Embed Size (px)
Citation preview

Susceptibility of brainstem to kindling and transfer to the
forebrain*yAnn Lam, *Naomi Whelan, and *yMichael E. Corcoran
*Department of Anatomy and Cell Biology; and yNeural Systems and Plasticity Research Group,
University of Saskatchewan, Saskatoon, Saskatchewan, Canada
SUMMARY
Purpose: The kindling of seizures with stimulation of
brainstem sites has been reported inconsistently in the lit-
erature. The characteristics of the kindling observed,
involving high intensities of stimulation and immediate
onset of generalized tonic–clonic convulsions, raise ques-
tions regarding the nature of kindling from these sites.
Methods: We implanted chronic electrodes in either the
nucleus reticularis pontis oralis (RPO), mesencephalic
reticular formation (MRF), dorsal periaqueductal gray
(dPAG), or ventrolateral periaqueductal gray (vlPAG) in
male Long-Evans rats, with a recording electrode in the
amygdala. Rats received conventional high-frequency
kindling stimulation once daily for 30 days. To test for
transfer, we kindled the amygdala beginning 7 weeks after
the last brainstem kindling trial.
Results: Tonic–clonic seizures were evoked by stimula-
tion from all brainstem sites. Seizures were brief and
were associated with characteristic low-amplitude high-
frequency afterdischarge (AD). Kindling of the dPAG
resulted in the development of classic AD and increased
AD duration. Prior kindling of the dPAG facilitated
subsequent kindling of the amygdala; however, no trans-
fer was observed with prekindling of other brainstem
sites.
Discussion: The variability in the response to kindling
stimulation suggests that certain brainstem sites are
resistant to kindling, whereas other sites are more sus-
ceptible to kindling but are still relatively resistant in
comparison to sites in the forebrain. The development of
classic AD in later trials of dPAG stimulation suggests
that epileptogenesis can occur even in the initial absence
of classic AD when low-amplitude high-frequency AD is
present.
KEY WORDS: Afterdischarge, Periaqueductal gray, Amy-
gdala, Mesencephalic reticular formation, Tonic–clonic
seizures.
Kindling was first described with stimulation in theforebrain, whereby repeated application of short trains ofelectrical stimulation led to development of increasinglycomplex convulsions and a permanent increase in seizuresusceptibility (Goddard, 1967; Goddard et al., 1969). Laterwork revealed that the stimulation must trigger afterdis-charge (AD) to produce kindling (Racine, 1972a), and hencekindling was characterized by additional criteria: (1) a longlasting decrease in the AD threshold (ADT); (2) an increasein AD duration and propagation to distant sites; and (3)progressive changes in motor seizures (Racine, 1972a,b).Kindling also involves transfer, wherein kindling of one sitecan lead to facilitation of subsequent kindling from asecondary site.
Variability of the characteristics of kindling occurs withstimulation of structures within and outside of the temporallobe. Examples of such patterns exist with kindling of thedorsal hippocampus (McIntyre & Kelly, 1993), olfactorybulb (Cain, 1977), piriform cortex, and perirhinal cortex(Mohapel et al., 1996). In such cases progression throughindividual stages may be accelerated, or absent altogether.Examination of the susceptibility to kindling of nonlimbicsites (e.g., claustrum) or sites outside of the forebrain (e.g.,brainstem) may help us understand their potential role inseizures.
The involvement of brainstem sites in seizure develop-ment and generalization has been investigated through avariety of interventions. For example, administration ofconvulsive drugs to thalamic and brainstem sites producesAD and convulsions (Elazar & Feldman, 1987; Peterson,1995; Ishimoto et al., 2000; Raisinghani & Faingold, 2003;Ishimoto et al., 2004). Furthermore, lesions of brainstemnuclei can disrupt development of audiogenic seizures(Browning, 1986). However, the responsiveness of brain-stem sites to electrical stimulation is less clear, and previous
Accepted February 2, 2010; Early View publication April 2, 2010.Address correspondence to Michael E. Corcoran, Neural Systems and
Plasticity Research Group, University of Saskatchewan, #342 – 111Research Drive, Saskatoon, SK, Canada S7N 3R2. E-mail: [email protected]
Wiley Periodicals, Inc.ª 2010 International League Against Epilepsy
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x
FULL-LENGTH ORIGINAL RESEARCH
1736

studies examining the susceptibility of brainstem sites tokindling have yielded conflicting results. Some researchers(Chiba & Wada, 1995; Chiba et al., 1996; Omori et al.,2001) have reported that classic AD, similar to that seen withforebrain stimulation and conventional kindling, occurs withstimulation of brainstem sites (e.g., periaqueductal gray,deep mesencephalic nucleus of the reticular formation) inrats. In contrast, others (Burnham et al., 1981; Chiu & Burn-ham, 1982) have reported convulsions without classic AD,and the absence of kindling with stimulation of brainstemsites.
In the present study, we investigated the susceptibility ofvarious brainstem sites to kindling and examined the subse-quent transfer to the amygdala. Our goal was to resolve theinconsistencies in the literature by selecting the previouslystudied regions including the dorsal periaqueductal gray(dPAG) and the mesencephalic reticular formation (MRF).Both sites have been shown to project to the amygdala(Robbins et al., 1990; Vianna & Brandao, 2003), which iswell known for its susceptibility to kindling. To furtherexamine the sensitivity of the brainstem to kindling, weselected two additional sites: the ventrolateral periaqueduc-tal gray (vlPAG) because of its reciprocal connectivity tothe dPAG and amygdala (Krettek & Price, 1978; Li et al.,1990), and the nucleus reticularis pontis oralis (RPO), whichhas been implicated in regulating convulsions in tonic–clo-nic seizures (Browning, 1986; Peterson, 1995). We hoped toobtain a better understanding of the brainstem’s involve-ment in development of generalized seizures.
Methods
General methodologySubjects were male Long-Evans rats from Charles Rivers
Laboratories (St. Constant, Quebec, Canada), weighing350–480 g at the time of surgery. They were housed ingroups of three prior to surgery, and individually thereafter.Food and water were freely available. Lights followed a12:12 h light–dark schedule, with lights on at 7 a.m. Ratswere individually handled for 3–5 min daily for 5 daysbefore surgery. Housing and experimental procedures werein accordance with the guidelines of the Canadian Councilof Animal Care, and procedures were approved by the Uni-versity of Saskatchewan Animal Care Committee.
