Embed Size (px)
Citation preview

[ 30 ]
TERMINAL DEGENERATION IN THE DIENCEPHALONAFTER ABLATION OF FRONTAL CORTEX
IN THE CAT
By J. AUERDepartment of Anatomy, University of Ottawa, Canada
Corticofugal connexions arising from the frontal lobe have been reported coursingto various nuclei in the diencephalon. Particular reference should be made in thisrespect to the work of Le Gros Clark (1932), Le Gros Clark & Boggon (1935),Mettler (1935), Meyer (1949) and Wall, Glees & Fulton (1951). The earlier findingsstem from Marchi material. Meyer studied human material with a modified Biel-schowsky technique (Glees, 1946), and so did Wall and his co-workers in the rhesusmonkey. However, only few observations are in accordance with the recent dataderived from neurophysiological experiments (Niemer & Jimenez-Castellanos,1950; Stratford, 1954; Jasper, Ajmone-Marson & Stoll, 1952; Livingston, Hernandez-Peon & French, 1953). It should be emphasized that particularly the so-calledunspecific thalamic nuclei have been little investigated with a view to revealingterminals from cortical efferents. Neither the Marchi technique nor the modifiedBielschowsky method (Glees, 1946) seem to be suitable for this purpose. The formerdoes not show unmyelinated fibres which are abundant in those nuclei while thelatter does allow precise detection, only after considerable exploration of the variousfields.The present study was primarily carried out in order to examine cortical efferents
from the frontal lobe to the hypothalamus. It had been impossible in recent studies(Auer & di Virgilio, 1953) to verify several connexions inferred (Ward & McCulloch,1947) and described earlier (Le Gros Clark, 1948; Meyer, 1949). In view of the factthat a bouton technique (Auer & di Virgilio, 1953) was used primarily for thispurpose, it was thought advisable to study the same problem with a selective silverimpregnationtechnique (Nauta & Gygax, 1954) for degenerating fibres and terminals.However, observations on all specific and non-specific diencephalic nuclei, par-ticularly those nuclei that have recently been discussed as the diffuse thalamicprojection system (Jasper, 1949; Hanbury & Jasper, 1953; Starzl & Magoun, 1951),were included in the investigation. It was considered important to study thecounterpart of the problem of the connexions from the thalamus to the cortexwhich have recently been clarified by the investigations of Rose (1952) and Powell &Cowan (1954). Therefore, the experiments described below deal with observationson efferents to (a) the specific thalamic nuclei, (b) the unspecific thalamic nuclei,(c) the subthalamus and (d) the hypothalamic nuclei. The subthalamus is also in-cluded in view of its intimate relationship with the nucleus reticularis which, asrecent evidence (Droogleever Fortuyn, 1950) has shown again, belongs to theunspecific thalamic nuclei.

Terminal degeneration in the diencephalon
MATERIAL AND METHODS
A series of five cats from a larger group has been used for this investigation. Theoperations were performed under antiseptic conditions and nembutal anaesthesia.A round opening was made with a drill in the roof of the right frontal sinus. Thepostero-inferior wall of this sinus was then removed in order to expose the frontalpart of the brain. The right cruciate sulcus could usually be identified easily and a
70 72
75 76
Text-fig. 1. Site of cortical lesion in cats 73, 70, 72, 75 and 76.
lesion was inflicted with a long thin thermocauter blade to the area anterior to thissulcus. As far as possible the attempt was made to restrict the lesion to the gyrusproreus and its immediate surrounding. The amount of bleeding encountered wasusually negligible and the exposed area was closed by a filling of bone wax. A doublelayer of sutures was used to close the surface wound. Infections were never seen andthe animals were all sacrificed by the fifth or sixth day after the operation. Func-tional changes during the post-operative period were never observed, due mainly tothe fact that the lesions were unilateral. Text-fig. 1 illustrates the different sites ofthese lesions in cats, 73, 70, 72, 75 and 76.
