Embed Size (px)
Citation preview

of July 1, 2018.This information is current as
DermatitisNC/Nga Mice, a Model of Human Atopic
inApplication of CpG-Oligodeoxynucleotides Suppression of Skin Lesions by Transdermal
Joe Inoue and Yukihiko Aramaki
http://www.jimmunol.org/content/178/1/584doi: 10.4049/jimmunol.178.1.584
2007; 178:584-591; ;J Immunol
Referenceshttp://www.jimmunol.org/content/178/1/584.full#ref-list-1
, 9 of which you can access for free at: cites 38 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 1, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Suppression of Skin Lesions by Transdermal Applicationof CpG-Oligodeoxynucleotides in NC/Nga Mice, a Modelof Human Atopic Dermatitis1
Joe Inoue and Yukihiko Aramaki2
Atopic dermatitis (AD) is a pruritic inflammatory skin disease characterized by an elevation of the total IgE level in plasma, theinfiltration of mast cells and eosinophils, and the expression of cytokines by Th2 cells. NC/Nga mice kept in conventional conditionsare known to develop skin lesions resembling human AD. We examined in this study the alterations of immune response in NC/Ngamice kept in conventional conditions, following transdermal application of CpG-oligodeoxynucleotides (ODN), which plays acritical role in immunity via the augmentation of Th1-type and suppression of Th2-type responses. CpG-ODN remarkablychanged the immune response from type Th2 to Th1 as determined from cytokine mRNA and Ab levels. The serum IgE level wasdecreased and the expression of IgG2a was up-regulated. The application of CpG-ODN to the skin also decreased inflammatoryinfiltration of mast cells, and suppression in the skin lesions was observed. Furthermore, the generation of regulatory T cells, whichare considered immune suppressive T cells, was observed in the skin on treatment with CpG-ODN. These results suggestedCpG-ODN is effective for immunotherapy in patients with AD, which is characterized by Th2-dominated inflammation. TheJournal of Immunology, 2007, 178: 584–591.
A topic disorders have a complex and chronic pathogenesisthat provides many potential cellular and molecular tar-gets for therapeutic intervention, but may also include
redundant pathways mediating disease (1, 2). In atopic dermatitis(AD),3 the skin lesions show a chronically relapsing inflammatorydisorder with prurience and eczema usually associated with ele-vated serum IgE levels (3, 4). The skin lesions of AD patients arecharacterized by the presence of inflammatory infiltrates consistingof T lymphocytes, monocytes/macrophages, eosinophils, and mastcells (5, 6).
Dysregulated Th1 or Th2 responses are thought to be central tothe pathology of diseases such as AD and asthma, which are char-acterized by Th2-dominated allergic inflammation (7–9). Th2-likeimmune responses mediated by IL-4, IL-5, and IL-13 are key tothe pathogenesis of atopic disorders (8, 9) because the up-regula-tion of IgE production, one of the major causes of atopic inflam-mation, has been extensively regulated with Th2 cytokines, IL-4and IL-13. Th2 cell numbers are increased in lesional tissue ofpatients who suffer from atopic diseases and patients with ADfrequently show elevated IgE levels in response to many kinds ofallergens such as mite Ag and house dust (10, 11). Recently, manyexperiments have shown that the T cells constitutively expressingCD25, regulatory T cells (Tregs), are stably anergic and suppres-
sive, and regulate various autoimmune diseases and allergic dis-eases (12–14). Therefore, it is very important to consider the bal-ance of Th1, Th2, and Tregs in patients with immune diseasesespecially in allergic diseases.
Both bacterial dinucleotides flanked by certain bases (CpG-motif)and synthetic oligodeoxynucleotides (ODN) containing a CpG-motif(CpG-ODN) activate cells such as B cells, macrophages, and den-dritic cells through TLR9 (15–17). Signaling through TLR9 leadsto the secretion of proinflammatory cytokines such as IL-1, IL-6,TNF-�, and IL-12 (15–17). IL-12 acts on T cells and NK cellsinducing the production of cytokines, primarily IFN-�. Conse-quently, CpG-ODN could be useful as an adjuvant for cellular andhumoral immunity (18). It is also important that CpG-ODN-acti-vated dendritic cells induced the generation of Tregs with a strongimmune-suppressive function (19). So we expect that the ability ofCpG-ODN to shift the immune response from type Th2 to Th1with the generation of Tregs can be exploited to develop an im-munotherapy for patients with diseases like AD that are associatedwith a Th2-predominant immune response.
NC/Nga mice are an inbred strain established from Japanesefancy mice in 1957 by Kondo et al. (20). When kept under con-ventional conditions, they started to scratch themselves at �8 wk,and their skin became dry and scaly. Within the next severalweeks, the mice developed lesions on the ears, back, neck, andface. Immunohistochemical examination of the lesions in conven-tional NC/Nga mice reveal hyperkeratosis, acanthosis, and para-keratosis, all of which resemble the typical features of the skinobserved in patients with AD (9, 21). The lesions show lympho-cyte infiltration, macrophage infiltration, and mast cell and eosin-ophil degranulation. In addition, the level of IgE in the serumgradually increases (22). All these immunological, histological,and biochemical changes in conventional NC/Nga mice resemblehuman AD.
We have reported that administration of CpG-ODN through thebarrier-disrupted skin may shift the immune response from typeTh2 to Th1 and drastically attenuated the production of IgE inmice undergoing an IgE-type immune response (23, 24). In this
School of Pharmacy, Tokyo University of Pharmacy and Life Science, Tokyo, Japan
Received for publication December 7, 2005. Accepted for publication October16, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by Grant-in-aid 14657594 for Scientific Researchfrom the Ministry of Education, Science, Sports, and Culture of Japan (to Y.A.).2 Address correspondence and reprint requests to Dr. Yukihiko Aramaki, School ofPharmacy, Tokyo University of Pharmacy and Life Science 1432-1 Horinouchi,Hachioji, Tokyo 192-0392, Japan. E-mail address: [email protected] Abbreviations used in this paper: ODN, oligodeoxynucleotide; AD, atopic derma-titis; Treg, regulatory T cell; LN, lymph node.
