Embed Size (px)
Citation preview

Journal of Comparative and Physiological Psychology1980, Vol 94, No 2,289-304
Subtotal Lesions of the Visual Cortex ImpairDiscrimination of Hidden Figures by Cats
Paul Cornwell, William Overman, and Alfonso CampbellPennsylvania State University
Cats with partial or nearly total ablation of areas 17, 18, and 19 were assessedon the discrimination of hidden figures and other visually guided behaviors todetermine whether such insults produce deficits like those that follow lateralstriate lesions in monkeys. Cats with destruction limited to the representa-tion of central vision (Group M) were impaired at discriminating patternscomplicated by extraneous cues, but they were less impaired than cats withmore complete lesions (Group MS). The deficit was not a general one in visu-al learning since animals in both Groups M and MS learned simple patterndiscriminations as rapidly as controls. It is suggested that the loss of genicu-locortical functions representing central vision produces similar deficits mcats and monkeys but that to have this effect in cats, damage must extend be-yond area 17.
A number of common features charac-terize the cortical visual systems of mam-mals, (a) Most have a core of primary visualkoniocortex, with a relatively thick fourthlayer (area 17), which is surrounded by oneor more belts having a less prominent fourthlayer (Gray, 1924; Hall & Diamond, 1968;Kaas, Hall, & Diamond, 1972; Sanides &Hoffmann, 1969). (b) The core cortex is atarget of the thalamocortical projectionsfrom the dorsal division of the lateral ge-niculate nucleus (LGNd), and (c) the beltcortex (areas 18 and 19 and other regions)receives projections from the pulvinar andlateral posterior nuclei (Diamond, 1976;Diamond & Hall, 1969; Hughes, 1977; Jones,1974; M. Wilson, 1978). (d) The pattern ofcorticofugal projections from core and beltcortex to the thalamus and superior collic-ulus is similar in most mammals, as are (e)
This research was supported by Grants MS 04726 andNS 10819 from the U.S Public Health Service. Theauthors wish to thank Bryan Kolb, Richard Ravizza,and J. M. Warren for their critical review of the manu-script. Parts of this research were reported at themeeting of the Eastern Psychological Association,Philadelphia, April 1974.
William Overman is now at the University of NorthCarolina at Wilmington, and Alfonso Campbell is nowat the University of Michigan.
Requests from outside North America for reprintsshould be sent to Paul Cornwell, 130 Moore Building,Pennsylvania State University, University Park,Pennsylvania 16802.
many corticocortical connections (Jones,1974; Lund, Henry, MacQueen, & Harvey,1979). (f) In all mammalian species studiedone finds multiple orderly representationsof the visual field in both the thalamus andthe cortex; there are at least 13 separaterepresentations of the visual field at thecortical level in cats (Tusa, Palmer, & Ros-enquist, 1978), and the situation is probablyat least as complex in primates (M. Wilson,1978; Zeki, 1974,1975).
Species differ, however, in other aspectsof the cortical visual system. A major ana-tomical difference between monkeys andcats, for example, is in the relation of LGNdto the cerebral cortex. Although LGNdprojects to the visual core in both domesticcats and rhesus monkeys, it sends additionalprojections to widespread areas of the visualbelt in cats, but not in monkeys (Garey, 1970;Hollander & Vanegas, 1977; Rosenquist,Edwards, & Palmer, 1974; M. Wilson &Cragg, 1967). Thus, the only route to thebelt cortex from LGNd in monkeys isthrough area 17 (Kuypers, Szwarcbart,Mishkin, & Rosvold, 1965; Martinez-Millan& Hollander, 1975; Zeki, 1975). In contrast,the belt cortex in cats receives two imputsfrom LGNd, one directly and the other viaarea 17 (Hubel & Wiesel, 1965; Rosenquistet ah, 1974).
Physiological experiments yield resultsthat are completely consonant with these
Copyright 1980 by the American Psychological Association, Inc 0021 -9940/80/9402-0289$00 75
289

290 P. CORNWELL, W OVERMAN, AND A. CAMPBELL
anatomical differences. Inactivation of area17 in monkeys abolishes the ability of cellsin area 18 to respond to visual stimuli(Schiller & Malpeli, 1977), but such treat-ments have far less effect on the activity ofcells in area 18 in cats (Donaldson & Nash,1975; Dreher & Cottee, 1975; Sherk, 1978).
The primary goal of the present study wasto relate these anatomical and physiologicaldifferences to differences in the behavioraleffects of lesions in the visual cortex ofmonkeys and cats. One major differencebetween monkeys and cats in the effects ofstriate lesions on behavior is already wellknown. Complete destruction of area 17greatly impairs the discrimination of visualpatterns by monkeys; only with prolongedtraining do striatectomized monkeys revealthe capacity to discriminate figures thatdiffer in contour but are equal in luminousflux (Humphrey, 1974; Keating, 1975; Klii-ver, 1942; Pasik & Pasik, 1971; Weiskrantz,1963). Cats with lesions confined to area 17,in contrast, are little affected in postopera-tive tests of pattern vision (Doty, 1971;Sprague, Levy, DiBerardino, & Berlucchi,1977). However, cats with destruction of theprojection fields of LGNd in both core andbelt cortices are severely impaired in visuallyguided behavior (Cornwell, Warren, &Nonneman, 1976; Dalby, Meyer, & Meyer,1970; Murphy, Mize, & Schechter, 1975;Spear & Braun, 1969; Sprague et al., 1977;Wetzel, 1969; Wetzel, Thompson, Horel, &Meyer, 1965; Wood, Spear, & Braun, 1974).They, like striatectomized monkeys, even-tually learn to discriminate simple patterns,but they have serious trouble differentiatingshapes of equal luminous flux that are alsoequal or nearly equal in contour and com-plexity (Cornwell, Overman, & Ross, inpress; Ritchie, Meyer, & Meyer, 1976). Theeffects of removing all or nearly all of theprojection field of LNGd are, therefore, verysimilar in monkeys and cats, but the relativesize of the lesions needed to accomplish thisdisruption is far greater in cats than inmonkeys.
The effect of removing only the cortexrepresenting central vision has been exam-ined systematically in rhesus monkeys butnot in cats. In monkeys the central 6-8° ofthe visual field is represented on the lateral
striate (LS) cortex (Daniel & Whitteridge,1961). Removal of this region does not se-riously impair rhesus monkeys on many sortsof pattern discrimination tasks, but monkeyswith LS lesions are defective in performanceon tasks that require the discrimination offine differences in visual size (Butter &Doehrman, 1968; W. Wilson & Mishkin,1959), spatial frequency (Weiskrantz &Cowey, 1963), and flicker rates (Mishkin &Weiskrantz, 1959). Monkeys with LS le-sions are also impaired on patterned strings(W. Wilson & Mishkin, 1959) and hiddenfigures (Butter, 1969,1972,1979) tasks, andthe relatively minor decreases in acuity seeminsufficient to account for these impairments(Weiskrantz & Cowey, 1963).
Nothing is known precisely about the ef-fects of the lesions confined to the corticalrepresentation of the central part of the vi-sual field in cats. Several investigators agreein finding that small remnants of the ge-niculocortical system can mediate a sub-stantial degree of visual capacity in cats(reviewed by Frommer, 1978). This work isof limited value for comparative purposes,however, since the lesions were not restrictedto the representation of central vision andperformance of the subjects was not testedon tasks like those used to assess the conse-quences of LS lesions in monkeys.
The specific aim of the present experi-ment was to ascertain whether the deficitsresulting from destruction of the geniculo-cortical representation of central vision incats resemble those observed in monkeyswith LS lesions. Since LGNd projects bothto belt and core cortex in cats, it was neces-sary to ablate cortex in areas 18 and 19 aswell as in area 17 to destroy most of thegeniculocortical system representing centralvision. This allowed comparisons with be-havioral findings with monkeys having LSlesions, in which the total geniculocorticalsystem representing central vision was de-stroyed by lesions restricted to the striatecortex.
Three groups of cats were studied: (a)normal controls, (b) cats with lesions in-tended to destroy the total projection ofLGNd in areas 17, 18, and 19, and (c) catswith lesions restricted to the representationof central vision in areas 17,18, and 19. This

VISUAL CORTEX AND HIDDEN FIGURES 291
design permitted comparison between catswith defects in central vision and monkeyswith LS lesions. It also provided the op-portunity to compare the performance ofcats with definitely subtotal and with nearlycomplete ablations of the projection field ofLGNd. The tasks (Table 1) were chosen tomaximize comparability with tests used withmonkeys having LS lesions; the major com-parison was between performance on con-ventional tests of pattern vision and per-formance on tasks that involved spatialmasking. The chief finding was that catswith lesions limited to the representation ofcentral vision, like monkeys with LS lesions,were disrupted very little in the conventionaltests but were greatly impaired on those thatinvolved spatial masking.
