuata
trichura, is likely to be extinct. Murray’s Pipistrelle Bat Pipistrellus murrayi was once widespread but is now presumed extinct (Beeton et al. 2010). Yellow Crazy Ants are considered a threat to both the Pipistrelle Bat (Schultz and Lumsden 2004) and the Christmas Island Shrew (Schultz 2004). 3.3 (b) Hydrology, including water flows
Most rainforest on Christmas Island occurs over ancient limestone reefs of marine origin, which sit perched on basaltic bedrock. The soils are extremely porous, and Christmas Island has little flowing water. The few streams and soaks occur on the eastern and western coasts of the island, where the interface of the limestone and basalt occurs above sea level.

Environment Protection and Biodiversity Conservation Act 1999
Page 31 of 44
3.3 (c) Soil and Vegetation characteristics
The Christmas Island environment can be classified into 12 broad environments:
1) marine – ocean water, sand flats, caves, coral reefs and walls,
2) shoreline rock platforms,
3) beaches,
4) sea cliffs,
5) terrace forest,
6) shallow soil rainforest on the higher terraces,
7) limestone scree slopes and pinnacles, cliffs
8) deeper plateau and terrace soils rainforest,
9) mangrove forest,
10) perennially wet areas,
11) karst, comprising caves, overhangs, rock crevices, and sinkholes, and
12) mining field.
The biological control agent will be released at multiple locations in environments 5, 6 and 8, but will hopefully spread to 7, 9, 10 and 12 as well. 3.3 (d) Outstanding natural features
There are several mapped caves in karst environments on the island. These will not be affected by the introduction of a biological agent. 3.3 (e) Remnant native vegetation (from Draft Christmas Island Management Plan, Commonwealth of Australia 2012)
The island’s remoteness, climate and the influence of land crabs have resulted in the development of distinct tropical rainforest vegetation, including some species that have evolved to be taller and larger than examples of the same species found elsewhere.
There are at least 17 endemic plant species, including a rare fern Asplenium listeri, a tall treelike pandanus Pandanus elatus, and a palm Arenga listeri. There are also relict populations of mangrove species and cycads which have been left isolated by the island’s tectonic uplift. A number of these species are listed as threatened under the EPBC Act.
Many of the world’s tropical rainforests, particularly on islands in the Indian Ocean, the Pacific Ocean and South-East Asian regions, are under threat from human activities. This makes the park’s and the island’s rainforest ecosystems of increasingly significant conservation value.
Vegetation and species of significance will not be adversely affected by the introduction of the biological control agent.
3.3 (f) Gradient (or depth range if action is to be taken in a marine area)
N/A 3.3 (g) Current state of the environment Include information about the extent of erosion, whether the area is infested with weeds or feral animals and whether the area is covered by native vegetation or crops.

Environment Protection and Biodiversity Conservation Act 1999
Page 32 of 44
N/A 3.3 (h) Commonwealth Heritage Places or other places recognised as having heritage values
As for Section 3.1b
3.3 (i) Indigenous heritage values
Christmas Island was never been settled by Indigenous peoples prior to European settlement.
3.3 (j) Other important or unique values of the environment Describe any other key features of the environment affected by, or in proximity to the proposed action (for example, any national parks, conservation reserves, wetlands of national significance etc).
As for Section 1.4
3.3 (k) Tenure of the action area (eg freehold, leasehold)
Commonwealth Land
3.3 (l) Existing land/marine uses of area
National Park and Crown Land
3.3 (m) Any proposed land/marine uses of area
There are no additional proposed land uses for the area