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

study, we examined the alterations in immune response in con-ventional NC/Nga mice, which spontaneously develop AD-likesymptoms and high Th2-immune responses following a applica-tion of CpG-ODN to the skin. We also investigated the generationof Tregs in the skin lesions following the application of CpG-ODN. CpG-ODN remarkably changed the immune response fromtype Th2 to Th1 as determined from cytokine mRNA and Ab lev-els. CpG-ODN induced Tregs to appear in the skin lesions. Theseresults suggested that CpG-ODN could change cytokine produc-tion and generate Tregs in conventional NC/Nga mice, and may beeffective for immunotherapy in patients with AD.
Materials and MethodsAnimals and reagents
NC/Nga mice (male, 6-wk-old) were purchased from Nihon SLC Co. Thesequence for control ODN (non-CpG-ODN) was 5�-TCCATGAGCTTCCTGAGTCT-3� and CpG-ODN was 5�-TCCATGACGTTCCTGATGCT-3�, and HPLC-purified phosphorothioate ODN was obtained fromSigma-Genosys. Rhodamine (Rho)-labeled CpG-ODN was also obtainedfrom Sigma-Genosys.
Animal experiment
Animal use and relevant experimental procedures were approved by theTokyo University of Pharmacy and Life Science Committee on the Careand Use of Laboratory Animals (permission no. 2004-003). Mice housedunder conventional conditions were anesthetized with an i.p. injection ofNembutal (1.5 mg/mouse) and their back was shaved. After the applicationof CpG-ODN or Rho-labeled CpG-ODN onto the shaved skin, the skin wasexcised at specific points in time, embedded in Tissue-Tek OTC compound(Miles), and snap-frozen in liquid nitrogen. The skin was then cut with acryostat into 10-�m vertical sections. To analyze the distribution of Rho-CpG-ODN, for the observation of skin hypertrophy, and for immune stain-ing for CD4�CD25� cells, the dermal skin was fixed with 10% formalin at4°C for 16 h, embedded, snap-frozen, and cut as described. The sectionswere examined by confocal laser scanning microscopy (MicroRadiance;Bio-Rad) or with an optical microscope following H&E staining and to-luidine blue staining. The number of infiltrated cells was discussed permicroscope field, and two skin sections from each mouse were used. Anti-CD4 mAb, anti-c-kit mAb, anti-foxp3 mAb, and anti-CD25 mAb, usingAlexa Fluor 488- and Alexa Fluor 594-conjugated secondary Ab, and nu-clear staining with 4�,6�-diamidino-2-phenylindole were used for immunestaining and the sections were examined by fluorescent microscope.
Cytokine determination
Mice housed under conventional conditions were anesthetized with an i.p.injection of Nembutal (1.5 mg/mouse) and their back was shaved. After theapplication of CpG-ODN (50 �g) or non-CpG-ODN (50 �g) onto theshaved skin, the skin was excised and homogenated with the Polytronhomogenizer in 1 ml of saline containing protease inhibitors (0.17 mg/mlPMSF, 0.02 mg/ml leupeptin, and 0.01 mg/ml aprotinin). The homogenatewas centrifuged at 2200 � g for 5 min and the supernatant was collected.The level of IL-12p70 in the skin was determined with a sandwich ELISAusing pairs of purified capture and biotinylated detection mAbs (all ob-tained from BD Pharmingen) recognizing murine IL-12p70, according tothe manufacturer’s directions.
Ab determination
Blood samples from mice treated with CpG-ODN (50 �g) or non-CpG-ODN (50 �g) every week from 11 to 18 wk were collected on specific daysfrom the retro-orbital plexus and sera were pooled. The levels of IgG1,IgG2a, and IgE in sera were determined with a sandwich ELISA usingpairs of purified capture and biotinylated detection mAbs (all obtainedfrom BD Pharmingen) recognizing murine IgG1, IgG2a, and IgE, accord-ing to the manufacturer’s instructions.
RT-PCR
The changes in cytokine and chemokine mRNA expression in lymph node(LN) and skin were determined by RT-PCR. Total RNA (1 �g) was iso-lated with Isogen solution (Nippon Gene) as reported previously (25).cDNA was synthesized using SuperScript III. Then, cDNAs were amplifiedwith primers specific for each cytokine. The primers are: IL-4 (forward)5�-AGTTGTCATCCTGCTCTTCTTT-3�, (reverse) 5�-GACTGGGACTCATTCATGGTGC-3�; IL-5 (forward) 5�-CAAAAAGAGAAGTGTGG
CGAGG-3�, (reverse) 5�-TAGATAGGAGCAGGAAGCCC-CG-3�; IL-10(forward) 5�-G-GACAACATACTGCTAACCGACTC-3�, (reverse) 5�-AAAATCACTCTTCACCTGCTC-CAC-3�; IL-12 p40 (forward) 5�-CAGAAGCTAACCATCTCCTGGTTTG-3�, (reverse) 5�-TCCGGAGTAATTTGGTGGTTCACAC-3�; IL-13 (forward) 5�-GCAACGGCAGCATGGTATGGAG-3�, (reverse) 5�-TGGTATAGGGGAGGCTGGAGAC-3�;IFN-� (forward) 5�-CTCAAGTGGCATAGATGT-3�, (reverse) 5�-GAGATAATCTGGCTCTGCAGGATT-3�; MDC (forward) 5�-TCTGATGCAGGTCCCTATGGT-3�, (reverse) 5�-TTATGGAGTAGCTTCTTCAC-3�; TARC, (forward) 5�-CAGGAAGTTGGTGAGCTGGTATA-3�,(reverse) 5�-TTGTGTTCGCCTGTAGTGCATA-3�; TNF-� (forward)
FIGURE 1. Th2-predominant immune responses in conventional(CONV) NC/Nga mice. a, The macroscopic appearance in NC/Nga micekept under conventional conditions. Blood samples from conventional NC/Nga mice were collected from the retro-orbital plexus, and sera werepooled. b, Serum IgE levels in conventional NC/Nga mice were determinedwith ELISA. c, The mRNA levels of cytokines in the draining LN inconventional NC/Nga mice were evaluated by RT-PCR. Each value rep-resents the mean � SD for five mice. ��, p � 0.01 compared with micekept under specific pathogen-free conditions.
