Embed Size (px)
Citation preview

Estuarine, Coastal and Shelf Science (1983) 16, 217- 227
The Abundance and Distribution of Free-living Nematodes from. Two Canadian Pacific Beaches
Jyotsna Sharmaa and John M. Webster Department of Biological Sciences, Simon Fraser University, Burnaby, Vancouver, British Columbia, Canada VJA 15%
Received 22 October 1981 and in revisedform 16 March 1982
Keywords: Xematoda; littoral; distribution environmental conditions; Canada west coast
Free-living marine nematodes at two littoral sites near Vancouver, Canada were studied over a I -year period. Sixty-four species were found at a Belcarra Park site, with a maximum nematode density of 6.7 x lo5 m -a and 24 species at Iona Island, with a maximum density of 1.6 x 10~ m -2. At both sites most of the nematodes occurred in the upper 2 cm of the 6 cm depth of sediment examined, and the vertical distribution of most species remained constant during the year. The distribution of the nematodes in relation to some physical parameters is discussed. The occurrence of certain species suggests that they have a worldwide distribution.
Introduction
Free-living nematodes are the major component of the meiofauna in most marine benthic habitats, from the littoral to the deep sea. In a recent review of their role in the littoral ecosystem, Platt & Warwick (1980) conclude that ‘any general assessment of intertidal habitats is incomplete if the nematode fauna is not taken into consideration’.
Though several publications on the natural history and ecology of the North American Pacific coast are available, there are only a few taxonomic descriptions on free-living nematodes from the Canadian Pacific coast (Nelson et al., 1971, 1972). The taxonomic relationships of two of the cyatholaimids discussed in this paper, Paracyatholaimuspugettensis and P. vancouverensis, are discussed by Sharma & Vincx (1982) and Paracanthonchus caecus is described as P. canadensis n. sp. by Vincx et al. (in press). This paper describes the composition of the nematode fauna over a I-year period at two contrasting sites near Vancouver, British Columbia. The density of nematodes and vertical distribution of the dominant species in the surficial sediment is recorded and discussed in relation to some of the physical parameters.
Materials and methods
Samples were taken during 1976 at two sites in the area of Vancouver, British Columbia, namely Iona Island (49’1o’N, 123’15’W) at 2.13 m above mean tide level and Belcarra Park (49’19’N, 123”56’W) at 0.76 m above mean tide level.
“Present address: Instituut voor Dierkunde, Rijksuniversiteit Gent, K. L. Ledeganckstraat-36, gooc-Gent, Belgium.
2x7 o2?2-7714/83/o20217$-11 $03.00/o ‘G 1983 Academic Press Inc. (London) Limited

218 J. Sharma &‘J. M. Webster
At each site, three replicate samples were taken within a r-m2 quadrat on six occasions during the year using a hand-held plexiglass corer (2.4 cm diameter). In addition, approximately 500 g of sediment were collected from the quadrat for organic carbon measurement and sediment grain size analysis. The upper 6 cm of the core were sliced into 2 cm sections and nematodes extracted in the following way (based on Hopper & Meyers, 1967): the sediment was first passed through a ~.o-mm sieve to remove coarse particles and then through a 44-pm sieve to separate the silt-clay particles. Sediment retained on this sieve was centrifuged twice in a I : I Karo syrup-sea water mixture (x80 g for 2 min) and the supernatant poured through a 44-pm sieve. Organisms retained on the fine sieve were fixed in hot 5% formalin-sea water and the nematodes were processed through glycerin (Seinhorst, 1959) for identification.
Temperature and salinity measurements of the overlying water were taken at the site of sampling using a Yellow Springs Instrument S-C-T meter. The sediment’s grain size was determined by the technique of Buchanan (1971) and the sorting determined by the phi quartile deviation. Organic carbon was measured with a Perkin-Elmer Elemental analyzer.
A three-level nested analysis of variance was performed on the total number of species and log @+I transformed nematode density with the components of site, time and depth. A three-way analysis of variance on the log n+~ transformed data of the 17 dominant species was done with the components of time, depth and developmental stage.
