Embed Size (px)
Citation preview

Accepted Manuscript
Title: Testing Central and Inner Asian admixture amongcontemporary Hungarians
Author: Andras Bıro Tibor Feher Gusztav Barany HorolmaPamjav
PII: S1872-4973(14)00247-6DOI: http://dx.doi.org/doi:10.1016/j.fsigen.2014.11.007Reference: FSIGEN 1270
To appear in: Forensic Science International: Genetics
Received date: 16-8-2014Revised date: 5-11-2014Accepted date: 7-11-2014
Please cite this article as: A. Biro, T. Feher, G. Barany, H. Pamjav, Testing Centraland Inner Asian admixture among contemporary Hungarians, Forensic ScienceInternational: Genetics (2014), http://dx.doi.org/10.1016/j.fsigen.2014.11.007
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

Page 1 of 19
Accep
ted
Man
uscr
ipt
1
Testing Central and Inner Asian admixture among contemporary Hungarians1
2
Authors:3
András Bíró 1, Tibor Fehér 2, Gusztáv Bárány2 and Horolma Pamjav2*4
1Department of Anthropology, Hungarian Natural History Museum, Budapest H-1088, 5
Hungary6
2Institute of Forensic Medicine, Network of Forensic Science Institutes, Ministry of Justice,7
Budapest, Hungary8
*Corresponding author: Horolma Pamjav ([email protected]), Tel: 361-457-01-83, Fax: 9
361-457-018210
Address: Institute of Forensic Medicine, Network of Forensic Science Institutes, Ministry of 11
Justice, Budapest, Hungary12
1536 Budapest, PO 216. Hungary13
14
1.15
Highlights16966 samples tested from Central/Inner Asian) and Hungarian-speaking populations.17The possible paternal genetic contribution from Central/Inner Asian populations to 18contemporary Hungarian speaking populations ranges between 5 - 7.4%. 19Present-day Hungarian speakers are genetically very similar to neighbouring populations, 20isolated Hungarian speaking groups having relatively higher presence of Central and Inner 21Asian genetic elements.22
2.23
3.24
4.25
5.26
27

Page 2 of 19
Accep
ted
Man
uscr
ipt
2
6. Introduction27
For centuries, great efforts were made by Hungarian historians to study the earliest 28
period of their national history. While the academic mainstream was clearly in favour of the 29
Hungarian language belonging to the Uralic family, many other researchers favour the theory 30
of a closer relationship with the Turkic language family and Turkic peoples. Anthropological 31
analysis of bones originating in the 10th century showed characteristics of Central Asian 32
origin [1, 2, 3]. Archaeological remains of weapons, haversacks, belt mountings, and 33
ornaments on clothing also showed similarities to those of Central and Inner Asia [4, 5, 6, 7]. 34
Hungarian archaeologists and ethnographers showed that there are similarities in the traditions 35
of the ancient Hungarians and various Central and Inner Asian cultures [8, 9, 10]. These were 36
in the areas of burial, belief, and figurative arts. Therefore, an origin of the Hungarian 37
language and early culture in a region ranging from Asia to Siberia is suggested, but a specific 38
origin has been difficult to identify. 39
Finns were thought to be close genetic relatives of Hungarians. However, based on 40
studies done with mtDNA, Y chromosome STRs and SNPs, they seem to have little 41
genetically in common with Hungarians [11, 12]. And this is despite the fact that they also 42
speak a non-Indo-European Finno-Ugric language. A genetic relationship was proven 43
between two Hungarian ethnic groups, the Csangos and Seklers. Both groups showed genetic 44
affiliations with certain Central Asian and European populations. These findings could have 45
supported theories about a partially Asian origin of Hungarian population [11]. However,46
most of the Central Asian-Hungarian Y-chromosomal relationship was based on the high 47
frequency of haplogroup R1a-M198 among Kyrgyz and a small Hungarian sample, without 48
knowing the deep structure of this haplogroup. Since then, first Pamjav et al. [13], and then in 49
a more comprehensive analysis by Underhill et al. [14] it was shown that there is a clear SNP-50
based distinction between Eastern European (Z282, Z280, M458) and Central Asian (Z93) 51
R1a-M198 males.52

Page 3 of 19
Accep
ted
Man
uscr
ipt
3
It was also noted that Hungarians show very limited or no presence of Haplogroup N-53
M231 – including subclade N1c-Tat –, which is frequent among other Uralic-speaking 54
populations [15, 16]. However, the potential genetic relationship with Turkic and Inner Asian55
peoples has been less researched, although this relationship could shed light on the genetic 56
basis of the alternative Turkic (Turanian) theory. Different Turkic-speaking populations have 57
widely differing Y-chromosomal gene pools. They range from N1c-Tat dominated Yakuts 58
through C3-M47 dominated Kazakhs, and Q-M25 dominated Turkmens to genetically more 59
diverse Uzbeks, Azeri and Anatolian Turks (Table S1). Therefore, we chose not to focus only 60
on haplogroup frequencies, but on analysing haplotype structure. We have undertaken a 61
survey of 966 samples from Europe and Asia. This study is expected to provide insights 62
relevant to the Central and Inner Asian genetic contribution into Hungarian speaking 63
populations. It will also provide insight into how the genetic variation is distributed in the 64
contemporary Hungarian, Central and Inner Asian population gene pool studied. 65
7. Materials and Methods66
7.1. DNA samples67
To analyze the genetic relationship of present-day Hungarians with present-day Central 68
and Inner Asians, we tested 522 samples from Hungarian-speaking populations (33269
Hungarians from Hungary, 95 Sekler from Romanian Transylvania, 95 Csango from 70
Romanian Moldova), 115 Uzbek samples from various parts of Uzbekistan (Ferghana Valley, 71
Tashkent, Khwarezm, Samarqand, Surkhodarya, Karakalpakstan), 8 samples from 72
Kazakhstan’s Aqtöbe region, 127 Mongolian and 88 Buryat Mongolian samples from 73
Mongolia. Archaic Sekler and Csango populations were included to increase the matching 74
potential, and we also collected additional samples from tribes whose self-designation may 75
have connection to the ethnonym Magyar, i.e. 61 Madjars from Uzbekistan and 45 Madjars 76
from Kazakhstan. Out of the 966 samples, the 45 Kazakh Madjars [17], and 215 Hungarian77
samples [12] were published before, but tested for further SNPs and samples in this study.78

