Embed Size (px)
Citation preview

i
STUDIES ON THE EPIDEMIOLOGY OF
ONCHOCERCIASIS AND MORPHOMETRIC
CHARACTERISTICS OF THE SIMULIUM DAMNOSUM
COMPLEX IN NKPOLOGU AND OGURUGU, UZO-UWANI
L.G.A., ENUGU STATE, NIGERIA.
BY
EBIDO, CHIKE. C.
PG/M.Sc/06/42105
A PROJECT REPORT SUBMITTED FOR THE DEGREE
OF MASTEROF SCIENCE (M.SC) IN ZOOLOGY
DEPARTMENT OF ZOOLOGY
UNIVERSITY OF NIGERIA
NSUKKA
NOVEMBER, 2009

ii
CERTIFICATION
Ebido, Chike C., a postgraduate student in the Department of Zoology with Registration
number PG/M.Sc/06/42105 has satisfactorily completed the course and research work
requirements for the award of Master Degree (M.Sc.) in Zoology. The work embodied in this
project report is original and has not been submitted in part or in full for any other diploma
or degree.
_________________ __________________
Ebido, C.C. Dr. P.O. Ubachukwu
(Student) (Supervisor)
_________________ __________________
Dr J.E. Eyo (External Examiner)
(Head of Department)

iii
DEDICATION
This work is dedicated to the millions of Africans who are being plagued
by the evils of sickness, sorrow and poverty … Your redemption draws
near.

iv
ACKNOWLEDGEMENTS
My gratitude goes to God Almighty – the source of all knowledge, He is ever faithful. My
supervisor, Dr, P. O. Ubachukwu was very supportive and generous with ideas; May God
bless you. My appreciation also goes to all the lecturers of Zoology department especially
Prof. F. C. Okafor, Rev. (Dr.) I. C. Okoye and Dr. J. E. Eyo for their kind advice and
encouragement. My parents, Engr. and Mrs J. N. Ebido were very helpful financially and
morally; God will not forget your labour of love. I will not forget Dr. Dan Ozoko, Ify Ozor
and other members of the REO MINISTRIES team for their encouragement and support.
Special thanks also go to my aubty, Mrs F. A. Ebido and my colleagues Cyril, Sowechi,
Ogoo and Chika. Chuks, Lanre, Uzy, Stanley, Ngozi, Amara, Ozioma, Dominic and others
too numerous to mention; you are not forgotten. May God bless you richly.

v
LIST OF TABLES
TABLE 1: Prevalence of Nodules in Nkpologu … .. .. .. .. 28
TABLE 2: Prevalence of Nodules in Ogurugu … .. .. .. .. 28
TABLE 3: Prevalence rate of Leopard Skin in Nkpologu … .. .. .. 32
TABLE 4: Prevalence of Leopard Skin in Ogurugu … .. .. .. .. 32
TABLE 5: Prevalence of Lizard Skin in Nkpologu … .. .. .. .. 35
TABLE 6: Prevalence of Lizard Skin in Ogurugu … .. .. .. .. 35
TABLE 7: Prevalence of Onchodermatitis in Nkpologu … .. .. .. 38
TABLE 8: Prevalence of Onchodermatitis in Ogurugu … .. .. .. 38
TABLE 9: Prevalence of Impaired Sight in Nkpologu … .. .. .. 40
TABLE 10: Prevalence of Impaired Sight in Ogurugu … .. .. .. 40
TABLE 11: Prevalence of Blindness in Nkpologu … .. .. .. 41
TABLE 12: Prevalence of Blindness in Ogurugu . … .. .. .. 42
TABLE 13: Mectizan Distribution In Nkpologu And Ogurugu Communities … 43
TABLE 14: Mean Length of the various fly parts measured … .. .. .. 44
TABLE 15: Mean ratios of the various fly parts measured … .. .. .. 45
TABLE 16: Mean Length of the various parts (Savanna and Forest) … .. 46
TABLE 17 Mean ratios of the various fly parts (Savanna and Forest) … .. 46
TABLE 18: Discriminant Function Analysis (Variables Entered/Removeda,b,c,d) .. 47
TABLE 19: Discriminant Function Analysis (Eigenvalues) … .. 48
TABLE 20: Discriminant Function Analysis (Classification Results a) … .. 48

vi
LIST OF FIGURES
Figure 1: Map of Uzo-Uwani showing Ogurugu and Nkpologu … .. ... 25
Figure 2: Prevalence of Nodules in Nkpologu and Ogurugu… .. .. .. 30
Figure 3: Prevalence of Leopard Skin in Nkpologu and Ogurugu … .. .. 33
Figure 4: Prevalence of Lizard Skin in Nkpologu and Ogurugu … .. .. 36
Figure 5: Prevalence of Onchodermatitis in Nkpologu and Ogurugu … .. 39
Figure 6: Prevalence of Impaired Sight in Nkpologu and Ogurugu … .. .. 41
Figure 7: Prevalence of Onchocercal Blindness in Nkpologu and Ogurugu … .. 43

vii
LIST OF PLATES
Plate 1: Head Nodules in an Individual … .. .. .. .. .. 28
Plate 2: Leopard Skin in an Individual … .. .. .. .. .. 31
Plate 3: Lizard Skin in an Individual … .. .. .. .. .. 34
Plate 4: Onchodermatitis in an Individual … .. .. 37
Plate 5: Simulium damnosum … .. .. .. .. .. .. .. 44

viii
TABLE OF CONTENTS
Title page ... .. .. .. .. .. .. .. .. .. i Certification ... .. .. .. .. .. .. .. .. .. ii Dedication ... .. .. .. .. .. .. .. .. .. iii Acknowledgements... .. .. .. .. .. .. .. .. .. iv List of tables ... .. .. .. .. .. .. .. .. .. v List of figures ... .. .. .. .. .. .. .. .. .. vi List of plates ... .. .. .. .. .. .. .. .. .. vii Table of contents... .. .. .. .. .. .. .. .. .. ix Abstract ... .. .. .. .. .. .. .. .. .. x
CHAPTER ONE
INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction ... .. .. .. .. .. .. .. 1 1.2 Review of literature ... .. .. .. .. .. .. 3 1.2.1 Human Onchocerciasis … .. .. .. .. .. .. 4 1.2.2 Transmission dynamics of Onchocerciasis… .. .. .. .. 5 1.2.3 Biting activity of Simulium (BLACKFLIES) … .. .. .. 7 1.2.4 Host choice/specificity ... .. .. .. .. .. .. 9 1.2.5 Adult Morphology … .. .. .. .. .. .. 10 1.2.6 Larval Structure and Function … .. .. .. .. .. 14 1.2.7 Phenotypic Plasticity … .. .. .. .. .. .. 16 1.2.8 Cytotaxonomy of the Simulium damnosum complex. .. .. .. 19 1.2.9 Control Measures … .. .. .. .. .. .. 21 1.3 Objectives of the Study … … … … .. .. .. 23 CHAPTER TWO
MATERIALS AND METHODS … .. .. .. .. .. .. 24 2.1 Study Area … .. .. .. .. .. .. .. 26 2.2 Epidemiological Studies …. .. .. .. .. .. .. 26 2.2.1 Questionnaire Administration … .. .. .. .. .. 26 2.2.2 Clinical Examination … .. .. .. .. .. .. 26 2.2.3 Visual Acuity Testing … .. .. .. .. .. .. 26 2.3 Fly Collection … .. .. .. .. .. .. .. 27 2.4 Morphometric Studies on Blackflies … .. .. .. .. 27 2.5 Statistical Analysis … .. .. .. .. .. .. 27
CHAPTER THREE
RESULTS ... .. .. .. .. .. .. .. .. .. 28 3.1 Epidemiology of Onchocerciasis in Nkpologu and Ogurugu Communities 28 3.1.1 Presence of Palpable Nodules … .. .. .. .. .. 28 3.1.2 Presence of Leopard Skin … .. .. .. .. .. 31 3.1.3 Presence of Lizard Skin … .. .. .. .. .. .. 34 3.1.4 Presence of Onchodermatitis … .. .. .. .. .. 37 3.1.5 Visual Acuity Testing … .. .. .. .. .. .. 39

ix
3.1.5.1 Impaired Sight … .. .. .. .. .. .. .. 39 3.1.5.2 Blindness … .. .. .. .. .. .. .. 41 3.1.6 Mectizan Distribution … .. .. .. .. .. .. 43 3.2 Morphometric Studies on Blackflies … .. .. .. .. 44
CHAPTER FOUR
DISCUSSION AND CONCLUSION ... .. .. .. .. .. 49 4.1 Epidemiology of Onchocerciasis in Nkpologu and Ogurugu ... .. 49 4.2 Morphometric Studies on Blackflies … .. .. .. .. 54 4.3 Summary and Recommendations … .. .. .. .. .. 56
REFERENCES APPENDIX

x
ABSTRACT
Studies on the epidemiology of onchocerciasis and the morphometric characteristics
of the Simulium vectors were carried out in Ogurugu and Nkpologu in Uzo-Uwani L.
G. A. of Enugu State, Nigeria, between September 2008 and January, 2009. The
subjects used for epidemiological studies were randomly sampled using Rapid
Assessment Methods while Simulium flies were collected using human baits. The
two communities sampled were found to be endemic for onchocerciasis endemicity
(palpable nodules, skin depigmentation). Nkpologu had a higher prevalence rate of
nodules (5.8%) and leopard skin (12.5%) than Ogurugu (nodules = 2.6%; leopard
skin = 4.0%). Also, the two communities were found to be meso-endemic for
onchocerciasis based on blindness rates, with Nkpologu having a higher prevalence
of blindness than Ogurugu (Nkpologu = 3.8%; Ogurugu = 1.6%). This could be
attributed to the better distribution systems of Mectizan in Ogurugu than Nkpologu.
Furthermore, the morphometric studies carried out on the Simulium damnosum
populations in Nkpologu showed a higher abundance of savanna flies (54) than
forest flies (21) among those sampled. The classification was done using the thorax
length/antenna length (TL/AL) ratio. The morphometric study could not be carried
out in Ogurugu because of the absence of blackflies due to the treatment of the river
with chemicals which killed the blackfly larvae. Discriminant function analysis
reveals that 96.0% of originally grouped cases were correctly classified. The
implications of the results are discussed.

xi
CHAPTER ONE
INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction
Human onchocerciasis is a severely debilitating and blinding disease caused
by infection with the parasitic nematode O. volvulus (Leuckart) (Nematoda:
Filarioidea). In West Africa, Onchocerciasis is transmitted exclusively by sibling
species of the S. damnosum Theobald complex (Diptera: Simuliidae) with blindness
rates reaching as high as 15% in the savanna where up to 100% of adults are
infected. Infection rates can be similarly high in the forest bioclimatic zone and
although the ocular manifestation is usually very much less than in the savanna
(with blindness rates typically around 0.5%, (Duke, 1990), onchocercal skin disease
can be a significant socioeconomic problem (Remme, 1995).
Blackfiles (Diptera: Simuliidae) are found from Arctic to tropical ecosystems,
where they have significant economic impact on humans and animal production,
and may reduce the fitness of wildlife (Crosskey, 1990; Adler et al., 2004). About
1800 species of blackflies are recognized worldwide (Crosskey, 2002) and they
often reach very high abundances (Wotton, 1988) suggesting that their impact on
wild animals in terrestrial landscape is significant. Blackflies are among the insects
of foremost medical importance around the world, as they are vectors of filarial
nematode causing onchocerciasis in inhabitants of tropical regions (Molloy, 1982).
They also have a veterinary importance, because they transmit Protozoa and
Nematoda to animals, and are therefore a nuisance, reducing the productivity of
livestock and other farm animals (National Research Council of Canada, 1982).
In the African savanna, Simulium damnosum Theobald sensu lato which
transmit Onchocerca volvulus is also know to naturally transmit other species of

xii
Onchocerca from animals (Duke, 1967; Renz et al., 1989) including O. ochengi from
cattle (Omar, et al., 1979; Wahl, 1996) and O. ramachandrini (Duke, 1967) from the
Wart hog, Phacochoerus aethiopicus.
Accurate identification of Simuliidae is becoming increasingly significant as
more species become the target of pest and vector control operations, or the subject
of epidemiological studies on the transmission of the blackfly-borne parasites of man
(O. volvulus and Mansonella ozzardi). The characterization and differentiation of
species are the most important practical functions of contemporary taxonomy. At
present, the overwhelming majority of blackflies are still recognized and defined on
characters of their external (“hard-parts”) morphology, even recently described
species.
Species limits cannot be determined yet from experimental genetics because
of the lack of success in cross mating blackflies in the laboratory. Thus, most so-
called species in the taxonomic literature are morphospecies, differentiated from
other such taxa by morphological discontinuities deemed to be evidence of
specificity in one or more life-history stages and/or (in the case of adults) in either or
both sexes (Crosskey, 1981).
Prior to 1966, S. damnosum Theobald was considered to be a single, but
morphologically and biologically variable species. However, 25 cytotypes in this
complex have been discovered as a result of further chromosomal studies (Dunbar,
1969; Dunbar and Vajime, 1971, 1972; Vajime and Dunbar, 1975). These cytotypes
and their distributions were summarized in tabular form in a report by WHO (1978).
The West African members of this complex include S. damnosum Theobald s.s, S.
squamosum Enderlein, S. sirbanum, S. sanctipauli, S. soubrense, S. yahense, S.
dieguerense, and S. sudanense (Vajime and Dunbar, 1975). The latter two species
are rare and of uncertain status.

xiii
A detailed morphological study of the adults of the six common sibling species of the
S. damnosum complex occurring in West Africa has been in progress since 1976. Its
purpose was to try to find external characters that could be relatively easily
observed and used with a high degree of reliability to separate these species.
According to current opinion, S. squamosum, a member of this complex
described by Enderlein in 1921, is a distinct species both on the basis of
chromosomal and external morphological features. Even though there is great
similarity between S. squamosum and S. damnosum s.s., there are specific external
characters that can be used to separate the majority of adult specimens of these two
species. Presumably, it should also be possible to separate most, if not all, of the
other known siblings in the complex.
1.2 Review of Literature
Onchocerciasis is being controlled in Nigeria by the antifilarial drug ivermectin
(Edungbola, 1991) distributed in communities identified by Rapid Epidemiological
Mapping procedures (Ngoumou and Walsh, 1993) as being ‘at risk’. Rapid
Epidemiological Mapping aims to assess the current status of onchocerciasis in a
community from the current frequency of easily assessed symptoms, but it is less
efficient at quickly detecting any trend of change in prevalence or severity of the
disease. Early warning of any such change might be obtained by regular
identification of the sibling species comprising the biting fly population (Mafuyai et
al., 1996).
The most practical procedure routinely available for distinguishing between
the species of adult females belonging to the S. damnosum complex is the
assessment of micro-morphological characters (Dang and Peterson, 1980), best
known from the area of the World Health Organization Onchocerciasis Control

xiv
Programme (WHO/OCP) involving West African countries to the West of Nigeria
(Wilson et al., 1993, 1994).
1.2.1 Human Onchocerciasis
Onchocerciasis is the most debilitating and most dreadful human filariasis,
which is caused by the parasitic filarial worm Onchocerca volvulus. It is transmitted
to humans through the bites of blackflies of the genus Simulium that breed in fast
flowing rivers hence the name river blindness. According to estimates of the WHO
(1995a), there are more than 120million people at risk of infection worldwide, 18
million people are infected, 800,000 visually impaired and 270,000 blind.
In Nigeria, onchocerciasis is widespread and a cause of blindness in most
rural communities. Of all the countries of the world, Nigeria has the largest number
of persons with onchocerciasis accounting for about a third of the global prevalence
(Edungbola, 1991), with 40 million Nigerians at the risk of infections and about 7
million Nigerians infected (Nwoke, 1990). The adult worms are located under the
skin in nodules, usually knotted together in pairs. Their distribution over the body is
largely determined by the biting pattern of the insect vector. In Central America, the
nodules are usually found above the waist, where as in Africa they are mainly found
below the waist (WHO, 1995a). The nodules formed around the adult worms are as
a result of the host reaction. The nodules are prominent when they occur over bone
joints and/or the skull region; these are also used as rapid assessment diagnostic
features for community diagnosis (Edungbola and Parakoyi, 1991).
Dermal manifestations of onchocerciasis have very serious social, cultural
and psychological implications for affected persons. People with severe onchocercal
dermatitis are discriminated against, humiliated by peers, avoided by friends, and
stigmatized by society. Hanging groins, hernias and scrotal elephantiasis have also

xv
been associated with onchocerciasis with attendant psychosocial maladjustment
(Nwoke, 1990). In a study to assess the epidemiological status of Uzo-uwani Local
Government Area of Enugu State, Nigeria, Ubachukwu (2004) concluded that the
area could be said to be mesoendemic for onchocerciasis based on the rapid
indicators of onchocerciasis endemicity used especially the prevalence of palpable
nodules. The study reported that there was consistently a higher prevalence of
every examined manifestation of onchocerciasis in males than in females. This was
because, males right from childhood, were involved more in outdoor activities such
as farming, swimming etc. than females who are involved more in in-door activities
such as cooking.
1.2.2 Transmission Dynamics of Onchocerciasis
It is generally accepted that man-fly contact must be reduced to a very low
level to interrupt transmission of onchocerciasis or to reduce it to levels at which
serious clinical signs of the disease are unlikely to occur. Duke et al. (1975) and
Thylefors et al. (1978) found that there was a correlation between the intensity of
human infection in the Sudan savanna of West Africa and the amount of man-fly
contact (Annual Biting Rate, ABR) and also the amount of transmission (Annual
Transmission Potential, ATP).
According to Renz and Wenk (1987), the number of flies counted per hour
was divided by the number of collectors working simultaneously to give the hourly
biting rate per man. The daily biting rate (DBR) was then the sum of 13-hourly
values from catches between 06:00 and 18:30 hours. The monthly and Annual
Biting Rates (MBR, ABR) were calculated according to Walsh et al. (1979), the ABR
being the sum of 12 monthly biting rates during one hydrological year, from the
beginning of the rainy season in May to the end of the following dry season in April.

xvi
The Annual Biting Rate (ABR) is important as an indicator of the risk of
disease transmission especially during a Simulium control campaign (Walsh et al.,
1978). Consequently, an ABR of 1000 represents the maximal tolerable biting rate in
a controlled area (Walsh et al., 1979). Also, the Monthly Transmission Potential
(MTP) could be calculated as the arithmetic mean of the number of L3 larvae per fly
dissected multiplied by the MBR. The sum of the MTPs for the 12 months of the year
gave the Annual Transmission Potential (ATP). This estimate follows the procedure
described by Walsh et al., (1978) and Renz (1987).
In a study on the intensity of natural transmission of O. ochengi and O.
vovulus by anthropophilic and zoophilic S. damnosum s.I. in the Guinea savanna of
Cameroon, Achukwi et al. (2000) reported that the ATP of O. ochengi at the river
was 5 times higher than that of O. volvulus in this study area. This followed previous
observations that the S. damnosum s.I. biting density on cattle was about twice that
of human bait (Wahl, 1996; Wahl et al., 1998).
1.2.3 Biting Activity of Simulium (Blackflies)
Blackflies bite by day and in the open. Biting inside buildings and vehicles, or
after dark, is exceptional. Dalmat (1955) has recorded flies of the S. ochraceum
complex feeding in dimly lit rooms and on dark verandahs after nightfall.
Nevertheless, the general rule is that simuliidae are daytime and exophilic (outdoor)
biters. Due to the influence of climate on development and adult fly emergence,
there are seasonal cycles of biting activity in most parts of the world-but especially in
tropical areas with well-defined wet and dry seasons and in high latitudes with cold
winters and hot summers (Crosskey, 1990).

