Embed Size (px)
Citation preview

Thalamocortical Connections of theParabelt Auditory Cortex in Macaque
Monkeys
T.A. HACKETT, I. STEPNIEWSKA, AND J.H. KAAS*
Department of Psychology, Vanderbilt University, Nashville, Tennessee, 37240
ABSTRACTThe auditory cortex of macaque monkeys contains a core of primary-like areas sur-
rounded by a narrow belt of associated fields that encompass much of the superior temporalplane in these animals. Adjacent to the lateral belt on the superior temporal gyrus is aparabelt region that contains at least two subdivisions (rostral and caudal). In a previousstudy (Hackett et al. [1998] J. Comp. Neurol. 394:475–495), we determined that the parabelthas topographic connections with the belt areas surrounding the core, but minimal connec-tions with the core itself. In this study, we describe the thalamocortical connections of theparabelt auditory cortex based on multiple injections of neuronal tracers into this region ineach of five macaque monkeys. Injections confined to the parabelt labeled large numbers ofneurons in the dorsal (MGd) and magnocellular (MGm) divisions of the medial geniculatecomplex (MGC), suprageniculate (Sg), limitans (Lim), and medial pulvinar (PM) nuclei. Onlywhen injections encroached on the lateral belt cortex were substantial numbers of labeledneurons found in the ventral (MGv) division of the MGC, consistent with the absence ofsignificant connections between the parabelt and core fields. The rostrocaudal topography ofthe parabelt region was maintained in the thalamocortical connections, supporting theparcellation of this region of cortex. The results suggest that the parabelt region represents athird level of auditory cortical processing, which is not influenced by direct inputs fromprimary cortical or subcortical auditory structures. J. Comp. Neurol. 400:271–286,1998. r 1998 Wiley-Liss, Inc.
Indexing terms: association cortex; thalamus; medial geniculate; pulvinar
Our studies of auditory cortex in monkeys, in conjunc-tion with those from other laboratories, have led us topropose three levels of auditory processing in the temporallobe (Hackett et al., 1998; Kaas and Hackett, 1998). At thefirst level, three tonotopically organized fields make up anauditory core. These fields include the traditional primaryfield, AI, a more rostral or rostrolateral field, R, and arostrotemporal field, RT. Each of these fields receivesauditory information from several nuclei of the medialgeniculate complex and distributes in an extremely localmanner to a narrow surrounding belt of approximatelyseven auditory areas, at least some of which also receiveparallel inputs from the medial geniculate complex. Allbelt areas project in turn to a wide adjoining band of cortexin the superior temporal gyrus that we have termed theparabelt. Whereas the parabelt may have functional subdi-visions, for the present purposes, it is characterized as acontinuous zone of temporal cortex with inputs from thebelt, but not from the core. The question we address hereis: what are the thalamic inputs to the parabelt? Althoughthis question can be answered in part by referring to the
results of previous studies of temporal lobe connections(e.g., Mesulam and Pandya, 1973; Trojanowski and Jacob-son, 1975; Burton and Jones, 1976; De Vito and Simmons,1976; Fitzpatrick and Imig, 1978; Pandya et al., 1994;Molinari et al., 1995), these studies related results todifferent concepts of cortical organization, rather than theframework outlined above. By placing injections of differ-ent distinguishable tracers in rows along the rostrocaudalextent of the parabelt region, we could determine whetherthe parabelt region, defined as auditory by inputs from thebelt, also receives auditory inputs from the thalamus. Wecould also determine whether the parabelt cortex receivesother significant thalamic inputs, such as from the medial
Grant sponsor: NINDS; Grant number: NS 16446; Grant sponsor:NIDCD Research Fellowship; Grant number: DC 00249.
*Correspondence to: Jon H. Kaas, Ph.D., Department of Psychology,Vanderbilt University, 301 Wilson Hall, 111 21st Avenue South, Nashville,TN 37240. E-mail: [email protected]
Received 18 February 1998; Revised 19 June 1998; Accepted 22 June1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 400:271–286 (1998)
r 1998 WILEY-LISS, INC.

pulvinar, which is known to project broadly to the temporalcortex (Clark and Northfield, 1937; Simpson, 1952; Whit-lock and Nauta, 1956; Locke, 1960; Siqueria, 1965; Tro-janowski and Jacobson, 1975; De Vito and Simmons, 1976;Pandya et al., 1994). The results would also indicatewhether thalamic structures project to parabelt cortex intopographic patterns, whether the complete parabelt re-gion shares inputs from any specific thalamic nuclei, andwhether parts of the parabelt region differ in thalamicinputs.
To facilitate the interpretation of results, it was impor-tant to identify thalamic nuclei effectively in the experimen-tal cases. Thus, we used several histological and histo-chemical procedures to help define thalamic nuclei. Inparticular, some sections were processed for calbindin andparvalbumin, in part because the distribution of theseproteins can be quite variable among different thalamicnuclei (Jones and Hendry, 1989; Molinari et al., 1995).Recently, nuclei in the medial geniculate complex havebeen shown to contain variable populations of parvalbumin-positive cells that relay to the middle layers of specificcortical fields, and generally complementary populationsof calbindin-positive cells that project more broadly tosuperficial layers of cortex (Hashikawa et al., 1990, 1991,1995; Molinari et al., 1995). Thus, it is useful to examinethe connections of types of cells within nuclei, as well asthe collective connections of each nucleus. Some of theseresults have been briefly presented elsewhere (Hackett etal., 1996).
MATERIALS AND METHODS
In five macaque monkeys (three M. mulatta, threeM. nemestrina), microinjections of four to seven anatomi-cal tracers were made at equal intervals along the dorsalsuperior temporal gyrus within 3 mm of the lateral sulcusin the left hemisphere. The procedures for making tracerinjections and evaluating their locations have been de-scribed in detail in our previous paper on cortical connec-tions based on the same cases (Hackett et al., 1998). Thetracers used were horseradish peroxidase conjugated towheat germ agglutinin (WGA-HRP) and the following
fluorescent tracers: Fast Blue (FB), Diamidino Yellow(DY), Fluoro-Ruby (FR), Fluoro-Gold (FG), fluorescein(FS), rhodamine green beads (GB), and rhodamine redbeads (RB). Because the tracers vary in sensitivity, theamounts and solution concentrations were varied accord-ingly (typically, 0.01–0.05 ml at 2% for WGA-HRP; 0.1–0.3µl at 2–10% for fluorescent tracers). The postinjectionsurvival period was 7 days when WGA-HRP was used(three cases) and 12 days when only fluorescent tracerswere injected (two cases). At the end of the survival period,a lethal dose of pentobarbital was administered. Justbefore cardiac arrest, the animal was perfused through theheart with warm saline followed by cold 2% paraformalde-hyde dissolved in phosphate buffer. Immediately afterperfusion the brains were removed and photographed. Thethalamus was separated from the brain, soaked in 30%sucrose solution overnight, and then cut in the frontalplane at 40 µm on a freezing microtome. Alternate series ofsections were processed for: 1) fluorescent microscopy;2) WGA-HRP using a low artifact tetramethyl benzidine(TMB) procedure (Gibson et al., 1984); 3) cytochromeoxidase (Wong-Riley, 1979); 4) acetylcholinesterase (Gen-eser-Jensen and Blackstad, 1971); 5) myelin (Gallyas,1979); 6) parvalbumin immunohistochemistry; 7) calbin-din immunohistochemistry; or 8) staining for Nissl sub-stance with cresyl violet.
Cells labeled with fluorescent tracers were plotted underultraviolet illumination with a Leitz microscope coupled toan X-Y plotter (Bioquant System, R & M Biometrics,Nashville, TN). Cells labeled with WGA-HRP were plottedunder darkfield illumination with a drawing tube. Photo-graphs and drawings of each section were made notingarchitectonic boundaries, the location of blood vessels, andthe distribution of labeled cells. A composite drawing wasmade from adjacent sections processed for label, cyto-chrome oxidase, acetylcholinesterase, parvalbumin, calbi-ndin, and Nissl by aligning common architectonic featuresand blood vessels. Composite images were composed of thereconstructions by using Canvas 3.5.4 software installedon a Power Macintosh 7200/75 computer. The final draw-ings were analyzed to reveal: 1) the individual connectionpatterns of each tracer injection; 2) the connection pat-
Abbreviations
AD anterodorsal medial geniculate nucleusAI auditory area I (core)AS arcuate sulcusBrCs brachium of the superior colliculusCiS circular sulcusCP caudal parabelt area (CP)CS central sulcusDY Diamidino YellowFB Fast BlueFR Fluoro-RubyGB rhodamine green beadsHb habenular nucleusHbL lateral habenular nucleusHbM medial habenular nucleusHRP horseradish peroxidaseILS inferior limiting sulcusIPS intraparietal sulcusLim limitans nucleusLS lateral sulcusLuS lunate sulcusM magnocellular medial geniculate nucleusMD mediodorsal nucleusMGC medial geniculate complexMGd dorsal medial geniculate complex
MGad anterodorsal medial geniculate nucleusMGm magnocellular medial geniculate nucleusMGpd posterodorsal medial geniculate nucleusMGv ventral medial geniculate nucleusPA anterior pulvinar nucleusPD posterodorsal medial geniculate nucleusPI inferior pulvinar nucleusPL lateral pulvinar nucleusPM medial pulvinar nucleusPP peripeduncular nucleusPo posterior nucleusPS principal sulcusR rostral area (core)RB rhodamine red beadsRP rostral parabelt area (parabelt)RT rostrotemporal area (core)Sg suprageniculate nucleusSTG superior temporal gyrusSTS superior temporal sulcusTpt temporal parietotemporal areaWGA wheat germ agglutininV ventral medial geniculate nucleusZ zone of transition
272 T.A. HACKETT ET AL.

