Embed Size (px)
Citation preview

Kidney International, Vol. 50 (1996), pp. 1212—1223
Superinduction of IL-6 synthesis in human peritoneal mesothelialcells is related to the induction and stabilization of IL-6 mRNA
JANUSZ WITowsKI, ACHIM JORRES, GERALD A. COLES, JOHN D. WILLIAMs, and NICHOLAS TOPLEY
Institute of Nephrolo, University of Wales College of Medicine, and Cardiff Royal Infirma,y, c'ardiff Wales, United Kingdom; and Abteilung für InnereMedizin mit Schwerpunkt Nephrologie und Intemistische Intensivmedizin, Virchow-Klinikum, Humboldt Universität zu Berlin, Berlin, Germany
Superinduction of IL-6 synthesis in human peritoneal mesothelial cellsis related to the induction and stabilization of IL-6 mRNA. The initiationof peritonitis is accompanied by a massive increase in intraperitoneallevels of IL-6. The source of this cytokinc and the mechanism by which itslevels are increased so dramatically are unknown. We examined themechanism of IL-6 secretion by HPMC exposed to the milieu present inthe peritoneal cavity during the initiation of inflammation. Exposure ofHPMC to spent peritoneal dialysis effluent (PDE) or IL-113 resulted in atime- and dose-dependent increase in IL-6 secretion. After 24 hours theIL-6 release (pg4g cell protein) was increased from 5.0 0.8 in controlcells to 16.0 2.4 and to 83.8 17.4 in HPMC treated with PDE andIL-lp (1000 pg/mI), respectively (N = 5, P < 0.05). If, however, PDE andIL-113 were combined, then the secretion of IL-6 was synergisticallyincreased to 747.7 349.9 (P < 0.05 vs. expected additive value). Thesame effect was evident when PDE was combined with TNFcs or mixedwith peritoneal macrophage conditioned medium. These changes were nota reflection of HPMC proliferation as estimated by 3H-thymidine incor-poration. The "superinduction" of IL-6 release was associated both withthe induction and stabilization of IL-6 mRNA. Competitive PCR demon-strated that the amount of IL-6 mRNA (fM/jLg total RNA) was increasedfrom 0.35 0.13 in control cells to 11.66 3.89 and to 10.83 5.17 inHPMC treated with PDE and IL-I j3 (100 pg/mI), respectively (N = 5, P <0.05). The combination of PDE + IL-113 synergistically increased IL-6mRNA levels to 56.33 8.79 (P < 0.05 vs. additive value). RNA stabilityexperiments using actinomycin D revealed that the half life of IL-fl mRNA(hours) was increased from 2.8 hours in control cells to 6.7 and 9.4 inHPMC exposed to PDE and IL-I 13, respectively. The combination of PDEtogether with IL-1$ resulted in a prolonged stabilization of IL-6 mRNAwith levels remaining constant over the 12 hours of the experiment. Thesedata demonstrate that HPMC IL-fl synthesis can be synergistically ampli-fied in the presence of peritoneal dialysis effluent and PM0-derivedcytokines. The results suggest that the peritoneal mesothelium may beresponsible for the dramatic rise in IL-6 levels seen during the initialstages of CAPD peritonitis.
Continuous ambulatory peritoneal dialysis (CAPD) has becomean established form of therapy for patients with end-stage renalinsufficiency. The successful outcome of APD depends largelyon the long-term preservation of the peritoneal membrane as adialyzing organ. The integrity of the peritoneal membrane may,however, be easily jeopardized by peritonitis, which still remains amajor complication in patients undergoing CAPD. In severe cases
Received for publication March 21, 1996and in revised form April 30, 1996Accepted for publication May 2, 1996
© 1996 by the International Society of Nephrology
the consequence of repeated episodes of peritonitis may beirreparable damage to the peritoneal membrane, loss of ultrafil-tration, and treatment failure [1j.
Examination of intraperitoneal levels of inflammatory media-tors prior to and during peritonitis have revealed that the intra-peritoneal levels of prostaglandins (PGE7 and 6-keto-PGF1,) andeytokines (IL-i, TNFa, IL-6, IL-8, and IFN-y) are significantlyelevated during acute phase of peritonitis and subsequently returnto basal values when the inflammatory process subsides [2—9].Moreover, it has been demonstrated that this rise (specifically inTNFcs levels) may even precede the occurrence of clinical signs ofinfection [5, 101. Thus the specific time courses of cytokineappearance in peritoneal dialysis effluent may identify differentstages of the intraperitoneal inflammatory response.
In 1990 Goldman et al [6] and then several other groups [7,11—131 observed that the initiation of peritonitis in CAPD patientswas accompanied by a massive increase (100- to 10,000-fold) inintraperitoneal levels of interleukin-6 (IL-6). The source of thiscytokine, its function within peritoneal cavity, and the mechanismby which its levels increased so dramatically have not been fullyidentified. The observation that the JL-6 dialysate/serum ratios inperitonitis CAPD patients were much higher than could hepredicted on the basis of simple diffusion of IL-6 from circulationto the peritoneal cavity provided evidence that IL-6 might beproduced intraperitoneally [14]. This focused attention on perito-neal macrophages (PM0), which are commonly viewed as the firstline of defence against invading microorganisms and as such arebelieved to be a major source of inflammatory mediators initiatingthe inflammatory response. It has been demonstrated that PMØfrom infection-free CAPD patients release significant quantitiesof eicosanoids and are capable of generating considerableamounts of cytokines when challenged with S. epidermidis isolates[15, 161.
More recently, however, increasing evidence suggests that thereis another likely candidate as a potent source of intraperitonealIL-6. The peritoneal mesothelium has been shown to act not onlyas an inert lining of the peritoneal cavity, but also to possess thecapacity to produce cytokines. It has been demonstrated thatcultured human peritoneal mesothelial cells (HPMC) secretebiologically active IL-6, and that the IL-6 synthesis can bemarkedly augmented in response to pro-inflammatory cytokinesIL-113 and TNFcs as a result of specific induction of IL-6 mRNA[17].
1212
In this study we aimed to characterize the mechanism of IL-6

Witowski et al: Superinduction of peritoneal IL-6 synthesis 1213
secretion by HPMC at both the protein and mRNA levels in themilieu mimicking that of peritoneal inflammation. Our datademonstrate that HPMC IL-6 synthesis can be synergisticallyamplified in the environment consisting of spent peritoneal dia-lysate and PM0-derived cytokines. This observation indicates thatthe peritoneal mesothelium may be responsible for the dramaticrise in IL-6 levels seen during the initial stages of peritonitis.
Methods
All chemicals, unless otherwise stated, were purchased from theSigma Chemical Company (Poole, Dorset, UK). Recombinanthuman IL-1j3 was obtained from Advanced Protein Products Ltd(Brierley Hill, UK). Its biological activity (expressed as ED50) wasbelow 0.1 ng/ml as determined by the uptake of thymidine bymurine C3H/HeJ thymocytes. Human TNFs was obtained fromBASF AG (Ludwigshafen, Germany). Its specific activity was 8 Xi0 U/mg protein as assessed in a 48 hour L929 bioassay in theabsence of actinomycin D. Human recombinant IL-I receptorantagonist (IL-Ira) was kindly provided by Dr. P. Scholtz (Scher-ing AG, Berlin, Germany). All cytokine preparations werebatched and stored at —70°C and freshly thawed for eachexperiment. Endotoxin contamination of recombinant materialwas below 0.7 pg/sg protein as assessed by limulus amoebocytelysate assay (Kabi Vitrum, Stockholm, Sweden). All tissue cultureflasks and multi-well plates were obtained from Falcon (Becton-Dickinson UK Ltd, Oxford, UK).
Isolation and culture of human peritoneal mesothelial cells
Human peritoneal mesothelial cells (HPMC) were obtainedfrom the omental tissue of consenting non-uremic patients under-going elective abdominal surgery. The cells were isolated andcharacterized essentially as described previously [18]. The identityof mesothelial cells was confirmed by their uniform cobblestoneappearance at confluence, by the presence of surface microvilli,and by the uniform positive staining for cytokeratins 8 and 18. Theendothelial origin of the cells cultured was excluded by the lack ofstaining for the factor VIJI-related antigen and by the absence ofendothelin-1 secretion [19]. The presence of contaminating mac-rophages was excluded following assessment of Fe receptorexpression (using a red cell rosetting assay that was negative in allmesothelial cell cultures examined). Cells were maintained in theEarle's buffered M 199 culture medium (Sera Lab/JRH Bio-sciences Ltd, Crawley, UK) supplemented with penicillin (100U/mI), streptomycin (100 sg/ml), L-glutamine (2 mM) (GibcoBRL, Life Technologies Ltd, Paisley, UK), transferriri (5 JLg/ml),insulin (5 sg/ml), hydrocortisone (0.4 jtg/ml) (all from Sigma) and10% vol/vol fetal calf serum (FCS) (Sera Lab Ltd). HPMCcultures were maintained at 37°C in a humidified 5% CO2atmosphere.
Cells were passaged using a trypsin:EDTA solution (0.125%wt/vol:0.01% wt/vol). All data presented are from experimentsperformed with cells from the second passage that had previouslybeen growth arrested as described below. Cells used were fromthe second passage and not later to minimize the number ofsenescent cells that appeared from the third passage onwards [17,18].
Establishment of growth anested HPMC
After HPMC reached confluence in multi-well plates, the 10%FCS-containing medium was removed and cells were washed
three times with the culture medium containing 0.1% vol/vol FCS(rest medium). Then HPMC were incubated in the rest mediumfor 48 hours prior to stimulation. Previous experiments haddemonstrated that at this time point the cells were maintained ina viable (as assessed by the lack of LDH release and constantcellular ATP levels) but non-proliferative state (data not shown).Under these conditions the cells could be maintained for up to 96hours without any significant loss of viability [17].
Peritoneal dialysis effluentThe samples of peritoneal dialysis effluent (PDE) were ob-
tained from consenting non-infected CAPD patients (N = 9)using the following technique. Four hours after intraperitonealinfusion of 2.0 liters of fresh peritoneal dialysis solution (BaxterHealthcare Ltd, Thetford, Norfolk, UK), containing 1.36% wt/volglucose and no other additives, the spent dialysate was drainedinto the dialysis bag. Then the effluent was transferred to steriletubes, immediately centrifuged (4000 X g for 30 mm) and filteredthrough 0.2 jsm pore size filter (Minisart, Sartorius Ltd, Epsom,UK) to remove any cellular material. Equal volumes of samplesfrom individual patients were pooled and stored in aliquots at—70°C until required.
