Embed Size (px)
Citation preview

Structure
Article
Structural Basis for Ligand Recognitionand Activation of RAGEMichael Koch,1 Seth Chitayat,2 Brian M. Dattilo,2 Andre Schiefner,3 Joachim Diez,4 Walter J. Chazin,2,*and Gunter Fritz1,5,*1Department of Biology, University of Konstanz, 78457 Konstanz, Germany2Departments of Biochemistry and Chemistry, Center for Structural Biology, Vanderbilt University, Nashville, TN 37232-8725, USA3Lehrstuhl fur Biologische Chemie, Technische Universitat Munchen, 85350 Freising-Weihenstephan, Germany4Swiss Light Source at Paul Scherrer Institute, 5232 Villigen PSI, Switzerland5Department of Neuropathology, University of Freiburg, 79106 Freiburg, Germany
*Correspondence: [email protected] (W.J.C.), [email protected] (G.F.)DOI 10.1016/j.str.2010.05.017
SUMMARY
The receptor for advanced glycation end products(RAGE) is a pattern recognition receptor involvedin inflammatory processes and is associated withdiabetic complications, tumor outgrowth, and neuro-degenerative disorders. RAGE induces cellular sig-naling events upon binding of a variety of ligands,such as glycated proteins, amyloid-b, HMGB1, andS100 proteins. The X-ray crystal structure of theVC1 ligand-binding region of the human RAGE ecto-domainwas determined at 1.85 A resolution. The VC1ligand-binding surface was mapped onto the struc-ture from titrations with S100B monitored by hetero-nuclear NMR spectroscopy. These NMR chemicalshift perturbations were used as input for restraineddocking calculations to generate a model for theVC1-S100B complex. Together, the arrangement ofVC1 molecules in the crystal and complementarybiochemical studies suggest a role for self-associa-tion in RAGE function. Our results enhance under-standing of the functional outcomes of S100 proteinbinding to RAGE and provide insight into mecha-nistic models for how the receptor is activated.
INTRODUCTION
The receptor for advanced glycation end products (RAGE) is
a member of the immunoglobulin (Ig) superfamily of cell surface
receptors (Neeper et al., 1992; Schmidt et al., 1992). RAGE
signaling plays a central role in the inflammatory response (Hof-
mann et al., 1999), mediating aspects of innate immunity (Tian
et al., 2007), acute and chronic inflammatory disorders (Orlova
et al., 2007), and certain cancers (Taguchi et al., 2000). Ligand
binding to the ectodomain of RAGE is tightly coupled to the
recruitment of intracellular factors such as Diaphanous-1 to the
cytoplasmic domain, ultimately leading to the proliferation of
intracellular signals (Hudson et al., 2008b). Activated RAGE is
known to recruit extracellular signal–regulated kinase–1 and –2
1342 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
(ERK-1/2) (Ishihara et al., 2003), resulting in downstream activa-
tion of NF-kB via the MAP kinase pathway.
RAGE mediates physiological and pathological effects
through interaction with a diverse set of ligands, which remark-
ably, are each associated with a specific disease. The first
identified RAGE ligand was advanced glycation end products
(AGEs), which form by nonenzymatic glycation of proteins and
lipids and accumulate as a result of normal aging and inflamma-
tory processes, particularly in diabetes (Yamagishi et al., 2003).
RAGE is up-regulated in Alzheimer disease, and soluble
amyloid-b has been shown to bind to the receptor inducing
oxidative stress in neurons and the production of proinflamma-
tory cytokines in microglia (Yan et al., 2003). RAGE in the
endothelium mediates amyloid-b transport across the blood
brain barrier into the central nervous system, promoting
amyloid-b plaque formation (Deane et al., 2003). Activation of
RAGE on neurons by high mobility group protein 1 (HMGB1)
induces reorganization of the cytoskeleton and neurite extension
(Hori et al., 1995). However, in cancer cells, RAGE-HMGB1 inter-
action stimulates tumor invasiveness and growth (Taguchi et al.,
2000). Moreover, HMGB1-DNA complexes trigger the associa-
tion of RAGE with Toll-like receptor 9, which is essential for the
immune response to pathogens (Tian et al., 2007). Several
S100 proteins bind and activate RAGE. In fact, the first native
RAGE ligand discovered was EN-RAGE (S100A12), which initi-
ates and sustains the inflammatory response (Hofmann et al.,
1999). S100B binding to RAGE is among the best characterized,
having been shown to inducemultimerization of the receptor and
promote neurite extension and neuronal survival (Huttunen et al.,
2000; Leclerc et al., 2007; Ostendorp et al., 2007).
The specific association of RAGE with disease pathogenesis
has resulted in a growing interest in RAGE as a therapeutic
target. The initial steps toward validating RAGE as a target
involved the use of the extracellular ligand-binding region of
RAGE (sRAGE), which effectively competes with the receptor
for ligands. For example, sRAGE blockage of receptor activation
in cell culture has been useful for studies in mouse models for
various diseases (Deane et al., 2003; Park et al., 1998; Yan
et al., 2003). Amodel for advanced atherosclerosis shows severe
plaque development in the aorta, the formation of which is sup-
pressed in mice treated with exogenous sRAGE (Park et al.,
1998). In a mouse model for enterocolitis, administration of
sRAGE prevented immune cell infiltration into the colon and
All rights reserved

Structure
Structure of Human RAGE
sepsis, presumably by blocking the S100A12-RAGE interaction
and consequent signaling (Hofmann et al., 1999). Alternative
splicing of RAGE mRNA results in an endogenous secretory
form of RAGE (esRAGE). Levels of esRAGE are largely
decreased in inflammatory and neurodegenerative disorders
(Emanuele et al., 2005; Ghidoni et al., 2008; Sternberg et al.,
2008; Wittkowski et al., 2007), implying a regulatory function
for esRAGE.
In order to better understand the molecular basis for RAGE
signaling and its role in cellular physiology, and to lay a founda-
tion for exploiting RAGE as a therapeutic target, we have
undertaken a comprehensive analysis of the structure and
ligand-binding properties of human sRAGE. sRAGE is
composed of a V-type immunoglobulin (Ig) domain and two
C-type Ig domains (C1 and C2) (Schmidt et al., 1992). Previous
characterization of sRAGE revealed the C-terminal Ig domain
(C2) to be structurally independent of the first N-terminal two Ig
domains (V and C1), which together form an integrated structural
unit, VC1 (Dattilo et al., 2007). VC1 has been shown to bind
AGE-modified BSA and S100B (Allmen et al., 2008; Dattilo
et al., 2007). Here, we report a high-resolution crystal structure
of the tandem VC1 domain pair and link its unique structural
features to its binding properties. Heteronuclear NMR spectros-
copy is used to map the VC1-binding site for S100B and the
S100B-binding site for VC1. The NMR data are then used in
combination with molecular docking calculations to generate
a model of the complex. These results shed new light on models
for RAGE activation.
RESULTS
VC1 Forms an Integrated Structural UnitThe tandem RAGE VC1 domains were crystallized and the X-ray
structure was refined at 1.85 A resolution to a Rcryst of 20.9%
and Rfree of 24.0% (Table 1). VC1 is found to form a bent elon-
gated structure with an angle of 145� between the two Ig
domains (Figure 1A). A specific interface between the domains
is observed, which includes several interdomain hydrogen
bonds and hydrophobic interactions (Figure 1A). Among these
H-bonds, Gln119 in V engages in a pseudo b sheet hydrogen-
bonding pattern with Tyr150 in C1, and the carboxylate oxygen
atom of Glu94 in V forms an H-bond with the hydroxyl hydrogen
of Tyr150 in C1. In addition, a water molecule bridges the amide
nitrogen of Ile120 in C1 domain with the side chains of Gln119
and Arg29 in V. Hydrophobic contacts at the interface are medi-
ated between side chain atoms of Pro215 from the C1 F-G loop
and the side chain of Tyr118, as well as between the side chain of
Tyr150 from the C1 B-C loop and Ile91. Notably, all residues at
the VC1 interface are strictly conserved (see Figure S1 available
online). These characteristics confirm that the two domains form
an integrated structural unit (Dattilo et al., 2007).
