Embed Size (px)
Citation preview

1
Structural basis for outer membrane sugar uptake in pseudomonads
Bert van den Berg
UMass Medical School, Program in Molecular Medicine, Worcester MA 01605
e-mail: [email protected]
Keywords: OprB, outer membrane channel, sugar transport, crystal structure, pseudomonads.
Background: OprB channels mediate outer membrane carbohydrate uptake in pseudomonads. Results: X-ray crystal structures of Pseudomonas putida F1 OprB have been determined. Conclusions: OprB forms a 16-stranded, monomeric β-barrel with a constriction formed by loops L2 and L3. OprB lacks a greasy slide for substrate binding.
Significance: The structure explains the substrate specificity of OprB for monosaccharides.
http://www.jbc.org/cgi/doi/10.1074/jbc.M112.408518The latest version is at JBC Papers in Press. Published on October 12, 2012 as Manuscript M112.408518
Copyright 2012 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

2
SUMMARY Substrate-specific outer membrane (OM) channels of Gram-negative bacteria mediate uptake of many small molecules, including carbohydrates. The mechanism of sugar uptake by enterobacterial channels such as E. coli LamB (maltoporin) has been characterized in great detail. In pseudomonads and related organisms, sugar uptake is not mediated by LamB, but by OprB channels. Beyond the notion that OprB channels seem to prefer monosaccharides as substrates, very little is known about OprB-mediated sugar uptake. Here I report the X-ray crystal structure of an OprB channel from Pseudomonas putida F1. The structure shows that OprB forms a monomeric, 16-stranded β-barrel with a constriction formed by extracellular loops L2 and L3. The side chains of two highly conserved arginine residues (Arg83 and Arg110) and a conserved glutamate (Glu106) line the channel constriction and interact with a bound glucose molecule. Liposome swelling uptake assays show a strong preference for monosaccharide transport over disaccharides. Moreover, substrates with a net negative charge are disfavored by the channel, likely due to the negatively charged character of the constriction. The architecture of the eyelet and the absence of a greasy slide provide an explanation for the observed specificity of OprB for monosaccharides rather than the oligosaccharides preferred by LamB and related enterobacterial channels. Gram-negative bacteria have substrate-specific channels in the OM for the selective uptake of solutes from the external environment (1). A number of such channels have been studied in detail by genetic, biochemical and structural approaches. This has led to the elucidation of substrate uptake mechanisms for e.g. FadL-mediated uptake of long-chain fatty acids in E. coli (2) and for
phosphate uptake by P. aeruginosa OprP (3). The best-studied OM uptake channels are the carbohydrate-selective channels LamB and the closely related sucrose channel ScrY that are found within Enterobacteriaceae. Genetic and biochemical analyses of LamB identified the involvement of some of the extracellular loops in binding and orientation of the substrate (4,5). Subsequent crystallographic and mutagenesis studies clarified the actual transport step through the constriction, or "eyelet", of the channel. Important roles were discovered for the "greasy slide", a striking, helical row of aromatic residues protruding from the wall of the barrel and lining the channel (6,7). The aromatic rings of the greasy slide interact via van der Waals interactions with the glycosyl moieties of the substrate. In addition, hydrogen bonds between ionizable residues and substrate hydroxyl groups are found to be important for substrate translocation (8). In common with the general, large channel porins, most of these charged residues are located within the extracellular loop L3 that folds inwards and constricts the channel. The architecture of the LamB channel explains the pronounced preference of the Enterobacterial sugar channels for oligosaccharides over monosaccharides. Pseudomonads and related soil bacteria are characterized by a lack of large-channel porins in the OM. Instead of porins, a large number (~ 30) of substrate-specific channels mediates uptake of nutrients and ions (9,10). Despite their metabolic versatility, Pseudomonads have only a limited ability to metabolize sugars (11). The exception is glucose metabolism, which is biochemically diverse (12). OprB is an OM channel that is part of a high-affinity uptake system for glucose that was first described for Pseudomonas aeruginosa by Hancock et al. (13). Subsequent in vitro and in vivo studies showed a substrate preference of OprB for monosaccharides (14-16), which is very
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

3
different from LamB and related channels. Beyond the substrate specificity, however, not much is known about OprB, mainly due to a lack of structural information. Here we present two crystal structures of P. putida OprB, one at low pH and one at high pH in the presence of glucose. The high pH structure shows that OprB forms a monomeric 16-stranded β-barrel with a novel pore constriction involving loops L2 and L3. The absence of a greasy slide and the architecture of the eyelet provide an explanation for the preference of OprB channels for monosaccharide substrates. EXPERIMENTAL PROCEDURES Cloning, Expression and Purification of proteins. The mature parts of OprB and OprP proteins were amplified from genomic DNA and cloned into a modified E. coli expression vector pB22 (17,18) that expresses OM proteins as a fusion with the signal sequence of the E. coli YtfM OM protein. Upon expression and signal