Results
Environmental parameters Seawater temperature tended to be lower throughout the year at Iona Island than at Belcarra Park (Table I). Salinity at both sites was highest in February and lowest in the summer especially at Iona Island (Table I). The mean grain size composition of the sediment (Table I) was higher at Belcarra Park than at Iona Island but the sediments at Iona Island were better sorted than those at Belcarra Park. By the scale of Folk & Ward (1957), the Iona Island sediment was in the range of well to moderately sorted while the Belcarra Park sediment was moderately to poorly sorted. The amount of organic carbon associated with sediments was
TABLE I. Temperature and salinity of water overlying sediments and the percentage silt-clay content, mean grain size, sorting and percentage carbon content of sediments at Belcarra Park and Iona Island sites over I year
Belcarra Park 8 Feb. 7 Apr. 20 May 20 Aug. IO Nov. IO Dec.
Temperature (“C) Salinity (x0) o/O silt-clay Mean grain size (mm) Sorting 0/O carbon
IO’0 23’5
-
- 2’1
16.5 ‘4’5 20’0 10’0 6.5 - 19’0 21’0 18.0 14’5 5.6 7.6 15‘2 15.6 12’0
0’574 0.574 0’392 0.709 0.a13
0.81 1’10 0.80 1’90 “55 “3 4’1 I .6 2’2 2’2
Iona Island II Jan. 7 Mar. 19 Apr. 31 May 15 Oct. 20 Dec.
Temperature (“C) Salinity (x0) % silt-clay Mean grain size (mm) Sorting yO carbon
3’0 9’5 - ‘4’5 10’0 1’0
23.0 16.0 - 12.5 7.0’0 I 8.0 4’4 4’4 6.0 8.0 40.0 25’2
0.420 0.483 0.467 0’595 0’234 0.268 0’45 0’55 0.68 0’50 1’00 0.85 0’5 0.8 0’7 I.4 0.8 0.4

TABL
E 2.
Nu
mbe
r of
sp
ecie
s an
d ne
mat
ode
dens
ity
per
18
cm2
at
the
Belca
rra
Park
and
Iona
Isl
and
sites
at
thr
ee
dept
hs
sam
pled
over
I
year
. Ea
ch
value
is
th
e m
ean
of
three
re
plica
te
sam
ples&
s.D.
(mea
n va
lues
rndl
are
in
pare
nthe
ses)
Balca
rra
Park
Dept
h (cm
)
o-2
8 Fe
brua
ry 7
April
20
May
I Au
gust
IO
Nove
mbe
r IO
De
cem
ber
SP.
Dens
ity
SP.
Dens
ity
Sp.
Dens
ity
SP.
Dens
ity
Sp.
Dens
ity
Sp.
Dens
ity
21
51of
253
31
422&
135
3o
38o~
t235
36
85
2f715
29
23
2415
5 27
36
2 +7
4 (2
.83
X 10
~)
(2.3
4 x
IO’)
(2.1
I
x 10
5)
(4’73
x
IO51
(1
.29X
Io5)
(2
.01
x 10
6)
2-4
I2
256&
187
25
257x
t181
22
Io9f
47
26
3o5+
355
22
228
4~ 17
7 I2
16
oh81
(1
.42
X Io
4)
(I.43
x
106)
(6.
10
X IO
”) (1.
69
X 10
~)
(1.27
X
10”)
(8.90
X
IO’)
4-6
IO
70+4
8 22
12
6~t25
23
85
145
15
43
*4r
I2
ro
5f13
o 19
37
xt16
(3;;;
x 104
) (7.
00
x IO
”) (4
.70
x 10
4)
(2.40
X
100
(5.80
x
10~)
(2
.10
x IO
’)
Tota
l 80
5 57
3 11
99
407
560
(4.53
x
107
(4’47
x
107
(3.18
X
10~)
(6.
66
x 10
5)
(2.26
X
IO’)
(3.11
X
Io5)
---~ Iona
Isl
and
II Ja
nuar
y 7
Mar
ch
19
April
31
May
15
Octob
er
20
Dece
mbe
r -
Dept
h (cm
) Sp
. De
nsity
SP
. De
nsity
Sp
. De
nsity
Sp
. De
nsity
Sp
. De
nsity
SP
. De
nsity
o-2 2-4
4-6
Tota
l
8 31
&12
6 43
dz
41
II 59
i26
I4
Io9i3
1 7
193
zt74
IO
197*
5o
(I.70
x
100
(2.40
X
10~)
(3
.30
x 104
) (6.
05
x 10
~)
(1.07
X
10~)
(1.
09
x 10
~)
3 23
*4
3 IS
&II
5 r4
f3 (1.
30X
10~)
(8
.30
x 10
8)
(7.7
0X
103)
I0
54
*45
2 76
f34
3 75
177
(3
’00
x IO
’) (4
.20
X 10
~)
(4’17
X
IO’)
3 II&
IO
2 91
7 4
8zt4
5 12
59
2 26
&s
2 IS
+19
(6.10
X 10
~)
(5’00
x
103)
64
67
(44;
x ro
3)
(6.70
x
10~)
(1.