FIGURE 2. Localization of CpG-ODN to the skin lesions of conven-tional (CONV) NC/Nga mice. Rhodamine-labeled CpG-ODN (50 �g)was applied to the skin and the localization of Rho-labeled CpG-ODNin the skin was determined at 12 h by confocal laser scanning micros-copy. High-density fluorescence of the skin is indicated (open arrows).
585The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
5�-CACCACGCTCTTCTGTCTACTGAAC-3�, (reverse) 5�-CCGGACTCCGTGATGTCTAAGTACT-3�; COX-2 (forward) 5�-CAGAACCGCATTGCCTCTG-3�, (reverse) 5�-TTGAAGGTGTCGGGCAGC-3�; TGF-�1 (for-ward) 5�-CTTTAGGAAGGACCTGG-GTT-3�, (reverse) 5�-CAGGAGGCGACAATCATGTT-3�; foxp3 (forward) 5�-CAGCTGCCTACAGTGCCCCTAG-3�, (reverse) 5�-CATTTGCCAGCAGTGGGTAG-3�; and�-actin (forward) 5�-GCACCACACCTTCTACAATGAG-3�, (reverse) 5�-TTGGCATAGAGGTCTTTACGGA-3�. �-Actin primers were used as an in-ternal control. Primers designed based on the mouse sequences were obtainedfrom Sigma-Genosys. �-Actin primers were used as an internal control.
Statistical analysis
The paired Student’s t test was used to compare paired groups. ANOVAwas used for multigroup analysis. Values of p � 0.05 were considered toindicate lack of significance.
ResultsTh2-predominant immune response in conventionalNC/Nga mice
When NC/Nga mice were kept under conventional conditions, theystarted to scratch themselves at �8 wk, and their skin became dryand scaly (21). Within the next several weeks, the mice developedlesions on the ears, back, neck, and face (Fig. 1a). In AD patients(3, 4), skin lesions show a chronically relapsing inflammatory dis-order with prurience and eczema usually associated with elevated
serum IgE levels and a Th2-predominant immune response. Inconventional NC/Nga mice, serum IgE levels were extremely high(Fig. 1b). mRNA levels of cytokines and chemokines in the drain-ing LN were evaluated by RT-PCR, and Th2-type cytokines andchemokines were expressed but not IFN-�, which is classified as aTh1-type cytokine as shown in Fig. 1c. From these findings, weconfirmed that the conventional NC/Nga mice used in this exper-iment show high Th2-like immune responses, and the up-regula-tion of IgE production regulated by Th2 cytokines IL-4 and IL-13is one of the major causes of atopic inflammation.
Localization of CpG-ODN in the skin of conventionalNC/Nga mice
In AD patients, a deficiency in the barrier function of the skin atthe lesional site was reported (26). Therefore environmental aller-gens, such as mites and house dusts, easily penetrate such suscep-tible skin. So we first examined the penetration of CpG-ODN bythe barrier-disrupted AD skin in conventional NC/Nga mice. Rho-damine-labeled CpG-ODN (Rho-CpG-ODN) was applied to theshaved back and its localization was examined by confocal laserscanning microscopy. In the skin of NC/Nga mice kept under spe-cific pathogen-free conditions, fluorescent signals for rhodaminewere observed faintly in the stratum corneum, suggesting that
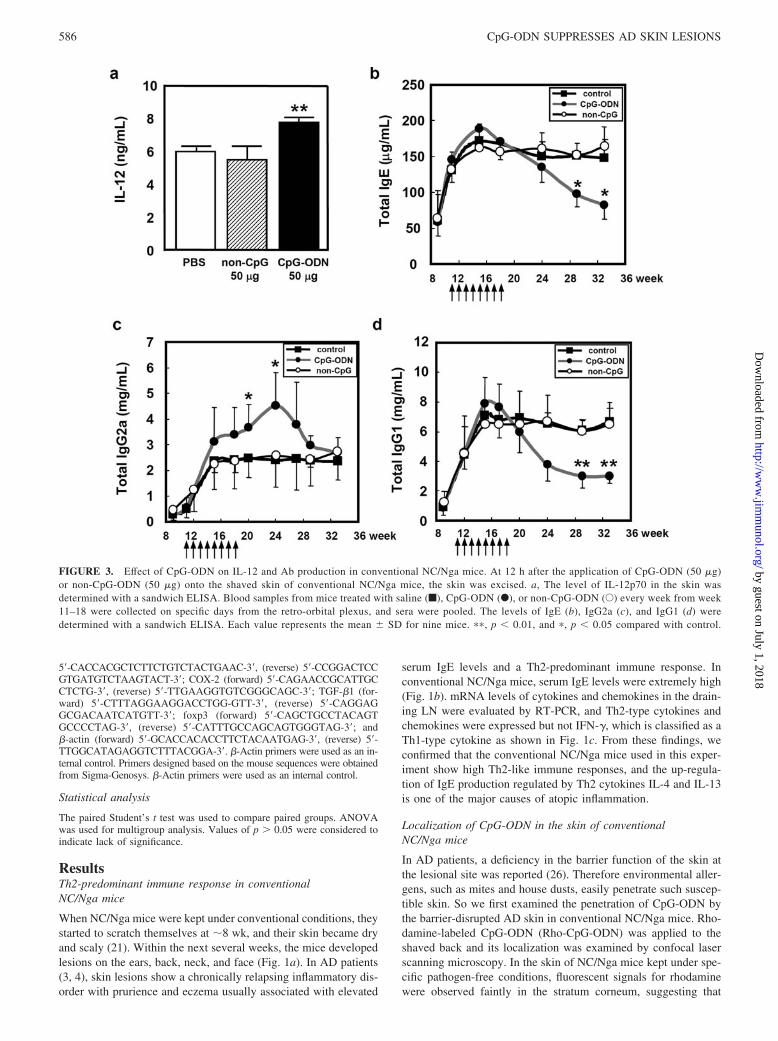
FIGURE 3. Effect of CpG-ODN on IL-12 and Ab production in conventional NC/Nga mice. At 12 h after the application of CpG-ODN (50 �g)or non-CpG-ODN (50 �g) onto the shaved skin of conventional NC/Nga mice, the skin was excised. a, The level of IL-12p70 in the skin wasdetermined with a sandwich ELISA. Blood samples from mice treated with saline (f), CpG-ODN (F), or non-CpG-ODN (E) every week from week11–18 were collected on specific days from the retro-orbital plexus, and sera were pooled. The levels of IgE (b), IgG2a (c), and IgG1 (d) weredetermined with a sandwich ELISA. Each value represents the mean � SD for nine mice. ��, p � 0.01, and �, p � 0.05 compared with control.
