Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 251, No. 16, Issue of August 25. pp. 5035-5042, 1976
Printed in U.S.A.
The Allosteric Regulation of Hexokinase C from Amphibian Liver *
(Received for publication, March 1, 1976)
TITO URETA
From the Departamento de Biologia, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile
A type C hexokinase (ATP:n-hexose-6-phosphotransferase, EC 2.7.1.1) was partially purified from the liver of the frog Calyptocephalella caudiuerbera. The enzyme is inhibited by glucose levels in the range of normal blood sugar concentrations. The extent of the inhibition by glucose depends on the concentration of ATP, being most marked between 1 and 5 mM ATP. Fructose, although a substrate, was not inhibitory of its own phosphorylation.
The inhibitory effect of high glucose levels exhibited a strong, reversible pH dependence being most marked at pH 6.5. At pH 7.5 the inhibition by high glucose levels was a function of the enzyme concentration, the effect being stronger at high enzyme concentrations, whereas no inhibition was observed when assaying very diluted preparations. At all enzyme concentrations studied, high levels of glucose caused no inhibition at pH 8.5, whereas at pH 6.5 strong inhibition was always observed.
Short times of photooxidation of hexokinase C as well as incubation with low concentrations of p-chloromercuribenzoate resulted in the loss of the inhibition by excess of glucose.
Glucose 6-phosphate was found to be a strong inhibitor of hexokinase C but only at high glucose levels. The inhibitory effect of glucose-6-P follows sigmoidal kinetics at low (about 0.02 mM) glucose concentrations, the Hill coefficient being 2.3. The kinetics of the inhibition became hyperbolic at high (>0.2 mM) glucose levels.
These results suggest that the inhibition of hexokinase C by excess glucose is due to the interaction of glucose with a second, aldose-specific, regulatory site on the enzyme. The modification of the inhibitory effect by ATP, glucose-6-P, enzyme concentration, and pH, all of them at physiological levels, indicates a major role for hexokinase C in the regulation of glucose utilization by the liver.
The ATP-dependent phosphorylation of glucose in verte- brates is catalyzed by four isozymes named hexokinases A, B, C, and D, (ATP:n-hexose-6-phosphotransferases, EC 2.7.1.1).’ Their kinetic and physicochemical properties, tissue distribu- tion, phylogenetic, developmental, and adaptive behavior have been studied with some detail (for reviews see Refs. 1 to 6).
One of the four glucose phosphotransferases, hexokinase C, ’ was found to be inhibited by its own substrate (7-9). Since the inhibition occurs in the normal range of blood sugar concentra-
*This work was supported by Grant 70A from the Servicio de Desarrollo Cientffico y Creacibn Artistica, Universidad de Chile, and by the Multinational Project on Biochemistry, Chile, of the Organiza- tion of American States.
’ A logical nomenclature for the hexokinase isozymic system awaits further knowledge of the structural basis of the isozymes. We have dealt with this problem elsewhere (1) proposing the provisional names hexokinase C for the isozyme inhibited by excess glucose, and hexokinase D (the so-called glucokinase) for the isozyme with a high K, (-6 mM) for glucose, irrespective of their chromatographic or electrophoretic mobilities, or the number of isozymes actually found in a given tissue or organism. Hexokinase A and hexokinase B are those isozymes eluting in the first and second position of the DEAE cellulose chromatograms, respectively, provided they are not inhibited by excess glucose or do not have a high K, for glucose.
tions this property may be considered as a regulatory mecha- nism probably involved in the channeling of glucose to the several pathways starting at the glucose-6-P crossroad. The “antiparallel” behavior of hexokinase D, i.e. the fact that its velocity increases precisely at the same sugar concentrations (K, for glucose = 6 mM), supports the notion that the inhibition by glucose of hexokinase C is relevant to the control of sugar utilization.
Further work on the regulation of hexokinase C activity has not progressed because it is present in very low amounts in rodent liver. Recently however, a major hexokinase presenting the same inhibition feature of rat hexokinase C has been found in the liver of some anurans.’ This work describes the partial purification of this isozyme and the study of the substrate inhibition. The data obtained suggest the existence of two catalytically different forms of hexokinase C (inhibited and noninhibited forms), the interconversion of which can be brought about by several physiological factors, indicating that this isozyme is a major regulatory entity in the hepatic utilization of glucose.
‘T. Ureta, J. C. Slebe, R. Radojkovit, C. Lozano, and N. Diaz, manuscript in preparation.
5035
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from
5036 Allosteric Regulation of Hexokinase C from Amphibian Liver
MATERIALS AND METHODS
Animals-An endemic Chilean leptodactylid frog, Calyptocephalel- 24 c
la caudiuerbera ( = Caudiuerbera caudiuerbera (lo)), was used throughout this study. The frogs were caught in the vicinities of Santiago and maintained in aquaria with cold tap water.
Enzyme Preparation-The frogs were killed by decapitation without anesthesia, thoroughly bled, and the liver excised and weighed. All further operations were performed at O-4” unless otherwise stated. The pooled livers were finely minced with scissors and squeezed through a Harvard tissue press. The 50% homogenates (w/v) were prepared using a Potter-Elvehjem apparatus in a medium containing 0.01 MTris’-HCl (pH 7),0.001 M EDTA (Tris buffer). The homogenates were centrifuged at 105,000 x g for 60 min in a Spinco ultracentrifuge. The red supernatant liquid was then chromatographed in a DEAE-cellulose column equilibrated with the Tris buffer and the glucose-phos- phorylating isozymes eluted with a linear gradient from 0 to 0.5 M KC1 in the same buffer (8). The procedure can be performed with as little as 506 mg of liver or scaled up to 100 g of material. The active fractions obtained from the columns were concentrated by the addition of moist DEAE-cellulose to the dialyzed enzyme solution until all the activity was retained by the resin. After addition of a small amount of 0.5 M KC1 in Tris buffer the eluted activity was recovered by filtration.
