Embed Size (px)
Citation preview

The Acute Regulation of Intestinal Chylomicron Secretion
by Glucagon-Like Peptides
by
Joanne Hsieh
A thesis submitted in conformity with the requirements
for the degree of Doctorate of Philosophy
Graduate Department of Biochemistry
University of Toronto
© Copyright by Joanne Hsieh (2012)

ii
The Acute Regulation of Intestinal Chylomicron Secretion by Glucagon-
Like Peptides
Joanne Hsieh
Doctorate of Philosophy
Graduate Department of Biochemistry
University of Toronto
2012
ABSTRACT
Postprandial overproduction of apolipoprotein B48 (apoB48)-containing lipoproteins has
been observed in states of insulin resistance and is important to the sequelae of cardiovascular
disease, but little is understood about factors that regulate their secretion. The glucagon-like
peptides (GLPs) are released from ileal enteroendocrine L-cells following lipid ingestion. I
hypothesized that the GLPs could acutely affect the production of apoB48-containing
triglyceride (TG)-rich lipoproteins (TRL) in the small intestine. Using the Syrian golden
hamster, I first characterized the gross effects of the GLPs on TRL secretion in response to an
oral fat load and then continued to dissect the mechanisms of these changes using primary
intestinal cell cultures and a variety of knockout mouse models. An exogenous GLP-1 receptor
(GLP-1R) agonist was found to acutely inhibit chylomicron secretion in both hamsters and
mouse models, and extending the bioactivity of endogenously-secreted GLP-1 with a dipeptidyl
peptidase-4 inhibitor had suppressive effects in insulin-resistant fructose-fed hamsters. The
insulinotropic and delayed gastric emptying functions do not completely account for the
hypolipidemic effect of GLP-1R agonism, and the effect of the GLP-1R agonist exendin-4 could

iii
be seen directly in the apoB48 secretion of primary enterocytes. In contrast, the sister peptide
GLP-2 was a potent acute stimulator of chylomicron secretion in hamsters and mice. The
hyperlipidemic effect of GLP-2 could be attributed to an increased rate of luminal FA uptake
mediated by the posttranslational modification of the FA transporter CD36, and CD36-deficient
mice were found to be refractory to the stimulatory effects of GLP-2. The activity of nitric oxide
synthase was also found to be essential to the hyperlipidemic action of GLP-2. I identified a set
of intercellular communications that could contribute in mediating the action of GLP-2, in which
GLP-2 secreted from the enteroendocrine L-cell stimulates intestinal subepithelial
myofibroblasts to release vascular endothelial growth factor, which directly activated the
enterocyte to secrete apoB48. In summary, this thesis demonstrates that two co-secreted
postprandial hormones have considerable but completely opposite influences on chylomicron
production. Changing the balance of the GLPs’ actions in vivo could provide a therapeutic
strategy to combat postprandial dyslipidemia.

iv
ACKNOWLEDGMENTS
This Ph.D. thesis was a long endeavour that could not have been accomplished without
the support and input of many people. It began with my parents Tse Kao and Yu Sze in
supporting my decision to move from beautiful British Columbia to Toronto to pursue my
studies at the University of Toronto. I have to thank my husband Edward who, despite not being
trained in science, was indispensable to the completion of the thesis. He was my motivation,
respite from the frustrations of problematic experiments, and even in a couple of instances, my
financial bailout when annual tuition fees were due. I would also like to express gratitude to my
sisters Vivian and Yvonne, and my parents-in-law Thomas and Rebecca, for all making this
journey a little easier.
My supervisor Dr. Khosrow Adeli always managed to provide project funding, review
manuscripts, input on oral presentations, and mentorship on my academic career all while having
the busiest professional and travel schedule of anyone I personally know. Much of the
confidence I have come to acquire over the years have come from his encouragement. I came
with an interest in lipid biochemistry, and I can honestly say I am leaving the lab with a love for
lipids. My committee members Dr. Amira Klip and Dr. Patricia Brubaker have provided
invaluable suggestions for experiments during each and every one of my supervisory committee
meetings, and I’d hate to imagine where I would be without the immense knowledge of GLP
physiology and intestinal biology of Dr. Brubaker. The technical support of Angelo Izzo with
the conditioned media studies presented in Chapter 4 is also very much appreciated. Dr. Daniel
Drucker has also been an integral part of this thesis with his involved collaboration in these
projects, and the successful publication of the papers in this thesis would not have been possible
with some of the experiments conducted by Dr. Christine Longuet.
Many people over the years have commented that the lab has an exceptional
environment, and I couldn’t agree more. My fellow lab members were not only brainstormers,
troubleshooters, and extra hands during technically-demanding experiments; they were my
closest friends in Toronto. A list of everyone would necessitate its own thesis chapter, but
standouts have included past (Julie Tsai, Angela Rutledge, Elaine Xu, Rita Kohen, Bolin Qin,
Diana Wong, Joanna Nelken, Jennifer Webb and Amanda Hayashi) and present (Chris Baker,

v
Mark Naples, Rianna Zhang, Mark Dekker, Wei Qiu, Qiaozhu Su, and Man Khun Chan)
members. In addition, my classmates in the Molecular Structure and Function department of The
Hospital for Sick Children could always be counted on for fun, relaxation, and reagents in a
pinch.
Finally, I would like to express my gratitude to the Department of Biochemistry of the
University of Toronto and The Hospital for Sick Children. My thesis project has been
financially supported by The Hospital for Sick Children’s Restracomp program, the National
Sciences and Engineering Research Council of Canada Canada Graduate Scholarship M, and the
Frederick Banting and Charles Best Canada Graduate Scholarship D and an operating grant from
the Canadian Institutes of Health Research.

vi
TABLE OF CONTENTS
Abstract ........................................................................................................................................... ii
Acknowledgments.......................................................................................................................... iv
Table of Contents ........................................................................................................................... vi
List of Tables ................................................................................................................................. xi
List of Figures ............................................................................................................................... xii
List of Abbreviations ................................................................................................................... xiv
Chapter 1: Introduction ..............................................................................................................1
1.1 Clinical significance.............................................................................................................2
1.2 Intestinal Fat Absorption and Chylomicron Assembly ........................................................3
1.2.1 Dietary lipid uptake..................................................................................................3
1.2.2 Chylomicron assembly and secretion ......................................................................5
1.2.3 Naturally occurring mutation in fatty acid transporters .........................................11
1.2.4 Insulin action and insulin resistance ......................................................................14
1.2.5 Hormonal regulation of chylomicron secretion .....................................................16
1.2.6 Other fates of dietary lipid in the intestine.............................................................20
1.2.7 Chylomicron catabolism ........................................................................................21
1.3 Glucagon-Like Peptides .....................................................................................................23
1.3.1 Glucagon-like peptide structure .............................................................................23
1.3.2 Intestinal GLP secretion .........................................................................................25
1.4 Biology of GLP-1 ..............................................................................................................27
1.4.1 Physiological functions of GLP-1 ..........................................................................27
1.4.2 GLP-1 receptor.......................................................................................................27

vii
1.4.3 GLP-1 and lipid metabolism ..................................................................................28
1.5 Biology of GLP-2 ..............................................................................................................30
1.5.1 Biological functions of GLP-2 ...............................................................................30
1.5.2 GLP-2 and intestinal nutrient absorption ...............................................................31
1.5.3 GLP-2 receptor.......................................................................................................35
1.6 Hypothesis..........................................................................................................................37
Chapter 2: The glucagon-like peptide-1 receptor regulates postprandial lipoprotein
synthesis and secretion ..............................................................................................................38
2.1 Summary ............................................................................................................................39
2.2 Introduction ........................................................................................................................40
2.3 Materials and Methods .......................................................................................................42
2.3.1 Animals ..................................................................................................................42
2.3.2 Assessment of intestinal lipoprotein production by in vivo Triton-WR1339
infusion ..................................................................................................................42
2.3.3 Isolation of triglyceride-rich lipoproteins (TRL) ...................................................43
2.3.4 Chemiluminescent immunoblotting .......................................................................43
2.3.5 Ex vivo metabolic labeling of intact primary enterocytes ......................................44
2.3.6 Immunoprecipitation, SDS-PAGE, and fluorography ...........................................44
2.3.7 Plasma measurements ............................................................................................45
2.3.8 Fast protein liquid chromatography of plasma lipoproteins ..................................45
2.3.9 Statistical analysis ..................................................................................................45
2.4 Results ................................................................................................................................46
2.4.1 A DPP-4 inhibitor attenuates dyslipidemia in fructose-fed hamsters ....................46
2.4.2 A DPP-4 inhibitor decreases intestinal production of TRL-TG and TRL-
cholesterol ..............................................................................................................49
2.4.3 Pharmacological activation of the GLP1R mimics the effects of sitagliptin on
intestinal lipid absorption .......................................................................................53

viii
2.4.4 Endogenous GLP-1R signalling is required for control of postprandial lipemia ..57
2.4.5 Intact GLP-1R signalling is required for the hypolipidemic action of
sitagliptin................................................................................................................57
2.4.6 Exendin-4 directly reduces enterocyte ApoB48 secretion .....................................60
2.5 Discussion ..........................................................................................................................62
Chapter 3: GLP -2 increases intestinal lipid absorption and chylomicron production via
CD36 ................................................................................................................................65
3.1 Summary ............................................................................................................................66
3.2 Introduction ........................................................................................................................67
3.3 Materials and Methods .......................................................................................................69
3.3.1 Antibodies and chemicals ......................................................................................69
3.3.2 Animals ..................................................................................................................69
3.3.3 Determination of triglyceride-rich lipoprotein apoB48 secretion in vivo in
hamsters .................................................................................................................69
3.3.4 Determination of TG-rich lipoprotein apoB48 secretion in vivo in mice ..............70
3.3.5 Isolation of TRL .....................................................................................................70
3.3.6 FPLC of plasma lipoproteins .................................................................................70
3.3.7 Density ultracentrifugation of plasma lipoproteins ................................................71
3.3.8 Metabolic labelling of primary jejunal fragments ex vivo .....................................71
3.3.9 Monitoring labelled triolein secretion in vivo ........................................................71
3.3.10 In situ apical membrane protein biotinylation .......................................................72
3.3.11 Statistical analysis ..................................................................................................72
3.4 Results ................................................................................................................................73
3.4.1 GLP-2 acutely increases circulating levels of apoB48-containing TRL in
hamsters .................................................................................................................73
3.4.2 GLP-2 accelerates secretion of luminal fatty acids................................................78
3.4.3 Acute GLP-2 treatment increases apoB48-containing TRL production in mice ...81

ix
3.4.4 GLP-2-stimulated chylomicron secretion requires CD36 ......................................83
3.5 Discussion ..........................................................................................................................86
Chapter 4: Intercellular communications mediating GLP-2-stimulated chylomicron
secretion ................................................................................................................................90
4.1 Summary ............................................................................................................................91
4.2 Introduction ........................................................................................................................92
4.3 Materials and Methods .......................................................................................................94
4.3.1 Animals ..................................................................................................................94
4.3.2 Determination of dietary fat absorption in vivo .....................................................94
4.3.3 Determination of postprandial TRL production in vivo .........................................95
4.3.4 Determination of intestinal apoB48 secretion ex vivo ...........................................95
4.3.5 ISEMF-conditioned media .....................................................................................96
4.3.6 Steady state labelling of apoB48 production ex vivo .............................................96
4.3.7 Other biochemical measurements ..........................................................................96
4.3.8 Kinex phosphoprotein screen .................................................................................97
4.3.9 Statistical analysis ..................................................................................................97
4.4 Results ................................................................................................................................98
4.4.1 In vivo role of NO in GLP-2-stimulated postprandial TRL secretion ...................98
4.4.2 Ex vivo role of NO in apoB48 secretion ..............................................................100
4.4.3 Role of eNOS in GLP-2-stimulated chylomicron secretion ................................102
4.4.4 Role of ISEMFs in GLP-2-stimulated apoB48 secretion ....................................104
4.4.5 VEGF action in enterocyte apoB48 secretion ......................................................106
4.4.6 VEGF as a mediator linking ISEMFs and enterocytes ........................................108
4.5 Discussion ........................................................................................................................110
Chapter 5: Discussion and conclusions .................................................................................115

x
5.1 Summary of Results .........................................................................................................116
5.2 GLP-1 as a Direct Regulator of Intestinal Lipoprotein Metabolism................................117
5.3 CD36 as a Hormone-Sensitive FA Transporter in the Gut ..............................................118
5.4 VEGF and NO as Modulators of Intestinal Function ......................................................120
5.5 Concerted GLP-1 and GLP-2 Action ...............................................................................122
5.6 Future Directions .............................................................................................................128
5.7 Conclusions ......................................................................................................................129
Permission to Publish Copyrighted Material ...............................................................................130
References ....................................................................................................................................148

xi
LIST OF TABLES
Table 1-1. Biological functions of GLP-1 and GLP-2 ................................................................ 34
Table 2-1. Body mass and blood glucose following sitagliptin treatment. .................................. 48
Table 2-2. TRL fraction measurements in chow-fed mouse studies ........................................... 51

xii
LIST OF FIGURES
Figure 1-1. Dietary lipid absorption and chylomicron assembly in the enterocyte. ...................... 9
Figure 1-2. Insulin signalling pathway depicted with perturbations known to occur in the
enterocyte during insulin resistance. ............................................................................................. 18
Figure 1-3. Differential proglucagon polypeptide processing in pancreatic alpha-cells and
intestinal enteroendocrine L-cells ................................................................................................. 24
Figure 2-1. Changes in plasma lipids following administration of sitagliptin ............................. 47
Figure 2-2. Changes in TRL lipid mass following chronic sitagliptin administration ................ 50
Figure 2-3. Changes in postprandial lipid secretion following a single acute administration of
sitagliptin in chow-fed mice.......................................................................................................... 52
Figure 2-4. Pharmacological activation of the GLP1R mimics the effect of sitagliptin on post-
prandial lipid excursion................................................................................................................. 55
Figure 2-5. Role of glucose-modulating hormones in mice ........................................................ 56
Figure 2-6. Role of GLP-1 in postprandial lipemia and sitagliptin action................................... 58
Figure 2-7. Direct effects of exendin-4 on intestinal apoB48 metabolism ex vivo ...................... 61
Figure 3-1. In vivo effects of GLP-2 on apoB48-containing TRL secretion ............................... 75
Figure 3-2. Plasma lipoprotein profiling by salt-density sedimentation in hamsters .................. 76
Figure 3-3. Ex vivo effects of GLP-2 on apoB48-containing lipoprotein secretion .................... 77
Figure 3-4. GLP-2 accelerates secretion of luminal fatty acids in hamsters ............................... 79
Figure 3-5. GLP-2 enhances intestinal CD36 action in hamsters ................................................ 80

xiii
Figure 3-6. Stimulatory effects of GLP-2 on intestinal lipid absorption and apoB48-chylomicron
production in wild type mice ........................................................................................................ 82
Figure 3-7. Lack of GLP-2 mediated stimulation of intestinal lipoprotein secretion in Cd36-/-
mice ............................................................................................................................................... 84
Figure 4-1. The role of nitric oxide in apoB48 production in hamsters in vivo ........................... 99
Figure 4-2. The effect of an NO donor on apoB48 production in hamster enterocytes ex vivo 101
Figure 4-3. Effect of GLP-2 on dietary fat absorption and postprandial lipoprotein secretion in
C57Bl/6J and eNOS KO mice .................................................................................................... 103
Figure 4-4. Effect of ISEMF conditioned media on apoB48 production in primary hamster
enterocytes .................................................................................................................................. 105
Figure 4-5. The role of VEGF in intestinal apoB48 secretion in mice ...................................... 107
Figure 4-6. The role of VEGF in GLP-2-stimulated ISEMF-mediated intestinal apoB48
secretion in enterocytes ............................................................................................................... 109
Figure 4-7. Intercellular interactions resulting in GLP-2-stimulated chylomicron secretion .... 111
Figure 5-1. Physiological contributions of GLP-1 and GLP-2 to chylomicron secretion ......... 126

xiv
LIST OF ABBREVIATIONS
ABCG5 ATP binding cassette , sub-family G, member 5
ACAT2 acyl-CoA:cholesterol acyltransferase 2
ACC acetyl-CoA carboxylase
AMPK adenosine monophosphate-activated protein kinase
apoAI apolipoprotein AI
apoAIV apolipoprotein AIV
apoB48 apolipoprotein B48
apoCII apolipoprotein CII
apoCIII apolipoprotein CIII
BBM brush border membrane
CD36 cluster of differentiation CD36
CPT-1 carnitine palmitoyltransferase-I
CVD cardiovascular disease
DAG diacylglycerol
DGAT acyl-CoA:diacylglycerol acyltransferase
DPP-4 dipeptidyl peptidase-4
EGF epidermal growth factor
eNOS endothelial nitric oxide synthase
ER endoplasmic reticulum
ERK1/2 extracellular related kinase 1/2
FA fatty acid
FAS fatty acid synthase
FATP4 fatty acid transporter 4
FFA free fatty acids
FPLC fast protein liquid chromatography
GIP glucose-dependent insulinotropic peptide
GLP-1 glucagon-like peptide-1
GLP-1R glucagon-like peptide-1 receptor
GLP-2 glucagon-like peptide-2
GLP-2R glucagon-like peptide-2 receptor
GLUT2 glucose transporter 2
GPR119 G protein-coupled receptor 119
GPR120 G protein-coupled receptor 120
GPR40 G protein-coupled receptor 40
GSK-3 glycogen synthase kinase-3
GSNO S-nitroso-L-glutathione
HDL high density lipoprotein
HFD high fat diet
IGF-1 insulin-like growth factor-1
IL-6 interleukin-6
iNOS inducible nitric oxide synthase

xv
ISEMF intestinal subepithelial myofibroblast
JNK c-Jun N-terminal kinase
KGF keratinocyte growth factor
LDL low density lipoprotein
L-FABP liver fatty acid binding protein
L-NAME NG-nitro-L-arginine methyl ester
L-NMMA L-NG-monomethyl-L-arginine
LpL lipoprotein lipase
MAG monoacylglycerol
MAPK mitogen activated protein kinase
MGAT acyl-CoA:monoacylglycerol acyltransferase
MTP microsomal triglyceride transfer protein
NPC1L1 Niemann-Pick C1-like 1
NTS nucleus of the solitary tract
PC1/3 prohormone convertase 1/3
PCTV prechylomicron transport vesicle
PI3-K phosphatidylinositol-3-kinase
PKA protein kinase A
PKC protein kinase C
PPAR peroxisome proliferator-activated receptor
SCD-1 stearoyl-CoA desaturase-1
SGLT-1 sodium-glucose cotransporter-1
SNARE soluble N-ethylmaleimide sensitive factor attachment protein
SR-BI scavenger receptor class B type I
SREBP-1c sterol responsive element binding protein-1c
STZ streptozotocin
T2DM type 2 diabetes mellitus
TCA trichloroacetic acid
TG triglyceride
TGF- transforming growth factor-
TNF- tumour necrosis factor-
TRL triglyceride-rich lipoprotein
VAMP vesicle-associated membrane protei
VEGF vascular endothelial growth factor
VEGFR2 vascular endothelial growth factor receptor 2
VIP vasoactive intestinal peptide
VLDL very low density lipoprotein

1
Chapter 1: INTRODUCTION
Figure 1-2 is reproduced from:
Hsieh J, Hayashi AA, Webb J, Adeli K. Postprandial dyslipidemia in insulin resistance:
mechanisms and role of intestinal insulin sensitivity. Atheroscler Suppl. 2008;9(2):7-13.
Copyright 2008, Elsevier.
Author contribution: J. Hsieh produced all text and figures in this chapter.

2
1.1 Clinical significance
Cardiovascular disease (CVD) continues to be the largest cause of mortality in the world
(1). While CVD is traditionally thought of as a problem limited to developed nations, the
prevalence of risk factors for CVD, such as obesity, is rising worldwide (2). Low density
lipoprotein (LDL) has long been targeted as a causative factor of atherosclerosis, but growing
evidence suggest that intestinally-derived lipoproteins also play a role in the pathogenesis of
atherosclerotic heart disease (3-6). There is pathological and experimental evidence to indicate
that intestinally-derived apolipoprotein B48 (apoB48)-containing lipoproteins accumulate in the
arterial wall to deposit cholesterol (7; 8) and induce endothelial dysfunction (9). Postprandial
hyperlipidemia is also an inherent feature of diabetic dyslipidemia (10), thereby including it as
part of a major complication of a disease that has been identified as a growing epidemic.
Moreover, given typical Western eating patterns are based on three meals a day, humans spend
the majority of their day with some degree of postprandial lipemia (11). Understanding the
mechanisms regulating postprandial lipid metabolism is thus clearly important in elucidating the
pathways underlying the pathogenesis of atherosclerosis and its cardiovascular complications.
However, our knowledge of lipid handling in the intestine pales in comparison to what is known
about hepatic lipoprotein metabolism, and even more lacking is our understanding of how
intestinal lipoprotein secretion is regulated.

3
1.2 Intestinal Fat Absorption and Chylomicron Assembly
1.2.1 Dietary lipid uptake
The digestion of ingested fat in the intestine begins with its lipolysis by pancreatic juices
and emulsification by bile in the duodenum and upper jejunum. The action of pancreatic lipase
and colipase on triglyceride (TG) results in free fatty acid (FFA) and sn2-monoacylglycerol
(MAG), and the micellization of these products with bile acids and phospholipids prepares lipid
for uptake across the apical membrane of the enterocyte (reviewed in (12)). Because of the high
efficiency of fat absorption in the mammalian gut, and the high luminal concentrations in the
postprandial state, fatty acids (FA) and MAG were thought to enter the enterocyte by simple
diffusion across the phospholipid bilayer. However, there is a large body of evidence in different
cell lines and organs that indicate FA transport is also a saturable protein-facilitated process.
For the intestine, a number of proteins have been suggested to transport FA at the apical
membrane. Cluster of differentiation 36 (CD36)/fatty acid translocase is a widely expressed
scavenger type B receptor, with a hairpin topology that has short cytoplasmic tails but the large
extracellular loop contains a stretch of hydrophobic residues that could serve to bind fatty acids.
CD36 has three extracellular disulphide bridges and 10 putative N-linked glycosylation sites that
confer this 53 kDa protein an apparent mass of 88 kDa (reviewed in (13)). CD36 expression has
been documented to be on the cell surface in caveolae, where its fatty acid transport activity
occurs (14; 15), and is important for FA transport into myocytes (16), adipocytes (17), and
hepatocytes (18). Because CD36 has been demonstrated to bind multiple ligands in other tissues,
including oxidized LDL, thrombospondin, and malaria-infected erythrocytes, it is not surprising
it exhibits some ligand promiscuity in the intestine and has been implicated in the transport of
both long chain FA and cholesterol. The role of CD36-mediated FA uptake is most prominent in
the proximal intestine (19), although its expression along the gastrointestinal tract has been
reported to be highest in the ileum (20). Also in the proximal intestine, CD36 participates in the
uptake of the less efficiently absorbed cholesterol (21). CD36 transport activity appears to be
selective to very long chain FA, especially those over 24 carbons in length (22), while Cd36-/-
mice remain capable of absorbing most dietary FA. With regards to the absorption of the more
common 18 carbon-long oleic acid, CD36 plays a key role in its output from the intestinal

4
epithelium, where CD36 deficiency resulted in lipid trapping in the intestinal epithelium (21;
23). Also in the gastrointestinal tract, CD36’s affinity for long chain FA is thought to be
involved in conferring a gustatory disposition for lipids. CD36 is expressed in taste buds and
CD36-deficient mice do not exhibit any preference for long chain FA-enriched solutions (24).
Another scavenger type B receptor, scavenger receptor type B-I (SR-BI) has been
implicated in intestinal lipid absorption. SR-BI is known for its role in high density lipoprotein
(HDL)-binding and reverse cholesterol transport, but has been found to be concentrated in the
microvilli of enterocytes (25) and has a structure very similar to CD36. Intestine-specific
overexpression of SR-BI has been shown to increase absorption of both oleic acid and
cholesterol (26). However, SR-BI is typically regarded as a cholesterol transporter (27), given
its documented high affinity for cholesterol in the brush border membrane (BBM) (28). During
the absorptive state, SR-BI shows signs of internalization and trafficking to intracellular lipid
droplets in the enterocyte (29). Although SR-BI increases cholesterol across the BBM, it does
not form the rate limiting step in cholesterol absorption (30). Nieman-Pick C1 Like 1 (NPC1L1)
is also integral to cholesterol transport, as it was identified as the pharmacological target of
ezetimibe, a small molecule that inhibits intestinal cholesterol absorption (31). A deficiency in
NPC1L1 was clearly associated with a drastic reduction in dietary sterol absorption (32). Like
SR-BI, intracellular movements are essential to NPC1L1-mediated cholesterol transport, as the
mechanism of NPC1L1 action involves vesicular endocytosis, and ezetimibe affects its cycling
between endosomal compartments and the plasma membrane (33). In what is beginning to
appear as a common feature of membrane lipid transporters, NPC1L1 has been localized to
specific lipid raft domains (34).
Even if appreciable amounts of dietary FA likely pass through the apical membrane by
simple diffusion, the efficiency of absorption would suggests that is vectorial in nature, that is,
lipid traverses only in the apical to basolateral direction in enterocytes, which implicates a
protein-mediated mechanism. There are six members in the fatty acid transport protein (FATP)
family, but only FATP1 and FATP4 have been reported to be detected in the small intestine (35;
36). FATP4 is abundantly expressed in the small intestine in the microvilli (36) and endoplasmic
reticulum (ER) (37) of epithelial cells. Deleting one allele of Fatp4 almost halves the FA uptake
in enterocytes (38), but FATP4 also exhibits acyl-CoA synthetase activity, with greater

5
specificity for very long chain FA (39). It is argued that the acyl-CoA synthetase activity drives
FA uptake (37), presumably because the conjugation of the coenzyme A helps to maintain the
FA inside enterocytes. Mice with two mutant alleles of Fatp4 suffer from neonatally lethal skin
defects, but rescued Fatp4-/-
mice that only express FATP4 under a keratinocyte-specific
promoter do not have compromised dietary FA absorption and uptake, although FATP4 may be
necessary to target FA for secretion as TG on a high fat diet (40). FATP1 also possesses acyl-
CoA synthetase activity, and its expression in cells drives oleic acid import and assimilation into
TG (41). As for FATP1, though expressed in the small intestine in moderate amounts (35), no
function has been ascribed to it in the organ. Liver fatty acid binding protein (L-FABP) is found
in jejunal BBM and can bind FA starting from 16 carbons in length (42). Hepatocytes also
express high amounts of L-FABP, where its presence facilitates FA uptake (43). L-FABP is also
found in the cytoplasm, where it accelerates the intracellular movement of its bound FA, possibly
directing the FA for esterification (44). Each L-FABP molecule can bind two FA molecules
(45), and L-FABP efficiently sequesters the bulk of luminal-derived FA in the enterocyte
cytoplasm (46). These studies depict L-FABP as an intestinal sink for FA, which may contribute
to the directionality of lipid transport. Indeed, while there is no fat malasbsorption, Lfabp-/-
mice
exhibit delayed TG appearance in the plasma following an oral fat challenge, and this is
accompanied by increased lipid accumulation in the proximal intestine (47). Intestinal fatty acid
binding protein (I-FABP) in expressed specifically in the intestine, but has been deemed non-
essential for dietary lipid absorption (48). However, I-FABP does appear to play a role in
targeting dietary-derived FA to TG synthesis as opposed to -oxidation, while the directing of
dietary-derived MAG to TG synthesis is done by L-FABP (49). Despite their striking structural
similarities, the functions of L-FABP and I-FABP do not appear to have overlapping functional
role, as the other protein is not grossly upregulated when one is knocked out (50; 51).
1.2.2 Chylomicron assembly and secretion
Assembling a very large particle (up to 500 nm in diameter) comprised chiefly of
hydrophobic molecules in the aqueous environment of the cell requires a complex and
coordinated array of activities. To summarize, the chylomicron assembly process begins with
FA uptake at the apical membrane, which needs to be reesterified and targeted to the secretory
pathway. The lipids are then assembled with the nascent aggregate-prone apoB48 polypeptide,

6
followed by further expansion of the lipid core. To continue along the secretory pathway, the
large cargo necessitates specialized export machinery to exit the ER, followed by further
maturation before leaving the enterocyte at the basolateral membrane. Details of these processes
are outlined below.
Once FA is made available inside the enterocyte by diffusion and/or one or more of the
transporters described above, it is assimilated into TG at the ER membrane. In the intestine,
most TG synthesis follows the acyl-CoA:monoacylglycerol acyltransferase (MGAT)/acyl-
CoA:diacylglycerol acyltransferase (DGAT) pathway, where fatty acyl-CoA is sequentially
added to MAG to form diacylglycerol (DAG), then to DAG for the final TG product. The acyl
CoA likely arises from the activity of FATP4 at the ER, as discussed above. MGAT2
predominates in the proximal small intestine (52), while MGAT3 is more abundant in the distal
small intestine (53). These two isoforms appear to be capable of compensating for each other’s’
activity, for while MGAT2-deficient mice do not exhibit gross fat malabsorption, the kinetics of
absorption are delayed distally (54). Both DGAT1 and DGAT2 are expressed in the small
intestine, but currently data is available only for DGAT1. While DGAT1 accounts for 89% of
TG synthesis initiated from MAG in the intestinal mucosa (55), it is dispensable for esterifying
dietary FA with DAG to form TG (56). However, DGAT1 does play an important role in dietary
nutrient handling, for intestine-specific DGAT1 expression in Dgat-/-
mice was sufficient to
confer susceptibility to high fat diet-induced obesity (57). The TG synthesized by DGAT1 may
be more preferentially targeted for chylomicron assembly, as Dgat1-/-
mice exhibit large neutral
lipid droplets that persist in the enterocyte following feeding (56). Cholesterol is also esterified
at the ER, and the action of acyl CoA:cholesterol acyltransferase 2 (ACAT2) is essential to
intestinal cholesterol absorption (58).
Newly-synthesized lipids are required to cotranslationally lipidate the nascent apoB48
polypeptide, the large, amphipathic, nonexchangeable structural protein of the chylomicron. The
apoB48 arises from posttranscriptional editing of apoB mRNA by apoB mRNA editing enzyme
catalytic polypeptide 1, a cytidine deaminase that introduces a premature stop codon to yield a
truncated apoB (59). ApoB gene transcription is generally considered to be constitutive
(reviewed in (60)), but transforming growth factor (TGF-) can activate ApoB gene
transcription through SMADs in the human epithelial colorectal adenocarcinoma cell line, Caco-

7
2 (61). Also, recently there is evidence of a p53 response element in the ApoB promoter (62) and
interleukin-6 (IL-6) can increase apoB mRNA levels (63). The translation of apoB mRNA is
also subject to regulation in the liver, given the highly structured 5’ and 3’ untranslated regions
(64-67), but such studies have not yet been performed in the intestine.
Currently, a three-step model is proposed for chylomicron assembly (68). The first step
is the formation of a “primordial lipoprotein” which begins with the recruitment of phospholipids
to the N-terminal domain of apoB48 both intrinsically and by microsomal triglyceride transfer
protein (MTP) (69). Newly-synthesized apoB48 initially resides in the smooth ER membrane,
but moves to the lumen in an MTP-dependent step (70). At this point, the phospholipid-rich
apoB48 particle is still TG-poor and high density lipoprotein (HDL)-sized. In the liver,
insufficient lipids for continued lipidation would result in the apoB polypeptide being targeted
for degradation and prevent VLDL secretion. However, there is no strong evidence for apoB48
degradation in the intestine. There is evidence in models of intestinal lipoprotein oversecretion,
such as the fructose-fed hamster or Psammomys obesus, of attenuated degradation of newly-
synthesized apoB48 (71; 72). While inhibiting proteasomal degradation with N-carbobenzoxyl-
L-leucinyl-L-leucinyl-L-norleucinal (MG132) raised intracellular levels of newly-synthesized
apoB48, it did little to drive apoB48-containing particle secretion in chow-fed hamster
enterocytes (71). In the Caco-2 cell model, apoB is normally not degraded unless an MTP
inhibitor is added and only minimal levels of intracellular apoB are found conjugated to ubiquitin
(73). Imaging of this cell model has suggested that immature apoB48-containing lipoproteins
pass through the trans-Golgi and stored in an apical compartment, from which they proceed
basolaterally upon exposure to luminal lipid micelles (74). When lipid micelles are included ex
vivo, apoB48 can be recovered quantitatively from primary murine enterocytes, suggesting that
apoB48 degradation is not an active pathway in the postprandial condition (75). Moreover,
apoB48-containing particles exhibit far greater heterogeneity in size than apoB100 particles,
suggesting that less lipid is permissive to the secretion of the truncated form of apoB (75). After
all, under fasting conditions, the enterocyte can take up circulating albumin-bound nonesterified
FA, which will be referred to henceforth as free fatty acids (FFA), to form and secrete small
HDL-sized particles (76). The second step is the formation of lipid droplets, comprised of
neutral lipids synthesized by the enzymes outlined above. The third step is core expansion, with

8
MTP facilitating the incorporation of lipids into the nascent lipoprotein. Apolipoprotein AIV
(apoAIV) is thought to be involved in this step (77). Core expansion occurs in the smooth ER
lumen and, probably by virtue of the preformed lipid droplets, is a rapid step that allows the
apoB48 particle to acquire the bulk of its TG (78). This step also allows each apoB48
polypeptide to transport ten-fold more TG in the postprandial state compared to fasting (79).
By the time it is ready to exit the ER, the apoB48-containing lipoprotein has almost
attained the size of a mature chylomicron. To shuttle this immensely-sized cargo between the
ER and Golgi, a specialized COPII vesicle called the prechylomicron transport vesicle (PCTV)
has been identified. Notably, the PCTVs also contain the GTPase Sar1, which is necessary for
fusion with the Golgi, thus providing the mechanistic explanation for chylomicron retention
disease, a disorder of fat absorption characterized by malnutrition and hypocholesterolemia (80).
Other COPII proteins Sec23 and more importantly Sec24 are necessary for docking onto the cis-
Golgi (81), followed by fusion that is mediated by a v-N-ethylmaleimide-sensitive factor
attachment protein receptor (SNARE) complex comprised of vesicle associated membrane
protein 7 (VAMP7), syntaxin 5, Bet1 and vti1a (82). Interestingly, PCTV budding involves
factors that are involved in fatty acid uptake at the apical membrane, L-FABP and CD36 (83;
84), thus highlighting the mobility in the subcellular localization of these proteins. Notably, I-
FABP was not observed to have the same PCTV-budding activity as L-FABP (84). There is
evidence that PCTV budding can be regulated. Protein kinase C (PKC), which can be
activated by a variety of lipid species including ceramide, phosphorylates a 9 kDa ER-associated
protein to drive PCTV budding (85). In the Golgi, chylomicron maturation is completed with the
inclusion of the exchangeable apolipoproteins, apolipoprotein AI (apoAI), apoliprotein CII
(apoCII), and apolipoprotein CIII (apoCIII), followed by secretion as a particle encased in a
phospholipid monolayer and with a density less than 1.006 g/mL (86). Dietary lipid uptake and
chylomicron assembly and secretion are summarized in Figure 1.1.

9
Figure 1-1. Dietary lipid absorption and chylomicron assembly in the enterocyte.
Following intraluminal hydrolysis, FA or MAG is made available at the BBM by the action of
CD36, followed possibly by desorption from the membrane bilayer facilitated by L-FABP/I-
FABP. SR-BI appears to play a role in cholesterol uptake at the BBM but the rate-limiting step
involves the endocytosis of cholesterol with NPC1L1. SREBP-1c action induces expression of
ACC and FAS, which produce FA that can also be incorporated into chylomicron particle. FA is
activated to fatty acyl-CoA at the ER by FATP4, and then subsequently used to esterify MAG by
MGAT2. TG synthesis culminates with the transfer of a fatty acyl-CoA to DAG. Cholesterol
transported into the cell by the action of SR-BI and NPC1L1 and destined for secretion is also
esterified through the action of ACAT2 at the ER. Meanwhile, the nascent apoB48 polypeptide

10
acquires lipid cotranslationally with the help of MTP in the ER lumen. The primordial,
phospholipid-rich apoB48 particle continues to acquire additional TG and cholesteryl ester from
lipid droplets, with MTP facilitating lipid transfer between these unilamellar entities. The
apoB48 particle also acquires apoAIV in the ER, which aids in the lipoprotein’s core expansion.
By the time the apoB48 particle is ready to exit the ER, it is nearly the size of a mature
chylomicron, so a specialized transport vesicle called the PCTV is necessary to accommodate
this immensely-sized cargo. PCTV budding from the ER is a process that is L-FABP- and
CD36-dependent, while COPII proteins and Sar1 GTPase are necessary for docking and fusion at
the cis-Golgi. The apoB48 particle undergoes the final stages of maturation once the
exchangeable apolipoproteins are added as it progresses through the Golgi.

11
1.2.3 Naturally occurring mutation in fatty acid transporters
Mutations resulting in either defective CD36 expression or function are quite common in
the human population. Most reported deletions and insertions result in frameshift mutations that
reveal a premature stop codon or exon-skipping, often producing a truncated or unstable product,
thereby creating a phenotype of CD36 deficiency (87). The C268T substitution occurs with over
50% frequency among mutated Cd36 alleles in the Japanese population (88). Replacement of
Pro90 with Ser leads to an 81 kDa protein product indicative of incomplete posttranslational
glycosylation, and this mutated CD36 is not efficiently expressed on monocyte cell surface but
rather degraded in the cytoplasm (89). The Pro90Ser mutation was associated with higher FFA
levels (90), but not the insulin resistance, elevated fasting TG, and greater postprandial apoB48
levels observed in older individuals with rare Cd36 mutations (91; 92). Similar abnormalities
were observed in CD36-null mice (93). Another haplotype, 30294G>C, was also linked to FFA
elevation in non-diabetic Italians (94), but the effect of this polymorphism in the 3’ UTR has not
been fully characterized. Given the ubiquitous expression and ligand-promiscuity of CD36, the
lipid and lipoprotein abnormalities in these individuals could be a result of CD36 deficiency in
multiple tissues. For example, subjects with two mutated alleles have compromised myocardial
long chain FA uptake (95), and this defective uptake in the heart and peripheral tissues may
contribute to the greater circulating FFA levels. Consequently, this FFA elevation inhibits
lipoprotein lipase action (discussed in ‘Chylomicron catabolism’) , thereby contributing to the
persistence of chylomicron remnants (96). Moreover, CD36-mediated LCFA uptake in
myocytes helps maintain insulin sensitivity (97), which may provide an explanation for the
insulin resistance observed in CD36-deficient individuals. Lymphatic cannulations revealed
intestinal overproduction of apoB48-containing lipoproteins in CD36-deficient mice which may
explain the lipoprotein abnormalities (98), which is in disagreement with other published studies
in these mice. However, intestinal insulin resistance may be contributing to the chylomicron
overproduction (discussed later in ‘Hormonal regulation of chylomicron secretion’), as genes
involved in de novo lipogenesis were upregulated (98). A nonsense mutation in Cd36 that
presented with autosomal dominant diabetes was identified in the French population (99). The
mechanism for the diabetes is unresolved, as the truncated product, while capable of trafficking
to the cell surface, was incompetent in binding modified LDL (99) but still included the putative

12
FA binding domain (100). However, the loss of the second transmembrane domain would have
prevented CD36 from assuming the hairpin conformation and possibly affected the folding of the
FA binding domain.
Metabolic abnormalities have also been noted for naturally occurring polymorphisms in
L-FABP. A mutation in the highly conserved Thr94 position was found in the French Canadian
population, in which the Ala94 allele was found to occur with 32.3% frequency (101). Even
after adjustments for age, body mass index, smoking habit, menopausal status, and ApoE
genotype, the Thr94Ala mutation was significantly associated with higher fasting TG and LDL-
cholesterol in women (102). Thr94 is in the FA-binding -barrel of L-FABP, but it comprises a
-strand involved in hydrogen bonds atypical of a -barrel (45). Chang liver cells transfected
with the Thr94Ala variant have been noted to transport less FA (103), and McA-RH7777 rat
liver cells overexpressing wild type L-FABP secrete more apoB100 (104), so the etiology of the
fasting hyperlipidemia observed in Thr94Ala individuals remains unclear. However, Ala94
carriers who have a greater percentage of caloric intake from fat have lower plasma apoB levels
than Thr94 homozygotes (101), so the gene-diet interaction in this particular context may be due
to compromised FA transport in the intestine. Carriers of the Ala94 allele were also more likely
to have plasma TG above the therapeutic target following treatment with the PPAR agonist
fenofibrate (105). While the 5’ flanking region of the Lfabp gene contains a PPAR response
element, intestinal L-FABP expression appears to be regulated specifically by PPAR (106), so
the fenofibrate response of Ala94 carriers was likely attributable to aberrant lipid metabolism
occurring at the hepatic level.
Though the absence of IFABP in mice does not affect their ability to amass lipids in their
tissues, a naturally occurring single nucleotide polymorphism in IFABP is accompanied with
observable metabolic phenotypes in humans. A polymorphism in codon 54 was first discovered
in Pima Indians in an attempt to identify the genetic factor determining insulin action in the
exceptionally type 2 diabetes-prone population (107), and then later detected in many other
ethnicities. The IFABP Ala54Thr missense mutation confers two-fold greater affinity for long
chain FA (107), possibly due to stabilization of noncovalent interactions in the lipid entry portal
which may prohibit FA dissociation from the protein (108). Non obese and non-diabetic male
Japanese-American carriers of the Thr54 alleles have higher fasting TG (109). Type 2 diabetic

13
Caucasian male Thr54 homozygotes have fasting and postprandial hypertriglyceridemia, with the
postprandial TG attributed to elevated chylomicron levels (110). In a French-Canadian
paediatric population, Thr54 homozygotes had a steeper increase in fasting apoB with rising TG
levels, an interaction that was not even associated with MTP genotype (111). Consistent with
these observations in humans, Caco-2 cells transfected with this mutant IFABP have enhanced
long chain FA uptake and TG secretion (112). The Thr54 allele may thus render an individual
to be more sensitive to the FA content of their diet. Young healthy normolipidemic subjects
more readily exhibited signs of insulin resistance following 28 days of a saturated fat-enriched
diet than Ala54 homozygotes (113). Pima Indian Thr54 carriers mount a greater insulin response
to a high fat meal yet have higher postprandial FFA concentrations suggesting insulin resistance
(114). However, a study in Korean men showed that neither heterozygotes nor homozygotes of
the Thr54 allele had different peak serum 3H-activity following ingestion of
3H-labelled oleic
acid (115). Likewise, the Thr54Ala polymorphism did not affect the postprandial mean TG
following a fat tolerance test in males from the European Atherosclerosis Study (116).
Therefore, rather than being essential to intestinal lipid absorption, IFABP may be important in
adapting to dietary stresses.
Naturally occurring FATP4 polymorphisms are rare in the human population, considering
that even missense mutations in the ER localization signal, AMP binding domain, or the
noncatalytic C-terminal domain all result in icthyosis prematurity syndrome, which is
characterized by premature birth, neonatal asphyxia, and lifelong skin abnormalities (117). The
defective barrier function observed in infants with these FATP4 polymorphisms is reminiscent of
the phenotype of Fatp4-/-
mice (118). A missense mutation in exon 3 of Fatp4 was found to
occur in Swedish men with an allele frequence of 0.05. The Gly209Ser polymorphism resulted
in lower body mass index, plasma TG, VLDL-TG, insulin, HOMA index, and systolic blood
pressure. And oral fat tolerance test revealed that Ser209 carriers and homozygotes also tended
to have lower plasma TG, chylomicron apoB48, VLDL1-apoB100, VLDL2-apoB100 and FFA
concentrations in the postprandial state, which suggests FATP4 is involved in intestinal lipid
handling (119). A structural model of FATP4 suggested the variable residue 209 is flanked by
proline residues and located within an exposed hydrophobic loop, implicating this region in
protein-protein interactions, although the interacting partner was not identified (119). While

14
FATP1 has not been functionally characterized in the small intestine, there is evidence that a
common intronic single nucleotide polymorphism may affect postprandial lipid metabolism. 50-
year-old Swedish male homozygotes of the A/A genotype in intron 8 have significantly higher
plasma TG concentrations in response to an oral fat tolerance test, with the TG appearing to
reside in the chylomicron and larger VLDL1 fractions, concomitant with a failure to suppress
FFA levels in the postprandial state (120). However, with such a postprandial lipid profile and
the high FATP1 expression in adipose tissue (35), it was likely the postprandial dyslipidemia
was due to defective uptake of FA released by LpL-mediated hydrolysis of circulating TRL.
This FA would have inhibited further LpL action and/or induced hepatic VLDL production.
1.2.4 Insulin action and insulin resistance
Since the discovery of insulin 90 years ago, it was clear the pancreas-derived hormone
works to decrease blood glucose while having other anabolic effects (121). The actions of
insulin proceed via its binding and the autophosphorylation of the insulin receptor on tyrosine
residues, which then recruits insulin receptor substrates (IRS) that also become phosphorylated
on tyrosine residues. The phosphotyrosine residues on IRS-1/2 recruit phosphatidylinositol-3-
kinase (PI3-K) via the p85 regulatory subunit. The p110 catalytic subunit of PI3-K
phosphorylates phosphatidylinositol (4,5)-disphosphate, which consequently recruits Akt/protein
kinase B to the plasma membrane for phosphorylation by phosphoinositide-dependent kinase-1
(PDK1). The insulin receptor can also activate the mitogen-activated protein kinase/ERK kinase
(MEK)/extracellular related kinase (ERK) pathway by recruiting the adaptor protein Src
homology 2 domain-containing protein (Shc) and protein growth factor receptor-bound protein 2
(Grb2), which brings the Ras GTPase activator Son of Sevenless (SOS) to the membrane (122).
Through a combination of glucose transporter 4 (GLUT4)-mediated glucose uptake in muscle
and fat (123), suppressed gluconeogenesis in the liver (124), and stimulated glycogen storage
(124), insulin counteracts postprandial hyperglycemia. Insulin also inhibits the secretion of its
counterpart in glucose homeostasis, glucagon, from pancreatic -cells (125). While the effects
on glycemia are among its most immediately appreciable effects, insulin is integral to signalling
nutrient abundance in the postprandial state and is thus an important regulator of lipid
metabolism. In fat tissue, insulin stimulates the assimilation of carbohydrate-derived substrates
into TG stores and inhibits lipolysis (126). In adipocytes, lipid droplet-associated proteins such

15
as perilipin A (PLINA) and hydrolases such as hormone sensitive lipase (HSL) are among the
substrates for protein kinase A (PKA). The phosphorylation of PLINA recruits HSL to lipid
droplets, and the phosphorylated HSL associates with FABP4 for activation. Together with the
activities of adipose triglyceride lipase (ATGL) and monoacylglycerol lipase (MGL), the
activated HSL releases nonesterified FA (FFA) and glycerol from adipose storage depots into the
circulation. Insulin signalling suppresses FFA release in the postprandial state by activating
phosphodiesterase 3B to degrade cyclic adenosine monophosphate (cAMP) and remove the
stimulatory signal for PKA (127). Insulin also promotes FA synthesis through the induction of
lipogenic genes by activating sterol regulatory element binding protein-1c (SREBP-1c) in an
Akt2-dependent manner (128). The sterol regulatory element is found in the promoters of fatty
acid synthase (FAS) and acetyl-CoA carboxylase (ACC) (129). Moreover, insulin’s
lipogenic effects can be achieved more acutely by the phosphorylation of ACC (130).
Insulin is also a strong lipogenic signal in the liver. The liver-specific transcript insulin-
induced gene 2a (INSIG-2a) is an ER-localized protein present in limiting amounts that
sequesters SREBP cleavage-activating protein (SCAP). In an Akt2-dependent manner, insulin
suppresses Insig2a expression, thereby releasing SCAP from the ER and allowing it to escort
SREBP-1c to the Golgi for proteolytic processing and activation (131). The products of de novo
lipogenesis are then used to lipidate VLDL (132). Despite the stimulation of lipogenesis, insulin
actually reduces hepatic VLDL secretion (133-136). While the lowering effect is partly
attributable to diminished FFA supply due to suppressed lipolysis in adipose tissue, insulin
appears to have effects on apoB metabolism itself (137). Insulin may decrease VLDL secretion
by excluding forkhead box O1 (FoxO1) from the nucleus to inhibit Mttp transcription (138),
enhancing apoB degradation (139), and inhibiting ApoB mRNA translation (64), all in a PI3-K-
dependent manner.
The etiology of insulin resistance is under debate, but there is evidence that inflammatory
signalling that arise from tumour necrosis factor- (TNF-) action through its receptor TNFR
may impair insulin signalling through the action of c-Jun N-terminal kinase (JNK) which
negatively affects IRS-1 activity. Also, protein tyrosine phosphatase-1B (PTP-1B) attenuates
insulin action by dephosphorylating the insulin receptor tyrosine residues (122). In insulin
resistance, there is impaired glucose tolerance, hyperinsulinemia and hypertriglyceridemia. The

16
paradox in insulin resistance is that while glucose uptake and suppression of gluconeogenesis is
blunted, the lipogenic effect of insulin proceeds unabated (140). Coupling this selective insulin
resistance in lipoprotein-producing organs with defective insulin action in peripheral issues sets
the state for dyslipidemia. Both increased FA flux due to dysregulated lipolysis in adipose
depots and increased de novo lipogenesis provide more substrate for VLDL lipidation (141; 142).
Meanwhile in the liver, other steps regulating TRL secretion become refractory to insulin action
such as MTP expression (143; 144) and apoB degradation (145; 146). VLDL overproduction
strains an overburdened lipolytic system, in which the increased FFA inhibits lipoprotein lipase
(LpL) and prevent TRL clearance (147). In the postprandial state, the problem is amplified with
the influx of chylomicrons competing for common lipolytic pathways (further discussed in
‘Chylomicron catabolism’). However, the nature of dysregulated lipoprotein overproduction in
the intestine in insulin resistant states is less well understood.
1.2.5 Hormonal regulation of chylomicron secretion
The regulation of intestinal lipoprotein production by endocrine signals is a relatively
new notion, as apoB48 secretion was traditionally regarded as a constitutive process. Initially,
there were multiple human studies to suggest aberrant postprandial intestinal lipoprotein
metabolism in insulin resistance (148-151). Indeed, a kinetic study indicated that
hyperinsulinemic insulin resistant patients have increased apoB48 production rates, as opposed
to defective TG-rich lipoprotein (TRL) clearance (152). Prior to this observation, evidence from
multiple rodent models emerged to indicate that insulin resistance is associated with TRL
overproduction from the intestine and have provided clues regarding the mechanism. The
fructose-fed hamster, a model of diet-induced insulin resistance and hepatic VLDL
overproduction (145), secretes not only more apoB48-containing lipoproteins, but larger particles
as well (71; 145). The apoB48 overproduction can be partially normalized with the insulin-
sensitizing agent rosiglitazone, which restored intestinal MTP expression to the levels of chow-
fed hamsters (153). The enterocyte is itself sensitive to insulin, and fructose-feeding blunts
insulin signaling as evidenced by decreased IRS-1 tyrosine phosphorylation, an increase in PTB-
1B expression, and attenuated Akt response to insulin while maintaining mitogen-activated
protein kinase (MAPK) signalling (154). The consequence is increased SREBP-1c activation,
which may explain the increased lipogenesis observed by Haidari et al (71). A summary of the

17
signaling derangements that could lead to apoB48 oversecretion in insulin-resistant enterocytes
is depicted in Figure 1-2. Fructose-feeding is also associated with upregulated DGAT2
expression to accompany the increased intestinal lipid content (155) and the enterocyte
endoplasmic reticulum appears to be modified, as PCTV’s isolated from fructose-fed hamster
microsomes had increased chaperone content to suggest possible ER stress (156).

