Embed Size (px)
Citation preview

www.elsevier.com/locate/ydbio
Developmental Biology 267 (2004) 153–164
Temporal regulation of cerebellar EGL migration through a switch in
cellular responsiveness to the meninges
Yan Zhu,1 Tao Yu, and Yi Rao*
Department of Anatomy and Neurobiology, Washington University School of Medicine, St. Louis, MO 63110, USA
Received for publication 2 November 2002, revised 1 October 2003, accepted 28 October 2003
Abstract
We have studied the temporal and spatial control of cell migration from the external germinal layer (EGL) in the mammalian cerebellum
as a model for cortical migration. Our results have demonstrated that embryonic EGL cells do not migrate into internal layers because they
respond to a diffusible attractant in the meninges, the nonneural tissues covering the nervous system, and to a repellent in the
neuroepithelium. Two developmental changes are important for postnatal EGL migration: the disappearance of the repellent in the inner
layers and a switch in cellular responsiveness of EGL cells so that the postnatal EGL cells respond to the repellent, but not the attractant in the
meninges. Besides revealing the signaling role of meninges in cortical development, our study suggests that an active mechanism is required
to prevent cell migration, and that mechanisms of cell migration should be studied even in the absence of apparent changes in cell positions.
We propose a model for the developmental control of neuronal migration in the cerebellar cortex.
D 2003 Elsevier Inc. All rights reserved.
Keywords: Cell migration; Neuronal migration; Cortex; Cerebellum; Neural development
Introduction
Neuronal migration is essential for the formation of
cerebral and cerebellar cortices in the mammalian brain. A
classic model for cortical lamination involves the migration
of cells from the external germinal layer (EGL) into the
internal granule layer (IGL) in the cerebellum (Altman and
Bayer, 1997; Hatten, 1999; Hatten and Heintz, 1995; Pearl-
man et al., 1998; Rakic, 1971a,b, 1990; Rice and Curran,
1999).
In the embryo, EGL cells are derived from cells in the
upper rhombic lip (URL) (Alder et al., 1996; Altman and
Bayer, 1997; Hatten, 1999; Wingate and Hatten, 1999).
Cells in the URL migrate tangentially to form the EGL
(Wingate and Hatten, 1999). Once in the EGL, the URL
derived cells do not migrate towards the IGL in the embryo
(Altman and Bayer, 1997; Hatten, 1999). Postnatally, EGL
cells are induced to proliferate by the sonic hedgehog
protein secreted from the Purkinje cells underlying the
0012-1606/$ - see front matter D 2003 Elsevier Inc. All rights reserved.
doi:10.1016/j.ydbio.2003.10.037
* Corresponding author. Department of Anatomy and Neurobiology,
Washington University School of Medicine, Box 8108, 660 South Euclid
Avenue, St. Louis, MO 63110. Fax: +1-314-362-3446.
E-mail address: [email protected] (Y. Rao).1 Current address: HHMI at UCLA, 675 Charles E. Young Drive South,
5-784 MRL, Los Angeles, CA 90095.
EGL cells (Wechsler-Reya and Scott, 1999). Postnatal
EGL cells migrate radially from the more superficial layer
through the Purkinje cell layer to IGL, giving rise to the
granule cells (Rakic, 1971a, 1990). At the cellular level, the
migration of EGL cells towards the IGL in vivo is thought to
be dependent on radially aligned glial fibers (Rakic, 1971a).
Mutations in several genes affect neuronal positioning in
the cerebellum of rodents (Dhavan and Tsai, 2001; Rice and
Curran, 1999, 2001; Ross and Walsh, 2001). These include
reeler (reviewed in Rice and Curran, 2001), scrambler
(Howell et al., 1997; Sheldon et al., 1997; Sweet et al.,
1996; Ware et al., 1997), disabled-1 (Dab-1), weaver
(Hatten et al., 1984), cdk5 and its activator p35 (Chae et
al., 1997; Dhavan and Tsai, 2001; Nikolic et al., 1996; Tsai
et al., 1994). The reeler gene encodes a secreted protein
Reelin (D’Arcangelo et al., 1995). Reelin binds to the very
low density lipoprotein (VLDL) and ApoE2 receptors and
activates a pathway involving Dab-1 (D’Arcangelo et al.,
1999; Hiesberger et al., 1999; Trommsdorff et al., 1999). In
the neocortex, the precise functional role of the Reelin
pathway is still not clear (Rice and Curran, 2001), although
one possibility is that Reelin provides a stop signal for
neurons migrating from the ventricular zone to the cortical
plate (Pearlman et al., 1998). In the cerebellum, the Reelin
pathway does not seem to act on the EGL cells; Reelin is
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164154
secreted by granule precursor cells and acts on Purkinje cells
to promote the formation of the Purkinje cell layer (D’Ar-
cangelo et al., 1995; Howell et al., 1997; Sheldon et al.,
1997).
Despite extensive studies, the understanding of mecha-
nisms controlling the spatial and temporal aspects of EGL
migration is still limited. For temporal control, there have
been no experimental studies designed to address the
question why the embryonic EGL cells do not migrate into
the IGL whereas the postnatal EGL cell do. For spatial
control, it is not known what directs postnatal EGL cells
migrate towards the IGL. We have now investigated the
mechanisms underlying temporal and spatial control of EGL
migration by studying the migration of embryonic and
postnatal EGL cells in rodents. The study documents the
developmental properties of both guidance sources and
targets neurons, and provides a framework for understand-
ing molecular basis of EGL migration. Our analyses have
revealed attractive and repulsive forces that control the
migration of EGL cells. Our result uncovered a signaling
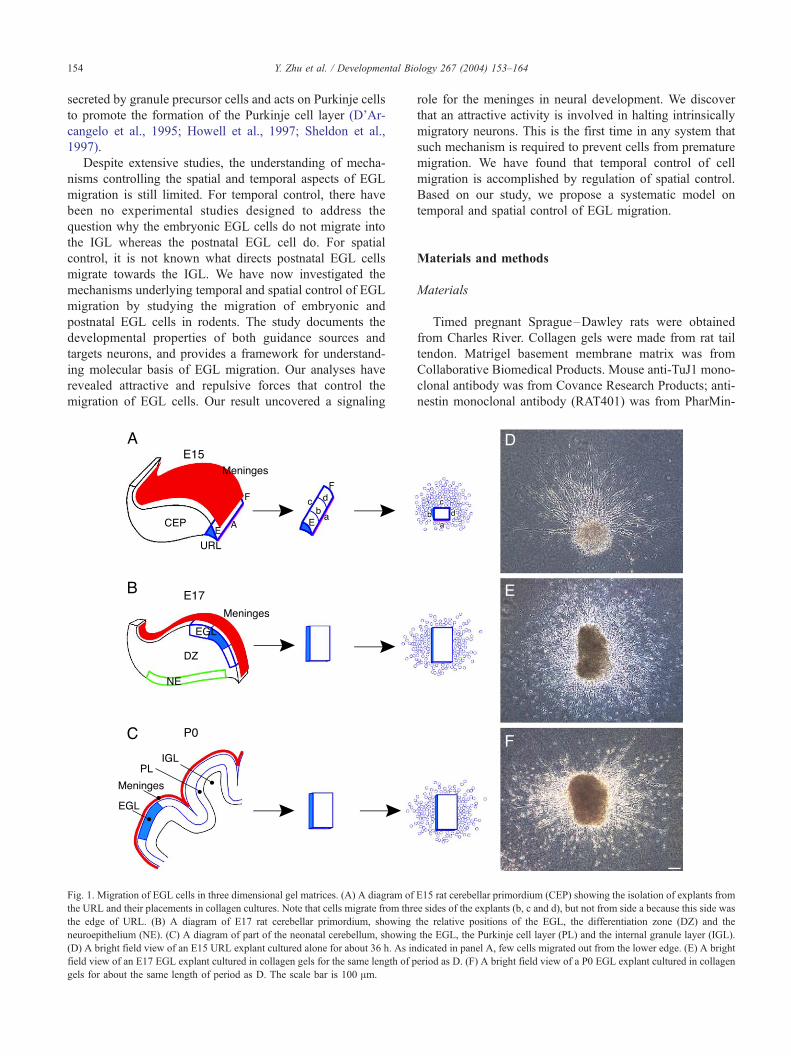
Fig. 1. Migration of EGL cells in three dimensional gel matrices. (A) A diagram of
the URL and their placements in collagen cultures. Note that cells migrate from thre
the edge of URL. (B) A diagram of E17 rat cerebellar primordium, showing
neuroepithelium (NE). (C) A diagram of part of the neonatal cerebellum, showing
(D) A bright field view of an E15 URL explant cultured alone for about 36 h. As in
field view of an E17 EGL explant cultured in collagen gels for the same length of p
gels for about the same length of period as D. The scale bar is 100 Am.
role for the meninges in neural development. We discover
that an attractive activity is involved in halting intrinsically
migratory neurons. This is the first time in any system that
such mechanism is required to prevent cells from premature
migration. We have found that temporal control of cell
migration is accomplished by regulation of spatial control.