586 CpG-ODN SUPPRESSES AD SKIN LESIONS
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

CpG-ODN did not penetrate the s.c. layer (Fig. 2). In the conven-tional NC/Nga mice, however, red fluorescence generated fromrhodamine was observed around the corium at 12 h after the ap-plication, indicating that CpG-ODN, having a m.w. of �6000 anda negative charge, easily penetrated the skin. Interestingly, the highdensity of fluorescence in the skin was observed (Fig. 2, openarrows). From these findings, the barrier function of conventionalNC/Nga mouse skin was disrupted as in AD patients.
Effect of CpG-ODN on production of IL-12 and Abs inconventional NC/Nga mice
We next examine the effect of CpG-ODN on the production ofIL-12 at 12 h. CpG-ODN significantly enhanced the production ofIL-12 in the skin, which is known to induce Th1-predominant im-mune responses as shown in Fig. 3a. To examine the changes inlevels of Abs, 50 �g of CpG-ODN or non-CpG-ODN was appliedto the skin lesions every week eight times from week 11, which iswhen the lesions first develop. Total IgE levels decreased from 24wk and a significant difference from the control and non-CpG-ODN was observed at 28–33 wk (Fig. 3b). In contrast, IgG2alevels increased significantly at 20–24 wk in mice applied CpG-ODN (Fig. 3c). The total amount of IgG1 that is characterized asa Th2-type Ab decreased drastically as did the profile of IgE pro-duction (Fig. 3d). These results suggest that the application ofCpG-ODN changes immune responses in conventional NC/Ngamice, which show a Th2-predominant reaction.
FIGURE 4. Effect of CpG-ODNon cytokine mRNA expression inconventional NC/Nga mice. LN frommice treated with saline or CpG-ODN(50 �g) every week from week 11 to18 were collected at 33 wk, and thecytokine and chemokine mRNA ex-pression in LN was determined byRT-PCR analysis (a), and normalizedby �-actin levels (b). �-Actin primerswere used as an internal control.
FIGURE 5. Macroscopic observations of skin lesions in CpG-ODN-treatedNC/Nga mice. Conventional NC/Nga mice treated with saline, CpG-ODN(50 �g) or non-CpG-ODN (50 �g) every week from week 11 to 18 wereexamined at 33 wk. The number of eczematous lesions was counted.Each value represents the mean � SD for nine mice.
587The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
588 CpG-ODN SUPPRESSES AD SKIN LESIONS
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Changes in immune responses in CpG-ODN-treatedconventional NC/Nga mice
Next we examined the changes in mRNA levels of cytokines andchemokines in the draining lymph node (LN) at 33 wk becauseproduction of IgE decreased significantly compared with the controlat that time (Fig. 3). As shown in Fig. 4, mRNA levels of Th2-typecytokines and chemokines were decreased in CpG-ODN-treated NC/Nga mice. Conversely, mRNA levels of Th1-type cytokines, IFN-�and IL-12, increased slightly in CpG-ODN-treated NC/Nga mice.Interestingly, the expression of IFN-� mRNA in LN was also ob-served in untreated mice (control). mRNA levels of TNF-� andCOX-2, inflammatory markers, decreased in CpG-ODN-treated
mice. These findings suggest that transdermal application of CpG-ODN changes the immune response of NC/Nga, which is predom-inantly of type Th2. The decrease in IL-4 and IL-13 mRNA ex-pressions in CpG-ODN-treated mice is suppose to be a cause ofsuppression in IgE production. Interestingly, CpG-ODN did notinduce strong Th1 immune responses and inflammatory markerexpression in 33 wk.