Method
Subjects
The 14 adult laboratory-reared mongrel cats had ailserved previously in an experiment involving reversalsof a probability learning problem with spatial cues(Warren, Note 1), but they had no preoperative trainingon any visual discrimination task
Apparatus
All discrimination training was conducted in a mod-ified Grice box described in detail elsewhere (Cornwell,Overman, & Ross, 1978). In brief, the apparatus con-sisted of a starting box and a choice compartment whichled to two goal boxes that were separated by an opaquepartition In each goal box was a vertically mountedstimulus plaque which subtended 9°-25° of visual anglewhen viewed from the choice point The cat had to gobehind one of the plaques to obtain a hidden piece ofpork kidney or tuna fish. In all but two of the tasks, thetwo goal boxes were painted white and the rest of theapparatus was painted gray For two tasks (Tasks 12and 13) the box was painted gray throughout
The visual cliff (Task 3) has been described in detailpreviously (Cornwell, Overman, Levitsky, Shipley, &Lezynski, 1976) Visual tracking (Task 17) was studiedin the home cage with a tassel of brown yarn, 4 cm indiameter, tied to the end of a wand 93 cm long and madeof coat-hanger wire
Shape and Pattern Stimuli
Hexagonal and hourglass shapes were cut from 2-cm-thick lumber, painted black, and mounted verticallyon Plexiglas bases (Figure 1) The original trainingshapes were each 80 cm2; those used to assess shapediscrimination in the presence of irrelevant size cueswere 80 cm2, 40 cm2, and 20 cm2. L-shapes, each 12.5
6a
LJFigure 1 Stimuli used for hidden figures test (Tasks5-6e), simple pattern discrimination (Task 8), and or-ientation discrimination (Task 9), drawn to scale(Only the L-shapes had a third dimension to them.)
X 11 5 cm, were cut from 4-cm-thick lumber and werepainted black All pattern stimuli were made fromblack construction paper mounted on white plywoodplaques and are displayed to scale in Figure 1 Thestimuli for the landmark discriminations were a pair ofvertically oriented gray triangles cut from 2-cm-thicklumber, 10 5 cm in altitude and 12 cm at the base Thelandmark was a 16 oz Coca Cola bottle, 28 cm high andpainted flat black.
A silhouette of a cat was cut from black constructionpaper and is shown in Figure 5, which also shows acontrol pattern made by rearrangement of fragmentsof the cat figure. Each silhouette was mounted on asheet of gray cardboard 70 X 98 cm and placed behinda sheet of nonreflective glass attached to one wall of aroom at floor level by a wooden frame The floor of theroom was 1.8 X 2 4 m and was marked off into 12 num-bered squares.
Procedure
Table 1 shows the order of presentation for the 2preoperative and 15 postoperative tests, along with thenumber of trials administered per session for each of thetasks. Several of the postoperative tests contained oneor more subtests, and these too are listed in Table 1 Onall discrimination learning tasks but the final two(Tasks 14 and 15), the criterion for acquisition was 90%or more correct responses. Table 1 indicates, for eachtask, whether this criterion needed to be met for one orfor two consecutive sessions
Visual cliff (Task 3) Each cat was given five trialsper day on the visual cliff for 4 consecutive days De-

292 P. CORNWELL, W. OVERMAN, AND A. CAMPBELL
tails of the testing procedure employed are given else-where (Comwell et al., 1976).
Visual discrimination training (Tasks 1, 2, and4-15) Training occurred under 15-24-hr food depriva-tion and was conducted in a Grice box in which allstimuli were displayed with reflected light. A trialbegan when the experimenter lifted the opaque door tothe starting compartment and, 3 sec later, raised thetransparent door to release the cat. The reward wasplaced behind one of the stimuli, and the other stimulusremained unbaited. A noncorrection procedure wasused throughout.
The position of the positive stimulus was varied fromtrial to trial according to a balanced irregular sequence.On those tasks that required the discrimination of formor pattern in the presence of irrelevant variations in thesize of the stimuli, each of the three sizes of the rein-forced shape was paired with each size of the nonrein-forced shape on 4 trials in the 36-tnal session. OnTasks 9-11, the same stimulus of each pair was rein-forced for all subjects with the reinforced member of thepairs as follows Task 8, grid; Task 9, right-facing L,Task 10, inverted triangle; and Task 11, oblique line.
Many of the discriminations were related to the initial
discrimination of the hexagon versus hourglass acquiredpreoperatively. Approximately half of the cats in eachgroup were trained with the hexagon positive and theother half with the hourglass positive. On all subse-quent tests, the basic shape of the reinforced stimulusremained the same for each cat. Thus, cats with thehexagonal form positive on the initial discriminationwere trained with an outline hexagonal positive duringtesting with stripes superimposed on the patterns (Task6) and were trained with the circle positive in the taskthat required the discrimination of a circle from an I(Task 7).
During all discrimination training except Task 12, asimultaneous procedure was used, in which both dis-criminanda were presented on each trial with one re-inforced. Task 12 involved the successive discrimina-tion of a circle and an I-pattern and was done in a Gricebox with gray goal compartments. Here, a pair of cir-cles was presented on half of the trials, with only re-sponses to the left member of the pair reinforced. Onthe other half of the trials a pair of Is was present, andonly responding to the stimulus on the right was rein-forced. The order of stimulus presentation followed abalanced irregular sequence.
Table 1Order of Presentation of Tasks Pre- and Postoperatively
Task Trials/session Criterion sessions
Preoperative1 Shape2 Shape: Size irrelevant
Postoperative3 Visual cliff4. Shape: Size irrelevant5 Pattern6. Outline pattern
a Outline onlyb. 2 superimposed stripesc. 4 superimposed stripesd. 6 superimposed stripese 10 superimposed stripes
7. Pattern: Size irrelevant (circle vs I)8. Pattern- Grid vs square bulls-eye9. Orientation: L vs. J
10. Orientationa. Upright vs inverted triangleb. Above triangles surrounded by squares
11. Orientationa. Oblique vs. horizontal lineb. Above lines surrounded by linesc. Above lines surrounded by squares
12. Pattern: Successive13. Outline pattern: Retention14. Landmark
a. Acquisition"b-d. Reversals"
15. Outline patterns: Superimposed stripesb
16. Cat model17. Visual tracking
2536
53625
2525252525362525
2525
2525252525
5050402.5 min8
22
22
22222211
11
11111
* Criterion was 90% correct responses in each of two consecutive blocks of 25 trials.b Tested a fixed 200 trials (not to a criterion).

VISUAL CORTEX AND HIDDEN FIGURES 293
Task 13 was also conducted in the Grice box withsolid gray goal compartments. In this retention test,the same outline patterns were employed as were usedin Test 6a, but here the white stimulus plaques werepresented against a gray background rather than a whiteone. One subject from Group C and one from GroupMS (Subject MS-1) were not tested on this particulartask.
The procedure in the landmark discrimination (Task14) was the same as that used previously by Campbell(1978). All testing was conducted in the apparatus withwhite goal compartments, and all 14 subjects weretested. First, the cat was reinforced for approachingthe member of a pair of identical gray triangles that wasadjacent to the landmark. Subjects were given 50 trialsper day, and the criterion was 90% correct responses ineach of two consecutive blocks of 25 trials. When cri-terion was attained, the cat was required to approachthe triangle that did not have the bottle next to it (Re-versal 1) Training to approach the stimulus unac-companied by the landmark continued until the initialcriterion was again attained, at which time the cat wasonce more required to approach the triangle next to thebottle (Reversal 2) A third reversal of the landmarkdiscrimination to the same criterion completed testingon this task.
The final test of the discrimination learning series(Task 15) used the outline stimuli with superimposedstripes employed previously (Tasks 6b-6e). Each of thefour stimulus pairs (2,4,6, or 10 stripes) was presentedon 10 trials of of each 40-tnal session The order otstimulus presentation was randomized, with the re-striction that no stimulus pair could appear more thanthree times in a row. All cats received a total of 200trials over five consecutive testing sessions
Responses to cat model {Task 16) Testing consistedof three habituation sessions followed by four sessionswith test patterns. Each cat was first habituated in-dividually to the small unfamiliar room for 2.5 min perday for 3 consecutive days During this baseline periodthe cat's behavior was monitored with a time samplingprocedure by two observers through a one-way glasswindow. One observer recorded the position of the catin the room and the orientation of its head at the end ofeach 2.5-sec period, marked by a beep audible to bothof the observers and to the cat The second observerrecorded the behavior of the cat at the end of each2.5-sec observation period The procedure for classi-fying the behaviors was the same as that employedpreviously by Nonneman and Kolb (1974) and consistedof behaviors including piloerection, crouching, andsniffing the region where the cat model appeared. An"interest and affect" score was obtained by taking thesum of the number of intervals during which one ormore of these behaviors were.observed.
