Embed Size (px)
Citation preview

Structure and Expression of Fibrillin-2, A Novel Microfibrillar Component Preferentially Located in Elastic Matrices Hui Zhang, Stephen D. Apfelroth,* Wei Hu, Elaine C. Davis,* Chiara Sanguineti,§ Jeffrey Bonadio,§ Robert P. Mecham,* a n d F rancesco R a m i r e z
Brookdale Center for Molecular Biology, Mt. Sinai School of Medicine, New York 10029-6574; * Department of Pathology, New York University School of Medicine, New York 10016; ~ Departments of Cell Biology and Medicine, Washington University Medical Center, St. Louis, Missouri 63110; § Department of Pathology, University of Michigan Medical Center, Ann Arbor, Michigan 48109
A b s t r a c t . During the previous cloning of the fibrillin gene (FBN1), we isolated a partial eDNA coding for a fibrillin-like peptide and mapped the corresponding gene (FBN2) to human chromosome 5. (Lee, B., M. Godfrey, E. Vitale, H. Hori, M. G. Mattei, M. Sarfarazi, P. Tsipouras, E Ramirez, and D. W. Hol- lister. 1991. Nature [Lond.]. 352:330-334). The study left, however, unresolved whether or not the FBN2 gene product is an extracellular component structur- ally related to fibrillin. Work presented in this report clarifies this important point. Determination of the en- tire primary structure of the FBN2 gene product demonstrated that this polypeptide is highly homolo-
gous to fibrillin. Immunoelectron microscopy localized both fibrillin proteins to elastin-associated extracellular mierofibrils. Finally, immunohistochemistry revealed that the fibrillins co-distribute in elastic and non- elastic connective tissues of the developing embryo, with preferential accumulation of the FBN2 gene prod- uct in elastic fiber-rich matrices. These results support the original hypothesis that the fibriUins may have dis- tinct but related functions in the formation and main- tenance of extracellular microfibrils. Accordingly, we propose to classify the FBN1 and FBN2 gene products as a new family of extracellular proteins and to name its members fibrillin-1 and fibrillin-2, respectively.
I T is now well established that fibriUin is the defective gene product responsible for the ophthalmic, cardio- vascular, and skeletal manifestations of the Marfan
syndrome (MFS) ~ (Dietz et al., 1991, 1992a,b, 1993a,b; Kainulainen et al., 1992; Hewett et al., 1993). FibriUin was originally isolated from the medium of cultured fibroblasts using an antibody raised against pepsin-treated and salt- precipitated proteins from human amnion (Sakai et al., 1986). Biochemical analyses estimated fibrillin to be a 350- kD acidic glycoprotein with a large number of cysteine residues (Sakai et al., 1991). Additionally, fibrillin was shown to be an integral component of the non-collagenous microfibrils with an average diameter of 10 nm (Sakai et al., 1986, 1991). These extracellular aggregates are present in both elastic and non-elastic tissues (Low, 1962; Cleary and Gibson, 1983; Mecham and Heuser, 1991). Microfibrils of elastic tissues have been shown to assemble into a scaffold upon which elastin is subsequently deposited, whereas in
Address all correspondence to E Ramirez, Brookdale Center for Molecular Biology, P.O.B. 1126, 1 Gustave L. Levy Place, New York, NY 10029.
1. Abbreviations used in this paper: AMP, associated microfibril protein; CB, calcium-binding; CCA, congenital contractural arachnodactyly; MAGP, microfibril-associated glycoprotein; MSF, Marfan syndrome.
non-elastic tissues they are believed to serve an anchoring function (for reviews see Mecham and Heuser, 1991; Rami- rez et al., 1993). Relevant to MFS pathology, microfibrils are a major structural component of the suspensory liga- ments of the eye, the aortic wall, and the perichondrium and periosteum (Mecham and Heuser, 1991; Ramirez et al., 1993).
Cloning experiments have established that the primary structure of fibrillin consists mainly of repeats similar to the calcium-binding (CB) sub-class of the EGF motif (Maslen et al., 1991; Lee et al., 1991; Corson et al., 1993; Pereira et al., 1993). These experiments identified a partial eDNA potentially coding for a fibriUin-like peptide, thus suggesting that fibrillin may represent a novel gene family (Lee et al., 1991). The gene coding for this putative fibrillin-like tran- script was mapped to human chromosome 5. Based on their respective chromosomal locations, the two fibrillin tran- scripts were provisionally termed Fib 15 and Fib 5 and the corresponding genes FBN1 and FBN2 (Lee et al., 1991). In- terestingly, the latter was genetically linked to the MFS-like condition, congenital contractural arachnodactyly (CCA) (Lee et al., 1991; Tsipouras et al., 1992). Patients affected by this rare connective tissue disorder share some of the skeletal features of MFS, but display joint contractures in
© The Rockefeller University Press, 0021-9525/94/03/855/9 $2.00 The Journal of Cell Biology, Volume 124, Number 5, March 1994 855-863 855

place of loose jointedness (Beals and Hecht, 1971). Aside from lacking the cardiovascular and ocular manifestations of MFS, these patients exhibit characteristically misshaped ex- ternal ears (Be~s and Hecht, 1971).
The association between phenotypically overlapping con- ditiom and two potentially related macromolecules prompted us to suggest that Fib 5 and Fib 15 may have distinct but re- lated extracellular functions (Lee et al., 1991). This postu- late is supported by results presented in this study which demonstrates that Fib 5 is a microfibriUar constituent, whose structure and expression are related to those of Fib 15. Based on this evidence, we propose to classify the fibrillins as a new family of extracellular proteins and to name its members fibrillin-1 (Fib 15) and fibrillin-2 (Fib 5).
Materials and Methods
Cloning Experiments and Northern Analysis A eDNA library in the phage vector larnbda-Zapll (Stratagene, La Jolla, CA) was generated using as a template poly-A + RNA purified from cul- tured MG-63 cells. This human osteosareoma cell line was previously shown to produce significant amounts of fibrillin (Sakai et al., 1986). Gen- eration, screehing, identification and purification of recombinant phages were accompfished by standard procedures (Sambrook et al., 1989). The 5' furthest eDNA was generated using the 5' RACE system for rapid am- plificatiun of eDNA ends, according to the manufacturer's recommenda- tions (Life Technologies Inc., Gaithersbu~, MD). Positive eDNA inserts were sequenced using the dideoxynucleotide chain termination proo~ure on double-stranded DNA (Zagursky et al., 1986). Sequences were obtained by multiple sequencing of both strands of eDNA inserts; they were analyzed using the computer program MacVector (International BioTechnologies, Inc., New Haven, CT). For Northern analysis, '~10/~g of total RNA was fractionated on a 0.8% agarose gel in formamide/formaldehyde, transferred onto a nylon membrane (Amersham Corp., Arlington Heights, IL), and hy- bridized at high stringency to fibrillin cDNA probes uniformly labeled by random priming (Sambrook et al., 1989). To insure specificity, we chose as probes twv eDNA inserts of comparable size, '~1 kb; they are: clone F-35, which covers the 3' non-coding region of the FBNI mRNA (Pcreim et al., 1993), and the internal EcoRI insert of clone A06-13 (see Fig. 1). The latter sequence exhibits 40% homology to the equivalent region of the FBN1 gene, whereas there is only 6% homology in the untranslated seg- ment of the two transcripts.