Peritoneal macrophage (PMØ) conditioned medium
PMØ were harvested from peritoneal dialysis effluent of infec-tion-free CAPD patients as described in detail elsewhere [151.More than 95% of the cells remaining adherent to the Petri dishes(Becton-Dickinson UK Ltd) were consistently found to be PMØas assessed morphologically following staining with the NeatDifferential Haematology Stain (Guest Medical Ltd., Sevenoaks,UK). Conditioned media were collected from PMØ culturesfollowing a three hour incubation in the culture medium alone(PM0-CM) or from PMØ stimulated with a strain of Staphylo-coccus epidermidis (S. epidermidis DM), isolated from the perito-neal dialysis effluent of a CAPD patient with peritonitis (S.epi/PMØ-CM), as previously described [16].
Induction of IL-6 production by HPMCHPMC were grown to confluence in 24-well plates and growth
arrested as previously described. Then cells were washed threetimes with the rest medium and incubated for 24 hours at 37°Cwith either the control medium (containing 0.1% FCS) or pooledPDE, both in the presence or absence of IL-113. In separateexperiments HPMC were exposed to the peritoneal macrophageconditioned-media (PM0-CM or S.epiPMø-CM) combined witheither the control medium or PDE. In the inhibition studiesHPMC were pre-treated with the transcription inhibitor actino-mycin D (Sigma) for 30 minutes at 37°C prior to stimulation.
At designated time intervals the HPMC supernatants wereremoved, centrifuged at 12000 >< g and stored at —70°C untilassayed. The cells were washed twice with phosphate bufferedsaline, pH 7.3 (PBS Dulbecco's; Gibco BRL, Life TechnologiesLtd, Paisley, UK) and solubilized with 0.1 N NaOI-I. Total cellularprotein was then analyzed by the modified Bradford method [201using bovine serum albumin (Sigma) as the standard. Repeatedcell counts revealed that 1 jig of cellular protein was equivalent to3.76 0.56 X i0 cells (N = 20). All data for JL-6 production areexpressed as pg/jig of cellular protein. In all experiments the PDE(and the respective control medium) was supplemented withheparin to a final concentration of 5 U/mI to avoid the clot
1214 Witowski et a!: Superinduction of peritoneal IL-6 synthesis
Table 1.
F: 5'-GGAGCAATGATCTTGATC11T-3R: 5 -TCCTGAGGTACGGGTCCTTCC-3'F: 5'-TACATCCTCGACGGCATCTC-3'R: 5'-GCTACAmGCCGAAGAGCC-3'F: 5 '-TACATCCTCGACGGCATCTCGCCCCAGCCACCCATITGTA-3'R: 5 '-GCTACATTTGCCGAAGAGCCGCTCCCTGCCCAVTTCTGTC-3'
formation. Incubation of HPMC with recombinant cytokines,PDE or PM0-conditioned media for up to 48 hours did notreduce cell viability as assessed by the LDH release (data notshown).
IL-6 production measurements
The concentration of JL-6 in HPMC supernatants was mea-sured using a specific sandwich-ELISA as previously described[211. The specific release of IL-6 from HPMC was calculatedfollowing the subtraction of background IL-6 levels detected ineither control medium, PDE, PM0-CM or S.epiPMø-CM fromthose recorded in the postculture supernatants.
RNA isolation, reverse transcription (RT) and polymerase chainreaction (FCR)
HPMC were grown to confluence in six-well plates and ren-dered quiescent as previously described. Then cells were washedthree times with a rest medium and exposed to either the controlmedium (containing 0.1% FCS) or PDE and incubated in thepresence or absence of 1L-l (100 pg/mi) for up to 24 hours. Atdefined time periods total cellular RNA was extracted fromHPMC following lysis with 4 M guanidine isothiocyanate andcentrifugation through 5.7 M caesium chloride/0.1 vi EDTAsolution [221.
The total RNA was reverse transcribed into eDNA using therandom hexamers method as previously described [17, 23]. Briefly,the reaction mixture contained 1 j.d random hexamers (100 jiM;Pharmacia Biosystems Ltd, Milton Keynes, UK), 5 pd dNTPs (2.5mM, Gibco BRL), 2 jil lOx PCR buffer (100 mvt Tris/HC1, 500 mMKC1, 15 mvi MgCI2 and 0.01% wt/vol gelatin), 2 p.l dithiothreitol(0.1 M, Gibco BRL), 1 jil RNasin ribonuclease inhibitor (Pro-mega, Southampton, UK), 1 pJ M-MLV° Superscript reversetranscriptase (200 units; Gibco BRL) and 1 jig of total RNA.
PCR amplification was performed in a total volume of 50 p.l (2j.tl of reverse transcription product and 48 jil of master mixcontaining 36.25 jii H20, 1.25 jrl 5-primer (20 jiM), 1.25 jil3'-primer (20 jiM), 4 jil dNTPs, 5 j.d lOx PCR buffer and 0.25 j.tlTaq polymerase (2.5 U, Amplitaq©, Perkin Elmer Ltd, Bea-consfield, UK) using a Perkin Elmer 480 Thermocycler (PerkinElmer Cetus, Applied Biosystems Ltd, Warrington, UK). ThePCR protocol was as follows: first cycle —94°C for three minutes,55°C for one minute, 72°C for one minute; second to 25th cycles—94°C for 40 seconds, 55°C for one minute, 72°C for one minute.The final cycle was 94°C for 40 seconds and 60°C for 10 minutes.PCR was performed for 23 cycles for a-actin and 25 cycles forIL-6. Preliminary experiments revealed that under these condi-tions the PCR products were generated during the exponentialphase of amplification. One-tenth of the PCR reaction from bothtest (IL-6) and control (a-actin) products were mixed and sepa-
Time, hours
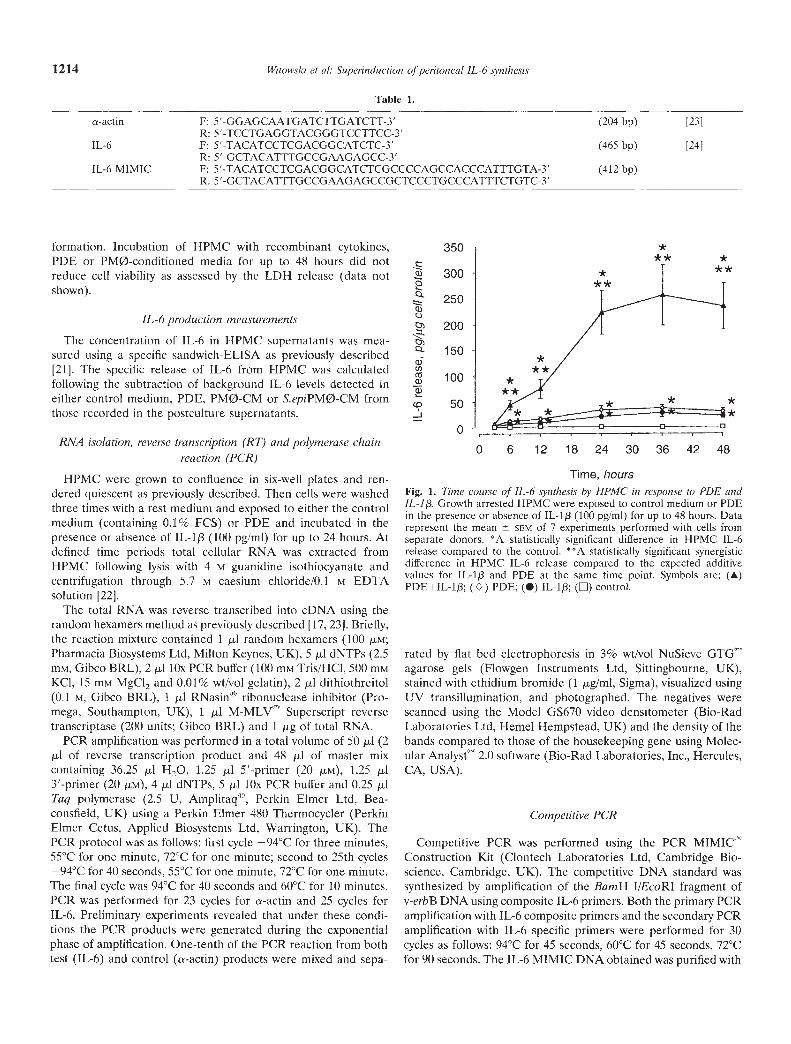
Fig. 1. Time course of JL-6 synthesis by HPMC in response to PDE andIL-I/3 Growth arrested HPMC were exposed to control medium or PDEin the presence or absence of IL-113 (100 pg/mI) for up to 48 hours. Datarepresent the mean SEM of 7 experiments performed with cells fromseparate donors. °A statistically significant difference in HPMC IL-6release compared to the control. °°A statistically significant synergisticdifference in HPMC IL-6 release compared to the expected additivevalues for TL-1 and PDE at the same time point. Symbols are: (A)PDE+IL-1J3; () PDE; (•) IL-i/I; () control.
rated by flat bed electrophoresis in 3% wt/vol NuSieve GTG'agarose gels (Flowgen Instruments Ltd, Sittingbourne, UK),stained with ethidium bromide (1 jig/mI, Sigma), visualized usingUV transillumination, and photographed. The negatives werescanned using the Model GS670 video densitometer (Bio-RadLaboratories Ltd, Hemel Hempstead, UK) and the density of thebands compared to those of the housekeeping gene using Molec-ular Analyst° 2.0 software (Bio-Rad Laboratories, Inc., Hercules,CA, USA).
Competitive PCR
Competitive PCR was performed using the PCR MIMIC'Construction Kit (Clontech Laboratories Ltd, Cambridge Bio-science, Cambridge, UK). The competitive DNA standard wassynthesized by amplification of the BamH I/EcoRl fragment ofv-erbB DNA using composite IL-6 primers. Both the primary PCRamplification with IL-6 composite primers and the secondary PCRamplification with IL-6 specific primers were performed for 30cycles as follows: 94°C for 45 seconds, 60°C for 45 seconds, 72°Cfor 90 seconds. The IL-6 MIMIC DNA obtained was purified with
a-actin
IL-6
IL-6 MIMIC
(204 hp)
(465 bp)
(412 bp)
[23]
[24]
***
***
***q)
Q)C.)
0Cu
ci)a)
(0-J
350 -
300 -
250 -
200 -
150 -
100
50
01
**
**
0 6
*
12 18 24 30 36 42 48
0Control 0.1 1 101001000 PDE 0.1 1 10 100 1000
lL-lp, pg/mi PDE + IL-1f, pg/miB
150***
125 12CI.