peptide cleavage, the proteins have the following N-terminal extension: ANVRLQHHHHHHHLE. For protein expression, BL21(DE3) T1 phage-resistant cells (New England Biolabs, Ipswich, MA) were transformed with pB22 constructs. The cells were grown to OD600 ~ 0.5 at 37ºC and then induced with 0.1% arabinose at 20°C for ~16 h. Cells were harvested by centrifugation at 4200 rpm for 12 min (Beckman Coulter, J6-MC). Cell pellets were suspended in TSB (20 mM Tris, 300 mM NaCl, pH 8.0) and cells were lysed in a microfluidizer (Emulsiflex C-3; Avestin). Cell lysates were centrifuged at 42,000 rpm for 45 min (45Ti rotor; Beckman). Total membrane pellets were solubilized in 0.5% lauryl-sarcosine for 30 min at room temperature with stirring, to extract inner membrane proteins. Upon centrifugation for 30 min at 42,000 rpm the pellets were extracted with 2-3% (w/v) of Elugent (Calbiochem) by stirring at 4 ºC for 1 hr. The extract was re-centrifuged at 42,000 rpm for 30 min and the supernatant was
applied to a 10 ml nickel column. The column was washed with 10-15 column volumes (CV) TSB containing 0.2% b-decyl-maltopyranoside (DM) and 20 mM imidazole. Proteins were eluted with 3 CV TSB containing 0.2% DM and 200 mM imidazole. Purification was done by gel filtration chromatography in 10 mM Tris/50-100 mM LiCl or NaCl/0.12% DM pH 8. For crystallization studies, this column was followed by a second gel filtration column in 10 mM Tris/100 mM LiCl or NaCl/0.3-0.4% C8E4 pH 8. The purified protein was concentrated to ~10 mg/ml using 50 kDa molecular weight cutoff filters (Amicon) and directly flash-frozen in liquid nitrogen. Seleno-methionine (SeMet) substituted protein was produced in BL21(DE3) T1 phage-resistant cells by inhibition of the methionine biosynthesis pathway (19), and purified as described above without using EDTA or reducing agents. Methionine mutants of F1OprB-1 were obtained using the QuikChange mutagenesis kit (Stratagene). Two mutants were made: one with two extra methionines (L220M/V222M) for a total of four (4Met). The other mutant contained an additional two methionine residues (L415M/I417M) to give a total of six (6Met). Crystallization of OprB and structure solution. Before crystallization, 50 mM D-Glucose in crystallization buffer was added to the protein from a 0.5 M stock solution and incubated for 1 hr on ice prior to setting up trays. Concentrations of glucose higher than 50 mM were also tried, but these were detrimental for obtaining crystal form 2. For both crystal forms, initial crystallization trials were performed at 293 K using a Gryphon crystallization robot (Art Robbins Instruments) and by sitting-drop vapor diffusion, using various in-house screens. The initial hits were optimized by fine-screening with larger drops using hanging drop vapor diffusion. Crystal form 1 was obtained from the 6Met mutant, and grew as large plates
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

4
from 50 mM Na-citrate/0.2 M Na-formate/35-40% PEG400 pH 4. The space group of this crystal form is I222, and there is one molecule within the asymmetric unit (Vm = 3.5; 65% solvent). Crystal form 2 was obtained from the 4Met mutant, using 50 mM Tris-HCl/0.2 M KH2PO4/10% PEG 4K pH 8.5. The space group of these crystals is P3121, with one molecule of OprB within the asymmetric unit (Vm = 4.3; 71% solvent). For phasing, SAD data sets at the selenium peak were collected for both crystal forms at beamline X6A at the National Synchrotron Light Source at Brookhaven National Laboratory. Data were indexed and scaled with HKL2000 (20). Initial phasing and automatic model building was done by AUTOSOL (21) within PHENIX (22), using the data for crystal form 2 (4Met). This resulted in ~50% of the residues being built. Four (out of four) selenium atoms were located in the asymmetric unit. Further model building was performed manually using the program COOT (23). The protein model was refined with PHENIX. Statistics for the collected data and refinement are described in Table 1. One Ramachandran outlier is present in the refined 4Met OprB structure (Ala3, with Phe4 and Asp73 is generously allowed regions). For the less well-ordered 6Met structure, two residues (Phe126, Asn144) are within the disallowed region of the Ramachandran plot (Glu8, Trp71, Trp171 and Glu196 are in generously allowed regions). Liposome Swelling Assays. Osmotically active liposomes were prepared as described previously (24). Briefly, 5 mg of egg phosphatidylcholine (Avanti Polar Lipids, Alabaster, Al) and 2 mg of dicetylphosphate (Sigma, St. Louis, Mo) in chloroform were dried under a nitrogen stream, followed by drying overnight in an evacuated dessicator. The dried lipid film was resuspended in 0.2 ml water after which 20-30 µg of C8E4-purified OprB was added (2 µl or less in volume). The suspension was then sonicated in a bath sonicator for 15-20 mins (Model G112SPIT,