31

The five specimens were fixed in neutral formalin after perfusion with physio-logical saline. Serial sections were subsequently stained with the Nauta-Gygaxtechnique (1954) for selective impregnation of degenerating fibres of passage andterminals. This method has proved to be much more reliable than that used earlierfor detection of terminal boutons (Auer & di Virgilio, 1953). The latter method isonly of value for the study of a restricted area where changes are anticipated before-hand. Some brains stained with this method were used for control of the findings inthe present series. Orientation of the sections was greatly facilitated by the use of anatlas on the diencephalon of the cat (Jasper & Ajmone-Marsan, 1954).
OBSERVATIONS
The brain of cat 73 shows the most extensive lesion in the cortex, penetratingslightly into the white matter. Practically the whole anterior sigmoid gyrus hasbeen damaged and also the most anterior tip of the coronary gyrus and the orbitalgyrus. As Text-fig. 1 illustrates a narrow strip of the gyrus proreus was also includedin the lesion. The degenerating fibres can be followed in the internal capsule and canbe seen entering into the diencephalon via the anterior thalamic peduncle as well asthrough the inferior thalamic peduncle. The fibres of the former are piercing the headof the reticular nucleus. This nucleus shows a large number of degenerating terminals(see P1. 1). However, some fibres are seen traversing this nucleus and they graduallydisappear in more caudal areas of the thalamus, mainly in the intralaminar nuclei.This cat which, as has been said above, contains the most extensive cortical lesionof the series, does' not show any terminal degeneration in the nucleus ventralisanterior. The anterior group of nuclei is also entirely devoid of signs of fibre andterminal degeneration. The traversing fibres reach the intralaminar nuclei of whichthe nucleus centralis lateralis and the nucleus paracentralis should be emphasizedas the end-stations. Other fibres reach the nucleus ventralis lateralis in which thereis extensive terminal degeneration. More posteriorily the nucleus ventralis postero-medialis (Text-fig. 2; P1. 2, fig. 2) the nucleus ventralis medialis and on the samelevel the caudal extent of the nucleus ventralis lateralis show extensive terminaldegeneration. Most of these terminals arise from fibres entering through the lateralpart of the reticular nucleus. The medial nuclei at this level have been explored indetail. The nucleus dorsalis medialis as well as the nucleus centralis medialis(Text-fig. 2) have suffered less from terminal degeneration than, for instance, thenucleus ventralis posteromedialis. The nucleus dorsalis medialis possesses only afew degenerating terminals, apparently entering into the nucleus from the internalmedullary lamina. It is interesting to note that terminal degeneration in the nucleuscentralis medialis was also observed in similar experiments after the use of a boutontechnique (Auer & di Virgilio, 1953). Analysis of observations in the subthalamicarea and the centrum medianum is somewhat more difficult. Degenerating fibres ofpassage are present on each side of the zona incerta (Text-fig. 2; P1. 2, fig. 7). Thelatter area possesses degenerating terminals. This is also seen in the subthalmicnucleus and the entopeduncular nucleus. It is a matter of some speculation atpresent as to how the centrum medianum receives its degenerating fibres. It wouldappear that most of them enter via the fasciculi thalamicus and lenticularis. These
32 J. Auber

Terminal degeneration in the diencephalonbundles also send many fibres towards the reticular formation immediately caudalto the centrum medianum (Text-fig. 2).
Since only the caudal strip of the gyrus proreus has been damaged the hypo-thalamus does not show conspicuous changes. However, the medial forebrain
A73-S 24
C73-S 30
Text-fig. 2. Diagram of localization of degeneration in caudal diencephalon of cat 73. Crossesrepresent degenerating' terminals; dots represent degenerating fibres of passage. Dottedarrow, to tegmentum; lined arrow, to hypothalamus.
bundle contains scattered fibres of passage showing degeneration, and degeneratingterminals are seen originating from these fibres. An undeniable verification ofconnexions to the ventromedial nucleus and the mamillary nuclei as inferred fromphysiological experiments in the monkey (Ward & McCulloch, 1947)was notobtainedin this cat (Text-fig. 2).