40x
10~)
(8
.30
x 10
~)
I75
295.
28
7 (3
.60
X ro
4)
(3’70
x
IO’)
(4.50
x
IO’)
(9’70
x
104)
(1.
64x
IO?
(1.59
x IO
51

220 J. Sharma &J. M. Webster
higher at Belcarra Park than at Iona Island (Table I). The high carbon content at Belcarra Park in May was associated with the presence ‘of an abundance of visible detritus and a substantial diatom population. No measurements were made on the redox potential but the redox discontinuity layer was visible at a depth of x*5-3*0 cm at Iona Island and 2-o-3.0 cm at Belcarra Park.
The nematode fauna The nematode density at Belcarra Park was significantly (P<o.og) higher than at Iona Island throughout the year (Table 2). The maximum and minimum densities in the upper 6 cm of the sediment at BeIcarra Park were 6.7 x ro5 mm2 and 2.3 x 10~ mm2 during August and November, respectively. The maximum and minimum at Iona Island, however, were 1.6 x 10~ me2 and 3.6 x 10~ rnp2 during October and February, respectively. At both sites total nematode density was highest in the upper 2 cm of the sediment, and although absolute density changed with time there was no significant change in the relative vertical distribution (Table 2).
The number of species recorded for Belcarra Park (64) was significantly greater than at Iona Island (24) (Table 3). Significantly more species (P<o.og) occurred in the summer at both sites than in the winter (Table 2). The proportional vertical distribution of species in the top 3 cm of sediment (Table 2) did not change during the year. Significantly (P<o.og) more species were found in the top 2 cm of sediment and few species were found below 4 cm, especially at Iona Island.
TABLE 3. List of nematode species occurring at the Belcarra Park and Iona Island sites during the year of sampling
Species Feeding
category” Belcarra
Park Iona
Island
Leptolaimus sp. Areolaimus elegans A. boomerangi Axonolaimus spitwsus Odontophora peritricha Parascolaimus sp. Paraterschellingia sp. Siphonolaimus sp. Solenolaimus sp. Desmolaimus zeelandicus Metalinhomoeus setosus Terschellingia longicaudata Eleutherolaimus stensoma Didelta sp. Linhomoeus sp. L. undulatus Paralinhomoeus buculentus Cylindrotheristus resimus Diplolaimella sp. Mesotheristus circumscriptus Monhystera refringens Paramonhystera mutilus Paramonhystera sp. Steineria gerlachi Theristus modicus Cobbia urinator Sphaerolaimus sp. Aponchium sp.
IA 1A IA IB IB IB IA 2B IA IB IB IA IB IB 2A 2A IB IB IB IB IB IB IB IB IB 2A 2B 2B
+++ +++ +++ +++ +++ ++ ++ +++ + +++ +++ +++ +++ ++ +++ +++ ++ + +++ + t-t+ +++ +++ +++ +++ +++ +++ +
+
+++
+
+++
+++
+
+++
+++

Free-living nematodes at two Canadian Pacific beaches 221
TABLE 3 continued
Species Feeding
category’ Belcarra
Park Iona
Island
Ehynchonema sp. SB Microlaimus dentatus 2A M. texianus ZA Monoposthia costata 2A Sabatieria americana IB S. ancudiana IB S. pulchra IB Chromadorina germanica 2A Euchromadora sp. 2A Denticulella sp. IA Innocuonema clivosum 2A Neochromadora appiana 2A Spilophorella paradoxa 2A Neotonchus sp. 2A Parapomponema websteria 2B Acanthonchus sp. 2A Paracanthonchus caecus 2A Paracyatholaimus pugetten Gs 2A P. vancouverensis 2A Cyatholaimus sp. 2A Hals3oanolaimus sp. 2B Bathylaimus sp. IB Paratripyloides sp. IB Triploides gracilis IB Cytolaimium sp. IA Bhabdocoma sp. 1A Oxystomina sp. IA Halalaimus sp. IA Anticoma pellucida IB Lauratonema pugiunculus IB Mesacanthion sp. 2B Viscosia carnleyensis 2B V. tumidula ZB Oxyonchus sp. 2B Anoplostoma sp. IB Onckolaimus skawensis 2B 0. vesicarius 2B Euystomina sp. 2B Symplocostoma sp. 2B Bhabditis marina IA Bolbella rp. 2B
+-t +-I+ ++ +t-+ +++ +++ +++ +++ +-t+ +++ +-I +++ + + +++ +++ +++ +++ ++ +++ +++ +++
t+t +-I-+ +++ +++ +++ ++ + ++ ++ +++ +++ t +++ ++
t +++ ++ +++
+++ +++ +++ + ++ +
+++
++ ++
+ +++
f, occurred in one sampling period; + -t, occurred in two sampling periods; + + +, occurred in more than two sampling periods. “Categorized as described by Wieser (1952) (see text).
