Embed Size (px)
Citation preview

Synthetic Collective IntelligenceRicard SoléDaniel R. AmorSalva Duran-NebredaN. Conde-PueyoMax CarbonllRaul Montañez
SFI WORKING PAPER: 2015-10-043
SFIWorkingPaperscontainaccountsofscienti5icworkoftheauthor(s)anddonotnecessarilyrepresenttheviewsoftheSantaFeInstitute.Weacceptpapersintendedforpublicationinpeer-reviewedjournalsorproceedingsvolumes,butnotpapersthathavealreadyappearedinprint.Exceptforpapersbyourexternalfaculty,papersmustbebasedonworkdoneatSFI,inspiredbyaninvitedvisittoorcollaborationatSFI,orfundedbyanSFIgrant.
©NOTICE:Thisworkingpaperisincludedbypermissionofthecontributingauthor(s)asameanstoensuretimelydistributionofthescholarlyandtechnicalworkonanon-commercialbasis.Copyrightandallrightsthereinaremaintainedbytheauthor(s).Itisunderstoodthatallpersonscopyingthisinformationwilladheretothetermsandconstraintsinvokedbyeachauthor'scopyright.Theseworksmayberepostedonlywiththeexplicitpermissionofthecopyrightholder.
www.santafe.edu
SANTA FE INSTITUTE

Synthetic collective intelligence
Ricard Sole∗,1, 2, 3 Daniel R. Amor,1, 2 Salva Duran-Nebreda,1, 2 Nuria Conde-Pueyo,1, 2 Max Carbonell,1, 2 andRaul Montanez1, 21ICREA-Complex Systems Lab, Universitat Pompeu Fabra, Dr Aiguader 88, 08003 Barcelona, Spain2Institut de Biologia Evolutiva, UPF-CSIC, Psg Barceloneta 37, 08003 Barcelona, Spain3Santa Fe Institute, 1399 Hyde Park Road, Santa Fe NM 87501, USA
Intelligent systems have emerged in our biosphere in different contexts and achieving differentlevels of complexity. The requirement of communication in a social context have been in all casesa determinant. The human brain, probably co-evolving with language, is an exceedingly successfulexample. Similarly, social insects complex collective decisions emerge from information exchangesbetween many agents. The difference is that such processing is obtained out of a limited individualcognitive power. Computational model and embodied versions using non-living systems, particu-larly involving robot swarms have been used to explore the potentiality of collective intelligence.Here we suggest a novel approach to the problem grounded in the genetic engineering of unicellularsystems, which can be modified in order to interact, store memories or adapt to external stimuliin collective ways. What we label as Synthetic Swarm Intelligence defines a parallel approach tothe evolution of computation and swarm intelligence and allows to explore potential embodiedscenarios for decision making at the microscale. Here, we consider several relevant examples ofcollective intelligence and their synthetic organism counterparts.
Keywords: Synthetic biology, swarm intelligence, evolution, social insects, cellular machines
Contents
I. Introduction 1
II. Natural and synthetic swarms 2
III. Quorum sensing and collective behaviour 4
IV. Majority detection and symmetry breaking 8
V. Self-regulation of division of labor 11
VI. Collective oscillations 13
VII. Discussion and conclusions 15
Acknowledgments 17
VIII. References 18
I. INTRODUCTION
Intelligent behaviour has been defined as the ability tosuccessfully operate in uncertain environments by adap-tations based on experience (Stenberg 2000). To succeed,intelligent agents have to be able to measure information,sense their inner and environment states, and performactions, receiving some kind of reward (Legg and Hut-ter 2006). Additionally, agents have to be able to storethese experiences and properly correlate experiences, be-coming memory an additional item essential for systemsadaptation and the development of an intelligent behav-ior. Our biosphere is currently populated by a plethora
∗Corresponding author
of information-processing entities. They are different in-stances of life forms. As pointed by John Hopfield, a cru-cial difference between physical and biological objects isthat the second perform computations (Hopfield 1994).Such special feature also distinguishes biological struc-tures as highly non-equilibrium ones, where informationplays a role far more relevant from pure energy and mat-ter.
Among the major transitions that have punctuated thehistory of life on our planet, several great steps involvednovel ways of dealing with information and adapting tothe external world by means of non-genetic mechanisms.These cellular, physiological, anatomical and behavioralinnovations became a key factor in dealing with com-plex environments. As organisms became capable of pro-cessing information in complex ways, they became ableof dealing with uncertainty. Intelligent behaviour waslargely responsible for the success of costly multicellu-lar systems as a consequence of their potential for be-ing autonomous agents capable of storing and processingepigenetic information. The result was, as Jablonka andLamb (2006) pointed out, the emergence of the neuralindividual:
”...with a high level of internal integrationand the ability to make rapid adaptive re-sponses. However, the emergence of the neu-ral individual meant more than a change inthe nature and speed of adaptation. Neu-ral processing led to behaviour based on sen-sory perception, and this in turn led to aform of communication between individualsthat did not require contact or the trans-mission of physical material from one to theother. This mode of information transmissionhad interesting consequences, one of which

2
was the ability of animals to learn from oth-ers through perceiving their behaviour or theoutcomes of their behaviour, i.e. it led to so-cial learning.”
Different kinds of neural structures can be identifiedwithin the long road from cellular systems with simplesensors to full-fledged brains (Rose 2005). After cellmembranes became complex enough, it became possi-ble to process information in terms of chemical signalsand internal metabolic responses. Later on, movement,coupled to these signalling phenomena allowed autonomyto develop beyond simple changes of metabolite concen-trations. The rise of multicellularity added a new po-tential for using specific types of cells facing the exter-nal world, whereas the rest could be freed from a directinteraction with the environment. As multicellular lifeforms further evolved, something close to ”behaviour”could be defined. With multicellularity, and with cellsalready in place having an internal and dynamical skele-ton (the so called cytoskeleton) a novel form of cellularstructure dominated the scene: neurone and neural netsbecame the controllers of behavioural patterns and therudimentary cellular computers. They paved the way toa rapid increase in complexity with the spatial segrega-tion of groups of specialised neurons (distributed over anumber of clusters known as ganglia) and eventually tothe complex brains.
Along with this neural individual, another major tran-sition of evolution led to the emergence of societiesformed by relatively simpler agents (such as ants, ter-mites or bees) capable of performing complex cognitiveactions at the collective level. Social insects belong, atthe individual level, to the ganglion-based organisation ofneural processing. But they reach a higher level of com-plexity by making these already sophisticated systems tointeract within a much more complex, parallel and spa-tially distributed entity: the super organism (Holdobllerand Wilson 2008). By means of their potential for gath-ering and storing information, systems displaying collec-tive intelligence are capable of making decisions abouttheir internal states, environmental resources, protectionagainst damage or foraging strategies (Deneubourg andGoss, 1989; Camazine et al., 2002). An example of thesesystems is given by army ants (figure 1a) which are blindand communicate mostly by means of pheromones, andyet involve millions of individuals that are capable of co-herently exploring vast areas in the rain forests.
In some ways, collective intelligent systems, particu-larly ant colonies, reminds us the way brains work (Hof-stadter, 1980; Gordon et al 1991*?, Sole and Goodwin2001). However, in such comparison ants must be seenas a fluid neural network (Sole et al 1993) since no stableconnections between pairs of ants exist. To a large extent,the cognitive potential associated to social insect be-haviour is tied to both individual and collective responsesto stimuli. On the one hand, ants and other social insectsare capable of responding in sharp, almost digital waysto external inputs. On the other hand, the emergence of
nest complexity comes from self-organization (Millonas1993; Gordon 1999, 2010; Detrain and Deneubourg, 2006;Sumpter 2006), since ants exploit bifurcations and non-linearities as their source of internal structuring and de-cision making (Deneubourg et al. 1989; Bonabeau 1996).
Collective intelligence has been studied under very dif-ferent approaches, including controlled experimental sys-tems, simulation and computational models. A great dealof knowledge has been reached though the study of phy-logenies and experimental observation or manipulation ofsocial insects. Moreover, computational and mathemat-ical modelling (Bonabeau et al 1999) along with swarmrobotics (figure 1b) have contributed to unravel the un-derlying rules associated to the origins of collective intel-ligence.
The emergent field of synthetic biology give us an es-sentially new way of exploring swarm intelligence. Thisfield provides the tools to modify and redesign livingsystems, instead of building artificial agents or makingmathematical models. We could properly claim thatwe can create living micro-robots by modifying the be-havioural repertoire of existing cells.
In this paper we outline this synthetic biology path,pointing a set of minimal circuits theoretically able toimplement collective intelligence in bacteria. We labelthis field as synthetic collective intelligence (SCI). Suchtype of collective response can be found in many otherdifferent contexts, some of which are related to bacterialconsortia (figure 1c) known as biofilms, where a collec-tive response leads to the formation of a super-cellularstructure. But can also involve tissue-like response phe-nomena, as it occurs with how pancreatic cells manageglucose regulation (figure 1d). In this case, specific cellsin tissues can also react in non-linear ways to the concen-tration of given molecules and allow the tissue to makedecisions on how to react to concentration changes.
As pointed by several authors, microbes are capableof integrating sensory information, store memories anddisplay behavioural control (Lambert and Kussell 2014,Vladimirov and Sourjik 2009). They thus incorporateseveral relevant components required to build or designcomplex decision making systems. This starting point al-lows to consider the engineering of microbes as an achiev-able goal. Below we show that several essential featuresexhibited by swarm systems can be mapped into existingor engineered microbial constructs, and how these en-gineered living swarms map different types of emergentcollective behaviour.
II. NATURAL AND SYNTHETIC SWARMS
Why using living systems and genetic engineering? Agood reason is to use embodied communicating agents:it has been shown that embodiment is a crucial compo-nent of biologically-inspired robotics (Pfeifer et al 2007).Embodiment here needs to be understood as the dynam-ically linked interactions between ant brains, their body
3
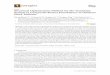
FIG. 1 Collective decision making in natural and artificial systems. Many different systems are capable of making decisionsthat are based on detecting and processing information in ways that require population responses. Social insects are the obviousexample, Alex as illustrated by army ants (a) where very large swarms emerge out from local interactions (Image by Alex Wild,from wikipedia commons). Swarm robotics (b) has taken some advantage of these collective patterns. In microbial systemscollective responses are often associated to the formation of so called biofilms (c) where spatially structured organisations emerge(image by Ronn Friedlander and Michael Bucaro). In physiology too organ-level functions, such as glucose regulation by thepancreas (d), result from sensing and responding to external signals (image by H. E. Farbung, from Wikimedia commons).
and the external world. Since designed microbial agentswill live and operate in a given spatial niche, the localnature of interactions and other physical constraints willplay a role. On the other hand, we also expect to finduniversal features that are shared by both insect coloniesand their synthetic microbial counterparts. In order toillustrate this point, we consider, in the following text,a selected list of well known cases associated to severalrelevant forms of collective behaviour. Our approachwill describe minimal mathematical models that capturethe key aspects of these case studies and finally provideexamples of engineered systems that would implementthose properties in synthetic biology.
Our aim is to define the minimal set of rules that per-vade collective behaviour, where a two-way communica-tion exists between individual and colony-level signals.As an example, if we take ants searching for resources,finding food by a single ant triggers an individual re-sponse that involves the release of a chemical signal thatother ants can detect. As other ants find the mark andfollow it, they reinforce the chemical field and eventu-ally a whole chemical pheromone trail will form. Thisbehaviour can be described as a closed loop connectingtwo different scales in an unbreakable fashion. Thereby,
the trail can be seen as a larger-scale structure that iscreated by ants while it also affects their individual be-haviour. This closed causality is a characteristic patternin most -if not all- self-organised systems. Such causalloop needs to be considered as a crucial part of our ap-proach to synthetic swarms.
Early experimental and theoretical work on the originsof swarm intelligence, as it occurs with other complexsystems, identified the presence of phase transition phe-nomena similar to those known in physics (Sole 2011).It seems clear that insect colonies are capable of shiftingbetween different global states (phases) when they re-spond to environmental changes. From a random walk-ing exploration of the environment to a coordinate ac-complishment of their tasks. Beyond the accuracy ofthis comparison, which has also been made in other ar-eas (Sole 2011), examples in insect colonies include thediversity of morphological casts (the abundance of dif-ferent function-related phenotypes in the colony), nestorganisation (Bonabeau et al 1997) or hierarchy (Osterand Wilson, 1978; Valverde et al 2009). But cells are alsoable to coordinate by diffusible signals. For instance, cellsoften respond to molecules in their environment via in-tracellular signalling pathways that eventually result in