SurgeryRats were anesthetized with 3% isoflurane and given a
subcutaneous injection of a preoperative analgesic, ketopro-fen (Anafen, 1 ml/kg). A bipolar nickel–chromium electrodewas stereotaxically implanted into one of the following sites:the RPO, 7.3 mm posterior, 2.0 mm lateral from bregma,7.5 mm ventral from the skull, n = 13; the MRF, 5.8 mmposterior, 1.7 mm lateral from bregma, 6.6 mm ventral fromthe skull; n = 10 (Ishimoto et al., 2000); the dPAG, 6.8 mmposterior, 2.2 mm lateral from bregma, 5.8 mm ventral from
the skull; n = 10 (Omori et al., 2001); and the vlPAG,7.8 mm posterior, 3.0 mm lateral from bregma, 6.7 mmfrom the skull; n = 11. A second bipolar electrode wasimplanted in the contralateral amygdala (2.6 mm posterior,4.5 mm lateral from bregma, 9.1 mm ventral from the skull).Four stainless steel screws were placed in the skull; electro-encephalography (EEG) was recorded from one pole of theelectrode referenced to a skull screw. Sham-kindled controlrats carrying electrodes implanted in the brainstem and theamygdala were connected to stimulation leads but did notreceive stimulation of the brainstem (n = 13). These ratswere used for behavioral and transfer experiments.
Electrical stimulation and kindling procedureOne week after surgery, rats received a train of conven-
tional high-frequency kindling stimulation in the brainstemonce daily. ADTs were determined at the brainstem elec-trode. Electrical stimulation was generated on a Grass S88stimulator (Astro Med, Inc., Warwick, RI, U.S.A.) and con-sisted of a 1-s train of balanced biphasic square-wave pulses;each pulse (positive or negative) was 1.0 ms in duration anddelivered at 60 pulse-pairs per second (pps). Data acquisi-tion for qualitative assessment of EEG employed Polyview2.3 (Astro Med, Inc.), collected at 100 samples per secondwith filters set at 3 Hz to 3 KHz. Higher sampling rates arecurrently being explored (see Electrographic Responses sec-tion in this article for further discussion). The initial stimula-tion current was set at 100 lA (base to peak) and increasedin increments of 200 lA at 1-min intervals until an AD wasevoked or tonic forelimb extension was observed. ADT wasdefined as the minimum intensity of stimulation producingAD outlasting the stimulation by at least 5 s.
Rats were stimulated once daily for 30 days at ADT. TheADT was measured again on the 10th, 20th, and 30th stimu-lations. Behavioral components during the seizure wererecorded in Polyview. The experimenters were trained to rec-ognize the behavioral components and recorded their onsetand offset using programmed markers on the keyboard.
TransferThe ADT of the amygdala was determined 7 weeks after
the last brainstem kindling session. During the 7 weeksbetween brainstem kindling and amygdaloid kindling, therats underwent a battery of behavioral tests. These testsincluded the elevated plus task, open field task, object recog-nition task, and water maze task. The results will be reportedelsewhere. With the exception of intensity, parameters ofstimulation were the same as with brainstem kindling. Theinitial stimulation current was set at 30 lA (base to peak)and increased in increments of 10 lA at 2-min intervals untilAD was evoked. If AD was triggered at 30 lA, the ADT wasremeasured the following day by setting the current at10 lA and increased in increments of 5 lA until AD wasevoked. Motor seizures were classified by the Racine scale:stage 1, mouth and facial movements; stage 2, head nodding;
1737
Characteristics of Brainstem Kindling and Transfer
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x
stage 3, unilateral forelimb clonus; stage 4, rearing andbilateral forelimb clonus; and stage 5, rearing and falling(Racine, 1972b). All rats were stimulated in the amygdalaat ADT twice daily with an interstimulation interval of atleast 4 h. Stimulation was discontinued after rats expressedfive stage 5 seizures or received 50 stimulations, whicheveroccurred first. Twenty-four hours after the last kindling ses-sion the amygdaloid ADTs were again measured.
HistologyAfter completion of testing, rats were euthanized and
their brains were sectioned at 40 lm on a freezing micro-tome. Sections were mounted on gelatin-coated slides andstained with the neuronal marker NeuN or with CresylViolet, and electrode placements were verified using a ratbrain stereotaxic atlas (Paxinos & Watson, 1997). Only ratswith correct placements were used for statistical analysis,and group sizes are reported in Fig. 2.
