Embed Size (px)
Citation preview

Synthetic pathways for aromatics
production from biobased feedstock
Harald Ruijssenaars
BIRD Engineering
Delft-Schiedam, The Netherlands

BIRD Engineering
Contract research company in Delft, The Netherlands• fermentation / medium optimization services
• strain development services (yeast; bacteria)
• non-food market (biofuels, bio-based chemicals)
Collaboration / Customers• (a.o.) Mascoma, Amyris, DSM, Tate&Lyle, Nedalco, Applikon, Heineken
• Delft University of Technology
• Knowledge networks: Kluyver Centre, BE-Basic, CLIB (Germany), SIM (USA)
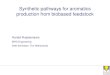
Substituted (hydroxy-)aromatics
•many & diverse applications, e.g., in plastics (LCP‟s),
resins, fibers
•fossil-based
•often difficult to synthesize chemically
•improved / novel functionality with biotech OH
R
OH
Rresins
polymers
specialties
........
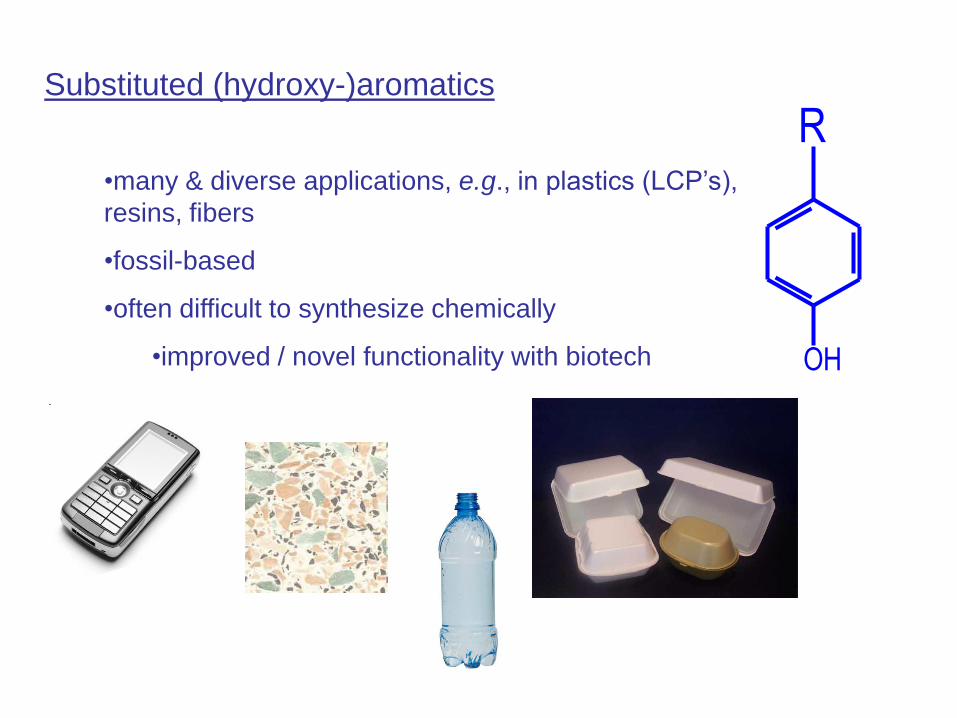
Biocatalytic production of aromatics - bioconversion
1. product toxicity
2. product formation
3. feedstock utilization
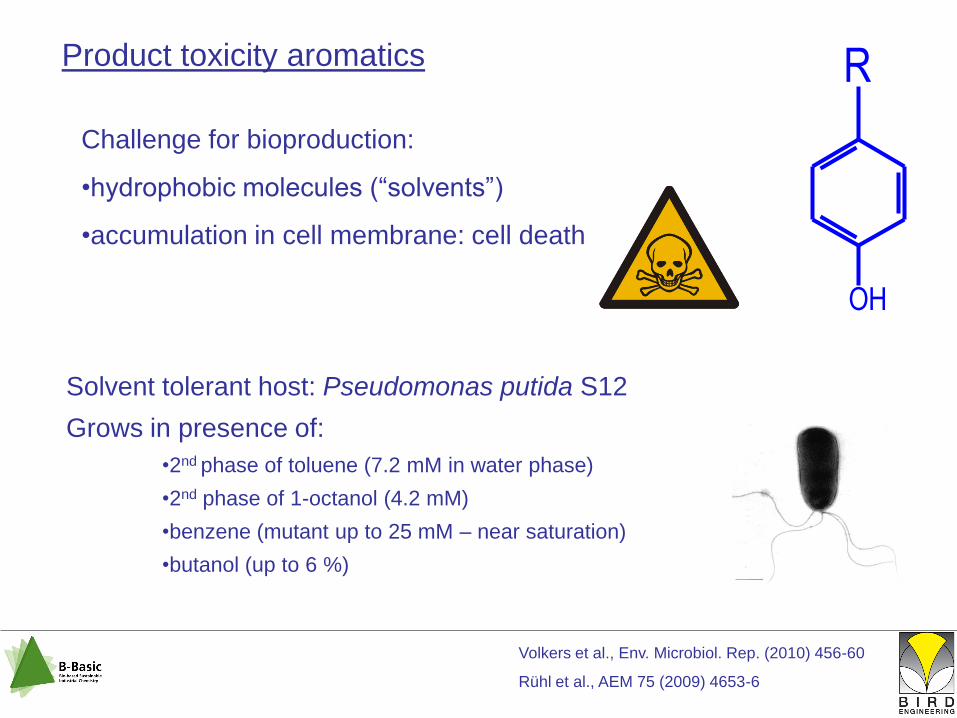
Product toxicity aromatics