100C,
75 I'if 1
50 * **a) * ** _**
25 * *
0 iaapiIitIIIIiIIControl 0.1 1 101001000 PDE 0.1 1 10 100 1000
TNFa, pg/mi PDE + TNFaI pg/mi
1000
A1200 ***
800
600
*
400
**
200 ***
******
Witowski et a!: Superinduction of peritoneal IL-6 synthesis 1215
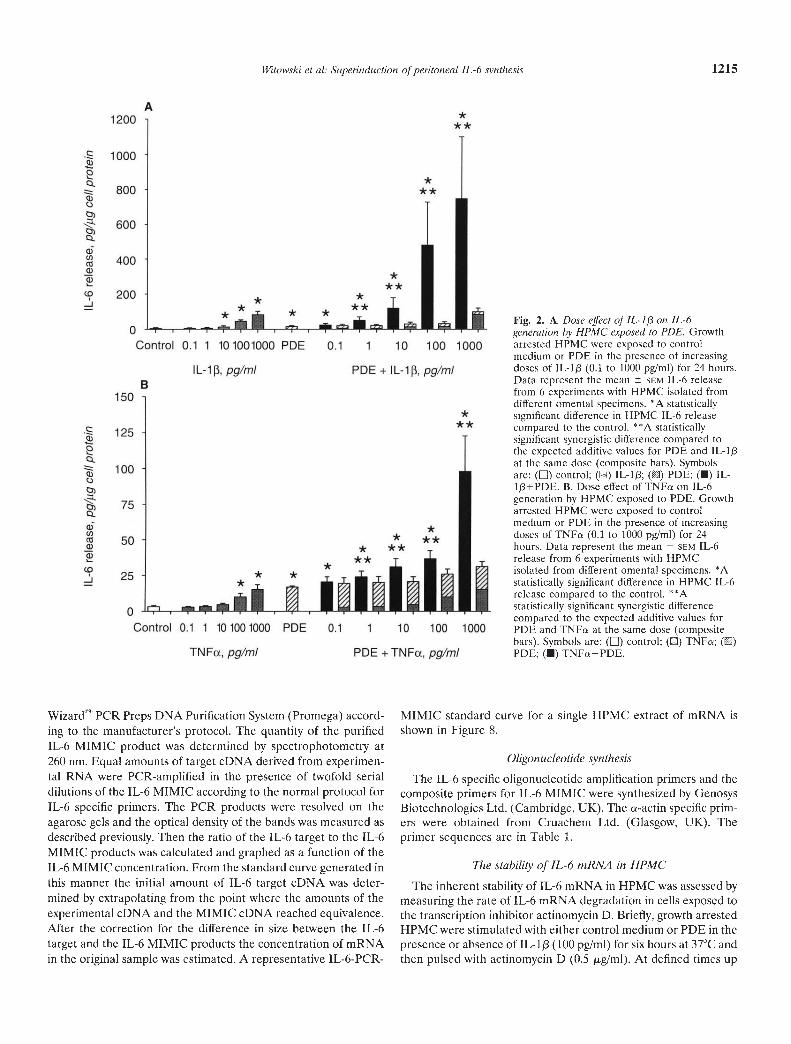
Fig. 2. A. Dose effect of IL-1/3 on JL-ógeneration by HPMC exposed to PDE. Growtharrested HPMC were exposed to controlmedium or PDE in the presence of increasingdoses of JL-l (0.1 to 1000 pg/mI) for 24 hours.Data represent the mean SCM IL-6 releasefrom 6 experiments with HPMC isolated fromdifferent omental specimens. *A statisticallysignificant difference in HPMC IL-6 releasecompared to the control. * *A statisticallysignificant synergistic difference compared tothe expected additive values for PDE and IL-lpat the same dose (composite bars). Symbolsare: () control; (13::) IL-1[3; () PDE; (•) IL-1+PDE. B. Dose effect of TNFa on IL-6generation by HPMC exposed to PDE. Growtharrested HPMC were exposed to controlmedium or PDE in the presence of increasingdoses of TNFs (0.1 to 1000 pg/mI) for 24hours. Data represent the mean SCM IL-6release from 6 experiments with HPMCisolated from different omental specimens. *Astatistically significant difference in HPMC IL-6release compared to the control. **Astatistically significant synergistic differencecompared to the expected additive values forPDE and TNFo at the same dose (compositebars). Symbols are: (LI) control; (El) TNFo; ()PDE; (•) TNFa+PDE.
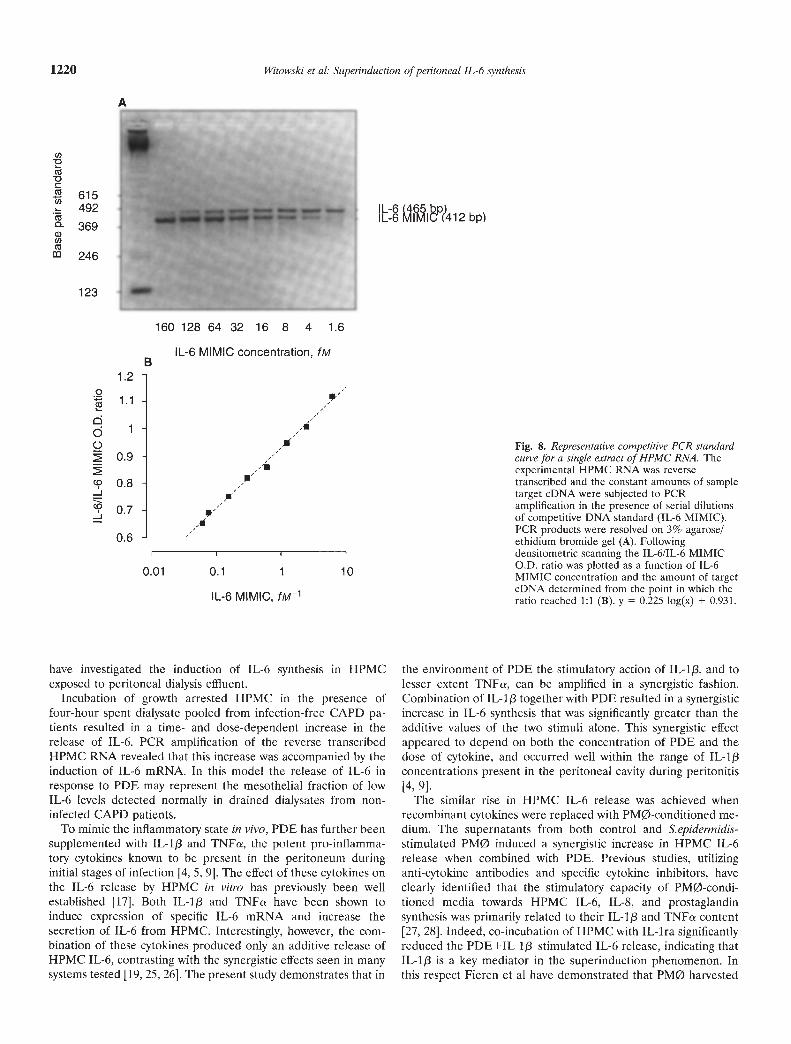
Wizard" PCR Preps DNA Purification System (Promega) accord-ing to the manufacturer's protocol. The quantity of the purifiedIL-6 MIMIC product was determined by spectrophotometry at260 nm. Equal amounts of target eDNA derived from experimen-tal RNA were PCR-amplified in the presence of twofold serialdilutions of the IL-6 MIMIC according to the normal protocol forIL-6 specific primers. The PCR products were resolved on theagarose gels and the optical density of the bands was measured asdescribed previously. Then the ratio of the IL-6 target to the IL-6MIMIC products was calculated and graphed as a function of theIL-6 MIMIC concentration. From the standard curve generated inthis manner the initial amount of IL-6 target eDNA was deter-mined by extrapolating from the point where the amounts of theexperimental eDNA and the MIMIC eDNA reached equivalence.After the correction for the difference in size between the IL-6target and the IL-6 MIMIC products the concentration of mRNAin the original sample was estimated. A representative IL-6-PCR-
MIMIC standard curve for a single HPMC extract of mRNA isshown in Figure 8.
Oligonucleotide synthesis
The IL-6 specific oligonucleotide amplification primers and thecomposite primers for IL-6 MIMIC were synthesized by GenosysBiotechnologies Ltd. (Cambridge, UK). The a-actin specific prim-ers were obtained from Cruachem Ltd. (Glasgow, UK). Theprimer sequences are in Table 1.
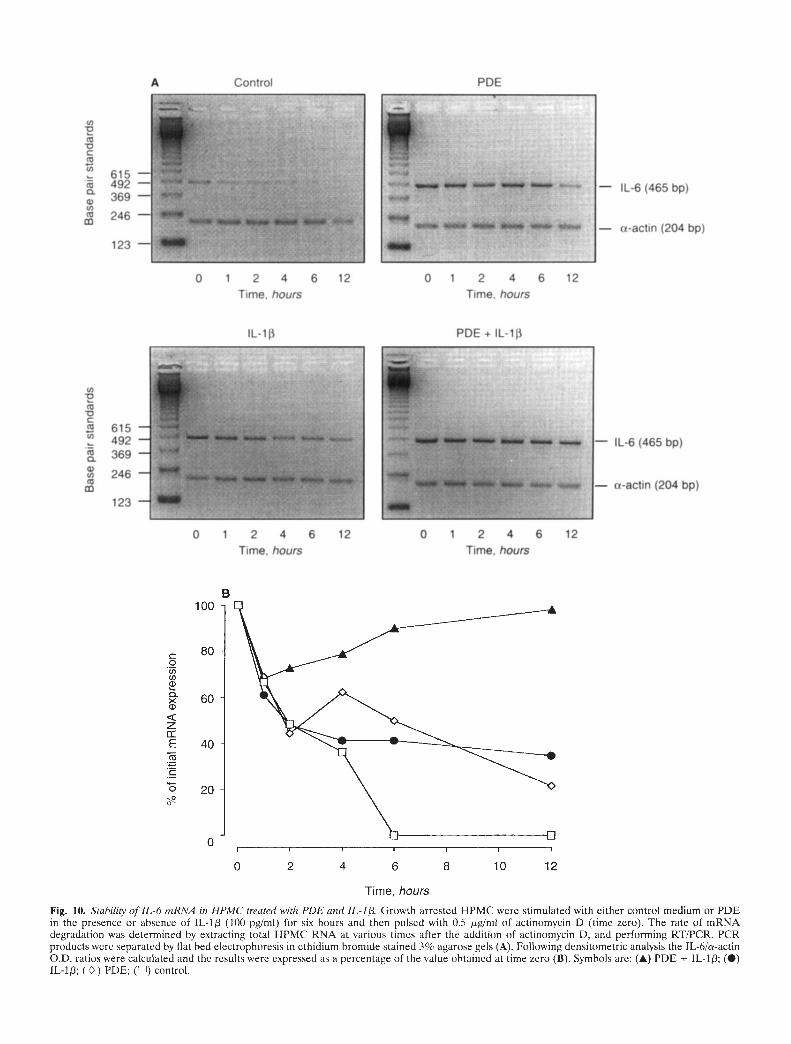
The stability of IL-6 rnRNA in IIPMC
The inherent stability of IL-6 mRNA in HPMC was assessed bymeasuring the rate of IL-fl mRNA degradation in cells exposed tothe transcription inhibitor actinomycin D. Briefly, growth arrestedHPMC were stimulated with either control medium or PDE in thepresence or absence of IL-113 (100 pg/mI) for six hours at 37°C andthen pulsed with actinomycin D (0.5 jig/mI). At defined times up
IL-6
rel
ease
, pg/
pg ce
//pro
tein
N)
N)
C.)