On the basis of sequence alignments, the N-terminal domain
of RAGE (V) had been assigned to the V-set type of Ig molecules.
However, once the structure was determined, it revealed
features typical of I-set topology (Harpaz and Chothia, 1994),
which is characteristic for cell adhesion molecules (Casasnovas
et al., 1997; Feinberg et al., 2001; Freigang et al., 2000; Kasper
et al., 2000; Su et al., 1998) or muscle proteins (Holden et al.,
1992; Zou et al., 2006). Strands A, C, C0, F, andG form one sheet,
Structure 18, 1342–1
whereas strands B, D, and E form the second sheet (Figure 1).
The region in the V domain corresponding to C00 strand in V-set
Ig domains instead forms an extended loop encompassing
three glycine residues (Gly68, Gly69, and Gly70) and two proline
residues (Pro66 and Pro71). The frequency of glycine and proline
residues is inconsistent with formation of a b strand. High
B-factors in this region suggest that the loop might be adopting
variable conformations, which is not unusual for a short Gly and
Pro-rich sequence. Although more correctly classified as an I
domain, the V nomenclature will be retained here in order to
maintain consistency with previous literature.
The C1-domain fits to the C1-set of Ig molecules, with strands
A, B, D, and E forming the back b sheet and strands C, C0, F, andG forming the front sheet of the b sandwich. However, the RAGE
C1-domain has a unique topology distinct from other Ig fold sets:
it contains two additional b strands, A0 and G0, which belong
neither to the front nor to the back sheet. Both strands form an
additional parallel b sheet stabilizing the C terminus of the
domain (Figures 1A and 1B). Further variation is observed for
strands D and E. In comparison to most C1-type Ig domains,
strand D is elongated by two residues encompassing Arg178
and Arg179, as well as the D-E loop, which is slightly longer
through the inclusion of Pro181-Gly184. The E-F loop, which
includes Thr195-Thr205, is also elongated by residues Arg198-
Asp201. This loop has a unique interaction that stabilizes the C
terminus of C1 involving a hydrogen bond between the Arg198
side chain and the Pro232 main chain carbonyl oxygen.
A well-ordered 10-residue C-terminal extension from C1
(Glu231-Leu240) is observed in the high-resolution structure
(Figures 1A and 1B). This extension constitutes at least in part
the linker to the C2 domains, which NMR studies have shown
is flexible (Dattilo et al., 2007). A solution NMR structure of the
isolated C2 domain has been determined (PDB 2ENS), but the
N terminus is not well defined. In our structure, the extension is
stabilized by a contact between Leu240 and the C1-domain of
a neighboring molecule in the crystal. Hence, some uncertainty
remains over the exact length of the linker between the C1 and
C2 domains.
VC1 Contains a Highly Basic SurfaceOne of the most unusual properties of RAGE is its ability to
engage very diverse ligands of different size, fold, or symmetry.
The structure of the VC1 ligand-binding domain provides a
means to assess what specific surface properties enable this
unique characteristic. Our analysis revealed large positively
charged patches on the V domain (Figure 2), which are formed
by highly conserved Arg and Lys residues. A patch of positively
charged electrostatic surface was also identified on the C1
domain. Thus, the entire V domain and a major part of C1 form
a large area of electropositive surface (Figure 2) that fits well to
the acidic (negative) character of the diverse RAGE ligands,
including AGE-modified proteins, amyloid-b, and S100 proteins.
Comparison of molecular surfaces displaying the isoelectro-
static field potential of structural homologs neural adhesion
molecules TAG-1/axonin-1 (Freigang et al., 2000) and N-CAM
(Kasper et al., 2000), as well as a structural model of the closest
homolog ALCAM, reveals that the electrostatic potential of VC1
is significantly stronger (Figure 2), which suggests RAGE
contains an electrostatic trap for negatively charged ligands.
352, October 13, 2010 ª2010 Elsevier Ltd All rights reserved 1343

Table 1. Crystallographic Data and Refinement Statistics
Native Peak Zn-MAD Inflection Remote
Data collection
Wavelength 1.0082 1.2820 1.2825 1.0082
Resolution (A) 47–1.85 (2.0–1.85)a 47–2.6 (2.7–2.6)a 47–2.6 (2.7–2.6)a 47–2.4 (2.1–2.0)a
Space group P21212 P21212 P21212 P21212
Cell dimensions (A) 74.85, 119.96, 28.89 74.76, 119.90, 28.87 74.76, 119.90, 28.87 74.76, 119.90, 28.87
Total observations 153,044 (23,037) 57,942 (15,395) 57,858 (15,401) 124,565 (33,842)
Unique reflections 28,559 (4,704) 6,158 (1,619) 6,138 (1,622) 15,819 (4,599)
Completeness (%) 99.7(99.8)a 99.9 (100.0)a 99.9 (100.0)a 99.7 (99.7)a
Redundancy 6.6 (6.0)a 3.7 (3.8)a 3.7 (3.8)a 3.7 (3.4)a
Rsymb (%) 8.2 (57.1)a 6.0 (19.1)a 5.5 (16.5)a 7.6 (32.9)a
I / sI 12.2 (2.6)a 16.8 (6.7)a 17.6 (7.4)a 11.3 (3.6)a
Phasing power acentric 1.391 1.382 0.419
FOM acentric/centric 0.63279/0.44639
Refinement
Resolution (A) 1.85 2.0 2.0 2.0
Rcrystc / Rfree
d (%) 20.9/24.0 (24.7/26.0)a
No. atoms
Protein 1938
Ligand/ion 4
Water 229
B-factors
Protein 25.6
Ligand/ion 44.8
Water 47.0
R.m.s deviations
Bond angles (�) 1.99
Bond lengths (A) 0.018
Ramachandran plot
Most favored (%) 87.8
Additionally allowed (%) 11.2
Generously allowed (%) 1.2aNumbers in parentheses apply to the highest-resolution shell.b Rsym = Shkl Sj j Ij � < I > j Shkl Sj j Ij j.c Rcryst = Shklj Fobs – Fc j / ShklFobs, where Fobs and Fc are observed and calculated structure factors, respectively.d Five percent randomly selected reflections were excluded from refinement and used for the calculation of Rfree.
Structure
Structure of Human RAGE
Self-Association of RAGERecently, it was shown that RAGE forms oligomers on the
plasma membrane (Xie et al., 2008), and it was suggested that
oligomerization is mediated by the C1 domain (Xie et al., 2007).
Indeed, in the crystal structure, there are side-by-side contacts
between VC1 molecules mainly involving residues in the C1
domain. The solvent accessible surface area buried by this
interface is 980 A2 (Figure 3A). The contact surface contains
several charged residues and is connected by a tight network
of hydrogen bonds and polar interactions (Figures 3B and 3C).