Page 4 of 19
Accep
ted
Man
uscr
ipt
4
The new samples published herein were sent to the YHRD and the accession numbers are the 79
following: Uzbekistan [Uzbek] YA003994, Uzbekistan [Madjar] YA003995, Mongolia 80
[Buryat] YA003996, Mongolia [Mongolian] YA003997 and Kazakhstan [Madjar, Aqtöbe] 81
YA003998. The new populations, as well as the previously published populations were 82
Hungarian [12], with accession number YP000175, Csango [Romanian] YP000227 [18] and 83
Sekler [Romanian] YP000257 [18]. These were used for comparison and can be referenced at 84
www.yhrd.org. The Y-SNP haplogroups for Sekler and Csango populations were tested by us 85
and included in our data.86
Each person gave their informed consent prior to their inclusion in the study. 87
7.2. Testing of Y-STR and Y-SNP markers88
DNA was amplified with the PowerPlex Y (Promega, USA) amplification kit including 12 89
Y-STR loci, according to the manufacturer’s instructions. Fragment sizes and allele 90
designations were determined with a 3130 Genetic Analyzer (Life Technologies, Foster City, 91
CA) using GeneMapper IDX 1.2.1. software.92
When testing Y-SNP markers, amplifications of 1-2 ng genomic DNA were performed in 93
an ABI 7500 Real-time PCR instrument with Taqman Assay (Life Technologies, Foster City, 94
CA) using the programs designed by the manufacturer. The relative fluorescence of the PCR 95
products were analyzed on an ABI 7500 with its’ SDS software, as described in the 96
manufacturer’s manual (Life Technologies, Foster City, CA). Fifty-five Y-chromosomal SNP 97
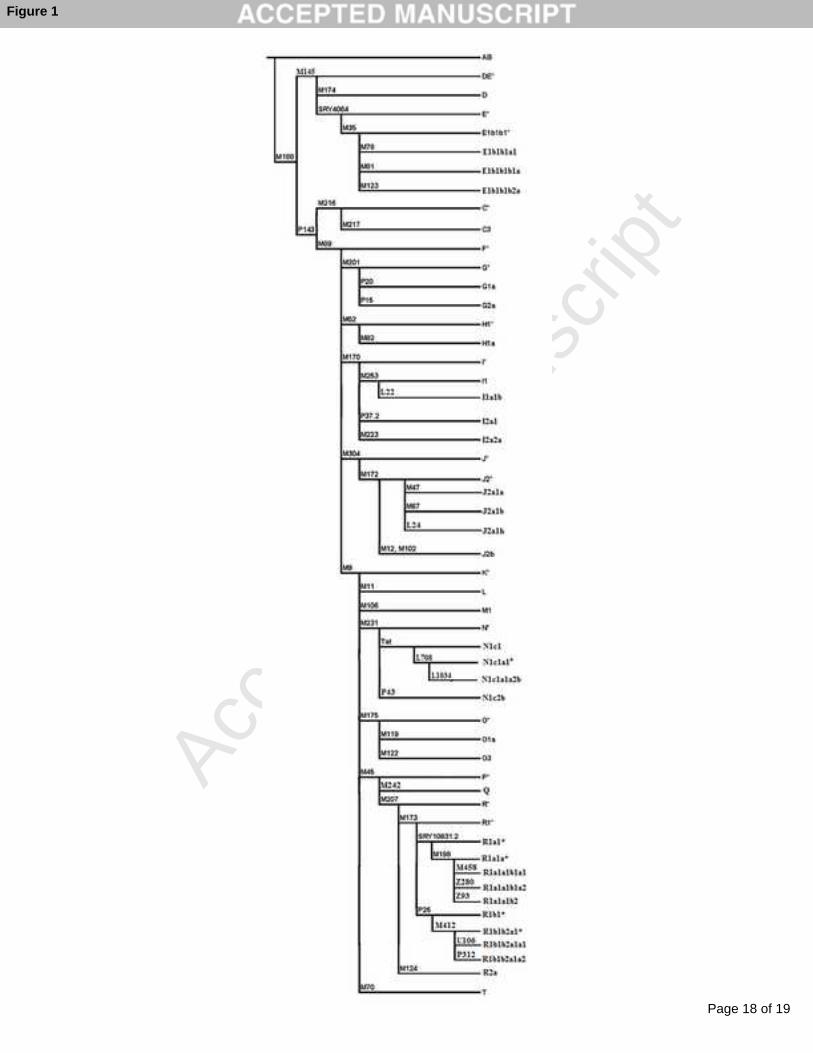
markers were tested with Taqman Assays (Fig. 1). The haplogroups tested and the markers 98
used in the study originated from YCC (Y-Chromosomal Consortium). The nomenclature of 99
haplogroups followed the ISOGG 2014 Y-DNA haplogroup tree due to recent, new additions 100
uncovered by YCC (Y-Chromosomal Consortium).101
A list of primers and Taqman probes for binary markers was previously published [19],102
but we now updated the list with new SNPs studied, as shown in Table S2. A new103