Challenge for bioproduction:
•hydrophobic molecules (“solvents”)
•accumulation in cell membrane: cell death
OH
R
Solvent tolerant host: Pseudomonas putida S12
Grows in presence of:
•2nd phase of toluene (7.2 mM in water phase)
•2nd phase of 1-octanol (4.2 mM)
•benzene (mutant up to 25 mM – near saturation)
•butanol (up to 6 %)
Volkers et al., Env. Microbiol. Rep. (2010) 456-60
Rühl et al., AEM 75 (2009) 4653-6
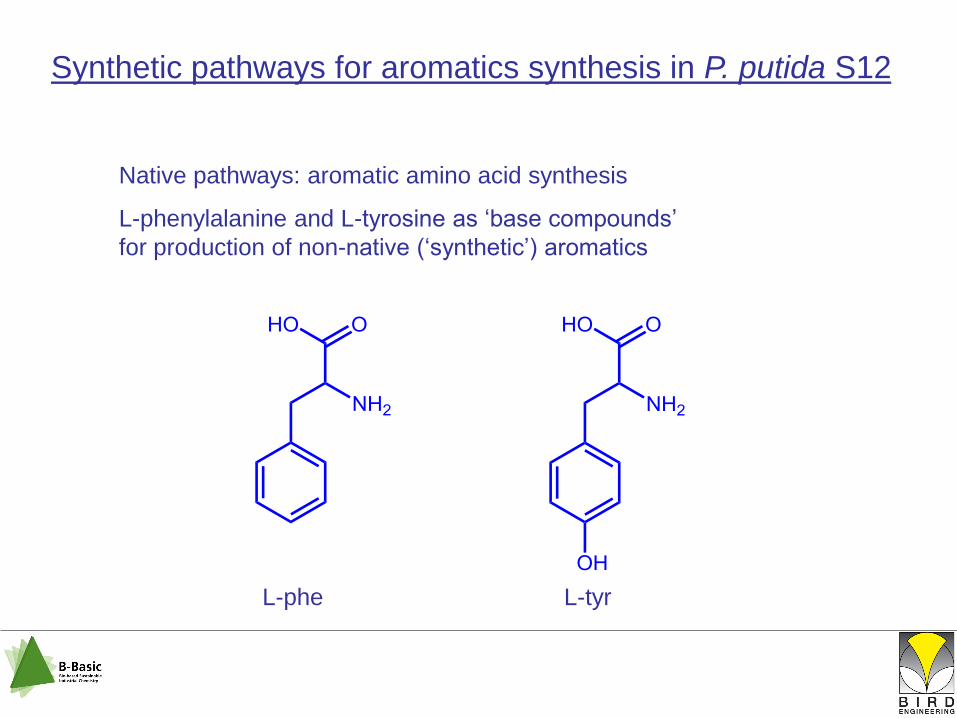
Synthetic pathways for aromatics synthesis in P. putida S12
Native pathways: aromatic amino acid synthesis
L-phenylalanine and L-tyrosine as „base compounds‟
for production of non-native („synthetic‟) aromatics
L-phe L-tyr
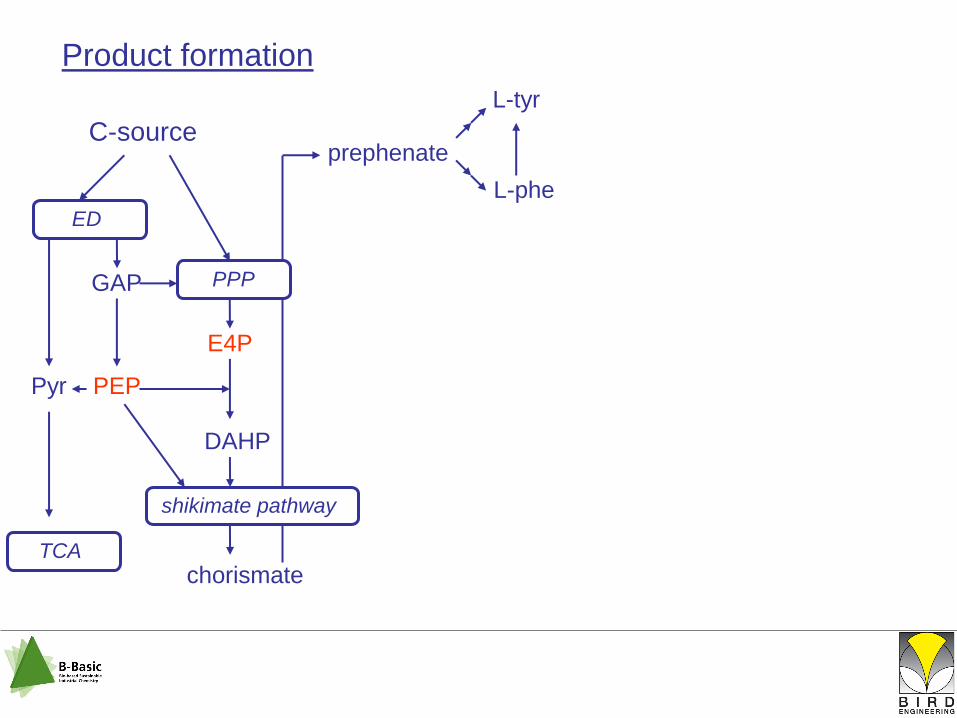
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
Product formation
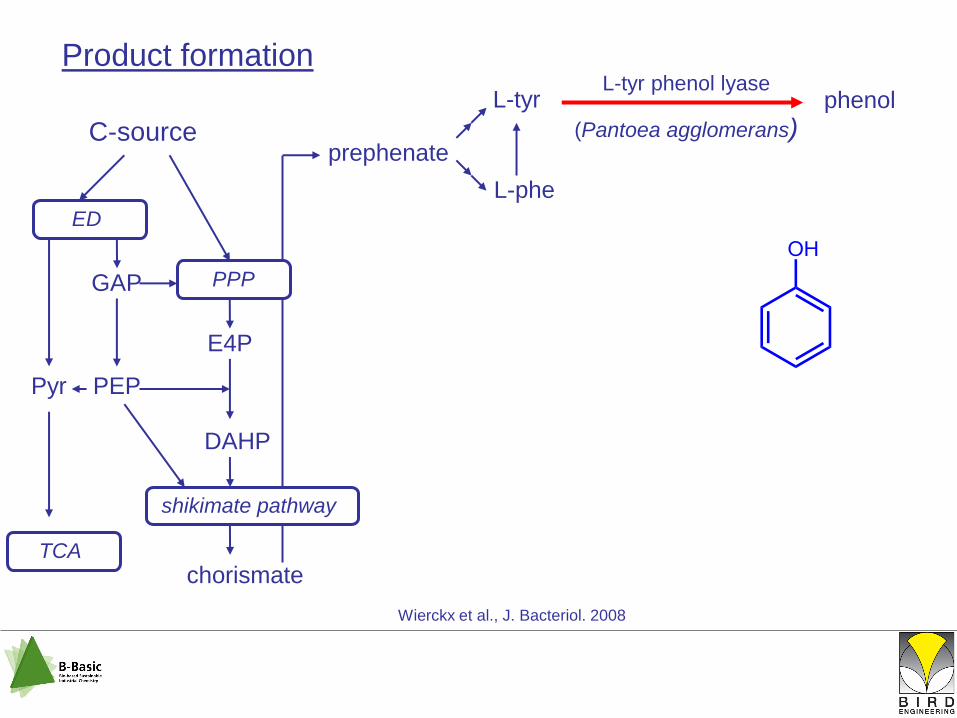
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
phenolL-tyr phenol lyase
(Pantoea agglomerans)
Wierckx et al., J. Bacteriol. 2008
Product formation
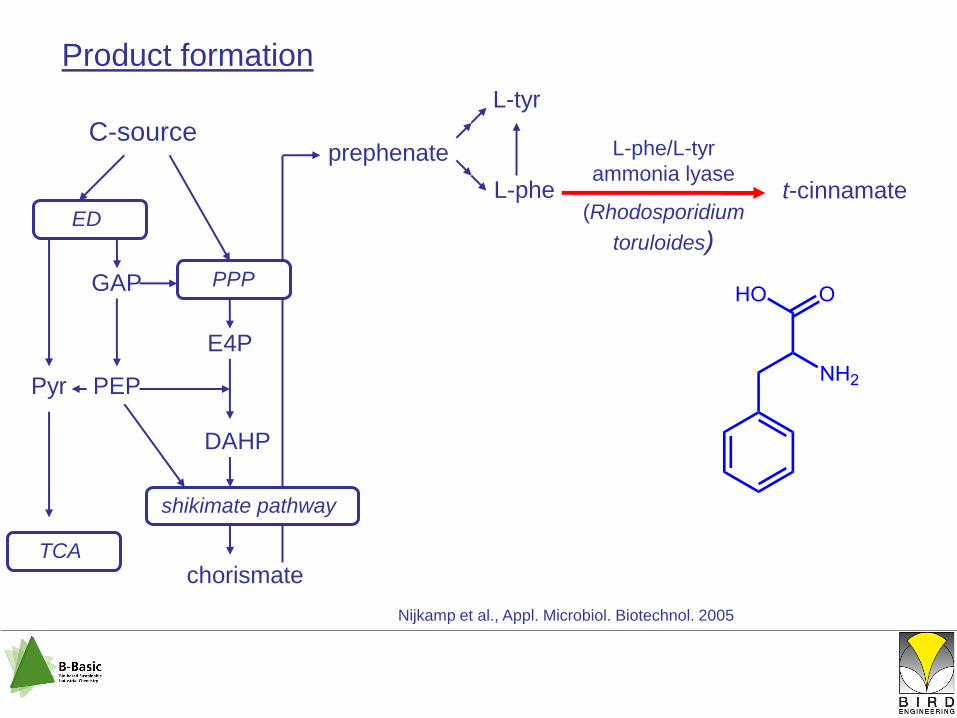
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
