Embed Size (px)
Citation preview

4/2008 4/2008
HO
NG
LI Structural and Functional Roles of K
CC
2 in Developing C
ortex
Structural and Functional Roles of KCC2 in Developing Cortex
Dissertationes bioscientiarum molecularium Universitatis Helsingiensis in Viikki
HONG LI
Institute of Biotechnology and
Department of Biological and Environmental SciencesFaculty of BiosciencesUniversity of Helsinki
Recent Publications in this Series:
15/2007 Päivi RamuOuter Membrane Protease/adhesin PgtE of S. enterica: Role in Salmonella-Host Interaction16/2007 Joni AlvesaloDrug Discovery Screening and the Application of Genomics and Proteomics in the Drug Development Process for Chlamydia pneumoniae17/2007 Tomi JukkolaFGFR1 Regulated Gene-Expression, Cell Proliferation and Differentiation in the Developing Midbrain and Hindbrain18/2007 Maarit HellmanStructural Biology of the ADF-H Domains19/2007 Rasa Gabrenaite-VerkhovskayaMovement-Associated Proteins of Potato Virus A: Attachment to Virus Particles and Phosphorylation20/2007 Henna ViholaStudies on Thermosensitive Poly(N-Vinylcaprolactam) Based Polymers for Pharmaceutical Applications21/2007 Terhi HakalaCharacterization of the Lignin-Modifying Enzymes of the Selective White-Rot Fungus Physisporinus Rivulosus22/2007 Ilya BelevichProton Translocation Coupled to Electron Transfer Reactions in Terminal Oxidases23/2007 Johan PahlbergSpectral Tuning and Adaptation to Different Light Environments of Mysid Visual Pigments24/2007 Beata Kluczek-TurpeinenLignocellulose Degradation and Humus Modifi cation by the Fungus Paecilomyces infl atus25/2007 Sabiruddin MirzaCrystallization as a Tool for Controlling and Designing Properties of Pharmaceutical Solids26/2007 Kaisa MarjamaaPeroxidases in Lignifying Xylem of Norway Spruce, Scots Pine and Silver Birch27/2007 Pekka NieminenMolecular Genetics of Tooth Agenesis28/2007 Sanna KoutaniemiLignin Biosynthesis in Norway Spruce: from a Model System to the Tree29/2007 Anne RantalaEvolution and Detection of Cyanobacterial Hepatotoxin Synthetase Genes30/2007 Tiina SikanenSU-8-Based Microchips for Capillary Electrophoresis and Electrospray Ionization Mass Spectrometry31/2007 Pieta MattilaMissing-In-Metastasis (MIM)Regulates Cell Morphology by Promoting Plasma Membrane and Actin Cytoskeleton Dynamics32/2007 Justus ReunanenLantibiotic Nisin and Its Detection Methods33/2007 Anton ShmelevFolding and Selective Exit of Reporter Proteins from the Yeast Endoplasmic Reticulum1/2008 Elina JääskeläinenAssessment and Control of Bacillus cereus Emetic Toxin in Food2/2008 Samuli HirsjärviPreparation and Characterization of Poly(Lactic Acid) Nanoparticles for Pharmaceutical Use3/2008 Kati HakalaLiquid Chromatography-Mass Spectrometry in Studies of Drug Metabolism and Permeability
Helsinki 2008 ISSN 1795-7079 ISBN 978-952-10-4489-2

Structural and Functional Roles of KCC2 in
Developing Cortex
HONG LI
Institute of Biotechnologyand
Department of Biological and Environmental SciencesFaculty of Biosciences
andFinnish Graduate School in Neuroscience
University of Helsinki
ACADEMIC DISSERTATION
To be presented, with the permission of the Faculty of Biosciences of the University of Helsinki, for public examination in auditorium 1041
at Biocenter 2 (Viikinkaari 7, Helsinki), on 22nd February 2008, at 12 noon.

Supervised by:
Docent Claudio Rivera, Ph.DInstitute of Biotechnology
University of HelsinkiFinland
and
Professor Kai Kaila, Ph.DDepartment of Biological and Environmental Sciences
Faculty of BioscienceUniversity of Helsinki
Reviewed by
Professor Dan Lindholm, MD, Ph.DMinerva Institute for Medical Research
BIOMEDICUM HelsinkiUniversity of Helsinki
and
Professor Kid Törnquist, Ph.D.Department of Biology
Åbo Akademi University
Opponent:
Professor Karl Åkerman, MD, Ph.DBiomedicum HelsinkiUniversity of Helsinki
ISBN 978-952-10-4489-2 (paperback)ISBN 978-952-10-4493-9 (PDF, http://ethesis.helsinki.fi )
ISSN 1795-7079 (paperback)ISSN 1795-8229 (PDF, http://ethesis.helsinki.fi )
Edita Prima OyHelsinki 2008

To Chunlin

Index:Abbreviations
List of original publications and manuscripts
Summary ....................................................................................................................... 9
Review of the literature .............................................................................................. 111. Neuronal network activity in developing cortex .................................................... 11 1.1 Neuronal transmitters in the central nervous system ....................................... 11 1.2 Synaptic properties of immature neurons ........................................................ 11 1.3 Network driven spontaneous activity in developing hippocampus ................. 12 1.4 Developmental roles of spontaneous activity .................................................. 132. GABAergic and glycinergic neurotransmission .................................................... 14 2.1 GABAA receptors ............................................................................................. 14 2.2 Glycine receptors ............................................................................................. 15 2.3 Developmental roles of GABAergic and glycinergic neurotransmission ........ 153. Cellular chloride homeostasis ................................................................................ 16 3.1 KCC1 ............................................................................................................... 18 3.2 KCC2 ............................................................................................................... 18 3.2.1 KCC2 and neuronal excitability .............................................................. 19 3.2.2 The regulation of KCC2 expression and activation ................................ 20 3.3 KCC3 ............................................................................................................... 21 3.4 KCC4 ............................................................................................................... 22 3.5 NKCC1 ............................................................................................................ 234. Glutamatergic neurotransmission .......................................................................... 24 4.1 Development of glutamatergic neurotransmission ......................................... 24 4.2 AMPA receptor interacting proteins ................................................................ 25 4.2.1 Proteins containing PDZ domains .......................................................... 25 4.2.2 Non-PDZ containing proteins ..................................................................26 4.3 Dendritic spines ............................................................................................... 27 4.3.1 Spinogenesis ........................................................................................... 28 4.3.2 Molecular mechanisms regulating dentritic spine morphology ............. 295. Excitatory and inhibitory synaptogenesis ................................................................ 30

Aims of the study ......................................................................................................... 33
Materials and methods ............................................................................................... 34
Results and discussion ................................................................................................ 351. Developmental expression of KCC2 follows neuronal maturation in vivo and in vitro .................................................................................................. 352. The role of KCC2 in spine formation .................................................................... 38 2.1 Aberrant morphology of dendritic spines in KCC2-/- cortical neurons and KCC2hy/null neurons ................................................................................... 38 2.2 Functional excitatory synapses are reduced in KCC2-/- neurons .................... 38 2.3 KCC2-ΔNTD lacking K-Cl transport activity restores normal spine morphology in KCC2-/- neurons ............................................... 39 2.4 Overexpression of KCC2 C-terminal domain in WT neurons induces elongation of dendritic protrusions ..................................................... 39 2.5 The interaction of KCC2-CTD with 4.1N underlies the function of KCC2 in spine morphology ..................................................... 403. Regulation of KCC2 expression during development and plasticity ..................... 41
Conclusions ................................................................................................................. 44
Acknowledgments ....................................................................................................... 45
Reference list ............................................................................................................... 46

Abbreviations aa amino acidACCPN Agenesis of the Corpus Callosum with Peripheral Neuropathy AE anion exchangerAMPA α-amino-3-hydroxy-5-methylisoxazole-4-propionic acidBDNF brain-derived neurotrophic factorCCC cation-chloride cotransporterCIP cation-chloride cotransporter interaction proteinCK creatine kinase CNS central nervous systemDRG dorsal root ganglionE embryonic dayE/I excitatory and inhibitory synapse ratioGABA γ-aminobutyric acidGDP Giant Depolarizing PotentialsGRIP glutamate receptor-interacting proteinIGF insulin growth factorKCC2-CTD K+-Cl- cotransporter isoform 2 C-terminalKCC K+-Cl- cotransporterKCC2-FL K+-Cl- cotransporter isoform 2 full lengthKCC2-ΔNTD K+-Cl- cotransporter isoform 2 N-terminal deletion LTP long-term potentialMAGUK membrane-associated guanylate kinasemGluR metabotropic glutamate receptorNKCC Na+-K+-2Cl- co-transportersNRSE neuronal-restrictive silencing elementNRSF neuron-restricted silencing factorNSF N-Ethylmaleimide-sensitive factorNT neurotrophic NCC Na-Cl cotransporterNEM N-ehyl-maleimideNMDA N-methyl-D-aspartateNR NMDA receptor subunitP postnatal

PDZ PSD-95/blg/ZO-1PIX Pak-interacting exchange factorPKA protein kinase APKC protein kinase CPLCγ-CREB phospholipase C gamma-cAMP response element-binding proteinPSD postsynaptic densityPTK protein tyrosine kinaseRVD regulatory volume decrease SAP synapse associated proteinShc/FRs-2 src homology 2 domain containing transforming protein/FGF receptor substrate 2 SPAK Ste20-related proline-alanine-rich kinaseTM transmembraneVGCC Voltage-gated calcium channelsWNK with no lysine (K) kinaseWT wild type4.1N protein 4.1 neuronal isoform

List of original publications and manuscripts
This thesis is based on the following original articles, which are referred to in the text by their Roman numerals (I-IV)
I Li H#, Tornberg J#, Kaila K, Airaksinen MS, Rivera C (2002) Patterns of cation-chloride cotransporter expression during embryonic rodent CNS development.
European Journal of Neuroscience 16: 2358-2370.
II Ludwig A, Li H, Saarma M, Kaila K, Rivera C (2003) Developmental up-regulation of KCC2 in the absence of GABAergic and glutamatergic transmission. European Journal of Neuroscience 18: 3199-3206.
III Li H, Khirug S#, Cai C#, Ludwig A, Blaesse P, Kolikova J, Afzalov R, Coleman SK, Lauri S, Airaksinen MS, Keinänen K, Khiroug L, Saarma M, Kaila K, Rivera C (2007) KCC2 interacts with the dendritic cytoskeleton to promote spine development.
Neuron 56: 1019-1033.
IV Rivera C, Voipio J, Thomas-Crusells J, Li H, Emri Z, Sipila S, Payne JA, Minichiello L, Saarma M, Kaila K (2004) Mechanism of activity-dependent downregulation of the neuron-specifi c K-Cl cotransporter KCC2.
Journal of Neuroscience 24: 4683-4691.
# denotes equal contribution

9
Summary
Summary
Cation chloride cotransporters (CCCs) are critical for controlling intracellular chloride homeostasis. They are involved in multiple cell functions including volume regulation, cell cycle control, and setting the reversal potential and the polarity of GABAA receptor-mediated currents. The CCC family is composed of four isoforms of K-Cl cotransporters (KCC1-4), two isoforms of Na-K-2Cl cotransporters (NKCC1-2), one Na-Cl cotransporter (NCC) and two structurally related proteins with unknown function, CCC8 also known as cation-chloride cotransporter interaction protein, CIP, and CCC9. KCC2 is a neuron-specifi c isoform, which plays a prominent role in controlling the intracellular Cl- concentration in neurons and is responsible for producing the negative shift of GABAA responses from depolarizing to hyperpolarizing during neuronal maturation. Transgenic mice lacking KCC2 do not survive after birth due to disturbance in respiratory rythmogenesis which points to a signifi cant role of KCC2 in CNS development and functions.
In the present studies we first used in situ hybridization to examine the developmental expression patterns of the cation-chloride cotransporters KCC1-4 and NKCC1. We found that they display complementary expression patterns during embryonic brain development. KCC1 mRNA was only detectable in the developing choroid plexus. KCC3 expression is lower in embryonic brain but is slightly up-regulated after birth. KCC4 is abundant in the ventricular zone but down-regulated perinatally. At E14.5, NKCC1 was highly expressed in proliferate zone of the subcortical region, but shift to the neuron specifi c class III β-
tubulin positive region of cortical plate at later embryonic stages. Most interestingly, KCC2 expression in the embryonic central nervous system strictly follows neuronal maturation and can be detected as earlier as E12.5, but only in the ventral part of spinal cord where the earliest neurogenesis takes place. The expression gradually spreads to higher brain regions like rhombencephalon, diencephalons, olfactory bulb and neocortical regions, in parallel with the maturation of these structures. In vitro data obtained from primary and organotypic neuronal cultures support this fi nding and revealed a temporal correlation between the expression of KCC2 and synaptogenesis. Then, when analyzing the subcellular distribution of KCC2 in primary cultures, we found that KCC2 is highly expressed in fi lopodia and mature spines as well as dendritic shaft.
The expression of KCC2 in dendritic spines is intriguing as these are the main postsynaptic targets for glutamatergic synapses. We investigated spine formation in KCC2-/- in vitro as well as in vivo in a compound heterozygous mice for KCC2 null and hypomorphic alleles (KCC2hy/
null) which express 17% of WT KCC2 protein levels. These studies revealed that KCC2 is a key factor in the maturation of dendritic spines. Dendritic protrusions in the KCC2-/- neurons were abnormally long and thin, often reminiscent of fi lopodia. Interestingly, this effect is not due to Cl- transport activity because the spine phenotype could be rescued not only by transfection of KCC2-/- neurons with full-length KCC2 construct (KCC2-FL) but also with an N-terminal deletion construct of KCC2 protein (KCC2-∆NTD) which, was found not to be capable of K-

10
Summary
Cl cotransport. Overexpression of the KCC2 C-terminal domain (KCC2-CTD) in wild type neurons induced a spine phenotype similar to that in KCC2-/- neurons. Moreover, we found that KCC2-CTD binds to the FERM domain of the cytoskeleton associated protein 4.1N. These results indicate a structural role for KCC2 in the development of functional glutamatergic synapses. In general, our data on the role of KCC2 in excitatory synapse formation and its well-known functional role in GABAergic response maturation suggest KCC2 as a synchronizer for the functional development of glutamatergic and GABAergic synapses in neuronal network.
We also examined the regulatory mechanisms of KCC2 expression during development and plasticity. In
primary and organotypic cultures, we demonstrated that synaptic activity of both the glutamatergic and GABAergic system is not required for up-regulation of KCC2 during development. But in acute mature hippocampal slices which undergo continuous synchronous activity induced by the absence of Mg2+ solution, KCC2 mRNA and protein expression were down-regulated in CA1 pyramidal neurons subsequently leading to a reduced capacity for neuronal Cl- extrusion. This effect is mediated by endogenous BDNF-TrkB down-stream cascades involving both Shc/FRS-2 and PLCγ-CREB signaling. BDNF-mediated changes in KCC2 expression indicate that KCC2 is significantly involved in the complex mechanisms of neuronal plasticity during development, and pathophysiological conditions.