4
altered concentration of extracellular molecular speciesthat triggered the pathway (Wiley et al., 2003, Pribyl etal., 2003; Freeman, 2000).
The analogy with phase transition phenomena is moreprofound that it might seem at first sight. The emergenceof collective behaviour requires from amplification pro-cesses that allow the system to shift from less-coherent,noisy behaviour (a disordered phase) to a coherent, moreorganised (ordered) phase. Existing studies and mod-els of different types of collective intelligence support theidea that the laws governing these transitions are ratheruniversal, as it occurs in physical systems (llibres?). Butwe know from the statistical physics of phase transitionsthat the way these changes occur obeys rather specifictrends. This is a signature of so called universal be-haviour (Sole and Goodwin, 2001, Sole 2011). More im-portantly, it is known from these studies that the globalbehaviour of the system is rather insensitive of the natureof the individual components. This means that, once thereally essential rules and interactions are in place, verydifferent systems sharing this minimal features will be-have in the same way. If this can be extrapolated tobiological systems, we would be able to engineer the syn-thetic counterparts of natural swarms.
In this paper we aim to address several importantquestions. Will synthetic (microbial-based) and natural(insect-based) systems behave in similar ways? Can novelforms of information processing be achieved by engineer-ing living cells? In the next sections we present severalexamples of proof-of-concept designs that can imitate theways social insects make decisions, exhibit division of la-bor, synchronise or simply aggregate as a consequenceof threshold-like phenomena. It will be shown that somecrucial components of future synthetic swarms are deeplyconnected to the universality of the rules pervading col-lective intelligence.
III. QUORUM SENSING AND COLLECTIVEBEHAVIOUR
Living cells both secrete and sense a wide variety ofchemical signals (Alberts 2002). Releasing molecules tothe external medium can be a consequence of metabolismbut also a response to internal and/or external signals.Similarly, cell membranes act as the interface with theexternal world, with a very large fraction of proteins act-ing as receptors and information sensors located at thisinterface (Alberts et al. 2002). Both components (figure2a,b) are much needed but none in itself is enough to trig-ger complex adaptive responses. In order to achieve thisgoal, cells need to do something more: create and sensethe same class of molecule, as ants following a pheromonepath. This is essential to support many instances of col-lective intelligence, since releasing a signal that others(including the releaser) can sense provides a feedback be-tween individual and system-level behaviour.
In order to engineer synthetic systems capable of deci-
sion making and collective behaviour, one special compo-nent can be used: a system capable of sensing the size ofthe population. In ant colonies, it has been shown thatthreshold responses are reached once critical numbers ofindividuals are attained. This is the case of foraging dy-namics of the Pharaoh’s ants (Beekman et al 2001) whichexhibit a marked transition from disordered to orderedpatterns of foraging as the colony size increases beyonda given threshold (cites, afegir Prigogine Ourder Out ofChaos). In this way, exploration can dominate while re-sources (and the sensing systems associated to them) arelow but a shift to active exploitation occurs once enoughindividuals are engaged in a given task. In many antspecies, foragers lay pheromone trails from food sourcesback to their nest. These trails allow nestmates to locateand exploit the source easily. As more ants collect foodthey reinforce the trail. Although the pheromone trailis volatile, it can be maintained if sufficient ants use it.This type of dynamics can be modelled, showing how thenumber of ants walking to a single food source dependson colony size.
In microbial species, a closely related phenomenon oc-curs. It involves a mechanism known as quorum sensing(QS) and is the key example that illustrates the existenceof communication phenomena in microbial populationsbeyond the single cell (Waters and Bassler 2005). QS al-lows bacteria to monitor the environment for other bac-teria and to alter behavioural responses on a population-wide scale in response to changes in the number and/orspecies present in a community. As a consequence, mi-crobial colonies can make collective decisions, from hostinfection (Jones et al. 1993) to the formation of a socalled biofilm (Davies et al. 1998). Most QS controlledprocesses are unproductive when undertaken by an in-dividual bacterium acting alone but become beneficialwhen carried out simultaneously by a large number ofcells. Thus, quorum sensing enables microbes to act (ef-fectively) as multicellular organisms. That is why QSpervades a large range of phenomena, from biofilm for-mation to the symbiotic coordination between legumi-nous plant and rhizobia (Gonzalez and Marketon 2003).In this section, we will analyse a toy model of QS thatenables us to introduce some of the basic methods toanalyse other, more complex mechanisms of collective be-haviour.
These basic mechanisms underlying QS are illustratedin Figure 2a-c. Three main components are required,namely: i) cell-level system of production of the key sig-nal, ii) a diffusion of this molecule through the cell mem-brane (in and out) and iii) cellular receptors actuallysensing the external (environmental) concentration of thesignal. By being both synthesized and detected, a min-imal network motif, a feedback control loop is created.QS signaling networks can be more complicated and evenperform sophisticated kind of computations (Waters andBassler 2005). Thereby, QS is postulated here as oneof the minimal requirements to implement swarm intelli-gence in microorganisms.