Results
Characteristics of kindlingCertain characteristics of the seizures were common to all
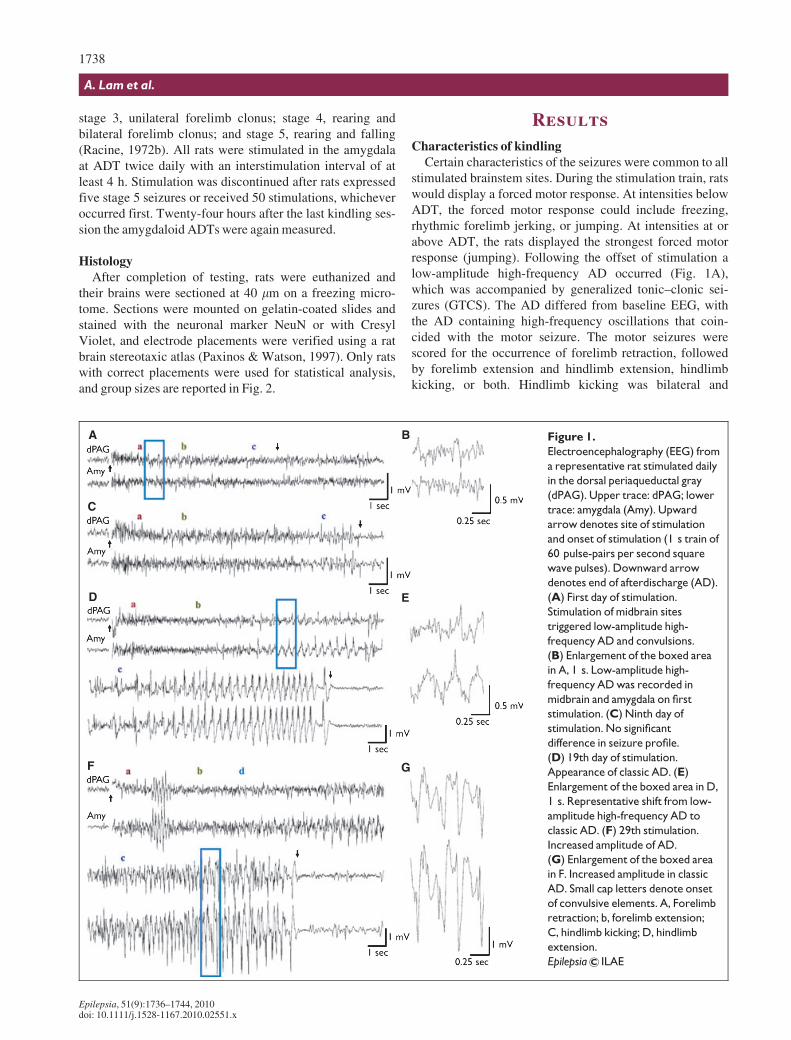
stimulated brainstem sites. During the stimulation train, ratswould display a forced motor response. At intensities belowADT, the forced motor response could include freezing,rhythmic forelimb jerking, or jumping. At intensities at orabove ADT, the rats displayed the strongest forced motorresponse (jumping). Following the offset of stimulation alow-amplitude high-frequency AD occurred (Fig. 1A),which was accompanied by generalized tonic–clonic sei-zures (GTCS). The AD differed from baseline EEG, withthe AD containing high-frequency oscillations that coin-cided with the motor seizure. The motor seizures werescored for the occurrence of forelimb retraction, followedby forelimb extension and hindlimb extension, hindlimbkicking, or both. Hindlimb kicking was bilateral and
A B
C
D E
F G
Figure 1.
Electroencephalography (EEG) from
a representative rat stimulated daily
in the dorsal periaqueductal gray
(dPAG). Upper trace: dPAG; lower
trace: amygdala (Amy). Upward
arrow denotes site of stimulation
and onset of stimulation (1 s train of
60 pulse-pairs per second square
wave pulses). Downward arrow
denotes end of afterdischarge (AD).
(A) First day of stimulation.
Stimulation of midbrain sites
triggered low-amplitude high-
frequency AD and convulsions.
(B) Enlargement of the boxed area
in A, 1 s. Low-amplitude high-
frequency AD was recorded in
midbrain and amygdala on first
stimulation. (C) Ninth day of
stimulation. No significant
difference in seizure profile.
(D) 19th day of stimulation.
Appearance of classic AD. (E)
Enlargement of the boxed area in D,
1 s. Representative shift from low-
amplitude high-frequency AD to
classic AD. (F) 29th stimulation.
Increased amplitude of AD.
(G) Enlargement of the boxed area
in F. Increased amplitude in classic
AD. Small cap letters denote onset
of convulsive elements. A, Forelimb
retraction; b, forelimb extension;
C, hindlimb kicking; D, hindlimb
extension.
Epilepsia ILAE
1738
A. Lam et al.
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x
synchronized, but occurred even after the end of AD. In allinstances, retraction and extension of limbs were bilateral.The duration of these motor components was variable acrossstimulation trials and groups, and the change in the totalduration of the motor seizures was found to be closely asso-ciated with changes in AD duration (data not shown). Themotor seizures resembled GTCS elicited by maximal elec-troshock (Kamei et al., 1978), pentylenetetrazol injection(Cain, 1980), and audiogenic stimulation (Browning, 1986).Initial and subsequent brainstem stimulations evoked AD inboth the brainstem and the amygdala (Fig. 1A, B) thatappeared to terminate simultaneously. In cases where clas-sic AD developed in later trials (Fig. 1C, D), secondary clas-sic AD or spindling also occurred simultaneously at bothrecording sites. In rare cases, rats of the dPAG group dis-played wet-dog shakes, after displaying classic AD.
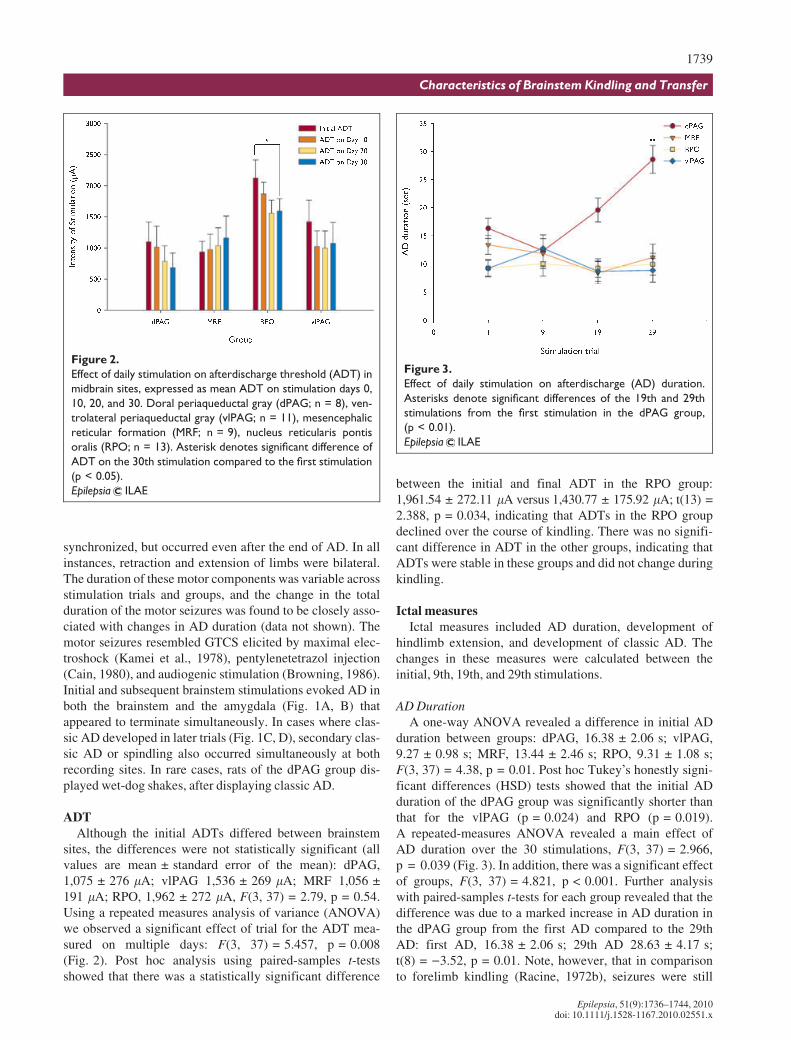
ADTAlthough the initial ADTs differed between brainstem
sites, the differences were not statistically significant (allvalues are mean € standard error of the mean): dPAG,1,075 € 276 lA; vlPAG 1,536 € 269 lA; MRF 1,056 €191 lA; RPO, 1,962 € 272 lA, F(3, 37) = 2.79, p = 0.54.Using a repeated measures analysis of variance (ANOVA)we observed a significant effect of trial for the ADT mea-sured on multiple days: F(3, 37) = 5.457, p = 0.008(Fig. 2). Post hoc analysis using paired-samples t-testsshowed that there was a statistically significant difference
between the initial and final ADT in the RPO group:1,961.54 € 272.11 lA versus 1,430.77 € 175.92 lA; t(13) =2.388, p = 0.034, indicating that ADTs in the RPO groupdeclined over the course of kindling. There was no signifi-cant difference in ADT in the other groups, indicating thatADTs were stable in these groups and did not change duringkindling.