18
Figure 1-2. Insulin signalling pathway depicted with perturbations known to occur in the
enterocyte during insulin resistance.
Certain components of the system are upregulated () or downregulated () with respect to
either mass or phosphorylation, as determined in the fructose fed hamster model. TNF-
signalling plays a role in inducing intestinal insulin resistance, and appears to be mediated by
both TNF- receptor 1 (TNFR1) and TNF- receptor 2 (TNFR2). Points at which TNF-
interrupts insulin action are shown, with a putative interaction between inflammatory signalling
(via JNK) and insulin signalling (on IRS-1 phosphorylation) represented by a dotted line.

19
Another rodent model of insulin resistance and type 2 diabetes is the sand rat
Psamommys obesus, which also oversecretes intestinally-derived apoB48-containing
lipoproteins. Aside from amplified apoB48 biogenesis and de novo lipogenesis, chylomicron
assembly is also modulated with heightened MGAT and DGAT activities, along with increased
L-FABP expression (72). ApoB48 oversecretion has also been observed in the leptin receptor-
deficient Zucker obese fa/fa rats, a phenomenon that was attributed to upregulated intestinal
MTP expression (157). Another insulin-resistant rodent, the JCR:LA-cp rat, has been
characterized with apoB48-containing TRL oversecretion (158). While the Zucker obese fa/fa
and JCR:LA-cp rats are of interest primarily for their insulin resistance, the defective leptin
action also complicates these models, as leptin itself affects intestinal lipid metabolim. For
example, leptin can acutely reduce intestinal apoAIV mRNA (159), thereby implicating a role
for the hormone in regulating chylomicron size, as apoAIV has been shown to facilitate
chylomicron core expansion (77). Moreover, db/db mice with a defective leptin receptor have
inefficient intestinal lipid secretion due to decreased MTP expression, in a manner that involves
a gut-intrinsic melanocortin pathway (160). The effect of diet-induced insulin resistance on
intestinal lipoprotein secretion in mice has not been characterized in great detail. The B6D2F1
mouse strain, which becomes diabetic within three weeks on a high fat diet (HFD), absorb and
secrete dietary fat as chylomicrons at a faster rate, which is then more efficiently cleared by
increased intestinal production of apoCII (161). Enterocytes isolated from HFD-fed C57BL/6
mice secrete newly-synthesized apoB48 at double the rate of enterocytes from chow-fed mice.
However, it cannot be discerned if this difference is attributed to the insulin resistance induced
by the diet, or the fat content of the diet itself, as bile salt lipid micelle supplementation can
ameliorate the differences in apoB48-containing particle secretion (75).
While the above mentioned studies look at a chronic deficiency in insulin action, there is
also evidence that insulin can acutely modulate intestinal lipoprotein secretion. Acute
hyperinsulinism delayed the postprandial rise in apoB48 by two hours in healthy men (162). The
ability of insulin to lower apoB48 output is partly attributable to its suppression of circulating
FFA (163), as an acute elevation in FFA is enough to significantly increase TRL-apoB48
secretion (164). Another hormone that can acutely modulate intestinal apoB48 metabolism
through the insulin pathway is the inflammatory cytokine TNF-. In hamsters, a 4 h infusion

20
suppressed intestinal insulin signalling while stimulating p38, extracellular related kinase 1/2
(ERK1/2), and c-jun N-terminal kinase JNK signalling, and increasing TRL-apoB48 secretion,
likely by raising MTP expression (165). There is evidence that other mitogenic signals can
regulate intestinal lipoprotein metabolism. Epidermal growth factor (EGF) has been shown to
stimulate apoB48 biogenesis in human fetal intestine (166). TGF- has also been shown to
augment apoB secretion in the Caco-2 cell model (167), but in addition to being an intestinal cell
line that secretes both apoB100 and apoB48 isoforms, observations in this model have been
inconsistent with the aforementioned studies in primary cultures. For example, TNF- was
noted to decrease apoB-containing lipoprotein secretion from Caco-2 cells (168), while the
aforementioned study on primary hamster enterocytes suggests that TNF- is a stimulatory
signal for secretion (165). Therefore, our current understanding of how endocrine factors can
regulate intestinal lipid metabolism is still in its infancy.
1.2.6 Other fates of dietary lipid in the intestine
Although most of the ingested lipid is packaged into chylomicrons for secretion, some of
the lipids are catabolized or even stored. Generally speaking, -oxidation occurs at a low rate in
enterocytes for in both fasting and fed states. Glutamine constitutes the primary fuel for
enterocytes, where it is catabolized to CO2 and alanine (169; 170). During food deprivation,
approximately 31% of FA taken up at the serosal side is oxidized in enterocytes, but this figure
drops to 12% for luminally-derived FA. The fed intestine oxidizes FA at half the rate, with only
12% of circulating FA and 5% of dietary FA being utilized for fuel (49). However, FA oxidation
can be induced in fat-enriched diets, perhaps as an early adaptation to increased caloric intake.
Polyunsaturated FA-enriched diet is associated with increases in intestinal -oxidation (171), and
the guts of both the obesity-resistant A/J and the obesity-prone C57BL/6J mouse strains have
upregulated -oxidation and carnitine palmitoyl transferase-1 (CPT-1) activity after only two
weeks on a HFD (172). While FA play only a minor role is providing energy for the enterocyte,
there is evidence that the metabolites of FA oxidation plays an important role in signalling
satiety. Inhibiting intestinal long-chain acyl-CoA dehydrogenase with an intrajejunal infusion of
mercaptoacetate stimulated food intake in sham-operated rats, but not in rats with
subdiaphragmatic vagal deafferentation, suggesting that vagal afferents are important in the
sensing of signals elicited by enterocytic FA oxidation (173).

21
Considering the enterocyte’s short lifespan, it seemed unlikely that this cell type would
store lipids for later use. However, it has been observed in humans that 13% of chylomicron TG
is derived from FA ingested the previous evening, indicating that dietary lipid continues to be
used for intestinal TG synthesis 18 h after ingestion (174). Glucose ingestion five hours after an
oral fat load is followed by a peak in chylomicron-TG and apoB48 45 min later, along with less
lipid staining in the jejunal mucosa, suggesting the lipoproteins were assembled from stored
lipids in the enterocytes (175). Indeed, imaging studies have shown that following lipid
ingestion, there is a cytoplasmic pool of lipid coincident with the lipid droplet protein tail-
interacting protein of 47 kDa (TIP47) that forms in enterocytes. The diameter of these
cytoplasmic lipid droplets increase up to 3 h following an oral gavage of oil, and then diminish in
size until the pool is depleted by 12 h (176). However, the acyltransferase responsible for
assimilating the TG in this temporary storage pool, or the hydrolase that mobilizes these stored
lipids for secretion, have yet to be identified.
1.2.7 Chylomicron catabolism
Chylomicrons are involved in ‘forward’ lipid transport – the delivery of lipids from the
intestine to peripheral tissues. After traversing the lympathic system, chylomicrons enter the
circulation where some remodelling occurs to allow these particles to deposit FA in peripheral
tissues. Upon reaching the circulation, apoAI and apoAIV along with some phospholipids are
rapidly transferred from nascent chylomicrons to circulating HDL particles (177). In exchange,
chylomicrons acquire apoCII and apoE from HDL, which are important for its catabolism and
removal (178; 179). The apoCII molecules on the chylomicron surface are necessary to activate
LpL bound to the vascular endothelial cell surface by heparin sulphate proteoglycans. The
enzymatic action of LpL is to hydrolyze 1(3)-ester linkages which liberates FA and 2-
monoacylglycerol from the TG transported in the chylomicron core (180). The heart is the main
site of hydrolysis and uptake of the dietary lipid transported by chylomicrons. In fact, whole
body chylomicron-TG clearance proceeds at only 40% of the wild type rate in heart-specific LpL
knockout mice (181). Chylomicrons therefore are critical to providing the predominant energy
substrate to the heart, where LpL deficiency results in cardiac dysfunction (182). Insulin raises
and depresses LpL activity in adipose tissue and skeletal muscle, respectively, thereby directing
chylomicron-TG away from muscle tissues and towards storage depots in the postprandial state

22
(183). However, the FA liberated by LpL is not efficiently taken up by local tissues, and about
36% escape into the systemic circulation. This “spillover” is consequently an important
contributor to postprandial FFA levels (184). The efficient lipolysis of chylomicron-TG is also
important to vascular health, as evidenced by glycosylphosphatidylinositol-anchored high-
density lipoprotein binding-protein 1 (GPIHBP1)-deficient mice. GPIHBP1 is expressed on
vascular endothelial cells and binds chylomicrons and LpL to mediate LpL’s hydrolytic activity.
GPIHBP1 deficiency resulted in severe chylomicronemia even on a low-fat diet (185), and
GPIHBP1 knockout mice spontaneously developed lipid-rich atherosclerotic lesions on a chow
diet (186). A chylomicron particle remains in circulation until 80% of its TG content has been
catabolized peripherally. The remaining chylomicron remnant still retains its apoB48
polypeptide, almost all of its original cholesteryl ester content, and surface apoE and is cleared
by the liver. Chylomicron remnants are endocytosed either by the low density lipoprotein
receptor (LDLR) or the low density lipoprotein receptor-related protein 1 (LRP1) via recognition
of apoE (187; 188). Notably, these receptors are also involved in the clearance of apoB100-
containing hepatic-derived lipoproteins, so chylomicron remnants compete for these receptors in
the postprandial state, which ultimately has implications for dyslipidemia (10). There is also
evidence of impaired hepatic chylomicron remnant clearance in LRP5-deficient mice fed a high
fat diet (189).

23
1.3 Glucagon-Like Peptides
1.3.1 Glucagon-like peptide structure
Glucagon-like peptide-1 (GLP-1) and glucagon-like peptide-2 (GLP-2) arise from
posttranslational proteolytic processing of the preproglucagon polypeptide by prohormone
convertase 1/3 (PC1/3) in intestinal enteroendocrine L-cells and in the brainstem. GLP-1 and
GLP-2 occur sequentially in preproglucagon, preceded by glicentin-related polypeptide and
glucagon with short intervening peptide sequences (190). The differential processing that occurs
in the gut and pancreas to produce different peptides from the same proglucagon polypeptide is
illustrated in Figure 1-3. Although PC1/3 cleaves at pairs of basic residues, GLP-1 is N-
terminally truncated, with the biologically active peptides being GLP-1(7-37) and more abundant
GLP-1(7-36) amide (191). GLP-2 is similarly-sized at 33 amino acids.
Human GLP-1 peptide sequence: HAEGTFTSDVSSYLEGGAAKEFIAWLVKGRG
Human GLP-2 peptide sequence: HADGSFSDEMNTILDNLAARDFINWLIQTKITD
The sequences of both GLPs are highly conserved across mammalian species, with the sequence
of GLP-1 being identical between humans, mice, rats, and other multiple other species. Both
GLPs have an alanine at the N-terminal penultimate position, rendering them susceptible to
degradation by dipeptidyl peptidase-4 (DPP-4), also known as CD26 (192). Exendin-4 which
was originally isolated from the venom of Heloderma suspectum has 53% amino acid sequence
identity with GLP-1. Exendin-4 also agonizes the GLP-1 receptor (GLP-1R) but is notable for
the fact that it is DPP-4-resistant (193). Therefore, exendin-4 serves as a peptide that exacts
similar functions as GLP-1, but in a more sustained manner. While exendin-4 is a GLP-1R
agonist, exendin9-39 is the antagonist (194). As for GLP-2, replacing the penultimate alanine
with glycine to make Gly2-GLP-2 is enough to confer resistance to DPP-4 (195). The N-
terminally truncated GLP-2, GLP-23-33
has been identified as a GLP-2 receptor (GLP-2R)
antagonist (196), but the peptide also exhibits weak agonist qualities at higher concentrations
(197). Although both have high amino acid identity with glucagon, hence their names, these
sister peptides exhibit divergent physiological functions, as will be further discussed below.

24
Figure 1-3. Differential proglucagon polypeptide processing in pancreatic alpha-cells and
intestinal enteroendocrine L-cells
The proglucagon polypeptide is a Type IV precursor protein that is processed at basic doublets
by proprotein convertases, while the C-terminal basic residues of the cleaved fragments are
removed by carboxypeptidase E (CpE). The pancreatic -cell-specific prohormone convertase 2
(PC2) cleaves to generate glucagon and the major proglucagon fragment. In ileal
enteroendocrine L-cells, prohormone covertase 1 (PC1, also known as PC3) simultaneously
cleaves the proglucagon polypeptide to generate GLP-1 and GLP-2, along with oxyntomodulin.
GLP-1 can exist in a glycine-extended form or amidated by peptidylglycine alpha-amidating
monooxygenase.

25
1.3.2 Intestinal GLP secretion
Because both GLPs are simultaneously processed from the same proglucagon
polypeptide, GLP-1 secretion is always mirrored by that of GLP-2. Therefore, for purposes of
discussion, it is assumed that documented observations regarding GLP-1 secretion would also
apply similarly to GLP-2. As postprandial hormones, the GLPs have low circulating levels
during fasting, with the low picomolar range for GLP-1 (198; 199), and values from 39 pM (200)
to over 200 pg/mL (201) for GLP-2 in healthy humans. Upon meal ingestion, GLPs are secreted
in a biphasic manner, with the first peak occurring quickly at about 15 min after meal ingestion,
followed by a larger and more prolonged elevation for the next 2 hours (198; 200; 202). The
first phase may arise from neural signals mediated by the vagus (203; 204) or cholecystokinin
and GIP originating from the proximal intestine (205). The second phase of secretion likely
results from nutrients directly interacting with enteroendocrine L-cells of the distal intestine.
The magnitude of the increase is dependent on both caloric content and nutrient composition
(206; 207). While the GLPs are found and cleared extensively in the circulation, there is some
evidence that the L-cells also secrete the peptides into the lymphatic system, where there is
protection from DPP-4-mediated degradation and the opportunity to interact with enteric neurons
(208).
It has been noted that the ingestion of fat is a stimulus for more prolonged and sustained
GLP secretion than carbohydrate or protein (199; 202). While sugars are very efficiently
absorbed in the proximal intestine, FA can persist more distally in the lumen of the
gastrointestinal tract, allowing them to be sensed by the enteroendocrine L cells in the ileum,
especially 60 min after ingestion during the second, more extended phase of GLP secretion
(209). Multiple studies assessed the role of FA saturation on GLP secretion, with
monounsaturated fat being more potent stimulus than saturated FA (210-212). A set of orphan G
protein-coupled receptors have been implicated in mediating FA-induced GLP-1 secretion. Two
Gq-coupled receptors, G protein-coupled receptor 40 (GPR40) which binds long chain FA, and G
protein-coupled receptor 120 (GPR120), which binds very long chain FA, have been shown to
increase GLP-1 secretion through the increase in cytosolic calcium. GPR40 is expressed on
enteroendocrine L cells and Grp40-/-
mice have impaired GLP-1 secretion 60 min following an
oral fat challenge (213). GPR120 signalling induces GLP-1 release from STC-1 and NCI-H716

26
intestinal cell lines (214), but a diet enriched in a natural agonist, -3 FA, failed to increase
postprandial GLP-1 in vivo (215). Oleoylethanolamide, a lipid-derived satiety factor generated
from CD36-mediated oleic acid transport (216), has also been shown to stimulate GLP-1
secretion through a G protein-coupled receptor 119 (GPR119)- and PKA-dependent pathway in
enteroendocrine L cells (217). PKC also has a role in oleic acid-induced GLP-1 secretion (209;
218). Moreover, a G protein-coupled receptor (GPCR) for bile acids, TGR5, is expressed on the
surface of enteroendocrine L cells and its activity mobilizes intracellular calcium for GLP-1
secretion (219). Therefore it is evident that multiple aspects of intestinal lipid metabolism are
intricately linked to postprandial GLP-1 and GLP-2 secretion.
Instances of pathology, particularly in insulin resistance and diabetes, have been
associated with modified GLP secretion. In obese women, there was an attenuated GLP-1
response to an oral carbohydrate load (220). The postprandial GLP-1 response, especially within
the first hour, correlated with insulin sensitivity in non-diabetic men (221), and a blunted GLP-1
response was noted even when insulin resistance was not associated with obesity (222). A defect
in GLP-1 secretion is also observed in type 2 diabetes (223), which appears to worsen with
disease severity (224). The importance of insulin sensitivity to GLP-1 levels may be attributable
to the stimulatory effect of insulin on enteroendocrine L cell GLP-1 secretion (225). Leptin is
also a stimulus for GLP-1 secretion and likewise leptin resistance has also been connected to
decreased GLP-1 levels in multiple rodent models, including high fat diet-fed mice (226).
Observations of attenuated GLP-1 secretion have also been noted in the JCR:LA-cp rats (227)
and high fat-fed Wistar rats (228). On the other hand, patients with type 1 diabetes have a
preserved GLP-1 response (207), and ostensibly normal levels of GLP-2, although studies in
streptozocin (STZ)-treated rodents suggest otherwise. The hyperphagia that accompanies STZ-
induced diabetes results in hypersecretion of both GLP-1 and GLP-2 to mediate the intestinal
growth (further discussed later) (229; 230).

27
1.4 Biology of GLP-1
1.4.1 Physiological functions of GLP-1
GLP-1 was discovered in the quest to identify the second incretin hormone to gastric
inhibitory peptide (GIP) (reviewed in (231)). As such, GLP-1 stimulates insulin secretion in a
glucose-dependent manner in response to enteral glucose (232; 233), but can also regulate
glycemic excursion in response to nonenteral glucose (234). In pancreatic islets, GLP-1
upregulates pro-insulin gene transcription (235; 236), sensitizes to glucose (237), suppresses
inflammation (238), restores protein synthesis in the recovery from ER stress (239), and of great
therapeutic interest, exerts protective and proliferative effects (240-244). GLP-1 also has a
glucagonostatic effect on pancreatic -cells, which elaborates on its glucose-lowering ability
(245-247). GLP-1 also has myriad extrapancreatic effects. To complement its ability to
augment insulin secretion, chronic GLP-1R agonism also has insulin-sensitizing effects on other
tissues (248; 249). Notably, exogenous GLP-1 can slow gastric emptying (250), to which part of
its postprandial glucose-lowering ability is attributable (251). In the central nervous system,
GLP-1 has the ability to limit food intake (252), which is of great interest in combating obesity.
Thus in studies looking at the chronic effects of GLP-1, it is essential to dissociate the other
physiological functions of GLP-1 from the weight loss induced by its anorectic ability. There is
also a growing appreciation for the cardiovascular effects of GLP-1, which includes increases in
arterial pressure and heart rate when it is administered peripherally (253) and centrally (254).
1.4.2 GLP-1 receptor
The GLP-1 receptor (GLP-1R) is a 7-transmembrane GPCR and classified as a part of the
family B secretin-receptor family. As to be expected of its insulinotropic function, the GLP-1R
is expressed on the pancreatic -cell, from which the cDNA was originally cloned (255). Much
of the current understanding of GLP-1R signalling comes from work in various -cell lines. The
GLP-1R is a Gs-coupled protein, and so its agonism is associated with an increase in cAMP
levels (256). Thus, it is no surprise that GLP-1 signalling can proceed through PKA (257; 258).
However, the cAMP formed from GLP-1R agonism can also bind cAMP-regulated guanine
nucleotide exchange factor-II (Epac2), which too can lead to a rise in intracellular calcium and
insulin secretion (259; 260). Consistent with its growth factor-like role on the -cell, GLP-1 also

28
activates PI3-K (261) and Erk (258). Many of the same pathways also appear to be applicable to
GLP-1 action in neurons (262; 263), along with PKA-mediated inhibition of 5’-adenosine
monophosphate-activated protein kinase (AMPK) (262).
The GLP-1R is also widely expressed in other tissues, including the lung, kidney, brain,
heart and throughout the gastrointestinal tract (264-266). Its expression in the central nervous
system has been localized to neurons of the nucleus of the solitary tract (NTS), paraventricular
nucleus, central nucleus of the amygdala, anterodorsal thalamic nucleus, and the arcuate nucleus
among other nuclei in the hypothalamus (252; 267), as well as other regions in the hypothalamus
(268). In the gastrointestinal tract, GLP-1R has been identified in vagal afferent nerves
terminating in the hepatic portal vein (269), and in myenteric neurons in the duodenum and colon
(270). The GLP-1R is also found in a specific population of enterocytes which number highest
in the jejunum, and the number of GLP-1R-positive enterocytes increases after ileal transposition
(271). It is not unlikely that GLP-1 exerts appreciable effects in the organ of its origin, given
that DPP-4 is highly expressed in the gastrointestinal tract and degrades the majority of GLP-1
before it enters the systemic circulation (reviewed in (272)).
1.4.3 GLP-1 and lipid metabolism
In adipose tissue, the conclusions are mixed on whether GLP-1 has an anabolic or
catabolic effect on lipid depots. It appears that at very high levels, GLP-1 has a lipolytic effect
on adipocytes associated with cAMP generation, while it is appears to have a direct lipogenic
effect at more physiological concentrations and exhibits synergism with insulin (273; 274).
Moreover, GLP-1R-deficient mice are relatively resistant to HFD-induced weight gain, thus
suggesting a role for GLP-1 signalling in lipid deposition, although the resistance to weight gain
may be attributable to increased energy expenditure arising from greater locomotor activity
(275). On the other hand, central GLP-1 indirectly decreases adiposity in a mechanism involving
the sympathetic nervous system. Through the action of -adrenergic receptors,
intracerebroventricular GLP-1 suppresses mRNA levels of the lipogenic genes FAS, ACC, and
stearoyl-CoA desaturase-1 (SCD-1) (276).
Noguieras et al also noted that central GLP-1R activation also decreased lipogenic gene
expression in the liver, although this was attributable entirely to the weight loss associated with

29
the anorexigenic effect (276). There is mounting evidence that peripheral GLP-1 can exert an
effect on hepatic lipid metabolism. GLP-1R mRNA has been detected in primary hepatocytes
and the receptor exhibits agonist-induced internalization (277), where it activates both Akt and
MAPK pathways (278). Exendin-4 directly reduced TG stores in these cells (277) because of
reduced lipogenesis, as evidenced by decreased SREBP-1c, ACC and SCD-1 mRNA (279).
Upregulated -oxidation may also play a part, as exendin-4 increased peroxisome proliferator
activator receptor (PPAR) and acetyl-CoA oxidase mRNA levels, which was also observed
in vivo in the livers of ob/ob mice (279), while 100 nM GLP-1 has been demonstrated to increase
carnitine palmitoyl transferase-1 (CPT-1) expression in primary hepatocytes. There is evidence
that GLP-1R action can lead to attenuated VLDL secretion, although whether this is
consequential to its insulin effect requires further investigation (280; 281).

30
1.5 Biology of GLP-2
1.5.1 Biological functions of GLP-2
As a consequence of its receptor’s limited expression throughout the body, the biological
actions of GLP-2 are restricted to the gastrointestinal tract, the central nervous system and
pancreatic -cells. The GLP-2 receptor (GLP-2R) has been identified in neurons in the NTS, a
structure in the brainstem that regulates food intake, and like GLP-1, central GLP-2 inhibits food
intake (282). Opposite to GLP-1 though, there are reports that GLP-2 has glucagonotropic
effects by acting directly on pancreatic -cells (283; 284). GLP-2 action in the gut has received
the most attention, and induction of small intestinal epithelial proliferation was the first function
identified for GLP-2 (285), followed by efforts to detail its ability to increase intestinal mucosal
mass. Chronic exposure to exogenous GLP-2 has proliferative and protective effects in crypt
cells, with the result of maintaining mucosal cellularity when faced with such challenges as
parenteral nutrition and radiation (reviewed in (286)). GLP-2 also has anti-apoptotic effects in
epithelial cells further up the villus, thereby increasing villus height upon repeated administration
(287; 288). Such trophic effects of exogenous GLP-2 are also observed, though to a lesser extent,
in the colon (289; 290). Studies done in vitro demonstrate that GLP-2 also promotes intestinal
epithelial restitution (291), and in vivo GLP-2 has been shown to improve intestinal epithelial
barrier function (292). In addition to preventing protein catabolism (287), GLP-2 stimulates
protein synthesis in the intestinal epithelium (293; 294). Physiologically, GLP-2 has been
demonstrated to be important to intestinal adaptive growth in response to feeding (196) and after
bowel resection (295).
A variety of other gut functions are modulated by GLP-2. In the stomach, GLP-2 inhibits
gastric acid secretion (284; 296), ghrelin secretion (297), and modestly slows antral emptying at
higher doses (298). The effect of GLP-2 on gastrointestinal motility is also observed in the small
intestine, where it slows transit time (299). Pharmacological amounts of GLP-2 have anti-
inflammatory effects in rodent models of intestinal injury, including limiting the infiltration of
immune cells and reducing pro-inflammatory cytokine levels (289; 300). At physiological levels,
GLP-2 also modulates the inflammatory response to reduce intestinal permeability (301; 302).

31
Moreover, pharmacological levels of GLP-2 acutely stimulates intestinal blood flow (303; 304),
specifically in the superior mesentery artery, as demonstrated in pigs, rats, and humans (305-307).
1.5.2 GLP-2 and intestinal nutrient absorption
Of interest is the ability of GLP-2 to promote nutrient absorption, because these effects
can typically be observed in an acute setting prior to any appreciable changes in mucosal
architecture. Though an increase in nutrient absorption could be attributable to greater digestive
enzyme activity in animals chronically-administered with GLP-2, such as sucrose-isomaltase in
rats (308; 309), such change in the chronic setting is likely secondary to the hyperplastic effect of
chronic GLP-2 treatment (310). The effect of GLP-2 on carbohydrate absorption has perhaps
received the most attention. GLP-2 can acutely increase enteral hexose transport, due to
upregulated sodium-dependent glucose transporter-1 (SGLT-1) and glucose transporter-2
(GLUT2) insertion into the BBM, the former being a PI3-K-dependent process (311; 312). An
effect of GLP-2 is also observed on the uptake of circulating glucose, possibly attributable to
increased GLUT2 expression on the basolateral membrane (303; 313; 314). The improvement in
dietary glucose uptake can still be observed in cases of chronic GLP-2 administration (315-317),
although it has also been observed that repeated exposure to exogenous GLP-2 does not increase
enteral hexose intake due to decreases in SGLT-1 and GLUT2 mRNA levels (303; 310). In
piglets receiving total parenteral nutrition, 6-day treatment with GLP-2 conditioned the intestine
to increase capacity for glucose and galactose transport, with increased SGLT-1 providing
greater apical transport activity and decreasing glucose metabolism to lactate (318). To
complement its protein anabolic effect, chronic GLP-2 treatment also stimulates amino acid
uptake in mice and pigs (310; 316). Intravenous delivery of GLP-2 for 6 days to fetal pigs
increased lysine uptake as determined by assays of intact intestinal tissue (319). A 14-day
infusion of GLP-2 by a subcutaneously-placed osmotic pump also dose-dependently upregulated
glycine absorption per small intestinal surface area (320). GLP-2’s influence on dietary lipid
absorption has mostly been investigated in a chronic setting, where adult mice exhibited greater
uptake of orally administered triolein (310). However, in suckling rats, GLP-2 increased FA
uptake in the ileum, but only in conjunction with dexamethasone (321). One acute study in
humans did note that GLP-2 infusion augmented postprandial TG and FFA, although the authors
surmised this was the result of decreased peripheral lipoprotein clearance as opposed to

32
modulated intestinal function (284). Therefore, there remains much to be characterized with
regards to GLP-2 action on intestinal lipid transport. Some functions of GLP-1 and GLP-2 are
summarized and compared and contrasted in Table 1-1.

33
Function GLP-1 GLP-2
Insulin secretion Stimulates (322; 323) No effect (323; 324)
Glucagon secretion Inhibits (194)
Inhibits in type 2 diabetes (245;
246)
Stimulates (283; 284)
No effect in type 2 diabetes
(246)
Intestinal growth Increases mucosal area in Goto-
Kakizaki rats (325)
No effect (285)
Colonic epithelial growth (326)
Crypt cell proliferation (327-
329)
Reduction in epithelial apoptosis
(287; 293)
Enhances barrier function (292;
330)
Pancreatic -cell growth Anti-apoptotic in humans (241)
Differentiation in humans (331;
332)
Neogenesis in rodents (243;
333)
Proliferation in rodents (240;
242-244)
No effect (334)
Gastric emptying Inhibits (198) Inhibits (298; 335)
Intestinal motility Inhibits (247; 336; 337) Inhibits in the presence of GLP-

34
1 (338)
Glucose uptake Potentiates insulin-stimulated
glucose uptake in peripheral
tissues (334; 339; 340)
Stimulates luminal glucose
uptake in the intestine (312)
Stimulates circulating glucose
uptake in the intestine (303)
Neurotropism Protective (341; 342)
Neurogenic (343; 344)
Protective (345)
Proliferative (346)
Food intake Centrally and peripherally
inhibits (252; 347)
Centrally inhibits (282)
Arterial blood flow Stimulates (253; 348) Stimulates mesenteric blood
flow (303; 349)
Heart rate Increases (253; 348) No effect (253; 305)
Bone resorption Inhibits (350) Inhibits (351; 352)
Table 1-1. Biological functions of GLP-1 and GLP-2
Differences and similarities in the biological actions of GLP-1 and GLP-2 as inferred from
studies with agonists, antagonists, and knockout mice. References included in the table are often
just a sampling of the studies that characterized these actions.

35
1.5.3 GLP-2 receptor
Efforts to detail the mechanisms of GLP-2-improved intestinal function have been
hindered largely by the highly specific expression of the GLP-2R in the intestine. Although
radiolabelled GLP-2 has been found to bind the villus epithelium in high density (353),
subsequent studies indicated that GLP-2R is conspicuously absent on the absorptive enterocyte.
Rather, to date the GLP-2R has been identified on intestinal subepithelial myofibroblasts
(ISEMF) (354), specific enteroendocrine cells (304; 355), enteric neurons (356; 357), and the
vagal afferent nerve (358). ISEMFs are positive for -smooth muscle actin and vimentin, but do
no express desmin like smooth muscle (359). ISEMFs are found throughout the lamina propria,
but GLP-2R-positive ISEMFs appeared to be limited to the population sitting under the basement
membrane of the intestinal epithelium at the villus tip (354). Moreover, GLP-2R expression is
limited to particular populations of these cell types, as it was colocalized with vasoactive
intestinal peptide (VIP)-positive and endothelial nitric oxide synthase (eNOS)-positive enteric
neurons, and specific enteroendocrine L-cells that have been detected to be positive for
serotonin, chromagranin A, in addition to enteroendocrine L-cells and K-cells (304; 355). These
cells are of relatively low abundance in the gastrointestinal tract and for this reason many earlier
studies on GLP-2R signalling has relied on transfected cell lines. The GLP-2R is a Gs-coupled
receptor, and so its agonism is associated with cAMP accumulation and PKA activation (360;
361). The cAMP accumulation is important to reducing apoptotic signalling, such as caspase-3
cleavage and mitochondrial cytochrome C release (362), while PKA activation is necessary to
inhibit the pro-apoptotic activities of Bad and glycogen synthase kinase 3 (GSK-3) (363). GLP-
2 also activates the PI3-K pathway in the intestinal epithelium, which may explain its
proliferative actions, although this likely occurs indirectly of GLP-2R signalling (293; 364). The
PI3-K/Akt pathway has also been implicated in the upregulation of insulin-like growth factor-1
(IGF-1) mRNA in intestinal subepithelial myofibroblasts (359), and protein synthesis in
transfected HEK-293 cells (365). The proliferative action of GLP-2 can also occur through
ERK1/2 signalling, stimulated putatively by the GLP-2R coupling to Gi/Go proteins (366). Like
the GLP-1R (367), the GLP-2R exhibits desensitization induced by its agonist and internalizes in
a manner requiring lipid rafts (368).

36
Despite the knowledge of GLP-2R signalling, given that the absorptive enterocyte does
not express the receptor, there is still much to understand about GLP-2’s mechanism of action on
the intestinal epithelium. GLP-2 must bind its receptor on a certain intermediary cell,
stimulating it to secrete a mediating signal that would act directly on the enterocyte to bring
about chylomicron overproduction. IGF-1/2 (369), keratinocyte growth factor (KGF) (370), and
EGF (328) have been suggested as mediators of the proliferative and protective effects of GLP-2
on the intestinal epithelium. Vascular endothelial growth factor (VEGF) has been implicated in
mediating intestinal epithelial restitution (371). VIP mediates the relaxation of fundic smooth
muscle induced by GLP-2 (372). To exact its anti-inflammatory effects, GLP-2 has also been
shown to activate VIP-positive enteric neurons in the submucosal plexus (300). However, the
peptide mediator for GLP-2’s stimulation of enteral nutrient uptake has not yet been identified.

37
1.6 Hypothesis
The intestine is evidently sensitive to hormonal signals, and its handling of dietary lipid is
a process that can be actively regulated. GLP-1 and GLP-2 are two postprandial hormones that
have been shown to have a number of biological effects in the gut. The fact that lipid ingestion
serves as a stimulus for the secretion of these two peptides hints at their potential role in
postprandial lipid metabolism. Moreover, GLP-1 can modulate lipid metabolism in other tissues
while GLP-2 has been shown to promote the absorption of various nutrients in the gut.
Therefore, my general hypothesis is that the GLPs are critical acute regulators of intestinal
lipid absorption and apoB48-containing lipoprotein secretion. This hypothesis was
examined in this thesis using multiple rodent models and primary intestinal cell cultures. There
was a particular reliance on the Golden Syrian hamster Mesocricetus auratus, which similar to
humans has minimal apoB mRNA editing in the liver and therefore secretes apoB48 exclusively
from the intestine (373). The use of the hamster model therefore allows for the ready delineation
between hepatic and intestinal contributions to circulating apoB levels in vivo.

38
Chapter 2: THE GLUCAGON-LIKE PEPTIDE-1
RECEPTOR REGULATES POSTPRANDIAL
LIPOPROTEIN SYNTHESIS AND SECRETION
Text and Figures in this chapter is reproduced from:
Hsieh J, Longuet C, Baker CL, Qin B, Federico LM, Drucker DJ, Adeli K. The glucagon-like
peptide 1 receptor is essential for postprandial lipoprotein synthesis and secretion in hamsters
and mice. Diabetologia. 2010;53(3):552-61. Copyright 2009, Springer
Author contributions:
Sitagliptin-treated hamster data was generated by J. Hsieh with the assistance of C.L. Baker. In
vivo mouse experiments and mouse hormones measurements were performed by C. Longuet
while biochemical analyses were performed by J. Hsieh. Exendin-4-treated hamster and
enterocyte data was generated by B. Qin, and J. Hsieh performed FPLC analysis. Data analysis
and writing were completed by J. Hsieh and C. Longuet.

39
2.1 Summary
Background: GLP-1 receptor agonists and DPP-4 inhibitors attenuate postprandial lipemia
through mechanisms that remain unclear. As dyslipidemia is a contributing risk factor for
cardiovascular disease in type 2 diabetes, we examined the mechanisms linking pharmacological
and physiological regulation of GLP-1 action to control of postprandial lipid metabolism.
Methods: Postprandial lipid synthesis and secretion was assessed in normal and fructose-fed
hamsters and in wildtype mice, treated with or without sitagliptin. ApoB48 synthesis and
secretion was also examined in primary enterocyte cultures. The importance of exogenous vs.
endogenous GLP-1 receptor signaling for regulation of intestinal lipoprotein synthesis and
secretion was assessed in mice and hamsters treated with the GLP-1R agonist exendin-4, the
GLP-1R antagonist exendin9-39 and in Glp1r+/+
vs. Glp1r-/-
mice.
Results: Sitagliptin decreased fasting plasma TG, predominantly in the VLDL fraction, as well as
postprandial TRL-TG, TRL-cholesterol and TRL-apoB48 in both hamsters and mice. GLP-1R
activation with exendin-4 alone also decreased plasma and TRL apoB48 in hamsters and mice
and reduced secretion of apoB48 in hamster enterocyte cultures. Conversely, blockade of
endogenous GLP-1R signaling using the antagonist exendin9-39 or genetic elimination of GLP-
1R signaling in Glp1r-/- mice enhanced TRL-apoB48 secretion in vivo.
Conclusions: Potentiation of endogenous incretin action via DPP-4 inhibition, or
pharmacological augmentation of GLP-1 receptor signaling reduces intestinal secretion of TG,
cholesterol and apoB48. Moreover, endogenous GLP-1R signaling is essential for the control of
intestinal lipoprotein biosynthesis and secretion.

40
2.2 Introduction
The growing incidence of type 2 diabetes (T2DM) is a major problem (374) and may be
associated with a variety of lipid abnormalities that pose as cardiovascular disease risk factors,
including hypertriglyceridemia, increased levels of LDL, and low levels of HDL (375). At the
top of this cascade are intestinally-derived, TG-rich apoB48-containing chylomicrons, which are
secreted following fat ingestion and peripherally catabolized to generate remnant particles (376).
Insulin resistance has been found to be positively correlated with remnant-like particle
cholesterol (377) and remnant-like particle TG is greatly increased in some diabetic populations
(378). Thus, it is of interest to identify therapeutic strategies that can limit postprandial TRL
secretion.
The ingestion of nutrients, including fat, provokes the secretion of gut-derived hormones,
including two incretins: GIP from duodenal K cells (379) and GLP-1 from ileal enteroendocrine
L cells (218). These peptides stimulate the secretion of insulin in a glucose-dependent manner,
and preserve pancreatic -cell function and mass in preclinical studies (reviewed in (380)). A
number of extra-pancreatic effects have also been demonstrated following exogenous
administration of these two hormones. GLP-1 slows gastric emptying and induces anorectic
effects (381), while GIP has anabolic effects on adipose tissue (382) and regulates adipokine
secretion (383). However, both GLP-1 and GIP are rapidly inactivated by DPP-4-mediated
cleavage, thus limiting the duration of action. Insulin resistance is associated with impaired
stimulated GLP release. In high fat diet-fed mice, ileal and colonic content of GLP-1 have been
reported to be higher (226; 384), although the L-cells exhibit a blunted GLP secretory response
to glucose stimulation (226). Insulin-resistant MKR mice have higher basal GLP-1 levels, but
likewise also exhibit impaired glucose-stimulated GLP secretion (225). Inducing insulin
resistance in murine GLUTag cells in vitro also compromised GLP-1 secretion in response to
insulin (225). While some clinical studies agree with the findings in these experimental models
(223; 385; 386), some studies have reported no diminution in GLP-1 secretion in type 2 diabetic
patients (387). Therefore, there is considerable interest in enhancing incretin action for the
treatment of type 2 diabetes.

41
DPP-4 inhibitors such as sitagliptin or vildagliptin protect endogenous GLP-1 and GIP
from degradation-terminal enzymatic inactivation, thereby prolonging their bioactivity. DPP-4
inhibition effectively lowers fasting and postprandial glycemia in patients with T2DM (388;
389). Given the increasing use of sitagliptin and vildagliptin for the treatment of T2DM, there is
considerable interest in understanding the effects of incretins and DPP-4 inhibitors on plasma
lipid profiles (390). Acute administration of GIP reduces circulating chylomicrons, likely
through promoting TG catabolism by adipose tissue (391; 392) and GLP-1 also attenuates
postprandial triglyceride secretion (393; 394), although the exact mechanisms underlying these
observations are not clear.
In the present study we studied the role of incretin action in controlling intestinal lipid and
lipoprotein metabolism using wildtype mice and a hamster model of insulin resistance and
postprandial dyslipidemia (146). The fructose-fed hamster exhibits intestinal overproduction of
TRL (71), along with aberrant insulin signalling in the absorptive enterocyte (154). Mice and
hamsters were treated with sitagliptin or exendin-4 to potentiate incretin action or with or
without exendin 9-39 to assess the importance of exogenous GLP-1R activation vs. endogenous
basal GLP-1R signaling for regulation of intestinal lipoprotein metabolism.

42
2.3 Materials and Methods
2.3.1 Animals
Male Syrian golden hamsters (Mesocricetus auratus) weighing 130-150 g were
purchased from Charles River (Montreal, PQ) and housed individually with room lighting set for
a 12:12-h light-dark cycle with ad libitum access to food and water. Animals were acclimatized
for 1 week prior to being given either a standard chow diet or a fructose-enriched diet (pelleted
hamster diet containing 60% fructose and 20% casein, Dyets Inc., Bethlehem, PA) for 10 day to
induce insulin resistance (145). The hamsters were then randomized to receive either sitagliptin
phosphate monohydrate (Merck, Whitehouse Station, NJ) (5 mg/kg) or water by oral gavage QD
in the afternoon under light inhalant anaesthesia. Chow-fed hamsters were given sitagliptin for
two weeks while fructose-fed hamsters were dosed for three weeks. Blood was collected by
retro-orbital bleeding at the baseline, endpoint, and midpoint of the dosing regimen to assess
ongoing effect of the treatment, in both the morning for ambient (fed) measurements and after a
6 hr fast. The hamsters were then sacrificed for the ex vivo protocol or underwent the in vivo
protocol. All animal protocols were approved by the animal ethics committee of the Hospital for
Sick Children, University of Toronto. Wild type C57BL/6J mice (Jackson laboratory), Glp1r-/-
and age and sex-matched Glp1r+/+
littermate controls fed a normal chow were used at the age of
10-12 weeks of age and maintained under a 12 h light/12 hr dark cycle. Treatments performed in
mice utilized a single dose of sitagliptin (10 mg/kg BW) or exendin-4 (24 nmol/kg) administered
as described in figure legends.
2.3.2 Assessment of intestinal lipoprotein production by in vivo Triton-WR1339
infusion
Hamsters were anesthetized with isoflurane administered through a vaporizer. A cannula
was inserted into the right jugular vein, exteriorized at the back of the neck, filled with
heparinized saline (40 IU/mL) and sealed. The hamsters were allowed to recover and fasted
overnight. Conscious hamsters were then given a 200 L olive oil load via oral gavage. 20 min
later, vehicle (saline), exendin-4 (5 nmol/kg) (Bachem, Torrance, CA), exendin(9-39) (50
nmol/kg) (Bachem), or both were administered via an intraperitoneal injection plus a separate

43
bolus of Triton WR-1339 diluted 20% (w/v) (0.5 g/kg) injected into the jugular vein to inhibit
lipoprotein catabolism and uptake (395; 396). Triton WR-1339 coats TRL particles to prevent
hydrolysis of the TG content by LpL, allowing for assessment of secretion without taking into
account any changes in peripheral catabolism. 300 L of blood was collected from the jugular
cannula into lithium heparin-coated tubes (BD Biosciences, Franklin Lakes, NJ) at baseline and
at 30, 60, and 120 min. The oral gavage, peptide administration, Triton infusion, and blood
collection were all performed on conscious animals in the absence of anesthetics. The jejunum
was excised under isoflurane anesthesia at the endpoint.
Mice fasted for 5 or 16 h were given 200 L olive oil orally. 20 min after gavage, a blood
sample was collected via the tail vein (time 0). Acute sitagliptin administration was given orally
to conscious mice 20 min prior to olive oil gavage. 20 min after fat load, Triton WR-1339 (0.5
g/kg BW of a 15% solution prepared in PBS) was injected via the tail vein with or without
exendin-4 (24 nmol/kg). Blood samples were collected via tail bleed without anesthestics for
plasma triglyceride and cholesterol assay and apoB48 immunoblotting. 90 min after Triton WR-
1339 injection, mice were euthanized and cardiac puncture was performed for plasma
triglyceride and cholesterol assay, apoB48 western blot analysis, and TRL purification as
described below.
2.3.3 Isolation of triglyceride-rich lipoproteins (TRL)
To isolate the TRL fraction of the plasma, blood samples were first centrifuged at 4°C for
15 min at rpm to separate the plasma layer. 150 L of plasma was overlayed with 4 mL of
potassium bromide solution (density = 1.006g/mL) in a 5 mL ultracentrifuge tube and
centrifuged at 35,000 rpm for 70 min at 15°C using a SW 55 Ti rotor (Beckman Coulter,
Mississauga, ON). The TRL fraction (Sf >400) was collected as the top 300 L of the tube.
2.3.4 Chemiluminescent immunoblotting
ApoB48 immunoblotting was performed on TRL fractions and diluted plasma (1:200) by
SDS-PAGE analysis as previously described (145). Following SDS-PAGE, the proteins were
transferred electrophoretically overnight at 4 °C onto PVDF membranes using a Bio-Rad wet
transfer system. The membranes were blocked with 5% fat-free milk solution and then incubated

44
with rabbit anti-hamster apoB antibody (1:1000) for 1-24 h. After several washes with TBST, the
membranes were incubated with peroxidase-conjugated secondary antibody (1:10,000 dilution)
for 1-2 hr. Membranes were then incubated in ECL detection reagents and exposed to Hyperfilm.
Quantitative analysis was performed using an imaging densitometer.
2.3.5 Ex vivo metabolic labeling of intact primary enterocytes
Primary enterocytes were isolated from hamster intestinal tissue as described previously,
which allows for over 90% viability for 4 h (71). This method developed by Perreault and
Beaulieu gives fully differentiated intestinal epithelial cells free of mesenchymal contamination
(397). Briefly, the jejunum was harvested from chow-fed hamsters, flushed with PBS to remove
luminal contents, and then cut longitudinally and into 5 mm strips. The jejunal pieces were
treated with Cell Recovery Solution (BD Biosciences) at 4°C for 45 minutes with agitation,
washed with ice-cold PBS, and then viable villi were collected. The villi suspension was washed
twice in ice-cold phosphate-buffered saline (180 × g, 5 min). After the final spin, villi were
resuspended in Dulbecco's modified Eagle's medium (DMEM, Wisent) supplemented with 1%
fetal bovine serum and placed in an incubator (37 °C, 5% CO2, 95% air, 100% humidity). The
experiment was performed with 8 mM glucose and 100 pM exendin-4 added to the media.
Primary hamster enterocytes were preincubated in methionine-free MEM at 37 °C for 45 min
and pulsed with 30-50 µCi/ml of [35
S]methionine for 30 min. The cells were then chased with
unlabelled 40 mM methionine-enriched DMEM.
2.3.6 Immunoprecipitation, SDS-PAGE, and fluorography
ApoB48 immunoprecipitation was performed as described previously (71).
Immunoprecipitates were washed and prepared for SDS-PAGE. Gels were then fixed, incubated
in Amplify (Amersham), dried and exposed to film at -80 °C for 1-4 days. The apoB48 bands
were excised and quantitative analysis was performed using a liquid scintillation counter. No
major differences were noticed in the trichloroacetic acid (TCA) protein precipitation counts
between the control and experimental groups. Media counts were normalized to initial cellular
TCA counts. Radioactivity incorporation into apoB48 was also visualized with a Phosphor
Imager.