Environment Protection and Biodiversity Conservation Act 1999
Page 33 of 44
4 Measures to avoid or reduce impacts Note: If you have identified alternatives in relation to location, time frames or activities for the proposed action at Section 2.3 you will need to complete this section in relation to each of the alternatives identified. Provide a description of measures that will be implemented to avoid, reduce, manage or offset any relevant impacts of the action. Include, if appropriate, any relevant reports or technical advice relating to the feasibility and effectiveness of the proposed measures. For any measures intended to avoid or mitigate significant impacts on matters protected under the EPBC Act, specify: what the measure is, how the measure is expected to be effective, and the time frame or workplan for the measure. Examples of relevant measures to avoid or reduce impacts may include the timing of works, avoidance of important habitat, specific design measures, or adoption of specific work practices. Provide information about the level of commitment by the person proposing to take the action to implement the proposed mitigation measures. For example, if the measures are preliminary suggestions only that have not been fully researched, or are dependent on a third party’s agreement (e.g. council or landowner), you should state that, that is the case. Note, the Australian Government Environment Minister may decide that a proposed action is not likely to have significant impacts on a protected matter, as long as the action is taken in a particular manner (section 77A of the EPBC Act). The particular manner of taking the action may avoid or reduce certain impacts, in such a way that those impacts will not be ‘significant’. More detail is provided on the Department’s web site. For the Minister to make such a decision (under section 77A), the proposed measures to avoid or reduce impacts must: clearly form part of the referred action (eg be identified in the referral and fall within the responsibility of the person
proposing to take the action), be must be clear, unambiguous, and provide certainty in relation to reducing or avoiding impacts on the matters
protected, and must be realistic and practical in terms of reporting, auditing and enforcement. More general commitments (eg preparation of management plans or monitoring) and measures aimed at providing environmental offsets, compensation or off-site benefits CANNOT be taken into account in making the initial decision about whether the proposal is likely to have a significant impact on a matter protected under the EPBC Act. (But those commitments may be relevant at the later assessment and approval stages, including the appropriate level of assessment, if your proposal proceeds to these stages).
Many parasitoid wasps are constrained by their evolution to be the enemies of a narrow range of hosts. In Section 2.5, it was demonstrated that the proposed biological control agent Tachardiaephagus somervillei is almost certainly a highly specialised enemy of scale insects in the family Kerriidae; all Tachardiaephagus species have a narrow host range and appear to be family specialists, known only to attack lac scale insects in the Kerriidae, the family to which the yellow lac scale Tachardina belongs (Table 2). Just two kerriid species occur on Christmas Island, the target Tachardina aurantiaca, and the false lobate lac scale Paratachardina pseudolobata. Both are introduced, exotic and invasive on Christmas Island. There is one host record of a closely related parasitoid Tachardiaephagus tachardiae from another Paratachardina species, P. lobata in India (Table 2). Even if the host range of Tachardiaephagus somervillei did extend to invasive Paratachardina on the island, this would have no negative consequences.
Based on the research conducted by La Trobe University the likelihood that Tachardiaephagus will have non-target impacts on any other species, including other scale insects, is considered remote. This view is supported by the Crazy Ant Scientific Advisory Panel (CASAP) (see Attachment 3). However, despite some spectacular successes elsewhere, the history of biocontrol has also been marred by a few failures when the biological control agent itself proved to be problematic. Most of these instances were not (unlike this proposal) based on pre-release research, involved the introduction of generalist or oligophagous predators prior to the establishment of regulatory frameworks for biological control introductions, or failed to consider the spread of the biological

Environment Protection and Biodiversity Conservation Act 1999
Page 34 of 44
control agent beyond the target area. In most cases, once a biological control agent has been released there is usually little that can be done to avoid or reduce unintended impacts. Clearly, such situations are best avoided entirely by making as sure as possible that Tachardiaephagus is as narrowly specific as the host records suggest (Table 2). Host specificity testing is required by Australia under the Biological Control Act 1984. However, no such regulatory framework has been identified that is currently applicable to the external territory of Christmas Island (end of Section 2.4). In the interim, La Trobe University will continue with plans to conduct pre-importation host-specificity trials with the assistance of cooperators at the Forest Research Institute of Malaysia at Kuala Lumpur and at Sarawak Forestry in Semenggoh.

Environment Protection and Biodiversity Conservation Act 1999
Page 35 of 44
5 Conclusion on the likelihood of significant impacts Identify whether or not you believe the action is a controlled action (ie. whether you think that significant impacts on the matters protected under Part 3 of the EPBC Act are likely) and the reasons why.
5.1 Do you THINK your proposed action is a controlled action?
No, complete section 5.2
Yes, complete section 5.3
5.2 Proposed action IS NOT a controlled action. Specify the key reasons why you think the proposed action is NOT LIKELY to have significant impacts on a matter protected under the EPBC Act.
The proposed biological control agent is almost certainly host-specific on one target invasive species
Non-target impacts, although highly unlikely, will almost certainly be restricted to one other exotic and invasive species, in the same subfamily as the target.
5.3 Proposed action IS a controlled action Type ‘x’ in the box for the matter(s) protected under the EPBC Act that you think are likely to be significantly impacted. (The ‘sections’ identified below are the relevant sections of the EPBC Act.) Matters likely to be impacted
World Heritage values (sections 12 and 15A)
National Heritage places (sections 15B and 15C)
Wetlands of international importance (sections 16 and 17B)
Listed threatened species and communities (sections 18 and 18A)
Listed migratory species (sections 20 and 20A)
Protection of the environment from nuclear actions (sections 21 and 22A)
Commonwealth marine environment (sections 23 and 24A)
Great Barrier Reef Marine Park (sections 24B and 24C)
Protection of the environment from actions involving Commonwealth land (sections 26 and 27A)
Protection of the environment from Commonwealth actions (section 28)
Commonwealth Heritage places overseas (sections 27B and 27C)
Specify the key reasons why you think the proposed action is likely to have a significant adverse impact on the matters identified above.