5
FIG. 2 An essential motif associated to collective intelligence implies the combination of (a) production and (b) sensing ofgiven molecules into a single feedback loop (c) where production and sensing are coupled. Such scenario provides the necessaryconditions for sensing -for example-the population density of neighbouring agents (cells, ants etc). Many social insects (asthe Pharao’s ants displayed in the right picture) display a characteristic transition from disorganised, low-density groups ofindividuals to ordered, collective patterns of organisation once the number of individuals crosses given thresholds. QS-relatedphenomena are responsible for this shifts in microorganism.
We can describe the threshold-like behaviour of a QSsystem using a simple mathematical model that cap-tures the essential nonlinearities associated to this phe-nomenon. Instead of looking at individuals and themolecules they send and detect, let us perform a meanfield approach. Mean field models are broadly used inphysics to reduce the complexity of many-body (or many-agents) problems, by looking at it on a coarse grainedlevel (Sole 2011). Let us here consider that only theaverage effect of individual (e.g. cellular or molecular)interactions is relevant. In this way, let us take the con-centration of the signalling molecule Q as the only statevariable of the system. This means that, according toour mean field approach, Q(t) is the only information weneed to describe the state of the system at time t. OurQ molecule could potentially govern a variety of cellularprocesses, from the induction of a reporter protein to thesecretion of a given pheromone or growth factor.
A simple equation can be written that is able to cap-ture the dynamics of Q over time:
dQ
dt= fµ(Q)− δQ, (1)
where fµ(Q) is a production term for the signal Q, and δstands for a degradation rate. We know from the observa-tion of natural systems that the production term displaya threshold-like phenomenon, which can be modelled bya so-called Hill-like function, namely
fµ(Q) =µρQ2
θ2 +Q2(2)
According to the equation (2), the concentration Q trig-
gers its own production with constant rate µ. Such trig-gering effect is proportional to the cell population densityρ. This dependence on ρ illustrates the role of cells assignal producers: the higher population density, the moreproduction of Q. The function fµ(Q) displays a specialshape, with low values for Q < θ and large values other-wise. Close to the threshold value θ it rapidly increases.The full equation of the QS field would thus read:
dQ
dt=
µρQ2
θ2 +Q2− δQ (3)
and roughly incorporates both the nonlinear, threshold-like production and the linear decay terms, respectively.
Despite its simplicity, the model properly displays theobserved threshold-like shift associated to increasing den-sity of individuals. Steady states Q∗ are obtained fromthe condition dQ/dt = 0 (i. e. when nothing changes)and it is easy to show that three points exist, namely:
Q∗ = 0 Q∗± =1
2
[µρ
δ±√(µρ
δ
)2− 4θ2
](4)
The first point is associated to a zero concentration field:production is unable to compensate degradation and noQS response would be observed. The two other pointsare related to non-zero QS responses, and they only existprovided that the density is larger than a critical value,namely a threshold density:
ρ > ρc =2θδ
µ(5)
6
1.0 1.5 2.0 2.5 3.0µ
0.0
1.0
2.0
Vege
tatio
n co
ver x
0.0 1.0 2.0¡
0.0
1.0
2.0
3.0
4.0
µ
a
Green
Dese
SWARM
1.0!!!!!!!!!!!!!!1.5!!!!!!!!!!!!!!!!2.0!!!!!!!!!!!!!!2.5!!!!!!!!!!!!!!!3.0
2.0
1.0
0.0
Q⇤
⇢
INDIVIDUAL Q+
Q�
-1 0 1 2 30 2
0
0 1 2 30 1 2 3
0
0.2
0.4
0.6
0.8
0 1 2 30 1 2 3-0.4-0.20
0.20.40.60.8
0 1 2 3 4 5 60 1 2 3 4 5 6-2
-1
0
1
�(Q
)
Q Q
�(Q
)
a
b c
d e
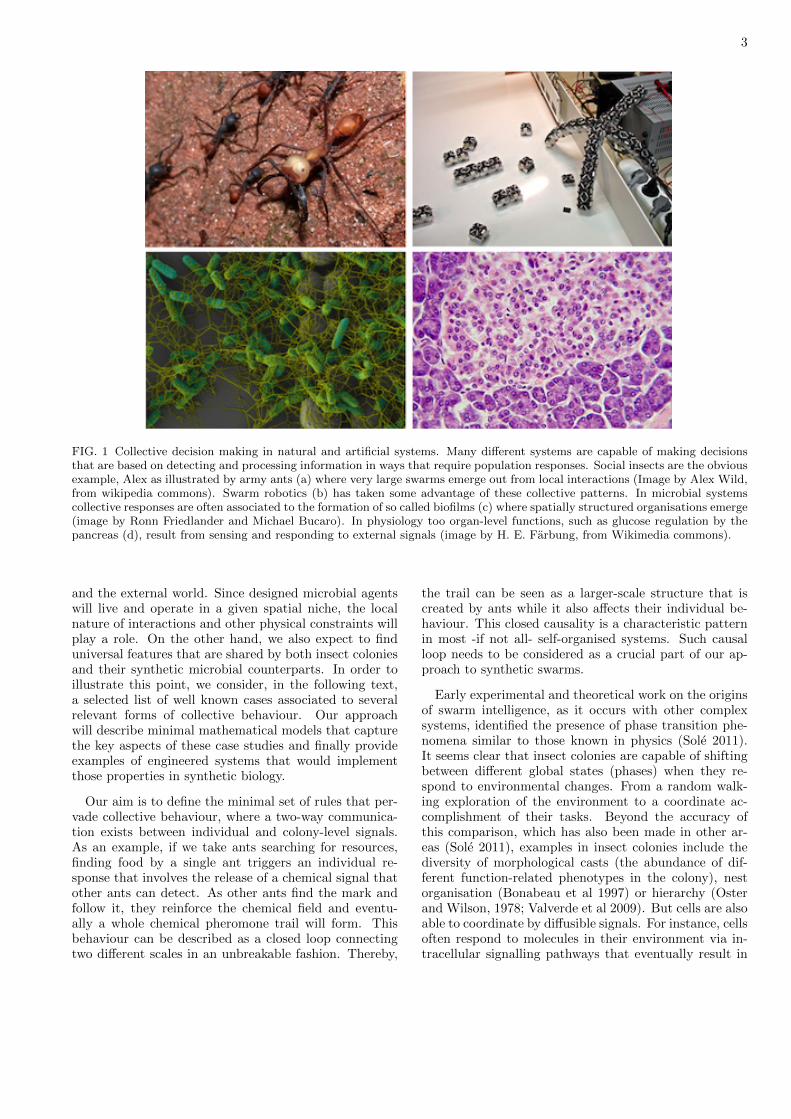
FIG. 3 Nonlinearities and breakpoints in the mean field modelfor quorum sensing. (a) Equilibrium concentration of thequorum sensing signal against the density of cells in thesystem (see text). The stable equilibrium values Q∗(ρ) (towhich close states converge) are indicated by the straight lineswhile the dashed line corresponds to the unstable equilibriumbranch. Beyond a critical density (see text) two branchesemerge, a stable Q+ and an unstable Q− one. A shift fromthe swarm phase (right) to an individual-level phase occurswhen we move through the upper branch Q+ until we reachthe minimal value at the critical point, here indicated by anopen circle. In (b-e) we show four examples of the potentialfunction for µ = 1, δ = 1 and θ = 1 and (b) ρ = 1.0, (c)ρ = 1.5, (d) ρ = 2.0 and (e) ρ = 2.5.
(otherwise they would be imaginary). It can be shownthat the first point is always stable, meaning that a smallincrease in Q is degraded towards the zero state. Instead,the two other points have different stability properties:Q∗+ is stable whereas Q∗− is unstable. This means that aninitial concentration close to the first will eventually tendto it while a small deviation from the second will grow,further increasing the distance from the unstable state.In general, the stability of the fixed points of a dynamicalsystem described by an equation dQ/dt = Γ(Q) can bedetermined (Strogatz 1994, Sole 2011) by evaluating the
sign of
λµ(Q∗) =
[dΓ(Q)
dQ
]Q∗
(6)
When λµ < 0 the point is stable, whereas it becomesunstable once λµ > 0. The case λµ = 0 corresponds to aso called marginally stable point.
These results are summarised in figure 3, where the socalled bifurcation diagram of our system is shown. Thisdiagram displays all the fixed points Q∗ against a key (bi-furcation) parameter. A shift occurs at the critical den-sity: a jump separates the zero state from Q = µρ/2δ,where a QS response would be observed for the first time.The existence of this gap implies that something needsto occur if we want to shift from the low (zero) state tothe QS state. This can occur, for example, if the un-derlying cells display stochastic fluctuations. In orderto address this component, we would need to include ex-plicitly the role played by noise, which has been shown tobe specially relevant in making swarms computationallyflexible (Deneubourg et al *** JTB, Bonabeau 1996). Analternative way of approaching this feature is to use thepotential Φ(Q), defined as an energy-like kind of func-tion. The key idea is that we can write our dynamicalsystem as derived from the potential, namely
dQ
dt= −∂Φ(Q)
∂Q(7)
where Φ(Q) is defined by
Φ(Q) = −∫
Γ(Q′)dQ′ (8)
It is not difficult to show that this function is such thatits minima and maxima correspond to the stable andunstable fixed points, respectively. This is a desirableproperty, that makes a strong connection with a physicalsystem where the state of a particle (for example, a pointmass under a gravitational field) rolls down on a surfaceuntil it reaches the minimum energy.
For our QS model, the potential function is obtainedby integrating the right-hand side of the QS dynamicalmodel (3), which gives:
Φ(Q) =δ
2Q2 − µρ
∫Q2
θ2 +Q2dQ (9)
=δ
2Q2 − µρ
[Q− θ arctan
(Q
θ
)](10)
Four different examples of this function are displayed infigure 3b-e. Here different values of ρ are used, two belowthe critical density and two beyond this value. The shapeof the potential is very revealing. For ρ < ρc we can seethat the only stable state is the Q∗ = 0 point, as ex-pected, but the shape of Φ(Q) is clearly deforming as weapproach ρc. Once this value is crossed, the deformationgrows and two valleys are observed. These correspond to

7
FIG. 4 Collective communication out of local production and sensing of signals. The top left diagram summarises the geneticlogic associated to a QS response. The specific inducible promoter (pQ) controls the expression of the Q molecule, that is inturn responsible for the self-detection feature associated to QS (see text). The sequence of a simulated growing colony has beenobtained by plotting the time evolution of a quorum sensing collective response in a stochastic, individual-based model [usingthe GRO package, (Jang et al. 2010)]. Here five consecutive snapshots of a two-dimensional simulation are shown. Each insilico cell can be described as the basic scheme provided in the upper diagram (a).
the minima associated to the two stable points obtainedfrom the linear analysis (indicated as circles in figures3d-e). But we can see that the trivial point Q∗ = 0 islocated at the bottom of a small valley (open circle) fromwhich a small perturbation might push it out (figure 3d).This tendency increases as we keep moving far from thebifurcation (figure 3e). This situation corresponds to aso called metastable state. For this state, we have thetwo conditions associated to a minimum, i. e.(
∂Φ(Q)
∂Q
)Q∗
= 0 (11)(∂2Φ(Q)
∂Q2
)Q∗
> 0 (12)
as expected. In this case we can say that
Φ(Q) > Φ(Q∗) (13)
when |Q − Q∗| is small (i.e. when we are really close tothe minimum), but we have
Φ(Q) < Φ(Q∗) (14)
otherwise (Dill and Bromberg 2011). In these cases,stability is usually a transient phenomenon, providedthat the fluctuations associated to the system are strongenough.
A broad, chemically diverse repertoire of QS signalmolecules has been described, of which the homoserinelactone (HSL) family have been the most intensivelystudied (Miller and Bassler 2001; Waters and Bassler2005; Goryachev 2009). Once the HSL concentration
reaches a critical threshold, a sensor molecule or responseregulator is activated, triggering the expression of QS-dependent target genes. This knowledge has been usedto modulate and modify the behaviour of many differentsynthetic systems. A minimal synthetic network that de-scribes the basic QS interactions is summarised in figure4(a). The circuit considers an inducible promoter (pQ)that controls de signalling inducer gene (QI) expressionto produce the signalling molecule (Q). This moleculecan pass through the cellular membrane and passivelydiffuse into the surrounding media, potentially reachingother bacteria. Inside cells, Q interacts specifically witha receptor protein (QR) in order to induces gene expres-sion at (pQ), thus controlling bith its own expression andthe expression of a green fluorescent protein.
An example of the dynamics of a growing bacterialcolony, rising from a unique cell integrating the syntheticcircuit described in figure 4(a), is shown by the snap-shots series in figures 4(b)-(f). The agent-based (Jang etal 2012) model we use here introduces additional degreesof complexity in comparison to the simple mean field ap-proach described above. A key difference is that we ex-plicitly consider the individual dynamics of cells. Thisincludes cellular growth and reproduction, as well as se-cretion and sensing processes regarding the AHL concen-tration Q. Moreover, spatial effects are considered, anddiffusion governs the dynamics of Q in the extracellularmedium. We also consider that the inducible promoterpQ exhibit a certain leakiness. Although such leakinessis not included in the minimal model described by Eq.(3), it is worth considering it in models for both bacte-ria (cells typically undergo a minimal HSL production

8
FIG. 5 Symmetry breaking in ant colonies. The image showsa late picture of the two-bridge experiment, with ants movingthrough one of two identical bridges between the nest and afood source (image courtesy of Guy Theraulaz). This typeof behaviour can be described by means of a simple modelwhere ants are capable of adding pheromones to each bridgewhile making their choices as a function of the pheromoneconcentration (see main text).
in the absence of signal) and ant swarms (in which thefirst individuals finding food have to be able to producethe signal from the zero state). In some cases, leakinessis introduced as those stochastic fluctuations that couldlead the system from the zero state to the active stablestate Q∗ (see figure 3 and its discussion above). Thetime series in figure 4(b)-(f) show that the cell colonyis over the critical density threshold [see Eq. (5)], sincethe active state (indicated by the production of a greenreporter protein) is achieved in the long run. However,according to our agent-based model, the colony has toreach a minimal number of cells so that the productionof the signal compensates both its diffusion and degra-dation. This explains the fact that the majority of cellsremain in the inactive state at initial stages [figures 4(b)and (c)]. After the minimal colony size is reached [fig-ure 4(c)], an increasing number of cells [figures 4(c)-(f)]switch to the active state. Such activation first appearsnear the center of the colony (where the concentration Qis higher).
IV. MAJORITY DETECTION AND SYMMETRYBREAKING
Because ants are capable of creating chemical fieldsthat can amplify existing signals (perhaps an incipienttrail) these global responses can be used to sense theenvironment and its variable clues in order to discrimi-nate, for example, between two nutrient sources select-ing the richest (Deneubourg and Goss 1989, Detrain andDeneubourg 2006, Garnier et al 2007). Similarly, ants
can use the QS-like mechanisms in order to determinethe shortest path to a given source among several exist-ing ones. In this way, colonies (not individuals) becomeable to decide between two food sources or two paths,finding the optimal solution. Such decision-making pro-cess is dramatically illustrated by the two-bridge experi-ment where the ant nest and a single nutrient source areconnected by two alternative paths (figure 5a).
Once ants start crossing the bridges, they will initiallydo so at random. We should expected at this point equalnumber of ants on each branch, i.e. ρ1 = ρ2. How-ever, once a successful ant has found the food sourceand gets back to the nest, it does so while releasing apheromone trail. Although this trail is faint, it can be de-tected by others, which will choose it and release furtherpheromones, thus amplifying the previous mark. Thepheromone trail also dissipates as molecules evaporate,but such process will be more effective in the longer trail,where more surface is available. As a result, the shortestpath is more likely to be used and is eventually chosen.Ants have computed the shortest path.
The previous experimental setup can be formalized byusing a very simple model involving (as it occurs in theQS case) Hill-like functions. If ρ1 and ρ2 indicate the con-centrations of trail pheromone in each branch, their dy-namics can be described (Nicolis and Denebourg, 1999)as a pair of coupled equations:
dρ1dt
= µq1P1(ρ1, ρ2)− νρ1 (15)
dρ2dt
= µq2P2(ρ1, ρ2)− νρ2 (16)
where µ is the rate of ants entering each branch fromthe nest, qi the rate of pheromone deposition in the i−thbranch and ν the rate of pheromone evaporation. Theprobabilities Pi(ρ1, ρ2) guide the choices made by ants(which bridge is chosen) given the pheromone concentra-tion. It has been shown that experimental observationsare well fitted by a threshold response function (Beckerset al., 1992; Deneubourg et al., 1990):
Pi(ρ1, ρ2) =(ρi +K)2
Θ(ρ1, ρ2)(17)
where Θ(ρ1, ρ2) =∑j=1,2(ρj + K)2 and i = 1, 2. The
parameter K measures the tendency for ants to explorea path that is free of pheromones (i.e., when ρi = 0), andthe exponent 2 is related to the steepness of the choicefunction (more abrupt choices could be modelled usinghigher exponents).
If we use an asymmetric system where branches aredifferent, (i.e. q1 6= q2) it can be shown that the ampli-fication process leads to the choice of the shortest path.Let us consider the symmetric case where q1 = q2 = qand thus the set of equations:
dρ1dt
= µq(ρ1 +K)2
Θ(ρ1, ρ2)− νρ1 (18)