terns of injections at similar or dissimilar locations;3) consistency across cases; and 4) aberrant or unusualpatterns. Photographic images were acquired by using aLeaf Lumina digital scanning camera mounted on a NikonE800 microscope. The digitized images were adjusted forbrightness and contrast, cropped, and pasted in the framewhere text was added with Adobe Photoshop 4.0 software.Except for contrast adjustment and cropping, the imageswere not altered in any way.
RESULTS
Thalamic architecture and subdivisions
The architecture and subdivisions of the posterior thala-mus were studied by comparing adjacent tissue sectionsstained for Nissl, acetylcholinesterase, cytochrome oxi-dase, calbindin, and parvalbumin. Tissue from all fourcases were used in the analysis and representative sec-tions were chosen for illustration. Figure 1 contains aseries of frontal sections through the posterior thalamuswhere labeled cells were found. The panels in the top row(Fig. 1A–E) are low magnification images of frontal sec-tions showing the medial pulvinar, suprageniculate/limitans, and medial geniculate complex. The medialgeniculate complex is shown at higher magnification in thebottom panels (Fig. 1F– J).
Medial geniculate complex. Within the medial genicu-late complex we identified several major subdivisions:ventral, magnocellular, posterodorsal, and anterodorsal.In Nissl-stained sections the ventral nucleus of the medialgeniculate (MGv) appeared as a tightly packed grouping ofsmall and medium-sized cells occupying the ventral andlateral portion of the complex (Fig. 1F). In some sections,such as those shown in Figure 1, cells within the MGv weresegregated into smaller groups, causing it to appearlobulated. The MGv stained darkly for acetylcholinester-ase, cytochrome oxidase, and parvalbumin (designated asV in Fig. 1G,H,J). Immunostaining for calbindin wasabsent (Fig. 1I).
The adjacent posterodorsal nucleus (MGpd) was popu-lated by small- and medium-sized cells spaced somewhatmore loosely than in the MGv (designated as PD in Fig.1F). The MGpd was moderately stained for acetylcholines-terase and cytochrome oxidase and moderately dark forparvalbumin. Reactivity for calbindin was generally weak,except at the extreme posterior pole of the MGpd whereimmunoreactivity was intense for calbindin and weak forparvalbumin. The anterodorsal nucleus (MGad, not shown)was populated by a relatively more heterogeneous cellpopulation characterized by numerous larger cells thatstained darkly for Nissl. The MGad stained moderatelydark for acetylcholinesterase, cytochrome oxidase, andparvalbumin. Immunoreactivity for calbindin was evenweaker than in the MGpd, although a small number ofcalbindin-positive cells were scattered throughout bothsubdivisions, becoming more numerous dorsomedially nearthe border with the ventrolateral Sg.
The magnocellular nucleus (MGm) was easily identifiedin Nissl preparations as an elongated area interspersedwith darkly stained large cells. The MGm occupied themedial border of the complex and extended from the Sg tothe MGv. The distribution of large cells in the MGm wastypically patchy in posterior sections where it was brokenup by groups of smaller cells and fibers of the brachium ofthe inferior colliculus (Fig. 1F). In anterior sections, where
the full dorsoventral extent of the MGm was visible, cellspacing in the MGm was more uniform. The largest cellswere more numerous dorsally. Staining for acetylcholines-terase and cytochrome oxidase was dark and patchy,reflecting the distribution of the larger cells. Immunostain-ing for calbindin was light to moderate among the smallercells, and many of the large cells stained positively,particularly in the dorsal half of the nucleus (Fig. 1I).Many of the largest cells throughout the MGm werestrongly immunoreactive for parvalbumin, as were a few ofthe smaller cells.
Positioned between the three primary nuclei of themedial geniculate complex, a poorly defined fourth regionwas inconsistently identified. Within this area, referred toelsewhere as the internal nucleus (e.g., DeVenecia et al.,1995) and the zone of transition (Molinari et al., 1995), thearchitectural properties of the surrounding three nucleioverlapped, blurring the borders between nuclei. In ourmaterial stained using the Nissl method, small, medium,and large cells were found within this zone (Z). Histochemi-cal staining for acetylcholinesterase and cytochrome oxi-dase was moderately dark to dark, and somewhat patchy.Immunostaining in the transitional zone was weak forcalbindin, but slightly stronger than in the MGv due to thepresence of scattered calbindin-positive cells. Immunoreac-tivity for parvalbumin was moderate, but clearly not asintense as in the MGv and MGpd (Fig. 1I,J). The borderbetween the transition zone and the ventral and magnocel-lular nuclei was generally more distinct than the borderwith the dorsal nucleus; therefore, this border was repre-sented as a dashed line in reconstructions depicting thethalamocortical connections (see Figs. 4–7).
Suprageniculate and limitans nuclei. The posteriorSg nucleus was populated almost entirely by closely spacedlarge cells posteriorly that were slightly larger, but moregenerously spaced anteriorly. In Figure 1A, the Sg appearsas patches of large cells grouped fairly close togetherdorsomedial to the medial geniculate complex. Neuropiland cell staining was dark for acetylcholinesterase andcytochrome oxidase. The number of parvalbumin- andcalbindin-positive cells in the Sg appeared to be about thesame, often occurring in patches of intense neuropil andcell staining (e.g., Fig. 1I).
In Nissl-stained sections the limitans nucleus appearedas an elongated strip of large and medium-sized cellsextending dorsomedially from the Sg along the ventrome-dial edge of the PM (Fig. 1A). Cells were tightly groupedalong the medial border of the limitans, appearing as adark band in acetylcholinesterase (Fig. 1B). Otherwise,the limitans neuropil stained moderately for cytochromeoxidase, parvalbumin, and calbindin. Like the Sg, the Limwas primarily populated by closely spaced large cells thatstained darkly for cytochrome oxidase, parvalbumin, andcalbindin.
Medial pulvinar. The cytoarchitecture revealed byNissl staining (Fig. 1A) showed that the medial pulvinar(PM) was primarily populated by medium-sized cells spacedwidely apart in a fairly uniform distribution. Cell packingwas denser along the dorsal and medial borders of the PM.A small number of much larger cells was found primarilyin the central and lateral PM. Neuropil and cell stainingfor acetylcholinesterase and parvalbumin in the PM wasvery weak dorsally and medially, where cell packingappeared greater in Nissl, becoming much darker ven-trally and laterally. In most sections, relatively few cells
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 273

Fig. 1. A–J: Coronal sections of the macaque monkey thalamus (case 96–1) at the level ofthe medial geniculate complex and medial pulvinar. Top row, low magnification images ofadjacent sections processed for Nissl (A), acetycholinesterase (AChE; B), cytochrome oxidase(CO; C), calbindin (D), and parvalbumin (E). Bottom row, higher magnification images of the
sections in the top row showing the medial geniculate complex and suprageniculate nucleus.Arrows denote location of the same blood vessels. Subdivisions are indicated in the Nissl-stained sections (A and F). For abbreviations, see list. Scale bars 5 2 mm in top row, 1 mm inbottom row.