Improvement of skin lesions in CpG-ODN-treated NC/Nga mice
In AD patients, the skin lesions show a chronically relapsing in-flammatory disorder with prurience and eczema (3, 4). We thusexamined the effect of CpG-ODN on the appearance of skin
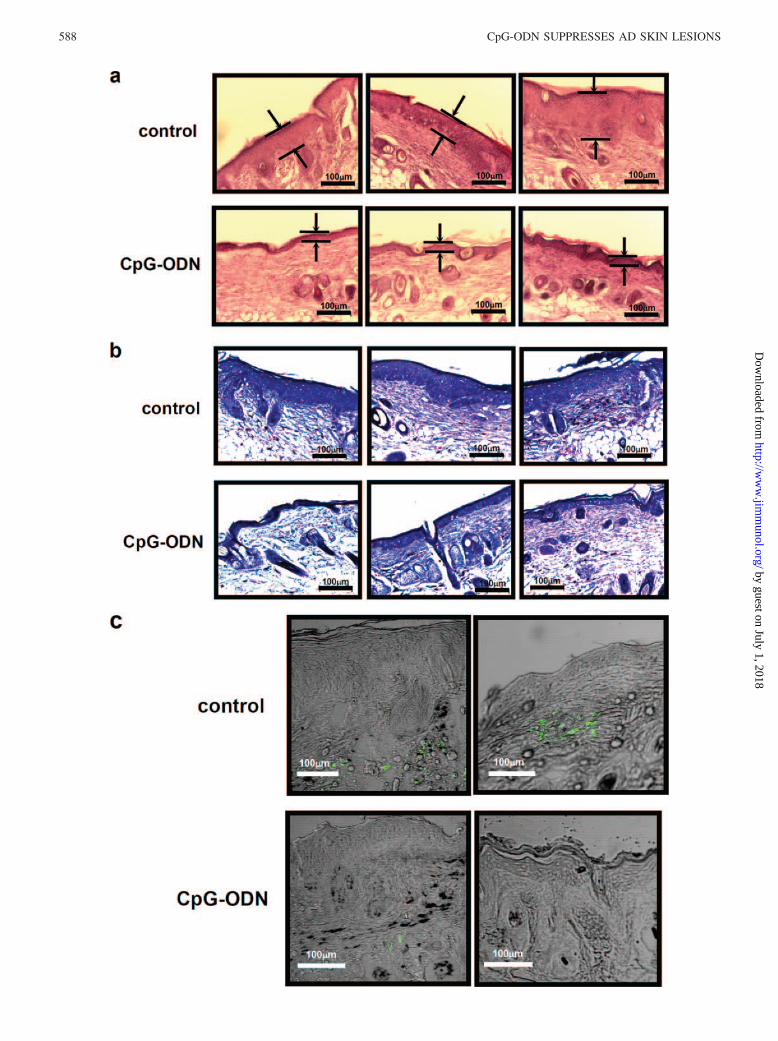
FIGURE 6. Histochemical examination of the lesions in CpG-ODN-treated NC/Nga mice. Skin sections from mice treated with saline or CpG-ODN (50�g) every week from week 11 to 18 were collected at 33 wk. Skin sections were stained with H&E (a), and truidin blue (b), a hypertrophy and mast cellinfiltration were examined with an optical microscope. c, Skin sections were also stained with anti-c-kit mAb (green) and then the localization offluorescence was examined with a fluorescence microscope.
FIGURE 7. Generation of Tregs in the skin of CpG-ODN-treated conventional NC/Nga mice. Skin from mice treated with saline or CpG-ODN (50 �g)every week from week 11 to 18 were collected at 33 wk. a, The TGF-�, IL-10, foxp3, and COX-2 mRNA expression in the skin was determined by RT-PCRanalysis. b and c, Skin sections were also stained with anti-CD4 mAb (green), anti-foxp3 mAb (green), anti-CD25 mAb (red), and 4�,6�-diamidino-2-phenylindole (blue), and the localization of fluorescence was examined with a fluorescent microscope. White arrows in b show CD4�CD25� double-positive cells, white arrows in c show foxp3� cells.
589The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

lesions in conventional NC/Nga mice. In untreated NC/Nga mice(control) and non-CpG-ODN-treated mice, eczematous injury wasobserved on the back, which resembles typical features of AD.Compared with control mice and non-CpG-ODN-treated mice, adrastic decrease in these injuries was observed in CpG-ODN-treated mice (Fig. 5). The number of these eczematous lesions wassignificantly decreased and 18.0 � 6.23 for control, 18.25 � 4.62for non-CpG-ODN-treated mice, and 6.13 � 4.19 for CpG-ODN-treated mice ( p � 0.05 compared with control and non-CpG-ODN-treated mice), respectively.
Improvements in epidermal skin lesions and mast cellsinfiltration to the skin in CpG-ODN-treated NC/Nga mice
Histochemical examination of the lesions in conventional NC/Ngamice reveals hyperkeratosis, acanthosis, and parakeratosis, all ofwhich are typical features of the skin of patients with AD. The infil-tration of lymphocytes, macrophages, mast cells and eosinophils wasobserved in the skin lesions. To analyze the effect of CpG-ODN onskin hypertrophy and granulocyte infiltration in conventional NC/Ngamice, CpG-ODN-treated skin was stained with H&E and truidin blue,and then examined with an optical microscope. As shown in Fig. 6a,the acanthosis was clearly suppressed in CpG-ODN-treated micecompared with control mice. Also, granulocyte infiltration, stained asred dots in Fig. 6b, was decreased in the CpG-ODN-treated mice. Thenumber of infiltrated cells was 36.33 � 15.86 for the control mice and8.72 � 3.59 for the CpG-ODN-treated mice ( p � 0.05 compared withcontrol). Skin sections were also stained with anti-c-kit mAb (green)and the localization of fluorescence was examined with a fluorescencemicroscope. As shown in Fig. 6c, mast cell infiltration was also de-creased in the CpG-ODN-treated mice. These results indicated thatCpG-ODN changes not only the Th2-predominant immune responsein conventional NC/Nga mice but also in skin conditions, skin hyper-trophy, and mast cell infiltration.