The guanidine of Arg178 side chain forms a salt bridge with the
carboxylate group of Glu1680 and a hydrogen bond to the
carbonyl oxygen of Pro1630 of the neighboring molecule
(Figure 3C). The Gln176 side chain oxygen forms two hydrogen
bonds to the N3 of Arg178, which positions the guanidine group
for optimal interaction with the neighboring molecule (Figure 3C;
1344 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
Figure S2). These interactions are reminiscent of the critical salt
bridge formed between Arg and Glu/Asp residues at the inter-
face between Ig domains in the active dimer of the receptor
tyrosine kinase KIT (Yuzawa et al., 2007) or between the
membrane-proximal Ig-domains of the VEGF receptor (Yang
et al., 2010). The extensive set of stabilizing interactions between
C1 domains in adjacent molecules in the crystal structure is
consistent with the proposal of RAGE aggregation in the plasma
membrane. In addition to H-bonds and salt bridges, a Zn2+ ion is
found at the intermolecular interface. The Zn2+ is coordinated by
residues His180, Glu182, and His1580 from the adjacent VC1
molecule (Figure 3B). These residues, like most involved in at
the C1-C1 interface, are conserved across species (Figure S1),
which implies a role for Zn2+ in receptor multimerization. We
note that Zn2+ can induce dimerization of the class II major
histocompatibility complex, leading to cooperative binding of a
All rights reserved

Figure 1. Structure of the Tandem VC1
Domains of RAGE
(A) Stereo ribbon diagram VC1 with the V domain
in green and the C1 domain in magenta. The two
Ig domains adopt a fixed orientation that is
stabilized by hydrogen bonds and hydrophobic
contacts.
(B) Topology diagram of VC1 with the strands
colored as in (A). The structure of the V domain
shows it belongs to the I-set of Ig domains,
whereas C1 remains in the C1 set. Note that a
unique parallel b sheet is formed by strands A0
and G0, which stabilizes the C terminus of the C1
domain.
See also Figure S1.
Structure
Structure of Human RAGE
superantigen ligand (Li et al., 2007). Moreover, homophilic inter-
actions of receptor Ig-type ectodomains play a central role in
ligand-binding and cellular activation (Stuttfeld and Ballmer-
Hofer, 2009; Xu and Jin, 2010; Yang et al., 2010).
In order to further investigate self-association of VC1 and a
potential role for Zn2+, we performed dynamic light scattering
experiments. The data show that VC1 is monomeric in the
absence of Zn2+ and at low pH. The tendency for VC1 to self-
Structure 18, 1342–1352, October 13, 2010 ª
associate is heavily influenced by Zn2+
and increased pH into the physiological
range (Figures 4A–4C). The degree of
self-association of VC1 is concentration
dependent (Figure 4D), which is consis-
tent with self-association being a specific
binding phenomenon. The concentration
dependence of self-association is pro-
moted by the presence of Zn2+ and
shows a saturation behavior that could
be fit by a hyperbolic function yielding
a Kd app of 3.8 ± 1.3 mM.
An NMR-Based Model for theComplex of VC1 with the RAGELigand S100BStructural details about ligand binding to
RAGE have been limited to a single study
thathighlightedseveral residues in theVdomain that are important
forbindingofAGEs (Matsumotoetal., 2008).Wehave investigated
the structural basis for binding of the ligand S100B, combining
two-dimensional heteronuclear NMR experiments to identify the
binding interface on both VC1 and S100B and HADDOCK-based
docking calculations to generate a model of the complex.
Tomap the S100B-binding site onRAGE, a series of 2D 15N-1H
HSQC NMR experiments were acquired as Ca2+-S100B was
Figure 2. Surface Analysis of VC1
Surface representation of the tandem VC1
domains and its closest structural homologs TAG
1/axonin-1 (Ig domains 3,4), neuronal cell adhe-
sion molecule (N-CAM Ig domains 2,3), and
activated leukocyte adhesion molecule (ALCAM
Ig domains 1,2). Positive electrostatic isopotential
surfaces at 0.8 kT/e are shown in blue. RAGE VC1
exhibits an unusually large positive electrostatic
potential. The isosurface shows a coherent poten-
tial across both Ig domains extending far into
space. The figure was prepared using PyMOL
(http://www.pymol.org) and APBS (Baker et al.,
2001).
See also Figure S2.
2010 Elsevier Ltd All rights reserved 1345
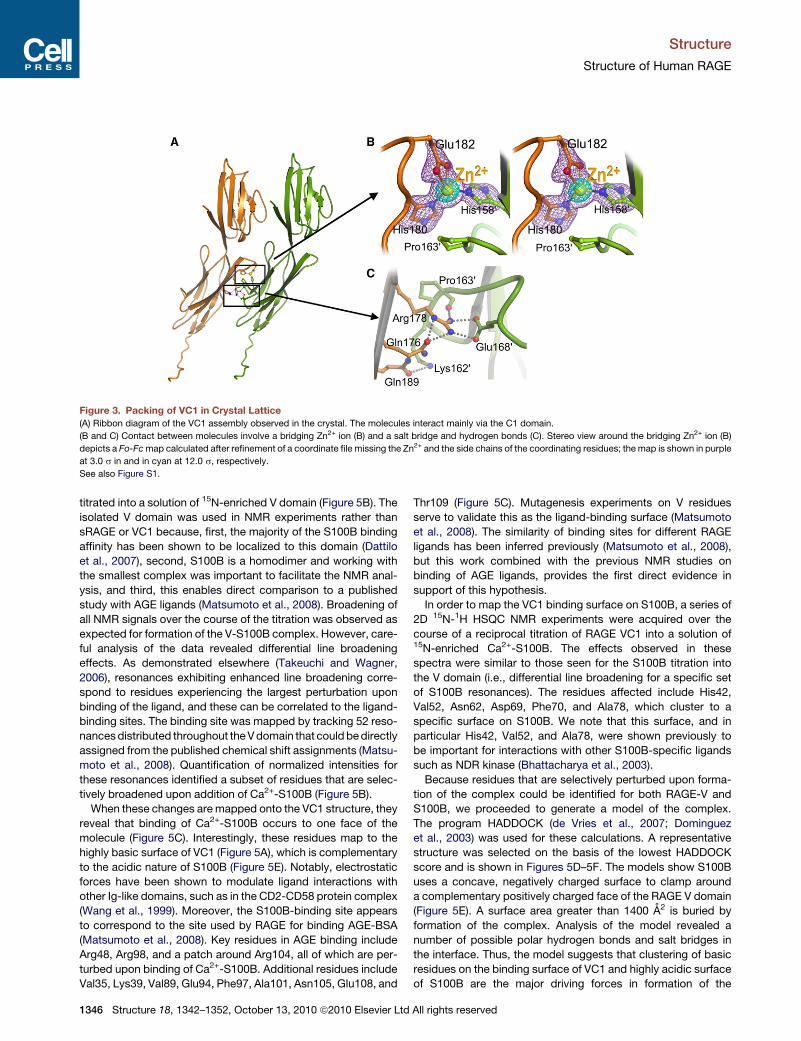
Figure 3. Packing of VC1 in Crystal Lattice
(A) Ribbon diagram of the VC1 assembly observed in the crystal. The molecules interact mainly via the C1 domain.
(B and C) Contact between molecules involve a bridging Zn2+ ion (B) and a salt bridge and hydrogen bonds (C). Stereo view around the bridging Zn2+ ion (B)
depicts a Fo-Fcmap calculated after refinement of a coordinate file missing the Zn2+ and the side chains of the coordinating residues; the map is shown in purple
at 3.0 s in and in cyan at 12.0 s, respectively.
See also Figure S1.