At the start of a trial the cat was placed in the dark-ened room, and within 2.5 sec the overhead lights wereturned on and the observation period was begun.During the initial three sessions the picture frame at-tached to one wall contained only a blank sheet of graycardboard. On Sessions 4 and 7 the silhouette of a catwas presented in place of the blank cardboard, and onSessions 5 and 6 the fragmented cat was presented(Figure 5)
Visual tracking (Task 17) Visual orienting and
tracking was assessed by moving a tassel of yarn towardthe center of the cat's visual field at about 5 rad persecond. One observer gained the subject's attention sothat its head was at approximately the center of thefront of the cage. A second observer then moved thetassel in from one corner of the cage in such a way thatthe individual strands of yarn jiggled slightly. Per-formance was rated independently by the two observerson a 5-point scale, with 0 indicating no orientationtoward the target and 4 representing immediate, accu-rate, and smooth tracking of the target Four trialswere conducted within a period of about 60 sec, one trialstarting from each of the four quadrants. A secondblock of four trials was administered 15 to 60 min later.The following day another eight trials were given in thesame way. Thus, a subject could obtain a maximumscore of 64 over the two sessions. The interjudgeagreement for the ranking of the total scores was nearly-perfect (p = .99). All nine operated subjects were as-sessed, but only two of the five controls were availablefor testing
Surgery
All subjects were operated on within 8 days of com-pleting their preoperative training. Cats were assignedto groups so that the median number of errors madeduring acquisition of the shape discrimination was ap-proximately equal for the three groups.
The surgery was conducted under sodium pento-barbital anesthesia (36 mg/kg), with sterile precautionsFive cats received control operations which involvedonly scalp incisions (Group C). In a second five animals(Group M) the marginal and posterior lateral gyri wereremoved by subpial aspiration from a point about 1 cmposterior to the ansate sulcus to the ventral limit of theposterior lateral gyrus. For these cats an attempt wasmade to spare all the cortex along the dorsal bank of thesplenial sulcus Thus, the lesions for Group M weredesigned to spare much of the visual cortex representingthe peripheral parts of the visual field (Kalia & Whit-teridge, 1973; Tusa et al., 1978).
In the four cats in Group MS the surgical procedurewas identical to that used for Group M, but in addition,an attempt was made to remove all the cortex on theupper bank of the splenial sulcus. In these cases mostof the white matter of the marginal and posterior lateralgyri was also removed. These lesions, then, were in-tended to remove area 17 completely and to includemost of areas 18 and 19 as well (Otsuka & Hassler, 1962;Sanides & Hoffmann, 1969). A 1-mo postoperativerecovery period was allowed for all cats before the re-sumption of behavioral testing.
Histology
At the completion of behavioral testing, the operatedcats were killed with an overdose of sodium pentobar-bital and were perfused intracardially with physiologicalsaline followed by 10% formalin in saline The brainswere photographed, dehydrated, embedded in celloidin,and cut in 30-jtm sections. Every 10th coronal sectionthrough the lesion and the thalamus was mounted andstained with cresyl violet. The lesions were recon-
294 P. CORNWELL, W OVERMAN, AND A. CAMPBELL
structed from the serial sections together with thephotographs of the brains, and the posterior region ofthe thalamus was analyzed for the location and severityof retrograde degeneration
Results
Histological
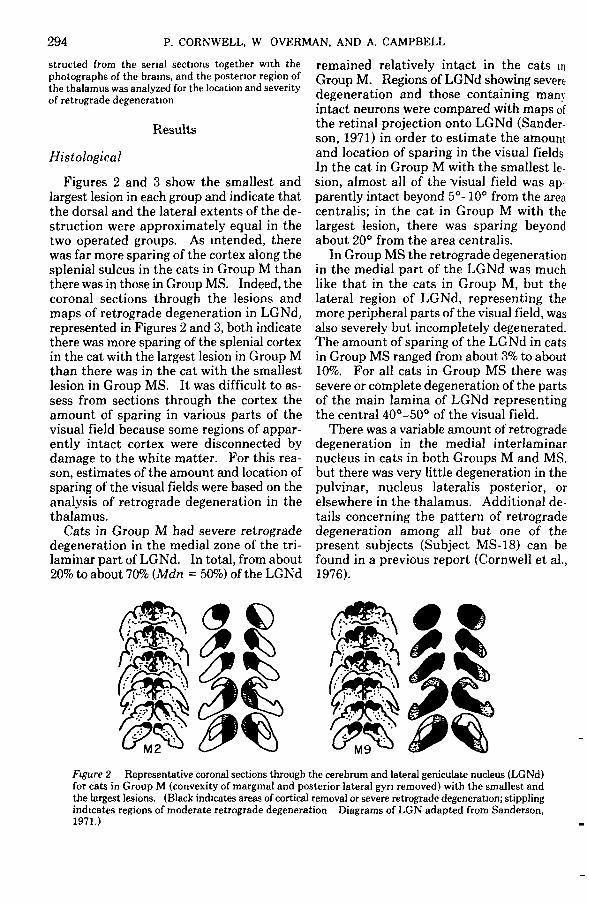
Figures 2 and 3 show the smallest andlargest lesion in each group and indicate thatthe dorsal and the lateral extents of the de-struction were approximately equal in thetwo operated groups. As intended, therewas far more sparing of the cortex along thesplenial sulcus in the cats in Group M thanthere was in those in Group MS. Indeed, thecoronal sections through the lesions andmaps of retrograde degeneration in LGNd,represented in Figures 2 and 3, both indicatethere was more sparing of the splenial cortexin the cat with the largest lesion in Group Mthan there was in the cat with the smallestlesion in Group MS. It was difficult to as-sess from sections through the cortex theamount of sparing in various parts of thevisual field because some regions of appar-ently intact cortex were disconnected bydamage to the white matter. For this rea-son, estimates of the amount and location ofsparing of the visual fields were based on theanalysis of retrograde degeneration in thethalamus.
Cats in Group M had severe retrogradedegeneration in the medial zone of the tri-laminar part of LGNd. In total, from about20% to about 70% (Mdn = 50%) of the LGNd
remained relatively intact in the cats inGroup M. Regions of LGNd showing severedegeneration and those containing manyintact neurons were compared with maps ofthe retinal projection onto LGNd (Sander-son, 1971) in order to estimate the amountand location of sparing in the visual fieldsIn the cat in Group M with the smallest le-sion, almost all of the visual field was ap-parently intact beyond 5°-10° from the areacentralis; in the cat in Group M with thelargest lesion, there was sparing beyondabout 20° from the area centralis.
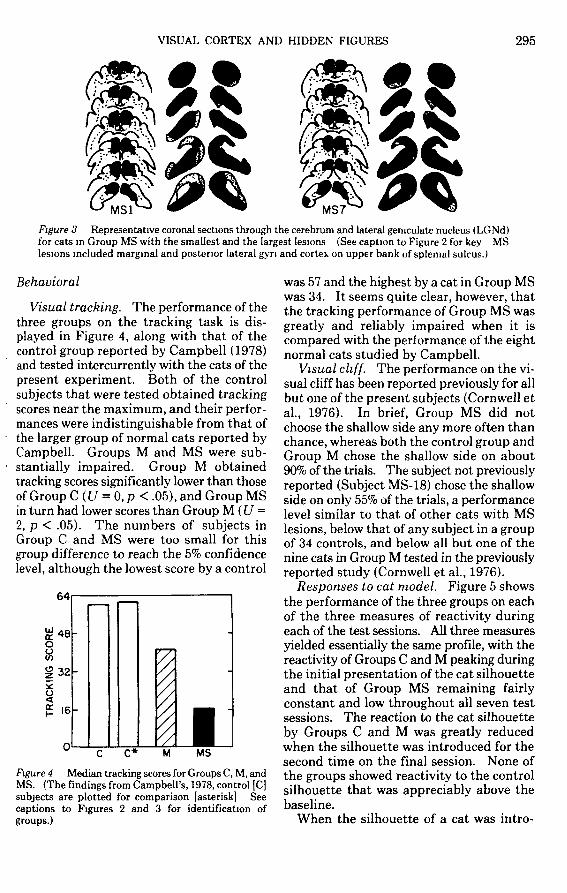
In Group MS the retrograde degenerationin the medial part of the LGNd was muchlike that in the cats in Group M, but thelateral region of LGNd, representing themore peripheral parts of the visual field, wasalso severely but incompletely degenerated.The amount of sparing of the LGNd in catsin Group MS ranged from about 3% to about10%. For all cats in Group MS there wassevere or complete degeneration of the partsof the main lamina of LGNd representingthe central 40°-50° of the visual field.
There was a variable amount of retrogradedegeneration in the medial interlaminarnucleus in cats in both Groups M and MS.but there was very little degeneration in thepulvinar, nucleus lateralis posterior, orelsewhere in the thalamus. Additional de-tails concerning the pattern of retrogradedegeneration among all but one of thepresent subjects (Subject MS-18) can befound in a previous report (Cornwell et al.,1976).
milFigure 2 Representative coronal sections through the cerebrum and lateral geniculate nucleus (LGNd)for cats in Group M (convexity of marginal and posterior lateral gyri removed) with the smallest andthe largest lesions. (Black indicates areas of cortical removal or severe retrograde degeneration; stipplingindicates regions of moderate retrograde degeneration Diagrams of LGN adapted from Sanderson,1971.)