45
2.3.7 Plasma measurements
Plasma TG and cholesterol was determined by an enzymatic-based colorimetric assay
(Randox, Crumlin, UK).
2.3.8 Fast protein liquid chromatography of plasma lipoproteins
200 L of hamster plasma was first filtered through a 0.45 M micro-spin polysulphone
filter (Alltech, Mandel Scientific, Lachine, PQ) to remove macroparticles. The plasma was then
subjected to gel filtration through a HR 10/300 GL Superose 6 column (Pharmacia, Uppsala,
Sweden) with a solution of 10 mM Tris, 150 mM NaCl, 2 mM CaCl2, 100 DTPA, 0.02%
NaN3, pH 7.4 was pumped through at a flow rate of 0.5 mL/min. Fractions were collected every
min.
2.3.9 Statistical analysis
All results are presented as mean ± SEM. Statistical comparisons were performed using
Student's t-test when comparing two groups and two-way ANOVA with Bonferroni’s post-test as
indicated in the text and figure legends.

46
2.4 Results
2.4.1 A DPP-4 inhibitor attenuates dyslipidemia in fructose-fed hamsters
As increased incretin action can attenuate postprandial plasma lipid accumulation (393;
394; 398), we first examined the consequences of DPP-4 inhibition on accumulation of TG and
cholesterol in hamsters fed a high fructose diet (60%), a regimen previously shown to result in
dyslipidemia and mild insulin resistance (399). Baseline values after a 5 h fast, obtained 10 days
after the beginning of high fructose diet, and prior to start of sitagliptin treatment, show a
significant increase in plasma of TG and cholesterol (Figure 2-1A,B; p = 0.0002 and 0.006
respectively), in the absence of changes in blood glucose or body weight (summarized in Table
2-1). After three weeks of sitagliptin treatment, plasma TG levels did not continue to rise
(Figure 2-1A).
Fast protein liquid chromatography (FPLC) was performed to separate the various
densities of lipid particles. In fructose-fed hamsters, levels of VLDL and LDL TG and
cholesterol were significantly higher than in control animals (data not shown). Sitagliptin
significantly attenuated the accumulation of VLDL-TG by 3-fold (Figure 2-1C). Although LDL
and HDL TG levels in fructose-fed hamsters were slightly decreased by sitagliptin treatment, this
effect was not statistically significant (Figure 2-1C). LDL and HDL TG levels were slightly but
not significantly decreased by sitagliptin treatment (Figure 2-1C). Whereas total plasma
cholesterol was not altered by sitagliptin, LDL-cholesterol trended towards reduction and a
significant decrease in VLDL- and HDL-cholesterol (Fig 2-1D) was observed in sitagliptin-
treated vs. control animals (p < 0.05).

47
Figure 2-1. Changes in plasma lipids following administration of sitagliptin
Blood samples were drawn from hamsters fed either regular chow or a high fructose diet
supplemented with vehicle or sitagliptin for 2 to 3 weeks. Plasma TG (A) and cholesterol (B)
levels were determined both prior to and after each dosing period. Lipoproteins were separated
from plasma by FPLC fractionation into VLDL/chylomicron remnants, LDL and HDL and levels
of TG (C) and cholesterol (D) were determined. n = 9-16 for plasma lipid parameters analyzed
by paired t-test; n= 4 for FPLC parameters analyzed by Student’s t-test, *p<0.05 fructose-fed
sitagliptin vs. fructose-fed vehicle, ***p<0.001 fructose-fed vehicle vs. chow-fed vehicle or
fructose-fed sitagliptin vs. chow-fed sitagliptin.

48
Chow fed
vehicle
Chow fed
sitagliptin
Fructose fed
vehicle
Fructose fed
sitagliptin
Baseline body
weight (g)
130.0 ± 3.1 129.3 ± 3.5 123.7 ± 1.9 124.6 ± 2.1
Endpoint body
weight (g)
141.4 ± 3.2 139.4 ± 3.7 129.0 ± 1.8 129.1 ± 1.0
Baseline fasting
glucose (mM)
4.7 ± 0.3 4.5 ± 0.1 5.3 ± 0.3 4.6 ± 0.2
Endpoint
fasting
glucose (mM)
4.7 ± 0.5 4.1 ± 0.2 5.5 ± 0.4 5.1 ± 0.3
Table 2-1. Body mass and blood glucose following sitagliptin treatment.
Body mass and blood glucose in chow-fed hamsters and fructose-fed hamsters chronically
treated with sitagliptin as described in methods. Values shown are mean ± SE, n = 5 – 10 per
group. None of the values were statistically significant as determined by two-way ANOVA.

49
2.4.2 A DPP-4 inhibitor decreases intestinal production of TRL-TG and TRL-
cholesterol
To understand how sitagliptin modulates plasma lipoprotein levels, we assessed intestinal
lipoprotein production and secretion after treatment of chow fed hamsters with sitagliptin for 2
weeks. In chow-fed hamsters, there was a trend for sitagliptin to reduce TRL apoB48 (Figure 2-
2C, p = 0.16) in TRL fraction after an acute fat load, while the accumulation of TRL-TG and
TRL-cholesterol was unaffected (Fig 2-2A and Fig 2-2B). In contrast, sitagliptin significantly
reduced the amount of TG in TRL fraction 90 min after the fat load (Figure 2-2E) in fructose-fed
hamsters and there was a trend for lowered TRL cholesterol (Figure 2-2D, p = 0.19) compared to
vehicle-treated high fructose fed animals. Slope calculations for TRL-cholesterol and TG
secretion in fructose-fed hamsters (for cholesterol, 0.280 ± 0.060 mg/dL min-1
vs. 0.428 ± 0.104
mg/dL min-1
, sitagliptin vs. control, p = 0.23 for the slope difference, and p = 0.037 for elevation
difference; for TG, 1.55 ± 0.30 mg/dL min-1
vs. 2.23 ± 0.33 mg/dL min-1
, sitagliptin vs. control,
p = 0.13 for the slope difference, while p = 0.012 for the elevation difference) suggested that
sitagliptin retards intestinal postprandial TG secretion. Because of normalization to very high
baseline TRL-apoB48 levels, the quantified postprandial rise in apoB48 in fructose-fed hamsters
appeared to be less pronounced compared to chow-fed hamsters (Figure 2-2F). While decreased
postprandial production of lipoproteins may play a role in reduced fasting lipoprotein levels
observed in Figure 2-1, the effect of hepatic production at this point cannot be excluded.
To determine if the effects of sitagliptin on plasma levels of cholesterol and TG were
restricted to hamsters, the effect of an acute dose of sitagliptin on chylomicron production was
assessed in chow-fed mice. Sitagliptin significantly decreased cholesterol in plasma (Figure 2-
3A) and in the TRL enriched fraction (Table 2-2) after an acute fat load; furthermore, plasma TG
(Figure 2-3B) and TRL-TG (Table 2-2) were significantly reduced 90 min after Triton injection
in sitagliptin-treated mice. The accumulation of ApoB48 in plasma was also decreased by
sitagliptin administration (Figure 2-3C).

50
Figure 2-2. Changes in TRL lipid mass following chronic sitagliptin administration
Hamsters were fed regular chow (A-C) or a high fructose diet (D-F) and administered either
vehicle or sitagliptin for 2 to 3 weeks following which hamsters were fat-loaded, administered
Triton WR-1339 and blood was drawn at 0, 30, 60, 90 and 120 minutes. Plasma was spun to
isolate TRL fraction and levels of cholesterol, TG and ApoB 48 were determined. Data shown
illustrates TRL levels of (A) cholesterol, (B) TG and (C) ApoB48 in chow fed hamsters with
representative apoB48 blots shown above. (D) Cholesterol (E) TG and (F) ApoB48 content of
the TRL fractions from fructose fed hamsters with representative apoB48 blots shown above. (n
= 4 for each group * p < 0.05 as analyzed by two-way ANOVA with Bonferroni’s post-test).

51
TRL TG (mmol/L) TRL-cholesterol
(mmol/L)
TRL-apoB48
mass
(% control)
A
Control
Sitagliptin
4.52 ± 1.04
2.10 ± 0.31a
1.01 ± 0.28
0.73 ± 0.07
100.00 ± 6.41
78.23 ± 10.52
B
Control
Insulin
D-Ala2-GIP
Exendin-4
1.61 ± 0.30
1.98 ± 0.67
3.57 ± 1.13
0.53 ± 0.11a
0.55 ± 0.14
n.d.
0.95 ± 0.32
0.19 ± 0.08
100.00 ± 2.6
n.d.
173.13 ± 26.09
59.78 ± 10.25
C
Control
Exendin-4
3.06 ± 0.64
1.13 ± 0.24a
n.d.
n.d.
100.00 ± 10.24
71.61 ± 17.41
D
Wild type
Glp1r-/-
1.78 ± 0.16
2.84 ± 0.22a
n.d.
n.d.
100.00 ± 8.53
121.32 ± 2.65a
Table 2-2. TRL fraction measurements in chow-fed mouse studies
TRL fraction lipid masses determined by colorimetric assays and TRL-apoB48 mass determined
by immunoblotting in chow-fed mouse studies, as described in methods and corresponding
figure legends. A: Mice were given sitagliptin 20 min prior fat load. Methods and plasma levels
of triglycerides, cholesterol and ApoB48 are reported in Figure 3. B: Mice were given insulin, D-
[Ala2] GIP or exendin-4 20 min after fat load. Plasma levels of triglycerides, cholesterol and
ApoB48 are reported in Figure 2-4A-E. C: Mice were given exendin-4 1h after fat load. Plasma
levels of TG and ApoB48 are reported in Figure 2-4F and 2-4G. D: Glp1r-/-
mice and littermate
controls were given an oral fat load to monitor intestinal lipid absorption. TG and ApoB48
plasma levels are reported in figure 5C and D. Values shown are mean ± SEM, a p < 0.05 vs.
control of respective group as determined by Student’s t-test, n.d. not determined.

52
Figure 2-3. Changes in postprandial lipid secretion following a single acute administration
of sitagliptin in chow-fed mice
Chow-fed wild type mice fasted for 5 h were given 10 mg/kg of sitagliptin prepared in water by
oral gavage. 20 min after sitagliptin treatment, mice were administered 200 L of olive oil and
mice were injected intravenously 20 minutes later with Triton WR1339 (0.5 g/kg ). Blood
samples were collected prior to IV injection (time 0), as well as 30, 60 and 90 min after injection
for (C) cholesterol, (D) TG and (B) ApoB48 measurements as described in methods, n = 4 per
group. * p < 0.05, *** p < 0.001 as determined by two-way ANOVA with Bonferroni’s post-test
analysis.

53
2.4.3 Pharmacological activation of the GLP1R mimics the effects of sitagliptin on
intestinal lipid absorption
As both GLP-1 and GIP have been involved in the regulation of plasma lipid levels and
are DPP-4 substrates, we measured intestinal lipid absorption in mice after acute administration
of the DPP-4 resistant analogues of GIP (D-Ala2-GIP) and GLP-1 (exendin-4). As shown in
figure 4, D-Ala2 GIP significantly increased levels of plasma triglyceride and ApoB48 after an
acute fat load (Figure 2-4A and 2-4B), and therefore could not have been responsible for the
lowering actions of sitagliptin on intestinal lipid absorption. In contrast, the GLP-1R agonist
exendin-4 reduced plasma and TRL-TG and ApoB48 accumulation (Figure 2-4D and 2-4E,
Table 2-2), and no effect was noted on plasma and TRL cholesterol (Figure 2-4C, Table 2-2).
Hence, activation of GLP1R, but not GIPR signaling, mimics the actions of sitagliptin on
intestinal lipid absorption.
Both insulin and glucagon have been involved in the regulation of lipemia and activation
of GLP1R signalling has been shown to stimulate insulin secretion and inhibit glucagon
secretion. Accordingly we measured plasma insulin and glucagon levels after an oral fat load,
with or without sitagliptin or exendin-4 treatment. Both sitagliptin and exendin-4 significantly
increased plasma insulin levels compared to controls (Figure 2-5A), but this increase was much
more rapid after exendin-4 administration. However, this transient difference in plasma insulin
levels does not likely account for the effect of sitagliptin or exendin-4 on intestinal lipid
absorption or for the difference in magnitude of the effect of those 2 treatments, as plasma
insulin levels are no longer significantly different between the groups 20 min after the fat load
(Figure 2-5A). More importantly, exogenous insulin administration had no effect on TG
accumulation (Figure 2-5B). Furthermore, neither sitagliptin nor exendin-4 had a significant
effect on plasma glucagon levels (Figure 2-5C). As expected, there was a trend for increased
plasma GLP-1 levels, as soon as 5 min following sitagliptin administration (Figure 2-5D).
As exendin-4 is a rapid and potent inhibitor of gastric emptying, which in turn might contribute
to decreased intestinal lipid absorption, we monitored intestinal lipid absorption when exendin-4
was administered 1 h after the oral fat load, to facilitate entry of the olive oil into the mouse

54
small bowel. Under these conditions, exendin-4 still significantly decreased triglyceride and
ApoB48 accumulation in plasma (Figure 2-4F and 2-4G) and TRL fraction (Table 2-2).

55
Figure 2-4. Pharmacological activation of the GLP1R mimics the effect of sitagliptin on
post-prandial lipid excursion
Chow-fed wild type mice fasted for 5h were gavaged with olive oil, and injected with Triton
WR-1339 (0.5 g/kg body weight) and D[Ala2]-GIP (24 nmol/kg) (A, B) or exendin-4 (24
nmol/kg) (C-G) 20 min (A-E) or 1h (F, G) after fat load. Blood samples were collected prior to
intravenous injection (time 0), as well as 30, 60 and 90 min after injection. (A, D, F) TG, (C)
cholesterol and (B, E, G) apoB48 secretion were measured, n = 3-6 per group, * p < 0.05 as
determined by two-way ANOVA with Bonferroni post-test.

56
Figure 2-5. Role of glucose-modulating hormones in mice
(A, C-E) Mice fasted for 5h were administered a single dose of sitagliptin at 10 mg/kg (A, C, D)
or exendin-4 24 nmol/kg (A, C, E), and 20 min later were challenged with an oral fat load.
Blood was collected by cardiac puncture 5 and 20 min after the olive oil gavage (25 and 40 min
after the initial sitagliptin dose). Hormones were measured at the indicated time after the olive oil
gavage. (A) Insulin (Mouse insulin ultrasensitive ELISA – Alpco Diagnostic) (C) Glucagon
(Mouse endocrine Lincoplex – Linco research) (D) Total GLP-1 (Mouse total GLP-1 ELISA –
Alpco Diagnostic) (E) Exendin-4 (Exendin-4 EIA kit – Phoenix Pharmaceuticals). n = 3 per
group, * p < 0.05 control vs. sitagliptin, ** p < 0.01 control vs. exendin-4 as determined by two-
way ANOVA. (B) Mice were challenged with an oral fat load, and then injected with insulin
(1.2 mU/kg BW) 20 min later concomitant with intravenous Triton WR-1339 (0.5 mg/kg BW)
administration to measure TG secretion, n = 6 per group, p > 0.05 as determined by two-way
ANOVA.

57
2.4.4 Endogenous GLP-1R signalling is required for control of postprandial
lipemia
We next investigated the role of physiological levels of GLP-1 in the regulation of
intestinal lipid absorption in both mice and hamsters after an acute fat load. Exendin-4 decreased
TG in the VLDL/chylomicron remnant-sized lipoproteins in chow-fed hamsters not given a bolus
of Triton WR-1339 (Figure 2-6A), and significantly reduced the levels of apoB48 in the TRL
fraction 90 min after a fat load (Figure 2-6B). The antagonist exendin9-39 blocked the exendin-
4-mediated reduction of TRL-apoB48, and exendin9-39 alone augmented levels of TRL-apoB48
120 minutes after fat load (Figure 2-6B), indicating that endogenous basal GLP-1R signalling
modulates postprandial lipemia.
Consistent with data obtained using the GLP1-R antagonist exendin9-39 in hamsters, TG
accumulation in plasma (Figure 2-6C) and TRL fraction (Table 2-2) was significantly enhanced
in Glp1r-/-
mice compared to Glp1r+/+
littermate controls despite the fact that Glp1r-/-
mice have a
gastric emptying rate similar to Glp1r+/+
littermate controls (400). There was also significantly
greater TRL apoB48 mass in GLP-1R-deficient mice (Table 2-2). Therefore, the modulatory
actions of basal GLP-1R signaling on intestinal lipid absorption are not due to regulation of
gastric emptying, and implicate the endogenous GLP-1 receptor in the control of intestinal lipid
absorption.
2.4.5 Intact GLP-1R signalling is required for the hypolipidemic action of
sitagliptin
To determine whether the actions of sitagliptin on postprandial lipid secretion required
GLP-1R signalling, the GLP-1R antagonist exendin9-39 was co-administered with sitagliptin 20
min before the fat load. Sitagliptin attenuated postprandial TG excursion in mice (Figure 2-6F)
but did not significantly affect plasma cholesterol levels (Figure 2-6E). Furthermore, the GLP-1R
antagonist exendin9-39 eliminated the hypolipidemic actions of sitagliptin on plasma TG levels
(Figure 2-6F). Unlike in hamsters (Figure 2-6B), exendin9-39 alone did not induce
hyperlipidemia in mice, which may have been attributable to a species-specific response.

58
Figure 2-6. Role of GLP-1 in postprandial lipemia and sitagliptin action
(A and B): Acute effect of the GLP-1R agonist exendin-4 and antagonist exendin9-39 on apoB48
metabolism in chow-fed hamsters. (A) Representative FPLC profile of plasma from chow-fed
hamsters (n = 3) injected intraperitoneally with exendin-4 without intravenous Triton WR-1339,
2 h following an oral gavage of olive oil. (B) Hamsters were challenged with a fat load with
Triton WR-1339 administration. TRL apoB48 mass was measured by immunoblotting. Data is
presented normalized to baseline values for each treatment. n = 3 per group, **p<0.01, exendin
9-39 or exendin 9-39 + exendin-4 vs. control, *** p <0.001 exendin-4 vs. control as determined
by two-way ANOVA with Bonferroni post-test. (C and D): Glp1r-/-
mice or littermate control
mice fasted for 16 h were given orally 200 L of olive oil. 20 min after gavage, mice were
injected IV with Triton WR1339 (0.5 g/kg). Blood samples were collected prior to IV injection
(time 0), as well as 30, 60 and 90 min after injection for triglyceride (C) and ApoB48 (D)

59
measurement as described in methods, n = 6 per group, * p<0.05, ** p<0.01 as determined by
two-way ANOVA with Bonferroni post-test. (E and F): Mice were fasted for 5 h and
administered sitagliptin (10 mg/kg) by oral gavage. GLP-1R signalling was blocked by co-
administering the GLP-1R antagonist exendin9-39 by intraperitoneal injection. 20 min later, the
mice were challenged with an oral fat load and given Triton WR-1339 by intravenous injection.
(E) Cholesterol and (F) TG secretion as measured as described in methods. n = 5-6 per group, *
p<0.05, ** p<0.01 sitagliptin vs. control or sitagliptin + exendin9-39 as determined by two-way
ANOVA with Bonferroni post-test.

60
2.4.6 Exendin-4 directly reduces enterocyte ApoB48 secretion
We next evaluated the effect of exendin-4 treatment on enterocytes isolated from chow-fed
hamsters. Pulse chase studies in primary enterocytes showed that cellular apoB48 was not
changed in exendin-4-treated hamsters (Figure 2-7A). In contrast, exendin-4 decreased the
amount of secreted apoB48 from enterocytes of fructose-fed hamsters (Figure 2-7B) (p < 0.01 at
60 min, p < 0.001 at 90 min). Calculations of total [35
S]-apoB48 indicate there was significantly
less apoB48 production overall with exendin-4 (not shown). However, the total amount of [35
S]-
apoB48 at 90 min relative to the starting amount (at time 0) was not different between control
and exendin-4-treated cells (Figure 2-7C). Therefore, the decreased apoB48 secretion appeared
to be due to decreased overall apoB48 production, as opposed to enhanced apoB48 degradation.
Based on [35
S]methionine incorporation, viability and global protein synthesis was not different
between exendin-4 and control groups. Hence GLP-1R activation directly regulates intestinal
apoB48 secretion ex vivo.

61
Figure 2-7. Direct effects of exendin-4 on intestinal apoB48 metabolism ex vivo
Enterocytes from chow-fed hamsters were analyzed. Cellular (A) and secreted (B) levels of
apoB 48 were determined in primary enterocytes metabolically labelled with [35
S]methionine in
media containing 100 pM exendin-4. Direct treatment with exendin-4 significantly decreased
secretion in newly-synthesized apoB48. (C) Total [34
S]-apoB48 expressed relative to the
amount at the beginning of the chase. Open circles = control, closed circles = exendin-4, n = 3
per group, **p<0.01, ***p<0.001 as determined by two-way ANOVA with Bonferroni post-test.

62
2.5 Discussion
DPP-4 inhibitors exert their actions in part via augmentation of GLP-1 action, which
leads to reduction of glucagon, increases in insulin, and reduced glycemia in human subjects
(401; 402). Moreover, GLP-1 administration reduced postprandial circulating lipid levels in
human subjects (393), although the underlying mechanisms remain uncertain. In the current
study, we found that sitagliptin decreased TG accumulation, specifically in the VLDL fraction, in
both hamsters and mice. Sitagliptin treatment also decreased apoB48 accumulation in the
plasma, specifically in the intestinally produced TRL fraction. Treatment with DPP-4 inhibitors
can significantly decrease postprandial TG, cholesterol and apoB48 levels in humans (403; 404)
however, DPP-4 inhibition has minimal effects on fasting lipid levels (403). Our data indicate
that a prominent aspect of sitagliptin’s effects on fasting plasma TG levels is the decrease in
levels of VLDL. This suggests that the major effect of augmenting GLP-1 action is the reduction
in the number and/or size of large TRL particles.
Our observations demonstrate that both sitagliptin and exendin-4 exert similar effects on
postprandial lipid profiles, consistent with the notion that increased GLP-1 action is likely
responsible for the reduced circulating lipid levels and reduced intestinally-derived TRL
observed following sitagliptin administration. Furthermore, although increased levels of insulin
may also reduce plasma TG levels and apoB 48 secretion from the intestine (405) administration
of exogenous insulin did not affect intestinal lipid secretion in mice (Figure 2-5B), excluding
insulin as a mediator for sitagliptin or exendin-4 effects on post-prandial lipemia. Modulated
insulin sensitivity and glycemia was also unlikely to play a role in sitagliptin’s effects as insulin
sensitivity was not noted to change following 3 weeks of treatment. While fructose-fed hamsters
exhibit glucose intolerance, fasting hyperinsulinemia, and fasting hyperlipidemia, they have not
been observed to have fasting hyperglycemia (145). Though sitagliptin does augment bioactive
GLP-1 levels, GLP-1 stimulates insulin secretion in a glucose-dependent manner. As glucose
levels were not different between control and sitagliptin-treated groups, insulin and therefore
glucose were not expected to be significantly different in the fasting state. Moreover,
pharmacological levels of GLP-1R agonism can result in improved insulin sensitivity (339; 406),
disrupting endogenous GLP-1R signalling did not adversely affect glucose utilization (407).

63
However, the sitagliptin apoB48 effect appeared to be attenuated in fructose-fed hamsters
(Figure 2-4) probably because hamsters are are still insulin-resistant and oversecrete apoB48. As
sitagliptin did not improve insulin sensitivity, apoB48 secretion was still refractory to the insulin
arm of GLP-1 action (154).Therefore, extending the bioactivity of endogenous GLP-1 with
sitagliptin was unlikely to yield great improvements in insulin sensitivity.
We also present data clarifying the putative role of GIP or glucagon in the actions of
sitagliptin on intestinal lipid secretion. Although glucagon has been reported to inhibit lipid
secretion from hepatocytes (408), circulating levels of glucagon were reduced following
sitagliptin administration to mice. Furthermore D-Ala2-GIP did not reduce but actually promoted
postprandial TG and apoB48 secretion. These experiments, taken together with data using
exendin9-39 and Glp1r-/-
mice implicate GLP-1 as the predominant mediator of sitagliptin action
on intestinal apoB48 secretion.
Our data also demonstrated overlapping effects of sitagliptin and exendin-4 treatment on
plasma and TRL lipoprotein levels. Sitagliptin decreased TG levels in the TRL fraction of
fructose-fed hamsters. Similarly, exendin-4 decreased apoB48 secretion in chow-fed hamsters
and in freshly-isolated primary enterocyte cultures. A discrepancy in plasma cholesterol was
noted in unchanged total cholesterol Figure 2-1B and the decreased cholesterol across almost all
lipoprotein fractions in Figure 2-1D measured in sitagliptin-treated fructose-fed hamsters. This
discrepancy in fasting cholesterol levels needs to be verified, possibly by assessing hepatic
cholesterol secretion. These observations suggest for the first time that GLP-1 directly regulates
lipoprotein assembly and/or secretory machinery in the enterocyte. However, additional work is
required to understand the molecular mechanism underlying the suppressive effect of GLP-1 on
apoB48-TRL production.
An important aspect of our studies is the demonstration that reduction or elimination of
GLP-1 receptor signalling in the absence of exogenous administration of DPP-4 inhibitors or
GLP-1R agonists results in detectable changes in postprandial lipoprotein profiles. Specifically,
administration of exendin9-39 alone resulted in increased levels of TRL apoB48 in hamsters,
whereas levels of apoB48 and triglyceride mass were increased in Glp1r-/-
compared to Glp1r+/+

64
mice. Hence, these findings establish an essential role for basal levels of GLP-1R signaling in the
control of intestinal lipoprotein synthesis/secretion in vivo.
In summary, our data supports an important role for the GLP-1 receptor signaling system
in regulating intestinal lipid and lipoprotein metabolism. Augmentation of GLP-1R signaling
lowers postprandial circulating levels of TG and cholesterol-rich TRL, and apoB48-containing
chylomicrons. The current study suggests that enhanced GLP-1 action, achieved via DPP-4
inhibition or use of GLP-1R agonists, may contribute to control of postprandial lipid excursion
through control of intestinal lipoprotein synthesis and secretion.

65
Chapter 3: GLP -2 INCREASES INTESTINAL LIPID
ABSORPTION AND CHYLOMICRON PRODUCTION VIA
CD36
Text and figures in this chapter were reproduced from:
Hsieh J, Longuet C, Maida A, Bahrami J, Xu E, Baker CL, Brubaker PL, Drucker DJ, Adeli K.
Glucagon-like peptide-2 increases intestinal lipid absorption and chylomicron production via
CD36. Gastroenterology. 2009;137(3):997-1005. Copyright 2009, Elsevier.
Author contributions:
J. Hsieh generated all of the hamster and Cd36-.-
mice data with assistance. C. Longuet provided
the data presented in Figure 3-6. Data analysis and writing was completed by J. Hsieh.

66
3.1 Summary
Background: Excessive postprandial lipemia is a prevalent condition that results from intestinal
oversecretion of apolipoprotein B48 (apoB48)-containing lipoproteins. Glucagon-like peptide-2
(GLP-2) is a gastrointestinal-derived intestinotropic hormone that links nutrient absorption to
intestinal structure and function. We investigated the effects of GLP-2 on intestinal lipid
absorption and lipoprotein production.
Methods: Intestinal lipid absorption and chylomicron production were quantified in hamsters,
wild-type (WT) mice, and Cd36-/-
mice infused with exogenous GLP-2. Newly synthesized
apoB48 was metabolically labelled in primary hamster jejunal fragments. Fatty acid absorption
was measured and putative fatty acid transporters were assessed by immunoblotting.
Results: Human GLP-2 increased secretion of the triglyceride (TG)-rich lipoprotein (TRL)-
apoB48 following oral administration of olive oil to hamsters; TRL-TG and cholesterol mass
each increased 3-fold. Fast protein liquid chromatography (FPLC) profiling indicated that GLP-2
stimulated secretion of chylomicron/very low density lipoprotein (VLDL)-sized particles.
Moreover, GLP-2 directly stimulated apoB48 secretion in jejunal fragments cultured ex vivo,
increased expression of fully glycosylated CD36, and induced intestinal absorption of
[3H]triolein. The ability of GLP-2 to increase intestinal lipoprotein production was lost in Cd36
-/-
mice.
Conclusions: GLP-2 stimulates intestinal apoB48-containing lipoprotein secretion, possibly
through increased lipid uptake, through a pathway that requires CD36. These findings suggest
that GLP-2 represents a nutrient-dependent signal that regulates intestinal lipid absorption and
the assembly and secretion of TRLs from intestinal enterocytes.

67
3.2 Introduction
Postprandial lipemia, particularly excessive accumulation of intestinally-derived
lipoproteins, is gaining recognition as a cardiovascular risk factor since the magnitude of
postprandial lipemia correlates with intima media thickness (3). ApoB48 serves as the main
structural apolipoprotein of the buoyant, TG-rich chylomicrons that deliver dietary lipids to
tissues. Interestingly, chylomicron remnants and apoB48 particles have been detected in
atherosclerotic plaques (409). In addition to augmenting the risk of atherosclerosis, postprandial
hypertriglyceridemia is also an important facet of the metabolic dyslipidemia observed in insulin
resistance (410), and there is evidence to suggest that intestinal cholesterol absorption is
upregulated in type 1 diabetes (411).
Despite increasing awareness that intestinally-derived lipoproteins may contribute to the
pathophysiology of atherosclerosis, our understanding of the cellular pathways involved in
chylomicron production and secretion remains limited. Chylomicron assembly is proposed to
occur in a three step manner: a) assembly of the primordial lipoprotein during the translocation
of apoB48 into the ER, b) accumulation of TG-rich lipid droplets in the lumen of smooth ER,
and c) core expansion achieved through additional lipidation of the primordial particle as it
proceeds through the secretory pathway to generate a large TRL. As the nascent apoB48
polypeptide traverses the secretory system, its lipidation is achieved through the action of MTP
(68). The intestine is capable of secreting small, dense, lipid-poor apoB48-containing
lipoproteins (76), but the secretion of TRL is dependent on TG availability, which can be derived
from the diet or intracellular de novo lipogenesis.
The observation that insulin can acutely inhibit intestinal apoB48 secretion (154), and
that insulin resistance is associated with intestinal apoB48 overproduction (71; 72), suggests that
intestinal lipid metabolism is sensitive to endocrine signals. GLP-2 is a 33 amino acid peptide
co-secreted with GLP-1 from enteroendocrine L cells in response to carbohydrate and fat
ingestion, is an attractive candidate for such an endocrine signal. GLP-2 is highly
intestinotrophic and prevents the intestinal mucosal hypoplasia observed with parenteral nutrition
(reviewed in (412)). GLP-2 exerts its actions through the GLP-2R, a G-protein-coupled receptor
that exhibits a highly restricted specific pattern of expression in enteroendocrine cells (355),

68
enteric neurons (304), ISEMFs (354) and the central nervous system (282). GLP-2R activation
results in cAMP accumulation and protein kinase A activation (366). Notably, GLP-2R mRNA
or protein has not been detected in absorptive enterocytes. Thus, GLP-2’s proliferative and
cytoprotective actions on the intestinal epithelium must occur indirectly likely involving GLP-
2R-dependent stimulation of downstream mediators such as KGF (354) and IGF-1 (327).
GLP-2 also exerts rapid actions converging on stimulation of nutrient absorption in the
gut. GLP-2 enhances hexose transport through upregulation of SGLT-1 (312) and GLUT-2
(311) in the BBM. Intriguingly, long chain fatty acids stimulate secretion of proglucagon-
derived peptides from the gut (212) and acute GLP-2 administration enhances lipid absorption in
normal human subjects (284). Nevertheless, the mechanism(s) through which GLP-2 modulates
intestinal lipid secretion and/or metabolism remain poorly understood. In this study, we utilized
both Syrian Golden hamster and mouse models to characterize the effect of GLP-2 on intestinal
lipid metabolism. As hamsters secrete apoB48 exclusively from the intestine, this facilitates
delineation of intestinal contributions to circulating lipoprotein levels. We present new evidence
implicating GLP-2 as a potent stimulator of apoB48-containing TRL secretion from the intestine,
and describe potential mechanisms via which GLP-2 augments the TG pool for secretion.

69
3.3 Materials and Methods
3.3.1 Antibodies and chemicals
Rabbit polyclonal anti-human CD36 antibody (Cayman Chemical, Ann Arbor, MI),
rabbit polyclonal anti-human FATP4 (kind gift from Dr. Paul A. Watkins, John Hopkins
University), and rabbit anti-MTP antibody (kind gift of Dr. André Theriault, University of
Hawaii), were used for immunoblots of protein from isolated hamster enterocytes.
3.3.2 Animals
Male Syrian Golden hamsters (Mesocricetus auratus) from 130 to 150 g (Charles River,
Montreal, Quebec, Canada) were housed individually in a 12:12 h light:dark cycle. Animals
were fed ad libitum with a standard chow diet. Male Cd36-/-
mice bred on the C57BL/6
background from 12 to 15 weeks of age were obtained from Dr. Maria Febbraio, Cleveland
Clinic via Dr. Kevin Kain, University of Toronto. Following at least a 1 week acclimatization
period, animals underwent the in vivo protocols described below. All procedures were approved
by the Hospital for Sick Children and Toronto General Hospital Animal Care Committees.
C57BL/6 male mice 8-12 weeks old (Charles River, Montreal, Quebec, Canada) were
maintained on standard rodent chow under a normal 12:12 h light:dark cycle..
3.3.3 Determination of triglyceride-rich lipoprotein apoB48 secretion in vivo in
hamsters
The in vivo protocol for Syrian Golden hamsters was performed as previously described
(165). Briefly, the right jugular vein was cannulated with a silastic catheter (VWR), filled with
heparinized saline (40 IU/mL in 0.9% NaCl), and exteriorized at back of neck under isoflourane
anesthesia. Animals were allowed to recover overnight and henceforth experiments were
performed on conscious hamsters. Following a 16 h overnight fast, a 400 L blood sample was
collected into heparinized tubes (Microtainer® PST tubes with lithium heparin, BD, Franklin
Lakes, NJ) from the jugular catheter as the baseline reading and succeeded by an oral gavage of
200 L olive oil. 20 min following the gavage, the hamster was administered human GLP-2(1-33)
(0.25 mg/kg) (Bachem Bioscience Inc, King of Prussia, PA) by intraperitoneal injection. This
dose of GLP-2 was a pharmacological dose. A subcutaneous injection of 5 g GLP-2 to mice

70
(approximately 0.19 mg/kg) induced jejunal crypt proliferation within 2 h of administration
(288), but the dose was increased as subcutaneous injections were more efficient at stimulating
an intestinotrophic response than an intraperitoneal injection (413). Following immediately, an
intravenous bolus of Triton WR-1339 (20% w/v in saline, 0.5 g/kg) (Tyloxapol, Sigma-Aldrich
Co., St. Louis, MO) was given. 400 L of blood was sampled at 30 min intervals until 120 min
post-gavage, at which point the intestine was excised under isoflurane anaesthesia and the
hamster was sacrificed.
3.3.4 Determination of TG-rich lipoprotein apoB48 secretion in vivo in mice
Analysis of intestinal triglyceride and ApoB48 secretion in mice was performed
following a similar procedure. After a 5 h fast, mice were administered 200 L olive oil by
gavage, and 20 min later Triton WR-1339 (15% in saline, 0.5 g/kg) with or without the GLP2
analogue hGly2-GLP-2 (0.25 mg/kg body weight) was injected intravenously (IV). 50 L blood
samples were collected via the tail vein prior to (time 0), 30 and 60 min after IV injection for
determination of plasma triglyceride and ApoB48. 90 min after IV injection, mice were
sacrificed and blood was collected by cardiac puncture.
3.3.5 Isolation of TRL
The plasma layer was separated by centrifuging the blood for 15 min in 4°C at 5,000
rpm. The plasma was supplemented with sodium azide and a cocktail of protease inhibitors
(Roche Diagnostics, Mannheim, Germany), and 150 L was layered under 4 mL of 1.006 g/mL
potassium bromide solution. The layered plasma was centrifuged at 10°C for 70 min at 35,000
rpm using a SW55Ti rotor (Beckmann Coulter, Mississauga, ON, Canada). The top 300 L was
collected as the triglyceride-rich lipoprotein (TRL) fraction.
3.3.6 FPLC of plasma lipoproteins
Blood was collected by cardiac puncture 2 hr following a gavage of 200 L olive oil.
200 L of plasma was filtered through a 0.45 M micro-spin polysulphone filter (Alltech,
Mandel Scientific, Lachine, PQ) and separated according to size through a HR 10/300 GL
Superose 6 column (Pharmacia, Uppsala, Sweden). A solution of 10 mM Tris, 150 mM NaCl, 2
mM CaCl2, 100 DTPA, 0.02% NaN3, pH 7.4 was pumped through at a flow rate of 0.5

71
mL/min and 0.5 mL fractions were collected. TG and cholesterol concentration of the fractions
were ascertained with one-step colorimetric assays (Roche Diagnostics).
3.3.7 Density ultracentrifugation of plasma lipoproteins
150 L plasma obtained from hamsters 40 min after fat loading was subject to density
ultracentrifugation over a KBr gradient as previously described for microsome luminal contents
(414). Concentrations of TG and total cholesterol (Roche Diagnostics) and free cholesterol and
phospholipids (Wako Diagnostics, Osaka, Japan) were assayed in each fraction using enzymatic-
based colorimetric assays.
3.3.8 Metabolic labelling of primary jejunal fragments ex vivo
Hamsters were given an oral gavage of 200 L olive oil. 60 min later, the jejunum was
excised under isoflurane anesthesia and the hamster was sacrificed. The jejunum was flushed
with ice-cold phosphate buffered saline (PBS, pH 7.4) and sliced longitudinally to reveal the
mucosa. 0.5 mm long fragments of jejunum were divided randomly among wells, with the
mucosal side facing up. The fragments were pre-pulsed for 45 min in DMEM lacking
methionine and cysteine and supplemented with 1% L-glutamine, 10% FBS, 1%
penicillin/streptomycin, and with/without the addition of 100 nM human GLP-2(1-33)
. The
fragments were then radioactively labeled with 225 Ci of [35
S]methionine for 30 min. After
washing with PBS, the fragments were chased in 40 mM methionine-enriched DMEM for 30, 60
and 90 min, following which tissues were homogenized with a Polytron homogenizer and media
were collected. Immunoprecipitation and SDS-PAGE was performed as previously described
(71). ATP binding cassette transporter G5 (ABCG5) was immunoprecipitated with a rabbit anti-
ABCG5 antibody (Santa Cruz Biotechnology, CA). Following SDS-PAGE, the gel was dried
and exposed to a phosphor imaging screen (Molecular Diagnostics). The screen was then read
with the Storm 840 Phosphorimager (Amersham).
3.3.9 Monitoring labelled triolein secretion in vivo
Hamsters with a jugular catheter were gavaged with 3 Ci [9,10-3H(N)] triolein mixed in
with 200 L olive oil and then injected intraperitoneally with 0.25 mg/kg GLP-2 20 min later.
Labelled triolein was used to ensure efficient absorption and reproducibility (415). 200 L of

72
blood was sampled from the jugular catheter into heparinized tubes at half hour intervals. The
activity of tritium in 20 L of plasma was determined by scintillation counting in triplicate.
After an overnight fast, mice were administered 3 Ci [9,10-3H(N)]triolein mixed in with 200 L
Intralipid (10%) and 50 L blood samples were collected via the tail vein.
3.3.10 In situ apical membrane protein biotinylation
To specifically label enterocyte brush border membrane proteins with biotin, intact
proximal jejunum was filled with a solution of 1.5 mg/mL sulfo-NHS-SS-Biotin (Pierce,
Rockford, IL) as described in (311). Enterocytes were isolated as previously described (71), and
lysed in 1% Triton X-100, 150 mmol/L NaCl, 50 mmol/L EDTA, 50 mmol/L Tris, pH 7.5.
Biotinylated proteins were pulled down by incubating 0.75 mg of total protein with 50 L
streptavidin agarose beads (Pierce, Rockford, IL) at 4°C overnight. The beads were washed, and
the biotin cleaved from the protein by boiling the sample in 8% SDS containing -
mercaptoethanol and dithiothreitol.
3.3.11 Statistical analysis
Two-way repeated measures ANOVA with the Bonferroni post-hoc analysis was performed on
time course experiments. For enterocyte expression of proteins and lipid synthesis assays, a
paired t-test was used. Results are expressed as mean ± SEM.

73
3.4 Results
3.4.1 GLP-2 acutely increases circulating levels of apoB48-containing TRL in
hamsters
Hamsters were administered human GLP-2(1-33)
(0.25 mg/kg) by intraperitoneal injection,
challenged with a fat load and the apoB48 mass in the TRL fraction of the plasma was monitored
by immunoblotting. An intravenous bolus of Triton WR-1339 was also introduced to inhibit
lipoprotein catabolism, thus maintaining the buoyancy of newly-secreted chylomicrons. As
shown in the photograph in Figure 3-1A, treatment with GLP-2 resulted in a marked lipemia that
was visibly noticeable, especially by 120 min post-gavage. Quantification of the apoB48 mass
by immunoblotting demonstrated that GLP-2 increased the mass of circulating apoB48-
containing TRL by 60 minutes (Figure 3-1B). The slopes were 4.36 ± 0.75 %·min-1
and 2.57 ±
0.9 %·min-1
for GLP-2- vs. vehicle-treated hamsters, p = 0.11 for slope difference, but p = 0.007
for elevation difference.
We next assessed whether GLP-2 produces changes in intestinal lipoproteins by both
FPLC analysis and salt-density sedimentation of plasma from hamsters challenged with a fat
load, but not given a bolus of Triton WR-1339. FPLC separates lipoproteins according to size
via gel filtration. 120 min after the gavage, levels of TG and cholesterol were higher in the
VLDL/chylomicron remnant-sized fractions in plasma from GLP-2 treated hamsters (Figure 3-
1C). No appreciable differences were observed in the LDL and HDL fractions, suggesting that
GLP-2’s stimulatory effect on circulating lipoprotein levels is limited largely to the TRL.
Complete profiling of plasma lipoproteins by sedimentation yielded similar observations, with
higher TG and free cholesterol concentrations in the large chylomicron, small chylomicron, and
large VLDL fractions isolated from GLP-2 treated hamster plasma (Figure 3-2). Free cholesterol
concentrations appeared higher than total cholesterol concentrations due to detection limit of the
assay kit. The concentration of TG in the TRL fraction of GLP-2-treated hamsters was 3-fold
higher than in control hamsters (Figure 3-1D). TRL cholesterol was also 3-fold higher in GLP-
2-treated hamsters (Figure 3-1E).
Metabolic labelling of primary hamster jejunal segments with [35
S]methionine was
performed to see if these in vivo observations could be recapitulated ex vivo. To rule out the

74
possibility that the effect of GLP2 on specific protein synthesis may be a result of GLP-2-
enhanced enterocyte survival, ApoB48 and ABCG5 synthesis data were normalized to total
protein synthesis. Addition of GLP-2 to the incubation media did not affect cellular levels of
apoB48 (Figure 3-3A), but resulted in significantly enhanced secretion of newly-synthesized
apoB48 (Figure 3-3B) that is reflected in significantly greater total levels of [35
S]-labelled
apoB48 (Figure 3-3C). This increase in apoB48 synthesis was specific, as GLP-2 did not
significantly affect ATP-binding cassette transporter G5 (ABCG5) synthesis (Figure 3-3D).
Percent viability of the absorptive enterocytes themselves could not be provided but based on
[35
S]methionine incorporation into proteins, control-treated fragments had 3.68 ±0.14 x 105
dpm/mg fragment while GLP-2-treated fragments had 5.25 ± 1.39 x 105 dpm/mg.

75
Figure 3-1. In vivo effects of GLP-2 on apoB48-containing TRL secretion
(A) Plasma collected from hamsters challenged with an oral gavage of olive oil coupled with an
intravenous bolus of Triton is shown. The time points represent the minutes after the initial fat
load. (B) Time-dependent increase of apoB48 mass in the plasma TRL fraction from chow-fed
hamsters given an oral olive oil gavage and Triton infusion, as determined by immunoblotting.
ApoB48 mass was normalized to baseline fasting levels determined immediately prior to the oil
gavage (time 0) and before GLP-2 injection (time 20 min). Representative apoB48 blots are
shown above the graphical representation. n = 7 per group, * p < 0.05. (C) Representative
FPLC lipoprotein profile of plasma from fat-loaded hamsters not given an intravenous bolus of
Triton WR-1339. Profiles are representative of 3 FPLC runs per group. (D) TG and (E)
cholesterol concentrations of the plasma TRL fraction, 120 min following fat load and without
Triton infusion. n = 4 per group, * p < 0.05.

76
Figure 3-2. Plasma lipoprotein profiling by salt-density sedimentation in hamsters
Hamsters were given an oral gavage of olive oil and followed by injection with GLP-2 or
PBS 20 min later, and no Triton WR-1339 was used in this experiment. Profiling was performed
on plasma collected 20 min after peptide injection. Representative of 3 density profiles depicting
TG (upper left), total cholesterol (lower left), free cholesterol (upper right), and phospholipids
(lower right) content of plasma lipoproteins separated by ultracentrifugation on a discontinuous
KBr gradient. Fractions are arranged as numbers 1 to 10 in order of increasing density.

77
Figure 3-3. Ex vivo effects of GLP-2 on apoB48-containing lipoprotein secretion
[35
S]-labeled apoB48 was measured during the cold methionine-enriched chase after a 30
min pulse in jejunal fragments freshly isolated from chow-fed hamsters and treated with or
without GLP-2 ex vivo, and normalized to total protein synthesis. Time ‘0’ refers to the end of
[35
S]methionine labeling and beginning of chase. (A) Intracellular levels of 35
S-apoB48 (B)
Secreted 35
S-apoB48 (C) Total levels of 35
S-apoB48. n=3 per group; * p < 0.05, *** p < 0.001.
(D) Cellular 35
S-ABCG5. n = 4 per group.
*

78
3.4.2 GLP-2 accelerates secretion of luminal fatty acids
To understand the mechanism by which GLP-2 induces apoB48-lipoprotein secretion, we
next examined the handling of luminally-derived FA. As a surrogate measure of dietary FA
transport, hamsters were challenged with a fat load that contained 3 Ci of [9,10-3H(N)]triolein,
but without Triton WR-1339 injection. By 60 min after the gavage, there was a 2-fold increase
in 3H radioactivity in the plasma of GLP-2-treated hamsters (Figure 3-4A). The majority of the
tritium label was incorporated into secreted TG, which accounted for the GLP-2-induced
increase in plasma 3H radioactivity at both 60 (Figure 3-4B) and 90 min (Figure 3-4C). No
difference in total intestinal 3H counts was detected (data not shown).
The expression of specific protein transporters implicated in intestinal fatty acid
absorption was examined by Western blot analyses. No difference was noted in the protein
levels of fatty acid transporter 4 (FATP4) (Figure 3-5A); similarly, intestinal MTP protein
expression was found not to change with acute GLP-2 treatment (Figure 3-5B). However, GLP-
2 significantly increased the expression of glycosylated CD36 (Figure 3-5C).
Immunohistochemical visualization of CD36 suggests this protein is expressed on hamster
intestinal villi tips and appears to be present on the apical membrane and intracellularly (Figure
3-5D), poised for luminal fatty acid uptake. To verify that high molecular weight CD36 is the
isoform that is expressed on the enterocyte apical membrane, a cell membrane-impermeable
biotinylating agent (sulfo-NHS-SS-biotin) was introduced into the intestinal lumens of both
control and GLP-2-treated hamsters in situ to specifically label apically expressed proteins of the
epithelium. As shown in the “PD” lane of Figure 3-5E, streptavidin affinity pulldown of
biotinylated proteins revealed only a higher molecular weight protein slightly larger than 88kDa
in enterocytes from both control and GLP-2-treated hamsters. However, this isoform of CD36
was not prominent in the supernatant and whole cell lysate samples under both conditions. Such
glycosylation behaviour of CD36 was observed in 3 animals per treatment group. Treatment
with PNGase F to remove all N-linked oligosaccharides yielded only the lower molecular weight
band across all lanes. Thus, a potential mechanism by which GLP-2 may promote fat absorption
is through increasing apical CD36 by means of advanced glycosylation.

