Embed Size (px)
Citation preview

STUDIES ON THE INFECTIVITY AND TRANSMISSION POTENTIAL OF
SIMULIUM DAMNOSUM COMPLEX IN UZO-UWANI LOCAL
GOVERNMENT AREA OF ENUGU STATE, NIGERIA
BY
ONAH, IKECHUKWU EUGENE
PG/M.Sc/09/51248
SUBMITTED TO
THE DEPARTMENT OF ZOOLOGY FACULTY OF BIOLOGICAL SCIENCES
UNIVERSITY OF NIGERIA, NSUKKA
IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE AWARD OF
MASTER OF SCIENCE (M. Sc) IN ZOOLOGY (ENTOMOLOGY)
SEPTEMBER 2011.

i
TITLE PAGE
STUDIES ON THE INFECTIVITY AND TRANSMISSION POTENTIAL OF
SIMULIUM DAMNOSUM COMPLEX IN UZO-UWANI LOCAL
GOVERNMENT AREA OF ENUGU STATE, NIGERIA

ii

iii
DEDICATION
This work is dedicated to my beloved mother Mrs A. C. Onah.

iv
ACKNOWLEDGEMENT
My profound gratitude is due to Almighty God who gave me the guide, protection and strength to
carry out this research project.
My profound gratitude is also due to Prof. P. O. Ubachukwu who made strenuous effort to see me
through in this project work. Her criticisms, suggestions and ingenuity are invaluable in making
this research work a success. I also wish to express my profound gratitude to Prof. J. E. Eyo
whose inspiration was crucial in the completion of this research work.
I also wish to register my sincere gratitude to Mr David Ekale, Mr Ngene Augustine of the
Department of Veterinary Medicine, University of Nigeria, Nsukka and Mr Haruna S. A. without
who this research work would not have been an easy task.
I also wish to express my gratitude to all the PG students and staff in the Department of Zoology,
University of Nigeria, Nsukka for their contributions and understanding.
I also want to acknowledge Eze Oluchi who laboured in typesetting the work.

v
ABSTRACT
The infectivity and transmission potentials of Simulium damnosum complex in Adani, Uzo-Uwani
Local Government Area of Enugu State, Nigeria were investigated from August 2010 to January
2011. A total of 548 adult female S. damnosum complex were caught using human baits and
dissected. Of this number, 248 flies were caught in the rainy season (August to October) while
300 flies were caught in the dry season (November to January). The relative abundance of flies in
Adani varied from 21 in December to 243 in January. The monthly catches between September
and October; and December and January were significantly different (P < 0.05). The monthly
population density of the flies ranged from 0.5Flies/Man/Hour (FMH) in December to 5.5FMH in
January. The diurnal biting pattern of S. damnosum complex in Adani showed a bimodal peak of
activity with the evening peak being higher than the morning peak except in October when the
morning peak was higher than the evening peak. The morning peaks were observed between
7.00am – 10.00am while the evening peaks occurred between 4.00pm – 6.00pm. The morning and
evening biting peaks in all the months were not significantly different (P > 0.05). Nulliparous flies
accounted for 75.7% of the total catch while 24.3% of the flies caught were parous. The infection
and infective bites, and transmission of Onchocerca volvulus during the study period was zero.
This suggests that there is low transmission of Onchocerca volvulus parasites in Adani. The flies
are presently more of nuisance biters rather than disease vectors since no stage of O. volvulus was
found in the flies dissected.

vi
TABLE OF CONTENTS
Title page - - - - - - - - - i
Certification - - - - - - - - - ii
Dedication - - - - - - - - - iii
Acknowledgment - - - - - - - - iv
Abstract - - - - - - - - - v
Table of contents - - - - - - - - vi
List of tables - - - - - - - - - ix
List of figures - - - - - - - - - x
List of plates - - - - - - - - - xi
CHAPTER ONE: INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction - - - - - - - - 1
1.2 Significance of the Study - - - - - - - 2
1.3 Objectives of the Study - - - - - - 3
1.4 Literature Review - - - - - - - - 4
1.4.1 Morphology of Simulium damnosum - - - - - 4
1.4.2 Classification of Simulium damnosum - - - - - 4
1.4.3 Life cycle of Simulium damnosum - - - - - 4
1.4.4 Habit of Simulium damnosum - - - - - - 6
1.4.4.1 Degree of Zoophily - - - - - - - 6

vii
14.4.2 Biting Site and Patterns of Simulium damnosum - - - 7
1.4.5 Medical importance of black flies. - - - - - 8
1.4.5.1 Annoyance. - - - - - - - - 8
1.4.5.2 Vector of human onchocerciasis - - - - - 9
1.4.6 Simulium damnosum and human onchocerciasis. - - - 10
1.4.6.1 Mode of transmission of human onchocerciasis - - - 10
1.4.6.2 Clinical manifestations of the disease - - - - 13
1.4.6.3 Prevalence of onchocerciasis and the disease burden - - 14
1.4.6.4 Factors influencing transmission of onchocerciasis - - - 17
1.4.6.5 Diagnosis of human onchocerciasis - - - - - 22
1.4.6.6 Treatment and control of human onchocerciasis - - - 22
CHAPTER TWO: MATERIALS AND METHODS
2.1 The study area - - - - - - - 25
2.2 Blackfly catching method - - - - - - 27
2.3 Dissection of the black flies - - - - - - 27
2.4 Calculation of transmission indices - - - - - 28
2.4.1 Monthly population density - - - - - 28
2.4.2 Daily Biting Rate (DBR) - - - - - - 28
2.4.3 Monthly Biting Rate (MBR) - - - - - 29

viii
2.4.4 Monthly Transmission Potential (MTP) - - - - 29
2.4.5 Proportion of parous flies - - - - - - 29
2.5 The diurnal biting pattern - - - - - - 30
2.6 Statistical Analysis. - - - - - - - 30
CHAPTER THREE: RESULTS
3.1 Relative abundance of flies in Adani - - - - - 31
3.2 The diurnal biting pattern of flies in Adani - - - - 33
3.3 Transmission indices of onchocerciasis in Adani - - - 40
CHAPTER FOUR: DISCUSSION
4.1 Discussion - - - - - - - - - 45
4.2 Conclusion and Recommendations - - - - - 53
REFERENCES - - - - - - - - 54
APPENDIX - - - - - - - - 60

ix
LIST OF TABLES
Table1. Diurnal biting activity of S. damnosum in Adani in August, 2010 - -34
Table 2. Diurnal biting activity of S. damnosum in Adani in September, 2010 -35
Table 3. Diurnal biting activity of S. damnosum in Adani in October, 2010- -36
Table 4. Diurnal biting activity of S. damnosum in Adani in November, 2010 -37
Table 5. Diurnal biting activity of S. damnosum in Adani in December, 2010 -38
Table 6. Diurnal biting activity of S. damnosum in Adani in January, 2011- -39
Table 7. Population density and Transmission indices of S. damnosum complex in Adani Uzo-
Uwani Local Government Area of Enugu State - - - - 41

x
LIST OF FIGURES
Fig.1: Life cycle of a Black fly - - - - - - 5
Fig.2: Life cycle of O. volvulus showing mode of transmission of the parasite- 12
Fig.3: Factors affecting vector capacity of Simuliidae to O. volvulus in
Latin America - - - - - - - - - 21
Fig.4: Map of Adani - - - - - - - - 26
Fig.5: Relative abundance of S. damnosum complex and mean monthly
rainfall in Adani from August 2010 to January 2011. - - - 32
Fig. 6: Diurnal biting pattern of S. damnosum complex in Adani in
August, 2010. - - - - - - - - 34
Fig.7: Diurnal biting pattern of S. damnosum complex in Adani
in September, 2010. - - - - - - - 35
Fig.8: Diurnal biting pattern of S. damnosum complex in Adani in October, 2010 36
Fig.9: Diurnal biting pattern of S. damnosum complex in Adani in
November, 2010. - - - - - - - 37
Fig.10: Diurnal biting pattern of S. damnosum complex in Adani in
December, 2010. - - - - - - - 38
Fig. 11: Diurnal biting pattern of S. damnosum complex in Adani in
January, 2011. - - - - - - - - 39

xi
LIST OF PLATES
Plate 1: Malpighian tubules exposed for determining parity. - - - -42
Plate 2: Normal ovary of S. damnosum displayed. - - - - -43
Plate 3: Infected ovary of S. damnosum. - - - - - -44

1
CHAPTER ONE
INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction
Onchocerciasis, also known as river blindness, caused by the filarial nematode Onchocerca
volvulus, is a chronic parasitic infection with public health and socioeconomic consequences of
considerable magnitude in many sub-Saharan African countries (Moyou-Somo et al., 1993; Opara
et al., 2005). It is an insect-borne disease transmitted by blackfly. It is the world’s second leading
infectious cause of blindness. In most of these countries, it constitutes a public health problem and
a serious obstacle to socio-economic development (Etyàale, 2001).
The disease affects about 17 to 18 million people in 37 countries of the world with approximately
123 million being at risk of infection (WHO, 1997). In Nigeria, onchocerciasis is widespread and
a cause of blindness in most rural communities. Of all the countries of the world, Nigeria has the
largest number of persons with onchocerciasis accounting for about a third of the global
prevalence with about 40 million at risk of infection (Edungbola, 1991). In Nigeria, Onchocerca
volvulus is transmitted primarily by the Simulium damnosum complex (Opara et al., 2008). The
prevalence of human onchocerciasis was shown to be directly related to the abundance of this
blackfly Simulium damnosum complex (Opara et al., 2005).
In West Africa and East Africa, the predominant species is Simulium damnosum complex, which
coexist with other species such as S. sirbanum, S. sanctipauli and S. soubrense (Atting et al.,
2005). Simulium neavei is a much less important vector which is responsible for transmission in
Congo, Zaire and Uganda (Service, 2004). In Nigeria, Simulium species are responsible for the
prevalence of onchocerciasis. Cytotaxonomic studies of the S. damnosum complex from different
parts of Nigeria have revealed the presence of the five cytospecies, these include: S. damnosum
sensu stricto, S. sirbanum, S. squamosum, S. yahense and S. soubrense (Atting et al., 2005). An
understanding of the transmission dynamics of onchocerciasis as in other forms of filariasis is

2
important in advancing knowledge of how vector capacity, vector abundance, survival rate,
feeding habit and behaviour influence the level of infection and disease in susceptible human
population. The knowledge of vectorial capacity would be of immense value in formulating the
most appropriate control strategies in a given locality (Opara et al., 2008)
1.2 Significance of the Study
Uzo-Uwani Local Government Area of Enugu State, Nigeria is a major food producing area in the
eastern part of Nigeria (Ubachukwu and Anya, 2001a).
Adani in Uzo-Uwani Local Government Area is popular because of the agricultural activities in
the area. Adani is one of the best known and major rice producing communities in South Eastern
Nigeria (Ezedinma, 2011). The agricultural activities in the area are intensified because of Obina
River. Besides serving as source of water for household use, the river is used for fishing and
irrigation purposes. Unfortunately, the river system that forms the basis of agriculture in the area
favours the breeding of blackflies in large numbers. This has been a serious constraint to further
agricultural development in the area.
Many works have been done on the infectivity and transmission dynamics of Simulium damnosum
in different parts of the world. Adewale et al. (1999) studied the infectivity and transmission
dynamics of Simulium damnosum s. l. around Owena Dam (Ondo State); Porter and Collins
(1988) studied the seasonality of adult blackflies and Onchocerca volvulus transmission in
Guatemala; Barbiero and Trpis (1984) studied the transmission of onchocerciasis by local
blackflies on the firestone rubber plantation harbel, Liberia; Opoku (2000) studied the ecology,
biting activity and parasite infectivity of the blackfly and onchocerciasis prevalence in the River
Birim catchment, Ghana; Opara et al. (2008) studied transmission dynamics of Simulium
damnosum in rural communities of Akwa Ibom State, Nigeria; Atting et al. (2005) studied
seasonal variations in human onchocerciasis transmission by black flies in a forest area of Cross
River State, Nigeria; Grillet et al. (2001) studied human onchocerciasis in Amazonian Area of
Southern Venezuela: Spatial and temporal variations in biting and parity rates of blackfly

3
(Diptera: Simulidae) vectors in Venezuela; Shelley (1991) studied transmission and control of
onchocerciasis in Latin America; Taye et al. (2000) studied onchocerciasis in Gilgel Ghibe River
valley southwest Ethiopia; Renz and Wenk (1987) studied the dynamics of transmission of
onchocerciasis in a Sudan-savanna area of North Cameroon and many others.
Despite the contributions of Adani in ensuring food security and other economic activities, the
menace of blackflies in the area has not been given a serious attention. Research on the dynamics
of Simulium population and vector infectivity and thus the level and magnitude of parasite
transmission in this area has not been carried out. It is only through this sort of research that the
attention of the Onchocerciasis Control Programme (OCP), the African Programme for
Onchocerciasis Control (APOC), World Health Organization (WHO) and other Governmental and
Non-Governmental and Development Organizations (NGDOs) can be attracted to come to the aid
of Adani people.
The capture and dissection of adult flies can be used as a means of following the dynamics of
Simulium population and vector infectivity and thus the level and magnitude of parasite
transmission (Opara, et al., 2005).
1.3 Objectives of the Study
The objectives of the study are:
1. To assess the population density of Simulium damnosum complex in Adani.
2. To determine the diurnal biting pattern of S. damnosum complex in the area
3. To evaluate the transmission indices of onchocerciasis in the area which include
i. Daily biting rate
ii. Monthly biting rate
iii. Monthly transmission potential
iv. Proportion of parous flies

4
1.4 Literature Review
1.4.1 Morphology of Simulium damnosum
The blackfly Simulium (Diptera: Simuliidae) is a small, sturdy, hump-backed Nematocera with
the immature stages limited to fluvial ecosystems, breeding in swiftly-running water (OCP, 1985;
Opoku, 2000). They are dark flies, 3-6mm long, with short, stout legs and long mandibles. The
females have dichoptic eyes while the eyes are holoptic in males. Antennae are 9-11 segmented,
without hairs in both sexes. The wings are large and broad. The anterior veins are stout while the
rest are weak. The males usually have the first tarsal joints swollen (Nayar et al., 1992).
1.4.2 Classification of Simulium damnosum
Simulium damnosum can be classified into the six obligate taxa thus: Phylum-Arthropoda, Class-
Insecta, Order-Diptera, Family-Simuliidae, Genus-Simulium and Species - Simulium damnosum
(Nayar et al., 1992).
1.4.3 Life cycle of Simulium damnosum
Blackflies breed in fast flowing waters of streams and rivers with high oxygen content. Eggs are
deposited in water in sticky masses or strings. The eggs hatch in 1 – 4 days depending on
temperature into larvae which spin silken thread from the salivary gland. The larva is sedentary
and holds unto substrates in water by the small silken pad and the hooked sucker at its posterior
end. The head bears a pair of cephalic fans or feeding brushes around the mouth used for filter
feeding. There are six or seven larval instars which develop in 7-12 days under suitable
conditions, but some species take longer. When the larva is ready to pupate, it spins a cocoon
around itself using the silken thread and attaches it to a substrate. The pupa does not move and
lasts from a few days to few months, respiring through the respiratory filaments at the anterior
end. The adult eventually crawls out from the pupal case and take to flight shortly after. Mating
usually occurs in flight and the females suck blood of mammals and birds for the maturation of
eggs (Crosskey, 1990; Nayar et al., 1992; Ubachukwu, 2009).

