Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Study of Infection with an Iranian Field-Isolated H9N2 Avian Influenza Virus inVaccinated and Unvaccinated Japanese QuailAuthor(s): Seyyed Mahmoud Ebrahimi, Soudeh Ziapour, Majid Tebianian, Mehran Dabaghian, andMashallah MohammadiSource: Avian Diseases, 55(2):195-200. 2011.Published By: American Association of Avian PathologistsDOI: http://dx.doi.org/10.1637/9538-092110-Reg.1URL: http://www.bioone.org/doi/full/10.1637/9538-092110-Reg.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

Study of Infection with an Iranian Field-Isolated H9N2 Avian Influenza Virus inVaccinated and Unvaccinated Japanese Quail
Seyyed Mahmoud Ebrahimi,A Soudeh Ziapour, Majid Tebianian, Mehran Dabaghian, and Mashallah Mohammadi
Department of Biotechnology, Razi Vaccine and Serum Research Institute (RVSRI), P.O. Box 31975/148, Karaj, Tehran, Iran
Received 27 September 2010; Accepted and published ahead of print 4 January 2011
SUMMARY. In the present study, we examined the mortality rate, egg production, and clinical signs of quail experimentallyinfected with a field isolate of A/Chicken/Iran/339/02 (H9N2) avian influenza virus obtained from an infected commercial layerfarm with severe morbidity and mortality. A total of 120 quail at 14 days old were randomly divided into four groups of vaccinated(B and C) and unvaccinated (A and D) birds. Vaccination was done on days 20 and 32, and viral inoculation of birds in groups Cand D was then carried out on day 43. For evaluation of viral transmission, at 24 hr postinoculation additional unvaccinated birdswere placed in direct contact with challenged birds. All the birds were evaluated for clinical signs, egg production, antibodyproduction, viral titration in lung homogenates, and viral transmission following inoculation. All unvaccinated-challenged birdswere infected and showed clinical signs, whereas the infection rate along with clinical signs of vaccinated-challenged birds reached30%–40%. Although vaccination induced high antibody titers, reduction in food and water consumption was evident in thisvaccinated-challenged group compared with the unchallenged control group. These results could indicate that inactivated vaccinedid not fully prevent the infection, although it was capable of protecting birds against clinical signs and significantly decreased viraltiters in lungs after intranasal challenge.
RESUMEN. Estudio de la infeccion de un aislamiento de campo iranı del virus de la influenza aviar subtipo H9N2 encodornices japonesas vacunadas y no vacunadas.
En el presente estudio, se estudio la tasa de mortalidad, la produccion de huevo, y los signos clınicos de codornices infectadasexperimentalmente con un aislamiento de campo del virus de la influenza aviar A/Pollo/Iran/339/02 (H9N2), obtenido de unagranja de ponedoras comerciales infectadas con morbilidad y mortalidad severas. Un total de 120 codornices a los 14 dıas de edadfueron asignadas aleatoriamente en cuatro grupos, dos de aves vacunadas (B y C) y dos de aves no vacunadas (A y D). La vacunacionse llevo a cabo en los dıas 20 y 32 de edad, y la inoculacion viral de las aves de los grupos C y D, se llevo a cabo en el dıa 43. Para laevaluacion de la transmision viral, a las 24 horas despues de la vacunacion, se colocaron aves no vacunadas adicionales en contactodirecto con las aves desafiadas. Todas las aves fueron evaluadas por los signos clınicos, la produccion de huevo, produccion deanticuerpos, la titulacion viral en homogeneizados de pulmon, y la transmision viral despues de la inoculacion. Todas las aves novacunadas y desafiadas se infectaron y mostraron signos clınicos, mientras que la tasa de infeccion junto con signos clınicos en lasaves vacunadas y desafiadas tuvo un rango de 30% a un 40%. Aunque la vacunacion indujo tıtulos altos de anticuerpos, lareduccion en el consumo de alimento y agua fue evidente en este grupo de aves vacunadas y desafiadas en comparacion con el grupocontrol no desafiado. Estos resultados podrıan indicar que la vacuna inactivada no impide completamente la infeccion, aunque fuecapaz de proteger las aves contra los signos clınicos y disminuyo significativamente el tıtulo viral en el pulmon despues del desafıointranasal.
Key words: H9N2, avian influenza, vaccination, egg production, quail
Abbreviations: AIV 5 avian influenza virus; EID50 5 median embryo infectious dose; GMT 5 geometric mean titer;H 5 hemagglutinin; HA 5 hemagglutination; HI 5 hemagglutination inhibition; N 5 neuraminidase; PBS 5 phosphate-bufferedsaline; pi 5 postinoculation; RBCs 5 red blood cells; SPF 5 specific pathogen free
Influenza viruses are single-stranded, negative-sense RNA virusesof the family Orthomyxoviridae. There are three types of influenza,A, B, and C, which differ in their pathogenicity and genomeorganization (28,29). Although the natural reservoir for all influenzaA viruses is aquatic birds, these viruses also infect and cause diseaseof varying severity in domestic poultry and several species ofmammals, including humans (5,7,8,28).