Page 5 of 19
Accep
ted
Man
uscr
ipt
5
downstream SNP marker, L24, was tested for J2*-M172 (xM47, M67, M12) samples to 104
obtain more resolution within the haplogroup as suggested by van Oven et al. [20].105
7.3. Data analysis106
To examine the STR variation within the haplogroups, networks were constructed using 107
the Network 4.6.1.2 program [21]. Repeats of the locus DYS389I were subtracted from the 108
locus DYS389II and, as is common practice, the locus DYS385 was excluded from the 109
network. Within the network program, the rho statistic was used to estimate the time to the 110
most recent common ancestor (TMRCA) of haplotypes within the compared haplogroups. 111
Evolutionary time estimates were calculated according to Zhivotovsky et al. [22] and STR 112
mutation rate was assumed to be 6.9x10-4 /locus/25 years. STR-based TMRCA estimates113
(Table S3) are not discussed in this paper due to their unreliability [14] and their irrelevance114
to the main purpose of the study.115
8. Results116
Based on ten Y-STR loci, networks were constructed within each of the haplogroups. 117
These haplogroups overlapped among populations studied. All haplotype and haplogroup 118
results can be found in Table S4. Haplogroup results are summarized in Table S5. For our 119
analysis, we only considered those haplogroups which occurred in more than one sample120
among both Hungarians and Central Asians (Uzbeks, Kazakhs, Madjars), or among 121
Hungarians and Inner Asians (Mongolians, Buryats). With this method, we identified 9122
haplogroups, which might indicate a genetic relationship between contemporary Magyars and 123
Altaic-speaking populations. They are E-M78, G2a-P15, J2*(xM47, M67 and M12), N1c-124
L708, Q-M242, R1a-M458, R1a-Z280, R1a-Z93 and R1b*-P25(xM412). To verify or confute 125
the relationships, we created median-joining networks on 10 loci for all “suspected” 126
haplogroups. Results are discussed only in the context of the potential matches between 127
Hungarian and Altaic-speaking populations, haplogroup by haplogroup. The haplogroups are 128
described as follows. 129