9
⇢
�(⇢
)
a
⇢
�(⇢
)
b
⇢
�(⇢
)
c
FIG. 6 Symmetry breaking in a synthetic microbial consortia. Three snapshots of the mean field prediction for the process areshown, using the potential function Φ(ρ) associated to the RFP dynamics. In (a) we represent the state of the ρ1) variable attwo slightly asymmetric initial conditions: the red and green balls indicate initial signal concentrations slightly favouring theproduction of RFP and GFP, respectively. In (b) and (c), the dynamics of the RFP is shown by the red (green) ball rollingdown on Φ towards the equilibrium state showing saturation (absence) of the RFP reporter. According to the mean field model,the absence of RFP (ρ1 = 0) corresponds to the equilibrium state exhibiting saturation of GFP (ρ2 = 1), and vice versa.
dρ2dt
= µq(ρ2 +K)2
Θ(ρ1, ρ2)− νρ2 (19)
This particular scenario might appear not so interest-ing, since there is no true optimal choice: both branchesare equally efficient. We would easily conclude that antswould choose both paths and that individuals will equallyspread over both sides. This is not the case: typically,an amplification phenomenon occurs, and neverthelessants eventually choose one of the two possible branches.Why is that? The phenomenon illustrates a very impor-tant class of phase transition phenomena: the so calledsymmetry breaking process amplifies initial fluctuationsin such a way that even small differences are inevitablyexpanded by some type of positive feedback loop.
It is not difficult to show using dρi/dt = 0 that onepossible solution to this system is the symmetric stateρ∗1 = ρ∗2 = ρ∗ when ants equally distribute themselvesover both branches. For this special case, we havePi(ρ
∗1, ρ∗2) = P (ρ∗) = µq/2 and thus a single equa-
tion dρ∗/dt = µq/2 − νρ∗ which gives a fixed pointρ∗ = µq/(2ν). This is our symmetric state to be bro-ken. The second state would correspond to the choice ofone of the branches (ρ∗1 6= ρ∗2). Since the total pheromoneconcentration is ρ1 + ρ2 = 2ρ∗ = µq/ν, we can see that(µq
ν− ρ∗i
)(ρ∗i +K)2 = ρ∗i
(µqν− ρ∗i +K
)2(20)
after some algebra, this gives the new fixed points ρ∗+ =(ρ∗1+, ρ
∗2−) and ρ∗− = (ρ∗1−, ρ
∗2+) with
ρ∗i+ =µq
2ν+
√(µq2ν
)2−K2 ρ∗i− =
µq
2ν−√(µq
2ν
)2−K2.
(21)This pair of fixed points will exist provided that µq/2ν >K which allows to obtain the critical point at
µc =2Kν
q(22)
indicating that there is a minimal rate of ants enter-ing the bridges required to observe the symmetry break-ing phenomena. Below this value, the only fixed point
is the symmetric case with identical flows of ants ineach branch. This symmetric model can be general-ized to (more interesting) assymmetric scenarios wherethe two potential choices are different (see Detrain andDeneubourg 2009 and references cited) either because thefood sources have different size or because paths have dif-ferent lengths and the shortest path need to be chosen.
Implementing the previous experiment using a syn-thetic construct is far from trivial, but a closely relatedproblem can be considered that allows to find an equiva-lent SCI formulation using synthetic biology. It also im-plies a decision between two potential solutions resultingfrom an amplification phenomenon. In computation the-ory, it is connected to the so called density classificationproblem (Mitchell et al 1994; Crutchfield and Mitchell1995). Imagine a system that receives two types of in-puts and one must determine which of them is stronger.Specifically, consider two given molecules M1 and M2
that can be detected by a cell or set of cells. The questionto be solved is what was the signal with greater initialconcentration [M ], that is deciding between
[M1]0 > [M2]0 (23)
or instead the opposite:
[M1]0 < [M2]0 (24)
where [Mk]0 indicate the initial (t = 0) concentrationsof each molecular signal. Since the original concentra-tions can be small or very similar, the problem is how toamplify the initial condition in such a way that the finalstate represents a decision between one of the two previ-ous inequalities. This problem has been addressed usingdeterministic models where a discrete set of elements areset initially in one of two alternative states (0, 1). Eachelement in the system is connected to a limited number ofneighbours and their state is synchronously updated fol-lowing simple rules. The problem can now be redefinedconsidering a system where two signals can be sensedfrom the environment by a strain of engineered cells. Letus assume that these cells, as it occurs with the quo-rum sensing mechanism described above, not only detect
10
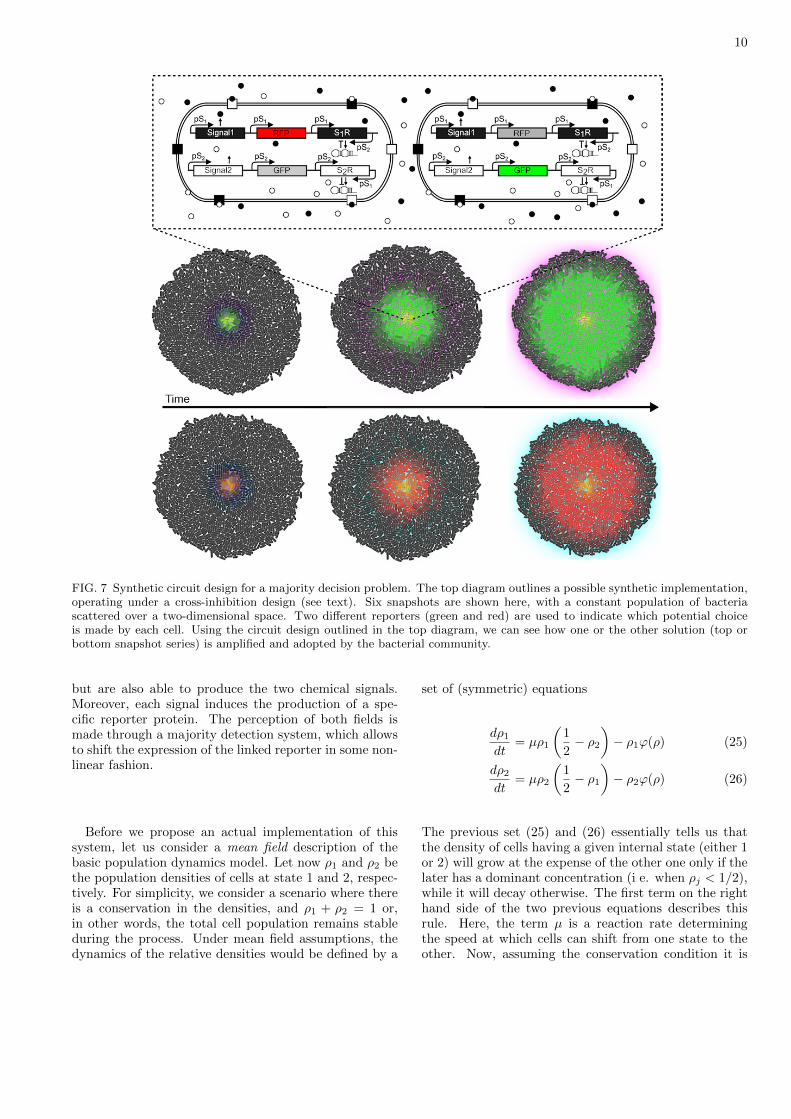
FIG. 7 Synthetic circuit design for a majority decision problem. The top diagram outlines a possible synthetic implementation,operating under a cross-inhibition design (see text). Six snapshots are shown here, with a constant population of bacteriascattered over a two-dimensional space. Two different reporters (green and red) are used to indicate which potential choiceis made by each cell. Using the circuit design outlined in the top diagram, we can see how one or the other solution (top orbottom snapshot series) is amplified and adopted by the bacterial community.
but are also able to produce the two chemical signals.Moreover, each signal induces the production of a spe-cific reporter protein. The perception of both fields ismade through a majority detection system, which allowsto shift the expression of the linked reporter in some non-linear fashion.
Before we propose an actual implementation of thissystem, let us consider a mean field description of thebasic population dynamics model. Let now ρ1 and ρ2 bethe population densities of cells at state 1 and 2, respec-tively. For simplicity, we consider a scenario where thereis a conservation in the densities, and ρ1 + ρ2 = 1 or,in other words, the total cell population remains stableduring the process. Under mean field assumptions, thedynamics of the relative densities would be defined by a
set of (symmetric) equations
dρ1dt
= µρ1
(1
2− ρ2
)− ρ1ϕ(ρ) (25)
dρ2dt
= µρ2
(1
2− ρ1
)− ρ2ϕ(ρ) (26)
The previous set (25) and (26) essentially tells us thatthe density of cells having a given internal state (either 1or 2) will grow at the expense of the other one only if thelater has a dominant concentration (i e. when ρj < 1/2),while it will decay otherwise. The first term on the righthand side of the two previous equations describes thisrule. Here, the term µ is a reaction rate determiningthe speed at which cells can shift from one state to theother. Now, assuming the conservation condition it is