t-cinnamate
L-phe/L-tyr
ammonia lyase
(Rhodosporidium
toruloides)
Nijkamp et al., Appl. Microbiol. Biotechnol. 2005
Product formation
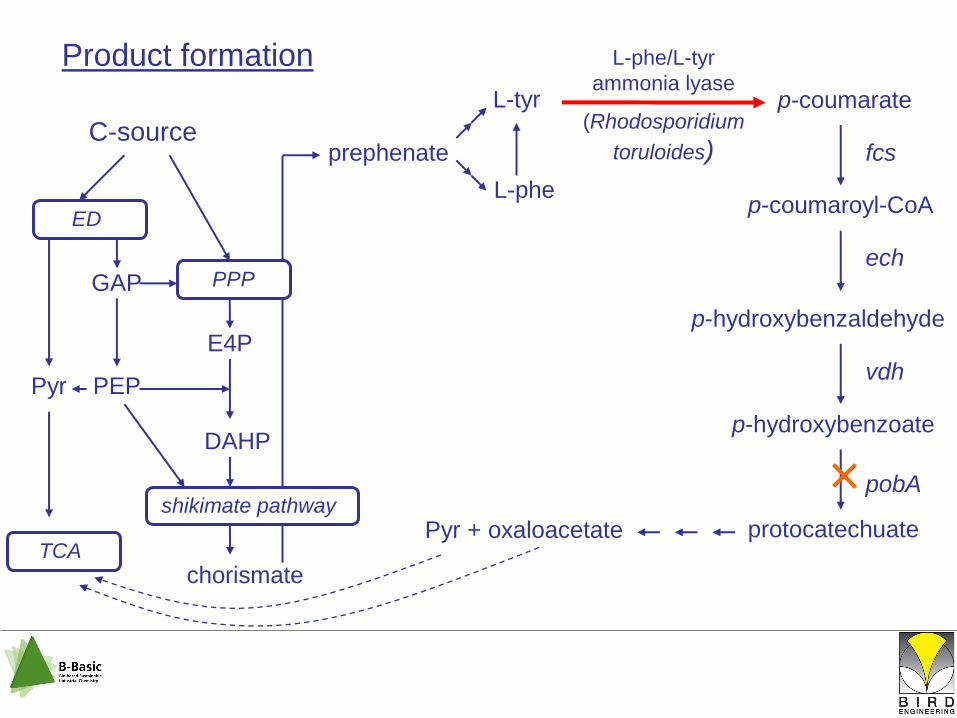
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
p-coumarate
L-phe/L-tyr
ammonia lyase
(Rhodosporidium
toruloides)
p-coumaroyl-CoA
p-hydroxybenzaldehyde
p-hydroxybenzoate
protocatechuatePyr + oxaloacetate
fcs
ech
vdh
pobA
Product formation
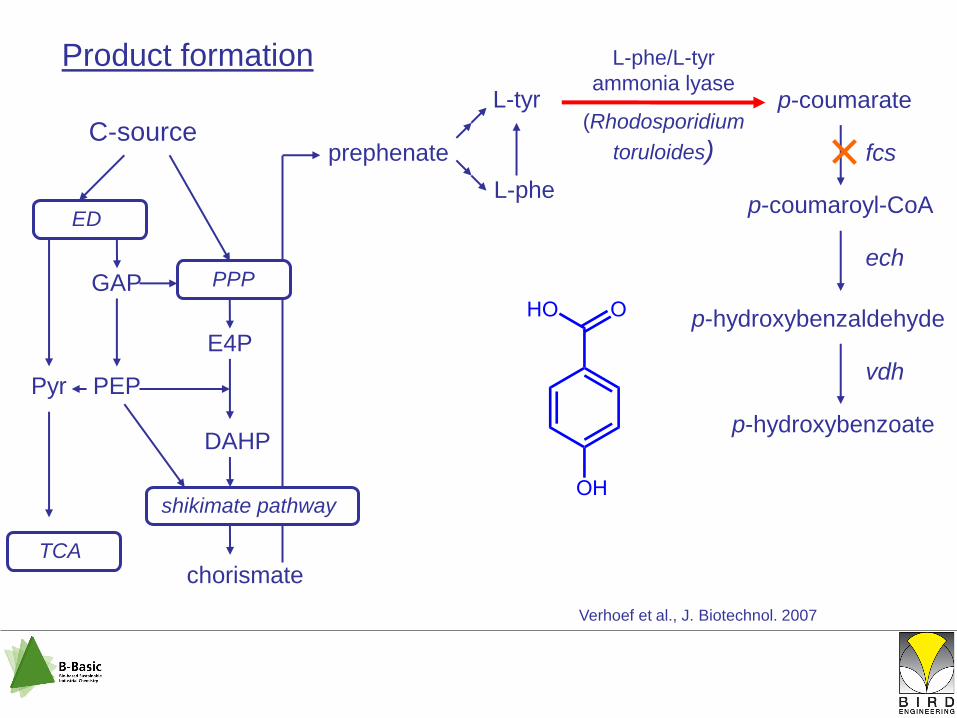
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
p-coumarate
L-phe/L-tyr
ammonia lyase
(Rhodosporidium
toruloides)
p-coumaroyl-CoA
p-hydroxybenzaldehyde
p-hydroxybenzoate
fcs
ech
vdh
Verhoef et al., J. Biotechnol. 2007
Product formation
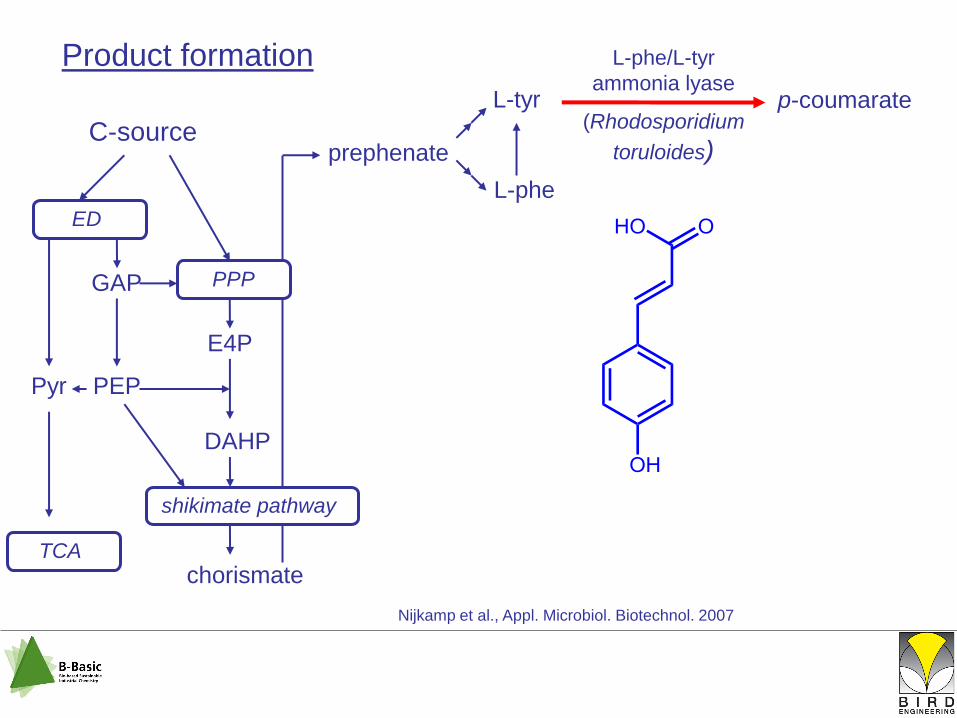
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
p-coumarate
L-phe/L-tyr
ammonia lyase
(Rhodosporidium
toruloides)
Nijkamp et al., Appl. Microbiol. Biotechnol. 2007
Product formation
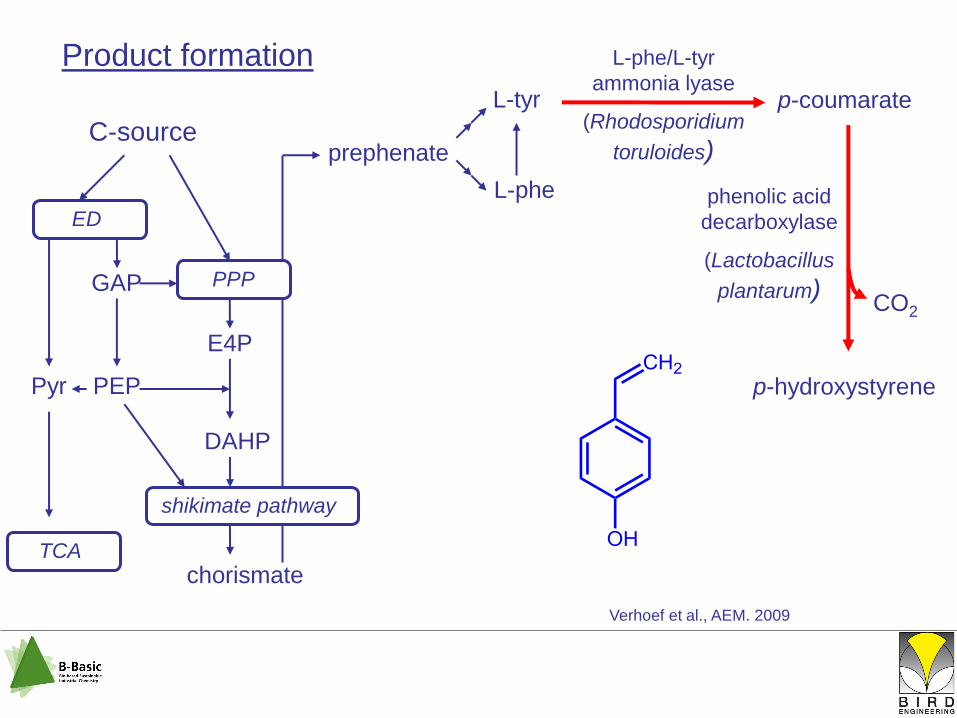
L-tyr
shikimate pathway
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
p-coumarate
L-phe/L-tyr
ammonia lyase