Those species which typically comprised over 10% of the samples at each site (25 at Belcarra and six at Iona) were examined in detail. The remaining species were excluded from this analysis as they occurred in too low numbers. The mean number of the most abundant species and their prevalence in the fauna was determined (Table 4). All species are categorized (Table 3) according to the feeding categories proposed by Wieser (1952), namely: IA, selective deposit feeders; IB, non-selective deposit feeders; 2A, epigrowth feeders; 2B, predators and omnivores. At Iona Island the six dominant species were non-selective deposit feeders (xB) and epigrowth feeders (2B) and the 25 dominant nematode species at Belcarra Park represented all four of Wieser’s feeding categories.

TABL
E 4.
Nu
mbe
r an
d pe
rcen
tage
ab
unda
nce
(in
pare
nthe
ses)
at
Be
lcarra
Pa
rk an
d Io
na
Islan
d sit
es
of
the
dom
inant
ne
mat
ode
spec
ies
in
the
uppe
r 6
cm
of
sedim
ent
(18.0
9 cm
*) ov
er
I ye
ar
(fkS.
D.
of
three
re
plica
tes)
. Th
e sig
nific
ance
lev
el of
th
e co
mpo
nent
s of
tim
e (T
), de
pth
(D)
and
deve
lopm
ent
stage
s (S
) ar
e al
so
given
’
Belca
rra
Park
8 Fe
b. 7
Apr.
20
May
I Au
g.
IO
Nov.
zo
Dec.
T D
S TD
Areo
laim
us
elega
ns
Desm
olaim
us
zeela
ndicu
s
Met
alinh
omoe
us
seto
sus
Ters
chel
lingi
a lon
gicau
data
Eleu
ther
olaim
us
steno
som
a
Para
linho
moe
us
bucu
lentu
s
Linho
moe
us
sp.
L.
undu
latus
Para
mon
ohys
tera
sp.
P.
mut
ila
Cobb
ia ur
inato
r
Ther
istus
m
odicu
s
Saba
tieria
sp
.
S.
ancu
diana
S.
amer
icana
S.
pulch
ra
11+8
(I.
4 98
zt43
(13.5)
0 24*12
(3.
2) 2If
I4 (2.8
) 0 2058
(3.3)
0 0 25*34
58
+21
(3.0)
(8.2
) IfI
41
*3I
(0.2
) (5.
6) 0
0
309f3
Io 21
*21
(34.8)
(3.
1) 0
0
0 0
ZIfI9
(1
.1)
4&s (043
) 30
*40
(3.3)
78f74
(8.
4) 12
*18 (1W
41
8 (0.
4) 9+
4 (1.2
) Ii2
(0.
3) 0 0 0
II&IO
(1.2
) II*
10
(1.8
) 7*3
(1
4)
108f9
6 (2
1.0)
51
f28
(9.0)
2Ozt2
7 W
I 29
1-24
(5.9)
27k-2
1 (5
.0)
6+x0
(1.7)
29i21
(4.
4) 7+
6 (0.
9) 31
4 (0
.5)
37zt2
9 (6.
4) IfI
(0.1
)
2f3
(0.4)
IIztI7
(1
.5)
46h5
3 (4.
4) 44
561
(2.8
) 9f7
(1
.0)
209*2
02
(15’
0)
28+2
0 (3'
1) 48
143
(3'9) 1z
.t2
(0.1
) 28
511~
390
(15.4)
30
f41
(3.0)
srf38
(5
’1)
33f33
(3.
0) 14
zt8
(1%
68
ct39
(89)
26
k36
(1.4)
II&
IO
(0.5
) 0
~&IO
(1.8
) 85
5 (1
.8)
11*1
6 (2.5
) 12
5*12
8
(17-J)
IIz
ti’
(2.5
) 2O
zk-13
(3.
