Embed Size (px)
Citation preview

STUDIES ON STREPTOCOCCUS PYOGENESV. BIOCHEMICAL AND MICROSCOPIC ASPECTS OF CELL LYsis AND
DIGESTION BY ENZYMES FROM Streptomyces albus'
HUTTON D. SLADE2 AND WILLIAM C. SLAMPDepartment of Microbiology, Northwestern University Medical School, Chicago, Illinois
Received for publication June 29, 1959
Studies on the lysis and enzymatic digestionof group A streptococci should provide informa-tion on the anatomy, structure, chemical com-position, and biological activities of the organism.The information obtained from such studies issignificant for an understanding of the patho-genesis of streptococcal infections.
Lysis is accomplished by (a) infection withbacteriophage (Evans, 1940, 1942), (b) the actionof phage lysates from group C streptococci(Maxted, 1957; Krause, 1957), and (c) the actionof enzymes from Streptomyces albus (Maxted,1948). In contrast to many gram-positive species,the cell wall of the streptococcus is resistant tothe action of lysozyme. Certain amino acids,however, are removed from the cell wall of themicroorganism by the action of trypsin (Salton,1953). Although this treatment rendered the cellpermeable to otherwise impermeable substrates(Slade, 1957) and removed M antigen, lysis didnot occur.
Little information is available on either themicroscopic changes which take place in thestructure of the bacterial cell, or the chemicalchanges which occur during lysis. The digestionof Streptococcus pyogenes by S. albus, involvedenzymes which liberated carbohydrate frompreparations of the cell wall of the microorganismand which also attacked casein (McCarty, 1952a).This report will present data on the microscopic,chemical, and biological aspects of the lysis anddigestion of the cell. Also, results on the liberationof spheroplast units from the streptococcus willbe given.
1 Supported by a grant-in-aid from the ChicagoHeart Association, National Heart Institute(H-3709), U. S. Public Health Service and theOffice of Naval Research (Nonr-1768 (00)).
2Established Investigator of the AmericanHeart Association.
METHODS
Culture of streptococci. The media used for thegrowth of both inocula and cells were brain heartinfusion broth (Difco) and Todd-Hewitt broth(Difco) supplemented with a glucose-phosphate-salts mixture (Hess and Slade, 1955).
Techniques for preparation of the media andinocula, and the centrifugation and washing ofthe cells were the same as used previously (Slade,1957).
Production of albus enzymes. S. albus was grownon the medium of McCarty (1952a) for 4 daysat 37 C. The pooled fluid from 36 Roux bottles(80 ml per bottle) was filtered through glass wooland coarse filter paper. The filtrate was cooledin ice, solid (NH4)2SO4 added to 70 per centsaturation, and the precipitated enzyme removedby filtration with Celite (1 g per L). The enzymewas extracted in 50 ml cold 0.1 M phosphatebuffer, pH 8.0, and separated from the Celiteby filtration. The Celite was washed twice withcold buffer, and the combined filtrates dialyzedovernight, and lyophilized. The material wassoluble in 0.1 M phosphate pH 8.0. One mg re-duced the optical density of a suspension ofstreptococci to 50 per cent of its original value(0.72 at 660 m,u) in 12 min at 37 C.
Cell lysis. The enzymatic digestion and lysisof the cells was followed by measurement of thechanges in optical density at 660 m,u duringincubation at 37 C. Cell suspensions were usuallyadjusted to give an original optical density valueof about 0.5. Control suspensions in water showeda drop in optical density which reached about20 per cent in 1 hr or more, depending on thestrain. The change with strain 12RN, however,amounted to almost 50 per cent. A similar de-crease in optical density also occurred in thepresence of molar sucrose, although the rate ofdecline was much lower.
103
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SLADE AND SLAMP
In the presence of sucrose the refractive indexof the medium was raised. This resulted in a de-pression of the optical density of the cell suspen-sion as compared to the control suspension inwater. Molar sucrose produced a depression of30 per cent. In most cases lysis in sucrose wascarried out in 0.2 M phosphate buffer, pH 8.0,containing 0.5 mg lyophilized enzyme, cells,sucrose, and water to 3 ml.