Structure
Structure of Human RAGE
titrated into a solution of 15N-enriched V domain (Figure 5B). The
isolated V domain was used in NMR experiments rather than
sRAGE or VC1 because, first, the majority of the S100B binding
affinity has been shown to be localized to this domain (Dattilo
et al., 2007), second, S100B is a homodimer and working with
the smallest complex was important to facilitate the NMR anal-
ysis, and third, this enables direct comparison to a published
study with AGE ligands (Matsumoto et al., 2008). Broadening of
all NMR signals over the course of the titration was observed as
expected for formation of the V-S100B complex. However, care-
ful analysis of the data revealed differential line broadening
effects. As demonstrated elsewhere (Takeuchi and Wagner,
2006), resonances exhibiting enhanced line broadening corre-
spond to residues experiencing the largest perturbation upon
binding of the ligand, and these can be correlated to the ligand-
binding sites. The binding site was mapped by tracking 52 reso-
nancesdistributed throughout theVdomain that could bedirectly
assigned from the published chemical shift assignments (Matsu-
moto et al., 2008). Quantification of normalized intensities for
these resonances identified a subset of residues that are selec-
tively broadened upon addition of Ca2+-S100B (Figure 5B).
When these changes aremapped onto the VC1 structure, they
reveal that binding of Ca2+-S100B occurs to one face of the
molecule (Figure 5C). Interestingly, these residues map to the
highly basic surface of VC1 (Figure 5A), which is complementary
to the acidic nature of S100B (Figure 5E). Notably, electrostatic
forces have been shown to modulate ligand interactions with
other Ig-like domains, such as in the CD2-CD58 protein complex
(Wang et al., 1999). Moreover, the S100B-binding site appears
to correspond to the site used by RAGE for binding AGE-BSA
(Matsumoto et al., 2008). Key residues in AGE binding include
Arg48, Arg98, and a patch around Arg104, all of which are per-
turbed upon binding of Ca2+-S100B. Additional residues include
Val35, Lys39, Val89, Glu94, Phe97, Ala101, Asn105, Glu108, and
1346 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
Thr109 (Figure 5C). Mutagenesis experiments on V residues
serve to validate this as the ligand-binding surface (Matsumoto
et al., 2008). The similarity of binding sites for different RAGE
ligands has been inferred previously (Matsumoto et al., 2008),
but this work combined with the previous NMR studies on
binding of AGE ligands, provides the first direct evidence in
support of this hypothesis.
In order to map the VC1 binding surface on S100B, a series of
2D 15N-1H HSQC NMR experiments were acquired over the
course of a reciprocal titration of RAGE VC1 into a solution of15N-enriched Ca2+-S100B. The effects observed in these
spectra were similar to those seen for the S100B titration into
the V domain (i.e., differential line broadening for a specific set
of S100B resonances). The residues affected include His42,
Val52, Asn62, Asp69, Phe70, and Ala78, which cluster to a
specific surface on S100B. We note that this surface, and in
particular His42, Val52, and Ala78, were shown previously to
be important for interactions with other S100B-specific ligands
such as NDR kinase (Bhattacharya et al., 2003).
Because residues that are selectively perturbed upon forma-
tion of the complex could be identified for both RAGE-V and
S100B, we proceeded to generate a model of the complex.
The program HADDOCK (de Vries et al., 2007; Dominguez
et al., 2003) was used for these calculations. A representative
structure was selected on the basis of the lowest HADDOCK
score and is shown in Figures 5D–5F. The models show S100B
uses a concave, negatively charged surface to clamp around
a complementary positively charged face of the RAGE V domain
(Figure 5E). A surface area greater than 1400 A2 is buried by
formation of the complex. Analysis of the model revealed a
number of possible polar hydrogen bonds and salt bridges in
the interface. Thus, the model suggests that clustering of basic
residues on the binding surface of VC1 and highly acidic surface
of S100B are the major driving forces in formation of the
All rights reserved

Figure 4. Self Association of RAGE VC1Monitored by Dynamic Light
Scattering
(A) VC1 exists as a monomer at pH 5.2.
(B and C) Increase of pH to 6.5 and 7.5 as well as the addition of Zn2+ leads to
a shift toward oligomeric states.
(D) VC1 oligomer formation at pH 7.6 was dependent on protein concentration
(-) and is strongly increased by the presence of Zn2+ (C). The Zn2+ and
concentration-dependent oligomerization was fit to a hyperbolic function
yielding an apparent dissociation constant (Kdapp) of 3.8 ± 1.3 mM.
See also Figure S2.
Structure
Structure of Human RAGE
Structure 18, 1342–1
complex. In order to validate the result obtained by HADDOCK,
we performed a second set of calculations without NMR
restraints using the computational docking program HEX
(Ritchie and Kemp, 2000; Ritchie et al., 2008). The models
obtained in this manner were remarkably similar to the models
obtained using the NMR restraints in HADDOCK (Figure S3).
Analysis of these models of the VC1-S100B complex reveals
a strong electrostatic component to binding, consistent with
the proposed electrostatic trap contributing significantly to
RAGE ligand binding.
DISCUSSION
Initiation of signal cascades by ligand-induced receptor oligo-
merization has been proposed as a general mechanism of
receptor activation. In some models, multimeric ligands activate
their receptors by recruitment of receptor molecules. Hence,
association of the extracellular ligand-binding domains drives
the colocalization of the cytoplasmic domains, which is needed
for signal transduction. In these models, the geometry of the
ligand will control activity because the cytoplasmic domains
need to be brought into close proximity in a specific orientation.
This model has been invoked for receptor tyrosine kinases (Yu-
zawa et al., 2007). However, it has also been shown that many
receptors, including TNFa-receptor (Chan, 2007; Chan et al.,
2000), interleukin-receptor (Kramer et al., 2006), EPO receptor
(Livnah et al., 1999), and EGF receptor (Stuttfeld and Ballmer-
Hofer, 2009; Yang et al., 2010), preassemble in the absence of
a ligand on the cell surface. In these cases, it is proposed that
the clustering of receptor molecules increases the affinity for
ligands and is a requirement for effective receptor signaling. It
is notable that the distance of two adjacent VC1 molecules in
the crystal structure is 20–25 A, close to the distance between
subunits in the receptor tyrosine kinase complex (Yuzawa
et al., 2007) and predicted in the active EGF receptor complex
(Burgess et al., 2003). Consistent with a recent FRET study (Xie
et al., 2008), our data suggest a similar preassembly model for
RAGE.
Preassembly has substantial implications for the mechanism
of RAGE activation by its diverse ligands. The molecular basis
for RAGE activation by its diverse set of ligands has remained
enigmatic; RAGE ligands exhibit different structure, size, and
symmetry or even no symmetry, as in the case of glycated
proteins or amyloid-b. Clearly, single copies of dissimilar ligands
cannot induce a regular arrangement of the intracellular receptor
domains as required for initiation of a signal cascade. The
common factor linking these ligands is their tendency to oligo-
merize. The structural and computational data provided here
strongly imply a critical role for the positive electrostatic potential
in ligand recognition and binding.
If RAGE assembles on the cell surface in the absence of a
ligand, ligand binding can be viewed as shifting the equilibrium
distribution between monomer and higher order oligomerization
states (Figure 6A). Thus, receptor assemblies would be stabi-
lized by the binding of multimeric ligands such as S100B,
S100A12, or polymeric glycated proteins. This explains why
RAGE activation by such ligands leads to a positive feedback
cycle to sustain elevated RAGE expression. Moreover, because
increased receptor levels at the cell surface will promote
352, October 13, 2010 ª2010 Elsevier Ltd All rights reserved 1347

Figure 5. Structural Model of the S100B-VC1 Complex
(A) Surface representation of VC1 with the electrostatic potential mapped on the surface. Blue represents positively charged areas, and red represents negative
charge.