Generation of Tregs in CpG-ODN-treated skin
To clarify the mechanisms of this effect of CpG-ODN, we first fo-cused on the change in cytokine levels of the skin. mRNA levels ofTGF-� and IL-10, which are cytokines considered to suppress inflam-mation especially in immune diseases, increased following transder-mal application of CpG-ODN (Fig. 7a). In contrast, the mRNAexpression of COX-2, a marker of inflammation, decreased. Further-more, we examined the mRNA expression of foxp3, a marker ofTregs, because Tregs produce immune suppressive cytokines TGF-�and IL-10. As shown in Fig. 7a, foxp3 was expressed in the CpG-ODN-treated skin, which also expressed TGF-� and IL-10. Thesefindings suggest that the up-regulation of TGF-� and IL-10 mRNAexpression results in Tregs being generated in the skin. Second, weanalyzed the induction and migration of Tregs into the skin. Skinsections were stained with anti-CD4 mAb (green) and anti-CD25mAb (red), a marker of Treg, and then examined with a fluorescentmicroscope. As shown in Fig. 7b, CD4� cells were detected in bothcontrol skin and CpG-ODN-treated skin as green fluorescence. Incontrast, CD4�CD25� double-positive cells were observed only inthe CpG-ODN-treated skin (Fig. 7b, white arrows). Skin sectionswere also stained with anti-foxp3 mAb (green) and foxp3� cells weredetected only in the CpG-ODN-treated skin (Fig. 7c, white arrows).These results indicated that CpG-ODN induces the induction and mi-gration of Tregs in the skin of conventional NC/Nga mice.
DiscussionThe ability of CpG-ODN to shift the immune response from typeTh2 to Th1 is expected to be used in immunotherapy for patientswith diseases like AD and asthma that are associated with a Th2-predominant immune response (27–29). Youn et al. (30) reported
that CpG-ODN reverses established allergen-induced airway re-modeling. It is also reported that CpG-ODN inhibits establishedTh2-type immune responses through production of IFN-� andIL-10 (31). In a model of AD, Kootiratrakarn et al. (32) showedthat CpG-ODN could improve an established eosinophilic inflam-mation in the skin. These studies suggested that CpG-ODN is apotent inducer of Th1 and improves allergic responses even inestablished Th2-type response models. Recently, it has been re-ported that CpG-ODN-activated dendritic cells induce the gener-ation of Tregs with a strong immune suppressive function (19). Inanother study, blocking the suppressor activity of Tregs and in-creasing Th2 cell frequency enhanced allergen-specific Th2 cellactivation ex vivo (14). We thus investigated whether the topicalapplication of CpG-ODN to conventional NC/Nga mice, a fre-quently used model of human AD with Th2-predominant immuneresponses, changes the immune response from type-Th2 to Th1and induces the generation of Tregs.
First we examined the penetration of CpG-ODN from the bar-rier-disrupted AD skin in conventional NC/Nga mice, and the pen-etration of CpG-ODN and the high density of fluorescence in theskin was observed (Fig. 2). We have reported that administrationof CpG-ODN through the barrier-disrupted skin also showed thelocalization of CpG-ODN in the skin, and the time-dependent mi-gration into the lower part of the skin was observed (24). Weexpected that dendritic cells may be the one of the reasons of thislocalization and may be connected with the immune responses. Infact, the application of CpG-ODN through the barrier-disruptedskin up-regulates the migration of CD11c� cell to LNs (data notshown). Thus, the activation of epidermal and dermal dendriticcells by CpG-ODN may be the one of the mechanisms of thechanging in the immune responses.
Our examination showed that the application of CpG-ODNchanged the immune response from type Th2 to type Th1 on thebasis of the Ig isotype in the sera (Fig. 3, b–d) and expression ofcytokine mRNA in LN (Fig. 4). Total IgE and IgG1 levels weredecreased from 24 wk and a significant difference was observed at28–33 wk (Fig. 3, b and d). In contrast, production of IgG2a wassignificantly increased at 20–24 wk (Fig. 3c). Interestingly, the in-crease in IgG2a production (20 wk) was slightly faster than the de-crease in IgE and IgG1 production (24 wk) (Fig. 3, b–d) and the levelof IgG2a returned to the control value from 24 wk. These findingsindicated that the Th1-type immune response was up-regulated byCpG-ODN until 24 wk (“early phase”), and also the balance ofTh1/Th2 was changed at this time. After 24 wk, these immuneresponses may be all suppressed with this balance maintained(“late phase”). Recently, it was reported that Tregs act as suppres-sor T cells that down-regulate other effecter T cells and inflam-mation. This down-regulation in “late phase” may be the result inthe induction of Tregs.
However, the Th1 cytokine IFN-� is found in the skin lesions atthe chronic stage of the disease and Th1 cells cause a deteriorationof disease at this stage (2). In our study, control animals will be inthe chronic stage at 33 wk because IFN-� was expressed in themice (Fig. 4). It is still unclear whether CpG-ODN heals the skinat chronic stages of the disease. Peng et al. (33) reported that CpG-ODN vaccinations may fail to down-regulate ongoing IgE re-sponses and the application induces a transient skin delayed-typehypersensitivity with production of IFN-�. Moreover, STAT-6-deficient NC/Nga mice, which fail to produce IgE and Th2 cyto-kines and highly express IFN-� in skin lesions, developed AD-likeskin lesions at an equivalent frequency to normal NC/Nga litter-mates (34). But another report suggested that CpG-ODN couldreserve the established eosinophilic inflammation in the skin with
590 CpG-ODN SUPPRESSES AD SKIN LESIONS
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

IFN-� production (32). The mechanisms of the effect of CpG-ODN on established Th2-immune responses are still unclear;therefore, further investigation is needed in chronically affectedNC/Nga mice.