11
Review of the literature
1. Neuronal network activity in developing cortex
1.1 Neuronal transmitters in the central nervous system In the mature brain, neuronal network activity is maintained by the coordinated action of excitatory and inhibitory neurotransmission. The most common excitatory neurotransmitter is glutamate which acts through ionotropic AMPA, kainate and NMDA receptors as well as metabotropic receptors (mGluRs). AMPA and kainate receptors conduct Na+ and K+ and mediate faster depolarizing responses, whereas NMDA receptors mediate slow postsynaptic depolarizing responses conducting Ca2+ in addition to Na+ and K+. (Kandel et al., 2000). In recent years, AMPA-receptor signaling has been intensively studied and it is known to have a major infl uence on excitatory synaptic strength. The activation of NMDA receptors is a prerequisite for induction of AMPA-mediated synaptic plasticity (e.g. long-term plasticity, LTP; Bliss and Collingridge, 1993; Kullmann et al., 2000). The metabotropic glutamate receptor subfamily (mGluR) is composed by three types of G-protein coupled receptors which are mainly different in their intracellular signaling mechanisms. The actions of glutamate on ionotropic receptors are always excitatory, but activation of metabotropic glutamate receptors can be both excitatory and inhibitory (Kandel et al., 2000). The major inhibitory neurotransmitters in the brain are GABA and glycine. They exert their physiological actions by binding to the ionotropic GABAA and Glycine receptors (GlyR) as well as the metabotropic GABA receptors GABAB.
As the ionotropic GABAA receptors are permeable for both Cl- and HCO3
-, the direction of the elicited postsynaptic net current is dictated by the electrochemical gradients and concentrations of these two ions (Kaila and Voipio, 1987; Kaila, 1994). The glycine receptors are best known for mediating inhibitory neurotransmission
in the spinal cord and brain stem. Recent evidence suggests it may also have other physiological roles, including excitatory neurotransmission in embryonic cortical neurons (Lynch, 2004).
1.2 Synaptic properties of immature neuronsThe synaptic organization and properties of the immature brain are different from those of adult ones. In this aspect, a prominent feature of an immature neuron is GABAergic and glycinergic signaling which is depolarizing at embryonic stage and early postnatal life (Cherubini et al., 1991; Ben Ari, 2001). Studies from various groups have demonstrated that GABA activates GABAA receptors causing depolarizing instead of hyperpolarizing responses in developing neurons. The depolarization action of GABA is due to an elevated intracellular Cl- concentration resulting from the late developmental expression of KCC2 (Rivera et al., 1999; Payne et al., 2003) and earlier expression of NKCC1 (Marty et al., 2002). Importantly, the depolarizing actions of GABAA during development can reach the threshold of voltage-gated Na+ channels and evoke action potentials (Chen et al., 1996; Dammerman et al., 2000; Owens et al., 1996). The depolarizing and sometimes even excitatory effect have been shown in developing neurons from various brain structures including cortex, hippocampus,
Review of the Literature

12
hypothalamus, spinal cord and olfactory bulb (LoTurco et al., 1995; Obrietan and van den Pol, 1995; Rivera et al., 1999; Rohrbough and Spitzer, 1996; Serafi ni et al., 1995).
Voltage-gated calcium channels (VGCCs) are another type of voltage-gated channels activated by GABA in developing neurons. Synaptic activation of GABA increases the intracellular Ca2+ (Ben Ari, 2002; Garaschuk et al., 1998; Leinekugel et al., 1995; Leinekugel et al., 1997; Owens and Kriegstein, 2002) through VGCC. This may underlie the mechanisms of trophic effects of GABA during development. GABA also acts in synergy with NMDA channels by reducing voltage-dependent Mg2+ block, thus leading to activation of NMDA receptors and a further rise of intracellular Ca2+.
Glutamatergic synaptic transmission also undergoes signifi cant changes at its earliest stage of development. In general, glutamatergic neurotransmission is thought to appear later than GABAergic transmission. By measurement of postsynaptic current, CA1 pyramidal neurons can be divided into silent, GABA-only neurons and GABA-glutamate neurons partly indicating their developmental stages (Ben Ari, 2002). Several studies in hippocampus (Durand et al., 1996; Liao and Malinow, 1996; Wu et al., 1996), cortex (Crair and Malenka, 1995; Isaac et al., 1997) have shown that at birth, glutamatergic transmission is largely NMDA receptor mediated but, over the course of development, the number of AMPA receptors increases and transformation of many of these synapses into the standard AMPA/NMDA synapses occurs. Interestingly, the emergence of functional ionotropic glutamatergic transmission coincides with that of earlier network-driven spontaneous activity in
developing hippocampus (Khazipov et al., 2001; Sipilä and Kaila, 2007).
1.3 Network driven spontaneous activity in developing hippocampusImmature neuronal network events are featured by spontaneous rhythmic activity in the central nervous system. In hippocampus, these earlier network events are termed Giant Depolarizing Potentials (GDPs) and can be observed in rats, mice, rabbits and primates using intracellular recordings (Ben Ari et al., 1989; Aguado et al., 2003; Khazipov et al., 2001; Menendez et al., 1998). These events can be detected by extracellular field-potential recordings, and they recruit both GABAergic interneurons and glutamatergic principal neurons (Ben Ari et al., 2001; Ben Ari et al., 2002; Hollrigel et al., 1998; Khazipov et al., 1997). These events have also been called “population bursts” (Lamsa et al., 2000), ”giant GABAergic potentials” (Strata et al., 1997) and “early network oscillations” (Garaschuk et al., 1998). GDPs are long-lasting (0.5-1 s) and recurrent (0.1-0.3 Hz) activity that occurs during fi rst two weeks of postnatal life during which postsynaptic GABAA receptors mediated action undergo developmental maturation from depolarizing to hyperpolarizing. Studying mechanisms that underlie the generation of GDPs has gained much progress in recent years. In general, the focus has been on the role of depolarizing GABAA receptor-mediated actions in the network and the intrinsic properties of pyramidal neurons during earlier development.
Earlier views on the mechanisms of GDP generation pointed to GABAergic neurons and synapses as the main generators. This was based on the experiments showing that GDPs are inhibited by GABAA antagonists (Ben
Review of the Literature

13
Ari et al., 1989; Garaschuk et al., 1998; Khalilov et al., 1999). However, glutamatergic synapses also contribute to their generation because they are fully blocked by ionotropic glutamate antagonists in many experimental conditions (Ben Ari et al., 1989; Bolea et al., 1999; Hollrigel et al., 1998; Khazipov et al., 2001; Lamsa et al., 2000). Recent data by Sipilä (Sipilä et al. 2005) suggest that the intrinsic bursting activities of CA3 pyramidal neurons in newborn are responsible for the generation of GDPs and recurrent glutamatergic connections between bursting CA3 pyramidal neurons are required for synchronization of field GDPs. The depolarizing actions of GABAergic transmission is to exert a temporally non-patterned, facilitator action via both synaptic and tonic (most likely extrasynaptic) GABAA-mediated input on the immature CA3 pyramidal neurons (Sipilä et al. 2005). Later, they found that the intrinsic burst activity of individual pyramidal neurons is driven by persistent Na+ current and terminated by a slow afterhyperpolarization mediated by a slow Ca2+-activated K+ current (Sipilä et al. 2006a). In addition, an inhibitory effect of GABAB receptors on presynaptic release of glutamate and GABA contributes to GDP termination (McLean et al., 1996). In conclusion these results indicate that glutamatergic transmission has an “instructive role”, whereas, GABA transmission has a facilitating and “permissive action” (Sipilä and Kaila, 2007).
1.4 Developmental roles of the spontaneous activitySpontaneous activity characterized by tight correlation in space and in time is thought to be critical for guiding neuronal circuit development. Periodic bursts of action
potentials lead to substantial increase in the intracellular calcium concentration in the participating neurons (Feller, 1999). There is growing evidence showing that synchronized changes in intracellular calcium can infl uence profoundly a variety of intracellular processes, including gene expression and the establishment of cell phenotype (Berridge, 1998). The roles of earlier spontaneous neuronal activity in neuronal development have been studied in various structures. In the spinal cord of Xenopus laevis embryos, patterns of spontaneous Ca2+ spikes generated by different classes of embryonic spinal neurons specify their expression of neurotransmitters (Borodinsky et al., 2004; Spitzer et al., 2004). In the visual system, activity-dependant neuronal wiring has been intensively studied (Penn and Shatz, 1999). Neuronal activities are required for establishment of the connectivity of eye-specific projection. After eye opening, axons from lateral genicuclate nucleus terminate in alternating ocular dominance columns in eye specific manner. The specifi city of these events is dependent on patterned and synchronized activity of the corresponding eye. The organization of retinal inputs to lateral geniculate nucleus in the thalamus is already formed before eye opening. The formation of retino-thalamic connectivity has been suggested to be determined by embryonic spontaneous, synchronized neural activity (Penn et al., 1998; Katz and Crowley, 2002). The retinal ganglion cells spontaneously generate these highly correlated bursts of action potentials before the retina can respond to light. Blockade of this endogenous activity, or biasing the competition in favor of one eye after birth, results in a severe disruption of the pattern of retinogeniculate connections (Penn and Shatz, 1999; Katz and Crowley, 2002).
Review of the Literature

14
A study on hippocampus developing circuit indicates that GDPs, paired with mossy fi ber stimulation, induce signifi cant increase in synaptic effi cacy (Kasyanov et al., 2004). But whether this may contribute to sculpting neuronal circuits, as demonstrated in the developing visual system and spinal cord, remains to be determined.
2. GABAergic and glycinergic neurotransmission
2.1 GABAA receptorsGABAA receptors are widely distributed in cerebral cortex, hippocampus, thalamus, brain stem and spinal cord. Modulation of their expression, cellular distribution and function has profound consequences for neuronal excitability. GABAA receptors function as pentameric anion channels. There are eight molecularly distinct subunit families for GABAA receptors α (1-6), β (1-3), γ (1-3), δ, π, ε, θ and ρ (1-3) (Steiger and Russek, 2004; Mehta and Ticku, 1999; Barnard et al., 1998; Frischy and Mohler, 1995; Sieghart, 1995; Wisden and Seeburg, 1992). Heteromeric combinations of these subunits give rise to receptors classes with different functional properties. Recent data has shown that the pentameric complexes of GABAA receptors usually contain 2α/2β/1γ-subunit variants (Klausberger et al., 2001; Baumann et al., 2001; Knight et al., 2000; Farrar et al., 1999). It has been concluded that the majority of GABAA receptors contain a single type of α- and β-subunit variants with the α1β2γ2 combination representing the largest population of GABAA receptors followed by α2β3γ2 and α3β3γ2 (Fritschy and Brunig, 2003). Pharmacological analyses of recombinant GABAA receptors suggest that structural heterogeneity of receptor subclasses determines the
functional properties of receptors. The α- and β- subunits participate in high affi nity GABA binding and are responsible for receptor assembly and membrane targeting (Connolly et al., 1996; Bureau and Olsen, 1990). Benzodiazepines which are among the most used drugs for anxiety treatment, bind at the interface of α- and γ- subunits and modulate their channel properties (Kaufmann et al., 2003; Sieghart, 1995; Steiger and Russek, 2004). Moreover, neurosteroids, which are positive allosteric modulators of recombinant and native GABAA receptors, are most potent on receptors containing the δ subunits (Fritschy and Brunig, 2003). The ρ-subunits are expressed primarily in the retina and formed homooligomeric receptors. In some literature, ρ-subunits were classified as GABAC receptors because of their differences in kinetics, sensitivity to GABA and pharmacological properties as compared with other subunits of GABAA receptors (Bormann, 2000; Feigenspan and Bormann, 1998; Johnston, 1996; Polenzani et al., 1991). Accumulated studies on subunit-specifi c properties of GABAA receptors in both recombinant and native receptors support the concept that GABAA receptor heterogeneity is a major factor determining the functional properties of GABAergic inhibitory synapses (Mohler et al., 1998; Sieghart, 2000).
Although GABAA receptors are targeted to inhibitory synapses on dendrites and cell bodies of neurons, they also occur in other places such as on synapses formed on axons and extrasynaptic membranes. GABAA receptors which are located at axon-axon synapses mediate presynaptic inhibition. Presynaptic inhibition operated by GABAA receptors are found not only in spinal cord but also in the brain, including hippocampal mossy fibers, brain stem,
Review of the Literature