(B) Overlay of the 600 MHz 15N-1H HSQC NMR spectrum of 15N-enriched V domain in the absence (black) and presence (red) of Ca2+-S100B.
(C) VC1 residues identified in (B) mapped on the VC1 structure. The V domain is colored in green, the C1 domain in light blue, and residues perturbed in the NMR
experiments are colored yellow.
(D) Docking of S100B on VC1 based on the chemical shift perturbations identified in (B).
(E) Surface representations of the structural model of the S100B-VC1 complex demonstrating the contributions of charge complementarity to binding. The
left-hand panel shows S100B engaged with VC1. The right-hand panel pulls apart the complex to reveal the intense negative potential of the binding face of
S100B and the intense positive potential of the binding face of VC1.
(F) Structural model of the S100B-VC1 complex with the S100B ribbon and VC1 displayed with the electrostatic potential surface.
See also Figure S3.
Structure
Structure of Human RAGE
preassembly, this would explain the hyperactivation of the RAGE
pathway in the development of chronic inflammatory and neuro-
degenerative disorders.
The importance of RAGE oligomerization is underscored by
a number of functional studies. A fusion of the RAGE cytoplasmic
domains to the C terminus of the homodimeric glutathione
S-transferase (GST) was able to bind the cytoplasmic RAGE
mediators Diaphanous-1 (Hudson et al., 2008b) and ERK kinase
(Ishihara et al., 2003) in vitro, implying that theGST-fusion protein
mimics the active receptor. Because the C termini in the GST
1348 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
homodimer are located in the same orientation and at a distance
of approximately 25 A, these observations imply the cytoplasmic
domains in the fusion are separated by the same distance as in
the activated receptor.
Although preassembly of RAGE might facilitate ligand binding
and increase its efficiency in signaling, preassembled receptors
could promote nonspecific activation with adverse conse-
quences. Therefore, preassembly of RAGE must be tightly regu-
lated. The major alternative splice product of RAGE comprises
the extracellular region and occurs as soluble protein (sRAGE)
All rights reserved

Figure 6. Models of RAGE Activation and Inhibition by
sRAGE
(A) Model in which RAGE preassembles in the plasma
membrane. Ligand binding to RAGE stabilizes oligomers,
which then can bind a signaling adaptor protein (gray sphere)
to the cytoplasmic region of RAGE.
(B) Diagram showing the action of sRAGE, in which interaction
with intact RAGE to form a hetero-oligomer limits binding of
intracellular adaptors and blocks signal transduction.
Structure
Structure of Human RAGE
(Hudson et al., 2008a), which represents an adaptable regulator
of RAGE activation. It has been proposed that soluble alternative
splice products of RAGE can act as decoy receptors, decreasing
the concentration of available ligands. Our data suggest a
different explanation for how sRAGE might interfere with RAGE
activation: both sRAGE and the splice variant lacking only the
intracellular domain (DN-RAGE) can form heterocomplexes
with full-length RAGE, resulting in nonfunctional assemblies
(Figure 6B). This notion is largely supported by the observation
that coexpression of a RAGE isoform devoid of its intracellular
domain has a dominant negative effect on RAGE signaling (Hut-
tunen et al., 1999; Taguchi et al., 2000). Dominant inhibition of
signaling by heterocomplex formation has also been observed
for tumor necrosis factor–related apoptosis-inducing ligand
receptor (Clancy et al., 2005) and Fas death receptor (Siegel
et al., 2000), where coexpression of truncated forms of the
receptors blocked signaling of full-length receptors by ligand-
independent association.
RAGE is a critically important mediator of the inflammatory
response and plays an important role in innate immunity. Thera-
peutic approaches to target RAGE will be valuable for the
treatment of diabetic complications, chronic inflammations,
and neurodegenerative disorders. The structure of VC1 and
NMR-based models of the VC1-S100B complex provide
detailed insights into the mechanism of ligand recognition and
binding, as well as how RAGE can be activated by the diverse
Structure 18, 1342–1352, October
range of ligands. Both structural and biochemical
data support the proposal that RAGE assembles
in the cytoplasmic membrane without the presence
of a ligand. These results define approaches to
block ligand binding or reduce self-association,
which represent a promising avenue for the pursuit
of therapeutic strategies based on interfering with
RAGE activation.
EXPERIMENTAL PROCEDURES
Protein Expression and Purification
Protein encompassing the V and C1 domains of RAGE and
S100 proteins were expressed in Escherichia coli and purified
as described previously (Allmen et al., 2008; Dattilo et al.,
2007; Ostendorp et al., 2005).
Crystallization, Data Collection, and Structure
Determination
Crystals were grown by hanging drop vapor diffusion at 298 K
bymixing 2 ml of protein with 2 ml of 0.1MNa cacodylate, 0.2M
Zn acetate (pH 6.5), and 11% PEG 8000. Crystals of a size
of 200 mm 3 100 mm 3 200 mm appeared after 10 to
20 days. The crystals were soaked for 1 min in three consec-
utive steps in mother liquor containing 4%, 8%, and 12% glycerol and were
flash-frozen in liquid nitrogen. Crystals diffracted to a resolution of 1.85 A.
Because Zn2+ turned out to be essential for crystallization, we assumed that
Zn2+ specifically interacts with RAGE VC1 and could, therefore, be used for
experimental phasing. Multiple anomalous dispersion data were collected at
a wavelength of 1.28 A at the Swiss Light Source (Villigen, Switzerland) beam-
line X06SA at 100 K. An additional data set collected at a wavelength of 1.0 A
was used for refinement. The data sets were reduced and scaled with the XDS
package (Kabsch, 1993). Crystals belonged to the space group P21212 with
unit cell parameters of 74.85, 119.96, and 28.89, as determined by XPREP
(2005 version, Bruker-AXS). The zinc substructure was determined using
SHELXD (Schneider and Sheldrick, 2002). Refinement of the zinc sites and
phase calculation was done with SHARP (Fortelle and Bricogne, 1997)
followed by solvent flattening and initial fragment building using RESOLVE
(Terwilliger, 2000). The final model was built in COOT (Emsley and Cowtan,
2004) and was refined using REFMAC5 (Murshudov et al., 1997; Winn et al.,
2001) with final Rwork and Rfree of 20.9% and 24.0%, respectively (Table 1).
Ninety-nine percent of the residues were located in the allowed regions, and
1% were located in the generously allowed regions of the Ramachandran
plot (Table 1).
Solution NMR Mapping Studies with Ca2+-Loaded S100B
Substoichiometric additions of Ca2+-loaded S100B were made into uniformly
labeled 15N-V in 10 mM Imidazole-d4 (pH 6.5), 100 mMNaCl, and 5mMCaCl2.
Similar additions of VC1 were made into a uniform solution of 15N-labeled
S100B in 20 mM KCl (pH 7.1), 2.5 mM Ca2+, and 5% D2O. Perturbations in
the NMR signals were monitored by 2D 15N-1H HSQC spectra using a Bruker
DRX500 spectrometer. Data were processed using Topspin 2.0b (Bruker) and
analyzed using NMRViewJ. The intensities (I) were extrapolated from peak
heights and plotted as a ratio (I/Io) versus the initial intensity (Io).