6. j : D
16
7 , vi
C
5
E 8
0
0. 20 40 60 80
Fractan rumba
Enzyme Assay-Hexokinase activity was measured by either of the following two procedures: Method a is measurement of glucose-6-P formation by coupling the phosphotransferase reaction with NADP and glucose-6-P dehydrogenase (11). NADPH formation at 30” was followed spectrophotometrically at 340 nm in a medium containing (final concentrations): 10 mM KCl, 100 mM Tris-HCl buffer (pH 7.5), 0.5 mM NADP, 0.2 i.u. of glucose-6-P dehydrogenase, substrates, and enzyme in a final volume of 0.5 ml. When fructose was used as a substrate, measurement of glucose-6-P was accomplished by the addition of 1 i.u. of phosphoglucose isomerase. Blanks with ATP or glucose omitted were routinely run. Method b is that measurement of ADP formation made by coupling the phosphotransferase reaction to pyruvate kinase and lactate dehydrogenase (8). NADH oxidation at 30” was followed spectrophotometrically at 340 nm in a medium containing (final concentrations): 10 mM KCl, 100 rnM Tris-HCl buffer (pH 7.5), 0.5 mM NADH, 1 i.u. of lactate dehydrogenase (containing pyruvate kinase), 2.5 mM phosphoenolpyruvate, substrates, and enzyme in a final volume of 0.5 ml. A similar system with sugar omitted was used as a blank. Modifications of these standard assay mixtures are indicated on the figures. The concentration of MgZ+ was adjusted in each case according to the ATP concentration, so that a level of free divalent ion of 1 mM was always present. The metal ion was added as the chloride salt. A Gilford spectrophotometer model 2400 with thermospacers was used. Full scale sensitivities of 0.25 to 2 optical density units and chart speeds from 1 to 4 inches per min were used. One unit of hexokinase activity is that amount of enzyme that would catalyze the phosphorylation of 1 rmol of glucose/min at 30”. Specific activity is defined as units/mg of protein. Protein was measured by the method of Miller (12) using bovine serum albumin as standard.
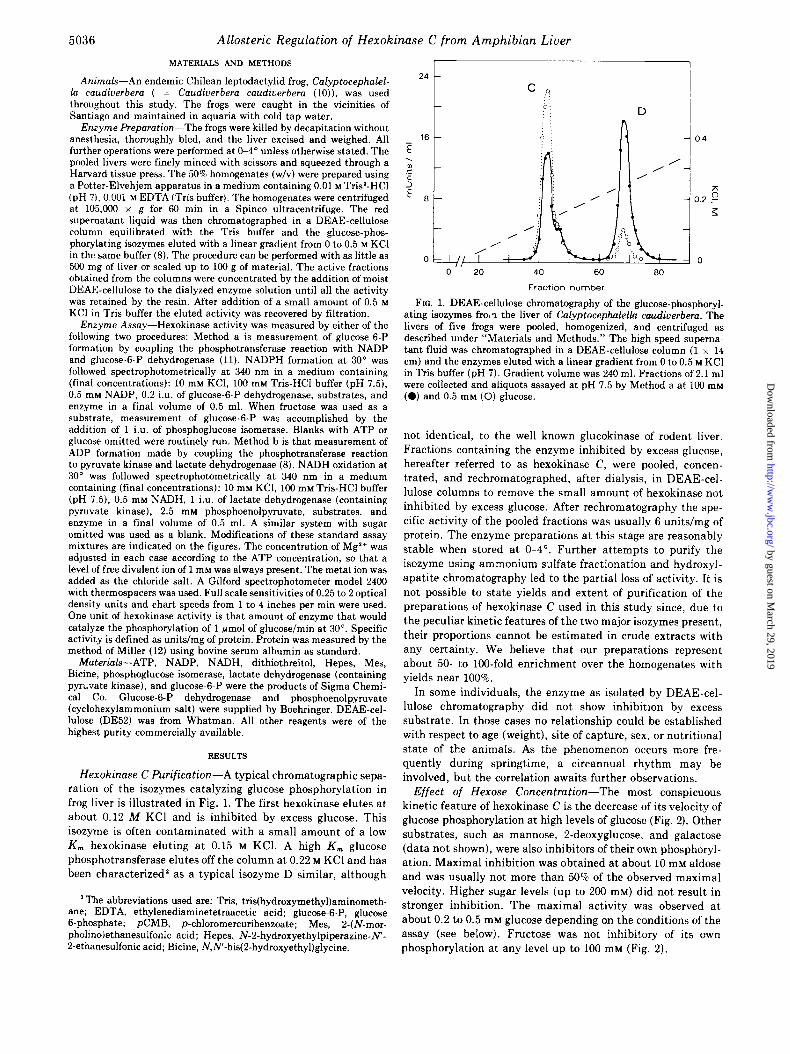
FIG. 1. DEAE-cellulose chromatography of the glucose-phosphoryl- ating isoxymes fro,? the liver of Calyptocephalella caudiuerbem. The livers of five frogs were pooled, homogenized, and centrifuged as described under “Materials and Methods.” The high speed superna- tant fluid was chromatographed in a DEAE-cellulose column (1 x 14 cm) and the enzymes eluted with a linear gradient from 0 to 0.5 M KC1 in Tris buffer (pH 7). Gradient volume was 240 ml. Fractions of 2.1 ml were collected and aliquots assayed at pH 7.5 by Method a at 100 mM (0) and 0.5 mM (0) glucose.
Materials-ATP, NADP, NADH, dithiothreitol, Hepes, Mes, Bicine, phosphoglucose isomerase, lactate dehydrogenase (containing pyruvate kinase), and glucose-6-P were the products of Sigma Chemi- cal Co. Glucose-6-P dehydrogenase and phosphoenolpyruvate (cyclohexylammonium salt) were supplied by Boehringer. DEAE-cel- lulose (DE52) was from Whatman. All other reagents were of the highest purity commercially available.
not identical, to the well known glucokinase of rodent liver.
Fractions containing the enzyme inhibited by excess glucose,
hereafter referred to as hexokinase C, were pooled, concen-
trated, and rechromatographed, after dialysis, in DEAE-cel-
lulose columns to remove the small amount of hexokinase not
inhibited by excess glucose. After rechromatography the spe-
cific activity of the pooled fractions was usually 6 units/mg of
protein. The enzyme preparations at this stage are reasonably
stable when stored at O-4”. Further attempts to purify the
isozyme using ammonium sulfate fractionation and hydroxyI-
apatite chromatography led to the partial loss of activity. It is
not possible to state yields and extent of purification of the
preparations of hexokinase C used in this study since, due to
the peculiar kinetic features of the two major isozymes present,
their proportions cannot be estimated in crude extracts with
any certainty. We believe that our preparations represent
about 50- to 100-fold enrichment over the homogenates with
yields near 100%.
