Embed Size (px)
Citation preview

Plant Physiol. (1983) 73, 94-990032-0889/83/73/0094/06/$00.50/0
Terpenoid Metabolism in Plastids1ISOLATION AND BIOCHEMICAL CHARACTERISTICS OF CAPSICUM ANNUUM CHROMOPLASTS
Received for publication September 23, 1982 and in revised form May 16, 1983
BILAL CAMARA, FRANgOISE BARDAT, ODETTE DOGBO, JUDY BRANGEON2, AND RENt MONEGERLaboratoire de Regulations Metaboliques et Diffe'renciation des Plastes, Universite Pierre et Marie Curie(Paris 6), 75230 Paris Cedex 05, France
ABSTRACI
A technique for the isolation and the purification of Capsicum annumL. var. Yolo Wonder chromoplasts is described. The degree of purity ofthe isolated chromoplasts is greatly improved by the absence of MgCk2in the extraction medium and in the gradient purification system, asshown by electron micrographs and the near absence of antimycin-insensitive NADH:cytochrome c reductase activity and succi-nate:cytochrome c reductase activity. Furthermore, phosphatidylserinewas absent and the phosphatidylethanolamine content was reduced by afactor of 5 in these preparations, which were active in the synthesis ofgalactolipids, prenylquinones, and carotenoids.
The differentiation of various types of plastids from the samepre-existing form (23, 27) involves complex physiological proc-esses in which the biogenesis and assembly of plastid terpenoidsplay an important role. The metabolism of these plastid com-ponents could be used as a biochemical parameter for develop-mental and functional studies. A prerequisite for this type ofinvestigation is a suitable model, as well as appropriate isolationand biochemical characterization of plastids. Capsicum fruitsprovide a useful model in that the chloroplast transforms into achromoplast during ripening. The latter organelle is the site ofactive terpenoid metabolism, including especially the synthesisof a wide range of carotenoids. In this paper, we describe amethod of isolating intact Capsicum fruit chromoplasts. Thelipid and carotenoid composition and related enzymic activitiesare reported.
MATERIALS AND METHODS
Radiochemicals. Uridine diphospho-D-[U-14C]galactose (340Ci/mol), 3-hydroxy-3-methyl[3-14C]glutaryl-CoA (26.2 Ci/mol),S-adenosyl-L-[methyl-'4C]methionine (58 Ci/mol) and [1-'4C]isopentenyl pyrophosphate (56 Ci/mol) were purchased fromAmersham.
Plant Material. Fruits of Capsicum annuum L. (var. YoloWonder) semiripened (orange) were used.
Isolation of Chromoplasts. The pericarp tissue (2 kg freshweight) ofthe fruit was shredded and infiltrated with the isolationmedium at 4C. Unless otherwise stated, the isolation mediumcontained: 10 mM MgCl2, 1 mM j3-mercaptoethanol, 1 mM
I Supported by grants from the Direction de la Recherche Universitaireand from the Centre National de la Recherche Scientifique (Al 03.1269).
2Presnt address: Laboratoire de Structure et Metabolisme des Plantes,Bit. 490, F. 91405 Orsay, France.
EDTA, 0.4 M sucrose, and 50 mm Tris-HCI (pH 8). The materialwas homogenized in a blender for 2 x 3 s and then filteredthrough four layers of Blutex (50 ,umesh wide). The cellulardebris was eliminated by 5 min of centrifugation at 150g in aprecooled Sorvall RC 2B equipped with a GSA rotor. The crudechromoplast fraction was pelleted by a 10-min centrifugation ofthe supematant at 2,000g. The sucrose density gradient usedpreviously for chromoplast isolation (7) was modified as de-scribed below in order to maximize chromoplast yield. The crudechromoplast suspension (2 ml) was layered on the top of acellulose nitrate tube containing the discontinuous gradient (9ml for each sucrose concentration), 1 mM MgC12 (unless other-wise stated), 1 mm ,3-mercaptoethanol, 0.46, 0.84, and 1.45 Msucrose buffered with 50 mm Tris-HCl (pH 7.6). After 1-hcentrifugation at 62,000g (Rmul,), in a precooled Beckman L3-50equipped with a SW 27 rotor, the chromoplasts banded at the0.46 to 0.84 M and 0.84 to 1.45 M interfaces. The two chloroplastbands were collected separately and used directly or after dilution(1 ml of chromoplast suspension + 1 ml of 50 mm Tris) andcentrifugation at 4,000g for 5 min. The chromoplasts werehomogenized in a tight fitting Potter-Elvehjem homogenizerbefore incubation.