xvii
As would be expected, host-seeking and biting activity occurs mainly within
certain optimal ranges of wind speed, temperature, light intensity and saturation
deficiency (McCreadie et al., 1986). Biting is generally inhibited by heavy rain and
strong blowing or gusting winds, but is not usually, affected much by light rain or
light winds; occasional drizzle has even been claimed to encourage the biting
activity of Himalayan species (Datta and Dasgupta, 1978).
Blackflies bite more or less from dawn to dusk, but seldom at the same
sustained level throughout the day. Biting tends to be strongly concentrated into
activity peaks with lulls between. For any given species, temperate or tropical, the
times when peaks and lulls occur are often more or less similar from day to day, but
between species, there are often differences in activity pattern (Crosskey, 1990).
Biting patterns have been most studied in the tropics, where season has the least
effect on day length. A bimodal pattern, with a morning peak about 07:00-09:00
hours and an afternoon or evening peak about 16:00-18.00 hours, is common and
has been reported for instance in the S. damnosum complex in West and East
Africa.
In a study on the daily biting activity pattern of S. damnosum complex in Uzo-
Uwani Local Government Area of Enugu State, Nigeria, Ubachukwu and Anya
(2001) reported that there were variations in the biting activity pattern within each
day and from day to day. The blackflies were generally bimodal in their diurnal biting
habits. Biting occurred from morning until evening but not at the same rate. Peak
biting periods occurred in the morning (9.00-10.00am) and in the evening (5.00-
6.00pm) with the evening peak significantly higher than the morning peak by a factor
of about 4. Unimodal patterns seem less frequent.
In a study carried out in Nkpologu in Uzo-Uwani Local Government Area of
Enugu State, Nigeria, Ubachukwu and Anya (2005) compared the biting densities of

xviii
Simulium flies in different seasons of the year. They reported that biting activity
occurred throughout the year but the biting densities varied with seasons, with the
dry season (without harmattan) having the lowest densities and the rainy season
having higher densities. This could be due to the provision of better breeding sites
for the blackflies as a result of increase in water levels and attachment objects that
usually occur during the rains (Crosskey, 1990). However, it was observed that the
highest fly densities were recorded during the harmattan season (Ubachukwu and
Anya, 2005). This could be attributed to the addition to local flies of migrating
savanna flies carried down into the area by the north-south harmattan winds blowing
from the sahara to the coasts (Boakye, 1999).
The numbers in which blackflies attack their hosts are enormously variable
and subject to many factors, such as emergence season, the proportion of blood
thirsty flies in the population and availability of suitable hosts in proximity to the
breeding sites or within range of blood-seeking appetitive flight (Crosskey, 1990).
Accurate assessment of biting rates (i.e. numbers of flies engorging blood or settling
to do so in a given time) is difficult on non-human hosts, especially as many flies are
often attracted to hosts such as poultry and livestock on which they land
intermittently without necessarily setting to bite.
1.2.4 Host Choice/Specificity
The Simuliidae are bloodsuckers on warm-blooded vertebrates and they bite
a wide range of birds and mammals. Blackfly larvae develop exclusively in running
waters and the annual number of blood-sucking blackflies that emerge from large,
unregulated boreal rivers is huge, possibly in the range of billions of individuals per
km of river. Therefore, investigating their impact on birds and mammals, including
blackfly species composition and host choice is important to describe the landscape-

xix
level interactions between an aquatic ecosystem and its terrestrial surroundings
(Polis et al. 1997; Malmqvist, 2002).
Blackflies and host have traditionally been linked by using various serological
methods (Hunter and Bayly, 1991), and by exposing potential hosts in cages insitu
(Hunter et al., 1993); these methods, however, are not suitable for wholesale
investigation of host choice for any blood-feeding insect in the wild. With the advent
of novel molecular techniques, better methods of studying blackfly-host interactions
have become available (Mukabana et al., 2002). In determining the vertebrate host
specificity of wild-caught blackflies, Malmqvist et al., (2003) demonstrated a
predominance of large hosts and marked discrimination between blackflies using
either avian or mammalian hosts.
Host choice is likely to be based on visual, olfactory and thermal cues,
providing the blackflies with information about host location and type (Sutcliffe,
1986), but may not always lead to specialization in a strict sense. For example,
blackflies may feed in a particular habitat such as the forest canopy or lakeshore,
with habitat taking precedence over the species of host. Recent studies on host
specialization in insects suggests that ecological host attributes, such as
microhabitat, phenology and host-finding constraints, may be decisive for host
preference (Janz and Nylin, 1997; Tompkins and Clayton, 1999; Stireman and
Singer, 2003). Since the most profitable hosts may be rare, acceptance of those of
lower rank may take place, which would also lead to the inclusion of a broader range
of host (Jaenike, 1990).
1.2.5 Adult Morphology
Adult female blackflies (Diptera: Simuliidae) have been well known as pests
to humans and animals, and they are also vectors of some parasites and

xx
pathogens, like Onchocerca spp. in humans and cattle and Leucocytozoon spp. in
birds (Mead et al., 1997; Stallings et al., 2002). Knowledge of the role of each vector
species in transmission is important for the rational design of disease-control
programmes (Mustapha et al., 2004). Unfortunately, the members of the S.
damnosum complex are extremely difficult to differentiate morphologically, and are
usually defined by specific chromosomal characters in the larvae (Vajime and
Dunbar, 1975, 1977; Crosskey, 1987).
However, it is still important to know the morphologies of the species of the S.
damnosum complex as this would aid in the control of the vector. From the studies
of Davies et al., (1988) and Wilson et al., (1993), the morphologies of adult S.
damnosum s.s. and S. squamosum are already well known. An adult female S.
yahense appears to be easily distinguishable from other adult female S. damnosum
s.I. because of the all-dark setae on the ninth abdominal tergite (Wilson et al., 1993;
Post et al., 2003; Fryauff and Trpis, 1986). In a similar study, Mustapha et al., (2004)
gave a morphological description of the S. mengense found in Cameroon. According
to this study however, the all-dark setae of the ninth abdominal tergite used to
distinguish the adult females of S. yahense from all other West African S.
damnosum s.I. vectors was rendered unusable in Southern Cameroon because the
adult female S. mengense examined also had this character.
Generally, in comparison with those of the other S. damnosum s.I. examined
by Mustapha et al., (2004), the fourth and fifth antennal segments of adult female S.
mengense are not compressed. The compression of the antennal segments is
known to be variable within the S damnosum cytospecies, however, and used alone
is not reliable in distinguishing individual specimens. The ratio of the length of the
thorax to the antenna is generally a more useful character (Garms, 1978). However,
since the range of the thorax length to antenna length for the adult female S.

xxi
mengense (1.7-2.3) overlaps with that of the adult female S. yahense of the ‘Bioko’
form (1.7-2.09) (Post et al., 2003), this ratio cannot be used to separate the two
species.
Although the basal tooth of the fore-tarsal claw of female S. mengense
appears distinctly longer and wider than that of the females of the ‘Bioko’ form, not
enough specimens have been examined to determine how this character varies
within S mengense, and so assess the value of the character for species
identification. In a comparative survey of the morphology of the fore-tarsal claw of
adult females, Kruger et al., (1998) found that nearly all the species they examined
had a smaller basal tooth than was observed in the new specimens of S. mengense
investigated. The two exceptions were S. soubrense and S. pandanophilum, which
had a basal tooth approximately the same size as, or larger than that of the S.
mengense, respectively.
Furthermore, Kruger (2006), in a study on the indicative property of tarsal
claws reported that females of some populations of the West African S. sanctipauli
subcomplex which is the only exclusive West African subcomplex of the S.
damnosum complex (Boakye, 1993), were found to have enlarged basal teeth of
their tarsal claws. This indicated zoo-/ornithophilic behaviour since tarsal claw
morphology is closely linked with ability of simuliids to penetrate either mammal fur
or avian feathers. In West Africa, the majority of populations and taxa of the S.
damnosum complex are incriminated vectors of O. volvulus (Boakye et al., 1998)
and hence are regarded as anthropophilic, but some observations exist that point
towards certain degrees of zoophily. Here, the character of interest was size of the
claw’s basal tooth relative to that of the entire claw. In order to measure this
morphological trait, a ratio (claw index) was introduced that consists of the distance
between the tarsal notch and the apical tip of the claw (Kruger, 2006).

xxii
In general, S. damnosum s.I. is known to exhibit small, conical teeth on claws
that are intermediate between the toothless claws of certain (mammalophilic)
Simulium species such as S. (metomphalus) hargreavesi (Freeman and De Meillion,
1953) and the prominently toothed claws found in many (ornithophilic) Simulim
species. However, a few members of the S. damnosum complex are also known for
their toothed claws. Such species are S. pandanophilum (Kruger et al., 1998), S.
juxtadamnosum and S. mengense (Mustapha et al., 2004).
The presence of hairs in the middle of the subcosta vein of adult female S.
mengense seems one of the best characters for distinguishing these flies from adult
females of the ‘Bioko’ form. In summary, the external morphology of the adult
females of S. mengense is generally similar to that of the ‘Bioko’ form of S. yahense
and only two characters (presence of hairs on the subcosta vein and basal-claw
morphology) might have value in distinguishing between the two (Mustapha et al.,
2004).
Also, in distinguishing S. mengense from the other S damnosum s. I. vectors,
it was noted that the colour of the wing tuft and of setae on the ninth abdominal
tergite are the most important characters because S. damnosum s.s. has
predominantly pale setae whilst S. squamosum has a mixture of pale and dark
(often mostly pale) wing tufts and pale setae on its ninth abdominal tergite (Garms,
1978; Wilson et al., 1993). As they are non-biting, the morphology of adult male S.
damnosum s.I. is important mainly as a confirmation of the identity of the females.
The male scutal pattern of S. mengense was constant in all of the adults examined
and belongs to the category-II group described by Meredith et al., (1983). The scutal
pattern of most males of the “Bioko” form is similar to type IIIh, the rest being
classified as type IIIg. The male scutal pattern, coupled with other differences
(presence of hairs on adult female subcosta vein and tufts of hair-like scales on

xxiii
larval thoracic cuticle) should enable S. mengense to be separated from the ‘Bioko’
form in the field.
In studying the Bioko form of S. yahense, Post et al., (2003) described the
adult females as having entirely dark wing tufts, entirely dark basicostal setae,
entirely dark postcranial hairs, a dark arculus and dark fore coxae. Furthermore, the
colour of the setae of the scutellum and ninth abdominal tergite was all dark and the
values of the thorax length ratios for females ranged from 1.77 to 2.07, most being
in the 1.90-2.00 range, consistent with other populations of S. yahense (Garms and
Zillman, 1984). The antenna of all females lacked compression, consistent with
membership of the forest group of flies and were uniformly dark, except for the basal
two segments and one-third to one-half of the third segment. Most of the males
studied has scutal patterns of type IIIh and the rest type IIIg, using the terminology
described by Meredith et al., (1983) and were distinguished from the members of
the S. sanctipauli subcomplex which usually have type I or type II scutal patterns.
In conclusion the most practical procedure routinely available for
distinguishing between species of adult females belonging to the S. damnosum
complex is the assessment of micro morphological characters (Dang and Peterson,
1980). Mafuyai et al., (1996) reported that for monitoring southward invasion by
savanna vectors of onchocerciasis in Nigeria, it is convenient and reasonably
reliable to identify adult females by means of morphometric differences between
sibling species of the S. damnosum complex.
1.2.6 Larval Structure and Function
Knowledge about all life stages of an organism is important to clarify
taxonomic problems, particularly in blackflies where there are many species
complexes. In common with the larvae of other Diptera, larval blackflies have no

xxiv
legs. Their general form is worm-like, but they are among the most easily
recognized insect larvae because of certain obvious structural features. The shape
is very important and shows three principal features: a pair of large cephalic fans on
the head and two “false feet” or pseudopods known as prolegs. The cephalic fans
are feeding organs and the prolegs, organs of attachment (Crosskey, 1990).
In larvae of some species, the body bears various modified cuticular hairs
and scales (macrotrichia) articulated to sockets in the cuticle. The cuticular
macrotrichia have been studied mainly in tropical African and Japanese species
(Crosskey, 1960; Matsuo and Uemoto, 1976). The macrotrichia vary greatly in size
and shape, some being small and simple hair-like setae, and others forked or multi-
branched hairs, fan-like scales or large blade-like or spatulate scales (Crosskey,
1990).
The ground colour of the head ranges from creamy white to honey yellow,
red-brown and blackish brown and can vary considerably in the same species. The
thorax and abdomen often appear to the naked eye yellowish or reddish, light to
dark brown, dark grey, purplish or almost black. The thoracic and abdominal colours
are quite different from the melanization responsible for head patterns and other
darkened parts of the cuticle and are caused by pigment chromatocytes.
According to Hinton (1959), the chromatocytes are usually closely gathered
into continuous cellular sheets on the upper body surface, but elsewhere they occur
scattered in an open meshwork connected to one another by long cytoplasmic
threads. Besides storing pigment granules that provide the larva with protective
colouration, the chromatocytes also contain fat globules. They were also believed to
be modified fat-body cells.
The external structures of the head comprise the cranium or head capsule
and the appendages and sense organs that are attached to it or borne on its

xxv
surface, viz the mouthparts, the antennae and the stemmata. The mouthparts of
normal larvae with cephalic fans have been much investigated, and good accounts
of them, based on conventional light microscopy, have been published by Chance
(1970) and others. The cephalic fans are large paired structures rooted anteriorly
and dorsally to the head above the mandibles and immediately in front of the
antennae. Embryologically, they begin as lateral outgrowths of the labral region of
the head (Craig, 1969; 1974).
In an earlier work, Dumbleton (1962) stated that most Simuliid larvae possess
mouth brushes, which sweep the food from the rays of the mouth brush into the
mouth. The larvae of the subfamily Gymnopaidinae (genera Gymnopais and
Twinnia) have neither and they feed by a browsing action of the mandibles with the
epipharyngeal hairbrushes acting as auxiliaries. The larvae of Cnephia crozetense
and S. oviceps both possess mouth brushes but these are much reduced in oviceps
and the short rays in Crozetense are toothed only at the apices.
1.2.7 Phenotypic Plasticity in Simulium Larvae
In a broad sense, phenotypic plasticity may be defined as environmentally
induced phenotypic variability of a genotype (Bradshaw, 1965; Stearns, 1989; West-
Eberhard, 1989), which may be adaptive (Houston and McNamara, 1992;
McNamara and Houston, 1996). Since no single phenotype is optimal under all
environmental conditions (McNamara, 1998), organisms often produce
environmentally specific phenotypes to adapt to variable conditions for a success in
habitats with high environmental variation (Agrawal, 2001; Piersma and Drent,
2003). Phenotypic plasticity is the conditional expression of a given genotype to
produce alternative phenotypes depending on the environment during ontogeny
(Nylin and Gotthard, 1998).

xxvi
Many small aquatic animals possess filters to capture food particles from
water. The performance of these feeding structures relies on the hydrodynamic
conditions around them. Cheer and Koehl (1987) developed a model for fluid motion
through a filter composed of a finite number of bristles and predicted that a simple
modification in the size of the filter may influence greatly flow moving through it.
Blackfly larvae are important components of streams and rivers worldwide. They
often are the predominant suspension feeders in these systems and their densities
may be astoundingly high, reaching 120 larvae cm-2 or more in Lake Outlet streams
(Wotton, 1987).
Current velocity is one of the most important physical factors for lotic animals
in its influence on morphological and behavioural adaptations, and ecological
distributions (Allen, 1995). Together with other aspects of fluid dynamics, it plays an
important role in the evolution of the feeding structures used by aquatic suspension
feeders (LaBarbera, 1991). Blackfly larvae attach on the substratum by a posterior
circlet of hooks anchored in a silken pad and use a pair of labral fans to remove
suspended food particles from the water.
Current velocity affects particle capture of the larvae. In fast currents, the
water flux past the larval fans is great and many particles are provided to the fans
per unit of time (Craig and Chance, 1982). At increasing current velocities, the drag
on the fans grows in the direction of flow (Vogel, 1994). In a previous study,
differences were found in the primary fan size, both between species and within
species, among populations inhabiting different kinds of running water habitats
(Zhang and Malmqvist, 1996). Larvae living in fast-flowing water had small fans,
stout fan rays and a short distance between individual rays, while those living in
slow-flowing water had larger fans with delicate, widely spaced rays.

xxvii
One important hydrodynamic force on blackfly larvae is drag. Drag on fans is
directly proportional to the fan area projected in the direction of flow (excluding
interray space) and the current velocity (Vogel, 1994). At high current velocities,
large fans with many rays and long fan stems experience higher drag than small
fans with few rays and short fan stems; therefore, the integrity of the large fans may
become disrupted (Lacoursiere and Craig, 1993), and suspension feeding
consequently reduced, or completely discontinued. The large fans of larvae reared
in a slow current had more delicate rays than those reared in fast currents. Drawing
from general hydrodynamic principles, skin friction (viscous forces) on fans is more
important than pressure drag at slow velocities (Vogel, 1994).
In an experiment, Zhang and Malmqvist (1997) showed a clear morphological
response in the fan structure of S. lundstromi to long-time exposure to different
velocities, which suggests that hydrodynamic factors may be powerful components
of natural selection in blackfly larvae. In a related study aimed at examining the
hypothesis that fan morphological plasticity of blackfly larvae could arise as a result
of interaction of the feeding structure and surrounding habitat, it was demonstrated
that both current velocity and food concentration can have strong overall effects on
the fan structure, growth rate and developmental time of S. noelleri blackfly larvae
(Zhang, 2006). Food concentration had stronger effects on ray numbers whereas
current velocity has stronger effects on fan area. This pattern is related to the
biological filtration theory (Cheer and Koehl, 1987).
Lucas and Hunter (1999) demonstrated that the ray number of S. rostratum
and S. decorum decreased with food supply increase in a laboratory experiment.
Morphological adaptations enable feeding at different flow regimes by balancing
increasing particle capture in slow currents and reducing drag force cost on fans in
fast currents (Zhang, 2000). The significance of interspecific and intraspecific

xxviii
variation in fan structure is obvious for blackfly larvae occupying various types of
lotic habitats (Zhang and Malmqvist, 1996; Malmqvist et al., 1999; Palmer and
Craig, 2000). This pattern can be explained using the theory of adaptive radiation,
which is the diversification of a taxon to exploit a variety of different resources by
differing morphological traits to utilize those resources (Huxley, 1942; Futuyma,
1986). The resource-based divergent selection on some phenotypic traits in different
environments is important for speciation to occur (Losos et al., 1997; McKinnon et
al., 2004).
Although blackfly species in different lotic habitats differ markedly in fan
structure (Zhang and Malmqvist, 1996; Palmer and Craig, 2000), the underlying
biomechanical factors influencing fan traits apply equally in different species (Cheer
and Koehl, 1987; Hart et al., 1991; Lacoursiere and Craig, 1993; Zhang and
Malmqvist, 1997).
In conclusion, experimental results illustrate a strong correlation between
environmental changes in food availability and current velocity, phenotypic plasticity;
particle flux rate (PFR) and growth rate of blackfly larvae. Compared to the current
velocity, food availability induced stronger
impact on fan phenotypic plasticity and growth (Zhang, 2006). This suggests that
food-current velocity induced phenotypic plasticity is a crucial feature for blackfly
larvae to occupy spatially and temporally heterogenous habitats. This is likely to be
one explanation for their success as members in streams and rivers throughout the
world.
1.2.8 Cytotaxonomy of The Simulium damnosum Complex
The ability to properly identify an insect of medical-veterinary importance is
the most important step in developing biological control for that particular insect.