In some individuals, the enzyme as isolated by DEAE-cel-
lulose chromatography did not show inhibition by excess
substrate. In those cases no relationship could be established
with respect to age (weight), site of capture, sex, or nutritional
RESULTS
Hexokinase C Purification-A typical chromatographic sepa-
ration of the isozymes catalyzing glucose phosphorylation in
frog liver is illustrated in Fig. 1. The first hexokinase elutes at
about 0.12 M KC1 and is inhibited by excess glucose. This
isozyme is often contaminated with a small amount of a low
K, hexokinase eluting at 0.15 M KCl. A high K, glucose
phosphotransferase elutes off the column at 0.22 M KC1 and has
been characterized’ as a typical isozyme D similar, although
3 The abbreviations used are: Tris, tris(hydroxymethyl)aminometh- ane; EDTA, ethylenediaminetetraacetic acid; glucose-6-P, glucose B-phosphate; pCMB, p-chloromercuribenzoate; Mes, 2-(N-mor- pholinolethanesulfonic acid; Hepes, N-2-hydroxyethylpiperazine-N’- 2-ethanesulfonic acid; Bicine, NJ’-bis(P-hydroxyethyl)glycine.
state of the animals. As the phenomenon occurs more fre- quently during springtime, a circannual rhythm may be
involved, but the correlation awaits further observations. Effect of Hexose Concentration-The most conspicuous
kinetic feature of hexokinase C is the decrease of its velocity of glucose phosphorylation at high levels of glucose (Fig. 2). Other substrates, such as mannose, 2-deoxyglucose, and galactose (data not shown), were also inhibitors of their own phosphoryl- ation. Maximal inhibition was obtained at about 10 mM aldose and was usually not more than 50% of the observed maximal velocity. Higher sugar levels (up to 200 mM) did not result in stronger inhibition. The maximal activity was observed at
about 0.2 to 0.5 mM glucose depending on the conditions of the assay (see below). Fructose was not inhibitory of its own
phosphorylation at any level up to 100 mM (Fig. 2).
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Allosteric Regulation of Hexokinase C from Amphibian Liver 5037
15
t
,’ /
/
0 0.001 0.01 0.1 1 10 100
Hexose, mM
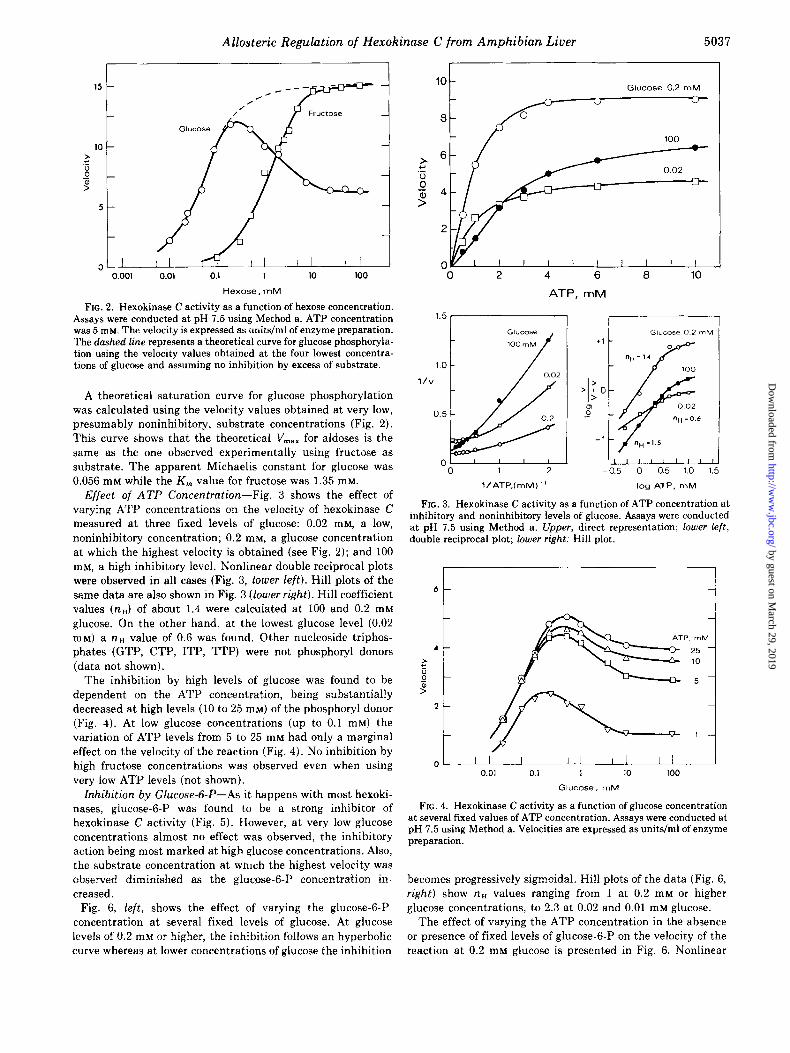
FIG. 2. Hexokinase C activity as a function of hexose concentration. Assays were conducted at pH 7.5 using Method a. ATP concentration was 5 rn~. The velocity is expressed as units/ml of enzyme preparation. The dashed line represents a theoretical curve for glucose phosphoryla- tion using the velocity values obtained at the four lowest concentra- tions of glucose and assuming no inhibition by excess of substrate.
A theoretical saturation curve for glucose phosphorylation
was calculated using the velocity values obtained at very low, presumably noninhibitory, substrate concentrations (Fig. 2).
This curve shows that the theoretical V,,, for aldoses is the same as the one observed experimentally using fructose as substrate. The apparent Michaelis constant for glucose was 0.056 mM while the K, value for fructose was 1.35 mM.
Effect of ATP Concentration-Fig. 3 shows the effect of varying ATP concentrations on the velocity of hexokinase C measured at three fixed levels of glucose: 0.02 mM, a low, noninhibitory concentration; 0.2 mM, a glucose concentration at which the highest velocity is obtained (see Fig. 2); and 100 mM, a high inhibitory level. Nonlinear double reciprocal plots were observed in all cases (Fig. 3, lower left). Hill plots of the same data are also shown in Fig. 3 (lower right). Hill coefficient values (n,) of about 1.4 were calculated at 100 and 0.2 mM glucose. On the other hand, at the lowest glucose level (0.02 mM) a na value of 0.6 was found. Other nucleoside triphos- phates (GTP, CTP, ITP, TTP) were not phosphoryl donors (data not shown).
The inhibition by high levels of glucose was found to be dependent on the ATP concentration, being substantially decreased at high levels (10 to 25 mM) of the phosphoryl donor (Fig. 4). At low glucose concentrations (up to 0.1 mM) the variation of ATP levels from 5 to 25 mM had only a marginal effect on the velocity of the reaction (Fig. 4). No inhibition by high fructose concentrations was observed even when using very low ATP levels (not shown).