Analytical Methods. Lipids, a-Tocopherol, and Plastoquinone-9 Analysis. The chromoplast suspension was extracted accordingto the technique described by Bligh and Dyer (2). The lipidextract was separated on a DEAE-cellulose column (9). Thecolumn was eluted successively with: chloroform, fraction I;chloroform:methanol (7:3, v/v), fraction II; chloroform:methanol (2:1, v/v) containing 0.05 M ammonium acetate and 2ml of concentrated NH40H/100 ml, fraction III.
Fraction II was chromatographed on Silica Gel G plates de-veloped with acetone:benzene:water (91:30:8, v/v/v) accordingto Pohl et al. (22). Fraction III was separated by two-dimensionalSilica Gel G TLC using chloroform:methanol:concentratedNH4OH (140:50:7, v/v/v) for the first dimension, and chloro-form:methanol:acetone:glacial acetic acid:water (100:20:40:20:10, v/v/v/v/v) for the second dimension. The separated lipidswere detected by spraying the plates with primuline (29) andidentified as previously described (9). Quantitative determinationwas carried out according to Kalra and Brooks (12) after analysisof the fatty acid methyl esters (9). The following mol wt wereused in the calculations: MGDG3 = 770; DGDG = 928; SL =
3 Abbreviations: MGDG, monogalactosyldiglyceride; DGDG, digalac-tosyldiglyceride; SL, sulfoquinovosyldiglyceride; PC, phosphatidylcho-line; PE, phosphatidylethanolamine; PI, phosphatidylinositol; PS, phos-phatidylserine; PG, phosphatidylglycerol; PA, phosphatidic acid; HMG-CoA, 3-hydroxy-3-methylglutaryl coenzyme A; PPO, 2,5-diphenyloxa-zole; dimethyl-POPOP, 1,4-bis[2-(4-methyl-5-phenyloxazolyl)]benzene;MVA, mevalonate; MVAP, mevalonate phosphate; MVAPP, mevalon-ate pyrophosphate.
94 www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1983 American Society of Plant Biologists. All rights reserved.
PLASTID TERPENOID METABOLISM
i
NW
2.I e~
B, %
I
.. tvT."
SI a-i .4 d4-_|~~~~~~~-d a
- 4*9.^
3m
'.1.... s
R.
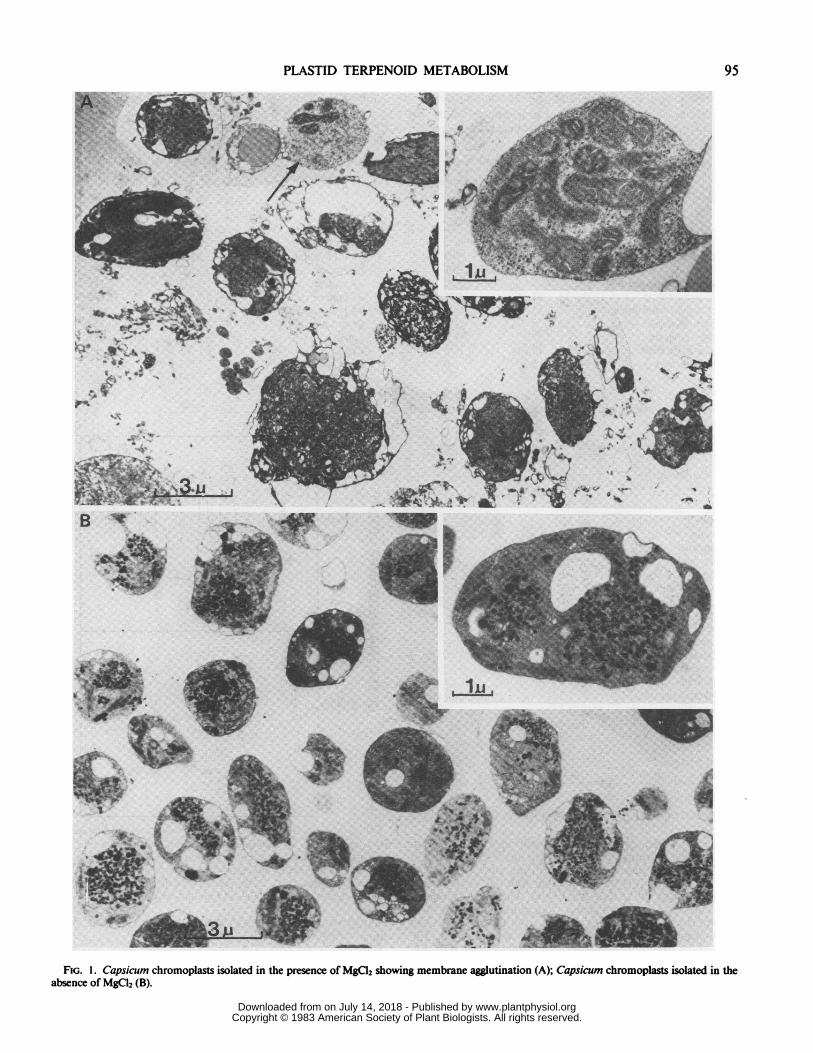
FIG. 1. Capsicum chromoplasts isolated in the presence of MgCl2 showing membrane agglutination (A); Capsicum chromoplasts isolated in theabsence of MgCI2 (B).