Ictal measuresIctal measures included AD duration, development of
hindlimb extension, and development of classic AD. Thechanges in these measures were calculated between theinitial, 9th, 19th, and 29th stimulations.
AD DurationA one-way ANOVA revealed a difference in initial AD
duration between groups: dPAG, 16.38 € 2.06 s; vlPAG,9.27 € 0.98 s; MRF, 13.44 € 2.46 s; RPO, 9.31 € 1.08 s;F(3, 37) = 4.38, p = 0.01. Post hoc Tukey’s honestly signi-ficant differences (HSD) tests showed that the initial ADduration of the dPAG group was significantly shorter thanthat for the vlPAG (p = 0.024) and RPO (p = 0.019).A repeated-measures ANOVA revealed a main effect ofAD duration over the 30 stimulations, F(3, 37) = 2.966,p = 0.039 (Fig. 3). In addition, there was a significant effectof groups, F(3, 37) = 4.821, p < 0.001. Further analysiswith paired-samples t-tests for each group revealed that thedifference was due to a marked increase in AD duration inthe dPAG group from the first AD compared to the 29thAD: first AD, 16.38 € 2.06 s; 29th AD 28.63 € 4.17 s;t(8) = )3.52, p = 0.01. Note, however, that in comparisonto forelimb kindling (Racine, 1972b), seizures were still
Figure 3.
Effect of daily stimulation on afterdischarge (AD) duration.
Asterisks denote significant differences of the 19th and 29th
stimulations from the first stimulation in the dPAG group,
(p < 0.01).
Epilepsia ILAE
Figure 2.
Effect of daily stimulation on afterdischarge threshold (ADT) in
midbrain sites, expressed as mean ADT on stimulation days 0,
10, 20, and 30. Doral periaqueductal gray (dPAG; n = 8), ven-
trolateral periaqueductal gray (vlPAG; n = 11), mesencephalic
reticular formation (MRF; n = 9), nucleus reticularis pontis
oralis (RPO; n = 13). Asterisk denotes significant difference of
ADT on the 30th stimulation compared to the first stimulation
(p < 0.05).
Epilepsia ILAE
1739
Characteristics of Brainstem Kindling and Transfer
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x
relatively short in duration. The increase in AD durationcoincided with the development of classic AD in nearly alldPAG-kindled rats after the 20th stimulation.
Display of hindlimb extensionThe display of full hindlimb extension was inconsistent
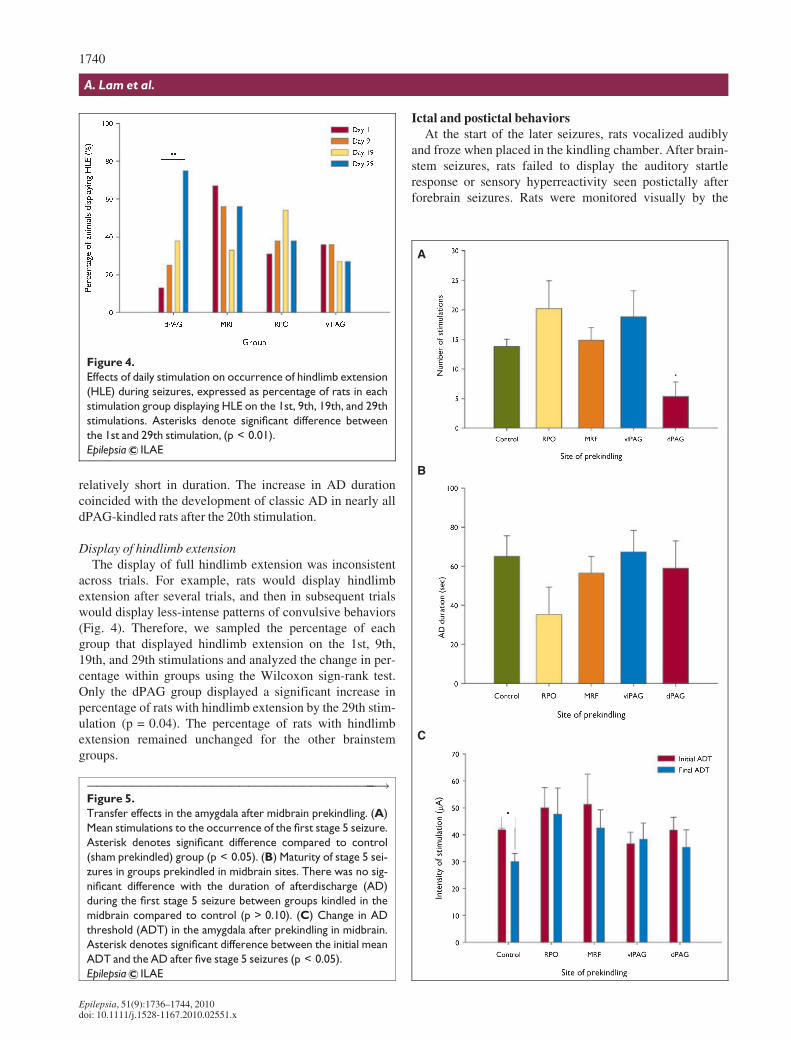
across trials. For example, rats would display hindlimbextension after several trials, and then in subsequent trialswould display less-intense patterns of convulsive behaviors(Fig. 4). Therefore, we sampled the percentage of eachgroup that displayed hindlimb extension on the 1st, 9th,19th, and 29th stimulations and analyzed the change in per-centage within groups using the Wilcoxon sign-rank test.Only the dPAG group displayed a significant increase inpercentage of rats with hindlimb extension by the 29th stim-ulation (p = 0.04). The percentage of rats with hindlimbextension remained unchanged for the other brainstemgroups.