C
O
01
0 01
0
01
0 01
no
0
0 0
0 0
0 0
0 I
a 0 01 0
—1
*
• 1*
* • __
____
_ (I
l
-s
1*
o N)
1*
0
1* **
1216 Witowski et a!: Superinduction of peritoneal IL-6 synthesis
ci>
I
350
to 12 hours after the addition of actinomycin D the total HPMCRNA was extracted, reverse transcribed and PCR amplified asdescribed above. Preliminary experiments revealed that HPMCcould be maintained in the presence of 0.5 jig/ml of actinomycinD for up to 12 hours without significant loss of cell viability asassessed by the level of cellular ATP (data not shown).
Effect of interleukin-1 receptor antagonist
In separate experiments growth arrested HPMC were co-incubated with PDE and/or IL-1/3 in the presence of increasingconcentrations of IL-ira (0.1 to 1000 ng/ml). After 24 hours ofexposure the supernatants were assayed for IL-6 and the datanormalized for cellular protein content.
Effect of cycloheximide
Growth arrested HPMC were pre-treated with 10 g/ml ofcycloheximide (Aldrich Chemical Company, Gillingham, Dorset,UK) for two hours at 37°C and then stimulated with PDE in thepresence or absence of IL-lp (100 pg/mI). After six hours ofexposure the total HPMC RNA was extracted, reverse transcribedand PCR amplified as described above.
HPMC proliferation
Proliferation of HPMC in response to PDE was measured by[3H]-thymidine incorporation assay. Cells were plated onto 96-well plates at a density of 1.25 X i0 cells/well, cultured for 24hours and then growth arrested as previously described. After thatsubconfluent HPMC were exposed to PDE, pulsed with [3H1-thymidine (as [methyl-3H]-thymidine; 2.5 pCi/ml; specific act. 82Ci/mM; Amersham international plc, Aylesbury, UK) and incu-bated in the presence or absence of IL-1/3 (1000 pg/mI) for 48hours at 37°C. At the end of the exposure the labeling fluids wereremoved and cells were washed twice with PBS and precipitatedwith 10% wt/vol trichioroacetic acid. The precipitate was dissolvedin 0.1 N NaOH and the released radioactivity was measured in abeta liquid scintillation counter (LKB Wallac, Turku, Finland).
Statistical analysis
All data were derived from experiments with HPMC preparedfrom at least five separate donors. All statistical analyses wereperformed using the Wilcoxon matched-pairs signed ranks test fornon-parametric data (Statview° SE; Abacus Concepts Inc.,Berkeley, CA, USA) with a P value of less than 0.05 beingconsidered as significant. All data are presented as mean (± SEM).
Results
Induction of HPMC IL-6 production
Incubation of growth arrested HPMC with PDE resulted in atime-dependent increase in IL-6 generation (Fig. 1). The timecourse and the magnitude of the IL-6 release was comparable tothat induced by 100 pg/mI lL-13. In each case the secretion of IL-6became significant above the control level after six hours andreached a plateau by 24 hours. The combination of PDE togetherwith IL-if3, however, induced an IL-6 production above the levelsgenerated by each stimulus alone, and significantly above thepredicted additive value. This synergistic increase in the HPMCIL-6 synthesis was first discernible at six hours, followed by largeincrements over the next i8 hours and a plateau by 24 hours.Repeated measurements over the whole study revealed that inHPMC from different donors the increase in 24-hour IL-6 releasein response to a combination of PDE and 100 pg/mI IL-1f3 rangedfrom 1.1- to 27.2-fold above expected additive levels (medianvalue 3.0-fold, N = 40).
The synergistic effect of the combined PDE and IL-lp stimu-lation was dependent on the dose of IL-1/3 (Fig. 2A). It becamesignificant above additive values when PDE was combined with Ipg/ml IL-113 and maximal synergy was obtained at the highestIL-1f3 dose tested (1000 pg/mI) when the 24-hour HPMC IL-6secretion was 747.74 349.93 pg/.tg cell protein compared to theadditive value of 99.81 19.16 pg/jig cell protein (N = 6, z =2.201, P < 0.027). The same effect was evident when IL-113 wasreplaced with TNFa, although the degree of stimulation was muchless pronounced (Fig. 2B). A 24-hour exposure of HPMC to 1000
300
250
200
150
100
50
0
Control Neat 2 5 10 20
Dilution factor
lL-13 Neat 2
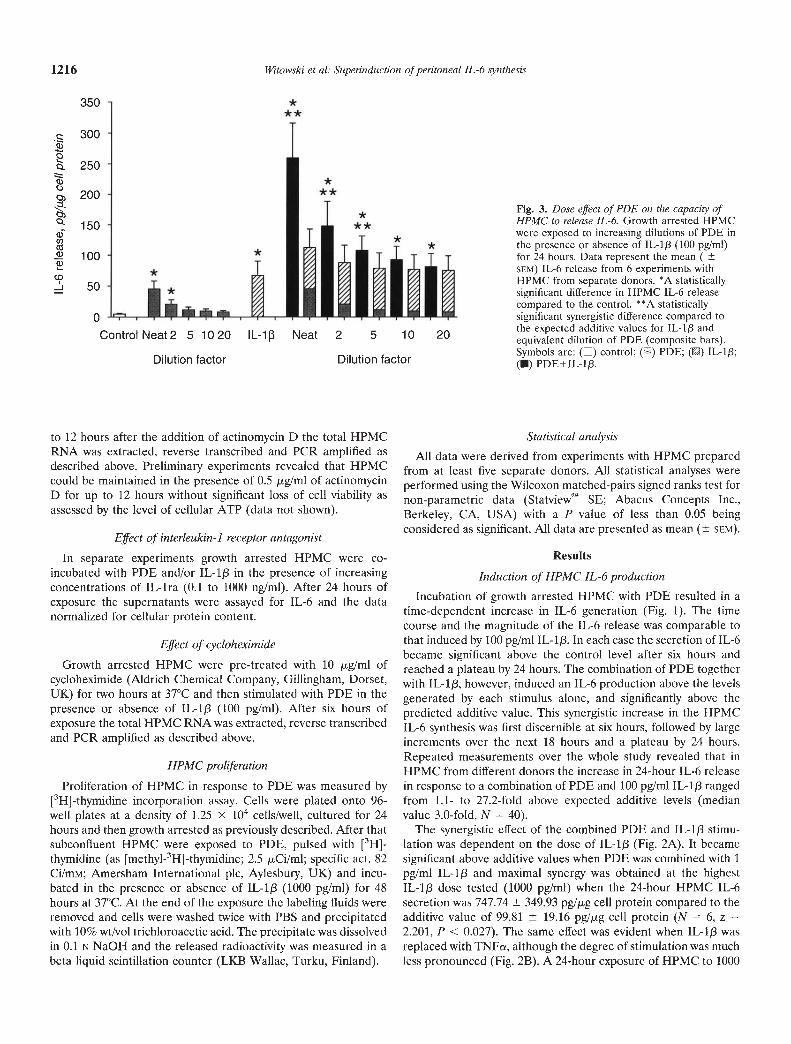
Fig. 3. Dose effect of PDE on the capacity ofHPMC to release IL-6. Growth arrested HPMCwere exposed to increasing dilutions of PDE inthe presence or absence of IL-1j3 (100 pg/mI)for 24 hours. Data represent the mean (SEM) IL-6 release from 6 experiments withHPMC from separate donors. *A statisticallysignificant difference in HPMC IL-6 releasecompared to the control. * °A statisticallysignificant synergistic difference compared tothe expected additive values for IL-lp andequivalent dilution of PDE (composite bars).Symbols are: () control; (LI) PDE; () IL-1f3;(U) PDE+IL-1j3.
5 10 20
Dilution factor
300
200
800*** 2600
400 I **60
CdC, 2ooLJJJA* **
0—
Control POE PM0 SFpiPM0 0 0.05 0.1 0.5 1 5
600
500
400***
100
**
I I S a
Witowski et a!: Superinduction of peritoneal IL-6 synthesis 1217
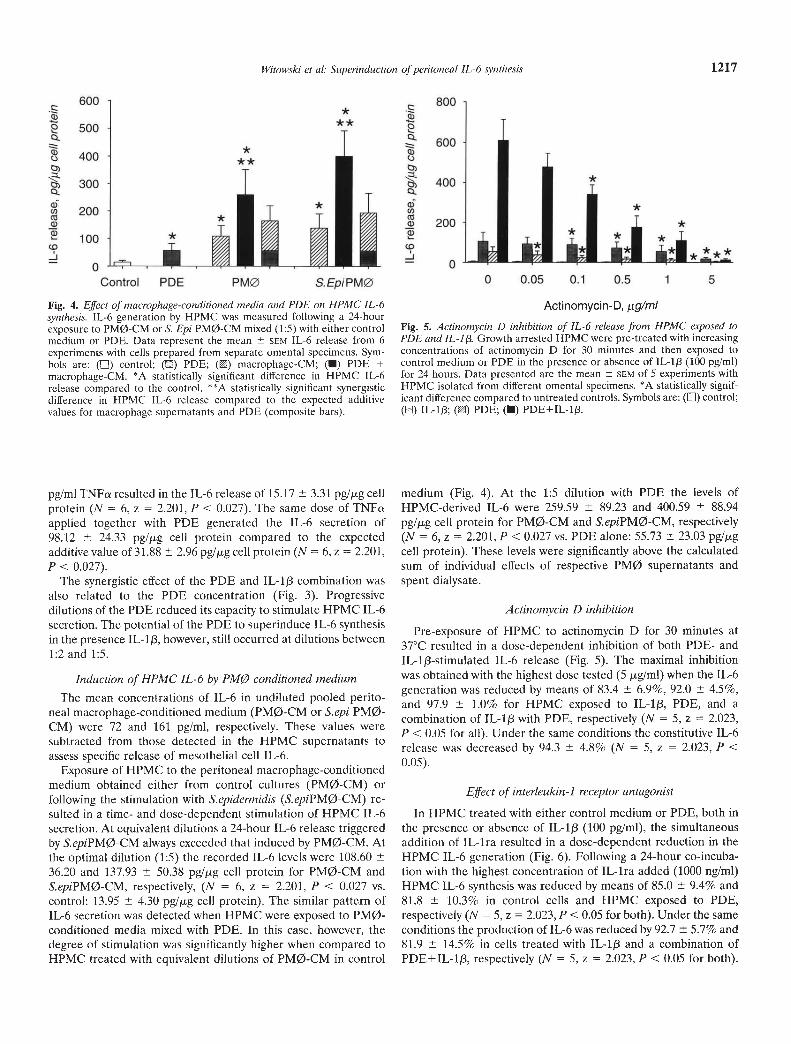
Fig. 4. Effect of macrophage-conditioned media and PDE on HPMC IL-6synthesis. IL-6 generation by HPMC was measured following a 24-hourexposure to PM0-CM or S. Epi PM0-CM mixed (1:5) with either controlmedium or PDE. Data represent the mean SCM IL-6 release from 6experiments with cells prepared from separate omental specimens. Sym-bols are: (LI) control; (LI) PDE; () macrophage-CM; (U) PDE +macrophage-CM. *A statistically significant difference in HPMC IL-6release compared to the control. **A statistically significant synergisticdifference in HPMC IL-6 release compared to the expected additivevalues for macrophage supernatants and PDE (composite bars).