were parvalbumin positive in the dorsal and medial por-tions of the PM. In contrast, numerous parvalbumin-positive cells were found in the ventral and central/lateralPM and throughout the PL (Fig. 1E). Staining for cyto-chrome oxidase was moderate and fairly uniform through-out the PM, but darker in regions that were also moredarkly stained in acetylcholinesterase. Calbindin immuno-reactivity was intense in the medial and central PM,becoming much lighter laterally. Staining for calbindinwas also very dense along the dorsal border of the PMwhere reactivity for parvalbumin, cytochrome oxidase,and acetylcholinesterase was the weakest. In Nissl-stained sections, cells were densely packed along thisborder in a band that ran from the PL to the medial edge ofthe PM. In the adjacent lateral pulvinar (PL) staining wasgenerally complementary to that found in the PM, appear-ing darker overall in acetylcholinesterase, cytochromeoxidase, and parvalbumin, but much lighter in calbindin.Although most of the cells in the PM were medium-sized, asmall number of much larger cells with long slenderprocesses stained darkly for calbindin (Fig. 2). These cellswere more numerous in the central and lateral PM, andoccasionally found in the medial third of the PM. In Figure2A a large calbindin-positive cell from the central PM isshown, surrounded by more weakly stained medium-sizedcells. This cell was typical of the large cells found in thelateral and central PM, which were not labeled by ourparabelt injections. In Figure 2B, two large cells from themedial PM are pictured. Note that these cells are slightlysmaller than those from the central/lateral PM. The largecells in the medial PM were frequently labeled by injec-tions of the parabelt.
Thalamic connections of the parabeltauditory cortex
The thalamocortical connections of the parabelt audi-tory cortex were determined by injecting retrograde trac-ers into the superior temporal gyrus near the ventral lip ofthe lateral sulcus (Fig. 3). The parabelt cortex occupies thesurface of the superior temporal gyrus lateral to theauditory core and belt regions on the superior temporalplane. Injections were judged as extending into the beltwhen significant numbers of neurons were labeled insidethe core and MGv, clearly within the parabelt when largenumbers of neurons were labeled in the belt and not thecore or MGv, and outside the parabelt when few or noneurons were labeled in the belt or parabelt, as deter-mined in this and previous studies (Hackett et al., 1998;Morel et al., 1993). The locations of labeled cells in thethalamus were plotted on drawings of adjacent sectionsshowing architectonic boundaries and subdivisions. Repre-sentative sections from four cases were included in Figures4–7. The strongest projections to the parabelt arose fromthe dorsal (MGad and MGpd) and magnocellular (MGm)nuclei of the medial geniculate complex, suprageniculate(Sg), limitans (Lim), and medial pulvinar (PM). Minorconnections were found with the peripeduncular nucleus(PP), and inconsistently with the posterior nucleus (Po)and lateral pulvinar (PL).
Medial geniculate complex
Ventral nucleus. Cell labeling in the MGv was nearlyabsent when tracer injections were limited to the parabeltand did not involve the cortex of the lateral belt on thesuperior temporal plane. In case 95–52 the FB (opentriangles), WGA-HRP (filled squares), and DY (open
squares) injections involved both the parabelt and thelateral belt adjacent to AI (Fig. 7). In the thalamus, theseinjections resulted in substantial cell labeling in the MGv(Fig. 7, sections 355, 361, 367). In the same case labeledcells associated with these three injections were also founddorsal to the MGd in a region corresponding in location tothe posterior nucleus (Po).
Dorsal nuclei. Connections of the dorsal medial genicu-late nuclei (MGpd and MGad) exhibited a rostrocaudaltopography with the parabelt, that is, rostral injections ofthe STG tended to label cells posteriorly and caudalinjections labeled cells anteriorly in the MGd. In case 96–1(Fig. 6), restriction of the parabelt injections to the rostralhalf of the STG resulted in topographic labeling in theposterior MGd, corresponding to MGpd, whereas the MGadwas virtually devoid of labeled cells. In addition to therostrocaudal topography, the distribution of labeled cells
Fig. 2. Large and medium-sized calbindin-positive cells in themedial pulvinar (PM). A: Central PM. B: Medial PM. The large cells inthe central PM were not labeled by parabelt injections and were largerthan those in the medial PM, which were sometimes labeled byparabelt injections. Scale bar 5 25 µm.
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 275

was somewhat patchy in the MGd, but less so than in thePM. For example, in case 94–64 (Fig. 4, sections 111 and121) labeled cells were distributed in patches that wereoften segregated from cells labeled by other injections. Asimilar pattern was found in case 96–1 (Fig. 6, section 97)and case 95–52 (Fig. 7, sections 361 and 367). In contrast,the distribution was more overlapping in other sections(e.g., case 95–10, Fig. 5, section 199; case 96–1, Fig. 6,section 109). Furthermore, as in case 94–64 section 111,labeled cells associated with the same injection weresometimes segregated into groups. Thus, whereas therewas a strong tendency for inputs to the parabelt from theMGd to be arranged rostrocaudally, evidence for topogra-phy in any other dimension (i.e., mediolateral) was equivo-cal.
Magnocellular nucleus. Cell labeling in the MGm wasgenerally strong throughout the subdivision, particularlyin the caudal two-thirds of the MGm. In the rostral third ofthe MGm, fewer numbers of labeled cells were found, andthese were usually associated with the most caudal injec-tion sites. Injections in the parabelt often labeled separatepatches of cells in the dorsal and ventral portions of theMGm. The patches of labeled cells sometimes reflected therostrocaudal sequence of injections in the cortex, but moreoften the patches contained labeled cells associated withthe same injection, indicating that separate zones in theMGm project in parallel to the same location in cortex. Therostrocaudal topography of connections found between theMGd and parabelt (i.e., caudal MGd projected to rostralparabelt, and rostral MGd projected to caudal parabelt)was partially maintained in the MGm, but the distribu-tions of labeled cells in the MGm overlapped more, as thefollowing examples illustrate. In case 94–64 (Fig. 4), cellslabeled by rostral and caudal parabelt injections over-lapped in sections 111 and 121 with little evidence ofsegregation. In case 96–1, cells labeled by the most rostraland most caudal injections overlapped in the most poste-
rior section analyzed (Fig. 6, section 97). A similar trendwas found in case 95–10 (Fig. 5). In case 95–52, thedistribution of labeled cells appeared to be slightly morepatchy than in the other cases, and even cells labeled byclosely spaced injections were segregated (Fig. 7, sections343 and 355). Note that the three most caudal injections inthis case also involved the cortex of the lateral belt, but it isunclear whether this contributed significantly to thegreater segregation. In case 94–64, only one labeled cellwas found in the MGm that was associated with the FastBlue injection (Fig. 4, open squares). This injection, madein area Tpt (Pandya and Sanides, 1973) caudal to theparabelt, was the most caudal injection made in the fivecases analyzed for this study and labeled many cells in thePM, PL, and Sg/Lim. The distribution of cells in the cortexlabeled by this injection was also unique, virtually circum-scribing the caudal belt areas (Hackett et al., 1998).
It was common to find cells in the MGm that weredouble-labeled by closely spaced injections. Figure 8 showsa cell in the MGm (case 96–1) double labeled by injectionsof FR (Fig. 8A) and DY (Fig. 8B). Just to the left of thedouble-labeled cell is a cell labeled only by FR (Fig. 8A).The injections were placed about 4 mm apart in theparabelt lateral to R (Fig. 6, open symbols). Note also thatthe double-labeled cell was not one of the largest cells (seethe larger cell positioned above the labeled cell in thephotograph), which are prominent in the MGm.
Suprageniculate and limitans nuclei. The topo-graphical relationship between the MGC and the parabeltappeared to be maintained in the Sg and Lim. In case94–64 (Fig. 4), numerous cells labeled by the rostral DYinjection (filled circles) were found in posterior sectionscontaining the Sg/Lim (sections 111, 121, and 131). Anteri-orly, a mixed population of cells labeled by rostral andcaudal injection sites was found, but the number of cellslabeled by the rostral injection was reduced (e.g., section141). A very similar trend was found in case 95–10 (Fig. 5),in which only labeled cells associated with the most caudalinjections were found in the anterior Sg/Lim. In case 96–1(Fig. 6) very few cells in the Lim were labeled by the threemost rostral injection sites (filled symbols), and apparentlynone in the Sg. This contrasts sharply with the relativelydense cell labeling in the PM and MGC associated withthese injections. In contrast, the number of labeled cells inthe Sg/Lim was much greater for the other, more caudal,injections in this case (open symbols). These injectionswere positioned lateral to the rostral subdivision (R) of theauditory core, whereas the three rostral injections wereplaced beyond the rostral border of R. In case 95–52(Fig. 7), the data were less complete for the Sg/Lim,making it difficult to substantiate the trends observed inthe other two cases. In some sections we observed thatlabeled cells in the Lim were concentrated in strips thatran along the lateral or medial edge of the nucleus (e.g.,Fig. 4, section 121; Fig. 5, section 199; Fig. 6, section 127).In a few sections labeled cells were distributed in twostrips, one lateral and the other medial (e.g., Fig. 4, section131; Fig. 6, section 115). In such sections the cells within a‘‘row’’ were positioned very close together. This arrange-ment of labeled cells may be related to the organization offibers passing through the nucleus.
Medial pulvinar. Labeled cells were generally concen-trated in the medial half of the PM. In posterior sections,labeled cells from only the most rostral parabelt injectionsoccupied the extreme ventromedial portion of the medial
Fig. 3. Dorsorostral view of the left macaque superior temporalregion showing the major regions of the auditory cortex (core, medial/lateral belt, parabelt) and locations of injection sites for each case. Theoverlying parietal cortex was removed to reveal the superior temporalplane and circular sulcus. The cortical surface was partially flattenedgraphically to enable visualization of areas that are hidden in sulci.For abbreviations, see list.
276 T.A. HACKETT ET AL.