73-S 27
33
3 Anat. 90

J. Auer
Cat 70The brain of cat 70 shows a cortical lesion which is slightly smaller than in cat 73.
The degeneration in the diencephalon, however, is very similar. Degeneratingterminals among the incoming fibres of passage are especially noted in the headof thereticular nucleus. The density of the degeneration, however, is slightly less than incat 73. The lateral part of the nucleus dorsalis medialis as well as the adjacent nuclei(PI. 1, fig. 3) of the internal medullary lamina present degenerating terminals as alsoseen in cat 73. The degenerating fibres directed towards the nucleus ventralis lateralis1~~~LA
73S1 7 70-S24Text-fig. 3. Diagram of localization of degeneration in diencephalon of cat 70. Crosses represent
degenerating terminals; dashes and dots represent degenerating fibres of passage.
and ventralis posterior traverse through the lateral part of the nucleus reticularisin which scattered terminal degeneration is noticeable. It should be pointed out thatthe head of the reticular nucleus, the lateral part of this nucleus and the continua-tion of this structure into the subthalamus contain degeneration throughout,degenerating fibres of passage as well as terminals. However, the greatest concentra-tion of terminal degeneration in all cats examined is present in the head of thereticular nucleus. The hypothalamus in this specimen is characterized by degenera-ting fibres traversing the medial forebrain bundle; this is particularly noticeablein the caudal part of the hypothalamus between the mamillary body and the internalcapsule (Text-fig. 3). The anterior part of the mamillary body contains severaldegenerating terminals. The origin of these terminals seems to lie in the medialforebrain bundle. Scattered terminal degeneration is present in the nucleus ventro-medialis. The question must be raised whether a few degenerating terminalsare significant enough to enable a conclusion to be made. However, comparisonwith the normal side shows a difference which renders it justifiable to mentionthe presence of degenerating terminals in some hypothalamic nuclei. The
34

Terminal degeneration in the diencephalonsubthalamus shows exactly the same sort of picture as described for cat 73. Thisregion, particularly the fields of Forel, are thoroughfares which are crowded withdegenerating fibres in all five of the cats.
Cat 72The brain of cat 72 has a lesion which completely destroyed the right gyrus
proreus. Only a small rostral strip of the anterior sigmoid gyrus was damaged bythe instrument. Some white matter deep to the cortex of the gyrus proreus was alsodamaged. The head of the reticular nucleus is again filled with numerous degenera-ting terminals and fibres of passage in degeneration. This part of the nucleus alsocontains a few degenerating terminals at the left side, which can only be due to thefact that a very small lesion was present at the medial side of the left gyrus proreus,inflicted by the tip of the blade. The degenerating terminals in the nuclei centralislateralis, paracentralis and centralis medialis, offer a picture similar to the onedescribed above (Text-fig. 4).
It would appear that a topic difference in efferents, i.e. a difference in distribution,from the gyrus proreus and the sensorimotor cortex, cannot be detected with themethod used; it remains to be seen whether such a difference exists. Only the con-centration of the degeneration seems to vary with the size of the lesion. A typicalpredilection is noted for the head of the reticular nucleus, which is the preferred sitein that nucleus for diffuse efferents from the frontal lobe.The inferior thalamic peduncle contains many degenerating fibres of passage which
seem to be directed towards the dorsomedial nucleus and the intralaminar nuclei(P1. 1, fig. 4). It seems impossible to accept the conclusion of Bodian (1940) thatthis peduncle carries mostly striate and pallidal connexions to the thalamus.The specific nuclei of the thalamus in this cat offer the same picture as described
for cats 70 and 73, with respect to the nucleus ventrolateralis and ventralis postero-medialis. However, the concentration of the degeneration is much less, due to thelesser involvement of the sensori-motor cortex in the lesion. This is not the case withthe dorsomedial nucleus, which shows the heaviest concentration of degeneratingterminals in the whole series (Text-fig. 4; P1. 2, fig. 5). The lateral half of the nucleusis particularly affected. The forward displacement of the lesion in this experimenthas definitely brought about a medial displacement of terminal degeneration in thespecific nuclei.The hypothalamus has been thoroughly investigated in this cat because of the
complete destruction of the gyrus proreus at the right side. The lateral hypothalamicarea possesses numerous degenerating terminals along the medial forebrain bundle.As in the former case a slight difference exists indeed between the right and leftinfundibular and mamillary regions. A few degenerating terminals are seen at theright side in the ventromedial, the lateral mamillary and the posterior hypothalamicnucleus. These terminals apparently arise from the medial forebrain bundle (Text-fig. 4).