Laboratory Supplies Company, Hicksville, NY). The liposomes were dried overnight in an evacuated dessicator. The dried film was gently resuspended in 0.4 ml of 10 mM HEPES, pH 7.0 containing 12 mM stachyose (Sigma). After standing at room temperature for 2 hrs, the liposomes were used in the assay. Control liposomes were prepared by addition of buffer without protein. For the uptake assays, 7-8 µl of the proteoliposomes were added to 100 µl of substrate solutions (7-15 mM) in 10 mM HEPES, pH 7.0. Liposome swelling was measured by monitoring changes in optical density at 400 nm at 17 s intervals for 600 s (TECAN, Safire, Männedorf, Switzerland). The substrate concentration iso-osmotic to the intra-liposomal milieu was determined by identifying the concentration of substrate that caused a change in OD400 of at most ± 0.5% upon dilution in control liposomes. Readings (4-6) were averaged and corrected for the average absorbance change in the control liposomes, and were normalized to the optical density at time zero for each time course. For testing the uptake activity of reduced OprB, the detergent-purified protein (pH 8) was first reduced in 5 mM DTT for 30 min at room temperature prior to reconstitution. All solutions used subsequently contained 0.5 mM DTT in order to keep the protein reduced. RESULTS Structure determination of OprB. The project was initiated starting with OprB from P. aeruginosa (PaOprB). There are two annotated orthologs in this organism, PA3186 and PA2291 (http://www.pseudomonas.com). These proteins differ only in one residue, and can therefore be considered identical. We obtained several crystal forms of PaOprB, but after optimization these did not diffract well enough for structure determination. We next switched to the biodegrader strain P. putida F1. This organism has two OprB orthologs, one with 80% identity to PaOprB (Pput 1057;
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

5
F1OprB-1) and another that is 68% identical to PaOprB (Pput 4276; F1OprB-2). Both OprB proteins expressed well in the OM of E. coli and both yielded a number of crystal forms. Since those of F1OprB-1 were the most promising, we focused on this ortholog. F1OprB-1 has only two methionines in a total of 421 residues (Mw ~47 kDa), and we therefore constructed two mutant proteins with additional methionine residues to facilitate structure determination by selenium-SAD. The methionines were introduced in predicted trans-membrane strands (25) to replace outward-facing hydrophobic residues that would be exposed to the OM. The first mutant protein has two additional methionine residues in predicted β-strand S8 (L220M/V222M) for a total of four (4Met). The second mutant protein has another two methionines within the C-terminal β-strand (L415M/I417M), for a total of six (6Met). After optimization of initial crystal hits for both proteins, two crystal forms were obtained that diffracted to 3.5-4 Å on a home source. The first crystal form obtained originated from the 6Met mutant. These crystals grew as large plates from 50 mM Na-citrate/0.2 M Na-formate/35-40% PEG400 pH 4 (crystal form 1). The second crystal form was obtained from the 4Met mutant. Interestingly, the crystallization conditions are completely different from those of the 6Met protein, with small diamond-shaped blocks growing from 50 mM Tris-HCl/0.2 M KH2PO4/10% PEG 4K pH 8.5 (crystal form 2). Selenium-SAD data at the selenium peak were collected for both crystal forms. As the resolution of the data was better for crystal form 2 (2.7 Å compared to 3.1 Å for crystal form 1) the F1OprB-1 structure was solved using crystal form 2. The final model has an R/Rfree of 18.8/23.9 % (see Experimental Procedures and Table 1). In the following we discuss the high-pH structure of OprB. The low pH structure, showing substantial disorder in the extracellular loops, is discussed in the Supplementary Materials.
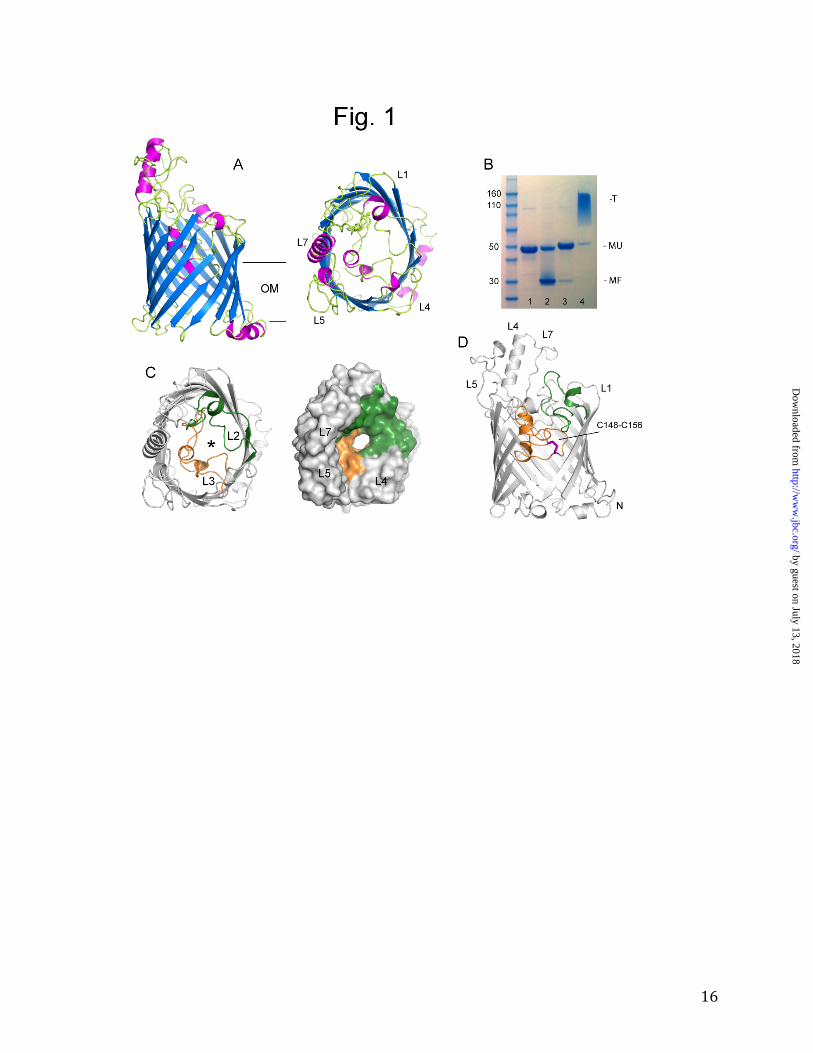
OprB forms a monomeric 16-stranded β-barrel. The available literature for OprB suggests that it forms a trimer, analogous to LamB and the general porins of E. coli (14-16, 26). These notions were not based on actual data, however. The crystal structure demonstrates that OprB in fact forms a monomer (Fig. 1A and Supplementary Fig. 1). The monomeric character of the protein is also evident from the behavior on SDS-PAGE (Fig. 1B, lanes 1 and 2). Without boiling (lane 2), most of the protein migrates at a lower apparent molecular weight (~30 kDa), indicative of a monomeric, folded structure. This behavior is consistent with that reported previously for several other OprB orthologs (14-16, 26). The minor fraction migrating at the predicted molecular weight (~47 kDa) likely means that the protein slowly unfolds at room temperature in the SDS sample buffer. Upon boiling (lane 1), all of the OprB unfolds and migrates at ~47 kDa, i.e. OprB is heat modifiable. By contrast, trimeric OM proteins such as P. aeruginosa OprP (3) and the general E. coli porins OmpF/C all migrate as trimers without boiling in SDS (Fig. 1B, lanes 3 and 4). Surprisingly, the OprB barrel consists of 16 β-strands, which differs from the 18 strands predicted by e.g. PRED-TMBB (25). Density is visible for the entire protein (Glu2-Phe421; numbering for the mature protein), with the exception of the N-terminal alanine residue and the N-terminal extension that includes the histidine tag (see Experimental Procedures). Several of the extracellular loops are quite long, most notably L5 and L7, with the latter containing an approximately 15-residue-long α-helix. Because of these two loops, the extracellular part of the protein is asymmetric, with a wall extending from the transmembrane part on only one side (Fig. 1A). The pore restriction ("eyelet") of the OprB channel is formed by the long inward-folded loops L2 and L3 (Fig. 1C). Loop L3 (residues 136-175) contributes to the eyelets
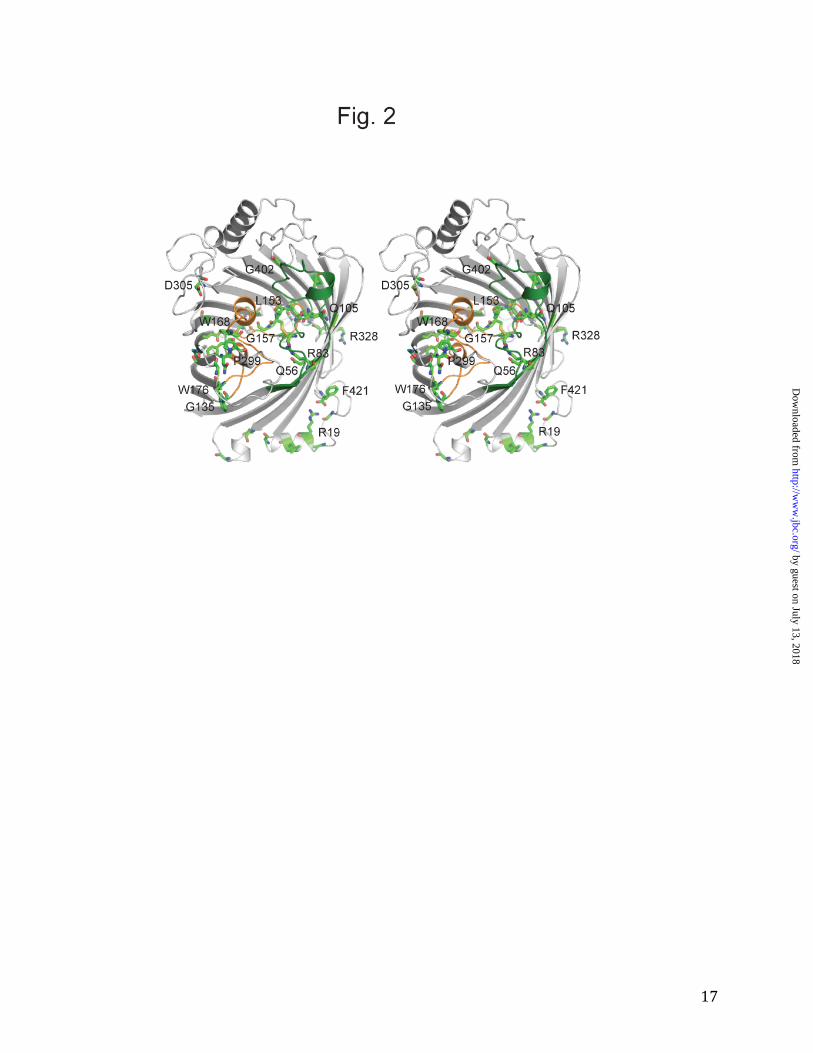
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

6