Inhibition by Glucose-&P-As it happens with most hexoki- nases, glucose-6-P was found to be a strong inhibitor of hexokinase C activity (Fig. 5). However, at very low glucose concentrations almost no effect was observed, the inhibitory action being most marked at high qlucose concentrations. Also, the substrate concentration at which the highest velocity was observed diminished as the glucose-6-P concentration in- creased.
Fig. 6, left, shows the effect of varying the glucose-6-P concentration at several fixed levels of glucose. At glucose levels of 0.2 mM or higher, the inhibition follows an hyperbolic
Glucose 0.2 mM
1.0
l/V
0.5
6 a 10
ATP, mM
0 0 1 2 -0.5 0 0.5 1.0 1.5
l/ATP,(mM)- log ATP. mM
FIG. 3. Hexokinase C activity as a function of ATP concentration at inhibitory and noninhibitory levels of glucose. Assays were conducted at pH 7.5 using Method a. Upper, direct representation; lower left, double reciprocal plot; lower right: Hill plot.
i
OL II- I I I I I I I I 0.01 0.1 1 10 100
Glucose, mM
FIG. 4. Hexokinase C activity as a function of glucose concentration at several fixed values of ATP concentration. Assays were conducted at pH 7.5 using Method a. Velocities are expressed as units/ml of enzyme preparation.
becomes progressively sigmoidal. Hill plots of the data (Fig. 6, right) show na values ranging from 1 at 0.2 mM or higher glucose concentrations, to 2.3 at 0.02 and 0.01 mM glucose.
The effect of varying the ATP concentration in the absence or presence of fixed levels of glucose-6-P on the velocity of the
curve whereas at lower concentrations of glucose the inhibition reaction at 0.2 mM glucose is presented in Fig. 6. Nonlinear
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from
5038 Allosteric Regulation of Hexokinase C from Amphibian Liver
0.01 0.1 1 IO 100
Glucose, mM
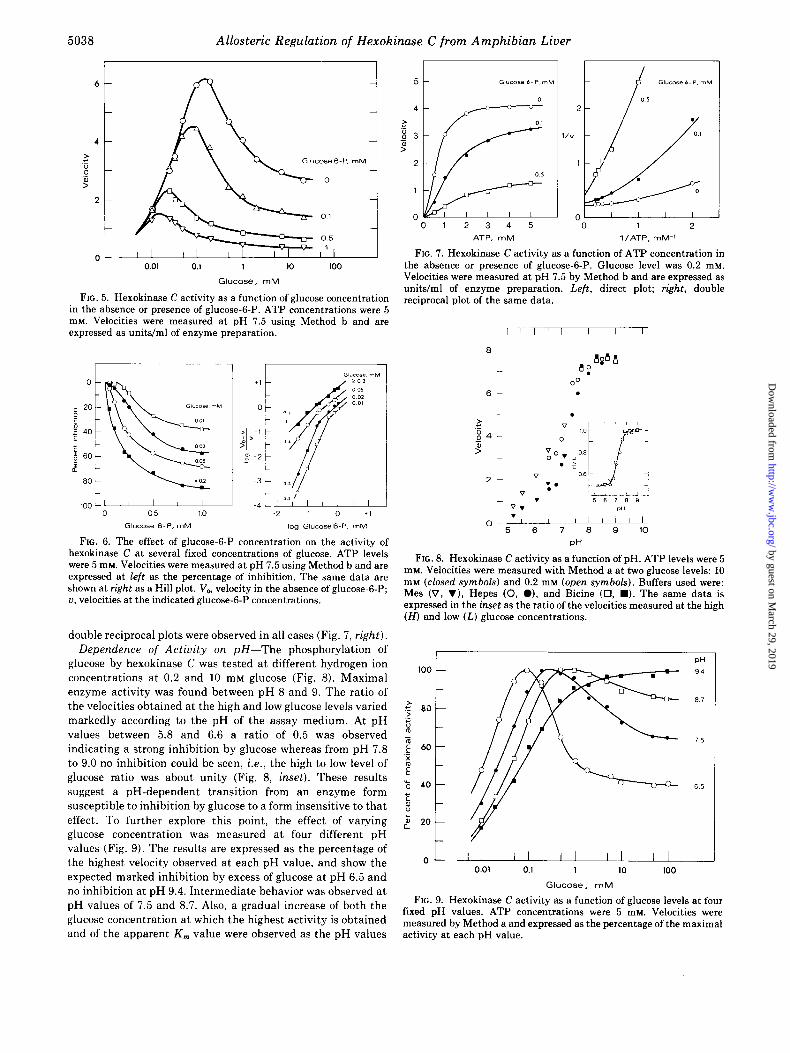
FIG. 5. Hexokinase C activity as a function of glucose concentration in the absence or presence of glucose-6-P. ATP concentrations were 5 rn~. Velocities were measured at pH 7.5 using Method b and are expressed as units/ml of enzyme preparation.
100 Ll I I I I 0 05 10
Glucose 6-P. rnM
c 4 LL
FIG. 6. The effect of glucose-6-P concentration on the activity of hexokinase C at several fixed concentrations of glucose. ATP levels were 5 mM. Velocities were measured at pH 7.5 using Method b and are expressed at left as the percentage of inhibition. The same data are shown at right as a Hill plot. V,, velocity in the absence of glucose-6-P; a, velocities at the indicated glucose-6-P concentrations.
double reciprocal plots were observed in all cases (Fig. 7, right). Dependence of Activity on pH-The phosphorylation of
glucose by hexokinase C was tested at different hydrogen ion concentrations at 0.2 and 10 mM glucose (Fig. 8). Maximal enzyme activity was found between pH 8 and 9. The ratio of the velocities obtained at the high and low glucose levels varied markedly according to the pH of the assay medium. At pH values between 5.8 and 6.6 a ratio of 0.5 was observed indicating a strong inhibition by glucose whereas from pH 7.8 to 9.0 no inhibition could be seen, i.e., the high to low level of glucose ratio was about unity (Fig. 8, inset). These results suggest a pH-dependent transition from an enzyme form susceptible to inhibition by glucose to a form insensitive to that effect. To further explore this point, the effect of varying glucose concentration was measured at four different pH values (Fig. 9). The results are expressed as the percentage of the highest velocity observed at each pH value, and show the expected marked inhibition by excess of glucose at pH 6.5 and no inhibition at pH 9.4. Intermediate behavior was observed at pH values of 7.5 and 8.7. Also, a gradual increase of both the glucose concentration at which the highest activity is obtained and of the apparent K, value were observed as the pH values
0 1 2 3 4 5
ATP, mM
0 1 2
l/ATP, mM-’
FIG. 7. Hexokinase C activity as a function of ATP concentration in the absence or presence of glucose-6-P. Glucose level was 0.2 mM. Velocities were measured at pH 7.5 by Method b and are expressed as units/ml of enzyme preparation. Left, direct plot; rig&, double reciprocal plot of the same data.