8) Ii2
(0.1
) 2I
f20 (2.7)
x0*1
6
(2.4)
28*18
(6.5)
29zk
-28
(3.8)
IOk7
(1
.8)
94f62
(19
.0)
I7fI4 (2
.5)
15+2
(6
.1)
0
8zt6
(1.
9) 42
*24
(7.3)
0 3954
4 (6.
4) 3*3
(0
.5)
241tI
I (4'
5) Ii2
(0.1
)
2lltI
(0
.5)
IIt1
(0.1
)
715 (1.4)
0 4*2
(0.7)
147+
67
(19.2)
0 0 8414
3 I
I (15
'71
+++
+++
+++
++-t
+++
+++
+++
+++
+--
+ 4.
.
+++
+++
n.s.
n.s.
n.s.
n.s.
n.s.
i i-
+
n.s.
+++
+++
+++
++i
+++
+++
+++
is+
i+i
+++
+++ KS
.
t-t+
+++
+++
+++
+++
-+-

S.
jubat
a
Neoc
hrom
ador
a ap
piana
Para
cyat
holai
mus
pu
gette
nsis
P.
vanc
ouve
rens
is
Neot
onch
us
sp.
Spilo
phor
ella
para
doxa
Tripy
loide
s gr
acilis
Visc
osia
ca
rnley
ensis
V.
tum
idula
0
39z!z
I4 (5.
9) 12
7+x2
0 (21
.0)
0 0 0 0 0 0
0
21xtI
3 (8
4 22
f21 (2.3)
31+3
6 (3.
3) 17
f20
(3.1)
56f5
2 (6
.0)
5zt6
(0
.6)
48h7
3 (4.
5) 8rk
I3 (1
’4
9ir2
(0.4) 6zt8
(1
.8)
22f32
(3.3)
6zt9 (043
) If1
(0
4 6f3
(1
’0)
0
241t1
6
(4.6)
51
f46
(9.0)
0 3rt2
(0.5
) 24
i26
(3-I)
26
+23
(x-7)
IIr
t5
(2’9)
9rt4
(0.7)
0
4Orfr3
4 (2.
7) 4x
*29
(4.8)
0 18&1
8
(4’3) ZjcI
(0
.5)
0 7f9
(1.0
) IO
*10 (2
.5)
2It:I
(0.
5) 18
lk25
(2
’1)
I9lfI2
(4.
6)
n.s.
+++
n.s.
+++
0 4613
2 (7.
9) 21
f23
++
+ ILS
.
(3.4)
0 ++
+ n.
s.
0 IfI (0.2
) 0 6+
4 n.
s. n.
s. n.
s. ++
i (1
.1)
14fIO
it-+
n.
s. ++
+ ++
i (2.
7)
Iona
Isl
and
II Ja
n.
7 Ma
r. 19
Apr
. 31
Ma
y 15
Oc
t. 20
De
c. T
D S
TD
Para
scola
imus
sp
. 74
8 3*2
7f6
3i2
0
0
@+2
(6
.0)
(6.4)
(1’4)
Ther
istus
sp
. 8=
t4 2f4
22
f9 IO
f17
0 5=
tfr8
+++
n.s.
++
+ n.
s. (13
.9)
(5.5
) (26
.2)
(6.1
) (2
.6)
Saba
tieria
pu
lchra
33
0 40
*41
IffIO
42
ytII
253fI
IO
209f2
3 ++
+ n.
s.
+++
+++
(48.0)
(50
.0)
(27.4)
(31
.3)
(84.0)
(73
.9)
Neoc
hrom
ador
a sp
. 3f
I If2
2&
I 16
-f14
0 If1
++
+ n.
s.
n.s.
n.
s.
(5.1
) (4.
7) (2.
3) (5
’4
(1-J
Pa
raca
ntho
nchu
s ca
nade
trtis
7*x
1418
X9
*12
2841
8 32
16
3813
n.
s. n.
s. ++
+ n.
s. (12
.0)
(22.4)
(8
.1)
0-w
(12.0)
(13
.1)
Tripy
loide
s gr
acilis
2*3
3f4
0
IIf
452
16f13
n.
s.
n.s.
n.
s. n.
s.
(z-6
) (5
.1)
(1243
) (1.
9) (5.
3)
“+
+ +
= PC
o.05
; n.
s. =
not
signi
fican
t.