(Rhodosporidium
toruloides)
Verhoef et al., AEM. 2009
phenolic acid
decarboxylase
(Lactobacillus
plantarum)
p-hydroxystyrene
CO2
Product formation
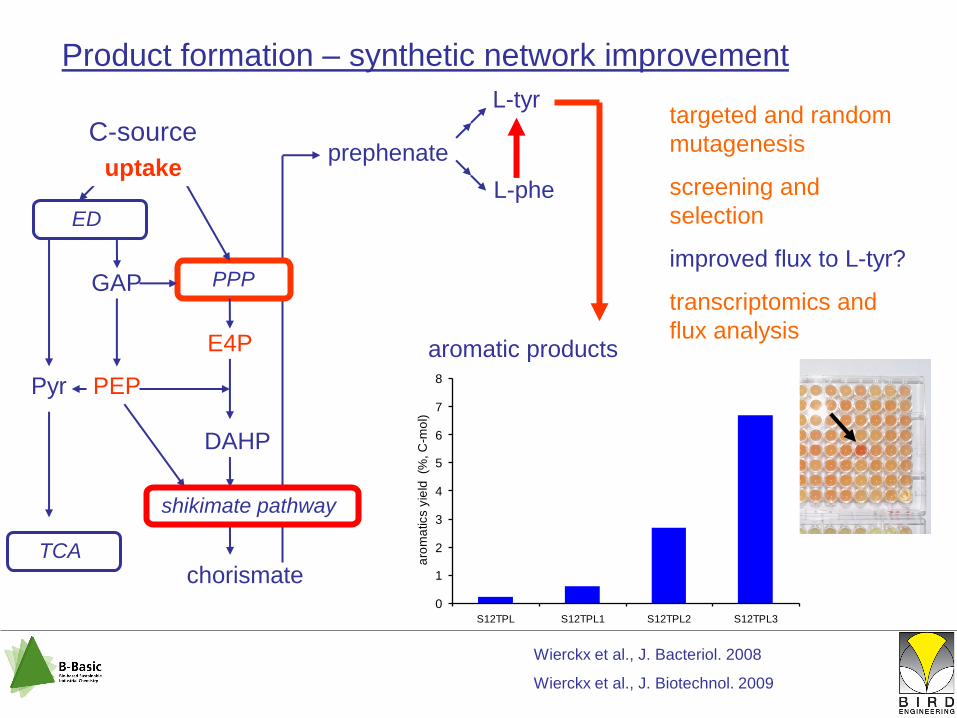
0
1
2
3
4
5
6
7
8
S12TPL S12TPL1 S12TPL2 S12TPL3
aro
ma
tics y
ield
(%
, C
-mo
l)
L-tyr
C-source
chorismate
ED
PPPGAP
E4P
DAHP
L-phe
prephenate
PEPPyr
TCA
Product formation – synthetic network improvement
targeted and random
mutagenesis
screening and
selection
improved flux to L-tyr?
transcriptomics and
flux analysis
Wierckx et al., J. Bacteriol. 2008
Wierckx et al., J. Biotechnol. 2009
uptake
shikimate pathway
aromatic products
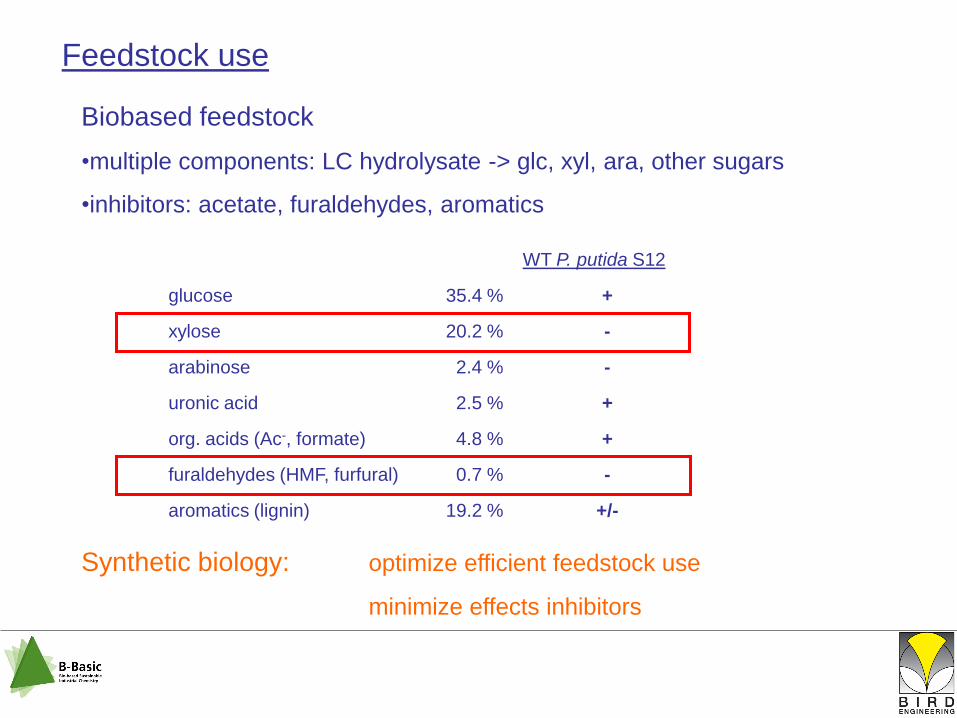
Feedstock use
Biobased feedstock
•multiple components: LC hydrolysate -> glc, xyl, ara, other sugars
•inhibitors: acetate, furaldehydes, aromatics
Synthetic biology: optimize efficient feedstock use
minimize effects inhibitors
WT P. putida S12
glucose 35.4 % +
xylose 20.2 % -
arabinose 2.4 % -
uronic acid 2.5 % +
org. acids (Ac-, formate) 4.8 % +
furaldehydes (HMF, furfural) 0.7 % -
aromatics (lignin) 19.2 % +/-
