Embed Size (px)
Citation preview

Ž .Brain Research 813 1998 406–410
Short communication
The alternation of optokinetic responses driven by moving stimuli in humans
Min Wei, Fuchuan Sun )
Laboratory of Neurobiology of Shanghai Institute of Physiology, and Laboratory of Visual Information Processing of Biophysics Institute,Chinese Academy of Sciences, 320 Yue-Yang Road, Shanghai 200031, China
Accepted 29 September 1998
Abstract
When the two eyes were exposed dichoptically to two oppositely moving patterns, the alternating OKN was elicited, in which the eyemovements tracked the two moving stimuli alternately. In the present experiment, two oppositely moving stimuli were periodicallyexchanged between the eyes, the results showed that the alternation of OKN direction was not affected by this exchange. It suggests thatalternating OKN was not controlled by rivalry between the two eyes but between two perceived motion patterns. q 1998 Elsevier ScienceB.V. All rights reserved.
Keywords: Eye movement; Optokinetic nystagmus; Alternating OKN; Binocular rivalry; Dichoptic presentation
A moving visual field evokes a series of tracking eyeŽ .movements known as optokinetic nystagmus OKN . When
the two eyes are exposed dichoptically to two patternsmoving in opposite directions, OKN alternates its directionperiodically; that is, eye movements track the two eyes’moving directions back and forth in alternating sequence.This alternating OKN was first reported by Enoksson as an
w xobjective indicator of binocular rivalry 6 , and has beeninvestigated more intensively for binocular rivalry in hu-
w xmans by Fox et al. 8 and in monkeys by Logothetis andw xSchall 14,15 . In their experiments, the direction of slow
phases of OKN was found correspondingly to the percep-tion of motion direction during rivalry. It has been pre-sumed that alternating OKN is the eye movement reflec-tion of dominance and suppression between the two eyes.However, neurophysiological results revealed that the nu-
Ž .cleus of the optic tract NOT and the dorsal terminalŽ .nucleus of the accessory optic tract DTN are essential
nuclei for generation of slow-phase eye movements inw xhorizontal OKN 1–4 . These nuclei receiving the motion
input from either eye are strongly direction-selective foroptokinetic stimuli, i.e., the NOT and DTN on the left sideof the brain prefer leftward movement and the right side
w xrightward 9–11 . These results indicate that the NOT andDTN on each side, although driven binocularly, are clus-
) Corresponding author. Fax: q86-21-64332445; E-mail:[email protected]
tered into motion direction-specific nuclei already. There-fore, it would be a rational assumption that alternatingOKN, tracking two motion directions alternately, might benot driven by rivalry between two eyes, but between twoperceived motion patterns. Logothetis et al. ingeniouslyemployed exchanging a pair of stationary gratings between
w xthe two eyes to investigate binocular rivalry 16 . In thepresent experiment, continuing our previous alternating
w xOKN work 7,20 , we manipulated the exchanging methodto attest the assumption of the mechanism of alternatingOKN.
Six volunteers participated in the experiment. All sub-jects had normal or correct-to-normal vision. Eye move-ments were measured with the magnetic scleral search coil
w xtechnique 5,19 . The annulus of silicone rubber with theŽ .induction coil Skalar Medical was adhered to the subject’s
w xright eye 5 . The head was stabilized by a bite bar. Eyemovement signals were sampled with a frequency 100 Hzand stored on a PC computer for off-line analysis.
