Embed Size (px)
Citation preview

ORIGINAL PAPER
Symbiosis of photosynthetic microorganisms with non-photosynthetic ones for the conversion of cellulosic massinto electrical energy and pigments
R. Navanietha Krishnaraj • Sheela Berchmans •
Parimal Pal
Received: 12 March 2014 / Accepted: 31 May 2014
� Springer Science+Business Media Dordrecht 2014
Abstract In this article, we report a three-compart-
ment microbial fuel cell (MFC) system for the
simultaneous degradation of cellulose and production
of natural pigments such as phycoerythrin and phyco-
cyanin along with bioelectricity generation. Oscilla-
toria annae, a freshwater cyanobacterium, was used
for the conversion of cellulose to reducing sugars,
which were fed as a substrate to a coculture of
Acetobacter aceti and Gluconobacter roseus for
current generation in a three-compartment MFC.
Carbon felt modified with a composite film containing
chitosan and sodium alginate served as the MFC
anode. The cellulose-fed three-compartment MFC
produced a maximum power output of 6.62 W m-3 at
17.55 A m-3.
Keywords Cellulose � Pigments � Bioelectricity �Microbial fuel cell
Introduction
Microbial fuel cells (MFCs) represent green tech-
nology bioelectrochemical systems in which the
electrigens oxidize the substrate in anaerobic condi-
tions and transfer the electrons to the anode (Logan
2009). MFCs have several operational and functional
advantages. However, the low power output and cost
factor remain its weaknesses (Dewan et al. 2010).
MFCs are likely to become viable if the economics
of running a sustainable fuel cell can be sorted out.
Further synthesis of novel chemical products along
with electricity production would be advantageous.
The use of a cheap and abundant biomass may help
to reduce the operational cost of the MFCs (Logan
and Rabaey 2012). Cellulosic biomass seems to be a
cheap and attractive carbon-neutral renewable
resource for use as a substrate in MFCs because of
its abundance. Use of cellulosic wastes in MFCs
could help to solve the current global energy crisis
and remediate the huge volume of cellulosic waste
(Rezaei et al. 2009).
The use of cellulosic biomass in any bioprocess
operation is mainly hindered by the recalcitrant
nature of cellulose, which makes the use of these
materials as substrates directly in MFC a challenge.
Hence, pretreatment is necessary for breaking the
structural and chemical complexity of the biomass
(Mosier et al. 2005). Cellulose can be hydrolytically
broken down into glucose by biological, physical or
chemical methods. Physical methods such as ball
Electronic supplementary material The online version ofthis article (doi:10.1007/s10570-014-0319-y) contains supple-mentary material, which is available to authorized users.
R. Navanietha Krishnaraj � S. Berchmans (&)
CSIR-Central Electrochemical Research Institute,
Karaikudi 630006, Tamil Nadu, India
e-mail: [email protected]
P. Pal
National Institute of Technology,
Durgapur 713209, West Bengal, India
123
Cellulose
DOI 10.1007/s10570-014-0319-y

milling and thermal methods are not suitable for real-
time applications. Chemical methods make use of
strong acids and solvents for the hydrolysis of
cellulose. Problems such as acid recovery, the
complicated recycling process and cost factors make
these methods unsuitable for commercial applica-
tions. If the acid is volatile, vacuum stripping
methods are also need to be practiced (Hendriks
and Zeeman 2009; Ladisch et al. 1978). Enzymatic
hydrolysis has several disadvantages, such as the
narrow temperature and pH range of the enzymes,
high costs, slow hydrolysis rate and need for
sophisticated facilities and huge reactors (Orozco
et al. 2007). Microbial hydrolysis offers many
advantages over other methods because of the higher
yields, low energy requirements, mild operating
conditions and ecofriendly nature. Certain species
of cyanobacteria possess excellent lignolytic and
cellulolytic activities. Metagenomic analysis of the
wood-decomposing microbial community revealed
the presence of a few cyanobacterial species in
association with other cellulolytic and lignolytic
organisms (Van der Lelie et al. 2012). As these
organisms have simple growth requirements, cultur-
ing the cyanobacteria in large amounts is less
expensive and can be used effectively for degrading
celluloses (Kumar Saha et al. 2003; Gupta et al.