79
Figure 3-4. GLP-2 accelerates secretion of luminal fatty acids in hamsters
(A) Rate of appearance of radioactivity in plasma following an oral gavage of 3 Ci [9,10-
3H(N)]triolein in 200L olive oil without an intravenous bolus of Triton WR-1339. n = 7 per
group, ** p < 0.01. Thin layer chromatography was performed on the hexane-soluble phase of
plasma to characterize the incorporation of the 3H label into secreted lipids at (B) 60 min and (C)
90 min. n = 4 per group, ** p < 0.01, *** p < 0.001.

80
Figure 3-5. GLP-2 enhances intestinal CD36 action in hamsters
Hamsters were given an oral gavage of olive oil and then injected with GLP-2. Enterocytes were
collected 60 min later for immunoblots. (A) FATP4 expression in hamster enterocytes. FATP4
immunoblot is shown above graphical representation. FATP4 expression was normalized to
cellular levels of -actin. n = 4 per group. (B) Immunoblot analysis of MTP protein in
enterocytes from hamsters treated with GLP-2 in vivo in comparison to control, normalized to -
actin. Immunoblots of individual animals in duplicate shown above. n = 3 per group. (C) Ratio
of fully glycosylated CD36 to unglycosylated CD36 as determined by immunoblotting. The top
band was visualized after a longer exposure than the bottom band. n = 4 per group, * p < 0.05.
(D) Immunohistochemistry of hamster intestinal villi CD36 expression with an anti-CD36
antibody, visualized with diaminobenzidine (DAB) deposition (brown) and counter-stained with
hematoxylin and eosin. (E) Intestinal apical membrane proteins were biotinylated in situ, subject
to streptavidin pulldown, and probed with anti-CD36 antibody by immunblotting. The
biotinylated proteins were also treated with PNGase F for 120 min. PD, biotinylated intestinal
apical proteins pulled down with streptavidin; Sup, supernatant after streptavidin pulldown;
WCL, whole cell lysate previous to addition of streptavidin-agarose beads. Molecular weight
markers are indicated on the left.

81
3.4.3 Acute GLP-2 treatment increases apoB48-containing TRL production in
mice
To determine whether GLP-2 exerts similar actions on intestinal lipid absorption and lipoprotein
production in mice, experiments were performed in wild type mice with hGly2-GLP-2, a human
DPP-4-resistant GLP-2 analogue. hGly2-GLP-2 rapidly increased the accumulation of plasma
TG in the presence of Triton WR-1339 (Figure 3-6A) in a linear manner (R2 = 0.9918 for control
mice and 0.9966 for GLP-2-treated mice), as well as the levels of TG and apoB48 in TRL
fraction (Figures 3-6B and 3-6C) in wild type mice after an oral gavage of olive oil in the
presence of Triton WR-1339. Furthermore, FPLC profiling showed that exogenous GLP-2
increased TG levels in the VLDL/chylomicron remnant fractions when lipoprotein clearance was
not inhibited (Figure 3-6D). These experiments demonstrate that the stimulatory effects of GLP-
2 on intestinal lipid absorption and chylomicron secretion are conserved in different species.

82
Figure 3-6. Stimulatory effects of GLP-2 on intestinal lipid absorption and apoB48-
chylomicron production in wild type mice
(A) Plasma TG accumulation after a fat load (200 L olive oil) and IV Triton WR-1339 injection
in wild type mice fasted for 5 h with or without Gly2-GLP-2 as described in methods. n = 4-6, **
p < 0.01. (B) TG and (C) apoB48 accumulation in TRL fraction purified 90 minutes after triton
WR-1339 injection as described in methods. n = 4-6 per group, ** p < 0.01. (D) Lipoprotein
profile of plasma pooled from 6 mice per group 90 min after the fat load.

83
3.4.4 GLP-2-stimulated chylomicron secretion requires CD36
The immunoblots shown in Figure 3-5 suggested that GLP-2 rapidly promotes intestinal
fat uptake by regulating the trafficking of CD36. To determine whether CD36 is required for
GLP-2-stimulated intestinal lipid absorption, we assessed GLP-2 action in Cd36-/-
mice.
Exogenous GLP-2 had no effect on levels of plasma triglycerides after an oral fat load in Cd36-/-
mice (Figure 3-7A). The time-dependent increase in plasma apoB48 following Triton WR-1339
injection indicate that Cd36-/-
mice are indeed competent in secreting postprandial apoB48-
containing lipoproteins (Figure 3-7A). Consistent with the lack of GLP-2 action on TG
absorption in Cd36-/-
mice, no significant differences were noticed among TG (Figure 3-7B),
cholesterol (Figure 3-7C), and apoB48 mass (Figure 3-7D) measurements on the TRL fraction,
even though there were significant increases in TRL-apoB48 in their littermate controls. Hence,
the actions of GLP-2 on the stimulation of chylomicron secretion require CD36. Additionally,
Gly2-GLP-2 failed to augment luminal [
3H]triolein uptake in Cd36
-/- mice, (Figure 3-7E).
As quantified by scoring ten different histological sections of the small intestine, there
was negligible oil red O staining in sections of proximal intestine from Cd36-/-
mice (Figure 3-
7F), consistent with observations of Nassir et al (19). There was however abundant oil red O
staining in the distal jejunum of the Cd36-/-
mice (data not shown), which could explain why
these mice were competent in TG secretion despite the lack of neutral lipid detected in their
proximal intestine. Finally, we observed increased lipid accumulation in the enterocytes of GLP-
2-treated wild type but not Cd36-/-
mice (Figure 3-7F).

84
Figure 3-7. Lack of GLP-2 mediated stimulation of intestinal lipoprotein secretion in Cd36-
/- mice
Cd36-/-
mice bred on the C57BL/6 background were challenged with an oral fat load, injected
intraperitoneally with hGLP-2(1-33)
and given an intravenous bolus of Triton WR-1339 to inhibit
lipoprotein catabolism. (A) Plasma TG following an oral fat load. n = 3-4 per group, * p <
0.05, wild type GLP-2 vs. wild type control, # p < 0.05, wild type GLP-2 vs. Cd36-/-
control. (B)
TG mass, (C) Cholesterol mass and (D) apoB48 mass in the TRL fraction; n = 3-4 per group, * p
< 0.05, wild type GLP-2 vs. wild type control, ** p < 0.01, wild type GLP-2 vs. Cd36-/-
groups.
(E) Plasma tritium levels in response to an oral gavage of 3 Ci [9,10-3H(N)]triolein in 200 L
olive oil, Gly2-GLP-2 injection, and no Triton WR-1339; n = 5 per group. (F) Representative

85
oil red O stains of the proximal intestine 90 min following an oral gavage, with quantification of
average area stained by oil red O, based on 10 separate fields of view per animal. n = 3 per
group, *** p < 0.001, Cd36-/-
mice vs. wild type mice.

86
3.5 Discussion
There is growing evidence that, in addition to its intestinotrophic effects, GLP-2 may act
as an endocrine signal controlling intestinal nutrient absorption. Although it has been previously
demonstrated that chronic GLP-2 administration enhanced [3H]triolein uptake into the circulation
(310), there is limited data available as to the role of GLP-2 in regulating acute intestinal
absorption, packaging, and secretion of chylomicron lipids. Long chain fatty acids have been
shown to regulate proglucagon-derived peptide secretion from ileal enteroendocrine L cells
(212), suggesting that these cells may act as important postprandial sensors for the presence of
luminal fatty acids and secrete factors such as GLP-1/GLP-2 to regulate intestinal lipid
absorption and/or metabolism. Data presented in this thesis establish a role for GLP-2 in the
regulation of intestinal lipid and lipoprotein metabolism. GLP-2 was found to acutely and rapidly
bring about changes in the intestine that ultimately result in an exaggerated postprandial lipemia.
The most pronounced changes in plasma lipids were noticed in the TRL fraction of circulating
lipoproteins, both in apoB48 mass and lipid content, which indicates that GLP-2 promotes the
lipidation of nascent apoB48 polypeptides. These data provide a mechanistic explanation for the
rise in postprandial TG observed in human subjects infused with GLP-2 (284). Since no change
was noted in MTP mass of GLP-2-treated enterocytes, GLP-2 likely acts to stimulate
chylomicron assembly and secretion by providing more lipid substrate for lipidation of the
growing chylomicron particle.
Increased GLP-2-stimulated packaging and secretion of diet-derived fatty acids as
chylomicrons was observed, as indicated by results from the [3H]triolein experiment. Since
GLP-2 has not been shown to affect pancreatic exocrine function (416), changes in pancreatic
lipase levels to accelerate the hydrolysis of luminal TG is likely not a contributing factor to GLP-
2-enhanced dietary lipid transport. Though Triton WR-1339 was not included in this
experiment, the elevation in plasma tritium was unlikely due to inhibited catabolism. GLP-2
infusion in humans has been reported to modestly elevate postprandial TG following a low
calorie test meal without a change in glycerol levels (284), signifying that changes in lipolysis
did not occur. Evidence is amassing that long chain fatty acid transport across the intestinal
BBM is protein-mediated. The role of FATP4 was considered, but no difference in enterocyte

87
FATP4 protein expression was noted between control and GLP-2-treated hamsters. Although
FATP4 has been proposed as an important mediator of intestinal fatty absorption (36), recent
evidence suggests that FATP4’s role lies within its acyl CoA-synthetase activity at the ER rather
than as a solute carrier at the BBM (37).
Our data suggest that GLP-2-accelerated secretion of dietary lipid can be attributed at
least in part to increased expression of fully glycosylated CD36. The highly glycosylated CD36
appears to be the isoform exclusively expressed on the enterocyte apical membrane as
determined by in situ biotinylation. CD36/fatty acid translocase, is essential for chylomicron
secretion (23), and appears to contribute significantly to intestinal cholesterol absorption (21).
Studies in CD36-deficient mice also established this scavenger receptor as an intestinal long
chain fatty acid transporter (22). It has been suggested that the carbohydrate moieties of CD36
are important for its subcellular localization. Deletions in the C-terminus of CD36
downregulates its expression on the cell surface, and this was accompanied by reduced
glycosylation (417). The effect of GLP-2 on CD36 was rapid, detected within 40 minutes of
peptide injection. Interestingly, CD36 translocation to the cell surface from intracellular
membranes is a process sensitive to metabolic signals in cardiac myocytes (418). GLP-2 has
previously been documented to influence the trafficking of another nutrient transporter, SGLT-1
from intracellular stores to the BBM of enterocytes (312).
Since Western blot analysis of enterocytes suggests that GLP-2 modifies CD36
glycosylation, studies in Cd36-/-
mice were performed to confirm the transporter’s role. In
contrast to the stimulatory effects of GLP-2 observed in wild type mice, the actions of GLP-2 on
intestinal lipid and lipoprotein secretion were absent in Cd36-/-
mice. The rate of TG,
cholesterol, and apoB48 secretion in GLP-2 and control-treated Cd36-/-
mice were all
superimposable. Since secretion rates were so similar to control-treated wild type mice, it
appears that CD36 is required for hormone-stimulated chylomicron secretion as opposed to being
essential in the basal uptake and secretion of dietary lipid. There is no evidence of fat
malabsorption in CD36-deficient mice based on fecal analysis (21), indicating that these mice are
competent in dietary FA absorption. TRL TG and apoB48 was higher in Cd36-/-
mice compared
to wild type (Figure 3-7), but this may be attributable to this mouse model’s elevated fasting
VLDL (93), due to impaired clearance of TG-rich VLDL-sized (96) and chylomicron-sized (419)

88
particles. Impairment in lipoprotein lipase-independent clearance pathways could also explain
the elevated TRL TG and apoB48 observed in Cd36-/-
mice (420). Enteroendocrine-regulated
post-translational modification of CD36 in the intestine may contribute to mechanisms
controlling lipid absorption. However, the pathways regulating CD36 movement to the apical
membrane in enterocytes have yet to be elucidated, and the intracellular compartments in which
CD36 resides are still unidentified. Interestingly, a large portion of the enterocyte’s CD36 is not
found at the plasma membrane, as shown by the strong signal of the 43-55 kDa-sized band in the
supernatant and whole cell lysate lanes of Figure 3-5E. CD36 has also been localized to the
Golgi apparatus in adipocytes (421), like MTP (422), so it may play a role in further lipidation of
the apoB48-containing particle in that compartment. Cd36-/-
mice have been reported to
accumulate neutral lipids in enterocytes during a high fat load (23), hinting at the importance of
intracellular movement of CD36 to target fatty acids for assembly and secretion as chylomicrons.
Figure 3-1B suggests there is an inflection point in increased TRL-apoB48 secretion in
GLP-2-treated hamsters at 60 min, after which the difference between the two groups
diminished, unlike the observations made in wild type mice in Figure 3-6. Since DPP-4-
sensitive native GLP-2 was administered to the hamsters, while DPP-4-resistant Gly2-GLP-2 was
used in mice in Figure 3-6, it can be inferred that sustained GLP-2 action is necessary to
maintain upregulated chylomicron secretion. In addition, given that GLP-2 resulted in an
inflection in TG secretion between 40 min and 60 min in both wild type and Cd36-/-
mice (Figure
3-7A), and the results from hepatic secretion studies (data not shown) refute differential hepatic
TG secretion, it suggests these kinetics of lipoprotein secretion may be a CD36-independent
actions of GLP-2 that can modulate fat absorption in vivo, perhaps due to alterations in gastric
(372) and intestinal (338) motility.
The striking absence of lipid accumulation in the proximal intestine following a fat load
is consistent with the observations of Nassir et al, who observed a 50% reduction in triolein-
derived lipid accumulation in the proximal mucosa 90 min after a fat load (19). On the other
hand, the lipid accumulation in the proximal intestine noted by Drover et al was observed
following administration of a FA analog, [125
I]15-(p-iodophenyl)-3-(R,S)-methyl pentadecanoic
acid (BMIPP), that is slowly oxidized (23). Moreover, CD36’s essentiality in uptake was
concluded for cholesterol and very long chain FA, two lipids that typically have less efficient

89
rates of absorption (21; 22). The role of CD36 therefore appears to differ with certain lipid
species, but this data implicates a role for CD36 in handling FA that are more abundant in the
diet in response to GLP-2. Since [3H]triolein was administered in vivo, a number of steps from
apical FA uptake to chylomicron assembly could contribute to the modulated appearance of
tritium in the plasma. Therefore, additional experiments are necessary to definitively test apical
FA uptake. One experiment could involve assessing the accumulation of labeled FA in isolated
brush border membrane vesicles. Another approach could involve gavaging [3H]triolein in vivo
while blocking chylomicron secretion with Pluronic L-81 administration (423), followed by
measuring intestinal tritium accumulation.
In conclusion, our data demonstrates that GLP-2 promotes assembly and secretion of
intestinally-derived apoB48-containing TRL through accelerated dietary fat absorption and
increased lipidation and secretion of apoB48-containing chylomicron particles. This suggests
that GLP-2 may facilitate the efficient absorption of dietary fats. The potent stimulatory role of
GLP-2 raises the intriguing possibility of a link between GLP-2 function and postprandial
dyslipidemia observed in conditions such as type 1 diabetes. Elevated levels of GLP-2 have been
reported and implicated in mediating the intestinal hyperplasia in the STZ-induced diabetic rat
model (230). This same animal model also exhibits excessive postprandial lipemia (424). GLP-
2 may thus provide a possible endocrine explanation for the intestinal apoB48-containing
lipoprotein oversecretion observed in pathological conditions.

90
Chapter 4: INTERCELLULAR COMMUNICATIONS
MEDIATING GLP-2-STIMULATED CHYLOMICRON
SECRETION
Author contributions:
With the exception of ISEMF treatment and conditioned media collection, all data was generated
by J. Hsieh. ISEMF treatment and conditioned media collection was performed by A. Izzo in Dr.
P. Brubaker’s lab.

91
4.1 Summary
Background: Glucagon-like peptide-2 (GLP-2) has been shown to stimulate TRL-apoB48 output
by promoting the uptake of dietary fatty acids (FA) via the action of CD36. However, the GLP-2
receptor (GLP-2R) is not expressed on the enterocyte, indicating that the hormone signals to the
intestinal epithelium indirectly via a paracrine mediator. We postulated that nitric oxide and
vascular endothelial growth factor (VEGF) may mediate the GLP-2 effects on enterocyte
function.
Methods: Syrian golden hamsters were pre-treated with L-NG-nitroargninine methyl ester (L-
NAME) and challenged to an oral fat load along with a GLP-2 injection to assess postprandial
TRL secretion. Similar studies were performed on endothelial nitric oxide synthase (eNOS)
knockout mice. Primary murine enterocytes were cultured in GLP-2-stimulated murine ISEMF-
conditioned media and apoB48 synthesis was steady-state labelled with [35
S]methionine.
Primary rodent enterocytes were also subjected to a pulse chase study in the presence of S-
nitroso-L-glutathione (GSNO) or vascular endothelial growth factor (VEGF).
Results: L-NAME pre-treatment blocked the stimulatory effect of GLP-2 on TRL-apoB48
particle number and dietary FA uptake, and significantly reduced total lipid mass in the TRL
fraction. eNOS-deficient mice were also resistant to the stimulatory effects of GLP-2 and
secreted smaller apoB48-containing TRL particles. Both GSNO and VEGF promoted apoB48
synthesis and secretion in primary enterocytes. Primary enterocytes also upregulated apoB48
production when cultured in GLP-2-stimulated ISEMF-conditioned media, a change that
diminished when a VEGF-neutralizing antibody was included in the media.
Conclusions: GLP-2 produced by enteroendocrine L-cells agonizes its receptor on ISEMFs,
which secrete VEGF as a paracrine factor that participates in stimulating enterocytes to secrete
chylomicrons. The generation of nitric oxide is important in this relay of intercellular
communication to fully lipidate apoB48 and stimulate TG-rich chylomicron particles.

92
4.2 Introduction
Postprandial TG-rich apoB48-containing lipoproteins are increasingly acknowledged for
their atherogenic role (8; 425-427), which renders their elevation a cause for concern. Intestinal
chylomicron overproduction has been recognized as a contributor to postprandial dyslipidemia in
insulin resistance and diabetes in humans and multiple animal models (71; 72; 152; 158; 428).
However, intestinal lipoprotein secretion has historically been regarded as a passive process and
consequently our understanding of the regulation of apoB48 secretion has lagged behind that of
hepatic apoB100 metabolism. Recently, it has become clear that intestinal apoB48 production is
acutely sensitive to endocrine signals including insulin (154; 163), GLP-1 (394; 429), and GLP-2
(430), but the mechanisms behind these observations are largely unknown. GLP-2 has been
noted to accelerate the uptake of dietary FA through upregulating the posttranslational
glycosylation of CD36 in the enterocyte, but the sequence of cellular events leading up to the
modification of CD36 have yet to be elucidated (430).
In addition to its well-characterized intestinotrophic actions (431), GLP-2 stimulates the
uptake of lipids, as noted above, and sugars in the gut (312; 314). However, the G protein-
coupled receptor for GLP-2 has a conspicuous general absence in the intestinal epithelium, the
site of most of its observed biological actions to date, but rather has been identified on ISEMFs
(354), specific enteroendocrine cells (304; 355), enteric neurons (304; 357), and the vagal
afferent nerve (432). Therefore, GLP-2 must bind its receptor on a certain intermediary cell,
stimulating it to secrete a mediating signal that would act directly on the enterocyte to bring
about chylomicron overproduction. Among others, IGF-1 (433), KGF (354), VEGF (434), and
EGF (328) have been suggested as mediators of the proliferative and protective effects of GLP-2
on the intestinal epithelium. We have demonstrated that intact fragments of jejunum treated with
GLP-2 exhibited increased newly-synthesized apoB48 secretion ex vivo (430), so ostensibly such
an intermediary cell would lie in close apposition to the absorptive enterocyte.
One biological action of GLP-2 is stimulating mesenteric blood flow in a NO-dependent
mechanism (303; 306). GLP-2’s ability to increase blood flow may have implications for
intestinal lipoprotein metabolism, given that apoB48 secretion can be induced by circulating
FFA (164; 435), and increased blood volume at the basolateral side of the epithelium may serve

93
to increase FA delivery to the enterocyte. However, it has been noted that GLP-2-stimulated
intestinal glucose uptake can also be blocked by the pan-specific NOS inhibitor L-NAME to an
extent that cannot be completely explained by changes in circulation, suggesting a direct role for
NO in enterocyte nutrient handling (303). GLP-2 has been shown to acutely stimulate intestinal
eNOS (304), and there is evidence that eNOS is a NOS isoform expressed in the intestinal
mucosa (436). A hormone that can stimulate NO production is VEGF, which has been shown to
be secreted by intestinal subepithelial myofibroblasts (434). VEGF is best known for its vascular
angiogenic effects, but has also been shown to play a role in lymphoangiogenesis (437). There
are multiple members in the VEGF family, with VEGF-A being ubiquitously expressed. VEGF-
A binds both VEGF receptor 1 (VEGFR1) and VEGFR2 as a homodimer, and exists in multiple
isoforms in which C-terminal extensions confer the ability to bind heparan sulfate proteoglycans
in the extracellular matrix (438). VEGFR2 is a receptor tyrosine kinase that autophosphorylates
upon ligand binding and has a well-documented ability to activate eNOS via either Akt or PKC
(439; 440). In this study we investigated the role of NO generation in GLP-2-stimulated dietary
fat absorption and TRL secretion. We employed the Syrian golden hamster and various mouse
models in in vivo studies along with intestinal cell cultures in ex vivo, and propose a relay of
intercellular communication that can regulate postprandial lipid metabolism in the gut.

94
4.3 Materials and Methods
4.3.1 Animals
Male Syrian golden hamsters (Mesocricetus auratus; Charles River, Montreal, QC)
weighing 110g, male C57Bl/6J mice at 12 weeks of age (Jackson Laboratory, Bar Harbor, ME),
and male Nos3-/-
mice at 12 weeks of age (B6.129P2-Nos3tm1Unc
/J strain; Jackson Laboratory)
were housed in 12:12 h light:dark cycle and fed standard rodent chow. For in vivo experiments,
hamsters were fasted overnight and mice were fasted for 5 h in the morning. Animals were not
fasted when intestine was being collected for ex vivo experiments. Upon sacrifice, all animals
were anaesthetized with 3% isoflurane and the small intestine was excised and flash frozen in
liquid nitrogen. All procedures were carried out in compliance with the guidelines of The
Hospital for Sick Children Animal Ethics Committee.
4.3.2 Determination of dietary fat secretion in vivo
In hamsters under isoflurane anaesthesia, a silastic catheter (VWR) was implanted into
the right jugular vein, exteriorized at the back of the neck, and filled with heparinized saline (40
IU/mL). After overnight recovery, the remainder of the experiment was performed on conscious
hamsters. A bolus of L-NG-nitroarginine methyl ester (L-NAME, Cayman Chemical, Ann Arbor,
MI) was infused via the jugular vein at a dose of 75 mol kg-1
to inhibit NOS. 10 min later, 200
L of blood was withdrawn from the jugular catheter into lithium heparain-coated tubes (BD,
Franklin Lakes, NJ) as the baseline (time ‘0 min’) sample followed by an oralgastric gavage of 3
Ci [9,10-3H(N)]triolein (PerkinElmer, Boston, MA) in 200 L olive oil (Fluka, Buchs,
Switzerland). At 20 min, the hamsters received an intraperitoneal injection of PBS vehicle or
human GLP-21-33
(Bachem, King of Prussia, PA) at a dose of 0.25 mg kg-1
. 200 L blood
samples were collected every half hour until sacrifice at 2 h. In mice, 50 L blood was collected
from the tail vein with EDTA-coated Microvette tubes (Sarstedt, Newton, NC) every 30 min
until sacrifice at 90 min, and Gly2-GLP-2 (Pepceuticals, Leicester, UK) was administered 20 min
after the oral fat load. Blood was centrifuged at 6,000 rpm for 10 min in 4°C to isolate plasma.
4 mL of Ready Safe scintillation fluid (Beckman Coulter, Fullerton, CA) was added to 20 L

95
plasma and counted for 1 min in a Beckman LS6500 IC scintillation counter to measure tritium
radioactivity.
4.3.3 Determination of postprandial TRL production in vivo
Hamsters received 20% Pluronic F-127 (Poloxamer 407, Sigma-Aldrich, St. Louis, MO)
at a dose of 1000 mg kg-1
by intraperitoneal injection to inhibit peripheral TRL catabolism.
Hamsters were pretreated with L-NAME as described above. 20 min later, conscious hamsters
were administered 200 L olive oil by oralgastric gavage and the sample collected immediately
prior to the gavage was considered the ‘0 min’ time point. 20 min following the gavage, GLP-2
or PBS vehicle was administered by intraperitoneal injection. 400 L blood was collected every
half hour until sacrifice at 2 h. In mice, 50 L blood was collected from the tail vein every 30
min until sacrifice at 90 min. 100 L plasma was supplemented with preservatives, layered
under 1.006 g mL-1
KBr solution in polyallomer Microfuge tubes (Beckman Coulter, Palo Alto,
CA), and centrifuged at 45,000 rpm in 15°C for 15 min in a TLA-45 rotor using an Optima TL
Ultracentrifuge (Beckman Coulter) to isolate the Sf > 400 fraction, followed by another 45 min
centrifugation to isolate the Sf 100-400 fraction. ApoB48 immunoblotting was performed as
previously described (430). Total lipid concentration was determined by Intralipid levels
measured with the Roche Cobas Integra 400 Plus Analyzer. This method measures lipaemia
according to the optical properties of the lipid emulsion Intralipid, and based on the ratio of
absorbance at 800 nm to 659 nm.
4.3.4 Determination of intestinal apoB48 secretion ex vivo
Primary enterocytes were isolated from murine small intestine and a pulse chase
experiment was performed as previously described (71; 430). Briefly, 0.5 cm fragments of
intestine were bathed in Cell Recovery Solution (BD Biosciences, Bedford, MA) at 4°C for 1 h
to loosen epithelial cells from the basement membrane. Enterocytes were pulsed in methionine-
and cystine-free DMEM (Wisent, St-Bruno, QC) containing 100 M S-nitroso-L-glutathione
(GSNO, Cayman Chemical), 100 ng mL-1
recombinant mouse VEGF-120 (R&D Systems,
Minneapolis, MN), or vehicle for 45 min. The enterocytes were then labelled with
[35
S]methionine (PerkinElmer) for 30 min, followed by a chase in DMEM supplemented with

96
0.5% BSA and bile salt lipid micelles prepared as previously described (441). ApoB48 in cell
lysates and media was immunoprecipitated with goat anti-human apoB antibody (Midland,
Boone, IA) and protein A from Staphylococcus aureus (Sigma-Aldrich, St. Louis, MO), and
resolved by SDS-PAGE. [35
S]methionine-labelled apoB48 was visualized by exposing dried
gels to a phosphor screen, visualized with Storm 840 Optical Scanner and quantified using
ImageQuant software (Molecular Dynamics, GE Healthcare Life Sciences, Piscataway, NJ).
4.3.5 ISEMF-conditioned media
Murine small ISEMFs were isolated and cultured as previously described (359). ISEMFs
were serum-starved in 0.5% BSA-containing DMEM for 24 h, and then stimulated with 10 nM
GLP-2 for 30 min. The media was passed through a 0.2-m filter to remove cellular debris
immediately prior to its use in the ex vivo steady state labelling experiment.
4.3.6 Steady state labelling of apoB48 production ex vivo
Primary murine enterocytes were suspended in ISEMF-conditioned media, supplemented
with bile salt lipid micelles, and labelled with 150 Ci [35
S]methionine. For neutralization
experiments, goat monoclonal anti-mouse VEGF (R&D Biosystems, Minneapolis, MN) or goat
IgG control was added for a final concentration of 0.2 ng mL-1
. Media and cell samples were
collected every 45 min for 135 min and newly-synthesized apoB48 production was quantified as
described above.
4.3.7 Other biochemical measurements
Intact jejunum was crushed to a powder using a mortar pestle and liquid nitrogen. Lipids
were extracted using a 2:1 mixture of chloroform:methanol, and TG mass was determined using
an enzymatic-based colourimetric kit (Randox, Crumlin, UK). Nitrate content of jejunum
homogenate was assessed by an assay that involved reducing with nitrate reductase and
visualizing nitrite colourimetrically with the Griess reagent (Cayman Chemical). ApoAIV levels
were determined by immunoblotting using a goat anti-apo AIV antibody (kind gift of Dr. Patrick
Tso, University of Cincinnati, OH). VEGF levels in ISEMF-conditioned media were determined
by ELISA (R&D Biosystems).

97
4.3.8 Kinex phosphoprotein screen
C57BL/6 mice were given 200 L olive oil by oralgastric gavage and then given an
injection of GLP-2 or PBS as described in experiments above. After 30 min, the jejunum was
excised and the mucosa was scraped off on ice with a glass microscope slide. Mucosa was
homogenized in lysis buffer (20 mM MOPS, 2 mM EDTA, 2 mM EGTA, 1% Triton X-100, and
1 mM DDT) supplemented with phosphatase and protease inhibitors. The homogenate was
cleared by centrifuging at 45,000 rpm in the Beckman Optima TL Ultracentrifuge. The cleared
homogenate was sent to Kinexus (Vancouver, BC) and analyzed by the Kinex Antibody
Microarray service.
4.3.9 Statistical analysis
Results are expressed and depicted as mean ± SEM. Secretion studies were analyzed by two-
way ANOVA with Bonferroni’s post-test to analyze individual time points. Other measurements
were analyzed either by two-tailed Student’s t-test or a one-way ANOVA.

98
4.4 Results
4.4.1 In vivo role of NO in GLP-2-stimulated postprandial TRL secretion
To determine whether GLP-2 can activate intestinal NOS in the hamster model in the
context of a fat load, nitrate, jejunal levels of a downstream metabolite of NO that occurs in
normoxic conditions were measured. There appeared to be an increase in jejunal nitrate in
association with GLP-2 treatment (Figure 4-1A). Next, functional studies were performed using
the pan-specific NOS inhibitor L-NAME. Hamsters were pretreated with L-NAME at a dose
sufficient to block GLP-2-induced circulation changes in rats (306) and induce hypertension in
Syrian golden hamsters (442), challenged with an oral fat load of olive oil and given an
intraperitoneal injection of Poloxamer 407 to inhibit peripheral TRL catabolism. GLP-2
increased apoB48 accumulation in the TRL fraction as previously described (Figure 4-1B), and
this increase was blocked by L-NAME pretreatment. Neither the L-NAME nor the GLP-2 + L-
NAME group had a difference TRL-apoB48 secretion relative to control. There was an increase
in postprandial TRL-lipids with an intraperitoneal injection of GLP-2 (slope 3.65 ± 0.73 × 10-3
g
L-1
min-1
vs. 1.63 ± 0.28 × 10-3
g L-1
min-1
, GLP-2 vs. control) and this was mitigated with L-
NAME pretreatment (slope 3.67 ± 0.15 × 10-4
g L-1
min-1
, p < 0.01 vs. GLP-2) (Figure 4-1C). L-
NAME alone also suppressed TRL-lipid mass relative to the control group (slope 3.60 ± 0.17 ×
10-4
g L-1
min-1
, p < 0.01 vs. control), altogether suggesting a basal role of NOS in lipidating
apoB48 particles. Because the source of fatty acid substrate could have been derived from the
circulation or the gavage sources in the previous experiment, a [3H]triolein label was included in
the oral gavage to label exogenously-derived fatty acids. As noted before, GLP-2 acutely
increased the appearance of tritium in the circulation, signifying increased uptake of luminal
fatty acids (slope 1.66 ± 0.25 × 102 dpm mL
-1 min
-1 vs. 1.11 ± 0.17 × 10
2 dpm mL
-1 min
-1, GLP-
2 vs. control) (Figure 4-1D). As with TRL-apoB48, L-NAME pretreatment abrogated the
stimulatory effect of GLP-2 on fatty acid uptake (slope 1.02 ± 0.13 × 102 dpm mL
-1 min
-1), but
alone did not have any effect relative to control (slope 1.38 ± 0.13 × 102 dpm mL
-1 min
-1).
Similar observations were made in the TRL tritium content (Figure 4-1E). The data suggests that
NOS is necessary for GLP-2-stimulated luminal FA uptake, but inhibition of all NOS isoforms
does not affect basal levels of uptake.

99
Figure 4-1. The role of nitric oxide in apoB48 production in hamsters in vivo
Hamsters were challenged with an oral fat load, and given an intraperitoneal injection of
Poloxamer 407 (1000 mg kg-1
) and GLP-2 (0.25 mg kg-1
). (A) Jejunal nitrate levels 2 h after the
fat load as determined using the Greiss reagent. n = 5-10 per group. (B) Increase in TRL-
apoB48 mass following the fat load. Representative apoB48 immunoblots are shown to the left.
n = 4 per group, * p < 0.05 GLP-2 vs. control, # p < 0.05 GLP-2 vs. GLP-2 + L-NAME, $ GLP-
2 vs. L-NAME. (C) Total TRL-lipid mass following the fat load. n = 5 per group, % p < 0.05
L-NAME vs. control, ** p < 0.01 GLP-2 vs. control, $$$ p < 0.001 GLP-2 vs. L-NAME, ### p <
0.001 GLP-2 vs. L-NAME + GLP-2. (D) Hamsters were challenged with an oral fat load of
olive oil containing 3 mCi [3H]triolein label. Circulating tritium levels was quantified as a
measure of luminal fatty acid uptake. n = 5 per group, # p < 0.05 GLP-2 vs. GLP-2 + L-NAME,
### p < 0.001 GLP-2 vs. GLP-2 + L-NAME, * p < 0.05 GLP-2 vs. control, ** p < 0.01 GLP-2
vs. L-NAME. (E) TRL tritium levels following the oral fat load containing the [3H]triolein
label. n = 5 per group.

100
4.4.2 Ex vivo role of NO in apoB48 secretion
Primary hamster enterocytes were isolated and exposed to a physiologically occurring
NO donor in the gut, GSNO at a dose sufficient to improve barrier function (443), which is a
described function of GLP-2. The enterocytes were also cultured in methionine- and cystine-free
media, followed by labelling with [35
S]methionine in a pulse chase study. GSNO trended to
increase intracellular amounts of [35
S]-apoB48 (Figure 4-2B) and significantly upregulated the
secretion of newly-synthesized apoB48 within 90 min of the chase (Figure 4-2C). Similar
observations were made with a chemically distinct NO donor, DEANONOate (data not shown),
thereby suggesting that NO has a direct role in modulating enterocyte apoB48 synthesis and
secretion.

101
Figure 4-2. The effect of an NO donor on apoB48 production in hamster enterocytes ex
vivo
Primary hamster enterocytes were treated with the NO donor GSNO or reduced glutathione
control, starved in methionine, cysteine, and serum-free DMEM supplemented in bile salt lipid
micelles, and then given [35
S]methionine to label protein synthesis. Samples were subject to
immunoprecipitation with a hamster anti-apoB antibody. (A) Representative SDS-PAGE of
immunoprecipitated [35
S]-apoB48 from cell and media. (B) Quantification of intracellular
amounts of newly-synthesized apoB48. (C) Quantification of secreted amounts of newly-
synthesized apoB48. n = 3 per group, * p < 0.05.

102
4.4.3 Role of eNOS in GLP-2-stimulated chylomicron secretion
Since NO can be generated by one of three NOS isoforms in the mammalian system, we
sought to identify the isoform involved in GLP-2’s action on chylomicron secretion. We
assessed the ability of GLP-2 to promote dietary fat absorption in the eNOS KO mouse model.
When challenged with an oral fat load containing [3H]triolein, GLP-2-treated eNOS KO mice
exhibited significantly less tritium secretion relative to their wild type counterparts (slope 3.43 ±
0.65 × 103 dpm mL
-1 min
-1 vs. 2.16 ± 0.31 ×10
3 dpm mL
-1 min
-1, wild type + GLP-2 vs. eNOS
KO + GLP-2) (Figure 4-3A), suggesting that eNOS is required to some degree to mediate the
lipid effect of GLP-2. ApoB48 mass in the TRL fraction was examined, and there was a
noticeable decrease in the largest lipoprotein fraction (Sf > 400) when mice were eNOS-deficient
(Figure 4-3B). No discernable difference was noted in the apoB48 content of the smaller Sf 100-
400 lipoprotein fraction (Figure 4-3C), suggesting that eNOS is required to maximize the amount
of lipid associated with each apoB48 polypeptide. The inability to fully lipidate apoB48
conferred by eNOS deficiency resulted in significantly more TG accumulation in the jejunum 90
min after the oral fat load (4.47 ± 0.20 mg TG/mg intestine vs. 2.77 ± 0.75 mg TG/mg intestine,
eNOS KO vs. wild type) (Figure 4-3D), indicating an inefficiency in exporting dietary fat out as
chylomicrons. The mechanism for this observation remains unexplained, as it was noted that
secretion of apoAIV, an intestine-specific apolipoprotein whose putative role is to expand the
chylomicron lipid core (77), was unchanged (Figure 4-3E). In summary, it appears that eNOS is
necessary for both GLP-2-stimulated TRL secretion and basally for proper lipidation of apoB48
in the intestine.

103
Figure 4-3. Effect of GLP-2 on dietary fat absorption and postprandial lipoprotein
secretion in C57Bl/6J and eNOS KO mice
Mice were challenged with an oral fat load containing 3 mCi [3H]triolein along with an
intraperitoneal injection of Poloxamer 407 (1000 mg kg-1
) and Gly2-GLP2 (0.25 mg kg
-1). (A)
Circulating tritium levels as an indicator of dietary fat absorption. n = 7 per group, * p < 0.05
wild type GLP-2 vs. eNOS KO GLP-2. (B) Sf > 400 TRL-apoB48 and (C) Sf 100-400 TRL-
apoB48 levels as assessed by immunoblot. n = 4 per group, *** p < 0.01 inter-genotype. (D)
TG accumulated in jejunum of mice 90 min following an oral fat load. (E) Sf > 400 TRL-
apoAIV and (F) Sf 100-400 TRL-apoAIV levels as determined by immunoblot.

104
4.4.4 Role of ISEMFs in GLP-2-stimulated apoB48 secretion
Next we sought to identify the GLP-2R-positive cell that could be secreting a paracrine
factor that stimulates chylomicron oversecretion in absorptive enterocytes. Cultured ISEMFs
were used as they are known to express the GLP-2R (359) and have been demonstrated to
increase Igf1 mRNA levels acutely in response to GLP-2. Conditioned media were obtained
from cultured murine intestinal SEMFs stimulated following treatment with GLP-2 (10 nM) for
30 min. Primary murine enterocytes were cultured in this conditioned media in the presence of
bile salt lipid micelles to mimic the lipid-rich environment of the postprandial lumen, and
apoB48 synthesis and secretion was steady-state labelled with [35
S]-methionine. There was a
trend for increased intracellular [35
S]-apoB48 levels (Figure 4A) and secreted [35
S]-apoB48
(Figure 4-4B) with significantly more total [35
S]-apoB48 in enterocytes that were incubated in
conditioned media collected from GLP-2-stimulated ISEMFs (p < 0.05, GLP-2 conditioned
media vs. control conditioned media) (Figure 4-4C). Enterocytes exposed to GLP-2 conditioned
media also had increased levels of glycosylated CD36 (Figure 4-4D). This would suggest that
ISEMF is at least one GLP-2R-positive gastrointestinal cell type that can release a mediator
capable of eliciting more chylomicron secretion in enterocytes.

105
Figure 4-4. Effect of ISEMF conditioned media on apoB48 production in primary hamster
enterocytes
Primary murine enterocytes were exposed to conditioned media collected from cultured murine
jejunal subepithelial myofibroblasts stimulated with 10 nM GLP-2 for 30 min. ApoB48
synthesis and secretion was steady-stated labelled with 150 mCi [35
S]methionine and
immunoprecipitated with an anti-apoB antibody. Representative SDS-PAGE of
immunoprecipitated [35
S]-labelled apoB48 shown above. (A) Amount of [35
S]-apoB48 within
the cell normalized to total protein synthesis as assessed by TCA precipitation. (B) Amount of
[35
S]-apoB48 secreted from enterocytes. (C) Total amounts of [35
S]-apoB48. n = 4 per group, *
p < 0.05 by two-way ANOVA. (D) Ratio of glycosylated CD36 to unglycosylated CD36 in
enterocytes incubated in conditioned media. n = 3 per group, * p < 0.05.

106
4.4.5 VEGF action in enterocyte apoB48 secretion
ISEMFs have been shown to secrete myriad peptide factors, including KGF and IGF-1
(354; 359). Both KGF and IGF-1 were tested and found not to promote apoB48 secretion in
primary hamster enteroctyes ex vivo (data not shown). Given the observations with L-NAME
and GSNO, we searched for a factor that could activate NOS, or particularly, eNOS. VEGF has
a well-documented ability to activate eNOS (444). In the Kinex phosphoprotein screen of
mucosal scraping lysates, it was noted that there was significantly increased Tyr1054
phosphorylation on VEGFR2 (VEGFR2) with GLP-2 administration in C57Bl/6 mice (Figure 4-
5A), indicating the receptor’s activation. However, such magnitude of increase in
phosphorylation may have been attributable to receptor activation in other contaminating
intestinal cell types that were removed in mucosal scraping. Primary murine enterocytes were
then isolated and exposed to 100 ng/mL VEGF-120 in a pulse chase study, while [35
S]-labelled
apoB48 was immunoprecipitated (Figure 4-5B). The inclusion of VEGF in the pulse media
trended towards an increase in the total accumulation of [35
S]-apoB48 over time, although this
effect was not significant for this sample size (Figure 4-5C).

107
Figure 4-5. The role of VEGF in intestinal apoB48 secretion in mice
(A) VEGFR2 Tyr1054 phosphorylation of intestinal mucosa scrapings of C57Bl/6 mice 1 h after
an oral fat load and GLP-2 administration. Phosphorylation was determined by Kinexus Kinex
phosphoprotein microarray analysis. n = 3 per group, * p < 0.05. (B) Primary murine
enterocytes were labelled with [35
S]methionine in a pulse chase experiment while cultured with
100 ng mL-1
VEGF-120. Representative SDS-PAGE of immunoprecipitated [35
S]-apoB48 from
three experiments. (C) Total amounts of [35
S]-apoB48, n = 3 per group.

108
4.4.6 VEGF as a mediator linking ISEMFs and enterocytes
The data presented in Figure 4-5 suggests that GLP-2 is capable of eliciting VEGF
secretion from this GLP-2R-positive intestinal cell, and may be a peptide communicator to
enterocytes. Therefore, a goat monoclonal antibody against murine VEGF was added to the
GLP-2-conditioned media to neutralize the peptide. Primary murine enterocytes were again
steady-state labelled and we assessed the anti-VEGF antibody’s ability to suppress the GLP-2-
conditioned media’s augmenting action on [35
S]-apoB48. Relative to the IgG control, VEGF
neutralization decreased intracellular [35
S]-apoB48 (Figure 5-6B), had a less pronounced effect
on secreted [35
S]-apoB48 (Figure 5-6C), but had very similar values to the control conditioned
media curve with regards to total [35
S]-apoB48 (Figure 5-6D).

109
Figure 4-6. The role of VEGF in GLP-2-stimulated ISEMF-mediated intestinal apoB48
secretion in enterocytes
(A) Primary murine enterocytes were steady-state labelled with [35
S]methionine in conditioned
media collected from jejunal SEMFs treated for GLP-2 for 30 min. VEGF in the media was
neutralized with goat anti-mouse VEGF antibody and compared to goat IgG control.
Representative SDS-PAGE of immunoprecipitated [35
S]-apoB48 from 3 experiments. (C)
Cellular amounts of [35
S]-apoB48. (D) Secreted amounts of [35
S]-apoB48. (E) Total amounts
of [35
S]-apoB48. n = 3 per group, * p < 0.05.
*

110
4.5 Discussion
Aside from the observation that GLP-2 acutely promotes CD36 glycosylation to facilitate
its expression at the apical brush border membrane, little is known as to how this postprandial
hormone promotes the secretion of apoB48-containing TRL secretion from the intestine. The
difficulty in determining the physiological mechanism lies in the absence of the GLP-2R on the
absorptive enterocyte. In the present study we found at least one intestinal cell type, the ISEMF
that can stimulate enterocytes with a paracrine mediator to promote dietary fatty acid uptake.
Based on studies using a NOS inhibitor, it is clear that NO generation in the intestine is
important to GLP-2-stimulated and basal chylomicron secretion. A search for a peptide that can
provoke NO production yielded VEGF, which was shown to induce newly-synthesized apoB48
production in enterocytes. We therefore propose that the schematic illustrated in Figure 4-7 is
one paracrine pathway that GLP-2 may use to promote chylomicron secretion in enterocytes.
The involvement of other cytokines and peptide hormones, however, cannot be excluded.

111
Figure 4-7. Intercellular interactions resulting in GLP-2-stimulated chylomicron secretion
We postulate that GLP-2 secreted by the enteroendocrine L-cell agonizes its G protein-coupled
receptor on SEMFs in the intestine. This induces SEMFs to secrete a number of cytokines,
including VEGF, which acts on its receptor VEGFR2 on the absorptive enterocyte. Stimulation
of VEGFR2 activates eNOS in the enterocyte, which leads to increases in dietary FA uptake and
subsequently chylomicron output. There is likely the involvement of additional factors secreted
by ISEMFs to bring about an increase in chylomicron production.