224 J. Sharna &‘J. M. Webster
IO 0
o- 6 o- 6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6
-6 O-6 Oe60-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6
IO
0 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6
O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 Fob 8 Apr 7 May20 Aq I Now IO Des 20 Rb8 Apr7 May2OAugl NovlOOa20 hb 8 APT 7 May 20 Auq I NW IO 0% 20
Parocanthonchus caecus
IO 0 Depth
O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 O-6 o - 6 bn)
Jonll Mor7 AprlS May31 Ocfl5 0.~20 Jonll Mar7 AprlS Mey 3, OsfI5 hc20 Jon II Mar 7 Apr 19 May 31 Od I3 Dee 20
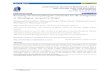
Figure I. Vertical distribution of males (m), females (0) and juveniles (illl) of nine dominant species at Belcarra Park (top three rows) and three dominant species at Iona Island (bottom row) over I year.
Some nematode species were lumped into a single species descriptor in the early samples
listed in Table 4. They are Sabatieria pukhra (including Sabatieria sp. females and juveniles), Paracyatholaimus pugettensis (includes P. vancouverensis) and Viscosia spp. (identified initially as oncholaimids). The vertical distribution of 12 of the most abundant species (nine from Belcarra Park and three from Iona Island) that show obvious changes in age structure over time, as expressed by the number of males, females and juveniles are shown in Figure I.
At Belcarra Park, large numbers of certain species were recorded only at certain times, e.g. Linhomoeus undulata, Desmolaimus zeelandicus and Eleutherolaimus stensbma (Table 4, Figure I). At Iona Island, S. pulchra occurred in larger numbers in the autumn months. Relatively high population densities were maintained over several months by Viscosik tumidula at Belcarra Park and by Paracanthonchus canadensis at Iona Island. There was a

Free-living nematodes at two Canadian Pacific beaches 225
significant difference in the variance of density over time of all species examined (Table 4) except V. carnleyensis, P. canadensis and Tripyloides gracilis.
There was a significant difference in the variance of the life stages of all species examined except P. pugettensis, P. vancouverensis, V. car¥.&, Neochromadora sp. and T. gracilis. Large numbers of juveniles were found during a restricted period of the year for Paralinhomoeus buculentus, L. undulatus and S. pulchra, while similar numbers of juveniles were found throughout several months for V. carnleyensis, P. pugettensis, P. vancouverensis, P. mutila, Cobbia urinator and P. canadensis.
None of the species was evenly distributed throughout the top 6 cm, and their vertical distribution remained fairly constant with the exception of Terschellingia longicaudata and Sabatieria sp. For example, during February and August most individuals of D. zeelandicus were found in the z-4-cm layer whilst they were dispersed more evenly throughout the top 6 cm for the remainder of the year. There was a significant difference in the variance of density at various depths for Areolaimus elegans. T. longicaudata and Theristus modicus.
A significant interaction between time and depth occurred for all species except E. stenosoma, Theristus sp., Neochromadora sp., P. canadensis and T. gracilis. A significant interaction between time and stage of development was recorded for Sabatieria sp. and S. pulchra but a significant interaction between depth and stage (but not between time, depth and stage of development) was seen for P. vancouverensis.
Discussion
The nematode faunas at Iona Island and Belcarra Park differ in terms of composition, numbers of individuals and number of species and this is probably partly attributable to the physicochemical nature of the substrate at the two sites.
The extreme variability noted in replicate samples at both sites is characteristic of the patchiness seen in littoral habitats. Sibert et al. (1977) reported standard deviations as large as 858 for a mean of 1943 nematodes from a mudflat in the Cowichan River estuary, British Columbia. Such patchy distribution is common among meiofaunal organisms and may often be due to the selective attraction of nematode species to scattered, dead and decaying organisms on a beach (Gerlach, 1978).
The large difference in nematode densities between the two sites and their association with grain size (Tables I and z) corroborates data from other studies. For example, densities of a.44 X 10~ m-* from an intertidal sandy beach (McIntyre & Murison, 1973), o-5 X IO’ m-’ from a fine sandflat (Ott, 1972) and 3.5 x 106 m--B from well sorted, medium size sediments (Platt, 1977a) have been reported. A study of the littoral benthic fauna at Iona Island referred to nematode densities on the north part of the causeway of 8.7 x 10~ m--2 (Anon., 1975) which is a similar order of magnitude to our records for the summer months (Table 2).
The large number of nematodes at Belcarra Park during the summer (Table 2) follows a pattern established for other locations (Tietjen, 1969; Skoolmun & Gerlach, 1971; Juario, 1975 ; Platt, 19773). Unfortunately, the summer samples are missing at Iona Island but the large number of nematodes occurring in October and December probably reflect the end of a summer peak population and also the considerably higher silt-clay composition of the October-December samples at this site (Table I). Such an environment is associated with large numbers of S. pulchra (Figure I) and Ward (1973) also found a greater abundance of Sabatieria sp. in areas of silty-clay sediments.