Cats 75 and 76The brains of these specimens may be described together as their lesions are
approximately of the same size; they are added to the description in order to provideadditional evidence to the data described for the other experiments. The brain of
3-2
35

36 J. Auercat 76 has the smaller lesion of the two, and it is doubtless because of this that alesser number of degenerating terminals is noticed in the head of the reticularnucleus. The concentration of degenerating terminals in the nucleus ventralis lateralisand nucleus ventralis posterior is less than in any of the other experiments. This also
CN
R+'IC~~~~~~~~~~
72-s4 +y/ < (72-sI
A B72-S4 (1 72-S7
LA
LP
C72-Sl18
Text-fig. 4. Diagram of degeneration in diencephalon of cat 72. Crosses represent terminaldegeneration, dashes and dots represent degenerating fibres of passage.
applies to the intralaminar nuclei and the nucleus centralis medialis. The number ofabnormal terminals in the dorsomedial nucleus is so few that the degeneration can-not be considered significant after comparison with the brains described above. Thebrain of cat 75 showed some change to the white matter which resulted in destruc-tion of more fibres arising from the gyrus proreus than would be expected from the

Terminal degeneration in the diencephalonsize of the lesion. This is doubtless why the dorsomedial nucleus shows more extensivedegeneration than in cat 76.Both brains have extensive degeneration in the subthalamus. Fibres of passage
in degeneration are noticed and can be followed to the centrum medianum and to themidbrain tegmentum (P1. 2, fig. 8). The grey matter of these areas obviously receivesfibres from the damaged cortical area as has been described above for the first threebrains.The hypothalamus shows some fibre degeneration only in cat 75; however, these
observations are less significant even than those described above. Therefore conclu-sions will not be drawn from these last two experiments.
DISCUSSION AND CONCLUSION
The foregoing observations provide ample evidence for the acceptance of numerousconnexions in the cat's brain from the sensori-motor cortex and the prefrontalcortex (gyrus proreus) to the reticular nucleus, the intralaminar nuclei, the centrummedianum and the midline nuclei of the thalamus.An anatomical substratum is hereby provided for the physiological data reported
by Livingston et al. (1953). Reference may be made also to afferent connexions,recently established, to the rostral telencephalon from some unspecific nuclei (Rose,1952; Powell & Cowan, 1954) in order to assume the existence of 'unspecific'thalamo-cortical circuits coursing through the diffusely projecting thalamic nuclei.How such circuits are to be related to the earlier established specific circuits (Hsiang-tung Chang, 1950) remains to be investigated. Anatomical data is now accumula-ting to show how these unspecific thalamic nuclei can be brought to bear upon thewhole cortex by stimulation, not only from afferent systems (Starzl, Taylor &Magoun, 1951), but also from the cortex; that is to say, the cortex of the frontallobe, as far as the present study is concerned. It may be asked, for instance, how thevisual cortex projects on these unspecific nuclei, in view of the hypothesis proposedby Livingston et al. (1953) on the role played by the system as a whole with regardto sustained attention. Such connexions from the visual cortex were not reported byNauta & Bucher (1954) in the rat. These authors, however, reported abundantconnexions from the visual cortex to the reticular formation of the brain stem.Similar connexions are also arising from the damaged cortex in the experimentsreported above, but their detailed description has not been added to this paper. Itseems difficult to maintain the rather artificial separation of the reticular formationof the midbrain from the intralaminar and midline nuclei of the thalamus, in thelight of the accumulating evidence that it forms a functional unit with these nucleiof the diencephalon. The connexions to the dorsomedial nucleus, already reportedby Le Gros Clark (1932, with the Marchi technique) and more recently by Meyer(1949, in leucotomy material with a modified Bielschowsky technique) have nowalso been revealed with the selective silver impregnation method used in this study.These connexions seem to enter with the anterior and inferior thalamic radiationtogether with most of the fibres directed to the nuclei centralis lateralis, para-centralis and centralis medialis. It seems important that, in the cat, lesions in thecortex of the gyrus proreus result in degeneration mainly in the lateral and ventral