5
Fig. 1: Life Cycle of a Black Fly (Source: Ottawa Rambling Club 2001).

6
1.4.4 Habits of Simulium damnosum
Both males and female blackflies feed on plant juices and naturally occurring sugary substances,
but only females take blood meals. In many species vision seems important in host location but
host odours also play a role. Biting occurs out of doors during the day. After feeding, blood-
engorged females shelter in vegetation, on trees and in other natural outdoor resting places until
the blood meal is completely digested. In the tropics this takes 2-3 days, while in non-tropical
areas it may take 3-8 days or longer, the speed of digestion depending mainly on temperature. A
few species can lay eggs without a blood meal (i.e. they are autogenous). Relatively little is
known about blackfly longevity, but it seems that adults of most species live for 3 – 4 weeks
(Service, 2004). The adults of the various species differ in flight range and biting habits but S.
damnosum is known to travel up to 100km during the first few days after emergence. In West
Africa, winds can carry adult S. damnosum up to 400-600 km. The long distances involved in
dispersal have great relevance in control programmes, because areas freed from blackflies can be
reinvaded from distant breeding places. In Central and South America, blackflies generally
disperse only about 2 – 15 km. The adult fly populations are often very large, and they may
emerge in enormous swarms that together with the irritating nature of their bites render riverine
areas almost uninhabitable (Metcalf and Novak, 1994; Service, 2004).
(a) Degree of zoophily: Simuliids bite both mammals and birds and frequently appear in swarms.
They bite the ears and udders of cattle and inside the ears, causing chronic inflammation of the
affected parts and irritation. Blood oozes out through the punctures and bleeding wounds caused
by several punctures cause death of cattle (Nayar et al., 1992). When feeding on animals, adults
crawl down fur of mammals, or feathers of birds, to bite the host’s skin (Service, 2004).
Many species of blackfly feed almost exclusively on birds (ornithophagic), others on non-human
mammalian hosts (zoophagic), while several species also bite people (anthropophagic). Some of
these, however, actually prefer large animals such as donkeys or cattle and bite humans only as a

7
poor second choice, whereas others appear to find humans equally attractive hosts; no species
feed exclusively on people (Service, 2004).
(b) Biting site and patterns of Simulium damnosum: Biting by blackflies occur out-doors at
almost any daylight hour but each species of blackfly may have its preferred times of biting. A
study conducted in Obubra Local Government Area of Cross River State, Nigeria revealed that
the biting density of Simulium vary with the time of the day (Iboh and Braide, 1987). Variations
have also been observed in the monthly proportions of S. damnosum s. l. biting various body
regions (Iboh and Braide, 1987). In addition, researchers working in different countries of the
world have observed that irrespective of the posture of the catchers, the legs and any exposed
parts of the body nearest to the ground are the preferred biting sites of Simulium (Iboh and Braide,
1987). A study in the forest area of Cross River State, Nigeria also revealed that the Daily Biting
Rate (DBR) and the Monthly Biting Rate (MBR) were higher in the rainy season than in the dry
season (Atting et al., 2005). However, Renz (1987a) reported that few flies were caught at Mayo
Galke causeway when the flow of water was more than 100m3sec
-1 at the height of the rainy
season. At the River Vina, near the Tonboro, (Renz, 1987a) observed that the biting densities
were at their lowest level during the rainy season in October 1976.
The diurnal biting cycle of black flies shows a bimodal peak of activity. There is usually a small
morning peak and a pronounced evening peak. The two peaks are separated by hours of low biting
intensity (Ubachukwu and Anya, 2001a; Opara et al., 2005). Early morning and late afternoon
hours were the most dangerous times with respect to the exposure to the bites of the flies and to
transmission of onchocerciasis (Renz, 1987a). Diurnal variations in the biting density have been
related to variations in the temperature (Leberre, 1966) and humidity (Hausermann, 1969) or to
the intensity of light (Kaneko et al., 1973). The variation in the biting activity of the flies delimits
the hours of maximum or minimum danger of transmission of onchocerciasis (Renz, 1987a).
According to Service (2004), many species including S. damnosum in Africa have bimodal biting
patterns, with a peak in the early morning and another in the afternoon or early evening. However,

8
in some species, such as S. ochraceum in Guatemala, biting continue more or less throughout the
day. Many species seem particularly active on cloudy, overcast days and in thundery weather.
Blackflies may exhibit marked preferences for feeding on different parts of the body; for example,
S. damnosum feeds mainly on the legs whereas S. ochraceum prefers to bite on the head and torso
(Service, 2004).
1.4.5 Medical Importance of Blackflies
(a) Annoyance: In both tropical and non-tropical areas of the world, blackflies can cause a very
serious biting problem. Blackflies are in many regions the most feared of all biting insects
because of the relentless and intolerable nature of their attacks, not only on man but on livestock,
poultry and wildlife too. The experience of being continually bitten, unable to step outside
without soon oozing blood from countless bites, is demoralizing (Crosskey, 1990).
Female blackflies, without even biting, can be almost unendurably annoying to man and deeply
distressing to domestic animals. Clouds of them may swarm about the body, especially the head
and face – when there is a great risk of their being inhaled into the lungs. Some species not
seriously interested in taking blood from man have the intensely irritating habit of landing
intermittently and crawling briefly on the skin, or darting into the ears and eyes, the mouth and
nostrils (Crosskey, 1990).
Crosskey (1990) stated that in Africa the highest biting rate of S. damnosum complex are usually
not above 30 – 60FMH in Savanna and 200FMH in forest areas, whereas biting densities of 100 –
1000FMH are commonplace with S. ochraceum complex in Guatemala and 300 – 450FMH
typical of S. oyapockense in the Amazon. Opara et al. (2008) recorded biting densities of
27.7FMH, 35.8 FMH and 33.5FMH in three different sampling points in Akwa Ibom State,
Nigeria. At Mayo Galke in the Sudan-savanna of North Cameroon, an Annual Biting Rate (ABR)
of as high as 83800flies/man/year was recorded in 1976 according to Renz (1987b).

9
The most aggravating man-biting blackflies, those potentially or actually of practical importance
to the health or the social economy are found in parts of North America, Central America, tropical
South America, Europe, USSR and New Zealand. Entomologists familiar with the African scene
are usually much surprised at the biting densities of vectors when they visit the New World foci.
In Southern Island, blackflies (Austrosimulium ungulatum) attack man in such vast numbers that
an unprotected person may experience as many as 1000 bites in an hour, an intolerable level of
attack that deter development of tourism in the region. It has also been reported that 6780 flies
were collected from the skin of one Amerindian volunteer in a single day, an average attack rate
of 565 FMH over 12 hours of catching (Crosskey, 1990). In some areas of North America
outdoor activities are almost impossible at certain times of the year due to the intolerable numbers
of biting simuliids (Service, 2004).
(b) Reaction to Simulium bites: Although the severity of the reaction to bites differs in different
individuals, localised swelling and inflammation frequently occurs, accompanied by intense
irritation lasting for several days or even weeks. Repeated biting by blackflies such as S.
erythrocephalum in Central Europe, S. posticatum in England, S. venustum and S. vittatum in
North America can cause headaches, fevers, swollen lymph glands and aching joints (Service,
2004). Blackflies produce irritations and itches and in sensitive individuals restlessness and
allergic reactions may prevail. They have also been reported to produce toxic saliva and their bites
lead to formation of vesicles, the resulting ulcers often prone to secondary infections (Igbinosa,
1990).
(C) Vector of human onchocerciasis: Blackflies are important in the transmission of
onchocerciasis – a form of filarial infection due to Onchocerca volvulus. S. damnosum and S.
neavei are of importance in Africa including Nigeria, whereas in Central America S. ochraceum
and S. metallicum are the important species (Igbinosa, 1990).

10
Human onchocerciasis is a parasitic disease of tropical Africa and the tropical Americas caused
by a filarial worm, Onchocerca volvulus. The infection is transmitted among humans by the biting
female black flies of the genus Simulium (Metcalf and Novak, 1994).
1.4.6 Simulium damnosum and Human Onchocerciasis
(a) Mode of transmission of human onchocerciasis: Onchocerca volvulus, the causal agent of
onchocerciasis is one of a large group of nematodes. The adult worms live encysted in fibrous
nodules. Each nodule contains between 2 to 3 female worms lying in a twisted, tangled mass.
Adult female worms have a life span of 8 to 10 years but may live up to 15 years during which
time each releases millions of first – stage larvae, also known as microfilariae. In hyperendemic
areas, the total microfilaria load in the body of affected individuals may be as high as 150 million
(Etya’ale, 2001).
Onchocerciasis is transmitted from one individual to another by a blackfly of the genus Simulium
(Metcalf and Novak, 1994; Etya’ale, 2001). Female black flies require a blood meal to produce
eggs, and it is during this meal that they may transmit or receive the onchocercal infection
(Etya’ale, 2001).
The female blackfly becomes infected when it bites an infected person and ingests microfilariae
along with its blood meal. Although many of the microfilariae are digested, a few may succeed in
penetrating through the fly’s gut and entering the thoracic muscle where they pass through three
larval stages over a period of about one week. Infective (third stage larvae) are formed which pass
from the fly’s blood into the proboscis and infect a subsequent human host as the fly feeds.
Infectivity is low and a victim must receive numerous bites before pairs of adult filariae can
survive and mate to produce a generalized infection (Metcalf and Novak, 1994; Etya’ale, 2001).
In the human body, the larvae form nodules in the subcutaneous tissue, where they mature to adult
worms. After mating, the female adult worm can release up to 1000 microfilariae a day. These

11
move through the body and when they die they cause a variety of conditions, including
blindness, skin rashes, lesions, intense itching and skin depigmentation (WHO, 2002).

12
Fig 2: Life cycle of O. volvulus showing mode of transmission of the parasite.
Source: Springer (2011).

13
O. volvulus is found in subcutaneous connective tissues (skin of man). The parasite is prevalent
in Nigeria, Ghana and is endemic in many other parts of the world but more in the tropics and
subtropics in areas with fast flowing rivers (Okafor, 1990).
(b) Clinical manifestations of the disease: There are very few external signs like the nodules.
The parasites wander about in connective tissues. The mature males and females collect in balls
bound together by fibrous tissue which forms the nodule typical of the infection. Mature worms
can also occur in the skin in the absence of nodules. The typical nodule is round and hard, 1-5 cm
in diameter (Okafor, 1990).
The people most at risk of onchocerciasis are those for reasons of occupation (example:
fishermen, farmers, sand diggers) spend long hours or live nearer to the breeding sites. Early
manifestations of the disease in infected persons usually appear one to three years after the
injection of infective larvae. The clinical features of onchocerciasis may be divided into two main
groups: (1) Non-Ocular and (2) Ocular (Etya’ale, 2001).
1. Non-Ocular Manifestation: These include itching, disfiguring of skin/skin
lesions. In addition, onchocerciasis could result in such important complications
as onchocercomata (nodules), hanging groin, hernia and elephantiasis (Metcalf
and Novak, 1994; Opoku, 2000; Etya’ale, 2001; Atting et al., 2005;
Ubachukwu, 2009).
2. Ocular Manifestations: These include sight impairment and blindness, serious
eye lesions, night blindness and visual field loss and constriction. Other ocular
manifestations include:
Punctate keratitis, leading on to sclerosing keratitis.
Early uveitis leading on to chronic uveitis.
Choroido-retinitis, leading on to choroido-retinal atrophy or
Optic nerve atrophy
Acute optic neuritis, leading on to optic atrophy (Etya’ale, 2001).

14
Live microfilariae cause no tissue reaction but dead ones cause an immediate tissue
reaction which is readily seen following treatment with the drug. These form the basis of
the Mazzotti reaction. At first there is an oedema and an infiltration of the dermis with
eosinophil cell, followed by a cell mediated response with macrophage cells, lymphocytes
and sometimes giant cells forming granuloma with subsequent fibrosis. Onchodermatitis
which is the later stage result in mottled depigmentation of the skin, as well as atrophy of
the skin leading to hanging groin. Retinopathy, retinitis and choroiditis involving optic
nerves are caused by Onchocerca volvulus which later leads to blindness or other ocular
lesions (Okafor, 1990).
In Nigeria, Okafor (1990) reported that ocular onchocerciasis is less prevalent and less
severe in the rainforest areas than in the savanna zone. This according to him is
attributable to two reasons:
(i) Three clinical classes of S. damnosum females are recognised viz: ‘’A’’, ‘’B’’ and
‘’C’’. It is reported that the ‘’C’’ females are found in the savanna zone while the ‘’A’’
females are found in the rainforest zone.
(ii) Two strains of O. volvulus exist. These two strains differ in their basic ability to
damage ocular tissues and use two different clinical classes of S. damnosum.
O. volvulus is the only filarial worm that develops to maturity and produce microfilariae in
the skin of man among the filarial worms.
(c) Prevalence of onchocerciasis, the disease burden and socio-economic consequences:
Onchocerciasis, a tropical parasitic disease, poses a serious public health problem and a serious
obstacle to socio-economic development in much of West Africa. It causes abandonment and /or
prevention of cultivation of fertile lands in endemic communities. As a result of this scourge,
whole communes in the northern Ghana had as at 1974 abandoned their fertile riverine villages
and moved interior into less fertile lands to scrape a living (OCP, 1980; Opoku, 2000; Atting et
al., 2005). About 125 million people world – wide are estimated to be at risk of onchocerciasis
and of these 96% are in Africa. Of the 37 countries where the disease is endemic, 30 are in sub-

15
Saharan Africa, six are in the Americas and one is in the Arabian Peninsula. A total of 18
million people are infected with the disease, of whom 99% live in Africa and at least one million
are either blind or severely visually disabled. To these are added each year an estimated 40, 000
new blind (Etya’ale, 2001).
In the Americas, onchocerciasis is localized in areas of Southern Mexico, Guatemala, Brazil,
Venezuela, Ecuador and Colombia. In Africa, onchocerciasis occurs throughout West Africa,
Central Africa, and much of East Africa from Ethiopia to Tanzania with isolated pockets of
infection in Malawi, Sudan and Southern Yemen, possibly extending into Saudi Arabia (Service,
2004).
The name “river blindness” suggests that onchocerciasis is essentially a focal disease. However,
where it exists, its impact on affected communities may be quite extensive and devastating. Thus,
in many hyperendemic areas with blinding onchocerciasis, almost every person will be infected
and half of the population will be made blind by the disease before they die. Once blind, affected
individuals have a life expectancy of only one third that of the sighted and most of them die
within ten years (Etya’ale, 2001).
Recent studies in Ethiopia, Nigeria and Sudan have also shown that onchocerciasis is responsible
for poor school performance and a higher dropout rate among infected children (due to itching,
lack of sleep, etc), while low productivity, low income and higher health-related cost are found
among infected adults (Etya’ale, 2001). The result of the effects of onchocerciasis manifestations
on academic performance showed that the higher the rate of visual impairment in a school, the
poorer the academic performance of such a school (Ubachukwu and Anya, 2003). In Nkpologu in
Uzo-Uwani Local Government Area of Enugu State, Ubachukwu and Anya (2001b) showed that
the loss on the productivity of farmers per man per season due to the bites of blackflies and
onchocerciasis was about N6, 500.00.
Onchocerciasis is usually a serious threat to public health and an impediment to socio-economic
development in areas with high intensity and high endemicity of the disease. Inhabitants of fertile

16
river valleys move to the less fertile upland country. Many young men migrate to urban areas,
reducing the productivity of the community and disrupting family life (Ubachukwu, 2006).
According to Kale (1998), the greatest burdens related to human onchocerciasis are the result of
the eye and skin lesions and severe itching produced by the microfilariae. He also said that the
skin lesions are a major socio-economic burden in terms of disability-adjusted life-years (DALY).
Employees’ classified as having a severe Onchocercal Skin Disease (OSD) earned 15 % less in
daily wages than those not infected. People with Onchocercal Skin Disease are stigmatized in
their communities. OSD limits the range of social involvement and can affect sexual life of
affected individuals. With reference to women and children, young females with OSD suffer
stigmatization more than young men. This affects their age of marriage and the kind of partners
they marry, limiting them to already married men, divorced men, elderly men, childless men, etc.
Severe itching that often accompanies OSD may reduce the period lactating mothers’ breastfeed
their babies. Children, particularly females, from households headed by individuals with
onchocerciasis, especially blindness and OSD are more at risk of being school dropouts.
Academic performance of school children with visual impairment is adversely affected
(Ubachukwu, 2006).
The socio-economic liabilities as a result of onchocerciasis are enormous. The blackfly vectors of
Onchocerca volvulus are a serious nuisance in the endemic communities because of the resultant
skin lesions from their bites. Susceptible persons may be uncomfortable for weeks with an almost
unbearable pruritus and scratching. In many individuals, this persists throughout the whole course
of the infection. Sometimes, the itching and scratching may be so severe as to cause insomnia
(Nwoke et al., 1987).The various skin changes associated with onchocerciasis such as rashes,
hypopigmentation and scaling, oedema and depigmentation have distressing effects on the
lifestyle of the infected individuals (Nwoke, 1986; Nwoke et al., 1987), sometimes constituting
destitute (Nwoke, 1990). The presence of hanging groin and elephantiasis of the genitalia
commonly seen in adult males and genital distortion seen in females nearly always results in the

17
infected individual’s unwillingness towards a free interaction within his or her locality. In
infected individuals with the pendulous sacs, sexual life is greatly affected if not completely
hindered (Nwoke, 1986; Nwoke et al., 1987; Nwoke, 1990). A study on perception and social
implication of onchocerciasis in Edo State, Nigeria, showed that attitude of non-affected towards
the affected is partially discriminatory and suspicious. The affected are socially withdrawn due to
frustration of their health condition (Wagbatsoma and Okojie, 2004). Jamnback (1973) reported
that in areas of major developmental projects, the incessant bites of blackflies could be a serious
threat to economic success. In Cote d`Ivoire, it was reported that farmers in the oncho-freed zones
around the Leraba, Bou and Fombou rivers could only work between 1000H and 1530H due to S.
damnosum s. l. biting nuisance (Boakye et al., 2009). This translates into 2.5 hours of work lost,
in addition to forcing the farmers to work during the hottest hours of the day. In Ghana, a possible
negative impact of the biting nuisance on tourism was observed around the Kakum National Park
during the rainy season in 1999 (Boakye et al., 2009).
Of all the countries of the world, Nigeria has the largest number of persons with onchocerciasis
accounting for about a third of the global prevalence with about 40 million at the risk of infection
(Edungbola, 1991).
(d) Factors that influence transmission of onchocerciasis: Many factors that affect or involve
both the parasite and simuliid host influence the transmission of onchocerciasis. In the parasite
they include the pathogenicity of the strain or population of O. volvulus, its accessibility to the
vector in terms of its distribution in infected individuals in relation to the preferred biting site of
the vector. In the vector, the factors that affect the efficiency of a simuliid species to transmit O.
volvulus may be divided into two groups: those that influence the ability of the fly to host the
parasite (host capacity) and those that interact with its host capacity and thereby influence the
ability of a species to transmit the parasite (vector capacity) (Shelley, 1991).
(i) Host capacity: Factors that affect host capacity are the presence or absence of a cibarial
armature in the fly, the presence or absence of intrinsic barriers to a synchronous development