Two synchronized moving gratings, generated by aw xcomputerized multi-function moving pattern generator 21 ,
were displayed on two tangential screens respectively asstimuli. The grating was organized with black and white
Ž 2 2strips the luminance was 0.1 cdrm and 4.46 cdrm ,.respectively; each strip was 28 wide , and extended 238
high by 308 wide at the view distance 46 cm. TwoŽoppositely moving gratings velocities ranged from 10 to
.508rs were presented to the subject’s two eyes dichopti-cally through a four-mirror stereoscope. And two stimulat-
Ž .ing modes were employed in the experiment: 1 the
0006-8993r98r$ - see front matter q 1998 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 98 01046-4
( )M. Wei, F. SunrBrain Research 813 1998 406–410 407
non-exchanging mode—two directions of oppositely mov-ing stimuli kept invariable for each eye during the experi-
Ž .ment; 2 the exchanging mode—two directions of oppo-sitely moving stimuli exchanged periodically between the
Žsubject’s two eyes illustrated in Fig. 2A,B,D,E, the upper.traces . The exchanging cycle was 400 ms, or 2 s, 10 s
Ž .i.e., the stimuli were exchanged every 200 ms, or 1 s, 5 s .If the alternating OKN was determined by the competitionof moving stimuli no matter which eye conveyed thesestimuli, the exchange of the stimuli should not influencethe alternation of OKN.
Ž .The stimuli were viewed monocularly patched one eyeor dichoptically. At the beginning of each trial, the subjectwas instructed to fixate four points sequentially for calibra-tion. Each trial lasted 30 s. We defined the ‘directionalduration’ for describing the time duration while the slowphases of OKN tracking the same direction in each alter-nating cycle.
In the non-exchanging mode, when the subject’s twoeyes were exposed dichoptically to two oppositely movingpatterns, all subjects exhibited the ‘alternating OKN’ eyemovement behavior, which tracked the two moving direc-
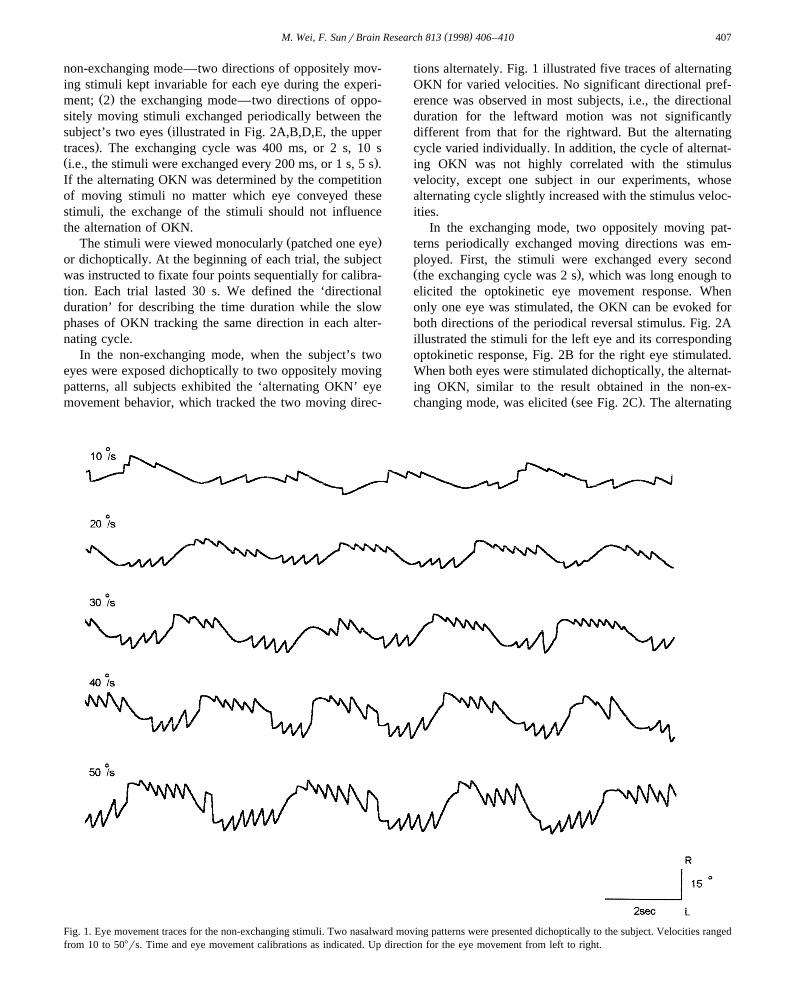
tions alternately. Fig. 1 illustrated five traces of alternatingOKN for varied velocities. No significant directional pref-erence was observed in most subjects, i.e., the directionalduration for the leftward motion was not significantlydifferent from that for the rightward. But the alternatingcycle varied individually. In addition, the cycle of alternat-ing OKN was not highly correlated with the stimulusvelocity, except one subject in our experiments, whosealternating cycle slightly increased with the stimulus veloc-ities.