11
not difficult to see that
ϕ(ρ) =∑i 6=j
µρi
(1
2− ρj
)and thus we have, from ρi = (1− ρj):
dρidt
= µρi
(ρi −
1
2
)− µρi
∑i 6=j
ρi
(1
2− ρj
)(27)
Equation (27) can be rewritten as the following cubicequation:
dρidt
= Γ(µ, ρi) = 2µ(1− ρi)(ρi −
1
2
)ρi (28)
which has three equilibrium points {ρ∗i |Γ(ρ∗i ) = 0} ={0, 1, 1/2}. The stability analysis of these points revealsthat ρ∗i = 0, 1 are stable whereas the coexistence one,ρ∗i = 1/2, is unstable.
This is an example of symmetry breaking and can beillustrated by calculating its associated potential functionΦ(ρ∗i ). In this case, we obtain for our problem a function:
Φ(ρi) = −µ(ρ4i − 2ρ3i + ρ2i
)(29)
that shows the expected two-well shape (restricted to0 ≤ ρi ≤ 1). As illustrated in figure 6, once an ini-tial fluctuation has favoured a given configuration overthe second, the amplification of the original fluctuationforces a collective decision.
How can we map this type of decision making processinto a synthetic biological circuit? The fundamental issuehere is to have a design that naturally incorporates twoalternative states in such a way that initial differencesget amplified by the nonlinear interactions. The basicprinciple is that we need a system involving two stablegenetic alternative states, each one acting exclusively andthus forbidding the alternative one. This can only beachieved if there is an interaction between the two actualgenes through negative feedback loops. In this scheme,two alternative states are possible because each releasedsignal inhibits the cell sensitivity to the other.
A simple solution is provided by the diagram shown infigure 7 (top diagram). Here, each cell has two differentbut equivalent signaling circuits able to sense both sig-naling molecules s1 and s2 via specific receptors. Signalactivated receptor promote the expression of three differ-ent genes, its own signal inducer SnI, that is the enzymethat synthesize the signal, its own receptor SnR and afluorescent protein, and the transcription of an antisenseRNA that specifically inhibit the opposite receptor bydegradation of the mRNA. Considering s1, explicit in-hibition of the receptor S2R brings the reduction of thesensitivity to s2 molecule (Carbonell et al. 2014) anda lower expression of its associated inhibitor. This alsoleads to a positive feedback on the sensitivity to the sig-nal s1, since the fall of S2R will lead to the weakening ofthe inhibition of S1R in a nonlinear manner.
The resulting dynamical behaviour of this system is il-lustrated in figure 7 using an agent-based model where aset of previously grown population of bacteria is used toperform the majority detection task. Here two input sig-nals are introduced near the centre of the colony. In thetwo different in silico experiments, one of the two signalsis slightly greater than the other. The genetic circuit istriggered while diffusion propagates the signals throughspace, and the dominant discloses as a fluorescent circleinvolving one of the two possible reporters. The two se-ries of snapshots shown in figure 7 illustrate the expectedcollective pattern. In one case, the initially more abun-dant signal triggers the expression of the green reporter(top snapshot series in figure 7) which spreads throughthe dish, whereas in the alternative scenario the balanceis just the oposite (bottom series in figure 7) and at theend of the simulation most cells already express only thered reporter.
The amplification phenomenon associated to our syn-thetic system has all the properties expected from thestatistical physics of symmetry breaking found in a broadrange of systems (Strocchi 2005; Sole 2011). It providesa clever illustration of the concept of symmetry breakingas a way of creating information (Haken 2010). In thiscontext, choosing between two different options needs asystem capable of amplifying fluctuations and exhibitingmultiple stable states.
V. SELF-REGULATION OF DIVISION OF LABOR
Among other fundamental features exhibited by sys-tems that display collective intelligence, division of la-bor is probably one of the most important in terms ofallowing efficient and robust decision making. Given arepertoire of tasks to be executed, several relevant fac-tors influence how efficiently are tasks performed. In antcolonies, specific tasks can be performed either by mor-phologically differentiated organisms (so that a relationexists between the phenotype and the task) or instead bythe same type of individuals, as it occurs in monomor-phic species. In the second case, some dynamical processmight be present in order to redistribute tasks amongindividuals as a function of the current demand. More-over, the lack of an external differential trait does notpreclude some internal feature, affecting potential be-havioural repertoires. An example is the presence of aresponse threshold that would affect the decision of eachindividual in relation with some task to be performed.
One example of caste-dependent task allocation isgiven by the dynamics of the ants of the genus Phei-dole, which exhibit a characteristic dimorphism into twoeasily distinguishable types of workers (namely, majorsand minors, which are shown in figure 8a, inset). Eachtype is associated to a preferential task: foraging or nestdefense. It has been shown (Detrain and Pasteels 1991,1992) that the probability that a given ant in any of thesegroups becomes involved in a given task is a nonlinear,

12
10-3 10-2 10-1 100 101 102
S
0.0
0.2
0.4
0.6
0.8
1.0
a
b
P(0
!T
k)
a b
+ +S
- -T1 T2
0 0
P [0 ! T2]P [0 ! T1]
↵
✓1 ✓2
C1 C2
⌘2⌘1
FIG. 8 Division of labor and its regulation. Here we illustrate this property with the two types of workers present in Pheidoleant colonies (a, inset) where individuals display a very different size (photograph by Alexander Wild). The probability of beingengaged in a given task is a nonlinear, Hill-like function, as indicated by the two curves. The curves are essentially identicalexcept for their shift associated to the presence of different thresholds. Given a specific task, the morphological class exhibitingthe lowest activation threshold is the one that preferentially works on it. The way castes and the stimulus S interact aresummarised in (b) using a simple block diagram. Namely, the stimulus (S) enhances the recruitment of the each caste (Ck) inthe aforementioned nonlinear way, with different thresholds of activation for each caste (θk). The recruited ants perform thetasks with a specific efficiency (ηk), lowering the levels of stimulus.
saturating function of the intensity of the stimulus. Thiswas shown experimentally by artificially altering the fre-quency of workers in different castes. Once a fraction ofworkers from one of the castes was removed, it was ob-served that, after some delay, individuals from the secondgroup became engaged in the task, successfully perform-ing it (Wilson 1984).
By decreasing the fraction of minors in Pheidolecolonies, it was possible to observe that the number ofmajors performing the required task remained low un-til a critical stimulus intensity is reached, followed by arapid increase in the fraction of majors involved. In otherwords, once the balance between castes is altered enough,the caste with higher threshold (which normally does notperform the required function) rapidly restores the ade-quate levels of task performance. This is a simple andinteresting scenario where we can see at work the robustresponse of a given system to changing conditions. Inthis section we aim to provide an equivalent engineereddesign of this response using modified cells.
The probability of switching towards a given task canbe described by a Hill-like response function using a fixedthreshold model (FTM). If we indicate by σk the stateof a given worker from the caste k, it moves from inac-tive (σ = 0) to active (indicated as σ = T ) with someprobability that depends on the intensity of the stimulusS. Once in the active state, it starts performing theirpossible tasks. Although different tasks Tk could be con-sidered, here we deal with the case of recruiting differentcastes for a specific task T. Here we will use a probabilityof transition described by a sigmoidal curve, namely:
P (0→ T ) =S2
θ2k + S2(30)
In figure 8a we show an example of the two transitionprobability curves. Since two different thresholds arepresent, the two curves appear displaced along the x-axis.
A simple model of task allocation considers two possi-ble types of workers that interact with S through a simplefeedback loop. In figure 8b we summarise the basic as-sumptions made in (Theraulaz et al 1998, Bonabeau etal 1996). We consider two castes indicated by C1 and C2
that are optimal for different tasks but, if necessary, bothcan perform the same task T. Specifically, individuals ofeach morphological class have a state σk ∈ {0, T} indicat-ing no activity and activity, respectively. The stimulus Sis constantly raised at a rate α and is also ”removed” at arate δ that depends on the efficiency at which the task Tis performed. The presence of this stimulus triggers thetransition from inactivity to activity, following the tran-sition probability given by equation (30). As summarisedin this diagram, the whole system operates under a neg-ative feedback control scheme with two regulatory com-ponents displaying different activation thresholds. Thismodel and variants of it were shown to account for thenonlinear response of different castes to the intensity ofthe stimulus
The caste-dependent task allocation scheme, as definedby the fixed threshold model, can be engineered follow-ing the design of the synthetic constructs presented infigure 9a-b. In these synthetic implementation, the twoant castes are substituted by similarly modified cells us-ing manipulated quorum sensing genes. Specifically, therecruiting factor for a single task is replaced by the quo-rum sensing signalling molecule HSL which is externallyintroduced in the system. These cells respond under aHill-like transfer function to the signal, albeit with differ-ent thresholds of activation and reaching different levels
13
a
c
b
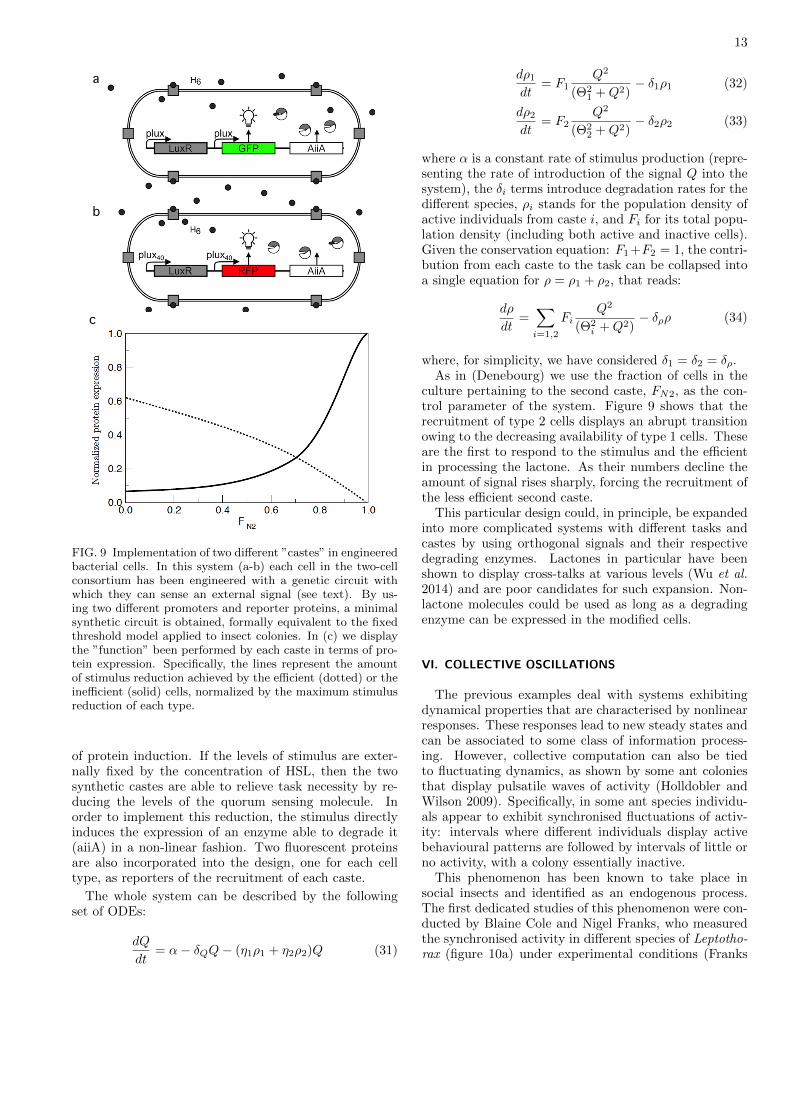
FIG. 9 Implementation of two different ”castes” in engineeredbacterial cells. In this system (a-b) each cell in the two-cellconsortium has been engineered with a genetic circuit withwhich they can sense an external signal (see text). By us-ing two different promoters and reporter proteins, a minimalsynthetic circuit is obtained, formally equivalent to the fixedthreshold model applied to insect colonies. In (c) we displaythe ”function” been performed by each caste in terms of pro-tein expression. Specifically, the lines represent the amountof stimulus reduction achieved by the efficient (dotted) or theinefficient (solid) cells, normalized by the maximum stimulusreduction of each type.
of protein induction. If the levels of stimulus are exter-nally fixed by the concentration of HSL, then the twosynthetic castes are able to relieve task necessity by re-ducing the levels of the quorum sensing molecule. Inorder to implement this reduction, the stimulus directlyinduces the expression of an enzyme able to degrade it(aiiA) in a non-linear fashion. Two fluorescent proteinsare also incorporated into the design, one for each celltype, as reporters of the recruitment of each caste.
The whole system can be described by the followingset of ODEs:
dQ
dt= α− δQQ− (η1ρ1 + η2ρ2)Q (31)
dρ1dt
= F1Q2
(Θ21 +Q2)
− δ1ρ1 (32)
dρ2dt
= F2Q2
(Θ22 +Q2)
− δ2ρ2 (33)
where α is a constant rate of stimulus production (repre-senting the rate of introduction of the signal Q into thesystem), the δi terms introduce degradation rates for thedifferent species, ρi stands for the population density ofactive individuals from caste i, and Fi for its total popu-lation density (including both active and inactive cells).Given the conservation equation: F1+F2 = 1, the contri-bution from each caste to the task can be collapsed intoa single equation for ρ = ρ1 + ρ2, that reads:
dρ
dt=∑i=1,2
FiQ2
(Θ2i +Q2)
− δρρ (34)
where, for simplicity, we have considered δ1 = δ2 = δρ.As in (Denebourg) we use the fraction of cells in the
culture pertaining to the second caste, FN2, as the con-trol parameter of the system. Figure 9 shows that therecruitment of type 2 cells displays an abrupt transitionowing to the decreasing availability of type 1 cells. Theseare the first to respond to the stimulus and the efficientin processing the lactone. As their numbers decline theamount of signal rises sharply, forcing the recruitment ofthe less efficient second caste.
This particular design could, in principle, be expandedinto more complicated systems with different tasks andcastes by using orthogonal signals and their respectivedegrading enzymes. Lactones in particular have beenshown to display cross-talks at various levels (Wu et al.2014) and are poor candidates for such expansion. Non-lactone molecules could be used as long as a degradingenzyme can be expressed in the modified cells.
VI. COLLECTIVE OSCILLATIONS
The previous examples deal with systems exhibitingdynamical properties that are characterised by nonlinearresponses. These responses lead to new steady states andcan be associated to some class of information process-ing. However, collective computation can also be tiedto fluctuating dynamics, as shown by some ant coloniesthat display pulsatile waves of activity (Holldobler andWilson 2009). Specifically, in some ant species individu-als appear to exhibit synchronised fluctuations of activ-ity: intervals where different individuals display activebehavioural patterns are followed by intervals of little orno activity, with a colony essentially inactive.
This phenomenon has been known to take place insocial insects and identified as an endogenous process.The first dedicated studies of this phenomenon were con-ducted by Blaine Cole and Nigel Franks, who measuredthe synchronised activity in different species of Leptotho-rax (figure 10a) under experimental conditions (Franks