18
cycle of the parasite in the fly, attractants in the saliva of flies that cause concentration of
microfilariae at the biting site and the speed of formation of the peritrophic membrane in the gut.
An additional factor not yet investigated is the production of acquired resistance to further filarial
infection by previously infected flies, as was shown to occur in two species of British blackflies
infected with the bovine filarial O. lienalis (Shelley, 1991).
Vector simulid species may be divided into two groups according to the effect that the presence or
absence of a cibarial armature has on the fly’s host capacity. In species with an armed cibarium,
host capacity is always low because the cibarial teeth damage many microfilariae ingested during
blood feeding by the fly and prevent their development to infective L3. High host capacity is only
found in species with no cibarial armature (S. callidum, S. exiguum, S. metallicum, S. guianense).
However, this has been studied extensively only in Latin American species of simuliids (Shelley,
1991).
However, a lack of cibarial teeth does not always signify high host capacity since intrinsic
barrier may exist that prevents full development of microfilariae to infective larvae. Thus, the
Cayapa cytospecies of S. exiguum is a high efficient host to O. volvulus in Ecuador with most
microfilariae developing rapidly to the L3 stage within 5-8 days. In Colombia, where a different
cytospecies of S. exiguum probably occur, the vector is less efficient since parasite development
in the fly is partially asynchronous resulting in a lower number of microfilariae reaching infective
larval stage in the same time period. Increased asynchrony of the development cycle results in an
even lower host capacity in S. exiguum s. l. of northern Venezuelan foci where a different
cytospecies probably occurs and where the development period of the few microfilariae that do
not become stunted is prolonged. Similarly, S. metallicum s. l. populations in Guatemala and
Mexico are poor hosts compared with populations in northern Venezuela, again probably
attributable to the presence of different cytospecies. Though a high proportion of microfilariae
eventually develop to L3 larvae in the northern Venezuela foci partial asynchrony of the
development cycle retards parasite development and here S. metallicum s. l. is a less efficient

19
vector than the Cayapa form of S. exiguum in Ecuador. Asynchrony in the parasite development
cycle in species with armed cibaria has only been recorded in S. haematopotum (Shelley, 1991).
The number of parasites ingested with the blood meal together with cibarial morphology
influences the final host capacity of the fly population. The number of microfilariae ingested is
related to two factors: parasite density in the host’s skin and the relative concentration effect
produced by different fly species. It has been demonstrated that a substance in the saliva of the fly
inoculated during feeding is responsible for attracting microfilariae to the site of the bite and this
effect varies with species (Shelley, 1991). A similar enhancing factor that increases the parasite’s
chances of transmission has been found in the saliva of phlebotomine vector of Leishmania. It has
been shown that in species with unarmed cibaria such as S. callidum, S. exiguum s. l., S.
metallicum s. l. and S. guianense, fly mortality rates increases with an increase in microfilarial
intake, directly linked to skin microfilarial densities and the concentration effect. The absence of
cibarial teeth in these species allows all ingested parasites to reach the mid gut intact and from
there invade other body organs causing the host to die from hyperparasitisation if microfilarial
intake is high. In contrast, the ingestion of large numbers of microfilariae by flies with armed
cibaria has a less drastic effect on fly mortality because the cibarial teeth damage many of the
parasites ingested during blood feeding and consequently prevent their further development. This
protective effect on the parasite has been most clearly demonstrated in the primary vector S.
ochraceum s. l. in Guatemala. The cibarial armature is, therefore, fundamental when considering
the effect that individual species have on disease endemicity and control. Species with no cibarial
teeth and high host capacities are able to transform a state of hypoendemicity to meso- and
hyperendemicity relatively rapidly (e.g. S. exiguum s. l. in Ecuador), whereas species with armed
cibaria, and consequently low host capacities, have little effect on parasite density and prevalence
rates over the short term (e.g. S. oyapockense s. l. in Brazil). The converse is probably true in
hyperendemic situations in that species with unarmed cibaria are unprotected from high
microfilarial intake and hence show higher mortality rates, whereas those species with armed

20
cibaria have lower mortalities and are more effective vectors when biting in high numbers
(Shelley, 1991).
(ii) Vectorial capacity: The vectorial capacity is defined as the product of all the intervening
factors that produce an infection in the vector and enable it to transmit the infection to a host
(Metcalf and Novak, 1994). This is determined by the interaction of host capacity with the other
factors affecting both fly and parasite such as seasonal biting densities, degree of zoophily, site of
biting on body and duration of bite, population density and longevity, dispersal and flight range
(Shelley, 1991; Metcalf and Novak, 1994; Grillet et al., 2001; Opara et al., 2005). Thus, a species
with high host capacity will not act as an efficient vector and will have a low annual transmission
potential (ATP) if man-vector contact is low because of low biting rates due to either small
population size or a predilection for zoophily. Conversely, a high ATP may occur in a species that
has a low host capacity if it is highly anthropophilic and has high man biting rates, if high
densities of parasites are available in man for transmission. In Africa the use ATPs for quantifying
transmission is routine, whereas in Latin America this method has been used only recently in
Guatemala and Mexico. Their quantification is important in the context of control programmes
(Shelley, 1991).

21
PARASITE IN MAN HOST CAPACITY OF FLY
Parasite density in man Concentration effect
Parasite prevalence in community Presence/absence cibarial teeth
Parasite distribution in body Speed of formation of peritrophic membrane
Synchrony of parasite cycle in fly
Acquired immunity to parasite
VECTOR CAPACITY
BIOLOGY OF FLY
Seasonal and daily biting densities
Degree of zoophily
Biting site on man
Duration of biting
Fig. 3: Factors affecting vector capacity of Simuliidae to O. volvulus in Latin America.
Source: Shelley (1991).

22
An understanding of the transmission dynamics of onchocerciasis as in other forms of filariasis
is important in advancing knowledge of how vector competence, behaviour, and abundance
influence the level of infection and disease in susceptible human population (Bockarie et al.,
1996). Quantitative analysis of potential vectors and their level of infection may also provide cost-
effective means of rapid assessment of the need for and success of various control measures.
(e) Diagnosis of human onchocerciasis: Diagnosis is made by finding either the microfilariae
in superficial skin shaving or punch biopsy, adult worms in histologic section of excised nodule or
characteristic eye lesions. Serologic testing is most useful for detecting infection in specific
groups, such as expatriates with a brief exposure history when microfilariae are not identifiable
(Fox, 2009). The presence of infection is confirmed by examination of snips for microfilariae
from thighs, buttocks and iliac crest or shoulder blade. Palpable nodules are also diagnostic
(Okafor, 1990).
Another potential diagnostic indicators of onchocerciasis according to Basile et al. (1995) and
Adeyeba and Adegoke (2002) is the Rapid Assessment Method (RAM) which according to them
include presence of subcutaneous nodules, depigmentation or leopard skin, microfilaruria,
diethylcarbamazine patch test positivity, excoriations and pruritus.
(f) Treatment and control of human onchocerciasis: Onchocerciasis control ideally aims at
reducing the parasite in man to a level at which it has no pathogenic effect and at which the vector
cannot be responsible for a rapid recrudescence of the disease. The level at which the disease is no
longer seriously pathogenic to man can be related to skin microfilarial density and the Annual
Transmission Potential (ATP). In Africa these levels have already been calculated and are used as
threshold targets in the Onchocerciasis Control Programme. However, in Latin America the
parasite level at which transmission only becomes sporadic has not yet been measured because of
the large number of vector species, each unique in its vector capacity and biology. It is postulated
that in foci in which species with unarmed cibaria and high host and vector capacities occur
recrudescence may be rapid even if parasite densities are low. Conversely, species with armed

23
cibaria do not produce significant changes in hypoendemic areas and only show high vector
capacities if biting rates and parasite levels are high. This evidence suggests that controls in areas
with vectors showing low host but high vector capacities needs to be indefinite, whereas
hyperendemic areas with vectors showing low host but high vector capacities reduction of the
parasite in man to a low, as yet undetermined critical level, and then cessation of control would be
sufficient (e.g. Central America) (Shelley, 1991).
The current strategy of controlling the disease in Africa relies mostly on the annual
chemotherapeutic treatment of the endemic communities through mass distribution of ivermectin.
Ivermectin has been proved to have microfilaricidal properties and substantially reduced the
burden in many affected communities (Dadzie et al., 1987; Yameogo et al., 1999).
The control of onchocerciasis today is based essentially on two strategies: Simulium vector control
and large-scale chemotherapy with ivermectin. Each may be used alone or in combination.
(i) Vector control: This is the chief strategy used in West Africa by the Onchocerciasis Control
Programme (OCP) since 1974. The main goal in vector control is to interrupt transmission of O.
volvulus by regular aerial spraying of all Simulium larval breeding sites and to maintain this for at
least 14 years until the infection has died out in human population. This strategy used alone at the
beginning and now in combination with ivermectin has been highly effective (Etya’ale, 2001).
Control measures include larviciding with chlorpyrifos methyl, Bacillus thuringiensis, use of
repellents such as deet and 2-ethylhexanediol; and use of pyrethroid impregnated clothing. Also
river channelling improvement to clean and deepen waterways with the removal of stones, logs,
and other obstruction that cause ripples attractive to blackfly larvae, the construction of outlets of
lakes, dams and spillways to provide clear, unobstructed drops into deep pools are effective in
vector control (Metcalf and Novak, 1994).
(ii) Chemotherapy: Ivermectin is the only chemotherapeutic agent recommended for use against
onchocerciasis. It is a semisynthetic, macrocyclic, lactone antibiotic widely used in the field of
veterinary medicine against a wide range of animal parasites. It is a microfilaricide with a very

24
wide therapeutic range (150-800 microgram/kg). However, because ivermectin has no
demonstrable direct effect on the adult worm, it must be given repeatedly for up to 12-15 years,
that is, the time it takes most adult worms to die (Etya’ale, 2001).
Nodulectomy or surgical removal of nodule is more effective with the head nodule of the Central
American onchocerciasis unlike the African forms where most nodules are in the lower
extremities and composed of dead or dying worms (Ubachukwu, 2009).

25
CHAPTER TWO
MATERIALS AND METHODS
2.1 The Study Area
Uzo-Uwani Local Government Area lies between longitude 60 55
1 and 7
0 20
1 East and between
latitude 60
301 and 7
0 00
1 North (Ubachukwu and Anya, 2003). It belongs to the forest-savanna-
mosaic zone of Nigeria. The area is traversed by many rivers and streams which belong to the
Anambra River system. These form the major breeding sites for the Simulium vector of human
onchocerciasis (Crosskey, 1981).
Adani is one of the communities in Uzo-Uwani Local Government Area of Enugu State. It lies
between latitude 60
441 and 6
0 52
1N and longitude 7
0 1
1 and 7
0 8
1E. The approximate population
of Adani for 7km radius from this point is 17, 992. It is bordered on the West by Ojjor, Igga,
Ogurugu and Umueje. On the North are Igulala, Okwoengele, Anokwo, Ameke, Alakija and
Akpatebe; on the East are Opandas, Ogbo-Uvuru, Adaba, Ukpabi-Nimbo, Ugwojoro, Nimbo, and
Ngalakwu. On the south are Omash and Ifute. The mean annual temperature is 22.50C (Google,
2010).
The people are primarily farmers being famous in rice and garri production.

26
. 5:. 4:

27
2.2 Blackfly Catching Method.
Biting adult female Simulium damnosum complex were collected using human baits (Taye et al.,
2000; Grillet et al., 2001; Ubachukwu and Anya, 2001; Opara et al., 2005; Opara et al., 2008). A
sampling point approximately 300 metres from the Obina River in Adani was used as fly
collection site. The site was sampled 4 times a month for six months (August 2010 to January
2011).
Fly catching was conducted between 7.00am and 6.00pm (11 hours) by two fly collectors
working alternately (Walsh et al., 1978; Taye et al., 2000; Opoku, 2000; Atting et al., 2005;
Opara et al., 2005; Opara et al., 2008). The fly collectors were dressed in knickers exposing the
lower legs (Atting et al., 2005). The collectors sat at a convenient place with the feet and the legs
below the knees exposed (Walsh et al., 1978). Any fly perching on the exposed parts was
collected before it started feeding by inverting a small glass tube over it and replacing the cap
immediately (Plate 3a and 3b) (Walsh et al., 1978; Atting et al., 2005; Opara et al., 2005). The
tubes containing the flies were labelled to indicate time and date of collection and the total
number of flies caught per day was recorded. The flies captured were packed in a cold box
containing ice packs to stop further development of microfilariae in the flies and then transported
to the laboratory for dissection (Opoku, 2000; Opara et al., 2005; Opara et al., 2008).
2.3 Dissection of the Blackflies
The flies captured were anaesthetized with chloroform and dissected fresh to determine their
physiological age (parity) (Atting et al., 2005). Before dissection for parity, each fly was placed
on a glass slide in normal saline solution (Plate 4) (Ibo and Braide, 1987; Renz and Wenk, 1987;
Mafuyai et al., 1997; Opoku, 2000; Okon et al., 2002; Atting et al., 2005).
The ovaries of the dissected flies were stretched and classified as parous or nulliparous after
observing other characters like absence or presence of fat bodies and the colour of malpighian
tubules. Flies were recorded as nulliparous if they have tightly coiled tracheal system and absence

28
of follicular relics (Corpora lutea). Nulliparous flies were discarded because they do not
transmit Onchocerca volvulus. On the other hand, flies were identified as parous indicating that
they had blood–fed and completed at least one gonotrophic cycle (Cupp and Collins, 1979;
Morkry, 1980; Hoc and Wilkes, 1995; Opara et al., 2008). The parous flies were further dissected
minutely to search for larvae of Onchocerca volvulus. In the second stage of dissection, the parous
flies were divided into head, thorax and abdomen. Each division was dissected by teasing it apart
in a normal saline using dissecting needles and dissecting microscope, and searched carefully for
O. volvulus larvae (Plate 5) (Nelson and Pester, 1962; Opoku, 2000; Opara et al., 2005; Opara et
al., 2008).
2.4 Calculation of Transmission Indices
The foci were on monthly population density of Simulium damnosum complex in the study site,
the daily biting rate, monthly biting rate, infective biting rate, the percentage of parous flies in the
total dissected (proportion of parous flies), and monthly transmission potential of Simulium
damnosum complex.
(a) Monthly Population Density: The monthly population density also called the biting density
of flies (Opara et al., 2005) was calculated through the fly per man hour (F.M.H) using the
formula: .hoursin x timedays catching ofNumber
month ain caught flies ofNumber FMH
(Crosskey, 1958; Davies et al., 1962; Gemade and Dipeolu, 1983)
(b) Daily Biting Rate (DBR)
The Daily Biting Rate (DBR) was calculated by using the formula
days catching ofNumber
month ain caught flies ofnumber TotalDBR
The Daily Parous Biting Rate was calculated by using the formula
days catching ofNumber
month ain caught flies parous ofnumber TotalDPBR
It is expressed as flies per person per day.

29
(c) Monthly Biting Rate (MBR)
The monthly biting rate (MBR1) was computed by using the formula:
days. catching ofNumber
month in the days ofnumber month ain caught flies ofNumber MBR1
(Walsh et al.,
1978; Grillet et al., 2001; Py-Daniel and Medeiros, 2004).
The Monthly Parous Biting Rate (MPBR) was calculated using the formula
MBR2 = days catching ofNumber
month thein days ofnumber caught flies parous ofnumber
(Py-Daniel and Medeiros, 2004). It is expressed as flies/person/month.
(d) Monthly Transmission Potential (MTP)
This index estimates the number of L3 (infective stage) that can be transmitted to a person
exposed to the vector during a one month period (Duke, 1968; Py-Daniel and Medeiros, 2004).
It is given by the formula:
xMTP workeddays ofNumber
larvaeinfectiveofnumberxmonththeindaysofNumber
dissected flies of No.
caught flies of No.
dissected. blackflies ofnumber Total
fliesblackby larvae stage L3 ofNumber x MBR 1 (Duke, 1968; Walsh et al., 1978;
Opoku, 2000; Py–Daniel and Medeiros, 2004). It is expressed as L3/person/month.
(e) Proportion of Parous flies
This represents the percentage of parous flies in the total dissected. It is given by the
dissected flies ofnumber Total
flies parous ofNumber :formular × 100
It is expressed as a percentage

30
2.5 The Diurnal Biting Pattern
A graph of the mean number of flies caught per hour against time in hour gives the diurnal biting
pattern of the flies for the different months in the study area.
2.6 Statistical Analysis. The data collected were subjected to analysis of variance (ANOVA). F-
LSD was used to compare the biting peaks for the different months and also to establish if there
were significant differences among the transmission indices for the different months.

31
CHAPTER THREE
RESULTS
3.1: Relative abundance of flies in Adani
A total of 548 Simulium damnosum complex were caught and dissected. Of this number, 248 were
caught in the rainy season (August, September and October) while 300 were caught in the dry
season (November, December and January). The monthly variation in the relative abundance of S.
damnosum complex at the site is shown in Figure 6. The highest number of flies was caught in
January (243) while the least number was caught in December (21). There was an increase in
population of flies from August to September and thereafter, there was a progressive decrease in
the number of flies from September to December and a sudden rise reaching a maximum in
January. The monthly population density of the flies ranged from 0.5 FMH in December to 5.5
FMH in January. The monthly catches between September and October; and between December
and January were significantly different (P < 0.05). However, the monthly catches between
August and September; October and November; and November and December were not
significantly different (P > 0.05).

32
0
50
100
150
200
250
300
August September October Novermber December January
Rela
tiv
e a
bu
nd
an
ce o
f f
lies
0
50
100
150
200
250
300
350
400
450
Mean
mo
nth
ly R
ain
fall
(m
m)
No of flies
Rainfall
Fig. 5: Relative abundance of S. damnosum complex and mean monthly rainfall in Adani
from August 2010 to January 2011.