Based on our study, we propose a systematic model on
temporal and spatial control of EGL migration.
Materials and methods
Materials
Timed pregnant Sprague–Dawley rats were obtained
from Charles River. Collagen gels were made from rat tail
tendon. Matrigel basement membrane matrix was from
Collaborative Biomedical Products. Mouse anti-TuJ1 mono-
clonal antibody was from Covance Research Products; anti-
nestin monoclonal antibody (RAT401) was from PharMin-
E15 rat cerebellar primordium (CEP) showing the isolation of explants from
e sides of the explants (b, c and d), but not from side a because this side was
the relative positions of the EGL, the differentiation zone (DZ) and the
the EGL, the Purkinje cell layer (PL) and the internal granule layer (IGL).
dicated in panel A, few cells migrated out from the lower edge. (E) A bright
eriod as D. (F) A bright field view of a P0 EGL explant cultured in collagen

Y. Zhu et al. / Developmental Biology 267 (2004) 153–164 155
gen International; monoclonal anti-GFAP antibody was
from Sigma (G 3893). Cy3-conjugated donkey anti-goat
antibodies and Cy3-conjugated goat anti-rabbit antibodies
were from Jackson ImmunoResearch Laboratories. Pertussis
toxin (PTX, P7208), Hoechst 33258, and paraformaldehyde
(PFA) were obtained from Sigma.
Explant assay for EGL migration
Cocultures of EGL explants were performed similarly as
previously reported with some modifications (Zhu et al.,
1999). Explants were isolated from the EGL of E17 rats or
the URL of E15 rats. Explants from postnatal day 0 (P0) and
P5 behaved similarly. For explants of postnatal EGL, para-
sagittal cerebellar slices of 300 Am were cut on a Vibratome,
and the EGL explants were dissected by a fine tungsten
needle from the anterior cerebellum. Meninges were taken
from the area directly covering the cerebellum. Isolated
regions of proper layers had been confirmed by immunos-
taining with anti-MATH1, anti-TuJ1, anti-Calbindin, and
anti-nestin antibodies on frozen sections of explants that
were fixed immediately after dissection. Aggregates of HEK
were prepared as described previously (Li et al., 1999; Zhu
et al., 1999). Explants and cell aggregates were positioned at
a distance of 300 to 400 Am in a 1:2 mixture of Matrigel and
collagen gel. For the embryonic EGL explants, few cells
migrated out of the edge of rhombic lip (side a in Figs. 1A
and D), and we therefore avoided to have this edge face
toward or away from the source: the explants were posi-
tioned in a way that either of the cutting edges of the
medial– lateral orientation in the developing cerebellum
faced the source (side b or d in Fig. 1A). For the postnatal
EGL explants, the explants were positioned in a way that
either of the vibratome-sectioned edges faced the source.
The cocultures were performed in 10% FCS (fetal calf
serum), 1�P/S (penicillin and streptomycin), DMEM (Dul-
becco’s minimal essential medium, Gibco) for 24 to 48 h in
5% CO2 at 37jC. To calculate P/D and D/P ratios, cultures
were fixed and stained with the nuclear dye. Pictures were
taken with a CCD camera and the images were analyzed
with Scion Image software (Release Beta 4.0.2, Scion
Corporation) to obtain the ratios of numbers of migratory
cells on the proximal and distal sides. Statistic analyses were
carried out with Student’s t test.
Immunostaining of the explants cultured in collagen
matrix was performed similarly as previously reported
(Zhu et al., 1999). Confocal images were taken on an
Olympus fluoview laser scanning microscope. SYTOX
Green nucleic acid stain (S-7020, Molecular Probes) instead
of Hoechst dye was used as the nuclear dye in the confocal
images.
PTX blocking experiment
Explants were dissected and then incubated in the culture
medium with 0.1 Ag/ml of PTX for 4 h at 37jC. The
controls were explants cultured at the same time in the
medium without PTX. After pretreatment, cocultures of the
EGL explants with meninges were performed as described
above except that PTX was added to both the mixture gel
and the culture medium. PTX in control cocultures did not
affect the production of secreted proteins from HEK cell
aggregates (data not shown).
Results
Migration of neurons from the EGL in three dimensional gel
matrices
It is known that cells in the URL migrate tangentially
into the EGL of the mammalian cerebellum (Adler et al.,
1996; Altman and Bayer, 1997; Hatten, 1999; Wingate and
Hatten, 1999). However, the embryonic EGL cells do not
migrate internally whereas the postnatal EGL cells do
migrate into the IGL (Altman and Bayer, 1997; Hatten,
1999). The mechanism for the lack of migration of embry-
onic EGL cells into the internal layers has not been
previously investigated. During the course of our studies
of EGL cell migration to address the question why embry-
onic EGL cells do not move into internal layers, we
established an in vitro culture assay with three dimensional
collagen matrices.
Three dimensional matrices are a reliable system for
studying neuronal migration (Letinic and Rakic, 2001;
Marin et al., 2001; Pleasure et al., 2000; Powell et al.,
2001; Wong et al., 2001; Wu et al., 1999; Zhu et al., 1999)
and were used here to culture explants of the URL and
different stages of EGL (Figs. 1 and 2). Cells could migrate
out of the URL explants from embryonic day 15 (E15) rats,
EGL explants of E17 and postnatal day 0 (P0) rats (Fig. 1).
Immunocytochemistry with the neuronal specific anti-TuJ1
antibody showed that these migrating cells were neurons
(Figs. 2B and E). Antibodies to the glial fibrillary acidic
protein (GFAP) can be used to examine postnatal glial cells
in the cerebellum, but they do not stain for embryonic glial
precursor cells. Antibodies to nestin could reveal glial
precursor cells as well as neuronal precursor cells in
embryonic and postnatal cerebellum (Hopfield and McKay,
1985; Sotelo et al., 1994). Results from immunocytochem-
istry with antibodies to nestin and GFAP showed that glial
cells and their precursor cells stayed within the explants and
did not migrate out of either embryonic or postnatal EGL
explants (Figs. 2H, K and N). All cells that migrated out of
the explants are TuJ1 positive neurons (Fig. 2E). These
results indicate that neurons in the EGL do not rely on glial
fibers for their migration.