Though sandy habitats generally have higher diversity (McIntyre & Murison, 1973 ; 40
species), the high silt content of the fine grained Iona Island sediments coupled with their

226 J. Sharma &f J. M. Webster
relative impoverishment, as shown by their low organic carbon content, results in low species diversity. The larger number of species found at Belcarra Park could be attributed to the heterogeneity of the sediments which have more interstitial niches available for the greater food content as reflected in the higher organic carbon than at Iona Island (Table I).
The variation in abundance of nematode species follows that described by Juario (1975)
from the German Bight with both epigrowth and deposit feeders found at Belcarra Park during the peak nematode population in spring and summer. In contrast, Tietjen (1969) found a large number of epigrowth feeders during spring and summer and a large number of deposit feeders and omnivores during fall and winter. The presence of different feeding types in the same microhabitat at the same time reflects the ability of nematode species to partition the available food (Lee et al., 1977). The peak abundance of several species in this study was associated with the presence of a large number of juveniles. This is also the case in the studies of Skoolmun & Gerlach (1971), Lorenzen (1974) and Juario (1975).
Most nematodes occurred in the upper 4 cm and significantly fewer at 4-6 cm below the surface (Table 2), which agrees with other reports for comparable habitats (Wieser & Kanwisher, 1961; Tietjen, 1969; Platt, 1977b). Occasional reports refer to free-living nematodes occurring at greater depths especially where the sediment is well organized: e.g. 12 cm in the summer and up to 32 cm in the winter on an intertidal, sandy beach (McIntyre & Murison, 1973), 12-13 cm in a Georgia salt marsh (Teal & Wieser, 1966) and 9 cm on a sandy beach in Hong Kong (Wormald, 1976). Although we detected no pattern in the seasonal fluctuations of total numbers of nematodes at different depths, Platt (1977b)
found that the number of nematodes in the deeper sediments was greater in winter than in the summer. He attributed this variation to the presence of a deeper redox discontinuity layer in winter. Though fluctuations were observed in the density of individual species and their life stages the patchiness of the environment coupled with infrequent sampling failed to give any significant interaction between fluctuations in density over time of the life stages and the depth.
Certain species have been shown to be characteristic of a deeper sulfide layer (Ott, 1972). In the present study the distribution of species such as T. longicaudata, S. pulchra and P. mutila below 2 cm (Figure I) may suggest that they have a greater tolerance of the anoxic conditions found in the deeper sediments. The near absence of species such as Neochromadora appiana and P. caecus below 2 cm could be an indication of their intolerance of oxygen- deficient sediments.
Finally, the occurrence and distribution of certain nematode species in the littoral sediments on the Canadian Pacific coast parallels that occurring in similar habitats in the eastern (Skoolmun and Gerlach, 1971; Platt, 1977a) and western (Tietjen, 1969) Atlantic Ocean and on the oriental coast of the Pacific Ocean (Wormald, 1976). A worldwide distribution of many of these nematode species is, therefore, becoming apparent.
Acknowledgements
We are grateful to Bruce E. Hopper (Canada Department of Agriculture, Ottawa) for his confirmation of species identification, to Nit Smol and Magda Vincx (Instituut voor Dierkunde, Gent) for comments on the manuscript and Carlo Heip (Instituut voor Dierkunde) for suggestions and help with the analysis of the distribution of dominant species.
The research was partly supported by an operating grant (No. A4679) from the Natural Sciences and Engineering Research Council of Canada.

Free-living nematodes at two Canadian Pacific beaches 227
References
Anon. 1975 Environmental studies at Iona Island. Report No. 2, the Greater Vancouver Sewerage Drainage District.
Buchanan, J. B. x971 Measurement of the physical and chemical environment-sediments. In Methods for the Study of Marine Benthos (Holme, N. A. & McIntyre, A. D., eds). I.B.P. Handbook No. 16, Blackwell Scientific Publications, London.
Folk, R. L. & Ward, W. C. 1957 Brazos River bar, a study in the significance of grain size parameters. Journal of Sedimentary Petrology 27, S-27.
Gerlach, S. A. 1978 Food chain relationships in subtidal silty and marine sediments and the role of meiofauna in stimulating bacterial productivity. Oecologiu 33, 55-69.
