Embed Size (px)
Citation preview

STUDIES IN POPULATION PHYSIOLOGY
VIII. THE EFFECT OF LARVAL POPULATION DENSITY ON THE POST-
EMBRYONIC DEVELOPMENT OF THE FLOUR BEETLE,
TRIBOLIUM CONFUSUM DT??AL
THOMAS PARK Department of Biology, School of Hygiene and Public Health, Johns Hopleilzs
University and H ~ l l Zoological Laboratory, The University of Chimgo
THREE FIGURES
INTRODUCTION
Experimental studies of insect populations recently have received considerable attention. Most of these investigations have emphasized primarily the effect of imago population density on fecundity, fertility and mortality. From this viewpoint, Pearl ( '32) has reported Drosophila findings ; MacLagan and Dunn ( '35) have discussed weevil populations ; Salt ('36) has dealt with certain insect parasites, and Chap- man ('28), Park ('37) and Park and Woollcott ('37) have emphasized populations of the flour beetle, Tribolium con- fusum. It is both natural and fortunate that reproduction and life-duration have been subj.ected to such intensive analysis, for, as Pearl ( '27) has pointed out, these variables constitute the two most basic factors of population dynamics- birth rate and death rate.
However, despite these and other studies there is real need for the investigation of population phenomena using, rather than the adult organism, the larval and pupal stage as the experimental unit. It is patent that in insect cultures, where a considerable portion of the life history is spent in post- embryonic development, the ' efficiency' of metamorphosis in a statistical sense is an important factor influencing the future
51
THE JOURXAI, OF EXPZRINENWL ZOOLORY, VOL. 79, KO. 1

52 THOMAS PARK
growth of the population. Using the flour beetle Tribolium confusum the present investigation attempts to form some concrete evaluation of this point by analyzing the effect of differential larval density upon 1) the mortality of larval development ; 2) the duration or rate of larval development; 3) the mortality of pupal development; 4) the duration of the subsequent pupal period, and 5) the weight of the sub- sequent pupal and imago stages.
METHODS AND PROCEDURE
The husbandry of Tribolium has been described elsewhere (Park, ’34) and it is merely necessary at this place to point out that the entire life cycle of the beetle is passed in flour. By sifting the medium the larvae, as well as the eggs, pupae and imagoes, can be collected for counting and observation. In the present experiments the larvae were counted on a large steel plate that had been enameled glossy black. This aided in observing the small, early instar stages and facilitated their handling. All the cultures were run in darkened incubators at a temperature of 28”C., and at a relative humidity of ap- proximately 40%. ‘Ceresota,’ a white flour, mas used as the medium.
The present investigation was organized about three dis- tinct experiments. The first experiment consisted of 127 small vials (approximately 76 x 30 mm.) each containing 10 gm. of flour and seeded initially with larvae ranging in number from one to sixty-four per container. The larvae were chosen from a large group of eggs and were selected at random at the time of hatching. This insured that no important age differences obtained between larvae at the start of the err- periment. All eggs came from the same in-bred stocks used in previous work. The larvae wer’e introduced into their respective bottles at the start of the experiment and placed in an incubator for 48 hours. The bottles were then removed, the flour sifted, and the larvae counted and examined. Dead larvae were recorded and replaced from extra bottles that exactly duplicated the experimental groups. After examina- tion and replacement the larvae were returned to the same

STUDIES IN POPULATION PHYSIOLOGY. VIII 53
flour and left alone for another 48 hours when the procedure was repeated. This was kept up during the period of metn- morphosis until pupation occurred. While in the pupating stage the bottles were examined for pupae every 12 hours so that the larval period could be timed accurately to the 4 day. As the pupae were found they were removed from their cultures, numbered, dated, sexed, weighed to the nearest 0.1 mg., and isolated singly into small shell vials containing 4 gm. of fresh flour. These vials were examined twice daily and at the time of emergence the length of the pupal period was noted and the young imago beetle was weighed. Pupal deaths were recorded but dead pupae were not replaced.
The first experiment, designated in this paper the ‘lO-agm.- not replaced series,’ was designed so that there were 64 bottles each containing 1 larva; 32 bottles each containing 2 larvae; 16 bottles with 4 larvae each; 8 bottles with 8 larvae each; 4 bottles with 16 larvae each; 2 bottles with 32 larvae each, and 1 bottle with 64 larvae. I n other words, each density series was represented by a total of 64 larvae. This total was maintained until pupation by making the replacements earlier mentioned.
The second analysis duplicated the first in experimental procedure except that 4 gm. of flour was used per bottle in- stead of 10 gm., and that the two were not run simultaneously. As previously the flour was not renewed. For experiment 2, hereafter designated the ‘4-gm.-not replaced series, the fol- lowing cultures were set up: 64 bottles with 1 larva each; 16 bottles 156th 4 larvae each; 4 bottles with 16 larvae each; 2 bottles with 32 larvae each; 2 bottles with 64 larvae each; 1 bottle containing 128 larvae, and 1 bottle with 256 larvae.
The third experiment also was similar to the others except that the 4 gm. of flour used in each bottle was replaced with fresh flour at the time of each count or every 48 hours. This experiment, the ‘4gm.-replaced series, ’ consisted of 64 bottles with 1 larva each; 16 bottles with 4 larvae each; 2 bottles with 32 larvae each; 2 bottles with 64 larvae each and 1 bottle with 128 larvae.

54 THOMAS PARK
The density relations for the three experiments are sum- marized in the following protocol :
I. The 10-=.-flour not replaced series (total, 127 bottles) A. 64 bottles: larval density 1. B. 32 bottles: larval density 2. C. 16 bottles : larval density 4. D. 8 bottles: larval density 8. E. 4 bottles: larval density 16. F. 2 bottles: larval density 32. G. 1 bottle: larval density 64.
11. The 4-gm.-flour not replaced series (total, 90 bottles) A. 64 bottles: larval density 1. B. 16 bottles: larval density 4. C. 4 bottles: larval density 16. D. 2 bottles: larval density 32. E. 2 bottles: larval density 64. F. 1 bottle: larval density 128. G. 1 bottle: larval density 256.