The leading processes of neurons migrating from E15
URL and postnatal EGL explants are quite long and give the
appearance that migrating neurons are associated with, or
perhaps even move along, the processes of other neurons
(Figs. 1D and F). However, although neurons from E17
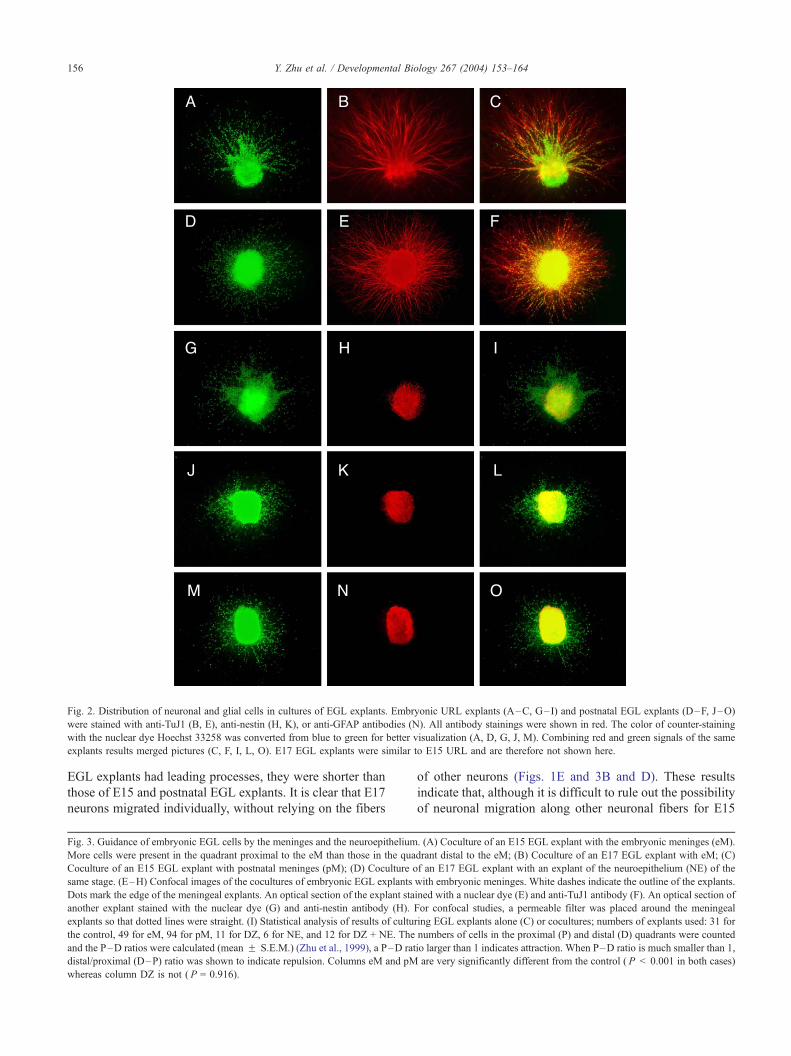
Fig. 2. Distribution of neuronal and glial cells in cultures of EGL explants. Embryonic URL explants (A–C, G–I) and postnatal EGL explants (D–F, J–O)
were stained with anti-TuJ1 (B, E), anti-nestin (H, K), or anti-GFAP antibodies (N). All antibody stainings were shown in red. The color of counter-staining
with the nuclear dye Hoechst 33258 was converted from blue to green for better visualization (A, D, G, J, M). Combining red and green signals of the same
explants results merged pictures (C, F, I, L, O). E17 EGL explants were similar to E15 URL and are therefore not shown here.
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164156
EGL explants had leading processes, they were shorter than
those of E15 and postnatal EGL explants. It is clear that E17
neurons migrated individually, without relying on the fibers
Fig. 3. Guidance of embryonic EGL cells by the meninges and the neuroepithelium
More cells were present in the quadrant proximal to the eM than those in the qua
Coculture of an E15 EGL explant with postnatal meninges (pM); (D) Coculture o
same stage. (E–H) Confocal images of the cocultures of embryonic EGL explants
Dots mark the edge of the meningeal explants. An optical section of the explant sta
another explant stained with the nuclear dye (G) and anti-nestin antibody (H).
explants so that dotted lines were straight. (I) Statistical analysis of results of cultu
the control, 49 for eM, 94 for pM, 11 for DZ, 6 for NE, and 12 for DZ + NE. The
and the P–D ratios were calculated (mean F S.E.M.) (Zhu et al., 1999), a P–D rat
distal/proximal (D–P) ratio was shown to indicate repulsion. Columns eM and pM
whereas column DZ is not ( P = 0.916).
of other neurons (Figs. 1E and 3B and D). These results
indicate that, although it is difficult to rule out the possibility
of neuronal migration along other neuronal fibers for E15
. (A) Coculture of an E15 EGL explant with the embryonic meninges (eM).
drant distal to the eM; (B) Coculture of an E17 EGL explant with eM; (C)
f an E17 EGL explant with an explant of the neuroepithelium (NE) of the
with embryonic meninges. White dashes indicate the outline of the explants.
ined with a nuclear dye (E) and anti-TuJ1 antibody (F). An optical section of
For confocal studies, a permeable filter was placed around the meningeal
ring EGL explants alone (C) or cocultures; numbers of explants used: 31 for
numbers of cells in the proximal (P) and distal (D) quadrants were counted
io larger than 1 indicates attraction. When P–D ratio is much smaller than 1,
are very significantly different from the control ( P < 0.001 in both cases)
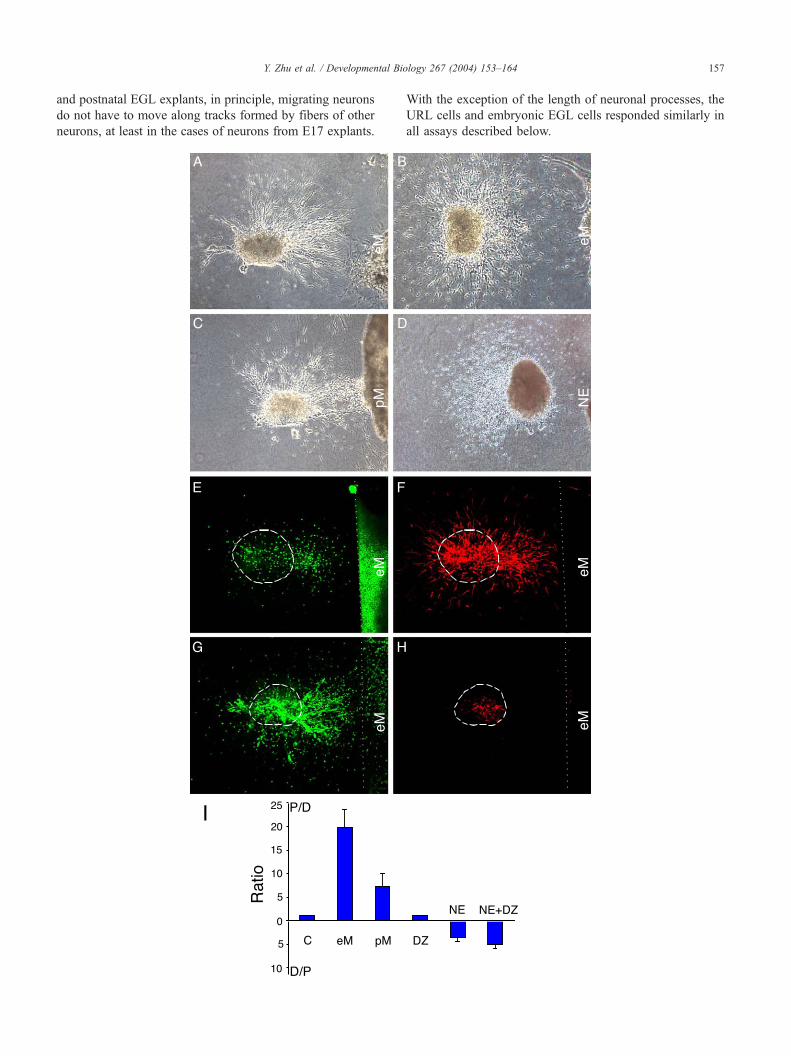
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164 157
and postnatal EGL explants, in principle, migrating neurons
do not have to move along tracks formed by fibers of other
neurons, at least in the cases of neurons from E17 explants.
With the exception of the length of neuronal processes, the
URL cells and embryonic EGL cells responded similarly in
all assays described below.