37

parts of this nuclear group. The present study failed to reveal changes in the medialpart. It should be mentioned here that the most recent evidence on efferent con-nexions from the gyrus proreus (Stratford, 1954) did not show abundant terminalsin these nuclei. However, these negative results may be due to the strychnineneuronography used by this author. The present findings are in accordance with thephysiological observations, also on cat material, carried out by Niemer & Jimenez-Castellanos (1950).The attention drawn to the nucleus ventralis anterior by some recent physio-
logical studies (e.g. Starzl & Magoun, 1951) could not be justified by the presentinvestigation. It is no doubt true that some fibres of passage traverse this nucleus,but terminal degeneration has not been observed in it.
It would be interesting to compare the present studies with material obtainedafter destruction of the head of the caudate nucleus. Degeneration in the intra-laminar nuclei and changes in the nucleus ventralis anterior would then seem to be inline with earlier investigations with physiological methods (Shimamoto & Verzeano,1954). At present one can only conclude that the nucleus ventralis anterior is not areception area for diffuse terminals from the frontal cortex, but it may be a relaynucleus between the thalamus and the caudate nucleus. The findings in the nucleusventralis lateralis and ventralis posterior are only a confirmation of earlier studies onthese nuclei, particularly by Gerebtzoff & Wauters (1941). It is, however, in thelight of the observations of Niemer & Jimenez-Castellanos (1950), interesting to notethat the nucleus ventralis posterior also receives connexions from the precruciate,i.e. the motor cortex.The fact that the subthalamus was never clear of degenerating fibres emphasizes
the conclusion that this area may be classified with the intralaminar and reticularnuclei as belonging to one system which receives diffusely arranged corticodien-cephalic fibres. It is quite correct to state with Niemer & Jimenez-Castellanos (1950)that the subthalamus represents a 'strategic bottleneck'. It should be emphasizedstrongly that the head of the reticular nucleus as well as its caudal continuation(Droogleever Fortuyn, 1950) show these degenerating terminals more abundantlythan any other part of the same extralaminar structures. It is not yet possible toanalyse further from which regions of the cortex the different elements of the sub-thalamus receive terminals, and similarly an analysis in this respect of the fasciculithalamicus and lenticularis cannot be offered. The lesions under consideration are inthe first place too extensive for this purpose. However, the distribution of fibres inthe whole area of the subthalamus after different ablations justify the conclusionthat the efferents observed here have a rather diffuse spread without any topicarrangement whatsoever. The efferents to the hypothalamus have caused con-siderable difficulties, particularly in view of the evidence of earlier anatomical andphysiological studies (Meyer, 1949; Ward & McCulloch, 1947). The present experi-ments caused degeneration in the medial forebrain bundle. This bundle showeddegenerating fibres of passage as well as degenerating terminals, particularly in cats70, 72 and 73. The number of degenerating terminals in the nucleus ventromedialis,the nucleus posterior and the nucleus mamillaris lateralis, observed in cats 70 and 72,is so small that one hardly feels justified in stating specifically that efferents to thesenuclei are present. If one compares the data of Niemer & Jimenez-Castellanos (1950)
38 J. Auzer

Terminal degeneration in the diencephalon3on the cat with the conclusions of Ward & McCulloch (1947) on ,the monkey onewould be inclined to assume that there is a difference in the number of connexions infavour of the monkey. Meyer's illustrations (1949) show even more connexions in theleucotomy material of humans. This would lead to the conclusion that the numberof cortical efferents to the hypothalamus has increased in higher mammals. Quantita-tively such an increase is perhaps to be expected, since the area from which corticalefferents to the hypothalamus may arise in monkey and man is considerably largerthan the gyrus proreus of the cat. This conclusion, however, needs further investiga-tion, particularly qualitatively.