2011). Acetic acid bacteria are ideal options for
bioelectricity generation because of their good oxi-
dizing ability for a wide range of substrates (Navan-
ietha Krishnaraj et al. 2013; Karthikeyan et al. 2009;
Navanietha Krishnaraj et al. 2014). The metal-
reducing characteristics of Gluconobacter roseus
have also been documented in the literature (Navan-
ietha Krishnaraj and Sheela 2013b). One report in the
literature describes the use of Enterobacter cloacae
for current generation along with degradation of
cellulose (Rezaei et al. 2009; Ren et al. 2007).
In this work, cellulolytic activity is provided by
cyanobacteria, and electrogenic activity is pro-
vided by the coculture of G. roseus and Aceto-
bacter aceti. Herein, we have also demonstrated
for the first time the symbiotic effect of a
photosynthetic organism and a coculture of non-
photosynthetic organisms for the effective degra-
dation of cellulose along with current generation
and the production of natural pigments, illustrating
the scope of using abundant cellulosic biomass for
energy production.
Experimental
MFC construction
A three-chambered MFC was constructed with a
Perspex sheet. The dimensions of the first chamber
were 5 cm 9 3 cm 9 4.5 cm, and the second and
third chambers had the same dimensions of
3 cm 9 3 cm 9 4.5 cm. The first and second cham-
bers were separated by a cellophane membrane. The
second and the third chambers were connected by a
2 cm 9 2-cm proton exchange membrane (Nafion
115). A. aceti (NCIM No. 2116) and G. roseus (NCIM
no. 2049) was procured from NCL, Pune, India.
Oscillatoria annae was procured from the National
Facility for Marine Cyanobacteria, Tiruchirappalli,
India. The first chamber contained cyanobacteria (1 g
wet weight) in BG-11 media (composition of BG-11
media is shown in the supplementary information); the
second chamber contained the anode with the biofilm
and the anolyte (phosphate buffer), and the third
chamber contained the carbon felt cathode and the
catholyte (3.3 g of potassium ferricyanide in 100 ml
of buffer).
Modification of the anode
The electrode modification procedure has already
been documented in the literature (Navanietha Krishn-
araj et al. 2013; Navanietha Krishnaraj and Sheela
2013a). Bare carbon felt (3.18 mm thick, procured
from Alfa Aesar) of 2 cm 9 2-cm dimensions was
modified by electrodepositing 1 % chitosan in 0.1 M
acetic acid at -10 V. The chitosan-deposited elec-
trode was left undisturbed until it became dry. Then,
an over layer of sodium alginate was formed by dip
coating in a 2 % sodium alginate solution and allowed
to dry further. Thus, the formed chitosan-sodium
alginate-coated felt was kept in 2 % CaCl2 overnight
for the formation of calcium alginate. Unmodified
carbon felt was used as a control for comparison.
Formation of the biofilm
The bare and the modified electrodes were kept in
phosphate buffer containing glucose (0.2 g/30 ml of
buffer) and a coculture of A. aceti and G. roseus (wet
weight of 0.1 g A. aceti and 0.1 g G. roseus) for
biofilm formation in stirred conditions until a stable
Cellulose
123

negative potential had been reached. Aneorobic con-
ditions were maintained during biofilm formation.
MFC operation and performance
The first chamber contained the cyanobacteria in BG
11 media. The anode compartment (second compart-
ment) of the three-chamber MFC was completely
deaerated with nitrogen gas. Cellulose acetate (0.03 g)
was used as the substrate. Modified carbon felt
(2.5 cm 9 2.5 cm) was used as the anode, and bare
carbon felt was used as the cathode. A data logger
(Agilent acquisition 34970A data acquisition) was
used to measure the voltage difference between the
anode and cathode across the fixed external resistance
at 5-min intervals. The data were collected automat-
ically by a data acquisition program and personal
computer. Then, the current (I) was calculated using
the formula I = V/R, where V is voltage and R the
applied resistance. Current density, j (A m-3), was
calculated using the formula j = I/v; power density,
P (W m-3), was calculated using the formula P = IV/
v, where v is the volume of the anolyte (15 ml).