Ictal and postictal behaviorsAt the start of the later seizures, rats vocalized audibly
and froze when placed in the kindling chamber. After brain-stem seizures, rats failed to display the auditory startleresponse or sensory hyperreactivity seen postictally afterforebrain seizures. Rats were monitored visually by the
Figure 4.
Effects of daily stimulation on occurrence of hindlimb extension
(HLE) during seizures, expressed as percentage of rats in each
stimulation group displaying HLE on the 1st, 9th, 19th, and 29th
stimulations. Asterisks denote significant difference between
the 1st and 29th stimulation, (p < 0.01).
Epilepsia ILAE
A
B
C
Figure 5.
Transfer effects in the amygdala after midbrain prekindling. (A)
Mean stimulations to the occurrence of the first stage 5 seizure.
Asterisk denotes significant difference compared to control
(sham prekindled) group (p < 0.05). (B) Maturity of stage 5 sei-
zures in groups prekindled in midbrain sites. There was no sig-
nificant difference with the duration of afterdischarge (AD)
during the first stage 5 seizure between groups kindled in the
midbrain compared to control (p > 0.10). (C) Change in AD
threshold (ADT) in the amygdala after prekindling in midbrain.
Asterisk denotes significant difference between the initial mean
ADT and the AD after five stage 5 seizures (p < 0.05).
Epilepsia ILAE
���������������������������������!
1740
A. Lam et al.
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x

experimenter for 1 min after the last motor seizure compo-nent. We observed porphyrin secretion around the eyes andnostrils during seizures, and piloerection and limb rigidityafter seizures. Although electrocardiography (ECG) was notrecorded, we noted apparent palpitations, prior to and duringhandling when rats were returned to their home cages. Ininstances in which hindlimb extension was present duringthe seizure, rats lost their righting reflex for up to 1 min afterthe seizure (data not shown). Because the postictal behav-iors were unanticipated at the time of the experiment, thedurations of these behaviors were not quantified. Hence, wereport these qualitative results only to raise the issue of pos-sible effects of brainstem seizures on interictal behavior.
TransferPrekindling of the RPO, vlPAG, or MRF failed to pro-
duce transfer to the forebrain, in that subsequent amygda-loid kindling proceeded at the rate seen in control rats.However, prekindling of the dPAG produced positivetransfer to amygdaloid kindling (Fig. 5A). The number ofamygdaloid stimulations to the first stage 5 seizurewere: dPAG, 5.33 € 2.49; vlPAG, 18.83 € 4.4; MRF, 14.88€ 2.15; RPO, 20.25 € 4.7; Control, 13.82 € 1.22; F(4, 37) =)3.065, p = 0.017.
The number of stimulations required for rats to developAD from both the amygdaloid and the brainstem sites wasalso significantly less with prekindling of the dPAG com-pared to control rats: dPAG, 3.33 € 1.33; vlPAG,11.67 € 2.51; MRF, 10.25 € 1.78; RPO, 11.25 € 6.80;Control, 9.73 € 0.97; F(4, 37) = 2.866, p = 0.038. Analy-sis of the first stage 5 seizure revealed that AD durationsdid not differ between experimental and control rats:dPAG, 59 € 13.98 s; vlPAG, 67.38 € 10.87 s; MRF,56.5 € 8.62 s; RPO, 35.38 € 8.26 s; Control, 65.09 €10.61 s; F(4, 37) = 1.406, p = 0.178 (Fig. 5B).
ANOVA of the latency to clonus and duration of clonusof the first stage 5 seizures revealed no difference betweenprekindled rats and control rats (data not shown). However,the variance of the initial amygdaloid ADTs and the finalamygdaloid ADTs in prekindled rats was consistentlygreater than control (Fig. 5C), despite similar electrodeplacements within the amygdala. Further analysis revealedthat only control rats showed a decrease in ADT (p = 0.04)after amygdaloid kindling. Therefore, we unexpectedlyobserved that prekindling of brainstem sites was associatedwith a failure of amygdaloid kindling to lead to a decreasein ADT.
Discussion
Electrographic responsesStimulation of all sites was followed by a pattern of low-
amplitude high-frequency activity, similar to the patternpreviously described in the bulbar reticular formation dur-ing amygdaloid kindling in baboons (Wada & Osawa,
1976), by Burnham et al. (1981) during MRF kindling inrats, and by Magoun (1950) in response to stimulation ofthe RF in cats. In an ongoing study employing waveletanalysis of the EEG acquired at high sampling rates, wehave confirmed that the epileptiform response associatedwith brainstem kindling comprises signals of a significantlyhigher frequency than background, and the response isqualitatively similar to the waveforms reported in thisarticle (Lam et al., 2009). High sampling rates for quanti-tative analysis are needed to properly capture the low-amplitude high-frequency activity observed in this studyand to address potential issues of aliasing.