Fig. 4. Reconstructions of coronal sections of the thalamus (case94–64) showing distribution of labeled cells after tracer injections intothe parabelt cortex. Inset: Location of injection sites. There was notransport to the thalamus from the red beads and green beads
injections. Open squares, Fast Blue; filled squares, Fluoro-Ruby; opentriangles, WGA-HRP; filled triangles, rhodamine green beads; opencircles, rhodamine red beads; filled circles, Diamidino Yellow. Forabbreviations, see list. Scale bar 5 2 mm.
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 277

Fig. 5. Reconstructions of coronal sections of the thalamus (case 95–10) showing distribution oflabeled cells after tracer injections into the parabelt cortex. Inset: Location of injection sites. Opensquares, Diamidino Yellow; open triangles, WGA-HRP; filled squares, rhodamine green beads; filledcircles, Fast Blue. For abbreviations, see list. Scale bar 5 2 mm.

Fig. 6. Reconstructions of coronal sections of the thalamus (case 96–1) showing distribution of labeledcells after tracer injections into the parabelt cortex. Inset: Location of injection sites. Open squares,Diamidino Yellow; open triangles, Fluoro-Ruby; filled squares, rhodamine green beads; filled triangles,rhodamine red beads; filled circles, Fast Blue. For abbreviations, see list. Scale bar 5 2 mm.
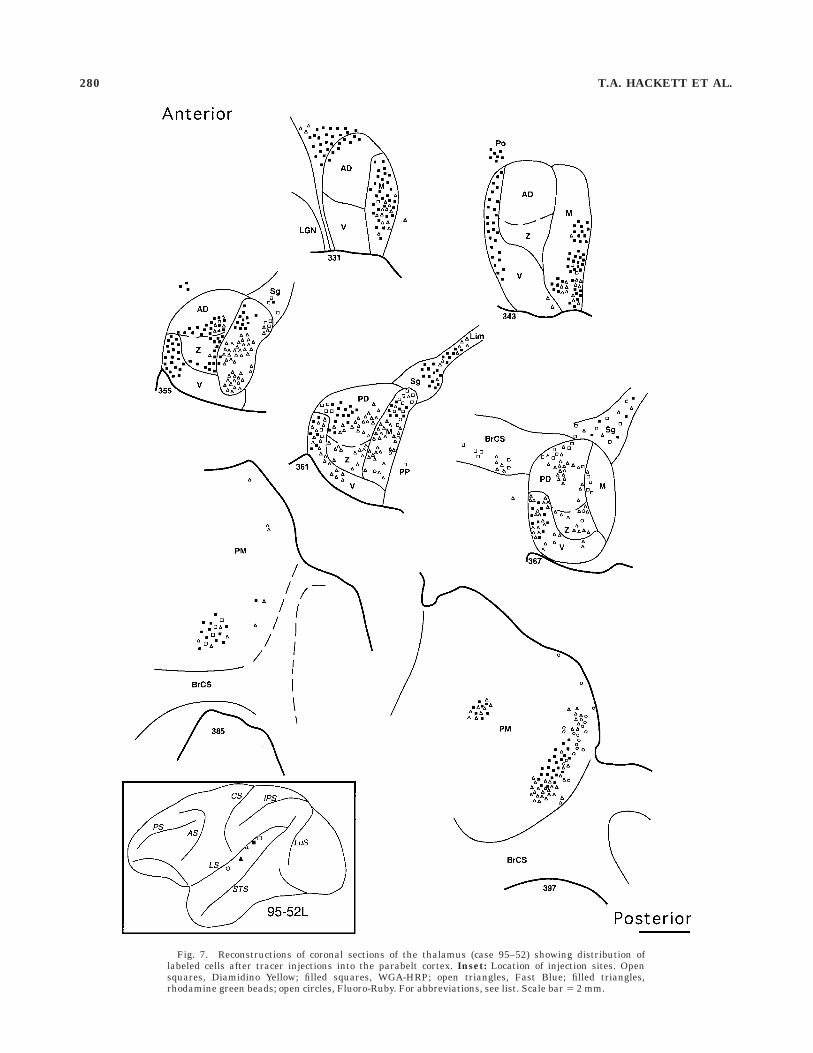
Fig. 7. Reconstructions of coronal sections of the thalamus (case 95–52) showing distribution oflabeled cells after tracer injections into the parabelt cortex. Inset: Location of injection sites. Opensquares, Diamidino Yellow; filled squares, WGA-HRP; open triangles, Fast Blue; filled triangles,rhodamine green beads; open circles, Fluoro-Ruby. For abbreviations, see list. Scale bar 5 2 mm.
280 T.A. HACKETT ET AL.

pulvinar bordering the limitans nucleus (Figs.4–7). Inmore anterior sections the distribution of labeled cellsspread dorsally and laterally, involving cells labeled by themore caudal parabelt injections. Labeled cells associatedwith widely spaced injection sites along the rostrocaudalaxis of the parabelt tended to remain segregated in themedial pulvinar, whereas closely spaced injection sitesresulted in overlapping distributions. Typically, cells la-beled by caudal injection sites occupied positions dorsaland lateral to cells labeled by rostral injections. In case96–1 (Fig. 6) labeled cells associated with the most rostralinjection (FB, filled circles) were clearly segregated fromcells associated with the three most caudal injections inPM. The DY (open squares) and FR (open triangles)injection sites, which were particularly close together,yielded very similar distributions of labeled cells, some ofwhich were labeled by both tracers (i.e., double-labeled). Incontrast, double-labeled cells associated with more widelyspaced injection sites were not observed in PM. Thus,segregated groups of neurons in the medial pulvinarprojected topographically to distinct locations along therostrocaudal axis of the parabelt auditory cortex, andcollateral branches from the axons of some cells withineach group projected to closely spaced sites in the cortex.
In one case, 94–64, patches of labeled cells extendedlaterally well into PL (Fig. 4). Typically, however, celllabeling did not extend laterally beyond the central PM.The overlapping distribution of cells often observed in thecentral PM tended to be associated with injections in thecaudal parabelt, whereas rostral parabelt injections la-beled cells nearer the medial and dorsal edge of the PM. Inmany sections, large scattered cells labeled by the mostrostral injection sites were found along the extreme dorso-medial edge of PM, coextensive with the narrow band ofintense calbindin immunoreactivity described above (e.g.,Fig. 6, sections 115, 127, 139; Fig. 7, section 397). Cellslabeled by more caudal injections occupied relatively ven-tral and lateral positions in medial PM.
In summary, the thalamocortical connections of theparabelt auditory cortex included the dorsal and magnocel-lular nuclei of the medial geniculate complex (i.e., MGd,MGm), suprageniculate, limitans, and medial pulvinarnuclei. The rostral parabelt and STG received projectionspredominantly from the caudal medial geniculate, whereasthe caudal parabelt and STG were preferentially con-nected with more rostral parts of the complex. The topo-graphic relationship was the least evident in the MGmwhere overlapping distributions of labeled cells and double-labeled cells were often found. Projections from the ventralnucleus (MGv) were only observed when the cortex of thelateral belt was involved in the injection; thus, the MGvdoes not appear to be significantly connected with theparabelt. The suprageniculate and limitans nuclei pro-jected widely to the parabelt and STG in a manner thatmaintained the rostrocaudal topography of the parabelt.The medial half of the medial pulvinar (PM) projectedstrongly to the parabelt. The rostrocaudal topography ofthe parabelt was maintained in the medial PM, with moreoverlap in the center of the nucleus. The lateral PM and PLhad limited connections with the parabelt auditory cortex.
DISCUSSION
In previous papers (Hackett et al., 1998; Kaas andHackett, 1998), we proposed that the auditory cortex inmacaque monkeys includes three major regions occupyingmuch of the superior temporal plane and dorsal superiortemporal gyrus. The koniocortical core region, which hasthree subdivisions (i.e., AI, R, RT), and the surroundingbelt cortex, which has as many as eight subdivisions (i.e.,C, CM, CL, ML, AL, RTL, RTM, RM), are largely buriedwithin the lateral sulcus on the superior temporal plane(Fig. 3). The core and belt regions are topographicallyinterconnected by dense connections between adjacentsubdivisions and weaker connections between nonadja-cent areas. The parabelt region is on the exposed surface ofthe dorsal superior temporal gyrus defined by topographicprojections that arise primarily from the belt of cortexsurrounding the primary core region, with little input fromthe core itself.
In the present study the thalamocortical connections ofthe parabelt region were examined by placing injections ofseveral retrograde tracers into the dorsal half of the STGwithin 3–5 mm of the lateral sulcus. Projections to theparabelt region from the thalamus originated principallyfrom the medial geniculate complex, suprageniculate, limi-tans, and medial pulvinar nuclei (Fig. 9). A topographicpattern of connections was reflected in these projectionssuch that rostral parabelt injections labeled more caudal
Fig. 8. Double-labeled cell in the magnocellular nucleus of themedial geniculate complex (MGm). A: Retrograde labeling of the somaand dendrites by Fluoro-Ruby. B: Retrograde labeling of the nucleusby Diamidino Yellow. Note that the double-labeled cell was not one ofthe largest cells in the MGm, one of which is positioned just above thedouble-labeled cell. Scale bar 5 25 µm.
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 281

