Embed Size (px)
Citation preview
Synthetic metabolic engineering-a novel,simple technology for designing a chimericmetabolic pathwayYe et al.
Ye et al. Microbial Cell Factories 2012, 11:120http://www.microbialcellfactories.com/content/11/1/120

Ye et al. Microbial Cell Factories 2012, 11:120http://www.microbialcellfactories.com/content/11/1/120
RESEARCH Open Access
Synthetic metabolic engineering-a novel,simple technology for designing a chimericmetabolic pathwayXiaoting Ye1, Kohsuke Honda1,2*, Takaaki Sakai1, Kenji Okano1, Takeshi Omasa1,3, Ryuichi Hirota4, Akio Kuroda4
and Hisao Ohtake1
Abstract
Background: The integration of biotechnology into chemical manufacturing has been recognized as a keytechnology to build a sustainable society. However, the practical applications of biocatalytic chemical conversionsare often restricted due to their complexities involving the unpredictability of product yield and the troublesomecontrols in fermentation processes. One of the possible strategies to overcome these limitations is to eliminate theuse of living microorganisms and to use only enzymes involved in the metabolic pathway. Use of recombinantmesophiles producing thermophilic enzymes at high temperature results in denaturation of indigenous proteinsand elimination of undesired side reactions; consequently, highly selective and stable biocatalytic modules can bereadily prepared. By rationally combining those modules together, artificial synthetic pathways specialized forchemical manufacturing could be designed and constructed.
Results: A chimeric Embden-Meyerhof (EM) pathway with balanced consumption and regeneration of ATP andADP was constructed by using nine recombinant E. coli strains overproducing either one of the seven glycolyticenzymes of Thermus thermophilus, the cofactor-independent phosphoglycerate mutase of Pyrococcus horikoshii, orthe non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase of Thermococcus kodakarensis. By couplingthis pathway with the Thermus malate/lactate dehydrogenase, a stoichiometric amount of lactate was producedfrom glucose with an overall ATP turnover number of 31.
Conclusions: In this study, a novel and simple technology for flexible design of a bespoke metabolic pathwaywas developed. The concept has been testified via a non-ATP-forming chimeric EM pathway. We designated thistechnology as “synthetic metabolic engineering”. Our technology is, in principle, applicable to all thermophilicenzymes as long as they can be functionally expressed in the host, and thus would be potentially applicable tothe biocatalytic manufacture of any chemicals or materials on demand.
BackgroundThe use of renewable feedstocks as a starting materialfor the production of a wide range of value-addedchemicals, the so-called “biorefinery”, has been one ofthe most outstanding issues in building of a sustain-able society [1,2]. Considerable research effort has beenexerted to improve the economy of current microbial-
* Correspondence: [email protected] of Biotechnology, Graduate School of Engineering, OsakaUniversity, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan2PRESTO, Japan Science and Technology Agency (JST), 4-1-8 Honcho,Kawaguchi, Saitama 332-0012, JapanFull list of author information is available at the end of the article
© 2012 Ye et al.; licensee BioMed Central Ltd.Commons Attribution License (http://creativecreproduction in any medium, provided the or
fermentation-based biorefinery processes. The optimiza-tion of the metabolic flux of microbial cells by enhan-cing the expression levels of desired genes and/or bydepleting those of undesired ones has emerged as apowerful strategy to improve microbial cells, the conceptso called “metabolic engineering” [3]. However, theseattempts often suffer from flux imbalances as artificiallyengineered cells typically lack the regulatory mechan-isms characteristic of natural metabolism [4]. One ofthe possible strategies to overcome this limitation is toavoid the use of living microorganisms and to use onlyenzymes involved in the metabolic pathway [5,6]. Thisin vitro production system would offer a number of
This is an Open Access article distributed under the terms of the Creativeommons.org/licenses/by/2.0), which permits unrestricted use, distribution, andiginal work is properly cited.

Thermophiles & Hyperthermophiles
Step 2: Expression in mesophiles (e.g. E. coli)
Step 3: Heat-treatment (typically at 70oC for 30 min)
Step 4: Construction of an in vitro metabolicpathway with the biocatalytic modules
Step 1: Selection of thermo-tolerant enzymesand construction of expression vectors
Figure 1 Schematic illustration of the basic procedure forsynthetic metabolic engineering.
Ye et al. Microbial Cell Factories 2012, 11:120 Page 2 of 11http://www.microbialcellfactories.com/content/11/1/120
potential advantages over the conventional fermentation-based production process, such as better process flexibil-ity, elimination of tight transcriptional regulation, andeasy optimization of production processes by alteringenzyme levels. The absence of a culture medium canmarkedly simplify the isolation and purification of theproduct of interest. Moreover, the elimination of micro-bial growth and byproduct formation would allow usto obtain stoichiometrical conversions as well as anability to perform thermodynamic predictions of produc-tion yield.Welch and Scopes reconstructed a glycolytic pathway
using individually purified yeast enzymes. The recon-structed pathway is capable of converting 1 M (18% [w/v])glucose to ethanol within 8 h with nearly 100% molaryield [7]. In their work, they demonstrated that theimbalance of ATP, which impedes the complete conver-sion of glucose to ethanol, can be prevented by addingan excess amount of arsenate to the reaction mixture.Glyceraldehyde-3-phosphate (GAP) dehydrogenase (GAPDH)can accept arsenate instead of phosphate to form 1-arseno-3-phosphoglycerate, which is simultaneously brokendown to arsenate and 3-phosphoglycerate (3-PG). A sim-ilar experiment using a cell-free extract of Zymomonasmobilis resulted in the conversion of 2 M glucose to3.6 M ethanol [8]. The final ethanol concentration (nearly20% [v/v]) was higher than any natural fermentation sys-tem can achieve. Nevertheless, little attention has beenpaid to the practical application of in vitro productionsystems mainly owing to the economical unprofitabilityof processes involving enzyme purification.Thermophilic enzymes have recently been increasingly