Influenza A viruses of the H9N2 subtype have become highlyprevalent in poultry in many countries, and although these virusesgenerally cause only mild to moderate disease, they have beenassociated with severe morbidity and mortality in poultry as a resultof co-infection with other pathogens (2,17).
Low pathogenicity avian influenza H9N2 infections have beenreported in the Middle East since 1998 and have also causedwidespread outbreaks in commercial chickens in Iran (16,17) andPakistan (1,12,13), often associated with serious disease problems.
In 1999, avian influenza H9N2 subtype, which is widespread inpoultry in Asia, was transmitted to humans and caused mildrespiratory disease (8,19). This virus isolate was similar to an H9N2virus isolated from a quail in Hong Kong in late 1997. Thetransmission of avian H5N1, H7N7, and H9N2 influenza virusesfrom some species of poultry, such as chickens and quail, to humansindicates that avian influenza viruses can directly infect humanswithout an intermediate host (7,8,10,23).
These direct transmissions of avian-origin influenza viruses tohumans indicate that these viruses have recognized SAa2-6 receptors;as a result, they may become pandemic and endanger public health(8,19).
These phenomena indicate that chickens and quail may play animportant role in the evolution of influenza viruses by acting asintermediate hosts, in which avian influenza viruses (AIVs) can beamplified and transmitted to other animal species (8,10).
In one study, 4-wk-old Japanese quail inoculated with AIVsrepresentative of subtypes H1 to H15, originally isolated from wildACorresponding author. E-mail: [email protected]
AVIAN DISEASES 55:195–200, 2011
195

waterfowl and shorebirds, were not able to transmit to contactcontrol quail placed in the same cage with inoculated quail (10).
The first reported outbreak in Italy showed quail to be infectedwith influenza A H9N2 virus, which caused respiratory disease andwas only lethal to young quail (14).
Moreover, it was reported that quail infected with an H5N3 virusthat was highly pathogenic to turkeys were resistant to disease;H5N3 caused mild clinical signs in quail, but the quail couldtransmit the lethal virus to chickens (26). These studies help not onlyto document both the variability in host adaptation and transmis-sibility among AIVs, but also to point to the significance of quail,which act as mixing vessels or disseminators of influenza viruses toother species. Considering the ever-increasing population size of thefarmed quail during the last two decades, and that the influenza viralreplication in quail is shown to be more rapid than other species,specific attention should be paid to the quail, which may have a rolein the spread of the disease (10,11,30).
Although quail, like many avian species, are said to be sensitive toavian influenza viruses and might act as mixing vessels ordisseminators of avian/mammalian reassortant influenza A viruses(10,14,18,21,26,27), related studies on the susceptibility of quail toinfluenza A viruses are indeed limited. So, we sought to examine themortality rate, egg production, and clinical signs of Japanese quailexperimentally infected with the field isolate of A/Chicken/Iran/339/02 (H9N2) avian influenza virus, which was obtained from aninfluenza-infected commercial layer farm with severe morbidity andmortality. The present study aims to support the hypothesis thatquail could play an important role in the evolution of influenzaviruses by acting as intermediate hosts in which avian influenzaviruses could be amplified without producing clinical signs ormortality.
MATERIALS AND METHODS
Specimen collection. Lungs and trachea were collected from acommercial layer farm at 53 wk of age, showing severe influenza clinicalsigns including respiratory distress, swelling of the infraorbital sinuses,and conjunctivitis accompanied with 100% morbidity and highmortality. There was also a 60% to 80% decrease in egg production,along with misshapen, whitish, and fragile eggs. The collected sampleswere stored at 270 C until use.
Virus isolation and identification. The isolation and identificationmethods used were based on classical laboratory methodology (25).Briefly, samples of lungs and trachea were pooled, suspended inphosphate-buffered saline (PBS) containing antibiotic solution, homog-
enized, clarified, and then inoculated into the allantoic cavity of 10-day-old specific-pathogen-free (SPF) embryonated chicken eggs. Allantoicfluid from these eggs was collected 2 days later and tested forhemagglutinating 1% v/v suspension of chicken red blood cells. Thehemagglutination inhibition (HI) tests for virus identification wereperformed using standard procedures (25). The H and N subtypes weredetermined using polyclonal chicken antisera as described previously(25). The titer of the propagated virus was then calculated as a medianembryo infectious dose (EID50) per milliliter by the Reed and Muenchmethod (22).
Identification of this field isolate was completed by using molecularcharacterization and intravenous index tests (with intravenous pathoge-nicity index , 1.2). Accordingly, this field-isolated virus was confirmedas the same low pathogenicity avian influenza (H9N2) that wascirculating in Iran. This virus isolate was named A/Chicken/Iran/339/02(H9N2) and then used as a challenge inoculum in the current study.