III. The I-gm.-flour replaced series (total, 85 bottles) A. 64 bottles : larval density 1. B. 16 bottles: larval density 4. C. 2 bottles : larval density 32. D. 2 bottles : larval density 64. E. 1 bottle: larval density 128.
Certain obvious differentials are set up between these ex- perimental series. All three vary the factor of larval density but in each the crowding has a different meaning. It will be one of the tasks of this paper to interpret these meanings as the discussion develops.
THE RELATION O F LARVAL MORTALITY TO DENSITY
From the population viewpoint two aspects of larval development are important. These are 1) larval mortality and 2) duration or rate of larval metamorphosis. I n a statisti- cal sense these two criteria appraise the ‘efficiency’ of the entire larval period. It is obvious that in cultures where development is uniformly rapid and where a minimal number of deaths occur the general growth of the population, other things being equal, will be favored. It is also obvious that in growing populations the larval density relationships are
STUDIES IN POPULATION PHYSIOLOGY. VIII 55
changing. Therefore, it becomes important to analyze experi- mentally such changes in the hope that they will aid in under- standing the mechanisms involved in the control of population trends.
There has been very little experimental work on the relation of population density to insect larval mortality. Park ('35) approached the problem and showed that Tribolium larvae
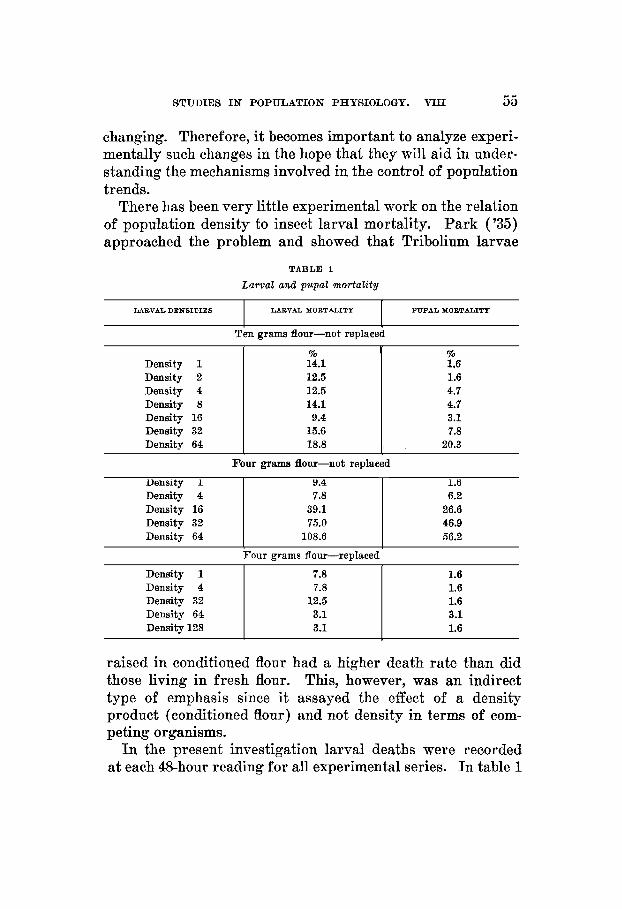
TBBLE 1
Larval and pupal mortality
LARVAL DENSITIES I LAEVAL MORTALITY I PUPAL MOBTALITY
T& gram flour-not replaced
Density 1 Density 2 Density 4 Density 8 Density 16 Density 32 Density 64
% 14.1 12.5 12.5 14.1 9.4 15.6 18.8
Four grams flour-not replaced
Density 16 Density 32 Density 64 108.6
% 1.6 1.6 4.7 4.7 3.1 7.8 20.3
1.6 6.2 26.6 46.9 56.2
Four grams flour-replaced
Density 1 Density 4 Density 32
7.8 7.8 12.5
1.6 1.6 1.6
Density 64 3.1 3.1 Density 128 3.1 1.6 I I
raised in conditioned flour had a higher death rate than did those living in fresh flour. This, however, was an indirect type of emphasis since it assayed the effect of a density product (conditioned flour) and not density in terms of com- peting organisms.
In the present investigation larval deaths were recorded a t each 48-hour reading for all experimental series. In table 1
56 THOMAS PARK
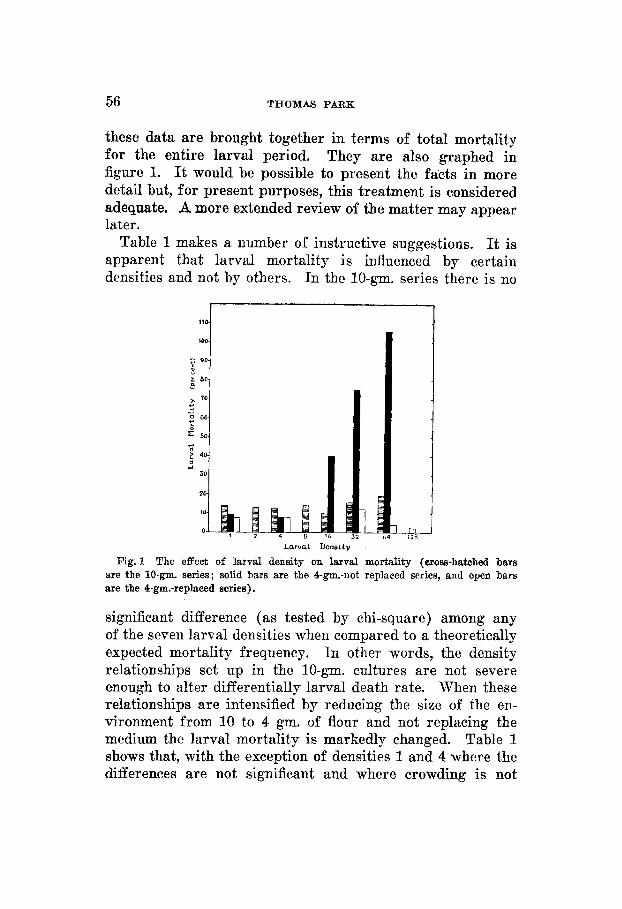
these data are brought together in terms of total mortality for the entire larval period. They are also graphed in figure 1. It would be possible to present the facts in more detail but, for present purposes, this treatment is considered adequate. A more extended review of the matter may appear later.