Y. Zhu et al. / Developmental Biology 267 (2004) 153–164158
Attractive and repulsive activities in the Meninges and in the
neuroepithelium
With the establish assay, we investigated whether the
migration of embryonic URL and EGL cells could be
affected by their neighboring tissues. The outer neighbor
of the EGL is the nonneuronal meninges, which cover the
nervous system. Layers internal to the EGL in the embry-
onic cerebellum include the differentiation zone (DZ) and
the neuroepithelium (NE) (Fig. 1B).
The intuitively obvious way for the neighboring
tissues to control EGL migration is to inhibit the
intrinsic motility of the embryonic EGL cells. However,
coculture experiments showed that cells migrated from
the embryonic EGL explants in the presence of the
neighboring tissues (Fig. 3), indicating that none of the
neighboring tissues could inhibit EGL migration. Inter-
estingly, cells from both the E15 URL (Fig. 3A) and
the E17 EGL (Fig. 3B) migrated towards the meninges,
as indicated by the presence of a larger number of cells
in the quadrant proximal to the meninges than that in
the distal quadrant (Figs. 3A, B and I). These results
indicate that the meninges attract EGL cells, and do not
inhibit EGL migration. Because the attraction was not
dependent on cell contact (Figs. 3A and B), the
attractive cue(s) in the meninges must be diffusible.
The neuronal nature of EGL cells attracted by the
meninges was shown by positive staining for the
TuJ1 antibody (Fig. 3F). The attractive effect of the
meninges is likely to be direct on the neurons, rather
than being indirectly through glial fibers because stain-
ing for glial precursor cells with an anti-nestin antibody
showed that glial precursor cells stayed within the
explants (Fig. 3H).
When EGL explants were cocultured with the NE, it was
found that the NE repelled the EGL cells (Figs. 3D and I).
The repellent(s) in the NE was also diffusible. The DZ
neither attracted nor repelled the EGL cells (Fig. 3I).
Explants containing both the NE and the DZ (labeled as
NE + DZ in Fig. 3I) were still repulsive to the EGL cells,
indicating that DZ did not mask the repulsive activity
diffusing from the NE.
In summary, the embryonic EGL cells respond to a
diffusible attractive activity in the meninges and a diffus-
ible repellent in the NE, suggesting that the strategic
positioning of the guidance cues may make the EGL cells
unable to move into the inner layers in the embryonic
cerebellum.
Fig. 4. Responses of postnatal EGL cells to guidance activities. (A) Cocultur
magnification view of the distal side of the EGL explant in A. (C) A high magnif
100 Am. (D) Coculture of a postnatal EGL explant with the embryonic meninges.
and internal granular layer. (F) Staining with the nuclear dye. (G) Staining of the sa
Staining of the same explant as that in H with the anti-nestin antibody. The scale ba
numbers of explants used: 28 in pEGL alone (C), 134 in eM, 215 in pM, and 22
control ( P < 0.001 in both cases), whereas column PL + IGL is not ( P = 0.773
Developmental changes in the responsiveness of EGL cells
to guidance cues in the meninges
The presence of attractive and repulsive activities in
tissues surrounding the EGL raises the question of how
postnatal EGL cells can migrate into the IGL. We first
investigated the activity in the inner layers. Postnatally, the
neuroepithelium is absent. But do the Purkinje cell layer
(PL) and the IGL have any attractive or repulsive activities?
Coculture experiments indicated that the PL and the IGL
were neither attractive nor repulsive to the postnatal EGL
cells (Figs. 4E and J).
We then examined the guidance activities in the meninges.
If the meninges are attractive to the EGL, how could EGL
cells move away from it? One possibility is that there is a
developmental change in the signal sent by the meninges, so
that only the embryonic, but not the postnatal, meninges
send the attractant. Alternatively, there could be a change in
the responsiveness of the EGL cells to the meningeal
signal(s), so that postnatal EGL cells do not respond to
the attractive signal. It is also possible that both of these
changes take place.
To distinguish among possibilities, we cocultured em-
bryonic and postnatal meningeal explants with EGL
explants of the same or different developmental stages.
Similar to the embryonic meninges (Figs. 3A and B), the
postnatal meninges also attracted the embryonic EGL cells
(Fig. 3C). On the other hand, the postnatal EGL cells were
repelled by both the postnatal meninges (Fig. 4A) and the
embryonic meninges (Fig. 4D). The repelled postnatal EGL
cells were neurons (Fig. 4G) whereas glial cells stayed
within the explants (Fig. 4I) and glial fibers grew a very
short distance outside the explants (Fig. 4I).
These results indicate that both attractive and repulsive
activities are present in the meninges and that these activ-
ities are present in both the embryonic and postnatal
meninges. By contrast, a critical developmental change
has taken place in the EGL cells: the embryonic EGL cells
respond predominantly to the attractant in the meninges,
whereas the postnatal EGL cells respond predominantly to
the repellent in the meninges.
Differential sensitivities of the attractive and repulsive
responses to PTX
Because it is known that the same cells can change their
responses to the same molecular cue (Ming et al., 1997;
Poznansky et al., 2000; Song et al., 1997, 1998) and because
e of a postnatal EGL explant with the postnatal meninges. (B) A high
ication view of the proximal side of the EGL explant in A. The scale bar is
(E) Coculture of a postnatal EGL explant with the postnatal Purkinje layer
me explant as that in F with anti-TuJ1. (H) Staining with the nuclear dye. (I)
r is 100 Am. (J) Statistical analysis with the response of postnatal EGL cells;
in PL + IGL. Columns eM and pM are very significantly different from the
).
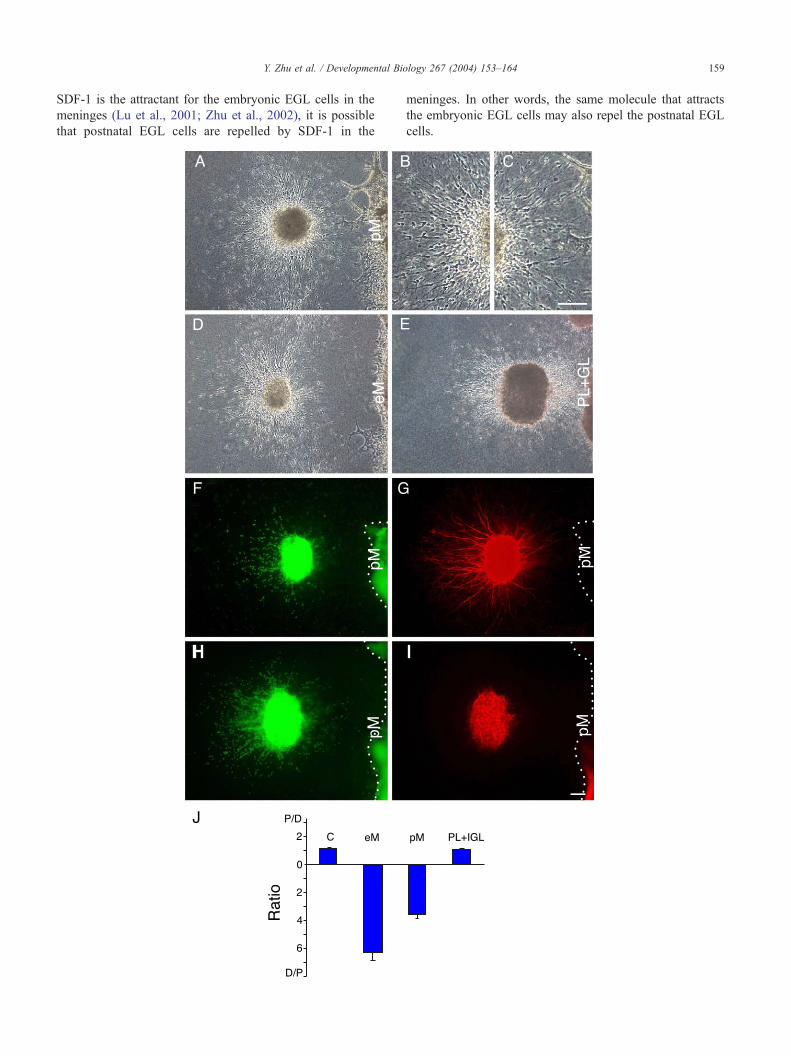
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164 159
SDF-1 is the attractant for the embryonic EGL cells in the
meninges (Lu et al., 2001; Zhu et al., 2002), it is possible
that postnatal EGL cells are repelled by SDF-1 in the
meninges. In other words, the same molecule that attracts
the embryonic EGL cells may also repel the postnatal EGL
cells.