used in bioindustrial processes [9,10]. RecombinantDNA techniques allow the heterologous overproductionof thermophilic enzymes in mesophilic microorganisms(e.g., Escherichia coli). The use of recombinant meso-philes having thermophilic enzymes at high tempera-tures results in the denaturation of indigenous enzymesand the minimization of unwanted side reactions. Con-sequently, highly selective thermophilic biocatalyticmodules comparative to the purified enzymes can bereadily obtained without costly and cumbersome proce-dures for enzyme purification [11]. The rational combin-ation of these biocatalytic modules makes it possible toconstruct in vitro synthetic metabolic pathways specia-lized for chemical manufacturing. More importantly, theexcellent stability of thermophilic enzymes can mitigatethe major disadvantage of in vitro enzymatic conver-sions, the inability of protein synthesis and renewal. Wedesignated this novel, simple technology as “syntheticmetabolic engineering”. To construct an artificial syn-thetic pathway by synthetic metabolic engineering, fourkey steps are included: 1) appropriate selection ofthermostable enzymes; 2) expression in mesophilic hosts
(e.g., E.coli); 3) preheating of the cell suspension at hightemperature (typically at 70°C for 30 min) to disrupt thecell membrane and to inactivate the indigenous hostenzymes; and 4) rational combination of those catalyticmodules at adequate ratio to achieve the stoichiometri-cal conversion (Figure 1).As can be seen from Welch and Scope’s work, the pre-
vention of cofactor depletion is a critical issue in con-structing a synthetic metabolic pathway [7]. ATP andNAD(P)H are the most important biological phosphateand electron donor, respectively, as they are required fornumerous enzymatic reactions in both anabolic andcatabolic metabolisms. One of the possible strategies toprevent cofactor depletion is the integration of suitablecofactor-regeneration enzymes into a synthetic pathway.For instance, ATP regeneration systems using thermo-philic polyphosphate kinase and polyphosphate have

Ye et al. Microbial Cell Factories 2012, 11:120 Page 3 of 11http://www.microbialcellfactories.com/content/11/1/120
been developed and applied to the production of D-ala-nyl-D-alanine [12] and fructose 1,6-bisphosphate [13].Thermophilic NAD(P)+-dependent 6-phosphogluconatedehydrogenase [14], glycerol dehydrogenase [15], andlactate dehydrogenase [16], are available for NAD(P)Hregeneration. However, these cofactor-regeneration systemsrequire an exogenous substrate serving as a phosphateand electron donor.Rapidly expanding genomics and metabolomics infor-
mation has revealed a great diversity of microbial meta-bolisms. In particular, although the central metabolicroutes of bacteria and eukaryotes are generally well-conserved, variant pathways consisting of several uniqueenzymes with unusual cofactor specificities have beendeveloped in archaea [17]. For instance, the modifiedEmbden-Meyerhof (EM) pathway of Pyrococcus furiosushas only four orthologues of the 10 glycolysis enzymesin bacteria and eukaryotes: triosephosphate isomerase(TIM), phosphoglycerate mutase (PGM), enolase (ENO),and pyruvate kinase (PK) [18]. Glucokinase (GK) andphosphofructokinase (PFK) of P. furiosus are ADP-dependent enzymes that are not related to the classicalATP-dependent kinases involved in the bacterial/eukaryotic EM pathway [19,20]. Although the conver-sion of GAP into 3-PG is catalyzed by the enzymecouple of the NAD+-dependent GAPDH and the ATP-generating phosphoglycerate kinase (PGK) in the clas-sical EM pathway, the GAP ferredoxin oxidoreductase(GAPOR) is responsible for the single-step phosphate-independent conversion of GAP into 3-PG in P. furiosus[21]. A variant enzyme, non-phosphorylating GAPDH(GAPN), that utilizes NAD+ and/or NADP+ as the cofac-tor for the phosphate-independent oxidation of GAP hasalso been found and characterized in several archaealstrains involving Thermoproteus tenax [22], Sulfolobussolfataricus [23], and Thermococcus kodakarensis [24].The substitution of GAPDH and PGK of the classicalEM pathway with archaeal GAPN theoretically enablesthe construction of a chimeric EM pathway, in whichthe consumption and regeneration rates of ATP andADP are balanced (Figure 2). Moreover, GAPN can by-pass the production of the extremely thermolabile inter-mediate 1,3-bisphosphoglycerate [23]. Meanwhile, theglucose oxidation via the chimeric EM pathway yields thereducing equivalents in the form of NAD(P)H. To demon-strate the feasibility of the chimeric EM pathway, aNADH-dependent malate/lactate dehydrogenase wasemployed to achieve the redox balance of the cofactor anddirect conversion from glucose to lactate.Lactate has been attracting a great attention for its
application in food, cosmetic, pharmaceutical and chem-ical applications [25]. However, in the conventional lac-tate fermentation process, fermentation broth containsimpurities such as color, residual sugars, nutrients, and
other organic acids, apart from cell mass [26]. Manystudies concerning lactate separation using differenttechniques such as direct distillation, solvent extraction,adsorption and electrodialysis, have been conducted inorder to reduce the operating cost [27]. Synthetic meta-bolic engineering without the use of culture mediumwould markedly simplify the isolation and purificationof lactate.In this work, the chimeric EM pathway was constructed
by synthetic metabolic engineering using a mixture of ninedifferent recombinant E. coli strains, each one of themoverproducing one of seven glycolytic enzymes of Ther-mus thermophilus, the cofactor-independent PGM (iPGM)of Pyrococcus horikoshii, or the GAPN of Thermococcuskodakarensis. By coupling this chimeric EM pathway withthe T. thermophilus malate/lactate dehydrogenase, a stoi-chiometric amount of lactate could be produced from glu-cose with an overall ATP turnover number of 31.
ResultsSelection of enzymes for chimeric EM pathwayGAPN, a key enzyme for constructing a chimeric EMpathway, was derived from the hyperthermophilicarchaeon T. kodakarensis. Although both NAD+ andNADP+ can serve as the electron acceptor, the Km ofThermococcus GAPN for NADP+ is two orders of mag-nitude lower than that for NAD+ [24]. However, thethermostability of NADP+ is considerably lower thanthat of NAD+, particularly under neutral and acidic con-ditions [28]. Owing to this fact, NAD+ was employed asthe redox cofactor for the construction of a chimericpathway. The enzyme can be strongly activated by theaddition of glucose-1-phosphate (G1P) [24]. Under theassay conditions employed in this study, GAPN exhib-ited the highest specific activity in the presence of G1Pat 100 μM or higher.The thermophilic bacterium T. thermophilus HB8 was
