Embed Size (px)
Citation preview

1
RESEARCH ARTICLE 1
The Arabidopsis Nodulin Homeobox Factor AtNDX Interacts with 2
AtRING1A/B and Negatively Regulates Abscisic Acid Signaling 3
Yujuan Zhu1, 2#
, Xiaoying Hu1,,# Ying Duan
1, 3#, Shaofang Li
1, Yu Wang
1, Amin Ur 4
Rehman1, Junna He
1, Jing Zhang
1, Deping Hua
1, Li Yang
1, Li Wang
1, Zhizhong Chen
1, 5
Chuanyou Li4, Baoshan Wang
5, Chun-Peng Song
6, Qianwen Sun
7, Shuhua Yang
1, 6
Zhizhong Gong1, *
7
1State Key Laboratory of Plant Physiology and Biochemistry, College of Biological Sciences, 8 China Agricultural University, Beijing, 100193, China 9
2Lingnan Guangdong Laboratory of Modern Agriculture, Genome Analysis Laboratory of the 10 Ministry of Agriculture, Agricultural Genomics Institute at Shenzhen, Chinese Academy of 11 Agricultural Sciences, Shenzhen, 518100, China 12
3 Key Laboratory of Biology and Genetic Improvement of Horticultural Crops of the Ministry 13 of Agriculture, Institute of Vegetables and Flowers, Chinese Academy of Agricultural 14 Sciences, Beijing, 100081, China 15
4State Key Laboratory of Plant Genomics, National Centre for Plant Gene Research, Institute 16 of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, 17 China 18
5Key Lab of Plant Stress Research, College of Life Science, Shandong Normal University, 19 Ji’nan 250000, China 20
6Collaborative Innovation Center of Crop Stress Biology, Henan Province, Institute of Plant 21 Stress Biology, Henan University, Kaifeng 475001, China 22
7Center for Plant Biology and Tsinghua-Peking Joint Center for Life Sciences, School of Life 23 Sciences, Tsinghua University, Beijing 100084, China 24
#: These authors contributed equally to this work. 25
*Corresponding author: [email protected] 26
27
Short title: AtNDX and PRC1 core components AtRING1A/B in ABA signaling 28
One-sentence summary: Arabidopsis nodulin homeobox protein AtNDX interacts with the 29 Polycomb Repressive Complex 1 (PRC1) core components AtRING1A and AtRING1B and 30 negatively regulates ABI4 expression during ABA signaling. 31
The author responsible for distribution of materials integral to the findings presented in this 32 article in accordance with the policy described in the Instructions for Authors 33 (www.plantcell.org) is Zhizhong Gong, [email protected]. 34
35
ABSTRACT 36
The phytohormone abscisic acid (ABA) and the Polycomb group (PcG) proteins have 37
key roles in regulating plant growth and development; however, their interplay and 38
underlying mechanisms are not fully understood. Here, we identified an Arabidopsis 39
thaliana nodulin homeobox (AtNDX) protein as a negative regulator in the ABA 40
signaling pathway. AtNDX mutants are hypersensitive to ABA, as measured by 41
Plant Cell Advance Publication. Published on January 9, 2020, doi:10.1105/tpc.19.00604
©2020 American Society of Plant Biologists. All Rights Reserved

2
inhibition of seed germination and root growth, and the expression of AtNDX is 42
downregulated by ABA. AtNDX interacts with the Polycomb Repressive Complex 1 43
(PRC1) core components AtRING1A and AtRING1B in vitro and in vivo, and 44
together, they negatively regulate the expression levels of some ABA-responsive 45
genes. We identified ABA-INSENSITIVE (ABI4) as a direct target of AtNDX. AtNDX 46
directly binds the downstream region of ABI4 and deleting this region increases the 47
ABA sensitivity of primary root growth. Furthermore, ABI4 mutations rescue the 48
ABA-hypersensitive phenotypes of ndx mutants and ABI4-overexpressing plants are 49
hypersensitive to ABA in primary root growth. Thus, our work reveals the critical 50
functions of AtNDX and PRC1 in some ABA-mediated processes and their regulation 51
of ABI4. 52
53
INTRODUCTION 54
The phytohormone abscisic acid (ABA) regulates a variety of developmental 55
processes including seed dormancy, seed germination, seedling growth, flowering and 56
plant senescence (Cutler et al., 2010; Zhu, 2016; Nonogaki, 2019). ABA also plays 57
crucial and generally protective roles in various abiotic stress responses including 58
drought, high salt, and low temperature (Cutler et al., 2010; Zhu, 2016; Qi et al., 59
2018). In the ABA signaling pathway, after perceiving ABA, ABA receptors 60
PYR/PYLs/RCARs interact with and inhibit the negative regulators of the clade 61
A-type protein phosphatases 2C (PP2Cs), which releases the inhibitory effect of these62
PP2Cs on downstream protein kinases such as SnRK2 (SUCROSE 63
NONFERMENTING1-RELATED PROTEIN KINASES2).2/2.3/2.6 (OST1, OPEN 64
STOMATA1), GUARD CELL HYDROGEN PEROXIDE-RESISTANT1 (GHR1) 65
and calcium-dependent protein kinases (CPKs) (Ma et al., 2009; Park et al., 2009; 66
Cutler et al., 2010; Zhu, 2016) (Hua et al., 2012; Qi et al., 2018). Some protein 67
kinases such as SnRKs then phosphorylate and activate downstream targets such as 68
the SLOW ANION CHANNEL-ASSOCIATED 1 (SLAC1), which regulates stomatal 69
movement, or transcription factors such as the basic leucine zipper (bZIP)-family 70
proteins ABFs (ABRE binding factors)/AREBs (including ABF1, ABF2/AREB1, 71
ABF3 and ABF4/AREB2) (Choi et al., 2000; Uno et al., 2000) and ABI5 (ABA 72
INSENSITIVE 5) (Finkelstein and Lynch, 2000), which regulate the expression of 73
ABA-responsive genes. 74

3
In addition to bZIP transcription factors, many other transcription factors such as 75
MYB, MYC, NAC (NAM, ATAF1/2 and CUC2), WRKY (The 76
tryptophan-arginine-lysine-tyrosine), ZFHD1 (ZINC-FINGER HOMEODOMAIN), 77
NF-Y (NUCLEAR FACTOR-Y) and ARF2 (AUXIN RESPONSE FACTOR 2) are 78
involved in the ABA signaling pathway (Ren et al., 2010; Fujita et al., 2011; Wang et 79
al., 2011; Rushton et al., 2012; Singh and Laxmi, 2015; Promchuea et al., 2017). 80
Early studies of ABA-insensitive (ABI) mutants in seed germination identified several 81
genes such as ABI1, ABI2, ABI3, ABI4, and ABI5. ABI1 and ABI2, which encode 82
PP2Cs, are key negative regulators in the early ABA signaling pathway. ABI3 83
encodes a transcription factor with a B3 domain that shares high sequence similarity 84
with maize (Zea mays) Viviparous1 (Giraudat et al., 1992). ABI4 encodes an 85
APETALA2 (AP2)-type transcription factor (Finkelstein et al., 1998), which can bind 86
to target genes by recognizing CE1-like cis-elements (CACCG and CCAC motif) and 87
act as either an activator or a repressor depending on the context (Kerchev et al., 2011; 88
Shu et al., 2013; Wind et al., 2013; Huang et al., 2017). ABI4 expression is hormone 89
regulated, and its protein stability is affected by ABA and gibberellic acid (GA) in 90
opposite ways (Shkolnik-Inbar and Bar-Zvi, 2010; Shu et al., 2016). Several 91
transcription factors have been reported to directly regulate ABI4 expression, such as 92
ABI4 itself and WRKY8, which positively regulate ABI4 expression (Bossi et al., 93
2009; Chen et al., 2013), and RAV1 (Related to ABI3/VP1) (Feng et al., 2014) and 94
BASIC PENTACYSTEINE (BPC) family proteins, which negatively regulate ABI4 95
(Mu et al., 2017). BPCs can bind the ABI4 promoter and recruit Polycomb Repressive 96
Complex 2 (PRC2), thus repressing ABI4 expression through the histone H3 lysine 27 97
trimethylation (H3K27me3) epigenetic modification (Mu et al., 2017). 98
Polycomb group proteins (PcGs) are the major epigenetic machinery executing 99
transcriptional repression and developmental regulation in animals and plants 100
(Calonje, 2014; Mozgova and Hennig, 2015; Zhou et al., 2018). The two best- 101
characterized PcG complexes to date are PRC1 and PRC2 (Mozgova and Hennig, 102
2015). PRC1 deposits histone H2A monoubiquitination (H2Aub) and mediates 103
chromatin compaction of its target genes (Calonje, 2014; Wang and Shen, 2018), and 104
PRC2 catalyzes H3K27me3 (Mozgova and Hennig, 2015). PRC1 components were 105
initially identified in Drosophila melanogaster by genetic approaches. The canonical 106
Drosophila PRC1 consists of Polycomb (Pc), Polyhomeotic (Ph), Posterior sex combs 107

4
(Psc), and dRing1, also known as Sex combs extra (Sce). These have multiple 108
homologs in mammals, resulting in different possible combinations of PcGs (Shao et 109
al., 1999; Francis et al., 2001). Arabidopsis thaliana has no Ph protein, and the LIKE 110
HETEROCHROMATIN PROTEIN 1 (LHP1), a plant homolog of animal HP1, 111
functions like Pc in recognizing and colocalizing with H3K27me3 genome-wide 112
(Turck et al., 2007; Zhang et al., 2007). EARLY BOLTING IN SHORT DAYS (EBS) 113
and SHORT LIFE (SHL) interact with EMBRYONIC FLOWER 1 (EMF1), a 114
functional equivalent of the Psc C-terminal region (Psc-CTR), to form a bivalent 115
chromatin reader complex for both H3K27me3 and H3K4me3 that mediate switching 116
between repressed and active chromatin during plant development (Li et al., 2018; 117
Yang et al., 2018). The most conserved PRC1 components in plants are the 118
RING-finger proteins RING1 and BMI1, corresponding to dRing1 and the N-terminus 119
of Psc in Drosophila, respectively (Molitor and Shen, 2013; Merini and Calonje, 120
2015). In Arabidopsis, two RING1 homologs (AtRING1A and AtRING1B) and three 121
BMI1 homologs (AtBMI1A, AtBMI1B and AtBMI1C) have been identified based on 122
a unique domain architecture that contains a conserved RING finger domain in the 123
N-terminal region and a ubiquitin-like domain named RAWUL (Ring-finger And 124
WD40 associated Ubiquitin-Like) in the C-terminal region (Sanchez-Pulido et al., 125
2008; Xu and Shen, 2008). AtRING1A and AtRING1B interact with themselves, with 126
each other, and with LHP1, EMF1, AtBMI1A, AtBMI1B, and AtBMI1C (Wang and 127
Shen, 2018). The two AtRING1A/B proteins and the three AtBMI1A/B/C proteins 128
have been shown to have E3 ligase activity in vitro and in vivo (Bratzel et al., 2010; 129
Bratzel et al., 2012). 130
Target selection by PcGs at various developmental stages is critical for the 131
functions of targets. In Arabidopsis, both cis-element and DNA binding factors are 132
required for recruiting PRC2 (Xiao et al., 2017). Besides the classical hierarchical 133
model in which PRC1 is recruited by the binding of Pc to the H3K27me3 marker 134
deposited by PRC2, numerous factors involved in the recruitment of PRC1 have 135
emerged in mammals (Blackledge et al., 2015). In plants, many proteins interact with 136
PRC1 components EMF1, LHP1, AtRING1A and AtBMI1A/B/C (Li et al., 2016; 137
Wang and Shen, 2018; Zhu et al., 2019). AtRING1A interacts with ALFIN1-like 138
PHD-domain H3K4me3-binding protein AL6 and directly regulates the expression of 139
ABI3 and DELAY OF GERMINATION 1 (DOG1 ) (Molitor et al., 2014), CLF 140

5
(CURLY LEAF, a PRC2 component catalyzing H3K27-methylation) (Xu and Shen, 141
2008), and FAS1 (Chromatin Assembly Factor-1 subunit) during DNA replication 142
(Jiang and Berger, 2017). The dissection of different PcG component regulons reflects 143
the important roles of PRC1 in repressing gene expression during seed maturation and 144
vegetative development, and suggests that DNA binding protein may play a crucial 145
role in recruiting PcG to specific targets in plants (Wang and Shen, 2018). Until now, 146
however, no transcription factor has been identified that interacts with AtRING1A/B 147
(Wang and Shen, 2018). 148
In our previous studies, we used a root-bending assay to study plant response to 149
ABA, and we identified several ABA overly-sensitive (abo) mutants that show 150
hypersensitivity to ABA with respect to inhibition of primary root growth (Liu et al., 151
2010; Ren et al., 2010; Wang et al., 2011; He et al., 2012; Yang et al., 2014; 152
Promchuea et al., 2017; Wang et al., 2018). The cloned genes include ABO3, which 153
encodes a WRKY transcription factor (Ren et al., 2010); ARF2 (Wang et al., 2011), 154
ABO5, and ABO8, which encode two pentatricopeptide repeat proteins localized in 155
mitochondria (Liu et al., 2010; Yang et al., 2014); ABO6, which encodes a 156
DEXH-box RNA helicase in mitochondria (He et al., 2012); and EAR1 (ENHANCER 157
OF ABA CO-RECEPTOR1), which encodes a novel protein that enhances the activity 158
of PP2Cs (Wang et al., 2018). 159
Here we characterized two mutant alleles of an Arabidopsis thaliana nodulin 160
homeobox homologous gene, AtNDX. NDX was previously isolated from a soybean 161
(Glycine max) nodule-specific expression library (Jorgensen et al., 1999) and 162
represents a small gene family in different plants. The Arabidopsis thaliana genome 163
has only one AtNDX gene. AtNDX can bind the single-stranded DNA at the 3′end of 164
Arabidopsis FLOWERING LOCUS C (FLC) and control expression of FLC and the 165
antisense transcript COOLAIR (Sun et al., 2013). Our results demonstrate that the 166
expression of AtNDX is downregulated by ABA and that AtNDX directly interacts 167
with AtRING1A and AtRING1B. These proteins co-regulate the expression of some 168
common ABA-responsive genes. Further, AtNDX directly binds the downstream 169
region of ABI4 and represses its expression, and mutation of ABI4 could recover the 170
ABA-hypersensitive phenotype of ndx mutants in both seed germination and primary 171
root growth. 172
173

6
RESULTS 174
AtNDX is a negative regulator of ABA-mediated inhibition of seed germination 175
and primary root growth. 176
A root-bending assay (Yin et al., 2009) was employed to screen for ABA 177
overly-sensitive mutants in an ethyl methyl sulfonate (EMS)-mutagenized 178
Arabidopsis M2 population, in which we identified two ABA overly-sensitive mutant 179
alleles, ndx-5 and ndx-6. Genetic analysis indicated that ndx-5 and ndx-6 are recessive 180
mutations in a single nuclear gene. ndx-5 and ndx-6 were backcrossed to wild-type 181
Columbia four times before performing the following analyses. 182
First, we quantified the ABA-induced inhibition of primary root growth in ndx 183
mutants and the wild type. The primary root growth was measured after 5-d-old 184
seedlings were moved from Murashige and Skoog (MS) medium to MS medium or 185
MS medium supplemented with different concentrations of ABA. The ndx mutants 186
and the wild type showed no difference in primary root growth on MS medium. 187
However, ABA-induced inhibition of root growth was significantly more severe in 188
ndx mutants than in the wild type. Here, we used ear1-1 for comparison of ABA 189
sensitivity (Figure 1A and 1B) (Wang et al., 2018). 190
Root growth integrates root cell division, differentiation, and elongation. 191
Therefore, we further determined the meristem zone (MZ) cell number and 192
differentiation zone (DZ) cell length in ndx-5 and wild type, which reflect the root cell 193
division activity and mature cell size, respectively. Consistent with the primary root 194
growth, both the MZ cell number and DZ cell length were inhibited by ABA to a 195
greater extent in the ndx mutant than in the wild type (Supplemental Figure 1A-1D). 196
Then we investigated the ABA sensitivity of the seed germination and seedling 197
establishment in ndx mutants and the wild type. With increasing concentrations of 198
ABA, the percentage of seedlings with green cotyledons in ndx mutants was reduced 199
significantly more than those of wild type, while there was no obvious difference 200
between ndx and wild type on MS medium without ABA (Figure 1C-1D, 201
Supplemental Figure 2A-2D). 202
We also detected the expression of ABA response genes, including ABI3 and 203
ABI4, and found the expression levels of these two genes were higher in ndx mutants 204
than the wild type (Supplemental Figure 2E). However, we did not find any difference 205

7
between the wild type and ndx mutants in a detached leaf water loss assay and 206
stomata number on leaves (Supplemental Figure 2F and 2G), suggesting that NDX is 207
not involved in water loss under drought stress. These results demonstrate that 208
AtNDX is a negative regulator of ABA signaling in seed germination and primary 209
root growth, but not in ABA-promoted stomatal closure. 210
Isolation of AtNDX by map-based cloning 211
We identified the mutation in ndx-5 by map-based cloning. AtNDX was delimited to 212
BAC clones T4I9 and F4C21 on chromosome IV (Supplemental Figure 3A). 213
Sequencing of putative genes in this region identified a G1911 to A1911 mutation 214
(counting from the first putative ATG of AT4G03090) in AT4G03090 (previously 215
named AtNDX, Supplemental Figure 3A) (Mukherjee et al., 2009). We compared the 216
sequences amplified from cDNAs of ndx-5 and the wild type and found that the point 217
mutation affected the acceptor splicing between the seventh intron and the eighth 218
exon and created a new acceptor site upstream of the original one, resulting in an 219
early putative stop codon in the new transcript (Supplemental Figure 3B). 220
Map-based cloning narrowed down the ndx-6 mutation to the same region as ndx-5. 221
Sequencing of AT4G03090 in ndx-6 revealed a mutation from G569 to A569, which is 222
suspected to change the donor splicing site between the third exon and third intron 223
from GT to AT. Transcript analyses indicated that cDNA from ndx-6 contained an 224
extra third intron that creates an early putative stop codon in the new transcript 225
(Supplemental Figure 3C). We also obtained a previously described AtNDX mutant, 226
enhancer of coolair1-4 (eoc1-4), here renamed ndx-4 (Sun et al., 2013). In the ndx-4 227
mutant, AtNDX is disrupted by a T-DNA insertion (Supplemental Figure 3A) (Sun et 228
al., 2013). ndx-4 showed similar ABA sensitivity to ndx-5 with respect to root growth 229
and seedling establishment (Figure 1A-1D). 230
Immunoblot assays using NDX antibodies indicated that the AtNDX level was 231
significantly reduced in the ndx-4, ndx-5 and ndx-6 mutants compared to the wild type, 232
GST-AtNDX-C from Escherichia coli was used as the positive control to evaluate the 233
specificity of NDX antibodies (Supplemental Figure 3E). Given that the phenotypes 234
of ndx-5 are weaker than those of ndx-6 for both flowering time (Supplemental Figure 235
3F) and ABA sensitivity, ndx-5 is assumed to produce a truncated protein with some 236
residual function. 237

8
An 8-kb fragment that includes the wild-type AT4G03090 gene and contains 238
about 3000 bp upstream of the first putative ATG to 1000 bp downstream of the 239
putative TGA stop codon complemented the ABA-sensitive phenotypes of ndx-5 in 240
several independent lines (two independent lines are shown in Supplemental Figure 241
4A and 4B). In addition, overexpressing AtNDX-MYC under control of a 35S promoter 242
or AtNDX-Flag driven by the native promoter also complemented the ABA-sensitive 243
root phenotype of ndx-5 or ndx-4 (Supplemental Figure 4C-4F). However, we found 244
that these overexpression lines were not more resistant to ABA than the wild type in 245
terms of primary root growth, suggesting that only a certain amount of AtNDX 246
protein is needed for its full functioning in ABA-mediated primary root growth, or the 247
protein did not accumulate much, as AtNDX is likely regulated by 26S proteasome 248
pathway as shown below. These results indicate that AT4G03090 is the AtNDX gene. 249
ABA negatively regulates the expression of AtNDX 250
To elucidate the role of AtNDX in the ABA response, we first examined the effect of 251
ABA treatment on AtNDX expression and AtNDX protein levels. Transcript analysis 252
by reverse transcription-quantitative PCR (RT-qPCR) showed that the expression of 253
AtNDX was significantly repressed by ABA (Figure 2A). We then assayed transgenic 254
plants transformed with the native AtNDX promoter (proAtNDX) driving a GUS 255
reporter gene and found that ABA treatment greatly reduced GUS expression as 256
observed by GUS staining (Figure 2B). We also observed GFP fluorescence in 257
transgenic plants carrying proAtNDX:AtNDX-GFP and found that 30 μM ABA 258
treatment significantly reduced GFP expression (Figure 2C and 2D). 259
To explore whether the AtNDX protein level is affected by ABA, we analyzed the 260
total protein extracts from seedlings with or without ABA treatment. Immunoblotting 261
analysis with anti-NDX antibodies indicated that endogenous AtNDX protein was 262
reduced under ABA treatment. When we used intracellular protein synthesis inhibitor 263
cycloheximide (CHX) or CHX plus ABA to treat the seedlings to block protein 264
biosynthesis in order to see the ABA effect on AtNDX protein, the protein reduction 265
caused by ABA was no longer detected (Figure 2E). These results indicate that ABA 266
negatively regulates AtNDX at the transcriptional level, but not at the 267
posttranscriptional level. At the same time, we found that AtNDX was accumulated 268
when seedlings were treated with the proteasome inhibitor MG132. This result 269
suggests that AtNDX is regulated by the 26S proteasome degradation pathway (Figure 270