Changes in EEG could be the result of shifts in nonepilep-tiform activity (i.e., arousal or motor activity), but severalaspects of the behavioral events following stimulation sug-gest a stronger case for identifying the low-amplitude high-frequency activity as epileptiform activity. For example,Moruzzi and Magoun demonstrated that stimulation of theMRF in lightly anaesthetized cats would result in ‘‘lowamplitude fast activity’’ (Moruzzi & Magoun, 1949); how-ever, in fully awake cats there was no difference in EEG.The rats in our preparation were awake and alert, and hencewe suggest that the low-amplitude high-frequency activityin response to the stimulation is not due to desynchronousactivity associated with changes in arousal. Furthermore,stimulation at subthreshold levels could evoke runningresponses without low-amplitude high-frequency AD, sug-gesting that the activity we observed is not associated withmotor artifacts. We, therefore, consider the low-amplitudehigh-frequency activity to be epileptiform AD, although itdoes differ markedly from the classic AD recorded in theforebrain.
AD durationIn the vlPAG, MRF, and RPO, the AD duration appeared
to remain stable over the course of the experiment, althoughsmall sample sizes may have contributed to the lack of dif-ference across trials. In contrast, repeated stimulation of thedPAG resulted in significant increases in the duration ofAD, and this was associated with development of classicAD. Classic AD, therefore, is correlated with and may benecessary for changes in AD duration, whereas the trigger-ing of only low-amplitude high-frequency AD seems tosignify the resistance of a site to kindling, insofar as pro-gressive increase in duration of AD is a characteristic ofkindling. Of interest, the AD duration of a brainstem sei-zure, even in the presence of classic AD, was also noticeablyshorter than with seizures kindled from forebrain sites suchas the hippocampus and the amygdala (Racine, 1972b).
Previous studies have reported a ‘‘silent’’ period duringaudiogenic seizures and brainstem seizures in the cortex(Omori et al., 2001). This period may be the low-amplitudehigh-frequency AD we have observed in cortical recordingduring brainstem kindling (Lam & Corcoran, 2005). We arecurrently employing wavelet analysis to characterize AD
1741
Characteristics of Brainstem Kindling and Transfer
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x

further, in order to provide additional insight into the cir-cuitry involved in generating very high frequency dis-charge, which has been implicated in epileptogenesis(Bragin et al., 1999; Jirsch et al., 2006).
Afterdischarge thresholdStimulation of the RPO but not the MRF, dPAG, or
vlPAG resulted in a significant decrease in ADT, a measurenot examined in previous studies (Chiba & Wada, 1995;Omori et al., 2001). Smaller changes in ADT from theMRF, dPAG, and vlPAG may not have been detected, dueto the large step-wise increases in intensities of stimulation,and these sites should therefore be reexamined in futurestudies.
We do not believe that the high intensities of stimulationrequired to trigger AD are a product of neuronal damage ordegeneration at the site of stimulation. Two lines of evi-dence argue against significant damage. First, the initialADT in the RPO was significantly higher than in thedPAG, and thus we should not have observed a decrease inADT in the RPO if the stimulation was producing damageor degeneration. Second, we have failed to observe fluoro-jade B positive staining in brainstem-stimulated tissue thatwould be indicative of damage or degeneration (data notshown).
ConclusionsStimulation of all brainstem sites at ADT triggered imme-
diate and severe motor seizures that resembled electrocon-vulsive shock-induced seizures and audiogenic seizures.However, repeated stimulation led to increasingly severemotor seizures only in the rats receiving stimulation of thedPAG, as indicated by an increase in the number of rats dis-playing hindlimb extension. Therefore, with the exceptionof the dPAG, most sites supported convulsive responses thatwere relatively implastic.
The convulsions and even the explosive jumping and roll-ing force motor reactions (during the stimulation) are alsoreminiscent of Pinel’s stage 7 and stage 8 seizures observedduring long-term kindling of the amygdala (Pinel & Rovner,1978), which have been suggested to be due to the involve-ment of brainstem nuclei. However, the forced motorresponses occurred during the 1 s of stimulation and notduring the seizure, and it is, therefore, unclear whether thesereactions are driven by the same mechanisms responsiblefor stage 7 seizures. On the other hand, the tonic responsesappear to be sustainable during the seizures. This may,therefore, imply that the tonic responses observed in long-term amygdaloid kindling involve recruitment of thesebrainstem nuclei. Tonic extension is also observed duringthe severe audiogenic seizures of the genetically epilepsyprone rat (GEPR-9), in which ‘‘rapid tonic firing greatlyaccelerated just prior to tonus’’ in the periaqueductal grey(Faingold, 1998). Thus, our findings may provide furtherevidence of the importance of recruiting the periaqueductal
grey to sustain tonic seizure components in a variety of tonicseizure types with or without brainstem origin.
Ictal and postictal behaviorsAbnormal changes in heart rate or rhythmicity could be
a sign of seizure effects on cardiovascular function,which could imply brainstem involvement in suddenunexpected death in epilepsy (SUDEP) (Stçllberger &Finsterer, 2004; Mukherjee et al., 2009). Therefore, studiesexamining the relation of ECG and EEG during the ictaland postictal periods of brainstem-related seizures would beinformative.
Porphyrin secretion (chromodacryorrhea) was not presentin all rats, and those rats that did secrete porphyrin did notdo so on every trial. However, this substance is a well-described sign of stress (Buccafusco, 1990; Rommereimet al., 1990), and we believe that it is indicative of stressfuleffects on the chronic brainstem seizures or activation of theautonomic nervous system, which receive input frombrainstem nuclei, such as the periaqueductal grey. Many ofthe postictal behaviors were unexpected and have not beennoted in the previous literature on brainstem kindling. Thesefindings could be useful in the understanding of the behav-ioral comorbidities of epilepsy.
TransferPrevious investigations of brainstem stimulation have not
consistently indicated whether transfer occurs. The differ-ences in the brainstem kindling literature in which transferis present (Wake & Wada, 1977; Hirsch et al., 1992) orabsent (Michalakis et al., 1998; Sheerin et al., 2004) appearto be due to variability in sites of stimulation. Herein wereport transfer to amygdaloid kindling with prekindling onlyof dPAG. In contrast, transfer was not observed by Omoriet al. with dPAG kindling (Omori et al., 2001). However,we kindled to 30 generalized seizures, 10–15 stimulationsmore than in previous brainstem kindling studies. In ourstudy, one difference between the characteristics of dPAGkindling and the other brainstem sites is the consistentdevelopment of classic AD within the entire group over 30stimulations. With stimulation of the MRF and vlPAG,some rats developed classic AD, but the effect was unstableand not present consistently. We did not find an effect ofstimulation of the brainstem on the duration of AD duringsecondary-site amygdaloid kindling, implying that therewas no effect on the severity of the amygdaloid seizures.