112
The decreased TRL-lipid associated with L-NAME pretreatment alone (Figure 4-1C)
suggests that NO that is constitutively produced, for example by neuronal NOS in the intestine,
has a role in regulating chylomicron metabolism. There is existing evidence that NO generation,
particularly by the eNOS isoform, is important to lipid metabolism in the liver. The eNOS-
deficient mice have abnormally high hepatic TG accumulation, but the authors propose the
observation was due to relief of NO-inhibited FA synthesis (445). Similarly, analysis of the S-
nitrosylated proteome in the liver reveals that the covalent modification occurs on proteins
involved in lipid metabolism, such as very long chain acyl-CoA dehydrogenase (446). A NO
donor has been implicated in promoting hepatic lipoprotein secretion by upregulating MTP
mRNA (447). While inhibiting NO synthesis could block the stimulatory effects of GLP-2 on
TRL secretion, the L-NAME data would suggest that NO has a basic role in intestinal lipoprotein
output. Though L-NAME pretreatment alone did not affect the number of apoB48 particles in
the TRL fraction (Figure 1B), there was a dramatic reduction in the lipid mass of the fraction
compared to control-treated hamsters (Figure 4-1C), while having no effect on the secretion of
exogenous labeled lipid (Figure 4-1D). Data from eNOS-deficient mice also support this notion,
as these mice secreted significantly fewer particles of Sf > 400 size (Figure 4-3B). Altogether,
this may suggest that NO is essential to secreting fully lipidated apoB48 particles but the lipid is
not sourced exogenously. The FA substrates for NO-mediated apoB48 assembly may come from
the circulation. FFA have been demonstrated to induce apoB48 secretion from isolated
enterocytes (76), and plasma FFA have been shown to promote apoB48 oversecretion in
hamsters (435) and humans (164). However, FFA derived from the circulation are unlikely to
account for the core size differences of apoB48 particles secreted by L-NAME-treated hamsters
and eNOS-deficient mice, as FA taken up at the basolateral surface are preferentially
incorporated into phospholipids, not TG, and catabolized by -oxidation (448). Another possible
source of lipid could come from mobilization of intestinal intracellular stores, as there is
evidence that FA from a meal is retained until release with Sf > 400 particles in the next meal
(174).
In Figure 4-3A, eNOS-deficient mice did not have altered postprandial appearance of
tritium in the plasma relative to control. While eNOS appears to be necessary for GLP-2-
increased chylomicron secretion, it may not be essential for dietary lipid transport per se. L-

113
NAME pre-treatment had no effect on the appearance of tritium in plasma relative to control
(Figure 4-1D), so constitutional nitric oxide generation was evidently not essential to this process
in the basal state. Following a fat load, eNOS-deficient mice accumulated TG in the jejunum
regardless of treatment (Figure 4-3D). Therefore, while eNOS possibly mediates the effect of
GLP-2-stimulated CD36 action, basal eNOS activity also plays an additional role later on in
chylomicron secretion pathway.
The VEGF data is still preliminary and requires confirmation. While VEGF
neutralization inhibited an increase in intracellular amounts of [35
S]-apoB48 (Figure 4-6B), there
was little effect on secretion. Therefore, VEGF may contribute to GLP-2-stimulated apoB48
biogenesis, but the secretion of chylomicrons involves another factor. To clarify the role of
VEGF in apoB48 production, basal and GLP-2-stimulated intestinal apoB48 levels can be
assessed in Vegf+/-
mice. The role of NO in VEGF’s actions on apoB48 can be confirmed by
administering eNOS knockout mice with VEGF-A in vivo, and assessing the effect of VEGF on
eNOS-deficient enterocytes ex vivo.
Though the ex vivo experiments with GSNO would suggest that NO can promote apoB48
secretion at the level of the enterocyte, NO generation in other cell types may contribute to the
mechanism of GLP-2 action. With low doses of GLP-1, NO has been implicated in the
inhibition of gastrointestinal motility (449). GLP-2 has been shown to potentiate the inhibitory
effect of GLP-1 (338), but GLP-2-suppressed gastric motility has been shown to occur via VIP
and independently of NOS activity (372). Given that GLP-2 has been co-localized to eNOS-
positive enteric neurons (304), and enteric neurons can secrete GSNO (443), it is possible that
the enteric nervous system plays a role in GLP-2-stimulated apoB48 secretion. Interestingly,
GLP-2’s intestinal epithelial restitution effects have been ascribed to VEGF secreted by ISEMF
(434), and such an intestinal epithelial wound healing effect is a biological function of GSNO
(443), so this functional overlap suggests that VEGF and NO act in the same pathway. Though
the anti-inflammatory effects described for GLP-2 include downregulation of inducible NOS
(iNOS) (300), inflammatory signalling has been shown to result in chylomicron oversecretion
(165). It may be worthwhile to investigate the specific role of other NOS isoforms in regulating
intestinal lipoprotein production. VEGFR2 signalling in the enterocyte itself also has the
potential to stimulate chylomicron secretion. Phospholipase C (PLC) directly binds a

114
phosphotyrosine residue on VEGFR2 to activate the MEK/ERK cascade (450), and ERK
activation has been associated with chylomicron overproduction in insulin-resistant enterocytes
(154). On the other hand, the adapator protein Shb binds another phosphotyrosine residue on
VEGFR2 that leads to PI3-K activation (451), with PI3-K being instrumental in lipid transporter
trafficking in Caco-2 cells (441).
The identification of a paracrine factor that can directly stimulate the enterocyte to
increase apoB48 output could serve as an important tool in understanding intestinal lipid
metabolism. In future studies, VEGF can be used in enterocyte cell models to elucidate the
molecular mechanisms behind the hyperlipidemia observed in vivo at the whole body level.

115
Chapter 5: DISCUSSION AND CONCLUSIONS
Author contribution: J. Hsieh produced all text and figures in this chapter.

116
5.1 Summary of Results
The present studies on GLP-1 and GLP-2 indicate that pharmacological levels of these
hormones are capable of acutely bringing about considerable changes in intestinal apoB48 output
in multiple rodent models. Though co-secreted in equimolar amounts, these gut peptides exact
complete opposite effects on intestinal lipoprotein secretion. Raising endogenous levels of GLP-
1 with a DPP-4 inhibitor or administrating pharmacological levels of a long-acting GLP-1R
agonist suppressed TRL-TG and TRL-apoB48 secretion in chow-fed hamsters, fructose-fed
hamsters, and mice challenged with an oral fat load within a 2 h period. The hypolipidemic
effect of the DPP-IV inhibitor sitagliptin was dependent on intact GLP-1R signalling, and was
independent of insulin secretion and prolonged GIP bioactivity. In contrast, pharmacological
levels of GLP-2 rapidly increased uptake of luminal FA to promote chylomicron secretion in
vivo in hamsters and mice, but not CD36-deficient mice. Increased dietary FA uptake was
achieved by GLP2-induced glycosylation of CD36 facilitating its insertion into the brush border
membrane of enterocytes, where it can perform its lipid transport function. NOS activity was
also found to be important in chylomicron secretion, as it was essential for basal levels of
apoB48 particle lipidation GLP-2-accelerated dietary FA uptake in vivo. GLP-2 appeared to
indirectly regulate the absorptive enterocyte through VEGF secreted by ISEMFs, thereby
establishing the involvement of paracrine communication in the physiological action of GLP-2.

117
5.2 GLP-1 as a Direct Regulator of Intestinal Lipoprotein
Metabolism
An exogenous GLP-1 agonist, exendin-4 acted to acutely suppress apoB48-containing
TRL secretion from the intestine in both hamsters and mice, thereby favourably moderating
postprandial lipemia. Our data suggested that endogenous levels of GLP-1 were also capable of
suppressing secretion, as long as its bioactivity was maintained by inhibiting DPP-4 activity.
The apoB48- and TG-lowering effect observed with exendin-4 is consistent with studies in
which GLP-1 was intravenously infused in rats (394) and humans (393). However, these studies
attributed the postprandial lipid lowering to decreased lymphatic flow rate (394), or speculated
that it was a result of increased peripheral clearance of TRL-TG (393). Moreover, delayed
gastric emptying could explain the blunted rise in TG in the study performed by Meier et al
(393). However the hypolipidemic effect was still observed when exendin-4 was injected 1 h
after the oral gastric gavage, which obviated any gastric emptying effects. Additionally, both
fructose-fed hamsters and mice secreted less TG following sitagliptin administration, and DPP-4
inhibition has been shown to have no effect on gastric emptying (452; 453). Though insulin
likely plays a role in the hypolipidemic action of GLP-1, there is evidence that GLP-1 can
directly modulate lipoprotein metabolism in the intestine. The decrease in apoB48 and TG
secretion persisted after differences in insulin levels diminished between sitagliptin-treated and
control mice, and insulin did not appear to have the same acute hypolipidemic effect in mice as it
does in hamsters (154) and humans (163). A single subcutaneous injection of the GLP-1R
agonist therapeutic exenatide has been shown to decrease serum TG and apoB48 in individuals
with impaired glucose tolerance challenged with a fat-enriched meal. The lipid lowering
occurred despite defective insulin action in these individuals, and was observed concurrently
with an unexpected drop in insulin and minimal alterations in circulating non-esterified FA
(454). The glucagonostatic activity of GLP-1 is unlikely to explain the changes in intestinal
lipoprotein secretion, as glucagon does not have an effect on VLDL1-apoB48 and VLDL2-
apoB48 production rates in healthy men (455). Last but not least, in pulse-chase experiments
exendin-4 inhibited newly-synthesized apoB48 secretion in primary enterocyte cultures. The
data presented herein indicate that GLP-1R signalling has a direct role in affecting enterocyte
lipid metabolism. This direct effect of GLP-1 may explain the improvement in postprandial

118
serum TG and apoB48 seen in exenatide-treated patients but not insulin glargine-treated patients,
especially since body weights were similar between the two groups (456). An interesting
observation was that exendin-4 could acutely raise HDL cholesterol levels which, given that
hamsters are cholesteryl ester transfer protein-expressing animals (457), could be a result of the
lower TRL TG allowing more cholesteryl ester to be retained in HDL particles.
5.3 CD36 as a Hormone-Sensitive FA Transporter in the Gut
The observation that GLP-2 can acutely affect the subcellular distribution of CD36 adds
the intestinal epithelium to the growing list of tissues in which the trafficking of CD36 can be
regulated by hormonal signals. Insulin has been recognized as a hormone that can direct CD36
to the plasma membrane in skeletal muscle (458), cardiomyocytes (418), and adipocytes (459).
In skeletal muscle and cardiomyocytes, the insulin-stimulated translocation of CD36 is PI3-K-
dependent, as it can be blocked by wortmannin or LY294002 (418; 458). Insulin-stimulated
CD36 translocation from subcellular compartments requires vesicle-associated membrane
proteins (VAMP). In addition to VAMP2 and VAMP5, VAMP4 appears to be specific to
insulin-stimulated CD36 trafficking (460). Notably, GLP-2-induced translocation of the glucose
transporter SGLT-1 is also a PI3-K-dependent process (312). It is also worth noting that the PI3-
K/Akt pathway is activated by VEGFR2 signalling (438), suggesting a signalling mechanism by
which the VEGF secreted by ISEMFs can direct CD36 insertion into the BBM.
Though the functional significance of greater CD36 surface expression is expected to be
the increased facilitated transport of dietary FA, apically-expressed CD36 may also serve as a
lipid sensor. Lingual CD36 is involved in the gustatory perception of fat and is responsible for
preference of lipid-rich foods (24). It has been noted that CD36 in rat enterocytes disappeared
from the apical membrane starting at 1 h after exposure to luminal lipids, coupled with protein
polyubiquitination and degradation. Moreover, in the same study, FA-induced ERK1/2
activation in the enterocyte paralleled CD36 levels, and coincided with increased apoB48 and
MTP levels (461). Since ERK1/2 activation is associated with apoB48-containing lipoprotein
overproduction in intestinal insulin-resistance (154), the result of GLP-2-stimulated apical CD36
expression may be the initiation of cellular signalling to promote chylomicron secretion. A
similar role has been proposed for a structurally-related scavenger receptor in the enterocyte, SR-

119
BI, which drives the trafficking of apoB48 towards the basolateral membrane for secretion
following micelle-stimulated MAPK signalling (462). In a corroborative study, SR-BI
knockdown by RNAi was shown to decrease apoB secretion from Caco-2 cells (441).
The biotinylation experiment demonstrated that the fully glycosylated form of CD36 is
specific to the BBM, and also indicated that the majority of enterocyte CD36 is unglycosylated
and intracellular. The intracellular compartment from which CD36 translocates is currently
unidentified. In 3T3-L1 adipocytes, surface CD36 is found specifically in cholesterol-enriched
detergent-resistant membranes, where it is important to its FA transport function (15). It has
been reported that CD36 functionally interacts with caveolin-1 in caveolae (463), where the
presence of sphingomyelin synthase 2 is also necessary for oleic acid uptake and lipid droplet
formation in hepatocytes (464). Interesting, the localization of CD36 in caveolae has
implications for eNOS regulation. The binding of LDL to CD36 lead to caveolae cholesterol
depletion, which in turn relocalized eNOS to intracellular compartments, rather than keeping
eNOS in close proximity to the signalling components such as receptors and Gq proteins that are
enriched in caveolae (465). However, CD36’s association with FA, in particular myristic acid,
has been linked with eNOS activation in a PI3-K-independent pathway (466). Therefore, it is
possible there is reciprocal regulation between CD36 and eNOS, and this can occur in a
substrate-specific manner. Endosomal alkalinization sustained CD36 expression at the plasma
membrane, thus indicating that CD36 is internalized into endosomal compartments (467). The
endosomal structure though, at least when CD36 is internalized upon binding oxidized LDL, is
transferrin- and caveolin-free, but colocalizes with glycosyl-phosphatidylinositol-anchored
protein decay factor (468). In hepatocytes however, CD36 colocalizes with caveolin-1 in
intracellular vesicles (464). Transfection of primary cultured human muscle cells with a CD36-
EGFP construct showed that CD36 was not coincident with the Golgi marker GM-130,
suggesting that unlike FATP4, CD36 is not found in the Golgi (469). CD36 has been detected in
the ER, where it appears to be essential for PCTV budding (83). The role of ER-localized CD36
may confer a functional importance to the large amount of unglycosylated intracellular CD36
detected in primary hamster enterocytes.
Mutation of several of the consensus sequence N-X-S/T sites indicated that N-linked
glycosylation is essential in targeting CD36 to the plasma membrane in HEK293T cells (470).

120
While our study confirms that glycosylation is important to CD36 trafficking in the intestine,
other posttranslational modifications contribute to CD36 processing and eventual localization in
the plasma membrane. CD36 contains two –SS- sites at both termini of the molecule, and
palmitoylation of these sites is necessary for CD36 to proceed from the ER to Golgi for further
maturation and for greater stability (471). Insulin has also been shown to affect covalent
modification of CD36 by suppressing polyubiquitination on lysine residues in the carboxy
terminus, thereby maintaining CD36 levels at the plasma membrane (472). There is also
emerging evidence that CD36 can be modulated according to its phosphorylation state. In the
intestine, there is evidence alkaline phosphatase is an enzyme that dephosphorylates CD36,
which could allow regulation of CD36 activity at the brush border membrane (473).
5.4 VEGF and NO as Modulators of Intestinal Function
Our studies show that VEGF is a paracrine factor secreted by subepithelial
myofibroblasts in response to GLP-2 that can enhance lipid transport in intestinal epithelial cells.
Though VEGF is more known for its angiogenic effects, there is abundant information that it has
trophic functions in the gastrointestinal tract. Upon an injury in the gastrointestinal tract such as
gastric ulceration, VEGF production is induced and can be detected in regenerated epithelial
cells (474). Gastric mucosal protection against ethanol is conferred after only a single oral dose
of VEGF, with the onset of angiogenesis occurring after three weeks of treatment (475).
Following small bowel resection, it was found that VEGF derived from salivary glands mediated
the adaptive villus heightening in the gut (476). Moreover, encapsulating VEGF in
biodegradable microspheres for sustained, controlled release increased villus height and crypt
depth 2 weeks following extensive bowel resection (477). Transgenic mice with VEGF
expression driven by the villin promoter have intestinal epithelial hyperproliferation and even
cyst formation in crypts when the mice were crossed with Multiple intestinal neoplasm (Min)
mice. Interestingly, these tumours were not highly vascularized, suggesting that VEGF has
neoplastic effects independent of angiogenesis (478). In addition, non-malignant human
colonocytes exhibited increased proliferation when co-cultured with fibroblasts and endothelial
cells, suggesting that communication between these cells types results in mucosal proliferation
independent of blood flow (478).

121
Like VEGF, NO has been implicated in improving intestinal function. Inhibiting all NOS
isoforms with NG-monomethyl-L-arginine (L-NMMA) exacerbates jejunal injury induced by
endotoxin, but the injury was improved with infusion of the NO donor S-nitroso-N-acetyl
penicillamine in rats (479). Very high levels of NO have been shown to reduce intestinal
epithelial cell viability, but such levels are achieved by the activity of iNOS, as opposed to the
constitutive, calcium-sensitive isoforms that include eNOS and neuronal NOS (480). Since
GLP-2 is generally considered anti-inflammatory and has been shown to lower iNOS expression
(302; 481), it is likely that the NO arising from GLP-2 action is generated by the constitutive
isoforms of NOS. Studies looking at the effect of dietary L-arginine on gut function may also
provide clues about the role of NO, since L-arginine is the substrate of NOS. In weaned pigs,
dietary L-arginine protected the intestinal epithelium from apoptosis and maintained cell
proliferation in response to lipopolysaccharide (482). In a neonatal pig model of necrotizing
enterocolitis, an intravenous infusion of L-arginine attenuated tissue damage while L-NAME
infusion compounded the problem with hemorrhagic congestion (483). Interestingly, L-arginine
supplementation was noted to stimulate mucosal growth independent of mesenteric arterial blood
flow, but this effect still proceeded in the presence of L-NAME (484). Therefore, it appears that
NO is more instrumental in intestinal repair and healing as opposed to basal levels of growth.
NO has also been implicated as a regulator of intestinal absorption. While the opposite has been
shown for high pathophysiological levels, physiological levels of NO appears to promote fluid
absorption in the gut and directly regulate ion secretion in the enterocyte by activating soluble
guanylate cyclase (485). Such a proabsorptive function has also been described for GLP-2,
which suppresses chloride secretion in the epithelium (486).
NO can regulate membrane nutrient transporters, but as is the case with intestinal growth
and absorption, there are differential effects according to its concentration. Inhibiting
constitutive NOS activity with L-NAME actually decreased SGLT-1’s affinity for glucose in
IEC-18 cells, while increasing Na/H exchange (NHE3) (487). Interestingly, inhibiting NOS
activity has been shown to inhibit SGLT-1 glycosylation, which may have implications for its
trafficking and activity (488). In muscle cells, high levels of NO generated by iNOS blunts
insulin-stimulated glucose uptake (489). However, lower levels of NO arising from eNOS action
have been implicated in stimulating nutrient transport, particularly in insulin-independent

122
pathways. Exercise training increases eNOS protein levels in skeletal muscle (490), and
inhibiting NOS with L-NAME completely prevented exercise-increased sarcolemma GLUT4
content and glucose transport in rats (491). Moreover, L-NMMA at a dose of 0.2 mg kg-1
min-1
decreased leg glucose uptake during exercise in humans, without any measurable difference in
blood flow (492). The NO donor sodium nitroprusside has been shown to increase glucose
uptake in 3T3-L1 adipocytes in a guanylate cyclase-dependent manner and without Akt
activation (493). In endothelial cells, peroxynitrite, which is generated by NO reacting with
superoxide anion, in low levels activates AMPK in a c-Src and PI3-K-dependent and
[AMP]/[ATP]-independent pathway (494). The ability of metformin, but not AICAR, to activate
AMPK is actually lost in the tissues of eNOS-knockout mice (494). Given the ability of AMPK
to promote CD36 trafficking to the sarcolemma in muscle cells (460), it may have a similar
effect in enterocytes. Also, considering that AMPK action in the liver leads to bile acid secretion
and subsequently dietary lipid emulsification (495), the possibility of AMPK-stimulated CD36
translocation in enterocytes could help to transport the emulsified lipids and thus complement the
activity of the liver to maximize calorie absorption during energy-depleted states.
5.5 Concerted GLP-1 and GLP-2 Action
Our findings indicate that two gut peptides that are co-secreted in equimolar amounts
paradoxically have completely opposite acute effects on intestinal apoB48 secretion: GLP-2
increased both apoB48 particle and size, while GLP-1 appears to completely blunt apoB48
secretion. It is not unlikely that GLP-1 and GLP-2 act via distinct pathways in modulating
intestinal lipoprotein metabolism. In GLP-1-treated rats, there was significantly greater
[3H]triolein-derived tritium accumulation in the mucosa of the proximal small intestine,
suggesting that there was no deficiency in the uptake of luminal FA (394). GLP-2 did not affect
MTP levels, while it is possible that the GLP-1-stimulated insulin may indirectly suppress MTP
gene expression possibly through forkhead box O1 (FoxO1) (496), thus inhibiting chylomicron
assembly in vivo. Labelling studies with [3H]oleic acid and [
3H]acetate indicated that GLP-2 had
no effect on de novo lipogenesis and cholesterogenesis (data not shown). There are multiple
reports that GLP-1 directly inhibits lipogenic genes in hepatocytes (279; 497), and since apoB48
overproduction has been coupled to SREBP-1c activation in enterocytes (154), the modulation of
lipid synthesis may be one pathway through which GLP-1 acts in the intestine.

123
Given that GLP-1 and GLP-2 are secreted in equimolar amounts, it may be expected that
the net physiological effect of GLP-1 and GLP-2 in vivo would be no change in postprandial
lipemia. However, the indirect mechanism of GLP-2’s hyperlipidemic action, in which GLP-2
stimulates intestinal subepithelial myofibroblasts (and possibly other GLP-2R-positive cell
types) to secrete factors that regulate the absorptive enterocyte, would provide an opportunity for
amplification of the GLP-2 signal in the intestine. In contrast, such biological amplification is
not operative when GLP-1 can directly agonize its receptor in the intestine. Therefore, it would
not be surprising if the net outcome of postprandial GLP secretion is enhanced dietary lipid
transport. Recent work in our lab has indicated that co-infusion of GLP-1 and GLP-2 at
physiological levels in chow-fed hamsters for 120 min results in postprandial hyperlipidemia,
indicating a predominantly GLP-2 effect. GLP-2-induced hyperlipidemia was also more
dominant in insulin-resistant fructose-fed hamsters, but was attenuated in the context of a mixed
meal challenge in which the oral fat load was given in conjunction with glucose. However, in
the presence of the DPP-4 inhibitor sitagliptin, the hypolipidemic effect of GLP-1 was
predominant (498). The diminution of the hyperlipidemic effect with the inclusion of glucose in
the oral load suggests that glucose-dependent insulinotropism, at least in hamsters, is one
pathway that mediates GLP-1’s hypolipidemic function. The involvement of insulin may also
explain the delayed peak postprandial TG concentration observed with glucose ingestion in
humans during a fat tolerance test where ostensibly both GLP-1 and GLP-2 were secreted (499).
This would have pathophysiological implications in states of defective insulin secretion or
compromised insulin sensitivity, where one arm of GLP-1 influence is unavailable and would
thus allow the GLP-2 effect to outweigh that of GLP-1. Hence in STZ-treated rats that are
hyperphagic and oversecrete GLP and cannot secrete insulin, the elevated plasma apoB48 levels
(500) could be attributed to a dominance of GLP-2. In B6D2F1 mice fed a HFD for 3 weeks to
induce insulin resistance, there is greater transport capacity of linoleic acid, along with elevated
CD36 expression and higher intestinal mitotic index (161), thereby consistent with a
preponderance of GLP-2 action. Moreover, in diseases with dampened GLP secretion such as
type 2 diabetes, decreased insulin sensitivity in these patients implies that the reduced GLP-1
levels would have an additionally compromised acute hypolipidemic effect. Meanwhile the
GLP-2 molecules, although also reduced in number, could stimulate chylomicron production to

124
their full extent. The differing magnitudes of effect may possibly explain the exacerbated
postprandial apoB48 secretion in type 2 diabetic individuals (501).
A major contribution of DPP-4-mediated GLP truncation to their net physiological effect
on apoB48 secretion is consistent with the conclusions that can be inferred from the sitagliptin-
treated fructose-fed hamsters and mice. Sitagliptin administration alone to hamsters and mice
attenuated postprandial chylomicron secretion, suggesting that protection of endogenously-
produced peptides from DPP-IV yielded a net hypolipidemic response. Though both GLP-1 and
GLP-2 are substrates for DPP-4 with similar Km values, DPP-4’s kcat for GLP-1 is 7.1 s-1
as
opposed to 0.87 s-1
for GLP-2, so the former is degraded 9 times faster (502). Infusing a DPP-4
inhibitor valine-pyrrolidide extended GLP-2’s plasma half-life only by about 3 min in pigs (503),
and one study showed that extending the bioactivity of endogenous GLP-2 with valine-
pyrrolidide did not affect intestinal morphology in mice or rats (504), so it is not unlikely that
there are physiological GLP-2 functions that do not intensify with protection from DPP-4-
mediated degradation. Moreover, the site of fat absorption along the longitudinal axis of the
intestine may explain the net physiological effect of the GLPs. Within the first half hour of meal
ingestion, bile salts can be detected in the duodenum and proximal jejunum, signifying that
nutrients have entered the small intestine and the gall bladder has emptied (505). The first phase
of GLP secretion also occurs within this time frame (198; 202). Given the absorption of ingested
glucose is almost complete within the first half hour, indicating that glucose is transported in
more proximal portions of the intestine (506), the insulinotropic effect of GLP-1 would therefore
be necessary to combat the ensuing hyperglycemia. On the other hand, dietary lipid is absorbed
more distally (507), likely to allow time for fat emulsification to smaller lipid droplets and
hydrolytic attack by pancreatic lipase in the intestinal lumen (508). The extended bioactivity of
GLP-2 relative to GLP-1 may therefore be more instrumental in intestinal lipid transport.
Therefore, in the physiological setting, the hypolipidemic effect of GLP-1 is limited largely by
its susceptibility to DPP-4, and this presents implications for therapeutic strategies to address
aberrant postprandial lipemia. It appears the hypolipidemic effect of GLP-1 benefits more from
protection from DPP-IV, so the DPP-4 inhibitor sitagliptin (marketed as Januvia) or the long-
acting GLP-1R agonists exenatide and liraglutide present therapies that can improve diabetic

125
dyslipidemia. A schematic of the physiological contributions of GLP-1 and GLP-2 to
postprandial chylomicron secretion is shown in Figure 5-1.

126
Figure 5-1. Physiological contributions of GLP-1 and GLP-2 to chylomicron secretion
The enteroendocrine L-cell cosecretes GLP-1 and GLP-2 in equimolar amounts. GLP-1 can
either directly inhibit TRL production in enterocytes. In hyperglycemia and intact insulin
signalling, GLP-1 stimulates insulin secretion from the pancreatic -cell, which can also
attenuate apoB48 output. The magnitude of GLP-1’s hypolipidemic effect is sensitive to DPP-4.
On the other hand, GLP-2 needs to proceed indirectly by first binding its receptor on ISEMFs,
and this elicits, among other factors, VEGF secretion. VEGF activates the VEGFR2 receptor
tyrosine kinase, which leads to NO activation in the enterocyte and subsequently CD36
activation and dietary FA uptake. The magnitude of GLP-2’s hyperlipidemic effect may depend
on DPP-4 activity as well, but likely to less extent.

127
Despite the complete opposite effects on dietary lipid transport, the biological functions
of GLP-1 and GLP-2 may in fact be complementary. The ileal brake is a primary function
ascribed to GLP-1 and GLP-2 (509), so that when luminal lipids are detected by enteroendocrine
L cells in the distal intestine, thus indicating inefficient proximal nutrient absorption, GLP-1 and
GLP-2 is secreted to slow gastric emptying and prevent duodenal delivery of nutrients. This
phenomenon is evident in MGAT2-deficient mice, in which the delayed kinetics of dietary FA
absorption is accompanied with a two-fold increase in GLP-1 on a high fat diet (54). Similarly,
increases in GLP-1 secretion have also been observed with rats fed a 35% fat diet and the
intestine-specific MTP inhibitor JTT-130 (510). The bulk of dietary FA is normally absorbed by
passive diffusion across the BBM, but by the time dietary lipids could be detected in the distal
intestine, the concentration of lipids in the lumen of the proximal intestine would be considerably
lower. In this case, it would be advantageous to have GLP-2 increase the expression of CD36 at
the apical membrane to provide facilitated transport of luminal FA and stimulate chylomicron
assembly. The stimulatory effect of GLP-2 on lipid transport may also serve to coordinate with
GLP-1 in mediating satiety. After all, central administration of GLP-2 has been shown to inhibit
feeding in rodents (282), and GLP-2 reduces the secretion of the orexigenic hormone ghrelin by
about 10% in humans (297). The GLP-2-increased presence of CD36 at the apical membrane
may be instrumental in transporting oleic acid as a substrate for the synthesis of
oleoylethanolamide (OEA), a satiety factor. Lastly, GLP-2-stimulated VEGF secretion may
have implications for the fate of the chylomicrons. VEGF-B has been shown to promote CD36
expression and lipid uptake in vascular endothelial cells, and these lipids are preferentially
targeted to muscle, heart, and brown adipose tissue where there are high levels of -oxidation
(511). Such a pathway helps to accelerate peripheral clearance of TRL (512), which in the case
of GLP-2-stimulated chylomicron secretion, may help to alleviate the consequences of
postprandial lipemia more rapidly. Moreover, insulin, which is secreted in response to GLP-1, is
well-known to stimulate lipoprotein lipase activity in adipocytes (513), which would help to
catabolize the lipids transported by chylomicrons. Therefore, while the net effect of GLP
secretion may enhance intestinal TRL secretion, the persistence of these lipoproteins in
circulation may not be of major concern.

128
5.6 Future Directions
There are still a lot of questions regarding the modulation of postprandial lipemia by GLP-
1 and GLP-2. For GLP-1, it remains to be determined the signal that directs dietary FA to be
stored in the mucosa as TG (394) rather than for secretion as chylomicrons. The signal could be
a result of direct GLP-1R signalling in the enterocyte or arise from other cell types such as
enteric neurons. Also of fascination is the fate of the dietary lipid that was not acutely secreted.
The possibilities include the FA was oxidized to produce a satiety signal, or mobilized later for
secretion as TRL in the early postprandial period of the next meal - the ‘second meal’ effect
(514). Therefore, it would be interesting to examine oxidative gene expression, mitochondrial
activity, and lipid droplet biology in exendin-4-treated enterocytes. To further explore the
mechanism of GLP-1-inhibted chylomicron secretion, it may be worthwhile to delineate the
contributions of insulin and direct GLP-1 action in enterocytes. Defining these contributions
may have implications for the use of existing therapies in ameliorating diabetic dyslipidemia.
As for GLP-2-stimulated chylomicron secretion, the involvements of other intestinal cell
types have not yet been thoroughly investigated. Enteric neurons are a rich physiological source
of NO donors and have stimulatory effects on intestinal epithelial cells (357), so this is one
intestinal cell type, in addition to subepithelial myofibroblasts, that could potentially signal the
enterocyte to increase chylomicron secretion. The role of enteric glia in GLP-2 action could be
considered using GFAP-HSVtk transgenic mice, which will have ablation of enteric glial cells
following a 14-day administration of ganciclovir (443). In addition, the molecular target of NO
in enterocytes can be identified by the trapping of S-nitrosylated and tyrosine nitrated proteins
and identification by mass spectrometric methods (515; 516). Lastly, mapping out the pertinent
arms of VEGFR2 signalling that drive chylomicron secretion could provide invaluable insight
into the basic regulation of lipoprotein metabolism in enterocytes, be it via the PI3-K/Akt
pathway, through activation of the MAPK cascade, or as a result of direct stimulation of eNOS
itself. Therefore, probing deeper in the mechanism of GLP-1 and GLP-2-regulated postprandial
lipemia could contribute greatly to the understanding of intestinal lipoprotein production.

129
5.7 Conclusions
GLP-1 and GLP-2 are important acute modulators of intestinal lipoprotein secretion and
the interplay of these two co-secreted hormones is important to appropriate handling of dietary
lipids to maximize absorption and to elicit signalling an appropriate postprandial response in
other tissues. While the net physiological and pathophysiological effect may be enhanced
chylomicron secretion due to GLP-2, the hypolipidemic action of GLP-1 presents a therapeutic
strategy to improve postprandial dyslipidemia.

130
PERMISSION TO PUBLISH COPYRIGHTED
MATERIAL

131
SPRINGER LICENSE
TERMS AND CONDITIONS
Aug 09, 2011
This is a License Agreement between Joanne Hsieh ("You") and Springer ("Springer")
provided by Copyright Clearance Center ("CCC"). The license consists of your order
details, the terms and conditions provided by Springer, and the payment terms and
conditions.
All payments must be made in full to CCC. For payment instructions, please see information listed at the bottom of this form.
1. License Number 2724810663588
2. License date Aug 09, 2011
3. Licensed content publisher Springer
4. Licensed content publication Diabetologia
5. Licensed content title The glucagon-like peptide 1 receptor is essential for
postprandial lipoprotein synthesis and secretion in hamsters and mice
6. Licensed content author J. Hsieh
7. Licensed content date Jan 1, 2009
8. Volume number 53
9. Issue number 3
10. Type of Use Thesis/Dissertation
11. Portion Full text
12. Number of copies 1
13. Author of this Springer article Yes and you are a contributor of the new work

132
14. Order reference number
15. Title of your thesis / dissertation
The Acute Regulation of Intestinal Chylomicron Secretion by Glucagon-Like Peptides
16. Expected completion date Nov 2011
17. Estimated size(pages) 200
18. Total 0.00 USD
19. Terms and Conditions
Introduction
The publisher for this copyrighted material is Springer Science + Business Media. By
clicking "accept" in connection with completing this licensing transaction, you agree that
the following terms and conditions apply to this transaction (along with the Billing and
Payment terms and conditions established by Copyright Clearance Center, Inc. ("CCC"), at
the time that you opened your Rightslink account and that are available at any time
at http://myaccount.copyright.com).
Limited License
With reference to your request to reprint in your thesis material on which Springer Science
and Business Media control the copyright, permission is granted, free of charge, for the use
indicated in your enquiry. Licenses are for one-time use only with a maximum distribution
equal to the number that you identified in the licensing process.
This License includes use in an electronic form, provided it is password protected or on the
university's intranet, destined to microfilming by UMI and University repository. For any
other electronic use, please contact Springer at ([email protected] or
The material can only be used for the purpose of defending your thesis, and with a
maximum of 100 extra copies in paper.
Although Springer holds copyright to the material and is entitled to negotiate on rights, this
license is only valid, provided permission is also obtained from the (co) author (address is
given with the article/chapter) and provided it concerns original material which does not
carry references to other sources (if material in question appears with credit to another
source, authorization from that source is required as well). Permission free of charge on this
occasion does not prejudice any rights we might have to charge for reproduction of our
copyrighted material in the future.
Altering/Modifying Material: Not Permitted

133
However figures and illustrations may be altered minimally to serve your work. Any other
abbreviations, additions, deletions and/or any other alterations shall be made only with prior
written authorization of the author(s) and/or Springer Science + Business Media. (Please
contact Springer at [email protected] or
Reservation of Rights
Springer Science + Business Media reserves all rights not specifically granted in the
combination of (i) the license details provided by you and accepted in the course of this
licensing transaction, (ii) these terms and conditions and (iii) CCC's Billing and Payment
terms and conditions.
Copyright Notice:
Please include the following copyright citation referencing the publication in which the
material was originally published. Where wording is within brackets, please include
verbatim.
"With kind permission from Springer Science+Business Media: <book/journal title,
chapter/article title, volume, year of publication, page, name(s) of author(s), figure
number(s), and any original (first) copyright notice displayed with material>."
Warranties: Springer Science + Business Media makes no representations or warranties
with respect to the licensed material.
Indemnity
You hereby indemnify and agree to hold harmless Springer Science + Business Media and
CCC, and their respective officers, directors, employees and agents, from and against any
and all claims arising out of your use of the licensed material other than as specifically
authorized pursuant to this license.
No Transfer of License
This license is personal to you and may not be sublicensed, assigned, or transferred by you
to any other person without Springer Science + Business Media's written permission.
No Amendment Except in Writing
This license may not be amended except in a writing signed by both parties (or, in the case
of Springer Science + Business Media, by CCC on Springer Science + Business Media's
behalf).
Objection to Contrary Terms
Springer Science + Business Media hereby objects to any terms contained in any purchase
order, acknowledgment, check endorsement or other writing prepared by you, which terms
are inconsistent with these terms and conditions or CCC's Billing and Payment terms and
conditions. These terms and conditions, together with CCC's Billing and Payment terms and
conditions (which are incorporated herein), comprise the entire agreement between you and
Springer Science + Business Media (and CCC) concerning this licensing transaction. In the
event of any conflict between your obligations established by these terms and conditions

134
and those established by CCC's Billing and Payment terms and conditions, these terms and
conditions shall control.
Jurisdiction
All disputes that may arise in connection with this present License, or the breach thereof,
shall be settled exclusively by the country's law in which the work was originally published.

135
ELSEVIER LICENSE
TERMS AND CONDITIONS
Aug 11, 2011
This is a License Agreement between Joanne Hsieh ("You") and Elsevier ("Elsevier")
provided by Copyright Clearance Center ("CCC"). The license consists of your order
details, the terms and conditions provided by Elsevier, and the payment terms and
conditions.
All payments must be made in full to CCC. For payment instructions, please see information listed at the bottom of this form.
20. Supplier Elsevier Limited
The Boulevard,Langford Lane Kidlington,Oxford,OX5 1GB,UK
21. Registered Company Number 1982084
22. Customer name Joanne Hsieh
23. Customer address The Hospital for Sick Children
24. Toronto, ON M5G 1X8
25. License number 2724820175995
26. License date Aug 09, 2011
27. Licensed content publisher Elsevier
28. Licensed content publication Atherosclerosis Supplements
29. Licensed content title Postprandial dyslipidemia in insulin resistan Mechanisms and role of intestinal insulin sensitivity
30. Licensed content author Joanne Hsieh,Amanda A. Hayashi,Jennifer Webb,Khosrow Adeli
31. Licensed content date September 2008
32. Licensed content volume 9

136
number
33. Licensed content issue number
2
34. Number of pages 7
35. Start Page 7
36. End Page 13
37. Type of Use reuse in a thesis/dissertation
38. Intended publisher of new work
other
39. Portion figures/tables/illustrations
40. Number of figures/tables/illustrations
1
41. Format both print and electronic
42. Are you the author of this Elsevier article?
Yes
43. Will you be translating? No
44. Order reference number
45. Title of your thesis/dissertation
The Acute Regulation of Intestinal Chylomicron Secretion by Glucagon-Like Peptides
46. Expected completion date Nov 2011
47. Estimated size (number of pages)
200
48. Elsevier VAT number GB 494 6272 12
49. Permissions price 0.00 USD

137
50. VAT/Local Sales Tax 0.0 USD / 0.0 GBP
51. Total 0.00 USD
52. Terms and Conditions
INTRODUCTION
1. The publisher for this copyrighted material is Elsevier. By clicking "accept" in
connection with completing this licensing transaction, you agree that the following terms
and conditions apply to this transaction (along with the Billing and Payment terms and
conditions established by Copyright Clearance Center, Inc. ("CCC"), at the time that you
opened your Rightslink account and that are available at any time
at http://myaccount.copyright.com).
GENERAL TERMS
2. Elsevier hereby grants you permission to reproduce the aforementioned material subject
to the terms and conditions indicated.
3. Acknowledgement: If any part of the material to be used (for example, figures) has
appeared in our publication with credit or acknowledgement to another source, permission
must also be sought from that source. If such permission is not obtained then that material
may not be included in your publication/copies. Suitable acknowledgement to the source
must be made, either as a footnote or in a reference list at the end of your publication, as
follows:
“Reprinted from Publication title, Vol /edition number, Author(s), Title of article / title of
chapter, Pages No., Copyright (Year), with permission from Elsevier [OR APPLICABLE
SOCIETY COPYRIGHT OWNER].” Also Lancet special credit - “Reprinted from The
Lancet, Vol. number, Author(s), Title of article, Pages No., Copyright (Year), with
permission from Elsevier.”
4. Reproduction of this material is confined to the purpose and/or media for which
permission is hereby given.
5. Altering/Modifying Material: Not Permitted. However figures and illustrations may be
altered/adapted minimally to serve your work. Any other abbreviations, additions,
deletions and/or any other alterations shall be made only with prior written authorization
of Elsevier Ltd. (Please contact Elsevier at [email protected])
6. If the permission fee for the requested use of our material is waived in this instance,
please be advised that your future requests for Elsevier materials may attract a fee.
7. Reservation of Rights: Publisher reserves all rights not specifically granted in the

138
combination of (i) the license details provided by you and accepted in the course of this
licensing transaction, (ii) these terms and conditions and (iii) CCC's Billing and Payment
terms and conditions.
8. License Contingent Upon Payment: While you may exercise the rights licensed
immediately upon issuance of the license at the end of the licensing process for the
transaction, provided that you have disclosed complete and accurate details of your
proposed use, no license is finally effective unless and until full payment is received from
you (either by publisher or by CCC) as provided in CCC's Billing and Payment terms and
conditions. If full payment is not received on a timely basis, then any license
preliminarily granted shall be deemed automatically revoked and shall be void as if never
granted. Further, in the event that you breach any of these terms and conditions or any of
CCC's Billing and Payment terms and conditions, the license is automatically revoked and
shall be void as if never granted. Use of materials as described in a revoked license, as
well as any use of the materials beyond the scope of an unrevoked license, may constitute
copyright infringement and publisher reserves the right to take any and all action to
protect its copyright in the materials.
9. Warranties: Publisher makes no representations or warranties with respect to the
licensed material.
10. Indemnity: You hereby indemnify and agree to hold harmless publisher and CCC, and
their respective officers, directors, employees and agents, from and against any and all
claims arising out of your use of the licensed material other than as specifically authorized
pursuant to this license.
11. No Transfer of License: This license is personal to you and may not be sublicensed,
assigned, or transferred by you to any other person without publisher's written permission.
12. No Amendment Except in Writing: This license may not be amended except in a
writing signed by both parties (or, in the case of publisher, by CCC on publisher's behalf).
13. Objection to Contrary Terms: Publisher hereby objects to any terms contained in any
purchase order, acknowledgment, check endorsement or other writing prepared by you,
which terms are inconsistent with these terms and conditions or CCC's Billing and
Payment terms and conditions. These terms and conditions, together with CCC's Billing
and Payment terms and conditions (which are incorporated herein), comprise the entire
agreement between you and publisher (and CCC) concerning this licensing transaction. In
the event of any conflict between your obligations established by these terms and
conditions and those established by CCC's Billing and Payment terms and conditions,
these terms and conditions shall control.
14. Revocation: Elsevier or Copyright Clearance Center may deny the permissions
described in this License at their sole discretion, for any reason or no reason, with a full
refund payable to you. Notice of such denial will be made using the contact information
provided by you. Failure to receive such notice will not alter or invalidate the denial. In

139
no event will Elsevier or Copyright Clearance Center be responsible or liable for any
costs, expenses or damage incurred by you as a result of a denial of your permission
request, other than a refund of the amount(s) paid by you to Elsevier and/or Copyright
Clearance Center for denied permissions.
LIMITED LICENSE
The following terms and conditions apply only to specific license types:
15. Translation: This permission is granted for non-exclusive world English rights only
unless your license was granted for translation rights. If you licensed translation rights
you may only translate this content into the languages you requested. A professional
translator must perform all translations and reproduce the content word for word
preserving the integrity of the article. If this license is to re-use 1 or 2 figures then
permission is granted for non-exclusive world rights in all languages.
16. Website: The following terms and conditions apply to electronic reserve and author
websites:
Electronic reserve: If licensed material is to be posted to website, the web site is to be
password-protected and made available only to bona fide students registered on a relevant
course if:
This license was made in connection with a course,
This permission is granted for 1 year only. You may obtain a license for future website
posting,
All content posted to the web site must maintain the copyright information line on the
bottom of each image,
A hyper-text must be included to the Homepage of the journal from which you are
licensing athttp://www.sciencedirect.com/science/journal/xxxxx or the Elsevier homepage
for books athttp://www.elsevier.com , and
Central Storage: This license does not include permission for a scanned version of the
material to be stored in a central repository such as that provided by Heron/XanEdu.
17. Author website for journals with the following additional clauses:
All content posted to the web site must maintain the copyright information line on the
bottom of each image, and
he permission granted is limited to the personal version of your paper. You are not
allowed to download and post the published electronic version of your article (whether
PDF or HTML, proof or final version), nor may you scan the printed edition to create an
electronic version,
A hyper-text must be included to the Homepage of the journal from which you are
licensing athttp://www.sciencedirect.com/science/journal/xxxxx , As part of our normal
production process, you will receive an e-mail notice when your article appears on
Elsevier’s online service ScienceDirect (www.sciencedirect.com). That e-mail will
include the article’s Digital Object Identifier (DOI). This number provides the electronic
link to the published article and should be included in the posting of your personal

140
version. We ask that you wait until you receive this e-mail and have the DOI to do any
posting.
Central Storage: This license does not include permission for a scanned version of the
material to be stored in a central repository such as that provided by Heron/XanEdu.
18. Author website for books with the following additional clauses:
Authors are permitted to place a brief summary of their work online only.
A hyper-text must be included to the Elsevier homepage at http://www.elsevier.com
All content posted to the web site must maintain the copyright information line on the
bottom of each image
You are not allowed to download and post the published electronic version of your
chapter, nor may you scan the printed edition to create an electronic version.
Central Storage: This license does not include permission for a scanned version of the
material to be stored in a central repository such as that provided by Heron/XanEdu.
19. Website (regular and for author): A hyper-text must be included to the Homepage of
the journal from which you are licensing
at http://www.sciencedirect.com/science/journal/xxxxx. or for books to the Elsevier
homepage at http://www.elsevier.com
20. Thesis/Dissertation: If your license is for use in a thesis/dissertation your thesis may
be submitted to your institution in either print or electronic form. Should your thesis be
published commercially, please reapply for permission. These requirements include
permission for the Library and Archives of Canada to supply single copies, on demand, of
the complete thesis and include permission for UMI to supply single copies, on demand,
of the complete thesis. Should your thesis be published commercially, please reapply for
permission.