13, 2010 ª2010 Elsevier Ltd All rights reserved 1349

Structure
Structure of Human RAGE
Protein Oligomerization Assay
Protein oligomerization assays of VC1 were performed at 298 K. A stock
solution of VC1 with 10 mg ml�1 in 10 mM sodium acetate buffer (pH 5.0)
was diluted into 20 mM Tris and 150 mM NaCl (pH 7.6). Large oligomers
formed at increasing VC1 concentrations and in the presence of Zn2+. The
oligomers were pelleted by centrifugation (10 min at 12,000 g). The amount
of oligomer formed was calculated by subtracting the amount of protein
remaining in the supernatant from total protein. VC1 concentration in the
supernatant was determined by the UV absorbance using an extinction coef-
ficient of 3278nm = 32500 M�1 cm�1. In experiments monitoring the effect of
Zn2+ ions, 200 mM ZnCl2 was present in the buffer.
Docking Calculations
Residues exhibiting significant NMR chemical shift perturbations were used
as input for HADDOCK calculations (Dominguez et al., 2003) to generate an
experimentally derived model of the S100B-VC1 complex. Specifically, those
residues in S100B and in the V-domain of RAGE whose intensities were
reduced by more than one standard deviation from the mean reduction in
intensity were selected. The active and passive categories of restraint were
assigned based on criteria outlined in HADDOCK2 (de Vries et al., 2007).
The calculation proceeded in a similar manner to that reported by Nordquist
et al. (2010). To improve the conformation sampling, twenty structures each
were generated from the X-ray crystal structures of S100B (PDB accession
number 2H61) and VC1 using room temperature MD simulations in AMBER 9
(Pearlman and Connelly, 1995). Using these starting structures, 1000 complexes
were generated from rigid-body docking, of which 200 were selected for further
refinement in explicit water. One hundred and four of the 200 structures fell into
11 clusters, of which the twowith the lowest target energieswere themost popu-
lated (cluster 1,25; cluster2,16).Cluster1had the lowestaverageHaddockscore
and highest average buried surface area, so the 20 structures with the lowest
energies were selected as the representative ensemble for further analysis.
Free docking calculations of Ca2+-loaded S100B with VC1 were performed
using HEX 5.0 (Mustard and Ritchie, 2005; Ritchie and Kemp, 2000) on amulti-
processor Linux workstation. HEX uses 3D parametric functions, which are
used to encode both surface shape and electrostatic charge and potential
distributions for docking calculations.
Modeling and Analysis
A homology model of the activated leukocyte adhesion molecule (ALCAM) Ig
domains 1 and 2 was constructed using VC1 as a template in MODELLER
(Eswar et al., 2007; Sali et al., 1995). Electrostatic potentials were calculated
with APBS (Baker et al., 2001). All molecular figures were generated using
PyMol (DeLano Scientific).
ACCESSION NUMBERS
Coordinates and structure-factor amplitudes have been deposited in the
Protein Data Bank with accession number 3CJJ.
SUPPLEMENTAL INFORMATION
Supplemental Information includes three figures and can be found with this
article online at doi:10.1016/j.str.2010.05.017.
ACKNOWLEDGMENTS
We thank thestaff at theSwissLightSourcebeamlineX06SA for excellent assis-
tance with data collection and Benjamin Chagot for technical expertise with
HADDOCK2.0.Financial supportwasprovidedby theDeutscheForschungsge-
meinschaft (grants FR 1488/3-1 and FR 1488/5-1 to G.F.) and the National
Institutes of Health (grant RO1 GM62112 to W.J.C.). S.C. was supported by
a Canadian Institutes of Health and Research postdoctoral fellowship.
Received: September 25, 2009
Revised: May 6, 2010
Accepted: May 15, 2010
Published: October 12, 2010
1350 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
REFERENCES
Allmen, E.U., Koch, M., Fritz, G., and Legler, D.F. (2008). V domain of RAGE
interacts with AGEs on prostate carcinoma cells. Prostate 68, 748–758.
Baker, N.A., Sept, D., Joseph, S., Holst, M.J., and McCammon, J.A. (2001).
Electrostatics of nanosystems: application to microtubules and the ribosome.
Proc. Natl. Acad. Sci. USA 98, 10037–10041.
Bhattacharya, S., Large, E., Heizmann, C.W., Hemmings, B., and Chazin, W.J.
(2003). Structure of the Ca2+/S100B/NDR kinase peptide complex: insights
into S100 target specificity and activation of the kinase. Biochemistry 42,
14416–14426.
Burgess, A.W., Cho, H.S., Eigenbrot, C., Ferguson, K.M., Garrett, T.P., Leahy,
D.J., Lemmon, M.A., Sliwkowski, M.X., Ward, C.W., and Yokoyama, S. (2003).
An open-and-shut case? Recent insights into the activation of EGF/ErbB
receptors. Mol. Cell 12, 541–552.
Casasnovas, J.M., Springer, T.A., Liu, J.H., Harrison, S.C., and Wang, J.H.
(1997). Crystal structure of ICAM-2 reveals a distinctive integrin recognition
surface. Nature 387, 312–315.
Chan, F.K. (2007). Three is better than one: pre-ligand receptor assembly in the
regulation of TNF receptor signaling. Cytokine 37, 101–107.
Chan, F.K., Chun, H.J., Zheng, L., Siegel, R.M., Bui, K.L., and Lenardo, M.J.
(2000). A domain in TNF receptors that mediates ligand-independent receptor
assembly and signaling. Science 288, 2351–2354.
Clancy, L., Mruk, K., Archer, K., Woelfel, M., Mongkolsapaya, J., Screaton, G.,
Lenardo, M.J., and Chan, F.K. (2005). Preligand assembly domain-mediated
ligand-independent association between TRAIL receptor 4 (TR4) and TR2
regulates TRAIL-induced apoptosis. Proc. Natl. Acad. Sci. USA 102, 18099–
18104.
Dattilo, B.M., Fritz, G., Leclerc, E., Vander Kooi, C.W., Heizmann, C.W., and
Chazin, W.J. (2007). The extracellular region of the receptor for advanced
glycation end products is composed of two independent structural units.
Biochemistry 46, 6957–6970.
de Vries, S.J., van Dijk, A.D., Krzeminski, M., van Dijk, M., Thureau, A., Hsu, V.,
Wassenaar, T., and Bonvin, A.M. (2007). HADDOCK versus HADDOCK: new
features and performance of HADDOCK2.0 on the CAPRI targets. Proteins
69, 726–733.
Deane, R., Du Yan, S., Submamaryan, R.K., LaRue, B., Jovanovic, S., Hogg,
E., Welch, D., Manness, L., Lin, C., Yu, J., et al. (2003). RAGE mediates
amyloid-b peptide transport across the blood-brain barrier and accumulation
in brain. Nat. Med. 9, 907–913.
Dominguez, C., Boelens, R., and Bonvin, A.M. (2003). HADDOCK: a protein-
protein docking approach based on biochemical or biophysical information.
J. Am. Chem. Soc. 125, 1731–1737.
Emanuele, E., D’Angelo, A., Tomaino, C., Binetti, G., Ghidoni, R., Politi, P.,
Bernardi, L., Maletta, R., Bruni, A.C., and Geroldi, D. (2005). Circulating levels
of soluble receptor for advanced glycation end products in Alzheimer disease
and vascular dementia. Arch. Neurol. 62, 1734–1736.
Emsley, P., and Cowtan, K. (2004). Coot: model-building tools for molecular
graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132.
Eswar, N., Webb, B., Marti-Renom, M.A., Madhusudhan, M.S., Eramian, D.,
Shen, M.Y., Pieper, U., and Sali, A. (2007). Comparative protein structure
modeling using MODELLER. Curr. Protoc. Protein Sci., Chapter 2, Unit 2 9.