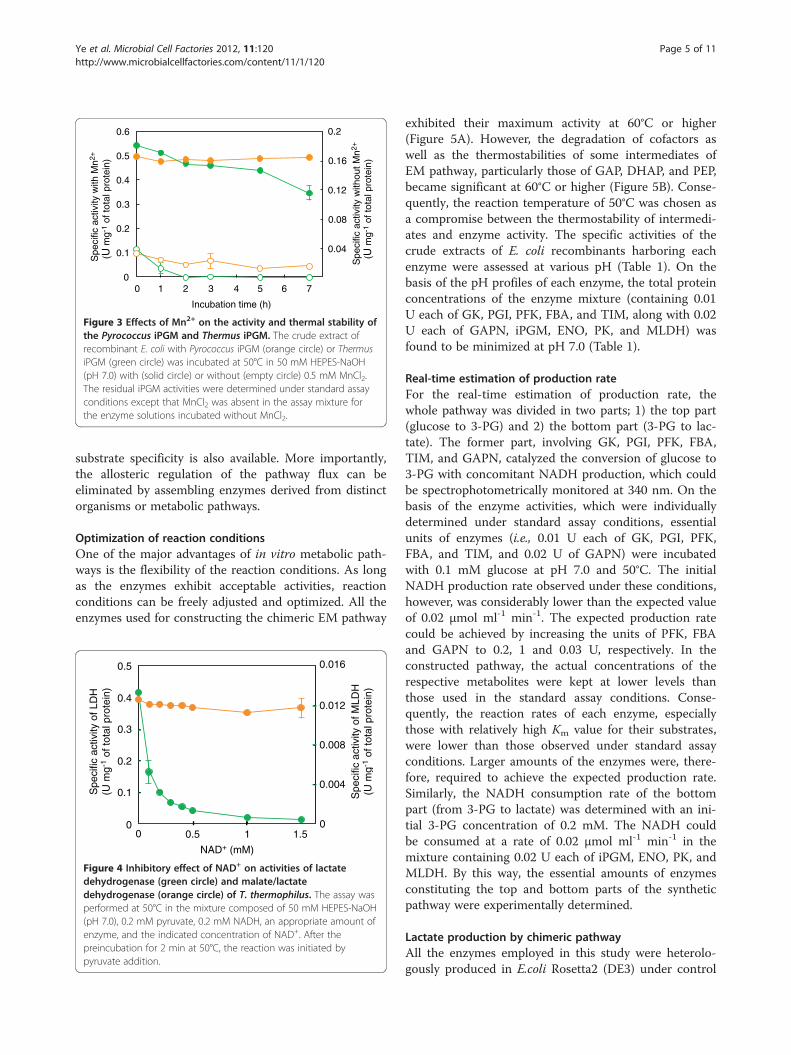
used as the source of other genes required for the con-struction of a chimeric pathway. Among them, thecofactor-independent phosphoglycerate mutase (iPGM)showed a relatively low thermal stability (Figure 3).Although the thermal inactivation of other enzymes wasinsignificant, the activity of the Thermus iPGM was al-most completely lost after the incubation of the enzymeat 50°C for 2 h. We then isolated and heterologouslyexpressed the gene encoding iPGM from a hyperther-mophilic archaeon, Pyrococcus horikoshii, exhibiting ahigher optimum growth temperature (98°C) [29] thanthat of Thermus thermophilus (70°C) [30]. Although thespecific activity and thermal stability of the PyrococcusiPGM were not markedly different from those of Ther-mus enzyme, they were considerably improved by theaddition of Mn2+ (Figure 3). This finding was in goodagreement with a previous report that the thermal

Bacterial/EukaryoticEM pathway
Glucose
G6P
F6P
FBP
GAP
1,3-BPG x 2
3-PG x 2
2-PG x 2
PEP x 2
Pyruvate x 2
3-PG x 2
2-PG x 2
PEP x 2
Pyruvate x 2
3-PG x 2
2-PG x 2
PEP x 2
Pyruvate x 2
DHAP
ATPADP
ATPADP
NAD(P)+ x 2NAD(P)H x 2
ADP x 2ATP x 2
ADP x 2ATP x 2
ADP x 2ATP x 2
GK
PGI
PFK
FBA
GAPDH
ENO
TIM
PGK
iPGM
PK
GAPOR GAPN
FBP
GAP DHAPTIM
Archaeal modifiedEM pathway
Glucose
G6P
F6P
ADPAMP
ADPAMP
AMP x 2ATP x 2
GK
PGI
PFK
FBA
ENO
iPGM
PK
NAD(P)+ x 2NAD(P)H x 2
NAD(P)+ x 2NAD(P)H x 2
NAD(P)+ x 2NAD(P)H x 2
Fdox x 2
Fdred x 2
ATPADP
GK
PGI
ATPADPPFK
iPGM
PK
FBA
GAPN
FBP
GAP DHAPTIM
ChimericEM pathway
Glucose
G6P
F6P
ENO
Lactate x 2
MLDH
Figure 2 Schematic illustration of design of chimeric Embden-Meyerhof (EM) pathway. The bacterial/eukaryotic classic EM pathway (left),modified EM pathway of Thermococcales archaea (center), and chimeric EM pathway constructed in this study (right) are illustrated. Reactionscatalyzed by enzymes uniquely involved in the archaeal modified EM pathway are indicated by green arrows. Abbreviations used: G6P, glucose-6-phosphate; F6P, fructose-6-phosphate; FBP, fructose-1,6-bisphosphate; GAP, glyceraldehyde-3-phosphate; DHAP, dihydroxyacetone phosphate;1,3-BPG, 1,3-bisphosphoglycerate; 3-PG, 3-phosphoglycerate; 2-PG, 2-phosphoglycerate; PEP, phosphoenolpyruvate; GK, glucose kinase; PGI,glucose-6-phosphate isomerase; PFK, 6-phosphofructokinase; FBA, fructose-bisphosphate aldolase; TIM, triosephosphate isomerase; PGM,phosphoglycerate mutase; ENO, enolase; PK, pyruvate kinase; MLDH, malate/lactate dehydrogenase, GAPOR, glyceraldehyde-3-phosphateferredoxin oxidoreductase; GAPN, non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; PGK, phosphoglycerate kinase; Fdox, oxidized ferredoxin; and Fdred, reduced ferredoxin.
Ye et al. Microbial Cell Factories 2012, 11:120 Page 4 of 11http://www.microbialcellfactories.com/content/11/1/120
stability of iPGM from the thermophilic archaeonArchaeoglobus fulgidus, which shares a 47.5% amino acidsequence identity with the Pyrococcus enzyme, can alsobe improved by an addition of Mn2+ [31]. On the basisof these results, we employed Pyrococcus iPGM and usedit in the presence of Mn2+.Some bacterial lactate dehydrogenases are inhibited by
ATP and/or NAD+, and thus play an important rolein the allosteric regulation of the flux of glycolysis [32].Although the inhibitory effect of ATP (up to 1 mM)was almost negligible, the addition of 0.1, 0.2, and 1 mMNAD+ caused 60, 76, and 95% decreases in the acti-vity of Thermus lactate dehydrogenase compared withthat under the standard assay conditions, respectively(Figure 4). Owing to the low affinity of GAPN to NAD+,
the chimeric pathway, therefore, required a certainamount of exogenous NAD+. This limitation led us tosearch for another enzyme that catalyzes the reductionof pyruvate to lactate. A putative NAD(P)H-dependentdehydrogenase, which was annotated as malate/lactatedehydrogenase (MLDH), was found in the gene-expression library of T. thermophilus HB8 [33]. The en-zyme was confirmed to show pyruvate reducing activitywith no inhibitory effect by NAD+ or other metabolicintermediates in this pathway (Figure 4). The naturalsubstrate of this enzyme was unknown and it mightmore preferably accept a metabolite other than pyruvatein physiological conditions. However, the in vitro syn-thetic pathway involves only a limited number of neces-sary metabolites, and thus an enzyme with a broad
00
0.1
0.2
0.3
0.4
0.5
0.6
6 754321
0.04
0.08
0.12
0.16
0.2
Spe
cific
act
ivity
with
Mn2+
(U m
g-1
of to
tal p
rote
in)
( U m
g-1
of to
tal p
rote
in)
Spe
cific
act
ivity
with
out M
n2+
Incubation time (h)
Figure 3 Effects of Mn2+ on the activity and thermal stability ofthe Pyrococcus iPGM and Thermus iPGM. The crude extract ofrecombinant E. coli with Pyrococcus iPGM (orange circle) or ThermusiPGM (green circle) was incubated at 50°C in 50 mM HEPES-NaOH(pH 7.0) with (solid circle) or without (empty circle) 0.5 mM MnCl2.The residual iPGM activities were determined under standard assayconditions except that MnCl2 was absent in the assay mixture forthe enzyme solutions incubated without MnCl2.
Ye et al. Microbial Cell Factories 2012, 11:120 Page 5 of 11http://www.microbialcellfactories.com/content/11/1/120
substrate specificity is also available. More importantly,the allosteric regulation of the pathway flux can beeliminated by assembling enzymes derived from distinctorganisms or metabolic pathways.
Optimization of reaction conditionsOne of the major advantages of in vitro metabolic path-ways is the flexibility of the reaction conditions. As longas the enzymes exhibit acceptable activities, reactionconditions can be freely adjusted and optimized. All theenzymes used for constructing the chimeric EM pathway
0.1
00 0.5 1 1.5
0
0.004
0.008
0.012
0.016
0.2
0.3
0.4
0.5
Spe
cific
act
ivity
of L
DH
(U m
g-1 o
f tot
al p
rote
in)
Spe
cific
act
ivity
of M
LDH
(U m
g-1 o
f tot
al p
rote
in)
NAD+ (mM)
Figure 4 Inhibitory effect of NAD+ on activities of lactatedehydrogenase (green circle) and malate/lactatedehydrogenase (orange circle) of T. thermophilus. The assay wasperformed at 50°C in the mixture composed of 50 mM HEPES-NaOH(pH 7.0), 0.2 mM pyruvate, 0.2 mM NADH, an appropriate amount ofenzyme, and the indicated concentration of NAD+. After thepreincubation for 2 min at 50°C, the reaction was initiated bypyruvate addition.