Antibody Generation Fibrillin antibodies were raised against recombinant peptides generated in the Escherichia coli-based expression system pET-3xa (Novagen, Inc., Madison, WI). To this end, the sequences coding for region C of the two fibrillins (see Fig. 3) were adapted to E. coli codon usage by ligating several overlapping synthetic oligonucleotides prior to subcloning into the expres- sion vector (Sambrook et al., 1989). Fusion products were expressed and isolated according to the manufacturer's protocol. Approximately 1 nag of gel-purified fusion protein was injected subcutaneously into rabbits. The in- jection was repeated at 2-wk intervals for a total of 12 wk and serum was collected by ear vein puncture. Pre-immune sera were also collected from the same rabbits to test for specificity.
Protein Analysis For metabolic labeling experiments, MG-63 cells were cultured in cysteine- free and serum-free DME medium containing 60 pCi/rnl ]3SSlcysteine (Amersbam Corp.) (Sakai et al., 1986). After an overnight incubation, the culture medium was harvested and placed immediately on ice with the addi- tion of 5 mM EDTA, 50 pM N-ethylmaleimide, and 50 ~tM phenyimethyl- sulfonyl fluoride (Sakai et al., 1986). Labeled cell cniture medium was incubated with each unpurified fibrillin antibody coupled to protein A-Seph- arose for 3 h at 4°C. This was followed by extensive washing with PBS, and subsequently with RIPA buffer (150 mM NaCI, 1% NP-40, 0.5% DOC, 0.1% SDS, and 50 mM Tris, pH 8.0). Bound material was eluted with gel electrophoresis loading buffer (1% SDS, Tris 10 raM, pH 8, 5% glycerol,
and 2.5 % B-mereaptcethanol) and analyzed by fraetionation on a 4 % poly- acrylamide gel. For Western blot analysis, unlabeled immunoprecipitates were electrophoresed onto nitrocellulnse paper after gel frnctionation (Harlow and Lane, 1988). The filter was blocked with 39[ BSA in 10 mM Tris, pH 8.0, 150 mM NaCI and 0.05% Tween-20, before incubation with each of the fibrillin antisera (Harlow and Lane, 1988). After bh incubation at room temperature the filter was washed, incubated for 30 rain with alka- line phospbatase-conjugated anti-rabbit IgG (Promega Corp., Madison, WI), washed again, and incubated with the substrate to visualize the antibody-bound electrophoretic band (Harlow and Lane, 1988). Pre- immune serum was included as a negative control sample.
Immunohistochemistry For indirect immunofluorescence studies, 20-wk gestation human tissues were immediately frozen, sectioned, fixed in 4% paraformaldehyde and processed as previously described (Sakai et al., 1986). Tissue specimens analyzed using the peroxidase anti-peroxidase method were initially fixed in paraformaldehyde, paraffin-embedded, and processed further using a commercial kit with chromogen AEC as a substrate (Zymed Laboratories, Inc., San Francisco, CA) following the manufacturer's recommendations. Tissue localization of elastic fibers was accomplished using standard Van Gieson elastic staining (Luna, 1968). All studies included preimmune se- rum as a negative control.
Immunoelectron Microscopy Fetal bovine tissues were fixed in 4 % paraformaldehyde in 0.1 M Sorensen's buffer (pH 7.4) for 4-6 h at 4°C and processed for reductive denaturation according to Gibson et al. (1989). 1issue specimens were dehydrated in a graded series of methanol to 90 % at progressively lower temperatures from 4°C to -20°C. Samples were then infiltrated and embedded with Lowicryl K4M (SPI Supplies, West Chester, PA), which was subsequently polymer- ized by ultraviolet illumination for 24 h at -35°C and an additional 48 h at -10°C. Thin sections of tissue were cut with a diamond knife on a Reichcrt ultracut microtome and placed on formvar-coated nickel grids. Af- ter a 15-rain blocking step on drops of 1% BSA in 50 mM Tris containing 100 mM NaC1 (pH 7.4), the grids were incubated with primary antibody diluted in blocking solution overnight at 4°C. Following extensive washing and a second blocking step, the grids were transferred to 15-pl drops of sec- ondary antibody for 1 h at room temperature. The secondary antibody, goat F(AlY) 2 anti-rabbit IgG conjugated to 10 nm colloidal gold (BioCell Re- search Lab., Cardiff, UK), was used at a 1:30 dilution in blocking solution. Immunolabeled sections were counterstained with methanolic urmwl ace- tate for 2 min followed by lead cilrate for 30 s. Controls inchded the use of non-immunized rabbit serum and omission of primary antibody.
Results
Hbriilins Are Structurally Related Proteins We previously reported the isolation of MF-23, a partial cDNA that codes for a pcptide consisting mostly of EGF-CB repeats (Lee et al., 1991). The conceptual product of this 2.8- kb clone is remarkably similar to the fibriMn protein origi- nally identified by Sakai et all. (1986), and subsequently characterized in full by molecular cloning experiments (Maslen et al., 1991; Lee et al., 1991; Corson et al., 1993; Pereira et al., 1993). To complete the characterization of Fib 5, appropriate sub-fragments of MF-23 were used to screen the MG-63 cDNA library. This and subsequent screenings led to the isolation of several overlapping cDNAs covering the entire coding sequence of Fib 5 and almost 1.5 kb of 3' untranslated sequence (Fig. 1). The experiments also re- vealed that the original MF-23 eDNA represents the reverse transcribed product of a partially spliced transcript with a long intronic sequence at its 3' end (Fig. 1). Overall, the Fib 5 and Fib 15 sequences are highly homologous (Fig. 1); in addition, Northern analysis showed that the two proteins are
The Journal of Ceil Biology, Volume 124, 1994 856

i A i i B : C : : D II E I
- ] ilillllilli
iliillli EB B II / . . . . . . . . . . . . . MF23
EB
c I A06-4 3 'A
X R A C E 1 I RP08 3 'B
B B i 15061 I I !
lkb
f Fibrillin-2
Figure L Restriction map of fibrillin-2 cDNAs with a sche- matic illustration of the fibril- lind and fibrillin-2 proteins. Letters indicate the five struc- turally distinct regions of the latter. Symbols designate the following structural elements: EGF-like repeat (cross-hatched rectangle); EGF-CB repeat (white rectangle); TGF-bp re- peat (white oval); Fib-motifs (other symbols). Note that re- gion C is depicted differently in the two fibrillins in order to emphasize its compositional diversity. A similar represen- tation of the translational prod- uct of the original clone MF- 23 is also shown for compari- son; the dotted line indicates the unspliced intron sequence
in MF-23. Letter on the cDNAs indicate the following restriction enzymes: Barn HI (B), Cla I (C), Eco RI (E) and Xho I (X). The open triangle and the pin-heads symbolize putative cell attaehmeat and glycosylation sites, respectively (see also Fig. 3).
encoded by transcripts of similar size (Fig. 2). Based on this evidence, Fib 15 and Fib 5 were re-named fibrillin-1 and fibrillin-2, respectively.
Fibrillin-1 is a multidomain protein consisting of five structurally distinct regions termed A, B, C, D, and E (Fig. 1) (Corson et al., 1993; Pereira et al., 1993). Although the precise start site of transcription remains to be determined, a consensus has been reached on the putative start site of translation (Corson et al., 1993). The predicted amino acid sequence of fibrillin-2 shows that this 2,889 amino acid poly- peptide can be divided into five regions analogous to do- mains A-E of fibrillin-1 (Fig. 1).
Pairwise alignment of the fibrillins further emphasizes the close kinship between these two gene products (Fig. 3). In region D, for example, fibrillin-1 and fibrillin-2 contain 41 EGF-CB repeats similarly interspersed among eight non- EGF-CB motifs (Fig. 1). Both fibrillins exhibit the same number of amino acids between the cysteinyl residues in nearly all of the corresponding 49 pairs of cysteine-rich repeats (Fig. 3). In addition, all but one of the putative N-gly-
Figure 2. Northern blot hy- bridization of MG-63 RNA to fibrillin-1 and fibrillin-2 probes. RNA size markers (in kb) are indicated on the left of the autoradiogram, while the estimated range of the two transcripts is shown on the right.
cosylation sites and the potential cell attachment sequence previously noted in fibrillin-1 (Maslen et al., 1991) are con- served in fibrillin-2 (Fig. 3). FibriUin-2 contains a second RGD sequence within another non-EGF-CB repeat of region D (Fig. 3). The overall level of amino acid homology of re- gion D is •81%.