pg/mi TNFa resulted in the IL-6 release of 15.17 3.31 pg/j.tg cell
protein (N = 6, z = 2.201, P < 0.027). The same dose of TNFaapplied together with PDE generated the IL-6 secretion of98.12 24.33 pg/jig cell protein compared to the expectedadditivevalue of 31.88 2.96 pg/jig cell protein (N= 6,z = 2.201,P < 0.027).
The synergistic effect of the PDE and IL-1f3 combination wasalso related to the PDE concentration (Fig. 3). Progressivedilutions of the PDE reduced its capacity to stimulate HPMC IL-6secretion. The potential of the PDE to superinduce IL-6 synthesisin the presence IL-113, however, still occurred at dilutions between1:2 and 1:5.
Induction of HPMC IL-6 by PMØ conditioned medium
The mean concentrations of IL-6 in undiluted pooled perito-neal macrophage-conditioned medium (PM0-CM or S.epi PM0-CM) were 72 and 161 pg/ml, respectively. These values weresubtracted from those detected in the HPMC supernatants toassess specific release of mesothelial cell IL-6.
Exposure of HPMC to the peritoneal macrophage-conditionedmedium obtained either from control cultures (PMØ-CM) orfollowing the stimulation with S.epidermidis (S.epiPMø-CM) re-sulted in a time- and dose-dependent stimulation of HPMC IL-6secretion. At equivalent dilutions a 24-hour IL-6 release triggeredby S.epiPMø-CM always exceeded that induced by PM0-CM. Atthe optimal dilution (1:5) the recorded IL-6 levels were 108.6036.20 and 137.93 50,38 pg/jig cell protein for PM0-CM andS.epiPMø-CM, respectively, (N = 6, z = 2.201, P < 0.027 vs.control: 13.95 4,30 pg/jig cell protein). The similar pattern ofIL-6 secretion was detected when HPMC were exposed to PM0-conditioned media mixed with PDE. In this case, however, thedegree of stimulation was significantly higher when compared toHPMC treated with equivalent dilutions of PM0-CM in control
Actinomycin-D, jig/mi
Fig. 5. Actinomycin D inhibition of JL-6 release from HPMC exposed toPDE and IL -1 13 Growth arrested HPMC were pre-treated with increasingconcentrations of actinomycin D for 30 minutes and then exposed tocontrol medium or PDE in the presence or absence of IL-1f3 (100 pg/mI)for 24 hours. Data presented are the mean SEM of 5 experiments withHPMC isolated from different omental specimens. *A statistically signif-icant difference compared to untreated controls. Symbols are: (El) control;(L:) IL-113; () PDE; (U) PDE+IL-1f3.
medium (Fig. 4). At the 1:5 dilution with PDE the levels ofHPMC-derived IL-6 were 259.59 89.23 and 400.59 88.94
pg/jig cell protein for PM0-CM and S.epiPMø-CM, respectively(N = 6, z = 2.201, P < 0.027 vs. PDE alone: 55.73 23.03 pg/jigcell protein). These levels were significantly above the calculatedsum of individual effects of respective PMØ supernatants andspent dialysate.
Actinomycin D inhibition
Pre-exposure of HPMC to actinomycin D for 30 minutes at37°C resulted in a dose-dependent inhibition of both PDE- andIL-1j3-stimulated IL-6 release (Fig. 5). The maximal inhibitionwas obtained with the highest dose tested (5 jig/mI) when the IL-6generation was reduced by means of 83.4 6.9%, 92.0 4.5%,and 97.9 1.0% for HPMC exposed to IL-1/3, PDE, and acombination of IL-1f3 with PDE, respectively (N = 5, z 2.023,P < 0.05 for all). Under the same conditions the constitutive IL-6release was decreased by 94.3 4.8% (N = 5, z 2.023, P <0.05).
Effect of interleukin-1 receptor antagonist
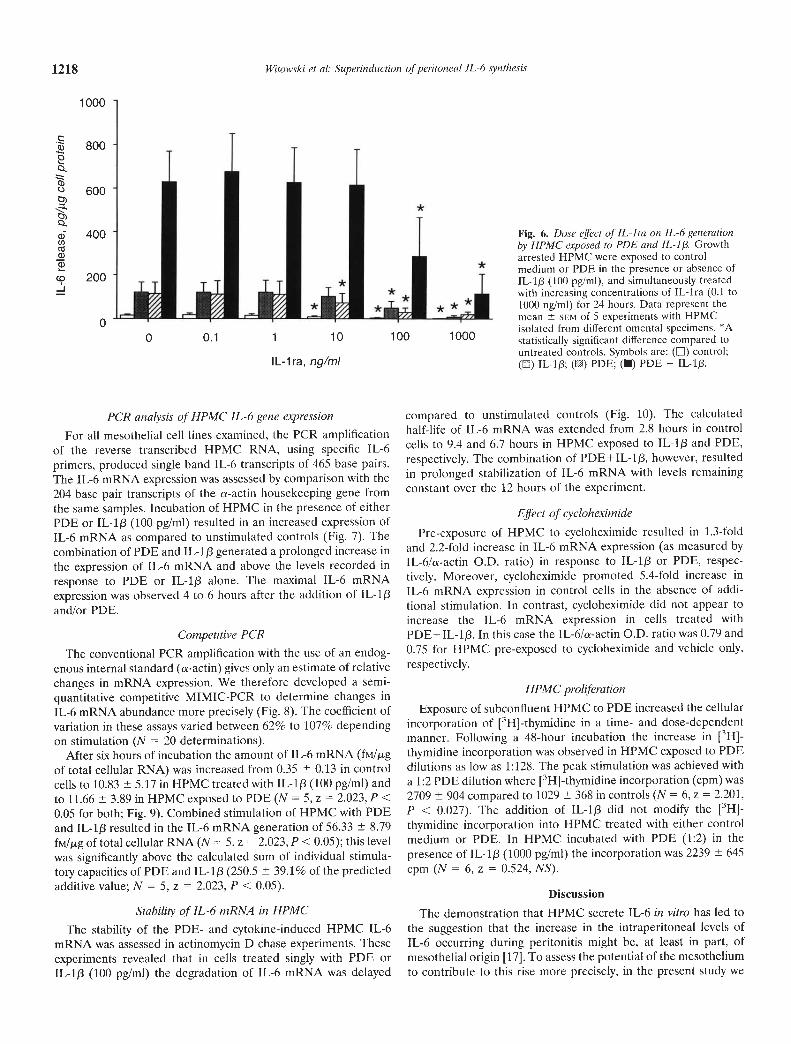
In HPMC treated with either control medium or PDE, both inthe presence or absence of IL-1j3 (100 pg/mI), the simultaneousaddition of IL-ira resulted in a dose-dependent reduction in theHPMC IL-6 generation (Fig. 6). Following a 24-hour co-incuba-tion with the highest concentration of IL-ira added (1000 ng/ml)HPMC IL-6 synthesis was reduced by means of 85.0 9.4% and81.8 10.3% in control cells and HPMC exposed to PDE,respectively (N = 5, z = 2.023, P < 0.05 for both). Under the sameconditions the production of IL-6 was reduced by 92.7 5.7% and81.9 14.5% in cells treated with IL-1/3 and a combination ofPDE+IL-lp, respectively (N = 5, z = 2.023, P < 0.05 for both).
*
800
0.
600
0 0.1 1 10 100 1000
IL-ira, ng/mI
*
1218 Witowski ci al: Superinduction of peritoneal JL-6 synthesis
a)
a)C.)
a))
a)C))Ca
a)Ca)
-J
1000
PCR analysis of HPMC IL-6 gene expressionFor all mesothelial cell lines examined, the PCR amplification
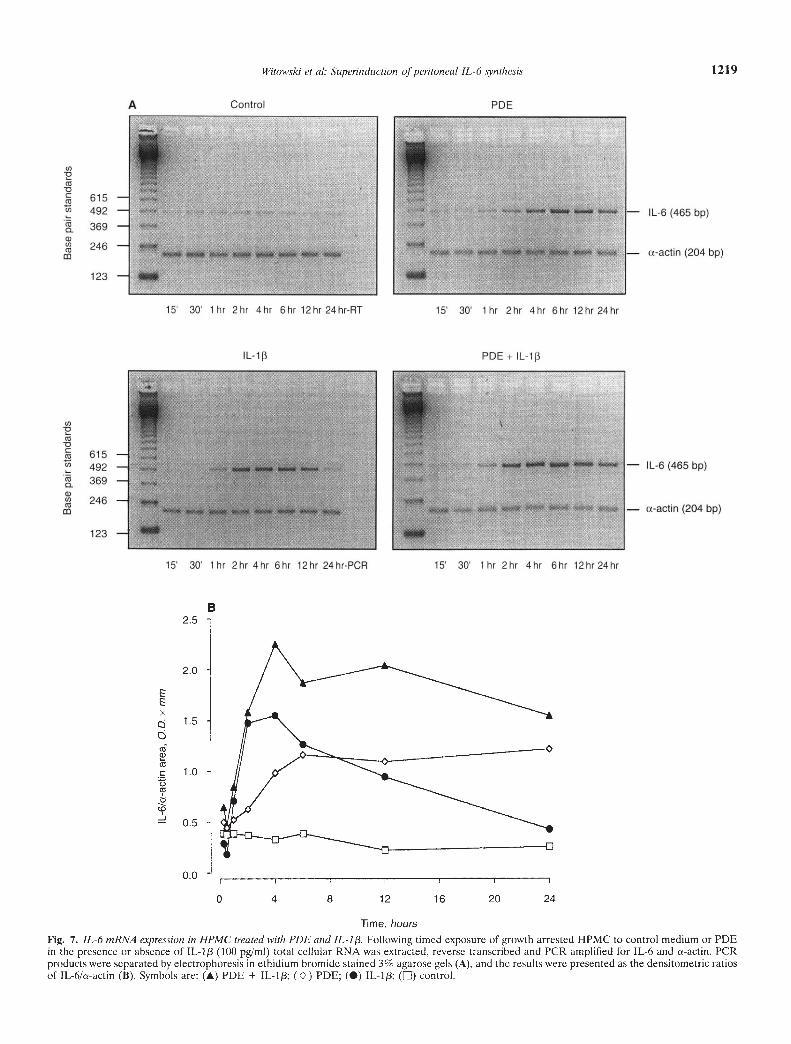
of the reverse transcribed HPMC RNA, using specific IL-6primers, produced single band IL-6 transcripts of 465 base pairs.The IL-6 mRNA expression was assessed by comparison with the204 base pair transcripts of the cs-actin housekeeping gene fromthe same samples. Incubation of HPMC in the presence of eitherPDE or IL-1/3 (100 pg/mI) resulted in an increased expression ofIL-6 mRNA as compared to unstimulated controls (Fig. 7). Thecombination of PDE and IL-1/3 generated a prolonged increase inthe expression of IL-6 mRNA and above the levels recorded inresponse to PDE or IL-1/3 alone. The maximal IL-6 mRNAexpression was observed 4 to 6 hours after the addition of IL-1/3and/or PDE.
