Embed Size (px)
Citation preview

© 2010 E. SchweizerbartÕ sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.deDOI: 10.1127/1869-6155/2010/0128-0015 1869-6155/2010/0128-0015 $ 04.50
Received January 16, 2009, in revised form August 3, 2009, accepted August 30, 2009
Plant Div. Evol. Vol. 128/3–4, 329–346E Stuttgart, September 17, 2010
Structural traits of some species of Hydrocotyle (Araliaceae) and their significance for constructing the generic system
By Alexandra I. Konstantinova and Elena Yu. Yembaturova
With 7 figures and 1 table
Abstract
Konstantinova, A.I. & Yembaturova, E.Yu.: Structural traits of some species of Hydrocotyle (Arali-aceae) and their significance for constructing the generic system. — Plant Div. Evol. 128: 329–346. 2010. — ISSN 1869-6155.
The characterization of Hydrocotyle species was traditionally almost entirely based on leaf characters, but these features are quite variable and directly dependent on age and ecological factors. However, we have shown that the fruits of representatives of these species, which display a certain amount of uniformity with regard to their macromorphology are very diverse anatomically. The objective evalu-ation of this kind of polymorphism in fruit anatomy, which is connected, first of all, with the size and shape of the cells constituting the crystalliferous layer in the pericarp, presence or absence of so-called “hydrocyte parechyma” complexes amongst the fruit tissues and other key features for Apiaceae sys-tematics, seems to be extremely important for developing well-grounded concepts of Hydrocotyle generic system.
Keywords: fruit anatomy, generic system, Hydrocotyle, morphology.
Introduction
The taxonomic history of Hydrocotyle L., as well as other representatives of Hydro-cotylaceae Hylander (1945), starts from the works of J. P. Tournefort, who mentioned several species of this genus (H. vulgaris L., H. asiatica L., H. umbellata L.) thus in-troducing its generic name, later validated by C. Linnaeus in 1753 (Shan & Liou 1964). Linnaeus also described H. americana L. and H. chinensis L., and new genera Solandra L. and Centella L. (a little later), which were included in the genus Hydrocotyle by C. Linnaeus filius as H. solandra L.f., H. villosa L.f., H. glabrata L.f. and along with H. virgata L.f., H. linifolia L.f., H. tridentata L.f., H. ranunculoides L.f. and H. erecta L.f., described by him (Richard 1820, Perez-Moreau 1948). Throughout the last centu-ries, various investigators had been discovering new species in this genus; the confu-sion in their synonymics, which arose later on, was initiated by their limited awareness

330 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
of the studies of both their precursors and contemporaries (Richard 1820). In the 13th edition of “Systema naturae…” of Linnaeus, published in 1791, J.F. Gmelin recorded 17 species of Hydrocotyle, and C.L. Willdenow, following a critical revision, increased the number of species up to 18, though rejecting some of previously described species and including the genus Spanathe Jacq. (described in 1789) in the genus Hydrocotyle, deprived of its generic status — as H. spanathe Willd. C.P. Thunberg, though holding the opinion that Spananthe is an autonomous genus, treated Hydrocotyle as consisting of 21 species (Richard 1820). The number of newly described species continued to grow steadily — thus, A. H. Ruiz & J. Pavon in “Flora Peruviana et Chilensis” (1802) described 9 new Hydrocotyle species (H. triflora Ruiz & Pavon , H. multiflora Ruiz & Pavon, H. globiflora Ruiz & Pavon, H. acutofolia Ruiz & Pavon, H. gracilis Ruiz & Pavon, H. incrassata Ruiz & Pavon et al.), and A. Michaux in “Flora boreali-ameri-cana…” (1803) presented the original description of H. lineata Michx.
In 1820 A. Richard clarified the synonyms and presented rather detailed descriptions of 60 Hydrocotyle species, placed by him in several sections. As he said (Richard 1820), at first, amazed at the huge interspecific distinctions, he had considered advisable to exclude several independent genera from Hydrocotyle, but later, after studying a large number of species thoroughly, he had quit that idea since he had observed how the dis-tinctions had dissolved in each other and had become less and less pronounced. Richard based his sectional divisions mostly on the difference in leaf blade shape and inflores-cence features. According to him, the distinctions in fruit structure of Hydrocotyle spe-cies are not significant enough to be used for setting intergeneric boundaries.
At the present time the number of species in Hydrocotyle is about 100 (Willis 1988) or even 130 (Pimenov & Leonov 1993). Moreover, the number of newly described species and varieties of Hydrocotyle (Mathias & Constance 1951, 1975, Constance & Dillon 1990) as well as the varieties acquiring specific status (Webb & Johnson 1982) continues to increase steadily. Although a huge gap in our knowledge of the genus pointed out by M. Mathias (1936) — the absence of full-valued monograph meeting all contemporary standards and embracing the entire genus — is unfortunately still to be filled. A colossal work was needed, nomenclatural aspects of which though were clarified by H. Eichler (Eichler 1987a, b, c) to a considerable extent.
Lately the concepts of placing Hydrocotyle and Trachymene Rudge close to each other within the сore Araliaceae as well as uniting Centella, Micropleura Lag., Platysace Bunge and Mackinlayeae (Plunkett & Lowry 2001, Chandler & Plunkett 2004, Plunkett et al. 2004a) within Apiaceae–Mackinlayoideae Plunkett & Lowry (Plunkett et al. 2004b) have been developed. As we believe, this concept needs to be thoroughly checked as it is at odds with the conclusions of the classical morphology and, in par-ticular, with carpological evidence on many genera Apiaceae-Mackinlayoideae, Apia-ceae-Azorelloideae and Araliaceae s. str. (Konstantinova, unpublished data).
As far as Hydrocotyle is concerned, it is noteworthy that molecular works, much to our regret, have always analyzed a very limited sampling of species of this genus (H. bowlesioides Math. & Const., H. verticillata Thunb., H. vulgaris — Plunkett et al. 2004a; H. bowlesioides, H. modesta Cham. & Schltdl., H. verticillata — Chandler & Plunkett 2004), and this does not give us an impression of Hydrocotyle diversity from the standpoint of recent molecular data.

A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 331
Thus, Hydrocotyle is a large, heterogenous genus, that is poorly defined based on macromorphological, generally vegetative characters alone. It is likely that a thorough work is needed in order to investigate the structure of reproductive sphere in this ge-nus’s species, in particular — not only macromorphological (shape, pubescence), but also anatomical details of pericarp structure. That is why the purpose of our research is to critically evaluate the advisability of such a study for this particular case and reveal the potential taxonomic value of fruit anatomy traits for establishing major groups within Hydrocotyle. Based on these traits, we attempted to line out possible carpologi-cal groups. We did not aim at finally defining these groups and determining their size and number as our limited sampling did not allow us to do that.
Materials and methods
The paper presents results of studying 16 species of the genus Hydrocotyle. The material for the re-search was obtained from the Herbarium of Botanical Institute of Russian Academy of Science (LE) and Herbarium of the Botanical Institute of Academia Sinica, Kunmimg, Yunnan, China (KUN).
The species were sampled the way that allows to, as much as possible, reflect the geographical spread of the genus Hydrocotyle: amongs the species there are cosmopolitan ones (H. bonariensis, H. verticillata), as well as species restricted to the Old World (H. sibthorpioides Lam., H. vulgaris) or to the New World (H. chamaemorus Cham. & Schltdl., Н.mexicana Cham. & Schltdl.). Also, some spe-cies are endemic to certain terrirories (H. poepigii DC. — endemic to Chile, H. hookeri Craib subsp. handelii (H. Wolff) M.F. Watson & M.L. Sheh, H. “dulongensis”, nom. unpubl.1 — to China).