Polarization studies were carried out in MFCs by
applying variable resistances between 10,000 and 100
X, and the final steady-state voltage was recorded for
each applied resistance as reported previously (Kart-
hikeyan and Sheela 2012). Ohmic resistance was
calculated from the slope of the polarization curve at
the linear (ohmic) region (Fan et al. 2008). A duplicate
experiment with an unmodified carbon felt anode was
carried out in an identical MFC for comparison. MFCs
with bare and modified electrodes were discharged
under a constant load, and their coulombic efficiencies
were calculated based on the following formula:
n ¼ Qobs=Qtheor � 100
where Qobs = the current gained under constant load
(C), and Qtheor = quantity of current expected from
the glucose consumption under constant load (C) (You
et al. 2006). Catalytic oxidation of fuel by the
microorganisms was analyzed by measuring COD
changes every 24 h (Logan et al. 2006). O. annae was
used for hydrolyzing the cellulose, and the kinetics of
hydrolysis of cellulose in the three-chambered MFC
were calculated by estimating the reducing sugar
levels (Miller 1959). After the completion of the
experiment, 5 ml of cyanobacterial culture from the
first chamber was centrifuged at 8,000 rpm. The
pigments were extracted, and the concentration of
the phycobiliproteins was calculated from the absorp-
tion spectrum of the supernatant in the wavelength
region of 250–700 nm. The C-phycocyanin, allophyc-
ocyanin and the C-phycoerythrin were quantified as
described elsewhere (Bennett and Bogorad 1973). The
acetone extract of the cyanobacterial biomass was
used to measure chlorophyll-a (663 nm) and carote-
noids (450 nm)(Mishra et al. 2012; McKinney 1941).
Electrochemical characterization of biofilm
Carbon felts of smaller dimensions (1 cm 9 0.4 cm)
were used for electrochemical characterization. A
brass rod was used for electrical contact. The electrode
was modified as described in ‘‘Modification of the
anode.’’ Bare and modified carbon felt electrodes in
the presence of biofilm were examined by cyclic
voltammetry in phosphate buffer in the potential
region from -1 to ?1.2 V at 50 mV s-1. A normal
calomel electrode (NCE) and Pt foil were used as the
reference electrode and the counter electrode,
respectively.
SEM analysis of microbial growth
The biofilm of the coculture formed over the bare and
modified felt was characterized by SEM analysis. A
piece of the bare and modified felt with biofilm felt
was carefully cut to 1 cm 9 1-cm dimensions in
aseptic conditions. The felt was gold sputter coated,
and the analyses of the biofilms were carried out using
the SEM Hitachi model-S-3000H unit.
Results and discussion
Modification of the electrode
Modification of the electrode was clearly investigated
using cyclic voltammetry analysis. The change in the
cyclic voltamogram indicates the effect of function-
alization of the carbon felt electrode with chitosan
and alginate. The cyclic voltamogram of bare carbon
felt, carbon felt modified with chitosan and carbon
felt modified with chitosan and sodium alginate
shows the increase in the surface area of the
electrode on modifying the electrode with chitosan
and alginate. The cyclic voltammograms of bare
Cellulose
123
carbon felt, carbon felt modified with chitosan and
carbon felt modified with chitosan and sodium
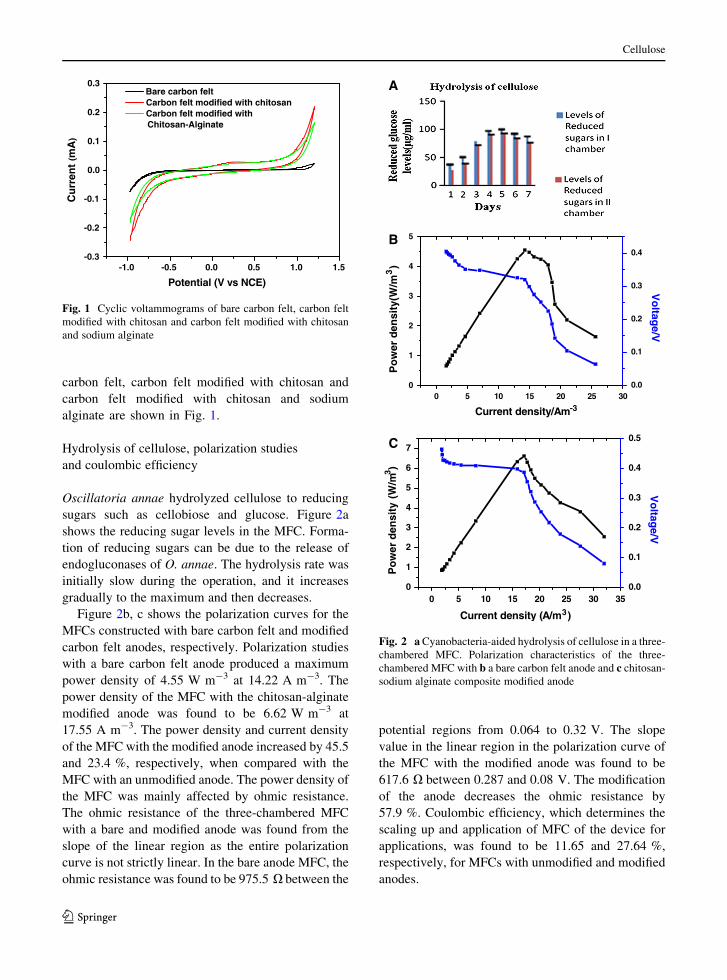
alginate are shown in Fig. 1.