9
2E). Here, GST-AtNDX-C was used as a positive control. 271
AtNDX interacts with PRC1 core components AtRING1A and AtRING1B 272
To further clarify the working mechanism of AtNDX, we attempted to identify the 273
proteins interacting with AtNDX through a yeast two-hybrid screening. A normalized 274
Arabidopsis yeast two-hybrid cDNA library was used as the prey, and screened with 275
binding domain (BD) fused with AtNDX as the bait. Two clones encoding AtRING1A 276
fragments and one clone encoding an AtRING1B fragment were identified among 277
about 100 sequenced clones encoding putative interactors. We then verified their 278
interaction with AtNDX using the full-length AtRING1A or AtRING1B in the yeast 279
two-hybrid assay (Figure 3A). 280
We next tested whether AtNDX and AtRING1A or AtRING1B interact in vivo by 281
using a co-immunoprecipitation (Co-IP) assay. Proteins were extracted from 282
protoplasts transiently expressing HA-Flag-AtNDX and AtRING1A- or 283
AtRING1B-GFP and used for Co-IP. The results indicated that AtRING1A-GFP and 284
AtRING1B-GFP co-immunoprecipitated HA-Flag-AtNDX (Figure 3B). Then, we 285
used GFP beads to immunoprecipitate proteins extracted from 10-d-old transgenic 286
seedlings (two lines) containing AtRING1A-GFP driven by a super promoter to detect 287
whether the endogenous AtNDX protein could coimmunoprecipitate with 288
AtRING1A-GFP. An immunoblot assay with anti-NDX antibodies showed that 289
endogenous AtNDX protein coimmunoprecipitated with AtRING1A-GFP (Figure 290
3C). 291
To examine the natural interaction between AtNDX and AtRING1A/B, we used 292
anti-NDX antibodies in immunoprecipitation experiments to identify 293
AtNDX-associated proteins from 10-d-old seedlings. Following mass spectrometry 294
analysis, we identified AtRING1A and AtBMI1A, suggesting that AtNDX is 295
associated with PRC1 core components in vivo (Figure 3D). Therefore, we tested 296
whether there were direct interactions between AtNDX and other PRC1 components 297
such as AtBMI1A, AtBMI1B, AtBMI1C, AtEMF1 or LHP1 using the yeast 298
two-hybrid assay. However, we did not find any interaction (Supplemental Figure 5). 299
To determine which parts of AtRING1A/B and AtNDX confer their physical 300
interaction, different deletions were tested in the yeast two-hybrid assay. Deletion 301
analysis revealed that the C-terminus of AtRING1A/B, containing the RAWUL 302

10
domain, is sufficient to bind AtNDX (Figure 3E), while the full-length AtNDX protein 303
is required for their interaction (Figure 3F). Together, our data demonstrate that 304
AtNDX interacts with AtRING1A/B in vitro and in vivo. 305
The atring1a atring1b double mutants are hypersensitive to ABA in primary root 306
growth and seedling establishment 307
To investigate the role of AtRING1A/B in the ABA response, we examined the 308
phenotype of atring1a and atring1b single and double mutants on MS medium 309
containing ABA. Single mutants for atring1 or atbmi1 do not show apparent 310
morphological phenotypes except for a late flowering phenotype of atring1a (Shen et 311
al., 2014). However, the atring1a/b and atbmi1a/b double mutants displayed strong 312
embryonic defects during post-germination (see Supplemental Figure 6 for mutant 313
information), which is consistent with the derepression of several key regulatory 314
genes implicated in embryogenesis and stem cell activity (Xu and Shen, 2008; Bratzel 315
et al., 2010; Chen et al., 2010). The expression of AtRING1A/B was not affected in 316
ndx mutants and the expression of AtNDX was not affected in the ring mutants 317
(Supplemental Figure 6B-6D). The atring1a and atring1b-2 single mutants displayed 318
similar ABA sensitivity to the wild type (Supplemental Figure 7A-7D and 8A-8D). 319
We then selected 5-day-old atring1a atring1b double mutants according to the 320
abnormal cotyledon phenotypes on MS medium and transferred them onto medium 321
supplemented with or without ABA for 4 days. Although atring1a atring1b mutants 322
displayed various root lengths on MS medium, their relative root growth (i.e., growth 323
on ABA medium relative to growth on ABA-free medium) was significantly more 324
sensitive to ABA than that of the wild type (Figure 4A-4C). 325
Since the atring1a atring1b double mutant is sterile, we were unable to test the 326
seed cotyledon greening phenotype on ABA-containing medium. We then used a 327
weak double mutant, ring1a-2 ring1b-3, in which a T-DNA fragment is inserted in the 328
promoter region of AtRING1A, resulting in reduced expression of AtRING1A 329
compared to the wild type and a more uniform phenotype (Supplemental Figure 6) (Li 330
et al., 2017). The atring1a-2 atring1b-3 double mutant was significantly more 331
sensitive to ABA with respect to inhibition of seedling establishment than the wild 332
type, and ear1-1 served as the ABA hypersensitive control (Figure 4D-4G). In 333
conclusion, these results revealed that AtRING1A and AtRING1B negatively regulate 334
ABA responses with respect to seedling establishment. 335

11
To examine the genetic interaction between AtNDX and AtRING1A/B, we 336
constructed ndx-5 atring1a and ndx-5 atring1b-2 double mutants and tested their 337
phenotypes on ABA medium. The ndx-5 atring1a mutant displayed slightly but not 338
significantly increased ABA sensitivity with respect to inhibition of root growth and 339
substantially increased ABA sensitivity with respect to inhibition of seedling 340
establishment (Supplemental Figure 7A-7D), while the ndx-5 atring1b-2 double 341
mutant showed similar sensitivity to the ndx-5 single mutant (Supplemental Figure 342
8A-8D). The ndx-5 atring1a double mutant even exhibited shorter roots and smaller 343
cotyledons on MS medium compared with wild type (Supplemental Figure 7C). The 344
additive effects of AtNDX and AtRING1A could be due to each protein having 345
specific additional targets. Our data suggest that AtRING1A plays a broader, more 346
important role than AtRING1B as demonstrated by its role in flowering control (Shen 347
et al., 2014). Collectively, the above results indicate that the AtNDX and 348
AtRING1A/B proteins are involved in some ABA-mediated responses. 349
AtNDX and AtRING1A/B co-regulate the expression of ABI4 and some 350
ABA-responsive genes 351
Because AtNDX and AtRING1A/B interact with each other in vivo and both mutants 352
are hypersensitive to ABA in seed germination, seedling establishment and root 353
growth, we speculated that they might regulate some common genes involved in ABA 354
responses. Therefore, we performed RNA-seq experiments to profile the 355
transcriptome of ndx-4, atring1a atring1b and the wild type under different conditions. 356
We used ndx-4 because it is a T-DNA insertion mutant with a cleaner genetic 357
background than the EMS mutants (ndx-5 and ndx-6). Total RNAs were isolated from 358
7-d-old seedlings treated with or without ABA for 3 h and used for mRNA-seq library 359
construction. We sequenced two biological replicates for each sample on an Illumina 360
NovaSeq 6000 platform and obtained more than 30 million paired-end clean reads for 361
each replicate. RNA-seq reads were mapped back to the TAIR10 genome, and gene 362
expression levels were quantified using the software cuffdiff (Trapnell et al., 2013). 363
Differentially expressed genes (DEGs) were filtered with FDR < 0.05 and fold 364
change >2 and required to have an expression higher than 1 FPKM (Trapnell et al., 365
2013). 366
Under normal conditions (i.e., no ABA treatment), 384 and 1798 genes were 367
upregulated in ndx-4 and atring1a atring1b, respectively, while 54 and 1409 genes 368

12
were downregulated in ndx-4 and atring1a atring1b, respectively, compared with the 369
wild type (Figure 5A, Supplemental Data Set 1, Sheet 1-11). Among these genes, 252 370
were co-upregulated and 22 were co-downregulated in the two mutants (Figure 5A). 371
We classified the genes co-regulated by AtNDX and AtRING1A/B under normal 372
conditions and found that these genes encode proteins with various functions, most of 373
them annotated as stimulus/stress responses and developmental processes (Figure 5B). 374
Under ABA treatment, 279 and 1467 genes were upregulated while 73 and 2382 genes 375
were downregulated in ndx-4 and atring1a atring1b, respectively, compared with the 376
wild type (Figure 5C). Among them, 163 genes were co-upregulated and 36 genes 377
were co-downregulated in the two mutants (Figure 5C). 378
To identify the genes that lead to ABA-hypersensitive phenotypes in the ndx and 379
atring1a atring1b mutants, we examined the overlap between the co-regulated genes 380
in the two mutants under normal conditions and the genes regulated by ABA signaling 381
in the wild type. In the wild type, 2269 genes were ABA-induced and 2774 genes 382
were ABA-repressed. Among the total 279 co-regulated genes in ndx-4 and atring1a 383
atring1b under normal conditions, 105 genes were responsive to ABA in the wild type, 384
including 80 ABA-induced genes and 25 ABA-repressed genes (Figure 5D, 385
Supplemental Data Set 1, Sheet 12). We then compared the expression levels of these 386
co-regulated and ABA-responsive genes under normal and ABA treatment conditions. 387
Heat map analysis indicated that the expression levels of most of these co-regulated 388
genes in ndx-4 and atring1a atring1b were higher than those in the wild type under 389
both normal conditions and ABA treatment (Figure 5E). 390
We used reverse transcription-quantitative PCR (RT-qPCR) to confirm the 391
expression of a sample of genes, including PLT5 (PLETHORA5), SUT4 (SUCROSE 392
TRANSPORTER4), CHI (CHALCONE ISOMERASE) and EM1 (LATE 393
EMBRYOGENESIS ABUNDANT1) from those that were both co-regulated and 394
ABA-responsive, and AT3G63410 from those that were neither co-regulated nor 395
ABA-responsive and found the results to be consistent with the RNA-seq data. SUP 396
(SUPERMAN) served as a control for no change in expression (Figure 5F). We 397
checked the expression levels of several major components in the ABA signaling 398
pathway and found that ABI3, ABI4, and ABI5 displayed much higher expression in 399
atring1a atring1b than in the wild type (Figure 5F), but were not identified by 400
RNA-seq owing to their low expression (ABI3, ABI4) or their insignificant difference 401

13
in expression between ndx-4 and wild type (ABI5). The RT-qPCR results indicated 402
that ABI3 and ABI4 were also expressed at higher levels in ndx-4 than in the wild type 403
in presence of ABA, but ABI5 was not (Figure 5F), suggesting that the expression of 404
ABI3 and ABI4 is modulated by AtNDX. 405
AtNDX is required for normal H2Aub levels in ABI4 and other ABA-responsive 406
genes 407
As core components of PRC1, AtRING1A/B mediate histone H2A 408
monoubiquitination (H2Aub) in vitro and in vivo (Bratzel et al., 2010; Li et al., 2017). 409
As AtNDX interacts with AtRING1A/B, we wanted to know whether the H2Aub level 410
would be affected by AtNDX. We performed immunoblotting analysis using total 411
proteins extracted from 7-d-old seedlings. Interestingly, we found that the total 412
H2Aub level was significantly reduced in each of the three ndx mutants (Figure 6A 413
and 6B). Since AtNDX did not affect the expression of AtRING1A/B (Supplemental 414
Figure 6), these results suggest that AtNDX is required for H2Aub modification, 415
which most likely relies on protein interaction between AtNDX and AtRING1A/B. 416
We hypothesize that if PRC1 works with AtNDX to regulate its target genes, the 417
H2Aub on some loci should be altered in the ndx mutants. Referring to published data 418
(Zhou et al., 2017), we identified 93 H2Aub-marked genes, representing about 84% of 419
the 105 co-regulated and ABA-responsive genes (Figure 6C). We then selected some 420
of these genes (Supplemental Figure 9) and checked their H2Aub levels by a 421
chromatin immunoprecipitation (ChIP) assay. Among these genes, the H2Aub levels 422
of PLT5, SUT4, CHI, EM1, and ABI4 were significantly reduced in both ndx-4 and 423
atring1a atring1b compared with the wild type, while the H2Aub level of ABI3 was 424
reduced slightly but not significantly. The H2Aub levels of AT3G63410 and ABI5 425
were apparently unchanged (Figure 6D), which is consistent with the lack of 426
difference in their expression levels in ndx-4 and atring1a atring1b relative to the 427
wild type (Figure 5F). SUP (SUPERMAN) was used as a negative control without 428
H2Aub. Collectively, our results demonstrate that AtNDX and AtRING1A/B affect 429
H2Aub levels of ABI4 and some ABA-responsive genes. These results suggest that 430
that H2Aub levels are negatively correlated with the expression levels of some genes, 431
but they are not necessarily linearly related, which is similar to several repressive 432
histone modifications reported before (Huang et al., 2013). 433
ABI4 is a direct target of AtNDX 434

14
In order to find the possible targets of AtNDX in ABA-regulated root growth, we 435
performed genetic analyses of several major components in the ABA signaling 436
pathway. We found that the abi4-1 mutation could rescue the ABA-hypersensitive 437
phenotype of the ndx-5 mutant with respect to both primary root growth and seedling 438
establishment (Figure 7A–7D). However, abi4-1 single mutants showed similar ABA 439
sensitivity in primary root growth as the wild type (Figure 7A, 7B). This result 440
suggests that as a positive regulator, ABI4 level in the wild type is not high enough to 441
inhibit the primary root growth under ABA treatment, but the higher expression of 442
ABI4 in ndx-5 mutants leads to more sensitivity to ABA in primary root growth, 443
compared with in the wild type. 444
To further confirm the results, we created two CRISPR/Cas9 abi4 mutants in ndx-4 445
and ndx-5 using an egg-specific promoter system (Wang et al., 2015), among which a 446
chimeric protein was produced in abi4-C1 and a truncated protein in abi4-C2. The 447
ABA-sensitive phenotypes of ndx-4 abi4-C1 and ndx-5 abi4-C2 were similar to those 448
of the wild type (Supplemental Figure 10A–10D), confirming that ABI4 acts at or 449
downstream of AtNDX in ABA signaling. By contrast, ndx-5 abi1-1 (Col-0), ndx-5 450
abi2-1 (Ler) double mutants and the ndx-5 snrk2.2/2.3 triple mutant displayed 451
intermediate ABA-sensitive phenotypes, suggesting that AtNDX is one of the 452
downstream targets of these components in the early ABA signaling pathway 453
(Supplemental Figure 11A–11F). The ndx-5 abi3-1 and ndx-5 abi5 double mutants 454
displayed similar primary root growth phenotypes to ndx-5, but exhibited 455
ABA-insensitive seedling establishment phenotypes similar to those of abi3-1 and 456
abi5 (Supplemental Figure 11G–11L). These findings suggest that ABI3 and ABI5 act 457
at or downstream of AtNDX in seedling establishment, but play different roles in 458
ABA-inhibited primary root growth. Indeed, recently it was found that ABI5 459
mutations can suppress the primary root ABA-sensitive phenotype of the glk1 460
(golden2-like1) glk2 wrky40 triple mutant in Arabidopsis, suggesting that ABI5 also 461
functions in primary root growth, although like abi4, abi5 single mutant did not show 462
any primary root growth phenotype to ABA (Ahmad et al., 2019). 463
Although studies have reported diverse functions of ABI4, including regulating 464
lateral root growth (Shkolnik-Inbar and Bar-Zvi, 2010) and increasing ABA 465
accumulation, decreasing GA content, and causing dwarfing when overexpressed 466
(Shu et al., 2016), few studies have examined the role of ABI4 in regulating primary 467

15
root growth. To test this, we produced estradiol-inducible ABI4-overexpression 468
transgenic lines and examined their phenotype on ABA medium. High expression of 469
ABI4 induced by addition of estradiol inhibited primary root growth on MS medium, 470
which is consistent with previous results on ectopic, constitutive ABI4 expression 471
(Soderman et al., 2000). ABI4 overexpression also caused increased sensitivity to 472
ABA in terms of primary root growth (Figure 7E and 7F). These results suggest that 473
ABI4 plays a positive role in ABA inhibition of primary root growth. 474
The above results prompted us to test whether AtNDX directly targets ABI4. To 475
search for AtNDX targets, we performed ChIP-seq analysis using anti-NDX 476
antibodies or using AtNDX-GFP complemented ndx seedlings with anti-GFP 477
antibodies, but failed to pull down enough genomic DNA for further analysis, 478
probably because of low antibody quality or a low amount of AtNDX protein. We 479
then used DNA affinity purification sequencing (DAP-seq) to examine AtNDX 480
binding (O'Malley et al., 2016). AtNDX is a homeodomain (HD)-containing protein 481
that belongs to a large family of transcription factors with the HD preferably binding 482
to a core TAAT motif (Mukherjee et al., 2009; Christensen et al., 2012). The 483
homeodomain of AtNDX is atypical and highly divergent (Mukherjee et al., 2009). 484
AtNDX has been reported to bind the single-stranded-DNA at the 3′ region of FLC in 485
a non-sequence-specific manner (Sun et al., 2013). We used the GST-fused 486
C-terminus of AtNDX protein (including the HD and NDXB domain) to pull down 487
fragmented genomic DNA in vitro as we could not purify full-length AtNDX from E. 488
coli (Sun et al., 2013). The DAP-seq analysis revealed that the AtNDX fragment 489
preferred to bind the promoter and terminal region of genes and that the AtNDX 490
binding sequence usually possessed a high AT ratio (Supplemental Figure 12A and 491
12B). 492
We noticed that there were both weak and strong binding signals close to the 493
ABI3/4/5 genes (Figure 8A). We further examined whether these loci were targeted by 494
AtNDX in vivo by using a ChIP-PCR assay. Here we used the ndx-1 (eoc1-1) mutant 495
complemented by AtNDX driven by its native promoter and fused with GFP in frame 496
(Sun et al., 2013). 7-day-old seedlings were used for a ChIP-PCR assay with GFP 497
antibodies, and pro35S:GFP transgenic seedlings were used as a control. We found 498
that a significant enrichment was detected around the ABI4 downstream region 499
(around 730-1013 bp from the putative stop codon), but not in the promoter region 500

16
(Figure 8B). For ABI3 and ABI5, we did not detect a clear AtNDX enrichment (Figure 501
8C). 502
We then confirmed the binding in vitro through an electrophoretic mobility shift 503
assay (EMSA) at the DAP-seq peak localized in the ABI4 downstream region (Figure 504
8D and 8E). We noticed that AtNDX-GST could bind to both single-stranded DNA 505
(ssDNA) and double-stranded DNA (dsDNA) of this region, but had stronger binding 506
affinity to dsDNA (Figure 8E). This differs from the results of a previous study, in 507
which AtNDX only bound to ssDNA (Sun et al., 2013), probably due to the different 508
probes used in two assays (Sun et al., 2013). These results suggest that AtNDX can 509
bind both ssDNA and dsDNA. The GST negative control did not show any binding 510
affinity. We observed that either TATA or ATTA mutations decreased the binding 511
affinity (Figure 8E). Moreover, AtNDX binding affinity seemed to be related to the 512
AT content of the sequence (Figure 8E). 513
As AtNDX can bind the downstream region of ABI4 and repress its expression, 514
we wanted to know whether deletion of the ABI4 downstream fragment would affect 515
ABI4 expression. We used CRISPR/Cas9 to create two deletion mutants (abi4-T1, 516
abi4-T2) in the downstream region of ABI4 (Wang et al., 2015). The two 517
CRISPR/Cas9 mutants were more sensitive to ABA than the wild type, but less 518
sensitive to ABA than ndx-4 with respect to primary root growth (Figure 8G-8F). 519
Consistently, we found that the expression of ABI4 under ABA treatment was 520
significantly increased in these two CRISPR/Cas9 lines (Figure 8I). These results 521
suggest that the downstream region of ABI4 plays a negative role in regulation of 522
ABI4 expression. 523
DISCUSSION 524
In this report, we provide several lines of evidence showing that AtNDX interacts 525
with the PRC1 core components AtRING1A and AtRING1B, and that they 526
co-regulate the H2Aub modification and expression of some ABA-responsive genes 527
(Figure 3, Figure 5 and Figure 6). Interestingly, ABA negatively regulates the 528
expression of AtNDX. Under normal conditions, a larger number of AtNDX and 529
AtRING1A/B co-regulated ABA-responsive genes were up-regulated than 530
down-regulated in ndx-4 and atring1a atring1b mutants relative to wild type, 531
suggesting that AtNDX and AtRING1A/B are required for repressing these genes 532

17
under the normal conditions. AtNDX and AtRING1A/B mutants are hypersensitive to 533
ABA in seed germination, seedling establishment and root growth, but not in ABA 534
promoted stomatal closure, indicating that they are negative regulators in some 535
ABA-mediated responses. We found that in ndx mutants, the H2Aub level was 536
apparently reduced but the expression levels of AtRING1A and AtRING1B were 537
unchanged, suggesting that H2Aub modification is mediated by AtNDX at the 538
posttranscriptional level. Furthermore, most of the AtNDX and AtRING1A/B 539
co-regulated ABA-responsive genes have H2Aub modification. We found an apparent 540
reduction in H2Aub in these co-regulated ABA-responsive genes, including ABI4, in 541
the ndx and atring1a atring1b mutant. So we speculate that AtNDX might relate to 542
the AtRING1A/B’s function in modifying the target genes, such as ABI4. 543
Under drought stress conditions, the upstream components in the ABA signaling 544
pathway such as ABA receptors, SnRKs, and PP2Cs usually modulate both the 545
expression of ABA-responsive genes at the transcriptional level in a relatively slow 546
response and guard cell movement at the posttranscriptional level in a quick response 547
(Cutler et al., 2010; Zhu, 2016; Qi et al., 2018). Here we found that ndx mutants 548
exhibited ABA super-sensitivity with respect to seedling establishment and primary 549
root growth. Our genetic analyses with classic ABA-responsive mutants indicate that 550
ndx mutation compromised the ABA-insensitive phenotypes of abi1-1 (Col-0) (Hua et 551
al., 2012), abi2-1 (Leung et al., 1997) and snrk2.2/2.3 (Fujii et al., 2007) double 552
mutants, further suggesting that AtNDX is one of downstream targets in ABA 553
signaling (Supplemental Fig. 11). During the seedling establishment stage, the 554
ABA-insensitive phenotypes of abi3 (Giraudat et al., 1992), abi4 (Finkelstein et al., 555
1998) and abi5 (Finkelstein and Lynch, 2000) were not changed by introducing the 556
ndx mutation, suggesting that ABI3/4/5 function genetically downstream of AtNDX. 557
The expression of ABI3 is increased in ndx, but we did not find that AtNDX could 558
directly bind to ABI3 in the ChIP-PCR analysis, suggesting either that AtNDX does 559
not directly target ABI3 or that our technique is not sensitive enough to detect the 560
binding. Indeed, the regulation network of ABI3, ABI4, and ABI5 is very complex 561
(Soderman et al., 2000; Feng et al., 2014). 562
Although previous studies had shown diverse functions of ABI4 (Wind et al., 563
2013; Feng et al., 2014; Li et al., 2014; Shu et al., 2016; Huang et al., 2017), its role in 564
regulating primary root growth in an abi4 mutant had not been revealed. Our genetic 565