SUMMARY
Ablations were made in the precruciate and prefrontal cortex of five cats. The catswere sacrificed five to six days post-operatively, the brains were fixed in neutralformalin, subsequently sectioned, and stained with a selective impregnation tech-nique for degenerating fibres of passage and terminals. Cortical efferents to thediencephalon have been observed and classified as follows:
(1) Diffuse efferents from the damaged areas are directed towards the intra-laminar nuclei, reticular nucleus and centrum medianum.
(2) The dorsomedial nucleus shows terminal degeneration to an amount propor-tional to the size of the lesion in the gyrus proreus.
(3) The subthalamus has terminal degeneration in the zona incerta, and the sub-thalamic nucleus. Moreover, this area appears to be a thoroughfare for fibres ofpassage to the midbrain reticular formation and probably also to the centrummedianum.
(4) The hypothalamus is characterized by terminal degeneration and degeneratingfibres of passage in the medial forebrain bundle. Little degeneration was observedin the ventromedial, the mamillary and the posterior hypothalamic nuclei.
Finally, confirmation has been obtained of cortical efferents to specific thalamicnuclei from the frontal lobe.
This work was supported by a grant in aid of research from the National ResearchCouncil of Canada.
REFERENCESAUER, J. & DI VIRGILIO, G. (1953). Some afferent connections of the hypothalamus in the cat.
Anat. Rec. 115, 277.AUER, J. & DI VIRGILIO, G. (1953). Demonstration of degenerating nerve fibers by a modified
Cajal technique. Stain Tech. 28, 141-146.BODIAN, D. (1940). Studies on the diencephalon of the Virginia opossum. II. The fiber connections
in normal and experimental material. J. comp. Neurol. 72, 207-297.DROOGLEEVER FORTUYN, J. (1950). On the configuration and the connections of the medio-ventral
area and the midline cells in the thalamus of the rabbit. Folia psychiat. neerl. 53, 213-254.GEREBTZOFF, M. A. & WAUTERS, A. (1941). Recherches sur l'ecorce cerebrale et le thalamus du
cobaye. II. Systematisation cortico-thalamique et voies efferentes de 1'ecorce cerebrate.Cellule, 48, 7-70.
GLEES, P. (1946). Terminal degeneration within the central nervous system as studied by a newsilver method. J. Neuropath. 5, 54-59.
HANBURY, J. & JASPER, H. H. (1953). Independence of diffuse thalamocortical projection systemshown by specific nuclear destruction. J. Neurophysiol. 16, 252-272.
39

40 J. AuerHSIANG-TUNG CHANG (1950). The repetitive discharge of cortico-thalamic reverberating circuit.
J. Neurophysiol. 13, 235-258.JASPER, H. H. (1949). Diffuse projection systems: the integrative action of the thalamic reticular
system. Electroenceph. clin. Neurophysiol. 1, 405-419.JASPER, H. H. & AJMONE-MARSAN, C. (1954). A Stereotaxic Atlas of the Diencephalon of the Cat.
Ottawa, Canada: Nat. Res. Council Publ.JASPER, H. H., AJMONE-MARSAN, C. & STOLL, J. (1952). Cortico-fugal projections to the brain stem.