Hydrolysis of cellulose, polarization studies
and coulombic efficiency
Oscillatoria annae hydrolyzed cellulose to reducing
sugars such as cellobiose and glucose. Figure 2a
shows the reducing sugar levels in the MFC. Forma-
tion of reducing sugars can be due to the release of
endogluconases of O. annae. The hydrolysis rate was
initially slow during the operation, and it increases
gradually to the maximum and then decreases.
Figure 2b, c shows the polarization curves for the
MFCs constructed with bare carbon felt and modified
carbon felt anodes, respectively. Polarization studies
with a bare carbon felt anode produced a maximum
power density of 4.55 W m-3 at 14.22 A m-3. The
power density of the MFC with the chitosan-alginate
modified anode was found to be 6.62 W m-3 at
17.55 A m-3. The power density and current density
of the MFC with the modified anode increased by 45.5
and 23.4 %, respectively, when compared with the
MFC with an unmodified anode. The power density of
the MFC was mainly affected by ohmic resistance.
The ohmic resistance of the three-chambered MFC
with a bare and modified anode was found from the
slope of the linear region as the entire polarization
curve is not strictly linear. In the bare anode MFC, the
ohmic resistance was found to be 975.5 X between the
potential regions from 0.064 to 0.32 V. The slope
value in the linear region in the polarization curve of
the MFC with the modified anode was found to be
617.6 X between 0.287 and 0.08 V. The modification
of the anode decreases the ohmic resistance by
57.9 %. Coulombic efficiency, which determines the
scaling up and application of MFC of the device for
applications, was found to be 11.65 and 27.64 %,
respectively, for MFCs with unmodified and modified
anodes.
-1.0 -0.5 0.0 0.5 1.0 1.5-0.3
-0.2
-0.1
0.0
0.1
0.2
0.3Bare carbon felt Carbon felt modified with chitosan Carbon felt modified with
Chitosan-Alginate
Cu
rren
t ( m
A)
Potential (V vs NCE)
Fig. 1 Cyclic voltammograms of bare carbon felt, carbon felt