p.
.^s-I~~~~~~~~,I. ;
, _ r * * # |i,
t .4
v~1,P,
,a1
95
%. .1 41. '46
w ~~~~-&L'. 10..
.00i6 .,.
.4
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1983 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 73, 1983
Table I. Lipid Content ofIsolated Capsicum ChromoplastsThe chromoplasts were purified by the same method (see the text)
except that 10 mM MgC2 was added in the medium in one case beforeextraction and purification.
Lipid Content
Lipid Molecule&s Chromoplasts Chromoplastsprepared withoutd
with MgCl2 without
Ag mg' proteinMonogalactosyldiglyceride 85 103Digalactosyldiglyceride 76 102Sulfoquinovosyldiglyceride 17 16Phosphatidylcholine 43 26Phosphatidylethanolamine 25 5Phosphatidylinositol 9 5Phosphatidylserine 1 0Phosphatidylglycerol 13 6Phosphatidic acid 6 0
Total lipid 275 263a Sterylglycoside and neutral lipids were detected, but their content
was not determined.
Table II. Carotenoid Composition ofCapsicum ChromoplastsConfiguration
Carotenoid Compositioncis trans
Phytoene 3.5 99.1 0.2Phytofluene 1.5 86.0 14.0{-Carotene 2.0 100a-Carotene tr,-Carotene 7.0,3-Cryptoxanthin 6.2Cryptocapsin 3.2Lutein 2.6Lutein epoxide 3.4Zeaxanthin 5.2Antheraxanthin 3.8Violaxanthin 8.5Capsanthin 35.3Capsanthin epoxide 1.8Capsorubin 9.3Neoxanthin 5.3Mutatoxanthin 1.4
Total carotenoid(Mg mg-' protein) 156
a tr, trace.
915; PC = 770; PE = 726; PI = 834; PS = 775; PG = 734; PA= 684.
Fraction I contained the bulk of a-tocopherol and plastoqui-none-9, which were analyzed after a preliminary separation on aSilica Gel G plate developed with petroleum ether, 40 to 50C,and ether (10:1, v/v) (15), and detected with 0.05% (w/v) rho-damine 6G in acetone:H20 (95:5, v/v). The plastoquinone-9band and the a-tocopherol band were eluted separately withacetone. The plastoquinone-9 band was purified by a two-stepmigration in carbon tetrachloride, on a Silica Gel G plate. TheAE 1%/I cm (ox-Red) 220 between the oxidized form (Ox) andthe reduced form (Red) was used for quantitative determination.The a-tocopherol band was purified on a Silica Gel G platedeveloped with chloroform:carbon tetrachloride (3:2, v/v). The
Table III. Incorporation of[1-'4CJlsopentenyl Pyrophosphate intoCarotenoids by Isolated Capsicum Chromoplasts
The chromoplasts were purified as described in Figure I and incubatedwith [1-'4C]isopentenyl pyrophosphate (see "Materials and Methods").