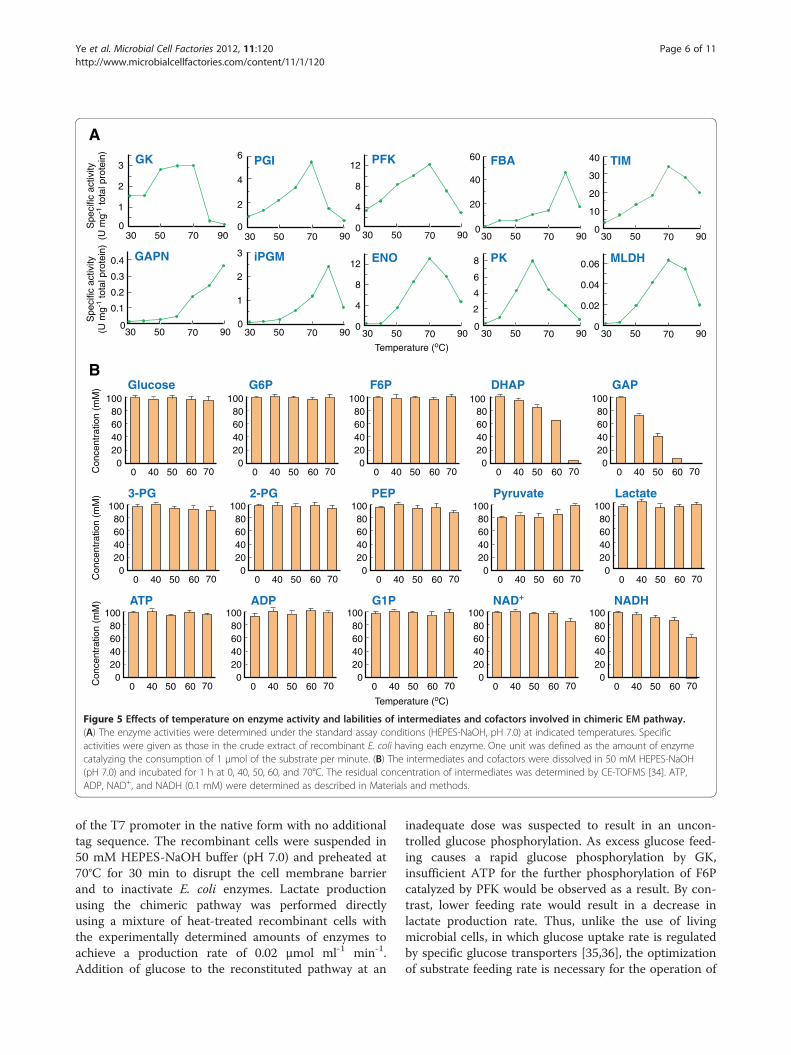
exhibited their maximum activity at 60°C or higher(Figure 5A). However, the degradation of cofactors aswell as the thermostabilities of some intermediates ofEM pathway, particularly those of GAP, DHAP, and PEP,became significant at 60°C or higher (Figure 5B). Conse-quently, the reaction temperature of 50°C was chosen asa compromise between the thermostability of intermedi-ates and enzyme activity. The specific activities of thecrude extracts of E. coli recombinants harboring eachenzyme were assessed at various pH (Table 1). On thebasis of the pH profiles of each enzyme, the total proteinconcentrations of the enzyme mixture (containing 0.01U each of GK, PGI, PFK, FBA, and TIM, along with 0.02U each of GAPN, iPGM, ENO, PK, and MLDH) wasfound to be minimized at pH 7.0 (Table 1).
Real-time estimation of production rateFor the real-time estimation of production rate, thewhole pathway was divided in two parts; 1) the top part(glucose to 3-PG) and 2) the bottom part (3-PG to lac-tate). The former part, involving GK, PGI, PFK, FBA,TIM, and GAPN, catalyzed the conversion of glucose to3-PG with concomitant NADH production, which couldbe spectrophotometrically monitored at 340 nm. On thebasis of the enzyme activities, which were individuallydetermined under standard assay conditions, essentialunits of enzymes (i.e., 0.01 U each of GK, PGI, PFK,FBA, and TIM, and 0.02 U of GAPN) were incubatedwith 0.1 mM glucose at pH 7.0 and 50°C. The initialNADH production rate observed under these conditions,however, was considerably lower than the expected valueof 0.02 μmol ml-1 min-1. The expected production ratecould be achieved by increasing the units of PFK, FBAand GAPN to 0.2, 1 and 0.03 U, respectively. In theconstructed pathway, the actual concentrations of therespective metabolites were kept at lower levels thanthose used in the standard assay conditions. Conse-quently, the reaction rates of each enzyme, especiallythose with relatively high Km value for their substrates,were lower than those observed under standard assayconditions. Larger amounts of the enzymes were, there-fore, required to achieve the expected production rate.Similarly, the NADH consumption rate of the bottompart (from 3-PG to lactate) was determined with an ini-tial 3-PG concentration of 0.2 mM. The NADH couldbe consumed at a rate of 0.02 μmol ml-1 min-1 in themixture containing 0.02 U each of iPGM, ENO, PK, andMLDH. By this way, the essential amounts of enzymesconstituting the top and bottom parts of the syntheticpathway were experimentally determined.
Lactate production by chimeric pathwayAll the enzymes employed in this study were heterolo-gously produced in E.coli Rosetta2 (DE3) under control
50 70 90300
1
GK
Temperature (oC)
Temperature (oC)
Spe
cific
act
ivity
(U m
g-1 to
tal p
rote
in)
Spe
cific
act
ivity
(U m
g-1 to
tal p
rote
in)
Con
cent
ratio
n (m
M)
Con
cent
ratio
n (m
M)
Con
cent
ratio
n (m
M)
2
3
50 70 90300
0.1
GAPN
0.2
0.3
0.4
50 70 90300
2
PGI
4
6
50 70 90300
4
PFK
8
12
50 70 90300
20
FBA
40
60
50 70 90300
20
10
TIM30
40
50 70 90300
1
iPGM
2
3
50 70 90300
4
ENO
8
12
50 70 90300
2
PK
4
6
8
50 70 90300
0.02
MLDH
0.04
0.06
Glucose G6P F6P DHAP GAP
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
3-PG 2-PG PEP Pyruvate Lactate
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
ATP ADP G1P NAD+ NADH
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
00 40 50 60 70
20406080
100
B
A
Figure 5 Effects of temperature on enzyme activity and labilities of intermediates and cofactors involved in chimeric EM pathway.(A) The enzyme activities were determined under the standard assay conditions (HEPES-NaOH, pH 7.0) at indicated temperatures. Specificactivities were given as those in the crude extract of recombinant E. coli having each enzyme. One unit was defined as the amount of enzymecatalyzing the consumption of 1 μmol of the substrate per minute. (B) The intermediates and cofactors were dissolved in 50 mM HEPES-NaOH(pH 7.0) and incubated for 1 h at 0, 40, 50, 60, and 70°C. The residual concentration of intermediates was determined by CE-TOFMS [34]. ATP,ADP, NAD+, and NADH (0.1 mM) were determined as described in Materials and methods.