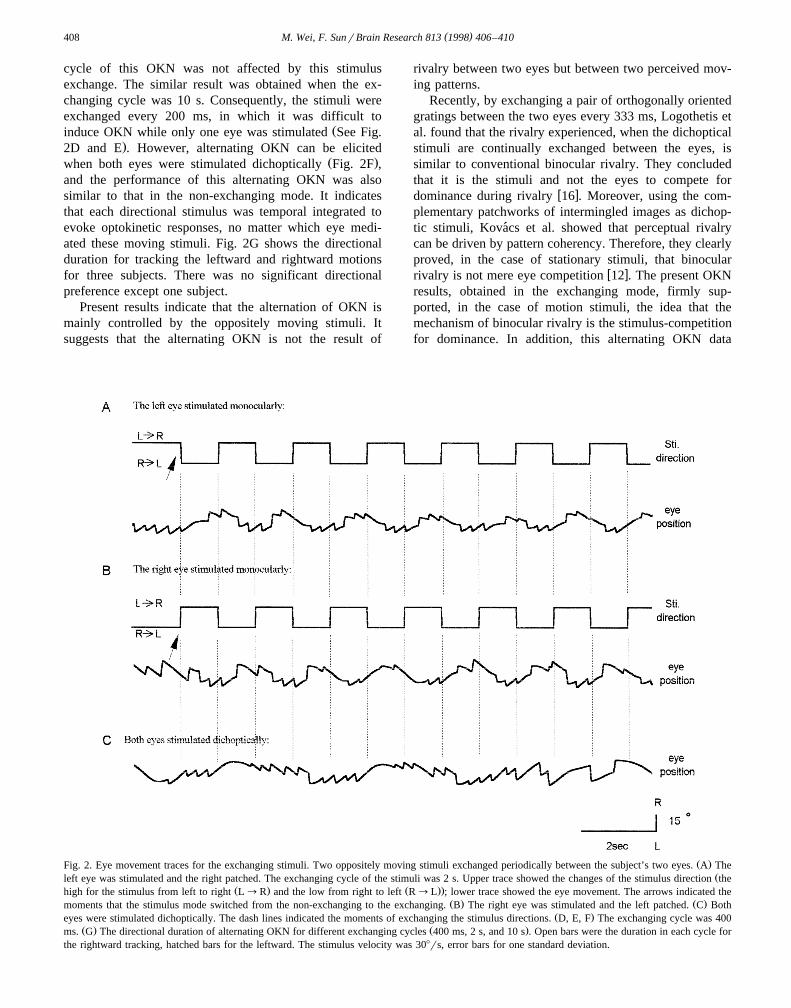
In the exchanging mode, two oppositely moving pat-terns periodically exchanged moving directions was em-ployed. First, the stimuli were exchanged every secondŽ .the exchanging cycle was 2 s , which was long enough toelicited the optokinetic eye movement response. Whenonly one eye was stimulated, the OKN can be evoked forboth directions of the periodical reversal stimulus. Fig. 2Aillustrated the stimuli for the left eye and its correspondingoptokinetic response, Fig. 2B for the right eye stimulated.When both eyes were stimulated dichoptically, the alternat-ing OKN, similar to the result obtained in the non-ex-
Ž .changing mode, was elicited see Fig. 2C . The alternating
Fig. 1. Eye movement traces for the non-exchanging stimuli. Two nasalward moving patterns were presented dichoptically to the subject. Velocities rangedfrom 10 to 508rs. Time and eye movement calibrations as indicated. Up direction for the eye movement from left to right.