Radioactivity IncorporatedCarotenoid at Incubation Times
I h 5h
dpmPhytoene 45,000 27,000cis-Phytofluene 22,000 16,000trans-Phytofluene 2,850 10,300{-Carotene 32,500 75,000a-Carotene 250 375,8-Carotene 15,300 32,740#-Cryptoxanthin 275 850Cryptocapsin 150 220Lutein 62 57Zeaxanthin 85 115Antheraxanthin 102 135Violaxanthin 140 320Capsanthin 34 163Capsorubin 28 87
4000
a. 20000
wC,)zo 0a.CDw
0 40000
ILL
D 2000
0
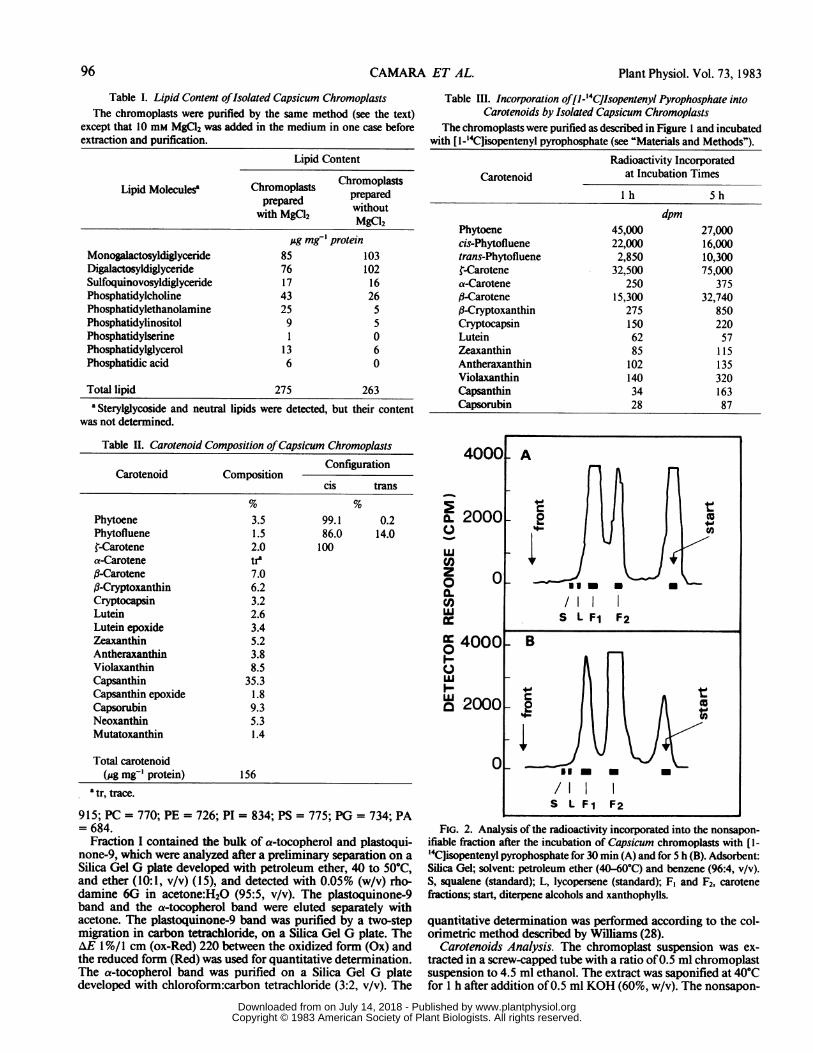
FIG. 2. Analysis of the radioactivity incorporated into the nonsapon-ifiable fraction after the incubation of Capsicum chromoplasts with [1-'4C]isopentenyl pyrophosphate for 30 min (A) and for 5 h (B). Adsorbent:Silica Gel; solvent: petroleum ether (40-60°C) and benzene (96:4, v/v).S, squalene (standard); L, lycopersene (standard); F, and F2, carotenefractions; start, diterpene alcohols and xanthophylls.
quantitative determination was performed according to the col-orimetric method described by Williams (28).
Carotenoids Analysis. The chromoplast suspension was ex-tracted in a screw-capped tube with a ratio of0.5 ml chromoplastsuspension to 4.5 ml ethanol. The extract was saponified at 40Cfor 1 h after addition of 0.5 ml KOH (60%, w/v). The nonsapon-
A
a t-
.
0
-; I 00~/IlS L FI F2
B
S L Fl F2
96 CAMARA ET AL.
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1983 American Society of Plant Biologists. All rights reserved.

PLASTID TERPENOID METABOLISM
ifiable material was extracted with peroxide-free ether, dried withNaCl, and applied to Silica Gel G plates previously adjusted topH 7 (to avoid isomerization of the carotenoid epoxy groups).The tanks were placed at 0 to 4°C and the development wasdone with petroleum ether (40-60°C) and acetone (60:40, v/v).This system allows the separation of carotenoids into differentfractions with increasing polarity: carotenes, monohydroxyxan-thophylls, monohydroxymonoketoxanthophylls, dihydroxyxan-thophylls, dihydroxymonoepoxyxanthophylls, dihydroxydike-toxanthophylls, and trihydroxyxanthophylls. The carotenes werefurther purified as described previously (3, 10). Each xanthophyllfraction was chromatographed on magnesium oxide:Kieselgur(1:1, w/w) plates developed with petroleum ether:acetone (60:40,v/v). A further purification of the xanthophylls was achieved onSilica Gel G plates with ethyl acetate:carbon tetrachloride (70:30,v/v).The quantitative determination was done according to Davies
(1 1). For total carotenoids, an average coefficient ofE 1%/1 cm= 2200 in benzene was adopted.