Enumeration of cell units. Counts were madein a Petroff-Hausser counting chamber. A smoothsuspension was obtained by repeated aspirationthrough a 20-gauge needle. A suspension in molarsucrose with an optical density of 0.25 was foundto contain approximately 65 cells per 0.0025 sqmm. In most cases it was necessary to wait severalminutes after the chamber was filled before countswere made. "Cell units" refers to intact cocci(with or without cell wall) which have not beenlysed.
Preparation of albus antibody. Antibody to thealbus enzymes was prepared by injection of theprecipitated enzyme into a rabbit. Six injectionsof 8 mg each were given intravenously every otherday. One week after the final injection the animalwas bled from the heart.
Preparation of ultrathin sections of streptococci.Samples were withdrawn from cell-enzyme mix-tures, cooled in ice, and centrifuged. Three ml of1.7 per cent osmic acid in 0.85 per cent NaClwas added to the packed cells. The suspensionwas held at room temperature for 17 hr.The cells were centrifuged and washed three
times in water and then dehydrated by one washeach in 50, 70, 80, and 95 per cent ethanol. Threewashes were then carried out in absolute ethanol.The cells were suspended in a small volume ofabsolute ethanol and an equal volume of meth-acrylate (9 parts butyl to 1 part methyl) wasadded. After centrifugation the cells were passedthrough three changes in methaerylate and finallyimbedded in methaerylate containing 1 per centbenzoyl peroxide. The preparations were heldat 55 C for 64 hr. Sections of the blocks were cuton a Porter-Blum microtome.
Chemical methods. Rhamnose was determinedby the method of Dische and Shettles (1948).All samples were hydrolyzed in H2SO4 at 100 Cas specified. The method of Palmer et al. (1937)was used to determine hexosamine. To obtainmaximum recovery of hexosamine from lyo-philized streptococci it was necessary to heat thecells in 4 N HCl in a sealed vial in boiling water
for 4 hr. Under these conditions the recovery ofthe glucosamine standard was 96 per cent. Totalnucleic acid was determined by the method ofSchneider (1945).
Lysates from the cells were centrifuged beforeany chemical determinations were made. Sampleswhich were removed up to 150 min of incubationcould be clarified by centrifugation at 10,000 X Gfor 20 min. Samples removed at approximately5 hr required 20,000 X G for 30 min.
Preparation of cell walls. Washed cells werepacked by centrifugation at 10,000 X G for 10min, drained, weighed, and made to a 15 percent suspension in water. A mixture of 5 ml cellsand 5 g Ballotini beads was placed in each cupof a Mickle shaker and shaken at maximumamplitude for approximately 20 min. Each batchof cells was checked by Gram stain to verifythat breakage was complete. Some strains re-quired more shaking than others. The beads wereremoved on a coarse glass filter, the cell wallssedimented and washed twice at 12,500 X G for10 min, and then lyophilized.Approximately 100 mg cell walls were sus-
pended in 8 ml of 0.1 M phosphate buffer (pH 7.7)containing 2 mg each of crystalline trypsin andribonuclease. Digestion was carried out at 37 Cfor 4 hr and the material centrifuged at 10,000 XG for 10 min. The walls were washed twice with10 ml water, digested for 21 hr at 37 C by 10 mgcrystalline pepsin in 8 ml of 0.02 N HCI, centri-fuged, washed twice, and lyophilized. All prepa-rations were examined in the electron microscopeto verify the absence of cytoplasmic material.
RESULTS
General aspects of lysis. Curve 1 of figure 1illustrates the optical density change when 0.20mg enzyme was incubated with 3 ml cell suspen-sion (strain T1). Curve 2 shows that the totaldecrease in the control approximated 20 per cent.It can be seen that 40 min elapsed before a sig-nificant difference appeared in the optical densitybetween the experimental and the control tubes.The decrease in the presence of enzyme was linearuntil a change of 65 per cent was reached. Therate then decreased and the final value of 95 percent was attained in about 200 min.