Page 6 of 19
Accep
ted
Man
uscr
ipt
6
Haplogroup E-M78130
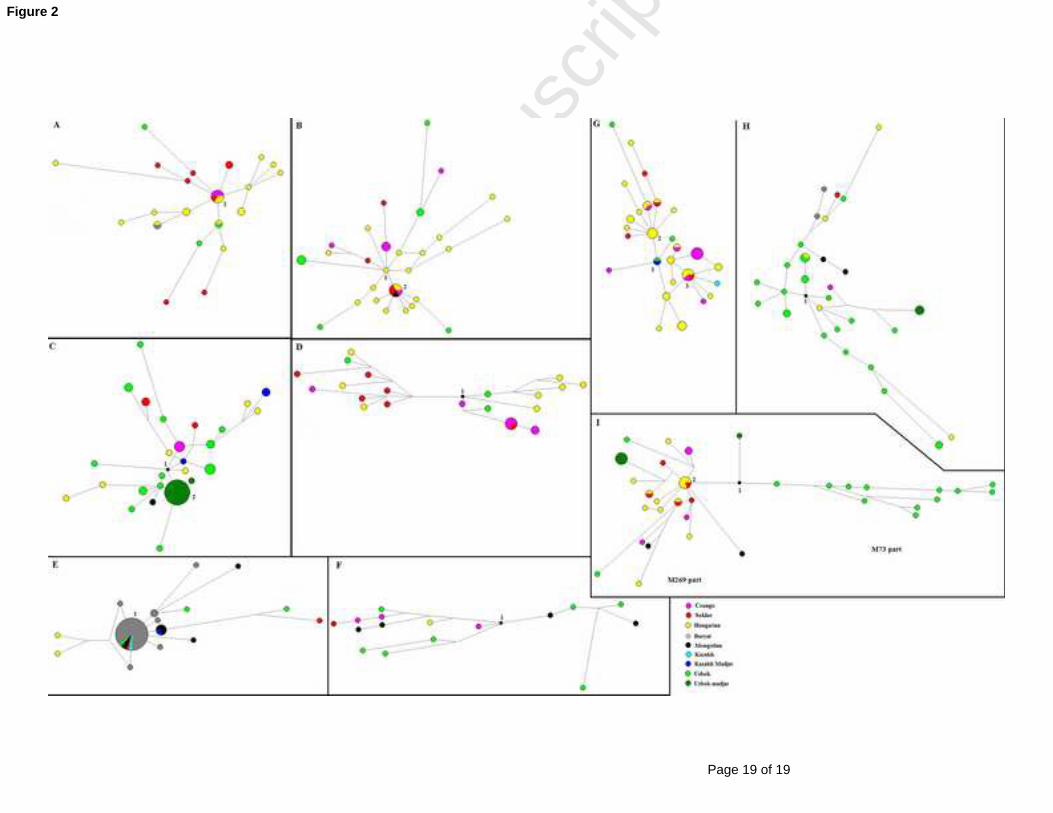
The median joining network (MJ) of 31 E-M78 haplotypes is shown in Fig. 2A. The 131
network shows a star-like pattern. The biggest cluster (cluster 1 in Fig. 2A) was the modal 132
haplotype shared by three Hungarian speaking population groups, which consisted of two133
Hungarian, one Sekler and three Csango males. There is a potential that two Seklers and two 134
Hungarians have a common origin with Uzbeks (1 from Tashkent, 1 from Khwarezm) on the 135
bottom of the network. 136
Haplogroup G2a-P15137
MJ network of 38 G2a-P15 haplotypes is depicted in Fig. 2B. The modal haplotype 138
cluster (cluster 2 in Fig. 2B) is shared by 4 populations including two Hungarians, two139
Seklers, one Csango and one Mongolian male. One Csango is on a common branch with three140
Uzbeks (on the top of the network). The three Uzbeks are from Khwarezm subregion.141
Haplogroup J2*-M172 and J2-L24142
MJ network of 51 J2*-M172 haplotypes is seen in Fig. 2C. There is no visible modal 143
haplotype cluster and it yielded a non-star like network. The lower part of the figure includes 144
all Uzbek Madjars, who are a homogenous population most likely affected by a founder effect145
or genetic drift. The biggest cluster (cluster 2 in fig. 2C) consists of 17 Uzbek Madjar males. 146
There are some other Uzbeks, one Mongolian and two Hungarians, which we consider to be 147
Central and Inner Asian. The upper part of the figure is less clear, but we can see that one148
Hungarian and two Seklers derive from the Kazakh Madjar in the centre (to the left), and two149
Hungarians who come from Uzbek haplotypes in the upper right part. So among J2*-M172 150
haplotypes, we consider 2 Seklers and 5 Hungarians to be of Central Asian admixture.151
Twenty-four J2-L24 haplotypes resulted in a non-star like network split into two parts,152
primarily based on DYS437 and DYS391 loci (Fig. 2D). In the network on the upper right 153
side, four Hungarians derive from an Uzbek (Ferghana) haplotype, so we considered them to 154
be of a Central Asian admixture.155

Page 7 of 19
Accep
ted
Man
uscr
ipt
7
The other part of the network (on the left side) included Hungarian speaking males,156
except for one Uzbek male and no shared haplotype was seen.157
Haplogroup N1c-Tat158
An MJ network of 54 N1c-Tat haplotypes is shown in Fig. 2E. The modal haplotype 159
cluster (cluster 1 in Fig. 2E) is shared by 5 population groups including 31 Buryat, two160
Mongolian, one Sekler, one Uzbek and one Kazakh chromosome. One Sekler matches the 161
Buryat-Mongolian modal haplotype and thus can be considered Inner Asian admixture in162
Hungary. Other Hungarian and Sekler haplotypes are very far from Altaic N1c haplotypes and 163
are therefore more likely of Uralic or Baltic origin. It has been noted that the most Buryat 164
males share the same haplotype, which is due to genetic drift. 165
Haplogroup Q-M242166
The network of Q-M242 haplotypes shows a non-star like and more diverse pattern,167
which makes it rather difficult to analyse the relationship (Fig. 2F). However, due to the pre-168
eminence of Q-M242 among Altaic Turkmens [23, 24] and its’ general absence in Europe and 169
Finno-Ugric speaking populations, we assume Central Asian admixture for all the 5 170
Hungarian speaking Q individuals.171
Haplogroup R1a-M458172
The network of 49 R1a-M458 haplotypes breaks down into two easily identifiable 173
star-like subclusters (Fig. 2G). The biggest cluster (cluster 3 in Fig. 2G) includes three174
Hungarians, one Sekler and one Csango male. The second largest cluster consists of four175
Hungarians (Fig. 2G, cluster 2). An interesting picture is noted in that one Uzbek from 176
Khwarezm and one Madjar from Kazakhstan are in the middle of the network connecting the 177
two separate clusters as the median haplotype (Fig.2G, cluster 1). One Csango descends from 178
this central haplotype and thus can be designated as Central Asian admixture. While R1a-179
M458 is generally considered as an Eastern European haplogroup, being especially frequent 180
among Western Slavs and to a lesser extent, Eastern Slavs [25], based on our result we cannot 181