Pairwise comparison of regions B, A, E, and C revealed several more similarities between fibrillin-1 and fibrillin-2, and at least one important difference. A high level of se- quence homology (87%) was noted in region B of the two molecules. Region A, on the other hand, is substantially less homologous (19%) and it contains a novel proline-rich se- quence, immediately after the postulated signal peptide cleavage site (Fig. 3). The significance of this finding, if any, is unclear at present. Despite the compositional difference, region A of fibrillin-2 is as basic as region A of fibriUin-1 (est. pIs, 12 versus 11.1). Region E of fibrillin-2 displays also relatively low homology (50 %) to the corresponding domain of fibrillin-1. In addition to two polylysine stretches at the carboxy terminus of region E, two cysteine residues found in this domain of fibriUin-1 are maintained in the same posi- tion in fibrillin-2 and in the context of a nearly identical se- quence (Fig. 3). Finally, region C shows the most striking and important sequence divergence in that this short se- quence is glycine rich in fibrillin-2 but proline rich in fibrillin-1.
FibriUin-2 Is a MicrofibriUar Component
The structural diversity of region C provided an experimen- tal means to characterize further the fibriUin proteins. Ac- cordingly, two polyclonal antibodies were raised against recombinant peptides of region C from fibrillin-1 and fibrillin-2 and used to immunoprecipitate radioactively la- beled proteins from the medium of cultured MG-63.
Consistent with the cloning and Northern blot data, elec- trophoretic analysis of the immunoisolates revealed that the antibodies interact with proteins of similar size, about 350
Zhang et al. Microfibrils Contain a Second FibriUin Protein 857

V i M.R}~GRL.LEIA.. ,LGFTVLLA . . . . . . . . . . . . . . . . . . . . . . . . . $¥TSHGADANLEAGNVKE..,TRAS[LkKRRGGCGHOALKGpNVCGSRyNAyC 67
[ I [ t l [ : : ] l , l l : l V [ F I , : : , : [ , : : l , [ 1 4 , : [ ] [ , : [ , [ : [ l [ l [ [ [ [ : : . [ I I MGRRRRLCLQLYFLWLGCV%'LW/tQGTAGQPQPPppKPpRPQPPPQOV~SRTA,GSDGGFLAPEyREEGAAVASRV~..GOQDVLBGpNvCGSRFHSYC 97
68 C~I (TLBGGN~IVP IC~C~DGFC~RPNMCTC~SG~IAP~G~R~ IQHC~RC~GG$CSDDH~LCQKGYIGT~CCQP~£~L~R~ 167 I IH IS I I I I I I I I I tH I : I qH l l l I I t lHH .HH. . . I I q : I I I : h : IHH I . I . I I I I I I I I I I I 1 . 1111111 .11 I I I I 1 : 1 . . 11
9S CP~4KTLPGGNCCIVP ICRt4SCt~GFCSRPNMCTC£SGQI~TC~K$~SVR~ .~NGG~.~ADDHC~CQKGYIGT¥CGQP~E~CIA `~ 1 9 6
168 C T Y G F - T G P ~ c ~ R D ¥ R T G ~ F 1 . V I ~ N ~ c ~ L ~ G ~ V C ~ K ~ L C ~ V i % . G R A k ~ G ~ P C E / ~ P A Q ~ H P C R R G ~ I ~ N I ~ C ~ D ` ~ E C ~ I ~ G 267
I . I I I I I I I l l l l l l l l l l l l : . l l l : l l l l . l l l l l l I I I I ) . I H I I I I I I I I I I I I Z l I I I I t l I I I I I I I I I I I I I ~ I H I I : I H I I I I I I I I t 9 7 ~ / X G F T G ~ C E R D Y a T G P C F ~ x / d h ~ x . ( ~ $ C G Q L ? G I V C T g T L C C % T T G R A x 4 G H ~ P A Q ~ Q ~ F I P N I ~ C ~ £ C ~ I P G I ~ h ~ C 296
268 $FgCKCP~GHKLNEV$QKCEDIDiX~$T~PG~CEGGECTNTVSSYFC~CPpG~YTSP~GTp~I~VRp~YCYTALTNG~CSNQLPQ~IT~AG~S 36~
I I H : H I I I I . I , , H t l I I I I I I . I I I I I I , H J A I I : I I I I II I : I I , I I . H H ~.1 I : . : l . l l l l . . : l t . . : L t l l l l l : : l l l l : 297 ~FECR~AGHKQ~E1q:QK~E~FX2~?G~CE~Ix;ECSN`I'vGS¥FCvCP~GYxtT~TDG~RC~D~R~F~GLVDGRCAQELt:~3~C~ 396
397 I ~ T ~ . P E A ~ P V p ~ $ E E Y R R L C t ` t ~ P ~ I P ~ S ~ S ~ S ~ * ~ Y ~ P ~ F ~ N ~ F ~ ~ ~ ~ 9 ~ a 1 t J V ~ . ~ . n ~ 1 ~ 7 ~ L 48~
446 ~V~YC~LVRY~RC~PTPGsY~%Cit~KGFQL~Ll~GEC~VDECE~t~EC~YTC~C~Y~TLTRT~I~ 543
.: I 1 1.1. ~I hHH~.:HHm. I:. I .MHHH..III..~:I:I,.III 1.1:11:1.1 I: .I HHI:III :hl 488 T~L~DICK~HANLCLNGRCIPTV~YRCEcNF£,YKQ{x~R`GDCtD'VDECT~t~FCTNGDCV~TPG~YYcKc~GF~RTPTK~cIDIDEcI~ 58~
544 G~aCINT~X;~F~Cvc~FHVTl~DGgNCED~'$D~I~NMCLNC/4CNN~D(;~?~CI~KPG~S~C~N~E~Y~ECF~L 643 [ [ [ : [ * [ H ~ : H I I I I I . ; I H H I I I I I . . I I I I I [ I I I I H H H H t l I [ [ [ . : [ [ [ ~ . I : : H : ~ [ H H H : ~ : I . : I I : I I : I ~[~
588 GRCVN~DG~FQC~CNAGF~LTTDGKNC.VDHDECTTTNM£LNGMCINEDG~f `KC I~PGFVLAPN~RYCT~H~ IN~PPGL 68~