Microscopic examination of the cell suspensionsduring the lytic process revealed that the cellunits became more refractile. Also, the chains ofcocei began to separate into pairs and single cellssoon after the enzyme was added. Under the con-
104 [VOL. 79
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

LYSIS AND DIGESTION OF STREPTOCOCCI
'J~4o-
a x0 4o a800o /1*o
AMen utessFigure 1. Percentage decrease in optical density (curve 1, experimental; curve 2, control) and decrease
in number of cell units per 0.0025 sq mm (curve 3) in suspension of Streptococcus pyogenes strain Ti at37 C in presence of albus enzyme preparation. Cells (6 mg) and enzyme (0.8 mg) were suspended in 3 ml0.15 M phosphate buffer, pH 8.0.
ditions described in figure 1 the process was com-plete within 15 min. The separation of the cellsinto singles and pairs is considered to be due toeither an enzyme in the albus preparation or anenzyme elaborated by the streptococcus. Evi-dence for the formation of such an enzyme fromStreptococcus faecalis was recently reported byLominski et al. (1958).Examination by Gram stain during the diges-
tion period showed a 50:50 ratio of gram-positiveto gram-negative cells at an optical densitychange of 25 per cent. At 50 per cent opticaldensity change all the cells were gram-negative.At the conclusion of the lytic period all materialstained gram-negative.A number of other group A strains (S43, N19,
T23, and 12RN) representing various serologicaltypes were examined. All followed a similar pat-tern, although in some cases a difference as muchas 4-fold existed in rate of lysis when tested withthe same enzyme preparation. No significantdifferences were noted between cells harvestedfrom the growth medium after 6 hr or 17 hr ofincubation. Cells stored for several days in the
refrigerator showed no change in their rate oflysis.The effect of various enzyme inhibitors on the
rate of lysis was determined. Arsenite, dinitro-phenol, azide, fluoride, iodoacetate, and mercuricchloride were tested in a concentration of 5,umoles per 3 ml cell suspension. The lattershowed 100 per cent inhibition, all others werewithout significant effect. A reduction in themercuric chloride concentration to 1 ,umoleper 3 ml decreased the inhibition about 80 percent.The C and M antigens of the group A strepto-
coccus are located in part on the surface of thecell. It was of interest to investigate the effectof adsorption of antibody to these antigens on therate of lysis. A volume of cell suspension in waterwas added to an equal volume of specific rabbitantiserum and NaCl added to 0.85 per cent. Themixture was incubated at 37 C for 4½f hr, thecells centrifuged, washed once, and resuspendedin phosphate buffer and enzyme. No change inrate of lysis was detected. This result indicatesthat in spite of adsorption of antibody protein
1960] 105
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SLADE AND SLAMP
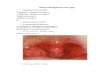
Figure 2. Control cells (strain Ti) of Streptococcus pyogenes removed at zero time. Ultrathin section.Cells harvested after 17 hr at 37 C.
to the cell surface antigen the lytic enzymes wereable to reach the cell wall substrate.Numbers of cell units. Optical density measure-
ments do not always provide a reliable indicationof the lysis of bacterial cells because of the light-scattering effect of the cell material released uponlysis. The reductionl in numbers of intact strepto-cocci was therefore followed by cell counts in acounting chamber under a phase contrast mag-nification of 970 X. Figure 1 (curve 3) shows thatthe number of cell units remained essentiallyconstant for the first 50 min. During this sameperiod, however, the optical density in the pres-ence of enzyme decreased 34 per cent whereasthe control showed a reduction of only 20 percent. The difference between these two values(14 per cent) was considered to represent a partial
removal of the cell walls, or other action on thecocci by the albus enzyme preliminary to lysis.This value has never been found to exceed 20per cent without a decrease in the original numberof cell units. Immediately after the 50-min period,the number of cell units began to decreasesharply, and the decrease continued for the next60 min. The over-all change was from 63 to 6units per 0.0025 sq mm.