Environment Protection and Biodiversity Conservation Act 1999
Page 36 of 44
6 Environmental record of the responsible party NOTE: If a decision is made that a proposal needs approval under the EPBC Act, the Environment Minister will also decide the assessment approach. The EPBC Regulations provide for the environmental history of the party proposing to take the action to be taken into account when deciding the assessment approach. Yes No 6.1 Does the party taking the action have a satisfactory record of responsible
environmental management?
Provide details
The Commonwealth Director of National Parks is the responsible party for this action. The DNP manages several Commonwealth reserves on behalf of the Australian Government, in accordance with the EPBC Act and relevant park Management Plans. These reserves include the Christmas Island National Park, and the World Heritage listed Kakadu and Uluru Kata-Tjuta National Parks.
On matters pertaining to the management of the YCA invasion on Christmas Island, the DNP considers advice from the Crazy Ant Scientific Advisory Panel, comprised of researchers with expertise in invasive species biology including invasive ants, expertise with the local Island environment, and expertise in policy development, implementation and ecosystem management.
The on-ground research and implementation of the biocontrol program will continue to be the responsibility of La Trobe University, funded by the DNP. The La Trobe team has a very long history of research on the island, and two members were part of the team with DNP/CINP that won the 2003 Banksia Award for environmental management.
6.2 Has either (a) the party proposing to take the action, or (b) if a permit has been applied for in relation to the action, the person making the application - ever been subject to any proceedings under a Commonwealth, State or Territory law for the protection of the environment or the conservation and sustainable use of natural resources?
If yes, provide details
6.3 If the party taking the action is a corporation, will the action be taken in accordance with the corporation’s environmental policy and planning framework?
N/A
If yes, provide details of environmental policy and planning framework
6.4 Has the party taking the action previously referred an action under the EPBC Act, or been responsible for undertaking an action referred under the EPBC Act?

Environment Protection and Biodiversity Conservation Act 1999
Page 37 of 44
Provide name of proposal and EPBC reference number (if known)
Aerial baiting of Yellow Crazy Ant supercolonies;
(a) EPBC Referral 2002/722 (b) EPBC Referral 2009/5016 (c) EPBC Referral 2012/6438

Environment Protection and Biodiversity Conservation Act 1999
Page 38 of 44
7 Information sources and attachments (For the information provided above)
7.1 References List the references used in preparing the referral. Highlight documents that are available to the public, including web references if relevant.
Abbott, K.L. (2004). Alien ant invasion on Christmas Island, Indian Ocean: The role of ant-scale associations in the dynamics of supercolonies of the yellow crazy ant, Anoplolepis gracilipes. Ph.D Thesis, Monash University, Melbourne, Australia. http://arrow.monash.edu.au/vital/access/manager/Repository/monash:6496 (accessed 21 January 2013)
Abbott, K.L. (2005). Supercolonies of the invasive yellow crazy ant, Anoplolepis gracilipes, on an oceanic island: forager activity patterns, density and biomass. Insectes Sociaux 52: 266–273.
Abbott, K.L and Green, P.T (2007). Collapse of an ant-scale mutualism in a rainforest on Christmas Island. Oikos 116: 1238-1246.
Beeton, B., Burbidge, A., Grigg, G., How, R., McKenzie, N., and Woinarski, J. (2010). Final Report of the Christmas Island Expert Working Group to Minister for the Environment, Heritage and the Arts. Canberra, A.C.T., Australia. http://www.environment.gov.au/parks/publications/christmas/final-report.html (accessed 21 January 2013)
Boland, C.R.J., Smith, M.J., Maple, D.J., Tiernan, B., Barr, R., Reeves, R., and Napier, F. (2011). Heli-baiting using low concentration fipronil to control invasive yellow crazy ant supercolonies on Christmas Island, Indian Ocean. Pp. 152-156 in Veitch, C.R., Clout, M.N., and Towns, D.R. (eds.). Island invasives: eradication and management. Proceedings of the International Conference on
Island Invasives. Gland, Switzerland: IUCN and Auckland, New Zealand. http://www.issg.org/pdf/publications/island_invasives/pdfhqprint/2boland.pdf (accessed 21 January 2013)
Butcher, R. and Hale, J. (2010). Ecological character description for The Dales Ramsar site. Unpublished report to the Department of the Environment, Water, Heritage and the Arts, Canberra, A.C.T., Australia. http://www.environment.gov.au/water/publications/environmental/wetlands/61-ecd.html (accessed 22 January 2013)
Centre for Environment Stress and Adaptation (2011). Monitoring of the 2009 aerial baiting of yellow crazy ants (Anoplolepis gracilipes) on non-target invertebrate fauna on Christmas Island. Unpublished Report for the Director of National Parks, Canberra, A.C.T., Australia. http://www.environment.gov.au/parks/publications/christmas/fipronil-report.html (accessed 22 January 2013)
Claussen, J. (2005). Native plants of Christmas Island. Flora of Australia, Supplementary Series 22. Australian Government Department of the Environment and Heritage, Australian Biological Resources Study. Canberra, A.C.T., Australia.
Commonwealth of Australia (2006). Threat Abatement Plan to Reduce the Impacts of Tramp Ants on Biodiversity in Australia and its Territories. Department of the Environment and Heritage, Canberra, A.C.T., Australia.