portions of these thalamic nuclei. The connections of eachof these nuclei are reviewed below, particularly as theyrelate to the results of the present study and to theorganization of the central auditory system in primates.
Medial geniculate complex
Ventral nucleus. In primates the MGv is principallyconnected with the primary (core) areas of the auditorycortex (AI, R, RT), and has weaker, or more diffuse,connections with the surrounding auditory belt (Walker,1937; Mesulam and Pandya, 1973; Pandya and Sanides,1973; Forbes and Moskowitz, 1974; Burton and Jones,1976; Fitzpatrick and Imig, 1978; Oliver and Hall, 1978b;Calford and Aitkin, 1983; Aitkin et al., 1988; Luethke etal., 1989; Morel and Kaas, 1992; Morel et al., 1993; Pandyaet al., 1994; Hashikawa et al., 1995; Molinari et al., 1995).In our material, input to the auditory parabelt cortex fromthe medial geniculate was limited to the MGd and MGm.Projections from the MGv were observed only when thecortex of the lateral belt was involved in the tracerinjection. The involvement of the belt cortex by the injec-tion was associated with the appearance of labeled cells inthe auditory core, which is unlabeled after injectionsrestricted to the parabelt (Hackett et al., 1998). Althoughthe thalamocortical connections of the parabelt regionhave not been systematically studied previously, tracerinjections in the superior temporal gyrus have not re-vealed significant connections with the MGv (Pandya etal., 1994). Only when these injections appeared to en-croach on the lateral belt or core were labeled cells found inthe MGv (Fitzpatrick and Imig, 1978; Morel and Kaas,1992; Morel et al., 1993; Molinari et al., 1995; Kosmal etal., 1997). Likewise, when lesions or injections of thethalamus are restricted to the MGv, degenerated termi-nals or labeled cells are limited to the areas in and aroundthe core on the superior temporal plane (Pandya andSanides, 1973; Mesulam and Pandya, 1973; Burton andJones, 1976; Hashikawa et al., 1995). In contrast, theparabelt region receives little or no direct input from thisnucleus. Two previous studies suggested that the thalamo-cortical connections of the MGv are restricted to the core(Burton and Jones, 1976; Molinari et al., 1995). A numberof other studies, including the present one, suggested thatthe MGv has strong connections with the core and signifi-
cant, though weaker, connections with areas in the sur-rounding belt region (Luethke et al., 1989; Morel andKaas, 1992; Morel et al., 1993; Pandya et al., 1994; Kosmalet al., 1997). The reasons for this discrepancy are unclear,but they may be related to differences in methodology orcriteria in determining the borders between areas in thecortex and thalamus. In any event, the lack of MGv inputsto the parabelt region, together with the paucity of connec-tions between the core and parabelt, indicates that audi-tory processing in the parabelt region is not directlyaffected by inputs from the primary lemniscal auditorypathways.
Magnocellular nucleus. The cortical connections ofthe MGm have not been well studied in primates, but theavailable data indicate that the MGm of primates hasreciprocal cortical connections with all three subdivisionsof the core, as well much of the lateral belt region(Mesulam and Pandya, 1973; Burton and Jones, 1976;Fitzpatrick and Imig, 1978; Aitkin et al., 1988; Luethke etal., 1989; Morel and Kaas, 1992; Morel et al., 1993; Pandyaet al., 1994; Molinari et al., 1995; Kosmal et al., 1997).There is also some evidence that the MGm is preferentiallyconnected with the medial belt areas (Mesulam and Pan-dya, 1973; Burton and Jones, 1976; Pandya et al., 1994),but the data are limited.
In our material labeled cells were found in the MGmafter all parabelt injections. In these projections a rostro-caudal pattern of connections between the MGm and theparabelt cortex was apparent, but was less robust com-pared with the other nuclei of the MGC. For example,rostral parabelt injections tended to label the caudal MGmand vice versa, but distributions of labeled cells from therostral and caudal parabelt sometimes overlapped inpatches, usually midway between the rostral and caudalthirds of the nucleus. Conversely, even labeled cells associ-ated with adjacent injections were segregated in patches,usually in the dorsoventral dimension. Thus, the topogra-phy of the connection patterns could not be explained byapplication of a simple rule, and may reflect diffuseprojections of the heterogeneous population of cells in theMGm (Jones and Hendry, 1989; Hashikawa et al., 1990,1991, 1995; Molinari et al., 1995). Unfortunately, theextent to which cell labeling in the MGm reflected theconnections of the parvalbumin- and calbindin-positivepopulations in our material is unknown. Parvalbumin-positive cells are thought to project to the middle layers ofspecific cortical fields, whereas calbindin-positive cellsproject more broadly to the superficial cortical layers(Hashikawa et al., 1990, 1991, 1995; Molinari et al., 1995).Based on the sizes and depths of our injections, it is likelythat both populations of cells were labeled in the MGm,possibly accounting for the complex pattern of connections.Clearly, it will be important in future studies to determinethe laminar projection patterns of the parvalbumin- andcalbindin-positive cells in the MGm. Nonetheless, thepresent results indicate that the MGm is strongly con-nected with the auditory parabelt cortex of the STG. Theoverall topographic patterns of these connections is consis-tent with previous observations that the rostral core andbelt regions of auditory cortex are connected with thecaudal MGm, whereas the caudal auditory cortex is con-nected with the rostral MGm (e.g., Luethke et al., 1989;Morel and Kaas, 1992; Pandya et al., 1994). Based onreported connections with tonotopic areas of the core, adorsal-to-ventral organization of high to low frequencies in
Fig. 9. Schematic summarizing the major thalamic connections ofthe auditory cortex. Top row, major auditory cortical regions: core, belt,parabelt. Bottom row, thalamic nuclei, which project to the auditorycortex. For abbreviations, see list.
282 T.A. HACKETT ET AL.