33
The rainfall data was supplied by the Centre for Basic Space Science, University of Nigeria,
Nsukka.
3.2: The diurnal biting pattern of flies in Adani
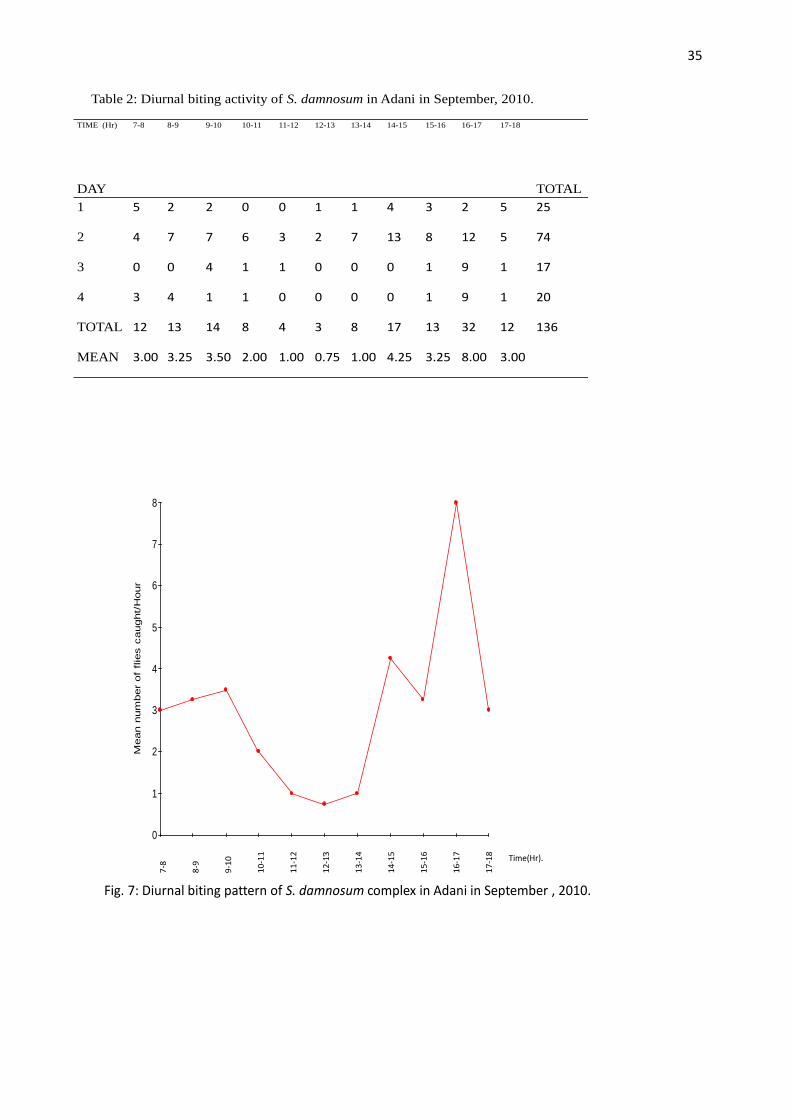
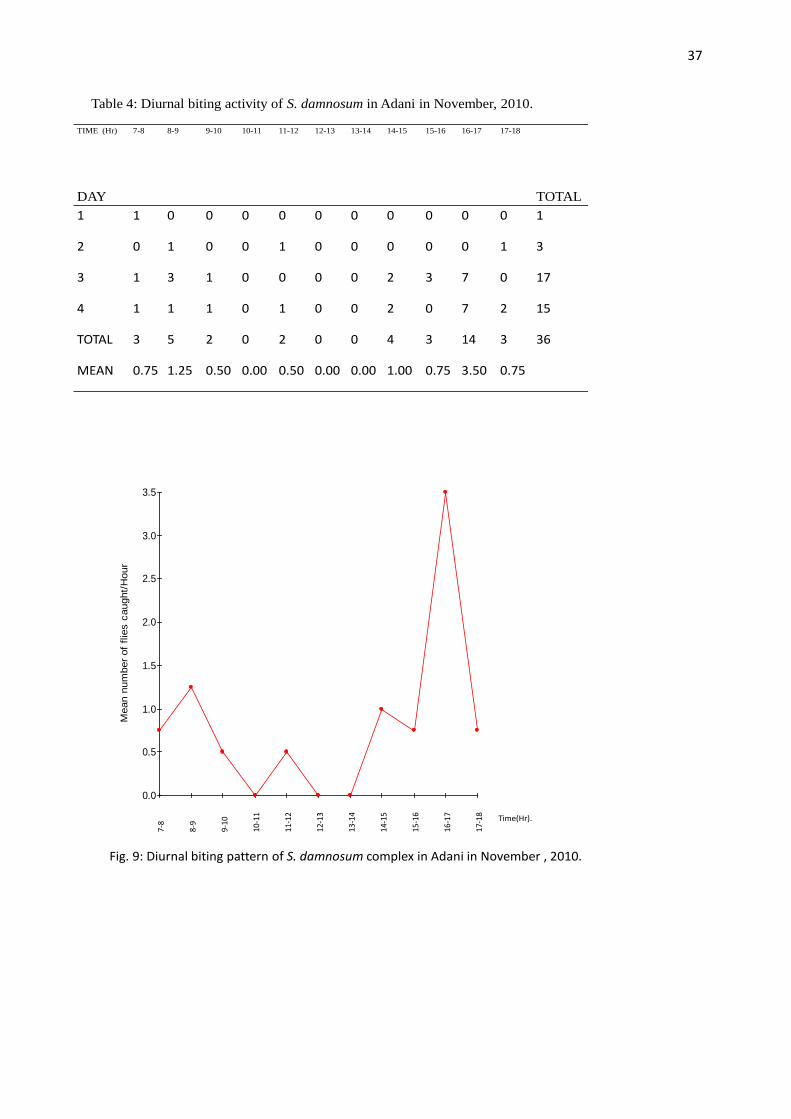
The diurnal biting patterns of S. damnosum complex in Adani showed a bimodal peak of activity
with the evening peak being higher than the morning peak (Fig. 6, 7, 8, 10, and 11) except in
October when the morning peak was higher than the evening peak (Fig. 9). The peaks were
separated by hours of low biting intensity. The morning peaks were observed between 7.00 am –
9.00 am in October, 8.00 am – 9.00 am in August and November, and 9.00 am – 10.00 am in
September, December and January. The evening peaks were observed between 5.00 pm – 6.00 pm
in August, December and January while in November and September it occurred between 4.00
pm – 5.00 pm. However, in October the evening peak was observed between 4.00 pm – 6.00 pm.
A paired t-test treatment of the mean biting peaks in all the months showed that the morning and
evening biting peaks were not significantly different (P > 0.05).

34
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 I 3 1 0 0 0 0 0 2 4 5 16
2 2 1 2 1 0 0 0 1 1 6 7 21
3 0 2 1 0 0 0 0 0 3 5 6 17
4 1 5 0 1 0 0 0 0 2 4 2 15
TOTAL 4 11 4 2 0 0 0 1 8 19 20 69
MEAN 1.00 2.75 1.00 0.5.0 0.00 0.00 0.00 0.25 2.00 4.75 5.00
Table 1: Diurnal biting activity of S. damnosum in Adani in August, 2010.
7-8
8-9
9-1
0
10
-11
11
-12
12
-13
13
-14
14
-15
15
-16
16
-17
17
-18
Fig. 6: Diurnal biting pattern of S. damnosum complex in Adani in August, 2010.
Time(Hr).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
Mean n
um
ber
of
flie
s c
aught/
Hour
.
35
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 5 2 2 0 0 1 1 4 3 2 5 25
2 4 7 7 6 3 2 7 13 8 12 5 74
3 0 0 4 1 1 0 0 0 1 9 1 17
4 3 4 1 1 0 0 0 0 1 9 1 20
TOTAL 12 13 14 8 4 3 8 17 13 32 12 136
MEAN 3.00 3.25 3.50 2.00 1.00 0.75 1.00 4.25 3.25 8.00 3.00
Table 2: Diurnal biting activity of S. damnosum in Adani in September, 2010.
7-8
8-9
9-1
0
10
-11
11
-12
12
-13
13
-14
14
-15
15
-16
16
-17
17
-18
Fig. 7: Diurnal biting pattern of S. damnosum complex in Adani in September , 2010.
Time(Hr).
0
1
2
3
4
5
6
7
8
Mean n
um
ber
of
flie
s c
aught/
Hour
.

36
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 6 4 0 0 0 0 1 4 4 1 4 24
2 1 1 1 0 0 0 0 0 0 1 0 4
3 1 2 2 0 0 0 0 0 1 1 0 7
4 1 2 2 0 0 0 0 0 0 2 1 8
TOTAL 9 9 5 0 0 0 1 4 5 5 5 43
MEAN 2.25 2.25 1.25 0.00 0.00 0.00 0.25 1.00 1.25 1.25 1.25
Table 3: Diurnal biting activity of S. damnosum in Adani in October, 2010.
7-8
8-9
9-1
0
10
-11
11
-12
12
-13
13
-14
14
-15
15
-16
16
-17
17
-18
Fig. 8: Diurnal biting pattern of S. damnosum complex in Adani in October , 2010.
Time(Hr).
0.0
0.5
1.0
1.5
2.0
2.5
Mean n
um
ber
of
flie
s c
aught/
Hour
.
37
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 1 0 0 0 0 0 0 0 0 0 0 1
2 0 1 0 0 1 0 0 0 0 0 1 3
3 1 3 1 0 0 0 0 2 3 7 0 17
4 1 1 1 0 1 0 0 2 0 7 2 15
TOTAL 3 5 2 0 2 0 0 4 3 14 3 36
MEAN 0.75 1.25 0.50 0.00 0.50 0.00 0.00 1.00 0.75 3.50 0.75
Table 4: Diurnal biting activity of S. damnosum in Adani in November, 2010.
7-8
8-9
9-10
10-1
1
11-1
2
12-1
3
13-1
4
14-1
5
15-1
6
16-1
7
17-1
8
Fig. 9: Diurnal biting pattern of S. damnosum complex in Adani in November , 2010.
Time(Hr).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Mean n
um
ber
of
flie
s c
aught/
Hour
.

38
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 0 1 0 0 0 0 0 0 1 1 0 3
2 0 0 1 0 0 0 0 0 0 0 4 5
3 1 1 1 0 0 0 0 0 1 0 3 7
4 0 0 1 0 0 0 0 0 0 5 0 6
TOTAL 1 2 3 0 0 0 0 0 2 6 7 21
MEAN 0.25 0.50 0.75 0.00 0.00 0.00 0.00 0.00 0.50 1.50 1.75
Table 5: Diurnal biting activity of S. damnosum in Adani in December, 2010.
7-8
8-9
9-1
0
10
-11
11
-12
12
-13
13
-14
14
-15
15
-16
16
-17
17
-18
Fig. 10: Diurnal biting pattern of S. damnosum complex in Adani in December , 2010.
Time(Hr).
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
Mean num
ber of flies caught/H
our
.

39
TIME (Hr) 7-8 8-9 9-10 10-11 11-12 12-13 13-14 14-15 15-16 16-17 17-18
DAY TOTAL
1 0 2 0 3 1 0 0 0 3 24 33 66
2 0 0 7 0 1 0 0 0 2 25 29 64
3 2 3 4 4 7 1 0 1 0 6 23 51
4 3 15 21 3 2 0 0 0 0 0 18 62
TOTAL 5 20 32 10 11 1 0 1 5 55 103 243
MEAN 1.25 5.00 8.00 2.50 2.75 0.25 0.00 0.25 1.25 13.75 25.75
Table 6: Diurnal biting activity of S. damnosum in Adani in January, 2010.
7-8
8-9
9-10
10-1
1
11-1
2
12-1
3
13-1
4
14-1
5
15-1
6
16-1
7
17-1
8
Fig. 11: Diurnal biting pattern of S. damnosum complex in Adani in January , 2011.
Time(Hr).
0
5
10
15
20
25
30
Mea
n nu
mbe
r of
flie
s ca
ught
/Hou
r
.

40
3.3: Transmission indices of onchocerciasis in Adani.
The Daily Biting Rate (DBR) varied from 5.3 flies per man per day in December to 60.8 flies per
man per day in January. The Monthly Biting Rate (MBR) was highest in January with 1883.3 flies
per man per month while the least Monthly Biting Rate (MBR) was recorded in December with
162.8 bites per man per month.
The parous flies were 133(24.3%) while the nulliparous flies were 415(75.7%) during the study
period. The infection and infectivity rate during the study was zero (Table 7). The
biting/population densities, biting rates, infection and infectivity rate, proportion of parous flies
and transmission potential of Onchocerca volvulus by S. damnosum during the study are as shown
in Table 7. Flies caught in August and September were all nulliparous. A paired t-test treatment of
the parity rates showed that there were significantly more parous flies than nulliparous flies in
October and November (P < 0.05) but no significance differences in December and January (P >
0.05).
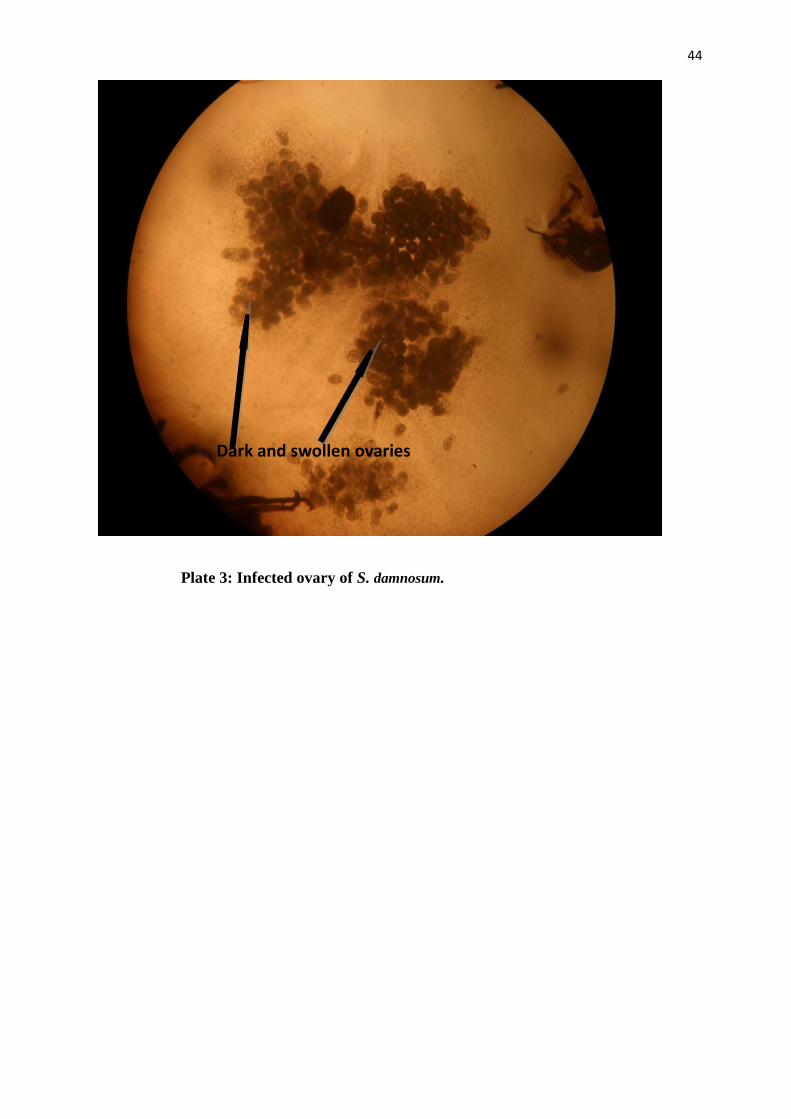
Infections that are not of O. volvulus were recorded as ‘Other infections’ (Table 7). Two of such
infections were seen in September and October (Plate 3). These are probably bacterial infections
that affect flies in the larval stage and carried to the adult.