18
studies show that ABI4 mutations can rescue the ABA-hypersensitive seedling 566
establishment and primary root growth phenotypes of ndx mutants, and that 567
overexpression of ABI4 in an inducible manner retarded primary root growth and 568
increased sensitivity to ABA. These results suggest that ABI4 functions downstream 569
of AtNDX in regulating primary root growth. Given that abi4-1 does not show any 570
primary root growth difference comparing with the wild type under ABA treatment, 571
we speculate one reason is that the expression of ABI4 is precisely and strictly 572
regulated during plant development. In the wild type after germination, ABI4 is 573
gradually silenced and stays at a very low expression level that is not enough to 574
inhibit primary root growth under ABA treatment. However, in the ndx mutants and 575
ABI4 overexpression lines, ABI4 is expressed to reach a high level that is sufficient to 576
inhibit the primary root growth. Therefore, we think the targets of ABI4 are probably 577
directly related to ABA response. 578
In our study, we found by DAP-seq and EMSA that AtNDX is able to bind the 579
downstream region of ABI4, while H2Aub is mainly distributed in the ABI4 gene body, 580
especially in the first exon (Zhou et al., 2017). How then does AtNDX promote 581
H2Aub modification and repress ABI4 expression? We speculate that AtNDX might 582
bind the downstream region of ABI4 and then form a DNA loop, allowing it to interact 583
with AtRNG1A/B in the coding region, promoting H2Aub modification and 584
suppressing ABI4 expression (Figure 9). One of the examples is a recent study on the 585
expression of GLABRA1 (GL1), in which the KANADI1-TARGET OF EAT1 (TOE1) 586
complex binds the 3′ downstream region of GL1 and then forms a loop in the 587
promoter region, repressing GL1 expression and inhibiting trichome formation (Wang 588
et al., 2019). Genome-wide Hi-C (a genome-wide chromatin conformation capture 589
[3C]) analysis in Arabidopsis indicates that promoter regions with repressive histone 590
modification markers such as H3K27me3 show a strong tendency to form 591
conformational linkages over long distances in gene bodies (Liu et al., 2016). ABI4 is 592
also regulated by BPC-recruited PRC2 in the promoter, which mediates H3K27me3 593
modification (Mu et al., 2017). Some components of PRC1 could interact with those 594
of the PRC2 complex to form a large complex (Wang and Shen, 2018). It is possible 595
that AtNDX interacts with AtRING1A/B to form a DNA loop after it binds to the 596
downstream region of ABI4 (Figure 9). In this study, we attempted to test this 597
hypothesis by the 3C technique, but failed due to a lack of suitable restriction enzyme 598

19
recognition sites in this region. Nevertheless, the deletion by CRISPR/Cas9 of the 599
ABI4 downstream region where AtNDX most likely binds increased ABI4 expression 600
and ABA sensitivity. The deletion lines were more ABA-sensitive in primary root 601
growth than the wild type, but less ABA-sensitive than ndx, suggesting that AtNDX 602
not only works in the downstream region of ABI4, but also, for example, works 603
together with PRC1 in the ABI4 coding region. Without ABA treatment, the 604
expression of ABI4 in two deletion lines was not increased, but increased in ndx 605
mutant, suggesting that AtNDX is required for repressing ABI4 expression under 606
normal condition (Figure 8I). 607
In our study, it was very hard to obtain a stably overexpressed ABI4 transgenic 608
line in later generations, suggesting that ABI4 can be easily silenced, likely due to its 609
sequence in coding region. A previous study showed that AtNDX could stabilize the 610
R-loop structure of FLC (Sun et al., 2013), and the present study reveals a novel role 611
for AtNDX working with PRC1 for gene repression, suggesting that AtNDX uses 612
different regulatory mechanisms to control gene expression. When comparing gene 613
expression patterns, the number of genes co-regulated by AtNDX and 614
AtRING1A/AtRING1B was not very large; one possible explanation is that some 615
genes may be regulated by AtNDX-mediated R-loop formation during transcription. 616
Recent studies have indicated that, like other epigenetic markers such as histone 617
modification and DNA methylation, R-loops play crucial roles in various cellular 618
processes including gene expression and DNA repair (Xu et al., 2017; Zhou et al., 619
2017; Crossley et al., 2019). In order to fully understand the functions of AtNDX, it 620
will be necessary to perform genome-wide profiles of R-loops and H2Aub in ndx and 621
atring1a atring1b mutants. However, more improved techniques are needed as 622
AtNDX protein availability is a limiting factor: the protein is not highly accumulated 623
even when the gene is driven by the 35S promoter (this study) (Sun et al., 2013). This 624
may be also the possible reason for why the overexpressed AtNDX lines did not show 625
ABA insensitivity while only rescued the ndx mutant phenotype. 626
Many studies have shown that epigenetic regulation is involved in plant abiotic 627
stress responses (Chinnusamy et al., 2008; Yuan et al., 2013). A recent study indicates 628
that PRC2 attenuates ABA-induced senescence in Arabidopsis in a long-term process 629
(Liu et al., 2018a). The PRC1 complex also represses the expression of some 630
important genes in phase transitions during different developmental stages (Calonje, 631

20
2014; Wang and Shen, 2018). The connection between ABA signaling and chromatin 632
modification has been explored in several previous studies (Chinnusamy et al., 2008; 633
Saez et al., 2008; Han et al., 2012; Peirats-Llobet et al., 2016; Zhao et al., 2017; Liu et 634
al., 2018b). For example, SWI3B, a subunit of the SWI/SNF chromatin-remodeling 635
complex, interacts with HYPERSENSITIVE TO ABA1 (HAB1) and positively 636
modulates ABA signaling (Saez et al., 2008). The SWI/SNF ATPase BRAHMA 637
(BRM) can directly bind to the transcription start site of ABI5 and maintain the 638
well-positioned nucleosome for repressing the expression of ABI5 (Han et al., 2012). 639
The activity of BRM is adversely regulated by phosphorylation and 640
dephosphorylation mediated by SnRK2s and PP2CA (Peirats-Llobet et al., 2016), 641
which would result in activation of ABI5 or maintaining the repression status of ABI5, 642
respectively. H3K4me3 and H3K27me3 marks were found to be involved in drought 643
stress memory (Liu et al., 2014). ARABIDOPSIS TRITHORAX4 (ATX4) and ATX5 644
directly regulate the expression of ABA-HYPERSENSITIVE GERMINATION 3 645
(encoding a PP2C) through H3K4 methylation by binding at the locus (Liu et al., 646
2018b). The finding in this study that AtNDX interacts with PRC1 core components 647
AtRING1A/B to mediate ABA signaling provides novel evidence for the direct 648
connection between ABA signaling and epigenetic regulation. 649
MATERIALS AND METHODS 650
Plant materials and growth conditions 651
The Arabidopsis thaliana seeds were surface-sterilized and sown on MS medium 652
containing 2% sucrose and 0.8% or 0.9% agar. After stratified at 4oC for 3 days, the 653
plates were transferred to a light incubator (PHILIPS F17T8/TL841 bulb) with a light 654
intensity of 60 μE m−2
s−1
under 22-h-light/2-h-dark at 22oC. The mutants used in this 655
study Atring1a, Atring1b-2 (Shen et al., 2014), Atring1a-/-
Atring1b+/-
(Xu and Shen, 656
2008), ring1a-2 ring1b-3 (Li et al., 2017), eoc1-4 is renamed as ndx-4 in this study 657
(Sun et al., 2013), abi1-1 (Kong et al., 2015), snrk2.2/2.3 (Wang et al., 2018), abi4-1 658
(Liu et al., 2010) were in the Col-0 background; abi2-1, abi3-1 (Wang et al., 2011) 659
were in the Landsberg erecta (Ler) background, abi5 were in the Wassileskija (Ws) 660
background. The transgenic plants AtNDX-FLAG (in eoc1-4, ndx-4 in this study) and 661
AtNDX-eGFP (in eoc1-1, ndx-1) renamed as proAtNDX: AtNDX-Flag and proAtNDX: 662
AtNDX-GFP were as described before (Sun et al., 2013). For transgenic plants 663
overexpressing AtNDX and AtRING1A, their CDSs were fused with the indicated tag 664

21
and driven by the super promoter (Ni et al., 1995) and cloned into the pCAMBIA1300 665
vector. For the β-estradiol inducible ABI4 overexpression lines, the coding sequence 666
of ABI4 was fused with GFP tag and cloned into the pER8 vector (Zuo et al., 2000). 667
The constructs were introduced into Agrobacterium tumefaciens strain GV3101 and 668
then transformed into the ndx-5 mutant or wild type by floral infiltration. The primers 669
used to examine these mutants and construct the transgenic plants are listed in 670
Supplemental Table 1. 671
To generate the ABI4 downstream deletion lines (abi4-T1 and abi4-T2), targeting the 672
fragment from 618 bp and 1007 bp downstream of ABI4 (counting after the ABI4 673
putative stop codon) was selected for CRISPR/Cas9 editing. The targets were cloned 674
into the pHSE401 vector as described previously (Wang et al., 2015) and the 675
constructs were transformed into Col-0 by floral dip. The transgenic T1 seeds were 676
screened on MS medium containing 25 mg/L hygromycin. To identify the deletion 677
lines, about 80 T1 plants were first screened by PCR then verified by sequencing, and 678
the homozygous mutants were used to generate T2. To generate ndx-4 abi4-C1 and 679
ndx-5 abi4-C2 mutants, the same method was used, except that targets at the coding 680
sequence of ABI4 were selected and the constructs were introduced into ndx-4 and 681
ndx-5 mutant. T2 plants were harvested individually, and T3 seeds were screened on 682
hygromycin medium for the non-hygromycin-resistant lines. The primers used to 683
examine these mutants, to construct the CRISPR/Cas9 vector and transgenic plants 684
are listed in Supplemental Tables 2–4. 685
Map-based cloning of AtNDX 686
The mutants ndx-5 and ndx-6 were isolated from approximately 20,000 EMS- (ethyl 687
methane sulfonate) mutated M2 seedlings based on the root bending assay. ndx-5 688
mutant plants were crossed with Landsberg erecta plants. A total of about 1,300 ndx-5 689
mutant plants were selected from the self-fertilized F2 population based on the 690
ABA-hypersensitive phenotype on 30 μM ABA. The simple sequence-length 691
polymorphism (SSLP) markers were used for mapping. The position of the AtNDX 692
mutation was determined within BAC clone T4I9. All genes in the BAC clone T4I9 693
were sequenced, and the mutation in the At4g03090 gene was identified in ndx-5. 694
ndx-6 was mapped to the same BAC clone as ndx-5, and a mutation in ndx-6 was 695
found in At4g03090. For the complementation, an 8 kb fragment that included about 696
3000 bp upstream of the first putative ATG to 1000 bp downstream of the putative 697

22
stop codon TGA of At4g03090 gene was amplified from genomic DNA and cloned 698
into pCAMBIA1391 vector. The constructs were introduced into Agrobacterium 699
tumefaciens strain GV3101 and then transformed into the ndx-5 mutant or wild type 700
by floral infiltration. The several independent transgenic lines obtained were able to 701
complement the ABA-hypersensitive phenotypes of ndx-5. 702
ABA-related phenotype analyses 703
For the root growth assay, seeds were germinated on the vertically oriented plates, 704
and 5-d-old seedlings were transferred to MS medium supplemented with different 705
concentrations of ABA for 3 or 4 days. Then the seedlings were photographed and the 706
root growth after transfer (below the dotted line) was measured by Image J. For the 707
calculation of relative root growth, the root growth of each genotype on MS medium 708
without ABA was set to 100%. For the seed germination greening ratio assay, seeds 709
were sown on MS medium supplemented with different concentrations of ABA and 710
grew for 2-11 days after sowing before photographed, and the ratio of seedlings with 711
ruptured endosperm, emerged radical and expended green cotyledons was calculated. 712
Observation of root meristem and mature epidermal cell 713
To measure of meristem cell number, a drop of transparent liquid was added to a glass 714
slide, and the root tips of the seedlings were immersed in the transparent liquid, then 715
covered with a glass cover and placed under a microscope (Olympus BX53). The cells 716
in the meristem were observed with a 20-fold objective lens and photographed. The 717
transparent liquid contains 60 mL deionized water, 7.5 g Gum Arabic, 100 g hydrated 718
trichloroacetaldehyde and 5 mL Glycerol. 719
To measure the length of mature epidermal cell, roots were treated as following steps: 720
(1) aqueous solution containing 4% HCl and 20% methanol at 57 °C for 15 min; (2)721
aqueous solution containing 7% NaOH and 60% ethanol at room temperature for 15 722
min; (3) 60% ethanol solution at room temperature for 10 min; (4) 40% ethanol 723
solution at room temperature for 10 min; (5) 20% ethanol solution at room 724
temperature for 10 min; (6) 10% ethanol solution at room temperature for 10 min; (7) 725
aqueous solution containing 25% glycerol and 5% ethanol at room temperature for 10 726
min; (8) store in 50% glycerol solution and observe the film. The films were observed 727
under a 10-fold objective lens and photographed, and the cell length was measured 728
using ImageJ software. For each root, the first five epidermal cells (near to the distal) 729

23
from the first cell with root hair were measured and used to calculate the average 730
length. 731
Physiological Experiments 732
For the water loss in detached leaves, shoots were cut from plants growing under 733
normal conditions for 4 weeks, and placed on a piece of weighing paper under a light 734
in a greenhouse, weighed immediately, every half an hour or one hour. The results are 735
shown as a percentage of the fresh weight. More than two independent experiments 736
were performed, each including three or four replicate shoots per line. 737
For stomatal density, epidermal strips were peeled from 4-weeks plants and the 738
mesophyll cells were removed with a small brush. The cleaned epidermal strips were 739
photographed under a microscope (Olympus BX53) with a 40-fold objective lens and 740
photographed. The stomata numbers were counted. 741
GUS staining 742
A 1609-bp fragment upstream from the start codon ATG of AtNDX was amplified by 743
PCR. The amplified fragment was cloned into the pCAMBIA1391 vector for a 744
transcriptional fusion of the AtNDX promoter with the GUS coding region. 7-d-old 745
homozygous transgenic plants carrying proAtNDX:GUS were treated with 0 μM or 30 746
μM ABA for 12 hours, and incubated in GUS staining buffer (0.1M PBS, 0.5 mM 747
K4Fe(CN)6, 0.5 mM K3Fe(CN)6, 1 mg/mL X-gluc, 0.1% Triton X-100) for 6 hours at 748
37 ºC. 749
Fluorescence microscopy 750
5-d-old proAtNDX:AtNDX-GFP transgenic seedlings grown on MS medium were 751
transferred to MS medium supplemented with 0 μM or 30 μM ABA for 12 hours. 752
Then seedlings were incubated in 10 μM propidium iodide for 3 min (to visualize the 753
cell) before imaged with the Carl Zeiss LSM710 confocal microscope. The roots were 754
photographed under the same setting and the fluorescence intensities of a same area in 755
the root meristem zone were measured by ImageJ software. 756
Yeast two-hybrid assay 757
For yeast two-hybrid screening, a normalized universal Arabidopsis cDNA library 758
(Clontech, 630487) was screened with the full length of AtNDX as bait. The 759
screening was performed according to the operating instruction. For construction the 760

24
bait and prey, the full length or truncated AtNDX and AtRING1A/B were fused into 761
pGBKT7 (BD) or pGADT7 (AD) vectors. The plasmids were co-transfected into 762
yeast strain AH109. Then the transfected yeast cells were plated on SD/-Leu/-Trp 763
medium (-LW) and SD/-Ade/-His/-Leu/-Trp medium (-LWHA) and cultured at 30oC 764
for 5 days. The primers used for vector construction are listed in Supplemental Table 765
3. 766
Co-IP and LC-MS/MS Assays 767
The Co-IP assays were performed as described previously (Wang et al., 2018). Briefly, 768
the total proteins of Arabidopsis protoplasts transfected with pro35S:HA-Flag-AtNDX 769
and proSuper:AtRING1A-GFP or proSuper:AtRING1B-GFP, or transgenic plants 770
harboring proSuper:AtRING1A-GFP were extracted and immunoprecipitated by GFP 771
beads (gta-20, ChromoTek). The primers used for vector construction are listed in 772
Supplemental Table 3. The immunoprecipitated proteins were detected with indicated 773
antibodies by immunoblotting analysis. The antibodies used were anti-GFP antibodies 774
(ab290, Abcom), anti-HA antibodies (H6908, Sigma-Aldrich), anti-NDX antibodies 775
(produced by Shanghai Youke Biotechnology Co., Ltd. ). Anti-NDX antibodies are 776
polyclonal antibodies prepared by immunizing rabbits with a peptide comprising the 777
589th to 879th amino acid of the AtNDX protein. For LC-MS/MS, total proteins were 778
extracted from wild-type and ndx-5 mutant 10-d-old seedlings, and 779
immunoprecipitated with anti-NDX antibodies, and the precipitated products were 780
subjected to LC-MS/MS analysis as described previously (Kong et al., 2015). 781
RNA-seq analyses. 782
7-d-old seedlings were treated with 1/2 liquid MS medium supplemented with 0 or 60 783
μM ABA for 3 hours. The total RNAs were extracted using RNeasy Plant Mini Kit 784
(Qiagen, 74104) and sent to the Berry Genomics Corporation for library construction 785
and sequencing. The sequencing was performed on the Illumina Novaseq6000 786
platform. Each sample had two biological replicates and obtained about 12G clean 787
bases each replicate. RNA-seq reads were collapsed into non-redundant reads and 788
mapped back to TAIR10 genome with Araport 11 annotation using STAR aligner with 789
maximum 8 mismatches per pair reads (Dobin et al., 2013). The gene expression level 790
was quantified using the available software cuffdiff. The differential expression genes 791
(DEGs) were filtered with FDR < 0.05 and Fold change >2 and required to have an 792

25
expression higher than 1 FPKM (Trapnell et al., 2013). The Venn diagrams were 793
drawn on the website http://bioinformatics.psb.ugent.be/webtools/Venn/ and the E 794
values were caculated through hypergeometric test by R package. The heatmap was 795
drawn based on the expression levels of indicated genes by the software Mev 4.9.0 796
with default parameters (Saeed et al., 2003). The raw data have been deposited in the 797
NCBI Sequence Read Archive and are accessible through SRA accession code 798
PRJNA556351 (https://www.ncbi.nlm.nih.gov/sra/PRJNA556351). 799
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) 800
7-d-old seedlings were treated with different concentrations of ABA for 3 hours, and 801
the total RNAs were extracted by HiPure Plant RNA Mini Kit (Magen, R4151). 4-μg 802
total RNAs were used for first-strand cDNA synthesis by Maxima H Minus First 803
Strand cDNA Synthesis Kit (Theeermo Scientific, K1682). RT-qPCR was performed 804
as described previously (Kong et al., 2015). The primers used for RT-qPCR were 805
listed in Supplemental Table 5. 806
Chromatin immunoprecipitation (ChIP) 807
The ChIP assays were performed as described previously (Zhang et al., 2016) with a 808
little modification. For H2Aub ChIP, 10-d-old seedlings were ground into fine 809
powder and cross-linked with 1% formaldehyde for 10 min. The nuclei were isolated 810
and the chromatin was sonicated by Bioruptor (Diagenode) and immunoprecipitated 811
by H2Aub antibody (Cell Signaling, D27C4) and Protein A+G Magnetic Beads 812
(Millipore, 16-663). For AtNDX-GFP ChIP, 7-d-old proAtNDX: AtNDX-GFP/ndx-1 813
and pro35S: GFP/Col-0 transgenic plants were used and the chromatin was 814
immunoprecipitated by GFP beads (gta-20, ChromoTek). The precipitated DNAs 815
were recovered by ChIP DNA Clean & Concentrator kit (Zymo Research, D5205) and 816
analyzed by RT-qPCR. The primers used for ChIP-qPCR are listed in Supplemental 817
Table 6. 818
Purification of recombinant protein, electrophoretic mobility shift assay (EMSA) 819
and DNA Affinity purification sequencing (DAP-seq) 820
The DNA-binding and NDXB domain (1800-2634 bp) in the C-terminal of AtNDX 821
was fused in frame with GST in the pGEX-2T vector and expressed in E. coli BL21 822
(DE3). The fused protein was induced by 1 mM IPTG and incubated overnight at 16 823
ºC. The recombinant protein was purified with Glutathione Sepharose 4B (GE 824