Ye et al. Microbial Cell Factories 2012, 11:120 Page 6 of 11http://www.microbialcellfactories.com/content/11/1/120
of the T7 promoter in the native form with no additionaltag sequence. The recombinant cells were suspended in50 mM HEPES-NaOH buffer (pH 7.0) and preheated at70°C for 30 min to disrupt the cell membrane barrierand to inactivate E. coli enzymes. Lactate productionusing the chimeric pathway was performed directlyusing a mixture of heat-treated recombinant cells withthe experimentally determined amounts of enzymes toachieve a production rate of 0.02 μmol ml-1 min-1.Addition of glucose to the reconstituted pathway at an
inadequate dose was suspected to result in an uncon-trolled glucose phosphorylation. As excess glucose feed-ing causes a rapid glucose phosphorylation by GK,insufficient ATP for the further phosphorylation of F6Pcatalyzed by PFK would be observed as a result. By con-trast, lower feeding rate would result in a decrease inlactate production rate. Thus, unlike the use of livingmicrobial cells, in which glucose uptake rate is regulatedby specific glucose transporters [35,36], the optimizationof substrate feeding rate is necessary for the operation of
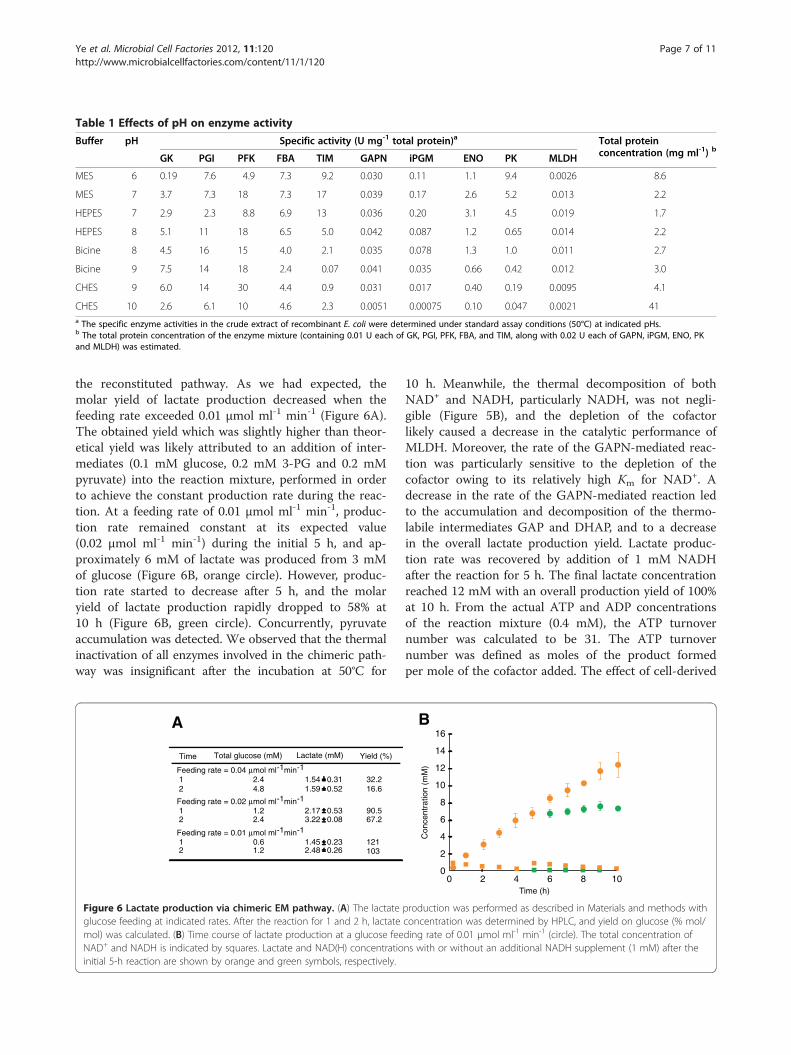
Table 1 Effects of pH on enzyme activity
Buffer pH Specific activity (U mg-1 total protein)a Total proteinconcentration (mg ml-1) bGK PGI PFK FBA TIM GAPN iPGM ENO PK MLDH
MES 6 0.19 7.6 4.9 7.3 9.2 0.030 0.11 1.1 9.4 0.0026 8.6
MES 7 3.7 7.3 18 7.3 17 0.039 0.17 2.6 5.2 0.013 2.2
HEPES 7 2.9 2.3 8.8 6.9 13 0.036 0.20 3.1 4.5 0.019 1.7
HEPES 8 5.1 11 18 6.5 5.0 0.042 0.087 1.2 0.65 0.014 2.2
Bicine 8 4.5 16 15 4.0 2.1 0.035 0.078 1.3 1.0 0.011 2.7
Bicine 9 7.5 14 18 2.4 0.07 0.041 0.035 0.66 0.42 0.012 3.0
CHES 9 6.0 14 30 4.4 0.9 0.031 0.017 0.40 0.19 0.0095 4.1
CHES 10 2.6 6.1 10 4.6 2.3 0.0051 0.00075 0.10 0.047 0.0021 41a The specific enzyme activities in the crude extract of recombinant E. coli were determined under standard assay conditions (50°C) at indicated pHs.b The total protein concentration of the enzyme mixture (containing 0.01 U each of GK, PGI, PFK, FBA, and TIM, along with 0.02 U each of GAPN, iPGM, ENO, PKand MLDH) was estimated.
Ye et al. Microbial Cell Factories 2012, 11:120 Page 7 of 11http://www.microbialcellfactories.com/content/11/1/120
the reconstituted pathway. As we had expected, themolar yield of lactate production decreased when thefeeding rate exceeded 0.01 μmol ml-1 min-1 (Figure 6A).The obtained yield which was slightly higher than theor-etical yield was likely attributed to an addition of inter-mediates (0.1 mM glucose, 0.2 mM 3-PG and 0.2 mMpyruvate) into the reaction mixture, performed in orderto achieve the constant production rate during the reac-tion. At a feeding rate of 0.01 μmol ml-1 min-1, produc-tion rate remained constant at its expected value(0.02 μmol ml-1 min-1) during the initial 5 h, and ap-proximately 6 mM of lactate was produced from 3 mMof glucose (Figure 6B, orange circle). However, produc-tion rate started to decrease after 5 h, and the molaryield of lactate production rapidly dropped to 58% at10 h (Figure 6B, green circle). Concurrently, pyruvateaccumulation was detected. We observed that the thermalinactivation of all enzymes involved in the chimeric path-way was insignificant after the incubation at 50°C for
Time
Feeding rate = 0.04 µmol ml-1 min-1
1 2
Feeding rate = 0.02 µmol ml-1 min-1
1 2
Feeding rate = 0.01 µmol ml-1 min-1
1 2
A
Total glucose (mM) Lactate (mM) Yield (%)
2.4 1.54 0.31 32.24.8 1.59 0.52 16.6
1.2 2.17 0.53 90.52.4 3.22 0.08 67.2
0.6 1.45 0.23 121 1.2 2.48 0.26 103
Figure 6 Lactate production via chimeric EM pathway. (A) The lactateglucose feeding at indicated rates. After the reaction for 1 and 2 h, lactatemol) was calculated. (B) Time course of lactate production at a glucose feeNAD+ and NADH is indicated by squares. Lactate and NAD(H) concentratioinitial 5-h reaction are shown by orange and green symbols, respectively.
10 h. Meanwhile, the thermal decomposition of bothNAD+ and NADH, particularly NADH, was not negli-gible (Figure 5B), and the depletion of the cofactorlikely caused a decrease in the catalytic performance ofMLDH. Moreover, the rate of the GAPN-mediated reac-tion was particularly sensitive to the depletion of thecofactor owing to its relatively high Km for NAD+. Adecrease in the rate of the GAPN-mediated reaction ledto the accumulation and decomposition of the thermo-labile intermediates GAP and DHAP, and to a decreasein the overall lactate production yield. Lactate produc-tion rate was recovered by addition of 1 mM NADHafter the reaction for 5 h. The final lactate concentrationreached 12 mM with an overall production yield of 100%at 10 h. From the actual ATP and ADP concentrationsof the reaction mixture (0.4 mM), the ATP turnovernumber was calculated to be 31. The ATP turnovernumber was defined as moles of the product formedper mole of the cofactor added. The effect of cell-derived
2
00
4
6
8
10
12
14
16
Con
cent
ratio
n (m
M)
Time (h)
B
2 4 6 8 10
production was performed as described in Materials and methods withconcentration was determined by HPLC, and yield on glucose (% mol/ding rate of 0.01 μmol ml-1 min-1 (circle). The total concentration ofns with or without an additional NADH supplement (1 mM) after the

Ye et al. Microbial Cell Factories 2012, 11:120 Page 8 of 11http://www.microbialcellfactories.com/content/11/1/120
endogenous ATP and ADP (0.37 μM) was considered tobe negligible.