15
cerebellum, and hypothalamus (Kullmann et al., 2005). Studies on cerebellar granule cells have shown that extrasynaptic receptors on dendrites and somas provide a persistent tonic current (Wall and Usowicz, 1997; Brickley et al., 1996; Kaneda et al., 1995). Tonic GABAA receptor mediated current has been also described in hippocampal dentate gyrus granule cells (Stell and Moday, 2002; Nusser and Mody, 2002) and inhibitory interneurons in the CA1 region (Semyanov et al., 2004; Semyanov et al., 2003). Interestingly, significant tonic GABAA receptor current is absent in adult hippocampal pyramidal neurons but can be detected only in early development (Demarque et al., 2002; Semyanov et al., 2004; Sipilä and Kaila, 2007) indicating a role of tonic GABAA response in development. Subunit requirements for the formation of extrasynaptic receptors are different in diverse neuronal populations. δ-subunit-containing receptors are probably the major contributors to non-desensitizing high affinity receptors mediating tonic current in cerebellar and dentate granule cells (Semyanov et al., 2004).
2.2 Glycine receptorsAlong with GABAA receptors, glycine receptors (GlyR) belong to the pentameric ligand-gated ion channel superfamily. So far, four different GlyR α subunits have been identified in vertebrates and only one GlyR β subunit has been identifi ed in mammals (Betz and Laube, 2006). The expression of the GlyR α subunits is developmentally and regionally regulated. In adult rodents, GlyR α1 mRNA and protein has been shown to be prominently expressed in spinal cord, brainstem and colliculi (Betz and Laube, 2006), whereas α2 transcripts were found only at low levels in adult hippocampus, cerebral
cortex and thalamus (Sato et al., 1992; Malosio et al., 1991). α3 transcripts were detected in spinal cord, cerebellum and olfactory bulb (Malosio et al., 1991). α4 mRNA is scarcely localized in mammalian CNS (Harvey et al., 2000). However, α2 and α4 transcripts are abundant at embryonic stages and decrease gradually after birth. Expression of α1 and α3 mRNA increases postnatally. GlyR β subunit is widely expressed throughout the embryonic and postnatal CNS (Betz and Laube, 2006). Subunits composition of functional GlyR undergo developmental shift from predominantly α2-homomers neonatal form to α1β-heteromers adult form (Becker et al., 1988). The α2-homomers GlyRs that are expressed at embryonic stages and in the neonate are thought to be extrasynaptic receptors because postsynaptic GlyR assembly requires the interaction of β subunit and tubulin-binding protein gephyrin (Meyer et al., 1995). In developing neocortex, GlyRs were found to be activated by nonsynaptic release of taurine stored in developing neocortex (Flint et al., 1998). The extrasynaptic GlyR mediated signaling could be important for neuronal differentiation and synaptogenesis (Lynch, 2004).
2.3 Developmental roles of GABA and glycine neurotransmissionGABAA and glycine receptors are generally known as inhibitory receptors in mature neurons because the Cl- equilibrium potential is usually close to the cell resting potential. However, in immature neurons, the intracellular Cl- concentration is high with the effect that GABAA and GlyR activation cause a strong depolarization. Depolarization that leads to activation of voltage-gated Ca2+ channels thus increasing the concentration of intracellular Ca2+
Review of the Literature

16
is a fundamental mechanism for trophic effects of GABA during development. The GABAA mediated depolarization can enhance both the expression and release of brain-derived neurotrophic factor (BDNF) (Marty et al., 1996). Despite its functionally excitatory role (see Sipilä and Kaila, 2007), the depolarization action of GABA and glycine in immature neurons can produce suppression of glutamatergic EPSCs because of the effective shunting of postsynaptic membrane. As demonstrated in the neonate hippocampus (Lamsa et al., 2000; Palva et al., 2000; Sipila et al., 2006a), activation of GABAA receptors by muscimol inhibits the spontaneous bursting of CA3 pyramidal neurons indicating that GABAergic currents can exert a shunting-type inhibitory action on pyramidal neurons already in an early developmental stage.
3. Cellular chloride homeostasis The chloride concentration within the cell is critical for multiple functions, for example: cellular excitability, intracellular pH (Kaila and Ransom, 1998), cell volume regulation and cell cycle control (Russell, 2000). Changing the intracellular Cl concentration of a neuron affects the polarity of GABAA and glycine receptor-mediated responses (Zhang and Jackson, 1995; Sutor and Luhmann, 1995; Thompson and Gahwiler, 1989; Misgeld et al., 1986; Kaila, 1994). The maintenance of chloride homeostasis is largely carried out by cotransporters and exchangers who selectively allow chloride and other coupled ions to pass through in and out of the cell (Kaila, 1994; Payne et al, 2003). Chloride cotransporters are secondary active membrane transporters which carry chloride crossing the membrane together with coupled cations or anions. Members of the cation-chloride cotransporter
(CCCs) family have been identified as important regulators of neuronal Cl- concentration (DeFazio et al., 2000; Kaila, 1994; Kakazu et al., 1999; Mercado et al., 2004; Payne, 1997; Payne et al., 2003; Sung et al., 2000) and play a key role in various aspects of neuronal development, plasticity and trauma (Payne et al., 2003).
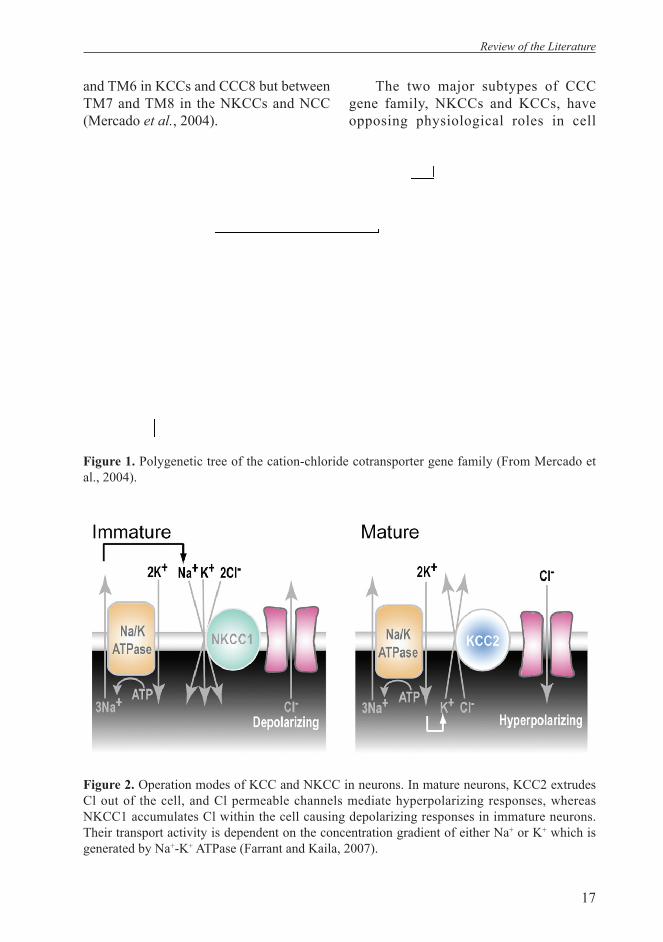
The CCC gene family consists of a series of membrane proteins that transport chloride ions together with Na+ and/or K+. They function as secondary active transporters using the transmembrane K+ and Na+ gradients generated by Na+/K+ ATPase to transport Cl in or out of cells, respectively (Kaila, 1994; Payne et al, 2003). Other secondary active transport proteins also participate in neuronal Cl- homeostasis, including Na+-dependent and Na+-independent anion exchangers (NDAE and AE) which exchange Cl- for HCO3
- (Grichtchenko et al., 2001; Romero et al., 1997). Cation-chloride cotransporter function is electroneutral and transport activity results in no change of the transmembrane potential (Mercado et al., 2004). To date, nine genes of the CCCs have been identifi ed: one Na+-Cl- co-transporter (NCC), two Na+-K+-2Cl- co-transporters (NKCCs) and four K+-Cl- co-transporters (KCCs). In addition, two structurally related proteins with unknown function: CCC8 (also known as cation-chloride cotransporter interaction protein, CIP, (Caron et al., 2000) and CCC9 (Mercado et al., 2004) (Figure 1). The hydropathy profi le of the predicted proteins reveals a similar basic membrane topology for the gene family. The proteins have a central hydrophobic core containing 12 α-helical transmembrane domains (TM), fl anked by a hydrophilic amino-and a larger carboxy-terminal domains (Delpire and Mount, 2002). A large N-linked glycosylated extracellular loop locates between TM5
Review of the Literature
17
and TM6 in KCCs and CCC8 but between TM7 and TM8 in the NKCCs and NCC (Mercado et al., 2004).
Figure 1. Polygenetic tree of the cation-chloride cotransporter gene family (From Mercado et al., 2004).
The two major subtypes of CCC gene family, NKCCs and KCCs, have opposing physiological roles in cell
Figure 2. Operation modes of KCC and NKCC in neurons. In mature neurons, KCC2 extrudes Cl out of the cell, and Cl permeable channels mediate hyperpolarizing responses, whereas NKCC1 accumulates Cl within the cell causing depolarizing responses in immature neurons. Their transport activity is dependent on the concentration gradient of either Na+ or K+ which is generated by Na+-K+ ATPase (Farrant and Kaila, 2007).
Review of the Literature

18
volume regulation and intracellular neuronal Cl homeostasis. NKCCs normally accumulate Cl within the cell whereas KCCs lower the intracellular Cl concentration (Payne et al., 2003; Russell, 2000) (Figure 2). NKCC1 is ubiquitously expressed, present in nonepithelial cells and basolateral membrane of epithelial cells (Xu et al., 1994). NKCC2, the apical isoform, is expressed exclusively in the thick ascending limb of Henle’s loop in the kidney (Gamba et al., 1994).
K+ coupled Cl- co-transporters (KCC) are encoded by four separate genes (KCC1 to KCC4). KCC1 is widely expressed and plays a role in cellular volume regulation. KCC2 is unique in the protein family being exclusively expressed in central neurons (Payne et al., 1996; Rivera et al., 1999). KCC3 is widely expressed in the CNS as well as other tissues (Boettger et al., 2002; Mercado et al., 2004; Mount et al., 1999; Race et al., 1999). KCC4 is expressed in kidney, heart and brain (Mercado et al., 2004; Mount et al., 1999; Le Rouzic et al., 2006). Except KCC2, the other KCCs are all swelling-activated, suggesting an important role in regulatory volume decrease (RVD) as shown in red blood cells where KCC1, KCC3 and KCC4 are co-expressed (Adragna et al., 2004). Particularly, NKCCs and KCCs display reciprocal regulatory mechanism. NKCC1 is activated by cell shrinkage and by a decrease in [Cl-]i (Lytle and McManus, 2002; Schomberg et al, 2003), and inhibited by the serine-threonine phosphatase-1 (PP1) (Darman et al., 2001; Matskevich et al., 2005), the kinase inhibitor staurosporine (Lytle, 1998; Matskevich et al., 2005), and the thiol-alkylating agent N-ehyl-maleimide (NEM) (Russell, 2000). In contrast, KCCs (but not KCC2) are activated by cell swelling and by increases in [Cl-]i (Lauf and Adragna, 2000; Lytle and McManus, 2002). KCCs
are activated by PP1, staurosporine, and NEM (Lauf and Adragna, 2000). More recently, WNK3, WNK4 and SPAK of the serine-threonine kinase protein family were found to participate in the reciprocal regulation of NKCC and KCC transport activity (Gagnon et al., 2006; Kahle et al., 2005; Piechotta et al., 2002).
Alternative splicing has also been found in this gene family. There are six alternative splicing forms of NKCC2 which differ in functional characteristics (Gimenez et al., 2002; Mount et al., 1999; Plata et al., 2001; Plata et al., 2002). Two C-terminal domain spliced isoforms in NKCC1 (Mercado et al., 2004) and fi ve N-terminal domain variants were found in KCC3 (Mercado et al., 2005; Pearson et al., 2001) for which the KCC3a is abundant in the brain whereas KCC3b is mainly present in kidney.
3.1 KCC1KCC1 was the fi rst K+-Cl- co-transporter to be cloned. The expression of KCC1 is ubiquitous. The physiology of KCC1 is best studied in red blood cells from various vertebrate species (Lauf and Adragna, 2000). KCC1 is considered to have a house-keeping role in cell volume maintenance and regulation. In adult central nervous system, KCC1 mRNA is expressed in both neurons and non-neuronal cells at relatively low levels, but is abundant in choroid plexus epithelial cells (Kanaka et al., 2001). During development, KCC1 is detected in choroid plexus as earlier as E14.5 with scarce distribution in neuronal structures (I). These results indicate that KCC1 may participate in the regulation of ionic composition of the cerebro-spinal fl uid.
3.2 KCC2KCC2 was cloned by Payne et al in 1996 and turned out to be the only neuronal
Review of the Literature