41
Table 7: Population density and Transmission indices of S. damnosum complex in Adani,
Uzo-Uwani Local Government Area of Enugu State
August
2010
September
2010
October
2010
November
2010
December
2010
January
2011
Person days worked 4 4 4 4 4 4
Total number of flies caught
and dissected
69 136 43 36 21 243
No(%) of parous flies 0 (0) 0 (0) 13 (30.2) 9 (25) 10 (47.6) 101 (41.6)
No(%) of nulliparous flies 69 (100) 136 (100) 30 (69.8) 27 (75) 11 (52.4) 142 (58.4)
Total No(%) of flies infected
with O. volvulus
0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
Flies(%) with L1 and L2 of
O. volvulus
0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
Flies(%) with L3 of
O. volvulus
0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
Biting density of flies 1.6 3.1 1.0 0.8 0.5 5.5
Daily Biting Rate(DBR) 17.3 34 10.8 9 5.3 60.8
Daily Parous Biting
Rate(DPBR)
0 0 3.3 2.3 2.5 25.3
Monthly Biting Rate(MBR1) 534.8 1020 333.3 270 162.8 1883.3
Monthly Parous Biting
Rate(MBR2)
0 0 100.8 67.5 77.5 1100.5
Monthly Transmission
Potential(MTP)
0 0 0 0 0 0
Other infections 0 0 2 2 0 0

42
Plate 1: Malpighian tubules exposed for determining parity.
Malpighian tubules
43
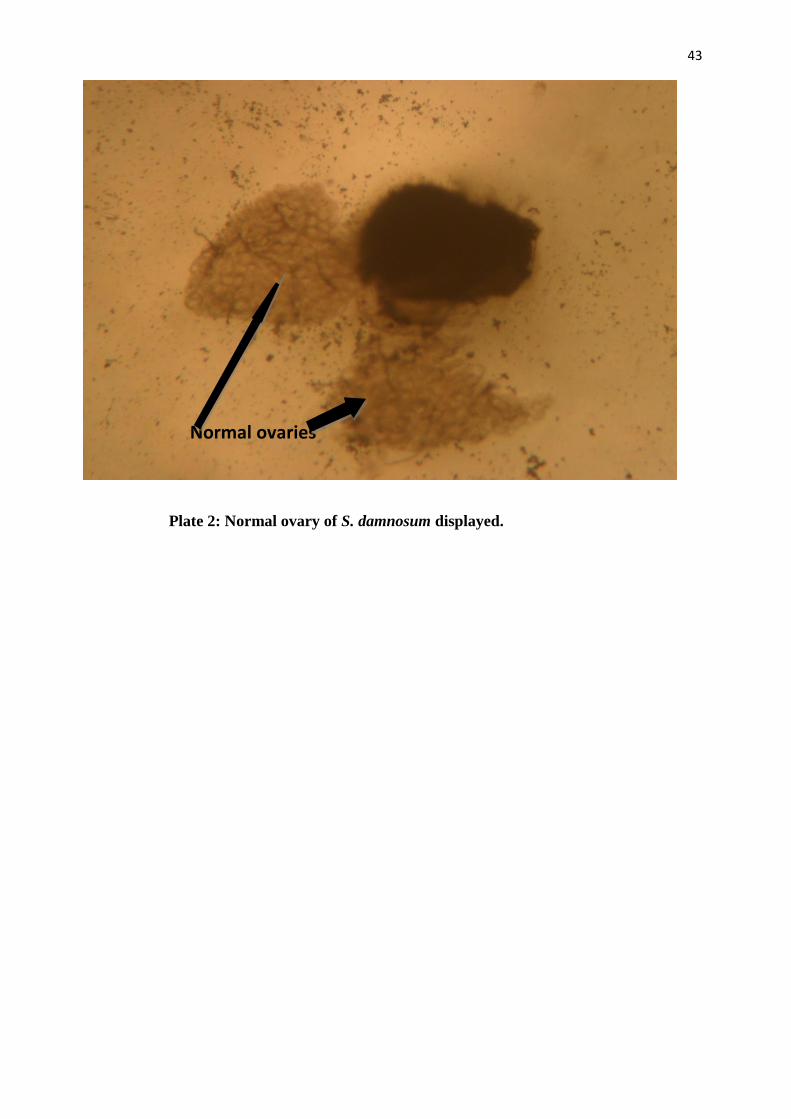
Plate 2: Normal ovary of S. damnosum displayed.
Normal ovaries
44
Plate 3: Infected ovary of S. damnosum.
Dark and swollen ovaries

45
CHAPTER FOUR
DISCUSSION
4.1 Discussion
The study showed a progressive decrease in fly relative abundance from September to December.
This is in agreement with the findings of Atting et al. (2005), Opara et al. (2005) and Opara et al.
(2008). The study recorded the highest number of flies in January. This agrees with the findings
of Ubachukwu and Anya (2005) who observed similar increase in fly population in Nkpologu,
Uzo-Uwani Local Government Area of Enugu State during the dry season with harmmattan but
contradicts the findings of Opara et al. (2005) who observed the highest number of flies in
September in Cross River State, Nigeria, Opara et al. (2008) who observed the highest number of
flies in September in Akwa Ibom State, Nigeria, Atting et al. (2005) who recorded the highest
population density of flies in August in Cross River State, Nigeria and that of Adeleke et al.
(2010) who observed the highest number of flies in September and October in Osun State,
Nigeria. The fly population in Adani was higher in the dry season. This agrees with Ubachukwu
and Anya (2005) but contradicts the findings of Crosskey (1990) and Atting et al. (2005) who
reported that the biting nuisance of the flies was greater in the rainy season. Opara et al. (2005)
also recorded a greater number of flies in the rainy season in Cross River State, Nigeria which
contradicts the present observation in Adani.
The observed decrease in fly population in Adani from September to October may be attributed to
continuous flooding of the river during the peak rainy season (August, September and October)
(Fig. 5) which may have resulted in dislodging and washing away of immature stages of
Simulium flies to long distances away from Adani. These may lead to a decrease in the population
of adult flies emerging. According to Opara et al. (2005), stormy weather may be a factor because
it may have washed away most of the breeding sites, thus resulting in a smaller fly population.
Also Atting et al. (2005) noted that the Kwa fall in Akwa Ibom southern Nigeria was completely
flooded in the month of October 1999. He argued that the flooding brought about a reduced

46
productivity of existing breeding sites during the rainy season hence decreasing the fly
population in the month. Also, Renz (1987a) reported that few flies were caught at Mayo Galke
causeway in northern Cameroon when the flow of water was more than 100m3sec
-1 at the height
of the rainy season. At the River Vina, near the Tonboro, he observed that the biting densities
were at their lowest level during the rainy season in October 1976.
The observed reduction in the abundance of flies during the months of November and December
may be attributed to the extensive bush burning that occurred all through the two months that
could lead to massive death and migration of the adult flies. Taye et al. (2000) attributed the
decline in population of S. neavei and S. ethiopiense in Ethiopia to recent degradation of
vegetation cover in the vicinity of their breeding sites. Also, McMahon et al. (1958) noted that
deforestation was the sole means used to eradicate S. neavei s.s. and onchocerciasis from some
foci in Kenya.
Changes in the distribution patterns of blackflies occur annually in association with the
pronounced seasonal climatic variations manifested in West Africa by the dry and wet seasons.
The seasonal changes is accompanied by shifts in wind direction and by the drying out or flooding
of rivers which provide the S. damnosum breeding sites (Boakye et al., 1998). Biting activity can
be highly seasonal in tropical regions with prolonged dry season, which eliminates or at least
severely reduces the available breeding habitats. In West Africa, biting activity of S. damnosum
complex ceases for several months in the long dry season of northern savannas when rivers stop
flowing or are reduced to trickles and resumes only when rivers run again in the rains. On the
other hand, where the climate is almost uniform throughout the year and rivers run perennially,
biting activity occur year-round in association with continuous breeding (Crosskey, 1990). The
observation of the highest fly densities during the harmattan season may lie, in addition to local
flies, of migrating savanna flies carried down into the area with the help of the north-south
harmattan winds blowing from the Sahara to the coasts (Boakye, 1999). Possible addition of these
migrating flies to the local flies during the harmattan season would most likely be responsible for

47
swelling the biting populations of the blackflies within this season (Ubachukwu and Anya,
2005). In Adani, Obina River flows perennially, thus, S. damnosum complex is assumed to breed
throughout the year. Hence, the observed sudden increase in fly population density in the month
of January 2011 (harmattan) may be attributed to flies that were carried by wind from other
breeding sites in addition to local production of flies plus flies that migrated from other breeding
sites where the river/stream dried up.
The diurnal biting pattern of the flies showed a bimodal peak of activity - a morning peak and a
more pronounced evening peak in all the months except in October where the morning peak was
higher than the evening peak. The morning peaks occurred between 7.00am - 10.00am and the
evening peaks occurred between 4:00pm - 6:00pm. These findings are in consonance with Opoku
(2000), Ubachukwu and Anya (2001a), Opara et al. (2005) and Opara et al. (2008) but contradicts
that of Adeleke et al. (2010) who observed three biting peaks in two different sampling points in
south western Nigeria and Barbiero and Trpis (1984) who observed a unimodal biting peak
activity pattern in Liberia. The cause of biting activity peaks is still poorly understood, but it has
been suggested that an innate clock rhythm may be involved (Crosskey, 1990). According to him
blackflies do not suck blood daily; hence, the biting cycle may be loosely described as a circadian
rhythm which by definition entails a biological rhythm on a one-day periodicity (Lincoln et al.,
1982). Diurnal variations in the biting density have been related to variations in the temperature
(Leberre, 1966) and humidity (Hausermann, 1969) or to the intensity of light (Kaneko et al.,
1973). The variation in the biting activity of the flies delimits the hours of maximum or minimum
danger of transmission of onchocerciasis (Renz, 1987a). However, Nwoke, (1988), Porter and
Collins (1988), Adewale et al. (1999) and Ubachukwu and Anya (2001a) reported that the biting
activities of S. damnosum s. l. is greatly influenced by illumination and temperature. It is possible
that the bimodal peaks observed might be due to decreased illumination and temperature during
these peak periods which is characteristics of climatic conditions of south-eastern Nigeria. The
diurnal biting cycle of the flies may have epidemiologic implications since their biting peaks
correspond to periods of peak human outdoor activities.

48
The relative abundance and biting patterns of the flies at Adani revealed that the control of
Simulium damnosum complex with regard to their relative population density and distribution
should be targeted at certain months and times of the day. For instance, larviciding will not be
feasible in the months of August, September and October due to heavy rainfall and flooding of
Obina River during these months. On the other hand larviciding and aerial spraying of insecticides
in January specifically in the morning and evening will go a long way in eradicating the flies and
reduction in their nuisance bites and transmission of onchocerciasis.
The biting density of flies in Adani ranged from 0.5 FMH in December to 5.5 FMH in January.
The daily biting rate ranged from 5.3 flies/man/day in December to 60.8flies/man/day in January.
The monthly biting rate of the flies ranged from 162.8flies/man/month in December to
1883.3flies/man/month in January. The biting density of flies recorded in Adani is far below the
maximum range given by Crosskey (1990) who stated that in Africa the highest biting rate of S.
damnosum complex are usually not above 30 – 60FMH in Savanna and 200FMH in forest areas,
whereas biting densities of 100 – 1000FMH are commonplace with S. ochraceum complex in
Guatemala and 300 – 450FMH typical of S. oyapockense in the Amazon. Opara et al. (2008)
recorded biting densities of 27.7FMH, 35.8FMH and 33.5FMH in three different sampling points
in Akwa Ibom State, Nigeria. Opara et al. (2005) also reported 0.5FMH in December and
21.6FMH in August in Cross River State, Nigeria. In Uzo-Uwani L.G.A. of Enugu State, the peak
and average biting rates according to Ubachukwu and Anya (2005) were 25FMH and < 3FMH
respectively during the harmattan season and 4FMH and < 1FMH respectively during the dry
season without harmattan. During the early rainy season the peak biting rate was 12FMH while
the average biting rate was about 2FMH. During the late rain, the peak biting rate was 6 FMH
while the average biting rate was 1FMH (Ubachukwu and Anya, 2005).
The monthly biting rate (MBR) in Adani compares with that of other investigators. Opara et al.
(2005) reported a range of 217flies/man/month in January to 7,354flies/man/month in August at
Agbokim Waterfall and 46.5flies/man/month to 6,893flies/man/month at Afi River both in Cross

49
River State, Nigeria. Opara et al. (2008) recorded a range of 201flies/man/month in December
to 1,286flies/man/month in October at Idomido, 317flies/man/month in December to
1,470flies/man/month in September at Obio Camp and 232flies/man/month in December to
1,387flies/man/month in September at Ikot Adaha, all in Akwa Ibom State, Nigeria. Adeleke et al.
(2010) observed a range of 47flies/man/month in January to 3,075flies/man/month in September
at Osun Budepo, 0flies/man/month in December and January to 450flies/man/month in September
at Osun Ogbere, and 28 flies/man/month in February to 2,250 flies/man/month in June at Osun
Eleja, all along Osun River in South-western Nigeria. At Mayo Galke causeway, Renz (1987a)
reported a bimodal biting pattern with peak Monthly Biting Rate (MBR) in the early rainy season
during the months of May to August (MBR 15273 in July 1976; 10256 in June 1977 and 10112 in
May 1978) and with a second peak in the cold dry season from December to January (16890 in
January 1977; 4624 in December 1978 and 10155 in January 1979). However, at Mayo Galke
(causeway a) in the Sudan-savanna of North Cameroon, an Annual Biting Rate (ABR) of as high
as 83800flies/man/year was recorded in 1976 according to Renz (1987b).
Flies caught during the dry season had higher parous rate than those caught in the rainy season.
This is in agreement with the findings of Opara et al. (2005), Opara et al. (2008) and Adeleke et
al. (2010) who recorded greater percentages of parous flies in the dry season. A high proportion
of parous flies may be an indication of ageing of local blackfly populations or the presence of
migratory female flies (Opara et al., 2005).
The result of the dissection showed that the majority of the flies caught were nulliparous and they
accounted for 75.7% of the total. This observation is consistent with the report of Mafiana (1988),
and Adeleke et al. (2010) who recorded 53.9%, 57.86% and 59.58% nulliparous flies in three
communities along Osun River in South Western Nigeria and Atting et al. (2005) who recorded
96.83% nulliparous flies in a forest area of Cross River State, Nigeria but contrary to the reports
of Usip et al. (1999), Okolo et al. (2004), Opara et al. (2005) who recorded 84.17% parous flies in
Cross River State, Nigeria, Opara et al. (2008) who recorded 86%, 89.6% and 88.3% parous flies

50
at three different sites in Akwa Ibom State, Nigeria and Taye et al.(2000) who recorded 74.7%
parous flies in south western Ethiopia. The high proportion of nulliparous flies in Adani may be a
reflection of local production of flies as reported by Opara et al. (2008).
The six months study did not record any Onchocerca volvulus infection in the flies dissected.
Hence, the infectivity rates and transmission of parasites in Adani was zero during the study
period. This finding is in consonance with Adeleke et al. (2010) who worked from February 2008
to June 2009 in three communities along Osun River, south western Nigeria. For one year of
collection and dissection of flies, no parasite was found and according to him there has been no
record of ivermectin distribution in those communities. In Adani, the inhabitants confirmed that
there have been many years (1996 – Date) of uninterrupted mass distribution of ivermectin till
date. According to the respondent the inhabitants were given 4 tablets annually but later they were
given 4 tablets biannually. That could be the reason for the absence of O. volvulus L1, L2 and L3
in the flies and suggests that transmission has been temporarily interrupted in Adani. The
observations may indicate low level of onchocerciasis transmission in Adani or low level of
microfilarial load in human reservoirs. This sort of observation according to Remme et al. (1990)
is expected in areas where the distribution of ivermectin is being carried out. Recent reports from
endemic communities in Mali and Senegal have indicated the possibility of elimination of
onchocerciasis after several treatments with ivermectin (Diawara et al., 2009). According to them,
15 to 17 years of annual or six-monthly ivermectin treatment was carried out in some
communities in these countries. Thereafter, a total of 157,500 blackflies were collected and
analyzed for the presence of Onchocerca volvulus larvae using a specific DNA probe. It was
observed that vector infectivity rates were all below 0.5 infective flies per 1,000 flies. Except for a
subsection of one focus, all infection and transmission indicators were below postulated
thresholds for elimination. This study provided the first empirical evidence that elimination of
onchocerciasis with ivermectin treatment is feasible in some endemic foci in Africa. Borsboom et
al. (2003) reported that ivermectin treatment has been very successful in eliminating
onchocerciasis as a public health problem. According to him, ivermectin treatment sharply

51
reduced prevalence and intensity of infection of Onchocerca volvulus in all of the river basins
studied in West Africa. Boussinesq et al. (1997) reported that repeated treatments with ivermectin
brought about a notable reduction in the transmission of Onchocerca volvulus in northern
Cameroon. According to him, after 8 annual treatments, the intensity and prevalence of
microfilariae decreased by more than 90% compared with the pre-treatment values. Cupp et al.
(1986) reported that treatment with ivermectin diminished rate of infectiousness after 6 months.
He suggested that ivermectin could be effective in interrupting transmission of Onchocerca
volvulus for epidemiological important period of time. Rodriguez-Perez and Reyes-Villanueva
(1994) observed that ivermectin mass treatment produced a reduction of 30% in the transmission
potential of Onchocerca volvulus in southern Mexico. Osei-Atweneboana et al. (2007) observed
that ivermectin treatment in Ghana showed 100% clearance of microfilariae in more than 99% of
the treated individuals after 30 days, but there was evidence of recrudescence after 90 days
resulting from reproduction by adult worms. Remme et al. (1989) reported that 3 days of
ivermectin treatment in Asubende, Ghana reduced the mean skin microfilarial load in treated
persons by more than 96% two months after treatment. Guderian et al. (1997) also observed that
the microfilarial density in the treated group declined from 19.3 to 0mf/mg after 7 years of
biannual treatment with ivermectin in the Rio Santiago focus in Ecuador. The rate of infection of
blackflies declined from 1.1% - 0.08% which according to him is below the vectorial capacity of
the Simulium vector, and no new nodules were detected after the treatment and no children under
5 years became infected over the observation period. Hence, from the foregoing it is possible to
eradicate onchocerciasis or prevent/interrupt the transmission of O. volvulus by mass distribution
of ivermectin in endemic areas and that could be the situation in Adani.
Atting et al. (2005) reported zero transmission of O. volvulus in the month of January in Cross
River State, Nigeria and concluded that the problem of onchocerciasis in the study area was more
of a biting nuisance by the vectors. Opara et al. (2005) also reported zero transmission of O.
volvulus in the months of January, February, March, November and December at Agbokim
Waterfalls and Afi River in Cross River State, Nigeria after five years of ivermectin intervention.