VISUAL CORTEX AND HIDDEN FIGURES 295
MS7Figure 3 Representative coronal sections through the cerebrum and lateral geniculate nucleus (LGNd)for cats in Group MS with the smallest and the largest lesions (See caption to Figure 2 for key MSlesions included marginal and posterior lateral gyn and cortex on upper bank of splenial sulcus.)
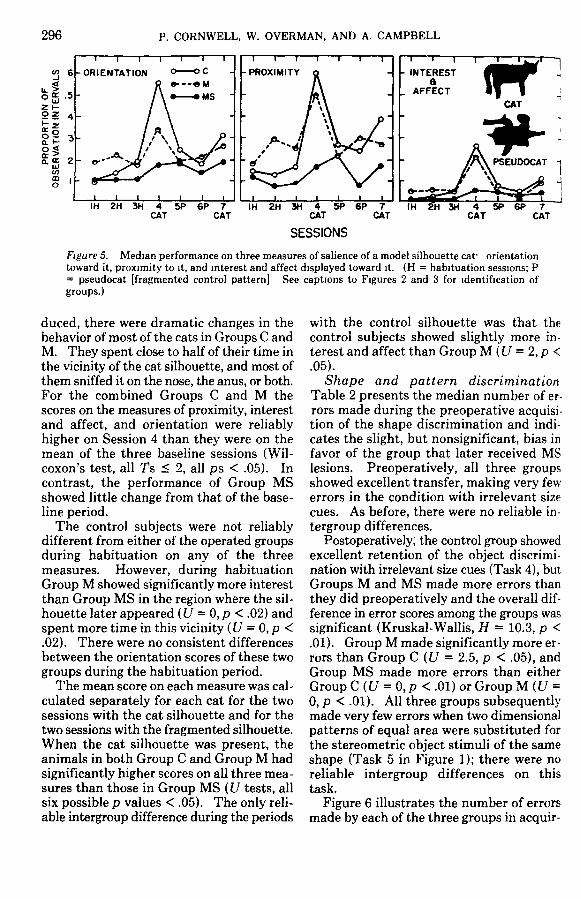
Behavioral
Visual tracking. The performance of thethree groups on the tracking task is dis-played in Figure 4, along with that of thecontrol group reported by Campbell (1978)and tested intercurrently with the cats of thepresent experiment. Both of the controlsubjects that were tested obtained trackingscores near the maximum, and their perfor-mances were indistinguishable from that ofthe larger group of normal cats reported byCampbell. Groups M and MS were sub-stantially impaired. Group M obtainedtracking scores significantly lower than thoseof Group C (U = 0,p < .05), and Group MSin turn had lower scores than Group M (U =2, p < .05). The numbers of subjects inGroup C and MS were too small for thisgroup difference to reach the 5% confidencelevel, although the lowest score by a control
64
£48O
H •
O
1 16
O L
C* M
Figure 4 Median tracking scores for Groups C, M, andMS. (The findings from Campbell's, 1978, control [C]subjects are plotted for comparison [asterisk] Seecaptions to Figures 2 and 3 for identification ofgroups.)
was 57 and the highest by a cat in Group MSwas 34. It seems quite clear, however, thatthe tracking performance of Group MS wasgreatly and reliably impaired when it iscompared with the performance of the eightnormal cats studied by Campbell.
Visual cliff. The performance on the vi-sual cliff has been reported previously for allbut one of the present subjects (Cornwell etal., 1976). In brief, Group MS did notchoose the shallow side any more often thanchance, whereas both the control group andGroup M chose the shallow side on about90% of the trials. The subject not previouslyreported (Subject MS-18) chose the shallowside on only 55% of the trials, a performancelevel similar to that of other cats with MSlesions, below that of any subject in a groupof 34 controls, and below all but one of thenine cats in Group M tested in the previouslyreported study (Cornwell et al., 1976).
Responses to cat model. Figure 5 showsthe performance of the three groups on eachof the three measures of reactivity duringeach of the test sessions. All three measuresyielded essentially the same profile, with thereactivity of Groups C and M peaking duringthe initial presentation of the cat silhouetteand that of Group MS remaining fairlyconstant and low throughout all seven testsessions. The reaction to the cat silhouetteby Groups C and M was greatly reducedwhen the silhouette was introduced for thesecond time on the final session. None ofthe groups showed reactivity to the controlsilhouette that was appreciably above thebaseline.
When the silhouette of a cat was intro-
296 P. CORNWELL, W. OVERMAN, AND A. CAMPBELL
IH 2H 3H 4 5P 6P 7CAT CAT
IH 2H 3H 4 5P 6P 7CAT CAT
SESSIONS
3H 4 5P 6P 7CAT CAT
Figure 5. Median performance on three measures of salience of a model silhouette cat- orientationtoward it, proximity to it, and interest and affect displayed toward it. (H = habituation sessions; P= pseudocat [fragmented control pattern] See captions to Figures 2 and 3 for identification ofgroups.)
duced, there were dramatic changes in thebehavior of most of the cats in Groups C andM. They spent close to half of their time inthe vicinity of the cat silhouette, and most ofthem sniffed it on the nose, the anus, or both.For the combined Groups C and M thescores on the measures of proximity, interestand affect, and orientation were reliablyhigher on Session 4 than they were on themean of the three baseline sessions (Wil-coxon's test, all Ts < 2, all ps < .05). Incontrast, the performance of Group MSshowed little change from that of the base-line period.
The control subjects were not reliablydifferent from either of the operated groupsduring habituation on any of the threemeasures. However, during habituationGroup M showed significantly more interestthan Group MS in the region where the sil-houette later appeared (U = 0, p < .02) andspent more time in this vicinity (U = 0, p <.02). There were no consistent differencesbetween the orientation scores of these twogroups during the habituation period.
The mean score on each measure was cal-culated separately for each cat for the twosessions with the cat silhouette and for thetwo sessions with the fragmented silhouette.When the cat silhouette was present, theanimals in both Group C and Group M hadsignificantly higher scores on all three mea-sures than those in Group MS (U tests, allsix possible p values < .05). The only reli-able intergroup difference during the periods
with the control silhouette was that thecontrol subjects showed slightly more in-terest and affect than Group M (U = 2, p <.05).
Shape and pattern discriminationTable 2 presents the median number of er-rors made during the preoperative acquisi-tion of the shape discrimination and indi-cates the slight, but nonsignificant, bias infavor of the group that later received MSlesions. Preoperatively, all three groupsshowed excellent transfer, making very few-errors in the condition with irrelevant sizecues. As before, there were no reliable in-tergroup differences.
Postoperatively; the control group showedexcellent retention of the object discrimi-nation with irrelevant size cues (Task 4), butGroups M and MS made more errors thanthey did preoperatively and the overall dif-ference in error scores among the groups wassignificant (Kruskal-Wallis, H = 10.3, p <.01). Group M made significantly more er-rors than Group C (U = 2.5, p < .05), andGroup MS made more errors than eitherGroup C (U = 0, p < .01) or Group M (U =0, p < .01). All three groups subsequentlymade very few errors when two dimensionalpatterns of equal area were substituted forthe stereometric object stimuli of the sameshape (Task 5 in Figure 1); there were noreliable intergroup differences on thistask.