26
Healthcare) following the instruction. EMSA was carried out using the LightShift™ 825
Chemiluminescent EMSA Kit (Thermo Scientific) according to the instructions. For 826
double strand probes, the 5’- biotin-labeled DNA primers were made by 20 min 100 827
oC denaturation and renaturation in room temperature, and purified by native 828
polyacrylamide gel electrophoresis and QIAEX II Gel Extraction Kit (QIAGEN). For 829
single strand probes, the primers were denatured for 20 min and then transferred to ice 830
immediately. Each 20-μL volume of binding reaction contained 10 fM Biotin-probe, 1 831
μg of protein. The probes were incubated with the GST-AtNDX-C protein at room 832
temperature for 30 min. The reaction products were separated by 6.5% native 833
polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane 834
and detected according to the instructions. The DAP-seq was performed according to 835
O'Malley et al. (O'Malley et al., 2016) with a little modification.The genomic DNA 836
was extracted from 7-d-old wild type seedlings and fragmented by Bioruptor 837
(Diagenode) to 200-500 bp. The GST-AtNDX-C protein was first incubated with 838
Glutathione Sepharose 4B beads for 2 h, and washed out the free protein. Then the 839
GST-AtNDX-C covered beads were incubated with 500 ng fragmented DNA for 1h. 840
After washed out the free DNA, the affinity purified DNA was eluted and used for 841
sequencing. Two repeats were sequenced and 5 M reads were obtained for each repeat. 842
The clean data were mapped to the Arabidopsis genome (TAIR10) using bowtie2 843
(Langmead and Salzberg, 2012) with the following parameters: -N 1 -p 5 -q; and the 844
NDX binding peaks were identified by MACS (version 1.4.2) software (Zhang et al., 845
2008). The raw data have been deposited in the NCBI Sequence Read Archive and are 846
accessible through SRA accession code PRJNA556351 847
(https://www.ncbi.nlm.nih.gov/sra/PRJNA556351). 848
849
Accession Numbers 850
Sequence data from this article can be found in the GenBank/EMBL data libraries 851
under the following accession numbers: AtNDX, AT4G03090; AtRING1A, 852
AT5G44280; AtRING1B, AT1G03770; AtBMI1A, AT2G30580; AtBMI1B, AT1G06770; 853
AtBMI1C, AT3G23060; AtEMF1, AT5G11530; LHP1, AT5G17690; PLT5, 854
AT5G57390 ; SUT4, AT1G09960; CHI, AT2G43570; EM1, AT3G51810; ABI3, 855
AT3G24650; ABI4, AT2G40220; ABI5, AT2G36270; ACT4, AT5G59370; SUP, 856
AT3G23130. Mutants used in this article can be obtained from the ABRC under the 857

27
following accession numbers: snrk2.2 (GABI-Kat 807G04), and snrk2.3 858
(SALK_107315). The RNA sequencing data and DAP sequencing data for this 859
research have been deposited in the NCBI Sequence Read Archive under accession 860
code PRJNA556351. 861
862
Supplemental Data 863
Supplemental Figure 1. Root cell division and elongation of ndx-5 mutant are 864
hypersensitive to ABA. Supports Figure 1. 865
Supplemental Figure 2. Seedling establishment and stomata movement analysis of 866
AtNDX. Supports Figure 1. 867
Supplemental Figure 3. Map-based cloning and mutation analysis of AtNDX. 868
Supports Figure 1. 869
Supplemental Figure 4. Complementation of ndx mutants. Supports Figure 1. 870
Supplemental Figure 5. AtNDX does not directly interact with other PRC1 871
components in yeast cells. Supports Figure 3. 872
Supplemental Figure 6. The transcriptional levels of AtNDX and AtRING1A/B in 873
corresponding mutants. Supports Figure 5. 874
Supplemental Figure 7. ndx-5 atring1a double mutant is more sensitive to ABA than 875
ndx-5 in seed germination but not in primary root growth. Supports Figure 5. 876
Supplemental Figure 8. ndx-5 atring1b-2 double mutant shows similar sensitivity 877
with ndx-5 to ABA in seedling establishment and primary root growth. Supports 878
Figure 5. 879
Supplemental Figure 9. ChIP-seq genome browser views from published results. 880
Supports Figure 6. 881
Supplemental Figure 10. Disruption of ABI4 by CRISPR/Cas9 rescues the 882
ABA-hypersensitive phenotype of ndx mutants. Supports Figure 7. 883
Supplemental Figure 11. Genetic analysis of AtNDX and ABA signaling pathway 884
members. Supports Figure 7. 885
Supplemental Figure 12. DAP-seq analysis shows that AtNDX prefers to bind the 886

28
promoter and terminal regions of genes that possess high AT ratio. Supports Figure 8. 887
888
Supplementary Tables 1-6. Primers used in this study. 889
Supplemental Data Set 1. Gene expression analysis from RNA-seq. Supports Figure 890
5. 891
892
893
ACKNOWLEDGMENTS 894
We thank Dr. Zhen Li at China Agricultural University for performing LC-MS/MS 895
analysis. We thank Dr. Lin Xu at Shanghai Institutes for Biological Sciences for 896
providing ring1a-/-
ring1b+/-
double mutants, Dr. Hao Yu at National University of897
Singapo for providing ring1a and ring1b-2 mutants, and Dr. Ligeng Ma at Capital 898
Normal University for providing ring1a-2 ring1b-3 double mutants. This work was 899
supported by grants from the National Major Project for Transgenic Organism 900
Breeding (Ministry of Agriculture and Rural Affairs of the People’s Republic of 901
China) (2016ZX08009002), Beijing Outstanding Univeristy Displine Program, and 902
National Science Foundation of China (31921001). 903
AUTHOR CONTRIBUTIONS 904
Z.G., and Y.Z. conceived and designed the experiments. Y.Z. performed most of905
experiments. X.H. analyzed the phenotypes of ABI4 deletion mutants and Y.D. 906
isolated the mutants and cloned AtNDX gene. S.L. performed the bioinformatic 907
analyses of RNA-seq and ChIP-seq data. Z.G., Y.Z. and X.H. wrote the manuscript. 908
All other authors participated in the discussion of the results and commented on the 909
manuscript. 910
911
912

29
References 913
914
Ahmad, R., Liu, Y., Wang, T.J., Meng, Q.X., Yin, H., Wang, X., Wu, Y.F., Nan, N., Liu, B., and Xu, Z.Y. 915 (2019). GOLDEN2-LIKE Transcription Factors Regulate WRKY40 Expression in Response to 916 Abscisic Acid. Plant physiology 179, 1844-1860. 917
Blackledge, N.P., Rose, N.R., and Klose, R.J. (2015). Targeting Polycomb systems to regulate gene 918 expression: modifications to a complex story. Nature reviews. Molecular cell biology 16, 919 643-649. 920
Bossi, F., Cordoba, E., Dupre, P., Mendoza, M.S., Roman, C.S., and Leon, P. (2009). The Arabidopsis 921 ABA-INSENSITIVE (ABI) 4 factor acts as a central transcription activator of the expression of its 922 own gene, and for the induction of ABI5 and SBE2.2 genes during sugar signaling. The Plant 923 journal : for cell and molecular biology 59, 359-374. 924
Bratzel, F., Lopez-Torrejon, G., Koch, M., Del Pozo, J.C., and Calonje, M. (2010). Keeping cell identity 925 in Arabidopsis requires PRC1 RING-finger homologs that catalyze H2A monoubiquitination. 926 Current biology : CB 20, 1853-1859. 927
Bratzel, F., Yang, C., Angelova, A., Lopez-Torrejon, G., Koch, M., del Pozo, J.C., and Calonje, M. (2012). 928 Regulation of the new Arabidopsis imprinted gene AtBMI1C requires the interplay of different 929 epigenetic mechanisms. Molecular plant 5, 260-269. 930
Calonje, M. (2014). PRC1 marks the difference in plant PcG repression. Molecular plant 7, 459-471. 931 Chen, D., Molitor, A., Liu, C., and Shen, W.H. (2010). The Arabidopsis PRC1-like ring-finger proteins 932
are necessary for repression of embryonic traits during vegetative growth. Cell research 20, 933 1332-1344. 934
Chen, L., Zhang, L., Li, D., Wang, F., and Yu, D. (2013). WRKY8 transcription factor functions in the 935 TMV-cg defense response by mediating both abscisic acid and ethylene signaling in 936 Arabidopsis. Proceedings of the National Academy of Sciences of the United States of 937 America 110, E1963-1971. 938
Chinnusamy, V., Gong, Z.Z., and Zhu, J.K. (2008). Abscisic acid-mediated epigenetic processes in plant 939 development and stress responses. Journal of integrative plant biology 50, 1187-1195. 940
Choi, H., Hong, J., Ha, J., Kang, J., and Kim, S.Y. (2000). ABFs, a family of ABA-responsive element 941 binding factors. The Journal of biological chemistry 275, 1723-1730. 942
Christensen, R.G., Enuameh, M.S., Noyes, M.B., Brodsky, M.H., Wolfe, S.A., and Stormo, G.D. (2012). 943 Recognition models to predict DNA-binding specificities of homeodomain proteins. 944 Bioinformatics 28, i84-89. 945
Crossley, M.P., Bocek, M., and Cimprich, K.A. (2019). R-Loops as Cellular Regulators and Genomic 946 Threats. Molecular cell 73, 398-411. 947
Cutler, S.R., Rodriguez, P.L., Finkelstein, R.R., and Abrams, S.R. (2010). Abscisic acid: emergence of a 948 core signaling network. Annual review of plant biology 61, 651-679. 949
Dobin, A., Davis, C.A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., Batut, P., Chaisson, M., and 950 Gingeras, T.R. (2013). STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15-21. 951
Feng, C.Z., Chen, Y., Wang, C., Kong, Y.H., Wu, W.H., and Chen, Y.F. (2014). Arabidopsis RAV1 952 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, 953 and ABI5 during seed germination and early seedling development. The Plant journal : for cell 954 and molecular biology 80, 654-668. 955
Finkelstein, R.R., and Lynch, T.J. (2000). The Arabidopsis abscisic acid response gene ABI5 encodes a 956 basic leucine zipper transcription factor. The Plant cell 12, 599-609. 957
Finkelstein, R.R., Wang, M.L., Lynch, T.J., Rao, S., and Goodman, H.M. (1998). The Arabidopsis 958 abscisic acid response locus ABI4 encodes an APETALA 2 domain protein. The Plant cell 10, 959 1043-1054. 960
Francis, N.J., Saurin, A.J., Shao, Z., and Kingston, R.E. (2001). Reconstitution of a functional core 961 polycomb repressive complex. Molecular cell 8, 545-556. 962
Fujii, H., Verslues, P.E., and Zhu, J.K. (2007). Identification of two protein kinases required for abscisic 963 acid regulation of seed germination, root growth, and gene expression in Arabidopsis. The 964 Plant cell 19, 485-494. 965
Fujita, Y., Fujita, M., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2011). ABA-mediated transcriptional 966

30
regulation in response to osmotic stress in plants. Journal of plant research 124, 509-525. 967 Giraudat, J., Hauge, B.M., Valon, C., Smalle, J., Parcy, F., and Goodman, H.M. (1992). Isolation of the 968
Arabidopsis ABI3 gene by positional cloning. The Plant cell 4, 1251-1261. 969 Han, S.K., Sang, Y., Rodrigues, A., Wu, M.F., Rodriguez, P.L., Wagner, D., and F2010, B. (2012). The 970
SWI2/SNF2 Chromatin Remodeling ATPase BRAHMA Represses Abscisic Acid Responses in the 971 Absence of the Stress Stimulus in Arabidopsis. The Plant cell 24, 4892-4906. 972
He, J.N., Duan, Y., Hua, D.P., Fan, G.J., Wang, L., Liu, Y., Chen, Z.Z., Han, L.H., Qu, L.J., and Gong, Z.Z. 973 (2012). DEXH Box RNA Helicase-Mediated Mitochondrial Reactive Oxygen Species Production 974 in Arabidopsis Mediates Crosstalk between Abscisic Acid and Auxin Signaling. The Plant cell 975 24, 1815-1833. 976
Hua, D.P., Wang, C., He, J.N., Liao, H., Duan, Y., Zhu, Z.Q., Guo, Y., Chen, Z.Z., and Gong, Z.Z. (2012). A 977 Plasma Membrane Receptor Kinase, GHR1, Mediates Abscisic Acid- and Hydrogen 978 Peroxide-Regulated Stomatal Movement in Arabidopsis. The Plant cell 24, 2546-2561. 979
Huang, C., Xu, M., and Zhu, B. (2013). Epigenetic inheritance mediated by histone lysine methylation: 980 maintaining transcriptional states without the precise restoration of marks? Philosophical 981 transactions of the royal society of London series B-bilogical sciences 368, 20110332. 982
Huang, X., Zhang, X., Gong, Z., Yang, S., and Shi, Y. (2017). ABI4 represses the expression of type-A 983 ARRs to inhibit seed germination in Arabidopsis. The Plant journal : for cell and molecular 984 biology 89, 354-365. 985
Jiang, D.H., and Berger, F. (2017). DNA replication-coupled histone modification maintains Polycomb 986 gene silencing in plants. Science 357, 1146-+. 987
Jorgensen, J.E., Gronlund, M., Pallisgaard, N., Larsen, K., Marcker, K.A., and Jensen, E.O. (1999). A 988 new class of plant homeobox genes is expressed in specific regions of determinate symbiotic 989 root nodules. Plant molecular biology 40, 65-77. 990
Kerchev, P.I., Pellny, T.K., Vivancos, P.D., Kiddle, G., Hedden, P., Driscoll, S., Vanacker, H., Verrier, P., 991 Hancock, R.D., and Foyer, C.H. (2011). The transcription factor ABI4 Is required for the 992 ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent 993 defense signaling pathways in Arabidopsis. The Plant cell 23, 3319-3334. 994
Kong, L., Cheng, J., Zhu, Y., Ding, Y., Meng, J., Chen, Z., Xie, Q., Guo, Y., Li, J., Yang, S., and Gong, Z. 995 (2015). Degradation of the ABA co-receptor ABI1 by PUB12/13 U-box E3 ligases. Nature 996 communications 6, 8630. 997
Langmead, B., and Salzberg, S.L. (2012). Fast gapped-read alignment with Bowtie 2. Nature methods 998 9, 357-U354. 999
Leung, J., Merlot, S., and Giraudat, J. (1997). The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and 1000 ABI1 genes encode homologous protein phosphatases 2C involved in abscisic acid signal 1001 transduction. The Plant cell 9, 759-771. 1002
Li, J., Wang, Z., Hu, Y., Cao, Y., and Ma, L. (2017). Polycomb Group Proteins RING1A and RING1B 1003 Regulate the Vegetative Phase Transition in Arabidopsis. Frontiers in plant science 8, 867. 1004
Li, P., Zhou, H., Shi, X., Yu, B., Zhou, Y., Chen, S., Wang, Y., Peng, Y., Meyer, R.C., Smeekens, S.C., and 1005 Teng, S. (2014). The ABI4-induced Arabidopsis ANAC060 transcription factor attenuates ABA 1006 signaling and renders seedlings sugar insensitive when present in the nucleus. PLoS genetics 1007 10, e1004213. 1008
Li, Z., Fu, X., Wang, Y., Liu, R., and He, Y. (2018). Polycomb-mediated gene silencing by the BAH-EMF1 1009 complex in plants. Nature genetics 50, 1254-1261. 1010
Li, Z.F., Li, B., Liu, J., Guo, Z.H., Liu, Y.H., Li, Y., Shen, W.H., Huang, Y., Huang, H., Zhang, Y.J., and Dong, 1011 A.W. (2016). Transcription factors AS1 and AS2 interact with LHP1 to repress KNOX genes in 1012 Arabidopsis. Journal of integrative plant biology 58, 959-970. 1013
Liu, C., Wang, C., Wang, G., Becker, C., Zaidem, M., and Weigel, D. (2016). Genome-wide analysis of 1014 chromatin packing in Arabidopsis thaliana at single-gene resolution. Genome research 26, 1015 1057-1068. 1016
Liu, C., Cheng, J., Zhuang, Y., Ye, L., Li, Z., Wang, Y., Qi, M., Xu, L., and Zhang, Y. (2018a). Polycomb 1017 repressive complex 2 attenuates ABA-induced senescence in Arabidopsis. The Plant journal : 1018 for cell and molecular biology 97, 368-377. 1019
Liu, N., Fromm, M., and Avramova, Z. (2014). H3K27me3 and H3K4me3 Chromatin Environment at 1020 Super-Induced Dehydration Stress Memory Genes of Arabidopsis thaliana. Molecular plant 7, 1021 502-513. 1022
Liu, Y., He, J.N., Chen, Z.Z., Ren, X.Z., Hong, X.H., and Gong, Z.Z. (2010). ABA overly-sensitive 5 (ABO5), 1023

31
encoding a pentatricopeptide repeat protein required for cis-splicing of mitochondrial nad2 1024 intron 3, is involved in the abscisic acid response in Arabidopsis. The Plant journal : for cell 1025 and molecular biology 63, 749-765. 1026
Liu, Y.T., Zhang, A., Yin, H., Meng, Q.X., Yu, X.M., Huang, S.Z., Wang, J., Ahmad, R., Liu, B., and Xu, Z.Y. 1027 (2018b). Trithorax-group proteins ARABIDOPSIS TRITHORAX4 (ATX4) and ATX5 function in 1028 abscisic acid and dehydration stress responses. New phytologist 217, 1582-1597. 1029
Ma, Y., Szostkiewicz, I., Korte, A., Moes, D., Yang, Y., Christmann, A., and Grill, E. (2009). Regulators 1030 of PP2C phosphatase activity function as abscisic acid sensors. Science 324, 1064-1068. 1031
Merini, W., and Calonje, M. (2015). PRC1 is taking the lead in PcG repression. The Plant journal : for 1032 cell and molecular biology 83, 110-120. 1033
Molitor, A., and Shen, W.H. (2013). The polycomb complex PRC1: composition and function in plants. 1034 Journal of genetics and genomics 40, 231-238. 1035
Molitor, A.M., Bu, Z., Yu, Y., and Shen, W.H. (2014). Arabidopsis AL PHD-PRC1 complexes promote 1036 seed germination through H3K4me3-to-H3K27me3 chromatin state switch in repression of 1037 seed developmental genes. PLoS genetics 10, e1004091. 1038
Mozgova, I., and Hennig, L. (2015). The polycomb group protein regulatory network. Annual review of 1039 plant biology 66, 269-296. 1040
Mu, Y., Zou, M.J., Sun, X.W., He, B.Y., Xu, X.M., Liu, Y.N., Zhang, L.X., and Chi, W. (2017). BASIC 1041 PENTACYSTEINE Proteins Repress ABSCISIC ACID INSENSITIVE4 Expression via Direct 1042 Recruitment of the Polycomb-Repressive Complex 2 in Arabidopsis Root Development. Plant 1043 Cell Physiol 58, 607-621. 1044
Mukherjee, K., Brocchieri, L., and Burglin, T.R. (2009). A comprehensive classification and 1045 evolutionary analysis of plant homeobox genes. Molecular biology and evolution 26, 1046 2775-2794. 1047
Ni, M., Cui, D., Einstein, J., Narasimhulu, S., Vergara, C.E., and Gelvin, S.B. (1995). Strength and 1048 Tissue-Specificity of Chimeric Promoters Derived from the Octopine and Mannopine Synthase 1049 Genes. The Plant journal 7, 661-676. 1050
Nonogaki, H. (2019). Seed germination and dormancy: The classic story, new puzzles, and evolution. 1051 Journal of integrative plant biology 61, 541-563. 1052
O'Malley, R.C., Huang, S.C., Song, L., Lewsey, M.G., Bartlett, A., Nery, J.R., Galli, M., Gallavotti, A., 1053 and Ecker, J.R. (2016). Cistrome and Epicistrome Features Shape the Regulatory DNA 1054 Landscape. Cell 166, 1598. 1055
Park, S.Y., Fung, P., Nishimura, N., Jensen, D.R., Fujii, H., Zhao, Y., Lumba, S., Santiago, J., Rodrigues, 1056 A., Chow, T.F.F., Alfred, S.E., Bonetta, D., Finkelstein, R., Provart, N.J., Desveaux, D., 1057 Rodriguez, P.L., McCourt, P., Zhu, J.K., Schroeder, J.I., Volkman, B.F., and Cutler, S.R. (2009). 1058 Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. 1059 Science 324, 1068-1071. 1060
Peirats-Llobet, M., Han, S.K., Gonzalez-Guzman, M., Jeong, C.W., Rodriguez, L., Belda-Palazon, B., 1061 Wagner, D., and Rodriguez, P.L. (2016). A Direct Link between Abscisic Acid Sensing and the 1062 Chromatin-Remodeling ATPase BRAHMA via Core ABA Signaling Pathway Components. 1063 Molecular plant 9, 136-147. 1064
Promchuea, S., Zhu, Y.J., Chen, Z.Z., Zhang, J., and Gong, Z.Z. (2017). ARF2 coordinates with 1065 PLETHORAs and PINs to orchestrate ABA-mediated root meristem activity in Arabidopsis. 1066 Journal of integrative plant biology 59, 30-43. 1067
Qi, J.S., Song, C.P., Wang, B.S., Zhou, J.M., Kangasjarvi, J., Zhu, J.K., and Gong, Z.Z. (2018). Reactive 1068 oxygen species signaling and stomatal movement in plant responses to drought stress and 1069 pathogen attack. Journal of integrative plant biology 60, 805-826. 1070
Ren, X., Chen, Z., Liu, Y., Zhang, H., Zhang, M., Liu, Q., Hong, X., Zhu, J.K., and Gong, Z. (2010). ABO3, 1071 a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance 1072 in Arabidopsis. The Plant journal : for cell and molecular biology 63, 417-429. 1073
Rushton, D.L., Tripathi, P., Rabara, R.C., Lin, J., Ringler, P., Boken, A.K., Langum, T.J., Smidt, L., 1074 Boomsma, D.D., Emme, N.J., Chen, X., Finer, J.J., Shen, Q.X.J., and Rushton, P.J. (2012). 1075 WRKY transcription factors: key components in abscisic acid signalling. Plant biotechnology 1076 journal 10, 2-11. 1077
Saeed, A.I., Sharov, V., White, J., Li, J., Liang, W., Bhagabati, N., Braisted, J., Klapa, M., Currier, T., 1078 Thiagarajan, M., Sturn, A., Snuffin, M., Rezantsev, A., Popov, D., Ryltsov, A., Kostukovich, E., 1079 Borisovsky, I., Liu, Z., Vinsavich, A., Trush, V., and Quackenbush, J. (2003). TM4: A free, 1080