It should be pointed out that the concentrationof both cells and enzyme must be carefully ad-justed if the above processes are to be seen.With an excess of enzyme the digestion pro-ceeded so rapidly that it was not possible, forphysical reasons, to detect the period of digestionin which the number of cell units remainedconstant.
[VOL. 79106
i
A.
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

LYSIS AND DIGESTION OF STREPTOCOCCI
Microscopic aspects of lysis. It was considerednecessary at this point in the investigation tomake electron microscopic observations at variousstages of the lytic process. It was clear that thinsections of the cells would be necessary in orderto obtain the information.
Preliminary examination of shadowed un-sectioned material removed from the cell-enzymesuspension at the end of the lytic period showedthe presence of large pieces of cell wall material.This observation indicated that enzymatic actionon the cell wall resulted in a rupture of the cellwith consequent loss of protoplasm.
Streptococci for microscopic examination weremixed with enzyme and buffer in a total volumeof 12 ml. The proportions of each were the sameas those used in the experiment described infigure 1. Optical density readings were made andsamples removed at appropriate intervals. Theoptical density changes given in figure 1 were used
as a guide to indicate the degree of lysis of thecells in the mixture.
Figure 2 illustrates control cells (no enzyme)which were removed at zero time. The charac-teristics of the normal cell are evident. In a fewcases the protoplast has shrunken and pulledaway from the cell wall (plasmolysis). All thecells eventually reached this stage as shown ina sample removed at the end of the experiment(90 min, figure 3). A few cells possessed rupturedcell walls due probably to autolysis or physicaldegeneration. It is evident, however, that thegreat majority of cells are intact even thoughmarked plasmolysis has occurred. The maximumoptical density decrease with most strains at thisstage was 20 per cent.
In contrast, streptococci which were attackedby albus enzyme presented a markedly differentappearance. Figure 4 illustrates cells removedat the beginning of the lytic phase (45 min,
Figure S. Control cells (strain T1) of Streptococcus pyogenes removed after 90 min in buffer. Ultra-thin section.
1960] 107
,.'z
Ei
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SLADE AND SLAMP
w t.1. --k* .: . t:.:.,:::::::. !' ......... .0i.h ... ..... . ..... ,65S: .. :-.:.:.. = t , :. :
*; .. 4 .:., ::e , ''::
.&.X ..
*. ,. i'* :0. : < :' ...........
Figure 4 Streptococcus pyogenes strain Ti after 45 min treatment with albus enzyme. Cells removedbefore lvtic phase. Ultrathin section.
figure 1). Many cells are partially emptied ofprotoplasm. In several cases (indicated byarrows) openings in the cell wall are clearly seen.Protoplasm in the form of strands has partlyemerged and can be seen in the vicinity of theopening in the cell wall. It should be noted thatthe opening in the wall is close to the point offormation of the new cross wall. This observationhas been made frequently in other preparations.The action of the enzyme very likely covers theentire surface of the cell but the cell wall may bethinner and consequently the break occurredfirst in the vicinity of the equatorial ring.
Incubation of the cocci for 90 min (figure 1)with albus enzyme resulted in the cells illustrated
in figure 5. The cells are for the most part fairlyfree of protoplasm and a considerable portionof the wall of each cell is present. This cell wallmaterial has always been observed at the end ofthe lytic phase with many group A strains ofvarious serological types. Continued incubationof the lysed suspension resulted in digestion ofthe cell wall material (see below).
Evidence to be presented will illustrate thatalbus culture filtrate also possessed an enzymewhich was able to destroy the protoplast mem-brane of the group A streptococcus. Figures 2 to5 do not provide information on the nature ofthe process, however it is logical to assume thatdigestion of the membrane began when damage
108 [VOL. 79
::... Impppr
.:. Im
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

LYSIS AND DIGESTION OF STREPTOCOCCI
to the cell wall was sufficient to allow the mem-brane enzyme to reach its substrate.