To make the preparations, mainly traditional anatomical procedures were applied: dry fruits were kept in a mixture of 96% ethanol, glycerol and water in equal proportions for several days. To deter-mine the degree of lignification in cell walls as well as to locate tannins and oils corresponding his-tochemical test reactions were conducted (Prozina 1969, O’Brien & McCully, 1981).Transverse sec-tions through the middle part of fruits were examined by means of light and scanning electron microscopes (Hitachi S–450a, CamScan S–2m) at different magnifications.
Results
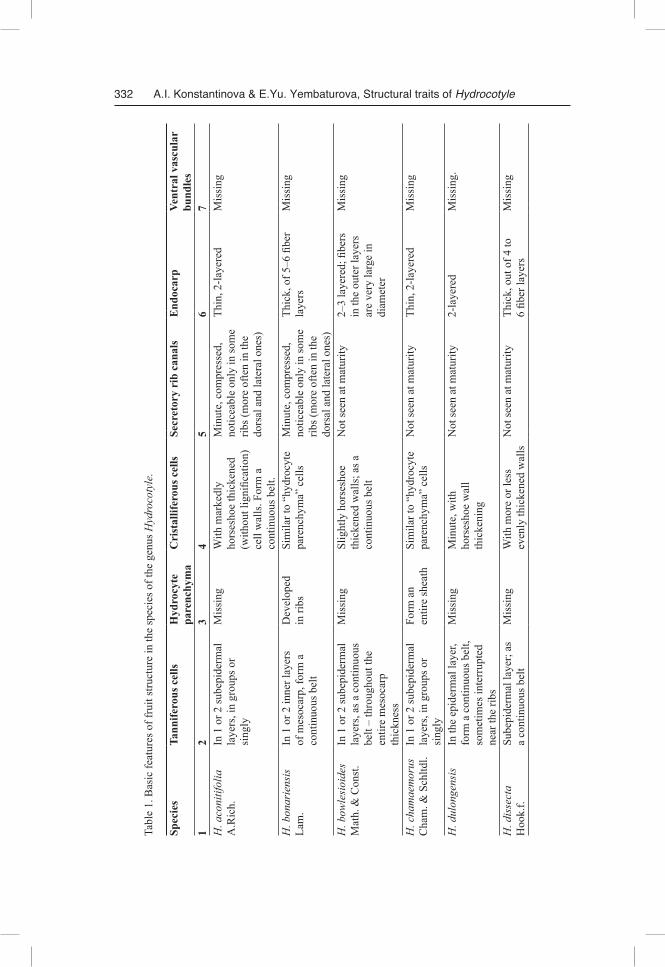
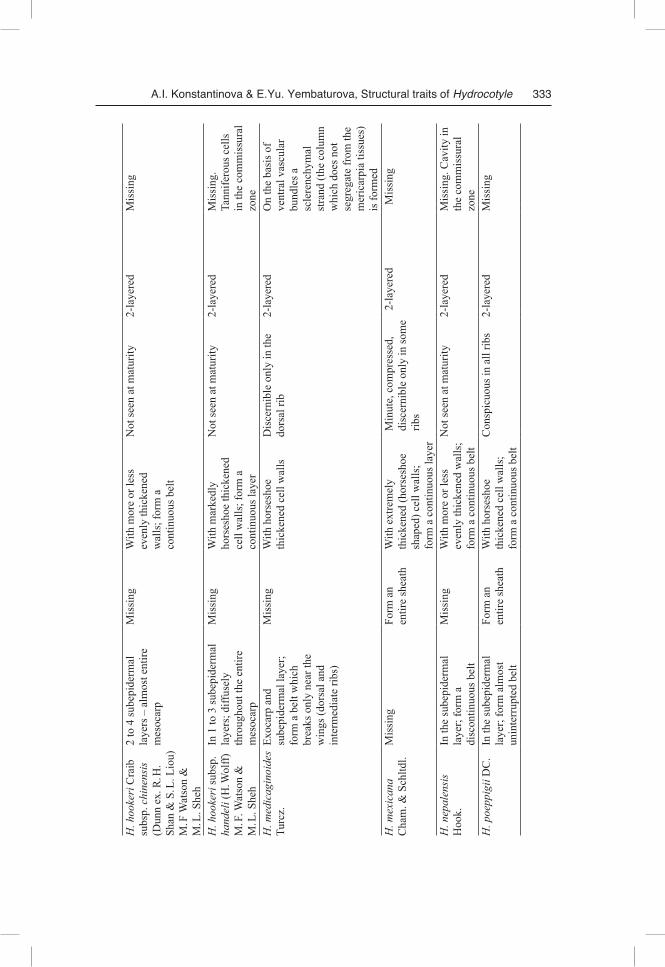
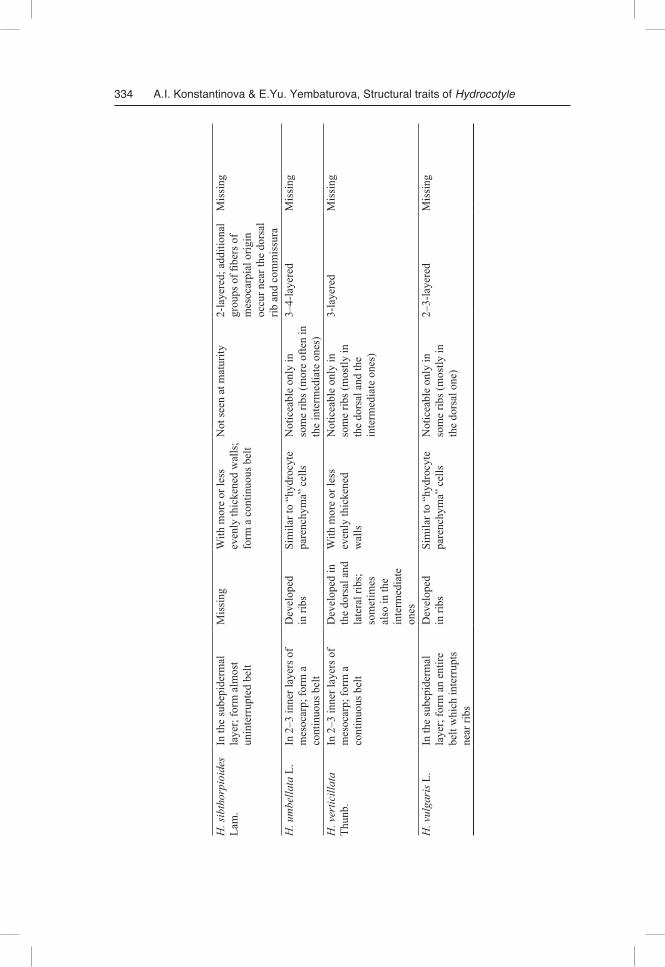
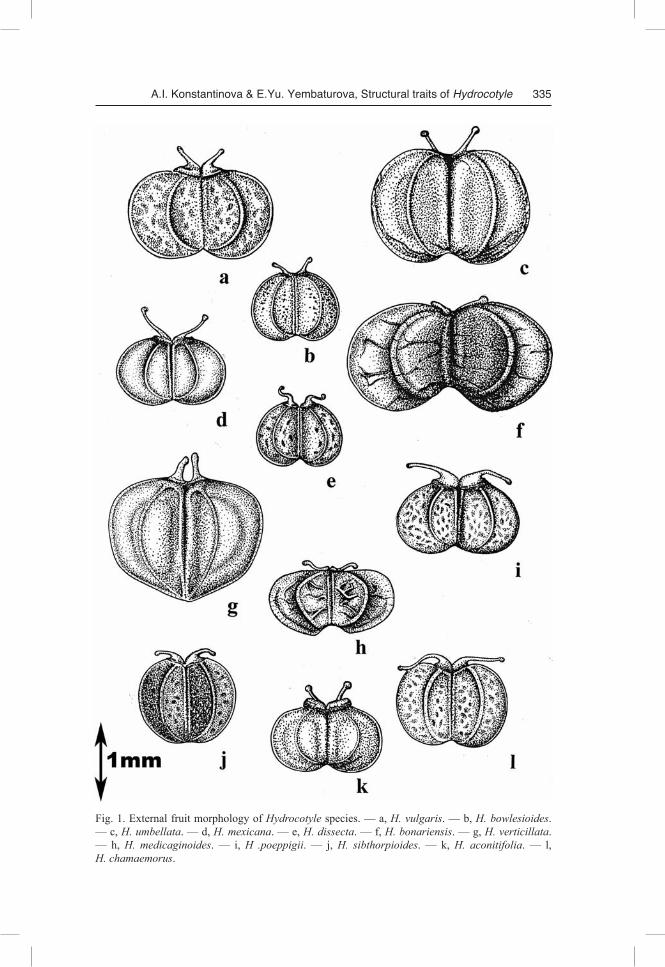
One of the primary methodological principles of our study was the strict necessity of obtaining comparable results. Therefore, even though our study has revealed signifi-cant diversity in morphological and anatomical features of fruit structure in the inves-tigated species, we still start from a general overview, which can be and should be given. The difference between Hydrocotyle species is presented in Table 1.