00 6 .
PH
FIG. 8. Hexokinase C activity as a function of pH. ATP levels were 5 mM. Velocities were measured with Method a at two glucose levels: 10 mt.r (closed symbols) and 0.2 mre (open symbols). Buffers used were: Mes (v, v), Hepes (0, l ), and Bicine (0, n ). The same data is expressed in the inset as the ratio of the velocities measured at the high (H) and low (L) glucose concentrations.
DH
OL - I I I I I I I I I I 0.01 0.1 1 10 100
Glucose, mM
FIG. 9. Hexokinase C activity as a function of glucose levels at four fixed pH values. ATP concentrations were 5 mM. Velocities were measured by Method a and expressed as the percentage of the maximal activity at each pH value.
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Allosteric Regulation of Hexokinase C from Amphibian Liver
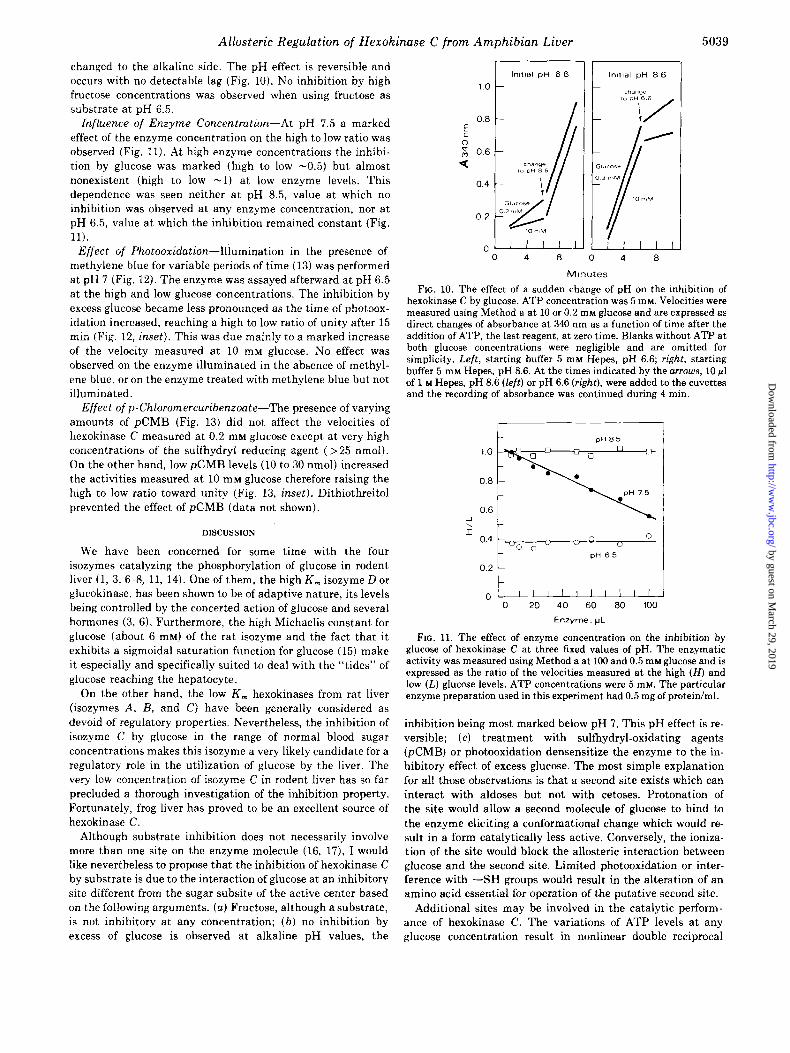
changed to the alkaline side. The pH effect is reversible and occurs with no detectable lag (Fig. 10). No inhibition by high fructose concentrations was observed when using fructose as substrate at pH 6.5.
Influence of Enzyme Concentration-At pH 7.5 a marked effect of the enzyme concentration on the high to low ratio was observed (Fig. 11). At high enzyme concentrations the inhibi- tion by glucose was marked (high to low -0.5) but almost nonexistent (high to low -1) at low enzyme levels. This dependence was seen neither at pH 8.5, value at which no inhibition was observed at any enzyme concentration, nor at pH 6.5, value at which the inhibition remained constant (Fig. 11).
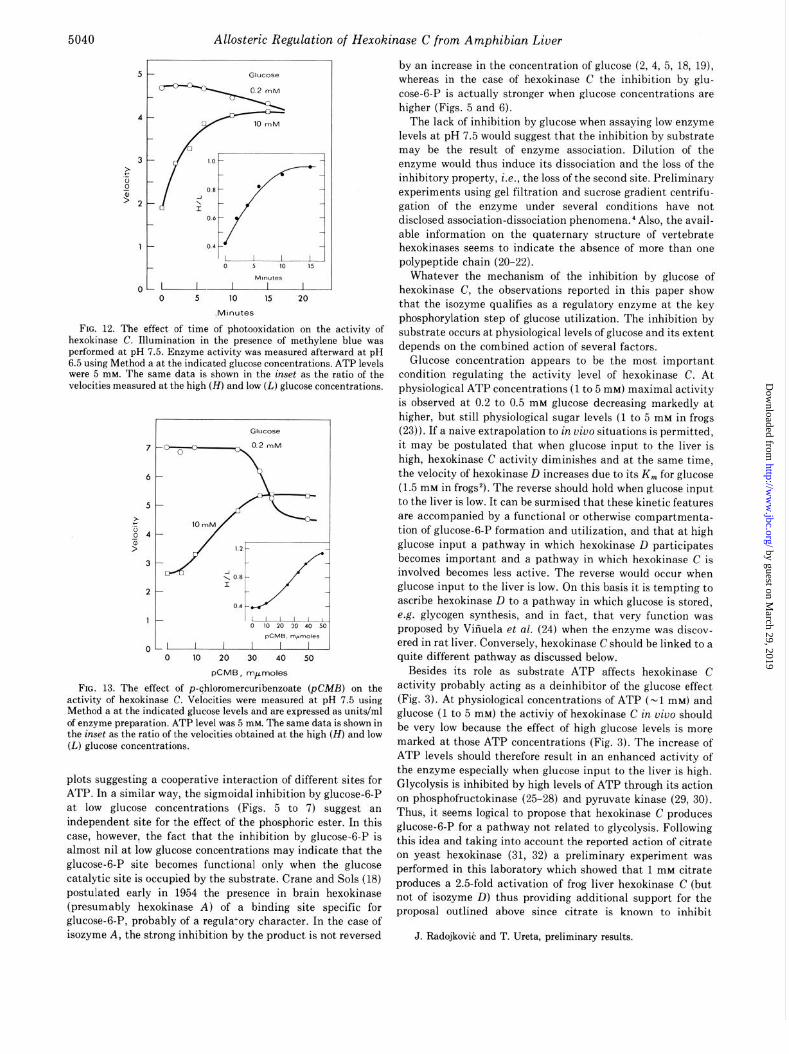
Effect of Photooxidation-Illumination in the presence of methylene blue for variable periods of time (13) was performed at pH 7 (Fig. 12). The enzyme was assayed afterward at pH 6.5 at the high and low glucose concentrations. The inhibition by excess glucose became less pronounced as the time of photoox- idation increased, reaching a high to low ratio of unity after 15 min (Fig. 12, inset). This was due mainly to a marked increase of the velocity measured at 10 mM glucose. No effect was observed on the enzyme illuminated in the absence of methyl- ene blue, or on the enzyme treated with methylene blue but not illuminated.