Birds and housing. A total of 120 two-week-old Japanese quails(Coturnix coturnix japonica) were obtained from commercial quailindustry in Yazd province of Iran and were fed ad libitum by standardgrower diet throughout our experiment according to the NationalResearch Council requirements (15). All birds were wing banded forindividual identification and were kept 2 wk for acclimatization beforevaccination. Birds were reared, before and after vaccination, in sixdifferent cages in two separate rooms (1 and 2), such that quail eggscould be collected, with similar environmental conditions, sameavailability of feeder and drinker conditions, with separate ventilationsystems, and offered feed and drinking water ad libitum at the animalresearch unit of Razi Vaccine and Research Institute. Before theexperiment, rooms and cages were vigorously washed and fumigatedusing formaldehyde.
Experimental design. The present study was focused on the level ofegg production, morbidity, and mortality rate in quail experimentallyinfected with an Iranian field-isolated avian influenza virus (H9N2).The aim was to design groups so that the male birds could be consideredas the sacrificed representatives for antibody reaction and female birds asthe representatives for egg production in each group.
For this purpose, a total of 120 quail at the age of 14 days wererandomly divided into four 25-bird groups (A–D), including 10 malesand 15 females with approximately the same weight in each group(Fig. 1). In order to examine the rate of viral transmission in birds ofvaccinated (C) and unvaccinated (D) groups after viral inoculation, two10-bird groups (Cc and Dd), including only unvaccinated leg-bandedmales in each group, were also considered as the corresponding groupsof contact birds, so that the birds of group Cc were in contact withvaccinated and challenged birds of group C and birds of group Dd werein contact with unvaccinated and challenged birds of group D as shownin Fig. 1.
Serum samples were collected from five birds of each group, bloodsamples (0.5 ml) were taken from wing veins, prior to first vaccination
Fig. 1. Experimental design used in inoculation study in quail. Groups A, B, C, and D indicate main groups of birds (n 5 25; 15 female and 10male birds in each group). Group Dd indicates a corresponding group of contact birds (n 5 10 only male birds) for birds of group D. Group Ccindicates a corresponding group of contact birds (n 5 10 only male birds) for birds of group C.
196 S. M. Ebrahimi et al.
(at 19 days of age) to ensure that the birds were serologically negative forAIVs. At 20 and 32 days of age, birds in group B in room 1 and group Cin room 2 were vaccinated subcutaneously in the neck with 0.5 ml ofkilled virus, A/Chicken/Iran/ZMT-101/98 (H9N2), in oil emulsionvaccine (Razi Institute, Tehran, Iran) against A/Chicken/Iran/339/02(H9N2). Birds of group A in room 1, group D in room 2, and contactbirds Cc and Dd were also inoculated subcutaneously with oil emulsionvaccine using negative allantoic fluid as unvaccinated groups. At 43 daysof age, quail in groups C and D in room 2 were challenged with 106.8
50% embryo infectious dose/bird of A/Chicken/Iran/339/02 (H9N2)virus intranasally. Twenty-four hours after viral challenge, unvaccinatedcontact birds of groups Cc and Dd (Fig. 1) were placed in the same cagewith inoculated birds of groups C and D, respectively. Feed and waterconsumption was not measured quantitatively, but it was checkedweekly. Birds were then weighed weekly and observed daily for clinicalsigns of disease. Blood samples from sacrificed birds were collected forserologic assay and their lungs for viral titer examination on 49 and56 days of age. Three male birds from unchallenged groups A and B andsix male birds containing three contact birds from challenged groups Dand C were selected as serologic assay representatives, and their lungswere stored at 270 C for virus titration. Fifteen female birds in eachgroup were also observed by monitoring their clinical signs and eggproduction to the end of our project. All surviving birds from bothchallenged and unchallenged groups on day 65 of age, 23 days after viralchallenge, were humanely sacrificed and subjected for serologicexamination, and their lungs were preserved at 270 C for later virustitration.
Serologic analysis. The hemagglutination (HA) and HI tests wereperformed as described previously (25). Briefly, samples of lungs takenfrom infected birds on days 6, 13, and 23 after viral challenge weresuspended in PBS containing antibiotic solution, homogenized,clarified, and then inoculated once into the allantoic cavity of 10-day-old SPF embryonated chicken eggs. Allantoic fluid from these eggs wascollected 2 days later and tested for hemagglutinating 1% v/v suspensionof chicken red blood cells (RBCs). The HA titer was the highest dilutionof the antigen that caused the agglutination of the RBCs. The HI titerwas done on the antisera taken, and it was the highest dilution of theantiserum that caused complete inhibition of agglutination by 4 HAU of
H9N2 antigen (Razi Institute, Tehran, Iran). All negative samples forthe HI test were set at 2 log2, and all negative samples for the virustitration test were set at 0.9 log10 EID50/ml.
Statistical analysis. Data were analyzed using SPSS (version 11)software. Antibody titers and the numbers of egg production wereanalyzed for significance (P , 0.05) by using the one-way analyses ofvariance when variance between groups was homogeneous anddistribution of the data was normal or a nonparametric test (Kruskal-Wallis) when normality test or homogeneity of variance test failed.