Biochemical aspects of lysis and digestion. Figure6 shows the rate of release of hexosamine andrhamnose from cells of strain Ti by albus enzyme(curves 1 and 3). These two carbohydrates makeup two-thirds of the weight of trypsin-treatedcell wall preparations from group A streptococci(McCarty, 1952b). It is evident that enzymaticdigestion began immediately. At the beginningof the lytic phase approximately 26 per cent of therhamnose and 38 per cent of the hexosamine had
been released. This was sufficient in the case ofmany cells to result in a release of cell protoplasm(figure 4). At this stage about 50 per cent of thetotal nucleic acid in the suspension was found inthe supernatant (curve 2, figure 6). At the endof the lytic period 45 per cent of the rhamnose,55 per cent of the hexosamine, and 70 per cent ofthe nucleic acid was free of the cell. Controlsuspensions showed no significant release ofeither rhamnose, hexosamine, or nucleic acid(Slade, 1957). The incubation with enzyme wascontinued for 5 hr but a complete solubilization
...... ....
e ^ . R,; ; . R: .. :B: . :. :o : : :*::''': .'; e'.'!t'R, :|- eg.-. SqS. '.:0 0: ! .i; AC.E
W -a6@>- ¢* otS I
'4'-M
.N
Figure 5. Streptococcus pyogenes strain Tl after 90 min treatment with albus enzyme. Cells removedat end of lytic phase. Ultrathin section.
1960] 1.09
NW:
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SLADE AND SLAMP
L8o
12Z
0/to 200 300
k.~ ~ ~
?)/flL tesFigure 6. Release of rhamnose (curve 3), hexosamine (curve 1), and nucleic acid (c;urve 2) from Strepto-
coccus pyogenes strain Ti at 37 C by albus enzyme preparation. Cells (59 mg) and enzyme (10 mg) weresuspended in 30 ml 0.15 M phosphate buffer, pH 8.0.
of the nucleic acid was not obtained. The nucleicacid remaining was probably associated withmacromolecular components of the cell wall.Similar results were obtained with strains T3,12RN, S43, and N19, except that in two casesthe nucleic acid values reached 82 per cent.The lysates after extended incubation always
contained a residue which varied in rhamnosecontent from 10 to 15 per cent. On the otherhand, hexosamine residues when present con-tained 5 per cent or less of carbohydrate. Therhamnose in the complete lysates was usually ina nondialyzable form whereas 18 to 24 per centof the hexosamine could be dialyzed.
Cell walls from strains Ti, S23, and S43 were
also examined during digestion by albus en-zymes. Similar curves as in figure 6 for the releaseof carbohydrate were obtained. Rhamnose resi-dues were encountered which are probably similarto those found by McCarty (1952b).The lysates from both whole cells and cell walls
did not contain M protein as judged by lack ofreaction with specific antisera. The albus prepa-rations very likely contain an enzyme capable ofdigesting the type-specific protein.
Lysis in sucrose. Sucrose solutions of highosmotic pressure will maintain the integrity ofprotoplasts upon their release from group Astreptococci (Gooder and Maxted, 1958; Freimeret al., 1959). It was considered of interest to in-
[VOL. 79110
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

LYSIS AND DIGESTION OF STREPTOCOCCI
vestigate the lysis of streptococci in sucrose byalbus enzyme. Cell suspensions of several strainswere tested in 0.25 to 2.0 M sucrose. Lysis oc-curred in every case although the rate was re-duced significantly in the higher concentrationsof sucrose. It seemed likely therefore that thealbus preparations contained an enzyme(s) whichdigested the protoplast membrane, in additionto those which attacked the cell wall.