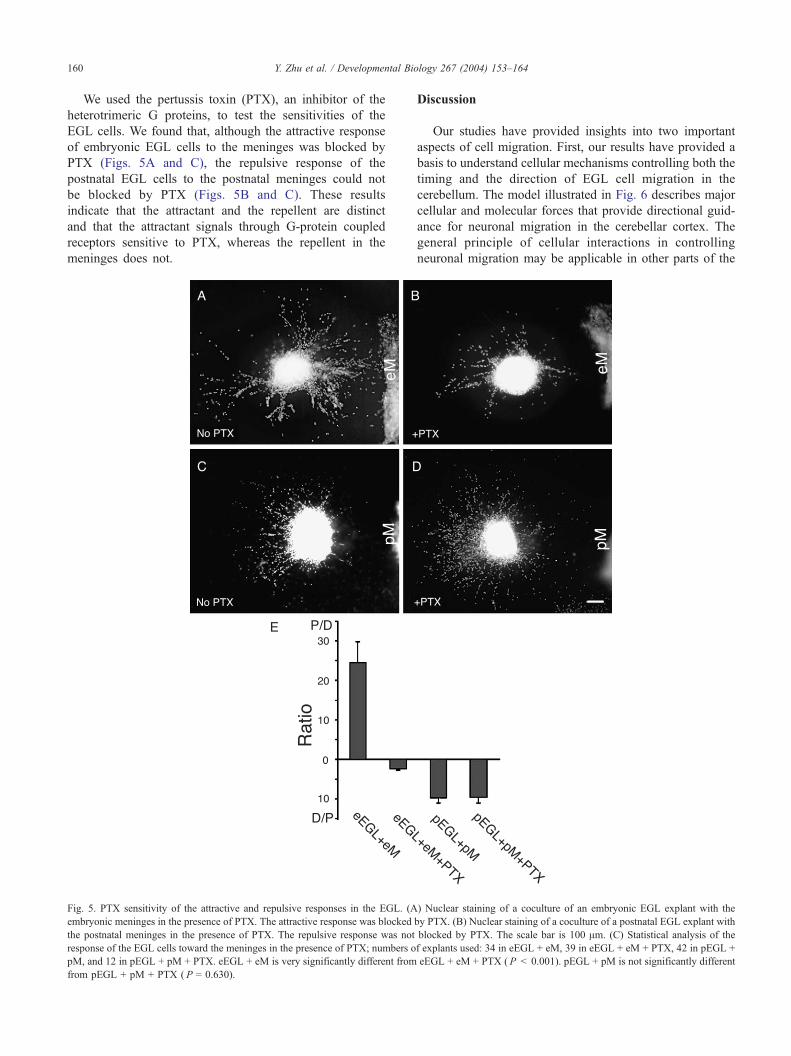
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164160
We used the pertussis toxin (PTX), an inhibitor of the
heterotrimeric G proteins, to test the sensitivities of the
EGL cells. We found that, although the attractive response
of embryonic EGL cells to the meninges was blocked by
PTX (Figs. 5A and C), the repulsive response of the
postnatal EGL cells to the postnatal meninges could not
be blocked by PTX (Figs. 5B and C). These results
indicate that the attractant and the repellent are distinct
and that the attractant signals through G-protein coupled
receptors sensitive to PTX, whereas the repellent in the
meninges does not.
Fig. 5. PTX sensitivity of the attractive and repulsive responses in the EGL. (A
embryonic meninges in the presence of PTX. The attractive response was blocked
the postnatal meninges in the presence of PTX. The repulsive response was not
response of the EGL cells toward the meninges in the presence of PTX; numbers o
pM, and 12 in pEGL + pM + PTX. eEGL + eM is very significantly different from
from pEGL + pM + PTX ( P = 0.630).
Discussion
Our studies have provided insights into two important
aspects of cell migration. First, our results have provided a
basis to understand cellular mechanisms controlling both the
timing and the direction of EGL cell migration in the
cerebellum. The model illustrated in Fig. 6 describes major
cellular and molecular forces that provide directional guid-
ance for neuronal migration in the cerebellar cortex. The
general principle of cellular interactions in controlling
neuronal migration may be applicable in other parts of the
) Nuclear staining of a coculture of an embryonic EGL explant with the
by PTX. (B) Nuclear staining of a coculture of a postnatal EGL explant with
blocked by PTX. The scale bar is 100 Am. (C) Statistical analysis of the
f explants used: 34 in eEGL + eM, 39 in eEGL + eM + PTX, 42 in pEGL +
eEGL + eM + PTX ( P < 0.001). pEGL + pM is not significantly different

Fig. 6. A model for mechanisms underlying temporal and spatial control of EGL cell migration. The left panel is a diagram of the embryonic cerebellum.
Diffusible attractant (+) and repellent (� ) are present in the embryonic meninges. The embryonic EGL cells respond to the attractive cue in the meninges.
There is a diffusible repulsive activity in the neuroepithelium (NE). The combination of the meningeal attractant and the neuroepithelial repellent prevents the
EGL cells from migrating into the internal layers of the embryonic cerebellum. PPL: prospective Purkinje layer. The right panel is a diagram of the postnatal
cerebellum. Both the attractive and repulsive cues are still present in the postnatal meninges. However, the postnatal EGL cells respond predominantly to the
repulsive cue in the meninges. The neuroepithelium is not present in the postnatal cerebellum and there is no repulsive activity in the Purkinje layer and the
IGL. EGL cells will thus migrate towards the IGL.
Y. Zhu et al. / Developmental Biology 267 (2004) 153–164 161
nervous system such as the neocortex. Secondly, our results
demonstrate that an active mechanism is required to keep
inherently mobile cells in place, which can be generally
important for controlling cell positioning not only in the
nervous system but also in other systems.
A model for temporal and spatial control of cell migration
in the cerebellum
We propose here the first mechanistic model for the
temporal and spatial control of cortical migration (Fig. 6).
Cortical migration is controlled by strategic positioning of
multiple attractive and repulsive cues and developmental
changes in the cellular responsiveness to these cues. In the
embryo, although the meninges contain both attractants and
repellents, the embryonic EGL cells respond predominantly
to the meningeal attractant. There is also a diffusible repel-
lent from the neuroepithelium. The combination of the
meningeal attractant and the neuroepithelial repellent pre-
vents the EGL cells from migrating into the internal layers.
Postnatally, we have shown that the inner layers are not
repulsive. However, both the attractant and the repellent are
still present in the postnatal meninges. Due to a change in
cellular responsiveness, postnatal EGL cells respond to the
repellent in the meninges, but not to the attractive activity.
In other parts of the nervous system, the model outlined
in Fig. 6 may also be applicable. For example, in the
neocortex, it is possible that cells from the ventricular zone
are repelled by cues in the ventricular zone and move
towards the outer layers by attractive cues in the outer
layers or in the meninges. In the spinal cord, the same
principle may also apply.
Molecular identities of the chemoattractive and
chemorepulsive factors
Our model for mechanisms underlying temporal and
spatial control of EGL cell migration provides a framework
to put molecular players involving in EGL migration into
proper context. We have uncovered both attractive and
repulsive activities in the meninges and a repulsive activity
in the neuroepithelium. The molecular identity of the
attractant in the meninges has been reported recently (Lu
et al., 2001; Zhu et al., 2002).
The role of SDF-1 as the meningeal attractant has been
proven by several findings. (1) Previous findings from mice
lacking SDF-1 and its receptor CXCR4 have led to the
observation of abnormal neuronal positioning in the IGL
(Ma et al., 1998; Tachibana et al., 1998; Zou et al., 1998).