Table 1 makes a number of instructive suggestions. It is apparent that larval mortality is influenced by certain densities and not by others. In the 10-gm. series there is no
110 I
2 4 8 16 32 64 128
Larval Densrty
Fig. 1 The effect of larval density on larval mortality (cross-hatched bars are the 10-gm. series; solid bars are the 4-gm.-not replaced series, and open bars are the 4-gm.-replaced series).
significant difference (as tested by chi-square) among any of the seven larval densities when compared to a theoretically expected mortality frequency. In other words, the density relationships set up in the 10-gm. cultures are not severe enough to alter differentially larval death rate. When these relationships are intensified by reducing the size of the en- vironment from 10 to 4 gm. of flour and not replacing the medium the larval mortality is markedly changed. Table 1 shows that, with the exception of densities 1 and 4 where the differences are not significant and where crowding is not

STUDIES I N POPULATION PHYSIOLOGY. VIII 57
severe, there is a proportionate increase in number of deaths as the cultures get more thickly populated. The mortality became so excessive in the 128 and 256 cultures that it was impossible to run them to completion and they had to be dis- continued. Even in the sixty-four group more larvae died than were initially introduced. It is plain that under the conditions obtaining in the ‘4-gm.-not replaced series ’ density is a potent factor in causing larval death.
The third experiment is important in that it suggests an interpretation of the density effect just described. Figure 1 shows that when the flour is renewed at each count the larval mortality stands in no consistent relation to density even though the actual degree of crowding exactly parallels that obtaining in the second experiment. How can this fact be interpreted? The obvious inference is that it is primarily the effect the larvae have on their medium which is important in increasing their death rate and not the behavioristic inter- actions between the organisms themselves. In other words, it is the ‘conditioning’ of the flour-its alteration through biological activity-that is the influential factor. It is inter- esting to note in passing that larval mortality is lower for the 4-gm.-replaced series than for the 10-gm. group, This difference, slightly significant, supports the view that the condition of the flour is more important in affecting larval mortality than the crowding of the beetles.
The main conclusions growing out of these data are 1) that density below a certain threshold is not effective in increasing larval mortality; 2) that density if severe enough may exert profound influences on larval death rate, and 3) that density operates primarily through the alteration or biological con- ditioning of the medium. This alteration certainly represents a reduction in the food content of the flour and may reflect other changes as well.
There is one puzzling point arising in the data that should be mentioned. In table 1 the mortality for the larvae reared in 10 gm. of flour at a density of 64 is 18.8% and that for the larvae reared in 4 gm. of unreplaced flour at a density of 16

58 THOMAS PARK
Density 1 Density 4 Density 16 Density 32 Density 64
is 39.1%. Each larva in the former unit has 0.15 am. of flour at its disposal while each larva of the latter has 0.25 In other words, the mortality is appreciably lower in the more crowded culture. This would not be anticipated either on the basis of the general results or on the grounds that reduction
55.5 31.5 38.3 f 0.41 4.9 rt 0.28 12.8 k0 .76 64 61.5 32.0 38.7 k 0.41 4.9 -C 0.28 12.6 2 0.74 64 75.5 37.0 52.0 2 0.95 11.3 zk 0.67 21.6 zk 1.28 64
166.0 51.5 92.1 f 2.53 30.1 t 1.78 32.6 k 2.10 64 222.0 63.5 141.4 2 2.22 37.8 t 1.59 26.4 f 1.11 128
TABLE 2
Protocol for duration of larval period I I I I
Density 1 Density 4 Density 32 Density 64 Density 128
MEAN LENGTH w * L N D ~ D COEFFICIENT NUYBm MT I LARVAL I I O F I OF ~~~i~~~ ~ i ~ i ~ ~ ~ PERIOD DEVIAT1oN VAEIABI&ITP LABVAE
46.5 30.5 34.0 t 0.28 3.3 20.19 9.7 20.59 64 45.5 27.5 33.8 zk 0.29 3.5 zk 0.20 10.3 k 0.61 64 46.0 29.0 35.8 k 0.20 3.5 zk 0.14 9.8 zk 0.41 64 57.0 29.5 36.9 k 0.28 4.7 2 0.19 12.8 2 0.53 128 63.0 30.0 40.3 2 0.35 6.0 f. 0.24 14.9 f 0.62 128
-VAL DENSXTIE 0
Ten grams flour-not replaced
Density 1 Density 2 Density 4 Density 8 Density 16 Density 32 Density 64
days 37.6 f 0.57 36.9 f 0.47 41.6 f 0.43 43.1 f 0.50 43.4 f 0.75 54.8 & 0.77 66.9 r+ 1.21
days 6.8 2 0.40 5.6 -+ 0.33 5.2 rt 0.30 6.0 rt 0.35 9.0 rt 0.53 9.2 t 0.54 .4.4 t 0.85
15.2 f 0.90 12.5 f 0.80 13.9 2 0.82 20.7 2 1.23 16.7 f 0.99 21.5 f 1.28
64 64 64 64 64 64 64
of the food content of the flour is the primary factor in in- creasing larval death rate. These particular data suggest that there may be influences operating other than the altera- tion of the medium. However, they do not invalidate the conclusion that conditioning is the most important single influence associated with increased larval mortality.

STUDIES IN POPULATION PHYSIOLOGY. \TI1 59
DERATION OF THE LARVAL PERIOD
As described in an earlier section the present experiments were set up so that the length of the larval period could be timed to the nearest Q day. These data are summarized statistically for all three experimental series in table 2. They are graphed in figure 2.
Considering the 10-gm. experiment it is apparent that there is some correlation between density and the time required for larval metamorphosis. However, the contrasts between the
1 2 4 8 3 Z 64 110
Larval Dens i ty
Fig. 2 The effect of larval density on rate of larval development (cross-hatched bars are the 10-gm. series, solid bars are the 4-gm.-not replaced series, and bpen bare are the .l-gm.-repla.eed series).
slightly crowded cultures are not great and no decided density differential is evident until the 16, 32 and 64 groups are compared. For example, there is no significant difference between the one and two densities although the larvae in these two populations do metamorphose faster than the larvae in any of the other cultures. Likewise, the 4, 8 and 16 groups are essentially similar to each other but definitely below the 32 and 64 populations.
'Measured in the conventional way by calculating the ratio of the difference between any two uncorrelated means to the probable error of that difference.