6t14 AVGLDGRVCVDT~TCYGGYK~RG~6~IKPLFGA~TKsECCCA~T~YAFG£~CQPCPAQN~AEY~s~G~NE~DIC~C~ 743
HI:HImHHHIIIH.I:I I::I: IIIIIIIIIII,,:I:~,HHHI~.IIII:::IIHM:~ .~.HHIIIIIIII:HIIH~ 688 A~C,M~yGRVCVDTHMR$TCYGG~KKGVC¢K~FPCA~TK~ECCCAN~DYGmg~¢)PC~A~$A~G~I~INE~m~D~IC~N~ 7 8 ~
~,, GTYKCI~SGy~S~INE~LNSL~D~TPGS~PKGFIYKPDLK~EDID~ESS~I~SpGSFICECSS~STLDPTKT 843
I,I:I IIIIII.I..I:II:II:H::~.~HHt ~Hh~: ~H~.~::::,: .~IIII:IIII.H:H.~:~. HI IIH.:~.~..~ ~88 GsY~NcNs~YEP~AsGR~C~I~ECLV'~R~LC{X̀2GLERNT~sY~?CPPGY~VRTET~TC£~Ng~g$NPcV2¢CAC~NNLG~gNCgCsPGag~$~G~ 88~
844 I C I E ~ K G T C W Q W ~ D G R C E I R ~ C C ~ T L K ~ C C C S ~ L G ~ w G ~ C T ~ C Q ~ D P I C ~ x ~ Y ~ R ~ x ~ T ~ D I ~ E C E V F F ~ T ~ S F ~ C ~ P S ~ L 942
888 ICI~$LKGTCWLNIQ~$R~EVNINGATLKS~CCA~LC4kAwG~C£~CE~A~GLAR~KGv~E~V~£~EVFPGv.~PNGRC¢N~KG~VHCEC~EGL~L 98~
944 ~TGRICLD~R~TCF~RY~£CT~P1AG~HBa~VCAAWG~CE~PM~N~`~LC~RG~FAT}<~EITNGKPFFKD~NECI~LMIP~HG 2042
9~8 DGTGRVCLDIR~QCyLKWDEDKCIHPVPGKFRMDAC~AVC, AAWGT EC£ECPKPGTMEY~TLCP~FAN~VL~RPFYKDINEC~YyG ~086
043 KCRNTIGSFKCRCD$GFALDSEEKb~C~D~DECR~S~DL`.'~GRCQ~?PCA3F~CKCDEGYE$GFM~NC~DECQRDPLLCRGGVCHNT~SYRCECPP 1~42
087 K~RNT~G~FK~R~NSG~ALDM~E~D~CR~PDLCGS~CvNT~G~V£~C~EGY~$GFI~NCMDID~CERNP~I~GGTCVNTEGSFQCDCP~ 1186
I~3 G~QLSP~CIDINECELsAHLCPNGRCVNL~GK¥QCACNPGY~i~TP~LFCvDIDEC~IMNGGCETFCr~SEGSYE~SCQFGFALMPDQRSCTDID£C 1242 II:III...I:IHH.Ih:H.H:IH:H.H~.HIII:.IH~ I,IIIII HHH:I IIHHHIIII .I:HJH.HI.IIIH
18~ G~EL~P~REDCVDIN~L~NLCRNGKCVNM~GTYCC~NPGYQAT~QCCTDI~IMNCI~CDTQCTN~£G~Y~C~G¥~LMPDGR~CAD~ 1286
2a3 EDNPNICDGGQCTNIPGEYR~LC¥DGFMASEDMKTCv~VN~CDLNPNIcLSGTCENT~G~VICHCDMGY~G~GKTCCTDIN~C~IGAHNt~J,KHAVCTNT 1342 I:II:HIIHHHHHHH1111111 Hm:HIIIIII.II,: I ,,I,'III~ ,::III.IIMHH::HHHHI:.II I I.
2B7 ~NN~D~C~TNI~G~YRC~Y~S~K~ECDLNSN~CM~£~NTKC~F~CHCQ~YSV~KG~1~VDECE~DM~CLNI 1386
343 AGSFKCSCSPCWI~/)G~KCTDLD~C~HMCSQHADCKNTMGSYRCLC~FLr;YTG~GVTC~D~DEC~NLNLCGNG~LNAPGGYRCECDMGWPSA~K ~442
:HHHI..IIII:HH.III~IHH~ H :1:1 H HIH I I': ,~I''.I:II .~:III:IIIIII.~I:H~H:HM..I:: 387 PG~FKCSCREG~4~GNG~K~DL~HQC~NA(2CVNT~G~¥RCAC~G~T~X3~C~V~CA£NINLCEN6QCLNV~GAYRCECFJ4GFTPA$D~R 1486
~ 3 A~I~EC~L2N~CVFGTC~N~PGLFR~ECE~GY~LDR&~DVN~C~DPTT~I~GNL~VN~PG~I~DCPPDFELNPTRVCCVDTR~GNcYL~]RP~C~ 1542
. l : l l l l l l : . ~ ] l l n l l : l l l l : l : l I : i I I i I l . l l l l l l : : l l I n . . : . : I I I I H . , ~: *111;1111 I I I I 1 . 1 I I I I 1 . : I I I ~87 ~CQD~E~$FQNICa¢sGTCNuLP~FH~GY£LDR~GG~d~D~NC7eNGI~RY~NC~FQUMPT~C~DNRVG~YL~FG~RG 1585
b~ 3 [2NGDTACSREI GVGVSKA $CCC S LGKAWGTpC£MC pAVhLI~.EYK [ LCPC~EGF RpNp ~ ~Vl IE:)) DECQEi P(;iL~;KC ~ ~'~D (;S~ QCRCP] Gy %'L NED 1642
r r..r..rrrrH~:.HHHIIIIHH H:H.H .HHrr' .... rr:rrH~rrHrH: , : '','I " IIII H
1643 ~RVCDDVN£c~ETPG~CGPG1X2¥~I~PPDY~$D~YKNYYAD~q`CIx;ELLF~ZKI@t~ctX:SYNIGP`A~PIp9T £741
H : [ : l : : [ [ . ~ [ : [ ~ 1 1 1 ~ [ [ [ : 1 [ [ [ [ [ ~ 1 [ : [ 1 [ [ ~ [ ~ ¢ [ [ [ 1 ~ [ : 1 : ~ [ ~ . [ : . [ 1 : . [ ~ [ [ : l [ : l l l t . [ ~ ; [ : [ H [ I h H * [ : [ 1686 TRICEDIDECFAqI~+~2CC4~GTCybr]?~ICPPEYML~IVN(~SFCyRSY..~Q_T'I'CENgLP?~C~K~NVGKAGNKPCEPCpT~T 1783
i~'12 DEFATLcGs~PGF'VID~Y`rGLP'VDIDEcR~IPGVCENG~vCINt`tVE~FRcECFVx;t~PtNDKLLvcEDID~P~Q~NTAG~Y~YR I840
1784 ~ F K T I ~ . N I p G ~ F D I H ~ B ~ I D ~ I ~ I ~ I ~ I ~ F ~ P ~ F S Y N D L L L ~ E D I D ~ S ~ D N ~ I N S P G S Y ~ K 1882
1841 FTS~DB~IPNICS~ID~SFyC~H~FK~D~LOIN~EB~TIGS~RCNHGFILS~ID~ 1940 : * . . I . I [[~ql [ [ [ [ : ~ I I [ ~tl ~[: ~ : [ I . I I I . . : H H I : ~ : : [ I I I * : I H [ I ~ : H : [ [ : H [ .11 I . H ~ H : [ : ~ [ { . I I .
2883 L$PNGACVDP3q~L£IPNVC~HG5CVDIX~G~YQCIC~NGFKA$Q~Cr~`M~F.CKRI~]~¢FVG~YN~LT~D~F~ 19@2
1941 ~ I N ~ S ~ N ~ Y ~ I N E C L L E P ~ S Y ~ I C P ~ Y S ~ K C ~ I D ~ E P E I ~ S ~ S F ~ C ~ 2040
:~,tl.~:l,:ll1.~ IIIIH:.IH:,I:I.elI: I .I.IIIIHI:H~IIHHII.=..M I~:II I:~:II :M.II.I:I.III 1983 ~RNGI~CFN~G$FK~LCNEGYELTFDG~W24CIDTN~CV~LPC,S~PGWCC~NLEG~FRC~C~PGYKVKs~NC~D~NE~N~LFG$CTNT~GGFQ CLC 2082
2041 PF~GF~L$~C~RRCQ~LRMsY~YAy`FEGCKC$sPK£KM~i~QDDCCA~KG£GWGDPCgLC~TEPDEAFP`Q~C~YGsGI~GPD~sAV~2`~DgC~(~`PDVCK 2138 ~ II I ) . . l l l l ~ I I : i ; . , l { . l l l l J l . [ . i . . l l l . : . I J i i i l H I I l i . z . z ] 1 . : : 1 1 1 , I . : . : . ; . I : : / I I I : : J .