RESULTS
Clinical signs and mortality. Following the inoculation of quailwith A/Chicken/Iran/339/02 (H9N2) virus isolate, no mortality wasobserved in challenged birds (groups C and D). Morbidity rate basedon AIV clinical signs following inoculation reached 30% to 40% ingroup C and 100% in group D. Moreover, unvaccinated contactbirds of groups Cc and Dd showed the same morbidity rate aftermixing with birds of groups C and D, respectively. Accordingly, incomparison with the control birds of groups A and B, there weresignificant decreases in egg production (Fig. 2) and subjectivereductions in food and water consumption of groups D and C,though the amount of this reduction was not quantified. Clinicalsigns such as depression, ruffled feathers, diarrhea, stopped singing,and nasal/eye discharges were observed from 2 to 6 dayspostinoculation (pi) in group D, and birds were quiet and reluctantto move. The clinical signs were mild in birds of group C (data notshown). As expected, birds of groups A and B were in good healthwithout any clinical signs and morbidity. By day 11 pi birds showedsome recovery, and on day 15 pi they were behaving normally andstarted to sing again. Likewise, their activity and appetite returned tonormal levels.
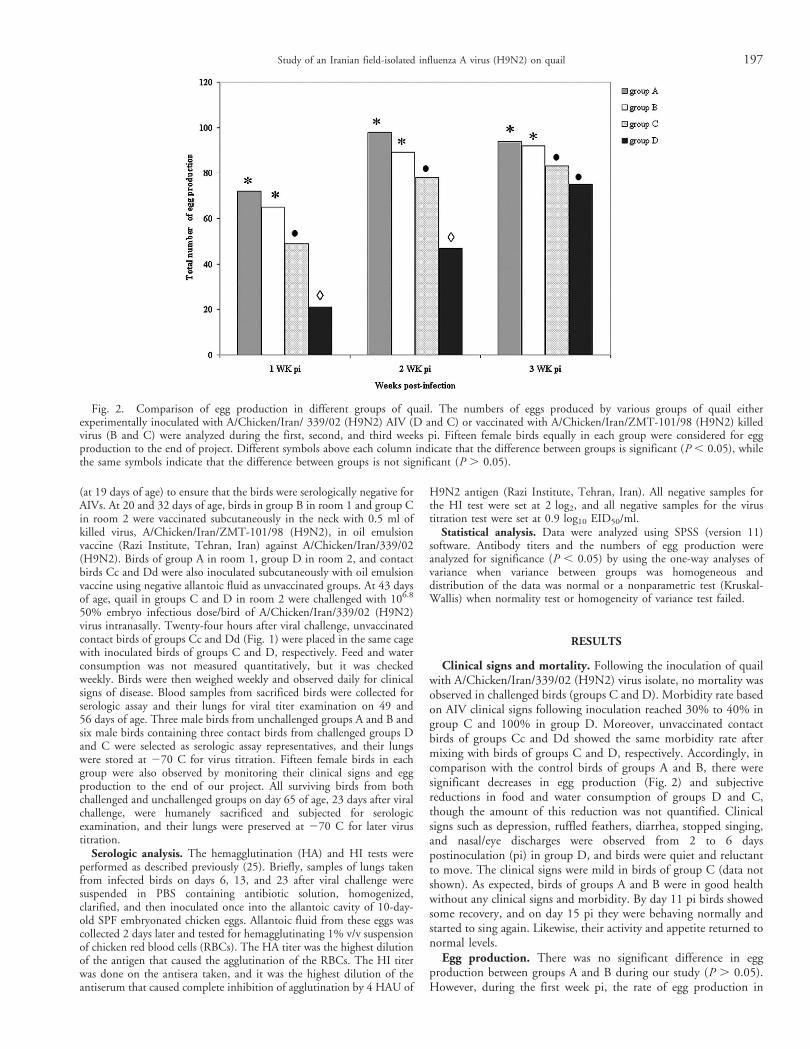
Egg production. There was no significant difference in eggproduction between groups A and B during our study (P . 0.05).However, during the first week pi, the rate of egg production in
Fig. 2. Comparison of egg production in different groups of quail. The numbers of eggs produced by various groups of quail eitherexperimentally inoculated with A/Chicken/Iran/ 339/02 (H9N2) AIV (D and C) or vaccinated with A/Chicken/Iran/ZMT-101/98 (H9N2) killedvirus (B and C) were analyzed during the first, second, and third weeks pi. Fifteen female birds equally in each group were considered for eggproduction to the end of project. Different symbols above each column indicate that the difference between groups is significant (P , 0.05), whilethe same symbols indicate that the difference between groups is not significant (P . 0.05).