60 THOMAS PAR.K
The picture is more diagrammatic for the 4-gm.-not re- placed series where, with the exception of the 1 and 4 densities, each group is significantly different and there is a proportional lengthening of larval development with crowding. This lengthening can be appraised roughly for both series by sub- tracting the mean length of the larval period for the single beetle cultures from the mean for some of the more crowded cultures. Certain of these differences appear in the following protocol :
Densitiea subtracted Diferences ( d a y s ) 10 gm. 4 gm.
64-1 29.3 103.1 32-1 17.2 53.8 16-1 5.8 13.7
These figures, all statistically significant, clearly bring out the effect of reducing the size of the environment. The larvae in the 10-gm. series metamorphose two to three times faster than those in the 4-gm. series. No data are available for the 128 and 256 densities for, as pointed out earlier, their mortality was so high and development so protracted that the experi- ments had to be discontinued.
One discrepancy in the data should be mentioned in passing. For the 10-gm. series the difference in rate of development between densities 1 and 4 is significant (5.6 times P.E.). The same difference for the 4-gm.-not replaced series is not signifi- cant (0.6 times P.E.). This is curious in the light of the general results and no explanation is apparent at the moment. It is a point, however, to be kept in mind since it is the type of effect that Allee ('31, '34) has shown to be frequently real and important in population dynamics.
The results of the third experiment in which the 4 gm. of flour was replaced at each reading are of an entirely different nature from those just described. The chief differences are 1) that the general rate of larval development has increased and 2) that density is only slightly correlated with meta- morphosis. The first point is supported by the fact that the mean rate of development for density 1 of the replaced series is significantly lower than that for the other two experiments.

STUDIES I N POPULATION PHYSIOLOGY. VIII 61
The second point may be brought out by subtracting the mean rate for density 1 (34.0) given in table 2 from the mean for density 64 (36.9). This difference, 2.9 days, is to be compared with the 103.1 days obtain for the same densities of the 4-gm.- not replaced series and demonstrates the importance of flour renewal for larval metamorphosis. In other words, as was the case for mortality, density is operating primarily through the medium and not through immediate competitive inter- actions between the larvae themselves.
However, there does appear to be a slight but consistent density effect exhibited by the 4-p.-replaced cultures. In table 2 it is noteworthy that, with the exception of densities 1 and 4, there is a gradual but significant increase in the dura- tion of metamorphosis with increased crowding. For ex- ample, the beet.les reared in the 128 and 64 densities took, respectively, 6.3 and 3.4 days longer to complete their de- velopment than those reared in the isolated cultures. Due to the way this experiment is designed it is impossible to state absolutely whether these small differences are attributable to real interactions between the larvae, or, whether the Tribolium in crowded cultures condition their environment sufficiently in a 48-hour period to affect adversely their development. Possibly the first view is supported by the fact that the larvae in the 32 density, where crowding in terms of 2-day periods is slight? have a significantly longer (5.2 times P.E.) period of growth than the single-individual controls.
The final point to be made in connection with larval meta- morphosis is concerned with variability as related to density. I n table 2 parameters, the standard deviation and coefficient of variation, are presented for each experimental unit. These constants demonstrate no particular trend for the 10-gm. series but do indicate for the 4-gm.-not replaced cultures that larval development becomes an increasing variable phenome- non as crowding gets more severe. There is also some sug- gestion that this type of effect is occurring in the 4-gm.-re- placed populations since the standard deviations increased consistently with density. However, it is obvious that these