32
open-source system for microarray data management and analysis. Biotechniques 34, 374-+. 1081 Saez, A., Rodrigues, A., Santiago, J., Rubio, S., and Rodriguez, P.L. (2008). HAB1-SWI3B Interaction 1082
Reveals a Link between Abscisic Acid Signaling and Putative SWI/SNF Chromatin-Remodeling 1083 Complexes in Arabidopsis. The Plant cell 20, 2972-2988. 1084
Sanchez-Pulido, L., Devos, D., Sung, Z.R., and Calonje, M. (2008). RAWUL: a new ubiquitin-like 1085 domain in PRC1 ring finger proteins that unveils putative plant and worm PRC1 orthologs. 1086 BMC genomics 9, 308. 1087
Shao, Z., Raible, F., Mollaaghababa, R., Guyon, J.R., Wu, C.T., Bender, W., and Kingston, R.E. (1999). 1088 Stabilization of chromatin structure by PRC1, a Polycomb complex. Cell 98, 37-46. 1089
Shen, L.S., Thong, Z.H., Gong, X.M., Shen, Q., Gan, Y.B., and Yu, H. (2014). The putative PRC1 1090 RING-finger protein AtRING1A regulates flowering through repressing MADS AFFECTING 1091 FLOWERING genes in Arabidopsis. Development 141, 1303-1312. 1092
Shkolnik-Inbar, D., and Bar-Zvi, D. (2010). ABI4 mediates abscisic acid and cytokinin inhibition of 1093 lateral root formation by reducing polar auxin transport in Arabidopsis. The Plant cell 22, 1094 3560-3573. 1095
Shu, K., Zhang, H., Wang, S., Chen, M., Wu, Y., Tang, S., Liu, C., Feng, Y., Cao, X., and Xie, Q. (2013). 1096 ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and 1097 gibberellins in arabidopsis. PLoS genetics 9, e1003577. 1098
Shu, K., Chen, Q., Wu, Y., Liu, R., Zhang, H., Wang, P., Li, Y., Wang, S., Tang, S., Liu, C., Yang, W., Cao, 1099 X., Serino, G., and Xie, Q. (2016). ABI4 mediates antagonistic effects of abscisic acid and 1100 gibberellins at transcript and protein levels. The Plant journal : for cell and molecular biology 1101 85, 348-361. 1102
Singh, D., and Laxmi, A. (2015). Transcriptional regulation of drought response: a tortuous network of 1103 transcriptional factors. Frontiers in plant science 6, 895. 1104
Soderman, E.M., Brocard, I.M., Lynch, T.J., and Finkelstein, R.R. (2000). Regulation and function of 1105 the Arabidopsis ABA-insensitive4 gene in seed and abscisic acid response signaling networks. 1106 Plant physiology 124, 1752-1765. 1107
Sun, Q., Csorba, T., Skourti-Stathaki, K., Proudfoot, N.J., and Dean, C. (2013). R-loop stabilization 1108 represses antisense transcription at the Arabidopsis FLC locus. Science 340, 619-621. 1109
Trapnell, C., Hendrickson, D.G., Sauvageau, M., Goff, L., Rinn, J.L., and Pachter, L. (2013). Differential 1110 analysis of gene regulation at transcript resolution with RNA-seq. Nature biotechnology 31, 1111 46-53. 1112
Turck, F., Roudier, F., Farrona, S., Martin-Magniette, M.-L., Guillaume, E., Buisine, N., Gagnot, S., 1113 Martienssen, R.A., Coupland, G., and Colot, V. (2007). Arabidopsis TFL2/LHP1 Specifically 1114 Associates with Genes Marked by Trimethylation of Histone H3 Lysine 27. PLoS genetics 3, 1115 e86. 1116
Uno, Y., Furihata, T., Abe, H., Yoshida, R., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000). 1117 Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent 1118 signal transduction pathway under drought and high-salinity conditions. Proceedings of the 1119 National Academy of Sciences of the United States of America 97, 11632-11637. 1120
Wang, K., He, J., Zhao, Y., Wu, T., Zhou, X., Ding, Y., Kong, L., Wang, X., Wang, Y., Li, J., Song, C.P., 1121 Wang, B., Yang, S., Zhu, J.K., and Gong, Z. (2018). EAR1 Negatively Regulates ABA Signaling 1122 by Enhancing 2C Protein Phosphatase Activity. The Plant cell 30, 815-834. 1123
Wang, L., Hua, D.P., He, J.N., Duan, Y., Chen, Z.Z., Hong, X.H., and Gong, Z.Z. (2011). Auxin Response 1124 Factor2 (ARF2) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response 1125 in Arabidopsis. PLoS genetics7, e1002172. 1126
Wang, L., Zhou, C.M., Mai, Y.X., Li, L.Z., Gao, J., Shang, G.D., Lian, H., Han, L., Zhang, T.Q., Tang, H.B., 1127 Ren, H., Wang, F.X., Wu, L.Y., Liu, X.L., Wang, C.S., Chen, E.W., Zhang, X.N., Liu, C., and Wang, 1128 J.W. (2019). A spatiotemporally regulated transcriptional complex underlies heteroblastic 1129 development of leaf hairs in Arabidopsis thaliana. The EMBO journal 38, e100063. 1130
Wang, Q., and Shen, W.H. (2018). Chromatin modulation and gene regulation in plants: insight about 1131 PRC1 function. Biochemical society transactions 46, 957-966. 1132
Wang, Z.P., Xing, H.L., Dong, L., Zhang, H.Y., Han, C.Y., Wang, X.C., and Chen, Q.J. (2015). Egg 1133 cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for 1134 multiple target genes in Arabidopsis in a single generation. Genome biology 16, 144. 1135
Wind, J.J., Peviani, A., Snel, B., Hanson, J., and Smeekens, S.C. (2013). ABI4: versatile activator and 1136 repressor. Trends in plant science 18, 125-132. 1137

33
Xiao, J., Jin, R., Yu, X., Shen, M., Wagner, J.D., Pai, A., Song, C., Zhuang, M., Klasfeld, S., He, C.S., 1138 Santos, A.M., Helliwell, C., Pruneda-Paz, J.L., Kay, S.A., Lin, X.W., Cui, S.J., Garcia, M.F., 1139 Clarenz, O., Goodrich, J., Zhang, X.Y., Austin, R.S., Bonasio, R., and Wagner, D. (2017). Cis 1140 and trans determinants of epigenetic silencing by Polycomb repressive complex 2 in 1141 Arabidopsis. Nature genetics 49, 1546-+. 1142
Xu, L., and Shen, W.H. (2008). Polycomb silencing of KNOX genes confines shoot stem cell niches in 1143 Arabidopsis. Current biology : CB 18, 1966-1971. 1144
Xu, W., Xu, H., Li, K., Fan, Y., Liu, Y., Yang, X., and Sun, Q. (2017). The R-loop is a common chromatin 1145 feature of the Arabidopsis genome. Nature plants 3, 704-714. 1146
Yang, L., Zhang, J., He, J.N., Qin, Y.Y., Hua, D.P., Duan, Y., Chen, Z.Z., and Gong, Z.Z. (2014). 1147 ABA-Mediated ROS in Mitochondria Regulate Root Meristem Activity by Controlling 1148 PLETHORA Expression in Arabidopsis. PLoS genetics 10, e1004791. 1149
Yang, Z.L., Qian, S.M., Scheid, R.N., Lu, L., Chen, X.S., Liu, R., Du, X.A., Lv, X.C., Boersma, M.D., Scalf, 1150 M., Smith, L.M., Denu, J.M., Du, J.M., and Zhong, X.H. (2018). EBS is a bivalent histone 1151 reader that regulates floral phase transition in Arabidopsis. Nature genetics 50, 1247-+. 1152
Yin, H., Zhang, X., Liu, J., Wang, Y., He, J., Yang, T., Hong, X., Yang, Q., and Gong, Z. (2009). Epigenetic 1153 regulation, somatic homologous recombination, and abscisic acid signaling are influenced by 1154 DNA polymerase epsilon mutation in Arabidopsis. The Plant cell 21, 386-402. 1155
Yuan, L., Liu, X., Luo, M., Yang, S., and Wu, K. (2013). Involvement of histone modifications in plant 1156 abiotic stress responses. Journal of integrative plant biology 55, 892-901. 1157
Zhang, J.X., Xie, S.J., Cheng, J.K., Lai, J.S., Zhu, J.K., and Gong, Z.Z. (2016). The Second Subunit of DNA 1158 Polymerase Delta Is Required for Genomic Stability and Epigenetic Regulation. Plant 1159 physiology 171, 1192-1208. 1160
Zhang, X., Germann, S., Blus, B.J., Khorasanizadeh, S., Gaudin, V., and Jacobsen, S.E. (2007). The 1161 Arabidopsis LHP1 protein colocalizes with histone H3 Lys27 trimethylation. Nature structural 1162 & molecular biology 14, 869-871. 1163
Zhang, Y., Liu, T., Meyer, C.A., Eeckhoute, J., Johnson, D.S., Bernstein, B.E., Nussbaum, C., Myers, 1164 R.M., Brown, M., Li, W., and Liu, X.S. (2008). Model-based Analysis of ChIP-Seq (MACS). 1165 Genome biology 9, R137. 1166
Zhao, Y., Gao, J.H., Kim, J.I., Chen, K., Bressan, R.A., and Zhu, J.K. (2017). Control of Plant Water Use 1167 by ABA Induction of Senescence and Dormancy: An Overlooked Lesson from Evolution. Plant 1168 and cell physiology 58, 1319-1327. 1169
Zhou, J.X., Liu, Z.W., Li, Y.Q., Li, L., Wang, B.J., Chen, S., and He, X.J. (2018). Arabidopsis PWWP 1170 domain proteins mediate H3K27 trimethylation on FLC and regulate flowering time. Journal 1171 of integrative plant biology 60, 362-368. 1172
Zhou, Y., Romero-Campero, F.J., Gomez-Zambrano, A., Turck, F., and Calonje, M. (2017). H2A 1173 monoubiquitination in Arabidopsis thaliana is generally independent of LHP1 and PRC2 1174 activity. Genome biology 18, 69. 1175
Zhu, J.K. (2016). Abiotic Stress Signaling and Responses in Plants. Cell 167, 313-324. 1176 Zhu, Y., Luo, X., Liu, X., Wu, W., Cui, X., He, Y., and Huang, J. (2019). Arabidopsis PEAPODs function 1177
with LIKE HETEROCHROMATIN PROTEIN1 to regulate lateral organ growth. Journal of 1178 integrative plant biology. 1179
Zuo, J., Niu, Q.W., and Chua, N.H. (2000). Technical advance: An estrogen receptor-based 1180 transactivator XVE mediates highly inducible gene expression in transgenic plants. The Plant 1181 journal : for cell and molecular biology 24, 265-273. 1182
1183
1184
1185

34
Figure 1. ndx mutants are hypersensitive to ABA in primary root growth and 1186
seedling establishment. 1187
(A) Primary root growth of ndx mutants is hypersensitive to ABA compared with the 1188
wild type. 5-d-old seedlings grown on MS medium were transferred to MS medium 1189
supplemented with 30 μM, 60 μM, 90 μM ABA for 4 days before photographed. Bar 1190
= 1 cm. The phenotype in 60 μM ABA was shown. ear1-1 served as a control. 1191
(B) Statistical analysis of relative root growth with different concentrations of ABA. 1192
The root growth of wild type and ndx mutants on MS medium without ABA was set 1193
to 100%. Error bars represent ± SE of fifteen seedlings from 3 plates in one 1194
representative experiment, and three independent experiments were done with similar 1195
results. 1196
(C) Seedling establishment of ndx mutants is hypersensitive to ABA. The seeds were 1197
germinated on MS medium supplemented with different concentrations of ABA for 9 1198
days before photographed. 1199
(D) The seedling greening ratio in (C). Error bars represent ± SE of about 90 seeds 1200
from three plates for one experiment, and three independent experiments were done 1201
with similar results. * represents significant difference compared with wild type, * P < 1202
0.05, ** P < 0.01, Student’s t-test. 1203
Figure 2. The expression of AtNDX is inhibited by ABA. 1204
(A) Relative transcript levels of AtNDX in 7-d-old wild type seedlings treated with 0 1205
μM or 30 μM ABA for 3 hours. Error bars represent ± SE of three technical replicates 1206
from one experiment, and three independent experiments were done with similar 1207
results. 1208
(B) Representative GUS staining results of proAtNDX:GUS transgenic plants treated 1209
with 0 μM or 30 μM ABA for 12 hours. 1210
(C) The fluorescence of AtNDX-GFP is reduced after ABA treatment. 5-d-old 1211
seedlings grown on MS medium were transferred to MS medium supplemented with 1212
30 μM ABA for 12 hours before observed. Bar = 50 μm. 1213
(D) Statistical analysis of the fluorescence of meristem zone in (C). The fluorescence 1214
was quantified by ImageJ software, and error bars represent ± SE of ten seedlings 1215
from 3 plates in one representative experiment, and three independent experiments 1216

35
were done with similar results. 1217
(E) Immunoblot assays indicate that AtNDX protein level is reduced after ABA1218
treatment. 7-d-old seedlings were treated by 0 μM (CK) or 50 μM ABA for 12 hours 1219
(top left), 100 μM CHX or 100 μM CHX +50 μM ABA for 12 hours (top right), 0 μM 1220
(CK) or 50 μM MG132 for 12 hours. Then the total proteins were extracted and 1221
detected with anti-NDX antibodies. GST-AtNDX-C purified from E. coli was used as 1222
a positive control. Actin protein serves as an loading control. The numbers indicate 1223
the relative gray value. 1224
Figure 3. NDX interacts with PRC1 core components AtRING1A and AtRING1B 1225
in vitro and in vivo. 1226
(A) AtNDX interacts with AtRING1A and AtRING1B in the yeast two-hybrid assay.1227
AD: Gal4 activation domain; BD: Gal4 DNA-binding domain. -LW: synthetic 1228
dropout medium without Leu and Trp; -LWHA: synthetic dropout medium without 1229
Leu, Trp, His and Ade. 1230
(B) Co-IP assay indicates that AtNDX interacts with AtRING1A and AtRING1B in1231
Arabidopsis protoplasts. The total proteins were isolated from protoplasts 1232
co-expressing HA-Flag-AtNDX and AtRING1A-GFP or AtRING1B-GFP proteins. 1233
Co-IP was carried out with GFP agarose beads and immunoblotting was conducted 1234
with anti-HA and anti-GFP antibodies. 1235
(C) Co-IP assay shows that AtNDX interacts with AtRING1A in two independent1236
transgenic lines. The total proteins were isolated from 10-day-old 1237
proSuper:AtRING1A-GFP transgenic seedlings (L1, L2) and Co-IP was carried out 1238
with GFP agarose beads. Immunoblotting was conducted with anti-NDX and 1239
anti-GFP antibodies. 1240
(D) Mass spectrometry analysis shows that AtRING1A co-purified with AtNDX. The1241
total proteins extracted from 10-d-old wild type and ndx-5 seedlings were 1242
immunoprecipitated with anti-NDX antibodies and the immunoprecipitated proteins 1243
were analyzed by LC-MS/MS. SC means sequence coverage of the protein. 1244
(E) The C-terminus of AtRING1A/B interacts with AtNDX in a yeast two-hybrid1245
assay. The structures of AtRING1A/B proteins and the diagrams of truncated proteins 1246
were shown on top. 1247

36
(F) The NDXA and NDXB domains of AtNDX are both required for interacting with1248
AtRING1A/B. The structure of AtNDX protein and the diagrams of truncated proteins 1249
were shown on top. 1250
Figure 4. The atring1a atring1b double mutants are hypersensitive to ABA in 1251
seedling establishment and primary root growth. 1252
(A) Primary root growth of atring1a atring1b (1a 1b) double mutant is hypersensitive1253
to ABA compared with the wild type. 4-d-old seedlings grown on MS medium were 1254
transferred to MS medium supplemented with 30 μM ABA for 3 days before 1255
photographed. Bar = 1 cm. 1256
(B) Statistical analysis of primary root growth in (A). Error bars represent ± SE of1257
forty five seedlings from three biological repeats. 1258
(C) Statistical analysis of relative root growth in (A). The root growth of wild type1259
and double mutant on MS medium without ABA was set to 100%. Error bars 1260
represent ± SE of forty five seedlings from three biological repeats. 1261
(D) Primary root growth of atring1a-2 atring1b-3 (1a-2 1b-3) double mutant is1262
hypersensitive to ABA compared with the wild type. 4-d-old seedlings grown on MS 1263
medium were transferred to MS medium supplemented with 60 μM ABA for 4 days 1264
before photographed. Bar = 1 cm. 1265
(E) Statistical analysis of relative root growth in different concentrations of ABA. The1266
primary root growth of the wild type and atring1a-2 atring1b-3 double mutant (1a-2 1267
1b-3) on MS medium without ABA was set to 100%. ear1-1 served as the positive 1268
control. Error bars represent ± SE of ten seedlings from one representative experiment, 1269
and three independent experiments were done with similar results. 1270
(F) Seedling establishment of atring1a-2 atring1b-3 double mutant (1a-2 1b-3) is1271
hypersensitive to ABA. The seeds were germinated on MS medium supplemented 1272
with 0.5 μM ABA for 9 days before photographed. 1273
(G) The seedling greening ratio in (F). Error bars represent ± SE of about 90 seeds1274
from three plates for one experiment, and three independent experiments were done 1275
with similar results. * Represents significant difference compared with the wild type, 1276
** P < 0.01, Student’s t-test. 1277
Figure 5. AtNDX and AtRING1A/B co-regulate ABI4 and a series of 1278

37
ABA-responsive genes. 1279
(A) Venn diagram shows the number of overlapped up- and down-regulated DEGs in 1280
ndx-4 and atring1a atring1b double mutant (1a 1b) compared with the wild type 1281
under control conditions. 1282
(B) GO annotation analysis of AtNDX and AtRING1A/B co-regulated genes. The 1283
abscissa represents a part of GO terms with significant enrichment, the ordinate 1284
represents the ratio of the genes enriched in the GO term to the total number of genes, 1285
and the number represents the E value of the significance analysis compared to the 1286
reference genome (TAIR10, 2017). 1287
(C) Venn diagram shows the number of overlapped up- and down-regulated DEGs in 1288
ndx-4 and atring1a atring1b double mutant (1a 1b) compared with the wild type 1289
under ABA treatment conditions. 7-d-old seedlings were treated with 0 or 60 μM 1290
ABA for 3 hours. 1291
(D) Venn diagram shows the overlapped number of co-up- and down-regulated genes 1292
under normal condition in ndx-4 and atring1a atring1b double mutant (1a 1b), which 1293
are ABA responsive genes in the wild type. 1294
(E) Heat map shows that the ABA responsive genes co-regulated by AtNDX and 1295
AtRING1A/B in (D) were mainly up-regulated in the ndx-4 and atring1a atring1b 1296
double mutant. The unit of expression is FPKM. 1297
(F) Relative expression level analyses of ABA related genes in the ndx-4 and atring1a 1298
atring1b double mutant (1a 1b). For each sample, 7-day-old seedlings were treated 1299
with 0 μM ABA (Control) or 60 μM ABA (ABA) for 3 hours, then the total RNAs 1300
were extracted. Error bars represent ± SE of three technical replicates from one 1301
experiment, and three independent experiments were done with similar results. * 1302
Represents significant difference compared with wild type, ** P < 0.01, Student’s 1303
t-test. 1304
Figure 6. AtNDX and AtRING1A/B regulate H2Aub levels of ABI4 and 1305
ABA-responsive genes. 1306
(A) Immunoblot assay indicates that the H2Aub levels in ndx mutants are reduced 1307
compared to the wild type. The nuclear proteins of 10-d-old seedlings were extracted 1308
and detected with H2Aub antibody, and H3 was used as an internal control. 1309

38
(B) Statistical analysis of relative band intensity in (A). The band intensity was 1310
quantified by ImageJ software, and the ratio of H2Aub to H3 in wild type was set to 1. 1311
Error bars represent ± SE of three biological replicates. ** P < 0.01, Student’s t-test. 1312
(C) Venn diagram shows the number of overlapped genes between AtNDX and 1313
AtRING1A/B co-regulated ABA responsive genes and H2Aub marked genes. 1314
(D) ChIP-qPCR analysis shows that the H2Aub levels of ABI4 and ABA responsive 1315
genes are reduced in ndx-5 and atring1a atring1b double mutant (1a 1b). The checked 1316
regions are labeled with the red lines below the gene diagrams in Supplemental Figure 1317
9. Error bars represent ± SE of three independent experiments. * Represent significant 1318
difference compared with wild type, *P < 0.05, **P < 0.01, Student’s t-test. 1319
Figure 7. ABI4 mutations rescue the ABA hypersensitive phonotype of ndx-5 and 1320
ABI4 overexpressing transgenic lines display ABA hypersensitive phonotype. 1321
(A) ndx-5 abi4-1 double mutant rescues the ABA hypersensitive phenotype of ndx-5 1322
in root growth. 5-d-old seedlings grown on MS medium were transferred to MS 1323
medium supplemented with 30 μM ABA for 4 days before photographed. Bar = 1 cm. 1324
(B) Statistical analysis of relative root growth with different concentrations of ABA 1325
treatment. The root growth of each genotype on MS medium without ABA was set to 1326
100%. Error bars represent ± SE of fifteen seedlings from one representative 1327
experiment, and three independent experiments were done with similar results. 1328
(C) ndx-5 abi4-1 double mutant rescues the ABA hypersensitive phenotype of ndx-5 1329
in seedling establishment. The seeds were germinated on MS medium supplemented 1330
with 0 or 0.5 μM ABA for 9 days before photographed. 1331
(D) The seedling greening ratio in (C). Error bars represent ± SE of about 90 seeds 1332
from three plates in one experiment, and three independent experiments were done 1333
with similar results. **P < 0.01, Student’s t-test. 1334
(E) Overexpression of ABI4 inhibits the root growth and confers ABA 1335
hypersensitivity. 5-days-old wild type and the transgenic wild type plants with 1336
inducible ABI4 overexpression were transferred to MS medium supplemented with 0 1337
or 30 μM ABA and without or with estradiol for 3 days before photographed. Bar = 1 1338
cm. 1339
(F) Statistical analysis of relative root growth in (E). The root growth of each 1340