Effect ofp-Chloronercuribenzoate-The presence of varying amounts of &MB (Fig. 13) did not affect the velocities of hexokinase C measured at 0.2 mM glucose except at very high concentrations of the sulfhydryt reducing agent (> 25 nmol). On the other hand, low pCMB levels (10 to 30 nmol) increased the activities measured at 10 mM glucose therefore raising the high to low ratio toward unity (Fig. 13, inset). Dithiothreitol prevented the effect of &MB (data not shown).
DISCUSSION
We have been concerned for some time with the four isozymes catalyzing the phosphorylation of glucose in rodent liver (1, 3, 6-8, 11, 14). One of them, the high K, isozyme D or glucokinase, has been shown to be of adaptive nature, its levels being controlled by the concerted action of glucose and several hormones (3, 6). Furthermore, the high Michaelis constant for glucose (about 6 mM) of the rat isozyme and the fact that it exhibits a sigmoidal saturation function for glucose (15) make it especially and specifically suited to deal with the “tides” of glucose reaching the hepatocyte.
On the other hand, the low K, hexokinases from rat liver iisozymes A, B, and C) have been generally considered as devoid of regulatory properties. Nevertheless, the inhibition of isozyme C by glucose in the range of normal blood sugar concentrations makes this isozyme a very likely candidate for a regulatory role in the utilization of glucose by the liver. The very low concentration of isozyme C in rodent liver has so far precluded a thorough investigation of the inhibition property. Fortunately, frog liver has proved to be an excellent source of hexokinase C.
Although substrate inhibition does not necessarily involve more than one site on the enzyme molecule (16, 17), I would like nevertheless to propose that the inhibition of hexokinase C by substrate is due to the interaction of glucose at an inhibitory site different from the sugar subsite of the active center based on the following arguments. (a) Fructose, although a substrate, is not inhibitory at any concentration; (b) no inhibition by excess of glucose is observed at alkaline pH values, the
0.8
: 0 $, 0.6
0.2
5039
0 4 8 0 4 8
M!nutes
FIG. 10. The effect of a sudden change of pH on the inhibition of hexokinase C by glucose. ATP concentration was 5 mM. Velocities were measured using Method a at 10 or 0.2 rnM glucose and are expressed as direct changes of absorbance at 340 nm as a function of time after the addition of ATP, the last reagent, at zero time. Blanks without ATP at both glucose concentrations were negligible and are omitted for simplicity. Left, starting buffer 5 mM Hepes, pH 6.6; right, starting buffer 5 mM Hepes, pH 8.6. At the times indicated by the arrows, 10 ~1 of 1 M Hepes, pH 8.6 (left) or pH 6.6 (right), were added to the cuvettes and the recording of absorbance was continued during 4 min.
1
0 20 40 60 80 100
Enzyme, p’L
FIG. 11. The effect of enzyme concentration on the inhibition by glucose of hexokinase C at three fixed values of pH. The enzymatic activity was measured using Method a at 100 and 0.5 mM glucose and is expressed as the ratio of the velocities measured at the high (H) and low (L) glucose levels. ATP concentrations were 5 mM. The particular enzyme preparation used in this experiment had 0.5 mg of protein/ml.
inhibition being most marked below pH 7. This pH effect is re- versible; (c) treatment with sulfhydryl-oxidating agents (pCMB) or photooxidation densensitize the enzyme to the in- hibitory effect of excess glucose. The most simple explanation for all those observations is that a second site exists which can interact with aldoses but not with cetoses. Protonation of the site would allow a second molecule of glucose to bind to the enzyme eliciting a conformational change which would re- sult in a form catalytically less active. Conversely, the ioniza- tion of the site would block the allosteric interaction between glucose and the second site. Limited photooxidation or inter- ference with --SH groups would result in the alteration of an amino acid essential for operation of the putative second site.
Additional sites may be involved in the catalytic perform- ance of hexokinase C. The variations of ATP levels at any glucose concentration result in nonlinear double reciprocal
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from
5040 Allosteric Regulation of Hexokinase C from Amphibian Liver
OL -
r”bnutes I I I
0 5 IO 15 20
Minutes
FIG. 12. The effect of time of photooxidation on the activity of hexokinase C. Illumination in the presence of methylene blue was performed at pH 7.5. Enzyme activity was measured afterward at pH 6.5 using Method a at the indicated glucose concentrations. ATP levels were 5 mM. The same data is shown in the inset as the ratio of the velocities measured at the high (H) and low (L) glucose concentrations.
pCMB, m,moles
FIG. 13. The effect of p-qhloromercuribenzoate (pCMB) on the activity of hexokinase C. Velocities were measured at pH 7.5 using Method a at the indicated glucose levels and are expressed as units/ml of enzyme preparation. ATP level was 5 mhr. The same data is shown in the inset as the ratio of the velocities obtained at the high (H) and low (L) glucose concentrations.
plots suggesting a cooperative interaction of different sites for ATP. In a similar way, the sigmoidal inhibition by glucose-6-P at low glucose concentrations (Figs. 5 to 7) suggest an independent site for the effect of the phosphoric ester. In this case, however, the fact that the inhibition by glucose-6-P is almost nil at low glucose concentrations may indicate that the glucose-6-P site becomes functional only when the glucose catalytic site is occupied by the substrate. Crane and Sols (18) postulated early in 1954 the presence in brain hexokinase (presumably hexokinase A) of a binding site specific for glucose-6-P, probably of a regulatory character. In the case of isozyme A, the strong inhibition by the product is not reversed
by an increase in the concentration of glucose (2, 4, 5, 18, 191,
whereas in the case of hexokinase C the inhibition by glu- cose-6-P is actually stronger when glucose concentrations are higher (Figs. 5 and 6).