62 THOMAS PAR.K
changes are of less degree than those exhibited by the former series.
The conclusions reached about larval metamorphosis on the basis of this analysis are essentially similar to those developed for mortality. They are 1) that, with the exception of very slightly crowded cultures, the duration of the larval period increases proportionately with density; 2) that this effect is due primarily to influences the larvae exert on the flour, and 3) that increased variability in terms of developmental time is frequently characteristic for larvae of crowded popula- tions.
PUPAL AND IMAGO WEIGHT
The weighing of each pupa and imago individually was justified on the ground that the present investigation is es- sentially a growth study. Thus weight becomes a variable worthy of appraisal. Also, since rate of metamorphosis was investigated, it was thought interesting to see if larvae that developed rapidly attained an ultimate pupal and imago weight that was different from that for slowly developing forms.
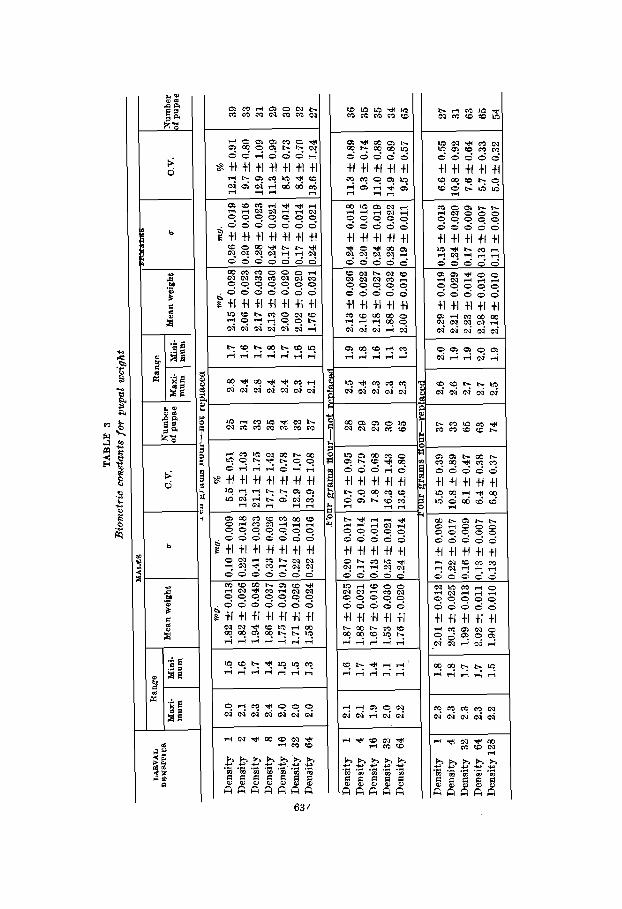
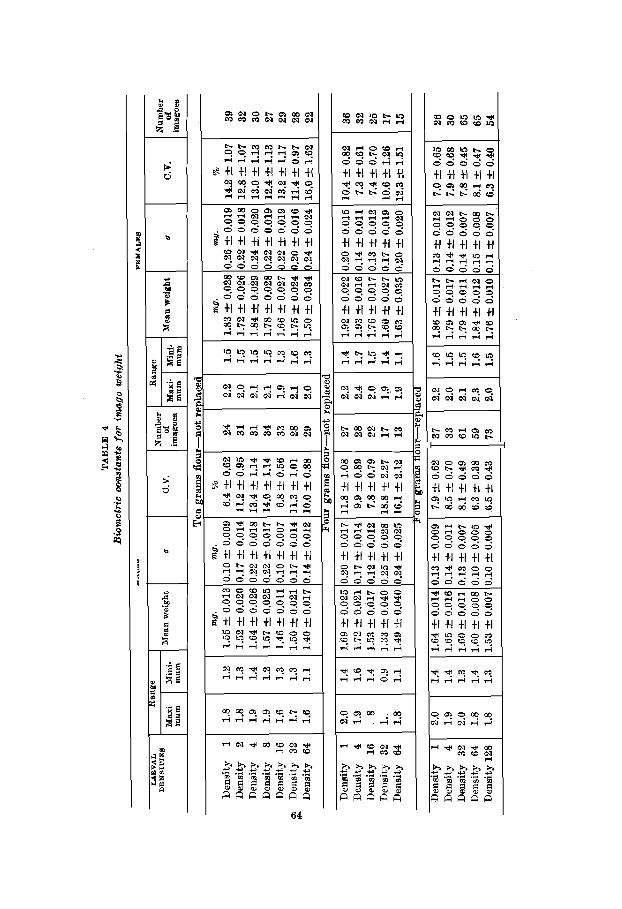
The data for pupal weight appear in table 3 and those for imago weight in table 4. The first point to be made is merely a substantiation of the fact (Brindley, '30; Park, '36) that female Tribolium weigh more than do males. The mean weights for male pupae and imagoes are consistently lower t*han those for females. For the pupae these sexual differences are significant in all experiments and there is only one border- line case for the imagoes where the difference is 2.7 greater than its probable error. It is very obvious 1) that there are real weight differences between the sexes and 2) that these differences are not appreciably altered by density. A glance at the standard deviations and coefficients of variability shows that no great or consistent differentials in variation exist between male and female pupae and imagoes.
The relation of larval density to pupal and imago weight is not as easily summarized as was larval mortality and rate of larval development. However, it seems possible to make certain positive points by grouping the data of tzbles 3 and 4.
WV
Ab
D
EN
SIT
IES
Den
sity
1
Den
sity
2
Den
sity
4
Den
sity
8
Den
sity
16
~
1
Den
sity
32
D
ensi
ty
64
W
MA
LE
S u8I.a
-a
C
U
Num
ber
c.v*
of
pupa
e
Rang
e
mum
m
um
- M
axi-
Min
i. M
ean
wei
ght
Num
ber
2.0
2.1
2.3
2.4
2.0
2.0
2.0
Den
sity
16
D
ensi
ty
32
Den
sity
64
mg.
mg.
1.82
t 0
.013
0.1
0 f O.O
OE
1.82
& 0
.026
0.2
2 f 0.
011
1.94
-C 0
.048
0.4
1 f 0.
032
1.86
+- 0
.037
0.3
3 f 0.
026
1.75
-C 0
.019
0.1
7 f 0.0
13
1.58
z!z
0.02
4 0.
22 f 0.
016 m',
1.5
1.6
1.7
1.4
1.5
1.3
1.5
1.71
-c
0.02
6 0.
22 2
0.0
18
Den
sity
I I
I I
12.1
+- 1
.03
21.1
f 1.
75
17.7
-C
1.4
2
r pr
ams
now
-not
10.7
& 0
.95
28
9.0 f 0.
79
29
7.8 f 0.
68
29
16.3
+- 1.
43
30
13.6
rlr 0
.80
65
Jur
gram
tiou
r-re
]
5.5
-C 0
.39
37
10.8
k 0
.89
33
8.1 f 0.
47
65
6.4 f 0.
38
63
6.8
-C 0
.37
74
uace
a
mn.
mg
. %
2.
8 1.
7 2.
15 f 0.
028
0.26
2 0
.019
12
.1 2
0.9
1 39
2.
4 1.
6 2.
06 4
0.0
23 0
.20
-+ 0.
016
9.7
4 0
.80
33
2.8
1.7
2.17
4 0
.033
0.2
8 4 0
.023
12
.9 -+
1.0
9 31
2.
4 1.
8 2.
13 -
C 0
.030
0.2
4 2
0.0
21
11
.3 f
0.99
29
2.
4 1.
7 2-
00 4
0.0
20
0.17
f 0.
014
8.5 f 0
.73
30
2.3
1.6
2.02
+. 0
.020
0.1
7 -C
0.0
14
8.4 5 0
.70
32
2.1
1.5
1.76
2 0
.031
0.2
4 f 0.
021
13.6
-C 1
.24
27
2.6
2.6
2.7
2.7
2.5
-
2.0
1.9
1.9
2.0
1.9 -
~
6.6
-C 0
.55
10.8
-t
0.92
7.
6 f 0.
64
5.7 f 0.
33
5.0 f 0.
32
27
31
63
65
54
LA
RV
AL
K
ange
N
umbe
r R
anee
M
axi.
yill
i-
Mea
n w
eigh
t U
'.V*
imag
oes
of
5;~
M
ni-
M
ean
wei
ght
C
C.V.
DE
NS
ITIE
S
mum
m
um
mum
Den
sity
1
Den
sity
2
Den
sity
4
Den
sity
8
Den
sity
16
g
Den
sity
32
D
ensi
ty
64
Num
ber
of
imag
oes
1.8
1.8
1.9
1.9
1.6
1.7
1.6
m0.
mo.
1.
83 -t
0.02
8 0.2
6 &
0.0
19
1.72
2 0
.026
0.
22 2
0.0
18
1.84
f 0.
029
0.24
& 0
.020
1.
78 2
0.0
28
0.22
2 0
.019
1.