of all known 16- and 18-stranded β-barrels, albeit in different ways compared to OprB. By contrast, the involvement of loop L2 (residues 81-116) in forming the eyelet is unique to OprB. The importance of loops L2 and L3 for channel function is clear from the large number of conserved residues stabilizing their conformations (Fig. 2 and Table 2). Interestingly, loop L2 is important for trimer stability in almost all trimeric OM channels (27,28), as it folds outwards and interacts extensively with a neighboring monomer. Thus, the conformation of L2 in OprB likely contributes to the monomeric character of this channel. However, since many residues outside of the L2 loop in porins form interface contacts (29) it is unlikely that the observed differences in oligomeric state are caused solely by differences in loop L2. The OprB pore is located approximately in the center of the protein and has an oval shape with dimensions of ~9 Å x 7 Å (atom-center to atom-center; hydrogens excluded). An interesting feature in the OprB structure is the disulfide bond formed by the two cysteine residues in the protein, Cys148 and Cys156 (Fig. 1D). This disulfide bond constrains the end of loop L3 and lines the pore. The fact that the cysteine residues are absolutely conserved in OprB orthologs (Fig. 2 and Supplementary Fig. 2) suggests that the disulfide could be important for transport. However, given the very similar substrate uptake mediated in liposome swelling experiments by reduced and oxidized OprB (Fig. 3), the data suggest that the disulfide may not be important for transport, but could play a role in proper folding and/or OM insertion of the channel. In terms of the global fold of OprB, a DALI search (30) identifies the phosphate channel OprP from P. aeruginosa as the closest structural homolog of OprB, with a Cα r.m.s.d of ~3 Å over approximately 300 residues (Z = 22.5). Slightly lower Z-scores are obtained for several general porins, such
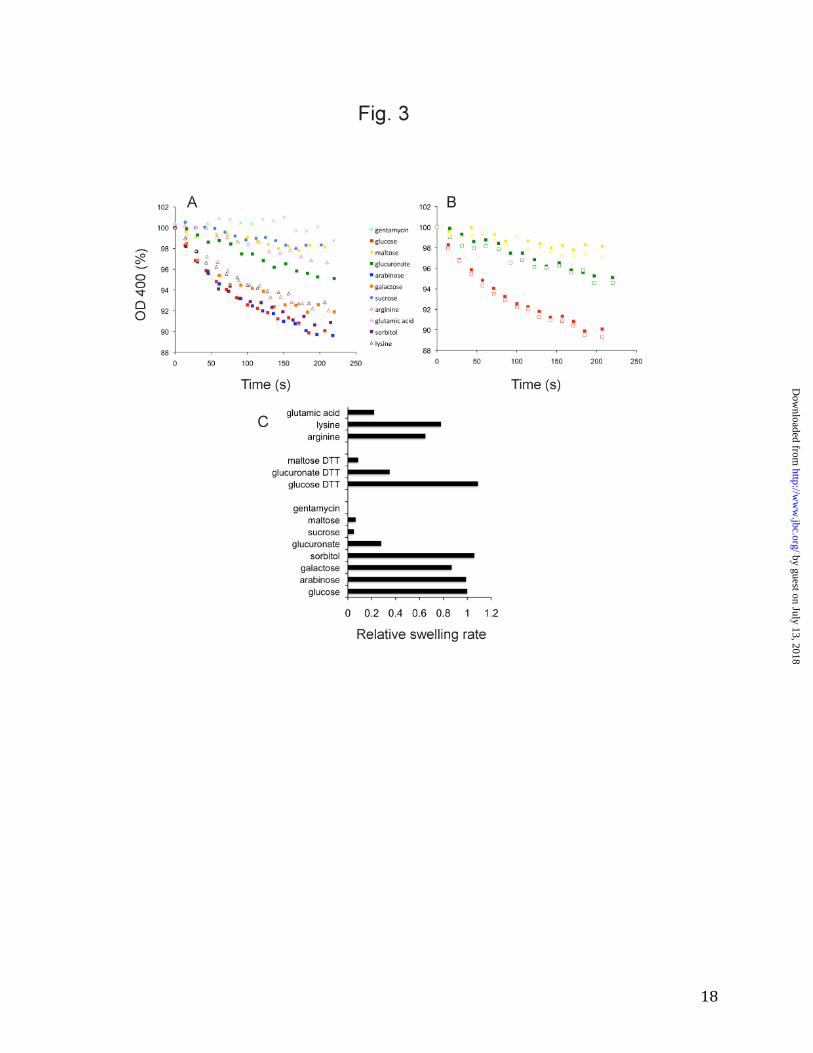
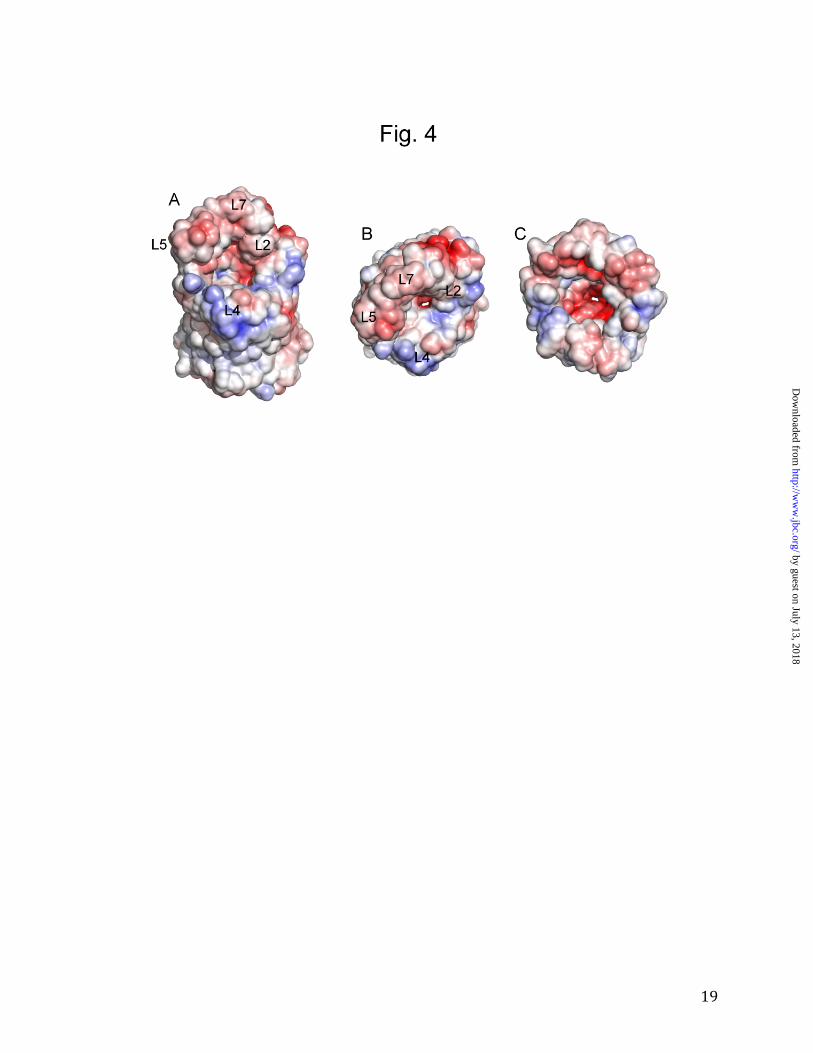
as E. coli OmpF and OmpC. Thus, despite sequence identities that are only 10-13%, OprB is a monomeric member of a large superfamily of trimeric 16-stranded β-barrels that includes the general porins of Enterobacteriaceae and P. aeruginosa OprP. OprB does not have a greasy slide for sugar binding. Previously, the OprB protein from P. aeruginosa was shown to be selective for a number of small sugars such as glucose, fructose, mannitol and glycerol, with glucose being a preferred substrate (13,15). Similar results were obtained for a P. putida OprB ortholog (14). We performed liposome swelling experiments for purified F1OprB-1 and obtained transport profiles that confirmed a preference for glucose and other monosaccharides (Fig. 3) and very low transport activities for disaccharides such as sucrose and maltose. Based on these results, all crystallization experiments with OprB were performed in the presence of 50 mM D-glucose. OprB did not mediate uptake of gentamycin, an aminoglycoside antibiotic containing three sugar molecules. Interestingly, despite the high concentration gradients in the assay (mM), the uptake of the carboxylated sugar glucuronate is ~ 4-fold slower than that of glucose, indicating that negatively charged substrates are poor substrates for OprB. In order to determine whether OprB can transport other small molecules besides sugars, we also tested uptake of lysine, arginine and glutamic acid (Fig. 3). The data show that OprB, at least in the presence of high substrate concentrations, also transports non-sugar substrates with a molecular weight similar to monosaccharides. Glutamic acid is a poor substrate, again demonstrating that the OprB channel is refractory to negatively charged substrates. These data are in excellent agreement with the electrostatic properties of the channel, which clearly show that the channel mouth is predominantly negatively charged (Fig. 4).
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

7
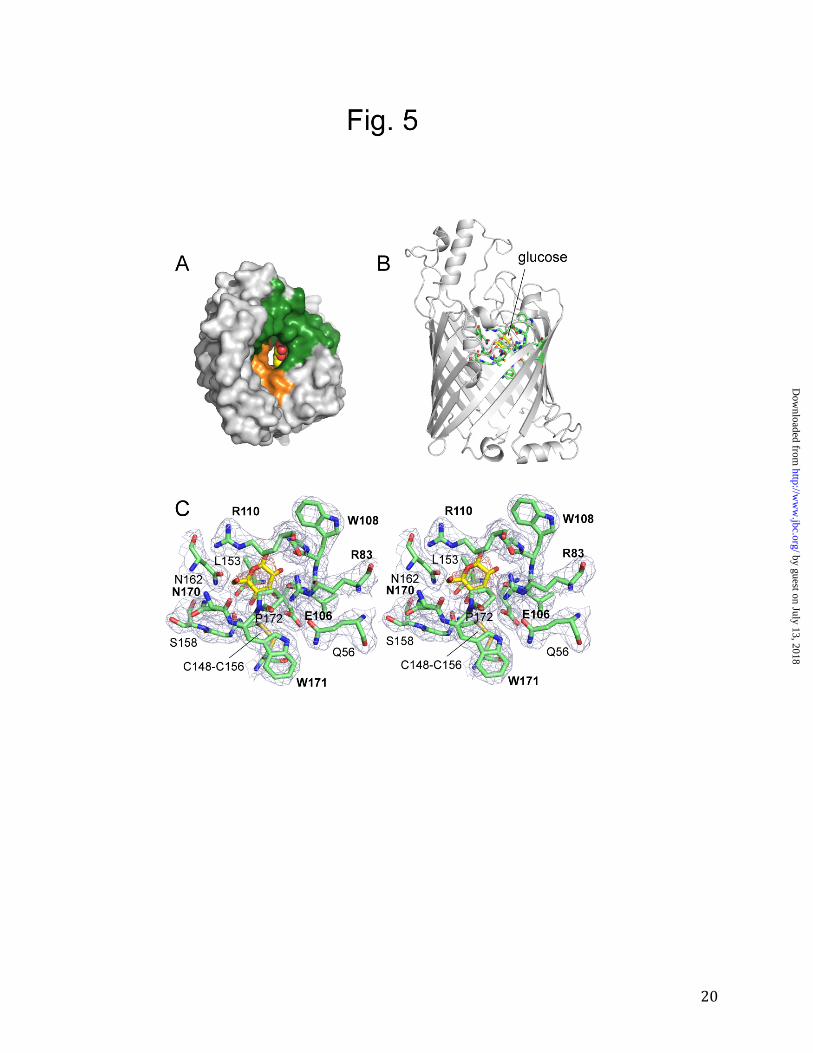
Inspection of the eyelet of the channel reveals density that is incompatible with the other small-molecule components of the crystallization solution, i.e. phosphate and Tris buffer. The density therefore corresponds most likely to a bound glucose molecule (Supplementary Fig. 3 and Fig. 5). The fact that the density is not very strong may be due to several factors. First, the resolution of the data is fairly limited (2.7 Å). Second, anomerization could have resulted in a mixed population of α- and β-D-glucose molecules that might be bound to the channel in a slightly different way. Finally, it is possible that the substrate affinity is sufficiently low for the glucose to have a partial occupancy in the channel even at 50 mM concentration. A previously reported half-saturation constant of 110 mM for glucose binding to P. putida (ATCC12633) OprB would be consistent with this possibility (14). In any case, additional support for the assignment of the channel density as glucose is provided by its similarity to the density of some of the glucose moieties of the β-decylmaltoside (DM) detergent molecules that are bound to the protein (Supplementary Fig. 4). The substrate in the channel is contacted by the residues Arg83, Glu106, Trp108, Gly109, Arg110, Asn170 and Trp171. Of these residues, Trp108 and Trp171 interact with the substrate only with their backbone atoms (Fig. 5C). This is a striking observation, since side chains of aromatic residues play crucial roles in substrate binding in other OM sugar channels (see Discussion). In fact, despite the presence of 57 aromatic residues (13.5% of the total), Trp171 is the only aromatic side chain that lines the channel and is positioned favorably to interact with passing substrate molecules. Thus, OprB does not have a greasy slide for sugar transport, and polar contacts between the channel and substrate dominate. These polar contacts consist of hydrogen bonds between the substrate and several side chains of OprB, i.e.