19
specific isoform in this protein family. Expression studies for KCC2 indicate that there is no or very low transcript levels detected in tissues outside CNS. Within the brain, no signal for KCC2 mRNA and protein could be found in white matter tracts or in other non-neuronal structures (Payne et al., 1996; Williams et al., 1999). Later studies have confi rmed high levels of expression of KCC2 in interneurons and pyramidal neurons of the cortex, hippocampus, cerebellum, retina, spinal cord and many nuclei in brain stem (Gulyas et al., 2001; Lu et al., 1999; Mercado et al., 2004; Mikawa et al., 2002; Rivera et al., 1999).
3.2.1 KCC2 and neuronal excitabilityThe neuronal-specific expression of KCC2 is signifi cant for maintaining the low intracellular chloride concentration required for hyperpolarization action of GABAA and glycine receptor (Hubner et al., 2001b; Payne et al., 2003; Rivera et al., 1999). However, in early developing neuron, GABA and glycine-mediated responses are depolarizing. KCC2 is developmentally up-regulated and causes the developmental shift in GABAA mediated responses from depolarizing to hyperpolarizing (Hubner et al., 2001b; Rivera et al., 1999). Overexpression
of KCC2 in immature neurons reduces
[Cl-]i (Lee et al., 2005). Reduction of KCC2 expression in pyramidal cells from semi-acute rat hippocampus slices using antisense oligonucleotides resulted in a marked positive shift in the reversal potential of GABA response (Rivera et al., 1999). KCC2 knock out mice die immediately after birth due to severe motor defi cits that abolished respiration. In spinal cord motoneurons of KCC2 knock out mice, unlike in wild type, GABA and glycine are excitatory at later embryonic stages (Hubner et al., 2001b). A KCC2
transgenic mouse that has been generated with only 5-10% expression of KCC2 exhibits frequent generalized seizures, and such mice die shortly after birth (Woo et al., 2002). The brains of these mice show a signifi cant loss of parvalbumin-positive interneurons, indicating brain injury. The regions most affected are the hippocampus and temporal and entorhinal cortices. Extracellular field potential measurements in the CA1 hippocampus revealed a state of hyperexcitability and an increased sensitivity to the application of picrotoxin, a blocker of the GABAA receptor, in homozygous hippocampal sections. The authors also demonstrated that adult heterozygote animals show increased susceptibility to epileptic seizures and increased resistance to the anticonvulsant effect of propofol (Woo et al., 2002). Hypomorphic KCC2 defi cient mice that retain 15-20% protein expression levels in the brain were viable and fertile but weighed 15-20% less than wild-type littermates at 2 weeks old and thereafter (Tornberg et al., 2005). The mice displayed increased anxiety-like behavior in several tests including elevated plus-maze and were more susceptible to pentylenetetrazole-induced seizures. Taken together, these results indicate that KCC2 plays an important role in controlling CNS excitability during both postnatal development and adult life.
Functional characterization of rat and human KCC2 shows that KCC2 has a higher K+ and Cl- affi nity than the other KCCs with extracellular Kms of ~5.2 and 9.2mM respectively (Mercado et al., 2004; Payne, 1997; Song et al., 2002). Extracellular K+ increases during pathophysiological neuronal activity, and under these conditions the activity of KCC2 may reverse as high K+ concentration changes the transport activity from effl ux
Review of the Literature

20
to influx (Payne, 1997; Payne et al., 2003).
Interestingly, investigation on mechanism of neuropathic pain revealed involvement of KCC2 in regulating pain sensitivity. Recent results show that neuropathic pain that follows peripheral nerve injury induced down-regulated expression of KCC2. The down-regulated KCC2 expression resulted in a shift in the Cl gradient (mediated by GABAA receptor and/or glycine receptor) in spinal superfi cial dorsal horn lamina I neurons which is responsible for nociceptive pathway transduction to brainstem (Coull et al., 2003). The shift in anion gradient was reported to happen trans-synaptically with respect to the injury site. The authors suggested that the change in the polarity of the GABAA receptor and glycine receptor mediated postsynaptic action provides a potential mechanistic basis for many of the reported consequences of central nociceptive sensitization (Coull et al., 2003). The down-regulation of KCC2 resulting in long-term increase in the excitability would remain in lamina I neurons and would be a cause of chronic pain (Mantyh and Hung, 2004).
Recently, an intriguing question arose from an ultrastructural study on KCC2 expression in rat hippocampus by Gulyas et al. (Gulyas et al., 2001). They found that KCC2 is highly expressed in the vicinity of synapses which are responsible of excitatory transmission. Their results were supported by electron-microscopy (EM) studies which showed that immunogold particles were preferentially found close to asymmetrical but not symmetrical synapses. KCC2 immunoreactivity was localized in dendritic spines of all principal cells and in the thorny excrescences of CA3 pyramidal cells. Dendritic spines receive only excitatory synapses. In contrast, the dendritic shaft, which receives
only symmetrical synaptic input, exhibited low levels of KCC2 (Gulyas et al., 2001). Moreover, parvalbumin (PV) -containing GABAergic interneurons which receive several times more excitatory inputs than any other interneuron and principal neuron, was covered by very high levels of KCC2 protein in their somata and dendrites. These results may implicate possible novel roles for KCC2. The authors speculated that KCC2 participates in Cl-driven volume regulation (see Payne et al, 2003) after excessive excitation-induced cell swelling in the vicinity of extrasynaptic GABAA receptors; the relationship of KCC2 with excitatory synapses needs further investigation. This is one of the aims of the present thesis.
3.2.2 Regulation of KCC2 expression and activationThe regulatory mechanisms of KCC2 expression and activation are also a major target of interest. The neuron-restricted expression pattern of KCC2 was suggested to be caused by the neuronal-restrictive silencing element (NRSE) in the mouse KCC2 gene (Karadsheh and Delpire, 2001). This element binds a transcription factor called neuron-restricted silencing factor (NRSF) that silences transcription of the gene in non-neuronal cells. However, Uvarov et al. (Uvarov et al., 2005) recently reported that KCC2 transgenic mice lacking NRSE showed CNS neuron-specific expression and developmental up-regulation of endogenous KCC2 gene that was similar to the wild type. Although exogenous expression of NRSF could down-regulate KCC2 expression in vitro, the in vivo results suggested that NRSE in KCC2 is not critical for KCC2 neuron-specifi c expression, and silencing elements other than NRSE are required to prevent the expression of KCC2 in most non-neuronal tissues (Uvarov et al., 2005).
Review of the Literature

21
BDNF is known to play a central role in the genesis and establishment of epileptic activity (Binder et al., 2001). BDNF can regulate KCC2 expression in two directions depending on different developmental stage. During early development, BDNF increases the expression of KCC2 as demonstrated by transgenic mice embryos with overexpression of BDNF (Aguado et al., 2003). Recently studies in acute hippocampus slices (Rivera et al., 2002) have shown that BDNF and NT-4 dramatically decrease KCC2 expression mediated by TrkB receptor activating, consequently impairing neuronal chloride extrusion capacity. After kindling-induced seizures in vivo, the expression of KCC2 is down-regulated with a spatial-temporal profi le similar to the up-regulation of TrkB and BDNF (Rivera et al., 2002). The data revealed a novel mechanism whereby BDNF/TrkB signaling suppresses chloride-dependent fast GABAergic inhibition which contributes to the role of BDNF/TrkB signal cascades in the induction and establishment of epileptic activity.
A recent study reported that de pola-rizing GABAergic activity upregulates the mRNA levels of KCC2 during development, thus promoting itself the switch from depolarization to hyperpolarization (Ganguly et al., 2001). However, these results were not supported by subsequent studies conducted by Titz et al., (2003) as well as by us (II, see results and discussion).
Recently, new data have been added to the functional regulation of KCC2 and its intracellular signal partners. KCC2 has a unique protein tyrosine kinase (PTK) consensus site (Payne et al., 1996), Kelsch and co-authors (Kelsch et al., 2001) reported that the activity of KCC2 during maturation of cultured hippocampus neurons is activated by co-application of IGF-1 and cytosolic tyrosine kinase, c-
src, and can be deactivated by membrane-permeable protein tyrosine kinase inhibitors, genistein and lavendustin A. Their results indicate that the development of neuronal K+/Cl- cotransport requires cooperation of growth factors with PTK-dependent phosphorylation.
Among KCCs, KCC2 is unique in mediating constitutive K-Cl transport activity under isotonic condition when heterologously expressed in Xenopus
oocytes (Song et al., 2002; Strange et al., 2000). In a recent publication the cytoplasmic domain (1021-1035aa) within KCC2 C-terminal was identified as the crucial domain for its constitutive activity (Mercado et al., 2006). Furthermore, swelling-activated K+-Cl- cotransport is suppressed by calyculin A, whereas isotonic transport mediated by KCC2
is completely resistant to this serine-threonine phosphatase inhibitor (Mercado et al., 2006). To date, little is know about the intracellular associating proteins of KCC2. Recently, by using yeast two-hybrid system, Inoue et al (Inoue et al., 2004) found that KCC2 interacts with a brain-type creatine kinase (CK). A functional analysis of this interaction in HEK293 cells showed that CK was able to activate KCC2 transport activity. Taken together, KCC2 is regulated by intracellular phophorylation cascades that need further investigations.
3.3 KCC3KCC3 was cloned independently by three groups in the same year (Hiki et al., 1999; Mount et al., 1999; Race et al., 1999). In initial studies, two KCC3 isoforms were found, generated by two alternative fi rst exons, KCC3a and KCC3b. KCC3a is widely expressed with abundant expression in brain, kidney, muscle, lung, and heart, whereas KCC3b is most abundant in kidney. No evidence about the functional
Review of the Literature

22
difference between these two isoforms is available. Later, a total of fi ve different N-terminal isoforms of the human KCC3 proteins have been described (Mercado et al., 2005) with no signifi cant functional difference. All fi ve isoforms of KCC3 are activated by cell swelling under hypotonic conditions, with no activity under isotonic conditions (Mercado et al., 2005). In the central nervous system, KCC3 protein was detectable in cerebral cortex, hippocampus, white matter, hypothalamus, brain stem, choroids plexus and spinal cord (Becker et al., 1988; Mercado et al., 2004; Boettger et al., 2003). The ontogeny of KCC3 correlates with the development of myelin (Pearson et al., 2001) and, notably, this isoform is also expressed in hippocampal and cerebellar neurons (Boettger et al., 2003).
Recently, KCC3 knock out mouse models were generated by two different groups. One group reported that the mice with targeted deletion of KCC3 have a locomotor defi cit, peripheral neuropathy and a sensorimotor gating defi cit, similar to a human disease, peripheral neuropathy associated with agenesis of the corpus callosum (ACCPN) (Howard et al., 2002). ACCPN is a severe sensorimotor neuropathy disease associated with mental retardation, dysmorphic features and complete or partial agenesis of the corpus callosum. The other group with KCC3 null mice supports the data on severe motor abnormalities that is correlated with a progressive neurodegeneration in the periphery and CNS which may be related with human ACCPN and Anderman syndrome, but also reported that the mice have reduced seizure threshold, EEG abnormalities, hippocampal and cerebellar neuronal degeneration, arterial hypertension and a slowly progressive deafness (Boettger et al., 2003).
3.4 KCC4KCC4 was identified by Mount et al. (Mount et al., 1999), at the same time as KCC3. The tissue distribution analysis shows that KCC4 is mainly expressed in the heart, kidney, scarcely in the brain. Later western blot analysis by KCC4 antibody (Karadsheh et al., 2004) revealed high expression in peripheral nerves and spinal cord, with low levels in whole brain. Within the brain, the cerebral cortex, hippocampus, and cerebellum revealed minimal KCC4 expression, whereas midbrain and brainstem demonstrated higher levels. From adult mouse brain immunostaining, KCC4 was observed in the apical membrane of choroid plexus epithelial cells as well as in cranial nerves. All other brain structures, e.g. cortex, hippocampus, cerebellum showed no KCC4 immunoreactivity, suggesting very low or absent expression of the cotransporter in these regions. KCC4 expression on the apical membrane of the choroid plexus, suggest that it may participate in K+ reabsorption (Karadsheh et al., 2004). The functional implication of KCC4 expression in peripheral neurons still remains to be determined.
Targeted disruption of KCC4 mice has been generated by Boettger et al. in 2002 (Boettger et al., 2002). The null mice developed distal tubular metabolic acidosis, indicating the prominent role of KCC4 in acid-base balance. KCC4-null mice exhibited no apparent alteration of vestibular and motor function but the mice are completely deaf because their outer hair cells, the cochlea and the organ of Corti are degenerated. The authors suggested an important role of KCC4 in maintaining extracellular homeostasis by salt uptake in Deiter’s cells (Boettger et al., 2002).
Review of the Literature