xxix
Correct identification is conducive to determining the types of problem they cause
and, subsequently, in implementing control programs (Mullen and Durden, 2002).
However, while the importance of identification cannot be stressed enough, the
ability to do so is not always so easily obtained. Taxonomy uses characters of the
organism to recognize the individuality of a species or the similarity of organisms
forming a taxon. Traditional taxonomic work has focused on the use of
morphological characters while more modern techniques have come to rely on
molecular data.
Identification to species level in blackflies is complicated by morphological
uniformity. Therefore, a broad range of data must be examined, compiled from
different morphological data during the life cycle stages, polytene chromosomes,
distributional and ecological data. While resolving taxonomic questions is the most
obvious use for cytotaxonomy, there are several other implications concerning the
field of medical entomology. Polytene chromosomes permit a high degree of
taxonomic resolution, as far as the population level (Adler et al., 2004). The
taxonomic resolution of species complexes currently depends mainly upon the
techniques of the cytotaxonomist studying the polytene chromosomes of the larval
simuliid salivary glands. In general, these are the only taxonomically usable
chromosomes in blackfly tissues, although Bedo (1976) has found easily
demonstrable polytene chromosomes in pupae and adults of some Australian
simuliidae.
Cytotaxonomy is now a well established major arm of simuliid taxonomy in
which one of its special virtues is the clarification of species-complexes. Rothfels
(1979) has provided a masterly up-to-date exposition of its capabilities and its
findings. The work of the cytologist on species complexes have been supplemented
by studies on isoenzymes, thus bringing a truly non-morphological approach to bear

xxx
on simuliid complexes. Potentially very important advances in the taxonomy of the
Simulium damnosum complex as a result of enzyme electrophoresis have been
reported.
The application of molecular techniques was first reported in West Africa
(Brockhouse et al., 1993, Tang et al., 1996) and was useful in separating four out of
five species of the S. damnosum complex from Eastern Africas (Kruger et al., 2000).
1.2.9 Control Measures
There are two main approaches to onchocerciasis control and an integration
of both may be the best tool to control the disease. One approach is the reduction of
the parasite burden in humans by chemotherapy using ivermectin; at present, this is
the main method of onchocerciasis control. Although chemotherapy has a beneficial
effect on disease pathology, it is not certain that used on its own, it can interrupt
parasite transmission (Boatin et al., 1998; Borsboom et al., 2003). The second
approach is to interrupt parasite transmission by long-term vector control or by
vector eradication, both of which have already been proven to be effective (Davies,
1994).
Several large-scale projects for larval blackfly suppression have been
conducted in different parts of the world. Since these began, the concept of pest
eradication has been modified to one of ‘control’. However, the success of vector
eradication has been seen to depend on two criteria:
(1) The accessibility of Larval breeding sites for larviciding, and
(2) The isolation of these sites from immigrant vectors (Garms et al., 1989).
The major control effort undertaken so far has been the Onchocerciasis
Control Programme (OCP) initiated in 1974 in West Africa (Samba, 1994) which was
mainly based on vector control with larvicides. From 1989, it was combined with the

xxxi
large scale, free-of-charge distribution of the microfilaricidal drug ivermectin to the
human population. Such distributions now cover 19 countries in Africa under the
African Programme for Onchocerciasis Control (APOC) (Benton, 1998).
Vector eradication has been achieved in some isolated foci in East Africa
(Davies, 1994) and for some Simulium species on the West African mainland, during
the very widespread and intensive activities of the World Health Organization’s
Onchocerciasis Control Programme in West Africa (Fiasorgbor et al., 1992). Few
other places have been known to meet the required criteria (Mustapha et al., 2004).
The distribution of breeding sites on Bioko has been described by McCall et
al., (1998) with special reference to their accessibility to possible vector control
operations but very little is known about the potential geographical isolation of the
Bioko population of vectors (Post et al., 1995). The major thrust of blackfly control
campaigns has been directed against the larvae, which are confined to a highly
restricted, easily treated habitat, running water. Adult blackfly control, although
effective in limited areas and for special purposes, is too costly for general use.
DDT, which had been the major chemical used in blackfly control, was banned
because of its adverse long-term consequences to the environment. Consequently,
two effective substitutes were discovered for blackfly control – temephos and
methoxychlor. It has been shown that both temephos and methoxychlor have been
effective against blackfly larvae (Wallace et al., 1976).
Blackflies have started developing some levels of resistance to temephos and
this necessitated research on a number of possible alternatives including other
larvicides and formulations and biological control agent (Lacey and Mulla, 1978;
Thompson and Adams, 1979). The clinical effects of the microfilaricidal drug
invermectin have been monitored but the parasitological effects have not been
assessed (Mas et al., 1995).

xxxii
There have been attempts at controlling onchocerciasis using biological
agents targeted against the Simulium larva. The principal agent used is Bacillus
thuringiensis var. israelensis (B.t.i). This is a safe and effective biological agent
ideally suited for integrated pest management programmes. The use of products
based on Bti has been effective in both small streams and large rivers (Molloy,
1990). However, it is still inadequate as there is still need to control adult flies as
well (Wegner, 2006).
There are undoubted clinical benefits of ivermectin treatment although it is
generally not clear whether ivermectin on its own can interrupt transmission of the
parasite (Boatin et al., 1998). Hence, the World Health Organization African
Programme for Onchcericasis Control (APOC) has a strategy not only to support
ivermectin distribution in onchocerciasis-endemic countries but also to effect focal
vector eradication where appropriate in isolated areas (Remme, 1995).
1.3 Objectives of the Study
The objectives of this study are
1. To examine the morphometric characteristics of the S. damnosum complex
in Ogurugu and Nkpologu.
2. To identify the sibling species present in these areas.
3. To compare the findings with those from cytological studies.
4. To relate fly types to the types of onchocerciasis present in the two areas.

xxxiii
CHAPTER TWO
MATERIALS AND METHOD
2.1 Study Area
The study areas were Nkpologu and Ogurugu in Uzo-Uwani Local Government Area
of Enugu State, Nigeria. Ogurugu lies between latitude 60 45′ and 60 50′ North and
longitude 60 55′ and 70 00′ East while Nkpologu lies between latitude 60 40′ and 60
50′ North and longitude 70 00′ and 70 16′ East. They both belong to the forest-
savanna mosaic vegetation zone of Nigeria (Crosskey, 1981). Ogurugu community is
traversed by rivers Eshi and Okpo while Nkpologu is traversed by Adada River.
These rivers are tributaries of the Anambra River. Farming is the major economic
activity of the area.
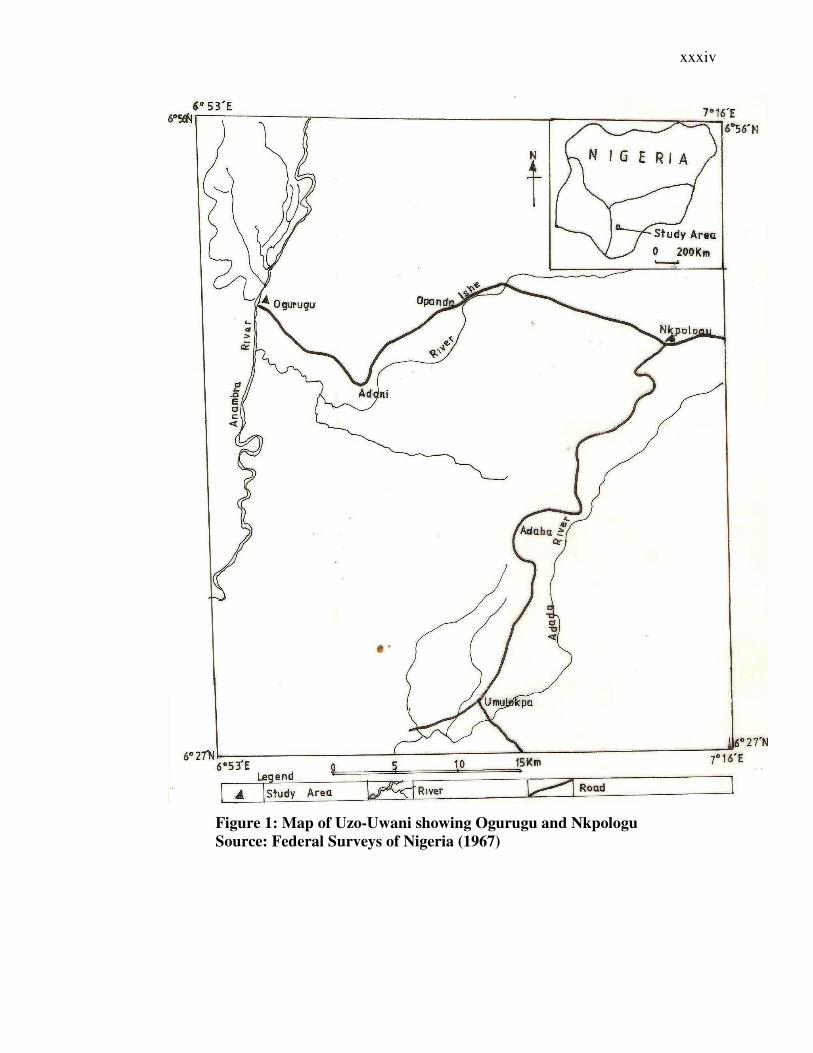
xxxiv
Figure 1: Map of Uzo-Uwani showing Ogurugu and Nkpologu
Source: Federal Surveys of Nigeria (1967)

xxxv
2.2 Epidemiological Studies
2.2.1 Questionnaire Administration
A survey to determine the epidemiology of onchocerciasis in the 2 communities was
conducted. Questionnaires were first pre-tested on students of Solid Rock
International Academy, Nkpologu on 23rd September, 2008. Questionnaires were
randomly administered on the residents of the communities. A total of 303 (121
males & 182 females) and 400 (175 males & 225 females) respondents were sampled
in Ogurugu and Nkpologu communities respectively. The questionnaire tested their
knowledge of blackflies, onchocerciasis and Mectizan.
2.2.2 Clinical Examination
The respondents were also physically examined to check for the presence of
onchocercal symptoms like nodules, leopard skin, lizard skin and onchodermatitis.
The necessary records were taken.
2.2.3 Visual Acuity Testing (VAT)
The standard Snellen illiterate ‘E’ optotype chart was used at 6 meters distance to test
both eyes together. The results were classified into Normal, Impaired or Blind
categories.
i. Normal sight: Ability to read 6/60 chart or show direction of alphabet at 6
meters.
ii. Impaired sight: inability to perform the above activities at 6m but at 3m.
iii. Blindness: Inability to carry out activities (i) above at 3m.

xxxvi
2.3 Fly Collection
Fly collection took place between December 2008 and January 2009. The flies were
usually collected during the daily peak biting periods (9.00am – 10.00am & 5.00pm –
6.00pm) as reported by Ubachukwu and Anya (2001).Two main locations were used
for fly collection in Nkpologu. The flies were collected using human bait as the
attractant. The human bait was treated with a dose of Mectizan as a preventive
measure. The human baits sat in the fly-catching locations with their lower legs
exposed. Blackflies landing on the baits for a blood meal were caught and preserved
in absolute ethanol for further studies in the laboratory.
2.4 Morphometric Studies On Blackflies
Seventy-five (75) flies were subjected to morphometric examinations. The Thorax
length, Antenna length, Wing length, Wing Width and Femur length were measured
and the data obtained were transformed into ratios. The preserved flies were first
rinsed with distilled water and fixed with glycerine. The slides were then viewed
under an electric binocular microscope and the different parts were measured using
an ocular and stage micrometer gauge and recorded.
2.5 Statistical Analysis
The results from the morphometric studies were subjected to multivariate analysis
(Discriminant Function Analysis) while the results from the epidemiological survey
were subjected to Chi Square test of independence.

xxxvii
CHAPTER THREE
RESULTS
3.1 Epidemiology of Onchocerciasis In Nkpologu And Ogurugu
Communities
The epidemiological status of both Nkpologu and Ogurugu communities were studied using
the 5 major symptoms of the disease – presence of palpable nodules, leopard skin, lizard
skin, onchodermatitis and measurement of visual acuity.
3.1.1 Presence of Palpable Nodules
Plate 1: Head Nodules in an Individual
The prevalence of palpable nodules was studied in both communities. In Nkpologu, the total
prevalence of nodules was 5.8%. The prevalence of nodules among males was 8.6% while
the females had a lower prevalence rate of 3.6%. The prevalence of nodules among the
different age groups are shown in Table 1 below. The differences in the prevalence rates of
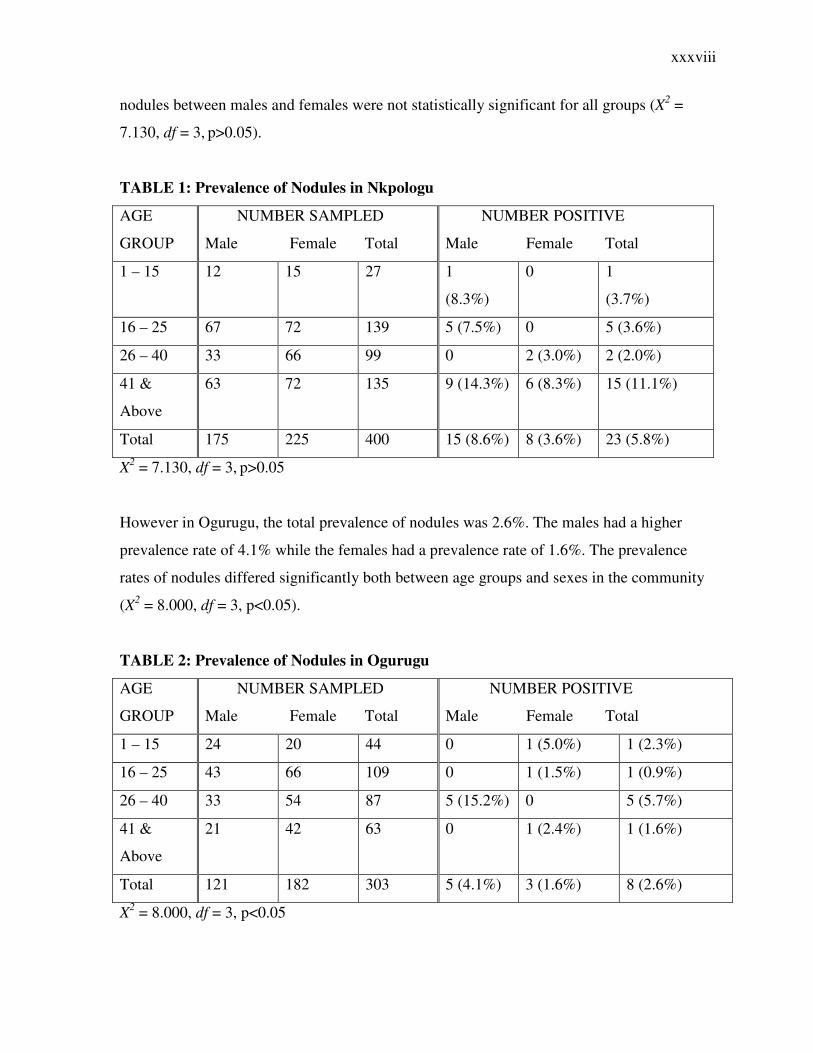
xxxviii
nodules between males and females were not statistically significant for all groups (X2 =
7.130, df = 3, p>0.05).
TABLE 1: Prevalence of Nodules in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 1
(8.3%)
0 1
(3.7%)
16 – 25 67 72 139 5 (7.5%) 0 5 (3.6%)
26 – 40 33 66 99 0 2 (3.0%) 2 (2.0%)
41 &
Above
63 72 135 9 (14.3%) 6 (8.3%) 15 (11.1%)
Total 175 225 400 15 (8.6%) 8 (3.6%) 23 (5.8%)
X2 = 7.130, df = 3, p>0.05
However in Ogurugu, the total prevalence of nodules was 2.6%. The males had a higher
prevalence rate of 4.1% while the females had a prevalence rate of 1.6%. The prevalence
rates of nodules differed significantly both between age groups and sexes in the community
(X2 = 8.000, df = 3, p<0.05).
TABLE 2: Prevalence of Nodules in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 24 20 44 0 1 (5.0%) 1 (2.3%)
16 – 25 43 66 109 0 1 (1.5%) 1 (0.9%)
26 – 40 33 54 87 5 (15.2%) 0 5 (5.7%)
41 &
Above
21 42 63 0 1 (2.4%) 1 (1.6%)
Total 121 182 303 5 (4.1%) 3 (1.6%) 8 (2.6%)
X2 = 8.000, df = 3, p<0.05
xxxix
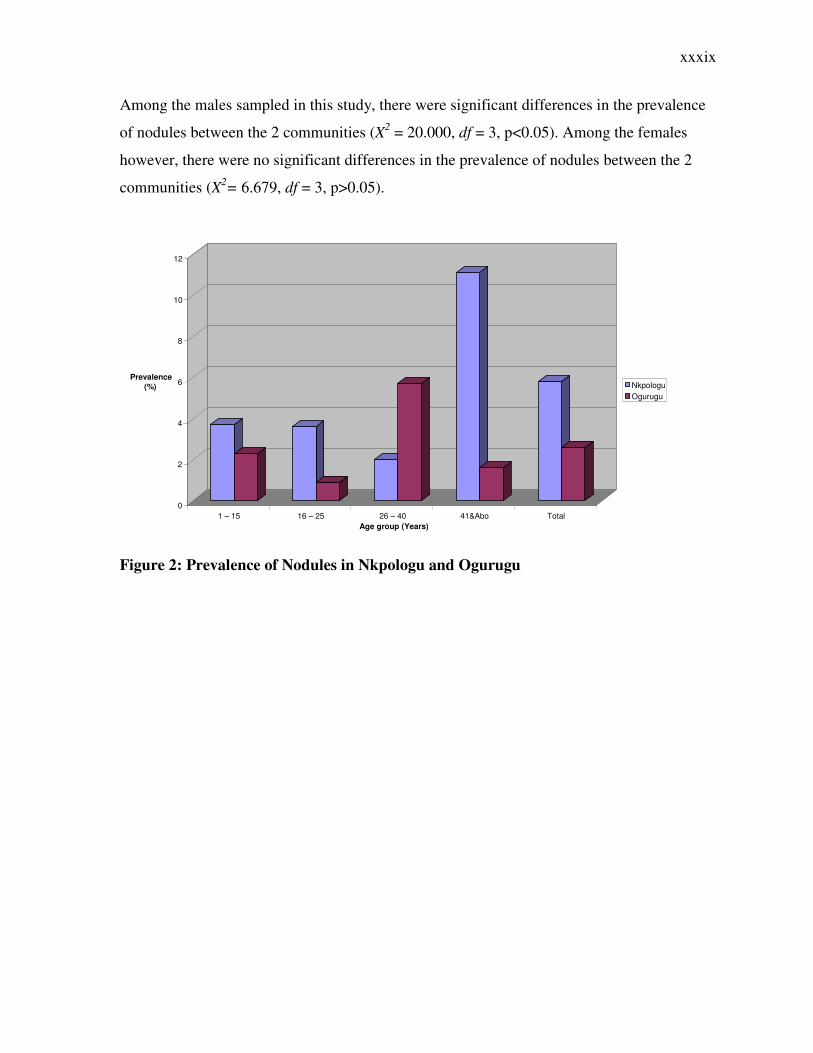
Among the males sampled in this study, there were significant differences in the prevalence
of nodules between the 2 communities (X2 = 20.000, df = 3, p<0.05). Among the females
however, there were no significant differences in the prevalence of nodules between the 2
communities (X2= 6.679, df = 3, p>0.05).
Figure 2: Prevalence of Nodules in Nkpologu and Ogurugu
0
2
4
6
8
10
12
Prevalence
(%)
1 – 15 16 – 25 26 – 40 41&Abo Total
Age group (Years)
Nkpologu Ogurugu