two arginine residues (Arg83 and Arg110), a glutamate (Glu106), and possibly an asparagine residue (Asn170). The guanidinium group of Arg83 faces the substrate at an angle almost perpendicular to the plane of the glucose ring and is therefore likely to be very important. This notion is supported by the fact that this residue is absolutely conserved in OprB orthologs (Fig. 2 and Supplementary Fig. 2). Glu106 is also likely to be important, with hydrogen bonds mediated both by the carbonyl oxygen and by the carboxyl side chain to the C-2 and C-3 hydroxyl groups of glucose. Accordingly, Glu106 is also highly conserved (Supplementary Fig. 2). Arg110 may be less important for substrate binding since its guanidinium group faces away from the substrate (Fig. 5C) and this residue is not conserved in more distant OprB orthologs (Supplementary Fig. 2). Besides these ionizable residues, another interesting eyelet residue is Leu153, the side chain of which points towards the substrate but does not contact it directly. Leu153 is also absolutely conserved (Fig. 2), and we propose that it may orient the substrate in a way that is productive for transport. A similar role may be performed by Pro172, which is located on the other side of the substrate relative to Leu153 (Fig. 5C). DISCUSSION Comparison of OprB with OprP. The phosphate channel OprP of P. aeruginosa is the closest structural homolog of OprB. The most striking difference between the two channels concerns the oligomeric state, with OprB being a monomer and OprP a trimer. In OprP, the N-termini of the monomers form an intricate "tricorn" motif that is likely to be a major factor stabilizing the trimer (3). Since the N-terminus of our OprB preparations contains an extension with a hepta-histidine tag, it is possible that this modification would prevent trimer formation. To investigate this possibility we expressed OprP in E. coli with
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

8
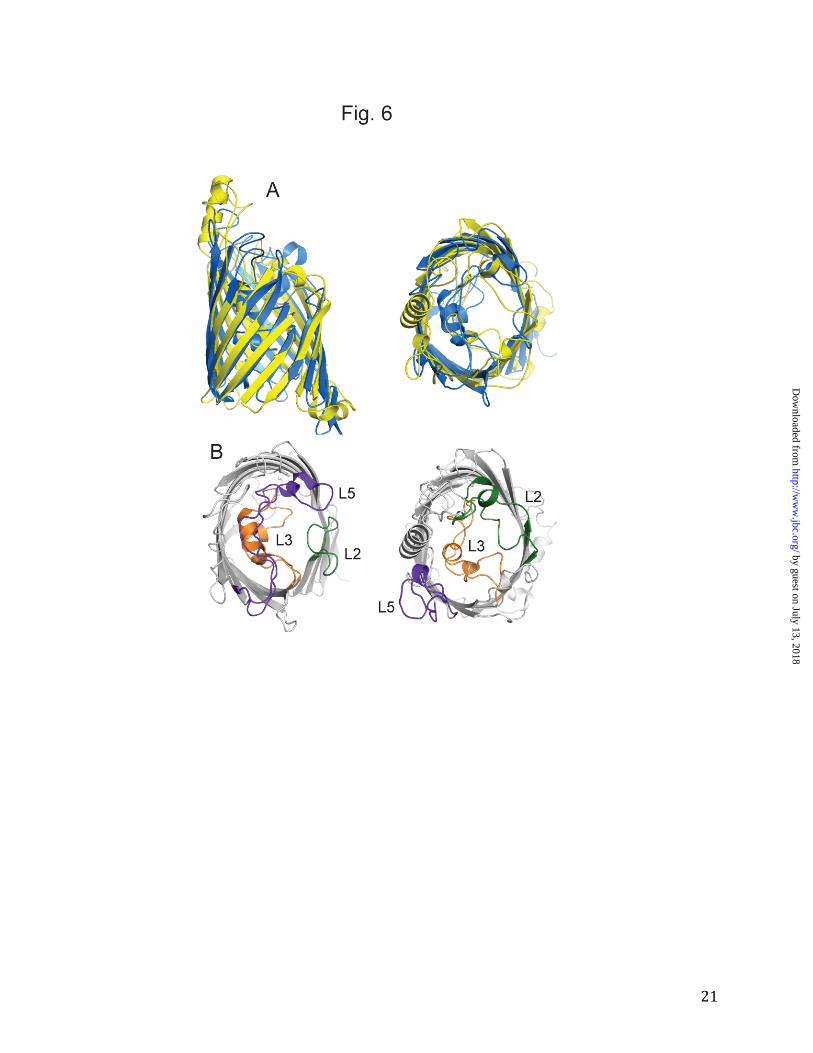
an identical N-terminal extension as OprB, including a histidine tag. Upon purification we find that OprP still forms trimers (Fig. 1B), demonstrating that (i) the E. coli OM protein integration machinery is capable of assembling trimers of P. aeruginosa OM proteins, and (ii) the N-terminal extension does not prevent trimer formation. We conclude from these data that OprB is likely to function as a monomeric channel. Our results are consistent with the recent characterization of an OprB ortholog purified from P. putida CSV86 (26). Here, the authors report a molecular mass of ~130 kDa for OprB using gel filtration chromatography, which they interpret as a trimer. However, taking into account that the protein was purified and analyzed in Triton X-100 which forms large micelles of ~90 kDa, these results in fact suggest that OprB is a monomer. The monomeric character of OprB is significant, since all other known 16-stranded β-barrels form trimers. A more detailed comparison between OprB and OprP shows that the trans-membrane parts of the barrels are very similar (Fig. 6), which is consistent with the DALI alignment of ~300 residues (~70% of the total sequence). The extracellular loops, on the other hand, are completely different between both channels. This can be illustrated by the architecture of the eyelet. In OprP, loop L2 is short and does not fold back into the channel. By contrast, the L2 loop in OprB is much longer and part of the eyelet. L3 has an important role in both proteins, and indeed in all known 16- and 18-stranded β-barrels, in constricting the channel. Finally, loop L5 has a remarkable structure in OprP, where it is very long and crosses the entire barrel, forming the entrance of the phosphate channel. In OprB, L5 is long as well, but it does not fold back inside the barrel (Fig. 6). It could be argued that this particular conformation may be a consequence of crystal packing contacts. While this may be the case,
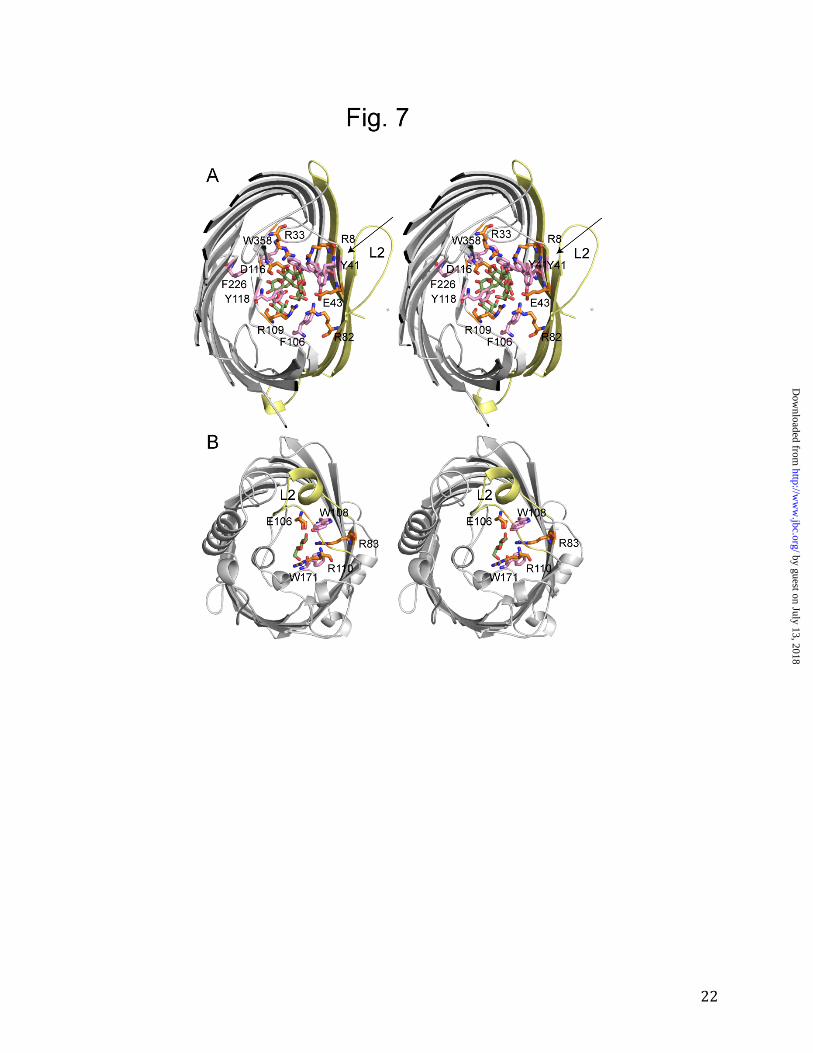
the distinct lack of conserved residues within L5 (Fig. 2) suggests that this loop does not play a role in channel formation in OprB. The OprB structure explains the preference of pseudomonads for monosaccharides. An interesting property of pseudomonads is that while they can utilize many compounds for growth, most of these have molecular masses smaller than ~200 Da. This preference can be partly explained by the absence of non-specific, large channel porins in pseudomonads. However, the preference for small growth substrates extends to sugars as well, as shown by the low uptake activities of F1OprB-1 for disaccharides (Fig. 3). How can this be reconciled with the fact that sugar channels from Enterobacteria (e.g. E. coli LamB) prefer oligosaccharides (31-33)? Using macroscopic conductance experiments and purified OprB, it was shown previously that the binding constants for glucose are approximately 100 mM both for LamB and P.putida (ATCC 12633) OprB (14,34). For LamB, maltose-oligosaccharides of increasing chain length bind progressively better to the channel, with the tightest binding observed for maltopentaose (~0.1 mM; 34). By contrast, for P. putida OprB the highest binding affinity is observed for maltose (~30 mM; 14). With longer carbohydrates binding becomes worse, and for compounds longer than three glucose units (maltotriose), binding is not measurable. From these data, Hancock and colleagues predicted that the binding site in OprB would be much shorter than that in LamB and related channels (14). A comparison of the OprB and the LamB structures reveals that this prediction was correct (Fig. 7). In LamB, malto-oligosaccharides bind via van der Waals interactions to the greasy slide, an extended binding site formed by a row of aromatic residues that protrude from the barrel wall and line the channel (Tyr41, Tyr6, Trp430, Trp358 and Phe226; Fig. 7A) (6). The interactions between the greasy slide and the substrates are made possible because of the concave shape
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

9