39
genotype on MS medium without ABA and estradiol was set to 100%. Error bars 1341
represent ± SE of ten seedlings from three plates for one representative experiment, 1342
and three independent experiments were done with similar results. **P < 0.01, 1343
Student’s t-test. 1344
Figure 8. ABI4 is a direct target of AtNDX. 1345
(A) DAP-seq genome browser views show AtNDX binding signals around ABI3/4/5 1346
genes. 1347
(B) and (C) ChIP-qPCR analyses show that AtNDX binds to the ABI4 downstream 1348
sequence in vivo. The letters in (A) indicate the tested regions. DNA fragments 1349
immunoprecipitated by GFP beads were quantified by qPCR and normalized to the 1350
internal control ACT4; the relative enrichment in AtNDX-GFP over GFP is presented. 1351
Error bars represent ± SE of three technical replicates from one experiment, and three 1352
independent experiments were done with similar results. * Represent significant 1353
difference, **P < 0.01, Student’s t-test. 1354
(D) The probes used in EMSA. The sequence of ABI4-c-F probe is from the center of 1355
ABI4-c region in (A). The red letter represent TATA motif; the blue letters represent 1356
ATTA motif; mut1 and mut2 probes show the corresponding mutation sequence; F, 1357
forward strand DNA. 1358
(E) EMSA shows that recombinant GST-AtNDX-C binds to ABI4 downstream 1359
sequence in vitro. The upper and lower arrows indicate the bound probe and the free 1360
probe, respectively; ds, double-stranded DNA, indicates a double-stranded probe 1361
formed by annealing; F and R represent forward and reverse single-stranded probes. 1362
GST protein was used as a negative control. 1363
(F) The diagram of the binding peak of ABI4 downstream by AtNDX in DAP-seq and 1364
target location of CRISPR/Cas9. The sequence alignments show that the fragments 1365
from ABI4 downstream bases 624 to 1000 (counting after the first base of stop codon 1366
in ABI4) in abi4-T1 and the bases 605 to 968 in abi4-T2 were deleted, and there are 1367
insert mutant in target 2 in abi4-T2. These mutations almost cover the whole binding 1368
peak sequence of AtNDX. 1369
(G) abi4-T1 and abi4-T2 lines confer more ABA sensitivity than the wild type, but 1370
less ABA sensitive than ndx-4. 4-d-old seedlings grown on MS medium were 1371
transferred to MS medium supplemented 0 or 30 μM ABA for 4 days before 1372

40
photographed. Bar = 1 cm. 1373
(H) Statistical analysis of relative root growth in (G). The root growth of each1374
genotype on MS medium without ABA was set to 100%. Error bars represent ± SE of 1375
twelve seedlings from three plates for one representative experiment, and three 1376
independent experiments were done with similar results. **P < 0.01, Student’s t-test. 1377
(I) Relative transcript levels of ABI4 in 8-d-old seedlings. Seedlings were grown on1378
MS medium for 4 day, then transferred to MS medium supplemented 0 or 30 μM 1379
ABA for 4 day before harvested. Error bars represent ± SE of three technical 1380
replicates. ** P<0.01, Student’s t-test. 1381
Figure 9. A proposed model for the role of AtNDX and AtRING1A/B in ABA 1382
signaling. 1383
Under normal conditions (-ABA), AtNDX binds to the downstream region of ABI4 1384
and interacts with AtRING1A/B in the coding region, which promotes H2Aub 1385
modification and suppresses ABI4 expression. Under stress condition (+ABA), the 1386
expression of AtNDX is repressed by ABA signaling, which results in activation of the 1387
ABI4 expression. 1388
1389
1390
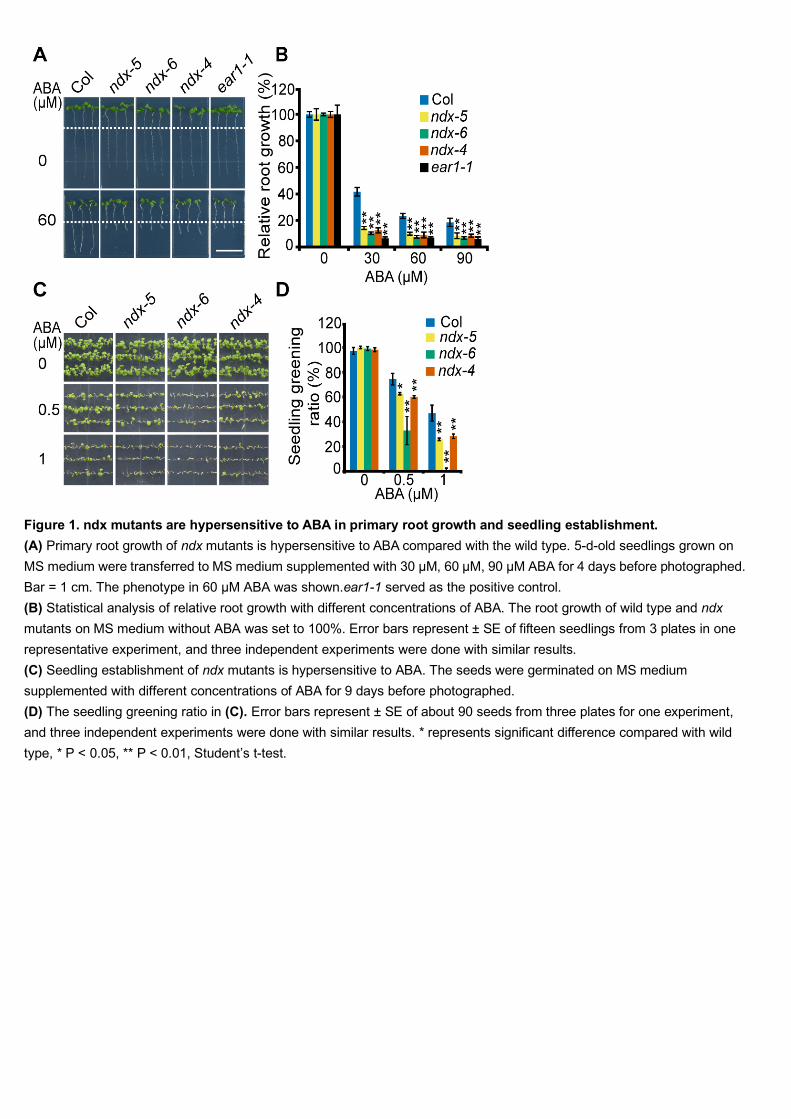
Figure 1. ndx mutants are hypersensitive to ABA in primary root growth and seedling establishment. (A) Primary root growth of ndx mutants is hypersensitive to ABA compared with the wild type. 5-d-old seedlings grown onMS medium were transferred to MS medium supplemented with 30 μM, 60 μM, 90 μM ABA for 4 days before photographed.Bar = 1 cm. The phenotype in 60 μM ABA was shown.ear1-1 served as the positive control.(B) Statistical analysis of relative root growth with different concentrations of ABA. The root growth of wild type and ndxmutants on MS medium without ABA was set to 100%. Error bars represent ± SE of fifteen seedlings from 3 plates in onerepresentative experiment, and three independent experiments were done with similar results.(C) Seedling establishment of ndx mutants is hypersensitive to ABA. The seeds were germinated on MS mediumsupplemented with different concentrations of ABA for 9 days before photographed.(D) The seedling greening ratio in (C). Error bars represent ± SE of about 90 seeds from three plates for one experiment,and three independent experiments were done with similar results. * represents significant difference compared with wildtype, * P < 0.05, ** P < 0.01, Student’s t-test.
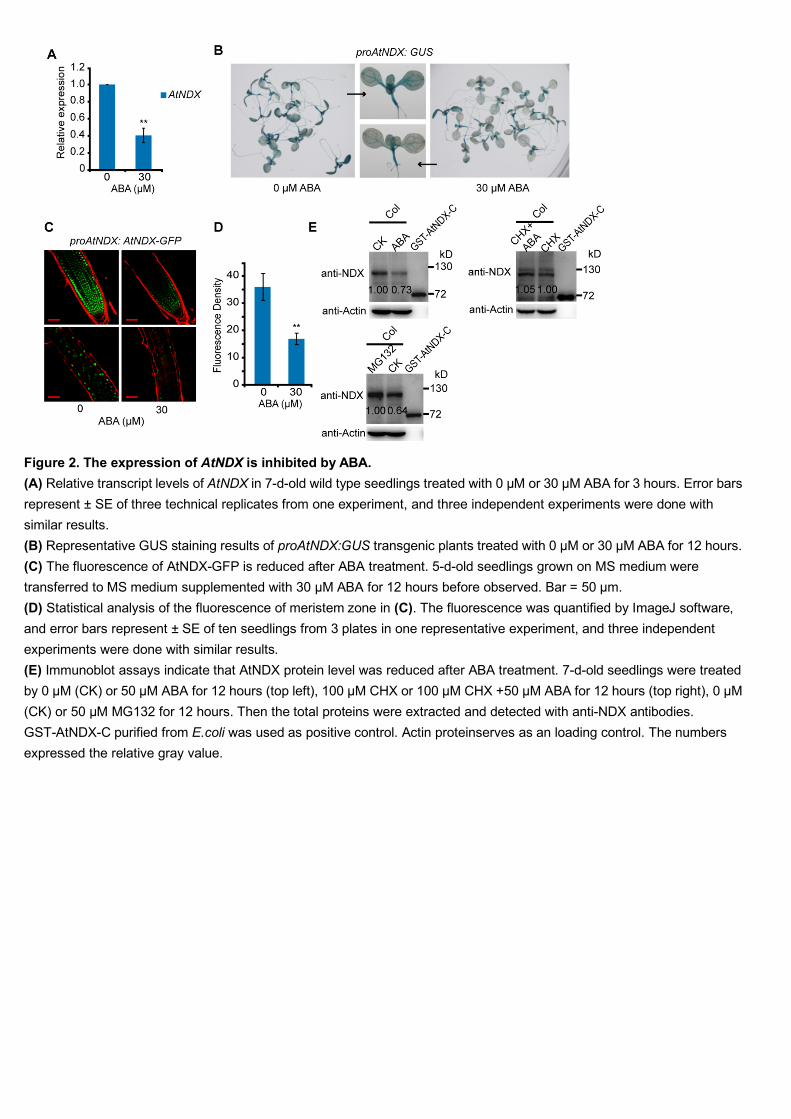
Figure 2. The expression of AtNDX is inhibited by ABA. (A) Relative transcript levels of AtNDX in 7-d-old wild type seedlings treated with 0 μM or 30 μM ABA for 3 hours. Error barsrepresent ± SE of three technical replicates from one experiment, and three independent experiments were done withsimilar results.(B) Representative GUS staining results of proAtNDX:GUS transgenic plants treated with 0 μM or 30 μM ABA for 12 hours.(C) The fluorescence of AtNDX-GFP is reduced after ABA treatment. 5-d-old seedlings grown on MS medium weretransferred to MS medium supplemented with 30 μM ABA for 12 hours before observed. Bar = 50 μm.(D) Statistical analysis of the fluorescence of meristem zone in (C). The fluorescence was quantified by ImageJ software,and error bars represent ± SE of ten seedlings from 3 plates in one representative experiment, and three independentexperiments were done with similar results.(E) Immunoblot assays indicate that AtNDX protein level was reduced after ABA treatment. 7-d-old seedlings were treatedby 0 μM (CK) or 50 μM ABA for 12 hours (top left), 100 μM CHX or 100 μM CHX +50 μM ABA for 12 hours (top right), 0 μM(CK) or 50 μM MG132 for 12 hours. Then the total proteins were extracted and detected with anti-NDX antibodies.GST-AtNDX-C purified from E.coli was used as positive control. Actin proteinserves as an loading control. The numbersexpressed the relative gray value.