Pyridine Nucleotides. The determination of pyridine nucleo-tides was carried out according to the procedure described byMoneger et al. (20).
Total protein was assayed by the method of Lowry et al. (19)after delipidation of the chromoplast suspension by extractionwith 4 volumes of acetone:ethanol (2:1, v/v) for 12 h at -20°C.
Reaction Mixtures for Subcellular Experiments. GalactolipidSynthesis. The reaction was performed at 25°C for various tissues.The reaction mixture contained: 0.13 M sorbitol, 50 mM Tris-HCI (pH 8), 5 mM MgCl2, 0,2 Mm UDP-[U-'4C]galactose (0.096MCi), and a chromoplast preparation (4.5 mg protein). The totalincubation volume was 1 ml for each tube. The reaction wasstopped by the addition of 2 ml of chloroform:methanol (2:1, v/v). The lipid extract was analyzed by procedures described forfraction II (above). The radioactivity of the lipid spots wasdetected by autoradiography on Kodirex films. The radioactivitywas eluted and determined with a toluene-based scintillationfluid, 3 g of PPO, and 0.5 g ofdimethyl-POPOP in 1 L oftoluene.
a-TocopherolSynthesis. The assay was based on the cyclizationof 2,3-dimethyl-5-phytylquinol to y-tocopherol followed bymethylation of the latter to yield a-tocopherol. The dimethyl-phenol used in the preparation of the dimethylphytylquinol wasoxidized and reduced with alkaline sodium dithionite solution(6). The resulting product was crystallized in ethanol. The di-methylquinol was condensed with isophytol, and the resultingproduct was purified and quantitated according to Camara et al.(6). The enzymic reaction was carried out at 25°C for differenttimes in an incubation medium (2.5 ml for each tube) containing0.13 M sorbitol, 50 mm Tris-HCl (pH 8), 100 gM 2,3-dimethyl-5-phytylquinol, 8 uM S-adenoSyl-L-methyl['4C]methionine (1.17,uCi), and a chromoplast preparation containing 4.8 mg of pro-tein. The reaction was terminated with 5 ml of chloro-form:methanol (2:1, v/v). The a-tocopherol was extracted andpurified as described above. The radioactivity was determined asfor other lipids (see above).Assayfor HMG-CoA Reductase. The reaction was carried out
at 300C for various times. The reaction medium contained: 10mM sorbitol, 50 mm Tris-HCl pH 7.6, 0.5% BSA, 10 mM DTT,4 mM NADPH, 19,uM [3-'4C]HMG-CoA (0.5 MCi), and a chrom-oplast preparation containing 4.5 mg of protein. The final incu-bation volume was 1 ml. The reaction was stopped by additionof 2 ml of 2 N HCI. The protein precipitate was centrifuged andthe supernatant was incubated at 37°C for 30 min to allowconversion of MVA to the lactone. An aliquot was chromato-graphed in a disposable pipette filled with Dowex 1-X8-100 resinprepared in the formate form. The elution of mevalonolactonewas performed with H20. An aliquot was counted using the
g), ethanol (300 ml), and toluene (1 L). The counting efficiencywas 80%.
Incorporation ofIsopentenyl Pyrophosphate into Carotenoids.The reaction was carried out at 25C in a medium containing:0.4 M sucrose, 50 mM Tris-HCI (pH 8), 10 mM DTT, 1.25 mMNADP, 1.25 mm FAD, 15 mM MgCl2, 5 mM MnCl2, 3 MM [1-
'4C]isopentenyl pyrophosphate (0.34 MCi), and a chromoplastpreparation containing 2.4 mg of protein. The final incubationvolume was 2 ml. The reaction was stopped by addition of 4 mlof ethanol. The carotenoids were extracted and analyzed aftersaponification as described above.