Environment Protection and Biodiversity Conservation Act 1999
Page 39 of 44
http://www.environment.gov.au/biodiversity/threatened/publications/tap/trampants.html (accessed 21 January 2013)
Davis, N.E., O’Dowd, D.J., Green, P.T., and Mac Nally, R. (2008). Effects of alien ant invasion on abundance, behaviour, and reproductive success of endemic island birds. Conservation Biology 22: 1165-1176.
Davis, N.E., O'Dowd, D.J., Mac Nally, R., and Green, P.T. (2010). Invasive ants disrupt frugivory by endemic island birds. Biology Letters 6: 85-88.
Detto, T. and Tiernan, B. (2011). The 2011 island-wide survey report on Yellow Crazy Ants and Red Crabs. Unpublished report to Christmas Island National Park. Christmas Island.
Drew, M.M., Harzsch, S., Stensmyr, M., Erland, S., and Hansson, B.S. (2010). A review of the biology and ecology of the Robber Crab, Birgus latro (Linnaeus, 1767) (Anomura:Coenobitidae). Zoologischer Anzeiger 249: 45-67.
Giraud, T., Pedersen, J.S., and Keller, L. (2002). Evolution of supercolonies: the Argentine ants of southern Europe. Proceedings of the National Academy of Science USA 99: 6075–6079.
Green, P.T. Comport, S., and Slip, D. (2004). The management and control of the invasive alien crazy ant (Anoplolepis gracilipes) on Christmas Island, Indian Ocean: the aerial baiting campaign, September 2002. Unpublished final report to Environment Australia and the Crazy Ant Steering Committee.
Green, P.T. and O’Dowd, D.J. (2009). Management of invasive invertebrates: lessons from the management of an invasive alien ant. Pp. 153-172 in Clout, M.N. and Williams, P.A. (eds). Invasive Species Management: A Handbook of Principles and Techniques. Oxford University Press, Oxford.
Green, P.T., O’Dowd, D.J., Abbott, K.L., Jeffery, M., Retallick, K., and Mac Nally, R. (2011). Invasional meltdown: invader-invader mutualism facilitates a secondary invasion. Ecology 92: 1758-1768.
Green, P.T., O’Dowd, D.J., and Lake, P.S. (1997) Control of seedling recruitment by land crabs in rain forest on a remote oceanic island. Ecology 78: 2474-2486.
Green, P.T., O’Dowd, D.J., Lake, P.S. (2001). From resistance to meltdown: secondary invasion of an island rain forest. Pp. 451-455 in Ganeshaiah, K.N., Uma Shankar, R. and Bawa, K.S. (eds.). Tropical Ecosystems: Structure, Diversity and Human Welfare. Proceedings of the International Conference on Tropical Ecosystems, Bangalore, India. Oxford-IBH, New Delhi.
Green, P.T., O’Dowd D.J., and Lake, P.S. (2008). Recruitment dynamics in a rainforest seedling community: context-independent impact of a keystone consumer. Oecologia 156: 373-385.
Hale, J. and Butcher, R. (2010). Ecological Character Description for Hosnie’s Spring Ramsar Site. Unpublished report to the Department of the Environment, Water, Heritage and the Arts, Canberra, A.C.T., Australia. http://www.environment.gov.au/water/publications/environmental/wetlands/40-ecd.html (accessed 21 January 2013)
Holmes, J. and Holmes, G. (2002). Conservation status of the flora of Christmas Island, Indian Ocean. Unpublished report to Environment Australia/Parks Australia North. Glenn Holmes and Associates, Atherton, Queensland.
Kuhlmann, U., Schafner, U. and Mason, P.G. (2006). Selection of non-target species for host specificity testing. Pp. 15-37 in Bigler, F., Babendreier, D. and Kuhlmann, U. (eds.). Environmental