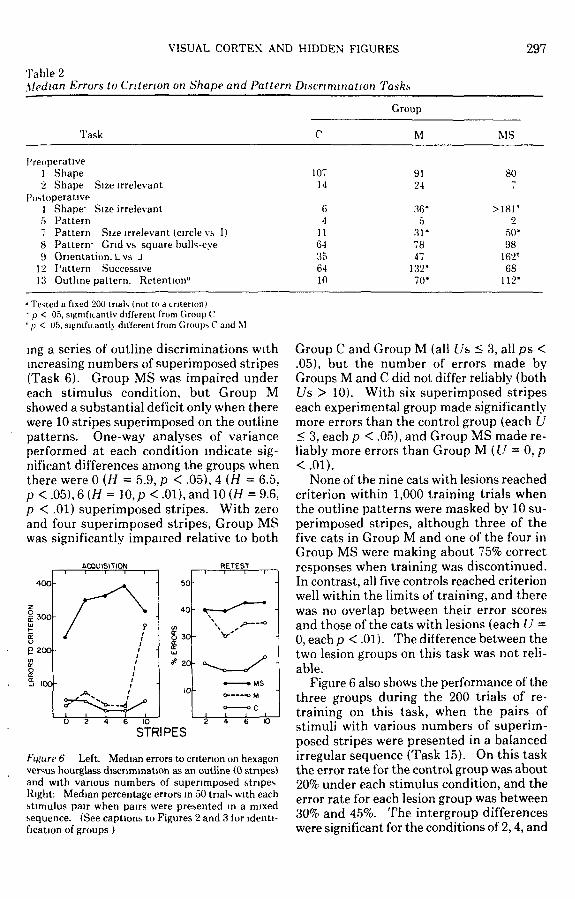
Figure 6 illustrates the number of errorsmade by each of the three groups in acquir-
VISUAL CORTEX AND HIDDEN FIGURES 297
Table 2.Median Errors to Criterion on Shape and Pattern Discrimination Tasks
Task
I'rooperative1 Shape2 Shape Size irrelevant
Postoperative1 Shape' Size irrelevant5 Pattern7 Pattern Size irrelevant (circle vs I)8 Pattern- Grid vs square hulls-eye9 Orientation. L vs J
12 Pattern Successive13 Outline pattern. Retention"
C
10714
64
1164356410
Group
M
9124
36*5
31*7847
132*70*
MS
80
>I81 '2
50*98
162'68
112*
" Tested a fixed 200 trials (not to a criterion)• p < 05, significantly different from Group V.1 p < 05, significanth different from Groups C and M
ing a series of outline discriminations withincreasing numbers of superimposed stripes(Task 6). Group MS was impaired undereach stimulus condition, but Group Mshowed a substantial deficit only when therewere 10 stripes superimposed on the outlinepatterns. One-way analyses of varianceperformed at each condition indicate sig-nificant differences among the groups whenthere were 0 {H = 5.9, p < .05), 4 (H = 6.5,p < .05), 6 (H = 10, p < .01), and 10 (H = 9.6,p < .01) superimposed stripes. With zeroand four superimposed stripes, Group MSwas significantly impaired relative to both
ACQUISITION RETEST
4 0 0 -
E 300-
P200-
6 10STRIPES
Figure 6 Left. Median errors to criterion on hexagonversus hourglass discrimination as an outline (0 stripes)and with various numbers of superimposed stripesRight: Median percentage errors in 50 trials with eachstimulus pair when pairs were presented in a mixedsequence. (See captions to Figures 2 and 3 for identi-fication of groups )
Group C and Group M (all /7s < 3, all ps <.05), but the number of errors made byGroups M and C did not differ reliably (bothUs > 10). With six superimposed stripeseach experimental group made significantlymore errors than the control group (each U< 3, each p < .05), and Group MS made re-liably more errors than Group M (U = 0, p<.01).
None of the nine cats with lesions reachedcriterion within 1,000 training trials whenthe outline patterns were masked by 10 su-perimposed stripes, although three of thefive cats in Group M and one of the four inGroup MS were making about 75% correctresponses when training was discontinued.In contrast, all five controls reached criterionwell within the limits of training, and therewas no overlap between their error scoresand those of the cats with lesions (each U =0, each p < .01). The difference between thetwo lesion groups on this task was not reli-able.
Figure 6 also shows the performance of thethree groups during the 200 trials of re-training on this task, when the pairs ofstimuli with various numbers of superim-posed stripes were presented in a balancedirregular sequence (Task 15). On this taskthe error rate for the control group was about20% under each stimulus condition, and theerror rate for each lesion group was between30% and 45%. The intergroup differenceswere significant for the conditions of 2,4, and
298 P CORNWELL, W. OVERMAN, AND A. CAMPBELL
6 superimposed stripes (each H > 7.7, eachp < .01) but not for the 10-stripes condition.With two, four, and six superimposed stripesboth Group M and Group MS made signifi-cantly more errors than Group C (each U <2.5, each p < .05), and with six stripes GroupMS made reliably more errors than Group M(U = \,p <.O5).
All subjects successfully discriminated acircle from an I pattern when variations insize were present but irrelevant (Task 7 inTable 2), and there was a reliable effect onthe lesions on the performance (H = 7.6. p< .05). On this task both Group M andGroup MS made significantly more errorsthan Group C (each U < 2, each p < .05).However, Groups M and MS were not sig-nificantly different from each other.
The performance of the three groups atdiscriminating a grid pattern from a square"bulls-eye" (Task 8) and a block L facingright from one facing left (Task 9) is given inTable 2. On neither task were there reliableoverall differences among groups (each H <5.2, each p > .05), but on the task involvingthe orientation of the L, Group MS madesignificantly more errors than either GroupC or Group M (both Us - 2, both ps <.05).
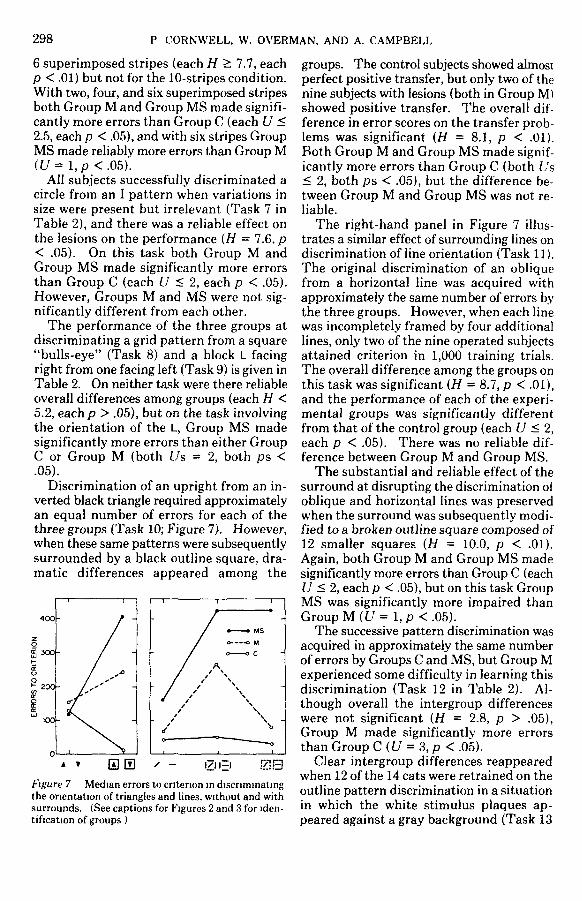
Discrimination of an upright from an in-verted black triangle required approximatelyan equal number of errors for each of thethree groups (Task 10; Figure 7). However,when these same patterns were subsequentlysurrounded by a black outline square, dra-matic differences appeared among the
100-
200-
Figure 7 Median errors to criterion in discriminatingthe orientation of triangles and lines, without and withsurrounds. (See captions for Figures 2 and 3 for iden-tification of groups )
groups. The control subjects showed almostperfect positive transfer, but only two of thenine subjects with lesions (both in Group M)showed positive transfer. The overall dif-ference in error scores on the transfer prob-lems was significant (H = 8.1, p < .01).Both Group M and Group MS made signif-icantly more errors than Group C (both L's< 2, both ps < .05), but the difference be-tween Group M and Group MS was not re-liable.
The right-hand panel in Figure 7 illus-trates a similar effect of surrounding lines ondiscrimination of line orientation (Task 11).The original discrimination of an obliquefrom a horizontal line was acquired withapproximately the same number of errors bythe three groups. However, when each linewas incompletely framed by four additionallines, only two of the nine operated subjectsattained criterion in 1,000 training trials.The overall difference among the groups onthis task was significant (H = 8.7, p < .01),and the performance of each of the experi-mental groups was significantly differentfrom that of the control group (each U < 2,each p < .05). There was no reliable dif-ference between Group M and Group MS.
The substantial and reliable effect of thesurround at disrupting the discrimination ofoblique and horizontal lines was preservedwhen the surround was subsequently modi-fied to a broken outline square composed of12 smaller squares (H = 10.0, p < .01).Again, both Group M and Group MS madesignificantly more errors than Group C (eachU < 2, each p < .05), but on this task GroupMS was significantly more impaired thanGroup M((7 = l ,p <.O5).
The successive pattern discrimination wasacquired in approximately the same numberof errors by Groups C and MS, but Group Mexperienced some difficulty in learning thisdiscrimination (Task 12 in Table 2). Al-though overall the intergroup differenceswere not significant (H = 2.8, p > .05),Group M made significantly more errorsthan Group C (U = 3, p < .05).