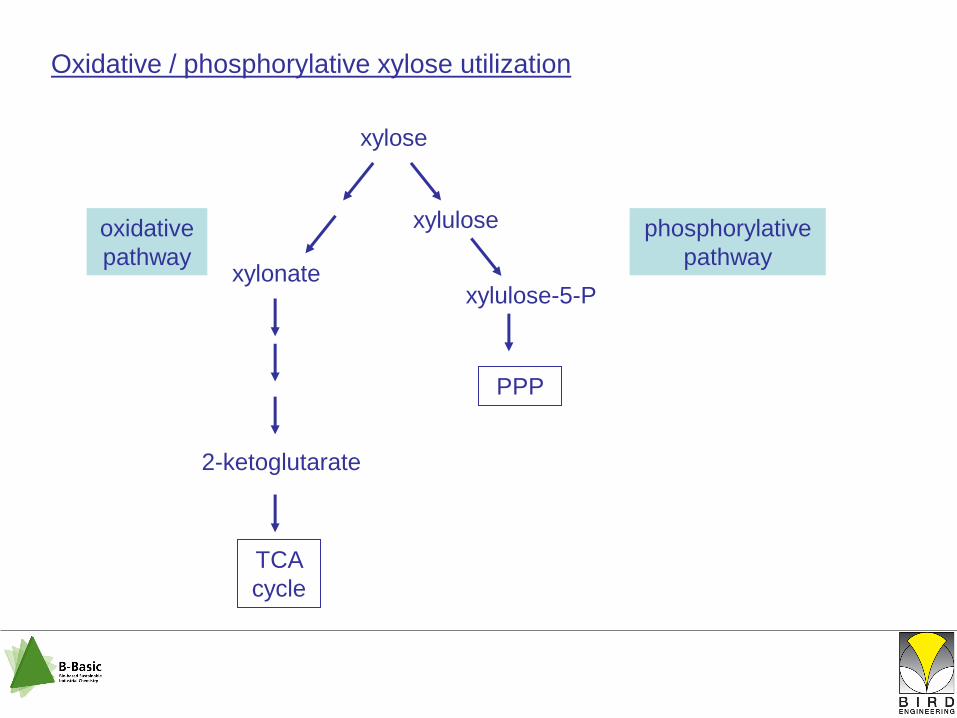
Oxidative / phosphorylative xylose utilization
xylose
xylonate
2-ketoglutarate
TCA
cycle
xylulose
xylulose-5-P
PPP
oxidative
pathway
phosphorylative
pathway
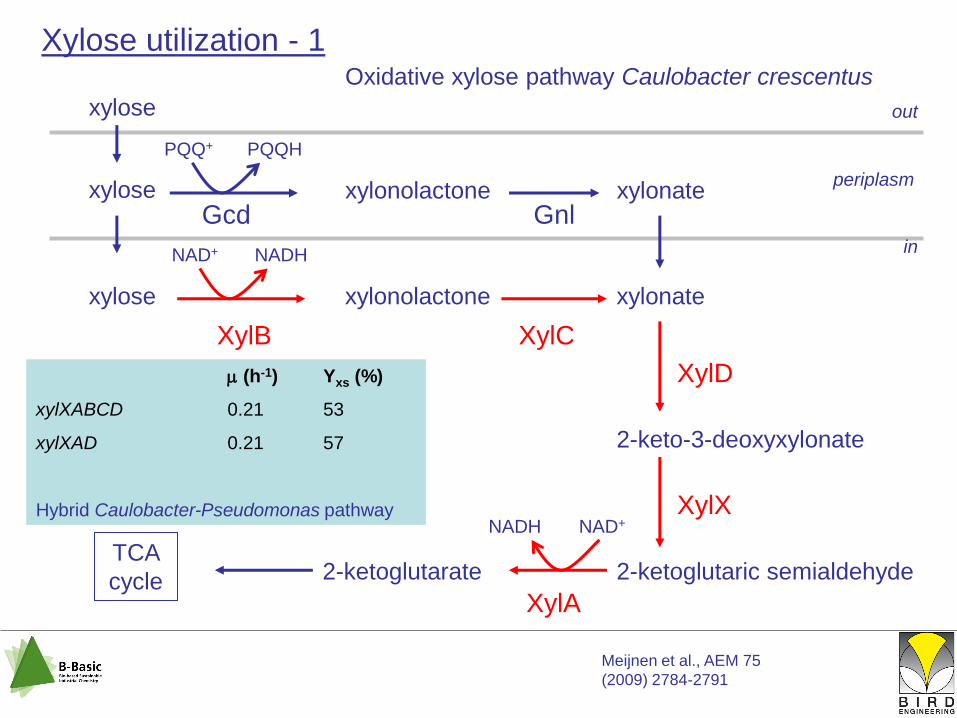
xylonate
2-keto-3-deoxyxylonate
2-ketoglutaric semialdehyde2-ketoglutarateTCA
cycle
NAD+NADH
XylD
XylX
XylA
xylose
periplasm
out
in
xylose
PQQ+ PQQH
Gcdxylonolactone xylonate
Gnl
Meijnen et al., AEM 75
(2009) 2784-2791
Xylose utilization - 1Oxidative xylose pathway Caulobacter crescentus
xylose xylonolactone
NAD+ NADH
XylB XylC
(h-1) Yxs (%)
xylXABCD 0.21 53
xylXAD 0.21 57
Hybrid Caulobacter-Pseudomonas pathway
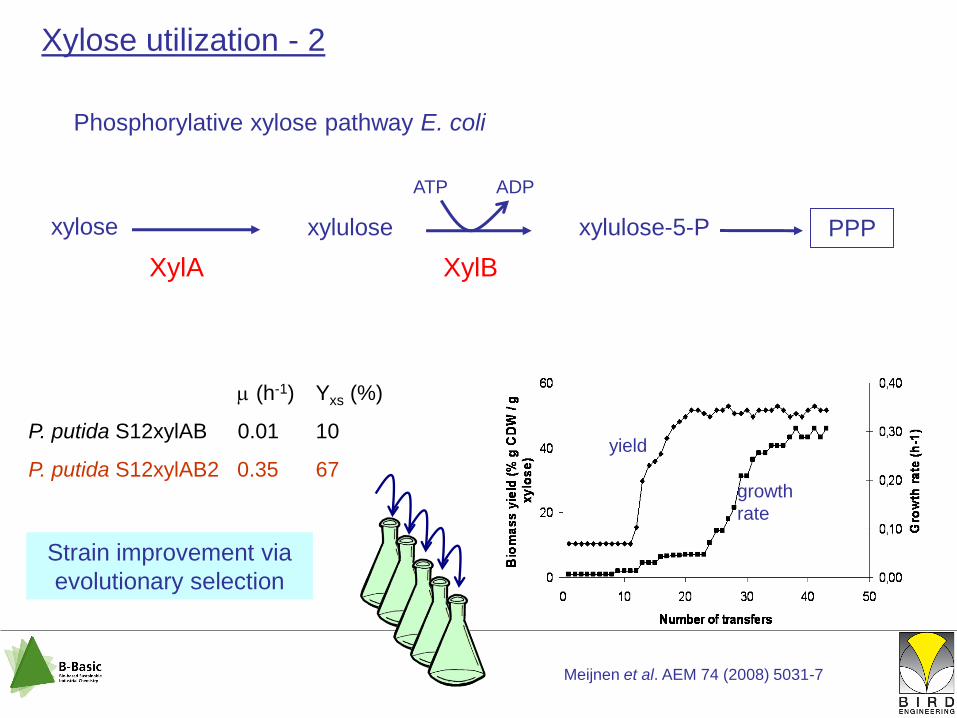
Phosphorylative xylose pathway E. coli
xylose xylulose xylulose-5-P PPP
ATP ADP
XylA XylB
Meijnen et al. AEM 74 (2008) 5031-7
(h-1) Yxs (%)
P. putida S12xylAB 0.01 10
P. putida S12xylAB2 0.35 67yield
growth
rate
Strain improvement via
evolutionary selection
Xylose utilization - 2
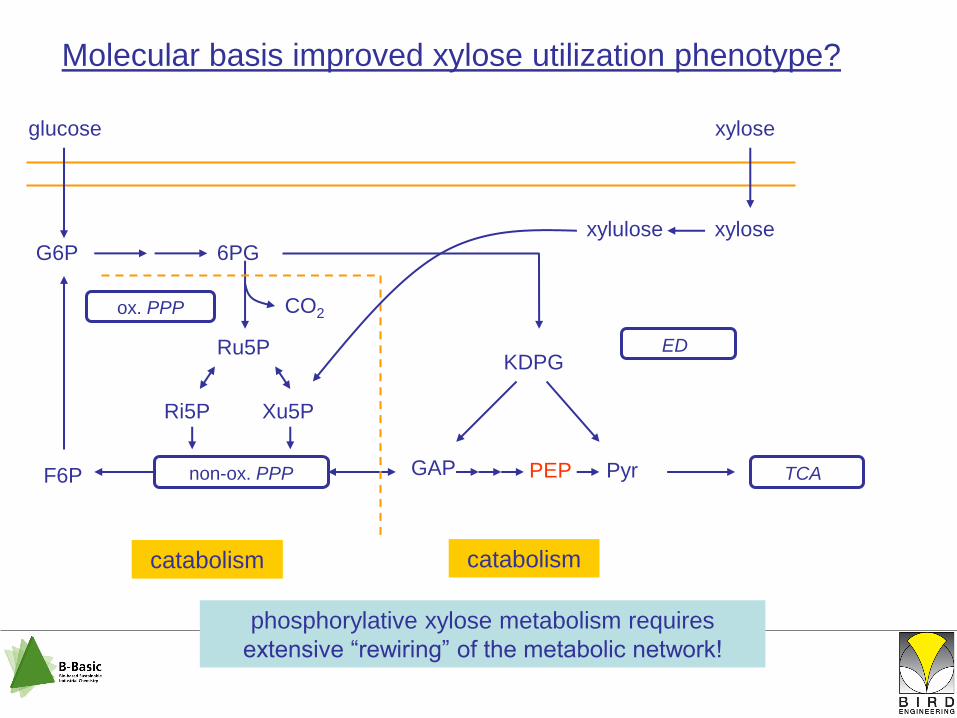
glucose
non-ox. PPP GAP
ED
PEP Pyr TCA
Molecular basis improved xylose utilization phenotype?