xl
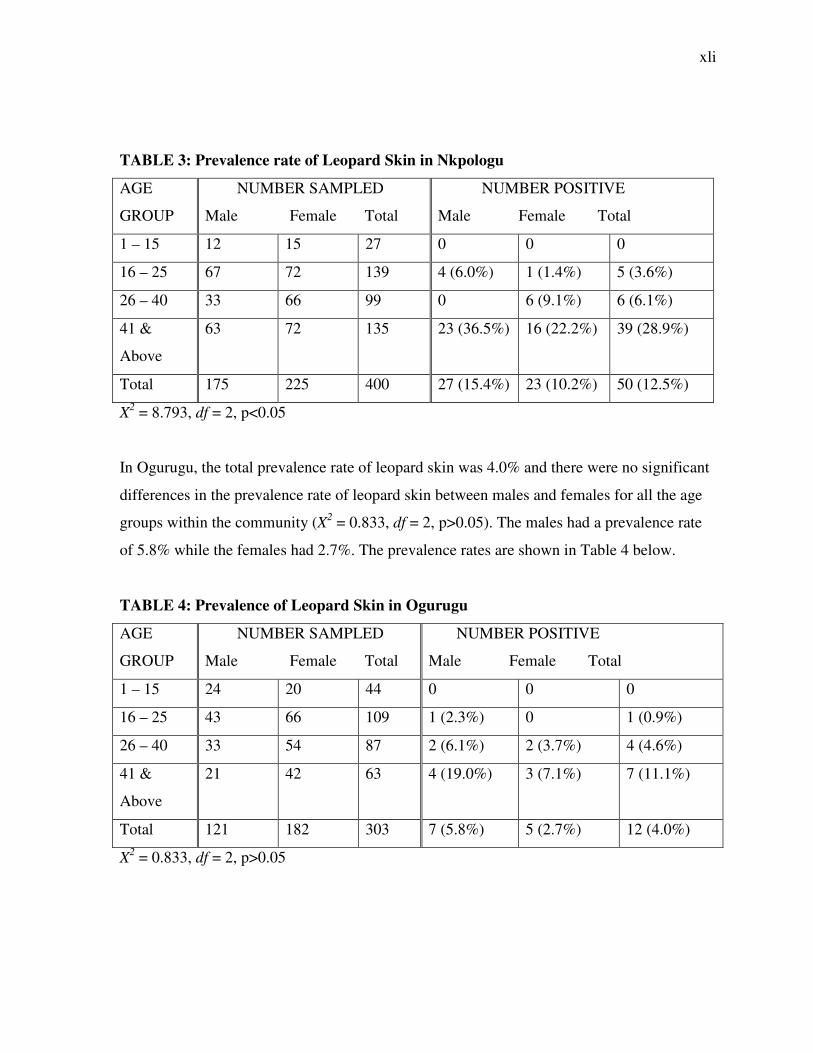
3.1.2 Presence of Leopard Skin
Plate 2: Leopard Skin in an Individual
The 2 communities were also studied for the presence of leopard skin. The result from
Nkpologu community shows that there was a total prevalence rate of 12.5%. The males had a
higher prevalence rate of 15.4% compared to the females which recorded a prevalence rate
of 10.2%. There were also significant differences in the prevalence rates of leopard skin
among the different age groups for the 2 sexes (X2 = 8.793, df = 2, p<0.05). Ages 41 & above
recorded the highest prevalence of 28.9%.
xli
TABLE 3: Prevalence rate of Leopard Skin in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 0 0 0
16 – 25 67 72 139 4 (6.0%) 1 (1.4%) 5 (3.6%)
26 – 40 33 66 99 0 6 (9.1%) 6 (6.1%)
41 &
Above
63 72 135 23 (36.5%) 16 (22.2%) 39 (28.9%)
Total 175 225 400 27 (15.4%) 23 (10.2%) 50 (12.5%)
X2 = 8.793, df = 2, p<0.05
In Ogurugu, the total prevalence rate of leopard skin was 4.0% and there were no significant
differences in the prevalence rate of leopard skin between males and females for all the age
groups within the community (X2 = 0.833, df = 2, p>0.05). The males had a prevalence rate
of 5.8% while the females had 2.7%. The prevalence rates are shown in Table 4 below.
TABLE 4: Prevalence of Leopard Skin in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 24 20 44 0 0 0
16 – 25 43 66 109 1 (2.3%) 0 1 (0.9%)
26 – 40 33 54 87 2 (6.1%) 2 (3.7%) 4 (4.6%)
41 &
Above
21 42 63 4 (19.0%) 3 (7.1%) 7 (11.1%)
Total 121 182 303 7 (5.8%) 5 (2.7%) 12 (4.0%)
X2 = 0.833, df = 2, p>0.05
xlii
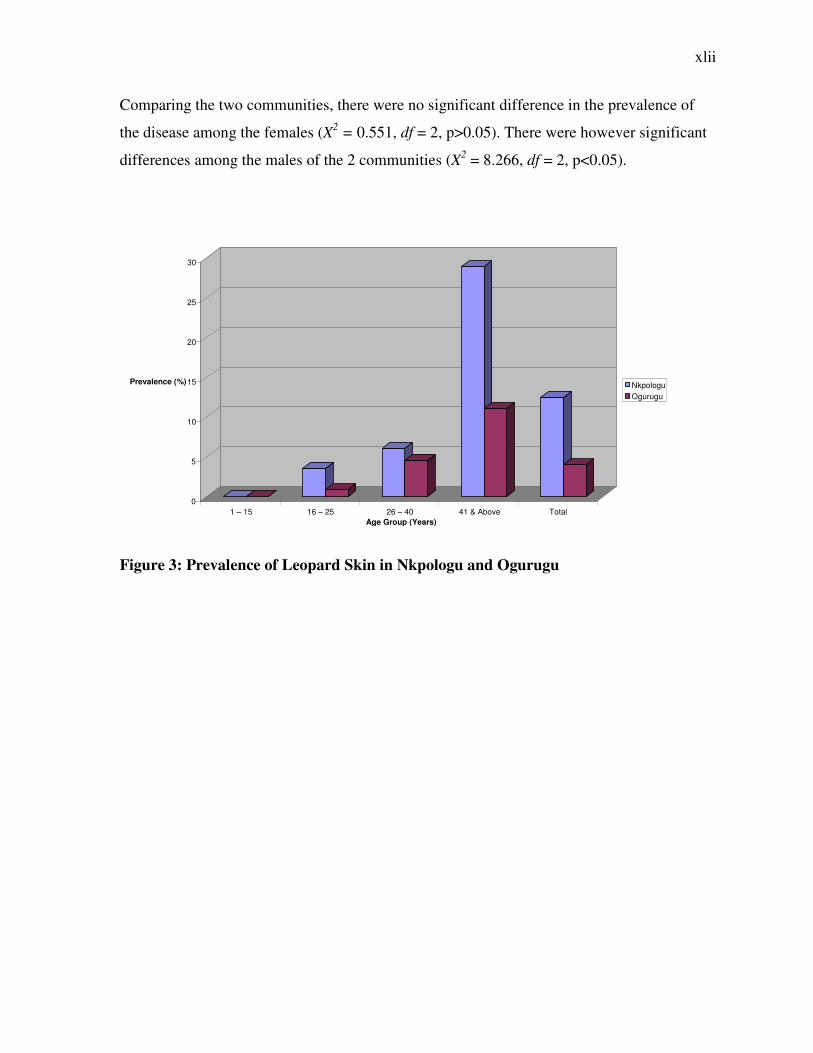
Comparing the two communities, there were no significant difference in the prevalence of
the disease among the females (X2 = 0.551, df = 2, p>0.05). There were however significant
differences among the males of the 2 communities (X2 = 8.266, df = 2, p<0.05).
Figure 3: Prevalence of Leopard Skin in Nkpologu and Ogurugu
0
5
10
15
20
25
30
Prevalence (%)
1 – 15 16 – 25 26 – 40 41 & Above Total
Age Group (Years)
Nkpologu Ogurugu
xliii
3.1.3 Presence of Lizard Skin
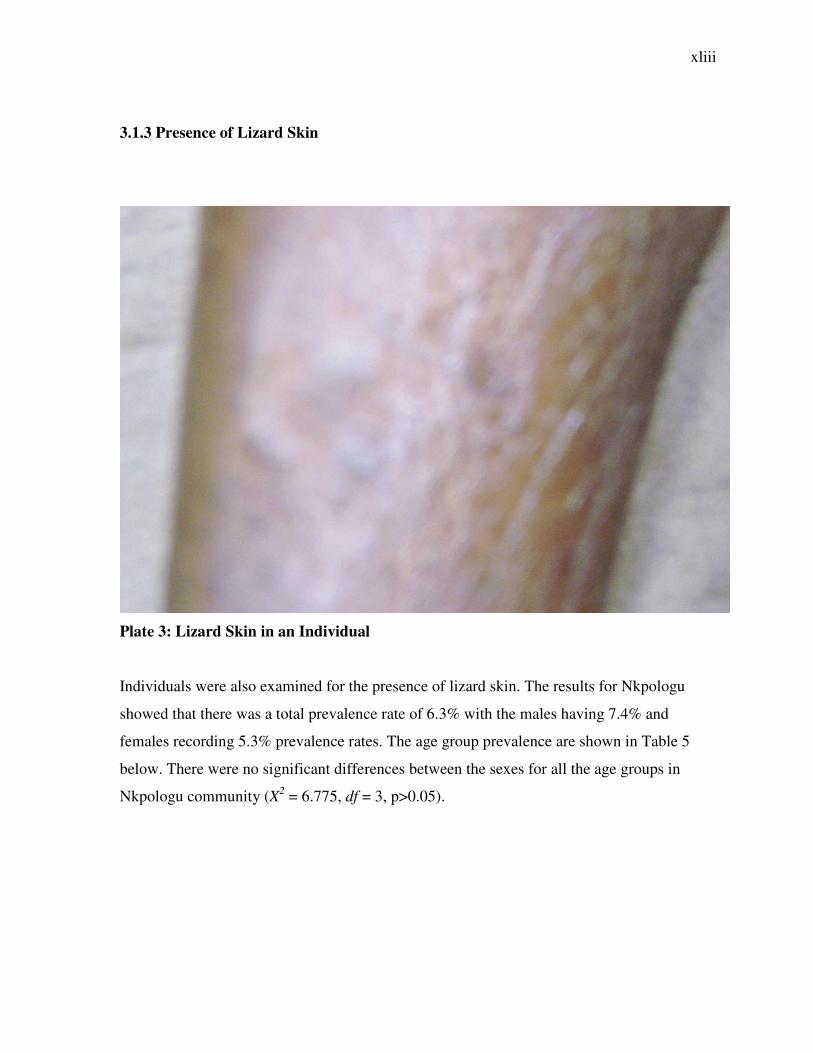
Plate 3: Lizard Skin in an Individual
Individuals were also examined for the presence of lizard skin. The results for Nkpologu
showed that there was a total prevalence rate of 6.3% with the males having 7.4% and
females recording 5.3% prevalence rates. The age group prevalence are shown in Table 5
below. There were no significant differences between the sexes for all the age groups in
Nkpologu community (X2 = 6.775, df = 3, p>0.05).
xliv
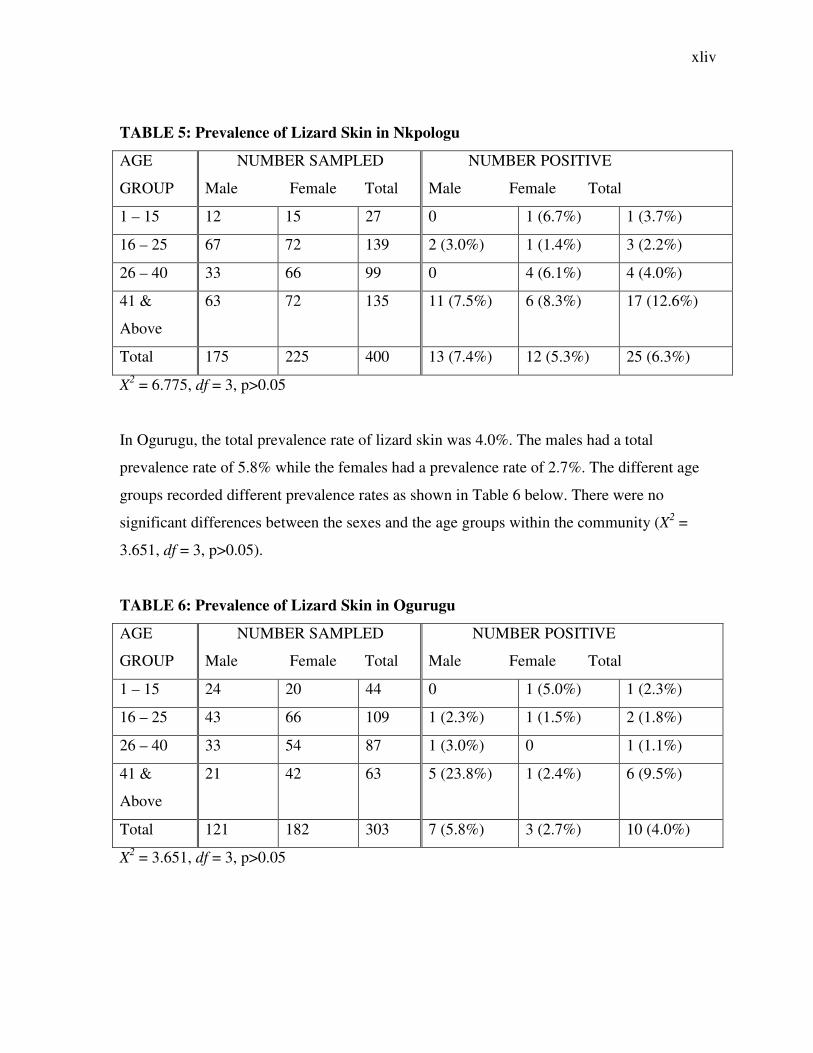
TABLE 5: Prevalence of Lizard Skin in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 0 1 (6.7%) 1 (3.7%)
16 – 25 67 72 139 2 (3.0%) 1 (1.4%) 3 (2.2%)
26 – 40 33 66 99 0 4 (6.1%) 4 (4.0%)
41 &
Above
63 72 135 11 (7.5%) 6 (8.3%) 17 (12.6%)
Total 175 225 400 13 (7.4%) 12 (5.3%) 25 (6.3%)
X2 = 6.775, df = 3, p>0.05
In Ogurugu, the total prevalence rate of lizard skin was 4.0%. The males had a total
prevalence rate of 5.8% while the females had a prevalence rate of 2.7%. The different age
groups recorded different prevalence rates as shown in Table 6 below. There were no
significant differences between the sexes and the age groups within the community (X2 =
3.651, df = 3, p>0.05).
TABLE 6: Prevalence of Lizard Skin in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 24 20 44 0 1 (5.0%) 1 (2.3%)
16 – 25 43 66 109 1 (2.3%) 1 (1.5%) 2 (1.8%)
26 – 40 33 54 87 1 (3.0%) 0 1 (1.1%)
41 &
Above
21 42 63 5 (23.8%) 1 (2.4%) 6 (9.5%)
Total 121 182 303 7 (5.8%) 3 (2.7%) 10 (4.0%)
X2 = 3.651, df = 3, p>0.05
xlv
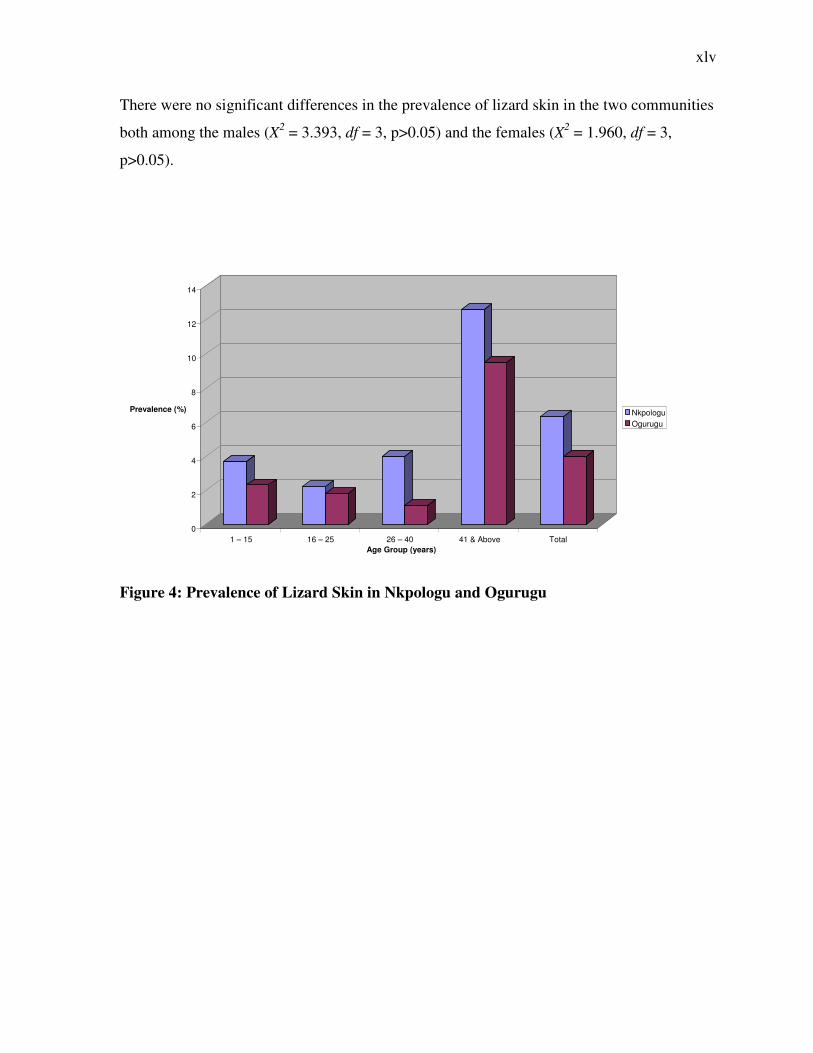
There were no significant differences in the prevalence of lizard skin in the two communities
both among the males (X2 = 3.393, df = 3, p>0.05) and the females (X2 = 1.960, df = 3,
p>0.05).
Figure 4: Prevalence of Lizard Skin in Nkpologu and Ogurugu
0
2
4
6
8
10
12
14
Prevalence (%)
1 – 15 16 – 25 26 – 40 41 & Above Total
Age Group (years)
Nkpologu Ogurugu
xlvi
3.1.4 Presence of Onchodermatitis
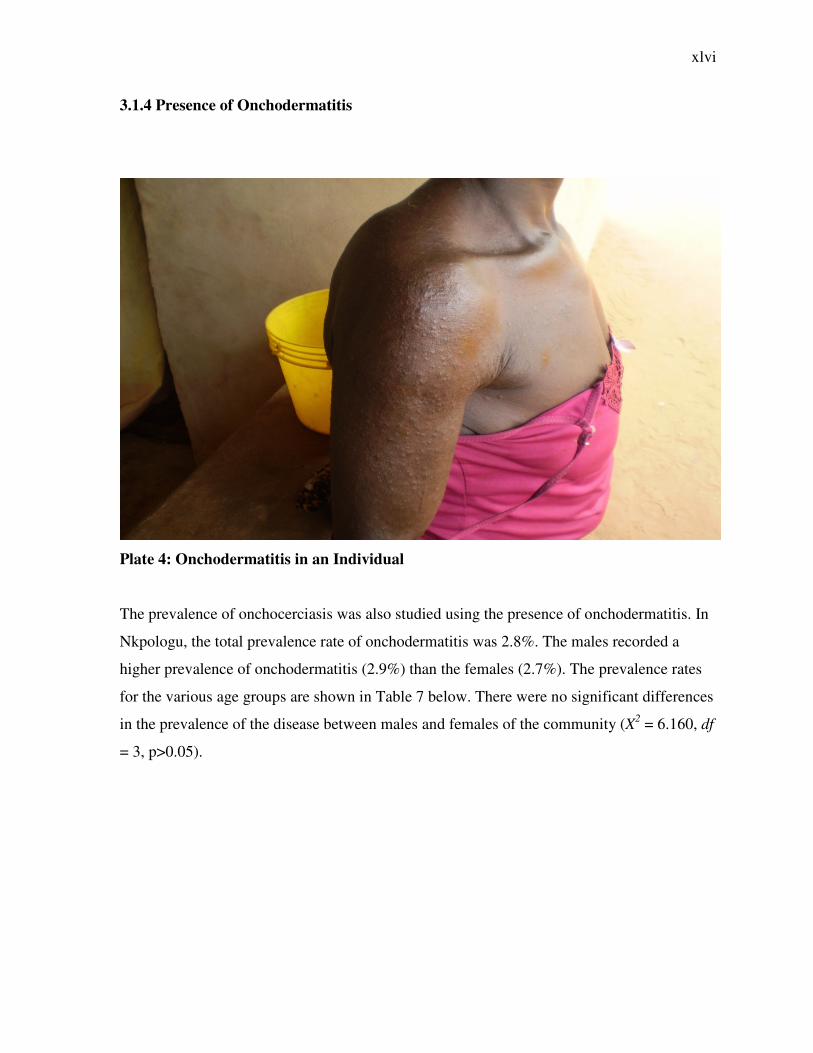
Plate 4: Onchodermatitis in an Individual
The prevalence of onchocerciasis was also studied using the presence of onchodermatitis. In
Nkpologu, the total prevalence rate of onchodermatitis was 2.8%. The males recorded a
higher prevalence of onchodermatitis (2.9%) than the females (2.7%). The prevalence rates
for the various age groups are shown in Table 7 below. There were no significant differences
in the prevalence of the disease between males and females of the community (X2 = 6.160, df
= 3, p>0.05).
xlvii
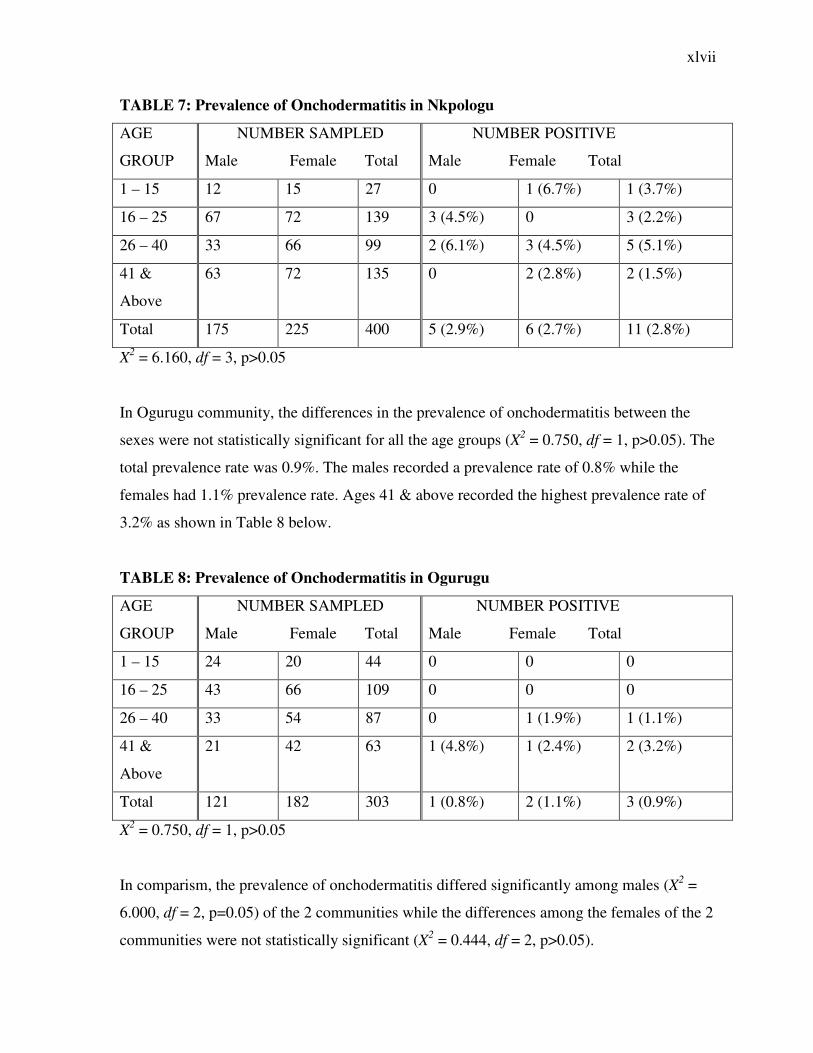
TABLE 7: Prevalence of Onchodermatitis in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 0 1 (6.7%) 1 (3.7%)
16 – 25 67 72 139 3 (4.5%) 0 3 (2.2%)
26 – 40 33 66 99 2 (6.1%) 3 (4.5%) 5 (5.1%)
41 &
Above
63 72 135 0 2 (2.8%) 2 (1.5%)
Total 175 225 400 5 (2.9%) 6 (2.7%) 11 (2.8%)
X2 = 6.160, df = 3, p>0.05
In Ogurugu community, the differences in the prevalence of onchodermatitis between the
sexes were not statistically significant for all the age groups (X2 = 0.750, df = 1, p>0.05). The
total prevalence rate was 0.9%. The males recorded a prevalence rate of 0.8% while the
females had 1.1% prevalence rate. Ages 41 & above recorded the highest prevalence rate of
3.2% as shown in Table 8 below.
TABLE 8: Prevalence of Onchodermatitis in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 24 20 44 0 0 0
16 – 25 43 66 109 0 0 0
26 – 40 33 54 87 0 1 (1.9%) 1 (1.1%)
41 &
Above
21 42 63 1 (4.8%) 1 (2.4%) 2 (3.2%)
Total 121 182 303 1 (0.8%) 2 (1.1%) 3 (0.9%)
X2 = 0.750, df = 1, p>0.05
In comparism, the prevalence of onchodermatitis differed significantly among males (X2 =
6.000, df = 2, p=0.05) of the 2 communities while the differences among the females of the 2
communities were not statistically significant (X2 = 0.444, df = 2, p>0.05).
xlviii
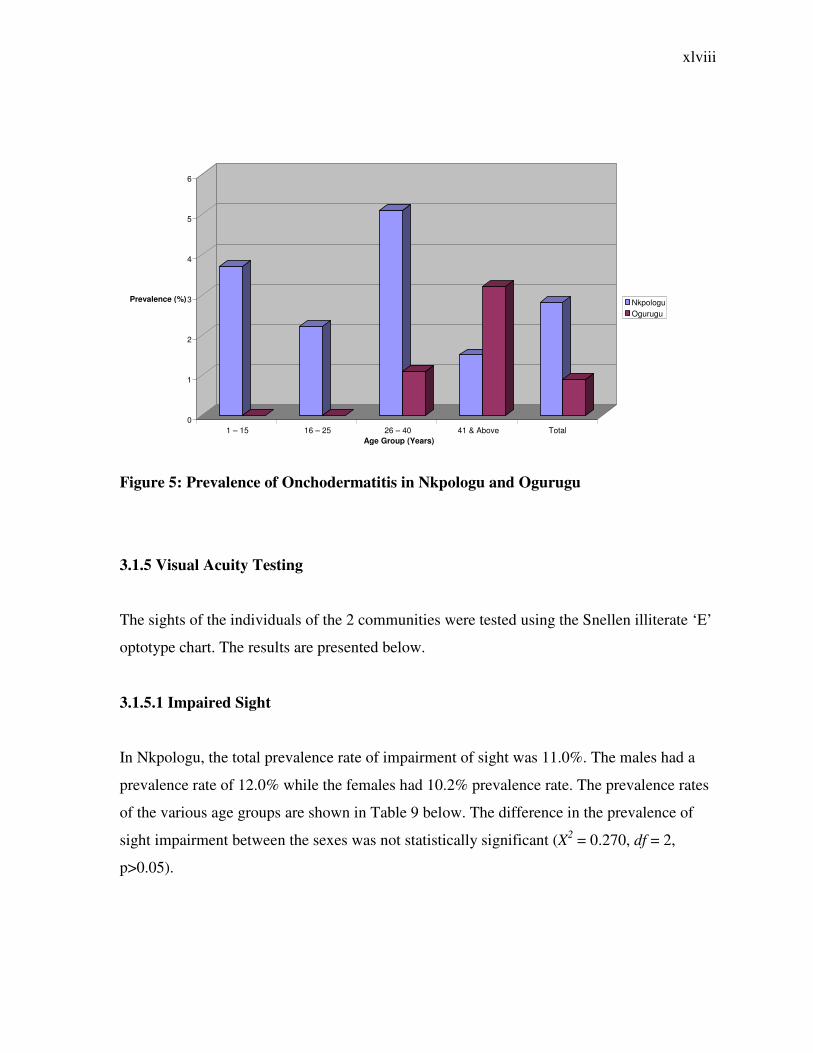
Figure 5: Prevalence of Onchodermatitis in Nkpologu and Ogurugu
3.1.5 Visual Acuity Testing
The sights of the individuals of the 2 communities were tested using the Snellen illiterate ‘E’
optotype chart. The results are presented below.
3.1.5.1 Impaired Sight
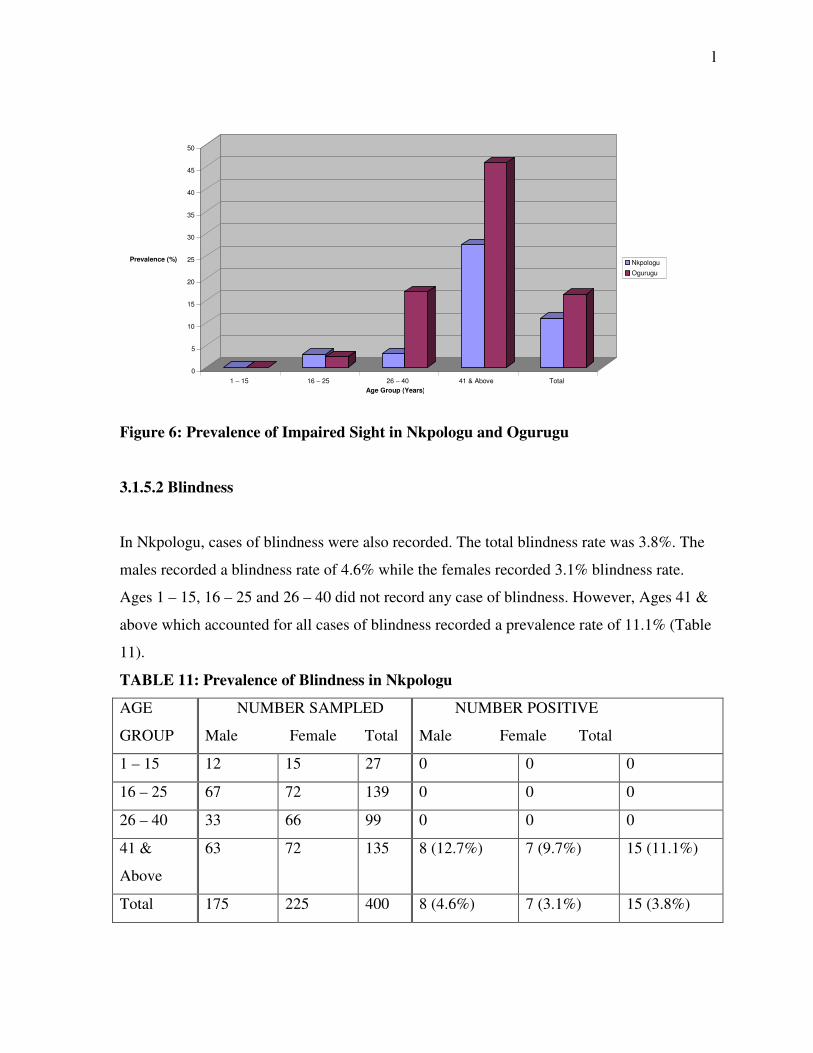
In Nkpologu, the total prevalence rate of impairment of sight was 11.0%. The males had a
prevalence rate of 12.0% while the females had 10.2% prevalence rate. The prevalence rates
of the various age groups are shown in Table 9 below. The difference in the prevalence of
sight impairment between the sexes was not statistically significant (X2 = 0.270, df = 2,
p>0.05).
0
1
2
3
4
5
6
Prevalence (%)
1 – 15 16 – 25 26 – 40 41 & Above Total Age Group (Years)
Nkpologu Ogurugu
xlix
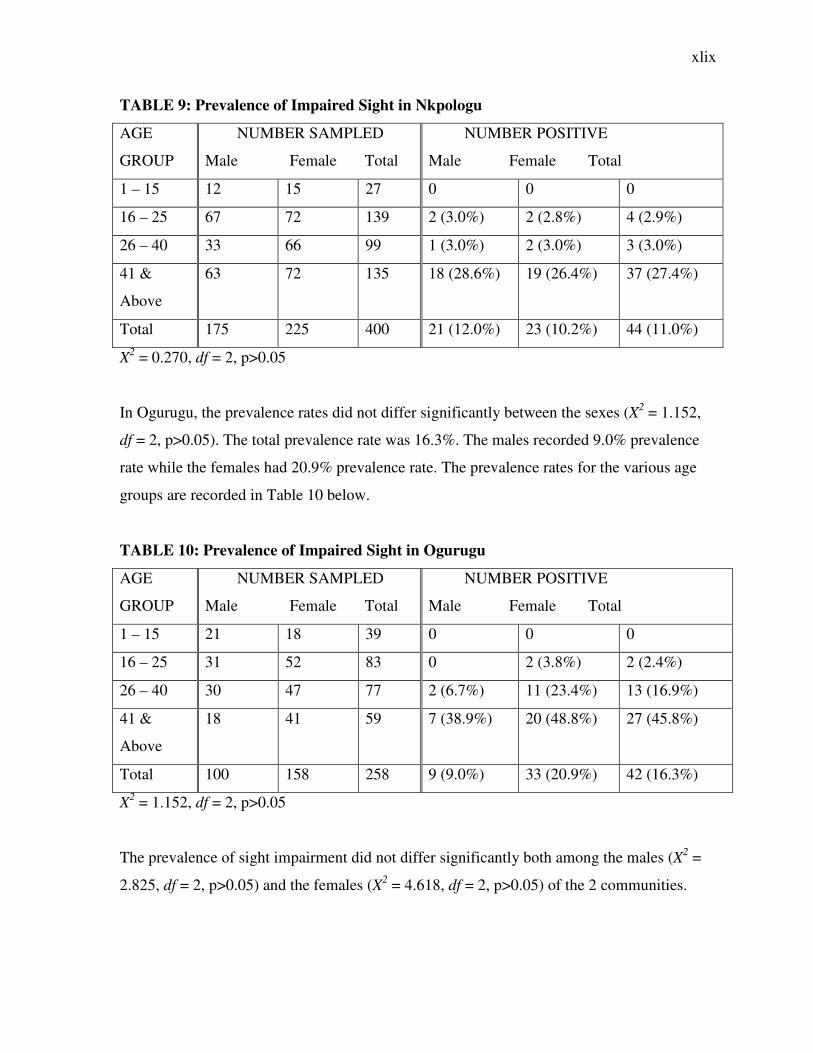
TABLE 9: Prevalence of Impaired Sight in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 0 0 0
16 – 25 67 72 139 2 (3.0%) 2 (2.8%) 4 (2.9%)
26 – 40 33 66 99 1 (3.0%) 2 (3.0%) 3 (3.0%)
41 &
Above
63 72 135 18 (28.6%) 19 (26.4%) 37 (27.4%)
Total 175 225 400 21 (12.0%) 23 (10.2%) 44 (11.0%)
X2 = 0.270, df = 2, p>0.05
In Ogurugu, the prevalence rates did not differ significantly between the sexes (X2 = 1.152,
df = 2, p>0.05). The total prevalence rate was 16.3%. The males recorded 9.0% prevalence
rate while the females had 20.9% prevalence rate. The prevalence rates for the various age
groups are recorded in Table 10 below.
TABLE 10: Prevalence of Impaired Sight in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 21 18 39 0 0 0
16 – 25 31 52 83 0 2 (3.8%) 2 (2.4%)
26 – 40 30 47 77 2 (6.7%) 11 (23.4%) 13 (16.9%)
41 &
Above
18 41 59 7 (38.9%) 20 (48.8%) 27 (45.8%)
Total 100 158 258 9 (9.0%) 33 (20.9%) 42 (16.3%)
X2 = 1.152, df = 2, p>0.05
The prevalence of sight impairment did not differ significantly both among the males (X2 =
2.825, df = 2, p>0.05) and the females (X2 = 4.618, df = 2, p>0.05) of the 2 communities.
l
Figure 6: Prevalence of Impaired Sight in Nkpologu and Ogurugu
3.1.5.2 Blindness
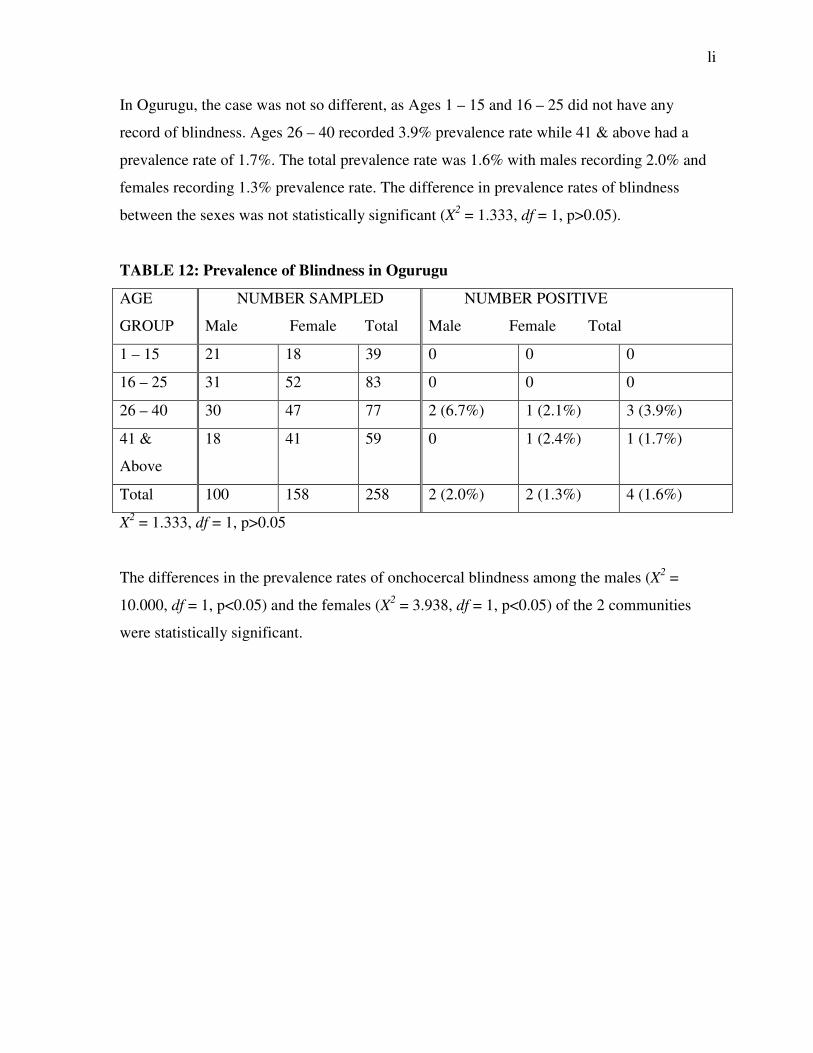
In Nkpologu, cases of blindness were also recorded. The total blindness rate was 3.8%. The
males recorded a blindness rate of 4.6% while the females recorded 3.1% blindness rate.
Ages 1 – 15, 16 – 25 and 26 – 40 did not record any case of blindness. However, Ages 41 &
above which accounted for all cases of blindness recorded a prevalence rate of 11.1% (Table
11).
TABLE 11: Prevalence of Blindness in Nkpologu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 12 15 27 0 0 0
16 – 25 67 72 139 0 0 0
26 – 40 33 66 99 0 0 0
41 &
Above
63 72 135 8 (12.7%) 7 (9.7%) 15 (11.1%)
Total 175 225 400 8 (4.6%) 7 (3.1%) 15 (3.8%)
0
5
10
15
20
25
30
35
40
45
50
Prevalence (%)
1 – 15 16 – 25 26 – 40 41 & Above Total
Age Group (Years)
Nkpologu
Ogurugu
li
In Ogurugu, the case was not so different, as Ages 1 – 15 and 16 – 25 did not have any
record of blindness. Ages 26 – 40 recorded 3.9% prevalence rate while 41 & above had a
prevalence rate of 1.7%. The total prevalence rate was 1.6% with males recording 2.0% and
females recording 1.3% prevalence rate. The difference in prevalence rates of blindness
between the sexes was not statistically significant (X2 = 1.333, df = 1, p>0.05).
TABLE 12: Prevalence of Blindness in Ogurugu
AGE
GROUP
NUMBER SAMPLED
Male Female Total
NUMBER POSITIVE
Male Female Total
1 – 15 21 18 39 0 0 0
16 – 25 31 52 83 0 0 0
26 – 40 30 47 77 2 (6.7%) 1 (2.1%) 3 (3.9%)
41 &
Above
18 41 59 0 1 (2.4%) 1 (1.7%)
Total 100 158 258 2 (2.0%) 2 (1.3%) 4 (1.6%)
X2 = 1.333, df = 1, p>0.05
The differences in the prevalence rates of onchocercal blindness among the males (X2 =
10.000, df = 1, p<0.05) and the females (X2 = 3.938, df = 1, p<0.05) of the 2 communities
were statistically significant.
lii
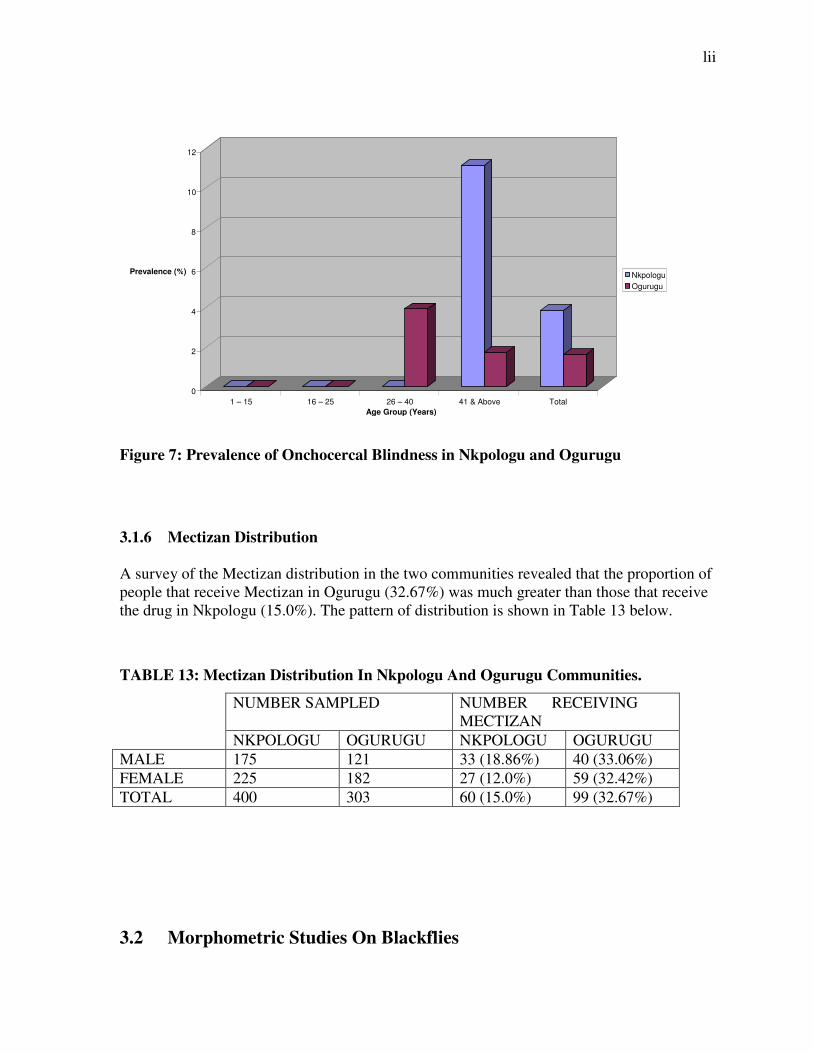
Figure 7: Prevalence of Onchocercal Blindness in Nkpologu and Ogurugu
3.1.6 Mectizan Distribution A survey of the Mectizan distribution in the two communities revealed that the proportion of people that receive Mectizan in Ogurugu (32.67%) was much greater than those that receive the drug in Nkpologu (15.0%). The pattern of distribution is shown in Table 13 below.
TABLE 13: Mectizan Distribution In Nkpologu And Ogurugu Communities.
NUMBER SAMPLED NUMBER RECEIVING MECTIZAN
NKPOLOGU OGURUGU NKPOLOGU OGURUGU
MALE 175 121 33 (18.86%) 40 (33.06%)
FEMALE 225 182 27 (12.0%) 59 (32.42%)
TOTAL 400 303 60 (15.0%) 99 (32.67%)
3.2 Morphometric Studies On Blackflies
0
2
4
6
8
10
12
Prevalence (%)
1 – 15 16 – 25 26 – 40 41 & Above Total
Age Group (Years)
Nkpologu Ogurugu