Figure 3. NDX interacts with PRC1 core components AtRING1A and AtRING1B in vitro and in vivo. (A) AtNDX interacts with AtRING1A and AtRING1B in the yeast two-hybrid assay. AD: Gal4 activation domain; BD: Gal4DNA-binding domain. -LW: synthetic dropout medium without Leu and Trp; -LWHA: synthetic dropout medium without Leu,Trp, His and Ade.(B) Co-IP assay indicates that AtNDX interacts with AtRING1A and AtRING1B in Arabidopsis protoplasts. The total proteinswere isolated from protoplasts co-expressing HA-Flag-AtNDX and AtRING1A-GFP or AtRING1B-GFP proteins. Co-IP wascarried out with GFP agarose beads and immunoblotting was conducted with anti-HA and anti-GFP antibodies.(C) Co-IP assay shows that AtNDX interacts with AtRING1A in two independent transgenic lines. The total proteins wereisolated from 10-day-old proSuper:AtRING1A-GFP transgenic seedlings (L1, L2) and Co-IP was carried out with GFPagarose beads. Immunoblotting was conducted with anti-NDX and anti-GFP antibodies.(D) Mass spectrometry analysis shows that AtRING1A co-purified with AtNDX. The total proteins extracted from 10-d-oldwild type and ndx-5 seedlings were immunoprecipitated with anti-NDX antibodies and the immunoprecipitated proteinswere analyzed by LC-MS/MS. SC means sequence coverage of the protein.(E) The C-terminus of AtRING1A/B interacts with AtNDX in a yeast two-hybrid assay. The structures of AtRING1A/Bproteins and the diagrams of truncated proteins were shown on top.(F) The NDXA and NDXB domains of AtNDX are both required for interacting with AtRING1A/B. The structure of AtNDXprotein and the diagrams of truncated proteins were shown on top.
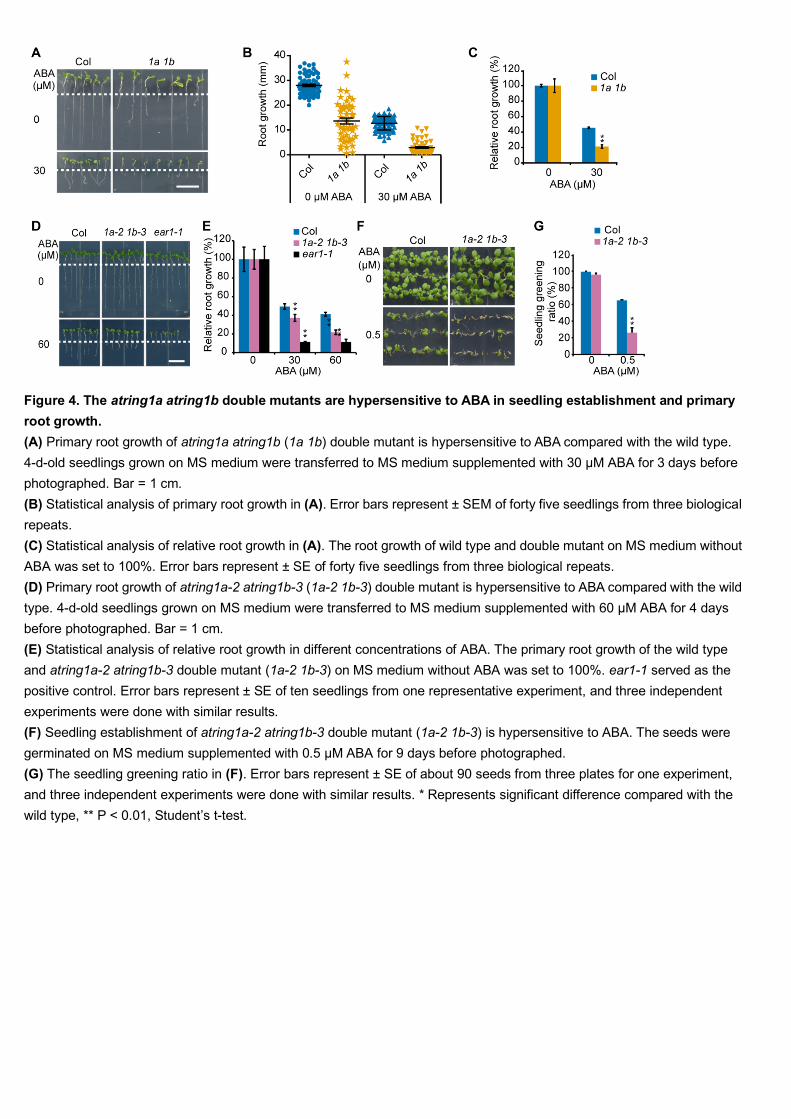
Figure 4. The atring1a atring1b double mutants are hypersensitive to ABA in seedling establishment and primary root growth. (A) Primary root growth of atring1a atring1b (1a 1b) double mutant is hypersensitive to ABA compared with the wild type.4-d-old seedlings grown on MS medium were transferred to MS medium supplemented with 30 μM ABA for 3 days beforephotographed. Bar = 1 cm.(B) Statistical analysis of primary root growth in (A). Error bars represent ± SEM of forty five seedlings from three biologicalrepeats.(C) Statistical analysis of relative root growth in (A). The root growth of wild type and double mutant on MS medium withoutABA was set to 100%. Error bars represent ± SE of forty five seedlings from three biological repeats.(D) Primary root growth of atring1a-2 atring1b-3 (1a-2 1b-3) double mutant is hypersensitive to ABA compared with the wildtype. 4-d-old seedlings grown on MS medium were transferred to MS medium supplemented with 60 μM ABA for 4 daysbefore photographed. Bar = 1 cm.(E) Statistical analysis of relative root growth in different concentrations of ABA. The primary root growth of the wild typeand atring1a-2 atring1b-3 double mutant (1a-2 1b-3) on MS medium without ABA was set to 100%. ear1-1 served as thepositive control. Error bars represent ± SE of ten seedlings from one representative experiment, and three independentexperiments were done with similar results.(F) Seedling establishment of atring1a-2 atring1b-3 double mutant (1a-2 1b-3) is hypersensitive to ABA. The seeds weregerminated on MS medium supplemented with 0.5 μM ABA for 9 days before photographed.(G) The seedling greening ratio in (F). Error bars represent ± SE of about 90 seeds from three plates for one experiment,and three independent experiments were done with similar results. * Represents significant difference compared with thewild type, ** P < 0.01, Student’s t-test.
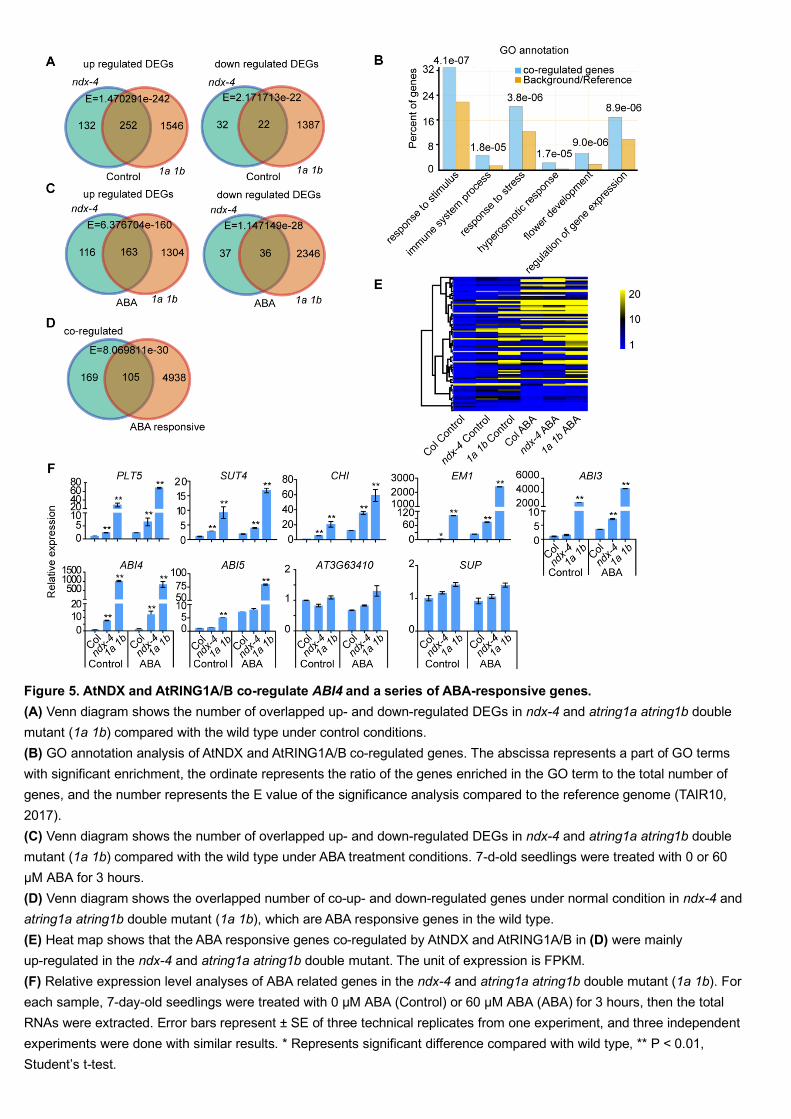
Figure 5. AtNDX and AtRING1A/B co-regulate ABI4 and a series of ABA-responsive genes. (A) Venn diagram shows the number of overlapped up- and down-regulated DEGs in ndx-4 and atring1a atring1b doublemutant (1a 1b) compared with the wild type under control conditions.(B) GO annotation analysis of AtNDX and AtRING1A/B co-regulated genes. The abscissa represents a part of GO termswith significant enrichment, the ordinate represents the ratio of the genes enriched in the GO term to the total number ofgenes, and the number represents the E value of the significance analysis compared to the reference genome (TAIR10,2017).(C) Venn diagram shows the number of overlapped up- and down-regulated DEGs in ndx-4 and atring1a atring1b doublemutant (1a 1b) compared with the wild type under ABA treatment conditions. 7-d-old seedlings were treated with 0 or 60μM ABA for 3 hours.(D) Venn diagram shows the overlapped number of co-up- and down-regulated genes under normal condition in ndx-4 andatring1a atring1b double mutant (1a 1b), which are ABA responsive genes in the wild type.(E) Heat map shows that the ABA responsive genes co-regulated by AtNDX and AtRING1A/B in (D) were mainlyup-regulated in the ndx-4 and atring1a atring1b double mutant. The unit of expression is FPKM.(F) Relative expression level analyses of ABA related genes in the ndx-4 and atring1a atring1b double mutant (1a 1b). Foreach sample, 7-day-old seedlings were treated with 0 μM ABA (Control) or 60 μM ABA (ABA) for 3 hours, then the totalRNAs were extracted. Error bars represent ± SE of three technical replicates from one experiment, and three independentexperiments were done with similar results. * Represents significant difference compared with wild type, ** P < 0.01,Student’s t-test.
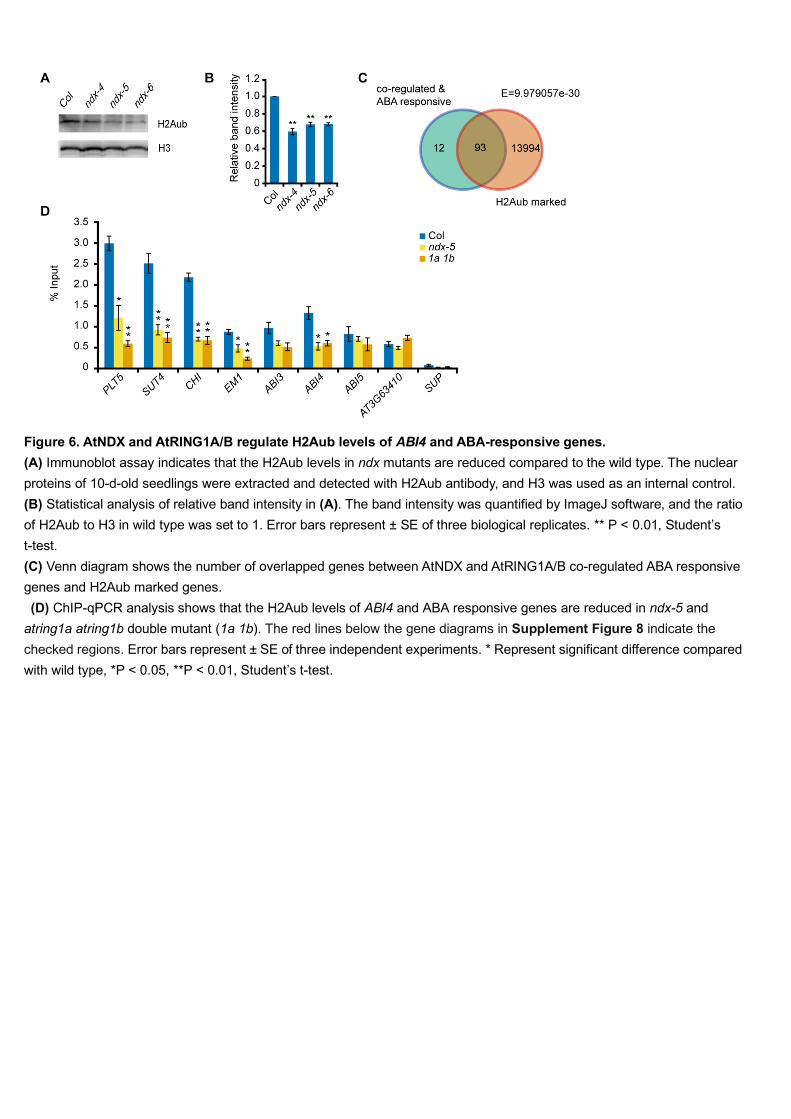
Figure 6. AtNDX and AtRING1A/B regulate H2Aub levels of ABI4 and ABA-responsive genes. (A) Immunoblot assay indicates that the H2Aub levels in ndx mutants are reduced compared to the wild type. The nuclearproteins of 10-d-old seedlings were extracted and detected with H2Aub antibody, and H3 was used as an internal control.(B) Statistical analysis of relative band intensity in (A). The band intensity was quantified by ImageJ software, and the ratioof H2Aub to H3 in wild type was set to 1. Error bars represent ± SE of three biological replicates. ** P < 0.01, Student’st-test.(C) Venn diagram shows the number of overlapped genes between AtNDX and AtRING1A/B co-regulated ABA responsivegenes and H2Aub marked genes.(D) ChIP-qPCR analysis shows that the H2Aub levels of ABI4 and ABA responsive genes are reduced in ndx-5 and
atring1a atring1b double mutant (1a 1b). The red lines below the gene diagrams in Supplement Figure 8 indicate the checked regions. Error bars represent ± SE of three independent experiments. * Represent significant difference compared with wild type, *P < 0.05, **P < 0.01, Student’s t-test.
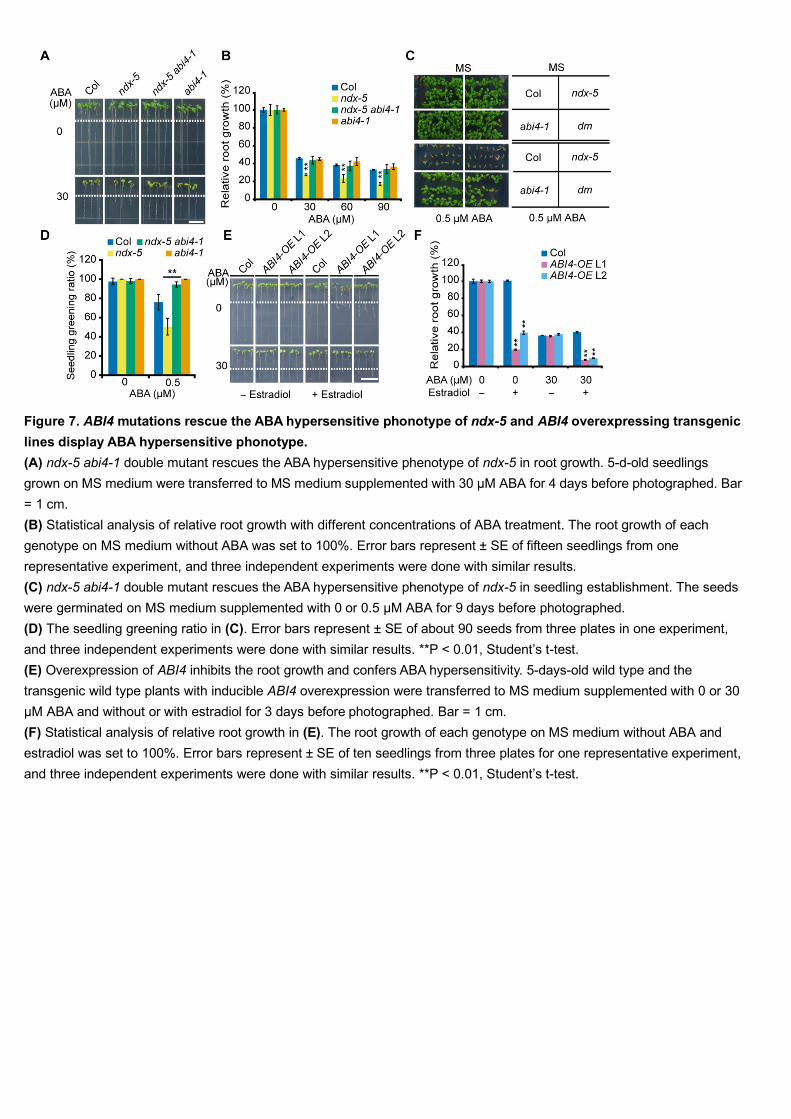
Figure 7. ABI4 mutations rescue the ABA hypersensitive phonotype of ndx-5 and ABI4 overexpressing transgenic lines display ABA hypersensitive phonotype. (A) ndx-5 abi4-1 double mutant rescues the ABA hypersensitive phenotype of ndx-5 in root growth. 5-d-old seedlingsgrown on MS medium were transferred to MS medium supplemented with 30 μM ABA for 4 days before photographed. Bar= 1 cm.(B) Statistical analysis of relative root growth with different concentrations of ABA treatment. The root growth of eachgenotype on MS medium without ABA was set to 100%. Error bars represent ± SE of fifteen seedlings from onerepresentative experiment, and three independent experiments were done with similar results.(C) ndx-5 abi4-1 double mutant rescues the ABA hypersensitive phenotype of ndx-5 in seedling establishment. The seedswere germinated on MS medium supplemented with 0 or 0.5 μM ABA for 9 days before photographed.(D) The seedling greening ratio in (C). Error bars represent ± SE of about 90 seeds from three plates in one experiment,and three independent experiments were done with similar results. **P < 0.01, Student’s t-test.(E) Overexpression of ABI4 inhibits the root growth and confers ABA hypersensitivity. 5-days-old wild type and thetransgenic wild type plants with inducible ABI4 overexpression were transferred to MS medium supplemented with 0 or 30μM ABA and without or with estradiol for 3 days before photographed. Bar = 1 cm.(F) Statistical analysis of relative root growth in (E). The root growth of each genotype on MS medium without ABA andestradiol was set to 100%. Error bars represent ± SE of ten seedlings from three plates for one representative experiment,and three independent experiments were done with similar results. **P < 0.01, Student’s t-test.
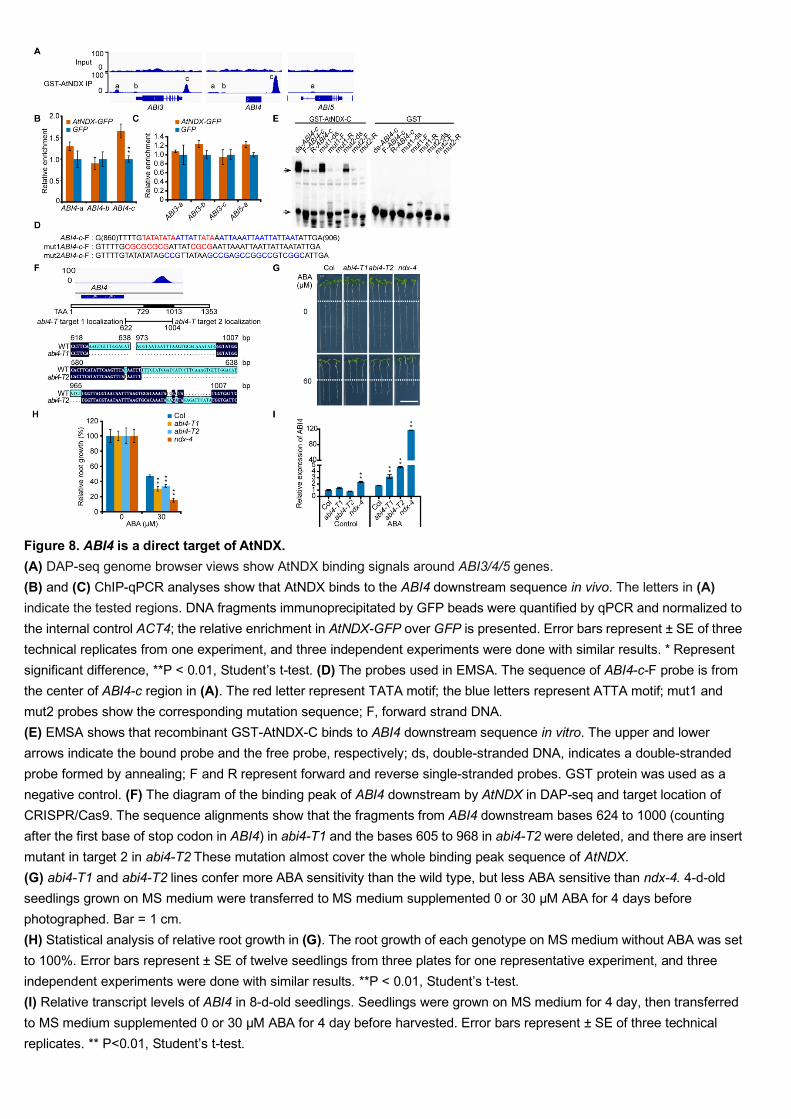
Figure 8. ABI4 is a direct target of AtNDX. (A) DAP-seq genome browser views show AtNDX binding signals around ABI3/4/5 genes.(B) and (C) ChIP-qPCR analyses show that AtNDX binds to the ABI4 downstream sequence in vivo. The letters in (A)indicate the tested regions. DNA fragments immunoprecipitated by GFP beads were quantified by qPCR and normalized tothe internal control ACT4; the relative enrichment in AtNDX-GFP over GFP is presented. Error bars represent ± SE of threetechnical replicates from one experiment, and three independent experiments were done with similar results. * Representsignificant difference, **P < 0.01, Student’s t-test. (D) The probes used in EMSA. The sequence of ABI4-c-F probe is fromthe center of ABI4-c region in (A). The red letter represent TATA motif; the blue letters represent ATTA motif; mut1 andmut2 probes show the corresponding mutation sequence; F, forward strand DNA.(E) EMSA shows that recombinant GST-AtNDX-C binds to ABI4 downstream sequence in vitro. The upper and lowerarrows indicate the bound probe and the free probe, respectively; ds, double-stranded DNA, indicates a double-strandedprobe formed by annealing; F and R represent forward and reverse single-stranded probes. GST protein was used as anegative control. (F) The diagram of the binding peak of ABI4 downstream by AtNDX in DAP-seq and target location ofCRISPR/Cas9. The sequence alignments show that the fragments from ABI4 downstream bases 624 to 1000 (countingafter the first base of stop codon in ABI4) in abi4-T1 and the bases 605 to 968 in abi4-T2 were deleted, and there are insertmutant in target 2 in abi4-T2 These mutation almost cover the whole binding peak sequence of AtNDX.(G) abi4-T1 and abi4-T2 lines confer more ABA sensitivity than the wild type, but less ABA sensitive than ndx-4. 4-d-oldseedlings grown on MS medium were transferred to MS medium supplemented 0 or 30 μM ABA for 4 days beforephotographed. Bar = 1 cm.(H) Statistical analysis of relative root growth in (G). The root growth of each genotype on MS medium without ABA was setto 100%. Error bars represent ± SE of twelve seedlings from three plates for one representative experiment, and threeindependent experiments were done with similar results. **P < 0.01, Student’s t-test.(I) Relative transcript levels of ABI4 in 8-d-old seedlings. Seedlings were grown on MS medium for 4 day, then transferredto MS medium supplemented 0 or 30 μM ABA for 4 day before harvested. Error bars represent ± SE of three technicalreplicates. ** P<0.01, Student’s t-test.
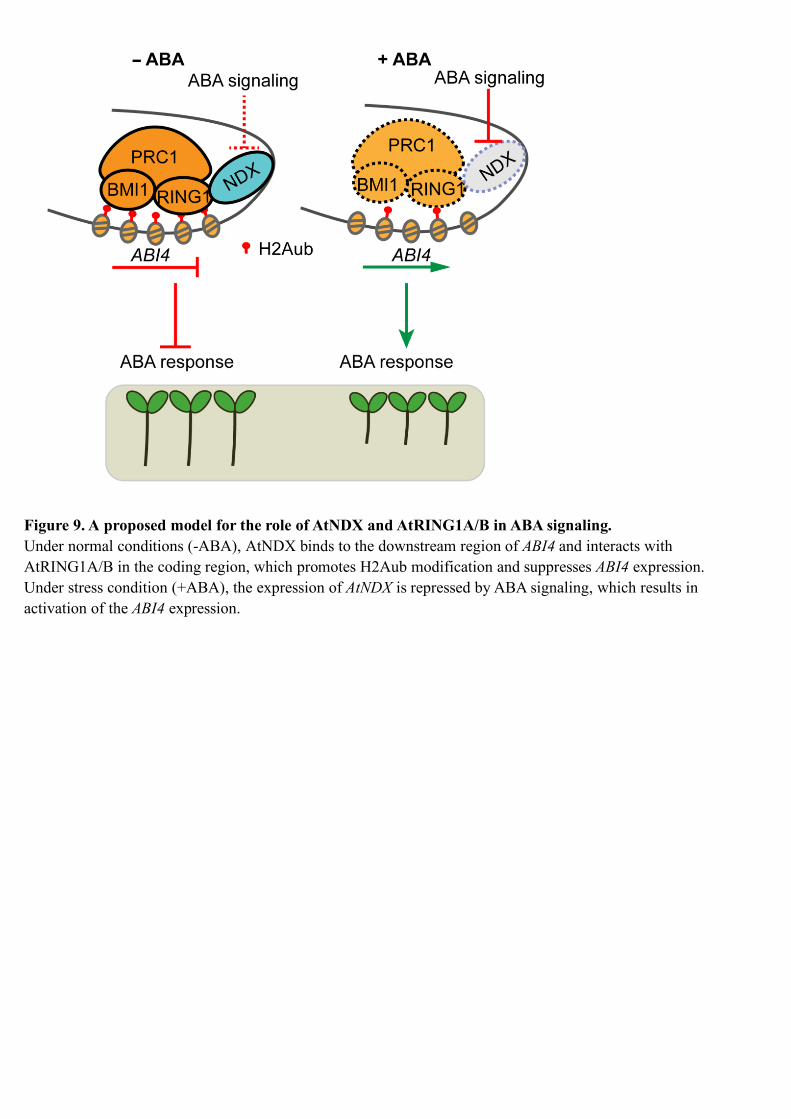
Figure 9. A proposed model for the role of AtNDX and AtRING1A/B in ABA signaling. Under normal conditions (-ABA), AtNDX binds to the downstream region of ABI4 and interacts with AtRING1A/B in the coding region, which promotes H2Aub modification and suppresses ABI4 expression. Under stress condition (+ABA), the expression of AtNDX is repressed by ABA signaling, which results in activation of the ABI4 expression.

Parsed CitationsAhmad, R., Liu, Y., Wang, T.J., Meng, Q.X., Yin, H., Wang, X., Wu, Y.F., Nan, N., Liu, B., and Xu, Z.Y. (2019). GOLDEN2-LIKE TranscriptionFactors Regulate WRKY40 Expression in Response to Abscisic Acid. Plant physiology 179, 1844-1860.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blackledge, N.P., Rose, N.R., and Klose, R.J. (2015). Targeting Polycomb systems to regulate gene expression: modifications to acomplex story. Nature reviews. Molecular cell biology 16, 643-649.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bossi, F., Cordoba, E., Dupre, P., Mendoza, M.S., Roman, C.S., and Leon, P. (2009). The Arabidopsis ABA-INSENSITIVE (ABI) 4 factoracts as a central transcription activator of the expression of its own gene, and for the induction of ABI5 and SBE2.2 genes duringsugar signaling. The Plant journal : for cell and molecular biology 59, 359-374.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bratzel, F., Lopez-Torrejon, G., Koch, M., Del Pozo, J.C., and Calonje, M. (2010). Keeping cell identity in Arabidopsis requires PRC1RING-finger homologs that catalyze H2A monoubiquitination. Current biology : CB 20, 1853-1859.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bratzel, F., Yang, C., Angelova, A., Lopez-Torrejon, G., Koch, M., del Pozo, J.C., and Calonje, M. (2012). Regulation of the newArabidopsis imprinted gene AtBMI1C requires the interplay of different epigenetic mechanisms. Molecular plant 5, 260-269.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Calonje, M. (2014). PRC1 marks the difference in plant PcG repression. Molecular plant 7, 459-471.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen, D., Molitor, A., Liu, C., and Shen, W.H. (2010). The Arabidopsis PRC1-like ring-finger proteins are necessary for repression ofembryonic traits during vegetative growth. Cell research 20, 1332-1344.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen, L., Zhang, L., Li, D., Wang, F., and Yu, D. (2013). WRKY8 transcription factor functions in the TMV-cg defense response bymediating both abscisic acid and ethylene signaling in Arabidopsis. Proceedings of the National Academy of Sciences of the UnitedStates of America 110, E1963-1971.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chinnusamy, V., Gong, Z.Z., and Zhu, J.K. (2008). Abscisic acid-mediated epigenetic processes in plant development and stressresponses. Journal of integrative plant biology 50, 1187-1195.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi, H., Hong, J., Ha, J., Kang, J., and Kim, S.Y. (2000). ABFs, a family of ABA-responsive element binding factors. The Journal ofbiological chemistry 275, 1723-1730.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Christensen, R.G., Enuameh, M.S., Noyes, M.B., Brodsky, M.H., Wolfe, S.A., and Stormo, G.D. (2012). Recognition models to predictDNA-binding specificities of homeodomain proteins. Bioinformatics 28, i84-89.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Crossley, M.P., Bocek, M., and Cimprich, K.A. (2019). R-Loops as Cellular Regulators and Genomic Threats. Molecular cell 73, 398-411.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cutler, S.R., Rodriguez, P.L., Finkelstein, R.R., and Abrams, S.R. (2010). Abscisic acid: emergence of a core signaling network. Annualreview of plant biology 61, 651-679.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dobin, A., Davis, C.A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., Batut, P., Chaisson, M., and Gingeras, T.R. (2013). STAR:ultrafast universal RNA-seq aligner. Bioinformatics 29, 15-21.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feng, C.Z., Chen, Y., Wang, C., Kong, Y.H., Wu, W.H., and Chen, Y.F. (2014). Arabidopsis RAV1 transcription factor, phosphorylated bySnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. The Plant

journal : for cell and molecular biology 80, 654-668.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Finkelstein, R.R., and Lynch, T.J. (2000). The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zippertranscription factor. The Plant cell 12, 599-609.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Finkelstein, R.R., Wang, M.L., Lynch, T.J., Rao, S., and Goodman, H.M. (1998). The Arabidopsis abscisic acid response locus ABI4encodes an APETALA 2 domain protein. The Plant cell 10, 1043-1054.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Francis, N.J., Saurin, A.J., Shao, Z., and Kingston, R.E. (2001). Reconstitution of a functional core polycomb repressive complex.Molecular cell 8, 545-556.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fujii, H., Verslues, P.E., and Zhu, J.K. (2007). Identification of two protein kinases required for abscisic acid regulation of seedgermination, root growth, and gene expression in Arabidopsis. The Plant cell 19, 485-494.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fujita, Y., Fujita, M., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2011). ABA-mediated transcriptional regulation in response to osmoticstress in plants. Journal of plant research 124, 509-525.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Giraudat, J., Hauge, B.M., Valon, C., Smalle, J., Parcy, F., and Goodman, H.M. (1992). Isolation of the Arabidopsis ABI3 gene bypositional cloning. The Plant cell 4, 1251-1261.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Han, S.K., Sang, Y., Rodrigues, A., Wu, M.F., Rodriguez, P.L., Wagner, D., and F2010, B. (2012). The SWI2/SNF2 Chromatin RemodelingATPase BRAHMA Represses Abscisic Acid Responses in the Absence of the Stress Stimulus in Arabidopsis. The Plant cell 24, 4892-4906.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
He, J.N., Duan, Y., Hua, D.P., Fan, G.J., Wang, L., Liu, Y., Chen, Z.Z., Han, L.H., Qu, L.J., and Gong, Z.Z. (2012). DEXH Box RNA Helicase-Mediated Mitochondrial Reactive Oxygen Species Production in Arabidopsis Mediates Crosstalk between Abscisic Acid and AuxinSignaling. The Plant cell 24, 1815-1833.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hua, D.P., Wang, C., He, J.N., Liao, H., Duan, Y., Zhu, Z.Q., Guo, Y., Chen, Z.Z., and Gong, Z.Z. (2012). A Plasma Membrane ReceptorKinase, GHR1, Mediates Abscisic Acid- and Hydrogen Peroxide-Regulated Stomatal Movement in Arabidopsis. The Plant cell 24, 2546-2561.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huang, C., Xu, M., and Zhu, B. (2013). Epigenetic inheritance mediated by histone lysine methylation: maintaining transcriptional stateswithout the precise restoration of marks? Philosophical transactions of the royal society of London series B-bilogical sciences 368,20110332.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huang, X., Zhang, X., Gong, Z., Yang, S., and Shi, Y. (2017). ABI4 represses the expression of type-A ARRs to inhibit seed germination inArabidopsis. The Plant journal : for cell and molecular biology 89, 354-365.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jiang, D.H., and Berger, F. (2017). DNA replication-coupled histone modification maintains Polycomb gene silencing in plants. Science357, 1146-+.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jorgensen, J.E., Gronlund, M., Pallisgaard, N., Larsen, K., Marcker, K.A., and Jensen, E.O. (1999). A new class of plant homeobox genesis expressed in specific regions of determinate symbiotic root nodules. Plant molecular biology 40, 65-77.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kerchev, P.I., Pellny, T.K., Vivancos, P.D., Kiddle, G., Hedden, P., Driscoll, S., Vanacker, H., Verrier, P., Hancock, R.D., and Foyer, C.H.