(2) The expression patterns of SDF-1 and CXCR4 are
consistent of a role for SDF-1 in anchoring the EGL cells
(Klein et al., 2001; Lu et al., 2001; Zhu et al., 2002; Zou et
al., 1998). (3) Transwell assays indicate that SDF-1 can
attract postnatal granule cells (Klein et al., 2001; Lu et al.,
2001). (4) In vitro explant coculture assays demonstrate that
embryonic EGL cells are attracted by SDF-1 whereas
postnatal EGL cells are not (Zhu et al., 2002). (5) Meninges
from SDF-1 knock-out mice no longer attract embryonic
EGL cells (Zhu et al., 2002). These results indicate that
SDF-1 is either the predominant or the sole attractant in the
meninges.
The repellent in the meninges is not known, but it is not
SDF-1 because its activity is still present in SDF-1 knock-
out mice (Zhu et al., 2002) and cannot be blocked by PTX
(Fig. 5).
Anchoring as a new mechanism for controlling cell
migration, and perhaps also for regulating axonal and
dendritic connections
Our finding of the use of an attractant to hold responsive
neurons in place is related to a question of general interest
for cell migration in all systems. Cell migration is usually
inferred from changes in cell positioning during develop-
ment or in adult life. In fact, lack of migration is most often
thought to imply the inability of cells to move, and there is

Y. Zhu et al. / Developmental Biology 267 (2004) 153–164162
little interest in studying cell migration when the apparent
positions of cells do not change. We propose a new
mechanism named anchoring for actively preventing inher-
ently mobile cells from migrating in a spatially or tempo-
rally erroneous manner.
Multiple potential mechanisms can be used to anchor
cells or neuronal processes. In theory, factors that make cells
lose motility transiently or permanently should anchor cells
for a corresponding period of time. Regulation of the
permissiveness of a migratory pathway could also be used
to anchor cells. Adhesive anchoring is another possible
means to anchor cells. This would involve increased cell
adhesion to substrates or neighboring cells.
Our results with the meninges indicate that an attractive
guidance cue plays an unusual role: not by directing cells to
their target region, but by anchoring the cells beneath the
meninges and preventing them from inward migration. This
mode of anchoring is termed attractive anchoring. There can
also be a mode of repulsive anchoring, whereby a repulsive
cue is placed to prevent cells from entering an otherwise
permissive pathway for migration. Thus, when attractive
and repulsive cues are operating in anchoring, the strategic
localization of the guidance cue is crucial.
If anchoring is a general mechanism inside and outside
the nervous system, a conceptual change is required. Not
only should we continue to investigate why cells move
towards their targets, but we will also have to study why
cells do not move away from certain regions. When cells are
apparently stationary for a short or a long time, they may
still be regulated by active mechanisms. The apparently
nonmigratory behavior of these cells or processes makes it
difficult to pinpoint where these mechanisms function in
normal development. Experimental manipulations will be
required to provide important starting points for exploring
functional roles of anchoring.
Similar mechanism may also exist in other neuronal
processes including axons and dendrites that are known to
undergo long- or short-range navigation. Because there are
common mechanisms regulating neuronal migration, and
axonal or dendritic projections (Polleux et al., 2000; Wu et
al., 1999; Yee et al., 1999; Zhu et al., 1999), it will be
interesting to examine whether anchoring can control the
beginning of outgrowth or temporary stops during axonal or
dendritic navigation. The most obvious step for anchoring to
function is at the termination of axonal or dendritic projec-
tion. Anchoring between the pre- and postsynaptic compo-
nents could be an important step in synaptogenesis. It is also
possible for anchoring to play roles in neural plasticity;
competition for anchoring cues may underlie some forms of
plasticity.
A signaling role for the meninges
While the intact meninges provide physical support to
the nervous system, traditionally they have not been thought
to play any important roles in neural development. Recent
work by Hatten et al. suggested that collagen IV expressed
in the pia could be a ligand for the discoidin domain
receptor 1 (DDR1) and that the interaction between collagen
IV and DDR1 might regulate axon extension of cerebellar
granule neurons (Bhatt et al., 2000).
Our results demonstrate that the meninges provide an
anchoring signal for cerebellar EGL cells. Since the meninx
covers other parts of the nervous system, it may also play
essential roles in the development of other regions. In cell
migration, those roles do not have to be anchoring cells near
the surface, they could also be attracting cells from the
deeper layers into more superficial layers, or repelling cells,
axons or dendrites from more superficial layers to deeper
layers. Although we only have evidence for its role in cell
migration, it may not be too speculative to think of other
possible roles for the meninges.
There are three layers in the meninges, the dura mater,
the arachnoid mater, and the pia, with the pia closest to the
EGL (Angelov and Vasilev, 1989; Kamiryo et al., 1990;
O’Rahilly and Mueller, 1986). Because it is technically not
possible to separate the layers and their precursors in
development (Angelov and Vasilev, 1989; Kamiryo et al.,
1990), we could not test the roles of individual layers of the
meninges for their signaling roles. However, the presence of
SDF-1 in all three layers suggests that there may not be
much difference (Zhu et al., 2002).
Questions about the precise role of glial fibers in neuronal
migration
The migration of EGL cells towards the IGL has long
been thought to depend on radial glial fibers (Hatten, 1999;
Hatten and Heintz, 1995; Rakic, 1971a,b, 1990). Because of
the linear shapes of glial processes, the mere action of
climbing along glial processes may greatly reduce the
possible migratory directions into two. We intended to
address whether radial glia also provide directional guidance
cues for neurons to move one direction vs. another one. Our
results with cells migrating out of EGL explants in vitro
raise questions about the precise role of the glial fibers in
neuronal migration.
In vitro, EGL cells can bind to and migrate along the
glial fibers (Edmondson and Hatten, 1987; Gregory et al.,
1988; Hatten and Liem, 1981; Hatten and Mason, 1990;
Mason et al., 1988). The binding of EGL cells to the glial
fiber requires the cell adhesion molecule astrotactin
(Edmondson et al., 1988; Stitt and Hatten, 1990; Fishell
and Hatten, 1991; Zheng et al., 1996). Because the same
neuron can migrate in both directions along the same glial
fiber (Edmondson et al., 1987; Gregory et al., 1988; Hatten
and Liem, 1981; Mason et al., 1988), the glial fibers
unlikely provide information to guide the direction of
neuronal migration under these conditions. In our studies
of EGL cell migration, the three dimensional matrices are
known to contain extracellular matrix proteins (Kleinman et
al., 1982). Neurons migrating from URL or EGL explants

Y. Zhu et al. / Developmental Biology 267 (2004) 153–164 163
were not associated with glial fibers. When neurons were
attracted or repelled by the meninges, they were not asso-
ciated with glial fibers. The glial fibers therefore are not
required in mediating EGL responses to the guidance cues,
which must act directly on neurons.
The conclusions reached from in vitro studies also agree
with in vivo results. Altman and Bayer (1997, pp. 341–347)
reported that migrating EGL cells appeared to move sepa-
rately in vivo and were not associated with glial or neuronal
fibers in postnatal rats of 5, 7 or 10 days of age. After P12,
neurons seem to migrate along the fibers of other neurons in
vivo and along glial fibers later (Altman and Bayer, 1997).
Recently, Nadarajah et al. (2001, 2002) report that, while
neuronal precursor cells in the rat neocortex after E15
prevalently move along glial fibers, early migrating neurons
are not associated with glial fibers.
Our results indicate that the presence of glial fibers is not
absolutely required for EGL neurons to migrate. In vitro
studies of live granule cells indicate that, when glass fibers
were coated with extracellular matrix proteins such as
laminin or fibronectin, granule cells can migrate along the
coated glass fibers (Fishman and Hatten, 1993). This sug-
gests that deposition of extracellular matrix proteins is
sufficient to replace glial fibers. Under our culture condi-
tions, migrating neurons from E15 URL and postnatal EGL
explants appear to be associated with tracks of neuronal
fibers. Although neurons from E17 EGL explants have
leading processes, cell bodies of individual neurons of
E17 explants were not associated with tracks of fibers of
other neurons. At least in the case of E17 neurons, the effect
of guidance cues is unlikely to be mediated by their action
on tracks of neuronal fibers.