2083 ~ P G F V L ~ I ~ P ` R C F D T R Q ~ F C F I ~ E N ~ X C ~ V P ~ T K A K C C C S K ~ E C ~ P C E L C ~ E V ~ P ~ L ~ L ~ S P ~ C S 2182
Z[39 ~ tNT~Sy~e~yTb . .AGN£CVD?DtCSVG~PCGbF~Ca2~VIGGFECTCeEGFEPGpMVTgCEDI~gCAQNPLL~yGsy~pvGy 2236
: ,HH IH# : I I I I I : ~? ,~ .# H H I I I 1 : I i I I I H I I . I I [ 1 :11 h ( : l l ( l l l l l h I I I I I I 1111111 :1 : I ~ : I l t l l l l l I 1 : 11 2153 NGQ~INTDG~RCECPMGYNLDYTGVRCVDTDEc~IGNPCG5Lq~TCTNVIG$F£CNCNEG[EPGPMMNCED~NF~AQNPLLSALRc~`~TFGSYF~TCPICY 2282
2237 VLR£DRBMcKDEDECEDGKHDCTEKQt'~CKNLIGTYMCICGPGYQRRP~GECCV~ENECQTKPGICE~RCLNT~GSY~ECN~VTA$P~LDN~ 233~
.I~I'.:HH III.I~ I~..:.~ ~IIHII:III~ II ,,IIIII,'III~.IHHI~III:~. HI IIH:II .I..,.H~III: 2283 AL~DQ~5C~KDLDECAEGLHDCE$RGMFCKNL1GTFMC1CPPGMARRPDGEGCv~ENECBTF`PGIC~R~IIGSY~N~FQ$$S~T~NBQ 2382
2337 GYcrT~VL~IGssNRNPV~sEcccD~GF~PHcEIcPr~G~W~KKLCwGRGE~TNCAD~DEck~l~HDvCF~NGrcVNDRGsYHc~cKTGYTPD 2436
~ . [ i . i i [ [ . : I ~ : ; I I . I I I I H I [ I [ I H [ [ [ h : H : I I : . I L . . : I & : I J I [ . ~ : [ : I H I I H I : . ; : [ l l : [ : ~ . . l l : : ~ : l l . [ l h l 2383 GLCFAi~/LQTIC~MA$$SRNLVTKsECCCDGGRGWGH~CELCPLPGTAQYKKICPHGPGYTTDGRDIDECKVMPN~INTMG$FRCFC~Y~D 2482
2437 ITGTSCVDLNF.CNQ~2I~PCNFIci(NTF~YQCsCPKGYILQKDGRsCK~t.DF~CATKQ~dNCQFt.CVNTZGGF~KCP~HT$CID~$DIN~G 2536
~.HH:II:H.~.HHI:HIIIHIIHHI:H:H~H:.HHH~.~HIHH HH~:HIIImlHIHH.HII~II.~: lJl 2483 I$GTSC~DLDEcsQSPK~Y~CI~TEGSY~sCPRGYVLQF/)GI(TC~C0t.DECQTKQHNCQF~CVNTLGGFTCRCP~HTACID~$QPL~G 2582
2537 $KGI(X'~qTPC~$FTCECQ~̀SLDQTGS$CED'~DEcEGNHRCQHGCC)N~IC4~RCSCP~YLQHY~ENEcLS~IC~SCHNT~$YK~PAG 26~6 : IH I IH t l I I ,H t l I I I I I hH , IH I IH : IH IH I I I I I ~ : I I I I I : ~ [ I I I ;H I I IHHHI I I . : : I 1 : 111 .1111~11~ I I * J
2583 GKGICQNTPGSF$CECQRGFSLDATGLNCEDVDECDGNHRCQHGC~NILGGyROGCp~GYIQHYI~'4NQ CVDENECSNP~G~$cYNT~$¥K~PSG 2682
Z637 FQYE(]F.$GGCQ~INEcGsA~APc$YGC$NTEGGYLCGCPPGYF1~IC'~GHCv$GHGr̀ t~RGNPEPPV$G~DN$L$P~Y~KING¥P~.~R$~ 2734 : : I H : : ~ : I : I H : h . . H . H H H H I I I , I I H H : I : , I , , H H H : . : I . . : . . I : ~ : I . m l I H I I I I ~ H . I : : l . : I t . :
7683 FSFDQF$SACHDVN£C$$SK~PCNyGCSNTEGGyLCGCppGyYRvc'QGHCVS~GFNKGQ YLSLDTEVDE£NALSP~Y~KINGYSK~$~$1H 2781
2735 Z~̀T~DA$NIEDQ$Er~F~A~iy~'$LA$W{)V~KTAIF/tFhLL~HV$N~'3/RIL£LLPALTTLTNHNRYL~E$~E~FFKINQ~GI$YLH~VAGT¥$~]ssT 28]~ I . I ; . . ; 1 : : l h l I ~ : . . . : I 1 : 1 1 : : . 1 : i l ~ l I 1 : . ] . 1 1 I I ; . . I I ; 1 : . } ; I ; I ; ; 1 : 1 1 1 1 .1~1 : : 1 / I . 1 : 1 . 1 .
218Z ~pDpTAVEQ . . . . . . . I$LE$VIX'~)$P~`~`~L~-'~RLGsKEHILELRPAIQPLNNI41RYvIS(~(̀ NDDSVfRz~QR~LSYLHT~u~GTYT~IT£] 28~4
283~ PLY/~,ifJ@£L.I'~bgOKYDKOYL~E~NLI~X/Ct . . . . . . . VLLH 287~ [ f l I I IH . .H : . : . 1~ [ [H : . ~ : I : ; I : : r .
2875 pLy~6~ELKKLEESNEDDyHLGELGEAL~MRLOK~LK$OGLILK 2918
Figure 3. Comparison of the amino acid sequences of fibrillin-1 (top) and fibrillin-2 (bottom) with residues numbered according to Pereira et al. (1993). Identities and conservative changes are indi- cated by vertical bars, whereas one and two dots denote decreasing levels of evolutionary distance between amino acids calculated ac- cording to Gribskov and Burgess (1986). Dashes were introduced for best alignment; the structural elements described in the text are underlined. The sequence of the glycine- and proline-rich regions used for the generation of the antibodies are italicized and the afore- mentioned residues are indicated in bold character. Triangles iden-
Figure 4. (A) SDS-PAGE of radioactively labeled immunoisolates using anti-fibrillin-1 (Ab-flC) or anti-fibrillin-2 (Ab-f2C) antibod- ies. (B) unlabeled immunoprecipitates using anti-fibrillin-2 (lane 1 ) or anti-fibriUin-1 (lane 2) antibodies immunoblotted against Ab- tiC. Underneath each lane is a silver staining of the gel showing equal loading of the two proteins. The estimated protein size is indi- cated on the fight side of each panel. The discrepancy in the relative mounts of fibrillin-2 mRNA (see Fig. 2) and protein, compared to fibrillin-1, might be due to differences in antibody affinities, epi- tope presentation, rate of mRNA translation and/or protein secre- tion and stability.
kD (Fig. 4 A). In addition, Western blot analysis of the im- munoisolates documented the antibody specificity (Fig. 4 B). To demonstrate that fibriUin-2 is actually associated with extracellular microfibrils, we used immunogold labeling at the ultrastrucmral level. In this analysis, both the anti-fibril- lin-2 antibody (Fig. 5 A) and the anti-fibdllin-1 antibody (Fig. 5 B) were specifically localized to microfibrils as- sociated with elastic fibers in fetal bovine ligamentum nuchae. Similar specific localization of the antibodies was also observed on microfibrils of elastic laminae in fetal bo- vine aorta (data not shown).