Competitive PCR
The conventional PCR amplification with the use of an endog-enous internal standard (a-actin) gives only an estimate of relativechanges in mRNA expression. We therefore developed a semi-quantitative competitive MIMIC-PCR to determine changes inIL-6 mRNA abundance more precisely (Fig. 8). The coefficient ofvariation in these assays varied between 62% to 107% dependingon stimulation (N = 20 determinations).
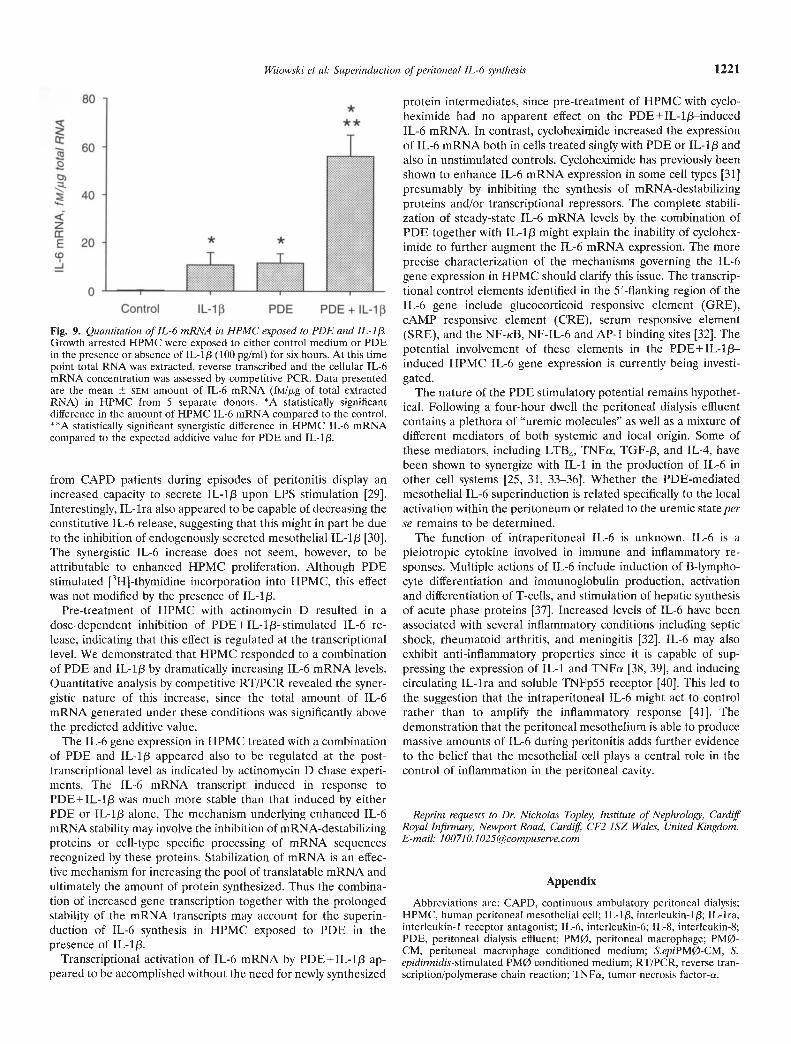
After six hours of incubation the amount of IL-6 mRNA (fM//kgof total cellular RNA) was increased from 0.35 0.13 in controlcells to 10.83 5.17 in HPMC treated with IL-i [3 (100 pg/mI) andto 11.66 3.89 in HPMC exposed to PDE (N = 5, z = 2.023, P <0.05 for both; Fig. 9). Combined stimulation of HPMC with PDEand IL-1/3 resulted in the IL-6 mRNA generation of 56.33 8.79fM//kg of total cellular RNA (N = 5, z 2.023, P < 0.05); this levelwas significantly above the calculated sum of individual stimula-tory capacities of PDE and IL-1!3 (250.5 39.1% of the predictedadditive value; N = 5, z = 2.023, P < 0.05).
Stability of IL-6 mRNA in HPMC
The stability of the PDE- and cytokine-induced HPMC IL-6mRNA was assessed in actinomycin D chase experiments. Theseexperiments revealed that in cells treated singly with PDE orlL-1/3 (100 pg/mI) the degradation of IL-6 mRNA was delayed
compared to unstimulated controls (Fig. 10). The calculatedhalf-life of IL-6 mRNA was extended from 2.8 hours in controlcells to 9.4 and 6.7 hours in HPMC exposed to IL-1/3 and PDE,respectively. The combination of PDE+IL-1/3, however, resultedin prolonged stabilization of IL-6 mRNA with levels remainingconstant over the 12 hours of the experiment.
Effect of cycloheximidePre-exposure of HPMC to cycloheximide resulted in 1.3-fold
and 2.2-fold increase in IL-6 mRNA expression (as measured byIL-6/a-actin O.D. ratio) in response to IL-l/3 or PDE, respec-tively. Moreover, cycloheximide promoted 5.4-fold increase inIL-6 mRNA expression in control cells in the absence of addi-tional stimulation. In contrast, cycloheximide did not appear toincrease the IL-6 mRNA expression in cells treated withPDE+IL-1/3. In this case the IL-6/a-actin O.D. ratio was 0.79 and0.75 for HPMC pre-exposed to cycloheximide and vehicle only,respectively.
HPMC proliferationExposure of subconfiuent HPMC to PDE increased the cellular
incorporation of [3H]-thymidine in a time- and dose-dependentmanner. Following a 48-hour incubation the increase in [3H1-thymidine incorporation was observed in HPMC exposed to PDEdilutions as low as 1:128. The peak stimulation was achieved witha 1:2 PDE dilution where [3H]-thymidine incorporation (cpm) was2709 904 compared to 1029 368 in controls (N =6, z = 2.201,P < 0.027). The addition of IL-1f3 did not modify the [3Hj-thymidine incorporation into HPMC treated with either controlmedium or PDE. In HPMC incubated with PDE (1:2) in thepresence of IL-i/I (1000 pg/mI) the incorporation was 2239 645cpm (N = 6, z = 0.524, A(S).
Discussion
The demonstration that HPMC secrete IL-6 in vitro has led tothe suggestion that the increase in the intraperitoneal levels ofIL-6 occurring during peritonitis might be, at least in part, ofmesothelial origin [171. To assess the potential of the mesotheliumto contribute to this rise more precisely, in the present study we
800
600
400
200
00 0.1 1 10 100
IL-ira, ng/mI
1000
Fig. 6. Dose effect of IL-ira on IL-b generationby HPMC exposed to PDE and IL-i /3. Growtharrested HPMC were exposed to controlmedium or PDE in the presence or absence ofIL-I/I (100 pg/mI), and simultaneously treatedwith increasing concentrations of IL-ira (0.1 to1000 nglml) for 24 hours. Data represent themean SEM of 5 experiments with HPMCisolated from different omental specimens. *Astatistically significant difference compared tountreated controls. Symbols are: () control;(E:) IL-I /3; () PDE; (I) PDE + IL-1/3.
4T615 —492 —369
tl)246
IL-i (3
15 30 1 hr 2hr 4hr 6hr l2hr 24hr-PCR
POE + IL-i I
15 30 1 hr 2hr 4hr 6hr i2hr24hr
IL-6 (465 bp)
A Control
123
PDE
____ — IL-6 (465 bp)
— a-actin (204 bp)
15 30 1 hr 2hr 4hr 6hr i2hr 24hr15 30' 1 hr 2hr 4hr 6hr l2hr 24hr-RT
615492369
246
123
KK
1a-actin (204 bp)
EEx
d
C0
(0-J
Witowski et al: Superinduction of peritoneal IL-6 synthesis 1219
B2.5
2.0
1.5
1.0
0.5
0.0I I I
0 4 8 12 16 20 24
Time, hours
Fig. 7. IL-h mRNA expression in HPMC treated with PDE and JL-1f3. Following timed exposure of growth arrested HPMC to control medium or PDEin the presence or absence of IL-lp (100 pg/mI) total cellular RNA was extracted, reverse transcribed and PCR amplified for IL-6 and a-actin. PCRproducts were separated by electrophoresis in ethidium bromide stained 3% agarose gels (A), and the results were presented as the densitometric ratiosof IL-6/a-actin (B). Symbols are: (A) PDE + IL-l13; () PDE; (•) IL-1; (LII) control.
Cl)
C
Cl)
a)(6)
0.01 0.1 1 10
IL-6 MIMIC, fM1
have investigated the induction of IL-6 synthesis in HPMCexposed to peritoneal dialysis effluent.
Incubation of growth arrested HPMC in the presence offour-hour spent dialysate pooled from infection-free CAPD pa-tients resulted in a time- and dose-dependent increase in therelease of IL-6. PCR amplification of the reverse transcribedHPMC RNA revealed that this increase was accompanied by theinduction of IL-6 mRNA. In this model the release of IL-6 inresponse to PDE may represent the mesothelial fraction of lowIL-6 levels detected normally in drained dialysates from non-infected CAPD patients.