Study of an Iranian field-isolated influenza A virus (H9N2) on quail 197
group D in comparison with those of control groups A and B wassignificantly reduced to 29% and 32%, respectively (P , 0.05;Fig. 2). During the second and third weeks pi, egg production ingroup D was moderately recovered, though it still was significantlyless than those of other groups (A–B; P , 0.05; Fig. 2). During thesecond week pi, egg production in group D was also significantly lessthan that of group C (P , 0.05); however, at the end of third weekpi, the rate of egg production in group D was less than that of groupC, but the difference between them was not significant(P . 0.05;Fig. 2). During the study, group C had a continuously significantlyhigher record (32%) of egg production compared with group D (P, 0.05) and lower production record (20.5% and 14.5%) comparedwith groups A and B, respectively (P , 0.05; Fig. 2).
Serologic infection and transmission evaluation. All birds wereseronegative to influenza A viruses prior to vaccination on day 20 ofage. Birds in the control group (A), which was placed in an isolateroom, were active, were eating and drinking normally, did not showclinical signs, and were seronegative throughout the study.
The serologic responses in the different groups of quail were eitherdue to their vaccination at 20 and 32 days of age (groups B and C) or
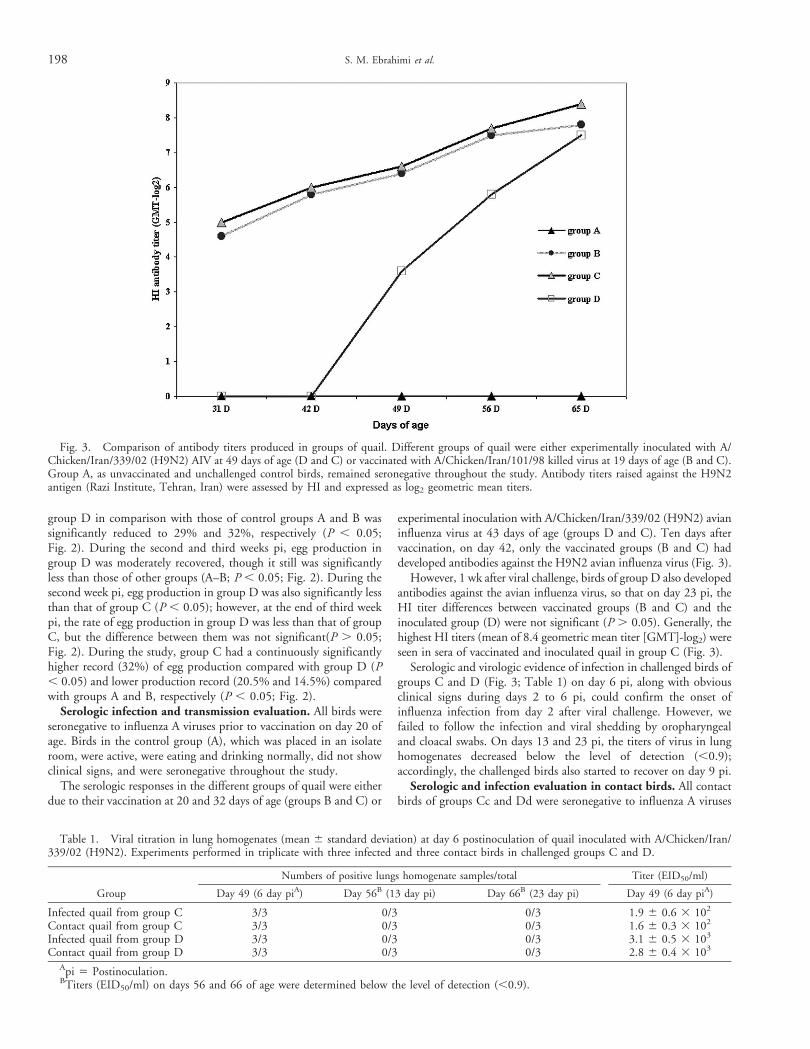
experimental inoculation with A/Chicken/Iran/339/02 (H9N2) avianinfluenza virus at 43 days of age (groups D and C). Ten days aftervaccination, on day 42, only the vaccinated groups (B and C) haddeveloped antibodies against the H9N2 avian influenza virus (Fig. 3).
However, 1 wk after viral challenge, birds of group D also developedantibodies against the avian influenza virus, so that on day 23 pi, theHI titer differences between vaccinated groups (B and C) and theinoculated group (D) were not significant (P . 0.05). Generally, thehighest HI titers (mean of 8.4 geometric mean titer [GMT]-log2) wereseen in sera of vaccinated and inoculated quail in group C (Fig. 3).
Serologic and virologic evidence of infection in challenged birds ofgroups C and D (Fig. 3; Table 1) on day 6 pi, along with obviousclinical signs during days 2 to 6 pi, could confirm the onset ofinfluenza infection from day 2 after viral challenge. However, wefailed to follow the infection and viral shedding by oropharyngealand cloacal swabs. On days 13 and 23 pi, the titers of virus in lunghomogenates decreased below the level of detection (,0.9);accordingly, the challenged birds also started to recover on day 9 pi.