G6P 6PG
KDPGRu5P
Ri5P Xu5P
F6P
ox. PPP CO2
catabolismanabolism
xylose
xylulose xylose
catabolism
phosphorylative xylose metabolism requires
extensive “rewiring” of the metabolic network!
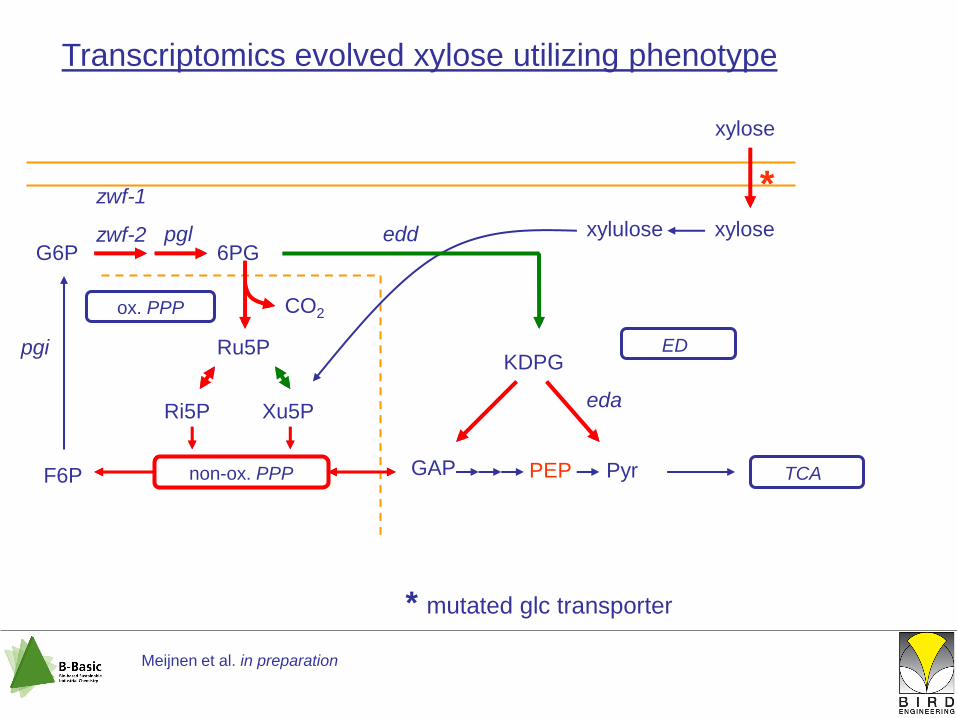
non-ox. PPP GAP
ED
PEP Pyr TCA
Transcriptomics evolved xylose utilizing phenotype
G6P 6PG
KDPGRu5P
Ri5P Xu5P
F6P
ox. PPP CO2
xylose
xylulose xylose
*
Meijnen et al. in preparation
* mutated glc transporter
edd
eda
pgi
pgl
zwf-1
zwf-2
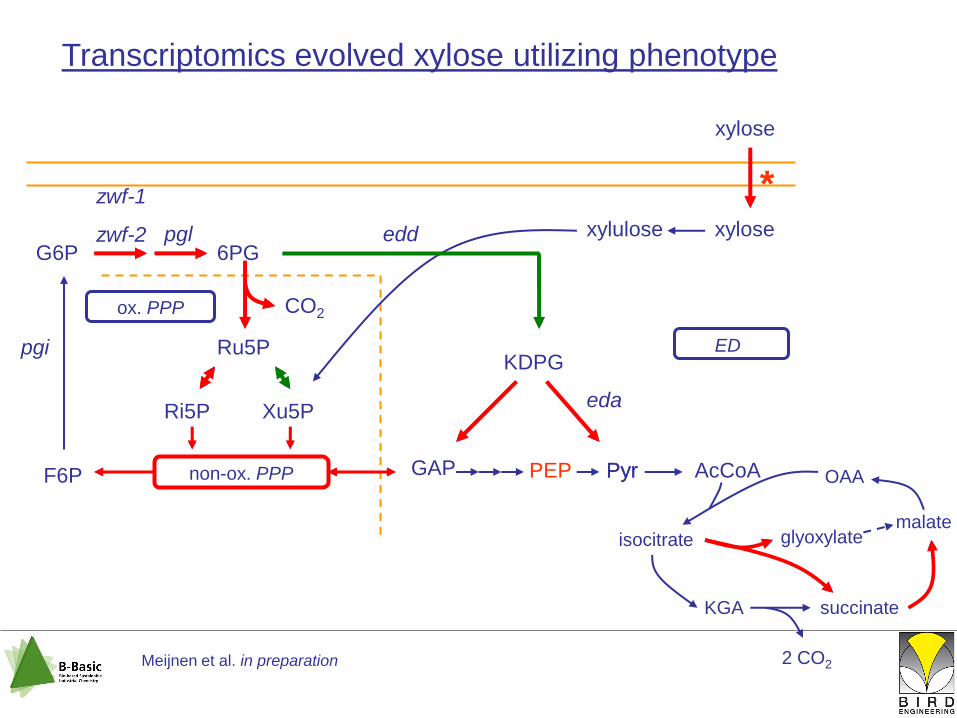
non-ox. PPP GAP
ED
PEP Pyr
Transcriptomics evolved xylose utilizing phenotype
G6P 6PG
KDPGRu5P
Ri5P Xu5P
F6P
ox. PPP CO2
xylose
xylulose xylose
*
Meijnen et al. in preparation
edd
eda
pgi
pgl
zwf-1
zwf-2
Pyr AcCoA
isocitrate
KGA succinate
malate
OAA
glyoxylate
2 CO2
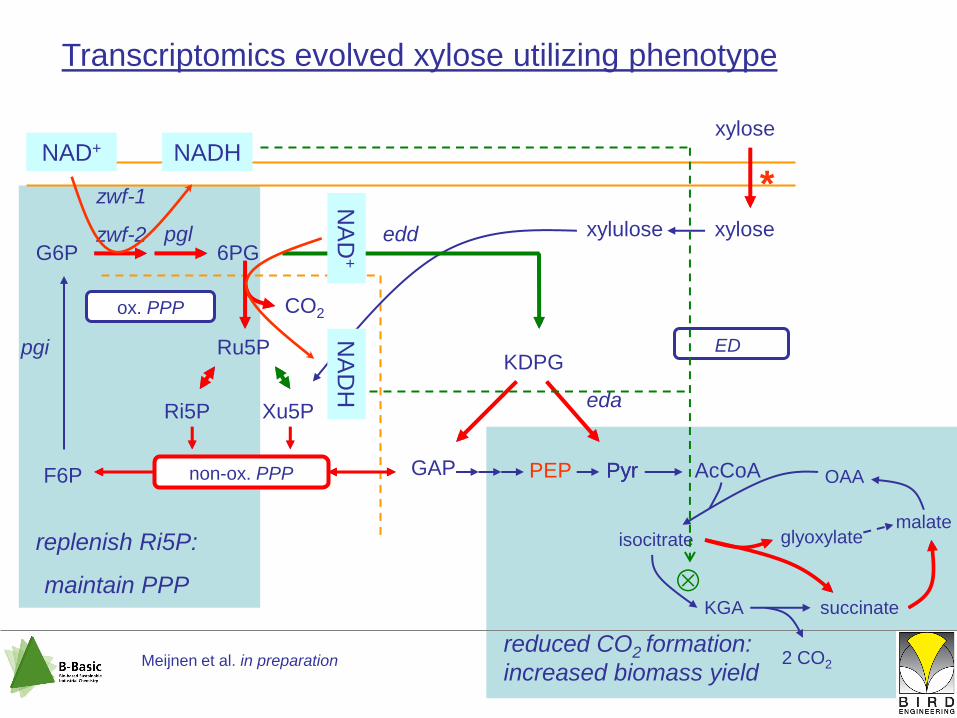
reduced CO2 formation:
increased biomass yield
replenish Ri5P:
maintain PPP
non-ox. PPP GAP
ED
PEP Pyr
Transcriptomics evolved xylose utilizing phenotype
G6P 6PG
KDPGRu5P
Ri5P Xu5P
F6P
ox. PPP CO2
xylose
xylulose xylose
*
Meijnen et al. in preparation
edd
eda
pgi
pgl
zwf-1
zwf-2
Pyr AcCoA
isocitrate
KGA succinate
malate
OAA
glyoxylate
2 CO2
NADHNAD+
NA
DH
NA
D+
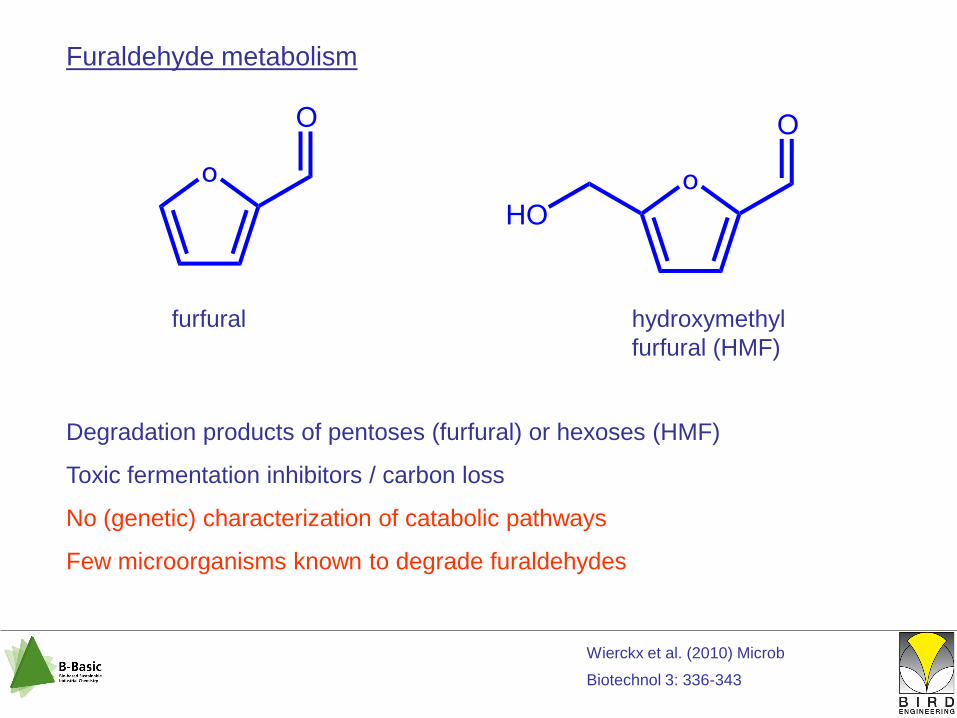
Furaldehyde metabolism
furfural hydroxymethyl
furfural (HMF)
Degradation products of pentoses (furfural) or hexoses (HMF)
Toxic fermentation inhibitors / carbon loss
No (genetic) characterization of catabolic pathways
Few microorganisms known to degrade furaldehydes
Wierckx et al. (2010) Microb
Biotechnol 3: 336-343