RESULTS AND DISCUSSION
Characteristics of Purified Chromoplasts. After centrifugation,two chromoplast bands were obtained: band A at the 0.46 to0.84 M interface and band B at the 0.84 to 1.45 M interface.Microsomal and mitochondrial contaminations were barely de-tectable in bands A and B as judged by antimycin-insensitiveNADH:Cyt c reductase (A: 5 nmol Cyt c reduced min -' mg-'protein; B: 3 nmol Cyt c reduced min-' mg-' protein) andsuccinate:Cyt c reductase (A: traces; B: traces). The two bandswere devoid of squalene synthetase activity when incubated withisopentenyl pyrophosphate. Phytoene synthetase was detected inband B and barely detectable in band A (results not published).Band A is probably enriched in broken chromoplasts which havelost their phytoene synthetase, since it has been shown recentlythat this enzyme is a soluble stromal protein (4, 5, 13). Therefore,only chromoplasts located at the 0.84 to 1.45 M interface (bandB) were used for the studies.A critical point we noted relevant to the isolation of chromo-
plasts was the effect of MgCl2 in the extraction medium and inthe gradient system. At the concentration used (10 mM), MgC12induced an agglutination of heterogeneous membranes (Fig. 1)which sometimes had the same sedimentation characteristics asisolated chromoplasts. When MgCl2 was excluded from theextraction medium and from the gradient, this agglutinationeffect was drastically reduced (Fig. 1). Similar results have beenobserved during plastid isolation (21, 24). The presence of MgCl2can lead to important changes in the apparent lipid composition(see below) ofthe isolated chromoplasts in relation to cytoplasmiccontamination. For this reason, MgCl2 was excluded from theextraction medium and the gradient system.
Lipid Composition and Galactosyltransferase Activities. Thelipid composition of the chromoplasts was qualitatively similarto that observed for chloroplasts, with galactolipids as the majorcomponent (Table I). The phospholipid content was more com-plex (Table I). Phosphatidylethanolamine and phosphatidylser-ine are not reported to be present in highly purified chloroplastpreparations, while they are present in chromoplasts so far ana-lyzed (16, 18). However, their presence is considered to representcytoplasmic contamination, and, in good agreement with this,we noted that phosphatidylserine disappeared while phosphati-dylethanolamine was reduced by a factor of 5 (Table I) in highlypurified chromoplasts obtained after elimination of MgCl2 fromthe extraction medium and from the gradient. When the chrom-oplasts were incubated with UDP-[U-'4C]galactose, the resultingradioactivity was distributed mainly between monogalactosyldi-glyceride and digalactosyldiglyceride. The galactosyltransferaseactivity was linear up to 20 min under the assay conditions: 0.4pmol min-' mg-' protein for monogalactosyldiglyceride; 0.04pmol min-' mg-' protein for digalactosyldiglyceride. A lowradioactivity was observed in the sterylglycoside (0.01 pmolmin-' mg-' protein); it could be due to an epimerase activity, orto a direct incorporation of galactose into the sterol moiety assuggested (17). The amount of radioactivity recovered in thesterylglycoside fraction after UDP-[U-'4C]Gal incorporation was
following scintillation cocktail: PPO (4 g), dimethyl-POPOP (0.1
97
lower than those observed for Narcissus (I 6, 17) and Tropaeolum www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from
Copyright © 1983 American Society of Plant Biologists. All rights reserved.

98 CAMARA ET AL.
(18) chromoplasts. The origin ofthe sterylglycoside in the chrom-oplast is unclear.
Plastoquinone-9 and a-Tocopherol Composition and Synthesis.The content of a-tocopherol ( 16 tg mg-' protein) is higher thanthat of plastoquinone-9 (5 ,ug mg-' protein). These results arecharacteristic for nonphotosynthetic tissues and corroborate thepreviously reported increase in a-tocopherol during the ripeningof Capsicum fruits ( 14).When isolated chromoplasts devoid of endogenous y-toco-
pherol are incubated in the presence of S-adenosyl-L-[methyl-'4C]methionine without addition of dimethylphytylquinol, theradioactivity incorporated into lipids is not located in a-toco-pherol (unpublished results). This may be explained by theabsence of endogenous dimethylphytylquinol and y-tocopherolin chromoplasts. Dimethylphytylquinol has been observed inyoung spinach leaves but not in matures leaves (26). Whendimethylphytylquinol was added to the incubation medium, thecyclization and the methylation reactions which are prerequisitesfor a-tocopherol formation were observed. The reaction waslinear up to 40 min. The activities (0.06 pmol min-' mg-'protein) of the preparations are higher than those reported forisolated spinach chloroplasts (26).
In the chromoplasts, the active synthesis and accumulation ofa-tocopherol, which may act as a protectant for membrane lipids,could consequently retard senescence. This is an important pointas chromoplasts contain highly unsaturated lipids.