Further investigations were carried out. Lysisin sucrose was prevented when, before the lyticphase, the cell suspension was cooled to 5 C.The temperature-dependent nature of the processindicated the final stage of lysis was enzymedependent. In addition it was considered likelythat an enzyme, if present, would produce anti-bodies that could be used to inhibit action of theenzyme. Lysis due to a nonenzymatic reactionwould not be inhibited by antibody. The injectionof albus preparations into rabbits was found toresult in the formation of antibodies which pre-vented lysis of group A streptococci.The following experiment was performed. The
cell units were harvested before the lytic periodto be certain that enzymatic digestion of the cellwall had occurred. The following materials wereused: 1.7 ml of 10 per cent cell suspension, 2.5ml of 2 M phosphate buffer, pH 8.0, 12.5 ml of 2 Msucrose, and 8 mg enzyme in 8.3 ml water. Themixture was incubated in a water bath at 37 C andplunged into ice when the difference in opticaldensity of the control and enzyme suspensionreached 15 per cent (figure 1).The cooled tubes were centrifuged for 30 min
at 4000 X G at 5 C. The material sedimented asa ball-shaped pellet of sticky consistency. It wasnecessary to draw the material through a hypo-dermic needle several times to obtain an evensuspension. The centrifugation was repeatedand the washed cell units resuspended in 3 mlof cold M sucrose.
Incubation of the treated suspension at 2 Cand 37 C in sucrose resulted in a slow lysis ofthe cell units at the higher but not the lowertemperature. This was considered to be due tothe presence of traces of cytoplasmic membraneenzyme adhering to the washed cell units. One-half ml antiserum in molar sucrose was thenadded and mixed with the suspension. The mix-ture was held at 3 C for ' hr, centrifuged asabove and resuspended in 1 ml of M sucrose.
Optical density measurements at 37 C showed
very little change in the preparation treated withantibody whereas the control preparation de-creased 70 per cent in 2}1 hr. These results in-dicate that lysis of the streptococei in sucrose ofsufficient molarity to prevent osmotic lysis wasprobably due to an enzyme active against theprotoplast membrane.
DISCUSSION
It is significant that complete removal of thecell wall is not necessary for lysis to occur. Inthis respect the action of S. albus enzymes onstreptococci may be similar to the autolysis ofB. cereus (Norris, 1957), although in the lattercase no chemical data were given as to degrada-tion of the cell wall. Streptococci harvested beforethe lytic phase may be similar to Escherichia coligrown in the presence of penicillin or in the ab-sence of diaminopimelic acid (McQuillen, 1958)in that they possess degraded cell wall. Brenneret al. (1958) have suggested the term "sphero-plast" be used to designate these structures. Ithas been found in the present work that ap-proximately 50 to 60 per cent of the spheroplastunits were gram-negative, most were viable andcapable of reproduction, and precipitin tests(Swift et al., 1943) for group- and type-specificantigens were positive. They were stable in waterfor many hours although about 30 per cent of thecell wall had been removed by enzyme action(figure 6). Further studies on the biological ac-tivity of these units will be of interest.
ACKNOWLEDGMENTS
The authors wish to express their appreciationto Dr. J. P. Marbarger and Miss Irena Kairysof the University of Illinois School of Medicinefor making available the electron microscope andfor generous assistance in the microscopic work.
SUMMARY
The process of lysis of cells of Streptococcuspyogenes by enzymes from Streptomyces albusis described. Results based on observations ofelectron microscope ultrathin sections, releaseof carbohydrate from the cell wall, and releaseof nucleic acid from the protoplast indicate thatenzymes active against the cell wall and theprotoplast membrane participate in the lyticprocess. Lysis occurs in molar sucrose solutionas well as in water. Anti-albus rabbit serumeffectively prevents the lysis in sucrose of strep-
19601 ill
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SLADE AND SLAMP
tococci possessing degraded cell walls. The prop-erties of these cells are similar to those of sphero-plasts.
Lysis is characterized by a period in which thenumber of cells remains constant while release ofcarbohydrate from the cell wall occurs. This isfollowed by a reduction to zero in numbers ofcells. At this stage large remnants of cell wall arepresent which contain approximately one halfof the carbohydrate of the original cell wall.Continued incubation results in digestion of thismaterial.
REFERENCES
BRENNER, S., DARK, F. A., GERHARDT, P., JEYNES,J. H., KANDLER, 0., KELLENBERGER, E.,KLEINBERGER-NOBEL, E., MCQUILLEN, K.,RuBIO-HUERTOS, M., SALTON, M. R. J.,STRANGE, R. E., TOMCSIK, J., AND WEIBULI,C. 1958 Bacterial protoplasts. Natuire,181, 1713-1715.