Feinberg, H., Mitchell, D.A., Drickamer, K., and Weis, W.I. (2001). Structural
basis for selective recognition of oligosaccharides by DC-SIGN and
DC-SIGNR. Science 294, 2163–2166.
Fortelle, E.l., and Bricogne, G. (1997). Maximum-likelihood heavy-atom
parameter refinement for multiple isomorphous replacement and multiwave-
length anomalous diffraction methods. Methods Enzymol. 276, 472–494.
Freigang, J., Proba, K., Leder, L., Diederichs, K., Sonderegger, P., and Welte,
W. (2000). The crystal structure of the ligand binding module of axonin-1/TAG-
1 suggests a zipper mechanism for neural cell adhesion. Cell 101, 425–433.
Ghidoni, R., Benussi, L., Glionna, M., Franzoni, M., Geroldi, D., Emanuele, E.,
and Binetti, G. (2008). Decreased plasma levels of soluble receptor for
All rights reserved

Structure
Structure of Human RAGE
advanced glycation end products in mild cognitive impairment. J. Neural
Transm. 115, 1047–1050.
Harpaz, Y., and Chothia, C. (1994). Many of the immunoglobulin superfamily
domains in cell adhesion molecules and surface receptors belong to a new
structural set which is close to that containing variable domains. J. Mol. Biol.
238, 528–539.
Hofmann, M.A., Drury, S., Fu, C., Qu, W., Taguchi, A., Lu, Y., Avila, C.,
Kambham, N., Bierhaus, A., Nawroth, P., et al. (1999). RAGE mediates a novel
proinflammatory axis: a central cell surface receptor for S100/calgranulin
polypeptides. Cell 97, 889–901.
Holden, H.M., Ito, M., Hartshorne, D.J., and Rayment, I. (1992). X-ray structure
determination of telokin, the C-terminal domain of myosin light chain kinase, at
2.8 A resolution. J. Mol. Biol. 227, 840–851.
Hori, O., Brett, J., Slattery, T., Cao, R., Zhang, J., Chen, J.X., Nagashima, M.,
Lundh, E.R., Vijay, S., and Nitecki, D. (1995). The receptor for advanced
glycation end products (RAGE) is a cellular binding site for amphoterin: medi-
ation of neurite outgrowth and co-expression of rage and amphoterin in the
developing nervous system. J. Biol. Chem. 270, 25752–25761.
Hudson, B.I., Carter, A.M., Harja, E., Kalea, A.Z., Arriero, M., Yang, H., Grant,
P.J., and Schmidt, A.M. (2008a). Identification, classification, and expression
of RAGE gene splice variants. FASEB J. 22, 1572–1580.
Hudson, B.I., Kalea, A.Z., Del Mar Arriero, M., Harja, E., Boulanger, E., D’Agati,
V., and Schmidt, A.M. (2008b). Interaction of the RAGE cytoplasmic domain
with diaphanous-1 is required for ligand-stimulated cellular migration through
activation of Rac1 and Cdc42. J. Biol. Chem. 283, 34457–34468.
Huttunen, H.J., Fages, C., and Rauvala, H. (1999). Receptor for advanced
glycation end products (RAGE)-mediated neurite outgrowth and activation of
NF-kappaB require the cytoplasmic domain of the receptor but different
downstream signaling pathways. J. Biol. Chem. 274, 19919–19924.
Huttunen, H.J., Kuja-Panula, J., Sorci, G., Agneletti, A.L., Donato, R., and
Rauvala, H. (2000). Coregulation of neurite outgrowth and cell survival by
amphoterin and S100 proteins through receptor for advanced glycation end
products (RAGE) activation. J. Biol. Chem. 275, 40096–40105.
Ishihara, K., Tsutsumi, K., Kawane, S., Nakajima, M., and Kasaoka, T. (2003).
The receptor for advanced glycation end-products (RAGE) directly binds to
ERK by a D-domain-like docking site. FEBS Lett. 550, 107–113.
Kabsch, W. (1993). Automatic processing of rotation diffraction data from
crystals of initially unknown symmetry and cell constants. J. Appl. Cryst. 26,
795–800.
Kasper, C., Rasmussen, H., Kastrup, J.S., Ikemizu, S., Jones, E.Y., Berezin, V.,
Bock, E., and Larsen, I.K. (2000). Structural basis of cell-cell adhesion by
NCAM. Nat. Struct. Mol. Biol. 7, 389–393.
Kramer, J.M., Yi, L., Shen, F., Maitra, A., Jiao, X., Jin, T., and Gaffen, S.L.
(2006). Evidence for ligand-independent multimerization of the IL-17 receptor.
J. Immunol. 176, 711–715.
Leclerc, E., Fritz, G., Weibel, M., Heizmann, C.W., and Galichet, A. (2007).
S100B and S100A6 differentially modulate cell survival by interacting with
distinct RAGE (receptor for advanced glycation end products) immunoglobulin
domains. J. Biol. Chem. 282, 31317–31331.
Li, H., Zhao, Y., Guo, Y., Li, Z., Eisele, L., and Mourad, W. (2007). Zinc induces
dimerization of the class II major histocompatibility complex molecule that
leads to cooperative binding to a superantigen. J. Biol. Chem. 282, 5991–6000.
Livnah, O., Stura, E.A., Middleton, S.A., Johnson, D.L., Jolliffe, L.K., and
Wilson, I.A. (1999). Crystallographic evidence for preformed dimers of erythro-
poietin receptor before ligand activation. Science 283, 987–990.
Matsumoto, S., Yoshida, T., Murata, H., Harada, S., Fujita, N., Nakamura, S.,
Yamamoto, Y., Watanabe, T., Yonekura, H., Yamamoto, H., et al. (2008).
Solution structure of the variable-type domain of the receptor for advanced
glycation end products: new insight into AGE-RAGE interaction. Biochemistry
47, 12299–12311.
Murshudov, G.N., Vagin, A.A., and Dodson, E.J. (1997). Refinement of macro-
molecular structures by the maximum-likelihood method. Acta Crystallogr.
D Biol. Crystallogr. 53, 240–255.
Structure 18, 1342–1
Mustard, D., and Ritchie, D.W. (2005). Docking essential dynamics eigenstruc-
tures. Proteins 60, 269–274.
Neeper, M., Schmidt, A.M., Brett, J., Yan, S.D., Wang, F., Pan, Y.C., Elliston,
K., Stern, D., and Shaw, A. (1992). Cloning and expression of a cell surface
receptor for advanced glycosylation end products of proteins. J. Biol. Chem.
267, 14998–15004.
Nordquist, K.A., Dimitrova, Y.N., Brzovic, P.S., Ridenour, W.B., Munro, K.A.,
Soss, S.E., Caprioli, R.M., Klevit, R.E., and Chazin, W.J. (2010). Structural
and functional characterization of the monomeric U-box domain from E4B.
Biochemistry 49, 347–355.
Orlova, V.V., Choi, E.Y., Xie, C., Chavakis, E., Bierhaus, A., Ihanus, E.,
Ballantyne, C.M., Gahmberg, C.G., Bianchi, M.E., Nawroth, P.P., and
Chavakis, T. (2007). A novel pathway of HMGB1-mediated inflammatory cell
recruitment that requires Mac-1-integrin. EMBO J. 26, 1129–1139.
Ostendorp, T., Heizmann, C.W., Kroneck, P.M., and Fritz, G. (2005).
Purification, crystallization and preliminary X-ray diffraction studies on human
Ca2+-binding protein S100B. Acta Crystallogr. Sect. F Struct. Biol. Cryst.
Commun. 61, 673–675.