the MGm has also been observed in New World monkeys(Luethke et al., 1989; Morel and Kaas, 1992); however,more variability in the connection patterns has beenobserved in studies involving macaque monkeys (Morel etal., 1993; Molinari et al., 1995).
An additional observation gleaned from the presentstudy was that numerous labeled cells were found in theMGm after all parabelt injections, but not after a singleinjection in Tpt (open squares, case 94–64), which labeledcells in the Sg/Lim and pulvinar. Several other studies(Pandya and Sanides, 1973; Mesulam and Pandya, 1973;DeVito and Simmons, 1976; Pandya et al., 1994) also foundno significant connections between the MGm and the gyralportion of Tpt. For example, after a large isotope injectionin Tpt, possibly limited to the exposed surface of the STG,labeled terminals were found in the pulvinar, supragenicu-late, and MGd, but not the MGm or MGv (Pandya et al.,1994). However, when the adjacent area paAc, just medialto Tpt on the superior temporal plane, was involved byinjections of Tpt in other cases, labeled terminals werethen found in the MGm. Furthermore, injections in theareas immediately rostral to Tpt, corresponding to theparabelt region (i.e., paAlt, Ts3) always labeled terminalsin the MGm. Overall, these findings indicate that Tpt canbe distinguished from the parabelt on the basis of itsthalamic and cortical connections (Hackett et al., 1998),and they suggest that Tpt is less directly involved inauditory processing, although it is certainly influenced byinputs from the auditory parabelt cortex.
Dorsal nuclei. Injections of the parabelt auditorycortex in the present account resulted in a topographicdistribution of labeled cells in the dorsal divisions (MGpdand MGad). Injections of the rostral parabelt labeled cellsposteriorly, in the MGpd, whereas injections of the caudalparabelt labeled cells anteriorly, primarily in the MGad.Overall, it appears that the extension of the AI/R bordermedially and laterally roughly serves as a line of demarca-tion for distinctions in corticocortical connections of thebelt and parabelt regions of the auditory cortex. Thus,injections in the parabelt caudal to lateral extension of theAI/R border tend to label cells in the caudal belt and caudalparabelt, whereas STG injections rostral to the border ofAI and R tend to label cells in the rostral belt and parabelt(Hackett et al., 1998). The pattern of thalamocorticalprojections observed in the present study reflects thistopography. A similar pattern has also been observed inthe thalamocortical connections of the lateral belt region(Burton and Jones, 1976; Morel and Kaas, 1992; Molinariet al., 1995). These studies have shown that cells in theMGpd project to the rostral lateral belt areas, whereas thecaudal lateral belt areas appear to be the primary targetsof the MGad. In the recent study by Molinari et al. (1995),injections of retrograde tracers were made in the rostraland caudal portions of the lateral belt region. Although noinjections were placed in the medial belt, it was clear thatthe MGad projected more strongly to the lateral auditorybelt caudal to the border of AI and R, whereas the MGpdprojected to the lateral belt cortex rostral to that border.
Suprageniculate and limitans nuclei
In primates, the suprageniculate nucleus (Sg) is locateddorsomedial to the MGC where it tends to merge with thedorsomedial edge of the MGad, and is continuous with theanterolateral expansion of the nucleus limitans (Lim;Olszewski, 1952; Jordan, 1973; Burton and Jones, 1976;
Jones, 1985; Morel et al., 1993). Most of the availableevidence suggests that subcortical projections to the Sgarise primarily from the deep layers of the superiorcolliculus (Morest, 1965; Benevento and Fallon, 1975;Casseday et al., 1976; Partlow et al., 1977; Oliver and Hall,1978a; Harting et al., 1980; Ledoux et al., 1987; Katoh etal., 1995), and that single neurons in the superior collicu-lus project bilaterally to the Sg (Lagares et al., 1994; Katohet al., 1995). Relatively minor projections to the Sg havebeen noted from the inferior colliculus (Katoh and Benedek,1995). Cortical targets of neurons in the Sg include theprefrontal cortex, superior temporal sulcus, and superiortemporal gyrus. In our material, most of the injections intothe parabelt labeled cells in the Sg/Lim. Injections thatfailed to label cells in the Sg/Lim were usually those thatalso exhibited poor transport of label to other areas of thebrain (Hackett et al., 1998). There was also consistentevidence for rostrocaudal topography in the connectionsbetween the parabelt and the Sg/Lim.
Existing descriptions of cortical connections with theSg/Lim vary widely, but they consistently associate thisregion of the thalamus with the auditory system. Walker(1935) found no degeneration in the Sg following hemide-cortication. Jones and Powell (1970a) reported antero-grade degeneration in Sg following lesions in SI, SII, andthe superior parietal lobe. Lesions of the posterior thala-mus involving the Sg resulted in degeneration in thetemporal operculum that could not be attributed specifi-cally to the Sg (Mesulam and Pandya, 1973). Yeterian andPandya (1989) reported topographic connections with theSTS that were very similar to those of the medial pulvinar(see below). Burton and Jones (1976) found topographi-cally organized labeling in the granular insula followingisotope injections restricted to the Sg and Sg/Lim complexin rhesus and squirrel monkeys. In their material, labelwas found in the lateral sulcus and caudal STG only whenan injection involved the MGC. In contrast, a number ofother studies at that time reported connections betweenthe Sg and STG in these primates (Forbes and Moskowitz,1974; Trojanowski and Jacobson, 1975; DeVito and Sim-mons, 1976). Auditory core and belt areas have Sg connec-tions, as well. In tamarins, Luethke et al. (1989) foundanterograde and retrograde labeling associated with injec-tions of WGA-HRP into AI, as well as cells labeled retro-gradely by fluorescent tracers. In owl monkeys, injectionsof tracers into AI and R of the core produced retrogradelylabeled cells in the Sg (Fitzpatrick and Imig, 1978; Moreland Kaas, 1992), although injections restricted to R wereassociated with much sparser labeling, compared with AI.Morel and Kaas (1992) also found labeled cells in the owlmonkey Sg associated with injections into the lateral beltand rostral superior temporal gyrus. Similar results werereported by Morel et al. (1993) in M. fascicularis, althoughlabeling in the Sg appeared to be heavier following injec-tion of the lateral belt compared with AI. Pandya et al.(1994) noted corticothalamic projections to the Sg/Limfrom the core, medial belt, lateral belt, and STG. A similarpattern of projections from the auditory cortex was re-ported by Molinari et al. (1995), except that they made noinjections into the STG.
Thus, it is clear that the Sg/Lim is broadly connectedwith the auditory structures of the brain. Based on thepattern of these connections, the Sg/Lim may be associatedwith the integration of multimodal inputs involved inmodality-independent spatial orientation of the head and
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 283

eyes (Katoh and Benedek, 1995). Therefore, it is probablysignificant that the Sg/Lim and the medial pulvinar (dis-cussed below) share similar subcortical and cortical connec-tions. Like the MGm, the Sg/Lim projects diffusely to thethree major regions of the auditory cortex (i.e., core, belt,and parabelt). The MGm and Sg/Lim also appear tocontain functionally distinct classes of cells that can bedistinguished on the bases of cortical and subcorticalconnections. As discussed earlier, calbindin- and parvalbu-min-positive cells in the MGm project to the auditorycortex in a laminar-specific manner. Thus, although thenotion is speculative, it is possible that distinct classes ofcells in the Sg/Lim project differentially to the auditorycortex as well.
Finally, different populations of cells in the Sg/Lim mayhave quite different cortical targets. Two primary popula-tions of large and small neurons have been identified in theSg of cats and opossums (Winer, 1985; Winer et al., 1988).In rats, Kurokawa and Saito (1995) found that a popula-tion of smaller neurons in the Sg projected to premotorfrontal cortex, whereas the overlapping population oflarger cells was retrogradely labeled by injections in theauditory association cortex of the temporal lobe (i.e., Te3).In cats, Katoh and Benedek (1995) reported that medialSC neurons tended to project to the posterior Sg, andneurons in the lateral SC to more anterior Sg. Further-more, efferents from the SC to the Sg arise from at leastfive types of cells in cats (Abramson and Chalupa, 1988;Katoh and Benedek, 1995). Thus, the architecture andconnections suggest that at least two functionally distinctclasses of neurons may exist in the Sg.
Medial pulvinar
The medial pulvinar (PM) is situated dorsomedially inthe thalamus forming much of the posterodorsal surface ofthe pulvinar (Jones, 1985). The PM is populated by widelyspaced small cells dispersed evenly throughout the nucleus(Olszewski, 1952; Burton and Jones, 1976; Cusick et al.,1993). The PM can be reliably distinguished from theadjacent nuclei of the pulvinar (e.g., PL, PI, PA) based ondifferences in architecture. In the present study, neuropiland cell staining in the PM was weak for acetylcholinester-ase, cytochrome oxidase, and parvalbumin, and muchstronger for calbindin. Calbindin immunoreactivity in thePM was uneven and banded, but more intense in dorsome-dial PM, gradually diminishing in intensity laterally andventrally. Nearer the lateral edge of the PM, and withinthe adjacent lateral pulvinar (PL), staining was darker inacetylcholinesterase, cytochrome oxidase, and parvalbu-min, and much lighter in calbindin (see also Heckers et al.,1992; Cusick et al., 1993; Gutierrez et al., 1995). Followingparabelt injections, cell labeling in the PM tended tooverlap with regions of darker immunoreactivity for calbin-din. Calbindin-positive cells reportedly project diffusely tolayer I of several cortical areas, whereas parvalbumin cellstend to project to the middle layers of specific cortical fields(Hashikawa et al., 1991, 1995; Molinari et al., 1995). Thus,the large numbers of calbindin-positive cells in the PM,and absence of parvalbumin-positive cells, may be consis-tent with the widespread cortical connections of thisnucleus.
The medial pulvinar (PM) is broadly connected withmany cortical areas, including the prefrontal cortex, poste-rior and inferior parietal cortex, superior and inferiortemporal cortex, insular cortex, and limbic cortex (for
reviews, see Kaas and Huerta, 1988; Yeterian and Pandya,1989; Romanski et al., 1997). Like the Sg/Lim complex,subcortical input to the PM arises primarily from the deeplayers of the superior colliculus (Benevento and Fallon,1975; Harting et al., 1980; Benevento and Standage,1983). Cortical connections with auditory-related areas ofthe superior temporal gyrus have been observed by severalinvestigators (Chow, 1950; Locke, 1960; Trojanowski andJacobson, 1975, 1976; Burton and Jones, 1976; DeVito andSimmons, 1976; Streitfeld, 1980; Yeterian and Pandya,1989; Morel and Kaas, 1992; Morel et al., 1993; Pandya etal., 1994). In addition, Locke (1960) and Pandya et al.(1994) reported connections between the PM and themedial belt. Fitzpatrick and Imig (1978) found a smallprojection from R, but not AI, to PM in owl monkeys.Finally, Akbarian et al. (1992) reported thalamic connec-tions to the parieto-insular vestibular cortex from the PM.
The results of the present study indicate that the PMprojects topographically to the dorsal STG in macaques.Rostral injections in the auditory parabelt tended to labelcells along the medial edge of the PM, whereas morecaudal injections revealed connections dorsally and later-ally, but typically not more lateral than the central third ofthe nucleus; thus the projections to the parabelt originatedin the medial and central PM, but not the lateral PM. Itmay be significant that the connections between the STGand the lateral PM tend to be restricted to the dorsal edgeof the PM where cells are larger and stain darkly. Other-wise, the rostrocaudal topography between the PM andSTG observed in our material was similar to that reportedfor the STS by Yeterian and Pandya (1989). In theirmaterial rostral injections into the dorsal bank of the STSlabeled cells in the medial PM (cases 1 and 2), whereasmore caudally placed injections in the dorsal bank of theSTS resulted in labeling in the central/lateral PM, as wellas PL (cases 3–8).
Comparison across studies suggests that the lateral PMis more strongly related to visual processing, by virtue ofits connections with posterior parietal and inferior tempo-ral cortex (DeVito and Simmons, 1976; Baleydier andMauguiere, 1985, 1987; Schmamann and Pandya, 1990;Baleydier and Morel, 1992; Hardy and Lynch, 1992; Cu-sick et al., 1993; Morecraft et al., 1993; Webster et al.,1993; Romanski et al., 1997), whereas the medial PM ismore strongly related to auditory processing, based onpreferential connections with auditory areas of the supe-rior temporal region established in the present study andseveral others (Simpson, 1952; Locke, 1960; DeVito andSimmons, 1976; Trojanowski and Jacobson, 1976; Burtonand Jones, 1976; Mauguiere and Baleydier, 1978; Streit-feld, 1980; Markowitsch et al., 1985; Moran et al., 1987;Pandya et al., 1994; Romanski et al., 1997). Most of thestudies cited above, among others (Jones and Powell,1970b; Trojanowski and Jacobson, 1976; Divac et al., 1977;Asanuma et al., 1985; Yeterian and Pandya, 1985, 1989;1991), have shown that the central PM has connectionswith auditory, somatic, visual, and multimodal corticalareas, suggesting that the central PM may subservemultimodal integrative functions more than the lateral ormedial portions of the nucleus, which appear to be some-what more modality specific. That is, the thalamocorticalconnection patterns reviewed here suggest that function-ally distinct groups of cells are segregated in the lateral(visual) and medial (auditory) PM, with more overlap inthe central third of the nucleus. Overall, the widespread
284 T.A. HACKETT ET AL.