23
3.5 NKCC1NKCC1 is a membrane transporter that generally promotes intracellular accumulation of Cl- driven by the Na+ gradient generated by the Na+-K+-ATPase. NKCC1 is expressed in the basolateral membrane of epithelial cells and in several nonepithelial cells including neurons and glia (Shillingford et al., 2002; MacVicar
Expression Function Phenotype ofKnock out mice
KCC1 Ubiquitously expressed Epithelium cells Non-epithelium cells Choroid plexus. Expressed in both neu-rons and glia (Wang et al., 2002; Lee et al., 2003).
1. Active potassium absorption (Sangan et al., 2000) 2. volume regulation, RVD, activate under hypotonic conditions.
No KO avaiable
KCC2 Exclusively expressed in neurons; No mRNA and protein were detected in white matter or other non-neuronal structures.
1. Extrude K and Cl out of cell 2. Activate under isotonic condition 3. Maintaining the low intracellular chloride concentration required for hyperpolarization of GABAA and glycine receptors.
Die immediately after birth due to respiratory failure. In spinal cord motor neuron, GABA and glycine are excitatory at E18 as compared to wild type (Hubner et al., 2001).
KCC3 KCC3a is abundantly expressed in brain, kidney, muscle, lung and heart; KCC3b is mainly expressed in kidney.
Activated by cell swelling under hypotonic condition, no activity under isotonic condition.
Locomotor defi cit, peripheral neuropathy and a sensorimotor gating defi cit, similar to a human disease (ACCPN) (Howard et al., 2002).
KCC4 Expressed in heart, kidney, scarcely in the adult brain; Higly expressed in ventricular zones of embryonic brain. Highly expressed in peripheral nerves, spinal cord, and apical membrane of choroid plexus.
Participates in potassium reabsorption. Activated by cell swelling under hypotonic condition, no activity under isotonic condition.
Distal tubular metabolic acidosis. Alteration of vestibular and motor functions. The mice are completely deaf (Bottger et al., 2002)
NKCC1 Ubiquitously expressed. Expressed in apical mem-brane of choroid plexus epithelial cells, young neu-rons, ventricular zones of LGE, MGE in embryonic brain. DRG neurons and oligodendrocytes.
Accumulates Cl into the cell. Participates in regulatory volume increase and epithelial chloride secretion.
shaker/waltzer phe-notype, complete deaf, reduced salivary secretion, increased intrarenal rennin level and alteration of blood pressure, male sterility (Flagella et al., 1999; Mercado et al., 2004)
et al., 2002; Hubner et al., 2001a; Marty et al., 2002). The expression of NKCC1 has been reported to be present in central neurons, oligodendrocytes, apical membrane of the choroids plexus and in dorsal-root-ganglion (DRG) neurons. In adult DRG neurons, GABA is depolarizing. Expression of NKCC1 may be critical for the appropriate response of GABA in
Table 1. CCC protein family: expression, function and phenotype differences of knock out mice
Review of the Literature

24
these cells that is necessary for normal sensory perception (Sung et al., 2000). Extensive studies on NKCC1 knock out mice revealed many physiological roles of this protein. The null mice develop an inner-ear dysfunction characterized by deafness and the shakerlwaltzer phenotype that indicates impaired epithelial secretion (Delpire et al., 1999; Flagella et al., 1999). Basolateral NKCC1 cotransport participates in the production and regulation of fl uid secretion by salivary glands demonstrated by a significant reduction in the total volume of saliva produced in the defi cient mice (Delpire and Mount, 2002). Homozygote NKCC1 males were completely infertile due to a defi cit in spermatocyte production (Delpire et al., 1999; Pace et al., 2000). The sensory perception abnormalities are probably secondary to the lack of NKCC1 function that disturbs normal GABA responses in sensory neurons. NKCC1 alone can account for accumulation of intracellular Cl- above electrochemical potential equilibrium in mouse DRG neurons that promotes GABA depolarization (Payne et al., 2003).
Recently, several studies have dem-onstrated that the depolarizing GABAA response is mediated by NKCC1 (Dzhala et al., 2005; Okabe et al., 2003; Yamada et al., 2001; Sipila et al. 2006b). As men-tioned above, in mature neurons, KCC2 produces efflux of Cl- and lower intra-cellular Cl concentration, GABA medi-ates hyperpolaring inhibitory responses, whereas in the immature brain, GABA is excitatory. This developmental switch of GABA response from depolarizing to hyperpolarizing can be explained by the different intracellular Cl concentration in immature neurons versus mature neurons. This difference in chloride homeostasis is considered to be produced by the up-regu-lation of KCC2 together with down-regu-
lation of NKCC1 (I,; Rivera et al., 1999; Payne et al., 2003). Thus, the cation-chlo-ride cotransporter family has signifi cant roles in brain development. The regulation of developmental expression of this pro-tein family still requires further studies.
4. Glutamatergic neurotransmissionAMPA receptors mediate the majority of the fast excitatory synaptic transmission (Hollmann and Heinemann, 1994). NMDA receptors are crucial for the induction of various forms of synaptic plasticity (Dingledine et al., 1999, Hollmann and Heinemann, 1994; Nicoll and Malenka, 1999; Rao and Finkbeiner, 2007). Although both are activated by glutamate, they differ in molecular composition and electrophysiological properties. AMPA receptors are tetrameric ion channels permeable to Na+ and K+ ions, consisting of four subunits, GluR1-4. Their subunit composition varies in different regions or under different conditions in the central nervous system. The NMDA receptor is permeable to Ca2+ ion in addition to Na+ and K+ ions and exhibits much slower voltage-dependent kinetics. Studies demonstrated that LTP and LTD in many brain regions depend on NMDA receptor activation (Nicoll and Malenka, 1999).
4.1 Development of glutamatergic neurotransmissionEarly glutamatergic synapses which are only composed of NMDA but not AMPA are termed as silent synapses (Wu et al, 1996). Silent synapses are only transiently found during maturation of glutamatergic neurotransmission. Some reports suggest that initially NMDA receptors present in neuron can induce subsequently AMPA receptor expression. In immature neuron, depolarization of GABAA receptors removes Mg2+ blockade
Review of the Literature

25
of NMDA receptors, leading to an increase in intracellular calcium which may play a role in AMPA receptor expression or traffi cking and activation (Shi et al., 1999; Constantine-Paton and Cline, 1998). There is evidence showing that with development, synapses that contained AMPA receptors gradually acquired more GluR1 subunits, whereas the number of NMDA subunit NR1 was relatively high and remained constant (Petralia et al., 1999). Smaller AMPA receptor-mediated responses observed earlier in development are due to a small fraction of synapses containing functional AMPA receptors. It still not clear whether AMPA receptors are already present early on at synapses but are electrically silent and subsequently activated or whether AMPA receptors are physically inserted into the postsynaptic membrane from intracellular pools (Rubio and Wenthold, 1999; Liao et al., 1999). The phenomenon of silent synapses and subsequently insertion of AMPA receptors not only exits during development of glutamatergic neurotransmission, but also account for a mechanism for adult form LTP. Accumulated data have shown that AMPA receptor traffi cking and synapse insertion play a significant role in LTP induction which underlies the mechanism of learning and memory (Malinow and Malenka, 2002).
4.2 AMPA receptor interacting proteinsA remarkable achievement in studying AMPA receptors in recent years is the discovery of large amounts of AMPA interacting proteins. Those proteins are responsible for AMPA receptors delivery, assembling, anchoring and clustering onto postsynaptic sites and removal in and out of synapses in response to neuronal activity. This dynamic behavior is essential for maintaining normal AMPA receptor mediated neuronal fi ring and an important
mechanism to modify synaptic strength during brain development and experience-dependent plasticity (Esteban, 2003).
Intracellular proteins which interact with AMPA receptor are classifi ed into two groups: those containing PDZ domains and those without PDZ domain.
4.2.1 Proteins containing PDZ domainsProteins containing PDZ domains play a general role in clustering and anchoring membrane proteins. They are mainly scaffolding proteins located at the postsynaptic density (PSD) which is found in excitatory postsynaptic sites. These proteins usually bind to the C-terminal of the interacting protein and they often associate with one another to form large protein complexes (Sheng and Sala, 2001; Craven and Bredt, 1998; Kornau and Seeburg, 1997). It has been proposed that PDZ containing proteins provide a mechanism for clustering ion channels and receptors in the plasma membrane (O’Brien et al., 1998). Glutamate receptor interacting protein (GRIP) was the fi rst protein shown to interact with AMPA receptors by yeast two-hybrid system. GRIP contains seven PDZ domains (Dong et al., 1997). The subcellular distribution of GRIP was originally described to be predominately in specific synapses and its immunoreactivity in primary cultures of rat hippocampal neurons is seen throughout the dendrites. GRIP protein is widely expressed in the brain throughout development and can be detected early in embryonic development (Dong et al., 1999). Biochemical studies show that PDZ domains bind to the C-terminal of GluR2 and GluR3, suggesting that GRIP could form supramolecular complexes of AMPA receptors.
Synapse-associated protein 97 (SAP97) is a member of the MAGUK family that contain many protein interac-
Review of the Literature

26
tion domains. This family of proteins has undergone extensive investigation and it is thought to have various roles including anchoring and trafficking of transmem-brane proteins (Fujita and Kurachi, 2000). Within the brain, the majority of SAP-97 can be detected at the postsynaptic density. But some of it can also be found cytoplas-mically and presynaptically (Aoki et al., 2001). It has been shown that SAP97 plays an important role in architecturing synaps-es (Thomas et al., 1997). GluR1 binds di-rectly to the PDZ domain of SAP97 (Cai et al., 2002; Leonard et al., 1998). This in-teraction allows GluR1 containing AMPA receptors to associate with many other proteins including PKA, PKC, calcineurin, calmodulin (Paarmann et al., 2002) and ki-nesin superfamily motor protein (Mok et al., 2002). SAP97 is capable of multi-merization with itself and other MAGUK proteins which is consistent with its role as anchoring protein (Feng et al., 2004; Kar-nak et al., 2002; Lee et al., 2002). Syn-aptic targeting of SAP97 is modulated by CaMKII (Mauceri et al., 2004).
The other PDZ-containing protein shown to interact with AMPA subunits through yeast-two hybrid screening is pro-tein interacting C-Kinase (PICK1) (Xia et al., 1999; Dev et al., 1999). A PDZ domain in PICK1 can interact with PKCα as well as GluR2 which implies that the two inter-actions are competitive. PICK1 has been shown to act as dimers and the dimers are capable of aggregating oligomeric target proteins. Indeed, it has been shown that PICK1 aggregates AMPA receptors in het-erologous expression systems (Xia et al., 1999; Dev et al., 1999).
4.2.2 Non-PDZ-domain containing proteinsApart from PDZ-domain-containing proteins, AMPA interacting proteins have expanded to a large scope of other proteins
which are indispensable for normal AMPA receptor function. Stargazin is a transmembrane protein isolated through its interaction with AMPA receptors (Vandernberghe et al., 2005). It is a member of a family of L-type calcium channel modulator γ subunits. These proteins are differentially expressed throughout the nervous system. Stargazin has a conserved N-terminal AMPAR binding domain and a C-terminal PDZ binding domain that can bind to PSD95, thus it plays a dual role in the traffi cking of AMPARs and synaptic assembling of these receptors (Tomita et al., 2005; Chen et al., 2000). Although PSD-95 does not directly associate with AMPA receptors it can influence the expression of these receptors through stargazin. Clustering the AMPA receptor complex is regulated by the palmitoylation state of PSD-95 and activity-dependent depalmitoylation results in the loss of synaptic AMPARs (Huang and El Husseini, 2005). The interaction between stargazin and PSD-95 also serves to link AMPARs to NMDARs and their signaling cascades (Lim et al., 2003; Chetkovich et al., 2002).
The other proteins that also interact with the intracellular C-terminal domain of AMPA receptors containing non-PDZ domain include N-Ethylmaleimide-sensitive factor (NSF) (Huang et al., 2005; Beretta et al., 2005; Gardner et al., 2005) and protein 4.1N (Shen et al., 2000). NSF has been reported to bind to a unique site located within the C-terminal of the GluR2 and GluR3 subunits of AMPA receptors (Song et al., 1998; Nishimune et al., 1998). NSF is a multihomomeric ATPase which plays essential role in protein traffi cking and membrane fusion (Song et al., 1998). Interestingly, NSF is strongly expressed in the postsynaptic density (Walsh and Kuruc, 1992). Blockade of the interaction of NSF with GluR2 by adenoviral
Review of the Literature

27
expression of the peptide or anti-NSF antibody results in a dramatic loss in the number of AMPA receptor aggregates and substantial decrease in evoked AMPA receptor mediated transmission (Beretta et al., 2005; Huang et al., 2005).
Protein 4.1 was originally isolated as cytoskeleton protein in erythrocytes (Hoover and Bryant, 2000). A neuronal homolog (4.1N) has been identifi ed that is expressed at high levels in the brain. 4.1N can be detected in the postsynaptic density of the neuron (Walensky et al., 1999). It has been found that both GluR1 and GluR4 can directly interact with 4.1N in the proximal region of their intracellular C-terminal (Coleman et al., 2003; Shen et al., 2000). The interaction between 4.1N and AMPARs is necessary for the surface expression of the receptors, links the receptors to actin cytoskeleton, and is involved in the formation of postsynaptic signaling complexes.
4.3 Dendritic spines Dendritic spines are small protrusions on the surface of dendrites and act as post-synaptic sites for most glutamatergic con-nections. AMPA and NMDA receptors usually are present within spines and form asymmetric synapses with presynaptic ter-minal. The morphological feature of den-dritic spines provides a postsynaptic bio-chemical and electrical compartment that separates the synaptic space from the den-dritic shaft and allows each spine to func-tion as a partially independent unit. Spine formation in pyramidal neurons occurs after birth, in consistency with ontogeny of synaptogenesis. Although spinogenesis is not exactly the same as synaptogenesis, there is a close relationship between the two as most spines are thought to serve as recipients of synaptic input. In rat neo-cortical pyramidal neurons, spine density increases continually during the fi rst post-
natal month, but subsequently is reduced with increasing age, refl ecting an initial overproduction and later elimination of synapses during early cortical develop-ment (Yuste and Bonhoeffer, 2004).
In addition to the change in density, spines undergo profound morphological rearrangement. The morphology of spines has been classifi ed as thin, stubby, mushroom and irregularly shaped head spines (Ethell and Pasquale, 2005) (Figure. 3). Early in development, stubby spines which lack clear necks are common. In mature neurons, mushroom spines which have more prominent necks and heads are more dominant. There is one type of dendritic protrusion, the fi lopodium, which occurs at a very early developmental stage, and is widely considered being a pre-form of dendritic spines. This transient structure is present mostly around the fi rst postnatal week, and is subsequently replaced by shaft synapses and stubby spines. Ultrastructural study on fi lopodia in rat neocortex showed that fi lopodia form synapses and receive up to 20% of the total synapses made on pyramidal neurons at this developmental stage. The data support the hypothesis that fi lopodia contribute to the generation of synapses on the dendritic shaft (Yuste and Bonhoeffer, 2004). Filopodia are highly dynamic. Imaging studies on dynamics of early dendritic protrusions from hippocampal slice cultures and dissociated cultures revealed that the appearance of shorter spine protrusions follows the disappearance of elongated fi lopodia. Highly motile dendritic fi lopodia become stabilized and transformed into spines. Several studies also postulate the existence of a ‘protospine’, the intermediate morphological structures that represent stabilized fi lopodia (Dailey and Smith, 1996; Ziv and Smith, 1996). It has also been shown by two-photon time lapse microscopy that stubby spines and other
Review of the Literature