Fruit morphology
The fruit consists of two similar mericarps, markedly compressed from both sides.Mericarp outline is usually lanceolate from the back edge (oval in H. medicagi
noides Turcz.) and semi-circular (H. aconitifolia A. Rich., H. hookeri Craib subsp.
1 The species was incorrectly described by H. Li in “Flora of Dulongjia Region (NW Yunnan)” in 1993.
332 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
Tabl
e 1.
Bas
ic fe
atur
es o
f fru
it st
ruct
ure
in th
e sp
ecie
s of t
he g
enus
Hyd
roco
tyle
.
Spec
ies
Tann
ifero
us c
ells
Hyd
rocy
te
pare
nchy
ma
Cri
stal
lifer
ous c
ells
Secr
etor
y ri
b ca
nals
End
ocar
pVe
ntra
l vas
cula
r bu
ndle
s1
23
45
67
H. a
coni
tifol
ia
A.R
ich.
In 1
or 2
sube
pide
rmal
la
yers
, in
grou
ps o
r si
ngly
Mis
sing
With
mar
kedl
y ho
rses
hoe
thic
kene
d (w
ithou
t lig
nific
atio
n)
cell
wal
ls. F
orm
a
cont
inuo
us b
elt.
Min
ute,
com
pres
sed,
no
ticea
ble
only
in so
me
ribs (
mor
e of
ten
in th
e do
rsal
and
late
ral o
nes)
Thin
, 2-la
yere
dM
issi
ng
H. b
onar
iens
is
Lam
.In
1 o
r 2 in
ner l
ayer
s of
mes
ocar
p, fo
rm a
co
ntin
uous
bel
t
Dev
elop
ed
in ri
bsSi
mila
r to
“hyd
rocy
te
pare
nchy
ma”
cel
lsM
inut
e, c
ompr
esse
d,
notic
eabl
e on
ly in
som
e rib
s (m
ore
ofte
n in
the
dors
al a
nd la
tera
l one
s)
Thic
k, o
f 5–6
fibe
r la
yers
Mis
sing
H. b
owle
sioi
des
Mat
h. &
Con
st.
In 1
or 2
sube
pide
rmal
la
yers
, as a
con
tinuo
us
belt
– th
roug
hout
the
entir
e m
esoc
arp
th
ickn
ess
Mis
sing
Slig
htly
hor
sesh
oeth
icke
ned
wal
ls; a
s a
cont
inuo
us b
elt
Not
seen
at m
atur
ity
2–3
laye
red;
fibe
rs
in th
e ou
ter l
ayer
s ar
e ve
ry la
rge
in
diam
eter
Mis
sing
H. c
ham
aem
orus
C
ham
. & S
chltd
l.In
1 o
r 2 su
bepi
derm
al
laye
rs, i
n gr
oups
or
sing
ly
Form
an
en
tire
shea
thSi
mila
r to
“hyd
rocy
te
pare
nchy
ma”
cel
lsN
ot se
en a
t mat
urity
Th
in, 2
-laye
red
Mis
sing
H. d
ulon
gens
isIn
the
epid
erm
al la
yer,
form
a c
ontin
uous
bel
t, so
met
imes
inte
rrup
ted
near
the
ribs
Mis
sing
Min
ute,
with
ho
rses
hoe
wal
l th
icke
ning
Not
seen
at m
atur
ity2-
laye
red
Mis
sing
.
H. d
isse
cta
H
ook.
f.Su
bepi
derm
al la
yer;
as
a co
ntin
uous
bel
tM
issi
ngW
ith m
ore
or le
ss
even
ly th
icke
ned
wal
lsN
ot se
en a
t mat
urity
Thic
k, o
ut o
f 4 to
6
fiber
laye
rsM
issi
ng
A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 333
H. h
ooke
ri C
raib
su
bsp.
chi
nens
is
(Dun
n ex
. R. H
. Sh
an &
S. L
. Lio
u)
M. F
Wat
son
&
M. L
. She
h
2 to
4 su
bepi
derm
al
laye
rs –
alm
ost e
ntire
m
esoc
arp
Mis
sing
With
mor
e or
less
ev
enly
thic
kene
d
wal
ls; f
orm
a
cont
inuo
us b
elt
Not
seen
at m
atur
ity2-
laye
red
Mis
sing
H. h
ooke
ri su
bsp.
ha
ndel
i (H
. Wol
ff)
M. F
. Wat
son
&
M. L
. She
h
In 1
to 3
sube
pide
rmal
la
yers
; diff
usel
y th
roug
hout
the
entir
e m
esoc
arp
Mis
sing
With
mar
kedl
y ho
rses
hoe
thic
kene
d
cell
wal
ls; f
orm
a
cont
inuo
us la
yer
Not
seen
at m
atur
ity2-
laye
red
Mis
sing
. Ta
nnife
rous
cel
ls
in th
e co
mm
issu
ral
zone
H. m
edic
agin
oide
s Tu
rcz.
Exoc
arp
and
su
bepi
derm
al la
yer;
fo
rm a
bel
t whi
ch
brea
ks o
nly
near
the
win
gs (d
orsa
l and
in
term
edia
te ri
bs)
Mis
sing
With
hor
sesh
oe
thic
kene
d ce
ll w
alls
Dis
cern
ible
onl
y in
the
dors
al ri
b2-
laye
red
On
the
basi
s of
vent
ral v
ascu
lar
bund
les a
sc
lere
nchy
mal
st
rand
(the
col
umn
whi
ch d
oes n
ot
segr
egat
e fr
om th
e m
eric
arpi
a tis
sues
) is
form
ed
H. m
exic
ana
C
ham
. & S
chltd
l.M
issi
ngFo
rm a
n
entir
e sh
eath
With
ext
rem
ely
th
icke
ned
(hor
sesh
oe
shap
ed) c
ell w
alls
; fo
rm a
con
tinuo
us la
yer
Min
ute,
com
pres
sed,
di
scer
nibl
e on
ly in
som
e rib
s
2-la
yere
dM
issi
ng
H. n
epal
ensi
s H
ook.