1Issue Distribution of the FibriUin Proteins The fibrillin antibodies were then used to compare and con- trast the tissue distribution of these macromolecules. As pre- viously shown by Sakai et al. (1986), we found that fibrillin-1 staining in the skin involves the discrete filaments that extend from the dermis to the epidermal junction (Fig. 6 A). Fibrillin-2 staining, on the other hand, was qualitatively, if not quantitatively, comparable to that of fibrillin-1, and rela- tively higher in the periappendiceal dermis around hair folli- cles and sweat glands (Fig. 6 B).
Immunostalning of fetal aorta and cartilaginous tissues highlighted the enrichment of fibrillin-2 in elastic fiber-rich matrices. While the anti-fibrillin-1 antibody reacted with the
tify putative signal peptide cleavage sites; asterisks indicate the coding boundaries of MF-23; and underlines highlight the struc- tural elements discussed in the text. The complete nucleotide se- quence of fibrillin-2 is available from EMBL/C, enBank/DDJB un- der accession number U03272.
The Journal of Cell Biology, Volume 124, 1994 858

Figure 5. Immunolocalization of fibrillin-2 (,4) and fibrillin-1 (B) on a developing elastic fiber in 170-d gestation bovine ligamentum nuchae. Gold particles label the peripheral mantle of microfibrils (MF) of the elastic fiber demonstrating the specific association of fibrillins with the microfibrils. Note that the central core of amor- phous elastin (E) is devoid of label. Bar, 0.2 ~m.
connective tissue across all three layers of the fetal aorta (Fig. 7 A), staining with the anti-fibriUin-2 antibody was more intense in the media where elastic fibers are most abun- dant (Fig. 7 B). In elastic cartilage, fibrillin-1 staining was strong to moderate in the connective tissue adjacent to the central elastic cartilaginous core, but the core itself remained urtreactive (Fig. 8 A). In contrast, this same core region stained strongly and evenly with the anti-fibrillin-2 antibody (Fig. 8 B). The same antibody also stained intensely the net- work of fibrils in the perichondrium directly adjoining the cartilage core (Fig. 8 B). In hyaline cartilage, fibriUin-1 epi- topes were distributed throughout the cartilage and sur- rounding matrices (Fig. 8 C), while fibriUin-2 epitopes were mostly identified in the peripheral areas of the cartilage and in the perichondrium (Fig. 8 D). Interestingly, staining with the anti-fibrillin-1 antibody was somewhat augmented in the hypertrophic zone of the osteogenic cartilage where fibril- lin-2 was nearly undeteetable (Fig. 8, E and F).
In the cornea, fibrillin-2 expression was found to be more restricted than that of fibrillin-1, notably to the anterior third of Descemet's membrane (Fig. 9, A and B). This morpholog- ically distinct portion of Descemet's membrane is organized in a characteristic succession of banded nodes intercon- nected by strands and is known to be rich in type II collagen fibrils (von der Mark et al., 1977; Hendrix et al., 1982). Fibrillin-1 was predominantly found in the outer layer and in-
Figure 6. Indirect immunofluorescence localization of fibrillins ¢pitopes in human fetal skin using anti-fibrillin-1 (.4) and anti-fibril- lin-2 (B) antibodies. Bar, 100/~m.
ner vascular choroid layer of the sclera, while fibrillin-2 was evenly distributed throughout (Fig. 9, D and E). A similar pattern of even, albeit low, fibrillin-2 distribution was ob- served in the lung where fibrillin-1 staining was significantly higher, particularly in the interlobular septa (Fig. 9, G and H). In contrast, the anti-fibrillin antibodies exhibited similar staining patterns in the basement membrane of the glomeruli and in the developing collecting ducts of the fetal kidney (Fig. 9, J and K).
Discussion
The term "rnicrofibrils" was originally used to identify mor- phologically similar matrix aggregates lacking the character- istic banding periodicity of interstitial collagen fibers (Low, 1962). This broad definition was subsequently narrowed to include only microfibrils with a diameter of 10 nm that may or may not be associated with elastic fibers (Cleary and Gib- son, 1983). The complete inventory of the macromolecular constituents of the microfibrils is presently unknown, chiefly because of the highly insoluble nature of these structures. Biochemical and immunochemical techniques have never- theless identified a number of distinct microfibrillar-associ- ated macromolecules (Gibson et al., 1989). Structural infor- mation about some of these microfibrillar components has
Zhang et al. Microfibrils Contain a Second FibriUin Protein 859

The Journal of Cell Biology, Volume 124, 1994 860
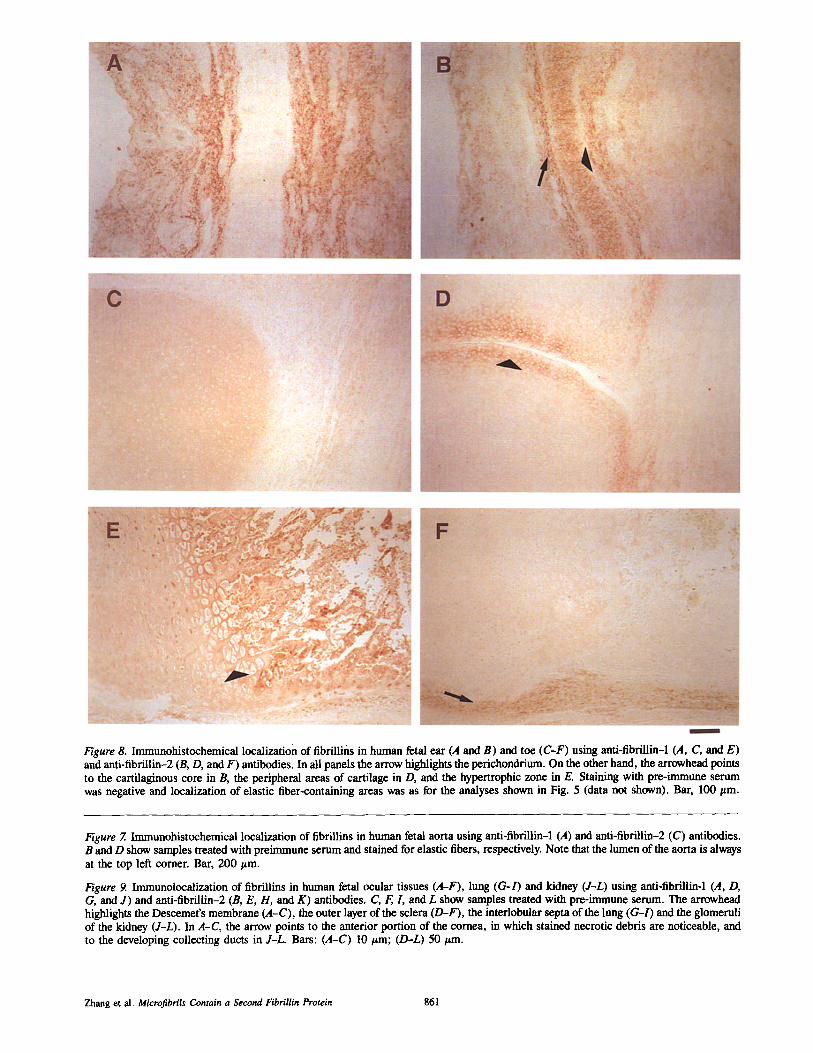
Figure 8. Immunohistochemical localization of fibrillins in human fetal ear (A and B) and toe (C-F) using anti-fibrillin-1 (,4, C, and E) and anti-fibrillin-2 (B, D, and F) antibodies. In all panels the arrow highlights the perichondrium. On the other hand, the arrowhead points to the cartilaginous core in B, the peripheral areas of cartilage in D, and the hypertrophic zone in E. Staining with pre-immtme serum was negative and localization of elastic fiber-containing areas was as for the analyses shown in Fig. 5 (data not shown). Bar, 100 gm.