52
Opara et al. (2008) reported zero transmission of O. volvulus during the dry periods –
November, December, January, February and March, and the early rainy periods – April and May
in Akwa Ibom State, Nigeria. Opoku (2000) also recorded zero infection and transmission of O.
volvulus in February in River Birim Catchment, Ghana. At Gandi (Village s) in the Sudan-
savanna of North Cameroon, an Annual Transmission Potential (ATP) of zero was reported by
Renz (1987b) with an Annual Biting Rate (ABR) of 2100 flies/man/year. An ATP of zero was
also recorded at Tchollire (village x) and Tonboro (village v and tributary u) in the Sudan-savanna
of North Cameroon (Renz, 1987b).
An annual transmission potential of zero has also been reported by Porter and Brandling-Bennet
(1988) and Brandling-Bennet et al. (1981) in Guatemala. Beltran et al. (1979) also reported that
mass treatment with ivermectin efficiently controlled and at least temporarily interrupted
transmission of O. volvulus by blackflies vectors in Liberia. He observed that the number of
infected flies with developing larvae (L1, L2 L3, stages) of O. volvulus in treated areas was reduced
by 93.4-95%; the number of infective flies with L3 larvae was reduced by 81.7-89.3%. Monthly
transmission potentials showed a similar decrease from 22.9 to 5.8 (74.6% reduction) in the
treated areas.
In addition, the relatively high number of nulliparous flies observed in the overall flies dissected
in the present study could also be a contributory factor since only aged flies have chances of
transmitting the parasite. Thus, as at now, the flies in Adani are more of biting nuisance than
disease vectors. The biting nuisances of the flies are also of socio-economic importance. Their
painful bites could serve as mechanical entry for viruses, bacteria, protozoan and nematodes
which the flies may carry on their bodies. The crawling effect of their movement in human hosts
could also constitute intolerable nuisance and distraction from work therefore reducing
productivity (Usip et al., 2008). Ubachukwu and Anya (2001b) also observed that farmers in
Nkpologu in Uzo-Uwani Local Government Area of Enugu State, Nigeria lost N6, 500.00 per
man per season due to the bites of black flies.

53
During the study, it was observed that actual man-fly contact was very low. This is because the
flies have a patchy distribution in Adani being encountered only in some farms and near
farmlands which are far away from dwellings. In addition, most compounds have wells where
they fetch water. Hence, inhabitants do not go to the River to fetch water but few who live near
the River do otherwise. Also, at the sampling point located approximately 300 metres from the
River, no anthropogenic activities was found in the River and its environs throughout the study
except for the Fulanis that took their cattle to the River to drink water during the dry season.
Hence, as Crosskey (1990) rightly observed, the biting rates inevitably exaggerates the level of
man-fly contact under natural conditions because people going about their ordinary activities in an
onchocerciasis area are unlikely to be contacted anywhere as often as the bait individuals who are
continuously exposed for a prolonged time with the deliberate intention of attracting flies.
4.2 Conclusion and recommendations
Although old infections (individuals harbouring adult O. volvulus) may still be found in Adani,
the transmission appears to have been temporarily halted by ivermectin distribution and the flies
are mere nuisance biters. Hence, no new infection may be recorded in Adani at present. I
recommend that vector control such as periodic aerial spraying of chemical pesticides and
larviciding with chlorpyrifos methyl, Bacillus thuringiensis, use of repellents such as deet and 2-
ethylhexanediol and use of pyrethroid impregnated clothing should be encouraged in Adani. Also
river channelling improvement to clean and deepen Obina River with the removal of stones, logs,
and other obstruction that cause ripples attractive to blackfly larvae, the construction of dams and
spillways to provide clear and unobstructed flow of the river should be encouraged. I recommend
that ivermectin distribution should continue in the area to avoid re-introduction of O. volvulus
parasites by flies migrating from other onchocerciasis endemic zones.

54
REFERENCES
Adeleke, M. A., Mafiana, C. F., Sam-Wobo, S. O., Olatunde, O. G., Ekpo, U. F., Akinwale, O. P.
and Toe, L. (2010). Biting behaviours of Simulium damnosum complex and Onchocerca
volvulus infection along the Osun River, Southwest Nigeria. Parasite and Vectors, 3:93 -
101.
Adewale, B., Mafe, M. A. and Oyerinde, J. P. O. (1999). Infectivity and transmission dynamics of
Simulium damnosum s. l. around Owena Dam (Ondo State). West African Journal of
Medicine, 18 (4): 257 - 260.
Adeyeba, O. A. and Adegoke, A. A. (2002). Onchocerciasis in communities in forest zone, south
west Nigeria: Prevalence and diagnostic method for rapid assessment. African Journal of
Clinical and Experimental Microbiology, 3(1): 29 – 32.
Atting, A. I., Ejezie, G. C., Braide, E. I., Opara, K. N. and Ekwe, A. (2005). Seasonal variations
in human onchocerciasis transmission by blackflies (Simulium damnosum s.l.) in a forest
area of Cross River State, Nigeria. African Journal of Applied Zoology and Environmental
Biology, 7: 14 - 18
Barbiero, V. K. and Trpis, M. (1984). Transmission of onchocerciasis by local blackflies on the
firestone rubber plantation harbel, Liberia. American Journal of Tropical Medicine and
Hygiene, 33:586 - 594.
Basile, K., Mather, F. J. and Barnett, L. C. (1995). Evaluation of alternate methods of rapid
assessment of endemicity of Onchocerca volvulus in communities in Southern Cameroon.
American Journal of Tropical Medicine and Hygiene, 53(3): 243 – 247.
Beltran, F., Gomez-Priego, A., Martinez-Chacon, J. F. and Ortega, M. I.
(1979). Perspectivas actuales para la vigilancia epidemiologica de la oncocercosis en
Mexico. Salud Publica Mexico, 21: 771 – 789.
Boakye, D. A. (1999). Insecticide resistance in the Simulium damnosum s.l. vectors of human
onchocerciasis: Distributional, Cytotaxonomic and Genetic Studies. Post-Doctoral Thesis,
University of Lieden, Netherlands.
Boakye, A. D., Back, C., Fiasorgbor, G. K., Sib, A. P. P. and Coulibay, Y. (1998). Sibling species
distribution of the Simulium damnosum complex in the West African Onchocerciasis
Control Programme area during the decade 1984-1993, following intensive larviciding since
1974. Medical and Veterinary Entomology,12: 345 – 358.
Boakye, D. A., Fokam, E., Ghansah, A., Amakye, J., Wilson, M. D. and Brown, C. A. (2009).
Cardiocladius oliffis (Diptera: Chironomidae) as a potential biological control agent against
Simulium squamosum (Diptera: Simuliidae).
http://www.parasitesandvectors.com/contents/2/1/20. Accessed 5th
January, 2011.
Bockarie, M., Kazura, J., Alexander, N., Dagoro, J., Bockarie, F., Perry, R. and Alpers, M.
(1996). Transmission dynamics of Wuchereria bancrofti in East Sepik Province, Papua,
New Guinea. American Journal of Tropical Medicine and Hygiene, 54: 577 - 581.
Borsboom, G. J. J. M., Boatin, B. A., Nagelkerke, N. J. D., Agoua, H., Akpobona, K. L. B., Alley,
E. W. S., Bissan, Y., Renz, A., Yameogo, L., Remme, J. H. F. and Habbema, J. D. (2003).
Impact of ivermectin on onchocerciasis transmission: Assessing the empirical evidence that
repeated ivermectin mass treatments may lead to elimination/eradication in West Africa.
Filarial Journal, 2: 8 - 16.

55
Boussinesq, M., Prodhon, J. and Chippaux, J. P. (1997). Onchocerca volvulus: Striking
decrease in transmission in Vina Valley (Cameroon) after eight annual large scale
ivermectin treatments. Transactions of the Royal Society of Tropical Medicine and Hygiene,
91(1): 82 – 86.
Brandling-Bennett, A. D., Anderson, J., Fuglsang, H. and Collins, R. (1981). Onchocerciasis in
Guatemala. Epidemiology in fincas with various intensities of infection. American Journal
of Tropical Medicine and Hygiene, 30: 970 – 981.
Crosskey, R. W. (1958). First result in the control of Simulium damnosum Theobald (Diptera:
Simuliidae) in Northern Nigeria. Bulletin of Entomological Research, 49: 715 - 735.
Crosskey, R. W. (1981). A review of Simulium damnosum s. l. and human onchocerciasis in
Nigeria with special reference to geographical distribution and development of a Nigerian
National Control Campaign. Tropenmed. Parasit., 32(1): 2 - 16.
Crosskey, R. W. (1990). The Natural History of Blackflies. First Edition. John Wiley and Sons,
New York.
Cupp, E. W. and Collins, R. C. (1979). The gonotrophic cycle of Simulium ochraceum. American
Journal of Tropical Medicine and Hygiene, 281: 422 - 426
Cupp, E. W., Bernado, M. J., Kiszewski, A. E., Collins, R. C., Taylor, H. R., Aziz, M. A. and
Greene, B. M. (1986). The effects of ivermectin on transmission of Onchocerca volvulus.
Science, 231(4739): 740 – 742.
Dadzie, K. Y., Bird, A. C., Awadzi, K., Schulzkey, H., Gillies, H. M. and Aziz, M. A. (1987).
Ocular findings in a double blind study of ivermectin versus diethylcarbamizine placebo in
the treatment of onchocerciasis. British Journa of Ophthalmology, 71:78 – 85.
Davies, J. B., Crosskey, R. W., Jonston, M. R. L. and Crosskey, M. E. (1962). The control of
Simulium damnosum at Abuja, Northern Nigeria (1955-1960). Bulletin of World Health
Organization, 27:491 - 510.
Diawara, L., Traore, M. O., Badji, L., Bissan, Y., Doumbia, K., Goita, S. F., Konate, L.,
Mounkoro, K., Sarr, M. D., Seck, A. F., Toe, L., Touree, S. and Remme, J. H. F. (2009).
Feasibility of onchocerciasis elimination with ivermectin treatment in endemic foci in
Africa: First evidence from studies in Mali and Senegal. Plos Neglected Tropical Diseases,
3 (7): 1 - 4.
Duke, B. O. L. (1968). Studies on factors influencing the transmission of onchocerciasis VI. The
infective biting potential of Simulium damnosum in different bioclimatic zones and its
influence on the transmission potential. Annals of Tropical Medicine and Parasitology, 63:
164 - 170.
Edungbola, L. D. (1991). Onchocerciasis control in Nigeria. Parasitology Today, 7(5): 97 - 99.
Etya’ale, M. E. (2001). Vision 2020: Update on onchocerciasis. Community Eye Health, 14(38):
19 – 21.
Ezedinma, C. (2011).Impact of trade on domestic rice production and the challenge of self
sufficiency in Nigeria. International Institute of Tropical Agriculture Ibadan, Nigeria.
http://www.warda.cgiar.org/workshop/RicePolicy/Chuma.E/Chuma.E.Nigeria.Paper.pdf
Accessed 5th January, 2011.

56
Fox, L. M. (2009). Onchocerciasis (River Blindness). Centers for Disease Control and
Prevention, Atlanta.
Gemade, E. I. I. and Dipeolu, O. O. (1983). Onchocerciasis in Benue State of Nigeria III: A study
of bionomics of Simulium damnosum (Theobald) complex at manor. International Journal
of Zoology, 10: 71 – 84.
Google (2010). Adani, Nigeria Page. http://www.fallingrain.com/world/NI/47/ Adani.html
Accessed 16th
September, 2010.
Grillet, M. E., Maria-Gloria, B., Vivas-Martinez, S., Nestor, V., Hortensia, F., Jose, C., Pablo, C.
and Carlos, B. (2001). Human onchocerciasis in the Amazonian Area of Southern
Venezuela: Spatial and temporal variations in biting and parity rates of Blackflies (Diptera:
Simuliidae) vectors. Journal of Medical Entomology, 38 (4): 520 - 530.
Guderian, R. H., Anselmi, M., Espinel, M., Mancero, T., Rivadeneira, G., Proano, R., Calvopina,
H. M., Vieira, J. C. and Cooper, P. J. (1997). Successful control of onchocerciasis with
community-based ivermectin distribution in the Rio Santiago focus in Ecuador. Tropical
Medicine and International Health, 2(10): 982 – 988.
Hausermann, W. (1969). On the biology of Simulium damnosum, Theobald, 1903, the main vector
of onchocerciasis in Mahenge Mountain,Ulange, Tanzania. Acta Tropica, 26: 29 – 69.
Hoc, T. Q. and Wilkes, T. J. (1995). Age determination in the blackfly Simulium woodi, a vector
of onchocerciasis in Tanzania. Medical and Veterinary Entomology, 9: 16 - 24.
Iboh, C. L. and Braide, E. I. (1987). The occurrence and distribution of blackflies (Simulium spp)
in Vakurr Clan of Obubra L.G.A, Cross River State, Nigeria. Ife Journal of Science, 2 (1-2):
1 – 5.
Igbinosa, I. B. (1990). Parasitic arthropods. In: 41 - 65, Ezigbo (ed). Parasitology for medical
students. New Frontier Publishing Limited, Lagos.
Jamnback, H. (1973). Recent developments in the control of black flies. Annual Review of
Entomology, 18: 281 – 304.
Kale, O. O. (1998). Onchocerciasis: the burden of disease. Annals o Tropical Medicine and
Parasitology, 92(1):101 - 115.
Kaneko, K., Satto, K. and Wonde, T. (1973). Observations on the diurnal rhythm of the biting
activity of Simulium damnosum in Omo-Gibe and Gojjeb rivers, south-west Ethiopia.
Japanese Journal of Sanitary Zoology, 24: 175 – 180.
Leberre, R. (1966). Contributions a`l`etude biologique et ecologique de Simulium damnosum
Theobald, 1903, (Diptera: Simuliidae). Memoires Orstom, 17: 204pp.
Lincoln, R. J., Boshall, G. A. and Clark, P. F. (1982). A dictionary of ecology, evolution and
systematic. Cambridge University Press, Cambridge, United Kingdom.
Mafiana, C. F. (1988). Studies on onchocerciasis and Simulium damnosum in Igbo-Uwa, Bendel
State, Nigeria. Ph.D Thesis University of Benin, Nigeria.
Mafuyai, H. B, Post, R. J., Molyneux, D. H. and Davies, D. H. (1997). First sibling species,
identification of Nigeria onchocerciasis vectors. Transactions of the Royal Society of
Tropical Medicine and Hygiene, 91: 90 – 91.
McMahon, J. P., Highton, R. B. and Goiny, H. (1958). The eradication of Simulium neavei from
Kenya. Bulletin of World Health Organization, 19: 75 – 76.

57
Metcalf, L. and Novak, R. J. (1994). Pest management strategies for insects affecting humans
and domestic animals. In: 587-628, Metcalf, R. L. and Luckmann, W. H. (Eds). Introduction
to Insect Pest Management. Third edition, Wile-Interscience Publication, New York.
Morkry, J. E. (1980). A method of estimating the age of field collected female Simulium
damnosum (Diptera: Simuliidae). Tropical Medical Parasitology, 31: 153 - 167.
Moyou-Somo, R., Eyong, P. A., Fobi, G., Dinya, J. S., Lafleur, C., Agnamey, P., Ngosso, A. and
Mpoudi, N. E. (1993). A study on onchocerciasis with severe skin and eye lesions in a
hyperendemic zone in the forest of South Western Cameroon. Clinical parasitologic and
entomologic findings. American Journal of Tropical Medicine and Hygiene, 48: 14 - 19.
Nayar, K. K., Ananthakrishnan, T. N. and David, B. V. (1992). General and Applied Entomology.
Ninth Edition Tata Mcgraw-Hill Publishing Company Limited, New Delhi.
Nelson, G. S. and Pester, F. R. N. (1962). The identification of infective filarial larvae. Bulletin of
World Health Organization, 27: 473 - 481.
Nwoke, B. E. B. (1986). Studies on the field epidemiology of human onchocerciasis on the Jos
Plateau, Nigeria. PhD Thesis, University of Jos, Nigeria.
Nwoke, B. E. B. (1988). Studies on the epidemiology of human onchocerciasis on the Jos Plateau,
Nigeria. The effects of climatic factors on the diurnal biting behaviour of Simulium
damnosum (Theobald) (Diptera: Simuliidae). Insect Science Appl., 91: 323 – 328.
Nwoke, B. E. B. (1990). The Socio-Economic Aspects of Human Onchocerciasis in Africa:
Present Appraisal. Journal of Hygiene, Epidemiology, Microbiology and Immunology,
34(1): 37 – 44.
Nwoke, B. E. B., Onwuliri, C. O. E., Iwuala, M. O. E., Ufomadu, G. O., Takahashi, H., Tada, I.
and Shiwaku, K. (1987). Studies on the field epidemiology of human onchocerciasis on the
Jos Plateau, Nigeria IV. Clinical manifestation, socio-economic importance and local
disease perception and treatment. Proceedings of Nigeria / Japan Joint Conference, Jos.
Onchocerciasis Control Programme (1980). World Health Organisation Onchocerciasis Control
Programme in the Volta River Basin Area. Onchocerciasis Control Programme (OCP)
Report (OCP/79.1 Rev. 4).
Onchocerciasis Control Programme (1985). Ten years of Onchocerciasis Control Programme in
West Africa. Review of the Works of Onchocerciasis Control Programme (OCP) in the
Volta River Basin Area from 1974 - 1984. (OCP/GVA/85.1B).
Okafor, F. C. (1990). Parasitic helminthes. In: 19 - 40, Ezigbo (ed). Parasitology for Medical
Students. New Frontier Publishing Limited, Lagos.
Okolo, G. C., Akpa, A. U., and Okonkwo, P. O. (2004). Studies on vectors of onchocerciasis,
Simulium species in Achi, Oji River Local Government Area, Enugu State, Nigeria. Nigerian
Journal of Entomology, 21: 84 - 93.
Okon, O. E., Atting, I. A., Akpanebong, U. E. and Oku, E. E. (2002). The biting behaviour of
black flies Simulium damnosum, the vector of human onchocerciasis in Akamkpa L.G.A
Cross River State, Nigeria. Bioscience Research Communications, 14 (4): 411 – 417.
Opara, K. N., Fagbemi, O. B., Ekwe, A. and Okenu, D. M. N. (2005). Status of forest
onchocerciasis in the lower Cross River Basin, Nigeria: Entomological profile after five
years of ivermectin intervention. American Journal of Tropical Medicine and Hygiene, 73
(2): 371 – 376.