28
type of spines can originate from fi lopodia in developing hippocampal neurons (Hering and Sheng, 2001). Interestingly, the opposite transformation (spine turning into fi lopodia) was also observed in the same study.
4.3.1 SpinogenesisUp to now, the mechanisms for spino-genesis are still not clear. Several models have been proposed based on studies in different type of neurons. One is referred to as the Sotelo model based on studies in Purkinje cells of cerebellum (Sotelo, 1990). The studies came from Weaver and Reeler mutant mice, both mutants have substantially loss of granule cells which are the presynaptic partners of around 90% of Purkinje cell spines. In these animals, Purkinje cells develop morphologically normal spines in the absence of presynaptic partner, even to the point of having normal postsynaptic specializations (Ethell and Pasquale, 2005). In normal developing cerebellar cortex, Larramendi and Victor already in late 1960th (Larramendi and Victor, 1967), pointed out that spines in the distal dendritic branches of Purkinje cells develop before they establish synaptic
contacts with parallel fi bers. Also at early postnatal stages (P0-P12), ‘naked’ spines without presynaptic terminals counterpart can be identifi ed (Yuste and Bonhoeffer, 2004). In this model, which was proposed by Sotelo (Sotelo, 1978; Yuste and Bonhoeffer, 2004), spine formation seems to be at least in some cases, an intrinsic, perhaps even cell-autonomous property of the neuron. The second model is based on terminal induced spine formation, proposed by Miller and Peters (Miller and Peters, 1981) according to their data from visual cortex studies. Synapses are made on the dendritic shaft fi rst, and can be recognized by axon terminal which is subsequently swollen as synaptic vesicles accumulate. Finally spines develop into thin or mushroom-shaped with a bulbous head and a clear neck (Miller and Peters, 1981; Yuste and Bonhoeffer, 2004). The hypothesis implies that presynaptic terminals act as inducers of spines from the dendritic shaft. However, more experiments are still needed to demonstrate this proposal, especially live imaging to show how spines emerge during development. The third is the fi lopodia model which proposed that fi lopodia act as spine precursors (Yuste
Figure 3. Classifi cation of dendritic spine morphology and ultrastructure. A. Morphological representation of fi lopodium and the most common types of dendritic spine. B. Dendritic spine ultrastructure. SV: synaptic vesicle; PSD: postsynaptic density (Adapted from Ethell and Pasquale, 2005).
Review of the Literature

29
and Bonhoeffer, 2004). Filopodia can catch axons, then engage in synaptic contact, and undergo a fi lopodia-to-spine transformation. This transformation involves a decrease in motility, substantial shortening, and enlargement of the distal portion of the fi lopodia to yield the shape of mature spines. Recently, live imaging of developing neurons in hippocampal tissue slices as well as in the neocortex in vivo has supported the hypothesis that synapse formation triggers the transformation of fi lopodia into spines (Dailey and Smith, 1996; Maletic-Savatic et al., 1999; Marrs et al., 2001; Okabe et al., 2001; Trachtenberg et al., 2002). This model also suggests that the high motility of fi lopodia is used to probe the space around for an appropriate contact site on the axon.
The different models outlined above summarize the descriptive data accumulated on how dendritic spines are formed, but the exact molecular mechanisms of spine formation are not clear. The studies on molecular signals that drive dendritic spine development and plasticity have expanded rapidly in recent years. Many of the signal cascades involved in these processes converge on the regulators of actin dynamics.
4.3.2 Molecular mechanisms regulating morphology of dendritic spinesActin is highly concentrated in spines, There are two types of actin, a soluble pool of monomeric G-actin and polymerized F-actin fi laments that confer the characteristic spine morphology (Ethell and Pasquale, 2005; Halpain, 2000; Rao and Craig, 2000). Multiple protein complexes and signal cascades contribute to actin rearrangement within spines. One major pathway involves the small guanosine triphosphatases (GTPases) of the Rho family. RhoA, Rac1 and Cdc42 of this
family have now been well characterized in the regulation of spine morphology (Ethell and Pasquale, 2005).
A dominant-negative form of Rac1 drastically decreases the number of both spines and synapses in cultured hippocampal slices and dissociated hippocampal cultures (Nakyama et al., 2000; Penzes et al., 2000). Overexpression of dominant-negative Cdc42 in dissociated hippocampal cultures inhibits spine morphogenesis (Irie and Yamaguchi, 2002). In another loss of function study on Drosophila visual system, there is significant reduction in the density of spine-like structures (Scott et al., 2003). Furthermore, overexpression of constitutively active RhoA in hippocampal slices promotes spine retraction and elimination (Govek et al., 2004; Tashiro et al., 2000). Inhibition of RhoA activity by expressing the C3 transferase results in more spines in some neurons and spines with long necks and resembling fi lopodia in other neurons (Ethell and Pasquale, 2005). These experiments clearly demonstrated the distinct effects of Rho GTPase family members in spine morphogenesis.
Rac1 and Cdc42 promote the development of new spines and maintenance of normal spine morphology and RhoA activity is necessary for shaping the rounded spine head and for mature spine morphogenesis. Therefore, the activities of Rac1, Cdc42 and RhoA more likely coordinate each other temporally and spatially which is critical for dendritic spine development and remodeling.
Molecules regulating activity of different Rho family GTPase are now being defi ned. Kalirin and Intersectin are multidomain exchange factors that have been shown to regulate actin reorganization in dendritic spines in response to cell surface signals by Eph receptor tyrosine
Review of the Literature

30
kinases (Irie and Yamaguchi, 2002; Penzes et al., 2000; Schmidt and Hall, 2002). Kalirin-5 and Kalirin-7 have been found to contain a PDZ domain-binding motif that targets them to the postsynaptic density (Johnson et al., 2000). Experiments using antisense approaches demonstrated that Kalirin expression is necessary for the regulation of dendritic spine morphology acting through Rac1 signaling to the actin cytoskeleton (Ma et al., 2001). Another Rac1 and Cdc42 exchange factor that has recently been implicated in dendritic spine morphogenesis is the β Pak-interacting exchange factor (βPIX), also known as cool-1 (Zhang et al., 2003). βPIX is transported to the synaptic region by binding to the adaptor protein, G protein-coupled receptor kinase-interacting protein 1 (GIT1). Disruption of synaptic localization of βPIX by mutating its GIT1 binding sites causes an increase in dendritic protrusions but a decrease in mature dendritic spines and synapses (Zhang et al., 2003).
Many actin-binding proteins that function down-stream of the GTPases of the Rho family have been shown to control the organization of the actin cytoskeleton in dendritic spines (Table 2).
The integrity of the postsynaptic density is important for normal spine morphology besides the proteins involved in Rho signaling pathways. Many of the postsynaptic density proteins have multiple PDZ domains that recognize short amino acid sequences at the carboxy terminus (PDZ binding motif) of other proteins (Kim and Sheng, 2004). They also bind to each other, contributing to the formation of diverse multiple protein complexes. PSD-95 is a major constituent of the postsynaptic density (Cho et al., 1992; Sheng, 2001). It directly binds to the NR2 subunit of the NMDA receptor. Overexpression of PSD-95 promotes
enlargement and maturation of spines by increasing AMPA receptor content (Cai et al., 2006; El Husseini et al., 2000). Lack of PSD-95 causes abnormalities in synaptic plasticity and learning. PSD-95 interacts with NMDA receptors at the cell surface, and downstream binds to guanylate kinase-associated protein (GKAP), which in turn binds to Shank. Shank associates with Homer proteins (Naisbitt et al., 1999; Tu et al., 1998). Consistent with its interaction with PSD-95, overexpression of Shank and Homer accelerates the maturation of spines and promotes the enlargement of mature spines (Sala et al., 2001). Shank is also found to interact with the actin-binding proteins Cortactin (Naisbitt et al., 1999), Abp1 (Qualmann et al., 2004), and the Rac1 and Cdc42 exchange factor βPIX (Park et al., 2003).
Although the dendritic spine is a very small structure, its molecular regulation involves the interactions of a huge collection of signal and structural proteins. Many of them still remain to be discovered. The intricate regulatory mechanisms of spines support their important role in brain plasticity and learning. More sophisticated work is needed to investigate how individual molecules interrelate with each other in the network and react to different stimulations.
5. Excitatory and inhibitory synaptogenesis
What are the molecular mechanisms for synapse formation is a critical question for understanding how neuronal circuitries are established. The mechanisms are still not very clear in CNS. An important step has been the cloning of gephyrin. Gephyrin is highly concentrated in the synaptic densities at glycinergic and some GABAergic synapses (Prior et al., 1992; Langosch et al., 1992; Betz et al., 1991;
Review of the Literature
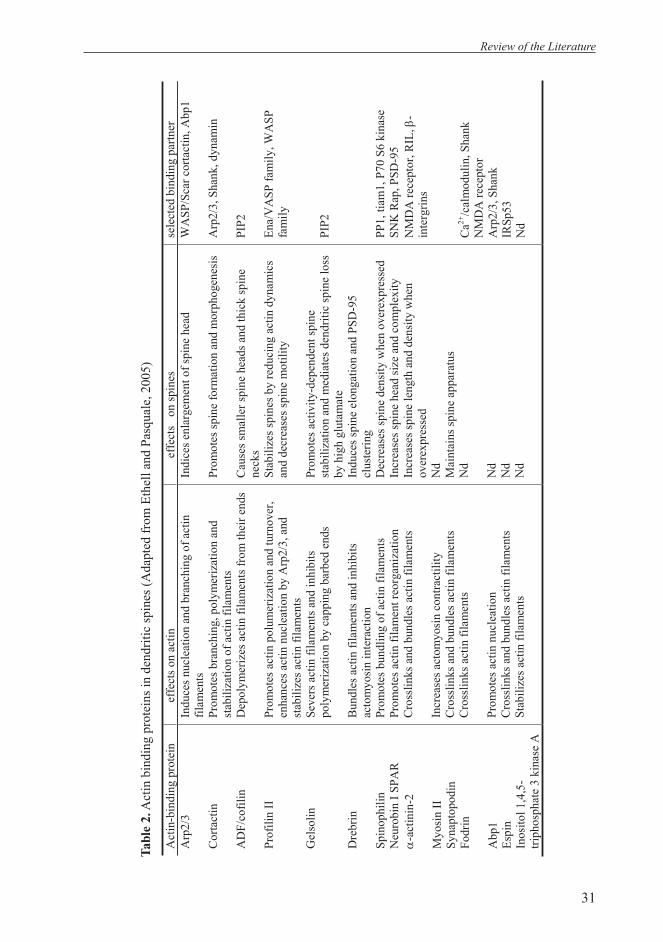
31
Review of the Literature
Act
in-b
indi
ng p
rote
in
effe
cts o
n ac
tinef
fect
s o
n sp
ines
sele
cted
bin
ding
par
tner
A
rp2/
3
Cor
tact
in
AD
F/co
filin
Prof
ilin
II
Gel
solin
Dre
brin
Spin
ophi
lin
Neu
robi
n I S
PAR
α
-act
inin
-2
Myo
sin
II
Syna
ptop
odin
Fo
drin
Abp
1 Es
pin
Inos
itol 1
,4,5
-tri
phos
phat
e 3
kina
se A
Indu
ces n
ucle
atio
n an
d br
anch
ing
of a
ctin
fil
amen
ts
Prom
otes
bra
nchi
ng, p
olym
eriz
atio
n an
d st
abili
zatio
n of
act
in fi
lam
ents
D
epol
ymer
izes
act
in fi
lam
ents
from
thei
r end
s
Prom
otes
act
in p
olum
eriz
atio
n an
d tu
rnov
er,
enha
nces
act
in n
ucle
atio
n by
Arp
2/3,
and
st
abili
zes a
ctin
fila
men
ts
Seve
rs a
ctin
fila
men
ts a
nd in
hibi
ts
poly
mer
izat
ion
by c
appi
ng b
arbe
d en
ds
Bun
dles
act
in fi
lam
ents
and
inhi
bits
ac
tom
yosi
n in
tera
ctio
n Pr
omot
es b
undl
ing
of a
ctin
fila
men
ts
Prom
otes
act
in fi
lam
ent r
eorg
aniz
atio
n
Cro
sslin
ks a
nd b
undl
es a
ctin
fila
men
ts
Incr
ease
s act
omyo
sin
cont
ract
ility
C
ross
links
and
bun
dles
act
in fi
lam
ents
C
ross
links
act
in fi
lam
ents
Prom
otes
act
in n
ucle
atio
n
Cro
sslin
ks a
nd b
undl
es a
ctin
fila
men
ts
Stab
ilize
s act
in fi
lam
ents
Indi
ces e
nlar
gem
ent o
f spi
ne h
ead
Prom
otes
spin
e fo
rmat
ion
and
mor
phog
enes
is
Cau
ses s
mal
ler s
pine
hea
ds a
nd th
ick
spin
e ne
cks
Stab
ilize
s spi
nes b
y re
duci
ng a
ctin
dyn
amic
s an
d de
crea
ses s
pine
mot
ility
Prom
otes
act
ivity
-dep
ende
nt sp
ine
stab
iliza
tion
and
med
iate
s den
driti
c sp
ine
loss
by
hig
h gl
utam
ate
Indu
ces s
pine
elo
ngat
ion
and
PSD
-95
clus
terin
g D
ecre
ases
spin
e de
nsity
whe
n ov
erex
pres
sed
Incr
ease
s spi
ne h
ead
size
and
com
plex
ity
Incr
ease
s spi
ne le
ngth
and
den
sity
whe
n ov
erex
pres
sed
Nd
Mai
ntai
ns sp
ine
appa
ratu
s N
d
Nd
Nd
Nd
WA
SP/S
car c
orta
ctin
, Abp
1
Arp
2/3,
Sha
nk, d
ynam
in
PIP2
Ena/
VA
SP fa
mily
, WA
SP
fam
ily
PIP2
PP1,
tiam
1, P
70 S
6 ki
nase
SN
K R
ap, P
SD-9
5 N
MD
A re
cept
or, R
IL, β
-in
terg
rins
Ca2+
/cal
mod
ulin
, Sha
nk
NM
DA
rece
ptor
A
rp2/
3, S
hank
IR
Sp53
N
d
Tab
le 2
. Act
in b
indi
ng p
rote
ins i
n de
ndrit
ic sp
ines
(Ada
pted
from
Eth
ell a
nd P
asqu
ale,
200
5)