The lack of inhibition by glucose when assaying low enzyme levels at pH 7.5 would suggest that the inhibition by substrate may be the result of enzyme association. Dilution of the enzyme would thus induce its dissociation and the loss of the inhibitory property, i.e., the loss of the second site. Preliminary experiments using gel filtration and sucrose gradient centrifu- gation of the enzyme under several conditions have not disclosed association-dissociation phenomena.’ Also, the avail- able information on the quaternary structure of vertebrate hexokinases seems to indicate the absence of more than one polypeptide chain (20-22).
Whatever the mechanism of the inhibition by glucose of hexokinase C, the observations reported in this paper show that the isozyme qualifies as a regulatory enzyme at the key phosphorylation step of glucose utilization. The inhibition by substrate occurs at physiological levels of glucose and its extent depends on the combined action of several factors.
Glucose concentration appears to be the most important condition regulating the activity level of hexokinase C. At physiological ATP concentrations (1 to 5 mM) maximal activity is observed at 0.2 to 0.5 mM glucose decreasing markedly at higher, but still physiological sugar levels (1 to 5 mM in frogs (23)). I f a naive extrapolation to in uiuo situations is permitted, it may be postulated that when glucose input to the liver is high, hexokinase C activity diminishes and at the same time, the velocity of hexokinase D increases due to its K, for glucose (1.5 mM in frogs*). The reverse should hold when glucose input to the liver is low. It can be surmised that these kinetic features are accompanied by a functional or otherwise compartmenta- tion of glucose-6-P formation and utilization, and that at high glucose input a pathway in which hexokinase D participates becomes important and a pathway in which hexokinase C is involved becomes less active. The reverse would occur when glucose input to the liver is low. On this basis it is tempting to ascribe hexokinase D to a pathway in which glucose is stored, e.g. glycogen synthesis, and in fact, that very function was proposed by Viiiuela et al. (24) when the enzyme was discov- ered in rat liver. Conversely, hexokinase C should be linked to a quite different pathway as discussed below.
Besides its role as substrate ATP affects hexokinase C activity probably acting as a deinhibitor of the glucose effect (Fig. 3). At physiological concentrations of ATP (~1 mM) and glucose (1 to 5 mM) the activiy of hexokinase C in oiuo should be very low because the effect of high glucose levels is more marked at those ATP concentrations (Fig. 3). The increase of ATP levels should therefore result in an enhanced activity of the enzyme especially when glucose input to the liver is high. Glycolysis is inhibited by high levels of ATP through its action on phosphofructokinase (25-28) and pyruvate kinase (29, 30). Thus, it seems logical to propose that hexokinase C produces glucose-6-P for a pathway not related to glycolysis. Following this idea and taking into account the reported action of citrate on yeast hexokinase (31, 32) a preliminary experiment was performed in this laboratory which showed that 1 mM citrate produces a 2.5fold activation of frog liver hexokinase C (but not of isozyme D) thus providing additional support for the proposal outlined above since citrate is known to inhibit
J. Radojkovii: and T. Ureta, preliminary results.
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Allosteric Regulation of Hexokinase C from Amphibian Liver 5041
glycolysis through its strong inhibitory action on phosphofruc- tokinase (33-37). The participation of hexokinase C in glyco- genesis seems doubtful as discussed in the previous paragraph. It would be worthwhile to search intermediates of the pentose phosphate or uranic acid pathways as activators or deinhibitors of the enzyme for a more precise assignment of the function of hexokinase C.
At the pH values expected to exist in the hepatocyte, isozyme C is inhibited by glucose but the effect becomes less marked at higher pH values (Figs. 8 to 10). In the absence of information about pH variations in the cell, the physiological meaning of this observation can only be uncertain. Negative cooperativity and slow transients markedly pH-dependent have been described for yeast hexokinases (31, 32).
Inhibition by excess glucose at pH values near 7.5 was found to be linearly dependent on hexokinase C concentration (Fig. 11). Thus, at low enzyme concentrations the high to low glucose ratio approaches unity to decrease to values of about 0.5 with a lo- to 20-fold increase in the amount of enzyme- protein. The concentration of hexokinase C in the hepatocyte is unknown but, as judged by other cases (38, 39) certainly much higher than the maximal ones used in this study. It can then be inferred that hexokinase C in oiuo is almost in a nonfunctional condition when high glucose levels prevail but that it should become very active at low glucose levels. A related phenome- non has been described for phosphofructokinase (40, 41) in which case the inhibition by excess ATP is observed at low enzyme concentrations, the effect being markedly lower at high enzyme levels. For this reason it has been proposed that ATP is likely to be a rather poor inhibitor of phosphofructokinase in vivo (41).
Sols (42) has remarked that although a considerable number of enzymes are inhibited by excess substrate, the inhibition is usually moderate (about 50%) and require levels of the inhibitory substrates well above the physiological range, or some otherwise markedly unphysiological condition. Among other enzymes (fructose bisphosphatase, glyceraldehyde-3- phosphate dehydrogenase, lactate dehydrogenase H) he listed liver hexokinase C as an example of a substrate-inhibited enzyme in which no regulatory sites should be postulated “on the flimsy evidence of inhibition.” Also, Cleland (43) has pointed out that inhibition by excess substrate often indicates that the enzyme is being assayed in the direction opposite to that operating in uiuo. The results now reported should be taken as enough evidence to exclude hexokinase C from the above mentioned list. In fact, the enzyme fully complies with the exacting criteria outlined by Sols to recognize allosteric interactions of substrates (42). Also, Cleland’s arguments should not be applied to hexokinases since in uiuo the reaction is far displaced to the synthesis of glucose-6-P.
A few observations have been performed on hexokinase C from some mammals5 which allow a preliminary comparison with the frog enzyme. Generally speaking, the same properties of the frog enzyme are present in the monkey, rat, cow, sheep, and goat enzymes. However, these enzymes do not show the marked pH-dependent changes characteristic of frog hexoki- nase C. In the case of the ruminant enzyme the inhibition by glucose at 5 mM ATP is very pronounced, the high to low glucose concentration ratio at pH 7.5 being about 0.2. Further studies on these enzymes are in progress in this laboratory and will be soon reported.
“T. Ureta and S. Zepeda, unpublished results.
While writing this report I learned of a paper by Siano et al. (44) on the kinetic mechanism of bovine liver hexokinase C. However, all the experiments reported by Siano et al. were performed at glucose concentrations below 0.15 mM and there- fore no comparisons with the present data are possible.