connections of the PM suggest roles in spatial orientationand attentional processes across modalities, but the func-tions of the PM are poorly understood at this time.
ACKNOWLEDGMENTS
This research was supported by NINDS grant NS 16446to J.H.K. and NIDCD Research Fellowship DC 00249 toT.A.H.
REFERENCES
Abramson, B.P. and L.M. Chalupa (1988) Multiple pathways from thesuperior colliculus to the extrageniculate visual thalamus of the cat. J.Comp. Neurol. 271:397–418.
Aitkin, L.J., M. Kudo, and D.R.F. Irvine (1988) Connections of the primaryauditory cortex in the common marmoset (Callithrix jacchus jacchus).J. Comp. Neurol. 269:235–248.
Akbarian, S., O.-J. Grusser, and W.O. Guldin (1992) Thalamic connectionsof the vestibular cortical fields in the squirrel monkey (Saimiri sci-ureus). J. Comp. Neurol. 326:423–441.
Asanuma, C., R.A. Andersen, and W.M. Cowan (1985) The thalamicrelations of the caudal inferior parietal lobule and the lateral prefrontalcortex in monkeys: Divergent cortical projections from cell clusters inthe medial pulvinar nucleus. J. Comp. Neurol. 241:357–381.
Baleydier, C. and F. Mauguiere (1985) Anatomical evidence for medialpulvinar connections with the posterior cingulate cortex, the retrosple-nial area, and the posterior parahippocampal gyrus in monkeys. J.Comp. Neurol. 232:219–228.
Baleydier, C. and F. Mauguiere (1987) Network organization of theconnectivity between parietal area 7, posterior cingulate cortex, andmedial pulvinar nucleus: A double fluorescent tracer study in monkey.Exp. Brain. Res. 66:385–393.
Baleydier, C. and A. Morel (1992) Segregated thalamocortical pathways toinferior parietal and inferotemporal cortex in macaque monkey. Vis.Neurosci. 8:391–405.
Benevento, L.A. and J.H. Fallon (1975) The ascending projections of thesuperior colliculus in the rhesus monkey (Macaca mulatta). J. Comp.Neurol. 160:339–362.
Benevento, L.A. and G.P. Standage (1983) The organization of projections ofthe retinorecipient and nonretinorecipient nuclei of the pretectal com-plex and layers of the superior colliculus to the lateral pulvinar andmedial pulvinar in the macaque monkey. J. Comp. Neurol. 217:307–336.
Burton, H. and E.G. Jones (1976) The posterior thalamic region and itscortical projection in new world and old world monkeys. J. Comp.Neurol. 168:249–302.
Calford, M.B. and L.M. Aitkin (1983) Ascending projections to the medialgeniculate body of the cat: Evidence for multiple, parallel auditorypathways through thalamus. J. Neurosci. 3:2365–2380.
Casseday, J.H., J.K. Harting, and I.T. Diamond (1976) Auditory pathwaysto the cortex in Tupaia glis. J. Comp. Neurol. 168:249–302.
Chow, K.L. (1950) A retrograde cell degeneration study of the corticalprojection field of the pulvinar in the monkey. J. Comp. Neurol.93:313–340.
Clark, W.E. Le Gros, and D.W.C. Northfield (1937) The cortical projection ofthe pulvinar in the macaque monkey. Brain 60:126–142.
Cusick, C.G., J.L. Scripter, J.G. Darensbourg, and J.T. Weber (1993)Chemoarchitectonic subdivisions of the visual pulvinar in monkeys andtheir connectional relations with the middle temporal and rostraldorsolateral visual areas, MT and DLr. J. Comp. Neurol. 336:1–30.
DeVenecia, R.K., C.B. Smelser, S.D. Lossman, and N.T. McMullen (1995)Complementary expression of parvalbumin and calbindin D-28k delin-eates subdivisions of the rabbit medial geniculate body. J. Comp.Neurol. 359:595–612.
DeVito, J.L. and D.M. Simmons (1976) Some connections of the posteriorthalamus in monkey. Exp. Neurol. 51:347–362.
Divac, I., J.H. LaVail, P. Rakic, and K.R. Winston (1977) Heterogeneousafferents to the inferior parietal lobule of the rhesus monkey revealedby retrograde transport method. Brain Res. 123:197–207.
Fitzpatrick, K.A. and T.J. Imig (1978) Projections of auditory cortex uponthe thalamus and midbrain in the owl monkey. J. Comp. Neurol.177:537–556.
Forbes, B.F. and N. Moskowitz (1974) Projections of auditory responsivecortex in the squirrel monkey. Brain Res. 67:239–254.
Gallyas, F. (1979) Silver staining of myelin by means of physical develop-ment. Neurol. Res. 1:203–209.
Geneser-Jensen, F.A. and T.W. Blackstad (1971) Distribution of acetylcho-linesterase in the hippocampal region of the guinea-pig. I. Entorhinalarea, parasubiculum, and presubiculum. Z. Zellforsch. Mikr. Anat.114:460–481.
Gibson, A.R., D.I. Hansma, J.C. Houk, and F.R. Robinson (1984) A sensitivelow artifact TMB procedure for the demonstration of WGA-HRP in theCNS. Brain Res. 298:235–241.
Gutierrez, C., A. Yaun, and C.G. Cusick (1995) Neurochemical subdivisionsof the inferior pulvinar in macaque monkeys. J. Comp. Neurol. 363:545–562.
Hackett, T.A., I. Stepniewska, and J.H. Kaas (1996) Thalamic connectionsof the parabelt auditory region in macaque monkeys. Soc. Neurosci.Abstr. 22(2):186.
Hackett, T.A., I. Stepniewska, and J.H. Kaas (1998) Subdivisions ofauditory cortex and ipsilateral cortical connections of the parabeltauditory cortex in macaque monkeys. J. Comp. Neurol. 394:475–495.
Hardy, S.G. and J.C. Lynch (1992) The spatial distribution of pulvinarneurons that project to two subregions of the inferior parietal lobule inthe macaque. Cereb. Cortex. 2:217–230.
Harting, J.K., M.F. Huerta, A.J. Frankfurter, N.L. Strominger, and G.J.Royce (1980) Ascending pathways from the monkey superior colliculus:An autoradiographic analysis. J. Comp. Neurol. 192:853–882.
Hashikawa, T., E. Rausell, M. Molinari, and E.G. Jones (1990) Chemicallydistinct medial geniculate cells project to different layers of monkeyauditory cortex. Soc. Neurosci. Abstr. 16:717.
Hashikawa, T., E. Rausell, M. Molinari, and E.G. Jones (1991) Parvalbu-min- and calbindin-containing neurons in the monkey medial genicu-late complex: Differential distribution and cortical layer specific projec-tions. Brain Res. 544:335–341.
Hashikawa, T., M. Molinari, E. Rausell, and Jones (1995) Patchy andlaminar terminations of medial geniculate axons in monkey auditorycortex. J. Comp. Neurol. 362:195–208.
Heckers, S., G. Changiz, and M.M. Mesulam (1992) Cholinergic innervationof the human thalamus: Dual origin and differential nuclear distribu-tion. J. Comp. Neurol. 325:68–82.
Jones, E.G. (1985) The Thalamus. New York: Plenum Press.Jones, E.G. and S.H.C. Hendry (1989) Differential calcium-binding protein
immunoreactivity distinguishes classes of relay neurons in monkeythalamic nuclei. Eur. J. Neurosci. 1:221–246.
Jones, E.G. and T.P.S. Powell (1970a) Connexions of the somatic sensorycortex of the rhesus monkey. III. Thalamic connexions. Brain. 93:37–56.
Jones, E.G. and T.P.S. Powell (1970b) An anatomical study of convergingsensory pathways within the cerebral cortex of the monkey. Brain93:793–820.
Jordan, H. (1973) Structure of the medial geniculate nucleus (MGN): A cyto-and myeloarchitectonic study in the squirrel monkey. J. Comp. Neurol.148:469–480.
Kaas, J.H. and T.A. Hackett (1998) Subdivisions of auditory cortex andlevels of processing in primates. Audiol. Neurootol. 3:73–85.
Kaas, J.H. and M.F. Huerta (1988) Subcortical visual system of primates. InH.P. Steklis (ed): Comparative Primate Biology, Vol. 4: Neurosciences.New York: Alan R. Liss, pp. 327–391.
Katoh, Y.Y. and G. Benedek (1995) Organization of the colliculo-supragenicu-late pathway in the cat: A wheat germ agglutinin-horseradish peroxi-dase study. J. Comp. Neurol. 352:381–397.
Katoh, Y.Y., G. Benedek, and S. Deura (1995) Bilateral projections from thesuperior colliculus to the suprageniculate nucleus in the cat: A WGA-HRP/double fluorescent tracing study. Brain Res. 669:298–302.
Kosmal, A., M. Malinowska, and D.M. Kowalska (1997) Thalamic andamygdaloid connections of the auditory association cortex of thesuperior temporal gyrus in rhesus monkey (Macaca mulatta). ActaNeurobiol. Exp. 57:165–188.
Kurokawa, T. and H. Saito (1995) Retrograde axonal transport of differentfluorescent tracers from the neocortex to the suprageniculate nucleus inthe rat. Hear. Res. 85:103–108.
Lagares, C., M. Caballero-Bleda, B. Fernandez, and L. Puelles (1994)Reciprocal connections between the rabbit suprageniculate pretectalnucleus and the superior colliculus: Tracer study with horseradishperoxidase and fluorogold. Vis. Neurosci. 11:347–353.
CONNECTIONS OF THE PARABELT AUDITORY CORTEX 285