Environment Protection and Biodiversity Conservation Act 1999
Page 40 of 44
Impact of Invertebrates for Biological Control of Arthropods: Methods and Risk Assessment. CABI Publishing, Delemont, Switzerland.
Lake, P.S. and O’Dowd, D.J. (1991). Red crabs in rainforest, Christmas Island: biotic resistance to invasion by an exotic snail. Oikos 62: 25-29.
Lowe, S. Browne, M., and Boudjelas, S. (2000). 100 of the world’s worst invasive alien species. Aliens 12: S1-S12. http://www.issg.org/database/species/search.asp?st=100ss (accessed 21 January 2013)
Lumsden, L. Schulz, M., Ashton, R., and Middleton, D. (2007). Investigation of the threats to the Christmas Island Pipistrelle. Unpublished report to the Department of Environment and Water Resources, Canberra, A.C.T., Australia. http::// http://ausbats.org.au/#/resources/4553704436 (accessed 21 January 2013)
Marr, R.M., O’Dowd, D.J., and Green, P.T (2003). Assessment of non-target impacts of Presto® 01 ant bait on litter invertebrates in Christmas Island National Park, Indian Ocean. Unpublished report to Parks Australia North, Darwin, NT.
Neumann, G., Follett, P.A., Hollingsworth, R.G., and de León, J. (2010). High host specificity in Encarsia diaspidicola (Hymenoptera: Aphelinidae), a biological control candidate against the white peach scale in Hawaii. Biological Control 54: 107-113.
Neumann, G., Green, P.T. and O’Dowd, D.J. (2011). First record of Pulvinaria urbicola (Hemiptera: Coccidae), a potentially damaging scale insect, on Christmas Island, Indian Ocean. Unpublished report to Parks Australia. http://www.environment.gov.au/parks/publications/christmas/pubs/pulvinaria-report-2011.docx (accessed 21 January 2013)
O’Dowd, D.J. and Green, P.T. (2000). Design and feasibility of an island-wide survey of the invasive alien ant Anoplolepis gracilipes and its impact on Christmas Island, Indian Ocean. Unpublished report to Parks Australia North, Darwin, NT.
O’Dowd, D.J. and Green, P.T. (2010). Invasional meltdown: do invasive ants facilitate secondary invasions? Pp. 271-272 in Lach, L., Parr, C. & Abbott, K. (eds.) Ant Ecology. Oxford University Press, Oxford.
O’Dowd, D.J. Green, P.T., & Lake, P.S. (2003). Invasional ‘meltdown’ on an oceanic island. Ecology
Letters 6: 812-817. http://wolfweb.unr.edu/~ldyer/classes/396/odowd.pdf (accessed 21 January 2013)
O’Dowd, D.J., Green, P.T., and Lake, P.S. (1999). Status, impact, and recommendations for research and management of exotic invasive ants in Christmas Island National Park. Unpublished report to Environment Australia, Canberra, A.C.T., Australia http://www.issg.org/database/species/reference_files/Christmas_Island_Report.pdf (accessed 21 January 2013).
O’Dowd, D.J., Green, P.T., Neumann, G., and Wittman, S. (2012). Executive summary: research and development for indirect biological control of the yellow crazy ant (Anoplolepis gracilipes) on Christmas Island, Indian Ocean. Unpublished report to the Director of National Parks and the Crazy Ant Scientific Advisory Panel, 27 pp.
Palmer, W.A., Heard, T.A., and Sheppard, A.W. (2010). A review of Australian classical biological control of weeds programs and research activities over the past 12 years. Biological Control 52: 271-287. https://docs.google.com/a/monash.edu/viewer?a=v&q=cache:0-

Environment Protection and Biodiversity Conservation Act 1999
Page 41 of 44
K3mgvVy6oJ:northeast.landcarevic.net.au/oln/projects/soil-health-program/reference-documents/biocontrol-12-year-review/at_download/file+&hl=en&gl=au&pid=bl&srcid=ADGEESgGf1MZIbSjBgRLOJrC7PanstLsXFnzXToIee3sUdK7Dn6ilFKnFls9q89PVBlxOSUNgJKVCjUOAKMyOHG2WDsP2FkvkoWpsPpgoJUntwXSJ6gKs63DvfDQ2aotz4z4J67pwN5K&sig=AHIEtbTK8Q9OLsVwz8aVroMgFYlNViLokA (accessed 21 January 2013)
Schultz, M. (2004). National Recovery Plan for the Christmas Island Shrew Crocidura attenuata
trichura. Commonwealth of Australia, Canberra. http://www.environment.gov.au/biodiversity/threatened/publications/recovery/c-attenuata-trichura/index.html (accessed 21 January 2013)
Schulz, M. and Barker, C. (2008). A Terrestrial Reptile Survey of Christmas Island, May-June 2008. Unpublished report for Parks Australia North, Darwin, NT.
Schulz, M. and Lumsden, L.F. (2004). National Recovery Plan for the Christmas Island Pipistrelle
Pipistrellus murrayi. Commonwealth of Australia, Canberra. http://www.environment.gov.au/biodiversity/threatened/publications/recovery/p-murrayi/pubs/p-murrayi.pdf (accessed 21 January 2013)
Secord, D. and Karieva, P. (1996). Perils and pitfalls in the host specificity paradigm. BioScience 46: 448-453.
Smith, M.J., Boland, C.R.J., Maple, D.J., Scroggie, M., Tiernan B., and Napier. F. (unpublished results) The Christmas Island Red crab (Gecarcoidea natalis): temporal and spatial patterns in burrow counts.
Smith. M.J., Cogger, H., Tiernan, B., Maple, D., Boland, C., Napier, F., Detto, T., and Smith, P. (2012). An oceanic island reptile community under threat: the decline of reptiles on Christmas Island, Indian Ocean. Herpetological Conservation and Biology 7: 206-218.
Suhr, E.L., O’Dowd, D.J., Mackay, D.A., and McKechnie, S.W. (2011). Genetic structure, behaviour and invasion history of the Argentine ant in Australia. Evolutionary Applications 4: 471–484.
Sunamura, E., Espadaler, X., Sakamoto, H., Suzuki, S., Terayama, M. and Tatsuki, S. (2009). Intercontinental union of Argentine ants: behavioral relationships among introduced populations in Europe, North America, and Asia. Insectes Sociaux 56: 143–147.
Thomas, M.L., Becker, K., Abbott, K., and Feldhaar, H. (2010). Supercolony mosaics: two different invasions by the yellow crazy ant, Anoplolepis gracilipes, on Christmas Island, Indian Ocean. Biological Invasions 12: 677-687.
Wetterer, J.K. (2005). Worldwide distribution and potential spread of the long-legged ant, Anoplolepis
gracilipes (Hymenoptera: Formicidae). Sociobiology 45: 77-97.
7.2 Reliability and date of information For information in section 3 specify: source of the information; how recent the information is; how the reliability of the information was tested; and any uncertainties in the information.
Information sources: Full references are provided in Section 7.1 for the cited sources in Section 3. In addition, other information came from the Draft Christmas Island National Park Management Plan