An unexpected finding was the increased variability inthe ADT in the amygdala after stimulation in all brainstemsites. It is known that there is variability in seizure thresholdwithin any given population, possibly due to individualresistance to seizures. Stimulation of the brainstem mightfurther accentuate the difference such that rats that havelower ADT in secondary sites are more susceptible to sei-zures in general, whereas those that have higher ADTs areless susceptible to seizures.
1742
A. Lam et al.
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x

Role of structure and connectivityThe differential responses of brainstem nuclei to kindling
stimulation may be a result of differences in afferent andefferent connections, or differences in network propertiesof the local cellular networks. The periaqueductal greycolumns receive efferents from sites implicated in seizures,such as the superior colliculus (i.e., in audiogenic seizures:[(Yang et al., 2003), and amygdala (Li et al., 1993)].Furthermore, the dlPAG and vlPAG columns have strongoutput to motor pathways of the spinal cord. However, thelack of kindling from stimulation of the RPO is surprising,given its efferent connections to forebrain sites. For exam-ple, along with the pedunculopontine nucleus, the RPO hasbeen implicated in the synchronized firing of ascendingpathways to the medial septal nucleus, which in turnconnects to the hippocampus, entorhinal cortex, and otherlimbic sites (Jackson & Bland, 2006). These sites are allsusceptible to kindling, and our results, therefore, mightsuggest that kindling does not occur only by overexcita-tion of efferents at the site of stimulation, but likely alsoinvolves the changing of local network properties (e.g.,neurochemical, gap junction, glial responses) to create aseizure focus.
Nevertheless, the anatomic connections of these brain-stem sites may play an important part in the chronic behav-ioral changes associated with epilepsy. For example, thevlPAG projects to the rostral ventral medulla, pontineadrenergic nuclei, and the pontobulbar reticular formation(Odeh & Antal, 2001), and the central amygdala (Adamecet al., 2001). These sites are implicated in the regulation ofanalgesia and defensive behaviors, which may be height-ened with chronic stimulation.
In summary we conclude that kindling can occur at brain-stem sites, but that the nuclei are not equally susceptible.The presence of low-amplitude high-frequency AD togetherwith the absence of classic AD in the early stimulation ofthe dPAG suggest that classic AD is not required for kind-ling. Further investigation of brainstem kindling may pro-vide insight into how these sites interact with others toproduce generalized seizures and are involved in the comor-bidities of epilepsy.
Acknowledgments
This work was financially supported by grants from the NaturalSciences and Engineering Research Council and the Canadian Institutesof Health Research awarded to MEC. We thank Jason Wagner and KenWolfe for technical assistance and Elan Ohayon for comments on themanuscript.
Disclosure
We confirm that we have read the Journal’s position on issuesinvolved in ethical publication and affirm that this report is consistentwith those guidelines. None of the authors has any conflict of interest todisclose.
References
Adamec RE, Blundell J, Collins A. (2001) Neural plasticity and stressinduced changes in defense in the rat. Neurosci Biobehav Rev 25:721–744.
Bragin A, Engel J Jr, Wilson CL, Fried I, Buzsaki G. (1999) High-fre-quency oscillations in human brain. Hippocampus 9:137–142.
Browning RA. (1986) Neuroanatomical localization of structures responsi-ble for seizures in the GEPR: lesion studies. Life Sci 39:857–867.
Buccafusco JJ. (1990) Participation of different brain regions in the anti-narcotic withdrawal action of clonidine in the dependent rat. Brain Res513:8–14.
Burnham WM, Albright P, Schneiderman J, Chiu P, Ninchoji T. (1981)Centrencephalic mechanisms in the kindling model. In Wada JA (Ed)Kindling 2. Raven Press, New York, pp. 161–178.
Cain DP. (1977) Seizure development following repeated electrical stimu-lation of central olfactory structures. Ann N Y Acad Sci 290:200–216.
Cain DP. (1980) Effects of kindling or brain stimulation on pentylenetetra-zol-induced convulsion susceptibility. Epilepsia 21:243–249.
Chiba S, Wada JA. (1995) Kindling of the interpeduncular nucleus and itsinfluence on subsequent amygdala kindling in rats. Epilepsia 36:410–415.
Chiba S, Omori N, Kamata S, Nunomura A, Mutoh F. (1996) Kindling ofthe mesencephalic reticular formation and its influence on subsequentamygdala kindling in rats. Epilepsia 37:116–117.
Chiu P, Burnham WM. (1982) The effect of anticonvulsant drugs on con-vulsions triggered by direct stimulation of the brainstem. Neurophar-macology 21:355–359.
Elazar Z, Feldman Z. (1987) Brainstem experimental seizures produced bymicroinjections of carbachol. Epilepsia 28:463–470.
Faingold CL. (1998) Locomotor behaviors in generalized convulsions arehierarchically driven from specific brain-stem nuclei in the networksubserving audiogenic seizure. Ann N Y Acad Sci 860:566–569.
Goddard GV. (1967) Development of epileptic seizures through brain stim-ulation at low intensity. Nature 214:1020–1021.
Goddard GV, McIntyre DC, Leech CK. (1969) A permanent change inbrain function resulting from daily electrical stimulation. Exp Neurol25:295–330.
Hirsch E, Maton B, Vergnes M, Depaulis A, Marescaux C. (1992) Positivetransfer of audiogenic kindling to electrical hippocampal kindling inrats. Epilepsy Res 11:159–166.
Ishimoto T, Omori N, Mutoh F, Chiba S. (2000) Convulsive seizuresinduced by n-methyl-d-aspartate microinjection into the mesencephalicreticular formation in rats. Brain Res 881:152–158.
Ishimoto T, Chiba S, Omori N. (2004) Convulsive seizures induced byalpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid microin-jection into the mesencephalic reticular formation in rats. Brain Res1021:69–75.