Page 8 of 19
Accep
ted
Man
uscr
ipt
8
exclude the possibility that R1a-M458 originates from Central Asia. These Khwarezm and 182
Madjar haplotypes may be the remnants of the ancestral population. Attributing one Kazakh,183
one Kazakh Madjar, and all three Uzbek R1a-M458 haplogroups to Slavic admixture seems 184
unlikely, especially given the nearly complete lack of other typically European haplogroups I-185
M170 and R1b-M412 among our Central and Inner Asian samples (Table S4).186
Haplogroup R1a-Z280187
The R1a-Z280 haplotypes produced a star-like network (figure not shown), with all 188
the Central Asians exactly matching Hungarian haplotypes (Table S5). Therefore, we189
assume that a genetic link was from Finno-Ugric or Slavic peoples to Central Asians [13].190
Haplogroup R1a-Z93191
Thirty-six R1a-Z93 haplotypes produced a non-star like and very diverse network192
(Fig. 2H). These included mostly Uzbek haplotypes found in the central area, who were 193
Hungarian-speaking, Uzbek Madjar, Buryat and Mongolian populations were branching off 194
towards the edges. While some R1a-Z93 haplotypes might be a result of Roma admixture195
[13], for the purpose of this study we assumed that all haplotypes of Hungarian-speaking 196
population groups to be Central/Inner Asian admixture.197
Haplogroup R1b-P25198
The network of 44 R1b-P25 (xM412) haplotypes clearly consists of two clusters (Fig. 2I). 199
On the left side, we find the star-like network of M269 haplotypes, while on the right side, the 200
typically Central Asian subgroup M73 is visible (Note: M269 and M73 markers were not 201
tested in this study, but a comparison with Myres et al. [26] STR-data suggest the202
connection). Among R1b-P25 haplotypes, no connection can be made, as Hungarian-203
speakers dominantly belong to the M269 branch, while Uzbeks belong to the M73 part. The 204
limited number of Central Asians (M269) is situated on the edges of the network, thus 205
representing external admixture rather than source. Cluster 2 (Fig. 2I) in the part of M269 206
consists of four Hungarians and one Sekler male. 207

Page 9 of 19
Accep
ted
Man
uscr
ipt
9
9. Discussion208
On examination of haplogroups with an N>1 frequency among both Hungarian-209
speaking European, and Altaic-speaking Central and Inner Asian populations, we showed that 210
the possible maximum Central/Inner Asian admixture among contemporary Hungarian 211
populations ranges around 5 -7.4%. We took into account only those haplotypes which could 212
derive from Central/Inner Asian haplotypes according to the MJ-networks. The admixture 213
was lowest among Hungarians from Hungary (5.1%), while somewhat higher among 214
Hungarian-speaking populations in Romania, notably Sekler (7.4%) and Csango (6.3%). The 215
average of these results was 5.7% among 522 Hungarian-speaking males (see Table S5). The216
reason of the difference might be the long-time isolation of Sekler and Csango groups, 217
resulting in lower admixture from neighbouring populations. However, we also must 218
acknowledge that these numbers represent an upper limit and that actual Central and Inner 219
Asian admixture might be somewhat lower. In these admixture cases, the genetic links are not 220
necessarily directly from Altaic populations to Hungarians, as both populations may have 221
received these genetic markers from a common third unidentified source (e.g. Middle East, 222
Caucasus, and East Slavs). Because of this possibility, further research is needed. We also 223
have to note that Central Asian admixture among Hungarians does not necessarily come from 224
Altaic-speakers. It may also come from ancient Iranian tribes who were later Turkicized by 225
Altaic conquerors. The main haplogroups responsible for the Central/Inner Asian admixture 226
among Hungarians are J2-M172 (xM47, M67, L24, M12), J2-L24, R1a-Z93, Q-M242 and E-227
M78.228
Earlier studies reported that Haplogroup E, J and their main subgroups spread from the 229
Middle East with the Neolithic agricultural revolution [27, 28, 29]. It spread towards both 230
Europe and Central Asia, thus some of the common haplotypes such as E-M78, J2-M172* 231
and J2-L24 may indicate a common Middle Eastern origin for both Hungarian-speaking and 232

Page 10 of 19
Accep
ted
Man
uscr
ipt
10
Central/Inner Asian samples. This is in contrast to the idea of a male migration from Asia 233
towards the Carpathian Basin.234
Based on the results of the Central and Inner Asian samples analysed in this study, we 235
could not find a strong genetic relationship with contemporary Hungarian-speaking 236
populations which would imply a common origin for these populations in the past. 237
Hungarians were heavily affected by neighbouring populations and also had effect on them. 238
We should also take into account that the East Central European region received Central and 239
Inner Asian genetic influence both before (Sarmatians, Huns, Avars, Onogur-Bulgars) and 240
after (Pechenegs, Yassic people, Cumans) the Hungarian settlement in the Carpathian Basin241
[3]. It is impossible to separate the genetic effects of these different migrations based on DNA 242
results from contemporary populations. The Central Asian gene pool also underwent 243
significant changes due to medieval Turkic and Mongolian invasions as well [3].244
Assuming a Central Asian origin for Eastern European subgroups of haplogroup R1a-245
M198, the share of Central Asian ancestry would significantly increase, up to almost 30%. 246
But then neighbouring Western and Eastern Slavic peoples would have an even higher Central 247
Asian admixture (50-60%) than that of Hungarians.248
Despite the similarity of tribal names among Kazakh and Uzbek Madjars, a significant 249
genetic connection could not be established; this based on our meticulously-selected samples, 250
which included pedigree analyses. Kazakh Madjars (dominated by Hg G1) differ significantly 251
from other Kazakhs and do not exhibit a relationship with Caucasian peoples (Hg G2). Uzbek 252
Madjars are more heterogeneous, although still dominated by Hgs C3 and D, which are 253
entirely absent from present-day Hungarian speakers. 254
Comprehensive surveys of more Central/Inner Asian and less-admixed population 255
groups in Hungary, including pedigree analyses, for deep-resolution haplogroups need to be 256
conducted in future studies to be able to draw more robust conclusions regarding the origins, 257
spread and genetic affiliations of contemporary populations.258