Figure Z Immunohistochemical localization of fibrillins in human fetal aorta using anti-fibrillin-1 (,4) and anti-fibrillin-2 (C) antibodies. B and D show samples treated with preimmune serum and stained for elastic fibers, respectively. Note that the lumen of the aorta is always at the top left corner. Bar, 200/zm.
Figure 9. Immunolocalization of fibrillins in human fetal ocular tissues (A-F), lung ((7-/) and kidney (J-L) using anti-fibrillin-1 (,4, D, G, and J) and anti-fibrillin-2 (B, E, H, and K) antibodies. C, E L and L show samples treated with pre-immune serum. The arrowhead highlights the Descemet's membrane (A-C), the outer layer of the sclera (D-F), the inteflobular septa of the lung (G-l) and the glomeruli of the kidney (J-L). In A-C, the arrow points to the anterior portion of the cornea, in which stained necrotic debris are noticeable, and to the developing collecting ducts in J-L. Bars: (A-C) I0 gm; (D-L) 50/zm.
Zhang et al. Microfibrils Contain a Second Fibrillin Protein 861

been recently gathered from the cloning of the genes coding for the 31-kD microfibril-associated glycoprotein (MAGP), the 58-kD associated microfibril protein (AMP) and the 350- kD fibrillin (Gibson et al., 1991; Chert et al., 1993; Horrigan et al., 1992; Maslen et al., 1991; Lee et al., 1991; Corson et al., 1993; Pereira et al., 1993). Contrary to previous evi- dence, cloning of a partial transcript suggested that fibrillin may represent a small family of extracellular proteins (Lee et al., 1991). Results presented in this paper confirm and extend this observation; in addition, they provide the first comparison of the tissue distribution of the fibrillin family members.
The sequencing data indicate that the structure of fibril- lin-2 replicates very closely that recently established for fibrillin-I (Maslen et al., 1991; Lee et al., 1991; Corson et al., 1993; Pereira et al., 1993). As a result, the struc- tural/functional criteria defining a fibriUin molecule are be- ginning to emerge. Accordingly, such a macromolecule is expected to be made of five structurally distinct regions preceded by a short signal peptide. Two of these regions are composed of cysteine-rich sequences that can be segregated into three distinct groups. The first and largest, comprises numerous EGF-CB repeats and fewer EGF-like repeats (Davis, 1990). The second group features an 8-eysteine mo- tif which was first described interspersed among the EGF- like repeats of the TGF-/31-binding protein, and is termed the TGF-bp motif (Kanzaki et al., 1990). The third class of fibrillin repeats is heterogeneous in composition and appar- ently unique to this protein (Corson et al., 1993; Pereira et al., 1993). We have referred to these repeats as the Fib motifs and recently postulated their evolution from various rear- rangements of ancestral exons coding for EGF-like and TGF- bp repeats (Pereira et al., 1993). We based our evolutionary considerations on structural homologies and the organiza- tion of the coding sequence of the FBN1 gene, which prelim- inary evidence suggests to be similar in the FBN2 gene.
In our previous work on fibrillin-1, we suggested that re- gion C may be of some potential relevance to microfibril as- sembly (Pereira et al., 1993). Based on its relative position and composition, we argued that region C may bend the fibril/in molecule and thus facilitate the postulated protein interactions of the cysteine-rich regions (Pereira et al., 1993). Such a scenario needs not to be changed for fibril- lin-2, since the sequence of region C could theoretically form a poly-glycine hinge and thus provide the protein with a high degree of flexibility at that position (Claassen and Gross- mann, 1991). Region E is also of potential relevance to microfibril assembly, for it contains the same two cysteines already noted in the corresponding segment of fibrillin-1 (Maslin et al., 1981). The maintenance of these residues in two different chains, together with the extensive identity of the sequence around them, may conceivably indicate that they participate in the polymerization of individual fibriUin monomers.
In general, the tissue distribution of fibrilfin-1 resembles that of fibrillin-2. There are however some notable differ- ences which are likely to imply distinct functions of the two glycoproteins in the assembly and maintenance of the microfibrillar network. The most obvious of these differ- ences is the preferential localization of fibrillin-2 to extracel- lular matrices that are rich in elastic fibers. This feature is best exemplified in the elastic cartilage of the external ear.
This preferential distribution of fibrillin-2 is also consistent with the abnormal presentation of the CCA auricle which is characterized by flattening of the helix and crumpled appear- ance of the antibelix with resulting partial obliteration of the concha (Beals and Hecht, 1971). Hyaline cartilage is another tissue that exhibits the intriguing feature of a differential dis- tribution of the two fibrillins, plausibly a reflection of distinct contributions of these proteins to the process of skeletogene- sis. Along this fine, it is of some interest to note the restricted localization of fibrillin-2 to the type II collagen-rich layer of Descemet's membrane.
It is likely that the contribution of fibrillin-2 to tissue in- tegrity will be eventually clarified with the analysis of natu- rally occurring mutations in humans and artificially gener- ated ones in transgenic mice. Along these lines, the complete characterization of the FBN2 gene and the detailed examina- tion of its early developmental pattern of expression are both necessary preconditions to undertaking these investigations. From this and related work, it is becoming apparent that the function of the 10-nm microfibrils is likely to depend on the integrity and proper spatio-temporal expression of several distinct gene products. More generally, the microfibrillar aggregates with or without elastin cores are increasingly be- coming appreciated as critical players in providing biome- chanical properties to a wide variety of tissues and organ systems.
This study could not have been completed without the assistance and gener- ous support of many of our colleagues, namely Drs. V. Frexiedch, R. Laz- zarini, M. Lebwhol, S. Morgello, and Mr. X. Li. We also wish to thank Ms. M. Sozomenu for the preparation of the manuscript.
This work was supported from grants of the National Institutes of Health (AR42044, HL26499 and AR40673) and the March of Dimes Birth Defects Foundation. This is article 129 from the Brookdale Center for Molecular
Biology.
Received for publication 22 September 1993, and in revised form 1 Decem-
ber 1993.
References
Beals, R. K., and F. Hecht. 1971. Congenital eontractural arachnodactyly: a heritable disorder of Connective tissue. J. Bone Join. Surg. 53-A:887-903.
Chert, Y., J. Franco, W. Yin, J. Germiller, U. Francke, and J. Bonadio. 1993. Structure, chromosomal localization and expression pattern of the marine Magp gene. J. Biol. Chem. 268:27381-27383.
Claassen, L. A., and L. Grossman. 1991. Deletion mutagenesis of Escherichia coli UvrA protein localizes domains for DNA binding, damage recognition, and protein-protein interactions. J. Biol. Chem. 266:11388-11394.
Cleary, E. G., and M. A. Gibson. 1983. Elastin-associated microfibrils and microfibrillar proteins. Int. Rev. Connect 1issue. Res. 10:97-209.
Corson, G. M., S. C. Chalberg, H. C. Dietz, N. L. Charbonneau, and L. Sakai. 1993. Fibrillin binds calcium and is coded by cDNAs that reveal a muhido- main structure and alternatively spliced exOns at the 5' end. Genomics. 17:476-484.
Davis, C. G. 1990. The many faces of epidern'ud growth factor repeats. New Biol. 2:410--419.