In th
e su
bepi
derm
al
laye
r; fo
rm a
di
scon
tinuo
us b
elt
Mis
sing
With
mor
e or
less
ev
enly
thic
kene
d w
alls
; fo
rm a
con
tinuo
us b
elt
Not
seen
at m
atur
ity2-
laye
red
Mis
sing
. Cav
ity in
th
e co
mm
issu
ral
zone
H. p
oepp
igii
DC
.In
the
sube
pide
rmal
la
yer;
form
alm
ost
unin
terr
upte
d be
lt
Form
an
en
tire
shea
thW
ith h
orse
shoe
th
icke
ned
cell
wal
ls;
form
a c
ontin
uous
bel
t
Con
spic
uous
in a
ll rib
s2-
laye
red
Mis
sing
334 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
H. s
ibth
orpi
oide
s La
m.
In th
e su
bepi
derm
al
laye
r; fo
rm a
lmos
t un
inte
rrup
ted
belt
Mis
sing
With
mor
e or
less
ev
enly
thic
kene
d w
alls
; fo
rm a
con
tinuo
us b
elt
Not
seen
at m
atur
ity2-
laye
red;
add
ition
al
grou
ps o
f fibe
rs o
f m
esoc
arpi
al o
rigin
oc
cur n
ear t
he d
orsa
l rib
and
com
mis
sura
Mis
sing
H. u
mbe
llata
L.
In 2
–3 in
ner l
ayer
s of
mes
ocar
p; fo
rm a
co
ntin
uous
bel
t
Dev
elop
ed
in ri
bsSi
mila
r to
“hyd
rocy
te
pare
nchy
ma”
cel
lsN
otic
eabl
e on
ly in
so
me
ribs (
mor
e of
ten
in
the
inte
rmed
iate
one
s)
3–4-
laye
red
Mis
sing
H. v
ertic
illat
a Th
unb.
In 2
–3 in
ner l
ayer
s of
mes
ocar
p; fo
rm a
co
ntin
uous
bel
t
Dev
elop
ed in
th
e do
rsal
and
la
tera
l rib
s;
som
etim
es
also
in th
e in
term
edia
te
ones
With
mor
e or
less
ev
enly
thic
kene
d
wal
ls
Not
icea
ble
only
in
som
e rib
s (m
ostly
in
the
dors
al a
nd th
e in
term
edia
te o
nes)
3-la
yere
dM
issi
ng
H. v
ulga
ris L
.In
the
sube
pide
rmal
la
yer;
form
an
entir
e
belt
whi
ch in
terr
upts
ne
ar ri
bs
Dev
elop
ed
in ri
bsSi
mila
r to
“hyd
rocy
te
pare
nchy
ma”
cel
lsN
otic
eabl
e on
ly in
so
me
ribs (
mos
tly in
th
e do
rsal
one
)
2–3-
laye
red
Mis
sing
A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 335
Fig. 1. External fruit morphology of Hydrocotyle species. — a, H. vulgaris. — b, H. bowlesioides. — c, H. umbellata. — d, H. mexicana. — e, H. dissecta. — f, H. bonariensis. — g, H. verticillata. — h, H. medicaginoides. — i, H .poeppigii. — j, H. sibthorpioides. — k, H. aconitifolia. — l, H. chamaemorus.
336 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
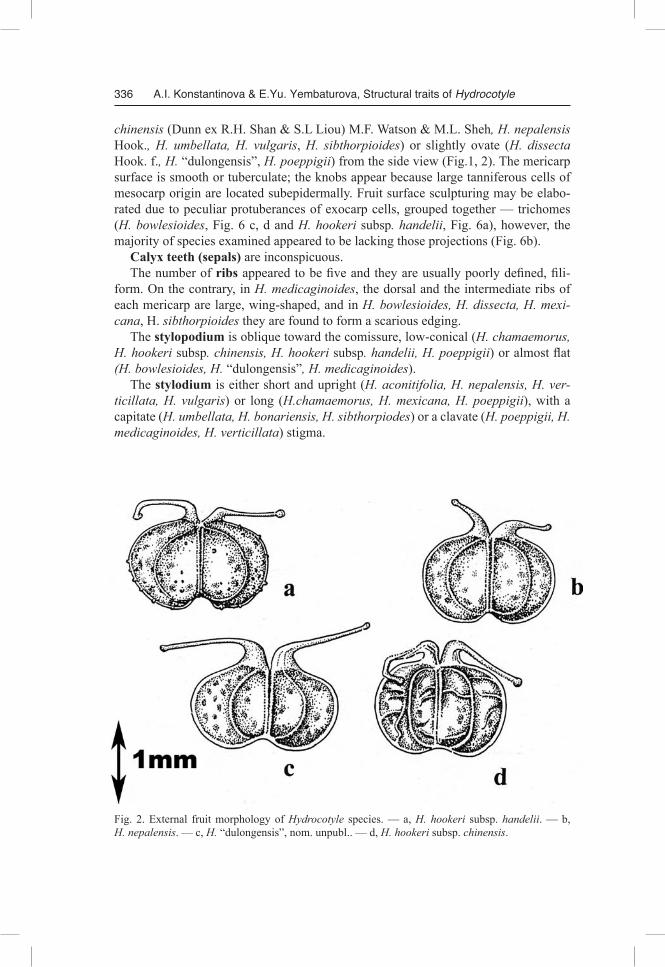
Fig. 2. External fruit morphology of Hydrocotyle species. — a, H. hookeri subsp. handelii. — b, H. nepalensis. — c, H. “dulongensis”, nom. unpubl.. — d, H. hookeri subsp. chinensis.
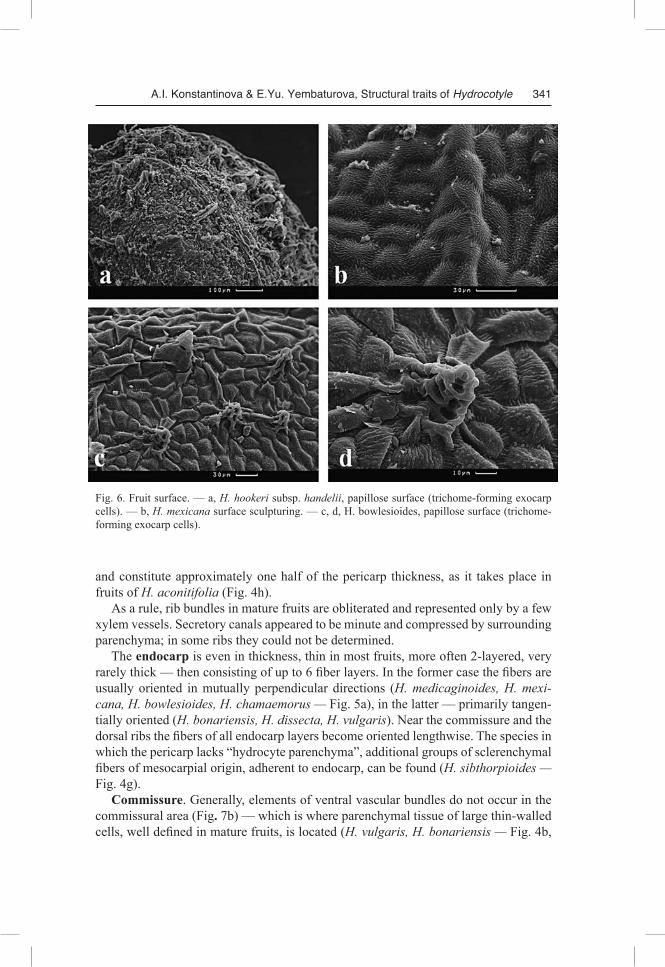
chinensis (Dunn ex R.H. Shan & S.L Liou) M.F. Watson & M.L. Sheh, H. nepalensis Hook., H. umbellata, H. vulgaris, H. sibthorpioides) or slightly ovate (H. dissecta Hook. f., H. “dulongensis”, H. poeppigii) from the side view (Fig.1, 2). The mericarp surface is smooth or tuberculate; the knobs appear because large tanniferous cells of mesocarp origin are located subepidermally. Fruit surface sculpturing may be elabo-rated due to peculiar protuberances of exocarp cells, grouped together — trichomes (H. bowlesioides, Fig. 6 c, d and H. hookeri subsp. handelii, Fig. 6a), however, the majority of species examined appeared to be lacking those projections (Fig. 6b).