141
ELSEVIER LICENSE
TERMS AND CONDITIONS
Nov 09, 2011
This is a License Agreement between Joanne Hsieh ("You") and Elsevier ("Elsevier")
provided by Copyright Clearance Center ("CCC"). The license consists of your order
details, the terms and conditions provided by Elsevier, and the payment terms and
conditions.
All payments must be made in full to CCC. For payment instructions, please see information listed at the bottom of this form.
Supplier Elsevier Limited The Boulevard,Langford Lane Kidlington,Oxford,OX5 1GB,UK
Registered Company Number 1982084
Customer name Joanne Hsieh
Customer address The Hospital for Sick Children
Toronto, ON M5G 1X8
License number 2736901210242
License date Aug 27, 2011
Licensed content publisher Elsevier
Licensed content publication Gastroenterology
Licensed content title Glucagon-Like Peptide-2 Increases Intestinal Lipid Absorption and Chylomicron Production via CD36
Licensed content author Joanne Hsieh,Christine Longuet,Adriano Maida,Jasmine Bahrami,Elaine Xu,Christopher L. Baker,Patricia L.
Brubaker,Daniel J. Drucker,Khosrow Adeli
Licensed content date September 2009
Licensed content volume number 137
Licensed content issue number 3
Number of pages 13
Start Page 997

142
End Page 1005.e4
Type of Use reuse in a thesis/dissertation
Intended publisher of new work other
Portion full article
Format both print and electronic
Are you the author of this Elsevier
article?
Yes
Will you be translating? No
Order reference number
Title of your thesis/dissertation The Acute Regulation of Intestinal Chylomicron Secretion by Glucagon-Like Peptides
Expected completion date Nov 2011
Estimated size (number of pages) 200
Elsevier VAT number GB 494 6272 12
Permissions price 0.00 USD
VAT/Local Sales Tax 0.00 USD / GBP
Total 0.00 USD
Terms and Conditions
INTRODUCTION
1. The publisher for this copyrighted material is Elsevier. By clicking "accept" in
connection with completing this licensing transaction, you agree that the following terms
and conditions apply to this transaction (along with the Billing and Payment terms and
conditions established by Copyright Clearance Center, Inc. ("CCC"), at the time that you
opened your Rightslink account and that are available at any time
at http://myaccount.copyright.com).
GENERAL TERMS
2. Elsevier hereby grants you permission to reproduce the aforementioned material subject
to the terms and conditions indicated.
3. Acknowledgement: If any part of the material to be used (for example, figures) has
appeared in our publication with credit or acknowledgement to another source, permission
must also be sought from that source. If such permission is not obtained then that material

143
may not be included in your publication/copies. Suitable acknowledgement to the source
must be made, either as a footnote or in a reference list at the end of your publication, as
follows:
“Reprinted from Publication title, Vol /edition number, Author(s), Title of article / title of
chapter, Pages No., Copyright (Year), with permission from Elsevier [OR APPLICABLE
SOCIETY COPYRIGHT OWNER].” Also Lancet special credit - “Reprinted from The
Lancet, Vol. number, Author(s), Title of article, Pages No., Copyright (Year), with
permission from Elsevier.”
4. Reproduction of this material is confined to the purpose and/or media for which
permission is hereby given.
5. Altering/Modifying Material: Not Permitted. However figures and illustrations may be
altered/adapted minimally to serve your work. Any other abbreviations, additions,
deletions and/or any other alterations shall be made only with prior written authorization of
Elsevier Ltd. (Please contact Elsevier at [email protected])
6. If the permission fee for the requested use of our material is waived in this instance,
please be advised that your future requests for Elsevier materials may attract a fee.
7. Reservation of Rights: Publisher reserves all rights not specifically granted in the
combination of (i) the license details provided by you and accepted in the course of this
licensing transaction, (ii) these terms and conditions and (iii) CCC's Billing and Payment
terms and conditions.
8. License Contingent Upon Payment: While you may exercise the rights licensed
immediately upon issuance of the license at the end of the licensing process for the
transaction, provided that you have disclosed complete and accurate details of your
proposed use, no license is finally effective unless and until full payment is received from
you (either by publisher or by CCC) as provided in CCC's Billing and Payment terms and
conditions. If full payment is not received on a timely basis, then any license preliminarily
granted shall be deemed automatically revoked and shall be void as if never

144
granted. Further, in the event that you breach any of these terms and conditions or any of
CCC's Billing and Payment terms and conditions, the license is automatically revoked and
shall be void as if never granted. Use of materials as described in a revoked license, as
well as any use of the materials beyond the scope of an unrevoked license, may constitute
copyright infringement and publisher reserves the right to take any and all action to protect
its copyright in the materials.
9. Warranties: Publisher makes no representations or warranties with respect to the
licensed material.
10. Indemnity: You hereby indemnify and agree to hold harmless publisher and CCC, and
their respective officers, directors, employees and agents, from and against any and all
claims arising out of your use of the licensed material other than as specifically authorized
pursuant to this license.
11. No Transfer of License: This license is personal to you and may not be sublicensed,
assigned, or transferred by you to any other person without publisher's written permission.
12. No Amendment Except in Writing: This license may not be amended except in a
writing signed by both parties (or, in the case of publisher, by CCC on publisher's behalf).
13. Objection to Contrary Terms: Publisher hereby objects to any terms contained in any
purchase order, acknowledgment, check endorsement or other writing prepared by you,
which terms are inconsistent with these terms and conditions or CCC's Billing and
Payment terms and conditions. These terms and conditions, together with CCC's Billing
and Payment terms and conditions (which are incorporated herein), comprise the entire
agreement between you and publisher (and CCC) concerning this licensing transaction. In
the event of any conflict between your obligations established by these terms and
conditions and those established by CCC's Billing and Payment terms and conditions, these
terms and conditions shall control.
14. Revocation: Elsevier or Copyright Clearance Center may deny the permissions
described in this License at their sole discretion, for any reason or no reason, with a full

145
refund payable to you. Notice of such denial will be made using the contact information
provided by you. Failure to receive such notice will not alter or invalidate the denial. In
no event will Elsevier or Copyright Clearance Center be responsible or liable for any costs,
expenses or damage incurred by you as a result of a denial of your permission request,
other than a refund of the amount(s) paid by you to Elsevier and/or Copyright Clearance
Center for denied permissions.
LIMITED LICENSE
The following terms and conditions apply only to specific license types:
15. Translation: This permission is granted for non-exclusive world English rights only
unless your license was granted for translation rights. If you licensed translation rights you
may only translate this content into the languages you requested. A professional translator
must perform all translations and reproduce the content word for word preserving the
integrity of the article. If this license is to re-use 1 or 2 figures then permission is granted
for non-exclusive world rights in all languages.
16. Website: The following terms and conditions apply to electronic reserve and author
websites:
Electronic reserve: If licensed material is to be posted to website, the web site is to be
password-protected and made available only to bona fide students registered on a relevant
course if:
This license was made in connection with a course,
This permission is granted for 1 year only. You may obtain a license for future website
posting,
All content posted to the web site must maintain the copyright information line on the
bottom of each image,
A hyper-text must be included to the Homepage of the journal from which you are
licensing athttp://www.sciencedirect.com/science/journal/xxxxx or the Elsevier homepage
for books athttp://www.elsevier.com , and
Central Storage: This license does not include permission for a scanned version of the

146
material to be stored in a central repository such as that provided by Heron/XanEdu.
17. Author website for journals with the following additional clauses:
All content posted to the web site must maintain the copyright information line on the
bottom of each image, and
he permission granted is limited to the personal version of your paper. You are not
allowed to download and post the published electronic version of your article (whether
PDF or HTML, proof or final version), nor may you scan the printed edition to create an
electronic version,
A hyper-text must be included to the Homepage of the journal from which you are
licensing athttp://www.sciencedirect.com/science/journal/xxxxx , As part of our normal
production process, you will receive an e-mail notice when your article appears on
Elsevier’s online service ScienceDirect (www.sciencedirect.com). That e-mail will
include the article’s Digital Object Identifier (DOI). This number provides the electronic
link to the published article and should be included in the posting of your personal
version. We ask that you wait until you receive this e-mail and have the DOI to do any
posting.
Central Storage: This license does not include permission for a scanned version of the
material to be stored in a central repository such as that provided by Heron/XanEdu.
18. Author website for books with the following additional clauses:
Authors are permitted to place a brief summary of their work online only.
A hyper-text must be included to the Elsevier homepage at http://www.elsevier.com
All content posted to the web site must maintain the copyright information line on the
bottom of each image
You are not allowed to download and post the published electronic version of your
chapter, nor may you scan the printed edition to create an electronic version.
Central Storage: This license does not include permission for a scanned version of the
material to be stored in a central repository such as that provided by Heron/XanEdu.

147
19. Website (regular and for author): A hyper-text must be included to the Homepage of
the journal from which you are licensing
at http://www.sciencedirect.com/science/journal/xxxxx. or for books to the Elsevier
homepage at http://www.elsevier.com
20. Thesis/Dissertation: If your license is for use in a thesis/dissertation your thesis may
be submitted to your institution in either print or electronic form. Should your thesis be
published commercially, please reapply for permission. These requirements include
permission for the Library and Archives of Canada to supply single copies, on demand, of
the complete thesis and include permission for UMI to supply single copies, on demand, of
the complete thesis. Should your thesis be published commercially, please reapply for
permission.
21. Other Conditions:
v1.6
If you would like to pay for this license now, please remit this license along with your
payment made payable to "COPYRIGHT CLEARANCE CENTER" otherwise you will be invoiced within 48 hours of the license date. Payment should be in the form of a check or money order referencing your account number and this invoice number RLNK11045309. Once you receive your invoice for this order, you may pay your invoice by credit card. Please follow instructions provided at that time.
Make Payment To: Copyright Clearance Center Dept 001 P.O. Box 843006 Boston, MA 02284-3006
For suggestions or comments regarding this order, contact RightsLink Customer Support:[email protected] or +1-877-622-5543 (toll free in the US) or +1-978-646-2777.
Gratis licenses (referencing $0 in the Total field) are free. Please retain this printable license for your reference. No payment is required.

148
REFERENCES
1. World Health Organization. The global burden of disease: 2004 update. 2008.
2. Finucane MM, Stevens GA, Cowan MJ, Danaei G, Lin JK, Paciorek CJ, Singh GM,
Gutierrez HR, Lu Y, Bahalim AN, Farzadfar F, Riley LM, Ezzati M. National, regional,
and global trends in body-mass index since 1980: systematic analysis of health
examination surveys and epidemiological studies with 960 country-years and 9.1 million
participants. Lancet 2011;377:557-567.
3. Boquist S, Ruotolo G, Tang R, Bjorkegren J, Bond MG, de Faire U, Karpe F, Hamsten A.
Alimentary lipemia, postprandial triglyceride-rich lipoproteins, and common carotid
intima-media thickness in healthy, middle-aged men. Circulation 1999;100:723-728.
4. McNamara JR, Shah PK, Nakajima K, Cupples LA, Wilson PW, Ordovas JM, Schaefer
EJ. Remnant-like particle (RLP) cholesterol is an independent cardiovascular disease risk
factor in women: results from the Framingham Heart Study. Atherosclerosis
2001;154:229-236.
5. Karpe F, Boquist S, Tang R, Bond GM, de Faire U, Hamsten A. Remnant lipoproteins
are related to intima-media thickness of the carotid artery independently of LDL
cholesterol and plasma triglycerides. J Lipid Res 2001;42:17-21.
6. Karpe F, de Faire U, Mercuri M, Bond MG, Hellenius ML, Hamsten A. Magnitude of
alimentary lipemia is related to intima-media thickness of the common carotid artery in
middle-aged men. Atherosclerosis 1998;141:307-314.
7. Pal S, Semorine K, Watts GF, Mamo J. Identification of lipoproteins of intestinal origin
in human atherosclerotic plaque. Clin Chem Lab Med 2003;41:792-795.
8. Proctor SD, Mamo JC. Intimal retention of cholesterol derived from apolipoprotein.
Arterioscler Thromb Vasc Biol 2003;23:1595-1600.
9. Doi H, Kugiyama K, Oka H, Sugiyama S, Ogata N, Koide SI, Nakamura SI, Yasue H.
Remnant lipoproteins induce proatherothrombogenic molecules in endothelial cells
through a redox-sensitive mechanism. Circulation 2000;102:670-676.
10. Mero N, Syvanne M, Taskinen MR. Postprandial lipid metabolism in diabetes.
Atherosclerosis 1998;141 Suppl 1:S53-S55.
11. Lopez-Miranda J, Marin C. Dietary, Physiological, and Genetic Impacts on Postprandial
Lipid Metabolism. 2010.
12. Mu H, Hoy CE. The digestion of dietary triacylglycerols. Prog Lipid Res 2004;43:105-
133.

149
13. Glatz JF, Luiken JJ, Bonen A. Membrane fatty acid transporters as regulators of lipid
metabolism: implications for metabolic disease. Physiol Rev 2010;90:367-417.
14. Pohl J, Ring A, Ehehalt R, Schulze-Bergkamen H, Schad A, Verkade P, Stremmel W.
Long-chain fatty acid uptake into adipocytes depends on lipid raft function. Biochemistry
2004;43:4179-4187.
15. Pohl J, Ring A, Korkmaz U, Ehehalt R, Stremmel W. FAT/CD36-mediated long-chain
fatty acid uptake in adipocytes requires plasma membrane rafts. Mol Biol Cell
2005;16:24-31.
16. Nickerson JG, Alkhateeb H, Benton CR, Lally J, Nickerson J, Han XX, Wilson MH, Jain
SS, Snook LA, Glatz JF, Chabowski A, Luiken JJ, Bonen A. Greater transport
efficiencies of the membrane fatty acid transporters FAT/CD36 and FATP4 compared
with FABPpm and FATP1 and differential effects on fatty acid esterification and
oxidation in rat skeletal muscle. J Biol Chem 2009;284:16522-16530.
17. Coburn CT, Knapp FF, Jr., Febbraio M, Beets AL, Silverstein RL, Abumrad NA.
Defective uptake and utilization of long chain fatty acids in muscle and adipose tissues of
CD36 knockout mice. J Biol Chem 2000;275:32523-32529.
18. Koonen DP, Jacobs RL, Febbraio M, Young ME, Soltys CL, Ong H, Vance DE, Dyck
JR. Increased hepatic CD36 expression contributes to dyslipidemia associated with diet-
induced obesity. Diabetes 2007;56:2863-2871.
19. Nassir F, Wilson B, Han X, Gross RW, Abumrad NA. CD36 is important for fatty acid
and cholesterol uptake by the proximal but not distal intestine. J Biol Chem
2007;282:19493-19501.
20. Masson CJ, Plat J, Mensink RP, Namiot A, Kisielewski W, Namiot Z, Fullekrug J,
Ehehalt R, Glatz JF, Pelsers MM. Fatty acid- and cholesterol transporter protein
expression along the human intestinal tract. PLoS One 2010;5:e10380.
21. Nauli AM, Nassir F, Zheng S, Yang Q, Lo CM, Vonlehmden SB, Lee D, Jandacek RJ,
Abumrad NA, Tso P. CD36 is important for chylomicron formation and secretion and
may mediate cholesterol uptake in the proximal intestine. Gastroenterology
2006;131:1197-1207.
22. Drover VA, Nguyen DV, Bastie CC, Darlington YF, Abumrad NA, Pessin JE, London E,
Sahoo D, Phillips MC. CD36 mediates both cellular uptake of very long chain fatty acids
and their intestinal absorption in mice. J Biol Chem 2008;283:13108-13115.
23. Drover VA, Ajmal M, Nassir F, Davidson NO, Nauli AM, Sahoo D, Tso P, Abumrad
NA. CD36 deficiency impairs intestinal lipid secretion and clearance of chylomicrons
from the blood. J Clin Invest 2005;115:1290-1297.

150
24. Laugerette F, Passilly-Degrace P, Patris B, Niot I, Febbraio M, Montmayeur JP, Besnard
P. CD36 involvement in orosensory detection of dietary lipids, spontaneous fat
preference, and digestive secretions. J Clin Invest 2005;115:3177-3184.
25. Cai SF, Kirby RJ, Howles PN, Hui DY. Differentiation-dependent expression and
localization of the class B type I scavenger receptor in intestine. J Lipid Res
2001;42:902-909.
26. Bietrix F, Yan D, Nauze M, Rolland C, Bertrand-Michel J, Comera C, Schaak S,
Barbaras R, Groen AK, Perret B, Terce F, Collet X. Accelerated lipid absorption in mice
overexpressing intestinal SR-BI. J Biol Chem 2006;281:7214-7219.
27. Hauser H, Dyer JH, Nandy A, Vega MA, Werder M, Bieliauskaite E, Weber FE,
Compassi S, Gemperli A, Boffelli D, Wehrli E, Schulthess G, Phillips MC. Identification
of a receptor mediating absorption of dietary cholesterol in the intestine. Biochemistry
1998;37:17843-17850.
28. Labonte ED, Howles PN, Granholm NA, Rojas JC, Davies JP, Ioannou YA, Hui DY.
Class B type I scavenger receptor is responsible for the high affinity cholesterol binding
activity of intestinal brush border membrane vesicles. Biochim Biophys Acta
2007;1771:1132-1139.
29. Hansen GH, Niels-Christiansen LL, Immerdal L, Danielsen EM. Scavenger receptor class
B type I (SR-BI) in pig enterocytes: trafficking from the brush border to lipid droplets
during fat absorption. Gut 2003;52:1424-1431.
30. Nguyen DV, Drover VA, Knopfel M, Dhanasekaran P, Hauser H, Phillips MC. Influence
of class B scavenger receptors on cholesterol flux across the brush border membrane and
intestinal absorption. J Lipid Res 2009;50:2235-2244.
31. Garcia-Calvo M, Lisnock J, Bull HG, Hawes BE, Burnett DA, Braun MP, Crona JH,
Davis HR, Jr., Dean DC, Detmers PA, Graziano MP, Hughes M, Macintyre DE, Ogawa
A, O'neill KA, Iyer SP, Shevell DE, Smith MM, Tang YS, Makarewicz AM, Ujjainwalla
F, Altmann SW, Chapman KT, Thornberry NA. The target of ezetimibe is Niemann-Pick
C1-Like 1 (NPC1L1). Proc Natl Acad Sci U S A 2005;102:8132-8137.
32. Altmann SW, Davis HR, Jr., Zhu LJ, Yao X, Hoos LM, Tetzloff G, Iyer SP, Maguire M,
Golovko A, Zeng M, Wang L, Murgolo N, Graziano MP. Niemann-Pick C1 Like 1
protein is critical for intestinal cholesterol absorption. Science 2004;303:1201-1204.
33. Ge L, Wang J, Qi W, Miao HH, Cao J, Qu YX, Li BL, Song BL. The cholesterol
absorption inhibitor ezetimibe acts by blocking the sterol-induced internalization of
NPC1L1. Cell Metab 2008;7:508-519.
34. Ge L, Qi W, Wang LJ, Miao HH, Qu YX, Li BL, Song BL. Flotillins play an essential
role in Niemann-Pick C1-like 1-mediated cholesterol uptake. Proc Natl Acad Sci U S A
2011;108:551-556.

151
35. Martin G, Nemoto M, Gelman L, Geffroy S, Najib J, Fruchart JC, Roevens P, de
Martinville B, Deeb S, Auwerx J. The human fatty acid transport protein-1 (SLC27A1;
FATP-1) cDNA and gene: organization, chromosomal localization, and expression.
Genomics 2000;66:296-304.
36. Stahl A, Hirsch DJ, Gimeno RE, Punreddy S, Ge P, Watson N, Patel S, Kotler M,
Raimondi A, Tartaglia LA, Lodish HF. Identification of the major intestinal fatty acid
transport protein. Mol Cell 1999;4:299-308.
37. Milger K, Herrmann T, Becker C, Gotthardt D, Zickwolf J, Ehehalt R, Watkins PA,
Stremmel W, Fullekrug J. Cellular uptake of fatty acids driven by the ER-localized acyl-
CoA synthetase FATP4. J Cell Sci 2006;119:4678-4688.
38. Gimeno RE, Hirsch DJ, Punreddy S, Sun Y, Ortegon AM, Wu H, Daniels T, Stricker-
Krongrad A, Lodish HF, Stahl A. Targeted deletion of fatty acid transport protein-4
results in early embryonic lethality. J Biol Chem 2003;278:49512-49516.
39. Hall AM, Wiczer BM, Herrmann T, Stremmel W, Bernlohr DA. Enzymatic properties of
purified murine fatty acid transport protein 4 and analysis of acyl-CoA synthetase
activities in tissues from FATP4 null mice. J Biol Chem 2005;280:11948-11954.
40. Shim J, Moulson CL, Newberry EP, Lin MH, Xie Y, Kennedy SM, Miner JH, Davidson
NO. Fatty acid transport protein 4 is dispensable for intestinal lipid absorption in mice. J
Lipid Res 2009;50:491-500.
41. Hatch GM, Smith AJ, Xu FY, Hall AM, Bernlohr DA. FATP1 channels exogenous FA
into 1,2,3-triacyl-sn-glycerol and down-regulates sphingomyelin and cholesterol
metabolism in growing 293 cells. J Lipid Res 2002;43:1380-1389.
42. Stremmel W, Lotz G, Strohmeyer G, Berk PD. Identification, isolation, and partial
characterization of a fatty acid binding protein from rat jejunal microvillous membranes.
J Clin Invest 1985;75:1068-1076.
43. Newberry EP, Xie Y, Kennedy S, Han X, Buhman KK, Luo J, Gross RW, Davidson NO.
Decreased hepatic triglyceride accumulation and altered fatty acid uptake in mice with
deletion of the liver fatty acid-binding protein gene. J Biol Chem 2003;278:51664-51672.
44. Murphy EJ. L-FABP and I-FABP expression increase NBD-stearate uptake and
cytoplasmic diffusion in L cells. Am J Physiol 1998;275:G244-G249.
45. Thompson J, Winter N, Terwey D, Bratt J, Banaszak L. The crystal structure of the liver
fatty acid-binding protein. A complex with two bound oleates. J Biol Chem
1997;272:7140-7150.
46. Alpers DH, Bass NM, Engle MJ, DeSchryver-Kecskemeti K. Intestinal fatty acid binding
protein may favor differential apical fatty acid binding in the intestine. Biochim Biophys
Acta 2000;1483:352-362.

152
47. Newberry EP, Xie Y, Kennedy SM, Luo J, Davidson NO. Protection against Western
diet-induced obesity and hepatic steatosis in liver fatty acid-binding protein knockout
mice. Hepatology 2006;44:1191-1205.
48. Vassileva G, Huwyler L, Poirier K, Agellon LB, Toth MJ. The intestinal fatty acid
binding protein is not essential for dietary fat absorption in mice. FASEB J
2000;14:2040-2046.
49. Lagakos WS, Gajda AM, Agellon L, Binas B, Choi V, Mandap B, Russnak T, Zhou YX,
Storch J. Different functions of intestinal and liver-type fatty acid-binding proteins in
intestine and in whole body energy homeostasis. Am J Physiol Gastrointest Liver Physiol
2011;300:G803-G814.
50. Agellon LB, Drozdowski L, Li L, Iordache C, Luong L, Clandinin MT, Uwiera RR, Toth
MJ, Thomson AB. Loss of intestinal fatty acid binding protein increases the susceptibility
of male mice to high fat diet-induced fatty liver. Biochim Biophys Acta 2007;1771:1283-
1288.
51. Xie Y, Newberry EP, Kennedy SM, Luo J, Davidson NO. Increased susceptibility to diet-
induced gallstones in liver fatty acid binding protein knockout mice. J Lipid Res
2009;50:977-987.
52. Cao J, Hawkins E, Brozinick J, Liu X, Zhang H, Burn P, Shi Y. A predominant role of
acyl-CoA:monoacylglycerol acyltransferase-2 in dietary fat absorption implicated by
tissue distribution, subcellular localization, and up-regulation by high fat diet. J Biol
Chem 2004;279:18878-18886.
53. Cheng D, Nelson TC, Chen J, Walker SG, Wardwell-Swanson J, Meegalla R, Taub R,
Billheimer JT, Ramaker M, Feder JN. Identification of acyl coenzyme
A:monoacylglycerol acyltransferase 3, an intestinal specific enzyme implicated in dietary
fat absorption. J Biol Chem 2003;278:13611-13614.
54. Yen CL, Cheong ML, Grueter C, Zhou P, Moriwaki J, Wong JS, Hubbard B, Marmor S,
Farese RV, Jr. Deficiency of the intestinal enzyme acyl CoA:monoacylglycerol
acyltransferase-2 protects mice from metabolic disorders induced by high-fat feeding.
Nat Med 2009;15:442-446.
55. Cheng D, Iqbal J, Devenny J, Chu CH, Chen L, Dong J, Seethala R, Keim WJ, Azzara
AV, Lawrence RM, Pelleymounter MA, Hussain MM. Acylation of acylglycerols by acyl
coenzyme A:diacylglycerol acyltransferase 1 (DGAT1). Functional importance of
DGAT1 in the intestinal fat absorption. J Biol Chem 2008;283:29802-29811.
56. Buhman KK, Smith SJ, Stone SJ, Repa JJ, Wong JS, Knapp FF, Jr., Burri BJ, Hamilton
RL, Abumrad NA, Farese RV, Jr. DGAT1 is not essential for intestinal triacylglycerol
absorption or chylomicron synthesis. J Biol Chem 2002;277:25474-25479.

153
57. Lee B, Fast AM, Zhu J, Cheng JX, Buhman KK. Intestine-specific expression of acyl
CoA:diacylglycerol acyltransferase 1 reverses resistance to diet-induced hepatic steatosis
and obesity in Dgat1-/- mice. J Lipid Res 2010;51:1770-1780.
58. Buhman KK, Accad M, Novak S, Choi RS, Wong JS, Hamilton RL, Turley S, Farese
RV, Jr. Resistance to diet-induced hypercholesterolemia and gallstone formation in
ACAT2-deficient mice. Nat Med 2000;6:1341-1347.
59. Powell LM, Wallis SC, Pease RJ, Edwards YH, Knott TJ, Scott J. A novel form of tissue-
specific RNA processing produces apolipoprotein-B48 in intestine. Cell 1987;50:831-
840.
60. Hussain MM, Kancha RK, Zhou Z, Luchoomun J, Zu H, Bakillah A. Chylomicron
assembly and catabolism: role of apolipoproteins and receptors. Biochim Biophys Acta
1996;1300:151-170.
61. Singh K, Batuman OA, Akman HO, Kedees MH, Vakil V, Hussain MM. Differential,
tissue-specific, transcriptional regulation of apolipoprotein B secretion by transforming
growth factor beta. J Biol Chem 2002;277:39515-39524.
62. Ashur-Fabian O, Har-Zahav A, Shaish A, Wiener Amram HW, Margalit O, Weizer-Stern
O, Dominissini D, Harats D, Amariglio N, Rechavi G. apoB and apobec1, two genes key
to lipid metabolism, are transcriptionally regulated by p53. Cell Cycle 2010;9:3761-3770.
63. Sparks JD, Cianci J, Jokinen J, Chen LS, Sparks CE. Interleukin-6 mediates hepatic
hypersecretion of apolipoprotein B. Am J Physiol Gastrointest Liver Physiol
2010;299:G980-G989.
64. Sidiropoulos KG, Meshkani R, Avramoglu-Kohen R, Adeli K. Insulin inhibition of
apolipoprotein B mRNA translation is mediated via the PI-3 kinase/mTOR signaling
cascade but does not involve internal ribosomal entry site (IRES) initiation. Arch
Biochem Biophys 2007;465:380-388.
65. Sidiropoulos KG, Zastepa A, Adeli K. Translational control of apolipoprotein B mRNA
via insulin and the protein kinase C signaling cascades: evidence for modulation of RNA-
protein interactions at the 5'UTR. Arch Biochem Biophys 2007;459:10-19.
66. Sidiropoulos KG, Pontrelli L, Adeli K. Insulin-mediated suppression of apolipoprotein B
mRNA translation requires the 5' UTR and is characterized by decreased binding of an
insulin-sensitive 110-kDa 5' UTR RNA-binding protein. Biochemistry 2005;44:12572-
12581.
67. Pontrelli L, Sidiropoulos KG, Adeli K. Translational control of apolipoprotein B mRNA:
regulation via cis elements in the 5' and 3' untranslated regions. Biochemistry
2004;43:6734-6744.
68. Hussain MM. A proposed model for the assembly of chylomicrons. Atherosclerosis
2000;148:1-15.

154
69. Jiang ZG, Liu Y, Hussain MM, Atkinson D, McKnight CJ. Reconstituting initial events
during the assembly of apolipoprotein B-containing lipoproteins in a cell-free system. J
Mol Biol 2008;383:1181-1194.
70. Cartwright IJ, Plonne D, Higgins JA. Intracellular events in the assembly of
chylomicrons in rabbit enterocytes. J Lipid Res 2000;41:1728-1739.
71. Haidari M, Leung N, Mahbub F, Uffelman KD, Kohen-Avramoglu R, Lewis GF, Adeli
K. Fasting and postprandial overproduction of intestinally derived lipoproteins in an
animal model of insulin resistance. Evidence that chronic fructose feeding in the hamster
is accompanied by enhanced intestinal de novo lipogenesis and ApoB48-containing
lipoprotein overproduction. J Biol Chem 2002;277:31646-31655.
72. Zoltowska M, Ziv E, Delvin E, Sinnett D, Kalman R, Garofalo C, Seidman E, Levy E.
Cellular aspects of intestinal lipoprotein assembly in Psammomys obesus: a model of
insulin resistance and type 2 diabetes. Diabetes 2003;52:2539-2545.
73. Liao W, Chan L. Apolipoprotein B, a paradigm for proteins regulated by intracellular
degradation, does not undergo intracellular degradation in CaCo2 cells. J Biol Chem
2000;275:3950-3956.
74. Morel E, Demignot S, Chateau D, Chambaz J, Rousset M, Delers F. Lipid-dependent
bidirectional traffic of apolipoprotein B in polarized enterocytes. Mol Biol Cell
2004;15:132-141.
75. Xie Y, Nassir F, Luo J, Buhman K, Davidson NO. Intestinal lipoprotein assembly in
apobec-1-/- mice reveals subtle alterations in triglyceride secretion coupled with a shift to
larger lipoproteins. Am J Physiol Gastrointest Liver Physiol 2003;285:G735-G746.
76. Guo Q, Avramoglu RK, Adeli K. Intestinal assembly and secretion of highly dense/lipid-
poor apolipoprotein B48-containing lipoprotein particles in the fasting state: evidence for
induction by insulin resistance and exogenous fatty acids. Metabolism 2005;54:689-697.
77. Lu S, Yao Y, Cheng X, Mitchell S, Leng S, Meng S, Gallagher JW, Shelness GS, Morris
GS, Mahan J, Frase S, Mansbach CM, Weinberg RB, Black DD. Overexpression of
apolipoprotein A-IV enhances lipid secretion in IPEC-1 cells by increasing chylomicron
size. J Biol Chem 2006;281:3473-3483.
78. Cartwright IJ, Higgins JA. Direct evidence for a two-step assembly of ApoB48-
containing lipoproteins in the lumen of the smooth endoplasmic reticulum of rabbit
enterocytes. J Biol Chem 2001;276:48048-48057.
79. Hayashi H, Fujimoto K, Cardelli JA, Nutting DF, Bergstedt S, Tso P. Fat feeding
increases size, but not number, of chylomicrons produced by small intestine. Am J
Physiol 1990;259:G709-G719.

155
80. Siddiqi SA, Gorelick FS, Mahan JT, Mansbach CM. COPII proteins are required for
Golgi fusion but not for endoplasmic reticulum budding of the pre-chylomicron transport
vesicle. J Cell Sci 2003;116:415-427.
81. Siddiqi S, Siddiqi SA, Mansbach CM. Sec24C is required for docking the
prechylomicron transport vesicle with the Golgi. J Lipid Res 2010;51:1093-1100.
82. Siddiqi SA, Siddiqi S, Mahan J, Peggs K, Gorelick FS, Mansbach CM. The identification
of a novel endoplasmic reticulum to Golgi SNARE complex used by the prechylomicron
transport vesicle. J Biol Chem 2006;281:20974-20982.
83. Siddiqi S, Saleem U, Abumrad NA, Davidson NO, Storch J, Siddiqi SA, Mansbach CM.
A novel multiprotein complex is required to generate the prechylomicron transport
vesicle from intestinal ER. J Lipid Res 2010;51:1918-1928.
84. Neeli I, Siddiqi SA, Siddiqi S, Mahan J, Lagakos WS, Binas B, Gheyi T, Storch J,
Mansbach CM. Liver fatty acid-binding protein initiates budding of pre-chylomicron
transport vesicles from intestinal endoplasmic reticulum. J Biol Chem 2007;282:17974-
17984.
85. Siddiqi SA, Mansbach CM. PKC zeta-mediated phosphorylation controls budding of the
pre-chylomicron transport vesicle. J Cell Sci 2008;121:2327-2338.
86. Iqbal J, Hussain MM. Intestinal lipid absorption. Am J Physiol Endocrinol Metab
2009;296:E1183-E1194.
87. Rac ME, Safranow K, Poncyljusz W. Molecular basis of human CD36 gene mutations.
Mol Med 2007;13:288-296.
88. Kashiwagi H, Tomiyama Y, Nozaki S, Kiyoi T, Tadokoro S, Matsumoto K, Honda S,
Kosugi S, Kurata Y, Matsuzawa Y. Analyses of genetic abnormalities in type I CD36
deficiency in Japan: identification and cell biological characterization of two novel
mutations that cause CD36 deficiency in man. Hum Genet 2001;108:459-466.
89. Kashiwagi H, Tomiyama Y, Honda S, Kosugi S, Shiraga M, Nagao N, Sekiguchi S,
Kanayama Y, Kurata Y, Matsuzawa Y. Molecular basis of CD36 deficiency. Evidence
that a 478C-->T substitution (proline90-->serine) in CD36 cDNA accounts for CD36
deficiency. J Clin Invest 1995;95:1040-1046.
90. Kajihara S, Hisatomi A, Ogawa Y, Yasutake T, Yoshimura T, Hara T, Mizuta T, Ozaki I,
Iwamoto N, Yamamoto K. Association of the Pro90Ser CD36 mutation with elevated
free fatty acid concentrations but not with insulin resistance syndrome in Japanese. Clin
Chim Acta 2001;314:125-130.
91. Miyaoka K, Kuwasako T, Hirano K, Nozaki S, Yamashita S, Matsuzawa Y. CD36
deficiency associated with insulin resistance. Lancet 2001;357:686-687.

156
92. Kuwasako T, Hirano K, Sakai N, Ishigami M, Hiraoka H, Yakub MJ, Yamauchi-
Takihara K, Yamashita S, Matsuzawa Y. Lipoprotein abnormalities in human genetic
CD36 deficiency associated with insulin resistance and abnormal fatty acid metabolism.
Diabetes Care 2003;26:1647-1648.
93. Febbraio M, Abumrad NA, Hajjar DP, Sharma K, Cheng W, Pearce SF, Silverstein RL.
A null mutation in murine CD36 reveals an important role in fatty acid and lipoprotein
metabolism. J Biol Chem 1999;274:19055-19062.
94. Ma X, Bacci S, Mlynarski W, Gottardo L, Soccio T, Menzaghi C, Iori E, Lager RA,
Shroff AR, Gervino EV, Nesto RW, Johnstone MT, Abumrad NA, Avogaro A, Trischitta
V, Doria A. A common haplotype at the CD36 locus is associated with high free fatty
acid levels and increased cardiovascular risk in Caucasians. Hum Mol Genet
2004;13:2197-2205.
95. Tanaka T, Nakata T, Oka T, Ogawa T, Okamoto F, Kusaka Y, Sohmiya K, Shimamoto
K, Itakura K. Defect in human myocardial long-chain fatty acid uptake is caused by
FAT/CD36 mutations. J Lipid Res 2001;42:751-759.
96. Goudriaan JR, den Boer MA, Rensen PC, Febbraio M, Kuipers F, Romijn JA, Havekes
LM, Voshol PJ. CD36 deficiency in mice impairs lipoprotein lipase-mediated triglyceride
clearance. J Lipid Res 2005;46:2175-2181.
97. Bastie CC, Hajri T, Drover VA, Grimaldi PA, Abumrad NA. CD36 in myocytes channels
fatty acids to a lipase-accessible triglyceride pool that is related to cell lipid and insulin
responsiveness. Diabetes 2004;53:2209-2216.
98. Masuda D, Hirano K, Oku H, Sandoval JC, Kawase R, Yuasa-Kawase M, Yamashita Y,
Takada M, Tsubakio-Yamamoto K, Tochino Y, Koseki M, Matsuura F, Nishida M,
Kawamoto T, Ishigami M, Hori M, Shimomura I, Yamashita S. Chylomicron remnants
are increased in the postprandial state in CD36 deficiency. J Lipid Res 2009;50:999-
1011.
99. Lepretre F, Vasseur F, Vaxillaire M, Scherer PE, Ali S, Linton K, Aitman T, Froguel P.
A CD36 nonsense mutation associated with insulin resistance and familial type 2
diabetes. Hum Mutat 2004;24:104.
100. Baillie AG, Coburn CT, Abumrad NA. Reversible binding of long-chain fatty acids to
purified FAT, the adipose CD36 homolog. J Membr Biol 1996;153:75-81.
101. Robitaille J, Brouillette C, Lemieux S, Perusse L, Gaudet D, Vohl MC. Plasma
concentrations of apolipoprotein B are modulated by a gene--diet interaction effect
between the LFABP T94A polymorphism and dietary fat intake in French-Canadian men.
Mol Genet Metab 2004;82:296-303.
102. Fisher E, Weikert C, Klapper M, Lindner I, Mohlig M, Spranger J, Boeing H,
Schrezenmeir J, Doring F. L-FABP T94A is associated with fasting triglycerides and
LDL-cholesterol in women. Mol Genet Metab 2007;91:278-284.

157
103. Gao N, Qu X, Yan J, Huang Q, Yuan HY, Ouyang DS. L-FABP T94A decreased fatty
acid uptake and altered hepatic triglyceride and cholesterol accumulation in Chang liver
cells stably transfected with L-FABP. Mol Cell Biochem 2010;345:207-214.
104. Linden D, Lindberg K, Oscarsson J, Claesson C, Asp L, Li L, Gustafsson M, Boren J,
Olofsson SO. Influence of peroxisome proliferator-activated receptor alpha agonists on
the intracellular turnover and secretion of apolipoprotein (Apo) B-100 and ApoB-48. J
Biol Chem 2002;277:23044-23053.
105. Brouillette C, Bosse Y, Perusse L, Gaudet D, Vohl MC. Effect of liver fatty acid binding
protein (FABP) T94A missense mutation on plasma lipoprotein responsiveness to
treatment with fenofibrate. J Hum Genet 2004;49:424-432.
106. Poirier H, Niot I, Monnot MC, Braissant O, Meunier-Durmort C, Costet P, Pineau T,
Wahli W, Willson TM, Besnard P. Differential involvement of peroxisome-proliferator-
activated receptors alpha and delta in fibrate and fatty-acid-mediated inductions of the
gene encoding liver fatty-acid-binding protein in the liver and the small intestine.
Biochem J 2001;355:481-488.
107. Baier LJ, Sacchettini JC, Knowler WC, Eads J, Paolisso G, Tataranni PA, Mochizuki H,
Bennett PH, Bogardus C, Prochazka M. An amino acid substitution in the human
intestinal fatty acid binding protein is associated with increased fatty acid binding,
increased fat oxidation, and insulin resistance. J Clin Invest 1995;95:1281-1287.
108. Zhang F, Lucke C, Baier LJ, Sacchettini JC, Hamilton JA. Solution structure of human
intestinal fatty acid binding protein with a naturally-occurring single amino acid
substitution (A54T) that is associated with altered lipid metabolism. Biochemistry
2003;42:7339-7347.
109. Nakanishi S, Yamane K, Kamei N, Okubo M, Kohno N. The effect of polymorphism in
the intestinal fatty acid-binding protein 2 gene on fat metabolism is associated with
gender and obesity amongst non-diabetic Japanese-Americans. Diabetes Obes Metab
2004;6:45-49.
110. Georgopoulos A, Aras O, Tsai MY. Codon-54 polymorphism of the fatty acid-binding
protein 2 gene is associated with elevation of fasting and postprandial triglyceride in type
2 diabetes. J Clin Endocrinol Metab 2000;85:3155-3160.
111. Stan S, Lambert M, Delvin E, Paradis G, O'loughlin J, Hanley JA, Levy E. Intestinal fatty
acid binding protein and microsomal triglyceride transfer protein polymorphisms in
French-Canadian youth. J Lipid Res 2005;46:320-327.
112. Baier LJ, Bogardus C, Sacchettini JC. A polymorphism in the human intestinal fatty acid
binding protein alters fatty acid transport across Caco-2 cells. J Biol Chem
1996;271:10892-10896.
113. Marin C, Perez-Jimenez F, Gomez P, Delgado J, Paniagua JA, Lozano A, Cortes B,
Jimenez-Gomez Y, Gomez MJ, Lopez-Miranda J. The Ala54Thr polymorphism of the

158
fatty acid-binding protein 2 gene is associated with a change in insulin sensitivity after a
change in the type of dietary fat. Am J Clin Nutr 2005;82:196-200.
114. Pratley RE, Baier L, Pan DA, Salbe AD, Storlien L, Ravussin E, Bogardus C. Effects of
an Ala54Thr polymorphism in the intestinal fatty acid-binding protein on responses to
dietary fat in humans. J Lipid Res 2000;41:2002-2008.
115. Kim CH, Yun SK, Byun DW, Yoo MH, Lee KU, Suh KI. Codon 54 polymorphism of the
fatty acid binding protein 2 gene is associated with increased fat oxidation and
hyperinsulinemia, but not with intestinal fatty acid absorption in Korean men.
Metabolism 2001;50:473-476.
116. Tahvanainen E, Molin M, Vainio S, Tiret L, Nicaud V, Farinaro E, Masana L, Ehnholm
C. Intestinal fatty acid binding protein polymorphism at codon 54 is not associated with
postprandial responses to fat and glucose tolerance tests in healthy young Europeans.
Results from EARS II participants. Atherosclerosis 2000;152:317-325.
117. Klar J, Schweiger M, Zimmerman R, Zechner R, Li H, Torma H, Vahlquist A, Bouadjar
B, Dahl N, Fischer J. Mutations in the fatty acid transport protein 4 gene cause the
ichthyosis prematurity syndrome. Am J Hum Genet 2009;85:248-253.
118. Herrmann T, van der HF, Grone HJ, Stewart AF, Langbein L, Kaiser I, Liebisch G,
Gosch I, Buchkremer F, Drobnik W, Schmitz G, Stremmel W. Mice with targeted
disruption of the fatty acid transport protein 4 (Fatp 4, Slc27a4) gene show features of
lethal restrictive dermopathy. J Cell Biol 2003;161:1105-1115.
119. Gertow K, Bellanda M, Eriksson P, Boquist S, Hamsten A, Sunnerhagen M, Fisher RM.
Genetic and structural evaluation of fatty acid transport protein-4 in relation to markers of
the insulin resistance syndrome. J Clin Endocrinol Metab 2004;89:392-399.
120. Gertow K, Skoglund-Andersson C, Eriksson P, Boquist S, Orth-Gomer K, Schenck-
Gustafsson K, Hamsten A, Fisher RM. A common polymorphism in the fatty acid
transport protein-1 gene associated with elevated post-prandial lipaemia and alterations in
LDL particle size distribution. Atherosclerosis 2003;167:265-273.
121. Macleod JJ. Insulin and diabetes: A general statement of the physiological and
therapeutic effects of insulin. Br Med J 1922;2:833-835.
122. Taniguchi CM, Emanuelli B, Kahn CR. Critical nodes in signalling pathways: insights
into insulin action. Nat Rev Mol Cell Biol 2006;7:85-96.
123. Bryant NJ, Govers R, James DE. Regulated transport of the glucose transporter GLUT4.
Nat Rev Mol Cell Biol 2002;3:267-277.
124. Michael MD, Kulkarni RN, Postic C, Previs SF, Shulman GI, Magnuson MA, Kahn CR.
Loss of insulin signaling in hepatocytes leads to severe insulin resistance and progressive
hepatic dysfunction. Mol Cell 2000;6:87-97.

159
125. Gromada J, Franklin I, Wollheim CB. Alpha-cells of the endocrine pancreas: 35 years of
research but the enigma remains. Endocr Rev 2007;28:84-116.
126. Bluher M, Michael MD, Peroni OD, Ueki K, Carter N, Kahn BB, Kahn CR. Adipose
tissue selective insulin receptor knockout protects against obesity and obesity-related
glucose intolerance. Dev Cell 2002;3:25-38.
127. Bezaire V, Langin D. Regulation of adipose tissue lipolysis revisited. Proc Nutr Soc
2009;68:350-360.
128. Krycer JR, Sharpe LJ, Luu W, Brown AJ. The Akt-SREBP nexus: cell signaling meets
lipid metabolism. Trends Endocrinol Metab 2010;21:268-276.
129. Wong RH, Sul HS. Insulin signaling in fatty acid and fat synthesis: a transcriptional
perspective. Curr Opin Pharmacol 2010;10:684-691.
130. Brownsey RW, Boone AN, Elliott JE, Kulpa JE, Lee WM. Regulation of acetyl-CoA
carboxylase. Biochem Soc Trans 2006;34:223-227.
131. Yecies JL, Zhang HH, Menon S, Liu S, Yecies D, Lipovsky AI, Gorgun C, Kwiatkowski
DJ, Hotamisligil GS, Lee CH, Manning BD. Akt stimulates hepatic SREBP1c and
lipogenesis through parallel mTORC1-dependent and independent pathways. Cell Metab
2011;14:21-32.
132. Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ. Sources of
fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty
liver disease. J Clin Invest 2005;115:1343-1351.
133. Lewis GF, Uffelman KD, Szeto LW, Steiner G. Effects of acute hyperinsulinemia on
VLDL triglyceride and VLDL apoB production in normal weight and obese individuals.
Diabetes 1993;42:833-842.
134. Chirieac DV, Chirieac LR, Corsetti JP, Cianci J, Sparks CE, Sparks JD. Glucose-
stimulated insulin secretion suppresses hepatic triglyceride-rich lipoprotein and apoB
production. Am J Physiol Endocrinol Metab 2000;279:E1003-E1011.
135. Patsch W, Franz S, Schonfeld G. Role of insulin in lipoprotein secretion by cultured rat
hepatocytes. J Clin Invest 1983;71:1161-1174.
136. Sparks CE, Sparks JD, Bolognino M, Salhanick A, Strumph PS, Amatruda JM. Insulin
effects on apolipoprotein B lipoprotein synthesis and secretion by primary cultures of rat
hepatocytes. Metabolism 1986;35:1128-1136.
137. Malmstrom R, Packard CJ, Caslake M, Bedford D, Stewart P, Yki-Jarvinen H, Shepherd
J, Taskinen MR. Effects of insulin and acipimox on VLDL1 and VLDL2 apolipoprotein
B production in normal subjects. Diabetes 1998;47:779-787.

160
138. Kamagate A, Qu S, Perdomo G, Su D, Kim DH, Slusher S, Meseck M, Dong HH. FoxO1
mediates insulin-dependent regulation of hepatic VLDL production in mice. J Clin Invest
2008;118:2347-2364.
139. Brown AM, Gibbons GF. Insulin inhibits the maturation phase of VLDL assembly via a
phosphoinositide 3-kinase-mediated event. Arterioscler Thromb Vasc Biol 2001;21:1656-
1661.
140. Brown MS, Goldstein JL. Selective versus total insulin resistance: a pathogenic paradox.
Cell Metab 2008;7:95-96.
141. Diraison F, Moulin P, Beylot M. Contribution of hepatic de novo lipogenesis and
reesterification of plasma non esterified fatty acids to plasma triglyceride synthesis
during non-alcoholic fatty liver disease. Diabetes Metab 2003;29:478-485.
142. Fabbrini E, Mohammed BS, Magkos F, Korenblat KM, Patterson BW, Klein S.
Alterations in adipose tissue and hepatic lipid kinetics in obese men and women with
nonalcoholic fatty liver disease. Gastroenterology 2008;134:424-431.
143. Kuriyama H, Yamashita S, Shimomura I, Funahashi T, Ishigami M, Aragane K, Miyaoka
K, Nakamura T, Takemura K, Man Z, Toide K, Nakayama N, Fukuda Y, Lin MC,
Wetterau JR, Matsuzawa Y. Enhanced expression of hepatic acyl-coenzyme A synthetase
and microsomal triglyceride transfer protein messenger RNAs in the obese and
hypertriglyceridemic rat with visceral fat accumulation. Hepatology 1998;27:557-562.
144. Bartels ED, Lauritsen M, Nielsen LB. Hepatic expression of microsomal triglyceride
transfer protein and in vivo secretion of triglyceride-rich lipoproteins are increased in
obese diabetic mice. Diabetes 2002;51:1233-1239.
145. Taghibiglou C, Carpentier A, Van Iderstine SC, Chen B, Rudy D, Aiton A, Lewis GF,
Adeli K. Mechanisms of hepatic very low density lipoprotein overproduction in insulin
resistance. Evidence for enhanced lipoprotein assembly, reduced intracellular ApoB
degradation, and increased microsomal triglyceride transfer protein in a fructose-fed
hamster model. J Biol Chem 2000;275:8416-8425.
146. Taghibiglou C, Rashid-Kolvear F, Van Iderstine SC, Le-Tien H, Fantus IG, Lewis GF,
Adeli K. Hepatic very low density lipoprotein-ApoB overproduction is associated with
attenuated hepatic insulin signaling and overexpression of protein-tyrosine phosphatase
1B in a fructose-fed hamster model of insulin resistance. J Biol Chem 2002;277:793-803.
147. De Man FH, Cabezas MC, Van Barlingen HH, Erkelens DW, de Bruin TW. Triglyceride-
rich lipoproteins in non-insulin-dependent diabetes mellitus: post-prandial metabolism
and relation to premature atherosclerosis. Eur J Clin Invest 1996;26:89-108.
148. Curtin A, Deegan P, Owens D, Collins P, Johnson A, Tomkin GH. Elevated triglyceride-
rich lipoproteins in diabetes. A study of apolipoprotein B-48. Acta Diabetol 1996;33:205-
210.

161
149. Lewis GF, O'Meara NM, Soltys PA, Blackman JD, Iverius PH, Druetzler AF, Getz GS,
Polonsky KS. Postprandial lipoprotein metabolism in normal and obese subjects:
comparison after the vitamin A fat-loading test. J Clin Endocrinol Metab 1990;71:1041-
1050.
150. Chen YD, Swami S, Skowronski R, Coulston A, Reaven GM. Differences in postprandial
lipemia between patients with normal glucose tolerance and noninsulin-dependent
diabetes mellitus. J Clin Endocrinol Metab 1993;76:172-177.
151. Rivellese AA, De Natale C, Di Marino L, Patti L, Iovine C, Coppola S, Del Prato S,
Riccardi G, Annuzzi G. Exogenous and endogenous postprandial lipid abnormalities in
type 2 diabetic patients with optimal blood glucose control and optimal fasting
triglyceride levels. J Clin Endocrinol Metab 2004;89:2153-2159.
152. Duez H, Lamarche B, Uffelman KD, Valero R, Cohn JS, Lewis GF. Hyperinsulinemia is
associated with increased production rate of intestinal apolipoprotein B-48-containing
lipoproteins in humans. Arterioscler Thromb Vasc Biol 2006;26:1357-1363.
153. Lewis GF, Uffelman K, Naples M, Szeto L, Haidari M, Adeli K. Intestinal lipoprotein
overproduction, a newly recognized component of insulin resistance, is ameliorated by
the insulin sensitizer rosiglitazone: studies in the fructose-fed Syrian golden hamster.
Endocrinology 2005;146:247-255.
154. Federico LM, Naples M, Taylor D, Adeli K. Intestinal insulin resistance and aberrant
production of apolipoprotein B48 lipoproteins in an animal model of insulin resistance
and metabolic dyslipidemia: evidence for activation of protein tyrosine phosphatase-1B,
extracellular signal-related kinase, and sterol regulatory element-binding protein-1c in the
fructose-fed hamster intestine. Diabetes 2006;55:1316-1326.
155. Casaschi A, Maiyoh GK, Adeli K, Theriault AG. Increased diacylglycerol acyltransferase
activity is associated with triglyceride accumulation in tissues of diet-induced insulin-
resistant hyperlipidemic hamsters. Metabolism 2005;54:403-409.
156. Wong DM, Webb JP, Malinowski PM, Xu E, Macri J, Adeli K. Proteomic profiling of
intestinal prechylomicron transport vesicle (PCTV)-associated proteins in an animal
model of insulin resistance (94 char). J Proteomics 2010;73:1291-1305.
157. Phillips C, Owens D, Collins P, Tomkin GH. Microsomal triglyceride transfer protein:
does insulin resistance play a role in the regulation of chylomicron assembly?
Atherosclerosis 2002;160:355-360.
158. Mangat R, Su J, Scott PG, Russell JC, Vine DF, Proctor SD. Chylomicron and apoB48
metabolism in the JCR:LA corpulent rat, a model for the metabolic syndrome. Biochem
Soc Trans 2007;35:477-481.
159. Morton NM, Emilsson V, Liu YL, Cawthorne MA. Leptin action in intestinal cells. J Biol
Chem 1998;273:26194-26201.