( )M. Wei, F. SunrBrain Research 813 1998 406–410408
cycle of this OKN was not affected by this stimulusexchange. The similar result was obtained when the ex-changing cycle was 10 s. Consequently, the stimuli wereexchanged every 200 ms, in which it was difficult to
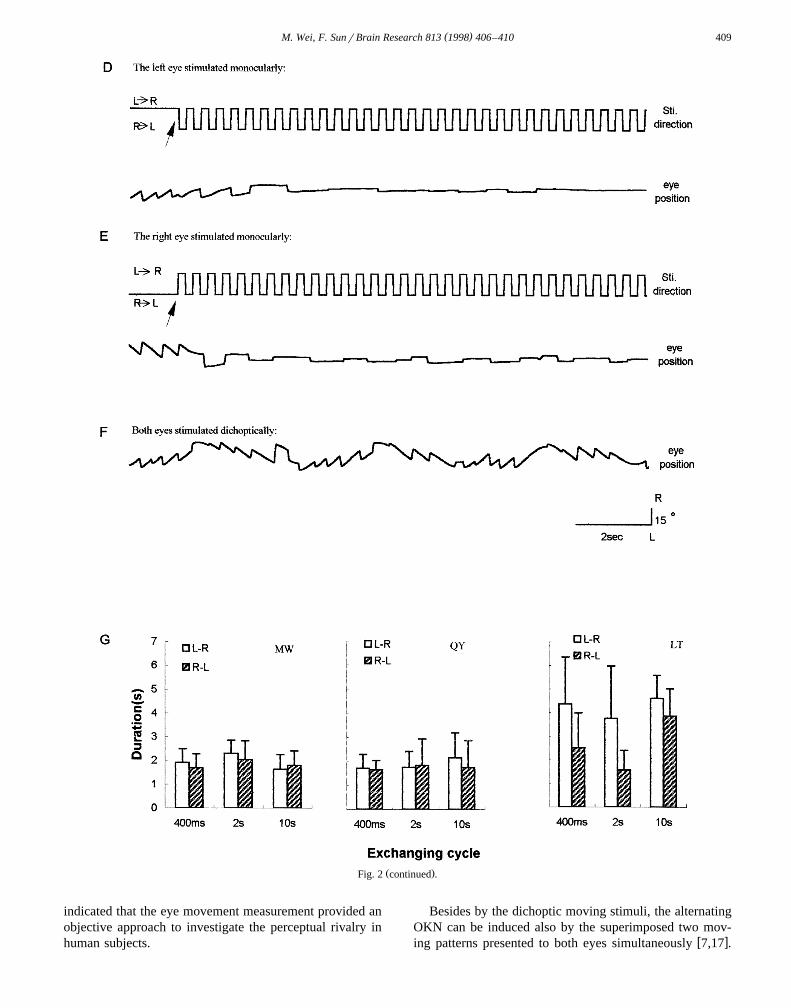
Žinduce OKN while only one eye was stimulated See Fig..2D and E . However, alternating OKN can be elicited
Ž .when both eyes were stimulated dichoptically Fig. 2F ,and the performance of this alternating OKN was alsosimilar to that in the non-exchanging mode. It indicatesthat each directional stimulus was temporal integrated toevoke optokinetic responses, no matter which eye medi-ated these moving stimuli. Fig. 2G shows the directionalduration for tracking the leftward and rightward motionsfor three subjects. There was no significant directionalpreference except one subject.
Present results indicate that the alternation of OKN ismainly controlled by the oppositely moving stimuli. Itsuggests that the alternating OKN is not the result of
rivalry between two eyes but between two perceived mov-ing patterns.