LeDoux, J.E., D. Ruggiero, R. Forest, R. Stornetta, and D.J. Reis (1987)Topographic organization of convergent projections to the thalamusfrom the inferior colliculus and spinal cord in the rat. J. Comp. Neurol.264:123–146.
Locke, S. (1960) Projection of the medial pulvinar of the macaque. J. Comp.Neurol. 115:155–169.
Luethke, L.E., L.A. Krubitzer, and J.H. Kaas (1989) Connections of primaryauditory cortex in the new world monkey, Saguinus. J. Comp. Neurol.285:487–513.
Markowitsch, H.J., D. Emmans, E. Irle, M. Streicher, and B. Preilowski(1985) Cortical and subcortical afferent connections of the primate’stemporal pole: A study of rhesus monkeys, squirrel monkeys, andmarmosets. J. Comp. Neurol. 242:425–458.
Mauguiere, F. and C. Baleydier (1978) Topographical organization ofmedial pulvinar neurons sending fibers to Brodman’s areas 7, 21, and22 in the monkey. Exp. Brain Res. 31:605–607.
Mesulam, M.M. and D.N. Pandya (1973) The projections of the medialgeniculate complex within the sylvian fissure of the rhesus monkey.Brain Res. 60:315–333.
Molinari, M., M.E. Dell’Anna, E. Rausell, M.G. Leggio, T. Hashikawa, andE.G. Jones (1995) Auditory thalamocortical pathways defined in mon-keys by calcium-binding protein immunoreactivity. J. Comp. Neurol.362:171–194.
Moran, M.A., E.J. Mufson, and M.M. Mesulam (1987) Neural inputs intotemporal polar cortex of the rhesus monkey. J. Comp. Neurol. 256:88–103.
Morecraft, R.J., C. Guela, and M.M. Mesulam (1993) Architecture ofconnectivity within a cingulo-fronto-parietal neurocognitive networkfor directed attention. Arch. Neurol. 50:279–284.
Morel, A. and J.H. Kaas (1992) Subdivisions and connections of auditorycortex in owl monkeys. J. Comp. Neurol. 318:27–63.
Morel, A., P.E. Garraghty, and J.H. Kaas (1993) Tonotopic organization,architectonic fields, and connections of auditory cortex in macaquemonkeys. J. Comp. Neurol. 335:437–459.
Morest, D.K. (1965) The lateral tegmental system of the midbrain andmedial geniculate body: Study with Golgi and Nauta methods in cat. J.Anat. 99:611–634.
Oliver, D.L. and W.C. Hall (1978a) The medial geniculate body of the treeshrew, Tupaia glis. I. Cytoarchitecture and midbrain connections. J.Comp. Neurol. 182:423–458.
Oliver, D.L. and W.C. Hall (1978b) The medial geniculate body of the treeshrew, Tupaia glis. II. Connections with the neocortex. J. Comp. Neurol.182:459–494.
Olszewski, J. (1952) The Thalamus of the Macaca mulatta. New York:Karger, 1952.
Pandya, D.N. and F. Sanides (1973) Architectonic parcellation of thetemporal operculum in rhesus monkey and its projection pattern. Z.Anat. Entwickl.-Gesch. 139:127–160.
Pandya, D.N., D.L. Rosene, and A.M. Doolittle (1994) Corticothalamicconnections of auditory-related areas of the temporal lobe in the rhesusmonkey. J. Comp. Neurol. 345:447–471.
Partlow, G.D., M. Colonnier, and J. Szabo (1977) Thalamic projections of thesuperior colliculus in the rhesus monkey, Macaca mulatta: a light andelectron microscopic study. J. Comp. Neurol. 171:285–318.
Romanski, L.M., M. Giguere, J.F. Bates, and P.S. Goldman-Rakic (1997)Topographic organization of medial pulvinar connections with theprefrontal cortex in the rhesus monkey. J. Comp. Neurol. 379: 313–332.
Schmamann, J.D. and D.N. Pandya (1990) Anatomical investigation ofprojections from thalamus to posterior parietal cortex in the rhesusmonkey: A WGA-HRP and fluorescent tracer study. J. Comp. Neurol.295:299–326.
Simpson, D.A. (1952) The projections of the pulvinar to the temporal lobe. J.Anat. 86:20–28.
Siqueria, E.B. (1965) The temporo-pulvinar connections in the rhesusmonkey. Arch. Neurol. 13:321–330.
Streitfeld, B.D. (1980) The fiber connections of the temporal lobe withemphasis on the rhesus monkey. Int. J. Neurosci. 11:51–71.
Trojanowski, J.Q. and S. Jacobson (1975) A combined horseradish peroxi-dase-autoradiographic investigation of reciprocal connections betweensuperior temporal gyrus and pulvinar in squirrel monkey. Brain Res.85:347–353.
Trojanowski, J.Q. and S. Jacobson (1976) Areal and laminar distribution ofsome pulvinar efferents in rhesus monkey. J. Comp. Neurol. 169:371–392.
Walker, A.E. (1935) Retrograde cell degeneration in the thalamus ofMacaca mulatta following hemidecortication. J. Comp. Neurol. 62:407–419.
Walker, A.E. (1937) The projection of the medial geniculate body to thecerebral cortex in the macaque monkey. J. Anat. 71:319–331.
Webster, M.J., J. Bachevalier, and L.G. Ungerleider (1993) Subcorticalconnections of inferior temporal areas TE and TEO in macaquemonkeys. J. Comp. Neurol. 335:73–91.
Whitlock, D.G. and W.S.H. Nauta (1956) Subcortical projections from thetemporal neocortex in Macaca mulatta. J. Comp. Neurol. 106:183–212.
Winer, J.A. (1985) The medial geniculate body of the cat. Adv. Anat.Embryol. Cell Biol. 86:1–97.
Winer, J.A., D.K. Morest, and I.T. Diamond (1988) A cytoarchitectonic atlasof the medial geniculate body of the opossum, Didelphys virginiana,with a comment on the posterior intralaminar nuclei of the thalamus. J.Comp. Neurol. 176:387–418.
Wong-Riley, M. (1979) Changes in the visual system of monocularly suturedor enucleated cats demonstratable with cytochrome oxidase histochem-istry. Brain Res. 171:11–28.
Yeterian, E.H. and D.N. Pandya (1985) Corticothalamic connections of theposterior parietal cortex in the rhesus monkey. J. Comp. Neurol.237:408–426.
Yeterian, E.H. and D.N. Pandya (1989) Thalamic connections of the cortexof the superior temporal sulcus in the rhesus monkey. J. Comp. Neurol.282:80–97.
Yeterian, E.H. and D.N. Pandya (1991) Corticothalamic connections of thesuperior temporal sulcus in rhesus monkeys. Exp. Brain. Res. 83:268–284.
286 T.A. HACKETT ET AL.