The precise roles of the glial fibers are thus not very
clear. Are they facilitating neurons to move faster? Is the
association of neurons and glial fibers used for coordinating
their differentiation than for migration? Are they limiting
neurons to migrate along one dimension among the three
dimensional space? It seems that ablation of specific glial
fibers along which neuronal migration could be traced in
live brain slices is one way to investigate the precise role of
glial fibers.
A related question is the subcellular structure in a single
migrating neuron that senses the guidance cues: is it the cell
body or the leading process? To answer this question, one
may have to develop a method to specifically deliver the
guidance cue to the cell body without affecting the leading
process or to the tip of the leading process without affecting
the cell body.
Acknowledgments
We are grateful to Huashun Li for participating in the
early phase of this project; to Jennifer Jones for assistance;
to Jane E. Johnson for the anti-Math1 antibody; to Guofa
Liu, Harumi Saito and Yong Yin for help; to Alan Pearlman,
Joshua Sanes, Joseph Price, Huashun Li and Bagi Nadaraja
for helpful discussions; to the NIH, the Klingenstein
foundation, and the John Merck fund for support (to Y. R.).
References
Alder, J., Cho, N.K., Hatten, M.E., 1996. Embryonic precursor cells from
the rhombic lip are specified to a cerebellar granule neuron identity.
Neuron 17, 389–399.
Altman, J., Bayer, S.A., 1997. Development of the Cerebellar System. CRC
Press, New York.
Angelov, D.N., Vasilev, V.A., 1989. Morphogenesis of rat cranial meninges.
A light- and electron-microscopic study. Cell Tissue Res. 257, 207–216.
Bhatt, R.S., Tomoda, T., Fang, Y., Hatten, M.E., 2000. Discoidin domain
receptor 1 functions in axon extension of cerebellar granule neurons.
Genes Dev. 14, 2216–2228.
Chae, T., Kwon, Y.T., Bronson, R., Dikkes, P., Li, E., Tsai, L.H., 1997.
Mice lacking p35, a neuronal specific activator of Cdk5, display cortical
lamination defects, seizures, and adult lethality. Neuron 18, 29–42.
D’Arcangelo, G., Miao, G.G., Chen, S.-C., Soares, H.D., Morgan, J.I.,
Curran, T., 1995. A protein related to extracellular matrix proteins
deleted in the mouse mutant reeler. Nature 374, 719–723.
D’Arcangelo, G., Homayouni, R., Keshvara, L., Rice, D.S., Sheldon, M.,
Curran, T., 1999. Reelin is a ligand for lipoprotein receptors. Neuron
24, 471–479.
Dhavan, R., Tsai, L.H., 2001. A decade of cdk5. Nat. Rev., Mol. Cell Biol.
2, 749–759.
Edmondson, J.C., Hatten, M.E., 1987. Glial-guided granule neuron migra-
tion in vitro: a high-resolution time-lapse video microscopic study.
J. Neurosci. 7, 1928–1934.
Edmondson, J.C., Liem, R.K., Kuster, J.E., Hatten, M.E., 1988. Astro-
tactin: a novel neuronal cell surface antigen that mediates neuron–
astroglial interactions in cerebellar microcultures. J. Cell Biol. 106,
505–517.
Fishell, G., Hatten, M.E., 1991. Astrotactin provides a receptor system for
CNS neuronal migration. Development 113, 755–765.
Fishman, R.B., Hatten, M.E., 1993. Multiple receptor systems promote
CNS neural migration. J. Neurosci. 13, 3485–3495.
Gregory, W.A., Edmondson, J.C., Hatten, M.E., Mason, C.A., 1988. Cy-
tology and neuron–glial apposition of migrating cerebellar granule cells
in vitro. J. Neurosci. 8, 1728–1934.
Hatten, M.E., 1999. Central nervous system neuronal migration. Annu.
Rev. Neurosci. 22, 511–539.
Hatten, M.E., Liem, R.H.K., 1981. Astroglia provide a template for the
positioning of developing cerebellar neurons in vitro. J. Cell Biol. 90,
622–630.
Hatten, M.E., Mason, C.A., 1990. Mechanisms of glial-guided neuronal
migration in vitro and in vivo. Experientia 46, 907–916.
Hatten, M.E., Liem, R.K., Mason, C.A., 1984. Defects in specific associ-
ations between astroglia and neurons occur in microcultures of weaver
mouse cerebellar cells. J. Neurosci. 4, 1163–1172.
Hiesberger, T., Trommsdorff, M., Howell, B.W., Goffinet, A., Mumby,
M.C., Cooper, J.A., Herz, J., 1999. Direct binding of Reelin to VLDL
receptor and ApoE receptor 2 induces tyrosine phosphorylation of the
adaptor protein Disabled-1 and modulates tau phosphorylation. Neuron
24, 481–489.
Hopfield, S., McKay, R.D., 1985. Identification of major cell classes in the
developing mammalian nervous system. J. Neurosci. 5, 3310–3328.
Howell, B.W., Hawkes, R., Soriano, P., Cooper, J.A., 1997. Neuronal
position in the developing brain is regulated by mouse disabled-1.
Nature 389, 733–737.
Kamiryo, T., Orita, T., Nishizaki, T., Aoki, H., 1990. Development of the
rat meninx: experimental study using Bromodeoxyuridine. Anat. Rec.
227, 207–210.

Y. Zhu et al. / Developmental Biology 267 (2004) 153–164164
Klein, R.S., Rubin, J.B., Gibson, H.D., DeHaan, E.N., Alvarez-Hernandez,
X., Segal, R.A., Luster, A.D., 2001. SDF-1a induces chemotaxis and
enhances Sonic hedgehog-induced proliferation of cerebellar granule
cells. Development 128, 1971–1981.
Kleinman, H.K., McGravey, M.L., Liotta, L.A., Robey, P.G., Tryggvason,
K., Martin, G.R., 1982. Isolation and characterization of type IV pro-
collagen, laminin, and heparan sulfate proteoglycan from the EHS sar-
coma. Biochemistry 21, 6188–6193.
Letinic, K., Rakic, P., 2001. Telencephalic origin of human thalamic GA-
BAergic neurons. Nat. Neurosci. 4, 931–936.
Li, H.S., Chen, J.H., Wu, W., Fagaly, T., Zhou, L., Yuan, W., Dupuis,
S., Jiang, Z.H., Nash, W., Gick, C., Ornitz, D.M., Wu, J.Y., Rao,
Y., 1999. Vertebrate slit, a secreted ligand for the transmembrane
protein roundabout, is a repellent for olfactory bulb axons. Cell 96,
807–818.
Lu, Q., Sun, E., Klein, R.S., Flanagan, J.G., 2001. Ephrin-B reverse signal-
ing is mediated by a novel PDZ-RGS protein and selectively inhibits G
protein-coupled chemoattraction. Cell 105, 69–79.
Ma, Q., Jones, D., Borghesani, P.R., Segal, R.A., Nagasawa, T., Kishi-
moto, T., Bronson, R.T., Springer, T.A., 1998. Impaired B-lymphopoi-
esis, myelopoiesis, and derailed cerebellar neuron migration in
CXCR4- and SDF-1-deficient mice. Proc. Natl. Acad. Sci. U. S. A.
95, 9448–9453.
Marin, O., Yaron, A., Bagri, A., Tessier-Lavigne, M., Rubenstein, J.L.,
2001. Sorting of striatal and cortical interneurons regulated by sema-
phorin–neuropilin interactions. Science 293, 872–875.
Mason, C.A., Edmondson, J.C., Hatten, M.E., 1988. The extending astro-
glial process: development of glial cell shape, the growing tip, and
interactions with neurons. J. Neurosci. 8, 3124–3134.