of the 18-stranded LamB β-barrel, bringing strands S1, S2 and S18 and their greasy slide residues Tyr6, Tyr41 and Trp420 in close proximity to the substrate (Fig. 7A). Besides the aromatics in the greasy slide, two additional aromatic residues in loop L3 (Phe106, Tyr118) provide potential interactions with the substrates. By contrast, in OprB there is a striking lack of aromatic residues lining the channel (Fig. 5 and Fig. 7B). Trp108 and Trp171 are the only two aromatics that could potentially interact with substrates, and they are not in close proximity to each other. Moreover, even if the wall of the channel would contain aromatic residues (as in LamB), the more circular shape of the OprB barrel would ensure that these would not be close to the substrate in the center of the channel. Thus, the shape of the barrel and the lack of aromatic residues in OprB result in a very different carbohydrate binding site compared to LamB and the sucrose channel ScrY (35). The shape of the barrel may also be a major contributing factor as to why OprB is a monomer. In LamB (as well as in OmpF/C), the large trimer interface is made possible by the shape of the barrel (Fig. 7A). A more
circular shaped barrel, as for OprB, would limit the extent of a trimer interface. In addition to substrate gliding over the aromatic residues of the greasy slide, hydrogen bonds between the hydroxyl groups of the substrate and ionizable residues of the channel are important for substrate transport in LamB ("polar track"; 8). Two arginine residues and one acidic residue interact with each of the two glucose molecules that are present within the eyelet of the LamB-maltohexaose structure (Fig. 7A). A very similar situation is encountered in OprB, with Arg83, Arg110 and Glu106 forming hydrogen bonds with the substrate (Fig. 5). However, there is only one set of ionizable residues in OprB, i.e. this channel has a polar patch rather than the polar track present within LamB. Together, the structural differences between the eyelets of OprB and LamB clearly explain their preferences for mono- and oligosaccharides, respectively. Interestingly, there are a number of Enterobacteriaceae (e.g. Klebsiella pneumoniae) that have both LamB and OprB orthologs in their genomes, a feature that presumably ensures the efficient uptake of both small and large sugar molecules.
REFERENCES
1. Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. (2003) Microbiol Mol Biol Rev. 67, 593-656. 2. Hearn E.M., Patel D.R., Lepore B.W., Indic M., and van den Berg B. (2009) Transmembrane passage of hydrophobic compounds through a protein channel wall. Nature 458, 367-70. 3. Moraes T.F., Bains M., Hancock R.E., and Strynadka N.C. (2007) An arginine ladder in OprP mediates phosphate-specific transfer across the outer membrane. Nat Struct Mol Biol. 14, 85-87. 4. Klebba P.E., Hofnung M., and Charbit A. (1994) A model of maltodextrin transport through the sugar-specific porin, LamB, based on deletion analysis. EMBO J. 13, 4670-4675. 5. Klebba P.E., Newton S.M., Charbit A., Michel V., Perrin D., and Hofnung M. (1997) Further genetic analysis of the C-terminal external loop region in Escherichia coli maltoporin. Res Microbiol. 148, 375-387.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

10
6. Schirmer T., Keller T.A., Wang Y.F., and Rosenbusch J.P. (1995) Structural basis for sugar translocation through maltoporin channels at 3.1 A resolution. Science 267, 512-514. 7. Dutzler R., Wang Y.F., Rizkallah P., Rosenbusch J.P., and Schirmer T. (1996) Crystal structures of various maltooligosaccharides bound to maltoporin reveal a specific sugar translocation pathway. Structure 4, 127-134. 8. Dumas F., Koebnik R., Winterhalter M., and Van Gelder P. (2000) Sugar transport through maltoporin of Escherichia coli. Role of polar tracks. J Biol Chem. 275, 19747-19751. 9. Tamber S., Ochs M.M., and Hancock R.E. (2006) Role of the novel OprD family of porins in nutrient uptake in Pseudomonas aeruginosa. J Bacteriol. 188, 45-54. 10. Eren E., Vijayaraghavan J., Liu J., Cheneke B.R., Touw D.S., Lepore B.W., Indic M., Movileanu L., and van den Berg B. (2012) Substrate specificity within a family of outer membrane carboxylate channels. PLoS Biol. 10, e1001242. 11. Stanier, R.Y, Palleroni, N.J., and Doudoroff, M. The aerobic pseudomonads: a taxonomic study. (1966) J. Gen. Microbiol. 43, 159-271. 12. Daddaoua A., Krell T., and Ramos J.L. (2009) Regulation of glucose metabolism in Pseudomonas: the phosphorylative branch and entner-doudoroff enzymes are regulated by a repressor containing a sugar isomerase domain. J Biol Chem. 284, 21360-21368. 13. Hancock, R. E. W., and Carey A.M. (1980) Protein D1-A glucose inducible, pore-forming protein from the outer membrane of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 8, 105-109. 14. Saravolac E.G., Taylor N.F., Benz R., and Hancock R.E. (1991) Purification of glucose- inducible outer membrane protein OprB of Pseudomonas putida and reconstitution of glucose-specific pores. J Bacteriol. 173, 4970-4976. 15. Wylie J.L., and Worobec E.A. (1995) The OprB porin plays a central role in carbohydrate uptake in Pseudomonas aeruginosa. J Bacteriol. 177, 3021-3026. 16. Adewoye L.O., Tschetter L., O'Neil J., and Worobec E.A. (1998) Channel specificity and secondary structure of the glucose-inducible porins of Pseudomonas spp. J Bioenerg Biomembr. 30, 257-267. 17. Guzman L.M., Belin D., Carson M.J., and Beckwith J. (1995) Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol 177, 4121-4130. 18. Van den Berg B., Clemons W.M., Jr., Collinson I., Modis Y., Hartmann E., et al. (2004) X- ray structure of a protein-conducting channel. Nature 427, 36-44. 19. Van Duyne G.D., Standaert R.F., Karplus P.A., Schreiber S.L., and Clardy J. (1993) Atomic structures of the human immunophilin FKBP-12 complexes with FK506 and rapamycin. Journal of molecular biology 229, 105-124 20. Otwinowski Z., and Minor W. (1997) Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol 276, 307-326. 21. Terwilliger T.C., Adams P.D., Read R.J., McCoy A.J., Moriarty N.W., Grosse- Kunstleve R.W., Afonine P.V., Zwart P.H., and Hung L.-W. (2009) Decision-making in structure solution using Bayesian estimates of map quality: the PHENIX AutoSol wizard. Acta Cryst. D65, 582-601. 22. Adams P.D., Grosse-Kunstleve R.W., Hung L.W., Ioerger T.R., McCoy A.J., Moriarty N.W., Read R.J., Sacchettini J.C., Sauter N.K., and Terwilliger T.C. (2002) PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D58, 1948-1954.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

11