Calyx teeth (sepals) are inconspicuous. The number of ribs appeared to be five and they are usually poorly defined, fili-
form. On the contrary, in H. medicaginoides, the dorsal and the intermediate ribs of each mericarp are large, wing-shaped, and in H. bowlesioides, H. dissecta, H. mexicana, H. sibthorpioides they are found to form a scarious edging.
The stylopodium is oblique toward the comissure, low-conical (H. chamaemorus, H. hookeri subsp. chinensis, H. hookeri subsp. handelii, H. poeppigii) or almost flat (H. bowlesioides, H. “dulongensis”, H. medicaginoides).
The stylodium is either short and upright (H. aconitifolia, H. nepalensis, H. verticillata, H. vulgaris) or long (H.chamaemorus, H. mexicana, H. poeppigii), with a capitate (H. umbellata, H. bonariensis, H. sibthorpiodes) or a clavate (H. poeppigii, H. medicaginoides, H. verticillata) stigma.

A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 337
Fruit anatomy
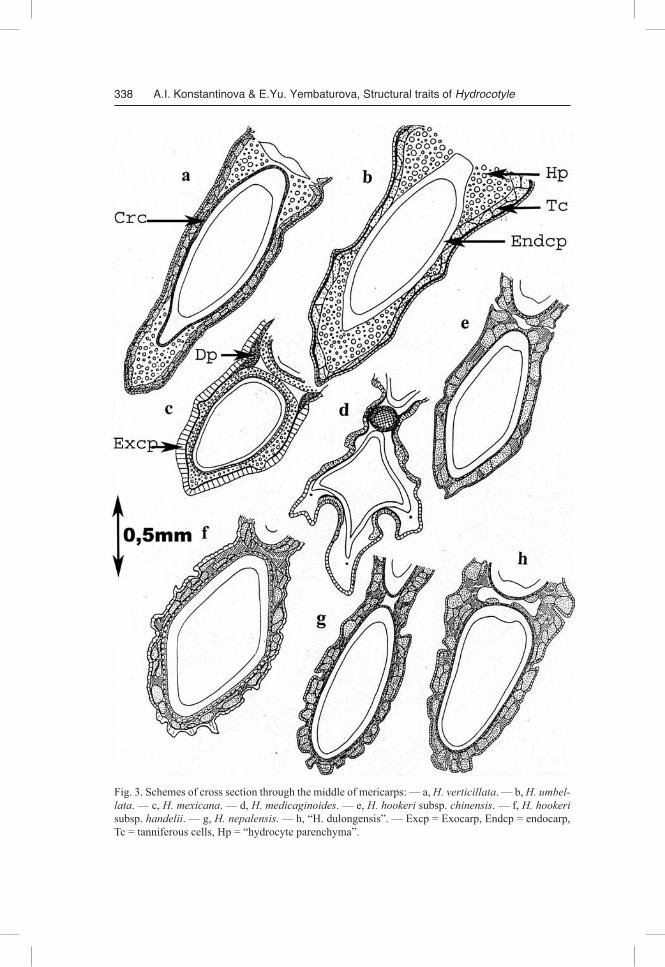
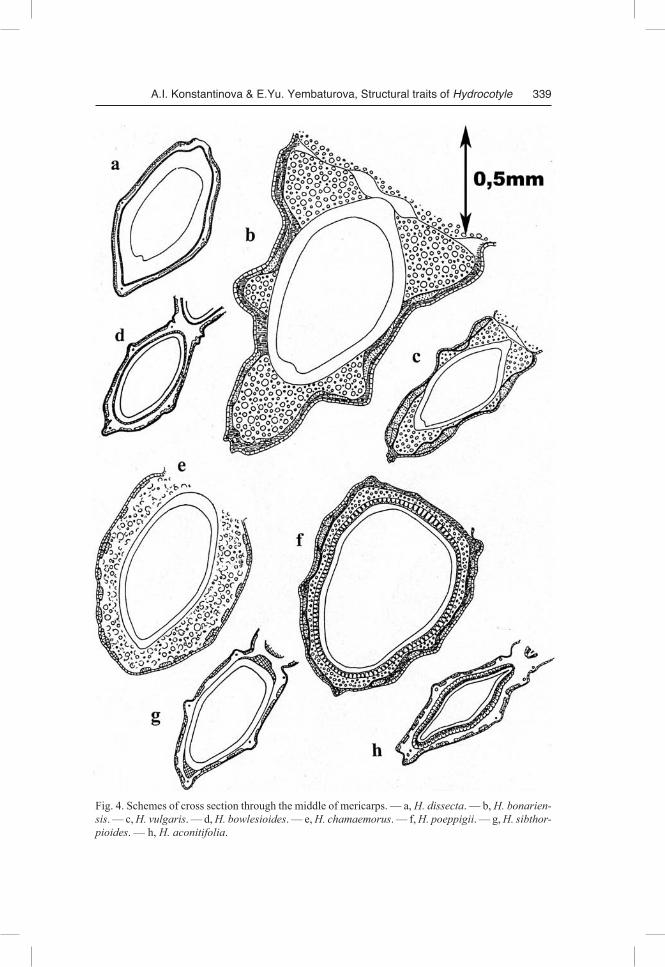
Outline of the transverse section (Fig. 3, 4). If cut in the middle, the mericarp in the cross section is oval (H. dissecta, H. “dulongensis”, H. bowlesioides, H. nepalensis), almost round (H. chamaemorus, H. poeppigii), shaped as an elongated triangle with the widest side between the marginal ribs (H. umbellata, H. bonariensis, H. vulgaris), or almost diamond-shaped (H. hookeri subsp. handelii, H. medicaginoides).
The exocarp is single-layered, consisting of cells slightly elongated tangentially. The exocarp cells in H. medicaginoides and H. mexicana are square or radially elon-gated. The outer cell walls, commonly slightly thickened, are covered with a thin cu-ticle layer. The exocarp cells can serve as reservoirs for tannins, like in H. medicaginoides (Fig. 3d), but more often mesocarp cells appear to be tanniferous (Fig.3a, b, e-h; 4a-h; 5b). In fruits of H. bowlesioides and H. hookeri subsp. handelii some trichome-forming exocarp cells (gathered in groups) have secondarily thickened walls and actu-ally appear to be “hydrocyte parenchyma” cells.