Jackson J, Bland BH. (2006) Medial septal modulation of the ascendingbrainstem hippocampal synchronizing pathways in the anesthetized rat.Hippocampus 16:1–10.
Jirsch JD, Urrestarazu E, LeVan P, Olivier A, Dubeau F, Gotman J. (2006)High-frequency oscillations during human focal seizures. Brain129:1593–1608.
Kamei C, Masuda Y, Oka M, Shimizu M. (1978) Effects of antiepilepticson both behavioral and electrographic seizure patterns induced by max-imal electroshock in rats. Epilepsia 19:625–636.
Krettek JE, Price JL. (1978) Amygdaloid projections to subcortical struc-tures within the basal forebrain and brainstem in the rat and cat. J CompNeurol 178:225–253.
Lam A, Corcoran ME. (2005) Convulsions induced by electrical stimula-tion of the brainstem in rats. Epilepsia 46:s113.
Lam A, Ohayon EL, Burnham WM, Corcoran ME. (2009) High frequenciesand chirps in the evolution of seizure transitions during brainstem kind-ling. Poster session presented at the Society for Neuroscience AnnualMeeting, October 17–21, 2009, Chicago, IL.
Li YQ, Jia HG, Rao ZR, Shi JW. (1990) Serotonin-, substance p- or leu-cine-enkephalin-containing neurons in the midbrain periaqueductalgray and nucleus raphe dorsalis send projection fibers to the centralamygdaloid nucleus in the rat. Neurosci Lett 120:124–127.
Li YQ, Takada M, Matsuzaki S, Shinonaga Y, Mizuno N. (1993) Identifica-tion of periaqueductal gray and dorsal raphe nucleus neurons projecting
1743
Characteristics of Brainstem Kindling and Transfer
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x

to both the trigeminal sensory complex and forebrain structures: a fluo-rescent retrograde double-labeling study in the rat. Brain Res 623:267–277.
Magoun HW. (1950) Caudal and cephalic influences of the brain stem reti-cular formation. Physiol. Rev. 30:459–474.
McIntyre DC, Kelly ME. (1993) Are differences in dorsal hippocampalkindling related to amygdala-piriform area excitability? Epilepsy Res14:49–61.
Michalakis M, Holsinger D, Ikeda-Douglas C, Cammisuli S, Ferbinteanu J,DeSouza C, DeSouza S, Fecteau J, Racine RJ, Milgram NW. (1998)Development of spontaneous seizures over extended electrical kindling.I. Electrographic, behavioral, and transfer kindling correlates. BrainRes 793:197–211.
Mohapel P, Dufresne C, Kelly ME, McIntyre DC. (1996) Differential sensi-tivity of various temporal lobe structures in the rat to kindling and statusepilepticus induction. Epilepsy Res 23:179–187.
Moruzzi G, Magoun HW. (1949) Brain stem reticular formation and activa-tion of the eeg. Electroencephalogr Clin Neurophysiol 1:455–473.
Mukherjee S, Tripathi M, Chandra P, Yadav R, Choudhary N, Sagar R,Bhore R, Pandey R, Deepak KK. (2009) Cardiovascular autonomicfunctions in well-controlled and intractable partial epilepsies. EpilepsyRes 85:261–269.
Odeh F, Antal M. (2001) The projections of the midbrain periaqueductalgrey to the pons and medulla oblongata in rats. Eur J Neurosci14:1275–1286.
Omori N, Ishimoto T, Mutoh F, Chiba S. (2001) Kindling of the midbrainperiaqueductal gray in rats. Brain Res 903:162–167.
Paxinos G, Watson C. (1997) The rat brain I stereotaxic coordinates. Aca-demic Press, San Diego.
Peterson SL. (1995) Infusion of NMDA antagonists into the nucleus reticu-laris pontis oralis inhibits the maximal electroshock seizure response.Brain Res 702:101–109.
Pinel JPJ, Rovner LI. (1978) Experimental epileptogenesis: kindling-induced epilepsy in rats. Exp Neurol 58:190–202.
Racine RJ. (1972a) Modification of seizure activity by electrical stimula-tion. I. After-discharge threshold. Electroencephalogr. Clin Neurophys-iol 32:269–279.
Racine RJ. (1972b) Modification of seizure activity by electrical stimula-tion. II. Motor seizure. Electroencephalogr Clin Neurophysiol 32:281–294.
Raisinghani M, Faingold CL. (2003) Identification of the requisite brainsites in the neuronal network subserving generalized clonic audiogenicseizures. Brain Res 967:113–122.
Robbins A, Schwartz-Giblin S, Pfaff DW. (1990) Ascending and descend-ing projections to medullary reticular formation sites which activatedeep lumbar hack muscles in the rat. Exp Brain Res 80:463–474.
Rommereim DN, Rommereim RL, Sikov MR, Buschbom RL, AndersonLE. (1990) Reproduction, growth, and development of rats duringchronic exposure to multiple field strengths of 60-hz electric fields.Fundam Appl Toxicol 14:608–621.
Sheerin AH, Nylen K, Zhang X, Saucier DM, Corcoran ME. (2004) Furtherevidence for a role of the anterior claustrum in epileptogenesis. Neuro-science 125:57–62.
Stçllberger C, Finsterer J. (2004) Cardiorespiratory findings in suddenunexplained/unexpected death in epilepsy (SUDEP). Epilepsy Res59:51–60.
Vianna DM, Brandao ML. (2003) Anatomical connections of the periaqu-eductal gray: specific neural substrates for different kinds of fear. BrazJ Med Biol Res 36:557–566.
Wada JA, Osawa T. (1976) Spontaneous recurrent seizure state induced bydaily electric amygdaloid stimulation in senegalese baboons (papiopapio). Neurology 26:273–286.
Wake A, Wada JA. (1977) Transfer and interference in amygdaloid kind-ling in cats. Can J Neurol Sci 4:5–11.
Yang L, Long C, Randall ME, Faingold CL. (2003) Neurons in the periaqu-eductal gray are critically involved in the neuronal network for audio-genic seizures during ethanol withdrawal. Neuropharmacology44:275–281.
1744
A. Lam et al.
Epilepsia, 51(9):1736–1744, 2010doi: 10.1111/j.1528-1167.2010.02551.x