Environment Protection and Biodiversity Conservation Act 1999
Page 42 of 44
(Commonwealth of Australia 2012), the Environment Protection and Biodiversity Conservation Act 1999 (Cwth), and the List of Commonwealth National Heritage places.
Information age: The information varies in age from 1991 through to 2012.
Information reliability: Studies published in scientific journals have been through a process of peer review and thus can be considered reliable. Unpublished reports have not been through this process, but can be considered as reliable because their authors are recognized as knowledgeable experts. This referral draws heavily on an unpublished report to the Director of Parks and the Crazy Ant Scientific Advisory Panel (O’Dowd et al. 2012). This report contains unpublished research, and in order to protect the publication rights of its authors this research should not be available to the public alongside the referral. The research was, however, formally peer-reviewed and endorsed by CASAP on 12 December 2012 (Attachment 3).
Information uncertainties: Uncertainties about the implementation and outcomes of the proposed action have been detailed in sections 2.1, 2.2, 2.4 and 2.5.
7.3 Attachments Indicate the documents you have attached. All attachments must be less than two megabytes (2mb) so they can be published on the Department’s website. Attachments larger than two megabytes (2mb) may delay the processing of your referral.
attached Title of attachment(s)
You must attach
figures, maps or aerial photographs showing the project locality (section 1)
Included within the referral document
figures, maps or aerial photographs showing the location of the project in respect to any matters of national environmental significance or important features of the environments (section 3)
Included within the referral document
If relevant, attach
copies of any state or local government approvals and consent conditions (section 2.5)
copies of any completed assessments to meet state or local government approvals and outcomes of public consultations, if available (section 2.6)
copies of any flora and fauna investigations and surveys (section 3)
technical reports relevant to the assessment of impacts on protected matters that support the arguments and conclusions in the referral (section 3 and 4)
report(s) on any public consultations undertaken, including with Indigenous stakeholders (section 3)

Environment Protection and Biodiversity Conservation Act 1999
Page 43 of 44
Other attachments Attachment 1. Indirect biological control of the yellow crazy ant on Christmas Island – host range testing of the proposed agent Tachardiaephagus
somervillei for the primary target Tachardina
aurantiaca. Attachment 2. Biological Control Implementation for Tachardina aurantiaca on Christmas Island. Attachment 3. CASAP advice to DNP on the feasibility of indirect biocontrol for the suppression of YCA supercolonies Attachment 4. Nomination of the Yellow lac scale as a biological control agent target

Environment Protection and Biodiversity Conservation Act 1999
Page 44 of 44
8 Contacts, signatures and declarations NOTE: Providing false or misleading information is an offence punishable on conviction by imprisonment and fine (s 489, EPBC Act). Under the EPBC Act a referral can only be made by: the person proposing to take the action (which can include a person acting on their behalf); or a Commonwealth, state or territory government, or agency that is aware of a proposal by a person to take an action,
and that has administrative responsibilities relating to the action1. Project title: Importation, rearing and release of Tachardiaephagus
somervillei (Hymenoptera: Encyrtidae) as a biological control agent for the yellow lac scale Tachardina aurantiaca (Hemiptera: Kerriidae) on Christmas Island, Indian Ocean
8.1 Person proposing to take action This is the individual, government agency or company that will be principally responsible for, or who will carry out, the proposed action. If the proposed action will be taken under a contract or other arrangement, this is:
the person for whose benefit the action will be taken; or the person who procured the contract or other arrangement and who will have principal control and
responsibility for the taking of the proposed action.
If the proposed action requires a permit under the Great Barrier Reef Marine Park Act2, this is the person requiring the grant of a GBRMP permission. The Minister may also request relevant additional information from this person. If further assessment and approval for the action is required, any approval which may be granted will be issued to the person proposing to take the action. This person will be responsible for complying with any conditions attached to the approval. If the Minister decides that further assessment and approval is required, the Minister must designate a person as a proponent of the action. The proponent is responsible for meeting the requirements of the EPBC Act during the assessment process. The proponent will generally be the person proposing to take the action3.
Name Peter Cochrane Title Commonwealth Director of National Parks. Organisation Director of National Parks (Christmas Island National Park) ACN / ABN (if applicable) 13 051 694 963 Postal address GPO Box 787 Canberra Act 2601
Telephone (02) 6274 2220 Email [email protected] Declaration
I declare that to the best of my knowledge the information I have given on, or attached to this form is complete, current and correct. I understand that giving false or misleading information is a serious offence.
1 If the proposed action is to be taken by a Commonwealth, state or territory government or agency, section 8.1 of this form should be completed. However, if the government or agency is aware of, and has administrative responsibilities relating to, a proposed action that is to be taken by another person which has not otherwise been referred, please contact the Referrals Business Entry Point (1800 803 772) to obtain an alternative contacts, signatures and declarations page. 2 If your referred action, or a component of it, is to be taken in the Great Barrier Reef Marine Park the Minister is required to provide a copy of your referral to the Great Barrier Reef Marine Park Authority (GBRMPA) (see section 73A, EPBC Act). For information about how the GBRMPA may use your information, see http://www.gbrmpa.gov.au/privacy/privacy_notice_for_permits. 3 If a person other than the person proposing to take action is to be nominated as the proponent, please contact the Referrals Business Entry Point (1800 803 772) to obtain an alternative contacts, signatures and declarations page.