Mesocarp. One or several subepidermal layers usually are located where large tannin-containing cells are concentrated. In some species these cells make a continu-ous belt girdling the entire perimeter of mericarp cross section and breaking only where the ribs are located (H. dissecta, H. medicaginoides, H. sibthorpioides — Fig. 3d, 4a, g); in the other ones they are sporadic (H. chamaemorus, H. poeppigii — Fig. 4e, f) or are completely missing (H. mexicana — Fig. 3c). In fruits of H. “dulongensis”, H. hookeri subsp. chinensis, H. hookeri subsp. handelii and H .nepalensis, the entire me-socarp zone consists of such tanniferous cells (Fig. 3e-h). In a number of species the major part of the mesocarp consists of roundish cells with slightly thickened lignified walls, forming thick strands under the rib vascular bundles (H. vulgaris, H. umbellata, H. bonariensis — Fig. 3a, b; 4b, c). These cells can constitute a continuous belt around the endocarp (H. mexicana, H. chamaemorus, H. poeppigii — Fig. 3c, 4e, f). Their walls possess many large pore canals (Fig. 7f). The researchers have interpreted such cell complexes differently: “tissu aerifer” (Briquet 1899), “pleenchyma” (Tamamshy-an & Vinogradova 1969), “aerofores” (Kozo-Polyanski 1916), “hydrocyte parenchy-ma” (Klimochkina 1950). This tissue appears at relatively late stages of fruit develop-ment between a bundle and crystalliferous layer and while the fruit continues to develop, it causes the bundle to move a significant distance away (Pervukhina 1950). Apparently it serves to lighten the entire fruit construction and, being a sort of a float, holds the hydrochorous fruit at water surface.
In the species whose pericarp lacks such a tissue (Fig. 3d-h, 4a, d, g, h), the most of the mesocarp commonly comprises parenchyma deformed in the course of fruit devel-opment (H. hookeri subsp. chinensis, H. dissecta, H. “dulongensis”, H. bowlesioides, H. hookeri subsp. handelii, H. nepalensis, H. sibthorpioides, H. medicaginoides).
The innermost mesocarp layer is represented by crystalliferous cells in all studied Hydrocotyle species; some researchers treated this layer as a part of the endocarp (Kli-mochkina 1950) or mesocarp (Tseng 1967). As we believe, in this case such terms as “exocarp”, “mesocarp” or “endocarp” could be used only in conditional (topographical) sense, because Apiaceae fruits develop from the inferior ovary, constituted by elements of different (hypanthial and carpellary) origin which congenially become inseparable
338 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
Fig. 3. Schemes of cross section through the middle of mericarps: — a, H. verticillata. — b, H. umbellata. — c, H. mexicana. — d, H. medicaginoides. — e, H. hookeri subsp. chinensis. — f, H. hookeri subsp. handelii. — g, H. nepalensis. — h, “H. dulongensis”. — Excp = Exocarp, Endcp = endocarp, Tc = tanniferous cells, Hp = “hydrocyte parenchyma”.
A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 339
Fig. 4. Schemes of cross section through the middle of mericarps. — a, H. dissecta. — b, H. bonariensis. — c, H. vulgaris. — d, H. bowlesioides. — e, H. chamaemorus. — f, H. poeppigii. — g, H. sibthorpioides. — h, H. aconitifolia.
340 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
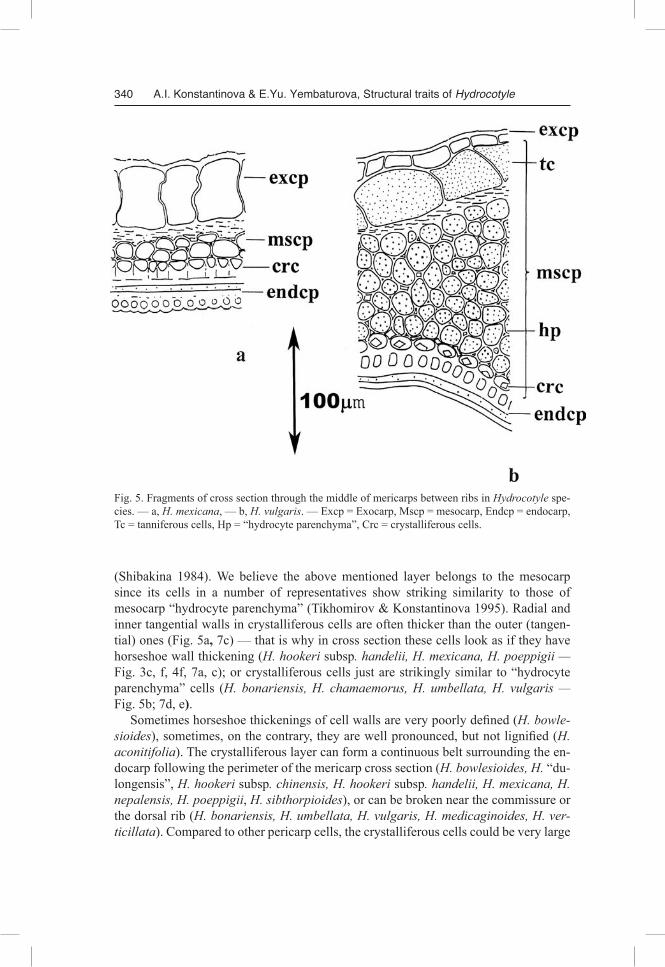
Fig. 5. Fragments of cross section through the middle of mericarps between ribs in Hydrocotyle spe-cies. — а, H. mexicana, — b, H. vulgaris. — Excp = Exocarp, Mscp = mesocarp, Endcp = endocarp, Tc = tanniferous cells, Hp = “hydrocyte parenchyma”, Crc = crystalliferous cells.
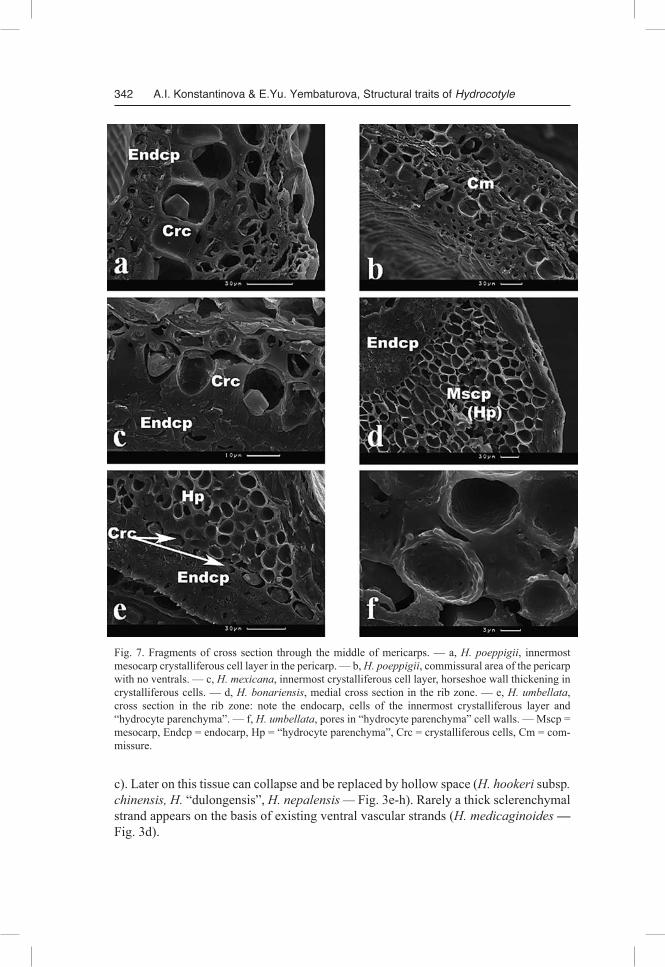
(Shibakina 1984). We believe the above mentioned layer belongs to the mesocarp since its cells in a number of representatives show striking similarity to those of mesocarp “hydrocyte parenchyma” (Tikhomirov & Konstantinova 1995). Radial and inner tangential walls in crystalliferous cells are often thicker than the outer (tangen-tial) ones (Fig. 5a, 7c) — that is why in cross section these cells look as if they have horseshoe wall thickening (H. hookeri subsp. handelii, H. mexicana, H. poeppigii — Fig. 3c, f, 4f, 7a, c); or crystalliferous cells just are strikingly similar to “hydrocyte parenchyma” cells (H. bonariensis, H. chamaemorus, H. umbellata, H. vulgaris — Fig. 5b; 7d, e).