Clear intergroup differences reappearedwhen 12 of the 14 cats were retrained on theoutline pattern discrimination in a situationin which the white stimulus plaques ap-peared against a gray background (Task 13
VISUAL CORTEX AND HIDDEN FIGURES 299
REVERSALS
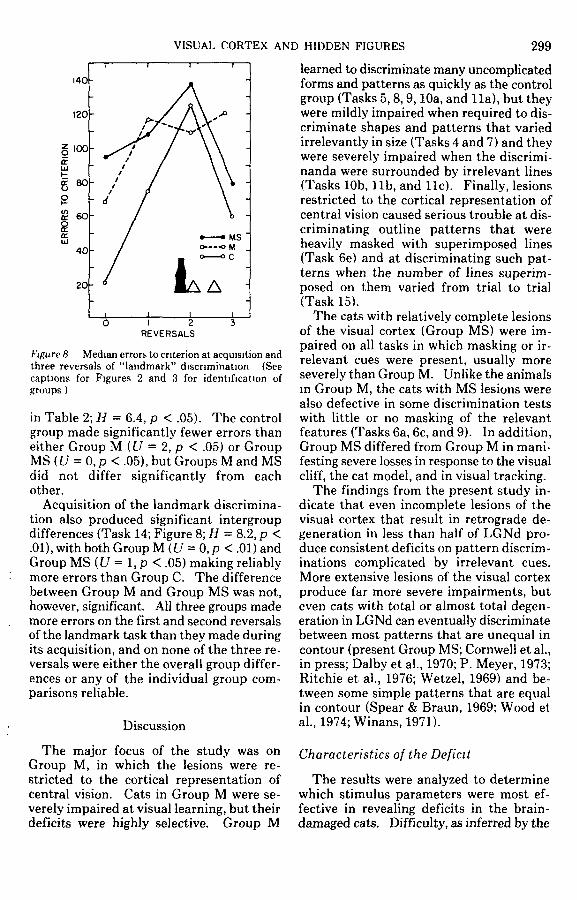
Figure 8 Median errors to criterion at acquisition andthree reversals of "landmark" discrimination (Seecaptions for Figures 2 and 3 for identification ofgroups )
in Table 2; H = 6.4, p < .05). The controlgroup made significantly fewer errors thaneither Group M (U = 2, p < .05) or GroupMS(t/ = 0,p <.05), but Groups M and MSdid not differ significantly from eachother.
Acquisition of the landmark discrimina-tion also produced significant intergroupdifferences (Task 14; Figure 8; H = 8.2, p <.01), with both Group M (U = 0, p < .01) andGroup MS (U = 1, p < .05) making reliablymore errors than Group C. The differencebetween Group M and Group MS was not,however, significant. All three groups mademore errors on the first and second reversalsof the landmark task than they made duringits acquisition, and on none of the three re-versals were either the overall group differ-ences or any of the individual group com-parisons reliable.
Discussion
The major focus of the study was onGroup M, in which the lesions were re-stricted to the cortical representation ofcentral vision. Cats in Group M were se-verely impaired at visual learning, but theirdeficits were highly selective. Group M
learned to discriminate many uncomplicatedforms and patterns as quickly as the controlgroup (Tasks 5,8,9,10a, and lla), but theywere mildly impaired when required to dis-criminate shapes and patterns that variedirrelevantly in size (Tasks 4 and 7) and theywere severely impaired when the discrimi-nanda were surrounded by irrelevant lines(Tasks 10b, l ib, and lie). Finally, lesionsrestricted to the cortical representation ofcentral vision caused serious trouble at dis-criminating outline patterns that wereheavily masked with superimposed lines(Task 6e) and at discriminating such pat-terns when the number of lines superim-posed on them varied from trial to trial(Task 15).
The cats with relatively complete lesionsof the visual cortex (Group MS) were im-paired on all tasks in which masking or ir-relevant cues were present, usually moreseverely than Group M. Unlike the animalsin Group M, the cats with MS lesions werealso defective in some discrimination testswith little or no masking of the relevantfeatures (Tasks 6a, 6c, and 9). In addition,Group MS differed from Group M in mani-festing severe losses in response to the visualcliff, the cat model, and in visual tracking.
The findings from the present study in-dicate that even incomplete lesions of thevisual cortex that result in retrograde de-generation in less than half of LGNd pro-duce consistent deficits on pattern discrim-inations complicated by irrelevant cues.More extensive lesions of the visual cortexproduce far more severe impairments, buteven cats with total or almost total degen-eration in LGNd can eventually discriminatebetween most patterns that are unequal incontour (present Group MS; Cornwell et al.,in press; Dalby et al., 1970; P. Meyer, 1973;Ritchie et al., 1976; Wetzel, 1969) and be-tween some simple patterns that are equalin contour (Spear & Braun, 1969; Wood etal., 1974; Winans, 1971).
Characteristics of the Deficit
The results were analyzed to determinewhich stimulus parameters were most ef-fective in revealing deficits in the brain-damaged cats. Difficulty, as inferred by the

300 P. CORNVVELL, W. OVERMAN, AND A CAMPBELL
rate at which normal cats learned each dis-crimination task, was not a consequentialfactor. Groups M and MS did not differfrom the control group on five of the six tasksthat were most difficult for the intact cats(Tasks 8,10a, 12,14b, 14c, and 14d); none ofthese problems entailed visual masking. Onthe eight tasks involving spatial masking(Tasks 6b, 6c, 6d, 6e, 10b, l ib, lie, and 15),most of the cats in Group MS failed all eight,and Group M was severely impaired at fiveof the eight. These findings corroborateprevious observations that the difficulty atdiscrimination learning by cats with com-plete or almost complete removal of visualcortex can be predicted from the disparity incontour and complexity between the twopatterns (Cornwell et al., in press; Dalby etal., 1970; Ritchie et al., 1976) and show thatcats with lesions confined to the corticalrepresentation of central vision have similarbut milder trouble in the discrimination ofmasked figures.
Cats in Groups M and MS had difficultyin relearning to discriminate outline patternswhen the goal box was gray and the stimulusplaques were white (Task 13). The poorperformance of Group MS probably reflects,at least in part, the initially poor perfor-mance on this same task when the stimuliwere presented in white goal boxes (Task6a). Group M, however, was as proficient atthe initial task as the control group. Thedeficit by Group M at relearning, then,might suggest a retention loss, but it alsomight indicate that the gray background ofthe goal box functioned as a masking stim-ulus, much like the borders used to surroundthe figures in Tasks 10 and 11.
The impairment by the operated cats ofGroup M and Group MS was not limited toproblems involving spatial masking. Thecats were also deficient in learning to dis-criminate objects (Task 4) and patternstimuli (Task 7) in the face of irrelevantvariations in the size of the relevant discri-minanda. Similar results were reportedpreviously from cats with MS lesions(Cornwell et al., 1976), and such deficits maybe similar to those created by large striateablations in tree shrews, in which there is anapparent inability to attend to one dimen-sion when a salient distracting cue is avail-
able (Killackey, Wilson, & Diamond.1972).
Groups M and MS also had difficulties onthe "landmark" problem (Task 14) whichfeatured spatial discontiguity between thecue to the correct response (bottle) and thetwo visually identical stimuli to which theresponse was to be made. Spatial separationof the loci of stimulus and response is a po-tent determinant of discrimination learningfor normal mammals (cf. D. Meyer, Treich-ler, & Meyer, 1965), and there is evidence inmonkeys that this effect is exaggerated b\damage to the cortical visual system (Butter& Hirtzel, 1970). The trouble of cats withlesions of the visual cortex on some taskswith spatial masking (e.g., Tasks 10b, lib,and lie) might be due, at least in part, to thefact that the irrelevant lines were added ata position nearer than the relevant cue to theplace of response. However, it seems pre-mature to attempt an explanation of theimpairments in cats with M or MS lesions ontests with masking lines, irrelevant cues, andspatial discontiguity in terms of a distur-bance in a single underlying process.
Comparison With Primates
The impairment in cats with incompletedestruction of the geniculocortical system isquite similar to that shown by primates withcomparable lesions. Monkeys with lateralstriate lesions are impaired at discriminatingbetween patterns obscured by extraneouslines (Butter, 1969, 1972, 1979), as arebushbabys with lateral striate lesions (At-encio, Diamond, & Ward, 1975). A deficithas also been shown in monkeys by the ad-dition of nonmasking extraneous cues suchas color (Gross, Cowey, & Manning, 1971).The findings for both cats and monkeyssuggest that one specific deficit followingpartial destruction of the geniculocorticalsystem is an inability to extract meaningfulvisual signals from visual noise.
Moreover, it is not easy to attribute thisdeficit in either species to simple sensorylosses, e.g., a large central scotoma plus re-duced visual acuity. Careful measurementsof visual acuity in monkeys with LS lesionsindicate that the elevation in threshold is notenough to account for the deficits in pattern

VISUAL CORTEX AND HIDDEN FIGURES 301
discrimination performance (Weiskrantz &Cowey, 1963). Even the most impairedmonkey of Weiskrantz and Cowey coulddetect lines with a spatial frequency of only1.66 minutes of arc. In the case of Butter's(1972, 1979) monkeys, a combination of acentral scotoma and reduced acuity cannotreadily explain the successful discriminationof pairs of letters, followed by poor perfor-mance when the same letters were sur-rounded by an annulus.