14
0 50 100 150 200 250 300 350time (min)
0.0
0.5
1.0
a
b
FIG. 10 (a) Some ant colonies, including some species of thegenus Leptothorax shown here are known to display coher-ent bursts of activity (picture by Bernhard Seifert). In (b)we show an example of the synchronised activity waves asrecorded from an experiment. The time series is a surrogateof the number of active individuals and despite its noisy ap-pearance, it has a marked periodicity (Sole et al 1991).
and Bryant 1987, Franks et al 1990, Cole 1991a, 1991b).In these colonies, it was found that a pulsatile, burstypattern was involved, as shown in figure 10b from arecorded experiment where the changes of pixels in con-secutive moments of time, giving a surrogate of the den-sity of active individuals moving around. These fluctu-ations have a ∼ 25 minute period. Using single ants, ithas been observed that individuals do not display thisregular pattern. Instead, they seem to be chaotic (Cole1991b) and thus this is not an example of a set of indi-vidual oscillators that get entrained due to some class ofcoupling among individuals (Strogatz 2003).
In these colonies, single individuals are known to be-come spontaneously active inside an inactive colony. Asthey move around, they contact other resting individualsand can make them awake. By increasing the density ofindividuals within a given arena, it is possible to studythe influence of the number of ants on the degree of syn-chronisation and spread of activity. It was found that adisordered burstiness is replaced by a more coherent col-lective activity pattern as density increases (Cole 1991a),thus indicating that the likelihood of finding active indi-viduals grows as more interactions are allowed to happen.In fact, if the density of individuals is forced beyondthe one found in non-manipulated colonies, oscillationsalso become more coherent, more than observed. Thisseems to indicate something very interesting, namely thatcolonies might self-regulate their density in such a waythat a special, intermediate density is achieved.
According to Robinson (1992), self-synchronization fa-cilitates the sampling of any information an individ-
ual may need from other individuals. Assuming thatants cannot be active all the time, why would self-synchronized behaviour be a better (simpler) strategythan, say, random (in the sense of nonsynchronized) ac-tivity patterns? We will need further assumptions toanswer this question: the obvious one of locality (an in-dividual is able to get only local information) and sec-ond, the quite reasonable (and biologically plausible, seeabove) assumption that the unique interaction allowedto an inactive individual is to be ”awaked” by other(s)individual(s); an inactive individual does not carry anyinformation, namely, it is equivalent to a ”sleeping” in-dividual. Now, in this context, it would be clear why weshould obtain an increase of efficiency with synchronizedpatterns of activity: it would maximize the number ofsimultaneously active neighbours of an active individual.A detailed study of the two alternatives (Delgado andSole, 2000) shows that a fluctuating system actually out-performs the alternative (not observed) constant-activityscenario.
The simplest mean field model that can be formu-lated is closely related to the propagation of epidemics(Cole 1991c, Sole 2011). Let us now consider our pop-ulation composed by two groups, namely active and in-active ants, whose numbers will be indicated by A andI, respectively. Using the normalized values x = A/Nand y = I/N , and assuming that the density of occupiedspace is ρ ∈ [0, 1], the mean field equation for the fractionof active ants is:
dx
dt= αx(ρ− x)− γx (35)
An important difference in relation to epidemic spread-ing is that the density of ants ρ is now a key controlparameter.
The equation displays two equilibrium points. Thefirst corresponds to no-activity at all x∗0 = 0 while thesecond is associated to the presence of a given level ofactivity, namely
x∗1 = ρ− γ
α(36)
The colony will display non-zero (i. e. x∗1 will be stable)activity provided that λµ(0) = αρ − γ > 0 which, interms of the density of ants corresponds to the conditionρ > ρc = γ/α.
The deterministic model predicts that no activity willbe present at subcritical densities, whereas experimentsindicate that activity is present, largely due to randomindividual activations, at small densities. Moreover, ithas been found that fluctuations become very large closeto the critical point, consistently with real data (Sole etal., 1993; Miramontes, 1995). This is illustrated by theshape of the associated potential, which now reads:
Φρ(x) = αx3
3− (αρ− γ)
x2
2(37)
Three examples of this potential are shown in figure 5cfor a subcritical,near-critical and supercritical cases. As