Serologic and infection evaluation in contact birds. All contactbirds of groups Cc and Dd were seronegative to influenza A viruses
Table 1. Viral titration in lung homogenates (mean 6 standard deviation) at day 6 postinoculation of quail inoculated with A/Chicken/Iran/339/02 (H9N2). Experiments performed in triplicate with three infected and three contact birds in challenged groups C and D.
Group
Numbers of positive lungs homogenate samples/total Titer (EID50/ml)
Day 49 (6 day piA) Day 56B (13 day pi) Day 66B (23 day pi) Day 49 (6 day piA)
Infected quail from group C 3/3 0/3 0/3 1.9 6 0.6 3 102
Contact quail from group C 3/3 0/3 0/3 1.6 6 0.3 3 102
Infected quail from group D 3/3 0/3 0/3 3.1 6 0.5 3 103
Contact quail from group D 3/3 0/3 0/3 2.8 6 0.4 3 103
Api 5 Postinoculation.BTiters (EID50/ml) on days 56 and 66 of age were determined below the level of detection (,0.9).
Fig. 3. Comparison of antibody titers produced in groups of quail. Different groups of quail were either experimentally inoculated with A/Chicken/Iran/339/02 (H9N2) AIV at 49 days of age (D and C) or vaccinated with A/Chicken/Iran/101/98 killed virus at 19 days of age (B and C).Group A, as unvaccinated and unchallenged control birds, remained seronegative throughout the study. Antibody titers raised against the H9N2antigen (Razi Institute, Tehran, Iran) were assessed by HI and expressed as log2 geometric mean titers.
198 S. M. Ebrahimi et al.

prior to contact with birds of group C and D, respectively. Thecontact birds of groups Cc and Dd also developed antibodies againstinfluenza virus on days 56 (13 days pi, with means of 3.1 and 4.2GMT-log2, respectively) and 66 (23 days pi, with means of 4.3 and5.4 GMT-log2, respectively), so that the HI titer differences betweencontact birds of groups Cc and Dd were shown to be significant ondays 56 and 66 (P , 0.05). The serologic responses in the contactgroups were only due to influenza A virus, which was transmittedfrom challenged birds of groups C and D to unvaccinated birds ofcontact groups Cc and Dd placed in the same cage with inoculatedbirds. These results of serologic responses along with virologicevidence (Table 1) of infection in contact birds of groups Cc and Ddon day 6 pi can confirm transmission of influenza virus frominoculated quail to unvaccinated contact birds. Hence, our resultsshowed that the HI titer and viral load differences between these twounvaccinated contact groups were significant (P , 0.05), and thelevel of viral transmission from unvaccinated-challenged birds ofgroup D to related contact birds was shown to be more thanvaccinated-challenged birds of group C.
DISCUSSION
Influenza virus serotype H9N2 is classified as low pathogenicityvirus; however, this serotype has already caused high levels ofmortality in an outbreak in Pakistan during 1998 (12,13) and alsohas been reported as the causative agent of severe outbreaks in someregions of Iran (16,17). Considering this background, our study wascarried out to assay the susceptibility of Japanese quail (Coturnixcoturnix japonica) with the field isolate of AIV, A/Chicken/Iran/339/02 (H9N2) isolate.
In the present study, there was a significant difference in eggproduction between groups A and B with C and between groups Aand B with D at the first, second, and third weeks postinfection(Fig. 2). Moreover, the difference between groups C and D was alsosignificant at the first and second weeks pi. At the third week pi, therate of egg production in group D was less than that of group C, butthe difference between them was not significant (Fig. 2). This resultindicated that in spite of good potency of the applied vaccine towardthe induction of excellent protection against the morbidity andclinical signs in group C; the egg production was significantlyaffected in this experimentally inoculated group. In other words,consistent with previous findings (3,6,18,24), this result pointed tothe replication level of the challenge virus in the vaccinated birds andconfirmed that inactivated vaccine did not fully prevent theinfection, though it was capable of protecting against clinical signsand significantly decreased virus titers in lungs after intranasalchallenge. Moreover, this isolate of influenza virus was previouslyevaluated in unvaccinated quail (4) and showed no more mortalityin terms of degree of clinical signs than what we present in this work.
There were steady increases in antibody response in treatmentgroups (B, C, and D) during the first through third weeks pi(Fig. 3). Similar results regarding antibody response has beenobtained using low pathogenic H7N2 AIV virus (9).
Moreover, group C, which received both the vaccine and viruschallenge, had a slightly higher antibody response than groups B andD. This finding was also in accordance with a previous study inbroiler chickens challenged and vaccinated with H9N2 AIV (31).
Generally, similar results regarding antibody responses, clinicalsigns, and the effect of AIV on egg production in quail have beenobtained with H9N2 AIV by Nili et al. (18), in which clinical signswere observed from 6 days pi in challenged birds.
Morbidity rate of 100% along with clinical signs in unvaccinatedcontact birds (group Dd) exposed by contact with unvaccinated-challenged birds (group D) and morbidity rate of 30% to 40% ofunvaccinated contact birds (group Cc) exposed by contact withvaccinated-challenged birds (group C), indicated a lower potential oftransmission from the vaccinated birds than the unvaccinated onesowing to the reduction in virus shedding.