We are somewhat less confident thatchanges in acuity, combined with scotomata,cannot account for the findings in thepresent experiment. However, Smith (1938)reported visual acuity far less than 1° of vi-sual angle in striatectomized cats whenacuity was assessed with optokinetic nys-tagmus. Berkley and Sprague (1979) re-ported comparable grating acuity for catswith area 17 removed that were tested in anoperant discrimination task. In addition,three of our cats were able to discriminatedifferences in the pattern of lines that sub-tended only 1.5° (outline task); yet severedeficits followed when these patterns weretransected by lines of the same width andplaced at least 4° apart. It is hard to un-derstand how visual acuity could be goodenough to allow discrimination of orienta-tion, direction, or curvature of lines yet sopoor that discrimination becomes impossiblewhen equally thin lines transect the patternswith interline spacings that exceed 4° of vi-sual angle. Furthermore, the great impair-ment shown by Groups M and MS on thetask involving triangles surrounded by blackframes (Task 10b) does not lend itself to aneasy explanation based on scotomata andloss in acuity. The emerging picture forboth species, then, is that while subtotal le-sions of the visual cortex no doubt causesome sensory losses, the greater impairmentconcerns a deficit involving the extraction ofvisual signals from noise, an extreme de-pendence on stimulus-response contiguity,or other nonsensory losses.
In the present study, the lesions of thecat's cortical representation of central visionincluded parts of more than one cytoarchi-tectural area. In this respect the lesionswere considerably more extensive than le-sions of the LS cortex in monkeys, which are
confined to area 17. However, a reasonableargument can be made for the functionalequivalence of these two lesions in spite ofthe anatomical and physiological differencesin the organization of the cortical visualsystem in cats and monkeys. First, in catsthe destruction of the cortical representationof central vision in both areas 17 and 18 isnecessary to produce total degeneration inthe corresponding part of LGNd (Garey &Powell, 1967; Niimi & Sprague, 1970). Inmonkeys only the lateral part of area 17needs to be removed to produce such de-generation (Polyak, 1957, Figures 251 and255). Second, the activity of neurons in area18 in cats is not completely dependent uponthe integrity of area 17 (Sherk, 1978); in themonkey it is (Schiller & Malpeli, 1977).These facts suggest that the lesions of GroupM in the present study are functionallyequivalent to LS lesions in monkeys in thateach lesion disrupts the representation ofcentral vision in both the core of the visualcortex and its adjacent belt.
Evidence from a variety of other mammalsalso suggests that it is the functional dis-ruption of the belt cortex which is crucial inpattern vision. In tree shrews (Snyder &Diamond, 1968), rats (Hughes, 1977), andsquirrels (Levey, Harris, & Jane, 1973;Wagor, 1978), as in cats (Doty, 1971; Spragueet al., 1977), pattern vision is relatively un-affected by even complete destruction ofarea 17, but it is greatly impaired when thelesions invade the cortex of the belt. In catsand rats this effect could be due to the in-clusion of more of the total projection fieldof LGNd, but this explanation cannot applyto the effects of destruction of the belt cortexin tree shrews and squirrels, for in both ofthese animals LGNd projects only to area 17(Harting, Diamond, & Hall, 1973; Kaas et al.,1972). It has been suggested on the basis ofbehavioral evidence, however, that in treeshrews (Killackey et al., 1972) and in squir-rels (Wagor, 1978) the tectothalamocorticalprojection is sufficient to allow substantialfunction of the belt cortex, but we know of nostudies that have yet examined this possi-bility electrophysiologically.
Comparison of the substrate necessary forpattern vision in various mammals is com-plicated. In all mammals the visual field is

302 P CORNWELL, W. OVERMAN, AND A CAMPBELL
represented on the cortex many times, butperhaps only in monkeys and a few othermammals does the function of many of therepresentations depend upon the functionalintegrity of area 17. It may be that to abol-ish or seriously disrupt pattern vision, it isnecessary to have a malfunction in one ormore of the representations for central visionin the cortical visual belts. In the primates,but not in most other mammals, this can beaccomplished by lesions confined to area 17(Atencio et al., 1975). This idea is consistentwith the impairments in pattern vision bycats with lesions in area 19 and the belts justlateral to it (Berlucchi, Sprague, Antonini,& Simoni, 1979; Sprague et al., 1977) andalso with the deficits in pattern vision thatresult from foveal prestriate lesions inmonkeys (M. Wilson, 1978).
Reference Note
1 Warren, J M Spatial probability learning by naivecats Unpublished manuscript, 1979
References
Atencio, K W , Diamond, I. T., & Ward, J. P Behav -loral study of the visual cortex ol Galago senegal-ensis Journal of Comparative and PhysiologicalPsychology, 1975,8.9, 1109-1135
Berkley, M A., & Sprague, J M Striate cortex andvisual acuity functions in the cat Journal of Com-parative Neurology, 1979, 187, 679-702
Berlucchi, G , Sprague, J. M , Antonini, A , &. Simoni,A Learning and interhemispheric transfer of visualpattern discriminations following unilateral supra-sylvian lesions in split-chiasm cats ExperimentalBrain Research, 1979, 34, 551-574
Butter, C M. Impairments in selective attention tovisual stimuli in monkeys with inierotemporal andlateral striate lesions Brain Research, 1969, 12,374-383
Butler, C M Detection of masked patterns in mon-kevs with inierotemporal. stnate or dorsolateralfrontal lesions Neurupsvchologia, 1972, 10, 241-243
Butter, C M Contrasting effects of lateral striate andsuperior colliculus lesions on wsual discriminationperformance in rhesus monkeys. Journal of Com-parative and Physiological Psychology, 1979, .93,522-537.
Butter, C. M , & Doehrman, S R Size discriminationand transposition in monkeys with striate and tem-poral lesions Cortex, 1968, 4, 35-46
Butter, C. M , & Hirtzel, M Impairment in samplingvisual stimuli in monkeys with inferotemporal lesionsPhysiology and Behavior, 1970,5,369-370.
Campbell, A , Jr Deficits in visual learning produced
by posterior temporal lesions in cats. Journal ofComparative and Physiological Psycholog\, 197892, 45-57.
Cornwell, P., Overman, W., Levitsky, C , Shipley, J , <£.Lezynski, B Performance on the visual cliff by catswith marginal gyrus lesions Journal ofComparatwtand Physiological Psychology, 1976, .90, 996- 1010
Cornwell, P , Overman, W., & Ross, C Extent of re-covery from neonatal damage to the cortical visualsystem in cats Journal of Comparative and Phys-iological Psychology, 1978, 92, 255-270
Cornwell, P., Overman, W , & Ross, C Lesions of thevisual cortex impair the discrimination of hiddenfigures by cats. Physiology and Behavior, inpress.
Cornwell, P , Warren, J M , & Nonneman, A. Marginaland extramargmal cortical lesions and visual dis-crimination by cats Journal of Comparative andPhysiological Psychology, 1976,90, 989-995
Dalby, D A , Meyer, D R , & Meyer, P M Effects ofoccipital neocortical lesions upon visual discrimina-tions in the cat. Physiology and Behavior, 1970, 5,727-734
Daniel, P M , & Whittendge, D The representationof the visual field of the cerebral cortex in monkeysJournal of Physiology, 1961, 159, 203-221.
Diamond, I T Organization of the visual cortexComparative anatomical and behavioral studiesFederation Proceedings, 1976, 35, 60-67
Diamond, 1 T., & Hall, \V C. Evolution of neocortexScience, 1969, 164, 251-262
Donaldson, I M. L , & Nash, J R G The effect of achronic lesion in cortical area 17 on the visual re-sponses .of units in area 18 of the cat Journal ofPhysiology, 1975,2-/5, 325-332
Doty, R W Survival of pattern vision after removalof striate cortex in the adult cat Journal of Com-parative Neurology, 1971, 143, 341-370
Dreher, B , & Cottee, L Visual receptive-field prop-erties of cells in area 18 of cat's cerebral cortex beforeand after acute lesions in area 17 Journal of Neu-rophvsiology, 1975,38, 735-750
Frommer, G P Subtotal lesions' Implications forcoding and recovery of function In S Finger (Ed),Recovery from brain damage New York' PlenumPress, 1978
Garey, L J The termination of thalamo-cortical fibresin the visual cortex of the catand monkey. Journalof Physiology, 1970,2/0, 15P-17P
Garey, L J., & Powell, T P S The projection of thelateral gemculate nucleus upon the cortex in the catProceedings of the Royal Society, Series- B, 1967,169,107-126.
Gray, P A The cortical lamination pattern of theopossum, Didelphys virgimana Journal of Com-parative Neurology, 1924,37, 221-263
Gross, C. G , Cowey, A., & Manning, F J. Furtheranalysis of visual discrimination deficits followingfoveal prestriate and inferotemporal lesions inmonkeys Journal of Comparative and Physiolog-ical Psychology, 1971, 76, 1-7.
Hall, W. C., & Diamond, I T. Organization and func-tion of the visual cortex in hedgehog. I. Corticalcytoarchitecture and thalamic retrograde degenera-tion Brain, Behavior and Evolution, 1968, /,

VISUAL CORTEX AND HIDDEN FIGURES 303
181-214Hartmg, J K., Diamond, I T., & Hall, W C Antero-
grade degeneration study of the cortical projectionsof the lateral genieulate and pulvinar nuclei in thetree shrew (Tupaia gin) Journal of ComparativeNeurology, 1973, 150, 393-440
Hollander, H , & Vanegas, H. The projection from thelateral geniculate nucleus onto the visual cortex in thecat A quantitative study with horseradish-peroxi-dase Journal of Comparative Neurology, 1977, 17.1,519-536.