Macroscopic observation and histochemical examination re-vealed an improvement of the AD-like skin lesions in CpG-ODN-treated NC/Nga mice (Figs. 5 and 6). The clear expression of sup-pressive cytokines, TGF-� and IL-10, and generation of Tregs inthe CpG-ODN-treated skin were also observed (Fig. 7). It is reportedthat Tregs develop mainly in the presence of IL-10 and TGF-� (35).We have examined IL-10 production in the skin and LN, and ob-served an up-regulation in CpG-ODN-treated skin and LN (data notshown). It is also reported in NC/Nga mice that the s.c. injection ofrecombinant TGF-� suppressed eczematous skin lesions associ-ated with reduced serum IgE levels, and histological analysisshowed that TGF-� inhibited the infiltration of inflammatory cellssuch as mast cells and eosinophils into the skin (36). Furthermore,many studies have shown that CCR2, CCR4, CCR5, CCR6, E-selectin and P-selectin ligands, and �1 integrin were important forthe homing of Tregs to various peripheral sites (37). CpG-ODNinduces the production of various chemokines, ligands of thesereceptors, such as MCP-1, eotaxin, RANTES, TARC, and MIP-1and MIP-3, from APCs and keratinocytes (15, 16, 38, 39). Thesefindings strongly support our results and suggest that topically ap-plied CpG-ODN is a potent generator of Tregs in the skin.
The current findings are the first demonstration of treating ADwith CpG-ODN as the skin lesions first develop in NC/Nga mice.In summary, CpG-ODN remarkably changed the immune re-sponses. The serum IgE level was decreased and the production ofIgG2a was up-regulated. CpG-ODN also improved the skin lesionsand the inflammatory infiltration of mast cells decreased with thegeneration of Tregs in the skin. These results suggested that CpG-ODN is effective for immunotherapy in patients with AD, which ischaracterized by Th2-dominated inflammation. Additional studiesare required to determine whether clinical trials that use CpG-ODN reduce serum IgE levels and improve skin lesions with thegeneration of Tregs.
AcknowledgmentsWe are grateful to T. Yoshida, H. Kumamoto, M. Ishida, K. Kunii, andY. Saigo for technical assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Nakamura, H., M. Aoki, K. Tamai, M. Oishi, T. Ogihara, Y. Kaneda, and
R. Morishita. 2002. Prevention and regression of atopic dermatitis by ointmentcontaining NF-�B decoy oligodeoxynucleotides in NC/Nga atopic mouse model.Gene Ther. 9: 1221–1229.
2. Leung, D. Y., M. Boguniewicz, M. D. Howell, I. Nomura, and Q. A. Hamid.2004. New insights into atopic dermatitis. J. Clin. Invest. 113: 651–657.
3. Cooper, K. D. 1994. Atopic dermatitis: recent trends in pathogenesis and therapy.J. Invest. Dermatol. 102: 128–137.
4. Rudikoff, D., and M. Lebwohl. 1998. Atopic dermatitis. Lancet 351: 1715–1721.5. Soter, N. A. 1989. Morphology of atopic eczema. Allergy 44(Suppl. 9): 16–19.6. Uehara, M., R. Izukura, and T. Sawai. 1990. Blood eosinophilia in atopic der-
matitis. Clin. Exp. Dermatol. 15: 264–266.7. Grewe, M., C. A. Bruijnzeel-Koomen, E. Schopf, T. Thepen, A. G. Langeveld-
Wildschut, T. Ruzicka, and J. Krutmann. 1998. A role for Th1 and Th2 cells in theimmunopathogenesis of atopic dermatitis. Immunol. Today 19: 359–361.
8. Spergel, J. M., E. Mizoguchi, H. Oettgen, A. K. Bhan, and R. S. Geha. 1999.Roles of TH1 and TH2 cytokines in a murine model of allergic dermatitis. J. Clin.Invest. 103: 1103–1111.
9. Vestergaard, C., H. Yoneyama, M. Murai, K. Nakamura, K. Tamaki, Y.Terashima, T. Imai, O. Yoshie, T. Irimura, H. Mizutani, and K. Matsushima.1999. Overproduction of Th2-specific chemokines in NC/Nga mice exhibitingatopic dermatitis-like lesions. J. Clin. Invest. 104: 1097–1105.
10. van Bever, H. P. 1992. Recent advances in the pathogenesis of atopic dermatitis.Eur. J. Pediatr. 151: 870–873.
11. Nakayama, H. 1996. Atopic dermatitis. Dermatology 38: 484.12. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell de-
velopment by the transcription factor foxp3. Science 299: 1057–1061.13. Hori, S., T. Takahashi, and S. Sakaguchi. 2003. Control of autoimmunity by
naturally arising regulatory CD4� T cells. Adv. Immunol. 81: 331–371.14. Stassen, M., H. Jonuleit, C. Muller, M. Klein, C. Richter, T. Bopp, S. Schmitt,
and E. Schmitt. 2004. Differential regulatory capacity of CD25� T regulatorycells and preactivated CD25� T regulatory cells on development, functional ac-tivation, and proliferation of Th2 cells. J. Immunol. 173: 267–274.
15. Krieg, A. M. 2002. CpG motifs in bacterial DNA and their immune effects. Annu.Rev. Immunol. 20: 709–760.
16. Agrawal, S., and E. R. Kandimalla. 2003. Modulation of Toll-like receptor 9responses through synthetic immunostimulatory motifs of DNA. Ann. NY Acad.Sci. 1002: 30–42.
17. Hemmi, H., T. Kaisho, K. Takeda, and S. Akira. 2003. The roles of Toll-likereceptor 9, MyD88, and DNA-dependent protein kinase catalytic subunit in theeffects of two distinct CpG DNAs on dendritic cell subsets. J. Immunol. 170:3059–3064.
18. Klinman, D. M., D. Currie, I. Gursel, and D. Verthelyi. 2004. Use of CpG oli-godeoxynucleotides as immune adjuvants. Immunol. Rev. 199: 201–216.