Furaldehyde metabolism
Novel furfural / HMF degrading bacterium isolated: Cupriavidus basilensis HMF14
• Gram- bacterium
• Mesophilic (<38 ºC), neutrophilic aerobe
• Growth on HMF, furfural, aromatics, NO sugars
• PHA production
Identify HMF-furfural degradation genes by transposon mutagenesis
Wierckx et al. (2010) Microb
Biotechnol 3: 336-343
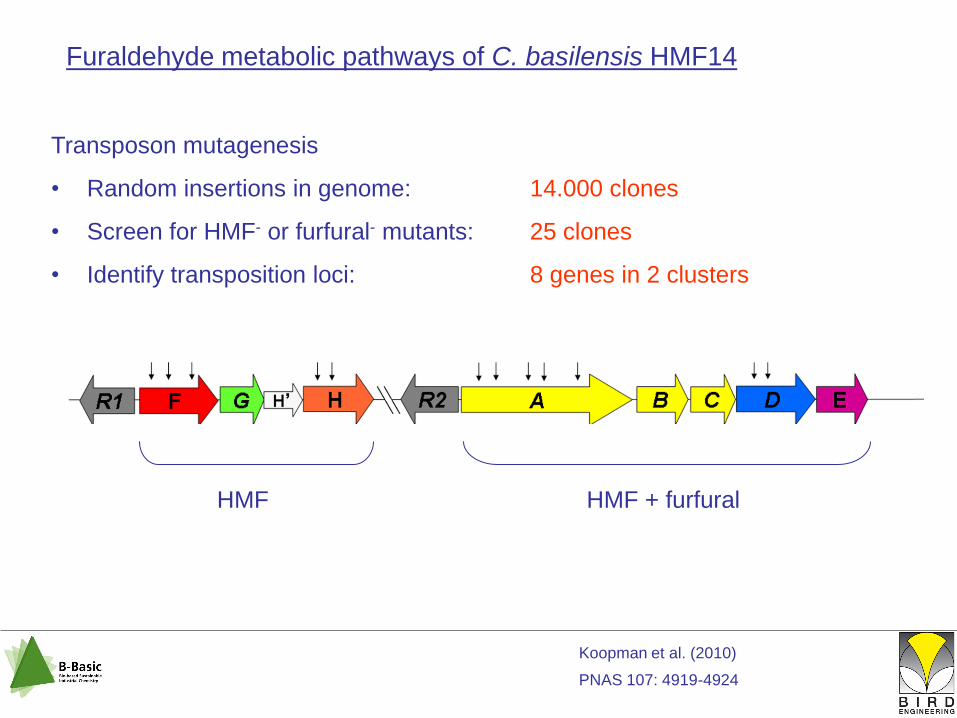
Furaldehyde metabolic pathways of C. basilensis HMF14
Transposon mutagenesis
• Random insertions in genome: 14.000 clones
• Screen for HMF- or furfural- mutants: 25 clones
• Identify transposition loci: 8 genes in 2 clusters
HMF HMF + furfural
Koopman et al. (2010)
PNAS 107: 4919-4924
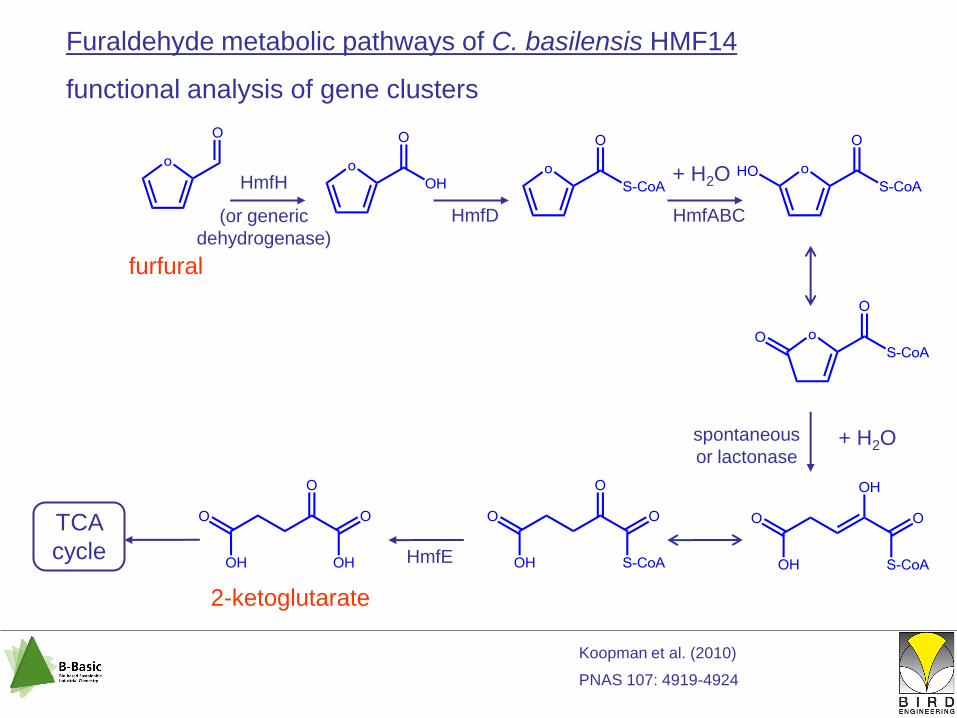
Furaldehyde metabolic pathways of C. basilensis HMF14
functional analysis of gene clusters
HmfH
(or generic
dehydrogenase)
HmfD HmfABC
HmfE
+ H2Ospontaneous
or lactonase
TCA
cycle
furfural
2-ketoglutarate
+ H2O
Koopman et al. (2010)
PNAS 107: 4919-4924
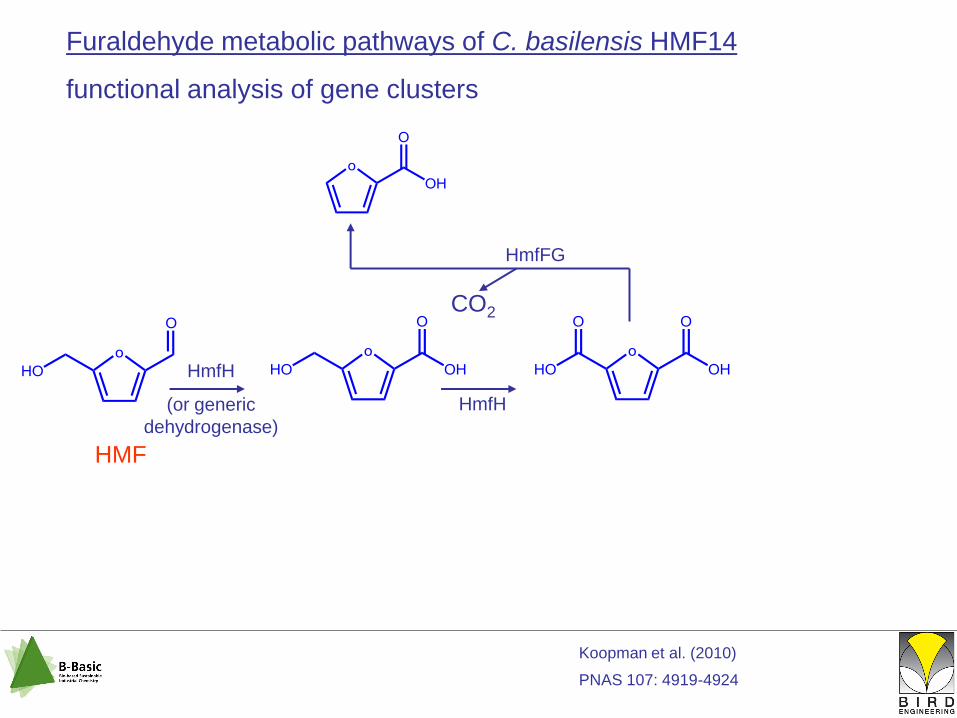
Furaldehyde metabolic pathways of C. basilensis HMF14
functional analysis of gene clusters
HmfH
(or generic
dehydrogenase)
HmfH
HmfFG
CO2
HMF
Koopman et al. (2010)
PNAS 107: 4919-4924
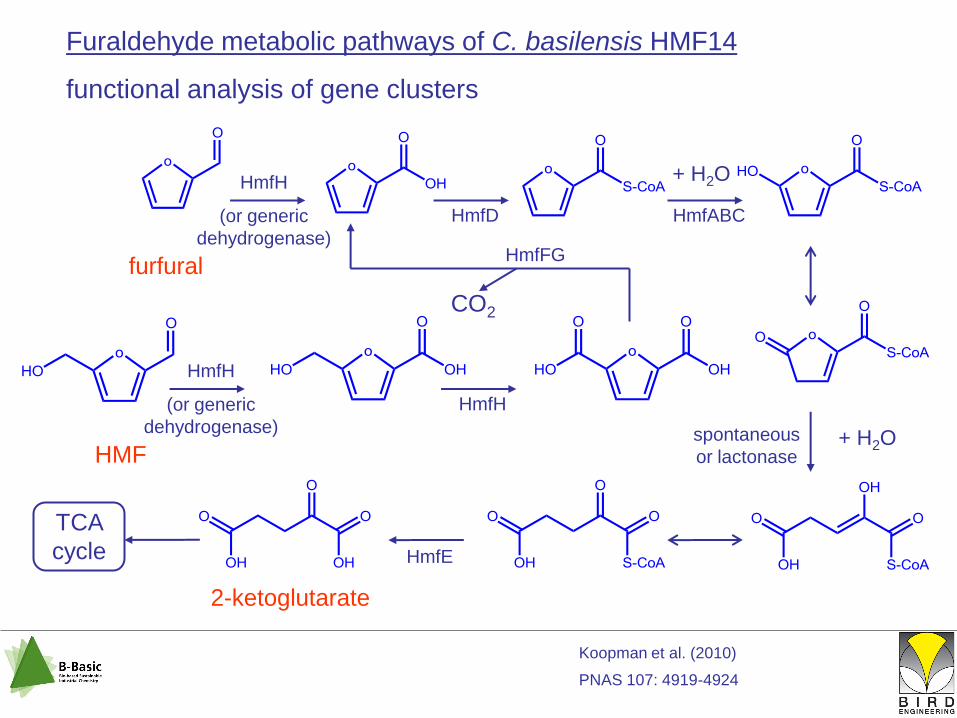
Furaldehyde metabolic pathways of C. basilensis HMF14
functional analysis of gene clusters
HmfH
(or generic
dehydrogenase)
HmfD HmfABC
HmfE
+ H2Ospontaneous
or lactonase
TCA
cycle
HmfH
(or generic
dehydrogenase)
HmfH
HmfFG
CO2
2-ketoglutarate
furfural
HMF
+ H2O
Koopman et al. (2010)