liii
The flies that were used in the study were collected from Nkpologu alone. No flies could be
collected at Ogurugu due to the use of chemicals in River Eshi by fishermen as reported by
the members of the community.
The mean lengths of the flies measured are as follows: Thorax length = 0.9821mm, Antenna
length = 0.3864mm, Wing length = 1.9739mm, Wing width = 0.8357mm and femur length =
0.5869mm. Table 14 shows the means and standard deviations of the various parts of the
flies measured.
Plate 5: Simulium damnosum
TABLE 14
PARAMETER MEAN ± STANDARD DEVIATION
Thorax Length (TL) 0.9821 ± 0.0904
Antenna Length (AL) 0.3864 ± 0.0736
Wing Length (WL) 1.9739 ± 0.1443
Wing Width (WW) 0.8357 ± 0.1800
Femur Length (FL) 0.5869 ± 0.0798
liv
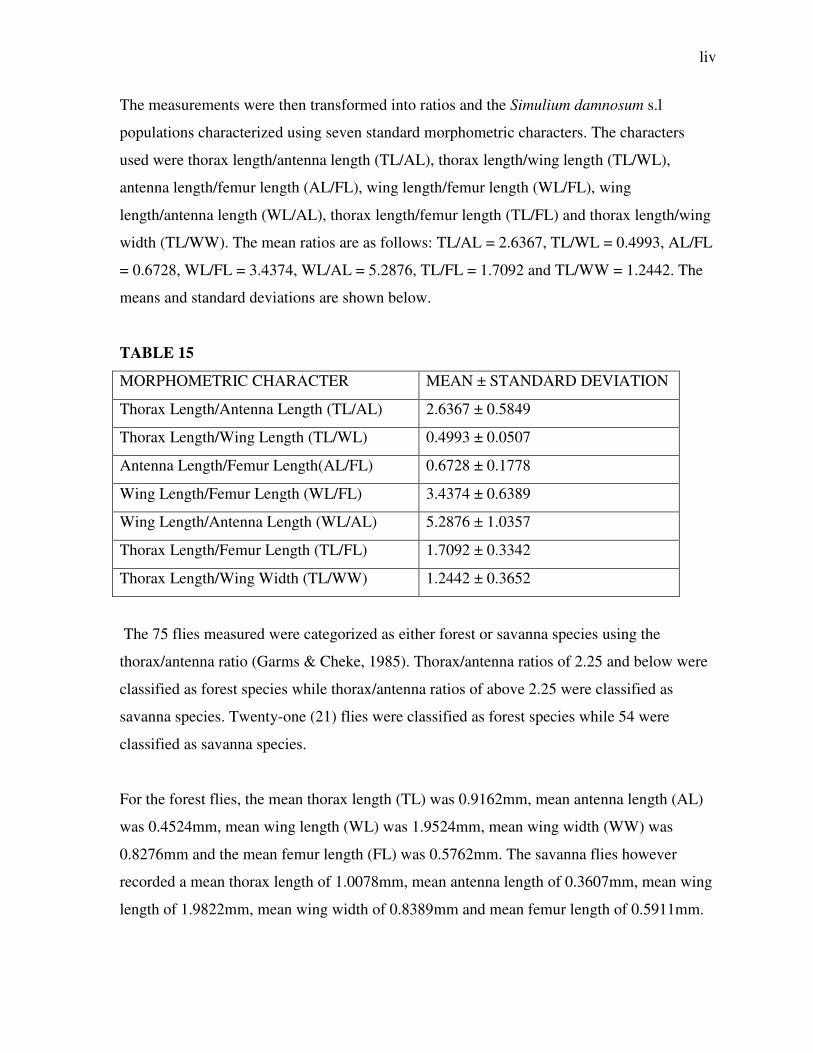
The measurements were then transformed into ratios and the Simulium damnosum s.l
populations characterized using seven standard morphometric characters. The characters
used were thorax length/antenna length (TL/AL), thorax length/wing length (TL/WL),
antenna length/femur length (AL/FL), wing length/femur length (WL/FL), wing
length/antenna length (WL/AL), thorax length/femur length (TL/FL) and thorax length/wing
width (TL/WW). The mean ratios are as follows: TL/AL = 2.6367, TL/WL = 0.4993, AL/FL
= 0.6728, WL/FL = 3.4374, WL/AL = 5.2876, TL/FL = 1.7092 and TL/WW = 1.2442. The
means and standard deviations are shown below.
TABLE 15
MORPHOMETRIC CHARACTER MEAN ± STANDARD DEVIATION
Thorax Length/Antenna Length (TL/AL) 2.6367 ± 0.5849
Thorax Length/Wing Length (TL/WL) 0.4993 ± 0.0507
Antenna Length/Femur Length(AL/FL) 0.6728 ± 0.1778
Wing Length/Femur Length (WL/FL) 3.4374 ± 0.6389
Wing Length/Antenna Length (WL/AL) 5.2876 ± 1.0357
Thorax Length/Femur Length (TL/FL) 1.7092 ± 0.3342
Thorax Length/Wing Width (TL/WW) 1.2442 ± 0.3652
The 75 flies measured were categorized as either forest or savanna species using the
thorax/antenna ratio (Garms & Cheke, 1985). Thorax/antenna ratios of 2.25 and below were
classified as forest species while thorax/antenna ratios of above 2.25 were classified as
savanna species. Twenty-one (21) flies were classified as forest species while 54 were
classified as savanna species.
For the forest flies, the mean thorax length (TL) was 0.9162mm, mean antenna length (AL)
was 0.4524mm, mean wing length (WL) was 1.9524mm, mean wing width (WW) was
0.8276mm and the mean femur length (FL) was 0.5762mm. The savanna flies however
recorded a mean thorax length of 1.0078mm, mean antenna length of 0.3607mm, mean wing
length of 1.9822mm, mean wing width of 0.8389mm and mean femur length of 0.5911mm.
lv
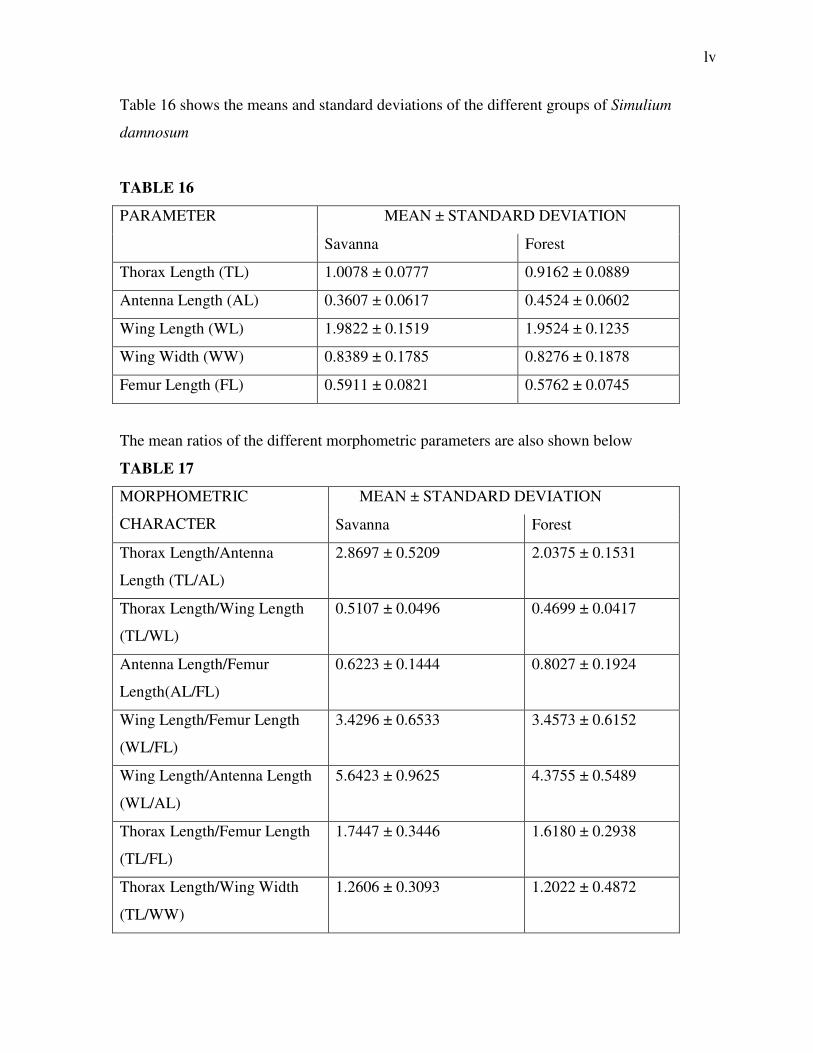
Table 16 shows the means and standard deviations of the different groups of Simulium
damnosum
TABLE 16
PARAMETER MEAN ± STANDARD DEVIATION
Savanna Forest
Thorax Length (TL) 1.0078 ± 0.0777 0.9162 ± 0.0889
Antenna Length (AL) 0.3607 ± 0.0617 0.4524 ± 0.0602
Wing Length (WL) 1.9822 ± 0.1519 1.9524 ± 0.1235
Wing Width (WW) 0.8389 ± 0.1785 0.8276 ± 0.1878
Femur Length (FL) 0.5911 ± 0.0821 0.5762 ± 0.0745
The mean ratios of the different morphometric parameters are also shown below
TABLE 17
MORPHOMETRIC
CHARACTER
MEAN ± STANDARD DEVIATION
Savanna Forest
Thorax Length/Antenna
Length (TL/AL)
2.8697 ± 0.5209 2.0375 ± 0.1531
Thorax Length/Wing Length
(TL/WL)
0.5107 ± 0.0496 0.4699 ± 0.0417
Antenna Length/Femur
Length(AL/FL)
0.6223 ± 0.1444 0.8027 ± 0.1924
Wing Length/Femur Length
(WL/FL)
3.4296 ± 0.6533 3.4573 ± 0.6152
Wing Length/Antenna Length
(WL/AL)
5.6423 ± 0.9625 4.3755 ± 0.5489
Thorax Length/Femur Length
(TL/FL)
1.7447 ± 0.3446 1.6180 ± 0.2938
Thorax Length/Wing Width
(TL/WW)
1.2606 ± 0.3093 1.2022 ± 0.4872
lvi
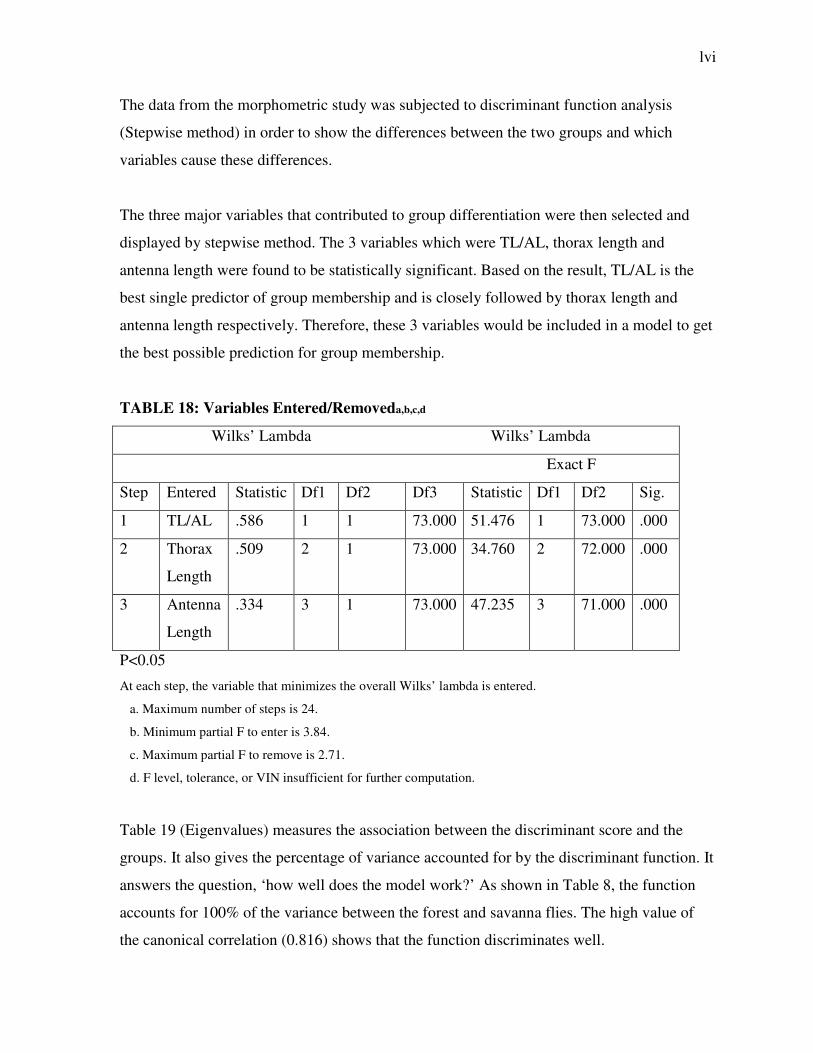
The data from the morphometric study was subjected to discriminant function analysis
(Stepwise method) in order to show the differences between the two groups and which
variables cause these differences.
The three major variables that contributed to group differentiation were then selected and
displayed by stepwise method. The 3 variables which were TL/AL, thorax length and
antenna length were found to be statistically significant. Based on the result, TL/AL is the
best single predictor of group membership and is closely followed by thorax length and
antenna length respectively. Therefore, these 3 variables would be included in a model to get
the best possible prediction for group membership.
TABLE 18: Variables Entered/Removeda,b,c,d
Wilks’ Lambda Wilks’ Lambda
Exact F
Step Entered Statistic Df1 Df2 Df3 Statistic Df1 Df2 Sig.
1 TL/AL .586 1 1 73.000 51.476 1 73.000 .000
2 Thorax
Length
.509 2 1 73.000 34.760 2 72.000 .000
3 Antenna
Length
.334 3 1 73.000 47.235 3 71.000 .000
P<0.05
At each step, the variable that minimizes the overall Wilks’ lambda is entered.
a. Maximum number of steps is 24.
b. Minimum partial F to enter is 3.84.
c. Maximum partial F to remove is 2.71.
d. F level, tolerance, or VIN insufficient for further computation.
Table 19 (Eigenvalues) measures the association between the discriminant score and the
groups. It also gives the percentage of variance accounted for by the discriminant function. It
answers the question, ‘how well does the model work?’ As shown in Table 8, the function
accounts for 100% of the variance between the forest and savanna flies. The high value of
the canonical correlation (0.816) shows that the function discriminates well.
lvii
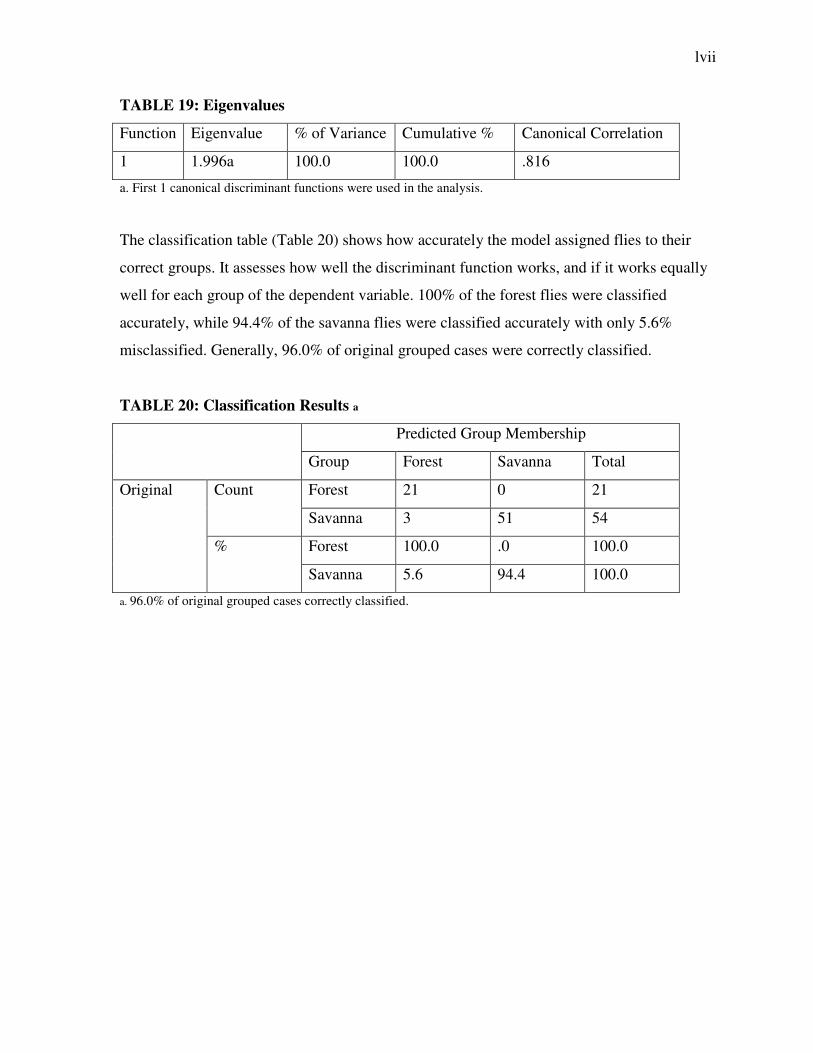
TABLE 19: Eigenvalues
Function Eigenvalue % of Variance Cumulative % Canonical Correlation
1 1.996a 100.0 100.0 .816
a. First 1 canonical discriminant functions were used in the analysis.
The classification table (Table 20) shows how accurately the model assigned flies to their
correct groups. It assesses how well the discriminant function works, and if it works equally
well for each group of the dependent variable. 100% of the forest flies were classified
accurately, while 94.4% of the savanna flies were classified accurately with only 5.6%
misclassified. Generally, 96.0% of original grouped cases were correctly classified.
TABLE 20: Classification Results a
Predicted Group Membership
Group Forest Savanna Total
Original Count Forest 21 0 21
Savanna 3 51 54
% Forest 100.0 .0 100.0
Savanna 5.6 94.4 100.0
a. 96.0% of original grouped cases correctly classified.