Results of this study could support the previous study in whichAsian quail H9N2 AIV could transmit from inoculated tosusceptible quail through either direct or indirect contact during 5to 7 days pi without any mortality (20).
The current study has demonstrated that quail infected with A/chicken/Iran/339/02 (H9N2), which caused severe health andeconomic problem in 53-wk-old commercial layers, showed onlymild disease signs and high egg production drops, with no rigorousmorbidity or mortality. This finding was generally in accordancewith the previous report, in which H5N9 virus, which is highlypathogenic to turkeys, caused mild clinical signs in quail (14).
Similarly, quail in Hong Kong have also shown a high incidenceof infection with influenza A, particularly H9N2 viruses, withoutany mortality and disease signs (26).
Although the ability of influenza viruses to replicate in quail andinfect them differs among influenza isolates, our field-isolated A/chicken/Iran/339/02 (H9N2) virus, which was obtained from a layerfarm with severe influenza clinical signs and high mortality, was ableto infect quail with clinical signs, transmit virus to contact birds, andaffect egg production significantly. This study could highlight thenecessity for more comprehensive surveillance and further evaluationof H9N2 viruses in quail, and the role of this reservoir bird species inthe ecology of influenza A virus needs to be better understood.
REFERENCES
1. Bano, S., K. Naeem, and S. A. Malik. Evaluation of pathogenicpotential of avian influenza virus serotype H9N2 in chickens. In: Proc. FifthInternational Symposium on Avian Influenza, Athens, GA. Avian Dis.47:817–822. 2003.
2. Brown, I. H., J. Banks, R. J. Manvell, S. C. Essen, W. Shell, M.Slomka, B. Londt, and D. J. Alexander. Recent epidemiology and ecology ofinfluenza A viruses in avian species in Europe and the Middle East. Dev.Biol. 124:45–50. 2006.
3. Capua, I., and S. Marangon. The use of vaccination as an option forthe control of avian influenza. Avian Pathol. 32:335–343. 2003.
4. Ebrahimi, S. M., H. Nili, and N. Sohrabi. Histopathologicalevaluation of A/Chicken/iran/339/02 (H9N2), an Iranian field isolate ofinfluenza virus, on Japanese quail (Coturnix coturnix japonica). World Appl.Sci. J. 9:226–229. 2010.
5. Fouchier, R. A., B. Olsen, T. M. Bestebroer, S. Herfst, G. F.Rimmelzwaan, and A. D. Osterhaus. Influenza A virus surveillance in wildbirds in Northern Europe in 1999 and 2000. Avian Dis. 47:857–860. 2003.
6. Halvorson, D. A. The control of H5 or H7 mildly pathogenic avianinfluenza: a role for inactivated vaccine. Avian Pathol. 31:5–12. 2002.
7. Koopmans, M., B. Wilbrink, M. Conyn, G. Natrop, H. van der Nat,H. Vennema, A. Meijer, J. van Steenbergen, R. Fouchier, A. Osterhaus, andA. Bosman. Transmission of H7N7 avian influenza A virus to human beingsduring a large outbreak in commercial poultry farms in the Netherlands.Lancet 363(9409):587–593. 2004.
8. Lin, Y. P., M. Shaw, V. Gregory, K. Cameron, W. Lim, A. Klimov,K. Subbarao, Y. Guan, S. Krauss, K. Shortridge, R. Webster, N. Cox, and A.Hay. Avian-to-human transmission of H9N2 subtype influenza A viruses:relationship between H9N2 and H5N1 human isolates. Proc. Natl. Acad.Sci. U. S. A. 97:9654–9668. 2000.
9. Lu, H., and A. E. Castro. Evaluation of the infectivity, length ofinfection, and immune response of a low-pathogenicity H7N2 avian
Study of an Iranian field-isolated influenza A virus (H9N2) on quail 199

influenza virus in specific-pathogen-free chickens. Avian Dis. 48:263–270.2004.
10. Makarova, N. V., H. Ozaki, H. Kida, R. G. Webster, and D. R.Perez. Replication and transmission of influenza viruses in Japanese quail.Virology 310:8–15. 2003.
11. Marangon, S., L. Bortolotti, I. Capua, M. Bettio, and M. DallaPozza. Low-pathogenicity avian influenza (LPAI) in Italy (2000–01):epidemiology and control. Avian Dis. 47:1006–1009. 2002.
12. Naeem, K., M. Naurin, S. Rashid, and S. Bano. Seroprevalence ofavian influenza virus and its relationship with increased mortality anddecreased egg production. Avian Pathol. 32:283–287. 2003.
13. Naeem, K., A. Ullah, R. J. Manvell, and D. J. Alexander. Avianinfluenza A subtype H9N2 in poultry in Pakistan. Vet. Rec. 145:560. 1999.
14. Nardelli, L., A. Rinaldi, H. G. Pereira, and G. Mandelli. Influenzavirus infection in Japanese quail. Arch. Exp. Vet. Med. 24:231–249. 1970.