Environment Protection and Biodiversity Conservation Act 1999
Page 45 of 44
I agree to be the proponent for this action. I acknowledge that I may be liable for fees related to my proposed action following the introduction of cost recovery under the EPBC Act.
Signature
Date 15 April 2013
8.2 Person preparing the referral information (if different from 8.1) Individual or organisation who has prepared the information contained in this referral form.
Name
1* - Drs Peter Green and Dennis O’Dowd 2* - Michael Misso
Title
1* - Senior Lecturer / Reader (retired) 2* - Manager, Christmas Island National Park
Organisation
1* - La Trobe University 2* - Parks Australia
ACN / ABN (if applicable)
1* - 64 804 735 113 2* - 13 051 694 963
Postal address
1* - Department of Botany, Biological Sciences Building 2 2* - Christmas Island National Park PO Box 867, Christmas Island, Indian Ocean.
Telephone
1* - 03 9479 3675 2* - 08 9168 8700
1* - [email protected] 2* - [email protected]
Declaration I declare that to the best of my knowledge the information I have given on, or attached to this form is complete, current and correct. I understand that giving false or misleading information is a serious offence.
Signature
Date
8/4/2013

Environment Protection and Biodiversity Conservation Act 1999
Page 46 of 44
REFERRAL CHECKLIST NOTE: This checklist is to help ensure that all the relevant referral information has been provided. It is not a part of the referral form and does not need to be sent to the Department. HAVE YOU:
Completed all required sections of the referral form?
Included accurate coordinates (to allow the location of the proposed action to be mapped)?
Provided a map showing the location and approximate boundaries of the project area?
Provided a map/plan showing the location of the action in relation to any matters of NES?
Provided complete contact details and signed the form?
Provided copies of any documents referenced in the referral form?
Ensured that all attachments are less than two megabytes (2mb)?
Sent the referral to the Department (electronic and hard copy preferred)?






PART A - Identifying and Analysing Project Management Risks Division Parks Australia Project Title Research and development for indirect biological control of the yellow crazy
ant (Anoplolepis gracilipes) on Christmas Island (Indian Ocean)
Branch /Section Christmas Island National Park Project Goal To suppress yellow crazy ant supercolonies on Christmas Island by the introduction of a host-specific parasitoid of its lac scale mutualist, Tachardina aurantiaca Date 21/11/2013
NOTE: In the Risk Assessment matrix below, the Consequence of The Risk can be judged in terms of:
1. Financial Consequences. For example, should the biocontrol program not proceed, the Consequence is recurrent funding for a
business-as-usual approach to YCA management dependent on surveillance, monitoring and control by toxic bait. 2. Indirect Biodiversity Consequences, which arise not from the implementation of an action itself or from its failure, but from the
knock-on effect of not implementing that action, or of it failing. For example, should the biocontrol program not proceed, the Consequence is continued negative impacts on elements of Island biodiversity, from either YCA or non-target impacts associated with the use of toxic bait.
3. Direct Biodiversity Consequences, which are a direct result of implementing an action. For example, off-target impacts of the
biocontrol agent imported to the island would have Direct Biodiversity Consequences.
For Risk 1 (below), No further action on the Biocontrol Program, we have combined both Financial and Indirect Biodiversity considerations when judging the severity of the Consequences. For Risks 2 – 5, we judged severity for Direct Biodiversity Consequences only.

In the Risk Assessment below,
1 2 3 4 5 6 7
Risk Ref/ID
The Risk
(What Can Happen?)
Source
(How can this Happen?)
Consequence
(What will happen if the risk occurs?)
Like
lihoo
d
Con
sequ
ence
Risk Level
1
No further action is taken to progress the biocontrol research and development project
a. Funding is insufficient to continue
a. Business as usual – management of YCA supercolonies continues to be reactive. Formation of new supercolonies cannot be prevented, and can only be controlled after they form. Continuing impacts of supercolony formation on biodiversity, especially red crabs and robber crabs. Continuing financial investment in the human resources and infrastructure (toxic bait and helicopter) to detect, map and control supercolonies. Continuing non-target impacts of the toxin, especially on robber crabs
Possible
Major
High
b. Research does not support the concept of indirect biocontrol – YCA is not dependent on honeydew, or suitable Biocontrol Agents (BCAs) are not available
b. As above Unlikely Major Medium