Carotenoid Composition and Synthesis. A typical carotenoidcomposition of chromoplasts is shown in Table II. There arenumerous differences in comparison to chloroplasts, includingthe presence of phytoene and phytofluene in chromoplasts. Fur-thermore, configuration analysis revealed that phytoene is pres-ent in the cis configuration (99%) while phytofluene is presentin the cis (86%) and trans (14%) configurations; {-carotene ispresent in the trans configuration (Table II). The biogeneticimplications ofthese data are discussed elsewhere (10). Addition-ally, the epoxyxanthophyll content is qualitatively and quanti-tatively higher in chromoplasts than in chloroplasts and themajor carotenoids in chromoplasts are the ketoxanthophyllscapsanthin and capsorubin.The ability of chromoplasts to synthesize MVA, an early
precursor in carotenoid biosynthesis, was tested by examiningthe enzymic reduction of HMG-CoA to MVA. Under the incu-bation conditions, the reaction was linear for 10 min (31 pmolmin-' mg-' protein). It can be concluded that HMG-CoA reduc-tase is active in the Capsicum chromoplast. These data, and theactivation ofMVA to MVAP and MVAPP previously reported(8), are of importance if one considers the high carotenoidcontent and the probably high carotenoid biosynthetic capacityof the chromoplast. The autonomy of the chromoplast revealedby this study does not exclude a possible cooperative relationbetween the chromoplast and the ER, which is known to inter-vene in MVA formation in plant cells. Schneider et al. (25) notedthat during chloroplast-chromoplast transformation the plastidenvelope showed an increased permeability to mevalonate.When chromoplasts are incubated with [1-'4C]IPP, most of
the radioactivity incorporated into carotenoids is found in thecarotenes (Table III); however, the carotenoids with an a ring(a-carotene and lutein) are poorly labeled. Radioactivity in thexanthophylls did not increase significantly by increasing theincubation period from 1 to 5 h. Possible explanations for thisphenomenon will be discussed elsewhere.Under our experimental conditions, no radioactivity was de-
tected in squalene or lycopersene (Fig. 2) in spite of the presenceof endogenous reduced pyridine nucleotides (NADPH, 57 pmolmg' protein; NADH, 1305 pmol mg:' protein). The radioactiv-ity is associated with carotenes, and with diterpene alcohols andxanthophylls which stay at the origin (Fig. 2). Therefore, lyco-
Plant Physiol. Vol. 73, 1983
persene is not synthesized by purified pepper chromoplasts andit is probably not involved in the synthesis of carotenes in thissystem. We conclude that, under our experimental conditions,phytoene rather than lycopersene is the first C40 precursor ofcarotenoids.
Finally, this article describes a technique for isolating intactCapsicum chromoplasts free from cytoplasmic contamination asshown by the absence ofphosphatidylserine and barely detectableamounts of phosphatidylethanolamine in the purified chromo-plast preparations. Moreover, it reports some preliminary studieson the enzymic activities of these plant organelles. These prepa-rations exhibit galactosyltransferase activity and are active in a-tocopherol synthesis from exogenous precursors. They performthe activation of MVA to MVAP and MVAPP, and the incor-poration of IPP into carotenoids, but not into detectable levelsof squalene or lycopersene. The results obtained, compared tothose described for chloroplasts or flower chromoplasts (1), showthat Capsicum chromoplasts are a useful system for the study ofterpenoid metabolism.
Acknowledgments-We thank C. Agnes and A. Nar.ot-Garcia for skilledtechnical assistance.
LITERATURE CITED
1. BEYER P, K KREUZ, H KLEINIG 1980 j3-Carotene synthesis in isolated chrom-oplasts from Narcissus pseudonarcissus. Planta 150: 435438
2. BLIGH EG, WJ DYER 1959 A rapid method of total lipid extraction andpurification. Can J Biochem Physiol 37: 911-917
3. CAMARA B, F BARDAT, R MONEGER 1982 Nature du premier precurseur enC,0 des carotenoides dans les chromoplastes de fruit de Poivron (Capsicumannuum L.) CR Acad Sci Paris III 294: 549-551
4. CAMARA B, F BARDAT, R MONEGER 1982 Sites de biosynthese des carotenoidesdans les chromoplastes de fruit de Poivron (Capsicum annuum L.) CR AcadSci Paris III 294: 649-652