66 1
0.02
7 0.
22 &
0.0
19
1.75
f 0.
024
0.20
& 0
.016
1.
50 2
0.0
34
.24 f 0.
024
1.2
1.3
1.4
1.2
1.3
1.3
1.1
YO
14.2
& 1
.07
12.8
? 1
.07
13.0
& 1
.13
12.4
2 1.
13
13.2
f 1
.17
11.4
-t
0.97
16
.0 -
C 1
.62
11.2
2 0
.95
13.4
-C
1.1
4 14
.0 f 1
.14
11.3
2 1
.01
Den
sity
1
I 2.0
I
1.4
I 1.69
-t
0.02
5 0.
20 f 0.
017
11.8
k 1
.08
27
Den
sity
4
1.9
1.6
1.72
2 0
.021
0.1
7 k 0
.014
9.9
k0
.89
28
D
ensi
ty
16
1.53
2 0
.017
0.1
2 &
0.0
12
7.8
zk 0
.79
Den
sity
32
I 1.8
I i:; I 1.33 &
0.0
40 1
0.25 f 0.
028
118.
8 f 2.
27
I 1'7" I
Den
sity
64
plae
ed
2.2
2.0
2.1
2.1
1.9
2.1
2.0
!pla
ce(
2.2
2.4
2.0
1.9
-
-
1.8
1.1
1.49
10
.04
0 0
.24
fO.0
25
16.1
-I
2.12
13
1.
9
1.5
1.5
1.5
1.5
1.3
1.6
1.3
39
32
30
27
29
28
22

STUDIES I N POPULATION PHYSIOLOGY. VIII 65
I f the mean weights of densities 1 through 16 and those of densities 32 and higher are averaged for all series it is ap- parent that crowding results in lighter pupae and imagoes in the 10-gm. and 4-gm.-not replaced experiments. This is especially true for the latter where the density relations are very stringent. Here, the ‘low density’ group weighed signifi- cantly more than the ‘high density’ group for both males and females as well as pupae and imagoes. For the 10-gm. series the same trend is present but the differences are not as great and have less statistical reliability. There is no evidence that weight has been altered by crowding in the 4-gm.-replaced cultures.
On the basis of these data the following conclusions can be drawn: 1) pupal and imago weight is reduced if the larvae are reared under conditions of extreme crowding and if the flour is not renewed during the larval period; 2) this reduction affects males and females alike; 3) weight is not as responsive to density changes as larval mortality and development, and 4) the conditioning of the flour is of primary importance in bringing about weight reduction in crowded cultures. There is no clear-cut evidence to the effect that variability in weight is proportional to density.
DURATION AND MORTALITY O F THE P U P A L PERIOD
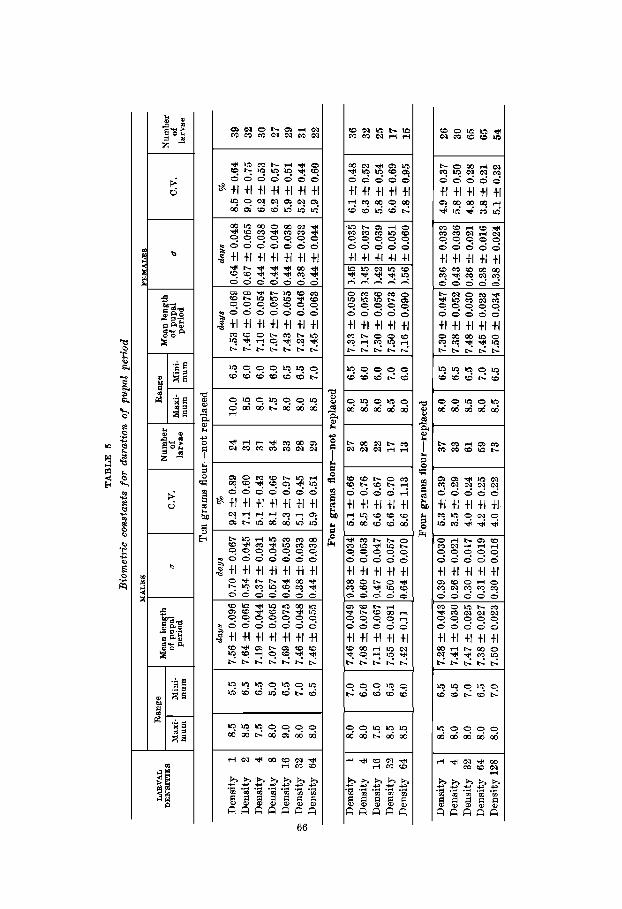
The duration and mortality of the pupal period were studied as final aspects of the post-embryonic development of Tri- bolium. It will be recalled that the pupal period was timed to the nearest + day and that all pupae, irrespective of their larval history, were isolated individually into vials. The biometric constants describing these results are summarized in table 5.
There is some evidence that pupal mortality is correlated with high larval density. From table 1 and figure 3 it is apparent for the 4-,gm.-not replaced series that there has been a high death rate among the pupae of the 16,32 and 64 density groups. The mortality of both sexes for these three cultures is 26.6%, 46.9% and 56.2%, respectively. The control (one
TA
BL
E 5
Bio
met
ric
coas
tant
s fo
r du
ratio
n o
f nu
oal
veri
od
Ran
ge
mum
m
um
IAF
.VA
D
DE
NSI
TIE
S M
axi.
Min
i. M
ean
leng
th
Num
ber
U
C.V
. of
O
f pu
pal
larv
ae
peri
od
Den
sity
1
Den
sity
2
Den
sity
4
Den
sity
8
Den
sity
16
o,
Den
sity
32
D
ensi
ty
64
Q,
davs
7.5
6 f 0.
096
7.64
f 0.
065
7.19
& 0
.044
7.
07 f 0.
065
7.69 f 0.
073
7.46
-C 0
.048
7.
46 k
0.0
55
8.5
8.5
7.5
8.0
9.0
8.0
8.0
dW
8
0.70
2 0
.067
0.
54 &
0.0
45
0.37
f 0.
031
0.57
k 0
.045
0.
64 f 0.
053
0.38
f 0.
033
0.44
f 0.
038
5.5
6.5
6.5
5.0
6.5
7.0
6.5
days
7.5
3 ?