23. Emsley P., and Cowtan K. (2004) Coot: model-building tools for molecular graphics. Acta Crystallogr D60, 2126-2132 24. Nikaido H., Rosenberg E.Y., and Foulds J. (1983) Porin channels in Escherichia coli: studies with beta-lactams in intact cells. J Bacteriol 153, 232-240. 25. Bagos P.G., Liakopoulos T.D., Spyropoulos I.C., and Hamodrakas S.J. (2004) PRED- TMBB: a web server for predicting the topology of beta-barrel outer membrane proteins. Nucleic Acids Res. 32(Web Server issue), W400-404. 26. Shrivastava R., Basu B., Godbole A., Mathew M.K., Apte S.K., and Phale P.S. (2011) Repression of the glucose-inducible outer-membrane protein OprB during utilization of aromatic compounds and organic acids in Pseudomonas putida CSV86. Microbiology 157, 1531-1540. 27. Phale P.S., Philippsen A., Kiefhaber T., Koebnik R., Phale V.P., Schirmer T., Rosenbusch J.P. (1998) Stability of trimeric OmpF porin: the contributions of the latching loop L2. Biochemistry. 37, 15663-15670. 28. Van Gelder P, Tommassen J. Demonstration of a folded monomeric form of porin PhoE of Escherichia coli in vivo. (1996) J Bacteriol. 178, 5320-5322. 29. Seshadri K., Garemyr R., Wallin E., von Heijne G., and Elofsson A. (1998) Architecture of beta-barrel membrane proteins: analysis of trimeric porins. Protein Sci 7, 2026-2032. 30. Holm L., and Rosenstrom P. (2010) Dali server: conservation mapping in 3D. Nucleic acids research 38, W545-549 31. Dargent B., Rosenbusch J., and Pattus F. (1987) Selectivity for maltose and maltodextrins of maltoporin, a pore-forming protein of E. coli outer membrane. FEBS Lett. 220, 136-142. 32. Schülein K., and Benz R. (1990) LamB (maltoporin) of Salmonella typhimurium: isolation, purification and comparison of sugar binding with LamB of Escherichia coli. Mol Microbiol. 4, 625-632. 33. Benz R., Francis G., Nakae T., and Ferenci T. (1992) Investigation of the selectivity of maltoporin channels using mutant LamB proteins: mutations changing the maltodextrin binding site. Biochim Biophys Acta. 1104, 299-307. 34. Benz, R., Schmid A., Nakae T., and Vos-Scheperkeuter. G.H. (1986). Pore formation by LamB of Escherichia coli in lipid bilayer membranes. J. Bacteriol. 165, 978-986. 35. Forst D., Welte W., Wacker T., and Diederichs K. (1997) Structure of the sucrose-specific porin ScrY from Salmonella typhimurium and its complex with sucrose. Nat Struct Biol. 5, 37-46. 36. The PyMOL Molecular Graphics System, Version 1.2r3pre, Schrödinger, LLC. 37. Dolinsky T.J., Nielsen J.E., McCammon J.A., Baker N.A. (2004) PDB2PQR: an automated pipeline for the setup, execution, and analysis of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Research 32 W665-W667 (2004) ACKNOWLEDGEMENTS I would like to thank the staff of NSLS beamline X6A for beam time and for assistance during data collection. I am also grateful to Deborah Hogan (Dartmouth Medical School) for her gift of P. aeruginosa genomic DNA. The research in my lab is supported by a grant from the NIH (RO1 GM085785).
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

12
FOOTNOTES The atomic coordinates and structure factors (codes 4GEY and 4GF4) have been deposited in the Protein Data Bank, Research Collaboratory for Structural Bioinformatics, Rutgers University, New Brunswick, NJ (http://www.rcsb.org/). FIGURE LEGENDS
Figure 1. Crystal structure of P. putida F1 OprB-1. A. Cartoon representation viewed from the
side and from the extracellular environment (right panel), with helices colored magenta and β-
strands blue. B. SDS-PAGE analysis of purified OprB and OprP. Lane 1; OprB boiled, lane 2;
OprB unboiled, lane 3; OprP boiled, lane 4; OprP unboiled. Molecular weight markers are
indicated on the left. On the right, the various oligomeric states are indicated: T, trimer; MU,
monomer unfolded; MF, monomer folded. C. Pore architecture of OprB, with loop L2 colored
green and loop L3 colored orange. The pore is indicated with an asterisk. The right panel shows
a surface view in the same orientation and with the same coloring as (A). D. Side slab view,
showing the location of the conserved disulfide bond (magenta sticks). For clarity, parts of trans-
membrane strands S3 and S4 have been omitted. The structural figures were made with PYMOL
(36).
Figure 2. Locations of conserved residues in OprB orthologs. Stereoview showing absolutely
conserved residues from a CLUSTALW alignment as green stick models (with oxygen atoms
colored red and nitrogen atoms blue). Selected residues have been labeled. Loop L2 is colored
green and loop L3 orange. The view is from the extracellular side.
Figure 3. Substrate specificity of F1OprB-1. A. Liposome swelling data for sugars and selected
amino acids. B. Comparison of substrate uptake mediated by reduced (open symbols) and
oxidized OprB (closed symbols). Red; glucose, green; glucuronate, yellow; maltose. C. Relative
swelling rates (24), determined from the first 60 s of the assays shown in A. The uptake mediated
by glucose has been set to 1.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

13
Figure 4. The channel constriction of F1OprB-1 is predominantly negatively charged. A,B.
Surface views (including hydrogen atoms) from the extracellular sides and from the periplasmic
side (C), showing positively charged surfaces in blue and negatively charged surfaces in red
(colored from -5kT/e to +5 kT/e). Positions of selected extracellular loops have been indicated.
The figure was made using the APBS plugin within PYMOL (36). The PQR file was generated
using the PDB2PQR server (37).
Figure 5. Interaction of glucose with F1OprB-1. A. Surface view from the extracellular milieu,
showing glucose bound in the OprB pore as a space-filling model (carbon atoms colored yellow
and oxygen atoms colored red). B. Overview from the side, showing the substrate bound in the
eyelet. C. Stereoview close-up of (B), showing the OprB eyelet residues. Amino acids that
contact the substrate are labeled in bold. Electron density from a 2Fo-Fc map (contoured at 1.0
σ) is shown as a blue mesh for the OprB residues and as a red mesh for the glucose substrate.
The Cys148-Cys156 disulfide bond is shown as well.
Figure 6. Comparison between OprB and OprP. A. Cartoon diagrams of a superposition of OprB
(yellow) and OprP (blue), viewed from the side and from the top (left panel). B. Views from the
extracellular side, showing the loops L2 (green), L3 (orange) and L5 (purple) within OprP (left
panel) and OprB (right panel).