To mimic the inflammatory state in vivo, PDE has further beensupplemented with IL-1/3 and TNFs, the potent pro-inflamma-tory cytokines known to be present in the peritoneum duringinitial stages of infection [4, 5, 9]. The effect of these cytokines onthe IL-6 release by HPMC in vitro has previously been wellestablished [17]. Both IL-Ip and TNFa have been shown toinduce expression of specific IL-6 mRNA and increase thesecretion of IL-6 from HPMC. Interestingly, however, the com-bination of these cytokines produced only an additive release ofHPMC IL-fl, contrasting with the synergistic effects seen in manysystems tested [19, 25, 261. The present study demonstrates that in
Fig. 8. Representative competitive PCR standardcurve for a single extract of HPMC RNA. Theexperimental HPMC RNA was reversetranscribed and the constant amounts of sampletarget eDNA were subjected to PCRamplification in the presence of serial dilutionsof competitive DNA standard (IL-6 MIMIC).PCR products were resolved on 3% agarose!ethidium bromide gel (A). Followingdensitometric scanning the IL-6/IL-6 MIMICO.D. ratio was plotted as a function of IL-6MIMIC concentration and the amount of targeteDNA determined from the point in which theratio reached 1:1 (B). y = 0.225 log(x) + 0.931.
the environment of PDE the stimulatory action of IL-1p, and tolesser extent TNFa, can be amplified in a synergistic fashion.Combination of IL-1f3 together with PDE resulted in a synergisticincrease in IL-6 synthesis that was significantly greater than theadditive values of the two stimuli alone. This synergistic effectappeared to depend on both the concentration of PDE and thedose of cytokine, and occurred well within the range of IL-I 13concentrations present in the peritoneal cavity during peritonitis[4, 9].
The similar rise in HPMC IL-6 release was achieved whenrecombinant cytokines were replaced with PM0-conditioned me-dium. The supernatants from both control and S.epidermidis-stimulated PMØ induced a synergistic increase in HPMC IL-flrelease when combined with PDE. Previous studies, utilizinganti-cytokine antibodies and specific cytokine inhibitors, haveclearly identified that the stimulatory capacity of PM0-condi-tioned media towards HPMC IL-fl, IL-8, and prostaglandinsynthesis was primarily related to their IL-1f3 and TNFa content[27, 28]. Indeed, co-incubation of HPMC with IL-ira significantlyreduced the PDE+IL-l/3—stimulated IL-6 release, indicating thatIL-I /3 is a key mediator in the superinduction phenomenon. Inthis respect Fieren et a! have demonstrated that PMØ harvested
1220 Witowski et al: Superinduction of peritoneal IL-6 synthesis
A
615492 IL-6 l??412 bpI L-6369
246
123
160 128 64 32 16 8 4 1.6
IL-6 MIMIC concentration, fMB
1.2
.2 I
. 1.1_I -U,
cid 1-0 I
0.9 -]
CD 0.8 4_J I
I /0.7-a/1
O.6-
80 *
60
**
2
40
zIE 20 * *Co __ __III
Control IL-iD PDE PDE + IL-ip
Wit owski et al: Superinduction of peritoneal IL-6 synthesis 1221
Fig. 9. Quantitation of !L-6 mRNA in HPMC exposed to PDE and IL-1/Growth arrested HPMC were exposed to either control medium or PDEin the presence or absence of IL-113 (100 pg/ml) for six hours. At this timepoint total RNA was extracted, reverse transcribed and the cellular IL-6mRNA concentration was assessed by competitive PCR. Data presentedare the mean SEM amount of IL-6 mRNA (fM/J.Lg of total extractedRNA) in HPMC from 5 separate donors. *A statistically significantdifference in the amount of HPMC IL-6 mRNA compared to the control.**A statistically significant synergistic difference in HPMC IL-6 mRNAcompared to the expected additive value for PDE and IL-1J3.
from CAPD patients during episodes of peritonitis display anincreased capacity to secrete IL-1f3 upon LPS stimulation [29].Interestingly, IL-ira also appeared to be capable of decreasing theconstitutive IL-6 release, suggesting that this might in part be dueto the inhibition of endogenously secreted mesothelial IL-1f3 [30].The synergistic IL-6 increase does not seem, however, to beattributable to enhanced HPMC proliferation. Although PDEstimulated [3H]-thymidine incorporation into HPMC, this effectwas not modified by the presence of IL-i/i.
Pre-treatment of HPMC with actinomycin D resulted in adose-dependent inhibition of PDE+IL-1f3—stimulated IL-6 re-lease, indicating that this effect is regulated at the transcriptionallevel. We demonstrated that HPMC responded to a combinationof PDE and IL-i/i by dramatically increasing IL-6 mRNA levels.Quantitative analysis by competitive RT/PCR revealed the syner-gistic nature of this increase, since the total amount of IL-6mRNA generated under these conditions was significantly abovethe predicted additive value.
The lL-6 gene expression in HPMC treated with a combinationof PDE and IL-i /3 appeared also to be regulated at the post-transcriptional level as indicated by actinomycin D chase experi-ments. The IL-6 mRNA transcript induced in response toPDE+IL-1f3 was much more stable than that induced by eitherPDE or IL-i/i alone. The mechanism underlying enhanced IL-6mRNA stability may involve the inhibition of mRNA-destahilizingproteins or cell-type specific processing of mRNA sequencesrecognized by these proteins. Stabilization of mRNA is an effec-tive mechanism for increasing the pooi of translatable mRNA andultimately the amount of protein synthesized. Thus the combina-tion of increased gene transcription together with the prolongedstability of the mRNA transcripts may account for the superin-duction of IL-6 synthesis in HPMC exposed to PDE in thepresence of IL-I/I.
Transcriptional activation of IL-6 mRNA by PDE+IL-i/3 ap-peared to be accomplished without the need for newly synthesized
protein intermediates, since pre-treatment of HPMC with cyclo-heximide had no apparent effect on the PDE+IL-1/3—inducedIL-6 mRNA. In contrast, cycloheximide increased the expressionof IL-6 mRNA both in cells treated singly with PDE or IL-I/i andalso in unstimulated controls. Cycloheximide has previously beenshown to enhance IL-6 mRNA expression in some cell types [31]presumably by inhibiting the synthesis of mRNA-destabilizingproteins and/or transcriptional repressors. The complete stabili-zation of steady-state IL-6 mRNA levels by the combination ofPDE together with IL-I/i might explain the inability of cyclohex-imide to further augment the IL-6 mRNA expression. The moreprecise characterization of the mechanisms governing the IL-6gene expression in HPMC should clarify this issue. The transcrip-tional control elements identified in the 5'-flanking region of theIL-6 gene include glucocorticoid responsive element (GRE),cAMP responsive element (CRE), serum responsive element(SRE), and the NF-KB, NF-IL-6 and AP-i binding sites [32]. Thepotential involvement of these elements in the PDE+ IL-i/i—induced HPMC IL-6 gene expression is currently being investi-gated.
The nature of the PDE stimulatory potential remains hypothet-ical. Following a four-hour dwell the peritoneal dialysis effluentcontains a plethora of "uremic molecules" as well as a mixture ofdifferent mediators of both systemic and local origin. Some ofthese mediators, including LTB4, TNFa, TGF-/3, and IL-4, havebeen shown to synergize with IL-i in the production of IL-6 inother cell systems [25, 31, 33—36]. Whether the PDE-mediatedmesothelial IL-6 superinduction is related specifically to the localactivation within the peritoneum or related to the uremic state perse remains to be determined.
The function of intraperitoneal IL-6 is unknown. IL-6 is apleiotropic cytokine involved in immune and inflammatory re-sponses. Multiple actions of IL-6 include induction of B-lympho-cyte differentiation and immunoglobulin production, activationand differentiation of T-cells, and stimulation of hepatic synthesisof acute phase proteins [37]. Increased levels of IL-6 have beenassociated with several inflammatory conditions including septicshock, rheumatoid arthritis, and meningitis [32]. IL-6 may alsoexhibit anti-inflammatory properties since it is capable of sup-pressing the expression of IL-i and TNFa [38, 39], and inducingcirculating IL-ira and soluble TNFp55 receptor [40]. This led tothe suggestion that the intraperitoneal IL-6 might act to controlrather than to amplify the inflammatory response [41]. Thedemonstration that the peritoneal mesothelium is able to producemassive amounts of IL-6 during peritonitis adds further evidenceto the belief that the mesothelial cell plays a central role in thecontrol of inflammation in the peritoneal cavity.
Reprint requests to Dr. Nicholas Topley, Institute of Nephrology, CardiffRoyal Infirmaty, Newport Road, Cardifl CF2 ISZ Wales, United Kingdom.E-mail: [email protected]
Appendix
Abbreviations are: CAPD, continuous ambulatory peritoneal dialysis;HPMC, human peritoneal mesothelial cell; IL-i/I, interleukin-1/3; IL-ira,interleukin-1 receptor antagonist; IL-6, interleukin-6; IL-8, interleukin-8;PDE, peritoneal dialysis effluent; PMØ, peritoneal macrophage; PM0-CM, peritoneal macrophage conditioned medium; S.epiPMø-CM, S.epidinnidis-stimulated PMØ conditioned medium; RT/PCR, reverse tran-scription/polymerase chain reaction; TNFa, tumor necrosis factor-a.
615492369
246
123 —.
A Control
Time, hours Time, hours
— IL-6 (465 bp)
— t-actin (204 bp)
lL-1 POE + lL-fli
0 1 2 4 6 12 0 1 2 4 6 12
Time, hours Time, hours
PDE
0 1 2 4 6 12 0 1 2 4 6 12
615492369
246
123
— iL-b (4bb D)
— a-actin (204 bp)
B100
800(I)(I)ci)0.><ci)
zE 40
20
0
0 2 4 6 8 10 12
Time, hours
Fig. 10. S/ability of IL-b rnRNA in HPMC treated with PDE and IL-I , Growth arrested HPMC were stimulated with either control medium or PDEin the presence or absence of IL-1 (100 pg/mI) for six hours and then pulsed with 0.5 jrg/nil of actinomycin D (time zero). The rate of mRNAdegradation was determined by extracting total J-TPMC RNA at various times after the addition of actinomycin D, and performing RT/PCR. PCRproducts were separated by flat bed electrophoresis in ethidium bromide stained 3% agarose gels (A). Following densitometric analysis the TL-6/a-actinO.D. ratios were calculated and the results were expressed as a percentage of the value obtained at time zero (B). Symbols are: (A) PDE + IL-1; (S)1L-1; () PDE; ([I]) control.