0.0
69
7.46
& 0
.079
7.
10 2
0.0
54
7.07
f 0.
057
7.43
f 0.
055
7.27
f 0.
046
7.45
f 0.
063
larv
ae
days
0.
64 -
t- 0.
048
0.67
f 0.
055
0.44
2 0
.038
0.
44 f 0.
040
0.44
f 0.
038
0.38
f 0.0
32
0.44
2 0.
044
Ten
gra
ms
flour
-not
re
plac
ed
Den
sity
1
Den
sity
4
Den
sity
16
D
ensi
ty
32
Den
sity
64
8.0
7.0
7.46
2 0.
049
0.38 f 0.
034
5.1 2
0.6
6 27
8.
0 6.
5 7.
33 k
0.0
50
D.45
t 0
.035
6.
1 f 0.
48
36
8.0
6.0
7.08
f 0.0
76
0.60 f 0.
053
8.5 2
0.7
6 28
8.5
6.
0 7.
17 k
0.0
53
3.45 t 0
.037
6.
3 2
0.5
2 32
7.5
6.
0 7.
11 f 0.
067
0.47 k 0
.047
6.
6 -C
0.67
22
8.0
6.0
7.3
0 k 0
.056
1.4
2 t 0
.039
5.
8 f 0.
54
25
8.5
6.5
7.55 2 0
.081
0.50
2 0
.057
6.
6 &
0.70
17
8.5
7.0
7.5
0 2
0.0
73
1.45
t0.0
51
6.0
f 0.6
9 17
8.
5 6.
0 7.
42 k
0.1
1 0.
64 k
0.0
70
8.6 &
1.1
3 13
8.0
6.
0 7.1
6 2 0
.090
1.5
6 k 0
.060
7.8
f 0.
95
15
%
9.2 f 0.
89
7.1 f 0.
60
5.1 2 0.
43
8.1 f 0.
66
8.3 -t 0
.97
5.1 5
0.4
5 5.
9 -t 0
.51
7.28
f 0.
043
7.41
f 0.
030
7.47
f 0.
025
7.38
f 0.
027
7.50
-t
0.02
3
24
31
3
1
34
33
28
29
0.39
2 0
.030
5.3
5 0
.39
37
0.26
f 0.
021
3.5 2 0
.29
33
0.30
k 0
.017
4.0
2
0.2
4 6
1
0.31
2 0
.019
4.2
& 0
.25
59
0.30
& 0
.016
4.0
2 0
.22
73
10.0
8.5
8.0
7.
5 8.0
8.0
8.5
8.0
8.0
8.5
8.0
8.5
6.5
6.0
6.0
6.0
6.5
6.5
7.0
6.5
7.30
20
.04
7 0
.36
0.03
3 4.
9 f 0.
37
26
6.5
7.38
2 0
.052
0.4
3 50
.036
5.
8 f 0.
50
30
6.5
7.48 f 0.
030
0.36
20.
021
4.8
0.28
65
7.0
7.4
5 &
0.0
23
0.28
-C 0
.016
3.8
-t
0.21
65
6.5
7.50
-C 0
.034
0.3
8 -C
0.0
24
5.1
2 0
.32
54
%
8.5 k
0.64
9.
0 f 0.
75
6.2 f 0.
53
6.2 f 0.
57
5.9 f 0.
51
5.2 f 0.
44
5.9 f 0.6
0
39
32
30
27
29
31
22
Den
sity
1
Den
sity
4
Den
sity
32
D
ensi
ty
64
Den
sity
128
8.5
6.5
8.0
6.5
8.0
7.0
8.0
6.5
8.0
7.0
STUDIES I N POPULATION PHYSIOLOGY. VIII 67
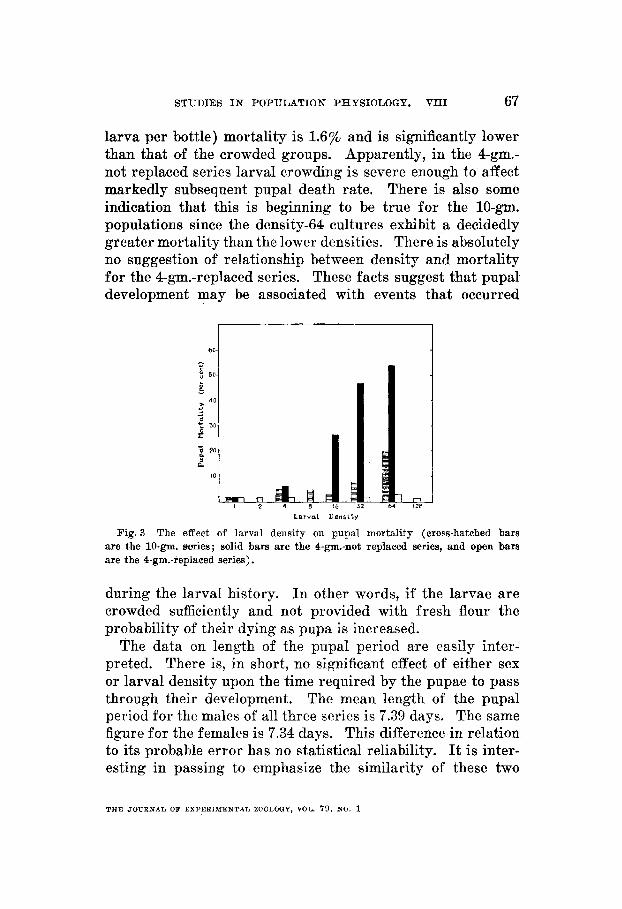
larva per bottle) mortality is 1.6% and is significantly lower than that of the crowded groups. Apparently, in the 4-gm.- not replaced series larval crowding is severe enough to affect markedly subsequent pupal death rate. There is also some indication that this is beginning to be true for the 10-gm. populations since the density-64 cultures exhibit a decidedly greater mortality than the lower densities. There is absolutely no suggestion of relationship between density and mortality for the 4-gm.-replaced series. These facts suggest that pupal development may be associated with events that occurred
L a r v a l D e n ~ i t y
Fig. 3 The effect of larval density 011 pupal mortality (cross-hatched bars are the 10-gm. series; solid bars are the 4-ga-not replaeed series, and open bars are the 4-gm.-replaced series).
during the larval history. In other words, if the larvae are crowded sufficiently and not provided with fresh flour the probability of their dying as pupa is increased.