Ming, G.-L., Song, H.-J., Berninger, B., Holt, C., Tessier-Lavigne, M., Poo,
M.-M., 1997. cAMP-dependent growth cone guidance by netrin-1.
Neuron 19, 1225–1235.
Nadarajah, B., Brunstrom, J.E., Grutzendler, J., Wong, R.O., Pearlman,
A.L., 2001. Two modes of radial migration in early development of
the cerebral cortex. Nat. Neurosci. 4, 143–150.
Nadarajah, B., Alifragis, P., Wong, R.O., Parnavelas, J.G., 2002. Ventricle-
directed migration in the developing cerebral cortex. Nat. Neurosci. 5,
218–224.
Nikolic, M., Dudek, H., Kwon, Y.T., Ramos, Y.F., Tsai, L.H., 1996. The
cdk5/p35 kinase is essential for neurite outgrowth during neuronal dif-
ferentiation. Genes Dev. 10, 816–825.
O’Rahilly, R., Mueller, F., 1986. The meninges in human development. J.
Neuropathol. Exp. Neurol. 45, 588–608.
Pearlman, A.L., Faust, P.L., Hatten, M.E., Brunstrom, J.E., 1998. New
directions for neuronal migration. Curr. Opin. Neurobiol. 8, 45–54.
Pleasure, S.J., Anderson, S., Hevner, R., Bagri, A., Marin, O., Lowenstein,
D.H., Rubenstein, J.L., 2000. Cell migration from the ganglionic emi-
nences is required for the development of hippocampal GABAergic
interneurons. Neuron 28, 727–740.
Polleux, F., Morrow, T., Ghosh, A., 2000. Semaphorin 3A is a chemo-
attractant for cortical apical dendrites. Nature 404, 567–573.
Powell, E.M., Mars, W.M., Levitt, P., 2001. Hepatocyte growth factor/
scatter factor is a motogen for interneurons migrating from the ventral
to dorsal telencephalon. Neuron 30, 79–89.
Poznansky, M.C., Olszak, I.T., Foxall, R., Evans, R.H., Luster, A.D., Scad-
den, D.T., 2000. Active movement of T cells away from a chemokine.
Nat. Med. 6, 543–548.
Rakic, P., 1971a. Neuron–glia relationship during granule cell migration in
developing cerebellar cortex. J. Comp. Neurol. 141, 283–312.
Rakic, P., 1971b. Guidance of neurons migrating to the fetal monkey neo-
cortex. Brain Res. 33, 471–476.
Rakic, P., 1990. Principles of neural cell migration. Experientia 46,
882–891.
Rice, D.S., Curran, T., 1999. Mutant mice with scrambled brains: under-
standing the signaling pathways that control cell positioning in the
CNS. Genes Dev. 13, 2758–2773.
Rice, D.S., Curran, T., 2001. Role of the Reelin signaling pathway in
central nervous system development. Annu. Rev. Neurosci. 24,
1005–1039.
Ross, M.E., Walsh, C.A., 2001. Human brain malformations and their
lessons for neuronal migration. Annu. Rev. Neurosci. 24, 1041–1070.
Sheldon, M., Rice, D.S., D’Arcangelo, G., Yoneshima, H., Nakajima, K.,
Mikoshiba, K., Howell, B.W., Cooper, J.A., Goldowitz, D., Curran, T.,
1997. Scrambler and yotari disrupt the disabled gene and produce a
reeler-like phenotype in mice. Nature 389, 730–733.
Song, H.-J., Ming, G.-L., Poo, M.-M., 1997. A cAMP-induced switching
of turning direction of nerve growth cones. Nature 388, 275–279.
Song, H.-J., Ming, G.-L., He, Z., Lehmann, M., McKerracher, L., Tessier-
Lavigne, M., Poo, M.-M., 1998. Conversion of neuronal growth cone
responses from repulsion to attraction by cyclic nucleotides. Science
281, 1515–1518.
Sotelo, C., Alvarado-Mallart, R.M., Frain, M., Vernet, M., 1994. Molecular
plasticity of adult Bergmann fibers is associated with radial migration of
grafted Purkinje cells. J. Neurosci. 14, 124–133.
Stitt, T.N., Hatten, M.E., 1990. Antibodies that recognize astrotactin block
granule neuron binding to astroglia. Neuron 5, 639–649.
Sweet, H.O., Bronson, R.T., Johnson, K.R., Cook, S.A., Davisson,
M.T., 1996. Scrambler, a new neurological mutation of the mouse
with abnormalities of neuronal migration. Mamm. Genome 7,
798–802.
Tachibana, K., Hirota, S., Iizasa, H., Yoshida, H., Kawabata, K., Kataoka,
Y., Kitamura, Y., Matsushima, K., Yoshida, N., Nishikawa, S., Kishi-
moto, T., Nagasawa, T., 1998. The chemokine receptor CXCR4 is
essential for vascularization of the gastrointestinal tract. Nature 393,
591–594.
Trommsdorff, M., Gotthardt, M., Hiesberger, T., Shelton, J., Stockinger,
W., Nimpf, J., Hammer, R.E., Richardson, J.A., Herz, J., 1999. Reeler/
disabled-like disruption of neuronal migration in knock-out mice lacking
the VLDL receptor and ApoE receptor 2. Cell 97, 689–701.
Tsai, L.H., Delalle, I., Caviness Jr., V.S., Chae, T., Harlow, E., 1994. p35 is
a neural-specific regulatory subunit of cyclin-dependent kinase 5. Na-
ture 371, 419–423.
Ware, M.L., Fox, J.W., Gonzale, J.L., Davis, N.M., Lambert de Rouvroit,
C., Russo, C.J., Chua Jr., S.C., Goffinet, A.M., Walsh, C.A., 1997.
Aberrant splicing of a mouse disabled homolog, mdab1, in the scram-
bler mouse. Neuron 19, 239–249.
Wechsler-Reya, R.J., Scott, M.P., 1999. Control of neuronal precursor pro-
liferation in the cerebellum by sonic Hedgehog. Neuron 22, 103–114.
Wingate, R.J.T., Hatten, M.E., 1999. The role of the rhombic lip in avian
cerebellum development. Development 126, 4395–4404.
Wong, K., Ren, X.R., Huang, Y.Z., Xie, Y., Liu, G., Saito, H., Tang, H.,
Wen, L., Brady-Kalnay, S.M., Mei, L., Wu, J.Y., Xiong, W.C., Rao, Y.,
2001. Signal transduction in neuronal migration: roles of GTPase acti-
vating proteins and the small GTPase Cdc42 in the Slit–Robo pathway.
Cell 107, 209–221.
Wu, W., Wong, K., Chen, J.H., Jiang, Z.H., Dupuis, S., Wu, J.Y., Rao, Y.,
1999. Directional guidance of neuronal Migration in the olfactory sys-
tem by the secreted protein Slit. Nature 400, 331–336.
Yee, K.T., Simon, H.H., Tessier-Lavigne, M., O’Leary, D.D.M., 1999.
Extension of long leading processes and neuronal migration in the
mammalian brain directed by the chemoattractant Netrin-1. Neuron
24, 607–622.
Zheng, C., Heintz, N., Hatten, M.E., 1996. CNS gene encoding astrotactin,
which supports neuronal migration along glial fibers. Science 272,
417–419.
Zhu, Y., Li, H.S., Zhou, L., Wu, J.Y., Rao, Y., 1999. Cellular and molecular
guidance of GABAergic neuronal migration from the striatum to the
neocortex. Neuron 23, 473–485.
Zhu, Y., Yu, T., Zhang, X.-C., Nagasawa, T., Wu, J.Y., Rao, Y., 2002. Role
of the chemokine SDF-1 as the meningeal attractant for embryonic
cerebellar neurons. Nat. Neurosci. 5, 719–720.
Zou, Y.R., Kottmann, A.H., Kuroda, M., Taniuchi, I., Littman, D.R., 1998.
Function of the chemokine receptor CXCR4 in haematopoiesis and in
cerebellar development. Nature 393, 595–599.