DiscussionMetabolic engineering has become a practical alternativeto conventional chemical conversion particularly forbiocommodity production processes; however, this ap-proach is often hampered by as yet unidentified inherentmechanisms of natural metabolism. One of the currentresearch directions in the field of metabolic engineering isto gain a deeper understanding of these underlying regula-tory networks by exploiting the information obtainedfrom a variety of “-omics” analyses [37]. By contrast, ourapproach, designated as “synthetic metabolic engineering”,provides a completely different means to overcome thislimitation by reconstituting only the pathway of interestusing thermo-tolerant biocatalytic modules. As well asbeing independent of transcriptional regulation, the as-sembly of enzymes derived from distinct organisms ormetabolic pathways can eliminate the effect of allostericregulation on the pathway flux.It is vital to keep both energy and redox cofactors
balanced to sustain overall reactions by synthetic meta-bolic systems because, unlike living biological systems,they are not equipped with complete enzyme apparatusto regenerate or resynthesize these cofactors. An injudi-cious “copy & paste” of natural pathways results in thedepletion of specific cofactors since the physiologicalroles of metabolisms include the energy generation (ca-tabolism) and energy-consuming synthesis of biomole-cules (anabolism), in which the cofactors serve as a“currency” for transferring energy and redox power. Inthis work, we constructed a glycolytic pathway with nonet ATP yield by chimerically integrating the archaealGAPN to a classical EM pathway. The synthetic pathwayproduced a stoichiometric amount of lactate from glu-cose with an overall ATP turnover number of 31. Notethat such a non-ATP-forming glycolysis pathway couldno longer play a physiological role (i.e., energy produc-tion particularly under anaerobic conditions) and thuswould not be applicable for a fermentative purposein vivo. In fact, although the GAPDH defect of E. colican be complemented with the Pisum sativum GAPNunder aerobic condition, this recombinant strain fails togrow anaerobically [38].Although endogenous ATP regeneration was demon-
strated in the present study, our results also indicatedthat the thermolability of NADH remained a major obs-tacle for the long-term operation of a synthetic meta-bolic system. A possible alternative to overcome thislimitation is the replacement of NADH with more stableand low-cost artificial biomimetic NADH analogs [39].Protein engineering approaches to improve the affinitiesof enzymes to these NADH biomimics may be also
required [40]. Another important issue that should beresolved is the increase in production rate. Althoughit was predicted to increase proportionally along with atotal enzyme concentration [41], the total amount ofenzymes required to obtain a desired rate is often notpractically achievable. On the other hand, naturallyoccurring cellular apparatus, in which a series ofmetabolic enzymes are packed in close proximity, canreduce the intermediate diffusion distance and thereforeincrease the overall reaction rate. The coexpression ofthermophilic enzymes constituting a synthetic path-way in a single recombinant is an effective strategy tospatially organize the enzymes and achieve a high pro-duction rate [42].
ConclusionsWe proposed a novel and simple technology, designatedas synthetic metabolic engineering and demonstrated itsapplication to the construction of the non-ATP-formingchimeric EM pathway. The synthetic pathway produceda stoichiometric amount of lactate from glucose with anoverall ATP turnover number of 31. The concept ofin vitro synthetic-pathway biotransformation is not newbut its feasibility in practical application has been largelyrestricted mainly owing to the prejudice that in vitrobiotransformation is too costly for producing low-valuebiocommodities. However, the comparative cost analysisbetween in vivo and in vitro fermentation processesdemonstrated that this interpretation is not necessarilytrue and that the development of stable standardizedenzyme modules will provide economical advantagesto the use of in vitro systems [5]. Synthetic metabolicengineering enables a one-step preparation of highlyselective and stable biocatalytic modules via a simpleheat-treatment of the recombinant mesophiles havingthermophilic enzymes. Most importantly, it is, in prin-ciple, applicable to all thermophilic enzymes as long asthey can be functionally expressed in the host, and thuswould be potentially applicable to the biocatalytic manu-facture of any chemicals or materials on demand.
Materials and methodsBacterial strain and plasmidThe expression vectors for genes encoding GK, PGI, PFK,FBA, TIM, ENO, PK, and MLDH of T. thermophilus HB8were obtained from the Riken Thermus thermophilus HB8expression plasmid set [33]. The expression vector forGAPN [24] was a kind gift from Professor H. Atomi ofKyoto University. The gene encoding iPGM was amplifiedby PCR from the chromosomal DNA of Pyrococcus hori-koshii OT3 (Takara Bio, Shiga, Japan) with the followingprimers: 50-TTCATATGGTGCTAAAGAGGAAAGGC-30
(the NdeI restriction site is underlined) and 50-TTGAATTCTCAAGCTCCAAATTTTTCGCTCCT-30 (the EcoRI

Ye et al. Microbial Cell Factories 2012, 11:120 Page 9 of 11http://www.microbialcellfactories.com/content/11/1/120
restriction site is underlined). The amplified DNA wasdigested with NdeI and EcoRI and inserted into the corres-ponding restriction sites of pET21a (Novagen, Madison,WI, USA).E. coli Rossetta2 (DE3) (Novagen) was used as a host
strain for gene expression. The recombinants were aer-obically cultivated in Luria-Bertani (LB) media contain-ing 100 μg/ml ampicillin and 34 μg/ml chloramphenicolat 37°C. Gene expression was induced by an addition of0.2 mM isopropylthiogalactoside (IPTG) to late-log cul-ture for 4 h. Cells were harvested by centrifugation andresuspended in 50 mM HEPES-NaOH buffer (pH 7.0).The cell suspensions were heated at 70°C for 30 minbefore being used for lactate production.
Enzyme assaysE. coli cell suspensions were disrupted with a UD-201ultrasonicator (Kubota, Osaka, Japan), and the crudelysate was heated at 70°C for 30 min. Cell debris anddenatured proteins were removed by centrifugation at15,000 rpm and 4°C for 10 min. The supernatant wasthen used as an enzyme solution. Protein concentrationwas measured with the Bio-Rad assay system (Bio-Rad,Hercules, CA, USA) using bovine serum albumin asthe standard.One unit of an enzyme was defined as the amount
consuming 1 μmol of the substrate per min under theassay conditions. The standard assay mixture for GKwas composed of 50 mM HEPES-NaOH (pH 7.0),0.1 mM glucose, 0.2 mM ATP, 5 mM MgCl2, 0.5 mMMnCl2, 1 mM NAD+, 1 mM G1P, 0.08 U of PGI, 0.2 Uof PFK, 1 U of FBA, 0.1 U of TIM, 0.02 U of GAPN, andan appropriate amount of GK. The mixture without glu-cose was preincubated at 50°C for 2 min, and then thereaction was initiated by an addition of 0.1 mM glucose.The reduction of NAD+ was monitored at 340 nm usinga UV-VIS spectrophotometer (Model UV-2450, Shimadzu,Kyoto, Japan). Similarly, the activities of PGI, PFK, FBA,TIM, and GAPN were spectrophotometrically assessedin the mixture containing the substrate for each enzyme(0.1 mM of glucose-6-phosphate, fructose-6-phosphate,fructose-1,6-bisphosphate, dihydroxyacetone phosphate,or 0.2 mM GAP, respectively) instead of glucose.iPGM activity was assayed at 50°C in a mixture con-
sisting of 50 mM HEPES-NaOH (pH 7.0), 0.2 mM3-phosphoglycerate, 5 mM MgCl2, 0.5 mM MnCl2,0.2 mM ADP, 0.2 mM NADH, 0.5 U of ENO, 0.5 Uof PK, 1.2 U of LDH, and an appropriate amount ofenzyme. ENO and PK were assayed in the same mannerusing 0.2 mM of 2-phosphoglycerate and phosphoenol-pyruvate as the substrate, respectively.The activities of LDH and MLDH were assessed at
50°C by mixing the enzymes with 50 mM HEPES-NaOH (pH 7.0), 5 mM MgCl2, 0.5 mM MnCl2, 0.2 mM
NADH, and 0.2 mM pyruvate. NADH oxidation wasmonitored at 340 nm.The heat-treated cell lysate of the recombinant E. coli
harboring an empty vector showed no detectable level ofenzyme activity under the assay conditions.