Page 11 of 19
Accep
ted
Man
uscr
ipt
11
Earlier studies have highlighted the very low genetic affinity of present-day 259
Hungarians with linguistically related Uralic peoples [16]. A recently published study showed 260
a limited genetic link between Hungarians and Ugric-speaking, Western Siberian Mansi based 261
on the new subhaplogroup N-L1034 defined by L1034 SNP mutation [30]. The typically262
Uralic haplogroup N accounted for only 1.7% among the Hungarian samples, this being even 263
lower than a potential Turkic admixture.264
The conclusion of this study is that present-day Hungarian speakers are genetically 265
very similar to neighbouring populations, isolated Hungarian speaking groups having266
relatively higher presence of Central and Inner Asian genetic elements. However, we could 267
not show any significant genetic correlation between Hungarian-speaking and Central/Inner 268
Asian samples which would explain the linguistic difference among Hungarians and 269
neighbouring populations. At the same time, the reliable historical and genetic conclusions 270
require an extension of the study to a significantly larger database with deep haplogroup 271
resolution, including ancient DNA data. 272
Conflict of interest273
The authors declare no conflict of interest.274
Acknowledgements275
We would like to say special thanks to Dr. Eva Susa (General Director of the Network of 276
Forensic Science Institutes) for her financial support. We thank sample donors and Betty-Jean 277
Sigethy and Rayn Hoyt for the English editing. We say special thanks to two unknown 278
reviewers for their constructive comments and suggestions.279
References280
[1] T. Tóth, A honfoglaló magyarság ethnogenezisének problemája, Anthropologiai 281
Közlemények. IX./ 4 (1965) 139-150.282
[2] K. Éry, Regionalis különbségek a magyarság X. századi embertani anyagában, 283
Anthropologiai Közlemények XXII. (1978) 77-86.284

Page 12 of 19
Accep
ted
Man
uscr
ipt
12
[3] P. Liptak, Magyar etnogenezis, In: Embertan és emberszármazástan, Budapest. 285
Tankönyvkiadó (1979) p 317-319.286
[4] I. Fodor, On the Problem of the Influence of Iranian Art upon Hungarian Art in the 287
Conquest Period, 10 th Century, Archeologiai Értesítő 100 (1973) p 32-41.288
[5] I. Fodor, Connections Between Early Turkish and Hungarian Art, In: Proseedings of the 9 289
th International Congress of Turkish Art. vol. II, Ankara (1995) p 133-140.290
[6] I. Fodor, The Ethnic Identity of the Early Hungarians and the Legend of the White Stag, 291
In:Csodaszarvas, vol. II. Molnár Kiadó, Budapest (2006) p 9-37.292
[7] A. Coq von Le, Bilderatla s zur Kunst- und Kulturgeschichte Mittel-Asiens (1925), Berlin293
[8] Плетнёва С.А.: От кочевий к городам. Салтово-Маяцкая культура. Материалы и 294
Исследования по археологии СССР 142. Москва (1967).295
[9] I. Dienes, A honfoglaló magyarok és ősi hiedelmeik, In: Urali népek, Budapest (1975) p 296
77-108.297
[10] I. Fodor, On Magyar-Bulgar-Turkish Contacts, In: Chuvash Studies. Ed: Róna-Tas A, 298
Budapest (1982) p 45-81.299
[11] A. Brandstätter, B. Egyed, B. Zimmermann, N. Duftner, Z. Padar, W. Parson, Migration 300
rates and genetic structure of two Hungarian ethnic groups in Transylvania, Romania, Ann 301
Hum Genet 71 (2007) 791-803.302
[12] A. Völgyi, A. Zalán, E. Szvetnik, H. Pamjav, Hungarian population data for 11 Y-STR 303
and 49 Y-SNP markers, Forensic Sci Int Genet 3 (2009) 27-28.304
[13] H. Pamjav, T. Fehér, E. Németh, Z. Pádár, New Y-Chromosome Binary Markers 305
Improve Phylogenetic Resolution Within Haplogroup R1a1, Am J Phys Anthropol (2012) 149 306
(4):611-615.307
[14] P.A. Underhill, G.D. Poznik, S. Rootsi, M. Järve, A.A. Lin, J. Wang, B. Passarelli, J. 308
Kanbar, N.M. Myres, R.J. King, J. Di Cristofaro, H. Sahakyan, D.M. Behar, A. 309
Kushniarevich, J. Sarac, T. Saric, P. Rudan, A.K. Pathak, G. Chaubey, V. Grugni, O. Semino, 310