Hubel, D H., & Wiesel, T. N Receptive fields andfunctional architecture in two nonstnate visual areas(18and 19)of thecat Journal of Neurophysiology,1965,28, 229-289
Hughes, H. C Anatomical and neuro-behavioral in-vestigations concerning the thalamocortical organi-zation of the rat's visual system Journal of Com-parative Neurology, 1977, 175, 311-336
Humphrey, N Vision in a monkey without stnatecortex A case study Perception, 1974, 3, 241-255
Jones, K G The anatomy of extrageniculatestnatevisual mechanisms In F O Schmitt & F G Worden(Eds ), The neurosciences Third study programCambridge, Mass.- MIT Press, 1974.
Kaas, J. H., Hall, W C, & Diamond, I. T Visual cortexof the grey squirrel (Sciurus carohnensis) Archi-tectonic subdivision and connections from the visualthalamus Journal of Comparative Neurology, 1972,145, 273-306
Kalia, M., & Whitteridge, D The visual areas in thesplemal sulcus of the cat. Journal of Physiology,1973,232,275-283
Keating, E G. Effects ot prestnate and stnate lesionson the monkey's ability to locate and discriminatevisual forms Experimental Neurology, 1975, -47,16-25
Killackey, H , Wilson, M , & Diamond, I T. Furtherstudies of the stnate and extrastriate visual cortex inthe tree shrew. Journal of Comparative and Phy.s-wlogieal Psychology, 1972,8/, 45-63
Klu\er. H Functional significance of the geniculo-striate system Biological Symposia, 1942, 7,253-299
Kuypers, H G J.M , Szwarcbart, M. K., Mishkin, M , &Rosvold, H E Occipitotemporal cortico-corticalconnections in the rhesus monkey. ExperimentalNeurology, 1965,11, 245-262
Levey, N H , Harris, J , & .Jane, J. A Effects of visualcortical ablation on pattern discrimination in theground squirrel (Citellus tridccemhneatus) Ex-perimental Neurology, 1973,39, 270-276
Lund,.I S., Henry, G H , MacQueen, C L ,& Harvey,A R Anatomical organization of the primary visualcortex (area 17) of the cat- A comparison with area17 of the macaque monkey Journal of ComparativeNeurology, 1979, 18 i, 599-618.
Martinez-Millan, L , & Hollander, H Cortico-corticalprojections from stnate cortex of the squirrel monkey(Saimiri sctureus) A radioautographic studyBrain Research, 1975,83, 405-417
Meyer, D. R , Treichier, F. R , & Meyer, P. M Dis-crete-trial testing techniques and stimulus variablesIn A. M Schrier, H.F Harlow,& F. Stollnitz(Eds ),
Behavior o\ Ntmhuman Primate* (Vol 1) NewYork. Academic Press, 1965
Meyer, P. M Recovery from neocortical damage.Introduction In C! M. French (Ed ), Corticalfunctioning in behavior Glenview, HI Scott,Foresman, 1973.
Mishkin, M , & Weiskrantz, 1. Effects of cortical le-sions in monkeys on critical flicker frequencsJournal of Comparative and Ph\*iological Psy-chology. 1959, ,52. 660- 669.
Murph\ . E H , Mize, R. R , t* Schechter, P B Visualdiscrimination following infant and adult ablation ofcortical areas 17, 18, and 19 in the cat ExperimentalNeuroltw, 1975, 49, 386-405
Nnmi, K ,& Sprague, .J M Thalamo-cortical organi-zation of the visual system in the cat Journal ofComparative Neurology, 1970. KiS, 219-250
Nonneman, A J.&Kolb, B K Lesions of hippocam-pus or prefrontal cortex alter species-tvpical behav-iors in the cat Behaviorul Biology, 1974, 12.•11-54
Otsuka, R , <& Hassler, R Uber aufbau und Gleiderungder corticalen Sehsphare be\ der Kat/e Arc hiv furP.sychiatrie und Zeitst hrift fur die gesamte Neuro-logie, 1962,203, 212-234.
Pasik, T., & Pasik, P The visual world ot monkeysdeprived of stnate cortex Effective stimulus pa-rameters and the importance of the accessory opticsystem Vision Research. 1971, Suppl 3,419-435
Polyak. S The vertebrate visual .system ChicagoUniversity of Chicago Press, 1957.
Ritchie, G D , Meyer, P M , & Meyer, 1) R Residualspatial vision of cats with lesions of the visual cortexExperimental Neurology, 1976, 53, 227-253
Rosenquist, A. 0 , Edwards, S B., & Palmer, L A. Anautoradiographic study of the projections of thedorsal lateral geniculale nucleus and the posteriornucleus in the cat Brain Research, 1974, HO,71 93
Sanderson. K .J The projection of the visual field tothe lateral genitulate and medial interlaminar nucleiin thecat Journal ofComparative Neurology, 1971,113, 101-118.
Sanides, F . i. Hoffmann, .) Cvto- and myeloarchi-tectureof the visual cortex of the < at and of the sur-rounding integration cortices. Journal fur Hirn-forshung, 1969, //, 79-104
Schiller, P. H , & Malpeh, J G The effect of strialecortex cooling on area 18 cells in the monkey BrainResearch. 1977, 126. 366-369
Sherk, H Area 18 cell responses in cat during revers-ible inactivation of area 17 Journal of Neuro-physiology, 1978 , II, 2 0 4 - 2 1 5
Smith, K U. Visual discrimination in the cat VIThe relation between pattern vision and visual acuityand the optic projection centers of the nervous sys-tem Journal of Genetic Psychology, 1938, 53,251-272.
Sn>der, M , & Diamond, I T The organization andfunction of the visual cortex in the tree shrew Brain,Behavior and Evolution, 1968. 7,244-288.
Spear, P D , & Braun, J J Pattern discriminationfollowing removal of visual neocortex in the catExperimental Neurology, 1969,2.5, 331-348
Sprague, J M , Levy, J., DiBerardino, A ,& Berlucchi,

304 P CORNWELL, \V OVERMAN. AND A CAMPBELL
G Visual cortical areas mediating form discrimi-nation in the cat Journal of Comparative Neurol-ogy, 1977, 172, 441-448
Tusa, R J., Palmer, L A , & Rosenquist, A C Theretinotopic organization of area 17 (striate cortex) inthe cat Journal of Comparative Neurology, 1978,777,213-236
Wagor, E Pattern vision in the grey squirrel alter vi-sual cortex ablation. Behavioral Biology, 1978.22,1-22
Weiskrantz, I. Contour discrimination in a youngmonkey with striate cortex ablation Neuropsv-chologia, 1963, 1, 145-164
Weiskrant/., L , & Cowey, A Striate cortex lesions andvisual acuity of the rhesus monkey Journal ofComparative and Physiological Psychology, 1961),,56, 225-231.
Wetzel, A. B Visual cortex lesions in the cat A studyof depth and pattern discrimination Journal ofComparative and Physiological Psychology, 1969,68, 580-588
Wetzel, A B , Thompson, V E , Horel, .1 A , & Meyer,P M Some consequences of perinatal lesions of thevisual cortex in the cat Psvihonomic Science, 1965,3,381-382
Wilson, M. Visual system Pulvinar-extrdstnatecortex In R B Masterton (Ed ), Handbook of be-havioral neurobiologx (\'o\ 1). New York Plenum
Press. 1978.Wilson, M E , & Cragg, B. G. Projections from the
lateral geniculate nucleus in the cat and monke>Journal of Anatomy, 1967, 101, 677-692.
Wilson, W A , & Mishkin, M Comparison of the ef-fects of inferotemporal and lateral occipital lesionson visually guided behavior in monkeys Journal »/Comparative and Physiological Psychology, 1959.52, 10-17.
Winans, S. S Visual cues used by normal and visual-decorticate cats to discriminate figures of equal lu-minous flux Journal of Comparative and Physio-logical Psychology. 1971,74, 167-178.
Wood.C C., Spear, P D,&Braun,J ,1 Effects of se-quential lesions of suprasy Ivian gyn and visual cortexon pattern discrimination in the cat Brain He-search, 1974,66, 443-466.
Zeki.S M Functional organization of a visual area inthe posterior bank of the superior temporal sulcus ofthe rhesus monkey. Journal of Physiology, 1974,236, 549-573
Zeki, S M The functional organization of projectionsfrom striate to prestnate visual cortex in the rhesusmonkey Cold Spring Harbor Symposia on Quan-titative Biology, 1975, 40, 591-600
Received April 25, 1979





