Ostendorp, T., Leclerc, E., Galichet, A., Koch, M., Demling, N., Weigle, B.,
Heizmann, C.W., Kroneck, P.M., and Fritz, G. (2007). Structural and functional
insights into RAGE activation by multimeric S100B. EMBO J. 26, 3868–3878.
Park, L., Raman, K.G., Lee, K.J., Lu, Y., Ferran, L.J., Jr., Chow,W.S., Stern, D.,
and Schmidt, A.M. (1998). Suppression of accelerated diabetic atheroscle-
rosis by the soluble receptor for advanced glycation endproducts. Nat. Med.
4, 1025–1031.
Pearlman, D.A., and Connelly, P.R. (1995). Determination of the differential
effects of hydrogen bonding and water release on the binding of FK506 to
native and Tyr82/Phe82 FKBP-12 proteins using free energy simulations.
J. Mol. Biol. 248, 696–717.
Ritchie, D.W., and Kemp, G.J. (2000). Protein docking using spherical polar
Fourier correlations. Proteins 39, 178–194.
Ritchie, D.W., Kozakov, D., and Vajda, S. (2008). Accelerating and focusing
protein-protein docking correlations using multi-dimensional rotational FFT
generating functions. Bioinformatics 24, 1865–1873.
Sali, A., Potterton, L., Yuan, F., van Vlijmen, H., and Karplus, M. (1995).
Evaluation of comparative protein modeling by MODELLER. Proteins 23,
318–326.
Schmidt, A.M., Vianna, M., Gerlach, M., Brett, J., Ryan, J., Kao, J., Esposito,
C., Hegarty, H., Hurley, W., and Clauss, M. (1992). Isolation and characteriza-
tion of two binding proteins for advanced glycosylation end products from
bovine lung which are present on the endothelial cell surface. J. Biol. Chem.
267, 14987–14997.
Schneider, T.R., and Sheldrick, G.M. (2002). Substructure solution with
SHELXD. Acta Crystallogr. D Biol. Crystallogr. 58, 1772–1779.
Siegel, R.M., Frederiksen, J.K., Zacharias, D.A., Chan, F.K., Johnson, M.,
Lynch, D., Tsien, R.Y., and Lenardo, M.J. (2000). Fas preassociation required
for apoptosis signaling and dominant inhibition by pathogenic mutations.
Science 288, 2354–2357.
Sternberg, Z., Weinstock-Guttman, B., Hojnacki, D., Zamboni, P., Zivadinov,
R., Chadha, K., Lieberman, A., Kazim, L., Drake, A., Rocco, P., et al. (2008).
Soluble receptor for advanced glycation end products in multiple sclerosis:
a potential marker of disease severity. Mult. Scler. 14, 759–763.
Stuttfeld, E., and Ballmer-Hofer, K. (2009). Structure and function of VEGF
receptors. IUBMB Life 61, 915–922.
Su, X.D., Gastinel, L.N., Vaughn, D.E., Faye, I., Poon, P., and Bjorkman, P.J.
(1998). Crystal structure of hemolin: a horseshoe shape with implications for
homophilic adhesion. Science 281, 991–995.
Taguchi, A., Blood, D.C., del Toro, G., Canet, A., Lee, D.C., Qu, W., Tanji, N.,
Lu, Y., Lalla, E., Fu, C., et al. (2000). Blockade of RAGE-amphoterin signalling
suppresses tumour growth and metastases. Nature 405, 354–360.
Takeuchi, K., andWagner, G. (2006). NMR studies of protein interactions. Curr.
Opin. Struct. Biol. 16, 109–117.
Terwilliger, T.C. (2000). Maximum-likelihood density modification. Acta
Crystallogr. D Biol. Crystallogr. 56, 965–972.
352, October 13, 2010 ª2010 Elsevier Ltd All rights reserved 1351

Structure
Structure of Human RAGE
Tian, J., Avalos, A.M., Mao, S.Y., Chen, B., Senthil, K., Wu, H., Parroche, P.,
Drabic, S., Golenbock, D., Sirois, C., et al. (2007). Toll-like receptor 9-depen-
dent activation byDNA-containing immune complexes ismediated byHMGB1
and RAGE. Nat. Immunol. 8, 487–496.
Wang, J.H., Smolyar, A., Tan, K., Liu, J.H., Kim, M., Sun, Z.Y., Wagner, G., and
Reinherz, E.L. (1999). Structure of a heterophilic adhesion complex between
the human CD2 and CD58 (LFA-3) counterreceptors. Cell 97, 791–803.
Winn,M.D., Isupov,M.N., andMurshudov, G.N. (2001). Use of TLS parameters
to model anisotropic displacements in macromolecular refinement. Acta
Crystallogr. D Biol. Crystallogr. 57, 122–133.
Wittkowski, H., Hirono, K., Ichida, F., Vogl, T., Ye, F., Yanlin, X., Saito, K., Uese,
K., Miyawaki, T., Viemann, D., et al. (2007). Acute Kawasaki disease is associ-
ated with reverse regulation of soluble receptor for advance glycation end
products and its proinflammatory ligand S100A12. Arthritis Rheum. 56,
4174–4181.
Xie, J., Burz, D.S., He, W., Bronstein, I.B., Lednev, I., and Shekhtman, A.
(2007). Hexameric calgranulin C (S100A12) binds to the receptor for advanced
glycated end products (RAGE) using symmetric hydrophobic target-binding
patches. J. Biol. Chem. 282, 4218–4231.
Xie, J., Reverdatto, S., Frolov, A., Hoffmann, R., Burz, D.S., and Shekhtman, A.
(2008). Structural basis for pattern recognition by the receptor for advanced
glycation end products (RAGE). J. Biol. Chem. 283, 27255–27269.
1352 Structure 18, 1342–1352, October 13, 2010 ª2010 Elsevier Ltd
Xu, Z., and Jin, B. (2010). A novel interface consisting of homologous immuno-
globulin superfamily members with multiple functions. Cell. Mol. Immunol. 7,
11–19.
Yamagishi, S., Takeuchi, M., Inagaki, Y., Nakamura, K., and Imaizumi, T.
(2003). Role of advanced glycation end products (AGEs) and their receptor
(RAGE) in the pathogenesis of diabetic microangiopathy. Int. J. Clin. Pharma-
col. Res. 23, 129–134.
Yan, S.S., Wu, Z.Y., Zhang, H.P., Furtado, G., Chen, X., Yan, S.F., Schmidt,
A.M., Brown, C., Stern, A., LaFaille, J., et al. (2003). Suppression of experi-
mental autoimmune encephalomyelitis by selective blockade of encephalito-
genic T-cell infiltration of the central nervous system. Nat. Med. 9, 287–293.
Yang, Y., Xie, P., Opatowsky, Y., and Schlessinger, J. (2010). Direct contacts
between extracellular membrane-proximal domains are required for VEGF
receptor activation and cell signaling. Proc. Natl. Acad. Sci. USA 107, 1906–
1911.
Yuzawa, S., Opatowsky, Y., Zhang, Z., Mandiyan, V., Lax, I., and Schlessinger,
J. (2007). Structural basis for activation of the receptor tyrosine kinase KIT by
stem cell factor. Cell 130, 323–334.
Zou, P., Pinotsis, N., Lange, S., Song, Y.H., Popov, A., Mavridis, I., Mayans,
O.M., Gautel, M., and Wilmanns, M. (2006). Palindromic assembly of the giant
muscle protein titin in the sarcomeric Z-disk. Nature 439, 229–233.
All rights reserved





