Page 13 of 19
Accep
ted
Man
uscr
ipt
13
L. Yepiskoposyan, A. Bahmanimehr, S. Farjadian, O. Balanovsky, E.K. Khusnutdinova, R.J. 311
Herrera, J. Chiaroni, C.D. Bustamante, S.R. Quake, T. Kivisild, R. Villems, The phylogenetic 312
and geographic structure of Y-chromosome haplogroup R1a, Eur J Hum Genet (2014) doi: 313
10.1038/ejhg.2014.50.314
[15] Z.H Rosser, T. Zerjal, M.E. Hurles, M. Adojaan, D. Alavantic, A. Amorim, W. Amos, 315
M. Armenteros, E. Arroyo, G. Barbujani, G. Beckman, L. Beckman, J. Bertranpetit, E. Bosch, 316
D.G. Bradley, G. Brede, G. Cooper, H.B. Côrte-Real, P. de Knijff, R. Decorte, Y.E. Dubrova, 317
O. Evgrafov, A. Gilissen, S. Glisic, M. Gölge, E.W. Hill, A. Jeziorowska, L. Kalaydjieva, M. 318
Kayser, T. Kivisild, S.A. Kravchenko, A. Krumina, V. Kucinskas, J. Lavinha, L.A. Livshits, 319
P. Malaspina, S. Maria, K. McElreavey, T.A. Meitinger, A.V. Mikelsaar, R.J. Mitchell, K. 320
Nafa, J. Nicholson, S. Nørby, A. Pandya, J. Parik, P.C. Patsalis, L. Pereira, B. Peterlin, G. 321
Pielberg, M.J. Prata, C. Previderé, L. Roewer, S. Rootsi, D.C. Rubinsztein, J. Saillard, F.R. 322
Santos, G. Stefanescu, B.C. Sykes, A. Tolun, R. Villems, C. Tyler-Smith, M.A. Jobling Y-323
chromosomal diversity in Europe is clinal and influenced primarily by geography, rather than 324
by language, Am J Hum Genet (2000) 67:1526-1543.325
[16] K. Tambets, S. Rootsi, T. Kivisild, H. Help, P. Serk, E.L. Loogväli, H.V. Tolk, M. 326
Reidla, E. Metspalu, L. Pliss, O. Balanovsky, A. Pshenichnov, E. Balanovska, M. Gubina, S. 327
Zhadanov, L. Osipova, L. Damba, M. Voevoda, I. Kutuev, M. Bermisheva, E. 328
Khusnutdinova, V. Gusar, E. Grechanina, J. Parik, E. Pennarun, C. Richard, A. Chaventre, 329
J.P. Moisan, L. Barác, M. Pericić, P. Rudan, R. Terzić, I. Mikerezi, A. Krumina, V. 330
Baumanis, S. Koziel, O. Rickards, G.F. De Stefano, N. Anagnou, K.I. Pappa, E. 331
Michalodimitrakis, V. Ferák, S. Füredi, R. Komel, L. Beckman, R. Villems. The western and 332
eastern roots of the Saami-the story of genetic "outliers" told by mitochondrial DNA and Y 333
chromosomes, Am J Hum Genet (2004) 74(4):661-682.334

Page 14 of 19
Accep
ted
Man
uscr
ipt
14
[17] A.Z. Bíró, A. Zalán, A. Völgyi, H. Pamjav, A Y-chromosomal comparison of the 335
Madjars (Kazakhstan) and the Magyars (Hungary), Am J Phys Anthropol (2009) 139(3): 305-336
310.337
[18] B. Egyed, S. Füredi, Z. Padar, Population genetic study in two Transylvanian populations 338
using forensically informative autosomal and Y-chromosomal STR markers, Forensic Sci Int 339
(2006)164(2-3):257-65.340
[19] Pamjav H, Zalán A, Béres J, Nagy M, Chang YM (2011) Genetic structure of the 341
paternal lineage of the Roma people. Am J Phys Anthropol.145(1):21-29.342
[20] M. van Oven, K. Toscani, N. van den Tempel, A. Ralf, M. Kayser, Multiplex 343
genotyping assays for fine-resolution subtyping of the major human Y-chromosome 344
haplogroups E, G, I, J, and R in anthropological, genealogical, and forensic investigations,345
Electrophoresis (2013) 34(20-21):3029-3038.346
[21] H.J. Bandelt, P. Forster, A. Röhl Median-joining networks for inferring intraspecific 347
phylogenie, Mol Biol Evol (1999)16: 37-48.348
[22] L.A. Zhivotovsky, P.A. Underhill , C. Cinnioğlu, M. Kayser, B. Morar ,T. Kivisild, R. 349
Scozzari, F. Cruciani, G. Destro-Bisol, G. Spedini, G.K. Chambers, R.J. Herrera , K.K. Yong, 350
D. Gresham, I. Tournev, M.W. Feldman, L. Kalaydjieva, The effective mutation rate at Y 351
chromosome short tandem repeats, with application to human population-divergence time., 352
Am J Hum Genet. 74 (2004) 50-61.353
[23] V. Grugni, V. Battaglia, B. Hooshiar Kashani, S. Parolo, N. Al-Zahery, A. Achilli, A. 354
Olivieri, F. Gandini, M. Houshmand, M.H. Sanati, A. Torroni, O. Semino, Ancient Migratory 355
Events in the Middle East: New Clues from the YChromosome Variation of Modern Iranians,356
PLoS ONE (2012) 7(7) e41252. doi:10.1371/journal.pone.0041252.357
[24] J. Di Cristofaro, E. Pennarun, S. Mazières, N.M. Myres, A.A. Lin, S.A. Temori, M. 358
Metspalu, E. Metspalu, M. Witzel, R.J. King, P.A. Underhill, R. Villems, J. Chiaroni, Afghan 359