Hopper, B. E. & Meyers, S. P. 1967 Population studies on benthic nematodes within a subtropical seagrass community. Marine Biology I, 85-96.
Juario, J. V. 1975 Nematode species composition and seasonal fluctuations of a sublittoral meiofauna community in the German Bight. Veriiffentlichungen des Institutsfiir Meeresforschung in Bremerhaven 15,283-3,37.
Lee, J. J., Tietjen, J. H., Mastropaolo, C. & Rubin, H. 1977 Food quality and the heterogenous spatial distribution of meiofauna. Helgol&nder Wissenschaften Meeresuntersuchungen 30, 272-282.
Lorenzen, S. 1974 Die nematoden Fauna der subtiloralin Region der Deutscher Bucht; inbesondire im Titan-Abwassergebiet bei Helgoland. Veriiffentlichungen des Instituts fiir Meeresforschung in Bremerhaven 2, I 25-130.
McIntyre, A. 1~. & Murison, D. J. 1973 The meiofauna of a flatfish nursery ground. Journal of the Marine Biological Association of the United Kingdom 53, 93-118.
Nelson, H., Webster, J. M. & Burr, A. H. 1971 A redescription of the nematode Oncholaimus vesicarius (Wieser, 1959) and observations on the pigment spots of this and of Oncholaimus skawensis Ditlevsen, 1921. Canadian Journal of Zoology 49, I 193-1 I 97.
Nelson, H., Hopper, B. E. & Webster, J. M. 1972 Enoplus anisospiculus, a new species of marine nematode from the Canadian Pacific Coast. CanadianJournal of Zoology 50, 1681-1684.
Ott, J. A. 1972 Determination of fauna boundaries of nematodes in an intertidal sand flat. Internationuir Revue der Gesamten Hydrobiologie und Hydrographie 57, 645-663.
Platt, H. 1977” Ecology of free-living nematodes from an intertidal sandflat in Strangford Lough, Northern Ireland. Estuarine and Coastal Marine Science 5, 685--693.
Platt, H. I977b Vertical and horizontal distribution of free-living marine nematodes from Strangford Lough, Northern Ireland. Cahiers de Biologie Marine 18, 261-273.
Platt, H. M. & Warwick, R. M. 1980 The significance of free-living nematodes to the littoral ecosystem. In The Shore Enoironment, Volume 2: Ecosystems (Price, J. H., Irvine, D. E. G. & Farnham, W. F., eds). Academic Press, London. pp. 772-759.
Seinhorst, J. W. 1959 A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 4, 67-69.
Sharma, J. & Vincx, M. 1982 Cyatholaimidae (Nematoda) from the Canadian Pacific Coast. Canadian Journal of Zoology 60, 271-280.
Sibert, J., Brown, T. J., Healey, M. C. & Kask, B. A. 1977 Detritus based food webs: exploitation by juvenile chum salmon (Onchorhynchus keta). Science 195, 645-650
Skoolmun, P. & Gerlach, S. A. 1971 Seasonal fluctuations of the intertidal nematode fauna in Weser estuary (German Bight). Ver+@ntlichungen des Znstituts fiir Meeresforschung in Bremerhaven 13, 119-138.
Teal, J. M. & Wieser, W. 1966 The distribution and ecology of nematodes in a Georgia salt marsh. Limnology and Oceanography II, 217-222.
Tietjen, J. H. 1969 The ecology of shallow water meiofauna in two New England estuaries. Oecologia 2,251-291.
Vincx, M., Sharma, J. & Smol, N. in press On the identity and nomenclature of ‘Paracanthonchus caecus (Bastian, I 85) ’ with a redefinition of the genus Paracanthonchus Micoletzky, I 924. Zoologica Scripte.
Ward, A. R. 1973 Studies on the sublittoral free-living nematodes of Liverpool Bay. I. The structure and distribution of the nematode population. Marine Biology 22, 53-66.
Wieser, W. 1952 Die Beziehungen zwischen mundhohlengestalt Ernarhungsweise und Vorkominen bei freilebenden marinen Nematoden. Arkiv fiir Zoologi 4, 439-484.
Wieser, W. & Kanwisher, J. 1961 Ecological and physiological studies on marine nematodes from a small salt marsh near Woods Hole, Massachusetts. Limnology and Oceanography 6, 262-270.
Wormald, A. P. 1976 Effects of a spill of marine diesel oil on the meiofauna at Picnic Bay, Hong Kong. Environmental Pollution II, I 17-130.