DISCHE, Z. AND SHETTLES, L. B. 1948 A specificcolor reaction of methylpentoses and aspectrophotometric method for their deter-mination. J. Biol. Chem., 175, 595-603.
EVANS, A. C. 1940 The potency of nascentstreptococcus bacteriophage B. J. Bacteriol,39, 597-604.
EVANS, A. C. 1942 Technique for the determina-tion of the sensitivity of a strain of strepto-coccus to bacteriophage of type A, B, C, or D.J. Bacteriol, 44, 207-214.
FREIMER, E. H., KRAUSE, R. M., AND MCCARTY,M. 1959 Protoplasts and L-forms of groupA streptococci. Bacteriol. Proc., 1959, 69.
GOODER, H. AND MAXTED, W. R. 1958 Proto-plasts of Beta-hemolytic streptococci. Na-ture, 182, 808-809.
HEss, E. L. AND SLADE, H. D. 1955 An electro-phoretic examination of cell free extracts fromvarious serological types of group A hemolyticstreptococci. Biochim. et Biophys. Acta,16, 346-353.
KRAUSE, R. M. 1957 Studies on bacteriophagesof hemolytic streptococci. I. Factors influ-encing the interaction of phage and sus-ceptible host cell. J. Exptl. Med., 106,365-384.
LOMINSKI, I., CAMERON, J., AND WYLLIE, G. 1958Chaining and unchaining Streptococcus fae-calis; a hypothesis of the mechanism ofbacterial cell separation. Nature, 181, 1477.
MAXTED, W. R. 1948 Preparation of strepto-coccal extracts for Lancefield grouping.Lancet, 2, 255-256.
MAXTED, W. R. 1957 The active agent in nas-cent phage lysis of streptococci. J. Gen.Microbiol., 16, 584-595.
MCCARTY, M. 1952a The lysis of group Ahemolytic streptococci by extracellular en-zymes of Streptomyces albus. I. Productionand fractionation of the lytic enzymes. J.Exptl. Med., 96, 555-568.
MCCARTY, M. 1952b The lysis of group Ahemolytic streptococci by extracellular en-zymes of Streptomyces albus. II. Nature ofthe cellular substrate attacked by the lyticenzymes. J. Exptl. Med., 96, 569-580.
MCQUILLEN, K. 1958 Bacterial "protoplasts":effects of diaminopimelic acid deprival andpenicillin addition compared in E. coli.Biochim. et Biophys. Acta, 27, 410-411.
NORRIS, J. R. 1957 A bacteriolytic principleassociated with cultures of Bacillus cereus.J. Gen. Microbiol., 16, 1-8.
PALMER, J. W., SMYTHE, E. M., AND MEYER,K. J. 1937 On glyco-proteins. IV. Estima-tion of hexosamine. J. Biol. Chem., 119,491-500.
SALTON, M. R. J. 1953 Studies on the bacterialcell wall. IV. The composition of the cellwalls of some gram-positive and gram-nega-tive bacteria. Biochim. et Biophys. Acta,10, 512-523.
SCHNEIDER, W. C. 1945 Phosphorus compoundsin animal tissues. I. Extraction and estima-tion of deoxypentosenucleic acid and pentose-nucleic acid. J. Biol. Chem., 161, 293-303.
SLADE, H. D. 1957 Studies on Streptococcuspyogenes. III. The effect of trypsin and acationic detergent on the structure, perme-ability and metabolism of the cell. J. Gen.Physiol., 41, 63-76.
SWIFT, H. F., WILSON, A. T., AND LANCEFIELD,R. C. 1943 Typing group A hemolyticstreptococci by M precipitin reactions incapillary pipettes. J. Exptl. Med., 78,127-133.
112 [VOL. 79
on February 3, 2019 by guest
http://jb.asm.org/
Dow
nloaded from