Page 15 of 19
Accep
ted
Man
uscr
ipt
15
Hindu Kush: where Eurasian sub-continent gene flows converge, PLoS One (2013) 360
8(10):e76748. doi: 10.1371/journal.pone.0076748.361
[25] P.A. Underhill, N.M. Myres, S. Rootsi, M. Metspalu, L.A. Zhivotovsky, R.J. King, A.A. 362
Lin, C.E. Chow, O. Semino, V. Battaglia, I. Kutuev, M. Järve, G. Chaubey, Q. Ayub, A. 363
Mohyuddin, S.Q. Mehdi, S. Sengupta, E.I. Rogaev, E.K. Khusnutdinova, A. Pshenichnov, O. 364
Balanovsky, E. Balanovska, N. Jeran, D.H. Augustin, M. Baldovic, R.J. Herrera, K. 365
Thangaraj, V. Singh, L. Singh, P. Majumder, P. Rudan, D. Primorac, R. Villems, T. Kivisild, 366
Separating the post-Glacial coancestry of European and Asian Y chromosomes within 367
haplogroup R1a, Eur J Hum Genet (2010)18(4):479-484.368
[26] N.M. Myres, S. Rootsi, A.A. Lin, M. Järve, R.J. King, I. Kutuev, V.M. Cabrera, E.K. 369
Khusnutdinova, A. Pshenichnov, B. Yunusbayev, O. Balanovsky, E. Balanovska, P. Rudan, 370
M. Baldovic, R.J. Herrera, J. Chiaroni, J. Di Cristofaro, R. Villems, T. Kivisild, P.A. 371
Underhill, A major Y-chromosome haplogroup R1b Holocene era founder effect in Central 372
and Western Europe, Eur J Hum Genet (2011) 19(1):95-101.373
[27] F. Di Giacomo, F. Luca, L. O. Popa, N. Akar, N. Anagnou, J. Banyko, R. Brdicka, G. 374
Barbujani, F. Papola, G. Ciavarella, F. Cucci, L. Di Stasi, L. Gavrila, M. G. Kerimova, D. 375
Kovatchev, A. I. Kozlov, A. Loutradis, V. Mandarino, C. Mammi, E. N. Michalodimitrakis, 376
G. Paoli, K. I. Pappa, G. Pedicini, L. Terrenato, S. Tofanelli, P. Malaspina, A. Novelletto, Y 377
chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe, Hum 378
Genet (2004) 115: 357–371.379
[28] O. Semino, Ch. Magri, G. Benuzzi, A. A. Lin, N. Al-Zahery, V. Battaglia, L. Maccioni, 380
C. Triantaphyllidis, P. Shen, P. J. Oefner, L. A. Zhivotovsky, R. King, A. Torroni, L. L. 381
Cavalli-Sforza, P. A. Underhill, and A. S. Santachiara-Benerecetti, Origin, Diffusion, and 382
Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of 383
Europe and Later Migratory Events in the Mediterranean Area, Am J Hum Genet (2004) 74 384
(5):1023–1034.385

Page 16 of 19
Accep
ted
Man
uscr
ipt
16
[29] F. Cruciani, R. La Fratta, P. Santolamazza, D. Sellitto, R. Pascone, P- Moral, E. Watson, 386
V. Guida, E. Beraud Colomb, B. Zaharova, J. Lavinha, G. Vona, R. Aman, F. Calí, N. Akar, 387
M. Richards, A. Torroni, A. Novelletto, and R. Scozzari, Phylogeographic Analysis of 388
Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and 389
Out Of Africa, Am J Hum Genet (2004). 74:1014–1022.390
[30] T. Fehér, E. Németh, A. Vándor, I. V. Kornienko, L. K. Csáji and H. Pamjav, Y-SNP 391
L1034: Limited genetic link between Mansi and Hungarian-speaking populations, Mol Genet 392
Genomics DOI 10.1007/s00438-014-0925-2.393
394

Page 17 of 19
Accep
ted
Man
uscr
ipt
17
Fig.1. A phylogenetic tree of the tested 53 Y-SNP loci394
395396
Fig.2.397
Median joining (MJ) networks of the Hungarian speaking, Central and Inner Asian 398
populations compared399
A. MJ network of Y-STRs within E-M78 haplogroup for the populations compared400
B. MJ network of Y-STRs within G2a-P15 haplogroup for the populations compared401
C. MJ network of Y-STRs within J2*-M172 haplogroup for the populations compared402
D. MJ network of Y-STRs within J2-L24 haplogroup for the populations compared403
E. MJ network of Y-STRs within N1c-Tat haplogroup for the populations compared404
F. MJ network of Y-STRs within Q-M242 haplogroup for the populations compared405
G. MJ network of Y-STRs within R1a-M458 haplogroup for the populations compared406
H. Median-joining network of Y-STRs within R1a-Z93 haplogroup for the populations 407
compared408
I. MJ network of Y-STRs within R1b-P25 haplogroup for the populations compared409
410The circle sizes are proportional to the haplotype frequencies. The smallest area is equivalent 411
to one individual.412
413414
Page 18 of 19
Accep
ted
Man
uscr
ipt
Figure 1
Page 19 of 19
Accep
ted
Man
uscr
ipt
Figure 2





