15
-0.5 0 0.5 1 1.5 2-0.5 0 0.5 1 1.5 2
0
2
4
6
-0.5 0 0.5 1 1.5 2-0.5 0 0.5 1 1.5 2-0.5
0
0.5
-0.5 0 0.5 1 1.5 2-0.5 0 0.5 1 1.5 2
-0.20
0.20.40.60.81
0 1 2 3 4 5 60 1 2 3 4 5 6
-4
-2
0
2
a b
c d
�(x
)
x x
�(x
)
FIG. 11 Four snapshots of the potential function associatedto the mean field model of ant colony synchronisation. Thepotentials have been calculated using increasing values of theparameter ρ using fixed values of the ant density parameter.Here we have α = γ = 1 and (a) ρ = 0.5, (a) ρ = ρc = 1.0, (a)ρ = 1.5 and (a) ρ = 3.0. The gray left areas indicate negative(and the unavailable) conditions.
expected, the potential becomes flatter as criticality isapproached.
The observed variance in colony fluctuations is a con-sequence of critical slowing down (see chapter 1) and forthis model it can be shown that the relaxation time τ(ρ),scales as
T (ρ) =1
α
∫ x(T (ρ))
x(0)
dx
x(ρ− ρc − x)∼ (ρ− ρc)−1 (38)
As we approach criticality, random activations are likelyto propagate for a while before global activation dies out,thus providing the source for higher activity levels andbroader fluctuations.
Although some previous studies have shown how toimplement a synthetic system capable of displaying peri-odic dynamics, all of them where based on the presenceof oscillators capable of synchronize leading to a collec-tive oscillation (Danino et al. 2010). This is the classicalproblem of entrainment oscillators through some kind ofcoupling. Here the lack of an individual-level oscillatorybehaviour requires a different approximation.
A possible way to engineer bacteria in such a way thatit behaves as an excitable element is by the implemen-tation of a motif able to release a long distance activa-tion signal, like HSL, and a inner cell inhibitory mecha-nism, like tagged protein degradation by ClpXP [figure11(a)]. The activator signal is HSL. HSL promote theexpression of its own biosynthetic enzyme LuxI and alsothe expression of a proteolytic protein, ClpX. The in-hibitory effect is implemented by means of ClpXP targetsequences in LuxR (the HSL receptor), LuxI (the HSLinducer) and the fluorescent protein GFP . Thereby, theequilibrium rate between leakiness of the Plux promoter
upstream of LuxI (activator) and ClpX (inhibitor), keepthe systems wondering around a low number of proteinsand molecules dynamic equilibrium, where the stochas-ticity is high (Raser and OShea 2005).
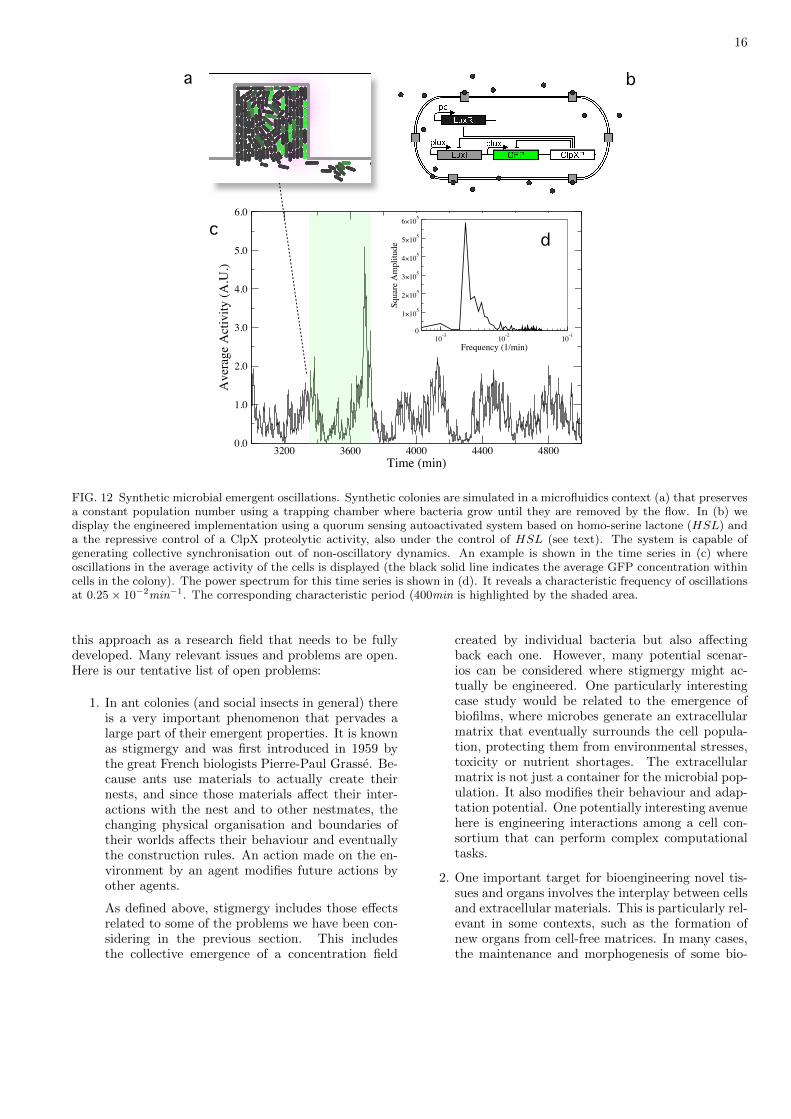
When one of those cell overcome the activation thresh-old, it will start to synthesize actively LuxI enzyme. Thiswill lead to the production and release of HSL, that willdiffuse in the media, activating the surrounding cells in apositive loop. However, HSL also promotes the expres-sion of ClpX protein, that will facilitate the degradationof LuxR, LuxI and GFP . This will drive the system tothe initial state of relaxation. As ClpXP has not a ClpXtarget sequence, it will persist more time in the cell giv-ing us a refractory period that each cell has to overcomebefore to the next activation loop.
An example of emergent collective oscillations in syn-thetic colonies is shown in figure 11(b). Here, eachcell in the colony carries the genetic construct describedabove. When a single cell is stochastically activated,it can activate its surrounding neighbours by releasingthe HSL signal, possibly generating a cascade of activ-ity that propagates in the space. Otherwise, the colonyremains at low activity levels. Such dynamics can betraced by plotting average GFP concentration displayedby cells in the colony [figure 11(b)]. Applying standardFast Fourier Transform techniques reveals a maximumsquare amplitude peak in figure (c), indicating that thesystem exhibits a characteristic frequency of oscillation.The shaded area in figure 11(b) shows the correspondingcharacteristic period of 400min and the power spectrumin figure 11(
A Fast Fourier Transform of this time series is shownin (c), where a peak is observed at 0.25 ? 10?2 min?1, re-vealing characteristic frequency of collective oscillations.
VII. DISCUSSION AND CONCLUSIONS
The widespread ecological success of insects resultingfrom the rise of swarm intelligence is undeniable. Sucha success required something more than simply get to-gether and evolved into a vast array of social structures.Ants and termites both display outstanding works as ar-chitects, but the blueprints for their buildings are notin their small brains. A termite nest has a scale that isseveral orders of magnitude larger than the size of theindividual workers. And these large scale structures pos-sess a rather complex and optimised structure. They re-sult from emergent patterns of construction out of simplerules. Similarly, the behavioural flexibility exhibited bytheir colonies and their potential for adapting to environ-mental uncertainty is equally remarkable. As a form ofintelligence, swarm intelligence has inspired generationsof computer scientists, theoreticians and robot designersalike.
In this paper we considered a novel approach to theproblem based on actually constructing living robots byusing genetic engineered microorganisms. We propose
16
3200 3600 4000 4400 4800Time (min)
0.0
1.0
2.0
3.0
4.0
5.0
6.0A
vera
ge A
ctiv
ity (A
.U.)
10-3 10-2 10-1
Frequency (1/min)
0
1×105
2×105
3×105
4×105
5×105
6×105
Squa
re A
mpl
itude
a b
c d
FIG. 12 Synthetic microbial emergent oscillations. Synthetic colonies are simulated in a microfluidics context (a) that preservesa constant population number using a trapping chamber where bacteria grow until they are removed by the flow. In (b) wedisplay the engineered implementation using a quorum sensing autoactivated system based on homo-serine lactone (HSL) anda the repressive control of a ClpX proteolytic activity, also under the control of HSL (see text). The system is capable ofgenerating collective synchronisation out of non-oscillatory dynamics. An example is shown in the time series in (c) whereoscillations in the average activity of the cells is displayed (the black solid line indicates the average GFP concentration withincells in the colony). The power spectrum for this time series is shown in (d). It reveals a characteristic frequency of oscillationsat 0.25 × 10−2min−1. The corresponding characteristic period (400min is highlighted by the shaded area.
this approach as a research field that needs to be fullydeveloped. Many relevant issues and problems are open.Here is our tentative list of open problems:
1. In ant colonies (and social insects in general) thereis a very important phenomenon that pervades alarge part of their emergent properties. It is knownas stigmergy and was first introduced in 1959 bythe great French biologists Pierre-Paul Grasse. Be-cause ants use materials to actually create theirnests, and since those materials affect their inter-actions with the nest and to other nestmates, thechanging physical organisation and boundaries oftheir worlds affects their behaviour and eventuallythe construction rules. An action made on the en-vironment by an agent modifies future actions byother agents.
As defined above, stigmergy includes those effectsrelated to some of the problems we have been con-sidering in the previous section. This includesthe collective emergence of a concentration field
created by individual bacteria but also affectingback each one. However, many potential scenar-ios can be considered where stigmergy might ac-tually be engineered. One particularly interestingcase study would be related to the emergence ofbiofilms, where microbes generate an extracellularmatrix that eventually surrounds the cell popula-tion, protecting them from environmental stresses,toxicity or nutrient shortages. The extracellularmatrix is not just a container for the microbial pop-ulation. It also modifies their behaviour and adap-tation potential. One potentially interesting avenuehere is engineering interactions among a cell con-sortium that can perform complex computationaltasks.
2. One important target for bioengineering novel tis-sues and organs involves the interplay between cellsand extracellular materials. This is particularly rel-evant in some contexts, such as the formation ofnew organs from cell-free matrices. In many cases,the maintenance and morphogenesis of some bio-

17
materials (such as bones) results from a a dynami-cal process of creation and destruction maintainedby interacting populations of cells specialised ineach part of the process. In social insects, this feed-back between individuals and the structures theybuild is known as stigmergy (Theraulaz et al).
3. Smart swarms capable of memory and informationprocessing could be useful in making complex deci-sions related to a number of biomedical issues wherethere is a need of detecting and targeting specificcells. Cancer is one example of this class. Ongo-ing work in this area has shown that engineeredmicrobes can be used to target tumor cells thanksto a combination of a special environment associ-ated to most solid tumours (i e. an acidic envi-ronment) that is suitable to microbes and a cy-tolytic mechanism that is triggered once a givendensity of microbial cells is reached. Here too aquorum sensing mechanism plays a specially rel-evant role. Further developments of this schemecould involve exploiting collective intelligence andcomplex decision-making designs.
4. Insect colonies, as discussed above, have been of-ten compared to a neural network. As it happenswith neural systems, individual agents inside an antcolony can exchange bits of information with theirnest mates through a range of chemical signals. Inour models and examples above we have assumed asingle class of signal as the vehicle of exchange butthere is a wide variety of QS messenger molecules(figure 13) and thus a potentially wide range of in-teractions (Williams et al 2014). This is somethingalso in common with ant colonies: multiple chem-ical signals appear to be associated to exchangesbetween nest mates. A theoretical framework thatcan take into account the capacity of a SCI systeminvolving multiple signals is much needed.
5. One potential scenario where SCI can play a rele-vant role involves the use of synthetic biology forecosystem repair (Sole 2015; Sole et al 2015). It hasbeen recently suggested that engineered microor-ganisms and their ecological interactions could beengineered to prevent ecosystem degradation andcollapse or even help in reducing the impact ofclimate change. The proposed engineering motifswere based in a simple set of interactions defined,for example, in terms of mutualistic links. Morecomplex scenarios could be considered here, takingadvantage of the views of SCI. Ecosystem bioengi-neering could be designed under more flexible con-ditions, allowing decision-making processes to takeplace in such a way that the engineered consortiacould adjust over time to different environmentalconditions.
As a final point, we can also ask ourselves to what ex-tent the previous examples are instances of how to map
FIG. 13 Structures of some representative quorum sens-ing signalling molecules. Here we display just seven exam-ples from the large set of QS candidate signalling molecules.Adapted from Williams et al (2014).
observed classes of swarm behaviour into an equivalentmicrobial counterpart. This is a correct statement atleast in some of the examples presented above, at leastunder the assumption that we have been searching forrather general, perhaps universal sets of rules that per-vade the nonlinearity of collective intelligence. But thereis nothing preventing us from moving beyond the bound-aries of the observed universe of actual forms of swarmintelligence.
Are there other, different potential ways of engineeringintelligent behaviour using synthetic biology? By differ-ent this means designs that lead to qualitatively differ-ent classes of complex decision-making departing frominsect colonies. This is a very important question, sincea negative answer could support the view that observedforms of a superorganism are attractors in the landscapeof the possible: nothing essentially different exists be-yond what evolutionary dynamics has been able to findand strong constraints operate limiting the repertoire ofpossible behaviours (Alberch 1989). But may be thereare other ways of building swarms through geneticallyengineering living entities in ways that depart (partiallyor completely) from the observable natural world. If thatwere the case, we could have evidence for alternative evo-lutionary scenarios for collective intelligence could havebeen possible.
Acknowledgments
We would like to thank Javier Macia and CarlosRodriguez-Caso for useful discussions on emergence inmicrobial and synthetic systems. This work was sup-