15. National Research Council. Nutrients requirements of poultry, 8threv. ed. National Academy Press, Washington, DC. pp. 11–15. 1984.
16. Nili, H., and K. Asasi. Natural cases and an experimental study ofH9N2 avian influenza in commercial broiler chickens of Iran. Avian Pathol.31:247–252. 2002.
17. Nili, H., and K. Asasi. Avian influenza (H9N2) outbreak in Iran. In:Proc. Fifth International Symposium on Avian Influenza, Athens, GA. AvianDis. 47:828–831. 2003.
18. Nili, H., K. Asasi, H. Dadras, and M. Ebrahimi. Pathobiology ofH9N2 avian influenza virus in Japanese quail (Coturnix coturnix japonica).In: Proc. Sixth International Symposium on Avian Influenza, Cambridge,UK. Avian Dis. 51:390–392. 2007.
19. Peiris, M., K. Y. Yuen, C. W. Leung, K. H. Chan, P. L. Ip, R. W.Lai, W. K. Orr, and K. F. Shortridge. Human infection with influenzaH9N2. Lancet 354:916–917. 1999.
20. Perez, D. R., W. Lim, J. P. Seiler, G. Yi, M. Peiris, K. F. Shortridge,and R. G. Webster. Role of quail in the interspecies transmission of H9influenza A viruses: molecular changes on HA that correspond to adaptationfrom ducks to chickens. J. Virol. 77:3148–3156. 2003.
21. Perez, D. R., R. J. Webby, E. Hoffmann, and R. G. Webster. Land-based birds as potential disseminators of avian mammalian reassortantinfluenza A viruses. Avian Dis. 47:1114–1117. 2003.
22. Reed, L. J., and H. A. Muench. Simple method for estimating fiftyper cent endpoints. Am. J. Hyg. 27:493–497. 1938.
23. Suzuki, Y., T. Ito, T. Suzuki, R. E. Holland, T. M. Chambers, M.Kiso, H. Ishida, and Y. Kawaoka. Sialic acid species as a determinant of thehost range of influenza A viruses. J. Virol. 74:11825–11831. 2000.
24. Swayne, D. E., M. L. Perdue, J. R. Beck, M. Garcia, and D. L.Suarez. Vaccine protects chickens against H5 highly pathogenic avianinfluenza in the face of genetic changes in field viruses over multiple years.Vet. Microbiol. 74:165–172. 2000.
25. Swayne, D. E., D. A. Senne, and C. W. Beard. Influenza. In: Alaboratory manual for the isolation and identification of avian pathogens. D.E. Swayne, J. R. Glisson, M. W. Jackwood, J. E. Pearson, and W. M. Reeds,eds. American Association of Avian Pathologists, Kennett Square, PA.pp. 150–155. 1998.
26. Tashiro, M., M. Reinacher, and R. Rott. Aggravation of pathoge-nicity of an avian influenza virus by adaptation to quails. Arch. Virol.93:81–95. 1987.
27. Wan, H., and D. R. Perez. Quail carry sialic acid receptorscompatible with binding of avian and human influenza viruses. Virology346:278–286. 2006.
28. Webster, R. G., S. Krauss, D. Hulse-Post, and K. Sturm-Ramirez.Evolution of influenza viruses in wild birds. J. Wildl. Dis. 43:1–6. 2007.
29. Wright, P. F., and R. G. Webster. Orthomyxoviruses. In: Fieldsvirology, 4th ed. D. M. Knipe, and P. M. Howley, eds. Lippincott Williams,Wilkins, Philadelphia, PA. pp. 1533–1579. 2001.
30. Xu, K. M., K. S. Li, G. J. D. Smith, J. W. Li, H. Tai, J. X. Zhang, R.G. Webster, J. S. M. Peiris, H. Chen, and Y. Guan. Evolution and molecularepidemiology of H9N2 influenza A viruses from quail in southern China,2000–2005. J. Virol. 81:2635–2645. 2006.
31. Ziegler, A. F., S. Davision, H. Acland, and R. J. Eckroade.Characteristics of H7N2 (nonpathogenic) avian influenza virus infectionin commercial Pennsylvania layers, 1997–1998. Avian Dis. 43:142–149.1999.
ACKNOWLEDGMENTS
We acknowledge assistance provided by Mr. Montazeri and Mr.Bagheri from the Department of Biotechnology, Razi Vaccine andSerum Research Institute. This research was supported by a grant fromRazi Vaccine and Serum Research Institute.
200 S. M. Ebrahimi et al.




![Public Awareness of Long COVID & Motivation to Get Vaccinated · 2021. 7. 19. · [Single select]Base: Unvaccinated and not too/not at all concerned about long COVID N=160; only fielded](https://img.pdfslide.us/doc/110x75/61479ef8afbe1968d37a2b2b/public-awareness-of-long-covid-motivation-to-get-vaccinated-2021-7-19.jpg)