The data on length of the pupal period are easily inter- preted. There is, in short, no significant effect of either sex or larval density upon the time required by the pupae to pass through their development. The mean length of the pupal period for the males of all three series is 7.39 days. The same figure for the females is 7.34 days. This difference in relation to its probable error has no statistical reliability. It is inter- esting in passing to emphasize the similarity of these two
T H E JOURNAL OF EXPERIMENTAL ZCOLOGY, VOL. 79, NO. 1

68 THOMAS PARK
means. Their difference, based on 580 males and 575 females, is of an order of only 0.05 of a day while the age of each individual pupa mas determined to 0.5 day. Clearly, this is a case where numbers and statistical treatment have refined the experimental technique.
To appraise the effect of larval density the data were grouped into ‘slightly crowded’ and ‘crowded’ cultures ex- actly as was done in dealing with pupal weight. These mean figures gave no indication that density played any role in altering the rate of pupal development. The means were remarkably similar. F o r example, for the 4-gm.-replaced series the ‘slightly crowded’ males took 7.48 days to pass through pupal growth while the ‘crowded’ males took 7.45 days. The females of both low and high density groups each required 7.48 days.
On the basis of these results, the conclusion is reached that it is immaterial in respect of rate of pupal development 1) whether the pupae have been reared as larvae in high or low densities and in fresh or conditioned flour or 2) whether they are males or females. I n other words, the time required for pupal growth is biologically closely fixed and is not subject to great variation.
DISCUSSION AND CONCLUSIONS
As a result of the analysis presented in this paper a number of factual conclusions have been reached regarding the effect of larval density on the post-embryonic development of Tribolium confusum. Many of these observations are of onto- genetic interest but not all are directly related to population dynamics. From the latter view the major point growing out of this investigation is that larval crowding can reduce drastically the efficiency of metamorphosis if the culture medium is not renewed. Interpreted in terms of population growth this means that crowding per se is not greatly detri- mental to larval and pupal development but is of utmost importance as the factor that controls the rate and degree of environmental conditioning. I n other words, the essential

STUDIES I N POPULATION PHYSIOLOGY. VIII 69
density effect is not the behavioristic interaction and com- petition between organisms but represents rather the cumula- tive influence these organisms exert on their environment. This does not mean that competition between larval Tribolium is never of population significance. It is possible that under different experimental situations a direct density effect could be demonstrated. I n fact, some evidence for this exists in the present data although here the effect, even if real, is of slight magnitude.
From the descriptive aspect the results suggest that Tribolium cultures will have their total population growth reduced, other things being equal, if the larvae are crowded and not supplied frequently with fresh flour. This reduction will be related to the density and to the conditioning of the medium. As the population grows and later generation larvae develop the effect should be more stringent provided the flour is not renewed. The inhibiting influence of conditioning on fecundity (Park and Woollcott, '37) would also become an increasingly important factor with age. This, combined with the metamorphosis mortality, retarded development and other possible factors not yet understood, should eventually bring about the extinction of the population.
SUMMARY
An analysis of the effect of differential larval density on the post-embryonic development of Tribolium confusum has shown that, 1) Larval and pupal mortality and duration of the larval period are increased by crowding if the culture medium is not renewed every 48 hours. Also, under these conditions pupae and imagoes reared from crowded larvae weigh less than do those from not-crowded cultures. 2) I f the culture medium is renewed the larvae develop as successfully and the pupal death rate is as low in crowded as in sparse popula- tions. Here, there is no demonstrable effect of density on pupal o r imago weight. 3) The duration of the pupal period is not significantly altered either by sex, density or changes in the medium. These facts are discussed in the light of

70 THOMAS PARK
population theory and some attempt is made to interpret the nature of the density effect.
I am indebted to Mr. B. A. Schneider for aid with the experimental routine and to Profs. Raymond Pearl, W. C. Allee and Sewall Wright for reading the manuscript.
LITERATURE CITED
ALLEE, W. C. 1931 Animal aggregations: a study in general sociology. Chicago:
Recent studies in mass physiology. Biol. Rev., vol. 9, pp. 1-48. BBINDLEY, T. A. 1930 The growth and development of Ephestia kuehniella
Zeller and Tribolium confusum Duval under controlled conditions of temperature and relative humidity. Annals Ent. SOC. Am., vol. 23,
UH~PMAN, R. N. 1928 The quantitative analysis of environmental factors.
MAOLAGAN, D. S., m EDWARD DTT” 1935 The experimental analysis of the growth of am inseet population- Proc. Royal SOL Edinburgh, vol. 55, pp. 126-139.
Observations on the general biology of the flour beetle, Tribolium confusum. Quart. Rev. Biol., vol. 9, pp. 36-54. 1935 Studies in population physiology. IV. Some physiological
eff eets of conditioned flour upon Tribolium confusum and its populations.
1936 Studies in population physiology. V. The oxygen eonsump- tion of the flour beetle, Tribolium confusw Duval. J. Cell. and Comp. Physiol., vol. 7, pp. 313-323. 1937 Experimental studies of insect populations. Am. Naturalist,
PARK, THOMAS, AND NANCY W o o ~ c o r r 1937 Studies in population physiology. VII. The relation of environmental conditioning to the decline of Tribolium confusum populations. Physiol. Zool., vol. 10, pp. 197-211.
PEBRL, RAYMOND 1927 The growth of populations. Quart. Rev. Biol., vol. 2, pp.
The influence of density of population upon egg production in Drosophila melanogaster. J. Exp. Zool., vol. 63, pp. 57-84.
@ A m , GEORGE 1936 Experimental studies in insect parasitism. Iv. The effect of superparasitism on populations of Trichogramma evanescens. J. Exp. Biol., vol. 13, pp. 363-376.
University of Chicago Press. 431 pp. 1934
pp. 741-757.
Ecology, VOI. 9, pp. 111-122.
PARK, THOMAS 1934
PhySiOl. ZOO^., V O ~ . 8, pp. 91-115.
V O ~ . 71, pp. 21-33.
532-548. 1932





