Sometimes horseshoe thickenings of cell walls are very poorly defined (H. bowlesioides), sometimes, on the contrary, they are well pronounced, but not lignified (H. aconitifolia). The crystalliferous layer can form a continuous belt surrounding the en-docarp following the perimeter of the mericarp cross section (H. bowlesioides, H. “du-longensis”, H. hookeri subsp. chinensis, H. hookeri subsp. handelii, H. mexicana, H. nepalensis, H. poeppigii, H. sibthorpioides), or can be broken near the commissure or the dorsal rib (H. bonariensis, H. umbellata, H. vulgaris, H. medicaginoides, H. verticillata). Compared to other pericarp cells, the crystalliferous cells could be very large
A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 341
Fig. 6. Fruit surface. — a, H. hookeri subsp. handelii, papillose surface (trichome-forming exocarp cells). — b, H. mexicana surface sculpturing. — c, d, H. bowlesioides, papillose surface (trichome-forming exocarp cells).
and constitute approximately one half of the pericarp thickness, as it takes place in fruits of H. aconitifolia (Fig. 4h).
As a rule, rib bundles in mature fruits are obliterated and represented only by a few xylem vessels. Secretory canals appeared to be minute and compressed by surrounding parenchyma; in some ribs they could not be determined.
The endocarp is even in thickness, thin in most fruits, more often 2-layered, very rarely thick — then consisting of up to 6 fiber layers. In the former case the fibers are usually oriented in mutually perpendicular directions (H. medicaginoides, H. mexicana, H. bowlesioides, H. chamaemorus — Fig. 5a), in the latter — primarily tangen-tially oriented (H. bonariensis, H. dissecta, H. vulgaris). Near the commissure and the dorsal ribs the fibers of all endocarp layers become oriented lengthwise. The species in which the pericarp lacks “hydrocyte parenchyma”, additional groups of sclerenchymal fibers of mesocarpial origin, adherent to endocarp, can be found (H. sibthorpioides — Fig. 4g).
Commissure. Generally, elements of ventral vascular bundles do not occur in the commissural area (Fig. 7b) — which is where parenchymal tissue of large thin-walled cells, well defined in mature fruits, is located (H. vulgaris, H. bonariensis — Fig. 4b,
342 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
Fig. 7. Fragments of cross section through the middle of mericarps. — a, H. poeppigii, innermost mesocarp crystalliferous cell layer in the pericarp. — b, H. poeppigii, commissural area of the pericarp with no ventrals. — c, H. mexicana, innermost crystalliferous cell layer, horseshoe wall thickening in crystalliferous cells. — d, H. bonariensis, medial cross section in the rib zone. — e, H. umbellata, cross section in the rib zone: note the endocarp, cells of the innermost crystalliferous layer and “hydrocyte parenchyma”. — f, H. umbellata, pores in “hydrocyte parenchyma” cell walls. — Mscp = mesocarp, Endcp = endocarp, Hp = “hydrocyte parenchyma”, Crc = crystalliferous cells, Сm = com-missure.
c). Later on this tissue can collapse and be replaced by hollow space (H. hookeri subsp. chinensis, H. “dulongensis”, H. nepalensis — Fig. 3e-h). Rarely a thick sclerenchymal strand appears on the basis of existing ventral vascular strands (H. medicaginoides — Fig. 3d).

A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 343
Seed coat. As in all Apiaceae, possessing unitegmal seeds, it is represented by ex-otesta — thin structureless lamina. The boundaries between its markedly deformed cells usually cannot be identified.
Discussion
Richard (1820) made intrageneric divisions, based mostly upon the characters of the vegetative sphere (primarily leaf shape) and noted how difficult it was to identify spe-cies on the basis of fruit traits. This is not surprising, since the fruits of Hydrocotyle are very small and can hardly be determined by macromorphological features. In addition, it is not always possible to assess the peculiarities of fruit structure in different species, because, even though flowers and fruits often coexist within an inflorescence for a long period of time, the plant does not stay in the flowering and fruiting stage all the time. That is why to date the identification of Hydrocotyle species is traditionally based upon the leaf characters (Shan 1936, Constance & Dillon 1990). However, the use of leaf blade shape as the key feature often does not seem advisable even for identification purposes due to its marked variability depending on the season and habitual properties of a particular individual, probably conditioned by ecological and age-specific fac-tors.
Without denying the evident significance of leaf blade morphology features and other traits of the vegetative sphere in different Hydrocotyle species, the authors wish to specifically emphasize the importance of carpological characters. Taking these traits into consideration is equally (if not more) important for building up the concept of intrageneric system.
As a result of our study of 16 Hydrocotyle species significant polymorphism of fruit anatomy within the genus was revealed. The studied representatives can be grouped according to the greatest similarity in fruit structure:1) species with “hydrocyte parenchyma” complexes in the mesocarp and crystallifer-
ous cells similar to those of this zone (H. bonariensis, H. chamaemorus, H. umbellata,, H. vulgaris);
2) species with “hydrocyte parenchyma” complexes in the mesocarp and crystallifer-ous cells strikingly different from those of these zone, with horseshoe-shaped (H. mexicana , H. poepigii) or relatively evenly thickened (H. verticillata) cell walls;
3) species lacking “hydrocyte parenchyma”, with a few tanniferous cells in outer pericarp layers, forming a broken belt around the main massif of the mesocarp parenchyma (H. aconitifolia, H. bowlesioides, H. dissecta, H. medicaginoides, H. sibthorpioides).
4) species lacking “hydrocyte parenchyma”, but with a great number of tanniferous cells, forming an unbroken or discontinuous belt around the endocarp or adjacent compressed layers of mesocarp parenchyma (H. nepalensis, H. hookeri subsp. handelii, H. dulongenis).Trying to evaluate the correlation between these groups, determined on the basis of
fruit anatomy traits, and the geographic distribution of species they consist of, the fol-lowing can be noted: if Group 1 and Group 3 include both cosmopolitan species and

344 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
those from the Old and the New World, Group 2 is composed of the New World spe-cies (with horseshoe-shaped cell walls) and Group 4, on the contrary — of those from the Old World. It is possible, however, that if the number of species sampled grew, these tendencies would not be so obvious.
Besides, some species investigated are characterized by particularly interesting car-pological features, such as:• the formation of wings on the basis of the dorsal and intermediate ribs (H. medi
caginoides).• the presence of extremely large crystalliferous cells with unevenly thickened (with
no lignification) walls; these cells constitute a considerable part of pericarp thick-ness (H. aconitifolia);
• the presence of special groups of lignified fibers of mesocarpic origin, adhering to the crystalliferous layer from the outside (H. sibthorpioides).According to the data obtained, an important conclusion can be drawn stating that
fruit structure features in Hydrocotyle representatives are of great interest for taxono-mists studying this genus; therefore, a detailed carpological investigation of all species is needed.
Conclusions
Fruit structure traits, so meaningful for compiling the taxonomic system of Apiaceae in general, have been neglected in the construction of the Hydrocotyle generic system. The present research was undertaken to fill (at least partly) the gap in our knowledge of the carpology of both Araliaceae at large and the genus Hydrocotyle in particular. It was revealed that by the nature of micromorphological (anatomical) features of fruit structure, various Hydrocotyle species demonstrated a wide range of interesting pecu-liar traits connected with the size and shape of cells of the pericarp crystalliferous layer, presence or absence of special “hydrocyte parenchyma”, location of tannin-con-taining cells, etc. On the basis of these traits we so far were able to reveal only 4 pro-visory carpological groups within Hydrocotyle. Generally, we highly evaluated the taxonomic potential of fruit anatomy traits in Hydrocotyle for furnishing the generic system hoping that this research will stimulate a series of carpological studies dedi-cated specifically to the genus Hydrocotyle. We also hope that the investigation of carpological polymorphism in Hydrocotyle, stated in the present paper will help to elucidate its phylogenetic relationships with other representatives of Araliaceae, on the one hand, and Apiaceae subfamily Mackinlayoideae, on the other hand.