Acknowledgments--I thank Dr. Frank Marcus for the sugges- tion of several key experiments. I have benefited by very helpful discussions with Dr. Catherine C. Allende, Dr. H. Niemeyer, Dr. J. Babul, and their collaborators. I am deeply indebted to Mr. J. C. Slebe, Mr. Carlos Lozano, and Mr. Rodrigo Bravo who collaborated in different stages of this work. The assistance of Miss Jasna Radojkovic in the completion of this research is gratefully acknowledged.
1.
2. 3.
4.
5.
6.
I.
8.
9.
10.
11.
12. 13. 14.
15.
16. 17. 18. 19.
20. 21.
22.
23. 24.
25.
26.
27.
28.
29.
30.
REFERENCES
Ureta. T. (1975) in Isozymes (Mark& C. L., ed) Vol. III, pp. 575- 601; Academic Press,New York
Walker, D. G. (1966) Essays Biochem. 2, 33-67 Niemeyer, H., and Ureta, T. (1972) in Molecular Basis of Biological
Actiuitv (Gaede. K.. Horecker, B. L., and Whelan, W. J., eds) PAABS Sympo&m Vol. 1, pp. 221-273, Academic Press, New York
Colowick, S. P. (1973) in The Enzymes (Boyer, P. D., ed) Vol. 9, Part B, pp. l-48, Academic Press, New York
Purich, D. L., Fromm, H. J., and Rudolph, F. B. (1973) Adu. Enzymol. Reiat. Areas Mol. Biol. 35,249-326
Niemeyer, H., Ureta, T., and Clark-Turri, L. (1975) Mol. Cell. Biochem. 6,109-126
Gonzalez, C., Ureta, T., Sanchez, R., and Niemeyer, H. (1964) Biochem. Biophys. Res. Commun. 16, 347-352
Gonzalez, C., Ureta, T., Babul, J., Rabajille, E., and Niemeyer, H. (1967) Biochemistry 6.460-468
Grossbard, L., and Schimke, R. T. (1966) J. Biol. Chem. 241, 3546-3560
Lynch, J. D. (1971) Misc. Publ. Mu. Nat. Hist., Univ. Kansas 53, l-238
Ureta, T., Gonzalez, C., Lillo, S., and Niemeyer, H. (1971) Comp. Biochem. Physiol. 40,71-80
Miller, G. L. (1959) Anal. Chem. 31, 964 Ray, W. J., Jr. (1966) Methods Enzymol. 11, 490-497 Ureta, T., Gonzalez, C., and Niemeyer, H. (1971) Comp.
Biochem. Physiol. 40,81-91 Niemeyer, H., Cardenas, M. L., Rabajille, E., Ureta, T., Clark-
Tnrri, L., and Penaranda, J. (1975) Enzyme 20.321-333 Alberty, R. A. (1956) Adu. Enzymol. 17, l-64 Ferdinand, W. (1966) Biochem. J. 98, 278-283 Crane, R. K., and Sols, A. (1954) J. Biol. Chem. 210, 597-606 Crane, R. K. (1962) in The Enzymes (Boyer, P. D., Lardy, H.,
and Myrback, K., (eds) Vol. 6, pp. 47-66, Academic Press, New York
Easterby, J. S. (1971) FEBS Lett. l&23-26 Chou, A. C., and Wilson, J. E. (1972) Arch. Biochem. Biophys. 151,
4655 Maccioni, R., and Babul, J. (1974) Abstracts of the 17th Annual
Meeting, Sociedad de Biologia de Chile, Concepcih, Chile, p. R-32
Penhos, J. C., and Ramey, E. (1973) Am. Zool. 13, 667-698 Viiiuela, E., Salas, M., and Sols, A. (1963) J. Biol. Chem. 238,
PC117551177 Mansour, T. E., and Mansour, J. M. (1962) J. Biol. Chem. 237,
629-634 Passonneau, J. V., and Lowry, 0. H. (1962) Biochem. Biophys.
Res. Common. 7. IO-15 Viriuela, E., Salas, M. L., and Sols, A. (1963) Biochem. Biophys.
Res. Commun. 12,140-145 Lowry, 0. H., and Passonneau, J. V. (1966) J. Biol. Chem. 241,
2268-2279 Reward, A. M., Hass. L. F. Jacobsen. D. D.. and Bayer. P. D.
-(1961) J. Biol: Chek 236,2277-2283 ” Carbonell, J., Feliu, J. E., Marco, R., and Sols, A. (1973) Eur. J.
Biochem. 37,148-156
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from

5042 Alloste~i~ Regulation of Herokinase C fron Amphibian Liuer
31. Kosow, D. P., and Rose, I. A. (1971) J. Biol. Chem. 246, 2618- 2625
32. Shill, J. P., and Neet, K. E. (1975) J. Biol. Chem. 250, 2259-2268 33. Parmeggiani, A., and Bowman, R. H. (1963) Biochem. Biophys.
Res. Commun. 12,X%-273 34. Viriuela, E., Salas, M. L., Salas, M., and Sols, A. (1964) Biochem.
Biophys. Res. Commun. 15,243-249 35. Williamson, J. R. (1965) J. Biol. Chem. 240,2308-2321 36. Salas, M. L., Viiiuela, E., Salas, M., and Sols, A. (1965) Biochem.
Biophys. Res. Common. 19,371-376 37. Passoneau, J. V., and Lowry, 0. H. (1963) Biochem. Biophys. Res.
Commun. 13,372-379
38. Srere, P. A. (1967) Science 158.936-937 39. Srere, P. A. (1970) Biochem. Med. 4,43-46 40. Hofer, H. W. (1971) Hoppe-Seyler’s Z. Physiol. Chem. 352, 997-
1004 41. Hulme, E. C., and Tipton, K. F. (1971) FEBS Lett. 12, 197-200 42. Sols, A. (1973) Mechunisms and Control Properties of Phospho-
transferuses, pp. 239-251, Akademie-Verlag, Berlin 43. Cleland, W. W. (1970) in The Enzymes (Bayer, P. D., ed) Vol. 2,
pp. l-65, Academic Press, New York 44. Siano, D. B., Zyskind, J. W., and Fromm, H. J. (1975) Arch.
Biochem. Biophys. 170,587-600
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from

T UretaThe allosteric regulation of hexokinase C from amphibian liver.
1976, 251:5035-5042.J. Biol. Chem.
http://www.jbc.org/content/251/16/5035Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/251/16/5035.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 29, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