Figure 7. Comparison between the carbohydrate OM channels LamB and OprB. A. Stereo
diagram showing maltohexaose bound to E. coli LamB (PDB ID; 1OMP). The substrate is
colored green (with oxygen atoms red). Aromatic residues within the eyelet and greasy slide are
shown in pink, and ionizable residues that form hydrogen bonds with the substrate are shown in
orange. The trimer interface is colored light yellow, with loop L2 labeled. B. Stereo diagram
showing OprB with bound glucose. The coloring is as in (A). For comparison with LamB, loop
L2 is colored light yellow and labeled.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from
14
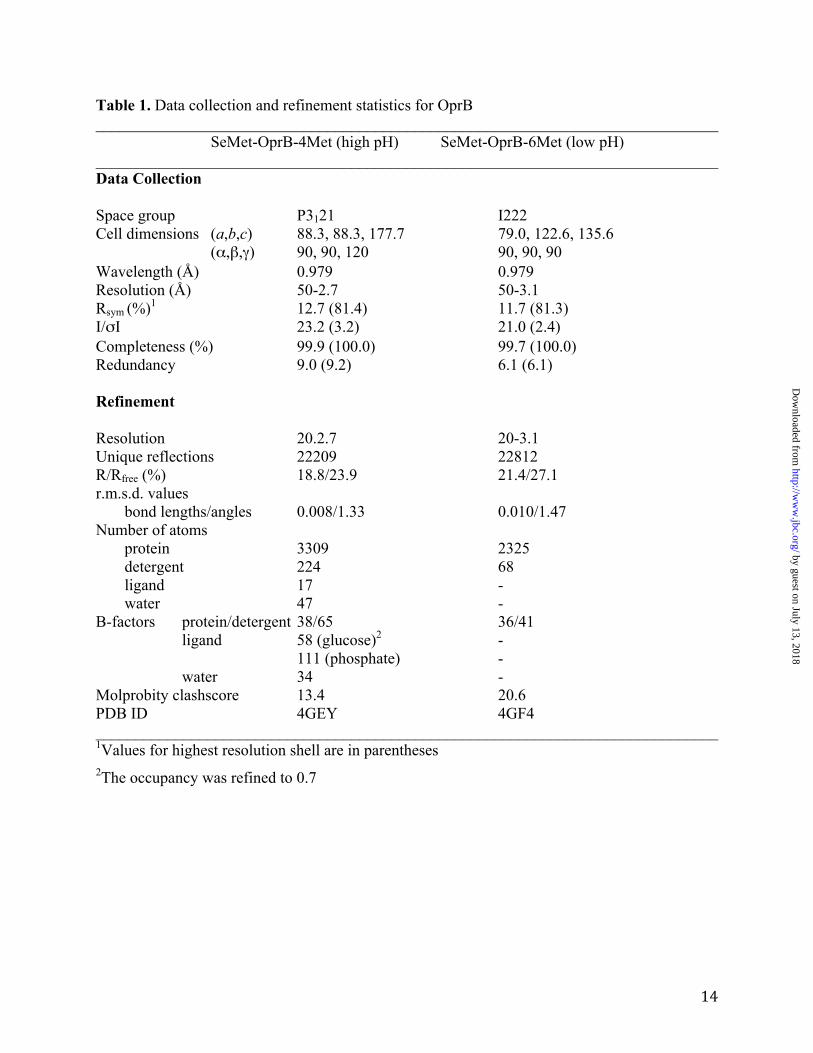
Table 1. Data collection and refinement statistics for OprB ______________________________________________________________________________ SeMet-OprB-4Met (high pH) SeMet-OprB-6Met (low pH) ______________________________________________________________________________ Data Collection Space group P3121 I222 Cell dimensions (a,b,c) 88.3, 88.3, 177.7 79.0, 122.6, 135.6 (α,β,γ) 90, 90, 120 90, 90, 90 Wavelength (Å) 0.979 0.979 Resolution (Å) 50-2.7 50-3.1 Rsym (%)1 12.7 (81.4) 11.7 (81.3) I/σI 23.2 (3.2) 21.0 (2.4) Completeness (%) 99.9 (100.0) 99.7 (100.0) Redundancy 9.0 (9.2) 6.1 (6.1) Refinement Resolution 20.2.7 20-3.1 Unique reflections 22209 22812 R/Rfree (%) 18.8/23.9 21.4/27.1 r.m.s.d. values bond lengths/angles 0.008/1.33 0.010/1.47 Number of atoms protein 3309 2325 detergent 224 68 ligand 17 - water 47 - B-factors protein/detergent 38/65 36/41 ligand 58 (glucose)2 - 111 (phosphate) - water 34 - Molprobity clashscore 13.4 20.6 PDB ID 4GEY 4GF4 ______________________________________________________________________________ 1Values for highest resolution shell are in parentheses 2The occupancy was refined to 0.7
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from
15
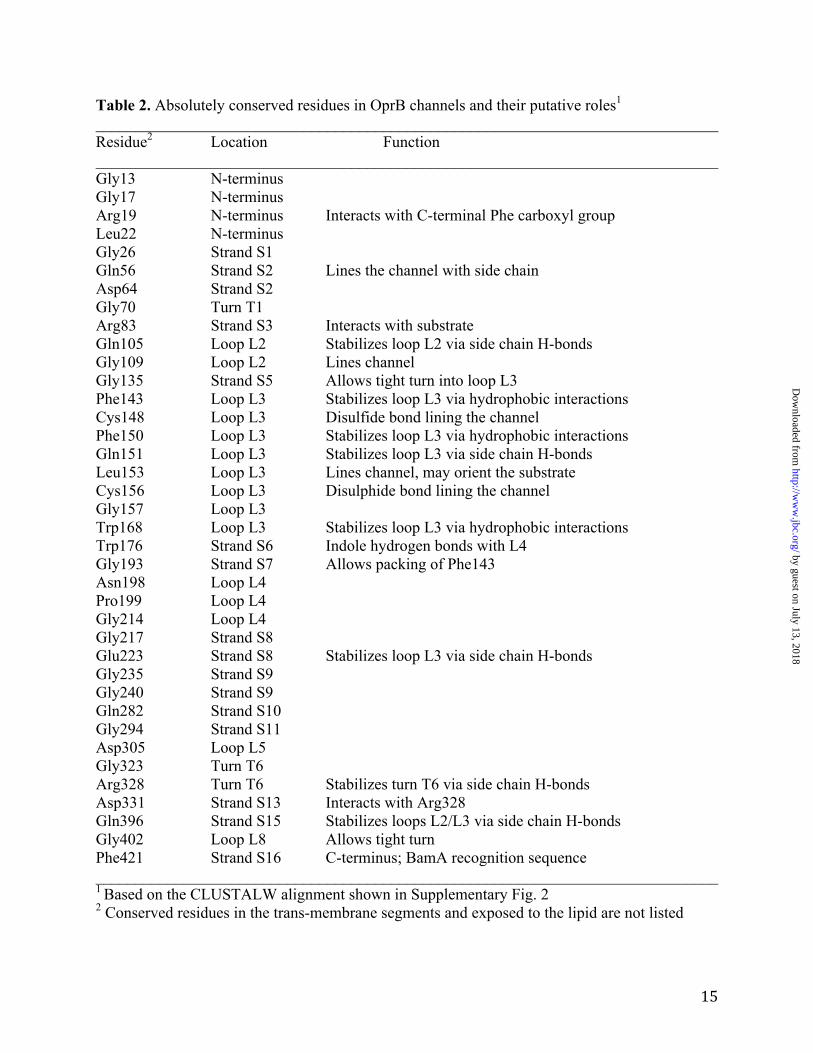
Table 2. Absolutely conserved residues in OprB channels and their putative roles1 ______________________________________________________________________________ Residue2 Location Function ______________________________________________________________________________ Gly13 N-terminus Gly17 N-terminus Arg19 N-terminus Interacts with C-terminal Phe carboxyl group Leu22 N-terminus Gly26 Strand S1 Gln56 Strand S2 Lines the channel with side chain Asp64 Strand S2 Gly70 Turn T1 Arg83 Strand S3 Interacts with substrate Gln105 Loop L2 Stabilizes loop L2 via side chain H-bonds Gly109 Loop L2 Lines channel Gly135 Strand S5 Allows tight turn into loop L3 Phe143 Loop L3 Stabilizes loop L3 via hydrophobic interactions Cys148 Loop L3 Disulfide bond lining the channel Phe150 Loop L3 Stabilizes loop L3 via hydrophobic interactions Gln151 Loop L3 Stabilizes loop L3 via side chain H-bonds Leu153 Loop L3 Lines channel, may orient the substrate Cys156 Loop L3 Disulphide bond lining the channel Gly157 Loop L3 Trp168 Loop L3 Stabilizes loop L3 via hydrophobic interactions Trp176 Strand S6 Indole hydrogen bonds with L4 Gly193 Strand S7 Allows packing of Phe143 Asn198 Loop L4 Pro199 Loop L4 Gly214 Loop L4 Gly217 Strand S8 Glu223 Strand S8 Stabilizes loop L3 via side chain H-bonds Gly235 Strand S9 Gly240 Strand S9 Gln282 Strand S10 Gly294 Strand S11 Asp305 Loop L5 Gly323 Turn T6 Arg328 Turn T6 Stabilizes turn T6 via side chain H-bonds Asp331 Strand S13 Interacts with Arg328 Gln396 Strand S15 Stabilizes loops L2/L3 via side chain H-bonds Gly402 Loop L8 Allows tight turn Phe421 Strand S16 C-terminus; BamA recognition sequence ______________________________________________________________________________ 1 Based on the CLUSTALW alignment shown in Supplementary Fig. 2 2 Conserved residues in the trans-membrane segments and exposed to the lipid are not listed
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Bert van den BergStructural basis for outer membrane sugar uptake in pseudomonads
published online October 12, 2012J. Biol. Chem.
10.1074/jbc.M112.408518Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2012/10/12/M112.408518.DC1
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from