32
Studer et al., 2006; Jacob et al., 2005; Li et al., 2005). Knock-out and overexpression studies demonstrated that gephyrin is responsible for clustering of postsynaptic receptors at inhibitory synapses (Kneussel and Betz, 2000). It links the receptors to the underlying cytoskeleton. For excitatory synapse formation, the molecules involved in spinogenesis that are described above also account for glutamatergic synapse formation, for example: glutamate receptors clustering for NMDA and AMPA receptors types are facilitated by a class of PDZ proteins, such as PSD95 and SAP97 (O’Brien et al., 1998; Cai et al., 2006). These proteins, interacting directly with glutamate receptor subunits and scaffold proteins such as Shank and Homer have been showed to promote excitatory synapse maturation (Kim and Sheng, 2004; El Husseini et al., 2000).
Intercellular adhesion is an additional important facet for central synapse formation. Matched adhesion molecules on pre- and postsynaptic membranes interact with each other providing tight association and specifi city between synaptic partners. For example, cadherins which bind homophilically are concentrated at synaptic sites in both pre-and postsynaptic membranes and link to the cytoskeleton via catenins (Bamji, 2005; Salinas and Price 2005; Goda, 2002). The presence of distinct cadherins at different synapses could play a role in the selectivity of synapse formation (Colman, 1997). β-neurexins and neuroligins (NLs) that bind to each other heterophilically are also present at synapses (Lise and El Husseini, 2006; Cline 2005; Hussain
and Sheng, 2005). Both bind to PDZ domain-containing proteins, such as PSD-95 (Levinson et al., 2005). Studying the physiological roles of these interactions yielded much progress as shown by recent publications (Chih et al., 2005; Graf et al., 2004; Prange et al., 2004). They provide strong evidence that β-neurexins-neuroligin signaling promotes synapse formation, both excitatory synapses and inhibitory synapses. Moreover, the association of neuroligin with PSD-95 may control the balance between excitatory and inhibitory synapses (Prange et al., 2004). Expression of NL1 alone induces the formation of both excitatory and inhibitory synapses (Chih et al., 2005). However, when coexpressed with PSD-95, the affect of NL1 was restricted to excitatory synapses. Another intriguing fi nding is that overexpression of PSD-95 was able to redistribute endogenous NL2 from inhibitory to excitatory synapses and enhances the formation of excitatory synapses at the expenses of inhibitory synapse formation (Graf et al., 2004). On the other hand, knock down of NLs either individually or collectively, results in a substantial decrease in inhibitory synaptic transmission, with relatively little effect on excitatory synaptic transmission, thus disturbing the excitatory/inhibitory (E/I) synaptic balance (Chih et al., 2005). These fi ndings provide a new clue to the mechanisms of controlling CNS synapse formation and E/I ratio. The regulation of expression and stoichiometry between cell adhesion molecules and scaffolding proteins will signifi cantly affect neuronal wiring.
Review of the Literature

33
Aims of the Study
Aims of the Study
The aims of this study are to investigate the functional role of KCC2 in neuronal development and the molecular mechanisms involved.
1. To characterize the expression patterns of KCC2 in developing rodent brain.2. To examine the dendritic morphology of primary cultured KCC2-/- neurons.3. To analyze the mechanisms that underlie the role of KCC2 in dendritic spine
formation4. To study the mechanisms that regulate KCC2 expression.
34
Materials and Methods
Materials and methods
Detailed information of materials and methods used in this thesis are described in the original papers (I-IV).
1. AnimalsEmbryos used for developmental expression and primary neuronal cultures are from timed-pregnant Sprague-Dawley rats and C56BL/6 mice from the local animal house (University of Helsinki). We used day 16 embryos in this study for primary neuronal culture. All experiments were approved by the Local Ethics Committee for Animal Research at the University of Helsinki
Primary neuronal and organotypic slice cultures of KCC2 -/- were obtained from crossing of heterozygote (KCC2+/-) females and heterozygote males (Tornberg et al. 2005).
2. Methods used and described in articles I-IV
Probes Accession/Source Nucleotides (Insert bp)
Mouse KCC1 AA185691 (EST) 3295-3746 (495bp)
Mouse KCC2 AA982489 (EST) 4605-5566 (1039bp)
Rat KCC2 Payne et al. (1996) 5-834 (830bp)
Mouse KCC3 BF020529 (EST) 2832-3348 (682bp)
Mouse KCC4 A1592647 (EST) 2508-3248 (745bp)
Mouse KCC4 RT-PCR 72-275 (203bp)
Mouse NKCC1 AA980272 (EST) 3498-4346 (964bp)
Mouse NKCC1 RT-PCR 2472-3006 (265bp)
In situ hybridization IImmunohistochemistry IRT-PCR IPrimary neuronal culture II, III, IVImmunocytochemistry II, III, IVFluorescence imaging of active synaptic terminals IIIElectrophysiology and local photolysis of caged GABA IIIAntibody production II, IIIImmunoprecipitation IIIOrganotypic slice culture IIITransfection III
2. ProbesThe probe used for in situ hybridizations are listed in article I:

35
Results and Discussion
1. Developmental expression of KCC2 follows neuronal maturation in vivo (I) and in vitro (II).
The expression of KCC2 in mature brain has been well characterized (Kanaka et al., 2001; Blaesse et al, 2006; Payne et al., 1996). Later, the analysis on the postnatal expression pattern in hippocampus led to the discovery of its significant role in the developmental shift of GABAA mediated responses from depolarizing to hyperpolarizing (Rivera et al., 1999). However, there is little information on the expression patterns of KCC2 in embryonic CNS. In order to cast light on this subject we used in situ hybridizations to show that KCC2 mRNA was already expressed in the ventral part of the cervical spinal cord as early as E12.5 in rat and mouse embryos. No signals were observed in higher brain regions at this stage (I, Fig. 2A). Similar results were also reported by Hubner et al (2001) that mouse spinal cord motoneurons express KCC2 as early as E12.5 and the expression spreads over the complete spinal cord by E18.5. As brain develops, the transcripts spread towards other differentiating parts of the CNS including the diencephalon, mesencephalon and rhombercephalon, coinciding with the up-regulated expression of KCC2 described previously (Lu et al., 1999; Rivera et al., 1999). In the diencephalon, high mRNA expression was restricted to the ventral thalamus and ventral lateral geniculate complex. Dorsal thalamus had a moderate expression level of the mRNA. In the brainstem, KCC2 expression was particularly high in tegmental region as well as in the caudal part of the spinal nucleus of trigeminal nerve (I, Fig 2B).
A strong hybridization signal of KCC2 transcripts was also found in the ventral parts of the developing olfactory bulb (I, Fig. 2Ad insert). Neurons in olfactory bulb mantle layer are born as earlier as E10 (Altman and Bayer, 1995). KCC2 is expressed in these early developing regions by E14.5, but not in the isocortex, which differentiates much later than the olfactory bulb and basal nuclei. It was not possible to detect KCC2 mRNA expression in isocortex of telencephalon (that develops into neocortex and hippocampus) until postnatal day 0 (P0) in mouse (I, Fig2Af). In newborn mouse and rat brain, KCC2 mRNA levels were high in the olfactory bulb, but very low in cortex and anterior thalamic regions of the rat. In new born mouse (but not in rat), the hippocampus and cerebellum already displayed low levels of KCC2 mRNA expression (I, Fig 2Al). In general, the development of KCC2 expression followed the rostral-caudal axis of neuronal maturation.
KCC2 mRNA was present within the areas that were labeled with the class III β-tubulin specifi c antibody TUJ-1 (a marker for differentiated neurons), for example, in the basal ganglia (Menezes & Luskin, 1994; Owens & Kriegstein, 1998; but see Menezes et al., 1995). But in the ventricular zone, where neurogenesis is ongoing, or in the intermediate zone, where neurons are migrating, there was no detectable KCC2 mRNA. By combining immunostaining and in situ hybridization, we found that wherever there was KCC2 hybridization signal, TUJ-1 immunostaining was positive. But not all migrating and postmitotic neurons labeled by TUJ1 express KCC2, for example those seen in the isocortex preplate (I, Fig.3). These results indicate that migrating and
Results and Discussion

36
postmitotic neurons have to develop into certain stage to express KCC2. We found also that, at a given embryonic age, the development of KCC2 expression was more advanced in mouse brain than in rat in according with more rapid development of the mouse embryo (I, Fig. 2A). These observations give additional support to KCC2 as an indicator of neuronal maturation.
Results on KCC1, KCC3, KCC4 and NKCC1 indicated that KCC1 expression was generally low in embryonic brain and show slightly up-regulation until birth (I; Fig. 1A). The expression of KCC1 was only detectable in the developing choroid plexus as early as E14,5. KCC3 mRNA was scarce in the cortical plate at E14.5 and slightly up-regulated at birth. Although the KCC3 expression level was low, we could confi rm its expression with RT-PCR from different brain regions like telencephalon, diencephalon and mesencephalon (I, Fig. 4). There was no hybridization signal for KCC3 in the ventricular layer and developing choroid plexus. In contrast, KCC4 mRNA was abundantly expressed in the ventricular zone and was down-regulated perinatally (I; Fig. 5). Detected as early as E12.5 in mouse embryos, KCC4 expression was associated with proliferative regions where neurogenesis takes place. It was also found that there was expression of KCC4 in other regions at E14.5 including the spinal nucleus of the trigeminal nerve in the rhombercephalon and vestibular ganglion. KCC4 expression was decreased at around P0, but it was detected in the choroid plexus and other epithelial layers as well as the peripheral ganglia. NKCC1 was highly expressed in the vimentin-positive radial glia of the proliferative zone of the subcortical region indicating that expression of NKCC1 was closely associated with neurogenesis (I;
Fig. 7). Neuronal progenitor cells in the ventricular zone go through several cell cycles to generate postmitotic neurons. An increase in cell size is an important variable in determining the timing of cell division (Albert et al., 1994; Gao and Raff, 1997). By controlling the cell size, NKCC1, a cotransporter known to mediate regulatory volume increase (Russell, 2000), might be involved in the control of the neuronal cell cycle. At later embryonic stages, there was a shift in NKCC1 expression to the neuron specifi c class III β-tubulin positive region of the cortical plate. Functional relevance for this expression shifting remains for further studies. An intensive signal of NKCC1 expression was observed in developing choroid plexus throughout development. As shown previously (Clayton et al., 1998), KCC1 was also expressed in the adult choroid plexus. Thus KCC1 and NKCC1 appear to regulate the ionic concentration of the cerebral fl uid and to set the internal Cl concentration within choroid plexus cells.
The distinct distributions of KCCs in embryonic brain indicate different roles of KCCs in neuronal development. In addition to above in vivo expression studies, we then focussed on the role of KCC2 and went on to further analyze the expression of KCC2 in hippocampal dissociated and organotypic cultures. The expression of KCC2 is developmentally up-regulated in both cases. In dissociated cultures, KCC2 immunoreactivity was fi rst observed after 3 days in vitro (DIV). The localization of the protein at this age was mainly somatic. KCC2 expression increased as the culture developed and spread from cell bodies to neurites. Double staining with MAP2 antibody showed that KCC2 positive neurites were exclusively dendrites (II; Fig. 2A). To quantify the
Results and Discussion
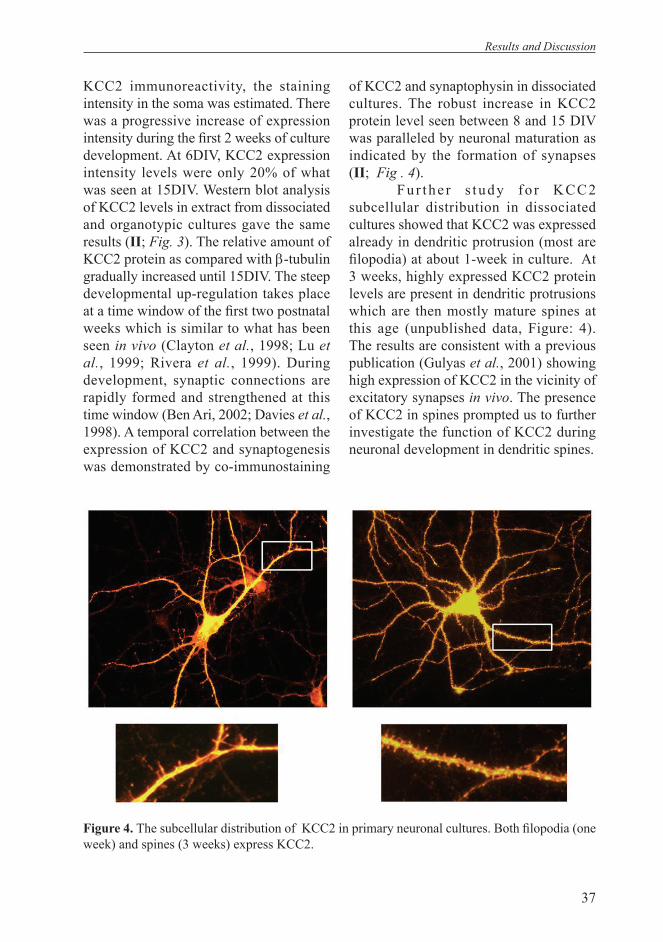
37
KCC2 immunoreactivity, the staining intensity in the soma was estimated. There was a progressive increase of expression intensity during the fi rst 2 weeks of culture development. At 6DIV, KCC2 expression intensity levels were only 20% of what was seen at 15DIV. Western blot analysis of KCC2 levels in extract from dissociated and organotypic cultures gave the same results (II; Fig. 3). The relative amount of KCC2 protein as compared with β-tubulin gradually increased until 15DIV. The steep developmental up-regulation takes place at a time window of the fi rst two postnatal weeks which is similar to what has been seen in vivo (Clayton et al., 1998; Lu et al., 1999; Rivera et al., 1999). During development, synaptic connections are rapidly formed and strengthened at this time window (Ben Ari, 2002; Davies et al., 1998). A temporal correlation between the expression of KCC2 and synaptogenesis was demonstrated by co-immunostaining
of KCC2 and synaptophysin in dissociated cultures. The robust increase in KCC2 protein level seen between 8 and 15 DIV was paralleled by neuronal maturation as indicated by the formation of synapses (II; Fig . 4).
Fu r the r s tudy fo r KCC2 subcellular distribution in dissociated cultures showed that KCC2 was expressed already in dendritic protrusion (most are fi lopodia) at about 1-week in culture. At 3 weeks, highly expressed KCC2 protein levels are present in dendritic protrusions which are then mostly mature spines at this age (unpublished data, Figure: 4). The results are consistent with a previous publication (Gulyas et al., 2001) showing high expression of KCC2 in the vicinity of excitatory synapses in vivo. The presence of KCC2 in spines prompted us to further investigate the function of KCC2 during neuronal development in dendritic spines.
Figure 4. The subcellular distribution of KCC2 in primary neuronal cultures. Both fi lopodia (one week) and spines (3 weeks) express KCC2.
Results and Discussion