58
Opara, K. N., Usip, L. P. and Akpabio, E. E. (2008). Transmission dynamics of Simulium
damnosum in rural communities of Akwa Ibom State, Nigeria. Journal of Vector Borne
Diseases, 45: 225 - 230.
Opoku, A. A. (2000). Some observations on the ecology, biting activity and parasite infectivity of
the blackfly (Simuliidae) and onchocerciasis prevalence in the River Birim Catchments.
Ghana Journal of Science, 40: 65 - 73.
Osei-Atweneboana, M. Y., Eng, J. K., Boakye, D. A., Gyapong, J. O. and Prichard, R. K. (2007).
Prevalence and intensity of Onchocerca volvulus infection and efficacy of ivermectin in
endemic communities in Ghana: a two-phase epidemiological study. Lancet, 369(9578):
2021 – 2029.
Ottawa Rambling Club (2001). The black flies. The Rambler Summer, 8:2 – 5.
Porter, C. H. and Brandling-Bennett, A. D. (1988). Vector density, parasite prevalence and
transmission of Onchocerca volvulus in Guatemala. American Journal of Tropical Medicine
and Hygiene, 39: 567 – 574.
Porter, C. H. and Collins, R. C. (1988). Seasonality of adult blackflies and Onchocerca volvulus
transmission in Guatemala. American Journal of Tropical Medicine and Hygiene, 37:153 -
167.
Py-Daniel, V. and Medeiros, J. F. (2004). Seasonality, parity rates and transmission indices of
Mansonella ozzardi (Manson) (Nematoda: Onchocercidae) by Cerqueirellum argentiscutum
(Shelley and Luna Dias) (Diptera: Simuliidae) in a lower Solimoes River community,
Amazonas, Brazil. Acta Amazonica, 34 (2):1 – 10.
Remme, J., Baker, R. H., De Sole, G., Dadzie, K. Y., Walsh, J. F., Adams, M. A., Alley, E. S. and
Avissey, H. S. (1989). A community trial of ivermectin in the onchocerciasis focus of
Asubende, Ghana. 1. Effects on microfilarial reservoir and the transmission of Onchocerca
volvulus. Tropical Medicine and Parasitology, 40(3): 367 – 374.
Remme, J., Sole, G. D. and Dadzie, K. Y. (1990). Large scale ivermectin distribution and its
epidemiological consequences. Acta Leidensia, 59:171 – 191.
Renz, A. (1987a). Studies on the dynamics of transmission of onchocerciasis in a Sudan-savanna
area of North Cameroon II. Seasonal and diurnal changes in the biting densities and in the
age-composition of the vector population. Annals of Tropical Medicine and Parasitology,
8(3): 229 – 237.
Renz, A. (1987b). Studies on the dynamics of onchocerciasis in a Sudan-savsnna area of North
Cameroon III. Infection rates of the Simulium vectors and Onchocerca volvulus transmission
potentials. Annals of Tropical Medicine and Parasitology, 81(3): 239 – 252.
Renz, A. and Wenk, P. (1987). Studies on the dynamics of transmission of onchocerciasis in a
Sudan-savanna area of North Cameroon I. Prevailing Simulium vectors, their biting rates and
age-composition at different distances from their breeding sites. Annals of Tropical
Medicine and Parasitology, 81 (3): 215 - 228.
Rodriguez-Perez, M. A. and Reyes-Villanueva, F. (1994). The effect of ivermectin on the
transmission of Onchocerca volvulus in southern Mexico. Salud Publica Mexico, 36(3):
281.
Service, M. W. (2004). Medical entomology for students. Third edition, Cambridge University
Press.

59
Shelley, A. J. (1991). Simuliidae and the transmission and control of human onchocerciasis in
Latin America. Cadornos de Saude Publica, 7 (3): 1 - 10.
Springer (2011). Life cycle of Onchocerca volvulus.
http://www.springerimages. com/Images/LifeScience/1-10.1007_0-306-48380-7_2972-0
Accessed 20th January, 2011.
Taye, A., Gebre-Michael, T. and Staticheff, S. (2000). Onchocerciasis in Gilgel Ghibe River
Valley Southwest Ethiopia. East African Medical Journal, 77 (2): 116 - 120.
Ubachukwu, P. O. (2006). Socio-economic impact of onchocerciasis with particular reference to
females and children: A review. Animal Research International, 3(2): 494 – 504.
Ubachukwu, P. O. (2009). Insects of medical importance. In: 139-192, Okafor, F. C. and Okoye,
I. C. (Eds). Parasitology for the Health Sciences. First Edition, Jolyn Publishers, Enugu.
Ubachukwu, P. O and Anya, A. O. (2001a). Studies on the diurnal biting activity pattern of
simulium damnosum complex (Diptera: simuliidae) in Uzo Uwani Local Government Area
of Enugu State, Nigeria. The Nigerian Journal of Parasitology, 22 (1 and 2): 163 - 168
Ubachukwu, P. O. and Anya, A. O. (2001b). Effects of blackfly bites and manifestations of
human onchocerciasis on the productivity of farmers. Journal of Tropical Agriculture, Food,
Environment and Extension, 2(1): 9 - 16.
Ubachukwu, P. O. and Anya, A. O. (2003). Effects of onchocerciasis manifestations on academic
performance. Journal of Biological Research and Biotechnology, 1(2): 77 - 85.
Ubachukwu, P. O. and Anya, A. O. (2005). Seasonal variations in the biting densities of Simulium
damnosum complex (Diptera: Simuliidae) in Enugu state, Nigeria: Implication for farmers.
Journal of Agriculture, Food, Environment and Extension, 6(1): 10 – 13.
Usip, L. P. E., Opara, K. N., Ibanga, E. S. and Atting, I. A. (1999). Longitudinal evaluation of
repellent activity of Ocimium gratissimum (Labatae) volatile oil against Simulium
damnosum. Memorio do Instituto Ostwaldo Cruz, 101: 201 - 205.
Wagbatsoma, V. A. and Okojie, O. H. (2004). Psychosocial effects of river blindness in Nigeria.
Journal of the Royal Society for the Promotion of Health, 124(3): 134 – 136.
Walsh, J. F., Davies, J. B., Le Berre, R. and Garms, R. (1978). Standardization of criteria for
assessing the effect of Simulium control in Onchocerciasis Control Programme.
Transactions of the Royal Society of Tropical Medicine and Hygiene, 72: 675 – 676.
World Health Organization (1997). Conquering suffering enriching humanity. World Health
Organization Report, Geneva.
World Health Organization (2002). Onchocerciasis. http://www.who.int/topics/onchocerciasis/en/
Accessed 30th
June, 2010.
Yameogo, L., Toe, L., Houghard, J. M., Boatin, B. A. and Unaash, T. R. (1999). Pool screening
polymerase chain reaction for estimating the prevalence of Onchocerca volvulus infection in
Simulium damnosum s. l. results of a field trial in an area subject to successful vector
control. American Journal of Tropical Medicine and Hygiene, 60:124 – 128.

60
APPENDIX
Oneway
Descriptives
N Mean Std. Deviation Std. Error
95% Confidence Interval
for Mean
Minimum Maximum
Lower Bound
Upper
Bound
TEMPERATUR
E
AUGUST 4 27.5525 1.35419 .67709 25.3977 29.7073 25.54 28.45
SEPTEMBER 4 28.3400 .41118 .20559 27.6857 28.9943 27.80 28.80
OCTOBER 4 29.3400 .16411 .08206 29.0789 29.6011 29.14 29.54
NOVEMBER 4 29.2250 1.11327 .55663 27.4535 30.9965 27.91 30.36
DECEMBER 4 29.9450 2.56570 1.28285 25.8624 34.0276 28.09 33.60
JANUARY 4 31.8925 1.57180 .78590 29.3914 34.3936 30.45 33.80
Total 24 29.3825 1.87991 .38373 28.5887 30.1763 25.54 33.80
NO INSECT
COLLECTED
AUGUST 4 17.2500 2.62996 1.31498 13.0652 21.4348 15.00 21.00
SEPTEMBER 4 34.0000 26.87006 13.43503 -8.7563 76.7563 17.00 74.00
OCTOBER 4 10.7500 8.99537 4.49768 -3.5636 25.0636 4.00 24.00
NOVEMBER 4 9.0000 8.16497 4.08248 -3.9923 21.9923 1.00 17.00
DECEMBER 4 5.2500 1.70783 .85391 2.5325 7.9675 3.00 7.00
JANUARY 4 60.7500 6.70199 3.35099 50.0856 71.4144 51.00 66.00
Total 24 22.8333 22.58639 4.61043 13.2959 32.3707 1.00 74.00
ANOVA
Sum of Squares df Mean Square F Sig.
TEMPEATURE Between Groups 44.315 5 8.863 4.316 .009
Within Groups 36.968 18 2.054
Total 81.283 23
NO OF INSECT COLLECTED Between Groups 8960.333 5 1792.067 11.633 .000
Within Groups 2773.000 18 154.056
Total 11733.333 23
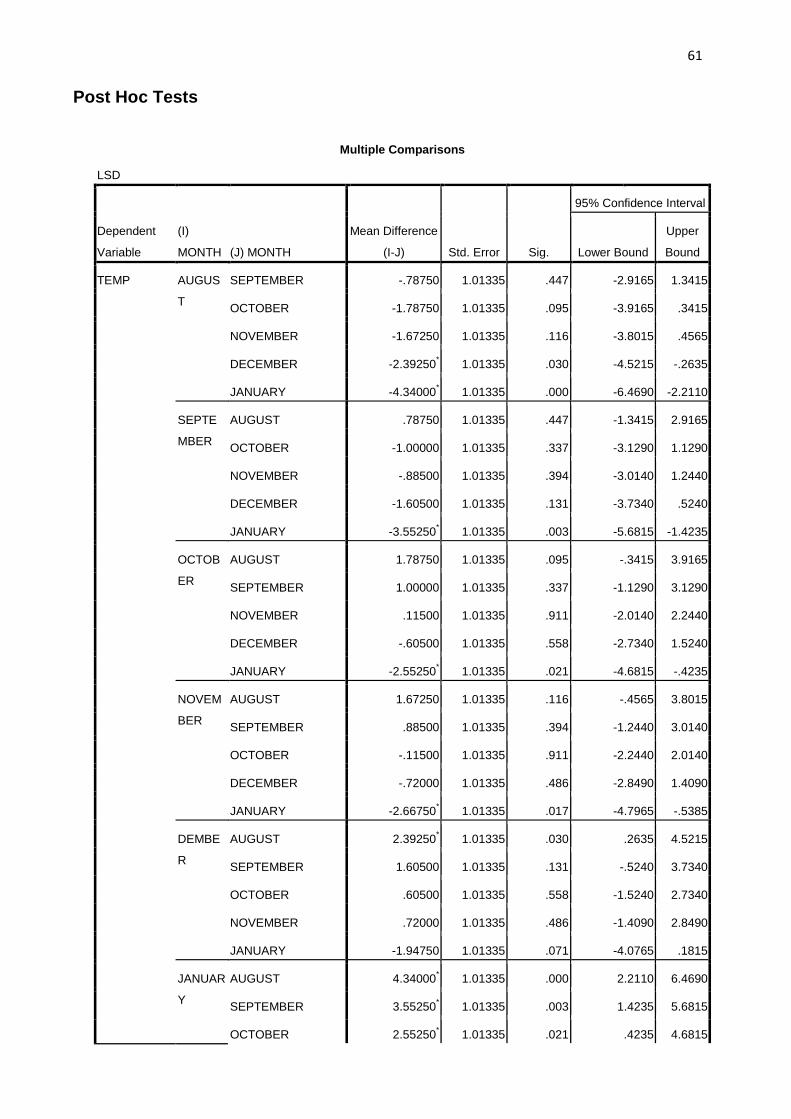
61
Post Hoc Tests
Multiple Comparisons
LSD
Dependent
Variable
(I)
MONTH (J) MONTH
Mean Difference
(I-J) Std. Error Sig.
95% Confidence Interval
Lower Bound
Upper
Bound
TEMP AUGUS
T
SEPTEMBER -.78750 1.01335 .447 -2.9165 1.3415
OCTOBER -1.78750 1.01335 .095 -3.9165 .3415
NOVEMBER -1.67250 1.01335 .116 -3.8015 .4565
DECEMBER -2.39250* 1.01335 .030 -4.5215 -.2635
JANUARY -4.34000* 1.01335 .000 -6.4690 -2.2110
SEPTE
MBER
AUGUST .78750 1.01335 .447 -1.3415 2.9165
OCTOBER -1.00000 1.01335 .337 -3.1290 1.1290
NOVEMBER -.88500 1.01335 .394 -3.0140 1.2440
DECEMBER -1.60500 1.01335 .131 -3.7340 .5240
JANUARY -3.55250* 1.01335 .003 -5.6815 -1.4235
OCTOB
ER
AUGUST 1.78750 1.01335 .095 -.3415 3.9165
SEPTEMBER 1.00000 1.01335 .337 -1.1290 3.1290
NOVEMBER .11500 1.01335 .911 -2.0140 2.2440
DECEMBER -.60500 1.01335 .558 -2.7340 1.5240
JANUARY -2.55250* 1.01335 .021 -4.6815 -.4235
NOVEM
BER
AUGUST 1.67250 1.01335 .116 -.4565 3.8015
SEPTEMBER .88500 1.01335 .394 -1.2440 3.0140
OCTOBER -.11500 1.01335 .911 -2.2440 2.0140
DECEMBER -.72000 1.01335 .486 -2.8490 1.4090
JANUARY -2.66750* 1.01335 .017 -4.7965 -.5385
DEMBE
R
AUGUST 2.39250* 1.01335 .030 .2635 4.5215
SEPTEMBER 1.60500 1.01335 .131 -.5240 3.7340
OCTOBER .60500 1.01335 .558 -1.5240 2.7340
NOVEMBER .72000 1.01335 .486 -1.4090 2.8490
JANUARY -1.94750 1.01335 .071 -4.0765 .1815
JANUAR
Y
AUGUST 4.34000* 1.01335 .000 2.2110 6.4690
SEPTEMBER 3.55250* 1.01335 .003 1.4235 5.6815
OCTOBER 2.55250* 1.01335 .021 .4235 4.6815
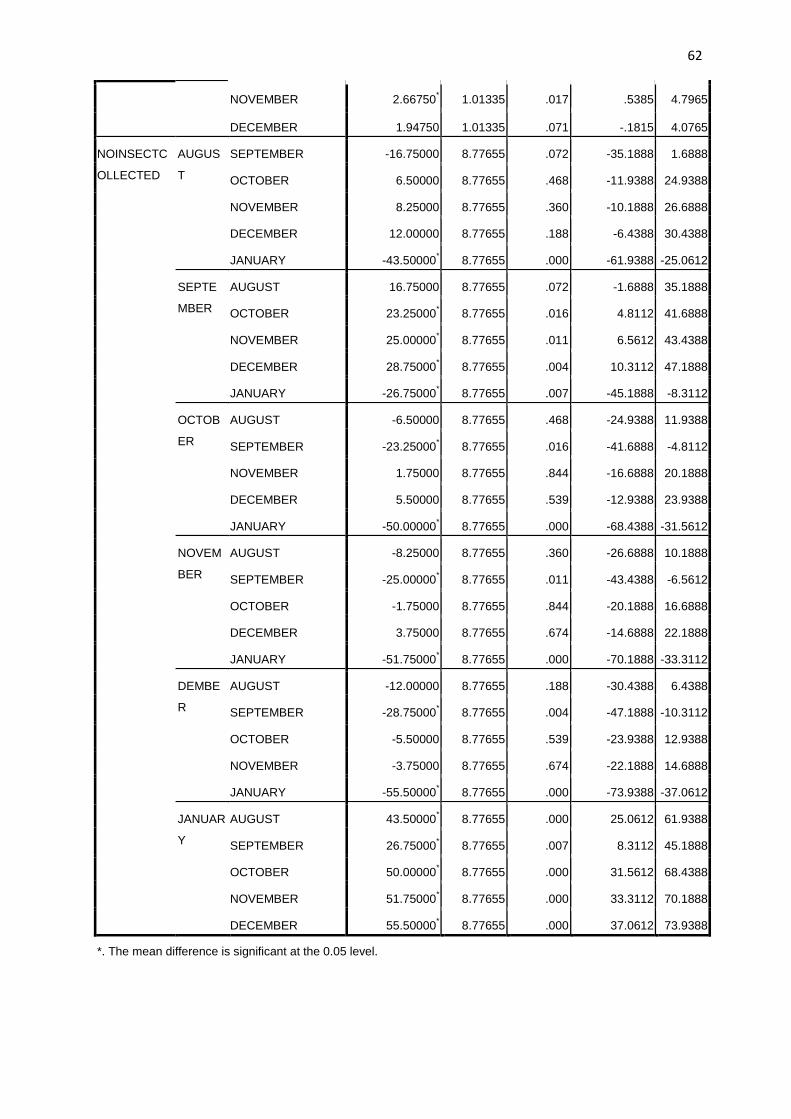
62
NOVEMBER 2.66750* 1.01335 .017 .5385 4.7965
DECEMBER 1.94750 1.01335 .071 -.1815 4.0765
NOINSECTC
OLLECTED
AUGUS
T
SEPTEMBER -16.75000 8.77655 .072 -35.1888 1.6888
OCTOBER 6.50000 8.77655 .468 -11.9388 24.9388
NOVEMBER 8.25000 8.77655 .360 -10.1888 26.6888
DECEMBER 12.00000 8.77655 .188 -6.4388 30.4388
JANUARY -43.50000* 8.77655 .000 -61.9388 -25.0612
SEPTE
MBER
AUGUST 16.75000 8.77655 .072 -1.6888 35.1888
OCTOBER 23.25000* 8.77655 .016 4.8112 41.6888
NOVEMBER 25.00000* 8.77655 .011 6.5612 43.4388
DECEMBER 28.75000* 8.77655 .004 10.3112 47.1888
JANUARY -26.75000* 8.77655 .007 -45.1888 -8.3112
OCTOB
ER
AUGUST -6.50000 8.77655 .468 -24.9388 11.9388
SEPTEMBER -23.25000* 8.77655 .016 -41.6888 -4.8112
NOVEMBER 1.75000 8.77655 .844 -16.6888 20.1888
DECEMBER 5.50000 8.77655 .539 -12.9388 23.9388
JANUARY -50.00000* 8.77655 .000 -68.4388 -31.5612
NOVEM
BER
AUGUST -8.25000 8.77655 .360 -26.6888 10.1888
SEPTEMBER -25.00000* 8.77655 .011 -43.4388 -6.5612
OCTOBER -1.75000 8.77655 .844 -20.1888 16.6888
DECEMBER 3.75000 8.77655 .674 -14.6888 22.1888
JANUARY -51.75000* 8.77655 .000 -70.1888 -33.3112
DEMBE
R
AUGUST -12.00000 8.77655 .188 -30.4388 6.4388
SEPTEMBER -28.75000* 8.77655 .004 -47.1888 -10.3112
OCTOBER -5.50000 8.77655 .539 -23.9388 12.9388
NOVEMBER -3.75000 8.77655 .674 -22.1888 14.6888
JANUARY -55.50000* 8.77655 .000 -73.9388 -37.0612
JANUAR
Y
AUGUST 43.50000* 8.77655 .000 25.0612 61.9388
SEPTEMBER 26.75000* 8.77655 .007 8.3112 45.1888
OCTOBER 50.00000* 8.77655 .000 31.5612 68.4388
NOVEMBER 51.75000* 8.77655 .000 33.3112 70.1888
DECEMBER 55.50000* 8.77655 .000 37.0612 73.9388
*. The mean difference is significant at the 0.05 level.