Arch. neurol. Psychiat., Chicago, 67, 155-171.LE GROS CLARK, W. E. (1932). Structure and connections of the thalamus. Brain, 55, 406-470.LE GROS CLARK, W. E. (1948). The connections of the frontal lobes in the brain. Lancet, 1, 353-56.LE GROS CLARK, W. E. & BOGGON, R. H. (1935). The thalamic connections ofthe parietal and frontal
lobes of the brain in the monkey. Phil. Trans. B, 224, 313-359.LIVINGSTON, R. B., HERNkNDEZ-PE'ON & FRENCH, J. D. (1953). Corticofugal projections to brain
stem activating system. Fed. Proc. 12, 89.METTLER, F. A. (1935). Corticofugal fiber connections of the cortex of Macaca muatta. The frontal
region. J. comp. Neurol. 61, 509-542.MEYER, M. (1949). A study of efferent connections of the frontal lobe in the human brain after
leucotomy. Brain, 72, 265-297.MORRISON, R. S. & DEMPSEY, E. W. (1942). A study of thalamocortical relations. Amer. J. Physiol.
135, 281-292.NAUTA, W. J. H. & BUCHER, V. M. (1954). Efferent connections of the striate cortex in the albino
rat. J. comp. Neurol. 100, 257-287.NAUTA, W. J. H. & GYGAX, P. A. (1954). Silver impregnation of degenerating axons in the central
nervous system. Stain Tech. 29, 91-93.NIEMER, W. T. & JIMENEZ-CASTELLANOS, J. (1950). Cortico-thalamic connections in the cat as
revealed by 'physiological neuronography', J. comp. Neurol. 93, 101-125.POWELL, T. P. S. & COWAN, W. M. (1954). The connections of the midline and intralaminar nuclei
of the thalamus of the rat. J. Anat., Lond., 88, 307-319.ROSE, J. E. (1952). The cortical connections of the reticular complex of the thalamus. Res. Publ.
Ass. nerv. ment. Dis. 30, 455-479.SHIMAMOTO, T. & VERZEANO, M. (1954). Relations between caudate and diffusely projecting thala-
mic nuclei. J. Neurophysiol. 17, 278-289.STARZL, T. E. & MAGOUN, H. W. (1951). Organisation of the diffuse thalamic projection system.
J. Neurophysiol. 14, 133-147.STARZL, T. E., TAYLOR, C. W. & MAGOUN, H. W. (1951). Collateral afferent excitation of reticular
formation of brain stem. J. Neurophysiol. 14, 479-497.STRATFORD, J. (1954). Cortico-thalamic connections from gyrus proreus and first and second
sensory areas. J. comp. Neurol. 100, 1-15.WALL, P. D., GLEES, P. & FULTON, J. F. (1951). Corticofugalconnectionsof posterior orbital surface
in Rhesus monkey. Brain, 74, 66-71.WARD, A. A. & MCCULLOCH, W. S. (1947). The projection of the frontal lobe on the hypothalamus.