Acknowledgements
The authors would like to express their sincere gratitude to the late Prof. Vadim N. Tikhomirov (Department of Higher Plants, Moscow State University) who initiated and profoundly supervised this research at its early stages. We wholeheartedly thank Prof. Alexander P. Melikian (Department of Higher Plants, Moscow State University) — we could not have done this work without his active participation and valuable remarks. Special thanks to Dr. Tatyana V. Lavrova for the material provided for research.

A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle 345
References
Briquet, J. 1899: Recherches anatomiques et biologiques sur le fruits du genre Oenanthe. — Bull. Herb. Boiss. 7 (6): 467–488.
Chandler G.T., Plunkett G.M. 2004: Evolution in Apiales: nuclear and chloroplast markers together in (almost) perfect harmony. — Bot. J. Linn. Soc. 144: 123–147.
Constance L., Dillon M.O. 1990: A new peltate Hydrocotyle (Umbelliferae) from northern Peru. — Brittonia. 42 (4): 257–259.
Eichler, H. 1987a: Nomenclatural and bibliographical survey of Hydrocotyle L. (Apiaceae). Part. I. — Feddes Repert. 98 (1–2): 1–51.
Eichler, H. 1987b: Nomenclatural and bibliographical survey of Hydrocotyle L. (Apiaceae). Part. II. — Feddes Repert. 98 (3–4): 145–196.
Eichler, H. 1987c: Nomenclatural and bibliographical survey of Hydrocotyle L. (Apiaceae). Part. III. — Feddes Repert. 98 (5–6): 273–351.
Hylander, N. 1945: Nomenklatorische und systematische Studien über nordische Gefässpflanzen. — Uppsala Univ. Årsskr. 7: 5–337.
Klimochkina, L.V. 1950: O type structury ploda roda Hydrocotyle L. [On the type of structure of the fruit in the genus Hydrocotyle]. — Acta V.L. Komarov Bot. Inst. Acad. Sci. URSS. 7 (1): 219–227. [In Russian].
Kozo-Polyanski, B.M. 1916: O nekotorykh novykh osnovaniyakh dlya diagnostiki Umbelliferae [On a few new bases for the Umbelliferae diagnostics]. — Acta Fl. Ross. 2 (1): 1–12. [In Russian]
Mathias, M.E. 1936: The genus Hydrocotyle in northern South America. — Brittonia. 2: 201–237.Mathias, M.E. & Constance, L. 1951: Supplementary notes on South American Hydrocotyle. –Bull.
Torrey Bot. Club. 78 (4): 300–309.Mathias, M.E. & Constance, L. 1975: A re-assessment of the South American Umbellifer Hydrocoty
le filipes Mathias. — Bol. Soc. Argentina Bot. 16 (3): 249–254.Michaux, A. 1803: Flora boreali-americana 1. — Paris & Strasbourg.O’Brien, T.P. & McCully, M.E. 1981: The Study of Plant Structure: Principles and Selected Methods.
–Termarcarphi and Pty. Ltd., Melbourne, 356 p.Pérez-Moreau, R.A. 1948: Las especies argentinas del género “Centella” (L.) emend. Urban. — Lil-
loa. 17: 541–553.Pervukhina, N.V. 1950: O philogeneticheskom znachenii nekotorykh priznakov stroenya ploda zon-
tichnykh. [On phylogenetic value of some characters of fruit structure of Umbelliferae]. –Acta V.L. Komarov Bot. Inst. Acad. Sci. URSS. 7: 82–120. [In Russian].
Pimenov, M.G. & Leonov, M.V. 1993: The genera of the Umbelliferae. — Royal Bot. Gardens & Moscow: Bot. Garden of MSU, Kew, 164 p.
Plunkett, G.M. & Lowry II, P.P. 2001: Relationships among “Ancient Araliads” and their significance for the systematics of Apiales. — Mol. Phyl. Evol. 19 (2): 259–276.
Plunkett, G.M., Wen, J. & Lowry II, P.P. 2004a: Infrafamilial classifications and characters in Aralia-ceae: Insights from the phylogenetic analysis of nuclear (ITS) and plastid (trnL-trnF) sequence data. — Pl. Syst. Evol. 245: 1–39.
Plunkett, G.M., Chandler, G.T., Lowry II, P.P., Pinney, S.M. & Sprenkle, T.S. 2004b: Recent advances in inderstanding Apiales and a revised classification. — S. Afr. J. Bot. 70 (3): 371–381.
Richard, A. 1820: Monographie du genre Hydrocotyle de la famille des Ombellifères. — Ann. Gén. Sci. Phys. 4: 145–224.
Prozina, M.N. 1960: Botanicheskaya mikrotekhnika. [Botanical microtechnique]. — Moscow: Vys-shaya shkola. [In Russian].
Ruiz, A.N. & Pavon, J. 1802: Flora peruviana et chilensis 3. — Madrid.Shan, R.H. 1936: Studies on Umbelliferae of China I. (Hydrocotyloideae, Saniculoideae). — Sinensia
7: 477–489.Shan, R.H. & Liou, S.L. 1964: On the Chinese species of Hydrocotyle Linn. — Acta Phytotaxon. Sin.
9 (2): 119–134.Shibakina, G.V. 1984: Kostyanka kak ekologicheskyi tip ploda I nekotorye voprosy terminologii pri

346 A.I. Konstantinova & E.Yu. Yembaturova, Structural traits of Hydrocotyle
opisanii plodov v semeistve Araliaceae [Drupe as an ecological fruit type and some questions on terminology used for describing fruits in the family Araliaceae] . — Bot. Journ. 69 (8): 1076–1083. [In Russian].
Tamamshyan, S.G. & Vinogradova, V.M. 1969: K systematike roda Grammosciadium DC. (Um-belliferae) [To the systematics of the genus Grammosciadium DC. (Umbelliferae)]. Journ. Bot. (Leningrad). 54 (8): 1197–1212. [In Russian].
Tseng, C.C. 1967: Anatomical studies of flower and fruit in the Hydrocotyloideae (Umbelliferae). — Univ. Calif. Publ. Bot. 42: 1–59.
Webb, C. J., Johnson P. N. 1982: Hydrocotyle (Umbelliferae) in New Zealand: the 3-foliate species. — New Zeal. J. Bot. 20 (2): 163–168.
Willis, J.C. 1988: A Dictionary of the Flowering Plants and Ferns. 8th ed. revised by H.K. Airy Shaw — Cambridge, 1294 p.
Addresses of the authors: Dr. Alexandra Konstantinova, Department of Higher Plants, Biological Faculty, M.V.Lomonosov
Moscow State University, 119992, Moscow, Russian Federation; e-mail: [email protected]; Dr. Elena Yu. Yembaturova Russian State Agrarian University — K.A. Timiryazev MSKHA, Depart-ment of Botany, ul. Timiryazevskaya, 49 127550 Moscow, Russian Federation; e-mail: [email protected]



























![Leste do brasil [eastern brasil] - araliaceae](https://img.pdfslide.us/doc/110x75/55a7a2461a28ab3f438b48cd/leste-do-brasil-eastern-brasil-araliaceae.jpg)