162
160. Iqbal J, Li X, Chang BH, Chan L, Schwartz GJ, Chua SC, Jr., Hussain MM. An intrinsic
gut leptin-melanocortin pathway modulates intestinal microsomal triglyceride transfer
protein and lipid absorption. J Lipid Res 2010;51:1929-1942.
161. Petit V, Arnould L, Martin P, Monnot MC, Pineau T, Besnard P, Niot I. Chronic high-fat
diet affects intestinal fat absorption and postprandial triglyceride levels in the mouse. J
Lipid Res 2007;48:278-287.
162. Harbis A, Defoort C, Narbonne H, Juhel C, Senft M, Latge C, Delenne B, Portugal H,
Atlan-Gepner C, Vialettes B, Lairon D. Acute hyperinsulinism modulates plasma
apolipoprotein B-48 triglyceride-rich lipoproteins in healthy subjects during the
postprandial period. Diabetes 2001;50:462-469.
163. Pavlic M, Xiao C, Szeto L, Patterson BW, Lewis GF. Insulin acutely inhibits intestinal
lipoprotein secretion in humans in part by suppressing plasma free fatty acids. Diabetes
2010;59:580-587.
164. Duez H, Lamarche B, Valero R, Pavlic M, Proctor S, Xiao C, Szeto L, Patterson BW,
Lewis GF. Both intestinal and hepatic lipoprotein production are stimulated by an acute
elevation of plasma free fatty acids in humans. Circulation 2008;117:2369-2376.
165. Qin B, Qiu W, Avramoglu RK, Adeli K. Tumor necrosis factor-alpha induces intestinal
insulin resistance and stimulates the overproduction of intestinal apolipoprotein B48-
containing lipoproteins. Diabetes 2007;56:450-461.
166. Levy E, Thibault L, Delvin E, Menard D. Apolipoprotein synthesis in human fetal
intestine: regulation by epidermal growth factor. Biochem Biophys Res Commun
1994;204:1340-1345.
167. Murthy S, Mathur SN, Varilek G, Bishop W, Field FJ. Cytokines regulate apolipoprotein
B secretion by Caco-2 cells: differential effects of IL-6 and TGF-beta 1. Am J Physiol
1996;270:G94-102.
168. Murthy S, Mathur SN, Field FJ. Tumor necrosis factor-alpha and interleukin-1beta inhibit
apolipoprotein B secretion in CaCo-2 cells via the epidermal growth factor receptor
signaling pathway. J Biol Chem 2000;275:9222-9229.
169. Newsholme EA, Carrie AL. Quantitative aspects of glucose and glutamine metabolism
by intestinal cells. Gut 1994;35:S13-S17.
170. Windmueller HG, Spaeth AE. Respiratory fuels and nitrogen metabolism in vivo in small
intestine of fed rats. Quantitative importance of glutamine, glutamate, and aspartate. J
Biol Chem 1980;255:107-112.
171. Mori T, Kondo H, Hase T, Tokimitsu I, Murase T. Dietary fish oil upregulates intestinal
lipid metabolism and reduces body weight gain in C57BL/6J mice. J Nutr
2007;137:2629-2634.

163
172. Kondo H, Minegishi Y, Komine Y, Mori T, Matsumoto I, Abe K, Tokimitsu I, Hase T,
Murase T. Differential regulation of intestinal lipid metabolism-related genes in obesity-
resistant A/J vs. obesity-prone C57BL/6J mice. Am J Physiol Endocrinol Metab
2006;291:E1092-E1099.
173. Langhans W, Leitner C, Arnold M. Dietary fat sensing via fatty acid oxidation in
enterocytes: possible role in the control of eating. Am J Physiol Regul Integr Comp
Physiol 2011;300:R554-R565.
174. Chavez-Jauregui RN, Mattes RD, Parks EJ. Dynamics of fat absorption and effect of
sham feeding on postprandial lipema. Gastroenterology 2010;139:1538-1548.
175. Robertson MD, Parkes M, Warren BF, Ferguson DJ, Jackson KG, Jewell DP, Frayn KN.
Mobilisation of enterocyte fat stores by oral glucose in humans. Gut 2003;52:834-839.
176. Zhu J, Lee B, Buhman KK, Cheng JX. A dynamic, cytoplasmic triacylglycerol pool in
enterocytes revealed by ex vivo and in vivo coherent anti-Stokes Raman scattering
imaging. J Lipid Res 2009;50:1080-1089.
177. Tall AR, Green PH, Glickman RM, Riley JW. Metabolic fate of chylomicron
phospholipids and apoproteins in the rat. J Clin Invest 1979;64:977-989.
178. Havel RJ, Kane JP, Kashyap ML. Interchange of apolipoproteins between chylomicrons
and high density lipoproteins during alimentary lipemia in man. J Clin Invest 1973;52:32-
38.
179. Blum CB. Dynamics of apolipoprotein E metabolism in humans. J Lipid Res
1982;23:1308-1316.
180. Goldberg IJ. Lipoprotein lipase and lipolysis: central roles in lipoprotein metabolism and
atherogenesis. J Lipid Res 1996;37:693-707.
181. Bharadwaj KG, Hiyama Y, Hu Y, Huggins LA, Ramakrishnan R, Abumrad NA,
Shulman GI, Blaner WS, Goldberg IJ. Chylomicron- and VLDL-derived lipids enter the
heart through different pathways: in vivo evidence for receptor- and non-receptor-
mediated fatty acid uptake. J Biol Chem 2010;285:37976-37986.
182. Augustus AS, Buchanan J, Park TS, Hirata K, Noh HL, Sun J, Homma S, D'armiento J,
Abel ED, Goldberg IJ. Loss of lipoprotein lipase-derived fatty acids leads to increased
cardiac glucose metabolism and heart dysfunction. J Biol Chem 2006;281:8716-8723.
183. Picard F, Naimi N, Richard D, Deshaies Y. Response of adipose tissue lipoprotein lipase
to the cephalic phase of insulin secretion. Diabetes 1999;48:452-459.
184. Miles JM, Park YS, Walewicz D, Russell-Lopez C, Windsor S, Isley WL, Coppack SW,
Harris WS. Systemic and forearm triglyceride metabolism: fate of lipoprotein lipase-
generated glycerol and free fatty acids. Diabetes 2004;53:521-527.

164
185. Beigneux AP, Davies BS, Gin P, Weinstein MM, Farber E, Qiao X, Peale F, Bunting S,
Walzem RL, Wong JS, Blaner WS, Ding ZM, Melford K, Wongsiriroj N, Shu X, de
Sauvage F, Ryan RO, Fong LG, Bensadoun A, Young SG. Glycosylphosphatidylinositol-
anchored high-density lipoprotein-binding protein 1 plays a critical role in the lipolytic
processing of chylomicrons. Cell Metab 2007;5:279-291.
186. Weinstein MM, Yin L, Tu Y, Wang X, Wu X, Castellani LW, Walzem RL, Lusis AJ,
Fong LG, Beigneux AP, Young SG. Chylomicronemia elicits atherosclerosis in mice--
brief report. Arterioscler Thromb Vasc Biol 2010;30:20-23.
187. Ishibashi S, Perrey S, Chen Z, Osuga J, Shimada M, Ohashi K, Harada K, Yazaki Y,
Yamada N. Role of the low density lipoprotein (LDL) receptor pathway in the
metabolism of chylomicron remnants. A quantitative study in knockout mice lacking the
LDL receptor, apolipoprotein E, or both. J Biol Chem 1996;271:22422-22427.
188. Beisiegel U, Weber W, Ihrke G, Herz J, Stanley KK. The LDL-receptor-related protein,
LRP, is an apolipoprotein E-binding protein. Nature 1989;341:162-164.
189. Fujino T, Asaba H, Kang MJ, Ikeda Y, Sone H, Takada S, Kim DH, Ioka RX, Ono M,
Tomoyori H, Okubo M, Murase T, Kamataki A, Yamamoto J, Magoori K, Takahashi S,
Miyamoto Y, Oishi H, Nose M, Okazaki M, Usui S, Imaizumi K, Yanagisawa M, Sakai
J, Yamamoto TT. Low-density lipoprotein receptor-related protein 5 (LRP5) is essential
for normal cholesterol metabolism and glucose-induced insulin secretion. Proc Natl Acad
Sci U S A 2003;100:229-234.
190. Bataille D. Pro-protein convertases in intermediary metabolism: islet hormones, brain/gut
hormones and integrated physiology. J Mol Med (Berl) 2007;85:673-684.
191. Orskov C, Rabenhoj L, Wettergren A, Kofod H, Holst JJ. Tissue and plasma
concentrations of amidated and glycine-extended glucagon-like peptide I in humans.
Diabetes 1994;43:535-539.
192. Deacon CF, Johnsen AH, Holst JJ. Degradation of glucagon-like peptide-1 by human
plasma in vitro yields an N-terminally truncated peptide that is a major endogenous
metabolite in vivo. J Clin Endocrinol Metab 1995;80:952-957.
193. Goke R, Fehmann HC, Linn T, Schmidt H, Krause M, Eng J, Goke B. Exendin-4 is a
high potency agonist and truncated exendin-(9-39)-amide an antagonist at the glucagon-
like peptide 1-(7-36)-amide receptor of insulin-secreting beta-cells. J Biol Chem
1993;268:19650-19655.
194. Schirra J, Sturm K, Leicht P, Arnold R, Goke B, Katschinski M. Exendin(9-39)amide is
an antagonist of glucagon-like peptide-1(7-36)amide in humans. J Clin Invest
1998;101:1421-1430.
195. Drucker DJ, Shi Q, Crivici A, Sumner-Smith M, Tavares W, Hill M, DeForest L, Cooper
S, Brubaker PL. Regulation of the biological activity of glucagon-like peptide 2 in vivo
by dipeptidyl peptidase IV. Nat Biotechnol 1997;15:673-677.

165
196. Shin ED, Estall JL, Izzo A, Drucker DJ, Brubaker PL. Mucosal adaptation to enteral
nutrients is dependent on the physiologic actions of glucagon-like peptide-2 in mice.
Gastroenterology 2005;128:1340-1353.
197. Thulesen J, Knudsen LB, Hartmann B, Hastrup S, Kissow H, Jeppesen PB, Orskov C,
Holst JJ, Poulsen SS. The truncated metabolite GLP-2 (3-33) interacts with the GLP-2
receptor as a partial agonist. Regul Pept 2002;103:9-15.
198. Naslund E, Bogefors J, Skogar S, Gryback P, Jacobsson H, Holst JJ, Hellstrom PM.
GLP-1 slows solid gastric emptying and inhibits insulin, glucagon, and PYY release in
humans. Am J Physiol 1999;277:R910-R916.
199. Herrmann C, Goke R, Richter G, Fehmann HC, Arnold R, Goke B. Glucagon-like
peptide-1 and glucose-dependent insulin-releasing polypeptide plasma levels in response
to nutrients. Digestion 1995;56:117-126.
200. Jeppesen PB, Hartmann B, Hansen BS, Thulesen J, Holst JJ, Mortensen PB. Impaired
meal stimulated glucagon-like peptide 2 response in ileal resected short bowel patients
with intestinal failure. Gut 1999;45:559-563.
201. Brubaker PL, Crivici A, Izzo A, Ehrlich P, Tsai CH, Drucker DJ. Circulating and tissue
forms of the intestinal growth factor, glucagon-like peptide-2. Endocrinology
1997;138:4837-4843.
202. Xiao Q, Boushey RP, Drucker DJ, Brubaker PL. Secretion of the intestinotropic hormone
glucagon-like peptide 2 is differentially regulated by nutrients in humans.
Gastroenterology 1999;117:99-105.
203. Rocca AS, Brubaker PL. Role of the vagus nerve in mediating proximal nutrient-induced
glucagon-like peptide-1 secretion. Endocrinology 1999;140:1687-1694.
204. Anini Y, Hansotia T, Brubaker PL. Muscarinic receptors control postprandial release of
glucagon-like peptide-1: in vivo and in vitro studies in rats. Endocrinology
2002;143:2420-2426.
205. Hansen L, Holst JJ. The effects of duodenal peptides on glucagon-like peptide-1 secretion
from the ileum. A duodeno--ileal loop? Regul Pept 2002;110:39-45.
206. Rijkelijkhuizen JM, McQuarrie K, Girman CJ, Stein PP, Mari A, Holst JJ, Nijpels G,
Dekker JM. Effects of meal size and composition on incretin, alpha-cell, and beta-cell
responses. Metabolism 2010;59:502-511.
207. Vilsboll T, Krarup T, Sonne J, Madsbad S, Volund A, Juul AG, Holst JJ. Incretin
secretion in relation to meal size and body weight in healthy subjects and people with
type 1 and type 2 diabetes mellitus. J Clin Endocrinol Metab 2003;88:2706-2713.
208. D'Alessio D, Lu W, Sun W, Zheng S, Yang Q, Seeley R, Woods SC, Tso P. Fasting and
postprandial concentrations of GLP-1 in intestinal lymph and portal plasma: evidence for

166
selective release of GLP-1 in the lymph system. Am J Physiol Regul Integr Comp
Physiol 2007;293:R2163-R2169.
209. Iakoubov R, Ahmed A, Lauffer LM, Bazinet RP, Brubaker PL. Essential Role for Protein
Kinase C{zeta} in Oleic Acid-Induced Glucagon-Like Peptide-1 Secretion in Vivo in the
Rat. Endocrinology 2011;152:1244-1252.
210. Thomsen C, Rasmussen O, Lousen T, Holst JJ, Fenselau S, Schrezenmeir J, Hermansen
K. Differential effects of saturated and monounsaturated fatty acids on postprandial
lipemia and incretin responses in healthy subjects. Am J Clin Nutr 1999;69:1135-1143.
211. Beysen C, Karpe F, Fielding BA, Clark A, Levy JC, Frayn KN. Interaction between
specific fatty acids, GLP-1 and insulin secretion in humans. Diabetologia 2002;45:1533-
1541.
212. Rocca AS, LaGreca J, Kalitsky J, Brubaker PL. Monounsaturated fatty acid diets improve
glycemic tolerance through increased secretion of glucagon-like peptide-1.
Endocrinology 2001;142:1148-1155.
213. Edfalk S, Steneberg P, Edlund H. Gpr40 is expressed in enteroendocrine cells and
mediates free fatty acid stimulation of incretin secretion. Diabetes 2008;57:2280-2287.
214. Hirasawa A, Tsumaya K, Awaji T, Katsuma S, Adachi T, Yamada M, Sugimoto Y,
Miyazaki S, Tsujimoto G. Free fatty acids regulate gut incretin glucagon-like peptide-1
secretion through GPR120. Nat Med 2005;11:90-94.
215. Oh DY, Talukdar S, Bae EJ, Imamura T, Morinaga H, Fan W, Li P, Lu WJ, Watkins SM,
Olefsky JM. GPR120 is an omega-3 fatty acid receptor mediating potent anti-
inflammatory and insulin-sensitizing effects. Cell 2010;142:687-698.
216. Schwartz GJ, Fu J, Astarita G, Li X, Gaetani S, Campolongo P, Cuomo V, Piomelli D.
The lipid messenger OEA links dietary fat intake to satiety. Cell Metab 2008;8:281-288.
217. Lauffer LM, Iakoubov R, Brubaker PL. GPR119 is essential for oleoylethanolamide-
induced glucagon-like peptide-1 secretion from the intestinal enteroendocrine L-cell.
Diabetes 2009;58:1058-1066.
218. Iakoubov R, Izzo A, Yeung A, Whiteside CI, Brubaker PL. Protein kinase Czeta is
required for oleic acid-induced secretion of glucagon-like peptide-1 by intestinal
endocrine L cells. Endocrinology 2007;148:1089-1098.
219. Thomas C, Gioiello A, Noriega L, Strehle A, Oury J, Rizzo G, Macchiarulo A,
Yamamoto H, Mataki C, Pruzanski M, Pellicciari R, Auwerx J, Schoonjans K. TGR5-
mediated bile acid sensing controls glucose homeostasis. Cell Metab 2009;10:167-177.
220. Ranganath LR, Beety JM, Morgan LM, Wright JW, Howland R, Marks V. Attenuated
GLP-1 secretion in obesity: cause or consequence? Gut 1996;38:916-919.

167
221. Rask E, Olsson T, Soderberg S, Johnson O, Seckl J, Holst JJ, Ahren B. Impaired incretin
response after a mixed meal is associated with insulin resistance in nondiabetic men.
Diabetes Care 2001;24:1640-1645.
222. Rask E, Olsson T, Soderberg S, Holst JJ, Tura A, Pacini G, Ahren B. Insulin secretion
and incretin hormones after oral glucose in non-obese subjects with impaired glucose
tolerance. Metabolism 2004;53:624-631.
223. Vilsboll T, Krarup T, Deacon CF, Madsbad S, Holst JJ. Reduced postprandial
concentrations of intact biologically active glucagon-like peptide 1 in type 2 diabetic
patients. Diabetes 2001;50:609-613.
224. Lugari R, Dei CA, Ugolotti D, Finardi L, Barilli AL, Ognibene C, Luciani A,
Zandomeneghi R, Gnudi A. Evidence for early impairment of glucagon-like peptide 1-
induced insulin secretion in human type 2 (non insulin-dependent) diabetes. Horm Metab
Res 2002;34:150-154.
225. Lim GE, Huang GJ, Flora N, LeRoith D, Rhodes CJ, Brubaker PL. Insulin regulates
glucagon-like peptide-1 secretion from the enteroendocrine L cell. Endocrinology
2009;150:580-591.
226. Anini Y, Brubaker PL. Role of leptin in the regulation of glucagon-like peptide-1
secretion. Diabetes 2003;52:252-259.
227. Reimer RA, Russell JC. Glucose tolerance, lipids, and GLP-1 secretion in JCR:LA-cp
rats fed a high protein fiber diet. Obesity (Silver Spring) 2008;16:40-46.
228. Gniuli D, Calcagno A, Dalla LL, Calvani R, Leccesi L, Caristo ME, Vettor R, Castagneto
M, Ghirlanda G, Mingrone G. High-fat feeding stimulates endocrine, glucose-dependent
insulinotropic polypeptide (GIP)-expressing cell hyperplasia in the duodenum of Wistar
rats. Diabetologia 2010;53:2233-2240.
229. Thulesen J, Hartmann B, Nielsen C, Holst JJ, Poulsen SS. Diabetic intestinal growth
adaptation and glucagon-like peptide 2 in the rat: effects of dietary fibre. Gut
1999;45:672-678.
230. Fischer KD, Dhanvantari S, Drucker DJ, Brubaker PL. Intestinal growth is associated
with elevated levels of glucagon-like peptide 2 in diabetic rats. Am J Physiol
1997;273:E815-E820.
231. Kieffer TJ, Habener JF. The glucagon-like peptides. Endocr Rev 1999;20:876-913.
232. Mojsov S, Weir GC, Habener JF. Insulinotropin: glucagon-like peptide I (7-37) co-
encoded in the glucagon gene is a potent stimulator of insulin release in the perfused rat
pancreas. J Clin Invest 1987;79:616-619.
233. Kreymann B, Williams G, Ghatei MA, Bloom SR. Glucagon-like peptide-1 7-36: a
physiological incretin in man. Lancet 1987;2:1300-1304.

168
234. Baggio L, Kieffer TJ, Drucker DJ. Glucagon-like peptide-1, but not glucose-dependent
insulinotropic peptide, regulates fasting glycemia and nonenteral glucose clearance in
mice. Endocrinology 2000;141:3703-3709.
235. Wang Y, Egan JM, Raygada M, Nadiv O, Roth J, Montrose-Rafizadeh C. Glucagon-like
peptide-1 affects gene transcription and messenger ribonucleic acid stability of
components of the insulin secretory system in RIN 1046-38 cells. Endocrinology
1995;136:4910-4917.
236. Fehmann HC, Habener JF. Insulinotropic hormone glucagon-like peptide-I(7-37)
stimulation of proinsulin gene expression and proinsulin biosynthesis in insulinoma beta
TC-1 cells. Endocrinology 1992;130:159-166.
237. Holz GG, Kuhtreiber WM, Habener JF. Pancreatic beta-cells are rendered glucose-
competent by the insulinotropic hormone glucagon-like peptide-1(7-37). Nature
1993;361:362-365.
238. Pugazhenthi U, Velmurugan K, Tran A, Mahaffey G, Pugazhenthi S. Anti-inflammatory
action of exendin-4 in human islets is enhanced by phosphodiesterase inhibitors: potential
therapeutic benefits in diabetic patients. Diabetologia 2010;53:2357-2368.
239. Yusta B, Baggio LL, Estall JL, Koehler JA, Holland DP, Li H, Pipeleers D, Ling Z,
Drucker DJ. GLP-1 receptor activation improves beta cell function and survival
following induction of endoplasmic reticulum stress. Cell Metab 2006;4:391-406.
240. Farilla L, Hui H, Bertolotto C, Kang E, Bulotta A, Di Mario U, Perfetti R. Glucagon-like
peptide-1 promotes islet cell growth and inhibits apoptosis in Zucker diabetic rats.
Endocrinology 2002;143:4397-4408.
241. Farilla L, Bulotta A, Hirshberg B, Li CS, Khoury N, Noushmehr H, Bertolotto C, Di
Mario U, Harlan DM, Perfetti R. Glucagon-like peptide 1 inhibits cell apoptosis and
improves glucose responsiveness of freshly isolated human islets. Endocrinology
2003;144:5149-5158.
242. Kim JG, Baggio LL, Bridon DP, Castaigne JP, Robitaille MF, Jette L, Benquet C,
Drucker DJ. Development and characterization of a glucagon-like peptide 1-albumin
conjugate: the ability to activate the glucagon-like peptide 1 receptor in vivo. Diabetes
2003;52:751-759.
243. Tourrel C, Bailbe D, Lacorne M, Meile MJ, Kergoat M, Portha B. Persistent
improvement of type 2 diabetes in the Goto-Kakizaki rat model by expansion of the beta-
cell mass during the prediabetic period with glucagon-like peptide-1 or exendin-4.
Diabetes 2002;51:1443-1452.
244. Wang Q, Brubaker PL. Glucagon-like peptide-1 treatment delays the onset of diabetes in
8 week-old db/db mice. Diabetologia 2002;45:1263-1273.

169
245. Hare KJ, Knop FK, Asmar M, Madsbad S, Deacon CF, Holst JJ, Vilsboll T. Preserved
inhibitory potency of GLP-1 on glucagon secretion in type 2 diabetes mellitus. J Clin
Endocrinol Metab 2009;94:4679-4687.
246. Lund A, Vilsboll T, Bagger JI, Holst JJ, Knop FK. The separate and combined impact of
the intestinal hormones, GIP, GLP-1, and GLP-2, on glucagon secretion in type 2
diabetes. Am J Physiol Endocrinol Metab 2011;300:E1038-E1046.
247. Schirra J, Nicolaus M, Roggel R, Katschinski M, Storr M, Woerle HJ, Goke B.
Endogenous glucagon-like peptide 1 controls endocrine pancreatic secretion and antro-
pyloro-duodenal motility in humans. Gut 2006;55:243-251.
248. Gedulin BR, Nikoulina SE, Smith PA, Gedulin G, Nielsen LL, Baron AD, Parkes DG,
Young AA. Exenatide (exendin-4) improves insulin sensitivity and {beta}-cell mass in
insulin-resistant obese fa/fa Zucker rats independent of glycemia and body weight.
Endocrinology 2005;146:2069-2076.
249. Zander M, Madsbad S, Madsen JL, Holst JJ. Effect of 6-week course of glucagon-like
peptide 1 on glycaemic control, insulin sensitivity, and beta-cell function in type 2
diabetes: a parallel-group study. Lancet 2002;359:824-830.
250. Nauck MA, Niedereichholz U, Ettler R, Holst JJ, Orskov C, Ritzel R, Schmiegel WH.
Glucagon-like peptide 1 inhibition of gastric emptying outweighs its insulinotropic
effects in healthy humans. Am J Physiol 1997;273:E981-E988.
251. Little TJ, Pilichiewicz AN, Russo A, Phillips L, Jones KL, Nauck MA, Wishart J,
Horowitz M, Feinle-Bisset C. Effects of intravenous glucagon-like peptide-1 on gastric
emptying and intragastric distribution in healthy subjects: relationships with postprandial
glycemic and insulinemic responses. J Clin Endocrinol Metab 2006;91:1916-1923.
252. Turton MD, O'Shea D, Gunn I, Beak SA, Edwards CM, Meeran K, Choi SJ, Taylor GM,
Heath MM, Lambert PD, Wilding JP, Smith DM, Ghatei MA, Herbert J, Bloom SR. A
role for glucagon-like peptide-1 in the central regulation of feeding. Nature 1996;379:69-
72.
253. Barragan JM, Rodriguez RE, Blazquez E. Changes in arterial blood pressure and heart
rate induced by glucagon-like peptide-1-(7-36) amide in rats. Am J Physiol
1994;266:E459-E466.
254. Cabou C, Campistron G, Marsollier N, Leloup C, Cruciani-Guglielmacci C, Penicaud L,
Drucker DJ, Magnan C, Burcelin R. Brain glucagon-like peptide-1 regulates arterial
blood flow, heart rate, and insulin sensitivity. Diabetes 2008;57:2577-2587.
255. Thorens B. Expression cloning of the pancreatic beta cell receptor for the gluco-incretin
hormone glucagon-like peptide 1. Proc Natl Acad Sci U S A 1992;89:8641-8645.

170
256. Drucker DJ, Philippe J, Mojsov S, Chick WL, Habener JF. Glucagon-like peptide I
stimulates insulin gene expression and increases cyclic AMP levels in a rat islet cell line.
Proc Natl Acad Sci U S A 1987;84:3434-3438.
257. Song WJ, Seshadri M, Ashraf U, Mdluli T, Mondal P, Keil M, Azevedo M, Kirschner
LS, Stratakis CA, Hussain MA. Snapin mediates incretin action and augments glucose-
dependent insulin secretion. Cell Metab 2011;13:308-319.
258. Gomez E, Pritchard C, Herbert TP. cAMP-dependent protein kinase and Ca2+ influx
through L-type voltage-gated calcium channels mediate Raf-independent activation of
extracellular regulated kinase in response to glucagon-like peptide-1 in pancreatic beta-
cells. J Biol Chem 2002;277:48146-48151.
259. Kashima Y, Miki T, Shibasaki T, Ozaki N, Miyazaki M, Yano H, Seino S. Critical role of
cAMP-GEFII--Rim2 complex in incretin-potentiated insulin secretion. J Biol Chem
2001;276:46046-46053.
260. Kang G, Chepurny OG, Holz GG. cAMP-regulated guanine nucleotide exchange factor II
(Epac2) mediates Ca2+-induced Ca2+ release in INS-1 pancreatic beta-cells. J Physiol
2001;536:375-385.
261. Buteau J, Roduit R, Susini S, Prentki M. Glucagon-like peptide-1 promotes DNA
synthesis, activates phosphatidylinositol 3-kinase and increases transcription factor
pancreatic and duodenal homeobox gene 1 (PDX-1) DNA binding activity in beta (INS-
1)-cells. Diabetologia 1999;42:856-864.
262. Hayes MR, Leichner TM, Zhao S, Lee GS, Chowansky A, Zimmer D, De Jonghe BC,
Kanoski SE, Grill HJ, Bence KK. Intracellular signals mediating the food intake-
suppressive effects of hindbrain glucagon-like peptide-1 receptor activation. Cell Metab
2011;13:320-330.
263. Ma X, Bruning J, Ashcroft FM. Glucagon-like peptide 1 stimulates hypothalamic
proopiomelanocortin neurons. J Neurosci 2007;27:7125-7129.
264. Dunphy JL, Taylor RG, Fuller PJ. Tissue distribution of rat glucagon receptor and GLP-1
receptor gene expression. Mol Cell Endocrinol 1998;141:179-186.
265. Bullock BP, Heller RS, Habener JF. Tissue distribution of messenger ribonucleic acid
encoding the rat glucagon-like peptide-1 receptor. Endocrinology 1996;137:2968-2978.
266. Campos RV, Lee YC, Drucker DJ. Divergent tissue-specific and developmental
expression of receptors for glucagon and glucagon-like peptide-1 in the mouse.
Endocrinology 1994;134:2156-2164.
267. Alvarez E, Martinez MD, Roncero I, Chowen JA, Garcia-Cuartero B, Gispert JD, Sanz C,
Vazquez P, Maldonado A, de Caceres J, Desco M, Pozo MA, Blazquez E. The expression
of GLP-1 receptor mRNA and protein allows the effect of GLP-1 on glucose metabolism
in the human hypothalamus and brainstem. J Neurochem 2005;92:798-806.

171
268. Schick RR, Zimmermann JP, vorm WT, Schusdziarra V. Peptides that regulate food
intake: glucagon-like peptide 1-(7-36) amide acts at lateral and medial hypothalamic sites
to suppress feeding in rats. Am J Physiol Regul Integr Comp Physiol 2003;284:R1427-
R1435.
269. Vahl TP, Tauchi M, Durler TS, Elfers EE, Fernandes TM, Bitner RD, Ellis KS, Woods
SC, Seeley RJ, Herman JP, D'Alessio DA. Glucagon-like peptide-1 (GLP-1) receptors
expressed on nerve terminals in the portal vein mediate the effects of endogenous GLP-1
on glucose tolerance in rats. Endocrinology 2007;148:4965-4973.
270. Amato A, Cinci L, Rotondo A, Serio R, Faussone-Pellegrini MS, Vannucchi MG, Mule
F. Peripheral motor action of glucagon-like peptide-1 through enteric neuronal receptors.
Neurogastroenterol Motil 2010;22:664-e203.
271. Klustaitis K. Activation of the central nervous system by circulating glucagon-like
peptide-1. -143. 2009. University of Cincinnati. Ref Type: Thesis/Dissertation
272. Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev 2007;87:1409-1439.
273. Villanueva-Penacarrillo ML, Marquez L, Gonzalez N, Diaz-Miguel M, Valverde I. Effect
of GLP-1 on lipid metabolism in human adipocytes. Horm Metab Res 2001;33:73-77.
274. Oben J, Morgan L, Fletcher J, Marks V. Effect of the entero-pancreatic hormones, gastric
inhibitory polypeptide and glucagon-like polypeptide-1(7-36) amide, on fatty acid
synthesis in explants of rat adipose tissue. J Endocrinol 1991;130:267-272.
275. Hansotia T, Maida A, Flock G, Yamada Y, Tsukiyama K, Seino Y, Drucker DJ.
Extrapancreatic incretin receptors modulate glucose homeostasis, body weight, and
energy expenditure. J Clin Invest 2007;117:143-152.
276. Nogueiras R, Perez-Tilve D, Veyrat-Durebex C, Morgan DA, Varela L, Haynes WG,
Patterson JT, Disse E, Pfluger PT, Lopez M, Woods SC, DiMarchi R, Dieguez C,
Rahmouni K, Rohner-Jeanrenaud F, Tschop MH. Direct control of peripheral lipid
deposition by CNS GLP-1 receptor signaling is mediated by the sympathetic nervous
system and blunted in diet-induced obesity. J Neurosci 2009;29:5916-5925.
277. Gupta NA, Mells J, Dunham RM, Grakoui A, Handy J, Saxena NK, Anania FA.
Glucagon-like peptide-1 receptor is present on human hepatocytes and has a direct role in
decreasing hepatic steatosis in vitro by modulating elements of the insulin signaling
pathway. Hepatology 2010;51:1584-1592.
278. Redondo A, Trigo MV, Acitores A, Valverde I, Villanueva-Penacarrillo ML. Cell
signalling of the GLP-1 action in rat liver. Mol Cell Endocrinol 2003;204:43-50.
279. Ding X, Saxena NK, Lin S, Gupta NA, Anania FA. Exendin-4, a glucagon-like protein-1
(GLP-1) receptor agonist, reverses hepatic steatosis in ob/ob mice. Hepatology
2006;43:173-181.

172
280. Parlevliet ET, Schroder-van der Elst JP, Corssmit EP, Picha K, O'Neil K, Stojanovic-
Susulic V, Ort T, Havekes LM, Romijn JA, Pijl H. CNTO736, a novel glucagon-like
peptide-1 receptor agonist, ameliorates insulin resistance and inhibits very low-density
lipoprotein production in high-fat-fed mice. J Pharmacol Exp Ther 2009;328:240-248.
281. Parlevliet ET, de Leeuw van Weenen JE, Romijn JA, Pijl H. GLP-1 treatment reduces
endogenous insulin resistance via activation of central GLP-1 receptors in mice fed a
high-fat diet. Am J Physiol Endocrinol Metab 2010;299:E318-E324.
282. Tang-Christensen M, Larsen PJ, Thulesen J, Romer J, Vrang N. The proglucagon-derived
peptide, glucagon-like peptide-2, is a neurotransmitter involved in the regulation of food
intake. Nat Med 2000;6:802-807.
283. de Heer J, Pedersen J, Orskov C, Holst JJ. The alpha cell expresses glucagon-like
peptide-2 receptors and glucagon-like peptide-2 stimulates glucagon secretion from the
rat pancreas. Diabetologia 2007;50:2135-2142.
284. Meier JJ, Nauck MA, Pott A, Heinze K, Goetze O, Bulut K, Schmidt WE, Gallwitz B,
Holst JJ. Glucagon-like peptide 2 stimulates glucagon secretion, enhances lipid
absorption, and inhibits gastric acid secretion in humans. Gastroenterology 2006;130:44-
54.
285. Drucker DJ, Erlich P, Asa SL, Brubaker PL. Induction of intestinal epithelial
proliferation by glucagon-like peptide 2. Proc Natl Acad Sci U S A 1996;93:7911-7916.
286. Rowland KJ, Brubaker PL. Life in the crypt: a role for glucagon-like peptide-2? Mol Cell
Endocrinol 2008;288:63-70.
287. Burrin DG, Stoll B, Jiang R, Petersen Y, Elnif J, Buddington RK, Schmidt M, Holst JJ,
Hartmann B, Sangild PT. GLP-2 stimulates intestinal growth in premature TPN-fed pigs
by suppressing proteolysis and apoptosis. Am J Physiol Gastrointest Liver Physiol
2000;279:G1249-G1256.
288. Tsai CH, Hill M, Asa SL, Brubaker PL, Drucker DJ. Intestinal growth-promoting
properties of glucagon-like peptide-2 in mice. Am J Physiol 1997;273:E77-E84.
289. Drucker DJ, Yusta B, Boushey RP, DeForest L, Brubaker PL. Human [Gly2]GLP-2
reduces the severity of colonic injury in a murine model of experimental colitis. Am J
Physiol 1999;276:G79-G91.
290. L'Heureux MC, Brubaker PL. Glucagon-like peptide-2 and common therapeutics in a
murine model of ulcerative colitis. J Pharmacol Exp Ther 2003;306:347-354.
291. Ramsanahie AP, Perez A, Duensing AU, Zinner MJ, Ashley SW, Whang EE. Glucagon-
like peptide 2 enhances intestinal epithelial restitution. J Surg Res 2002;107:44-49.

173
292. Benjamin MA, McKay DM, Yang PC, Cameron H, Perdue MH. Glucagon-like peptide-2
enhances intestinal epithelial barrier function of both transcellular and paracellular
pathways in the mouse. Gut 2000;47:112-119.
293. Burrin DG, Stoll B, Guan X, Cui L, Chang X, Holst JJ. Glucagon-like peptide 2 dose-
dependently activates intestinal cell survival and proliferation in neonatal piglets.
Endocrinology 2005;146:22-32.
294. Hadjiyanni I, Li KK, Drucker DJ. Glucagon-like peptide-2 reduces intestinal
permeability but does not modify the onset of type 1 diabetes in the nonobese diabetic
mouse. Endocrinology 2009;150:592-599.
295. Perez A, Duxbury M, Rocha FG, Ramsanahie AP, Farivar RS, Varnholt H, Ito H, Wong
H, Rounds J, Zinner MJ, Whang EE, Ashley SW. Glucagon-like peptide 2 is an
endogenous mediator of postresection intestinal adaptation. JPEN J Parenter Enteral Nutr
2005;29:97-101.
296. Wojdemann M, Wettergren A, Hartmann B, Hilsted L, Holst JJ. Inhibition of sham
feeding-stimulated human gastric acid secretion by glucagon-like peptide-2. J Clin
Endocrinol Metab 1999;84:2513-2517.
297. Banasch M, Bulut K, Hagemann D, Schrader H, Holst JJ, Schmidt WE, Meier JJ.
Glucagon-like peptide 2 inhibits ghrelin secretion in humans. Regul Pept 2006;137:173-
178.
298. Nagell CF, Wettergren A, Pedersen JF, Mortensen D, Holst JJ. Glucagon-like peptide-2
inhibits antral emptying in man, but is not as potent as glucagon-like peptide-1. Scand J
Gastroenterol 2004;39:353-358.
299. McDonagh SC, Lee J, Izzo A, Brubaker PL. Role of glial cell-line derived neurotropic
factor family receptor alpha2 in the actions of the glucagon-like peptides on the murine
intestine. Am J Physiol Gastrointest Liver Physiol 2007;293:G461-G468.
300. Sigalet DL, Wallace LE, Holst JJ, Martin GR, Kaji T, Tanaka H, Sharkey KA. Enteric
neural pathways mediate the anti-inflammatory actions of glucagon-like peptide 2. Am J
Physiol Gastrointest Liver Physiol 2007;293:G211-G221.
301. Bahrami J, Longuet C, Baggio LL, Li K, Drucker DJ. Glucagon-like peptide-2 receptor
modulates islet adaptation to metabolic stress in the ob/ob mouse. Gastroenterology
2010;139:857-868.
302. Cani PD, Possemiers S, Van de WT, Guiot Y, Everard A, Rottier O, Geurts L, Naslain D,
Neyrinck A, Lambert DM, Muccioli GG, Delzenne NM. Changes in gut microbiota
control inflammation in obese mice through a mechanism involving GLP-2-driven
improvement of gut permeability. Gut 2009;58:1091-1103.

174
303. Guan X, Stoll B, Lu X, Tappenden KA, Holst JJ, Hartmann B, Burrin DG. GLP-2-
mediated up-regulation of intestinal blood flow and glucose uptake is nitric oxide-
dependent in TPN-fed piglets 1. Gastroenterology 2003;125:136-147.
304. Guan X, Karpen HE, Stephens J, Bukowski JT, Niu S, Zhang G, Stoll B, Finegold MJ,
Holst JJ, Hadsell D, Nichols BL, Burrin DG. GLP-2 receptor localizes to enteric neurons
and endocrine cells expressing vasoactive peptides and mediates increased blood flow.
Gastroenterology 2006;130:150-164.
305. Bremholm L, Hornum M, Andersen UB, Holst JJ. The effect of glucagon-like peptide-2
on arterial blood flow and cardiac parameters. Regul Pept 2010;159:67-71.
306. Deniz M, Bozkurt A, Kurtel H. Mediators of glucagon-like peptide 2-induced blood flow:
responses in different vascular sites. Regul Pept 2007;142:7-15.
307. Stephens J, Stoll B, Cottrell J, Chang X, Helmrath M, Burrin DG. Glucagon-like peptide-
2 acutely increases proximal small intestinal blood flow in TPN-fed neonatal piglets. Am
J Physiol Regul Integr Comp Physiol 2006;290:R283-R289.
308. Kitchen PA, FitzGerald AJ, Goodlad RA, Barley NF, Ghatei MA, Legon S, Bloom SR,
Price A, Walters JR, Forbes A. Glucagon-like peptide-2 increases sucrase-isomaltase but
not caudal-related homeobox protein-2 gene expression. Am J Physiol Gastrointest Liver
Physiol 2000;278:G425-G428.
309. Scott RB, Kirk D, MacNaughton WK, Meddings JB. GLP-2 augments the adaptive
response to massive intestinal resection in rat. Am J Physiol 1998;275:G911-G921.
310. Brubaker PL, Izzo A, Hill M, Drucker DJ. Intestinal function in mice with small bowel
growth induced by glucagon-like peptide-2. Am J Physiol 1997;272:E1050-E1058.
311. Au A, Gupta A, Schembri P, Cheeseman CI. Rapid insertion of GLUT2 into the rat
jejunal brush-border membrane promoted by glucagon-like peptide 2. Biochem J
2002;367:247-254.
312. Cheeseman CI. Upregulation of SGLT-1 transport activity in rat jejunum induced by
GLP-2 infusion in vivo. Am J Physiol 1997;273:R1965-R1971.
313. Cheeseman CI, Tsang R. The effect of GIP and glucagon-like peptides on intestinal
basolateral membrane hexose transport. Am J Physiol 1996;271:G477-G482.
314. Cheeseman CI, O'Neill D. Basolateral D-glucose transport activity along the crypt-villus
axis in rat jejunum and upregulation induced by gastric inhibitory peptide and glucagon-
like peptide-2. Exp Physiol 1998;83:605-616.
315. Ramsanahie AP, Berger UV, Zinner MJ, Whang EE, Rhoads DB, Ashley SW. Effect of
glucagon-like peptide-2 (GLP-2) on diurnal SGLT1 expression. Dig Dis Sci
2004;49:1731-1737.

175
316. Sangild PT, Tappenden KA, Malo C, Petersen YM, Elnif J, Bartholome AL, Buddington
RK. Glucagon-like peptide 2 stimulates intestinal nutrient absorption in parenterally fed
newborn pigs. J Pediatr Gastroenterol Nutr 2006;43:160-167.
317. Drozdowski LA, Iordache C, Clandinin MT, Wild G, Todd Z, Thomson AB. A
combination of dexamethasone and glucagon-like peptide-2 increase intestinal
morphology and glucose uptake in suckling rats. J Pediatr Gastroenterol Nutr
2006;42:32-39.
318. Cottrell JJ, Stoll B, Buddington RK, Stephens JE, Cui L, Chang X, Burrin DG.
Glucagon-like peptide-2 protects against TPN-induced intestinal hexose malabsorption in
enterally refed piglets. Am J Physiol Gastrointest Liver Physiol 2006;290:G293-G300.
319. Sangild PT, Malo C, Schmidt M, Petersen YM, Elnif J, Holst JJ, Buddington RK.
Glucagon-like peptide 2 has limited efficacy to increase nutrient absorption in fetal and
preterm pigs. Am J Physiol Regul Integr Comp Physiol 2007;293:R2179-R2184.
320. Kato Y, Yu D, Schwartz MZ. Glucagonlike peptide-2 enhances small intestinal
absorptive function and mucosal mass in vivo. J Pediatr Surg 1999;34:18-20.
321. Iordache C, Drozdowski L, Clandinin MT, Wild G, Todd Z, Thomson AB. Treatment of
suckling rats with GLP-2 plus dexamethasone increases the ileal uptake of fatty acids in
later life. Am J Physiol Gastrointest Liver Physiol 2005;288:G54-G59.
322. Kjems LL, Holst JJ, Volund A, Madsbad S. The influence of GLP-1 on glucose-
stimulated insulin secretion: effects on beta-cell sensitivity in type 2 and nondiabetic
subjects. Diabetes 2003;52:380-386.
323. Schmidt WE, Siegel EG, Creutzfeldt W. Glucagon-like peptide-1 but not glucagon-like
peptide-2 stimulates insulin release from isolated rat pancreatic islets. Diabetologia
1985;28:704-707.
324. Orskov C, Holst JJ, Nielsen OV. Effect of truncated glucagon-like peptide-1
[proglucagon-(78-107) amide] on endocrine secretion from pig pancreas, antrum, and
nonantral stomach. Endocrinology 1988;123:2009-2013.
325. Simonsen L, Pilgaard S, Orskov C, Rosenkilde MM, Hartmann B, Holst JJ, Deacon CF.
Exendin-4, but not dipeptidyl peptidase IV inhibition, increases small intestinal mass in
GK rats. Am J Physiol Gastrointest Liver Physiol 2007;293:G288-G295.
326. Drucker DJ, DeForest L, Brubaker PL. Intestinal response to growth factors administered
alone or in combination with human [Gly2]glucagon-like peptide 2. Am J Physiol
1997;273:G1252-G1262.
327. Dube PE, Forse CL, Bahrami J, Brubaker PL. The essential role of insulin-like growth
factor-1 in the intestinal tropic effects of glucagon-like peptide-2 in mice.
Gastroenterology 2006;131:589-605.

176
328. Yusta B, Holland D, Koehler JA, Maziarz M, Estall JL, Higgins R, Drucker DJ. ErbB
signaling is required for the proliferative actions of GLP-2 in the murine gut.
Gastroenterology 2009;137:986-996.
329. Garrison AP, Dekaney CM, von Allmen DC, Lund PK, Henning SJ, Helmrath MA. Early
but not late administration of glucagon-like peptide-2 following ileo-cecal resection
augments putative intestinal stem cell expansion. Am J Physiol Gastrointest Liver Physiol
2009;296:G643-G650.
330. Cameron HL, Yang PC, Perdue MH. Glucagon-like peptide-2-enhanced barrier function
reduces pathophysiology in a model of food allergy. Am J Physiol Gastrointest Liver
Physiol 2003;284:G905-G912.
331. Abraham EJ, Leech CA, Lin JC, Zulewski H, Habener JF. Insulinotropic hormone
glucagon-like peptide-1 differentiation of human pancreatic islet-derived progenitor cells
into insulin-producing cells. Endocrinology 2002;143:3152-3161.
332. Hui H, Wright C, Perfetti R. Glucagon-like peptide 1 induces differentiation of islet
duodenal homeobox-1-positive pancreatic ductal cells into insulin-secreting cells.
Diabetes 2001;50:785-796.
333. Xu G, Stoffers DA, Habener JF, Bonner-Weir S. Exendin-4 stimulates both beta-cell
replication and neogenesis, resulting in increased beta-cell mass and improved glucose
tolerance in diabetic rats. Diabetes 1999;48:2270-2276.
334. Baumler MD, Nelson DW, Ney DM, Groblewski GE. Loss of exocrine pancreatic
stimulation during parenteral feeding suppresses digestive enzyme expression and
induces Hsp70 expression. Am J Physiol Gastrointest Liver Physiol 2007;292:G857-
G866.
335. Wojdemann M, Wettergren A, Hartmann B, Holst JJ. Glucagon-like peptide-2 inhibits
centrally induced antral motility in pigs. Scand J Gastroenterol 1998;33:828-832.
336. Giralt M, Vergara P. Sympathetic pathways mediate GLP-1 actions in the gastrointestinal
tract of the rat. Regul Pept 1998;74:19-25.
337. Tolessa T, Gutniak M, Holst JJ, Efendic S, Hellstrom PM. Inhibitory effect of glucagon-
like peptide-1 on small bowel motility. Fasting but not fed motility inhibited via nitric
oxide independently of insulin and somatostatin. J Clin Invest 1998;102:764-774.
338. Bozkurt A, Naslund E, Holst JJ, Hellstrom PM. GLP-1 and GLP-2 act in concert to
inhibit fasted, but not fed, small bowel motility in the rat. Regul Pept 2002;107:129-135.
339. Sandhu H, Wiesenthal SR, MacDonald PE, McCall RH, Tchipashvili V, Rashid S,
Satkunarajah M, Irwin DM, Shi ZQ, Brubaker PL, Wheeler MB, Vranic M, Efendic S,
Giacca A. Glucagon-like peptide 1 increases insulin sensitivity in depancreatized dogs.
Diabetes 1999;48:1045-1053.