Lactate productionThe reaction mixture (4 ml) was composed of 0.1 mMglucose, 0.2 mM 3-PG, 0.2 mM pyruvate, 5 mM MgCl2,0.5 mM MnCl2, 0.2 mM ATP, 0.2 mM ADP, 1 mM NAD+,0.2 mM NADH, 1 mM G1P, and 50 mM HEPES-NaOHbuffer (pH 7.0). The cell suspensions of E. coli producingGK, PGI, PFK, FBA, TIM, GAPN, iPGM, ENO, PK, andMLDH were preheated at 70°C for 30 min and thenadded into the reaction mixture at final concentrations of2, 1, 1, 1, 1, 26, 3, 3, 3, and 100 mg (wet weight cells)/ml,respectively. The reaction mixture was stirred in acontainer kept at 50°C, and glucose solution (40 mM)was added to the mixture at a flow rate of 1 μl min-1
(= 0.01 μmol ml-1 min-1) using a Shimadzu LC-20 ADsolvent delivery unit. Aliquots (50 μl) of the reactionmixture were withdrawn at 1 h intervals, diluted fourfoldwith distilled water, and centrifuged to remove the celldebris (15,000 rpm, 10 min). The supernatant was thenultrafiltered using Amicon 3 K (Millipore) and analyzed bya high-performance liquid chromatography (HPLC).
Analytic methodsLactate and pyruvate were quantified by HPLC on twotandemly connected ion exclusion columns (Shim-packSPR-H, 250 mm×7.8 mm, Shimadzu). The columnswere eluted at 50°C using 4 mM p-toluenesulfonic acidas a mobile phase at a flow rate of 0.3 ml min-1. Theeluent was mixed with a pH-buffered solution (16 mMBis-Tris, 4 mM p-toluenesulfonic acid, and 0.1 mMEDTA) supplied at a flow rate of 0.3 ml min-1, and thenanalyzed for lactate using a conductivity detector (CDD-20A, Shimadzu). NAD+ and NADH concentrations wereanalyzed colorimetrically using a NAD/NADH quan-tification kit (Biovision, Mountain View, CA, USA) inaccordance with the procedure provided in the kit. ATPand ADP concentrations were assessed quantitativelyusing the EnzyLight ADP/ATP ratio assay kit (BioAssaySystems, Hayward, CA, USA) according to the manufac-turer’s instructions.
Competing interestsThe authors declare that they have no competing interests.
Authors’ contributionsXY performed the experiments and wrote the manuscript. KH designed allthe experiments and wrote the manuscript. TS, KO and RH co-performed theexperiments on the enzyme characterization. TO and AK contributed generaladvice, particularly on the thermophilic microorganisms, and resourcesupport, as well as edited the manuscript. HO conceived the project andwrote the manuscript. All authors read and approved the final manuscript.

Ye et al. Microbial Cell Factories 2012, 11:120 Page 10 of 11http://www.microbialcellfactories.com/content/11/1/120
AcknowledgementsThis work was in part supported by the Japan Science and TechnologyAgency (JST), PRESTO program. This work was also partly supported by theJapan Society for the Promotion of Science (JSPS), Japanese-GermanGraduate Externship Program. We are grateful to H. Atomi (Kyoto University,Japan) for kindly donating the Thermococcus GAPN expression vector. Wethank T. Hirasawa (Osaka University, Japan) for the CE-TOFMS analysis. Wealso thank T. Pongtharangkul (Mahidol University, Thailand) for a criticalreading of the manuscript.
Author details1Department of Biotechnology, Graduate School of Engineering, OsakaUniversity, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan. 2PRESTO, JapanScience and Technology Agency (JST), 4-1-8 Honcho, Kawaguchi, Saitama332-0012, Japan. 3Department of Life System, Institute of Technology andScience, The University of Tokushima, 2-1 Minamijosanjimacho, Tokushima770-8506, Japan. 4Department of Molecular Biotechnology, HiroshimaUniversity, 1-3-1 Kagamiyama, Higashi-Hiroshima, Hiroshima 739-8530, Japan.
Received: 14 June 2012 Accepted: 31 August 2012Published: 6 September 2012
References1. Rohlin L, Oh MK, Liao JC: Microbial pathway engineering for industrial
processes: evolution, combinatorial biosynthesis and rational design.Curr Opin Microbiol 2001, 4:330–335.
2. Stephanopoulos G: Challenges in engineering microbes for biofuelsproduction. Science 2007, 315:801–804.
3. Stephanopoulos G, Sinskey AJ: Metabolic engineering-methodologies andfuture prospects. Trends Biotechnol 1993, 11:392–396.
4. Kwok R: Five hard truths for synthetic biology. Nature 2010, 463:288–290.5. Zhang YHP: Production of biocommodities and bioelectricity by cell-free
synthetic enzymatic pathway biotransformations: challenges andopportunities. Biotechnol Bioeng 2010, 105:663–677.
6. Hodgman CE, Jewett MC: Cell-free synthetic biology: thinking outside thecell. Metab Eng 2012, 14:261–269.
7. Welch P, Scopes RK: Studies on cell-free metabolism: Ethanol productionby a yeast glycolytic system reconstituted from purified enzymes.J Biotechnol 1985, 2:257–273.
8. Algar EM, Scopes RK: Studies on cell-free metabolism: Ethanol productionby extracts of Zymomonas mobilis. J Biotechnol 1985, 2:275–287.
9. Demirjian DC, Morís-Varas F, Cassidy CS: Enzymes from extremophiles. CurrOpin Chem Biol 2001, 5:144–151.
10. Niehaus F, Bertoldo C, Kähler M, Antranikian G: Extremophiles as a sourceof novel enzymes for industrial application. Appl Microbiol Biotechnol 1999,51:711–729.
11. Honda K, Maya S, Omasa T, Hirota R, Kuroda A, Ohtake H: Production of2-deoxyribose 5-phosphate from fructose to demonstrate a potential ofartificial bio-synthetic pathway using thermophilic enzymes. J Biotechnol2010, 148:204–207.
12. Sato M, Masuda Y, Kirimura K, Kino K: Thermostable ATP regenerationsystem using polyphosphate kinase from Thermosynechococcuselongatus BP-1 for D-amino acid dipeptide synthesis. J Biosci Bioeng 2007,103:179–184.
13. Iwamoto S, Motomura K, Shinoda Y, Urata M, Kato J, Takiguchi N, Ohtake H,Hirota R, Kuroda A: Use of an Escherichia coli recombinant producingthermostable polyphosphate kinase as an ATP regenerator to producefructose 1,6-diphosphate. Appl Environ Microbiol 2007, 73:5676–5678.
14. Wang Y, Zhang YHP: Overexpression and simple purification of theThermotoga maritima 6-phosphogluconate dehydrogenase in Escherichiacoli and its application for NADPH regeneration. Microb Cell Fact 2009,8:30.