Recently, by exchanging a pair of orthogonally orientedgratings between the two eyes every 333 ms, Logothetis etal. found that the rivalry experienced, when the dichopticalstimuli are continually exchanged between the eyes, issimilar to conventional binocular rivalry. They concludedthat it is the stimuli and not the eyes to compete for
w xdominance during rivalry 16 . Moreover, using the com-plementary patchworks of intermingled images as dichop-tic stimuli, Kovacs et al. showed that perceptual rivalry´can be driven by pattern coherency. Therefore, they clearlyproved, in the case of stationary stimuli, that binocular
w xrivalry is not mere eye competition 12 . The present OKNresults, obtained in the exchanging mode, firmly sup-ported, in the case of motion stimuli, the idea that themechanism of binocular rivalry is the stimulus-competitionfor dominance. In addition, this alternating OKN data
Ž .Fig. 2. Eye movement traces for the exchanging stimuli. Two oppositely moving stimuli exchanged periodically between the subject’s two eyes. A TheŽleft eye was stimulated and the right patched. The exchanging cycle of the stimuli was 2 s. Upper trace showed the changes of the stimulus direction the
Ž . Ž ..high for the stimulus from left to right L™R and the low from right to left R™L ; lower trace showed the eye movement. The arrows indicated theŽ . Ž .moments that the stimulus mode switched from the non-exchanging to the exchanging. B The right eye was stimulated and the left patched. C Both
Ž .eyes were stimulated dichoptically. The dash lines indicated the moments of exchanging the stimulus directions. D, E, F The exchanging cycle was 400Ž . Ž .ms. G The directional duration of alternating OKN for different exchanging cycles 400 ms, 2 s, and 10 s . Open bars were the duration in each cycle for
the rightward tracking, hatched bars for the leftward. The stimulus velocity was 308rs, error bars for one standard deviation.
( )M. Wei, F. SunrBrain Research 813 1998 406–410 409
Ž .Fig. 2 continued .
indicated that the eye movement measurement provided anobjective approach to investigate the perceptual rivalry inhuman subjects.
Besides by the dichoptic moving stimuli, the alternatingOKN can be induced also by the superimposed two mov-
w xing patterns presented to both eyes simultaneously 7,17 .

( )M. Wei, F. SunrBrain Research 813 1998 406–410410
Niemann et al. studied the behaviors of smooth-pursuit andalternating OKN more intensively with both the dichoptic
w xand the superimposed stimulation 18 . They found thedirectional preference for some subjects for both stimulusconditions. The similar result was also observed in a fewsubjects for our dichoptic experiment. Particularly, one
Ž .subject LT exhibited a rightward directional preferencefor the exchanging stimuli, and this result also explicitlyindicates that in the alternating OKN there is the direc-tional dominance, not the eye dominance.
Neurophysiological results, besides the NOT and DTNin the subcortical level, showed that some neurons in themonkey cortical area MT exhibited activity changes duringmotion rivalry that reflect the perceived direction of mo-
w x Žtion 14 . And neurons in other visual cortical areas V1,.V2, and V4 can respond to the perceived orientation of
w xrival gratings 13 . Many of these directional-preference ororientation-selective cells could be driven equally well
w xfrom either eye 13,14 . From this, it was suggested thatthe rivalry was between alternative stimuli representationsthat were encoded in the activity of neurons in different
w xvisual areas 13 . These findings provided a possibleneuro-physiological basis for our results: the alternatingOKN is controlled by rivalry between perceived motionsno matter which eye conveyed these stimuli. Further sys-tematic experiment is needed to verify this point.
Acknowledgements
This work was partially supported by the grants fromthe National Natural Science Foundation of China. Theauthors also gratefully acknowledge Dr. Lieyi Fang for thehelp in the equipment, and Qing Yang, Jing Tian, JianLiang Tong for the help in experiments.