1 2 3 4 5 6 7
Risk Ref/ID
The Risk
(What Can Happen?)
Source
(How can this Happen?)
Consequence
(What will happen if the risk occurs?)
Like
lihoo
d
Con
sequ
ence
Risk Level
2 The imported BCA is not as host specific as predicted
Insufficient knowledge of the host specificity of the BCA
a. Other scale insects; In addition to the target species Tachardina aurantiaca, the BCA parasitizes other species of scale insects on the Island. All known species are non-native.
Very Unlikely
Insignificant Minimal
b. Other arthropods, including endemic insects such as stick insects and butterflies
Very Unlikely
Major Low
3 There is an endemic or native scale insect in the Family Kerridae on Christmas Island that has not yet been discovered
Insufficient field survey effort The BCA will parasitize a native species of scale insect
Very Unlikely
Major Low
4 The BCA does not achieve island-wide population control of Tachardina aurantiaca to the low levels seen in Southeast Asia
a. The climate on Christmas Island is wrong for the BCA
a. The BCA will not establish Very Unlikely
Insignificant Minimal
b. The BCA is suited to parasitize an ecotype of Tachardina from one part of its native range, but a different ecotype occurs on
b. The BCA will not establish well on Christmas Island
Very Unlikely
Insignificant Minimal

1 2 3 4 5 6 7
Risk Ref/ID
The Risk
(What Can Happen?)
Source
(How can this Happen?)
Consequence
(What will happen if the risk occurs?)
Like
lihoo
d
Con
sequ
ence
Risk Level
Christmas Island
c. The BCA is a poor disperser, and cannot move much beyond its initial point of release
c. The BCA will only achieve local population control of Tachardina, at points of release
Very Unlikely
Insignificant Minimal
d. Tending of Tachardina by YCA reduces parasitization by BCA
d. The incidence of parasitization on Tachardina by the BCA will be low
Unlikely Insignificant Minimal
e. The BCA was infected with a hyperparasitoid at importation
e. The population densities of the BCA will never be high enough to achieve control Tachardina
Very Unlikely
Insignificant Minimal
f. Marietta leopardina, already present on Christmas Island, acts as a hyperparasitoid for the BCA once it is introduced
f. The population densities of the BCA will be too low to achieve control of the target
Possible Insignificant Low

1 2 3 4 5 6 7
Risk Ref/ID
The Risk
(What Can Happen?)
Source
(How can this Happen?)
Consequence
(What will happen if the risk occurs?)
Like
lihoo
d
Con
sequ
ence
Risk Level
5
The BCA does not reduce the incidence of supercolony formation
Despite a large contribution by Tachardina, there is sufficient honeydew contributed by soft scales to sustain existing supercolonies, and permit the formation of new ones.
Supercolonies continue to form across the island, requiring business as usual surveillance, monitoring and control using toxic bait and condsiderable human resources
Possible Insignificant Low

PART B – Risk Register & Corrective Strategies 1For justification of risk, see Executive Summary (ES)
Risk Short title Risk Level Justification for Likelihood & Consequences1
Options for Risk Mitigation
1a Insufficient funding High Christmas Island National Park Plan of Management Expert Working Group Final Report Reports and Publications by La Trobe University and Monash University
EA to provide sufficient funding for the Biocontrol Program
1b Negative research findings Medium ES A1-A4 ES B1-B5
None
2a Non-target scale insects Minimal ES B4 Pre-importation host specificity trials conducted under laboratory conditions in Malaysia.
2b Non-target other insects Low ES B4 Pre-importation host specificity trials conducted under laboratory conditions in Malaysia.
3 Undiscovered native/endemic scale insect
Low ES B2 None. Field surveys demonstrate to operational certainty there are no native or endemic scale insects on Christmas Island
4a Climate matching for the BCA Minimal ES B3 & B5 Select donor populations of the BCA from areas that match most closely the local climate on Christmas Island
4b Ecotype matching for Tachardina
Minimal ES B3 Select donor populations of the BCA from Tachardina hosts that match most closely the local type on Christmas Island
4c Dispersal Minimal ES B3 Create “propagule pressure” by releasing the BCA at multiple sites and multiple times across Christmas Island
4d Scale tending by YCA Minimal ES B4 & B5 Select donor populations of the BCA from sites where Tachardina is tended by high densities of mutualistic ants, especially Anoplolepis gracilipes
4e Hyperparasitism of the BCA Minimal ES B4 Use standard laboratory rearing techniques to ensure the BCA is free of any hyperparasitoids prior to its importation to Christmas Island
4f Marietta acts as a hyperparasitoid of BCA
Low ES B4 None. However, there is no evidence of that Marietta attacks female Tachardina, either as a primary parasitioid or in order to attack the larvae of other primary parasitoids inside female Tachardina.
5 Biocontrol of Tachardina insufficient to suppress YCA supercolonies
Low ES A4 Develop and implement a biocontrol program for soft scale insects on Christmas Island to complement the program under development for Tachardina. Coccophagus ceroplastes and Encyrtis inflelix, both parasitoids of coccid scale insects, already occur on Christmas Island. Their rearing and release would not require a referral under the EPBC Act.