lviii
CHAPTER FOUR
DISCUSSION AND CONCLUSION
4.1 Epidemiology Of Onchocerciasis In Nkpologu And Ogurugu
Communities
Both Nkpologu and Ogurugu communities were found to be endemic for onchocerciasis.
This is consistent with the report of Ubachukwu (2004), which stated that Uzo-Uwani Local
Government Area is meso-endemic for onchocerciasis based on the rapid indicators of
onchocerciasis endemicity used especially the prevalence of palpable nodules. Withworth
and Gemade (1999) found that prevalence of palpable nodules correlates more closely with
microfilarial prevalence and community microfilarial load than the prevalence of skin
depigmentation and can be used to replace skin snipping and parasitological examination.
Kale (1998) reported that the current standard practice is to examine a sample of 30 – 50
adult males per community for nodules. Using these parameters, the levels of endemicity
adopted by the African Programme for onchocerciasis Control (APOC) are hyper-endemic
(> 39% of adult males with nodules, equivalent to a prevalence of infection in the whole
community > 59%, with blindness rates usually above 10%), meso-endemic (a nodule rate of
20% - 39%, equivalent to a prevalence in the community of 40% - 59%) and hypo-endemic
(a nodule rate of < 20%, equivalent to a prevalence in the community of < 40%, with
blindness rates generally < 1%) (WHO, 1987).
In Nkpologu, the prevalence of palpable nodules for all ages was 5.8% while among the
adult male population, it was 9.1%. However, the blindness rate was 3.8%. This suggests
that Nkpologu is meso-endemic for onchocerciasis. On the other hand, Ogurugu had a
prevalence rate of 2.6% for all ages while the prevalence rate among adult males was 9.3%.
Blindness rate in Ogurugu was 1.6%. This also suggests meso-endemicity.