modified with chitosan and carbon felt modified with chitosan
and sodium alginate
0 5 10 15 20 25 300
1
2
3
4
5
Current density/Am-3
Po
wer
den
sity
(W/m
3)
0.0
0.1
0.2
0.3
0.4
Vo
ltage/V
B
0 5 10 15 20 25 30 350
1
2
3
4
5
6
7
Current density (A/m )3
Po
wer
den
sity
(W/m
)
3
C
0.0
0.1
0.2
0.3
0.4
0.5
Vo
ltage/V
A
Fig. 2 a Cyanobacteria-aided hydrolysis of cellulose in a three-
chambered MFC. Polarization characteristics of the three-
chambered MFC with b a bare carbon felt anode and c chitosan-
sodium alginate composite modified anode
Cellulose
123
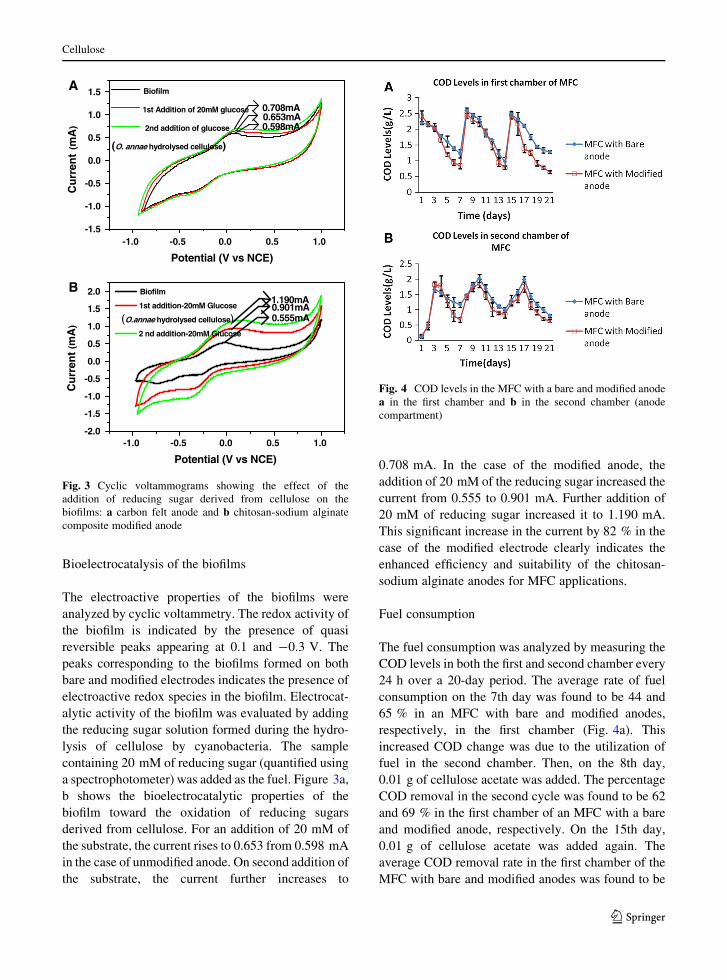
Bioelectrocatalysis of the biofilms
The electroactive properties of the biofilms were
analyzed by cyclic voltammetry. The redox activity of
the biofilm is indicated by the presence of quasi
reversible peaks appearing at 0.1 and -0.3 V. The
peaks corresponding to the biofilms formed on both
bare and modified electrodes indicates the presence of
electroactive redox species in the biofilm. Electrocat-
alytic activity of the biofilm was evaluated by adding
the reducing sugar solution formed during the hydro-
lysis of cellulose by cyanobacteria. The sample
containing 20 mM of reducing sugar (quantified using
a spectrophotometer) was added as the fuel. Figure 3a,
b shows the bioelectrocatalytic properties of the
biofilm toward the oxidation of reducing sugars
derived from cellulose. For an addition of 20 mM of
the substrate, the current rises to 0.653 from 0.598 mA
in the case of unmodified anode. On second addition of
the substrate, the current further increases to
0.708 mA. In the case of the modified anode, the
addition of 20 mM of the reducing sugar increased the
current from 0.555 to 0.901 mA. Further addition of
20 mM of reducing sugar increased it to 1.190 mA.
This significant increase in the current by 82 % in the
case of the modified electrode clearly indicates the
enhanced efficiency and suitability of the chitosan-
sodium alginate anodes for MFC applications.