19. Moseman, E. A., X. Liang, A. J. Dawson, A. Panoskaltsis-Mortari, A. M. Krieg,Y. J. Liu, B. R. Blazar, and W. Chen. 2004. Human plasmacytoid dendritic cellsactivated by CpG oligodeoxynucleotides induce the generation of CD4�CD25�
regulatory T cells. J. Immunol. 173: 4433–4442.20. Kondo, K., T. Nagami, and S. Todokoro. 1969. Differences in haematopoietic
death among inbred strains of mice. In Comparative Cellular and Species Radi-osensitivity. P. V. Bond, and T. Sugawara, eds. Igakusyoin, Tokyo, p. 20.
21. Vestergaard, C., H. Yoneyama, and K. Matsushima. 2000. The NC/Nga mouse:a model for atopic dermatitis. Mol. Med. Today 6: 209–210.
22. Matsuda, H., N. Watanabe, G. P. Geba, J. Sperl, M. Tsudzuki, J. Hiroi,M. Matsumoto, H. Ushio, S. Saito, P. W. Askenase, and C. Ra. 1997. Develop-ment of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Ngamice. Int. Immunol. 9: 461–466.
23. Inoue, J., S. Yotsumoto, T. Sakamoto, S. Tsuchiya, and Y. Aramaki. 2005.Changes in immune responses to antigen applied to tape-stripped skin with CpG-oligodeoxynucleotide in mice. J. Control. Release 108: 294–305.
24. Inoue, J., S. Yotsumoto, T. Sakamoto, S. Tsuchiya, and Y. Aramaki. 2005.Changes in immune responses to antigen applied to tape-stripped skin with CpG-oligodeoxynucleotide in NC/Nga mice. Pharm. Res. 22: 1627–1633.
25. Sakamoto, T., E. Miyazaki, Y. Aramaki, H. Arima, M. Takahashi, Y. Kato,M. Koga, and S. Tsuchiya. 2004. Improvement of dermatitis by iontophoreticallydelivered antisense oligonucleotides for interleukin-10 in NC/Nga mice. GeneTher. 11: 317–324.
26. Imokawa, G., A. Abe, K. Jin, Y. Higaki, M. Kawashima, and A. Hidano. 1991.Decreased level of ceramides in stratum corneum of atopic dermatitis: an etio-logic factor in atopic dry skin? J. Invest. Dermatol. 96: 523–526.
27. Kline, J. N., T. J. Waldschmidt, T. R. Businga, J. E. Lemish, J. V. Weinstock,P. S. Thorne, and A. M. Krieg. 1998. Modulation of airway inflammation by CpGoligodeoxynucleotides in a murine model of asthma. J. Immunol. 160: 2555–2559.
28. Kline, J. N., K. Kitagaki, T. R. Businga, and V. V. Jain. 2002. Treatment ofestablished asthma in a murine model using CpG oligodeoxynucleotides.Am. J. Physiol. 283: L170–L179.
29. Hussain, I., and J. N. Kline. 2003. CpG oligodeoxynucleotides: a novel thera-peutic approach for atopic disorders. Curr. Drug Targets Inflamm. Allergy 2:199–205.
30. Youn, C. J., M. Miller, K. J. Baek, J. W. Han, J. Nayar, S. Y. Lee, K. McElwain,S. McElwain, E. Raz, and D. H. Broide. 2004. Immunostimulatory DNA reversesestablished allergen-induced airway remodeling. J. Immunol. 173: 7556–7564.
31. Kitagaki, K., V. V. Jain, T. R. Businga, I. Hussain, and J. N. Kline. 2002. Im-munomodulatory effects of CpG oligodeoxynucleotides on established th2 re-sponses. Clin. Diagn. Lab. Immunol. 9: 1260–1269.
32. Kootiratrakarn, T., T. Fujimura, K. Sano, R. Okuyama, S. Aiba, H. Tagami, andT. Terui. 2005. Development of a novel Ag-specific immunotherapy using CpGoligodeoxynucleotides in a new, unique mouse cutaneous eosinophilic inflam-mation model. Eur. J. Immunol. 35: 3277–3286.
33. Peng, Z., H. Wang, X. Mao, K. T. HayGlass, and F. E. Simons. 2001. CpGoligodeoxynucleotide vaccination suppresses IgE induction but may fail to down-regulate ongoing IgE responses in mice. Int. Immunol. 13: 3–11.
34. Yagi, R., H. Nagai, Y. Iigo, T. Akimoto, T. Arai, and M. Kubo. 2002. Devel-opment of atopic dermatitis-like skin lesions in STAT6-deficient NC/Nga mice.J. Immunol. 168: 2020–2027.
35. Horwitz, D. A., S. G. Zheng, and J. D. Gray. 2003. The role of the combinationof IL-2 and TGF-� or IL-10 in the generation and function of CD4�CD25� andCD8� regulatory T cell subsets. J. Leukocyte Biol. 74: 471–478.
36. Sumiyoshi, K., A. Nakao, H. Ushio, K. Mitsuishi, K. Okumura, R. Tsuboi, C. Ra,and H. Ogawa. 2002. Transforming growth factor-�1 suppresses atopic derma-titis-like skin lesions in NC/Nga mice. Clin. Exp. Allergy 32: 309–314.
37. Huehn, J., and A. Hamann. Homing to suppress: address codes for Treg migra-tion. 2005. Trends Immunol. 26: 632–636.
38. Takeshita, S., F. Takeshita, D. E. Haddad, K. J. Ishii, and D. M. Klinman. 2000.CpG oligodeoxynucleotides induce murine macrophages to up-regulate chemo-kine mRNA expression. Cell. Immunol. 206: 101–106.
39. Miller, L. S., O. E. Sorensen, P. T. Liu, H. R. Jalian, D. Eshtiaghpour,B. E. Behmanesh, W. Chung, T. D. Starner, J. Kim, P. A. Sieling, et al. 2005.TGF-� regulates TLR expression and function on epidermal keratinocytes. J. Im-munol. 174: 6137–6143.
591The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