63
Correlations
Correlations
TEMPERATURE NO OF INSECTS COLLECTED
TEMP Pearson Correlation 1 .325
Sig. (2-tailed) .122
N 24 24
NOINSECTCOLLECTED Pearson Correlation .325 1
Sig. (2-tailed) .122
N 24 24
T-Test
Paired Samples Statistics
Mean N Std. Deviation Std. Error Mean
Pair 1 AUGUST EVENING 5.0000 4 2.16025 1.08012
AUGUST MORNING 2.7500 4 1.70783 .85391
Pair 2 SEPTEMBER EVENING 8.0000 4 4.24264 2.12132
SEPTEMBER MORNING 3.5000 4 2.64575 1.32288
Pair 3 OCTOBER EVENING 1.2500 4 .50000 .25000
OCTOBER MORNING 2.2500 4 1.25831 .62915
Pair 4 NOVEMBER EVENING 3.5000 4 4.04145 2.02073
NOVEMBER MORNING 1.2500 4 1.25831 .62915
Pair 5 DECEMBER EVENING 1.7500 4 2.06155 1.03078
DECEMBER MORNING .7500 4 .50000 .25000
Pair 6 JANUARY EVENING 25.7500 4 6.60177 3.30088
JANUARY MORNING 8.0000 4 9.12871 4.56435
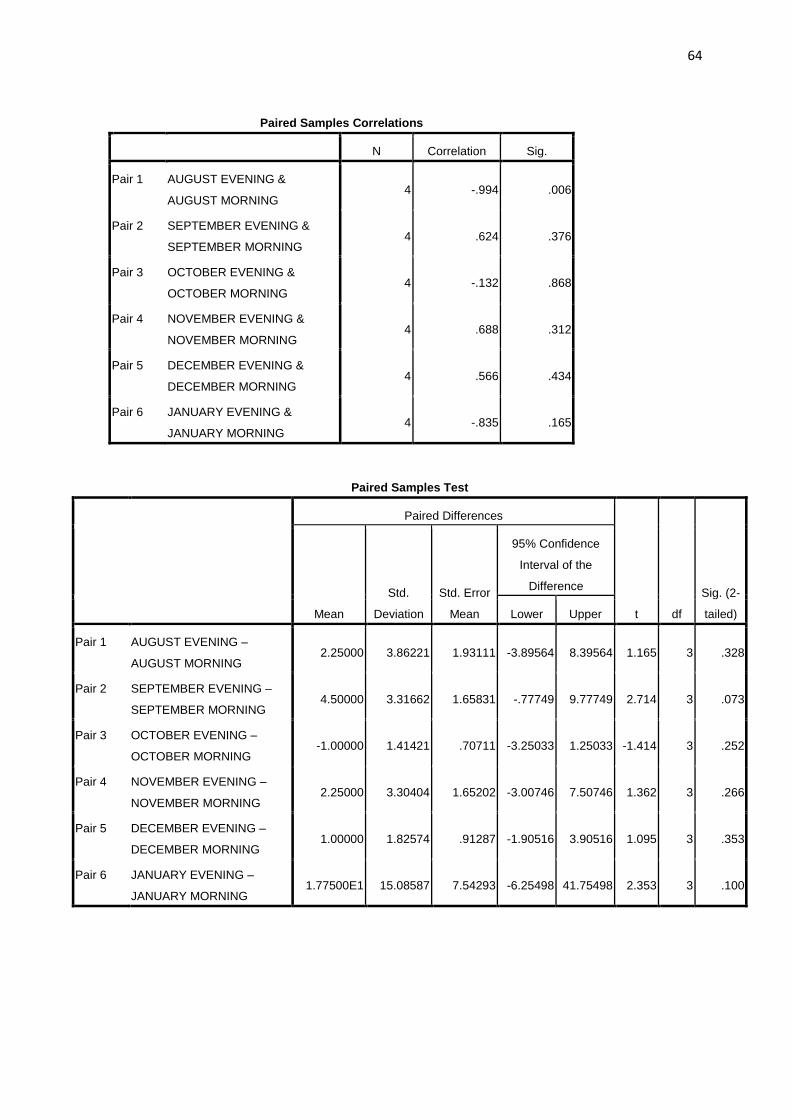
64
Paired Samples Correlations
N Correlation Sig.
Pair 1 AUGUST EVENING &
AUGUST MORNING 4 -.994 .006
Pair 2 SEPTEMBER EVENING &
SEPTEMBER MORNING 4 .624 .376
Pair 3 OCTOBER EVENING &
OCTOBER MORNING 4 -.132 .868
Pair 4 NOVEMBER EVENING &
NOVEMBER MORNING 4 .688 .312
Pair 5 DECEMBER EVENING &
DECEMBER MORNING 4 .566 .434
Pair 6 JANUARY EVENING &
JANUARY MORNING 4 -.835 .165
Paired Samples Test
Paired Differences
t df
Sig. (2-
tailed)
Mean
Std.
Deviation
Std. Error
Mean
95% Confidence
Interval of the
Difference
Lower Upper
Pair 1 AUGUST EVENING –
AUGUST MORNING 2.25000 3.86221 1.93111 -3.89564 8.39564 1.165 3 .328
Pair 2 SEPTEMBER EVENING –
SEPTEMBER MORNING 4.50000 3.31662 1.65831 -.77749 9.77749 2.714 3 .073
Pair 3 OCTOBER EVENING –
OCTOBER MORNING -1.00000 1.41421 .70711 -3.25033 1.25033 -1.414 3 .252
Pair 4 NOVEMBER EVENING –
NOVEMBER MORNING 2.25000 3.30404 1.65202 -3.00746 7.50746 1.362 3 .266
Pair 5 DECEMBER EVENING –
DECEMBER MORNING 1.00000 1.82574 .91287 -1.90516 3.90516 1.095 3 .353
Pair 6 JANUARY EVENING –
JANUARY MORNING 1.77500E1 15.08587 7.54293 -6.25498 41.75498 2.353 3 .100

65
T-Test
Paired Samples Statistics
Mean N Std. Deviation Std. Error Mean
Pair 1 AUGUST PAROUS .0000 4 .00000 .00000
AUGUST NULLIPAROUS 17.2500 4 2.62996 1.31498
Pair 2 SEPTEMBER PAROUS .0000 4 .00000 .00000
SEPTEMBER NULLIPAROUS 34.0000 4 26.87006 13.43503
Pair 3 OCTOBER PAROUS 3.2500 4 3.20156 1.60078
OCTOBER NULLIPAROUS 7.5000 4 5.80230 2.90115
Pair 4 NOVEMBER PAROUS 2.2500 4 2.62996 1.31498
NOVEMBER NULLIPAROUS 6.7500 4 5.56028 2.78014
Pair 5 DECEMBER PAROUS 2.5000 4 .57735 .28868
DECEMBER NULLIPAROUS 2.7500 4 1.50000 .75000
Pair 6 JANUARY PAROUS 23.5000 4 4.93288 2.46644
JANUARY NULLIPAROUS 37.2500 4 7.88987 3.94493
Paired Samples Correlations
N Correlation Sig.
Pair 1 AUGUST PAROUS & AUGUST
NULLIPAROUS 4 . .
Pair 2 SEPTEMBER PAROUS &
SEPTEMBER NULLIPAROUS 4 . .
Pair 3 OCTOBER PAROUS &
OCTOBER NULLIPAROUS 4 .996 .004
Pair 4 NOVEMBER PAROUS &
NOVEMBER NULLIPAROUS 4 .986 .014
Pair 5 DECEMBER PAROUS &
DECEMBER NULLIPAROUS 4 .192 .808
Pair 6 JANUARY PAROUS &
JANUARY NULLIPAROUS 4 -.535 .465

66
Paired Samples Test
Paired Differences
t df
Sig. (2-
tailed)
Mean
Std.
Deviation
Std. Error
Mean
95% Confidence Interval
of the Difference
Lower Upper
Pair 1 AUGUST PAROUS – AUGUST
NULLIPAROUS -1.72500E1 2.62996 1.31498 -21.43485 -13.06515 -13.118 3 .001
Pair 2 SEPTEMBER PAROUS –
SEPTEMBER NULLIPAROUS -3.40000E1 26.87006 13.43503 -76.75626 8.75626 -2.531 3 .085
Pair 3 OCTOBER PAROUS –
OCTOBER NULLIPAROUS -4.25000 2.62996 1.31498 -8.43485 -.06515 -3.232 3 .048
Pair 4 NOVEMBER PAROUS –
NOVEMBER NULLIPAROUS -4.50000 3.00000 1.50000 -9.27367 .27367 -3.000 3 .058
Pair 5 DECEMBER PAROUS –
DECEMBER NULLIPAROUS -.25000 1.50000 .75000 -2.63683 2.13683 -.333 3 .761
Pair 6 JANUARY PAROUS –
JANUARY NULLIPAROUS -1.37500E1 11.32475 5.66238 -31.77021 4.27021 -2.428 3 .093

67
Oneway
Descriptives
N Mean
Std.
Deviation Std. Error
95% Confidence
Interval for Mean
Minimu
m
Maxim
um
Lower
Bound Upper Bound
PAROUS AUGUST 4 .0000 .00000 .00000 .0000 .0000 .00 .00
SEPTEMBER 4 .0000 .00000 .00000 .0000 .0000 .00 .00
OCTOBER 4 3.2500 3.20156 1.60078 -1.8444 8.3444 1.00 8.00
NOVEMBER 4 2.2500 2.62996 1.31498 -1.9348 6.4348 .00 5.00
DECEMBER 4 2.5000 .57735 .28868 1.5813 3.4187 2.00 3.00
JANUARY 4 23.5000 4.93288 2.46644 15.6507 31.3493 18.00 29.00
Total 24 5.2500 8.74891 1.78586 1.5557 8.9443 .00 29.00
NULLIPARO
US
AUGUST 4 17.2500 2.62996 1.31498 13.0652 21.4348 15.00 21.00
SEPTEMBER 4 34.0000 26.87006 13.43503 -8.7563 76.7563 17.00 74.00
OCTOBER 4 7.5000 5.80230 2.90115 -1.7328 16.7328 3.00 16.00
NOVEMBER 4 6.7500 5.56028 2.78014 -2.0976 15.5976 1.00 12.00
DECEMBER 4 2.7500 1.50000 .75000 .3632 5.1368 1.00 4.00
JANUARY 4 37.2500 7.88987 3.94493 24.6955 49.8045 30.00 48.00
Total 24 17.5833 17.39044 3.54981 10.2400 24.9267 1.00 74.00
ANOVA
Sum of Squares Df Mean Square F Sig.
PAROUS Between Groups 1635.000 5 327.000 46.900 .000
Within Groups 125.500 18 6.972
Total 1760.500 23
NULLIPAROUS Between Groups 4381.833 5 876.367 6.128 .002
Within Groups 2574.000 18 143.000
Total 6955.833 23

68
Post Hoc Tests
Multiple Comparisons
LSD
Dependent Variable
(I)
GROUP
(J)
GROUP
Mean Difference
(I-J) Std. Error Sig.
95% Confidence Interval
Lower Bound Upper Bound
PAROUS AUGUS
T
SEPT .00000 1.86711 1.000 -3.9227 3.9227
OCT -3.25000 1.86711 .099 -7.1727 .6727
NOV -2.25000 1.86711 .244 -6.1727 1.6727
DEC -2.50000 1.86711 .197 -6.4227 1.4227
JAN -23.50000* 1.86711 .000 -27.4227 -19.5773
SEPT AUG .00000 1.86711 1.000 -3.9227 3.9227
OCT -3.25000 1.86711 .099 -7.1727 .6727
NOV -2.25000 1.86711 .244 -6.1727 1.6727
DEC -2.50000 1.86711 .197 -6.4227 1.4227
JAN -23.50000* 1.86711 .000 -27.4227 -19.5773
OCT AUG 3.25000 1.86711 .099 -.6727 7.1727
SEPT 3.25000 1.86711 .099 -.6727 7.1727
NOV 1.00000 1.86711 .599 -2.9227 4.9227
DEC .75000 1.86711 .693 -3.1727 4.6727
JAN -20.25000* 1.86711 .000 -24.1727 -16.3273
NOV AUG 2.25000 1.86711 .244 -1.6727 6.1727
SEPT 2.25000 1.86711 .244 -1.6727 6.1727
OCT -1.00000 1.86711 .599 -4.9227 2.9227
DEC -.25000 1.86711 .895 -4.1727 3.6727
JAN -21.25000* 1.86711 .000 -25.1727 -17.3273
DEC AUG 2.50000 1.86711 .197 -1.4227 6.4227
SEPT 2.50000 1.86711 .197 -1.4227 6.4227
OCT -.75000 1.86711 .693 -4.6727 3.1727
NOV .25000 1.86711 .895 -3.6727 4.1727
JAN -21.00000* 1.86711 .000 -24.9227 -17.0773
JAN AUG 23.50000* 1.86711 .000 19.5773 27.4227
SEPT 23.50000* 1.86711 .000 19.5773 27.4227
OCT 20.25000* 1.86711 .000 16.3273 24.1727
NOV 21.25000* 1.86711 .000 17.3273 25.1727

69
DEC 21.00000* 1.86711 .000 17.0773 24.9227
NULLIPAROUS AUGUS
T
SEPT -16.75000 8.45577 .063 -34.5149 1.0149
OCT 9.75000 8.45577 .264 -8.0149 27.5149
NOV 10.50000 8.45577 .230 -7.2649 28.2649
DEC 14.50000 8.45577 .104 -3.2649 32.2649
JAN -20.00000* 8.45577 .029 -37.7649 -2.2351
SEPT AUG 16.75000 8.45577 .063 -1.0149 34.5149
OCT 26.50000* 8.45577 .006 8.7351 44.2649
NOV 27.25000* 8.45577 .005 9.4851 45.0149
DEC 31.25000* 8.45577 .002 13.4851 49.0149
JAN -3.25000 8.45577 .705 -21.0149 14.5149
OCT AUG -9.75000 8.45577 .264 -27.5149 8.0149
SEPT -26.50000* 8.45577 .006 -44.2649 -8.7351
NOV .75000 8.45577 .930 -17.0149 18.5149
DEC 4.75000 8.45577 .581 -13.0149 22.5149
JAN -29.75000* 8.45577 .002 -47.5149 -11.9851
NOV AUG -10.50000 8.45577 .230 -28.2649 7.2649
SEPT -27.25000* 8.45577 .005 -45.0149 -9.4851
OCT -.75000 8.45577 .930 -18.5149 17.0149
DEC 4.00000 8.45577 .642 -13.7649 21.7649
JAN -30.50000* 8.45577 .002 -48.2649 -12.7351
DEC AUG -14.50000 8.45577 .104 -32.2649 3.2649
SEPT -31.25000* 8.45577 .002 -49.0149 -13.4851
OCT -4.75000 8.45577 .581 -22.5149 13.0149
NOV -4.00000 8.45577 .642 -21.7649 13.7649
JAN -34.50000* 8.45577 .001 -52.2649 -16.7351
JAN AUG 20.00000* 8.45577 .029 2.2351 37.7649
SEPT 3.25000 8.45577 .705 -14.5149 21.0149
OCT 29.75000* 8.45577 .002 11.9851 47.5149
NOV 30.50000* 8.45577 .002 12.7351 48.2649
DEC 34.50000* 8.45577 .001 16.7351 52.2649
*. The mean difference is significant at the 0.05 level.





