177
340. Mizuno A, Kuwajima M, Ishida K, Noma Y, Murakami T, Tateishi K, Sato I, Shima K.
Extrapancreatic action of truncated glucagon-like peptide-I in Otsuka Long-Evans
Tokushima Fatty rats, an animal model for non-insulin-dependent diabetes mellitus.
Metabolism 1997;46:745-749.
341. Li Y, Perry T, Kindy MS, Harvey BK, Tweedie D, Holloway HW, Powers K, Shen H,
Egan JM, Sambamurti K, Brossi A, Lahiri DK, Mattson MP, Hoffer BJ, Wang Y, Greig
NH. GLP-1 receptor stimulation preserves primary cortical and dopaminergic neurons in
cellular and rodent models of stroke and Parkinsonism. Proc Natl Acad Sci U S A
2009;106:1285-1290.
342. During MJ, Cao L, Zuzga DS, Francis JS, Fitzsimons HL, Jiao X, Bland RJ, Klugmann
M, Banks WA, Drucker DJ, Haile CN. Glucagon-like peptide-1 receptor is involved in
learning and neuroprotection. Nat Med 2003;9:1173-1179.
343. Bertilsson G, Patrone C, Zachrisson O, Andersson A, Dannaeus K, Heidrich J, Kortesmaa
J, Mercer A, Nielsen E, Ronnholm H, Wikstrom L. Peptide hormone exendin-4
stimulates subventricular zone neurogenesis in the adult rodent brain and induces
recovery in an animal model of Parkinson's disease. J Neurosci Res 2008;86:326-338.
344. Belsham DD, Fick LJ, Dalvi PS, Centeno ML, Chalmers JA, Lee PK, Wang Y, Drucker
DJ, Koletar MM. Ciliary neurotrophic factor recruitment of glucagon-like peptide-1
mediates neurogenesis, allowing immortalization of adult murine hypothalamic neurons.
FASEB J 2009;23:4256-4265.
345. Lovshin JA, Huang Q, Seaberg R, Brubaker PL, Drucker DJ. Extrahypothalamic
expression of the glucagon-like peptide-2 receptor is coupled to reduction of glutamate-
induced cell death in cultured hippocampal cells. Endocrinology 2004;145:3495-3506.
346. Velazquez E, Ruiz-Albusac JM, Blazquez E. Glucagon-like peptide-2 stimulates the
proliferation of cultured rat astrocytes. Eur J Biochem 2003;270:3001-3009.
347. Larsen PJ, Fledelius C, Knudsen LB, Tang-Christensen M. Systemic administration of
the long-acting GLP-1 derivative NN2211 induces lasting and reversible weight loss in
both normal and obese rats. Diabetes 2001;50:2530-2539.
348. Yamamoto H, Lee CE, Marcus JN, Williams TD, Overton JM, Lopez ME, Hollenberg
AN, Baggio L, Saper CB, Drucker DJ, Elmquist JK. Glucagon-like peptide-1 receptor
stimulation increases blood pressure and heart rate and activates autonomic regulatory
neurons. J Clin Invest 2002;110:43-52.
349. Bremholm L, Hornum M, Henriksen BM, Larsen S, Holst JJ. Glucagon-like peptide-2
increases mesenteric blood flow in humans. Scand J Gastroenterol 2009;44:314-319.
350. Yamada C, Yamada Y, Tsukiyama K, Yamada K, Udagawa N, Takahashi N, Tanaka K,
Drucker DJ, Seino Y, Inagaki N. The murine glucagon-like peptide-1 receptor is essential
for control of bone resorption. Endocrinology 2008;149:574-579.

178
351. Henriksen DB, Alexandersen P, Hartmann B, Adrian CL, Byrjalsen I, Bone HG, Holst JJ,
Christiansen C. Disassociation of bone resorption and formation by GLP-2: a 14-day
study in healthy postmenopausal women. Bone 2007;40:723-729.
352. Henriksen DB, Alexandersen P, Bjarnason NH, Vilsboll T, Hartmann B, Henriksen EE,
Byrjalsen I, Krarup T, Holst JJ, Christiansen C. Role of gastrointestinal hormones in
postprandial reduction of bone resorption. J Bone Miner Res 2003;18:2180-2189.
353. Thulesen J, Hartmann B, Orskov C, Jeppesen PB, Holst JJ, Poulsen SS. Potential targets
for glucagon-like peptide 2 (GLP-2) in the rat: distribution and binding of i.v. injected
(125)I-GLP-2. Peptides 2000;21:1511-1517.
354. Orskov C, Hartmann B, Poulsen SS, Thulesen J, Hare KJ, Holst JJ. GLP-2 stimulates
colonic growth via KGF, released by subepithelial myofibroblasts with GLP-2 receptors.
Regul Pept 2005;124:105-112.
355. Yusta B, Huang L, Munroe D, Wolff G, Fantaske R, Sharma S, Demchyshyn L, Asa SL,
Drucker DJ. Enteroendocrine localization of GLP-2 receptor expression in humans and
rodents. Gastroenterology 2000;119:744-755.
356. Guan X, Karpen HE, Stephens J, Bukowski JT, Niu S, Zhang G, Stoll B, Finegold MJ,
Holst JJ, Hadsell D, Nichols BL, Burrin DG. GLP-2 receptor localizes to enteric neurons
and endocrine cells expressing vasoactive peptides and mediates increased blood flow.
Gastroenterology 2006;130:150-164.
357. Bjerknes M, Cheng H. Modulation of specific intestinal epithelial progenitors by enteric
neurons. Proc Natl Acad Sci U S A 2001;98:12497-12502.
358. Nelson DW, Sharp JW, Brownfield MS, Raybould HE, Ney DM. Localization and
activation of glucagon-like peptide-2 receptors on vagal afferents in the rat.
Endocrinology 2007;148:1954-1962.
359. Leen JL, Izzo A, Upadhyay C, Rowland KJ, Dube PE, Gu S, Heximer SP, Rhodes CJ,
Storm DR, Lund PK, Brubaker PL. Mechanism of action of glucagon-like peptide-2 to
increase IGF-I mRNA in intestinal subepithelial fibroblasts. Endocrinology
2011;152:436-446.
360. Walsh NA, Yusta B, DaCambra MP, Anini Y, Drucker DJ, Brubaker PL. Glucagon-like
peptide-2 receptor activation in the rat intestinal mucosa. Endocrinology 2003;144:4385-
4392.
361. Yusta B, Somwar R, Wang F, Munroe D, Grinstein S, Klip A, Drucker DJ. Identification
of glucagon-like peptide-2 (GLP-2)-activated signaling pathways in baby hamster kidney
fibroblasts expressing the rat GLP-2 receptor. J Biol Chem 1999;274:30459-30467.
362. Yusta B, Boushey RP, Drucker DJ. The glucagon-like peptide-2 receptor mediates direct
inhibition of cellular apoptosis via a cAMP-dependent protein kinase-independent
pathway. J Biol Chem 2000;275:35345-35352.

179
363. Yusta B, Estall J, Drucker DJ. Glucagon-like peptide-2 receptor activation engages bad
and glycogen synthase kinase-3 in a protein kinase A-dependent manner and prevents
apoptosis following inhibition of phosphatidylinositol 3-kinase. J Biol Chem
2002;277:24896-24906.
364. Dube PE, Rowland KJ, Brubaker PL. Glucagon-like peptide-2 activates beta-catenin
signaling in the mouse intestinal crypt: role of insulin-like growth factor-I. Endocrinology
2008;149:291-301.
365. Shi X, Li X, Wang Y, Zhang K, Zhou F, Chan L, Li D, Guan X. Glucagon-like peptide-
2-stimulated protein synthesis through the PI 3-kinase-dependent Akt-mTOR signaling
pathway. Am J Physiol Endocrinol Metab 2011;300:E554-E563.
366. Koehler JA, Yusta B, Drucker DJ. The HeLa cell glucagon-like peptide-2 receptor is
coupled to regulation of apoptosis and ERK1/2 activation through divergent signaling
pathways. Mol Endocrinol 2005;19:459-473.
367. Baggio LL, Kim JG, Drucker DJ. Chronic exposure to GLP-1R agonists promotes
homologous GLP-1 receptor desensitization in vitro but does not attenuate GLP-1R-
dependent glucose homeostasis in vivo. Diabetes 2004;53 Suppl 3:S205-S214.
368. Estall JL, Yusta B, Drucker DJ. Lipid raft-dependent glucagon-like peptide-2 receptor
trafficking occurs independently of agonist-induced desensitization. Mol Biol Cell
2004;15:3673-3687.
369. Dube PE, Forse CL, Bahrami J, Brubaker PL. The essential role of insulin-like growth
factor-1 in the intestinal tropic effects of glucagon-like peptide-2 in mice.
Gastroenterology 2006;131:589-605.
370. Orskov C, Hartmann B, Poulsen SS, Thulesen J, Hare KJ, Holst JJ. GLP-2 stimulates
colonic growth via KGF, released by subepithelial myofibroblasts with GLP-2 receptors.
Regul Pept 2005;124:105-112.
371. Bulut K, Pennartz C, Felderbauer P, Meier JJ, Banasch M, Bulut D, Schmitz F, Schmidt
WE, Hoffmann P. Glucagon like peptide-2 induces intestinal restitution through VEGF
release from subepithelial myofibroblasts. Eur J Pharmacol 2008;578:279-285.
372. Amato A, Baldassano S, Serio R, Mule F. Glucagon-like peptide-2 relaxes mouse
stomach through vasoactive intestinal peptide release. Am J Physiol Gastrointest Liver
Physiol 2009;296:G678-G684.
373. Liu GL, Fan LM, Redinger RN. The association of hepatic apoprotein and lipid
metabolism in hamsters and rats. Comp Biochem Physiol A Comp Physiol 1991;99:223-
228.
374. Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes
epidemic. Nature 2001;414:782-787.

180
375. Ginsberg HN. Insulin resistance and cardiovascular disease. J Clin Invest 2000;106:453-
458.
376. Mero N, Syvanne M, Taskinen MR. Postprandial lipid metabolism in diabetes.
Atherosclerosis 1998;141 Suppl 1:S53-S55.
377. Ohnishi H, Saitoh S, Takagi S, Ohata J, Isobe T, Kikuchi Y, Takeuchi H, Shimamoto K.
Relationship between insulin-resistance and remnant-like particle cholesterol.
Atherosclerosis 2002;164:167-170.
378. Schaefer EJ, McNamara JR, Shah PK, Nakajima K, Cupples LA, Ordovas JM, Wilson
PW. Elevated remnant-like particle cholesterol and triglyceride levels in diabetic men and
women in the Framingham Offspring Study. Diabetes Care 2002;25:989-994.
379. Lu WJ, Yang Q, Sun W, Woods SC, D'Alessio D, Tso P. Using the lymph fistula rat
model to study the potentiation of GIP secretion by the ingestion of fat and glucose. Am J
Physiol Gastrointest Liver Physiol 2008;294:G1130-G1138.
380. Drucker DJ. The biology of incretin hormones. Cell Metab 2006;3:153-165.
381. Baggio LL, Huang Q, Brown TJ, Drucker DJ. Oxyntomodulin and glucagon-like peptide-
1 differentially regulate murine food intake and energy expenditure. Gastroenterology
2004;127:546-558.
382. Miyawaki K, Yamada Y, Ban N, Ihara Y, Tsukiyama K, Zhou H, Fujimoto S, Oku A,
Tsuda K, Toyokuni S, Hiai H, Mizunoya W, Fushiki T, Holst JJ, Makino M, Tashita A,
Kobara Y, Tsubamoto Y, Jinnouchi T, Jomori T, Seino Y. Inhibition of gastric inhibitory
polypeptide signaling prevents obesity. Nat Med 2002;8:738-742.
383. Hansotia T, Maida A, Flock G, Yamada Y, Tsukiyama K, Seino Y, Drucker DJ.
Extrapancreatic incretin receptors modulate glucose homeostasis, body weight, and
energy expenditure. J Clin Invest 2007;117:143-152.
384. Ellingsgaard H, Hauselmann I, Schuler B, Habib AM, Baggio LL, Meier DT, Eppler E,
Bouzakri K, Wueest S, Muller YD, Hansen AM, Reinecke M, Konrad D, Gassmann M,
Reimann F, Halban PA, Gromada J, Drucker DJ, Gribble FM, Ehses JA, Donath MY.
Interleukin-6 enhances insulin secretion by increasing glucagon-like peptide-1 secretion
from L cells and alpha cells. Nat Med 2011;17:1481-1489.
385. Muscelli E, Mari A, Casolaro A, Camastra S, Seghieri G, Gastaldelli A, Holst JJ,
Ferrannini E. Separate impact of obesity and glucose tolerance on the incretin effect in
normal subjects and type 2 diabetic patients. Diabetes 2008;57:1340-1348.
386. Toft-Nielsen MB, Damholt MB, Madsbad S, Hilsted LM, Hughes TE, Michelsen BK,
Holst JJ. Determinants of the impaired secretion of glucagon-like peptide-1 in type 2
diabetic patients. J Clin Endocrinol Metab 2001;86:3717-3723.

181
387. Vollmer K, Holst JJ, Baller B, Ellrichmann M, Nauck MA, Schmidt WE, Meier JJ.
Predictors of incretin concentrations in subjects with normal, impaired, and diabetic
glucose tolerance. Diabetes 2008;57:678-687.
388. Aschner P, Kipnes MS, Lunceford JK, Sanchez M, Mickel C, Williams-Herman DE.
Effect of the dipeptidyl peptidase-4 inhibitor sitagliptin as monotherapy on glycemic
control in patients with type 2 diabetes. Diabetes Care 2006;29:2632-2637.
389. Raz I, Hanefeld M, Xu L, Caria C, Williams-Herman D, Khatami H. Efficacy and safety
of the dipeptidyl peptidase-4 inhibitor sitagliptin as monotherapy in patients with type 2
diabetes mellitus. Diabetologia 2006;49:2564-2571.
390. Amori RE, Lau J, Pittas AG. Efficacy and safety of incretin therapy in type 2 diabetes:
systematic review and meta-analysis. JAMA 2007;298:194-206.
391. Wasada T, McCorkle K, Harris V, Kawai K, Howard B, Unger RH. Effect of gastric
inhibitory polypeptide on plasma levels of chylomicron triglycerides in dogs. J Clin
Invest 1981;68:1106-1107.
392. Kim SJ, Nian C, McIntosh CH. Activation of lipoprotein lipase by glucose-dependent
insulinotropic polypeptide in adipocytes. A role for a protein kinase B, LKB1, and AMP-
activated protein kinase cascade. J Biol Chem 2007;282:8557-8567.
393. Meier JJ, Gethmann A, Gotze O, Gallwitz B, Holst JJ, Schmidt WE, Nauck MA.
Glucagon-like peptide 1 abolishes the postprandial rise in triglyceride concentrations and
lowers levels of non-esterified fatty acids in humans. Diabetologia 2006;49:452-458.
394. Qin X, Shen H, Liu M, Yang Q, Zheng S, Sabo M, D'Alessio DA, Tso P. GLP-1 reduces
intestinal lymph flow, triglyceride absorption, and apolipoprotein production in rats. Am
J Physiol Gastrointest Liver Physiol 2005;288:G943-G949.
395. Taggart C, Gibney J, Owens D, Collins P, Johnson A, Tomkin GH. The role of dietary
cholesterol in the regulation of postprandial apolipoprotein B48 levels in diabetes. Diabet
Med 1997;14:1051-1058.
396. Zoltowska M, Ziv E, Delvin E, Sinnett D, Kalman R, Garofalo C, Seidman E, Levy E.
Cellular aspects of intestinal lipoprotein assembly in Psammomys obesus: a model of
insulin resistance and type 2 diabetes. Diabetes 2003;52:2539-2545.
397. Perreault N, Beaulieu JF. Primary cultures of fully differentiated and pure human
intestinal epithelial cells. Exp Cell Res 1998;245:34-42.
398. Ebert R, Nauck M, Creutzfeldt W. Effect of exogenous or endogenous gastric inhibitory
polypeptide (GIP) on plasma triglyceride responses in rats. Horm Metab Res
1991;23:517-521.
399. Taghibiglou C, Carpentier A, Van Iderstine SC, Chen B, Rudy D, Aiton A, Lewis GF,
Adeli K. Mechanisms of hepatic very low density lipoprotein overproduction in insulin

182
resistance. Evidence for enhanced lipoprotein assembly, reduced intracellular ApoB
degradation, and increased microsomal triglyceride transfer protein in a fructose-fed
hamster model. J Biol Chem 2000;275:8416-8425.
400. Baggio LL, Huang Q, Brown TJ, Drucker DJ. A recombinant human glucagon-like
peptide (GLP)-1-albumin protein (albugon) mimics peptidergic activation of GLP-1
receptor-dependent pathways coupled with satiety, gastrointestinal motility, and glucose
homeostasis. Diabetes 2004;53:2492-2500.
401. Bergman AJ, Stevens C, Zhou Y, Yi B, Laethem M, De Smet M, Snyder K, Hilliard D,
Tanaka W, Zeng W, Tanen M, Wang AQ, Chen L, Winchell G, Davies MJ, Ramael S,
Wagner JA, Herman GA. Pharmacokinetic and pharmacodynamic properties of multiple
oral doses of sitagliptin, a dipeptidyl peptidase-IV inhibitor: a double-blind, randomized,
placebo-controlled study in healthy male volunteers. Clin Ther 2006;28:55-72.
402. Nauck MA, Meininger G, Sheng D, Terranella L, Stein PP. Efficacy and safety of the
dipeptidyl peptidase-4 inhibitor, sitagliptin, compared with the sulfonylurea, glipizide, in
patients with type 2 diabetes inadequately controlled on metformin alone: a randomized,
double-blind, non-inferiority trial. Diabetes Obes Metab 2007;9:194-205.
403. Matikainen N, Manttari S, Schweizer A, Ulvestad A, Mills D, Dunning BE, Foley JE,
Taskinen MR. Vildagliptin therapy reduces postprandial intestinal triglyceride-rich
lipoprotein particles in patients with type 2 diabetes. Diabetologia 2006;49:2049-2057.
404. Boschmann M, Engeli S, Dobberstein K, Budziarek P, Strauss A, Boehnke J, Sweep FC,
Luft FC, He Y, Foley JE, Jordan J. Dipeptidyl-Peptidase-IV Inhibition Augments
Postprandial Lipid Mobilization and Oxidation in Type 2 Diabetic Patients. J Clin
Endocrinol Metab 2008.
405. Levy E, Sinnett D, Thibault L, Nguyen TD, Delvin E, Menard D. Insulin modulation of
newly synthesized apolipoproteins B-100 and B-48 in human fetal intestine: gene
expression and mRNA editing are not involved. FEBS Lett 1996;393:253-258.
406. Young AA, Gedulin BR, Bhavsar S, Bodkin N, Jodka C, Hansen B, Denaro M. Glucose-
lowering and insulin-sensitizing actions of exendin-4: studies in obese diabetic (ob/ob,
db/db) mice, diabetic fatty Zucker rats, and diabetic rhesus monkeys (Macaca mulatta).
Diabetes 1999;48:1026-1034.
407. Scrocchi LA, Marshall BA, Cook SM, Brubaker PL, Drucker DJ. Identification of
glucagon-like peptide 1 (GLP-1) actions essential for glucose homeostasis in mice with
disruption of GLP-1 receptor signaling. Diabetes 1998;47:632-639.
408. Longuet C, Sinclair EM, Maida A, Baggio LL, Maziarz M, Charron MJ, Drucker DJ. The
glucagon receptor is required for the adaptive metabolic response to fasting. Cell Metab
2008;8:359-371.
409. Nakano T, Nakajima K, Niimi M, Fujita MQ, Nakajima Y, Takeichi S, Kinosita M,
Matsushima T, Teramoto T, Tanaka A. Detection of apolipoproteins B-48 and B-100

183
carrying particles in lipoprotein fractions extracted from human aortic atherosclerotic
plaques in sudden cardiac death cases. Clin Chim Acta 2007.
410. Taskinen MR. Insulin resistance and lipoprotein metabolism. Curr Opin Lipidol
1995;6:153-160.
411. Gylling H, Tuominen JA, Koivisto VA, Miettinen TA. Cholesterol metabolism in type 1
diabetes. Diabetes 2004;53:2217-2222.
412. Estall JL, Drucker DJ. Glucagon-like Peptide-2. Annu Rev Nutr 2006;26:391-411.
413. Tsai CH, Hill M, Drucker DJ. Biological determinants of intestinotrophic properties of
GLP-2 in vivo. Am J Physiol 1997;272:G662-G668.
414. Guo Q, Avramoglu RK, Adeli K. Intestinal assembly and secretion of highly dense/lipid-
poor apolipoprotein B48-containing lipoprotein particles in the fasting state: evidence for
induction by insulin resistance and exogenous fatty acids. Metabolism 2005;54:689-697.
415. CHEARS WC, Jr., McCRAW BH, TYOR MP, RUFFIN JM. The I131 labeled triolein
absorption test: reproducibility and factors affecting blood levels. South Med J
1958;51:433-437.
416. Pedersen NB, Hjollund KR, Johnsen AH, Orskov C, Rosenkilde MM, Hartmann B, Holst
JJ. Porcine glucagon-like peptide-2: structure, signaling, metabolism and effects. Regul
Pept 2008;146:310-320.
417. Eyre NS, Cleland LG, Tandon NN, Mayrhofer G. Importance of the carboxyl terminus of
FAT/CD36 for plasma membrane localization and function in long-chain fatty acid
uptake. J Lipid Res 2007;48:528-542.
418. Luiken JJ, Koonen DP, Willems J, Zorzano A, Becker C, Fischer Y, Tandon NN, Van
Der Vusse GJ, Bonen A, Glatz JF. Insulin stimulates long-chain fatty acid utilization by
rat cardiac myocytes through cellular redistribution of FAT/CD36. Diabetes
2002;51:3113-3119.
419. Densupsoontorn N, Carpentier YA, Racine R, Murray FM, Seo T, Ramakrishnan R,
Deckelbaum RJ. CD36 and proteoglycan-mediated pathways for (n-3) fatty acid enriched
triglyceride-rich particle blood clearance in mouse models in vivo and in peritoneal
macrophages in vitro. J Nutr 2008;138:257-261.
420. Densupsoontorn N, Carpentier YA, Racine R, Murray FM, Seo T, Ramakrishnan R,
Deckelbaum RJ. CD36 and proteoglycan-mediated pathways for (n-3) fatty acid enriched
triglyceride-rich particle blood clearance in mouse models in vivo and in peritoneal
macrophages in vitro. J Nutr 2008;138:257-261.
421. Pohl J, Ring A, Korkmaz U, Ehehalt R, Stremmel W. FAT/CD36-mediated long-chain
fatty acid uptake in adipocytes requires plasma membrane rafts. Mol Biol Cell
2005;16:24-31.

184
422. Levy E, Stan S, Delvin E, Menard D, Shoulders C, Garofalo C, Slight I, Seidman E,
Mayer G, Bendayan M. Localization of microsomal triglyceride transfer protein in the
Golgi: possible role in the assembly of chylomicrons. J Biol Chem 2002;277:16470-
16477.
423. Fatma S, Yakubov R, Anwar K, Hussain MM. Pluronic L81 enhances triacylglycerol
accumulation in the cytosol and inhibits chylomicron secretion. J Lipid Res
2006;47:2422-2432.
424. Gleeson A, Anderton K, Owens D, Bennett A, Collins P, Johnson A, White D, Tomkin
GH. The role of microsomal triglyceride transfer protein and dietary cholesterol in
chylomicron production in diabetes. Diabetologia 1999;42:944-948.
425. Zilversmit DB. Atherogenic nature of triglycerides, postprandial lipidemia, and
triglyceride-rich remnant lipoproteins. Clin Chem 1995;41:153-158.
426. Cabezas MC, Erkelens DW. Triglycerides and atherosclerosis: to feast or fast. Neth J
Med 2000;56:110-118.
427. Karpe F, Steiner G, Uffelman K, Olivecrona T, Hamsten A. Postprandial lipoproteins and
progression of coronary atherosclerosis. Atherosclerosis 1994;106:83-97.
428. Phillips C, Bennett A, Anderton K, Owens D, Collins P, White D, Tomkin GH. Intestinal
rather than hepatic microsomal triglyceride transfer protein as a cause of postprandial
dyslipidemia in diabetes. Metabolism 2002;51:847-852.
429. Hsieh J, Longuet C, Baker CL, Qin B, Federico LM, Drucker DJ, Adeli K. The glucagon-
like peptide 1 receptor is essential for postprandial lipoprotein synthesis and secretion in
hamsters and mice. Diabetologia 2010;53:552-561.
430. Hsieh J, Longuet C, Maida A, Bahrami J, Xu E, Baker CL, Brubaker PL, Drucker DJ,
Adeli K. Glucagon-like peptide-2 increases intestinal lipid absorption and chylomicron
production via CD36. Gastroenterology 2009;137:997-1005, 1005.
431. Dube PE, Brubaker PL. Frontiers in glucagon-like peptide-2: multiple actions, multiple
mediators. Am J Physiol Endocrinol Metab 2007;293:E460-E465.
432. Nelson DW, Sharp JW, Brownfield MS, Raybould HE, Ney DM. Localization and
activation of glucagon-like peptide-2 receptors on vagal afferents in the rat.
Endocrinology 2007;148:1954-1962.
433. Dube PE, Forse CL, Bahrami J, Brubaker PL. The essential role of insulin-like growth
factor-1 in the intestinal tropic effects of glucagon-like peptide-2 in mice.
Gastroenterology 2006;131:589-605.
434. Bulut K, Pennartz C, Felderbauer P, Meier JJ, Banasch M, Bulut D, Schmitz F, Schmidt
WE, Hoffmann P. Glucagon like peptide-2 induces intestinal restitution through VEGF
release from subepithelial myofibroblasts. Eur J Pharmacol 2008;578:279-285.

185
435. Lewis GF, Naples M, Uffelman K, Leung N, Szeto L, Adeli K. Intestinal lipoprotein
production is stimulated by an acute elevation of plasma free fatty acids in the fasting
state: studies in insulin-resistant and insulin-sensitized Syrian golden hamsters.
Endocrinology 2004;145:5006-5012.
436. Korkmaz H, Bloch W, Bolck B, Labbe D, Addicks K, Arnhold S. The developmental
stage and cell type dependent phosphorylation of eNOS in murine enteric mucosa and
myenteric plexus. J Mol Histol 2007;38:227-235.
437. Nagy JA, Vasile E, Feng D, Sundberg C, Brown LF, Detmar MJ, Lawitts JA, Benjamin
L, Tan X, Manseau EJ, Dvorak AM, Dvorak HF. Vascular permeability factor/vascular
endothelial growth factor induces lymphangiogenesis as well as angiogenesis. J Exp Med
2002;196:1497-1506.
438. Olsson AK, Dimberg A, Kreuger J, Claesson-Welsh L. VEGF receptor signalling - in
control of vascular function. Nat Rev Mol Cell Biol 2006;7:359-371.
439. Fulton D, Gratton JP, McCabe TJ, Fontana J, Fujio Y, Walsh K, Franke TF,
Papapetropoulos A, Sessa WC. Regulation of endothelium-derived nitric oxide
production by the protein kinase Akt. Nature 1999;399:597-601.
440. Rask-Madsen C, King GL. Differential regulation of VEGF signaling by PKC-alpha and
PKC-epsilon in endothelial cells. Arterioscler Thromb Vasc Biol 2008;28:919-924.
441. Hayashi AA, Webb J, Choi J, Baker C, Lino M, Trigatti B, Trajcevski KE, Hawke TJ,
Adeli K. Intestinal SR-BI is upregulated in insulin resistant states and is associated with
overproduction of intestinal apoB48-containing lipoproteins. Am J Physiol Gastrointest
Liver Physiol 2011.
442. Sakai H, Hara H, Tsai AG, Tsuchida E, Intaglietta M. Constriction of resistance arteries
determines l-NAME-induced hypertension in a conscious hamster model. Microvasc Res
2000;60:21-27.
443. Savidge TC, Newman P, Pothoulakis C, Ruhl A, Neunlist M, Bourreille A, Hurst R,
Sofroniew MV. Enteric glia regulate intestinal barrier function and inflammation via
release of S-nitrosoglutathione. Gastroenterology 2007;132:1344-1358.
444. Garcia-Cardena G, Fan R, Shah V, Sorrentino R, Cirino G, Papapetropoulos A, Sessa
WC. Dynamic activation of endothelial nitric oxide synthase by Hsp90. Nature
1998;392:821-824.
445. Schild L, Dombrowski F, Lendeckel U, Schulz C, Gardemann A, Keilhoff G. Impairment
of endothelial nitric oxide synthase causes abnormal fat and glycogen deposition in liver.
Biochim Biophys Acta 2008;1782:180-187.
446. Doulias PT, Greene JL, Greco TM, Tenopoulou M, Seeholzer SH, Dunbrack RL,
Ischiropoulos H. Structural profiling of endogenous S-nitrosocysteine residues reveals

186
unique features that accommodate diverse mechanisms for protein S-nitrosylation. Proc
Natl Acad Sci U S A 2010;107:16958-16963.
447. Oliveira CP, Alves VA, Lima VM, Stefano JT, Debbas V, Sa SV, Wakamatsu A, Correa-
Giannella ML, de Mello ES, Havaki S, Tiniakos DG, Marinos E, de Oliveira MG,
Giannella-Neto D, Laurindo FR, Caldwell S, Carrilho FJ. Modulation of hepatic
microsomal triglyceride transfer protein (MTP) induced by S-nitroso-N-acetylcysteine in
ob/ob mice. Biochem Pharmacol 2007;74:290-297.
448. Storch J, Zhou YX, Lagakos WS. Metabolism of apical versus basolateral sn-2-
monoacylglycerol and fatty acids in rodent small intestine. J Lipid Res 2008;49:1762-
1769.
449. Tolessa T, Naslund E, Hellstrom PM. The inhibitory mechanism of GLP-1, but not
glucagon, on fasted gut motility is dependent on the L-arginine/nitric oxide pathway.
Regul Pept 2001;98:33-40.
450. Takahashi T, Yamaguchi S, Chida K, Shibuya M. A single autophosphorylation site on
KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA
synthesis in vascular endothelial cells. EMBO J 2001;20:2768-2778.
451. Holmqvist K, Cross MJ, Rolny C, Hagerkvist R, Rahimi N, Matsumoto T, Claesson-
Welsh L, Welsh M. The adaptor protein shb binds to tyrosine 1175 in vascular
endothelial growth factor (VEGF) receptor-2 and regulates VEGF-dependent cellular
migration. J Biol Chem 2004;279:22267-22275.
452. DeFronzo RA, Okerson T, Viswanathan P, Guan X, Holcombe JH, MacConell L. Effects
of exenatide versus sitagliptin on postprandial glucose, insulin and glucagon secretion,
gastric emptying, and caloric intake: a randomized, cross-over study. Curr Med Res Opin
2008;24:2943-2952.
453. Vella A, Bock G, Giesler PD, Burton DB, Serra DB, Saylan ML, Dunning BE, Foley JE,
Rizza RA, Camilleri M. Effects of dipeptidyl peptidase-4 inhibition on gastrointestinal
function, meal appearance, and glucose metabolism in type 2 diabetes. Diabetes
2007;56:1475-1480.
454. Schwartz EA, Koska J, Mullin MP, Syoufi I, Schwenke DC, Reaven PD. Exenatide
suppresses postprandial elevations in lipids and lipoproteins in individuals with impaired
glucose tolerance and recent onset type 2 diabetes mellitus. Atherosclerosis
2010;212:217-222.
455. Xiao C, Pavlic M, Szeto L, Patterson BW, Lewis GF. Effects of acute
hyperglucagonemia on hepatic and intestinal lipoprotein production and clearance in
healthy humans. Diabetes 2011;60:383-390.
456. Bunck MC, Corner A, Eliasson B, Heine RJ, Shaginian RM, Wu Y, Yan P, Smith U,
Yki-Jarvinen H, Diamant M, Taskinen MR. One-year treatment with exenatide vs. insulin

187
glargine: effects on postprandial glycemia, lipid profiles, and oxidative stress.
Atherosclerosis 2010;212:223-229.
457. Briand F. The use of dyslipidemic hamsters to evaluate drug-induced alterations in
reverse cholesterol transport. Curr Opin Investig Drugs 2010;11:289-297.
458. Luiken JJ, Dyck DJ, Han XX, Tandon NN, Arumugam Y, Glatz JF, Bonen A. Insulin
induces the translocation of the fatty acid transporter FAT/CD36 to the plasma
membrane. Am J Physiol Endocrinol Metab 2002;282:E491-E495.
459. Wang Y, Van Oort MM, Yao M, Van der Horst DJ, Rodenburg KW. Insulin and
Chromium Picolinate Induce Translocation of CD36 to the Plasma Membrane Through
Different Signaling Pathways in 3T3-L1 Adipocytes, and with a Differential
Functionality of the CD36. Biol Trace Elem Res 2010.
460. Schwenk RW, Dirkx E, Coumans WA, Bonen A, Klip A, Glatz JF, Luiken JJ.
Requirement for distinct vesicle-associated membrane proteins in insulin- and AMP-
activated protein kinase (AMPK)-induced translocation of GLUT4 and CD36 in cultured
cardiomyocytes. Diabetologia 2010;53:2209-2219.
461. Tran TT, Poirier H, Clement L, Nassir F, Pelsers MM, Petit V, Degrace P, Monnot MC,
Glatz JF, Abumrad NA, Besnard P, Niot I. Luminal lipid regulates CD36 levels and
downstream signaling to stimulate chylomicron synthesis. J Biol Chem 2011.
462. Beaslas O, Cueille C, Delers F, Chateau D, Chambaz J, Rousset M, Carriere V. Sensing
of dietary lipids by enterocytes: a new role for SR-BI/CLA-1. PLoS One 2009;4:e4278.
463. Ring A, Le Lay S, Pohl J, Verkade P, Stremmel W. Caveolin-1 is required for fatty acid
translocase (FAT/CD36) localization and function at the plasma membrane of mouse
embryonic fibroblasts. Biochim Biophys Acta 2006;1761:416-423.
464. Mitsutake S, Zama K, Yokota H, Yoshida T, Tanaka M, Mitsui M, Ikawa M, Okabe M,
Tanaka Y, Yamashita T, Takemoto H, Okazaki T, Watanabe K, Igarashi Y. Dynamic
modification of sphingomyelin in lipid microdomains controls development of obesity,
fatty liver, and type 2 diabetes. J Biol Chem 2011.
465. Kincer JF, Uittenbogaard A, Dressman J, Guerin TM, Febbraio M, Guo L, Smart EJ.
Hypercholesterolemia promotes a CD36-dependent and endothelial nitric-oxide synthase-
mediated vascular dysfunction. J Biol Chem 2002;277:23525-23533.
466. Zhu W, Smart EJ. Myristic acid stimulates endothelial nitric-oxide synthase in a. J Biol
Chem 2005;280:29543-29550.
467. Steinbusch LK, Wijnen W, Schwenk RW, Coumans WA, Hoebers NT, Ouwens DM,
Coumans WA, Hoebers NT, Diamant M, Bonen A, Glatz JF, Luiken JJ. Differential
regulation of cardiac glucose and fatty acid uptake by endosomal pH and actin filaments.
Am J Physiol Cell Physiol 2010;298:C1549-C1559.

188
468. Zeng Y, Tao N, Chung KN, Heuser JE, Lublin DM. Endocytosis of oxidized low density
lipoprotein through scavenger receptor CD36 utilizes a lipid raft pathway that does not
require caveolin-1. J Biol Chem 2003;278:45931-45936.
469. Garcia-Martinez C, Marotta M, Moore-Carrasco R, Guitart M, Camps M, Busquets S,
Montell E, Gomez-Foix AM. Impact on fatty acid metabolism and differential
localization of FATP1 and FAT/CD36 proteins delivered in cultured human muscle cells.
Am J Physiol Cell Physiol 2005;288:C1264-C1272.
470. Hoosdally SJ, Andress EJ, Wooding C, Martin CA, Linton KJ. The Human Scavenger
Receptor CD36: glycosylation status and its role in trafficking and function. J Biol Chem
2009;284:16277-16288.
471. Thorne RF, Ralston KJ, de Bock CE, Mhaidat NM, Zhang XD, Boyd AW, Burns GF.
Palmitoylation of CD36/FAT regulates the rate of its post-transcriptional processing in
the endoplasmic reticulum. Biochim Biophys Acta 2010;1803:1298-1307.
472. Smith J, Su X, El Maghrabi R, Stahl PD, Abumrad NA. Opposite regulation of CD36
ubiquitination by fatty acids and insulin: effects on fatty acid uptake. J Biol Chem
2008;283:13578-13585.
473. Lynes MD, Widmaier EP. Involvement of CD36 and intestinal alkaline phosphatases in
fatty acid transport in enterocytes, and the response to a high-fat diet. Life Sci
2011;88:384-391.
474. Suzuki N, Takahashi S, Okabe S. Relationship between vascular endothelial growth
factor and angiogenesis in spontaneous and indomethacin-delayed healing of acetic acid-
induced gastric ulcers in rats. J Physiol Pharmacol 1998;49:515-527.
475. Szabo S, Vincze A, Sandor Z, Jadus M, Gombos Z, Pedram A, Levin E, Hagar J, Iaquinto
G. Vascular approach to gastroduodenal ulceration: new studies with endothelins and
VEGF. Dig Dis Sci 1998;43:40S-45S.
476. Parvadia JK, Keswani SG, Vaikunth S, Maldonado AR, Marwan A, Stehr W, Erwin C,
Uzvolgyi E, Warner BW, Yamano S, Taichman N, Crombleholme TM. Role of VEGF in
small bowel adaptation after resection: the adaptive response is angiogenesis dependent.
Am J Physiol Gastrointest Liver Physiol 2007;293:G591-G598.
477. Lei NY, Ma G, Zupekan T, Stark R, Puder M, Dunn JC. Controlled release of vascular
endothelial growth factor enhances intestinal adaptation in rats with extensive small
intestinal resection. Surgery 2011;150:186-190.
478. Boquoi A, Jover R, Chen T, Pennings M, Enders GH. Transgenic expression of VEGF in
intestinal epithelium drives mesenchymal cell interactions and epithelial neoplasia.
Gastroenterology 2009;136:596-606.

189
479. Boughton-Smith NK, Hutcheson IR, Deakin AM, Whittle BJ, Moncada S. Protective
effect of S-nitroso-N-acetyl-penicillamine in endotoxin-induced acute intestinal damage
in the rat. Eur J Pharmacol 1990;191:485-488.
480. Tepperman BL, Brown JF, Whittle BJ. Nitric oxide synthase induction and intestinal
epithelial cell viability in rats. Am J Physiol 1993;265:G214-G218.
481. Moore BA, Peffer N, Pirone A, Bassiri A, Sague S, Palmer JM, Johnson DL, Nesspor T,
Kliwinski C, Hornby PJ. GLP-2 receptor agonism ameliorates inflammation and
gastrointestinal stasis in murine postoperative ileus. J Pharmacol Exp Ther 2010;333:574-
583.
482. Liu Y, Huang J, Hou Y, Zhu H, Zhao S, Ding B, Yin Y, Yi G, Shi J, Fan W. Dietary
arginine supplementation alleviates intestinal mucosal disruption induced by Escherichia
coli lipopolysaccharide in weaned pigs. Br J Nutr 2008;100:552-560.
483. Di Lorenzo M, Bass J, Krantis A. Use of L-arginine in the treatment of experimental
necrotizing enterocolitis. J Pediatr Surg 1995;30:235-240.
484. Puiman PJ, Stoll B, van Goudoever JB, Burrin DG. Enteral arginine does not increase
superior mesenteric arterial blood flow but induces mucosal growth in neonatal pigs. J
Nutr 2011;141:63-70.
485. Mourad FH, Turvill JL, Farthing MJ. Role of nitric oxide in intestinal water and
electrolyte transport. Gut 1999;44:143-147.
486. Baldassano S, Liu S, Qu MH, Mule F, Wood JD. Glucagon-like peptide-2 modulates
neurally evoked mucosal chloride secretion in guinea pig small intestine in vitro. Am J
Physiol Gastrointest Liver Physiol 2009;297:G800-G805.
487. Coon S, Kekuda R, Saha P, Talukder JR, Sundaram U. Constitutive nitric oxide
differentially regulates Na-H and Na-glucose cotransport in intestinal epithelial cells. Am
J Physiol Gastrointest Liver Physiol 2008;294:G1369-G1375.
488. Arthur S, Saha P, Kekuda R, Coon S, Sundaram U. 600 Constitutive Nitric Oxide (CNO)
Inhibits Na-Glucose Co-Transport By Altering the Glycosylation of the SGLT1 Co-
Transporter in the Intestinal Epithelial Cells. Gastroenterology 2008;134:A-83.
489. Kapur S, Bedard S, Marcotte B, Cote CH, Marette A. Expression of nitric oxide synthase
in skeletal muscle: a novel role for nitric oxide as a modulator of insulin action. Diabetes
1997;46:1691-1700.
490. Balon TW, Nadler JL. Evidence that nitric oxide increases glucose transport in skeletal
muscle. J Appl Physiol 1997;82:359-363.
491. Roberts CK, Barnard RJ, Scheck SH, Balon TW. Exercise-stimulated glucose transport in
skeletal muscle is nitric oxide dependent. Am J Physiol 1997;273:E220-E225.

190
492. Bradley SJ, Kingwell BA, McConell GK. Nitric oxide synthase inhibition reduces leg
glucose uptake but not blood flow during dynamic exercise in humans. Diabetes
1999;48:1815-1821.
493. Tanaka T, Nakatani K, Morioka K, Urakawa H, Maruyama N, Kitagawa N, Katsuki A,
Araki-Sasaki R, Hori Y, Gabazza EC, Yano Y, Wada H, Nobori T, Sumida Y, Adachi Y.
Nitric oxide stimulates glucose transport through insulin-independent GLUT4
translocation in 3T3-L1 adipocytes. Eur J Endocrinol 2003;149:61-67.
494. Zou MH, Kirkpatrick SS, Davis BJ, Nelson JS, Wiles WG, Schlattner U, Neumann D,
Brownlee M, Freeman MB, Goldman MH. Activation of the AMP-activated protein
kinase by the anti-diabetic drug metformin in vivo. Role of mitochondrial reactive
nitrogen species. J Biol Chem 2004;279:43940-43951.
495. Chopra AR, Kommagani R, Saha P, Louet JF, Salazar C, Song J, Jeong J, Finegold M,
Viollet B, DeMayo F, Chan L, Moore DD, O'Malley BW. Cellular energy depletion
resets whole-body energy by promoting coactivator-mediated dietary fuel absorption.
Cell Metab 2011;13:35-43.
496. Kamagate A, Qu S, Perdomo G, Su D, Kim DH, Slusher S, Meseck M, Dong HH. FoxO1
mediates insulin-dependent regulation of hepatic VLDL production in mice. J Clin Invest
2008;118:2347-2364.
497. Ben Shlomo S, Zvibel I, Shnell M, Shlomai A, Chepurko E, Halpern Z, Barzilai N, Oren
R, Fishman S. Glucagon-like peptide-1 reduces hepatic lipogenesis via activation of
AMP-activated protein kinase. J Hepatol 2010.
498. Hein GJ, Baker CL, Hsieh J, Adeli K. Effects of GLP-1 and GLP-2 Co-Administration on
Regulation of Intestinal Lipid Metabolism and Chylomicron Production in Syrian Golden
Hamsters. Diabetes 2011;60:0682-P.
499. Cohen JC, Berger GM. Effects of glucose ingestion on postprandial lipemia and
triglyceride clearance in humans. J Lipid Res 1990;31:597-602.
500. Lally S, Owens D, Tomkin GH. Genes that affect cholesterol synthesis, cholesterol
absorption, and chylomicron assembly: the relationship between the liver and intestine in
control and streptozotosin diabetic rats. Metabolism 2007;56:430-438.
501. Hogue JC, Lamarche B, Tremblay AJ, Bergeron J, Gagne C, Couture P. Evidence of
increased secretion of apolipoprotein B-48-containing lipoproteins in subjects with type 2
diabetes. J Lipid Res 2007;48:1336-1342.
502. Lambeir AM, Proost P, Scharpe S, De M, I. A kinetic study of glucagon-like peptide-1
and glucagon-like peptide-2 truncation by dipeptidyl peptidase IV, in vitro. Biochem
Pharmacol 2002;64:1753-1756.

191
503. Hansen L, Hare KJ, Hartmann B, Deacon CF, Ugleholdt RK, Plamboeck A, Holst JJ.
Metabolism of glucagon-like peptide-2 in pigs: role of dipeptidyl peptidase IV. Regul
Pept 2007;138:126-132.
504. Hartmann B, Thulesen J, Kissow H, Thulesen S, Orskov C, Ropke C, Poulsen SS, Holst
JJ. Dipeptidyl peptidase IV inhibition enhances the intestinotrophic effect of glucagon-
like peptide-2 in rats and mice. Endocrinology 2000;141:4013-4020.
505. Borgstrom B, Dahlqvist A, Lundh G, SjovallJ. Studies of intestinal digestion and
absorption in the human. J Clin Invest 1957;36:1521-1536.
506. Atkinson RM, Parson BJ, Smyth DH. The intestinal absorption of glucose. J Physiol
1957;135:581-589.
507. Ueno T, Hamanaka Y, Oka M, Suzuki T. Oral absorption tests: absorption site of each
substrate. Nutrition 1998;14:7-10.
508. Armand M, Pasquier B, Andre M, Borel P, Senft M, Peyrot J, Salducci J, Portugal H,
Jaussan V, Lairon D. Digestion and absorption of 2 fat emulsions with different droplet
sizes in the human digestive tract. Am J Clin Nutr 1999;70:1096-1106.
509. Brubaker PL, Drucker DJ, Asa SL, Swallow C, Redston M, Greenberg GR. Prolonged
gastrointestinal transit in a patient with a glucagon-like peptide (GLP)-1- and -2-
producing neuroendocrine tumor. J Clin Endocrinol Metab 2002;87:3078-3083.
510. Hata T, Mera Y, Ishii Y, Tadaki H, Tomimoto D, Kuroki Y, Kawai T, Ohta T, Kakutani
M. JTT-130, a novel intestine-specific inhibitor of microsomal triglyceride transfer
protein, suppresses food intake and gastric emptying with the elevation of plasma peptide
YY and glucagon-like peptide-1 in a dietary fat-dependent manner. J Pharmacol Exp
Ther 2011;336:850-856.
511. Hagberg CE, Falkevall A, Wang X, Larsson E, Huusko J, Nilsson I, van Meeteren LA,
Samen E, Lu L, Vanwildemeersch M, Klar J, Genove G, Pietras K, Stone-Elander S,
Claesson-Welsh L, Yla-Herttuala S, Lindahl P, Eriksson U. Vascular endothelial growth
factor B controls endothelial fatty acid uptake. Nature 2010;464:917-921.
512. Bartelt A, Bruns OT, Reimer R, Hohenberg H, Ittrich H, Peldschus K, Kaul MG,
Tromsdorf UI, Weller H, Waurisch C, Eychmuller A, Gordts PL, Rinninger F,
Bruegelmann K, Freund B, Nielsen P, Merkel M, Heeren J. Brown adipose tissue activity
controls triglyceride clearance. Nat Med 2011;17:200-205.
513. Sadur CN, Eckel RH. Insulin stimulation of adipose tissue lipoprotein lipase. Use of the
euglycemic clamp technique. J Clin Invest 1982;69:1119-1125.
514. Fielding BA, Callow J, Owen RM, Samra JS, Matthews DR, Frayn KN. Postprandial
lipemia: the origin of an early peak studied by specific dietary fatty acid intake during
sequential meals. Am J Clin Nutr 1996;63:36-41.

192
515. Raju K, Doulias PT, Tenopoulou M, Greene JL, Ischiropoulos H. Strategies and tools to
explore protein S-nitrosylation. Biochim Biophys Acta 2011.
516. Parastatidis I, Thomson L, Fries DM, Moore RE, Tohyama J, Fu X, Hazen SL, Heijnen
HF, Dennehy MK, Liebler DC, Rader DJ, Ischiropoulos H. Increased protein nitration
burden in the atherosclerotic lesions and plasma of apolipoprotein A-I deficient mice.
Circ Res 2007;101:368-376.












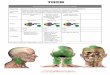








![Vascular Liver Disease and the Liver 2 Sinusoidal Endothelial ...play an important role in initiating atherosclero-sis [34, 35]. Decreased chylomicron remnant clearance with aging-related](https://img.pdfslide.us/doc/110x75/6047e1ed9bc5de65774b97b2/vascular-liver-disease-and-the-liver-2-sinusoidal-endothelial-play-an-important.jpg)