References
w x1 J.A. Buttner-Ennever, B. Cohen, A.K.E. Horn, H. Reisine, Efferent¨pathways of the nucleus of the optic tract in monkey and their role
Ž .in eye movement, J. Comp. Neurol. 373 1996 90–107.w x2 B. Cohen, H. Reisine, J.I. Yokota, T. Raphan, The nucleus of the
Ž .optic tract, Ann. New York Acad. Sci. 656 1992 277–296.
w x3 H. Collewijn, Oculomotor areas in the rabbit’s midbrain and pretec-Ž .tum, J. Neurobiol. 6 1975 3–22.
w x4 H. Collewijn, Direction-selective units in the rabbit’s nucleus of theŽ .optic tract, Brain Res. 100 1975 489–508.
w x5 H. Collewijn, F. van der Mark, T.C. Jansen, Precise recording ofŽ .human eye movements, Vision Res. 15 1975 447–450.
w x6 P. Enoksson, Binocular rivalry and monocular dominance studiedŽ .with optokinetic nystagmus, Acta Ophthalmologica 41 1963 544–
563.w x7 L.-Y. Fang, F. Sun, Alternating control of optokinetic nystagmus
eye-movements by two different isoluminant moving chromaticŽ . Ž .patterns, Acta Biophysica Sinica 10 3 1994 446–452.
w x8 R. Fox, S. Todd, L.A. Bettinger, Optokinetic nystagmus as anŽ .objective indicator of binocular rivalry, Vision Res. 15 1975
849–853.w x9 A.F. Fuchs, M.J. Mustari, F.R. Robinson, C.R.S. Kaneko, Visual
signals in the nucleus of the optic tract and their brain stemŽ .destinations, Ann. New York Acad. Sci. 656 1992 266–276.
w x10 K.-P. Hoffmann, Cortical versus subcortical contributions to theoptokinetic reflex in the cat, in: G. Lennerstrand, D.S. Zee, E.L.
Ž .Keller Eds. , Function Basic of Ocular Motility Disorders, Perga-mon, Oxford, 1982, pp. 303–310.
w x11 K.-P. Hoffmann, C. Distler, The role of direction selective cells inthe nucleus of the optic tract of cat and monkey during optokinetic
Ž .nystagmus, in: E.L. Keller, D.S. Zee Eds. , Adaptive Processes inVisual and Oculomotor System, Vol. 57, Pergamon, Oxford, 1986,pp. 261–266.
´w x12 I. Kovacs, T.V. Papathomas, M. Yang, A. Feher, When the brain´ ´changes its mind, interocular grouping during binocular rivalry,
Ž .Proc. Natl. Acad. Sci. U.S.A. 93 1996 15508–15511.w x13 D.A. Leopold, N.K. Logothetis, Activity changes in early visual
cortex reflect monkeys’ percepts during binocular rivalry, NatureŽ .379 1996 549–553.
w x14 N.K. Logothetis, J.D. Schall, Neuronal correlates of subjectiveŽ .visual perception, Science 245 1989 761–763.
w x15 N.K. Logothetis, J.D. Schall, Binocular motion rivalry in macaquemonkeys: eye dominance and tracking eye movements, Vision Res.
Ž .30 1990 1409–1419.w x16 N.K. Logothetis, D.A. Leopold, D.L. Sheinberg, What is rivalling
Ž .during binocular rivalry, Nature 380 1996 621–624.w x17 E.G. Merrill, L. Stark, Optokinetic nystagmus: double stripe experi-
Ž .ment, Quarterly Progress Report, Res. Lab. Electr. M.I.T. 70 1963357–359.
w x18 T. Niemann, U.J. Ilg, K.-P. Hoffmann, Eye movements elicited byŽ .transparent stimuli, Exp. Brain Res. 98 1994 314–322.
w x19 D.A. Robinson, A method of measuring eye movement using ascleral search coil in a magnetic field, IEEE Trans. Biomed. Elec-
Ž .tron. Biomed. Eng. 10 1963 137–145.w x20 M. Wei, F. Sun, The alternating optokinetic nystagmus during
simultaneous stimulation with the different moving patterns pre-Ž . Ž .sented to each eye respectively, Acta Biophysica Sinica 8 3 1992
434–442.w x21 H.M. Zhu, F. Sun, Computerized multi-function moving pattern
Ž .generator, J. Biomed. Eng. 9 1992 167–173.





