5. CAMARA B, F BARDAT, R MONEGER 1982 Sites of biosynthesis of carotenoidsin Capsicum chromoplasts. Eur J Biochem 127: 255-258
6. CAMARA B, F BARDAT, A SEYE, A D'HARLINGUE, R MONEGER 1982 Terpenoidmetabolism in plastids: localization of a-tocopherol synthesis in Capsicumchromoplasts. Plant Physiol 70: 1562-1563
7. CAMARA B, J BRANGEON 1981 Carotenoid metabolism during chloroplast tochromoplast transformation in Capsicum annuum fruit. Planta 151: 359-364
8. CAMARA B, J BRANGEON 1981 Lipid and carotenoid metabolism during plastiddevelopment in Capsicum annuum fruit. Proc Inter Congr Photosynth 5:267-275
9. CAMARA B, R MONtGER 1977 Les lipides du fruit vert et du fruit muir dePoivron (Capsicum annuum L.). Physiol Veg 15: 711-722
10. CAMARA B, C PAYAN, A ESCOFFIER, R MONtGER 1980 Les carotenes du fruitde Poivron (Capsicum annuum): mise en evidence du phytoene-trans et duphytofluene-trans; comparaison de la biosynthese des diflfrents carotenes.CR Acad Sci Paris D 291: 303-306
1 1. DAVIEs BH 1976 Carotenoid. InTW Goodwin ed, Chemistry and Biochemistryof Plant Pigments, Vol 2. Academic Press, London, pp 38-165
12. KALRA SK, JL BRooKs 1973 Lipids of ripening tomato fruit and its mitochon-drial fraction. Phytochemistry 12: 487492
13. KREUZ K, P BEYER, H KLEINIG 1982 The site of carotenogenic enzymes inchromoplasts from Narcissus pseudonarcissus L. Planta 154: 66-69
14. LICHTENTHALER HK 1967 Zur synthese der lipophilen Plastidenchinone undSekundarcarotinoide wahrend der Chromoplastenentwicklung. Ber DtschBot Ges 82: 483497
15. LICHTENTHALER HK 1977 Regulation of prenylquinone synthesis in higherplants. In M Tevini, HK Lichtenthaler, eds, Lipids and Lipid Polymers inHigher Plants. Springer-Verlag, Berlin, pp 218-258
16. LIEDVOGEL B, H KLEINIG 1977 Lipid metabolism in chromoplast membranesfrom the daffodil: glycosylation and acylation. Planta 133: 249-253
17. LIEDVOGEL B, H KLEINIG 1979 Galactolipid synthesis in chromoplasts in vitro.Planta 144: 467471
18. LIEDVOGEL B, H KLEINIG, JA THOMPSON, H FALK 1978 Chromoplasts ofTropaeolum majus L.: lipid synthesis in whole organelles and subfractions.Planta 41: 303-309
19. LOWRY OH, NJ ROSEBROUGH, AL FARR, RJ RANDALL 1951 Protein measure-ment with the Folin phenol reagent. J Biol Chem 193: 265-275
20. MONEGER R, J VERMEERSCH, D LECHEVALLIER, C RICHARD 1977 Microdosagedu NADP et du NAD reduits et oxydes dans les tissus foliaires et dans lesplastes isoles de spirodele et de ble. Physiol Veg 15: 29-62
21. NAKATANI HY, J BARBER 1977 An improved method for isolating chloroplastsretaining their outer membranes. Biochim Biophys Acta 461: 510-512
22. POHL P, H GLASL, H WAGNER 1970 Zur Analytik Pflanzlicher Glyko und
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1983 American Society of Plant Biologists. All rights reserved.

PLASTID TERPENOID METABOLISM
Phospholipoide und Ihrer Fettsaren. 1. Eine neue Dunschichtchromatogra-phische Methode zur Trennung Pflanzlicher Lipoide und quantitativenBestimmung Ihrer Fettsaure-Zusammensetzung. J Chromatogr 49: 488-492
23. POSSINGHAM JV 1980 Plastid replication and development in the life cycle ofhigher plants. Annu Rev Plant Physiol 31: 113-129
24. PRICE CA 1974 Plant cell fractionation. Methods Enzymol 3 1A: 505-5 1925. SCHNEIDER H, R HAMPP, H ZIEGLER 1977 Envelope permeability to possible
precursors of carotenoid biosynthesis during chloroplast chromoplast trans-formation. Plant Physiol 60: 518-520
26. SOLL J, G SCHULTZ 1980 2-Methyl-6-phytylquinol and 2,3-dimethyl-5-phy-tylquinol as precursors of tocopherol synthesis in spinach chloroplasts.Phytochemistry 19: 215-218
27. THOMSON WW, JM WHATLEY 1980 Development of nongreen plastids. AnnuRev Plant Physiol 31: 375-394
28. WILLIAMS JP 1968 Separation and estimation of quinones and a-tocopherolfrom Viciafaba leaves. J Chromatogr 36: 504-51 1
29. WRIGHT RS 1971 A reagent for a nondestructive location ofsteroids and somelipophilic materials on silica gel thin layer chromatography. J Chromatogr59: 220-221
99
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1983 American Society of Plant Biologists. All rights reserved.





