15. Yao S, Mikkelsen MJ: Metabolic engineering to improve ethanolproduction in Thermoanaerobacter mathranii. Appl Microbiol Biotechnol2010, 88:199–208.
16. Wichmann R, Vasic-Racki D: Cofactor regeneration at the lab scale. AdvBiochem Eng Biotechnol 2005, 92:225–260.
17. Verhees CH, Kengen SW, Tuininga JE, Schut GJ, Adams MW, de Vos WM,van der Oost J: The unique features of glycolytic pathways in Archaea.Biochem J 2003, 375:231–246.
18. Dandekar T, Schuster S, Snel B, Huynen M, Bork P: Pathway alignment:application to the comparative analysis of glycolytic enzymes. Biochem J1999, 343:115–124.
19. Kengen SWM, Tuininga JE, de Bok FAM, Stams AJM, de Vos WM:Purification and characterization of a novel ADP-dependent glucokinasefrom the hyperthermophilic archaeon Pyrococcus furiosus. J Biol Chem1995, 270:30453–30457.
20. Tuininga JE, Verhees CH, van der Oost J, Kengen SW, Stams AJ, de Vos WM:Molecular and biochemical characterization of the ADP-dependentphosphofructokinase from the hyperthermophilic archaeon Pyrococcusfuriosus. J Biol Chem 1999, 274:21023–21028.
21. Mukund S, Adams MW: Glyceraldehyde-3-phosphate ferredoxinoxidoreductase, a novel tungsten-containing enzyme with a potentialglycolytic role in the hyperthermophilic archaeon Pyrococcus furiosus.J Biol Chem 1995, 270:8389–8392.
22. Brunner NA, Brinkmann H, Siebers B, Hensel R: NAD+-dependentglyceraldehyde-3-phosphate dehydrogenase from Thermoproteus tenax.The first identified archaeal member of the aldehyde dehydrogenasesuperfamily is a glycolytic enzyme with unusual regulatory properties.J Biol Chem 1998, 273:6149–6156.
23. Ettema TJ, Ahmed H, Geerling AC, van der Oost J, Siebers B: The non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase (GAPN)of Sulfolobus solfataricus: a key-enzyme of the semi-phosphorylativebranch of the Entner-Doudoroff pathway. Extremophiles 2007, 12:75–88.
24. Matsubara K, Yokooji Y, Atomi H, Imanaka T: Biochemical and geneticcharacterization of the three metabolic routes in Thermococcuskodakarensis linking glyceraldehyde 3-phosphate and 3-phosphoglycerate. Mol Microbiol 2011, 81:1300–1312.
25. Dumbrepatil A, Adsul M, Chaudhari S, Khire J, Gokhale D: Utilization ofmolasses sugar for lactic acid production by Lactobacillus delbrueckiisubsp. delbrueckii mutant Uc-3 in batch fermentation. Appl EnvironMicrobial 2008, 74:333–335.
26. Joglekar HG, Rahman I, Babu S, Kulkarni BD, Joshi A: Comparativeassessment of downstream processing options for lactic acid. Sep PurifTech 2006, 52:1–17.
27. Kim YH, Moon SH: Lactic acid recovery from fermentation broth usingone-stage electrodialysis. J Chem Technol 2001, 76:169–178.
28. Wu JT, Wu LH, Knight JA: Stability of NADPH: effect of various factorson the kinetics of degradation. Clin Chem1986, 32:314–319.
29. González JM, Masuchi Y, Robb FT, Ammerman JW, Maeder DL,Yanagibayashi M, Tamaoka J, Kato C: Pyrococcus horikoshii sp. nov., ahyperthermophilic archaeon isolated from a hydrothermal vent at theOkinawa Trough. Extremophiles 1998, 2:123–130.
30. Oshima T, Imahori K: Description of Thermus thermophilus (Yoshida andOshima) comb. nov., a nonsporulating thermophilic bacterium from aJapanese Thermal Spa. Int J Syst Bacteriol 1974, 24:102–112.
31. Johnsen U, Schönheit P: Characterization of cofactor-dependent andcofactor-independent phosphoglycerate mutases from Archaea.Extremophiles 2007, 11:647–657.
32. Garvie EI: Bacterial lactate dehydrogenases. Microbiol Rev 1980, 44:106–139.33. Yokoyama S, Matsuo Y, Hirota H, Kigawa T, Shirouzu M, Kuroda Y,
Kurumizaka H, Kawaguchi S, Ito Y, Shibata T, Kainosho M, Nishimura Y, InoueY, Kuramitsu S: Structural genomics projects in Japan. Nat Struct Biol 2000,7:943–945.
34. Itoh A, Ohashi Y, Soga T, Mori H, Nishioka T, Tomita M: Application ofcapillary electrophoresis-mass spectrometry to synthetic in vitroglycolysis studies. Electrophoresis 2004, 25:1996–2002.
35. Meadow ND, Fox DK, Roseman S: The bacterial phosphoenolpyruvate:glycose phosphotransferase system. Annu Rev Biochem 1990, 59:497–542.
36. Postma PW, Lengeler JW, Jacobson GR: Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol Rev 1993,57:543–594.
37. Lee JW, Kim TY, Jang YS, Choi S, Lee SY: Systems metabolic engineeringfor chemicals and materials. Trends Biotechnol 2011, 29:370–378.
38. Valverde F, Losada M, Serrano A: Engineering a central metabolicpathway: glycolysis with no net phosphorylation in an Escherichia coligap mutant complemented with a plant GapN gene. FEBS Lett 1999,449:153–158.
39. Ansell RJ, Lowe CR: Artificial redox coenzymes: Biomimetic analogues ofNAD+. Appl Microbiol Biotechnol 1999, 51:703–710.

Ye et al. Microbial Cell Factories 2012, 11:120 Page 11 of 11http://www.microbialcellfactories.com/content/11/1/120
40. Ryan JD, Fish RH, Clark DS: Engineering cytochrome P450 enzymes forimproved activity towards biomimetic 1,4-NADH cofactors. Chem BioChem 2008, 9:2579–2582.
41. Tuesink B, Passarge J, Reijenga CA, Esgalhado E, van der Weijden CC,Schepper M, Walsh MC, Bakker BM, van Dam K, Westerhoff HV, Snoep JL:Can yeast glycolysis be understood in terms of in vitro kinetics of theconstituent enzymes? Testing biochemistry. Eur J Biochem 2000,267:5313–5329.
42. Conrado RJ, Varner JD, DeLisa MP: Engineering the spatial organization ofmetabolic enzymes: mimicking nature’s synergy. Curr Opin Biotechnol2008, 19:492–499.
doi:10.1186/1475-2859-11-120Cite this article as: Ye et al.: Synthetic metabolic engineering-a novel,simple technology for designing a chimeric metabolic pathway.Microbial Cell Factories 2012 11:120.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit





![Computational Approaches to Design and Test Plant ...Update on Plant Synthetic Metabolic Pathways Computational Approaches to Design and Test Plant Synthetic Metabolic Pathways1[OPEN]](https://img.pdfslide.us/doc/110x75/6118a3dece051064bf699784/computational-approaches-to-design-and-test-plant-update-on-plant-synthetic.jpg)





![Engineering of Metabolic Pathways Using Synthetic Enzyme Complexes1… · Engineering of Metabolic Pathways Using Synthetic Enzyme Complexes1[OPEN] Nicholas Smirnoff2,3 Biosciences,](https://img.pdfslide.us/doc/110x75/5eba4b9504ad29098d67bc72/engineering-of-metabolic-pathways-using-synthetic-enzyme-engineering-of-metabolic.jpg)

