Witowski et al: Superinduction of peritoneal IL-fl synthesis 1223
References
1. VERGER C, LUGER A, MOORE HL, NOLPH KD: Acute changes inperitoneal morphology and transport properties with infectious peri-tonitis and mechanical injury. Kidney mt 23:823—831, 1983
2. STEINHAUER HB, SCHOLLMEYER P: Prostaglandin mediated loss ofproteins during peritonitis in continuous ambulatory peritoneal dial-ysis. Kidney mt 29:584—590, 1986
3. STEINIIAUER HB, GÜNTER, B, SCHOLLMEYER P: Stimulation of pen-toneal synthesis of vasoactive prostaglandins during peritonitis inpatients on continuous ambulatory penitoneal dialysis. Eur J C/inInvest 15:1—5, 1985
4. MOUTABARRIK A, NAIcANI5HI I, NAMIKI M, TSUBAKIHARA Y: Inter-leukin-l and its naturally occurring inhibitor in penitoneal dialysispatients. C/in Nephrol 43:243—248, 1995
5. ZEMEL D, IMHOLZ ALT, DE WART DR, DINKLA C, STRUIJK DG,KREDIET RT: The appearance of tumor necrosis factor-a and solubleTNF-receptors I and II in penitoneal effluent during stable andinfectious CAPD. Kidney mt 46:1422—1430, 1994
6. GOLDMAN M, VANDENABEEI.E P, MOULART J, Aviicot:i Z,ABRAMOWICZ D, NORTIER J, VANHERWEGHEM JL, FIURS E: Intraper-itoneal secretion of interleukin-6 during continuous ambulatory pen-toneal dialysis. Nephron 56:277—280, 1990
7. BRAUNER A, HYLANDER B, WRETLIND B: Interleukin-6 and interleu-kin-8 in dialysate and serum from patients on continuous ambulatoryperitoneal dialysis. Am J Kid Dis 22:430—435, 1993
8. DASGUPTA MK, LARABIE M, HALLORAN PF: Interferon-gamma levelsin peritoneal dialysis effluents: Relation to peritonitis. Kidney Jot46:475—481, 1994
9. BRAUNER A, HYLANDER B, WRETLIND B: Tumor necrosis factor-a,interleukin-1, and interleukin-1 receptor antagonist in dialysate andserum from patients on continuous ambulatory peritoneal dialysis.Am J Kid Dis 27:402—408, 1996
10. ZEMEL D, BETJES MGH, DINKI.A C, STRUIJK DG, KREDIET RT:Analysis of inflammatory mediators and penitoneal permeability tomacromolecules shortly before the onset of overt peritonitis inpatients treated with CAPD. Pent Dial mt 15:134—141, 1995
11. LiN C-Y, UN C-C, HUANG T-P: Serial changes of intcrleukin-6 andinterleukin-8 levels in drain dialysate of urcmic patients with CAPDduring peritonitis. Nephron 63:404—408, 1993
12. NAKAHAMA H, TANAKA Y, SHIRAI D, MIYAZAKI M, IMAI N,YOKOKAWA T, OKADA M, KUBORI S: Plasma interleukin-6 levels incontinuous ambulatory penitoneal dialysis and hemodialysis patients.Nephron 61:132—134, 1992
13. ZEMEL D, KOOMEN GCM, HART AAM, TEN BERGE RJM, STRUIJKDG, KREDIET RT: Relationship of TNFa, interleukin-6, and prosta-glandins to penitoneal permeability for macromolecules during longi-tudinal follow-up of peritonitis in continuous ambulatory peritonealdialysis. I Lab C/in Med 122:686—696, 1993
14. ZEMEL D, TEN BERGE RJMGSD, BLOEMENA E, KOOMEN GCM,KREDIET RT: Interleukin-6 in CAPD patients without peritonitis:Relationship to the intrinsic permeability of the peritoneal membrane.C/in Nephrol 37:97—103, 1992
15. MACKENZIE RK, COLES GA, WIlLIAMS JD: Eicosanoid synthesis inhuman peritoneal macrophages stimulated with S. epidermidis. KidneyIn! 37:1316—1324, 1990
16. MACKENZIE RK, COLES GA, WILLIAMS JD: The response of humanpenitoneal macrophages to stimulation with bacteria isolated fromepisodes of continuous ambulatory peritoneal dialysis-related perito-nitis. J Infect Dis 163:837—842, 1991
17. TOPLEY N, JORRES A, LUTrMANN W, PErERsEN M, LANG M, THIER-AUsC1I K-H, MULLER C, COLES G, DAVIES M, WILLIAMS J: Humanperitoneal mesothclial cells synthesize IL-6: Induction by IL-i[3 andTNFa. Kidney In! 43:226—233, 1993
18. STYLIANOU E, JENNER LA, DAVIES M, COLES GA, WILLIAMS JD:Isolation, culture and characterization of human peritoneal mesothe-hal cells. Kidney In! 37:1563—1570, 1990
19. TOPLEY N, BROWN Z, JORRES A, WESTWICK J, COLES G, DAVIES M,WILLIAMS J: Human peritoneal mesothelial cells synthesize IL-8:Synergistic induction by intrleukin-1[3 and tumor necrosis factor a.Am J Pat/wI 142:1876—1886, 1993
20. RUDINBAUGH MG, CAMPBELL WH: Adaptation of the dye-bindingprotein assay to niicrotitre plates. Anal Biochem 147:144—151, 1985
21. JORRES A, TOPLEY N, STEENWEG L, MULLER C, KUI-Ic;LN E, GAHLGM: Inhibition of cytokine synthesis by peritoneal dialysate persiststhroughout the CAPD cycle. Am J Nephro/ 12:80—85, 1992
22. MANIATIS T, FRI1SCH E, SAMBROOK J: Extraction, Purification, andAnalysis of mRNA from Eucaiyotic Cells. New York, Cold SpringHarbour Laboratory, 1987, p 187
23. O'BISYAN JP, FRYE RA, COGSWELL PC, NEUBAUER A, KIICII B,PROKOP C, ESPINOZA R III, LE BMM, EARP HS, Liu ET: axi, atransforming gene isolated from primary human myeloid leukemiacells, encodes a novel receptor tyrosine kinase. Mol Cell Biol 11:5016—5031, 1991
24. ZILBERSTEIN A, RUGGIERI R, KORN JH, REVEL M: Structure and
expression of eDNA and genes for human interferon beta2 (IL-6) adistinct species inducible by growth stimulatory cytokines. Embo J5:2529—2537, 1986
25. HARIGAI M, HARA M, KITA! A, NORIOKA K, HIROSE T, HIR0SE W,SUZUKI K, KAWAKAMI M, MASUDA K, SHINMEI M: Interleukin I andtumor necrosis factor-a synergistically increase the production ofinterleukin 6 in human synovial fibroblasts. J Clin Lab Immunol34:107—1 13, 1991
26. TOPLEY N, FLOEGE J, WESSEL K, HASS R, RADEKE HH, KAEVER V,RESCH K: Prostaglandin E2 production is synergistically increased incultured human glomerulan mesangial cells by combinations of IL-Iand tumor necrosis factor-alpha. J Immuno/ 143:1989—1995, 1989
27. BETJE5 MGH, TUK CW, STRUIJK DG, KREDIET RT, ARISZ L, HARTM, BEELEN RH: lnterleukin-8 production by human peritoneal me-sothehial cells in response to tumor necrosis factor a, interleukin-1,and medium conditioned by macrophages co-cultured with Staphylo-coccus epidermidis. J Inf Dis 168:1202—1210, 1993
28. TOPLEY N, PETERSEN MM, MACKENZIE R, KAEVER V, NEUBAUER A,STYLIANOU E, COLES GA, DAVIES M, JORRES A, WILLIAMS JD:Human penitoneal mesothehial cell prostaglandin synthesis: Inductionof cyclooxygenase mRNA by peritoneal macrophage derived cyto-kines. Kidney hit 46:900—909, 1994
29. FIEREN MWJA, VAN DEN BEMD GJCM, BONTA IL: Endotoxin-
stimulated peritoneal macrophages obtained from continuous ambu-latory penitoneal dialysis patients show increased capacity to releaseinterleukin-lbeta in vitro during peritonitis. EurJ C/in Invest 20:453—457, 1990
30. DOUVDEVANI A, RAPOPORT J, KONFORTY A, ARGOV 5, OVNAT A,CHAIMOWITZ C: Human peritoneal mesothehial cells synthesisze IL-laand [3. Kidney Int 46:993—1001, 1994
31. ROLA-PLESZCZYNSKI M, STANKOVA J: Leukotriene B4 enhances inter-leukin-6 (IL-6) production and messenger RNA accumulation inhuman m000cytes in vitro: Transcriptional and postranscniptionalmechanisms. Blood 80:1004—10 11, 1992
32. HIRANO T, AKIRA 5, TAGA T, KIsHIM0ro T: Biological and clinicalaspects of interlcukin-6. Immunol Today 11:443—449, 1990
33. BRACH MA, DE Vos 5, ARNOLD C, GRUB H-J, MERTESMANN R,HERRMANN F: Leukotniene B4 trabscriptionally activates interleukin-6expression involving NF-eB and NF-1L6. Eur J Immunol 22:2705—2711, 1992
34. NAKAZATO Y, HAYASHIDA T, KANNO Y, OKAMIYA Y, SASAMURA H,SUZUKI H, SARUTA T: Synergistic effects of IL-h and IL-4 on NF-1L6
activities and IL-6 production in human mesangial cells. (abstract) JAm Soc Nephrol 6:804, 1995
35. MCGEE DW, BEAGLEY KW, AICHER WK, MCGEE JR: Transforminggrowth factor-p and IL-113 act in synergy to enhance IL-6 secretion bythe intestinal cell line, IEC-6. I Jmmuno/ 15 1:970—978, 1993
36. KUPPNER MC, MCKIILOP-SMITH SJVF: TGF-p and IL-p act insynergy to enhance IL-6 and IL-8 mRNA levels and lL-6 productionby human retinal pigment epithehial cells. Immunology 84:265—227,1995
37. Kii-nvio'ro T: The biology of intenleukin-6. Blood 74:1—10, 198938. ADERKA D, LE J, VILCEK J: IL-6 inhibits hipopolysaccharide-induced
tumor necrosis factor production in cultured human monocytes, U937cells, and in mice. I Immunol 143:3517—3523, 1989
39. SCIIINDLER R, MANCILLA J, ENDRES 5, GHORBANI R, CLARK SC,DINARELLO CA: Correlations in the production of interleukin-6. IL-iand TNFalpha in human blood mononuclear cells: IL-6 suppresses
IL-I and TNF. Blood 75:40—47, 1990
40. TILG H, TREIIU E, AIKINS MB, DINARELLO CA, Mu-B JW: Interlcu-kin-6 (IL-6) as an anti-inflammatory cytokine: Induction of circulating
IL-i receptor antagonist and soluble tumor necrosis factor receptorp55. Blood 83:113—118, 1994
41. TOPILY N, WILLIAMS JD: The role of the penitoneal membrane in thecontrol of inflammation in the penitoneal cavity. Kidney hit 46(Suppl48):S7l—S78, 1994





