lix
Different epidemiological patterns have been described within the same zone. There are two
fairly broad but distinct clinico-pathological patterns of onchocerciasis, particularly in West
Africa, based on the disease’s two predominant and major clinical complications: blindness
and skin disease. In West Africa, blindness rates are significantly higher in hyper-endemic
communities in the savanna than in communities with similar levels of infection in the rain
forest (Dadzie et al., 1990). Hence Nkpologu, which is closer to the rainforest vegetation
zone, has a higher prevalence of nodules (5.8%) and leopard skin (12.5%) than Ogurugu
(Nodules = 2.6%, Leopard Skin = 4.0%), which lies closer to the savanna zone.
Expectedly, Ogurugu has a higher prevalence of sight impairment (16.3%) than Nkpologu
(11.0%). However, the actual prevalence of blindness is greater in Nkpologu (3.8%) than in
Ogurugu (1.6%). This could be attributed to the patterns of Mectizan distribution in the 2
communities.
The proportion of individuals taking mectizan in Ogurugu was 19.5% while 6.8% of the
population in Nkpologu takes the drug. A single, annual dose of Mectizan of 150 - 200 µg/kg
has been found to effectively reduce the microfilarial density in the skin to near zero after
one month and to successfully maintain a low microfilarial level for up to 12 months.
Reduction of microfilarial density in the skin can significantly reduce parasite transmission.
It has been determined that Mectizan has a direct toxic effect on microfilariae and also
interferes with their development and release from the adult female worm uterus (Goa et al.,
1991). Mectizan also has a very beneficial effect on symptoms and clinical manifestations
due to onchocerciasis. It relieves the intense itching from the disease, and it clears
microfilaria from the eye, thus halting the progression towards blindness except in very
advanced cases (WHO, 1995a; Ogbuagu & Eneanya, 1998). Probably, the high level of
visual impairment observed in individuals in Ogurugu had been reversed as a result of better
distribution systems of Mectizan.
The major approach employed in the distribution of Mectizan in Uzo-Uwani Local
Government Area is the active or community mass treatment method. This is the method of
choice in meso- and hyper-endemic areas of onchocerciasis. In these areas, ivermectin is
given once a year for at least 14 years to all members of the community. Amazigo et al.

lx
(1998) reported that these community-based distributors can, with minimum but direct
supervision, lead to higher coverages, be more cost-effective and be more likely to be
sustained.
The crucial problem is that ivermectin leads to depletion of skin microfilariae (mf) for only a
few months, followed by a reappearance of mf within one year at levels of more than 20% of
that at pretreatment (Awadzi et al., 1995) and this mf (microfilaria) density seems sufficient
for transmission to continue. More recent meta-analysis of extensive data from former OCP
areas has shown that even semi-annual mass treatment of ivermectin for several years
followed by cessation e.g. due to political unrest, has resulted in high rates of recrudescence
(Borsboom et al., 2003).
A recent conference on the eradicability of onchocerciasis concluded that eradication is not
feasible with the present tools alone (Dadzie et al., 2003). In addition, sub-optimal efficacy
of ivermectin and/or ivermectin resistance in humans has been reported in onchocerciasis
patients in Ghana where despite multiple treatments with ivermectin, microfilaidermia
persisted in some patients (Awadzi et al., 2004).
Though for many years thought to be of relatively minor importance compared with
onchocercal eye disease, the skin lesions of onchocerciasis have recently been shown to be a
major socio-economic burden, in terms of disability-adjusted life-years. The prevalence of
onchocercal skin diseases (OSD) was also higher in Nkpologu than in Ogurugu as expected.
The prevalence of most types of onchocercal skin lesion show statistically significant
correlation with onchocercal endemicity, the correlation being stronger for reactive skin
lesions than for depigmentation (Kale, 1998). The demonstration of an excellent correlation
between the prevalence of palpable nodules in a community and the community microfilarial
load has led to the development of rapid, safe and non-invasive methods to assess and map
the levels of endemicity across whole countries. This has enabled mass treatment with
Mectizan to be targeted first at hyper-endemic communities. Troublesome itching is also an
important debilitating feature of onchocerciasis, especially in individuals with high levels of
infection and has been found to have a very strong correlation with endemicity (WHO,

lxi
1995b). Onchocercal skin disease is also a very important public health problem. Apart from
the undesirable and adverse, attendant, physical effects of skin disease, there are important
psychosocial consequences, often of a very stressful nature. Individuals with OSD may
suffer psychological disturbances such as sleeplessness, lack of concentration, poor self
esteem and other negative forms of stigmatization (WHO, 1995b).
It is noteworthy that the prevalence of onchocercal symptoms was consistently higher among
males than among females in both communities. This can be attributed to factors such as
occupation, intensity and frequency of man-fly contact and mode of dressing. Intensity of
exposure is a factor of the number of infective bites sustained by the individual and this, in
turn, depends as much on the transmission efficiency of the local vector (its ‘vectorial
capacity’), as it does on the proximity of the residence or work place to the breeding sites of
the vector. Ubachukwu (2004) reported that in this area, males, right from childhood, are
involved more in outdoor activities such as playing football, fishing, hunting, farming,
swimming etc. than females who are involved more in indoor activities such as cooking,
sweeping etc. Even when females are involved in outdoor activities, they spend less time
than the males and also cover themselves properly. Among the adult population, the males
are involved in occupations like full-time farming and fishing and generally spend longer
time outside than the females who combine farming with other occupations like trading.
Hence, the males are constantly being brought into close and intimate relationship with the
vector and are at relatively higher risk of infection (Kale, 1998). In addition, males usually
expose greater parts of their bodies than females while working.
The prevalence of onchocercal skin diseases (OSD) was also higher in Nkpologu than in
Ogurugu as expected. The prevalences of most types of onchocercal skin lesion show
statistically significant correlation with onchocercal endemicity, the correlation being
stronger for reactive skin lesions than for depigmentation (Kale, 1998). Troublesome itching
is also an important debilitating feature of onchocerciasis, especially in individuals with high
levels of infection and has been found to have a very strong correlation with endemicity
(WHO, 1995b). Onchocercal skin disease is also a very important public health problem.
Apart from the undesirable and adverse, attendant, physical effects of skin disease, there are

lxii
important psychosocial consequences, often of a very stressful nature. Individuals with OSD
may suffer psychological disturbances such as sleeplessness, lack of concentration, poor self
esteem and other negative forms of stigmatization (WHO, 1995b).
It is noteworthy that the prevalence of onchocercal symptoms was consistently higher among
males than among females in both communities. This can be attributed to factors such as
occupation, intensity and frequency of man-fly contact and mode of dressing. Ubachukwu
(2004) reported that in this area, males, right from childhood, are involved more in outdoor
activities such as playing football, fishing, hunting, farming, swimming etc. than females
who are involved more in indoor activities such as cooking, sweeping etc. Even when
females are involved in outdoor activities, they spend less time than the males and also cover
themselves properly. Among the adult population, the males are involved in occupations like
full-time farming and fishing and generally spend longer time outside than the females who
combine farming with other occupations like trading. Hence, the males are constantly being
brought into close and intimate relationship with the vector and are at relatively higher risk
of infection (Kale, 1998). In addition, males usually expose greater parts of their bodies than
females while working.
The prevalence of skin depigmentation, visual impairment and onchocercal blindness
increased with increasing age. This lays credence to the results from previous studies
(Nwoke et al., 1989; Amazigo et al., 1993; Ubachukwu, 2004) and bears witness to the
cumulative nature of onchocerciasis. The longer the exposure of individuals to vector
Simulium flies, the greater the accumulation of the parasites in the body. This results in
greater severity and manifestations of the disease in the upper age groups. Prevalence of
onchocercal infection is lowest in the first decade of life, after which it rises steeply to reach
a peak, usually in the third decade of life.
The economic impact of onchocerciasis is enormous. Among some adult populations living
in impoverished areas of developing countries, onchocerciasis is a leading cause of blindness
and skin disease. Although not fatal, blindness in onchocerciasis causes lowered expectation
of life (by 10 – 15 years). In some hyperendemic communities, every second person