J. Neurophysiol. 10, 309-315.
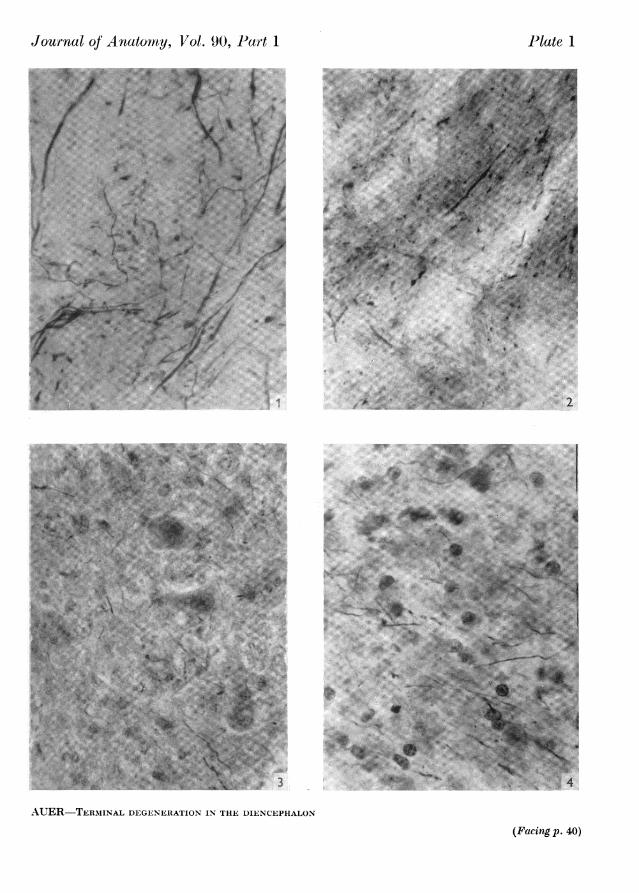
EXPLANATION OF PLATESPLATE 1
Fig. 1. Degeneration of terminals in the head of the nucleus reticularis (centre of picture). Section17, cat 73; x 1250.
Fig. 2. Terminal degeneration and degenerating fibres of passage in the head of the nucleusreticularis. Section 10, cat 72; x 1250.
Fig. 3. Terminal degeneration in the nucleus centralis lateralis (centre of picture). Section 17,cat 70; x1250.
Fig. 4. Scattered terminal degeneration in the nucleus paracentralis. Section 19, cat 72; x 1250.
Journal of Anatomy, Vol. 90, Part 1 Plate 1
_W 4 .jEi:, ..::4J. .
%r it 1. .- s
....I:,--... I. ..
..A....'
A I~ 2
.. .~~~~~~~~~~~.... :: ...iL ;
AUER-TERAIINAL DEGENERATION IN THE DIENCEPHALON
(Facing p. 40)
0W17

Journal of Anatomy, Vol. 90, Part 1
AUER-TERMINAL DEGENERIATION IN THE DIENCEPHALON
Plate 2

Terminal degeneration in the diencephalon 41
PLATE 2
Fig. 5. Degenerating terminals in the nucleus dorsomedialis thalami. Section 18, cat 72; x 1250.Fig. 6. Concentrated terminal degeneration in the nucleus ventralis posteromedialis. Section 28,
cat 73; x 625.Fig. 7. Degenerating terminals and fibers in the zona incerta. Section 28, cat 73; x 1250.Fig. 8. Degenerating fibers ofpassage entering midbrain tegmentum from pes pedunculi. Section 11,
cat 75; x 1250.
KEY TO LETTERINGanterior ectosylvian gyrus
anterior sylvian gyrus
anterior sigmoid gyrus
anterior suprasylvian gyrus
centrum medianumnucleus centralis lateralisnucleus centralis medialisnucleus caudatuscoronary gyrus
nucleus dorsalis medialisfornixlateral geniculate bodymedial geniculate bodyglobus pallidusgyrus proreus
hypothalamushabenula-hab.interped.tractinternal capsuleinferior thalamic pedunclenucleus lateralis anteriorgyrus lateralisnucleus lateralis posteriormamillary body
MESMFMSSMSYNNVMOBORBOSOTP
PCPESPSGPSSPSYRRFS
VAVLVM
VP
middle ectosylvian gyrus
medial forebrain bundlemiddle suprasylvian gyrus
middle sylvian gyrus
substantia nigranucleus ventromedialisolfactory bulborbital gyrus
olfactory striaoptic tractpes pedunculinucleus paracentralisposterior ectosylvian gyrus
posterior sigmoid gyrus
posterior suprasylvian gyrus
posterior sylvian gyrusnucleus reticularisreticular formation-midbrainsubthalamusnucleus ventralis anteriornucleus ventralis lateralisnucleus ventralis medialisnucleus ventralis posterior
AESASYASGASSCCLCMCNCORDMFGLGMGPGPrHHBICIPLALATLPM