Fuel consumption
The fuel consumption was analyzed by measuring the
COD levels in both the first and second chamber every
24 h over a 20-day period. The average rate of fuel
consumption on the 7th day was found to be 44 and
65 % in an MFC with bare and modified anodes,
respectively, in the first chamber (Fig. 4a). This
increased COD change was due to the utilization of
fuel in the second chamber. Then, on the 8th day,
0.01 g of cellulose acetate was added. The percentage
COD removal in the second cycle was found to be 62
and 69 % in the first chamber of an MFC with a bare
and modified anode, respectively. On the 15th day,
0.01 g of cellulose acetate was added again. The
average COD removal rate in the first chamber of the
MFC with bare and modified anodes was found to be
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5 Biofilm
1st Addition of 20mM glucose
2nd addition of glucose
(O. annae hydrolysed cellulose)
Cu
rren
t (m
A)
Potential (V vs NCE)
0.598mA0.653mA0.708mA
A
-1.0 -0.5 0.0 0.5 1.0
-1.0 -0.5 0.0 0.5 1.0-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0 Biofilm
1st addition-20mM Glucose
(O.annae hydrolysed cellulose) 2 nd addition-20mM Glucose
Cu
rren
t (m
A)
Potential (V vs NCE)
0.555mA0.901mA1.190mA
B
Fig. 3 Cyclic voltammograms showing the effect of the
addition of reducing sugar derived from cellulose on the
biofilms: a carbon felt anode and b chitosan-sodium alginate
composite modified anode
Fig. 4 COD levels in the MFC with a bare and modified anode
a in the first chamber and b in the second chamber (anode
compartment)
Cellulose
123

48 and 73 %, respectively. A similar study in the COD
levels was performed for the anolyte in the second
chamber and is shown in Fig. 4b. The COD levels
were initially very low in the second chamber of both
bare and modified anode MFCs. It slowly increased
and reached the maximum on the 3rd day after the
addition of substrate in the first chamber. Then, the
COD levels decreased on the 6th and 7th days. Then,
after the addition of substrate for the second cycle, the
COD levels increased after 2 days and decreased on
the 13th and 14th days, and this trend continued in the
third cycle. A similar pattern was observed in MFCs
with modified and unmodified anodes.
Production of cyanobacterial pigments
Different pigments were produced from the cyanobac-
terial biomass produced in the MFC. The biomass
contained 18.46783 mg l-1 of C-phycoerythrin,
13.09972 mg l-1 of allophycocyanin and
20.38901 mg l-1 of C-phycoerythrin. Chlorophyll-
a and carotenoid contents in the biomass were found
to be 0.376788 and 0.453 mg l-1 respectively. The
oxidation reactions in the second chamber did not affect
the production of pigments in the MFCs.
Morphological characterization of biofilms on bare
and modified electrodes
The morphological characterization of the biofilms on
bare and modified carbon felt was performed using
SEM. Figure 5a represents the biofilm on the bare
carbon felt anode. Figure 5b represents the SEM
image of the biofilms formed on the Chit–Alg/carbon
felt anode. It clearly shows the biocompatibility of the
chitosan-alginate composite enhances the biofilm
formation on the electrode.
Conclusion
The synergistic effect of two different types of
microorganisms has been capitalized for effective
degradation of cellulose resulting in the production of
pigments and electricity. The pigments could be of
substantial use in pharmaceuticals as antioxidants, as
food colorants, in cosmetics, in fluorescent labeling
and as a sensitizer for dye-sensitized solar cells. This
investigation brings out the scope of the effective
utilization of naturally abundant cellulosic biomass.
Acknowledgments The author, R. Navanietha Krishnaraj,
acknowledges CSIR, New Delhi, for the junior research
fellowship.
References
Bennett A, Bogorad L (1973) Complementary chromatic
adaptation in a filamentous blue green alga. J Cell Biol
58:419–435
Fig. 5 SEM image of the biofilm formed on a the bare carbon
felt anode and b carbon felt anode modified with chitosan–
sodium alginate composite
Cellulose
123

Dewan A, Donovan C, Heo D, Beyenal H (2010) Evaluating the
performance of microbial fuel cells powering electronic
devices. J Power Sources 195:90–96
Fan Y, Sharbrough E, Liu H (2008) Quantification of the
internal resistance distribution of microbial fuel cells.
Environ Sci Technol 42:8101–8107
Gupta V, Natarajan C, Kumar K, Prasanna R (2011) Identifi-
cation and characterization of endoglucanases for fungi-
cidal activity in Anabaena laxa (cyanobacteria). J Appl
Phycol 23:73–81
Hendriks AT, Zeeman G (2009) Pretreatments to enhance the
digestibility of lignocellulosic biomass. Bioresour Technol
100:10–18
Karthikeyan R, Sheela B (2012) Simultaneous degradation of
bad wine and electricity generation with the aid of the
coexisting biocatalysts Acetobacter aceti and Gluconob-
acter roseus. Bioresour Technol 104:388–393
Karthikeyan R, SathishKumar K, Murugesan M, Berchmans S,
Yegnaraman V (2009) Bioelectrocatalysis of Acetobacter
aceti and Gluconobacter roseus for current generation.