Dietz, H. C., G. R. Cutting, R. E. Pyeritz, C. L. Maslen, L. Y. Sakai, G. M. Corson, E. G. Puffenberger, A. Hamosh, E. J. Nanthakumar, S. M. Cur- ristin, G. Stetten, D. A. Meyers, and C. A. Franeomano. 1991. Mar'fan syn- drome caused by a recurrent de novo missense mutation in the fibrillin gene. Nature (Lond.). 352:337-339.
Dietz, H. C., R. E. Pyeritz, E. G. Puffenberger, R. J. Kendzior, G. M. Corson, C. L. Maslan, L. Y. Sakai, C. A. Frmacomano, and G. R. Cutting. 1992a. Marfan phenotype variability in a family segregating a rnissense mutation in the epidermal growth factor-like motif of the fibrillin gene. J. Clin. Invest. 89:1674-1680.
Dietz, H. C., J. Saralva, R. E. Pyeritz, G. R, Cutting, and C. A. Franeomano. 199"2b. Clustering of fibrillin (FBNI) miss~nse mutations in Marfan Syn- drome patients at cysteine residues in EGF-like domains. Hum. Murat. 1:366-3"/4.
Dietz, H. C., D. Valle, C. A. Francornano, R. J. Kendzior, Jr., R. E. Pyeritz,
The Journal of Cell Biology, Volume 124, 1994 862

and G. R. Cutting. 1993a. The skipping of constitutive exons in vivo induced by nonsense mutations. Science (Wash. DC). 259:680-683.
Dietz, H. C., L Mcl.ntosh, L. Y. Sakai, G. M. Corson, S. C. Chalberg, R. E. Pyefitz, and C. A. Francomano. 1993b. Four novel FBN1 mutations: Significance for mutant transcript level and EGF-like domain calcium bind- ing in the pathogenesis of Marfan syndrome. Genomics. 17:468-475.
Gibson, M. A., L. B. Sandberg, L. E. Grosso, and E. G. Cleary. 1991. Com- plementary DNA cloning establishes microfibril-associated glycoprotein (MAGP) to be a discrete component of elastin-associated microfihrils. J. Biol. Chem. 266:7596-7601.
Gibson, M. K., J. S. Kumarntilnke, and E. G. Cleary. 1989. The protein com- ponents of the 12-manometer microfibrils of elastic and nonelastic tissues. J. Biol. Chem. 264:4590-4598.
Gribskov, M., and R. R. Burgess. 1986. Sigma factors from E. coli, B. subtilis, phage SPO1 and phage T4 are homologous proteins. Nucleic Acids Res. 14: 6745-6763.
Hartow, E., and D. Lane. 1988. Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 471-510.
Hendrix, M, J. C., E, D. Hay, K. vonder Mark, and T. F, Lynsenmayer. 1982. Immunohistochemical localization of collagen types I and II in the develop- ing chick cornea and tibia by electron microscopy. Invest. Ophthalmol. Vis. Sci. 22:359-375.
Hewett, D. R,, J. R. Lynch, R. Smith, and B. C. Sykes. 1993. A novel fibrillin mutation in the Marfan syndrome which could disrupt calcium binding of the epidermal growth factor-like module. Hum. Genet. 2:475-477.
Horrigan, S. K., C. B. Rich, B, W. Streeten, Z. Y. Li, andJ. A. Foster. 1992. Characterization of an associated microfihril protein through recombinant DNA techniques. J. Biol. Chem. 267:10087-10095.
Kanzaki, T., A, Olofsson, A, Moren, C. Wemstedt, U. Hellrnan, K. Miya- zono, L. Claesson-Welsh, and C. H, Haldin. 1990. TGF-/~I binding protein: a component of the large latent complex of TGF-~I with multiple repeat se- quences. Cell. 61:1051-1061.
Kainulainen, K., L. Pulkkinen, A. Savolainen, 1. Kaitila, and L. Paltonen. 1990. Location on chromosome 15 of the gene defect causing Marfan syn- drome. N, Engl. J. Med. 323:935-939.
Kainulainen, K., L. Y. Sakai, A. Child, F. M. Pope, L. Puhakka, L. Ryhanen, A. Palotie, I. Kaitila, and L. Peltonen. 1992. Two mutations in Marfan syn- drome resulting in truncated fibrillin polypeptides. Proc. Natl. Acad. Sci. USA. 89:5917-5921.
Lee, B., M. D'Alessio, and F. Ramirez. 1991. Modifications in the organization and expression of collagen genes associated with skeletal disorders. Crit. Rev. Euk. Gene. Exp. 1:173-187.
Lee, B., M. Godfrey, E. Vitale, H. Hori, M. G. Mattei, M. Sarfarazi, P. Tsipouras, F. Ramirez, and D. W. Hollister. 1991. Linkage of Marfan syn- drome and a phenotypically related disorder to two flbrillin genes. Nature (Lond.). 352:330--334.
Low, F. N. 1962. Microfibrils: fine filamentous components of the tissue space. Anat. Rec. 142:131-137.
Luna, L. G. 1968. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology. McGraw-Hill Co., New York, NY. 251 pp.
Maslen, C. L., G. M. Corson, B. K. Maddox, R. W. Glanville, and L. Y. Sakai. 1991. Partial sequence of a candidate gene for the Marfan syndrome. Nature (Lond.). 352:334--337.
Mecbem, R. P., and J. E. Heuser. 1991. The Elastic Fiber. In Cell Biology of the ExtraceUular Matrix. 2rid edition. E. D. Hay, editor. Plenum Publish- ing Co., New York. 79-109.
Pureira, L., M. D'Alessio, F. Ramirez, L R. Lynch, B. Sykes, T. Pangilinan, and J. Bonadio. 1993. Genomic organization of the sequence coding for fihrillin, the defective gene product in Marfan syndrome. Hum. MoL Genet. 2:961-968.
Ramirez, F., L. Pereira, H. Zhang, and B. Lee. 1993. The fibrillin-Marfan syn- drome connection. BioEssays. 15:589-594.
Sakai, L. Y., D. R. Keene, and E. Engvall. 1986. Fibrillin, a new 350-kD gly- coprotein, is a component of extracellular microfihrils. J. Cell Biol. 103: 2499-2509.
Sakai, L. Y., D. R. Keene, R. W. Glanville, and H. P. B[tchinger. 1991. Purification and partial characterization of fibrillin, a cysteine-rich structural component of connective tissue microfibrils. J. Biol. Chem. 266:14763- 14770.
Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: A Lab- oratory Manual. 2rid edition. Cold Spring Harbor Laboratory Press, NY. 545 pp.
Tsipouras, P., R. Del Mastro, M. Sarfarazi, B. Lee, E. Vitale, A. Child, M. Godfrey, R. Devereux, D. Hewett, B. Steinmann, D. Viljoen, B. C. Sykes, M. Kilpatrick, and F. Ramirez. 1992. Linkage of Marfan syndrome, domi- nant ectopia lentis and congenital eontractural arachnodactyly to the fibrillin genes on chromosome 15 and 5. N. Eng. J. Med. 326:905-909.
yon der Mark, K., H. yon der Mark, R. Timpl, and R. L. Trelstad. 1977. Im- munofluorescent localization of collagen types I, II and Ill in the embryonic chick eye. Dev. Biol. 54:75-85.
Zagursky, R. L, M. L. Berman, K. Banmeister, and N. Ix)max. 1986. Rapid and easy sequencing of linear double stranded DNA and supercoiled plasmid DNA. Gene Anal. Tech. 2:89-94.
Zhang et al. Microfibrils Contain a Second Fibrillin Protein 863





