(2011). The transcription factor ABI4 Is required for the ascorbic acid-dependent regulation of growth and regulation of jasmonate-dependent defense signaling pathways in Arabidopsis. The Plant cell 23, 3319-3334.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kong, L., Cheng, J., Zhu, Y., Ding, Y., Meng, J., Chen, Z., Xie, Q., Guo, Y., Li, J., Yang, S., and Gong, Z. (2015). Degradation of the ABAco-receptor ABI1 by PUB12/13 U-box E3 ligases. Nature communications 6, 8630.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Langmead, B., and Salzberg, S.L. (2012). Fast gapped-read alignment with Bowtie 2. Nature methods 9, 357-U354.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Leung, J., Merlot, S., and Giraudat, J. (1997). The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encodehomologous protein phosphatases 2C involved in abscisic acid signal transduction. The Plant cell 9, 759-771.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, J., Wang, Z., Hu, Y., Cao, Y., and Ma, L. (2017). Polycomb Group Proteins RING1A and RING1B Regulate the Vegetative PhaseTransition in Arabidopsis. Frontiers in plant science 8, 867.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, P., Zhou, H., Shi, X., Yu, B., Zhou, Y., Chen, S., Wang, Y., Peng, Y., Meyer, R.C., Smeekens, S.C., and Teng, S. (2014). The ABI4-induced Arabidopsis ANAC060 transcription factor attenuates ABA signaling and renders seedlings sugar insensitive when present inthe nucleus. PLoS genetics 10, e1004213.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, Z., Fu, X., Wang, Y., Liu, R., and He, Y. (2018). Polycomb-mediated gene silencing by the BAH-EMF1 complex in plants. Naturegenetics 50, 1254-1261.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, Z.F., Li, B., Liu, J., Guo, Z.H., Liu, Y.H., Li, Y., Shen, W.H., Huang, Y., Huang, H., Zhang, Y.J., and Dong, A.W. (2016). Transcriptionfactors AS1 and AS2 interact with LHP1 to repress KNOX genes in Arabidopsis. Journal of integrative plant biology 58, 959-970.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, C., Wang, C., Wang, G., Becker, C., Zaidem, M., and Weigel, D. (2016). Genome-wide analysis of chromatin packing in Arabidopsisthaliana at single-gene resolution. Genome research 26, 1057-1068.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, C., Cheng, J., Zhuang, Y., Ye, L., Li, Z., Wang, Y., Qi, M., Xu, L., and Zhang, Y. (2018a). Polycomb repressive complex 2 attenuatesABA-induced senescence in Arabidopsis. The Plant journal : for cell and molecular biology 97, 368-377.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, N., Fromm, M., and Avramova, Z. (2014). H3K27me3 and H3K4me3 Chromatin Environment at Super-Induced Dehydration StressMemory Genes of Arabidopsis thaliana. Molecular plant 7, 502-513.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, Y., He, J.N., Chen, Z.Z., Ren, X.Z., Hong, X.H., and Gong, Z.Z. (2010). ABA overly-sensitive 5 (ABO5), encoding a pentatricopeptiderepeat protein required for cis-splicing of mitochondrial nad2 intron 3, is involved in the abscisic acid response in Arabidopsis. ThePlant journal : for cell and molecular biology 63, 749-765.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, Y.T., Zhang, A., Yin, H., Meng, Q.X., Yu, X.M., Huang, S.Z., Wang, J., Ahmad, R., Liu, B., and Xu, Z.Y. (2018b). Trithorax-groupproteins ARABIDOPSIS TRITHORAX4 (ATX4) and ATX5 function in abscisic acid and dehydration stress responses. New phytologist217, 1582-1597.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ma, Y., Szostkiewicz, I., Korte, A., Moes, D., Yang, Y., Christmann, A., and Grill, E. (2009). Regulators of PP2C phosphatase activityfunction as abscisic acid sensors. Science 324, 1064-1068.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Merini, W., and Calonje, M. (2015). PRC1 is taking the lead in PcG repression. The Plant journal : for cell and molecular biology 83, 110-120.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Molitor, A., and Shen, W.H. (2013). The polycomb complex PRC1: composition and function in plants. Journal of genetics and genomics40, 231-238.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Molitor, A.M., Bu, Z., Yu, Y., and Shen, W.H. (2014). Arabidopsis AL PHD-PRC1 complexes promote seed germination through H3K4me3-to-H3K27me3 chromatin state switch in repression of seed developmental genes. PLoS genetics 10, e1004091.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mozgova, I., and Hennig, L. (2015). The polycomb group protein regulatory network. Annual review of plant biology 66, 269-296.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mu, Y., Zou, M.J., Sun, X.W., He, B.Y., Xu, X.M., Liu, Y.N., Zhang, L.X., and Chi, W. (2017). BASIC PENTACYSTEINE Proteins RepressABSCISIC ACID INSENSITIVE4 Expression via Direct Recruitment of the Polycomb-Repressive Complex 2 in Arabidopsis RootDevelopment. Plant Cell Physiol 58, 607-621.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mukherjee, K., Brocchieri, L., and Burglin, T.R. (2009). A comprehensive classification and evolutionary analysis of plant homeoboxgenes. Molecular biology and evolution 26, 2775-2794.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ni, M., Cui, D., Einstein, J., Narasimhulu, S., Vergara, C.E., and Gelvin, S.B. (1995). Strength and Tissue-Specificity of ChimericPromoters Derived from the Octopine and Mannopine Synthase Genes. The Plant journal 7, 661-676.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nonogaki, H. (2019). Seed germination and dormancy: The classic story, new puzzles, and evolution. Journal of integrative plantbiology 61, 541-563.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
O'Malley, R.C., Huang, S.C., Song, L., Lewsey, M.G., Bartlett, A., Nery, J.R., Galli, M., Gallavotti, A., and Ecker, J.R. (2016). Cistrome andEpicistrome Features Shape the Regulatory DNA Landscape. Cell 166, 1598.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Park, S.Y., Fung, P., Nishimura, N., Jensen, D.R., Fujii, H., Zhao, Y., Lumba, S., Santiago, J., Rodrigues, A., Chow, T.F.F., Alfred, S.E.,Bonetta, D., Finkelstein, R., Provart, N.J., Desveaux, D., Rodriguez, P.L., McCourt, P., Zhu, J.K., Schroeder, J.I., Volkman, B.F., andCutler, S.R. (2009). Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. Science 324, 1068-1071.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Peirats-Llobet, M., Han, S.K., Gonzalez-Guzman, M., Jeong, C.W., Rodriguez, L., Belda-Palazon, B., Wagner, D., and Rodriguez, P.L.(2016). A Direct Link between Abscisic Acid Sensing and the Chromatin-Remodeling ATPase BRAHMA via Core ABA Signaling PathwayComponents. Molecular plant 9, 136-147.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Promchuea, S., Zhu, Y.J., Chen, Z.Z., Zhang, J., and Gong, Z.Z. (2017). ARF2 coordinates with PLETHORAs and PINs to orchestrate ABA-mediated root meristem activity in Arabidopsis. Journal of integrative plant biology 59, 30-43.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Qi, J.S., Song, C.P., Wang, B.S., Zhou, J.M., Kangasjarvi, J., Zhu, J.K., and Gong, Z.Z. (2018). Reactive oxygen species signaling andstomatal movement in plant responses to drought stress and pathogen attack. Journal of integrative plant biology 60, 805-826.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ren, X., Chen, Z., Liu, Y., Zhang, H., Zhang, M., Liu, Q., Hong, X., Zhu, J.K., and Gong, Z. (2010). ABO3, a WRKY transcription factor,mediates plant responses to abscisic acid and drought tolerance in Arabidopsis. The Plant journal : for cell and molecular biology 63,417-429.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rushton, D.L., Tripathi, P., Rabara, R.C., Lin, J., Ringler, P., Boken, A.K., Langum, T.J., Smidt, L., Boomsma, D.D., Emme, N.J., Chen, X.,Finer, J.J., Shen, Q.X.J., and Rushton, P.J. (2012). WRKY transcription factors: key components in abscisic acid signalling. Plantbiotechnology journal 10, 2-11.

biotechnology journal 10, 2-11.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Saeed, A.I., Sharov, V., White, J., Li, J., Liang, W., Bhagabati, N., Braisted, J., Klapa, M., Currier, T., Thiagarajan, M., Sturn, A., Snuffin,M., Rezantsev, A., Popov, D., Ryltsov, A., Kostukovich, E., Borisovsky, I., Liu, Z., Vinsavich, A., Trush, V., and Quackenbush, J. (2003).TM4: A free, open-source system for microarray data management and analysis. Biotechniques 34, 374-+.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Saez, A., Rodrigues, A., Santiago, J., Rubio, S., and Rodriguez, P.L. (2008). HAB1-SWI3B Interaction Reveals a Link between AbscisicAcid Signaling and Putative SWI/SNF Chromatin-Remodeling Complexes in Arabidopsis. The Plant cell 20, 2972-2988.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sanchez-Pulido, L., Devos, D., Sung, Z.R., and Calonje, M. (2008). RAWUL: a new ubiquitin-like domain in PRC1 ring finger proteins thatunveils putative plant and worm PRC1 orthologs. BMC genomics 9, 308.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shao, Z., Raible, F., Mollaaghababa, R., Guyon, J.R., Wu, C.T., Bender, W., and Kingston, R.E. (1999). Stabilization of chromatinstructure by PRC1, a Polycomb complex. Cell 98, 37-46.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shen, L.S., Thong, Z.H., Gong, X.M., Shen, Q., Gan, Y.B., and Yu, H. (2014). The putative PRC1 RING-finger protein AtRING1A regulatesflowering through repressing MADS AFFECTING FLOWERING genes in Arabidopsis. Development 141, 1303-1312.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shkolnik-Inbar, D., and Bar-Zvi, D. (2010). ABI4 mediates abscisic acid and cytokinin inhibition of lateral root formation by reducingpolar auxin transport in Arabidopsis. The Plant cell 22, 3560-3573.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shu, K., Zhang, H., Wang, S., Chen, M., Wu, Y., Tang, S., Liu, C., Feng, Y., Cao, X., and Xie, Q. (2013). ABI4 regulates primary seeddormancy by regulating the biogenesis of abscisic acid and gibberellins in arabidopsis. PLoS genetics 9, e1003577.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shu, K., Chen, Q., Wu, Y., Liu, R., Zhang, H., Wang, P., Li, Y., Wang, S., Tang, S., Liu, C., Yang, W., Cao, X., Serino, G., and Xie, Q. (2016).ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. The Plant journal : for cell andmolecular biology 85, 348-361.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Singh, D., and Laxmi, A. (2015). Transcriptional regulation of drought response: a tortuous network of transcriptional factors. Frontiersin plant science 6, 895.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Soderman, E.M., Brocard, I.M., Lynch, T.J., and Finkelstein, R.R. (2000). Regulation and function of the Arabidopsis ABA-insensitive4gene in seed and abscisic acid response signaling networks. Plant physiology 124, 1752-1765.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sun, Q., Csorba, T., Skourti-Stathaki, K., Proudfoot, N.J., and Dean, C. (2013). R-loop stabilization represses antisense transcription atthe Arabidopsis FLC locus. Science 340, 619-621.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Trapnell, C., Hendrickson, D.G., Sauvageau, M., Goff, L., Rinn, J.L., and Pachter, L. (2013). Differential analysis of gene regulation attranscript resolution with RNA-seq. Nature biotechnology 31, 46-53.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Turck, F., Roudier, F., Farrona, S., Martin-Magniette, M.-L., Guillaume, E., Buisine, N., Gagnot, S., Martienssen, R.A., Coupland, G., andColot, V. (2007). Arabidopsis TFL2/LHP1 Specifically Associates with Genes Marked by Trimethylation of Histone H3 Lysine 27. PLoSgenetics 3, e86.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Uno, Y., Furihata, T., Abe, H., Yoshida, R., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2000). Arabidopsis basic leucine zippertranscription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions.Proceedings of the National Academy of Sciences of the United States of America 97, 11632-11637.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, K., He, J., Zhao, Y., Wu, T., Zhou, X., Ding, Y., Kong, L., Wang, X., Wang, Y., Li, J., Song, C.P., Wang, B., Yang, S., Zhu, J.K., andGong, Z. (2018). EAR1 Negatively Regulates ABA Signaling by Enhancing 2C Protein Phosphatase Activity. The Plant cell 30, 815-834.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, L., Hua, D.P., He, J.N., Duan, Y., Chen, Z.Z., Hong, X.H., and Gong, Z.Z. (2011). Auxin Response Factor2 (ARF2) and Its RegulatedHomeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS genetics7, e1002172.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, L., Zhou, C.M., Mai, Y.X., Li, L.Z., Gao, J., Shang, G.D., Lian, H., Han, L., Zhang, T.Q., Tang, H.B., Ren, H., Wang, F.X., Wu, L.Y., Liu,X.L., Wang, C.S., Chen, E.W., Zhang, X.N., Liu, C., and Wang, J.W. (2019). A spatiotemporally regulated transcriptional complex underliesheteroblastic development of leaf hairs in Arabidopsis thaliana. The EMBO journal 38, e100063.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, Q., and Shen, W.H. (2018). Chromatin modulation and gene regulation in plants: insight about PRC1 function. Biochemicalsociety transactions 46, 957-966.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, Z.P., Xing, H.L., Dong, L., Zhang, H.Y., Han, C.Y., Wang, X.C., and Chen, Q.J. (2015). Egg cell-specific promoter-controlledCRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genomebiology 16, 144.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wind, J.J., Peviani, A., Snel, B., Hanson, J., and Smeekens, S.C. (2013). ABI4: versatile activator and repressor. Trends in plant science18, 125-132.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xiao, J., Jin, R., Yu, X., Shen, M., Wagner, J.D., Pai, A., Song, C., Zhuang, M., Klasfeld, S., He, C.S., Santos, A.M., Helliwell, C., Pruneda-Paz, J.L., Kay, S.A., Lin, X.W., Cui, S.J., Garcia, M.F., Clarenz, O., Goodrich, J., Zhang, X.Y., Austin, R.S., Bonasio, R., and Wagner, D.(2017). Cis and trans determinants of epigenetic silencing by Polycomb repressive complex 2 in Arabidopsis. Nature genetics 49, 1546-+.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu, L., and Shen, W.H. (2008). Polycomb silencing of KNOX genes confines shoot stem cell niches in Arabidopsis. Current biology : CB18, 1966-1971.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xu, W., Xu, H., Li, K., Fan, Y., Liu, Y., Yang, X., and Sun, Q. (2017). The R-loop is a common chromatin feature of the Arabidopsisgenome. Nature plants 3, 704-714.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang, L., Zhang, J., He, J.N., Qin, Y.Y., Hua, D.P., Duan, Y., Chen, Z.Z., and Gong, Z.Z. (2014). ABA-Mediated ROS in MitochondriaRegulate Root Meristem Activity by Controlling PLETHORA Expression in Arabidopsis. PLoS genetics 10, e1004791.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang, Z.L., Qian, S.M., Scheid, R.N., Lu, L., Chen, X.S., Liu, R., Du, X.A., Lv, X.C., Boersma, M.D., Scalf, M., Smith, L.M., Denu, J.M., Du,J.M., and Zhong, X.H. (2018). EBS is a bivalent histone reader that regulates floral phase transition in Arabidopsis. Nature genetics 50,1247-+.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yin, H., Zhang, X., Liu, J., Wang, Y., He, J., Yang, T., Hong, X., Yang, Q., and Gong, Z. (2009). Epigenetic regulation, somatic homologousrecombination, and abscisic acid signaling are influenced by DNA polymerase epsilon mutation in Arabidopsis. The Plant cell 21, 386-402.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yuan, L., Liu, X., Luo, M., Yang, S., and Wu, K. (2013). Involvement of histone modifications in plant abiotic stress responses. Journal ofintegrative plant biology 55, 892-901.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, J.X., Xie, S.J., Cheng, J.K., Lai, J.S., Zhu, J.K., and Gong, Z.Z. (2016). The Second Subunit of DNA Polymerase Delta Is Required

Zhang, J.X., Xie, S.J., Cheng, J.K., Lai, J.S., Zhu, J.K., and Gong, Z.Z. (2016). The Second Subunit of DNA Polymerase Delta Is Requiredfor Genomic Stability and Epigenetic Regulation. Plant physiology 171, 1192-1208.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, X., Germann, S., Blus, B.J., Khorasanizadeh, S., Gaudin, V., and Jacobsen, S.E. (2007). The Arabidopsis LHP1 proteincolocalizes with histone H3 Lys27 trimethylation. Nature structural & molecular biology 14, 869-871.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, Y., Liu, T., Meyer, C.A., Eeckhoute, J., Johnson, D.S., Bernstein, B.E., Nussbaum, C., Myers, R.M., Brown, M., Li, W., and Liu, X.S.(2008). Model-based Analysis of ChIP-Seq (MACS). Genome biology 9, R137.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhao, Y., Gao, J.H., Kim, J.I., Chen, K., Bressan, R.A., and Zhu, J.K. (2017). Control of Plant Water Use by ABA Induction of Senescenceand Dormancy: An Overlooked Lesson from Evolution. Plant and cell physiology 58, 1319-1327.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhou, J.X., Liu, Z.W., Li, Y.Q., Li, L., Wang, B.J., Chen, S., and He, X.J. (2018). Arabidopsis PWWP domain proteins mediate H3K27trimethylation on FLC and regulate flowering time. Journal of integrative plant biology 60, 362-368.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhou, Y., Romero-Campero, F.J., Gomez-Zambrano, A., Turck, F., and Calonje, M. (2017). H2A monoubiquitination in Arabidopsis thalianais generally independent of LHP1 and PRC2 activity. Genome biology 18, 69.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhu, J.K. (2016). Abiotic Stress Signaling and Responses in Plants. Cell 167, 313-324.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhu, Y., Luo, X., Liu, X., Wu, W., Cui, X., He, Y., and Huang, J. (2019). Arabidopsis PEAPODs function with LIKE HETEROCHROMATINPROTEIN1 to regulate lateral organ growth. Journal of integrative plant biology.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zuo, J., Niu, Q.W., and Chua, N.H. (2000). Technical advance: An estrogen receptor-based transactivator XVE mediates highly induciblegene expression in transgenic plants. The Plant journal : for cell and molecular biology 24, 265-273.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

DOI 10.1105/tpc.19.00604; originally published online January 9, 2020;Plant Cell
Song, Qianwen Sun, Shuhua Yang and Zhizhong GongZhang, Deping Hua, Li Yang, Li Wang, Zhizhong Chen, Chuanyou Li, Baoshan Wang, Chun-Peng Yujuan Zhu, Xiaoying Hu, Ying Duan, Shaofang Li, Yu Wang, Amin Ur Rehman, Junna He, Jing
Regulates Abscisic Acid SignalingThe Arabidopsis Nodulin Homeobox Factor AtNDX Interacts with AtRING1A/B and Negatively
This information is current as of February 12, 2020
Supplemental Data /content/suppl/2020/01/09/tpc.19.00604.DC1.html
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