Environ Sci Technol 43:8684–8689
Kumar Saha S, Uma L, Subramanian G (2003) Nitrogen stress
induced changes in the marine cyanobacterium Oscillato-
ria willei BDU 130511. FEMS Microbiol Ecol 45:263–272
Ladisch MR, Ladisch CM, Tsao GT (1978) Cellulose to sugars:
new path gives quantitative yield. Science 201:743–745
Logan BE (2009) Exoelectrogenic bacteria that power microbial
fuel cells. Nat Rev Microbiol 7:375–381
Logan BE, Rabaey K (2012) Conversion of wastes into bio-
electricity and chemicals by using microbial electro-
chemical technologies. Science 337:686–690
Logan BE, Hamelers B, Rozendal R, Schroder U, Keller J,
Freguia S, Aelterman P, Verstraete W, Rabaey K (2006)
Microbial fuel cells: methodology and technology. Environ
Sci Technol 40:5181–5192
McKinney G (1941) Absorption of light by chlorophyll solu-
tions. J Biol Chem 140:315–322
Miller GL (1959) Use of dinitrosalicylic acid reagent for
determination of reducing sugar. Anal Chem 31:426–428
Mishra SK, Shrivastav A, Maurya RR, Patidar SK, Haldar S,
Mishra S (2012) Effect of light quality on the C-phycoer-
ythrin production in marine cyanobacteria Pseudanabaena
sp. isolated from Gujarat coast, India. Protein Expr Purif
81:5–10
Mosier N, Wyman C, Dale B, Elander R, Lee YY, Holtzapple
M, Ladisch M (2005) Features of promising technologies
for pretreatment of lignocellulosic biomass. Bioresour
Technol 96:673–686
Navanietha Krishnaraj R, Sheela B (2013a) In vitro antiplatelet
activity of silver nanoparticles synthesized using the
microorganism Gluconobacter roseus: an AFM-based
study. RSC Adv 3:8953–8959
Navanietha Krishnaraj R, Sheela B (2013b) Developing a
chitosan–sodium alginate composite based anode for
enhanced power output in cellulose fed Microbial fuel cell.
In: 24th annual general meeting (AGM) of the Materials
Research Society of India (MRSI), 11–13 February 2013
Navanietha Krishnaraj R, Karthikeyan R, Sheela B, Chandran S,
Parimal P (2013) Functionalisation of electrochemically
deposited chitosan films with alginate and Prussian blue for
enhanced performance of Microbial fuel cells. Electrochim
Acta 112:465–472
Navanietha Krishnaraj R, Saravanan C, Parimal P, Sheela B
(2014) Molecular modeling and assessing the catalytic
activity of glucose dehydrogenase of Gluconobacter sub-
oxydans with a new approach for power generation in a
microbial fuel cell. Curr Bioinform. doi:10.2174/
1574893608666131217234633
Orozco A, Ahmad M, Rooney D, Walker G (2007) Dilute acid
hydrolysis of cellulose and cellulosic bio-waste using a
microwave reactor system. Process Saf Environ Prot
85:446–449
Ren Z, Ward TE, Regan JM (2007) Electricity production from
cellulose in a microbial fuel cell using a defined binary
culture. Environ Sci Technol 41:4781–4786
Rezaei F, Xing D, Wagner R, Regan JM, Richard TL, Logan BE
(2009) Simultaneous cellulose degradation and electricity
production by Enterobacter cloacae in a microbial fuel
cell. Appl Environ Microbiol 75:3673–3678
Van der Lelie D, Taghavi S, McCorkle SM, Li LL, Malfatti SA,
Monteleone D, Donohoe BS, Ding SY, Adney WS, Him-
mel ME, Tringe SG (2012) The metagenome of an anaer-
obic microbial community decomposing poplar wood
chips. PLoS ONE 7:e36740. doi:10.1371/journal.pone.
0036740
You S, Zhao O, Zhang J, Jiang J, Zhao S (2006) A microbial fuel
cell using permanganate as the cathodic electron acceptor.
J Power Sources 162:1409–1415
Cellulose
123