PNAS 107: 4919-4924

Furaldehyde metabolism in P. putida S12
μ (h-1) Yxs (%)
hmfABCDE 0.30 51 growth on furfural
hmfABCDE + hmfFGH 0.23 40 growth on HMF (and furfural)
WT P. putida S12 engineered P. putida S12
glucose 35.4 % + +
xylose 20.2 % - +
arabinose 2.4 % - +
uronic acid 2.5 % + +
org. acids (Ac-, formate) 4.8 % + +
furaldehydes (HMF, furfural) 0.7 % - +
aromatics (lignin) 19.2 % +/-+/-
Koopman et al. (2010)
PNAS 107: 4919-4924

Summary and conclusions - 1
Synthetic pathways constructed in P. putida S12 for:
•production of various aromatic products
•combined heterologous / endogenous activities / gene deletions
•utilization of xylose (+ arabinose)
•oxidative / phosphorylative
•utilization (detoxification) of furaldehydes
•novel pathway / genes isolated and characterized from
environmental isolate

Synthetic pathways may be:
•complete heterologous pathways
•hybrid heterologous / endogenous pathways
•“short-circuited” endogenous pathways
Synthetic pathways commonly need optimization
•improve metabolic flux towards (unnatural) product
•“rewiring” primary metabolic network
System-wide disturbance: optimization requires system-wide approach
•targeted / rational: extensive systems biology input (still underdeveloped)
•semi-targeted / random; classical strain improvement / evolutionary selection
combined with system-wide analysis: pragmatic
Summary and conclusions - 2

Acknowledgements
Nick Wierckx
Jean-Paul Meijnen
Frank Koopman
Suzanne Verhoef
TU Dortmund
Lars Blank, Andreas Schmid
TUDelft
Han de Winde