18
ported by an ERC Advanced Grant Number 294294from the EU seventh framework program (SYNCOM),by grants of the Botin Foundation, by Banco Santanderthrough its Santander Universities Global Division andby the Santa Fe Institute, where most of this work wasdone.
VIII. REFERENCES
1. Alberch P. 1989. The logic of monsters: evidencefor internal constraint in development and evolu-tion. Geobios. 22, 21-57
2. Anderson, P. W. 1972. More is different. Science177, 393-396.
3. Alberts, B. Johnson, A. Lewis, J. Raff, M. Roberts,K. and Walter, P. (2002) Molecular Biology of theCell, 4th edition. New York: Garland Science.
4. Camazine S, Deneubourg J-L, Franks N.R., SneydJ, Theraulaz G and Boabeau E 2003. Self-Organization in Biological Systems. Princeton UPress, Princeton.
5. Beckers, R., Deneubourg, J. L. and Goss, S. 1993.Modulation of trail laying in the ant Lasius niger(Hymenoptera: Formicidae) and its role in the col-lective selection of a food source. Journal of InsectBehavior 6, 751-759.
6. Beekman, M., Sumpter, D. J. T. and Ratnieks, F.L. W. 2001. Phase transition between disorderedand ordered foraging in Pharaoh's ants. Proc. NatlAcad. Sci. 98, 9703-9706.
7. Ben-Jacob, E. 2003. Bacterial Self-Organization:Co-Enhancement of Complexification and Adapt-ability in a Dynamic Environment. Phil. Trans.R. Soc. Lond. A, 361, 1283-1312.
8. Ben-Jacob, E., Becker, I., Shapiro, Y. 2004. Bac-terial Linguistic Communication and Social Intelli-gence. Trends Microbiol 12, 366-372.
9. Boi, S., Couzin, I. D., Del Buono, N., Franks, N.R. and Britton, N.F. 1998. Coupled oscillators andactivity waves in ant colonies. Proc. R. Soc. LondB 266, 371-378.
10. Bonabeau, E., Theraulaz, G. and Deneubourg, J.L.1996. Mathematical model of self-organizing hier-archies in animal societies. Bulletin of mathemati-cal biology. 58, 661-717.
11. Bonabeau, E., Theraulaz, G. Deneubourg, J.-L.1996. Quantitative study of the fixed thresholdmodel for the regulation of division of labour ininsect societies. Proc. Roy. Soc. London B 263,1565ı¿ 1
21569.
12. Bonabeau, E., Sobkowski, A., Theraulaz, G. andDeneubourg, J.L. 1997. Adaptive Task AllocationInspired by a Model of Division of Labor in SocialInsects. BCE Conference 36-45.
13. Bonabeau, E., Dorigo, M. and Theraulaz G. (1999)Swarm Intelligence: From Natural to Artificial Sys-tems. Oxford University Press.
14. Cole, B.J. 1991a. Short-term activity cycles inants: generation of periodicity by worker interac-tion. The American Naturalist 137, 244-259.
15. Cole, B.J. 1991b. Is animal behaviour chaotic? ev-idence from the activity of ants. Proc Roy Soc Lon-don B 244, 253-259.
16. Cole, B.J. 1991c. Short-term activity cycles inants: a phase response curve and phase resettingin worker activity. The Journal of Insect Behavior4, 129-137.
17. Danino, T. Mondragon-Palomino, O. Tsimring, L.Hasty, J. 2010. A synchronized quorum of geneticclocks. Nature. 463, 326-30
18. Davies, D.G. , Parsek, M.R., Pearson, J.P.,Iglewski, B.H., Costerton, J.W., Greenberg, E.P.1998. The involvement of cell-to-cell signals in thedevelopment of a bacterial biofilm. Science 280,295-298.
19. Delgado, J. and Sole, R. V. 1997. Noise InducedTransitions in Fluid Neural Networks. Phys LettA 229, 183-189.
20. Delgado, J. and Sole, R. V. 2000. Self-synchronization and Task Fulfilment in AntColonies. J Theor Biol 205, 433-441.
21. Deneubourg, JL., Pasteels, J.M. and Verhaeghe, J.C. 1983. Probabilistic behaviour in ants: a strategyof errors? J Theor Biol 105, 259-271.
22. Deneubourg, J. L. and Goss, S. 1989. Collectivepatterns and decision-making. Ethology, Ecologyand Evolution, 1, 295-311.
23. Deneubourg, J.L., Goss, S., Franks, N. and Pas-teels, J. M. 1989. The blind leading the blind: mod-eling chemically mediated army ant raid patterns.J Insect Behav 2, 719-725.
24. Detrain C. and Deneubourg JL. 2006. Self-organised structures in a superorganism: do antsbehave like molecules? Physics Life Revs 3, 162-187.
25. Franks, N. R. and S. Bryant. 1987. Rhythmicalpatterns of activity within the nests of ants. In:J. Eder and A. Rembold, Editors, Chemistry andbiology of social insects. pp 122-123. Springer

19
26. Garnier, S., Gautrais, J. and Theraulaz, G. 2007.The biological principles of swarm intelligence.Swarm Intell 1, 3-31.
27. Gordon, D. 1999. Ants at work. Free Press, NewYork.
28. Goss, S. and Deneubourg, J. L. 1988. Autocatalysisas a source of synchronised rhythmical activity insocial insects. Insectes Sociaux, 35, 310-315.
29. Goodwin, B. C. 1994. How the leopard changed itsspots. The evolution of complexity. Weidenfeld andNicolson, London UK.
30. Freeman, M. 2000. Feedback control of intercellularsignalling in development. Nature 408, 313-319.
31. Haken H. 2010. Information and self-organisation.Springer, Berlin.
32. Hofstadter DR. 1979. Godel, Escher, Back. BasicBooks, New York.
33. Holldobler, B. and Wilson, E. O. 2009. The Super-organism: The Beauty, Elegance and Strangenessof Insect Societies. Norton, New York.
34. Hopfield, J. J. 1994. Physics, computation and whybiology looks so different. J. Theor. Biol. 171, 53-60.
35. Huberman, B. and Hogg, T. 1987. Phase Transi-tions in Artificial Intelligence Systems. Artif. Intel.33, 155-171.
36. Jablonka E and Lamb MJ. 2006. The evolution ofinformation in the major transitions. J. Theor Biol239, 236-246.
37. Jang, S.S., Oishi, K.T., Egbert, R.G. and Klavins,E. 2012. Specification and simulation of multicelledbehaviors. ACS Synthetic Biology. 1(8), 365-374.
38. Jones, S., Yu, B., Bainton, N.J., Birdsall, M., By-croft, B.W., Chhabra, S.R., Cox, A.J., Golby, P.,Reeves, P.J., Stephens, S. 1993. The lux autoin-ducer regulates the production of exoenzyme viru-lence determinants in Erwinia carotovora and Pseu-domonas aeruginosa. EMBO J. 12(6), 2477-2482.
39. Lambert, G. and Kussell, E. 2014. Memory andFitness Optimization of Bacteria under FluctuatingEnvironments. PLoS Genet 10(9): e1004556.
40. Legg, S. and Hutter, M. (2006) A formal Measureof Machine Intelligence. In Proc. 15th AnnualMachine Learning Conference of Belgium and TheNetherlands. pg 73-80.
41. Millonas, M. 1993. Swarms, phase transitions, andcollective intelligence. In: Artificial Life III. C.Langton, editor. Addison-Wesley. Reading, MA.
42. Miramontes, O. 1995. Order-disorder transitions inthe behavior of ant societies. Complexity 1, 56-60.
43. Oster, G. F., and E. O. Wilson. 1978. Caste andEcology in the Social Insects. Princeton UniversityPress. Princeton.
44. Pribyl, M., Muratov, C.B. and Shvartsman, S.Y.2003. Long-range signal transmission in autocrinerelays. Biophys. J. 84, 883-896.
45. Raser, J.M. and O'Shea, E.K. 2005. Noise in GeneExpression: Origins, Consequences, and Control.Science 309, 2010-2013.
46. Rose S. 2006. The future of the brain. Oxford UPress, Oxford UK.
47. Sole, R.V., Miramontes, O. and Goodwin, B. C.1993. Oscillations and Chaos in Ant Societies. JTheor Biol 161, 343-357.
48. Sole, R.V., Manrubia, S. C., Luque, B., Delgado,J. and Bascompte, J. 1996. Phase transitions andcomplex systems. Complexity 1: 13-26.
49. Sole, R.V., Manrubia, S. C., Kauffman, S. A., Ben-ton, M. and Bak, P., 1999. Criticality and Scal-ing in Evolutionary Ecology, Trends in Ecology andEvolution 14: 156-160.
50. Sole, R.V. and Goodwin, B. C. 2001. Signs of Life:How Complexity Pervades Biology. Basic Books:New York.
51. Sole, R.V. 2011. Phase Transitions. Princeton U.Press, Princeton.
52. Sternberg, R. J. (200) Handbook of Intelligence.Cambrige university Press.
53. Strocchi, F. 2005. Symmetry breaking. Springer,Berlin.
54. Strogatz, S. 1994. Nonlinear Dynamics and Chaos.Addison Wesley, New York.
55. Strogatz, S. 2003. SYNC. The emerging science ofspontaneous order. Hyperion Books, New York.
56. Sumpter, D. 2006. The principles of collective an-imal behaviour. Phil. Trans. R. Soc. London, B361, 5-22.
57. Sutton, R. and Barto, A. .1998. ReinforcementLearning: An introduction. MIT Press, Cambridge,MA.
58. Theraulaz, G., Gautrais, J., Camazine, S. andDeneubourg , J.L. 2003. The formation of spatialpatterns in social insects: from simple behavioursto complex structures. Phil Trans Roy Soc A 361,1263-1282.

20
59. Theraulaz, G., Bonabeau, E. Deneubourg, J.-L.1998. Response threshold reinforcement and divi-sion of labour in insect societies. Proc. Roy. Soc.London B 265, 327ı¿ 1
2332.
60. Vladimirov, N. Sourjik, V. 2009. Chemotaxis: howbacteria use memory. Biol Chem. 390(11):1097-104.
61. Valverde, S., Corominas-Murtra, B., Perna, A.,Kuntz, P., Theraulaz, G. and Sole, R. V. 2009.Percolation in insect nest networks: evidence foroptimal wiring. Phys. Rev . E 79, 066106.
62. Vicsek, T. ed. 2001. Fluctuations and Scaling inBiology. Oxford U. Press, Oxford.
63. Waters CM. and Bassler BL. 2005. Quorum sens-ing: cell-to-cell communication in bacteria. Annu.Rev. Cell. Dev. Biol. 21, 319-46.
64. Wiley, H.S., Shvartsman, S.Y. and Lauffenburger,D. 2003. Computational modeling of the EGF-
receptor systems: a paradigm for systems biology.Trends Cell Biol. 13, 43-50.
65. Williams P, Winzer K, Chan WC amd Camara M(2014) Look whoı¿ 1
2 s talking: communication andquorum sensing in the bacterial world. Phil TransR Soc B 362, 1119-1134.
66. Wilson, E.O. 1971. The Insect Societies. HarvardUniv. Press. Cambridge, MA.
67. Wilson, E. 0. 1984 The relation between caste ra-tios and division of labour in the ant genus Pheidole(Hymenoptera: Formicidae). Behav.E col. Socio-biol.1 6, 89-98.
68. Wu, F. Menn, DJ. and Wang, X. 2014. Quorum-sensing crosstalk-driven synthetic circuits: fromunimodality to trimodality. Chem. Biol.21(12):1629-38.