lxiii
ultimately goes blind. In other communities, onchodermal skin lesions are an important
social problem. In 1990, onchocerciasis was responsible for the loss of 884,000 Disability
Adjusted Life Years (DALYs) (WHO, 1996).
Furthermore, a look at the cost benefit analysis of the various control programmes of
onchocerciasis gives an indication of the economic burden of the disease. The World bank
calculated the annual return on investment of the just concluded Onchocerciasis Control
Programme (attributable mainly to increased agricultural output) to be 20%, and it is
estimated that $3.7 billion will be generated from improved labour and agricultural
productivity (Hopkins & Richards, 1997). Also, an estimated 25 million hectares of unusable
arable land – enough to feed an additional 17 million people per annum – has become
available for agricultural production and resettlement (Samba, 1994).
4.2 Morphometric Studies On Blackflies
Human onchocerciasis is a severely debilitating, blinding disease caused by infection with
Onchocerca volvulus (Nematoda: Filarioidea). In West Africa, the parasite is transmitted by
blood-sucking adult females of sibling species of the Simulium damnosum complex (Diptera:
Simuliidae). The major patterns of epidemiological variation are related to the taxonomy of
the parasite and the vector (Post & Boakye, 1992), and so not all sibling species are equally
important. Although vector taxonomy is majorly based upon the analysis of the polytene
chromosomes from the larval silk glands, there is still a requirement for the identification of
the adult female, because it is this stage that actually transmits the parasite.
In this study, adult female Simulium damnosum were classified as either forest or savanna
species using the thorax/antenna ratio. Twenty-one flies were classified as forest while 54
were classed as savanna flies. The use of the thorax/antenna ratio in the differentiation of
forest and savanna flies has proved useful. No more reliable method has been found to
separate individual female flies of the S. damnosum complex (Wilson et al., 1993). The use

lxiv
of thorax and antenna lengths alone in a bivariate analysis resulted in 100% separation of the
savanna species and S. sanctipauli flies (Garms, 1978; Wilson & Baker, 1991).
Members of the Simulium damnosum Theobald complex are classified into forest and
savanna species based on the preferred breeding habitat. The savanna-species have been
known to be efficient vectors of the severely blinding savanna strain of Onchocerca volvulus
and inefficient vectors of the less blinding forest-strain and vice versa for forest species
(Duke, 1990). However, Toe et al. (1997) reported that this vector-parasite complex does not
exist in the transition zones between the forest and the savanna. Hence the co-existence of
both forest and savanna species in Nkpologu is not strange as this area belongs to the forest-
savanna-mosaic vegetation belt in south-eastern Nigeria (Crosskey, 1981). Such occurrences
have been reported in other parts of Africa (Opoku, 2006; Wilson et al., 2002; Kale, 1998).
In addition, this co-existence has been attributed to the incursion of savanna flies into
massively deforested sites (Boakye, 1999; Wilson et al., 2002). Hence, deforestation is a
very important factor in determining the kind of onchocerciasis prevalent in a particular area.
Infact, available data suggest that savanna species cannot only invade deforested areas but
also establish breeding populations that would persist (Post & Crosskey, 1985; Baker et al.,
1990). Onyenwe (2007) reported the occurrence of S. sirbanum from cytotaxonomic
examinations carried out in Ogurugu.
The importance of this study lies in the fact that it can be an efficient and fast indicator for
the detection of any trend of change in prevalence or severity of the disease. Such a change
might be brought about, for instance, by local deforestation and subsequent invasion by
savanna vector sibling species, which might bring the savanna strain of O. vovulus with
them. Early warning of any such change in fly population would be very important
especially in the detection of any trend of change in the prevalence or severity of
onchocerciasis (Ibeh et al., 2008).

lxv
Furthermore, the biggest problem in ascribing epidemiological importance to the different
cytotaxonomic entities of the members of the Simulium damnosum complex is that the
cytotaxonomic identifications can only be made from larvae, but it is the adult female that
actually transmits the parasite. The vagility of Damnosum species also makes it difficult to
say with certainty that flies biting alongside a river have emerged from that river. The
identification of adult flies biting man by reason of their proximity to breeding sites of
known species is therefore, at best uncertain. The admixture of a small proportion of migrant
individuals of a more efficient vector may grossly distort the apparent role of a particular
species in transmission (Ibeh et al., 2008).
4.3 Summary and Recommendations
Both Ogurugu and Nkpologu communities are shown to be endemic for onchocerciasis. The
prevalence in both communities is higher among males than females. The morphometric
studies on the Simulium vectors reveal a greater abundance of savanna flies as compared to
forest species in Nkpologu. This suggests that there might be a greater prevalence of sight-
related problems than onchocercal skin disease in the community. In addition, the prevalence
of onchocercal symptoms is higher in Nkpologu than in Ogurugu. This is as a result of the
greater proportion of individuals receiving Mectizan in Ogurugu than in Nkpologu.
It is recommended that both males and females should properly cover their bodies while
working outside in order to reduce man-fly contact to the minimal level. Also, the 2
communities, especially Nkpologu, should be educated on the need to increase the intake of
Mectizan in order to prevent further spread of the disease. A proper training of community-
based distributors would go a long way in increasing the reach of the drugs. Finally,
deforestation should be discouraged in Nkpologu in order to stop further incursion of the
savanna species of the Simulium damnosum complex and hence, stop the spread of the
severely blinding strain of Onchocerca volvulus.

lxvi
REFERENCES
Abiose A (1990). Onchocerciasis and blindness. Paper presented at the
Second National Conference on Onchocerciasis Research and Control in Kadunna, Nigeria. p. 20.
Adams, V.J., Lombard, C.J., Dhansay, M.A., Markus, M.B. and Fincham, J.E. (2004). Efficacy of albendazole against the whipworm Trichuris trichiura – a randomized, controlled trial. South African Medical Journal, 94: 972 – 976. Albonico, M., Bickle, Q., Ramsan, M., Montressor, A., Savioli, L. and Taylor, M. (2003). Efficacy of mebendazole and levamisole alone or in combination against intestinal nematode infections after repeated targeted mebendazole treatment in Zanzibar. Bulletin of the World Health Organization, 81: 343 – 352. Albonico, M., Smith, P.G., Ercole, E., Hall, A., Chwaya, H.M., Alawi, K.S. and Savioli, L. (1995). Rate of re-infection with intestinal nematodesafter treatment of children with mebendazole or albendazole in a highly endemic area. Transactions of the Royal Society
of Tropical Medicine and Hygiene, 89: 538 – 541.
Amazigo, U.O. and Obikeze, D.S. (1992). Social consequences of onchocercal skin
lesions on adolescent girls in rural Nigeria. WHO/TDR Discussion Paper. WHO, Geneva.
Asaolu, S.O. and Ofoezie, I.E. (2003). The role of health education and sanitation in the control of helminth infections. Acta Tropica, 86: 283 – 294. Barger, I.A. (1982). Helminth parasites and animal production. In: Symons, L.E.A., Donald, A.D. and Dineen, J.K. (eds). Biology and Control of Endoparasites. Academic Press, New York. Bethony, J., Brooker, S., Albonico, M., Geiger, S.M., Loukas, A., Diemert, D. and Hotez, P.J. (2006). Soil-transmitted helminth infections: ascariasis, trichuriasis and hookworm. Lancet, 367(9521): 1521 – 1532. Bleakley, H. (2003). Disease and development: evidence from hookworm eradication in the American south. Journal of the European Economic Association, 1: 376 – 386. Brooker, S., Bethony, J. and Hotez, P.J. (2004). Human hookworm infection in the 21st century. Advances in Parasitology, 58: 197 – 288.

lxvii
Brooker, S. and Michael, E. (2000). The potential of geographical information systems and remote sensing in the epidemiology and control of human helminth infections. Advances in Parasitology, 47: 245 – 287. Bundy, D.A.P. (1995). Epidemiology and transmission of intestinal helminthes In: Farthing,
M.J.G., Keusch, G.T. and Wakelin, D. (eds), Enteric Infection 2, Intestinal
Helminths, Chapman & Hall, New York. Bundy, D.A., Chan, M.S. and Savioli, L. (1995). Hookworm infection in pregnancy. Transactions of the Royal Society of Tropical Medicine and Hygiene, 89: 521 – 522. Cheng, T.C. (1986). General Parasitology, Carlifonia, Academic Press. Chiejina, S.N. and Ekwe, T.O. (1986). Canine toxocariasis and the associated environmental contamination of urban areas in Eastern Nigeria. Veterinary
Parasitology, 22(112): 157 – 161. Cooper, E. (1991). Intestinal parasitoses and the modern description of diseases of poverty. Transactions of the Royal Society of Tropical Medicine and Hygiene, 85: 168 – 170.
Coreil, J., Mayard, G., Louis-Charles, J. and Addiss, D. (1998). Filarial elephantiasis among Haitian women: social context and behavioural factors in treatment. Tropical Medicine and International Health, 3: 467 - 473.
Crompton, D.W. (1999). How much helminthiases is there in the world? Journal of
Parasitology, 85: 397 – 403.
Crompton, D.W. and Nesheim, M.C. (2002). Nutritional impact of intestinal
helminthiasis during the human life cycle. Annual Review of Nutrition, 22:
35 – 59. Crompton, D.W. and Savioli, L. (1993). Intestinal parasitic infections and urbanization. Bulletin of the World Health Organization, 71: 1 – 7. deSilva, N.R., Brooker, S., Hotez, P.J., Montressor, A., Engles, D. and Savioli, L. (2003). Soil-transmitted helminth infections: updating the global picture. Trends in Parasitology, 19: 547 – 551.
Dreyer, G., Medeiros, Z., Netto, M.J., Leal, N.C., de Castro, L.G. and Piessens, W.F. (1999). Acute attacks in the extremities of persons living in an area endemic for Bancroftian filariasis: differentiation of two syndromes. Transactions of the Royal
Soceity of Tropical Medicine and Hygiene, 93: 413-417.

lxviii
Dreyer, G., Noroes, J. and Addiss, D. (1997). The silent burden of sexual disability associated with lymphatic filariasis. Acta Tropica, 63: 57-60.
Edungbola, L.D. and Parakoyi, D.B. (1991). Onchocerciasis and Dracunculiasis: a review. Nigerian Journal of Parasitology, 12: 3 – 27.
Global Alliance to Eliminate Lymphatic Filariasis. www.filariasis.org/pdfs/MIP.pdf. Accessed 11th February, 2009.
Gyapong, M., Gyapong, J., Weiss, M. and Tanner, M. (2000). The burden of hydrocele on men in Northern Ghana. Acta Tropica, 77: 287 - 294.
Herdt, R.W. (1991). Research priorities for rice biotechnology. In: Khush, G.S. and Toenniessen, G.H. (eds). Pp 19 – 54. Rice Biotechnology. Commonwealth Agricultural Bureau International, Oxon, UK.
Hopkins, D.R. and Richards, F. (1997). Visionary campaign: eliminating river blindness.
In Bernsteid, E. (ed) Medical and Health Annual. Encyclopaedia Britannica, Chicago.
Hotez, P.J., Brindley, P.J., Bethony, J.M., King, C.H., Pearce, E.J. and Jacobson, J. (2008). Helminth infections: the great neglected tropical diseases. Journal of
Clinical Investigations, 118(4): 1311–1321.
Hotez, P.J., Brooker, S., Bethony, J.M., Bottazzi, M.E., Loukas, A. and Xiao, S.H. (2004). Current concepts: hookworm infection. New England Journal of Medicine, 351: 799 – 807.
Hotez, P.J., daSilva, N., Brooker, S. and Bethony, J. (2003). Soil Transmitted Helminth Infections: The Nature, Causes and Burden of the Condition. Working Paper No. 3, Disease Control Priority Project. Behesda, Maryland: Fogarty International Centre, National Institute.
Hotez, P.J. and Ferris, M.T. (2006). The antipoverty vaccines. Vaccine, 24: 5787 –
5799. Hotez, P.J., Molyneux, D.H., Fenwick, A., Kumaresan, J., Sachs, S.E., Sachs, J.D. and Savioli, L. (2007). Control of neglected tropical diseases. New England
Journal of Medicine, 357: 1018-1027 Ijagbone, I.F. and Olagunju, T.F. (2006). Intestinal helminth parasites in school children in Iragbiji, Boripe Local Government, Osun State, Nigeria. African Journal of Biomedical
Research, 9: 63 – 66.

lxix
Jithendran, K.P. (2000). Helminth parasites: a constraint in animal health management in Himachal Pradesh. ENVIS Bulletin, 8(2): 1 – 18. Jithendran, K.P. and Krishna, L. (1991). Liver fluke infection and its control. Livestock
Advisor, 16: 24 – 26.
Kumari, A.K., Harichandrakumar, K.T., Das, K.L. and Krishnamoorthy, K. (2005). Physical and psychosocial burden due to lymphatic fialriasis as perceived by patients and medical experts. Tropical Medicine and International Health, 10: 567-73.
Malgor, R., Oku, Y., Gallardo, R. and Yarzabal, I. (1996). High prevalence of Ancylostoma spp. Infection in dogs, associated with endemic focus of human cutaneous larva migrans, in Tacuarembo, Uruguay. Parasite, 3(2): 131 – 134. Miguel, E. A. and Kremer, M. (2003). Worms : identifying impacts on education and health in the presence of treatment externalities. Econometrica, 72(1): 159 – 217. Miller, J.E. and Horohov, D.W.(2006). Immunological aspects of nematode parasite control in sheep. Journal of Animal Science, 84: E124 – E132. Montressor, A., Crompton, D.W.T., Gyorkos, T.W. and Savioli, L. (2002). Helminth control
in school-age children: a guide for managers of control programmes. World Health Organization, Geneva.
Mujinja, P.G.M., Gasarasi, D.B., Premji, Z.G. and Nguma, J. (1997). Social and economic impact of lymphatic filariasis in Rufiji district, Southeast Tanzania. In: Lymphatic
filariasis research and control in Africa. Report on a workshop held in Tanga, Tanzania. Tanzania: Danish Bilharziasis Laboratory, Denmark & National Institute for Medical Research.
Nwoke, B.E.B. (1990). The socio-economic aspects of human onchocerciasis in Africa:
present appraisal. Journal of Hygiene, Epidemiology, Microbiology and Immunology,
59(1): 37 – 44.
OPEC Fund. Onchocerciasis Control programme nears completion. Available at:
http://www.opecfund.org/publications/ar01/boxes/box5.htm. Accessed 16th
May, 2008.
Perera, M., Whitehead, M., Molyneux, D., Weerasooriya, M. and Gunatilleke, G. (2007).
Neglected patients in neglected disease? Qualitative study of lymphatic filariasis.
Neglected Tropical Diseases, 1: e128.

lxx
Regassa, F., Sori, T., Dhuguma, R. and Kiros, Y. (2006). Epidemiology of gastrointestinal parasites of ruminants in Western Oromia, Ethiopia. International Journal of Applied Research in Veterinary Medicine, 4(1): 51 – 57. Ross, A.G., Bartley, P.B., Sleigh, A.C., Olds, G.R., Li, Y., Williams, G.M. and McManus, D.P. (2002). Schistosomiasis. New England Journal of Medicine, 346: 1212 – 1220. Sangster, N.C. (1999). Anthelminthic resistance: past, present and future. International
Journal of Parasitology, 29: 115 – 124. Sasser, J.N. and Freckman, D.W. (1987). A world perspective on nematology: the role of the society. In: Veech, J.A. and Dickson, D.W. (eds). Pp 7 – 14. Vista on nematology: a
commemoration of the twenty-fifth anniversary of the society of nematologists. Society of Nematologists, Hyattsville, Maryland. Schmitt, D.P. and Sipes, B.S. (1998). Plant-parasitic nematodes and their management. Plant Disease, 15: 1 – 4. Stear, M.J., Strain, S. and Bishop, S.C. (1999). Mechanisms underlying resistance to nematode infection. International Journal of Parasitology, 29: 51 – 56. Stephenson, L.S., Latham, M.C. and Ottesen, E.A. (2000). Malnutrition and parasitic helminth infections. Parasitology, 121(Supplement): S23 – S28. Stoltzfus, R.J., Dreyfuss, M.L., Chwaya, H.M. and Albonico, M. (1997). Hookworm control as a strategy to prevent iron deficiency anaemia. Nutrition Reviews, 55: 223 – 232.
Suma, T.K., Shenoy, R.K. and Kumaraswami, V. (2003). A qualitative study of the perceptions, practices and socio-pyschological suffering related to chronic brugian filariasis in Kerala, southern India. Annals of Tropical Medicine and Parasitolology, 97: 839 - 845.
Thomas, D. and Strauss, J. (1997). Health and wages: evidence on men and women in urban Brazil. Journal of Econometrics, 77: 159 – 185. Ubachukwu, P.O. (2006). Socio-economic impact of onchocerciasis with particular reference to females and children: a review. Animal Research International, 3(2): 494 – 504.

lxxi
Ukpai, O.M. and Ugwu, C.O. (2003). The prevalence of gastro-intestinal tract parasites in primary school children in Ikwuano Local Government Area of Abia State, Nigeria. Nigerian Journal of Parasitology, 240: 129 – 136.
Uneke, C.J., Eze, K.O., Oyibo, P.G., Azu, N.C. and Ali, E. (2007). Soil-transmitted helminth infection in school children in South-Eastern Nigeria: the public health implication. The Internet Journal of third World Medicine, 4(1): 1 – 10. Van der Werf, M.J., de Vlas, S.J., Brooker, S., Looman, C.W., Nagelkerke, N.J., Habbema, J.D. and Engels, D. (2003). Quantification of clinical morbidity associated with schistosome infection in sub-Saharan Africa. Acta Tropica, 86: 125 – 139. Wagbatsoma, U.A. and Aisien, M.B. (2000). Helminthiasis in selected children seen at the University of Benin City, Nigeria. Nigerian Journal of Parasitology, 113: 87 – 95.
WHO (1996). Investing in research and development. Report of the adhoc committee on
health research relating to future intervention options. World Health Organization,
TDR/Gen/96.1
WHO (2002). Lymphatic filariasis: the disease and its control. Fifth report of the WHO expert committee on filariasis. WHO, Geneva.
Witt, C. and Ottesen, E.A. (2001). Lymphatic filariasis: an infection of childhood. Tropical Medicine and International Health, 6: 582 - 606.
World Bank. (2003). School Deworming at a Glance. Public health at a glance series. http://www.worldbank.org/hnp World Health Organization. (2002). Prevention of Schistosomiasis and Soil-
Transmitted Helminthiasis. WHO Technical Series Report 912. WHO, Geneva.

lxxii





