38
2. The role of KCC2 in spine formation (III)
2.1 Aberrant morphology of dendritic spines in KCC2-/- cortical neurons and KCC2hy/null neuronsThe presence of KCC2 in spines raised the question whether KCC2 would have a novel function at excitatory synapses. We cultured cortical neurons from KCC2 knock-out embryos and examined the difference in morphology of dendritic spines in KCC2-/- neurons and wild type (WT) neurons. After about two weeks in culture, the dendritic spines of pyramidal-shaped cortical neurons from KCC2-/- mice were significantly longer than wild-type neurons. We also observed that many of these protrusions were clearly branched. Interestingly, there was no clear differences in the density of dendritic protrusions between KCC2-/- and wild type neurons (III Fig. 1A-F).
To test for the possibility that hyperexcitability induced by the lack of functional hyperpolarizing inhibition caused the abnormal spine morphology, we cultured KCC2-/- neurons in the continuous presence of TTX. There was no difference observed between the two cultures. In order to investigate a possible tonic GABAA mediated effect on spine morphology, we applied 10 uM bicuculline at 8DIV. However, bicucullin was not able to block the spine phenotype of KCC2 -/- neurons (III Fig. 1E, F) . These results indicate that the phenotype of dendritic protrusion in KCC2-/- neurons is not due to hyperexcitability or tonic actions of GABAA.
To examine the regulatory role of KCC2 in spine development in vivo, we used compound heterozygous mice for KCC2 null and hypomorphic alleles (KCC2hy/null) which express 17% of WT KCC2 protein levels but are viable
(Tornberg et al., 2005). Neocortical acute slices from these mice (P16) were intracellularly loaded with biocytin and dendritic protrusions were analyzed. The protrusions were significantly longer in KCC2hy/null than WT neurons. There was no difference in spine density observed between genotypes (III Fig. 1G-L).
2.2 Functional excitatory synapses reduced in KCC2-/- neuronsGlutamatergic synapses are formed on dendritic spines in neocortical neurons. To examine the maturation of excitatory synapses in KCC2-/- neurons, we used double immunostaining for presynaptic marker, VGLUT1, with either postsynaptic marker PSD-95, a postsynaptic density protein mainly present in mature excitatory synapses (Kim and Sheng, 2004) and Homer. Our results showed significant reductions in VGLUT1 positive PSD-95 clusters and VGLUT1 positive Homer clusters when compared KCC2-/- neurons with the wild type but no difference was observed in the soma (III, Fig. 2A-D). Another experiment was performed using the styryl dye SynaptoRed (III, Fig. 2E-G) to estimate active synapse number. When loaded onto the cultured neurons, we found that the number of active presynaptic elements targeting dendrites is significantly reduced in dendrites of KCC2-/- neurons. No clear difference was observed at neuronal soma. Consistent with the aberrant morphology of dendritic protrusions in the KCC2-/- neurons, the above data indicate a prominent reduction in the number of functional synapses. Further evidence supporting this conclusion was obtained by recording excitatory miniature postsynaptic currents (mEPSCs): there was a signifi cantly lower frequency of mEPSCs in KCC2-/- neurons but no difference in amplitude as compared to WT neurons (III, Fig. 2H-J)
Results and Discussion

39
2.3. KCC2-ΔNTD lacking K-Cl transport activity restores normal spine morphology in KCC2-/- neuronsIn order to examine whether a mechanism based on the regulation of neuronal [Cl]i underlies the action of KCC2 on the development of spine morphology, we constructed a N-terminal deleted KCC2 mutant (KCC2-ΔNTD) with the expectation that it is not capable of K-Cl cotransport. Previous work has shown that deletion of the N-terminal domain of the KCC1 results in a K-Cl cotransport mutant that is incapable of Cl- transport (Casula et al., 2001). To evaluate the chloride extrusion effi cacy of the KCC2 constructs used in the present study we employed a functional assay based on the somato-dendritic gradient of the reversal potential (EGABA) of GABAA receptor-mediated current responses induced along the dendrite by laser fl ash photolysis of caged GABA (Khirug et al., 2005) When comparing full length KCC2 (KCC2-FL) and KCC2-ΔNTD for their effi cacy to transport K-Cl in KCC2-/- neurons, we found that in KCC2-FL transfected neurons, the net Cl efflux was similar to more mature WT neurons (14-22 DIV), but in KCC2-ΔNTD transfected neurons, no difference was observed when compared with control EGFP transfected neurons (III; Fig. 3A, B). In addition to the electrophysiological assays above, we tested the functionality of the constructs using a classical 86Rb flux assay of K-Cl cotransport in HEK-293 cells. In full agreement with the results obtained in neurons, HEK-293 cells expressing the KCC2-FL construct induced a pronounced furosemide-sensitive 86Rb flux whereas cells expressing either KCC2-∆NTD or EGFP displayed small fluxes that were not signifi cantly above the baseline level. When transport activity was stimulated
with NEM, a well-known functional activator of K-Cl cotransport, only the cells that were transfected with KCC2-FL showed a robust increase in the 86Rb fl ux (III, Fig. 3C)
We ectopically expressed these two constructs in KCC2-/- neurons and examined their ability to restore altered spine morphology. KCC2-FL expressing neurons display normal spine morphology as we expected. Surprisingly, KCC2-∆NTD transfected neurons were able to fully restore normal spine morphology (III , Fig. 3D-F) and funct ional excitatory synapses as demonstrated by immunostaining of PSD95 and mEPSC recordings (III, Fig. 4A-E). The numbers of dendritic protrusions was not signifi cantly different between control and neurons transfected by each constructs. These results demonstrated that the role of KCC2 in the maturation of spines is not based on an effect on neuronal Cl- regulation and indicates a structural role that underlies the mechanism of KCC2 in spine formation.
We a lso t ransfec ted another cotransporter isoform, KCC3, as a control experiment to further demonstrated the specifi city of KCC2-FL and KCC2-∆NTD in rescuing the phenotype. The expression of KCC3-FL in KCC2-/- neurons was not able to restore normal spine morphology (III, Fig. 3D-F). The density of dendritic protrusions was not signifi cantly different between control neurons.
2.4. Overexpression of KCC2 C-terminal domain in WT neurons induces elongation of dendritic protrusions.We then used the KCC2 C-terminal domain (KCC2-CTD) to transfect wild type neurons. We assume that the effect of KCC2 in spine formation could be
Results and Discussion

40
mediated by its C-terminal interaction with other proteins and overexpression of KCC2-CTD may compete for the KCC2 intracellular interaction partner. Indeed, the phenotype in the transfected neurons was very similar to the phenotype of the KCC2 -/- neurons (III; Fig. 5A, B). There was signifi cant reduced functional presynaptic buttons as demonstrated by SynaptoRed loading experiment (III; Fig. 5D, E). No signifi cant change in the number of protrusions was observed in KCC2-CTD or KCC3-CTD transfected neurons (III; Fig. 5C). This experiment suggests that the C-terminal domain of KCC2 is necessary for normal dendritic spine morphogenesis. KCC2-CTD has a dominant negative effect, blocking the binding between KCC2 and an interaction protein that is important for normal spine development.
2.5. The interaction of KCC2-CTD with 4.1N underlies the function of KCC2 in spine morphologyConsidering the role of the KCC2 C-terminus in spine formation, we set out to fi nd the interacting partners of KCC2 C-terminal within spines. Dendritic spines are the main target of excitatory synapses, containing a variety of glutamate receptors, signaling molecules, and scaffold proteins in conjunction with actin-rich cytoskeleton (Ethell and Pasquale, 2005; Hering and Sheng, 2001). We screened mouse brain homogenate with a number of antibodies against glutamate receptor subunits (GluR1-4) and other proteins enriched in spines, including PSD-95, CRIPT, SAP97, GRIP, CaMKII and 4.1N, to test KCC2 immunoactivity from their immunoprecipitates. Out of these, the antibody against the cytoskeleton interacting protein 4.1N was the only one able to precipitate KCC2 (III; Fig. 6A).
4.1N is a neuronal isoform of the cytoskeleton associated protein 4.1 family that it is well-known to participate in stabilizing the spectrin/actin cytoskeleton (Hoover and Bryant, 2000). Thus, we went on to further analyze the interaction between 4.1N and KCC2. This interaction was further demonstrated by co-immunoprecipitation using antibodies against both the C-terminal and N-terminal epitopes of KCC2. Co-expression of KCC2 and myc-tagged 4.1N in HEK293 cells resulted in a clear myc-4.1N band in anti-KCC2 immunoprecipitates. Similarly, the anti-myc antibody could also bring down KCC2 (III; Fig. 6C, D).
The molecular interaction domains within KCC2 and 4.1N that are responsible for the interaction were further investigated by expressing a truncated form of KCC2 and 4.1N. We found that KCC2 C-terminal interacts with the FERM domain (Hoover and Bryant, 2000) of 4.1N (III; Fig. 6E, F).
In this study, we found a novel role of KCC2 in spine formation. And interestingly, the role is mediated by protein-protein interaction, independent of the well-established function of KCC2 as a K-Cl cotransporter. We showed that in the absence of KCC2, the morphology of dendritic protrusions is altered and functional synapses could not develop properly. We also demonstrated the 4.1N directly links the KCC2 C-terminus and the spine cytoskeleton. The actin-rich cytoskeleton of spines is critical for spinogenesis and also for the maintenance and plasticity of mature synapses (Halpain, 2000; Matus, 2000; Segal and Andersen, 2000; Yuste and Bonhoeffer, 2004). 4.1N is thought to have a central role in mediating structural interactions among the cytoskeleton, transmembrane proteins as well as adhesion molecules (Hoover
Results and Discussion

41
and Bryant, 2000). Both KCC2 and 4.1N are abundant in neurons during early development at the time of synaptogenesis and their expression is steeply correlated with the maturation of excitatory synapses (Gulyas et al., 2001; Rivera et al., 1999; Walensky et al., 1999).
The present fi nding of a novel role of KCC2 in spine formation shows that KCC2 participates in coordinating the functional maturation of excitatory and inhibitory neurotransmission. During a critical period of brain development, GABA responses shift from depolarizing and sometimes even excitatory to inhibitory, in parallel with the maturation of glutamatergic synapses. The dual role of KCC2 in chloride regulation and spine maturation during development is important in coordinating the two systems. Until now only some molecules have been shown to participate in balancing excitatory and inhibitory synapse formation, e.g. Neuroligin and its interaction with PSD-95. We provide evidence here that KCC2 acts as another molecular synchronizer having a significant role in functional neuronal network formation.
3. Regulation of KCC2 expression during development and plasticity (II, IV)Previously we have shown that KCC2 expression appears as earlier as 3DIV in dissociated cultures before synapse formation, and is up-regulated following neuronal maturation. To examine whether neuronal activity and synaptic transmission affect the developmental up-regulation of KCC2 in dissociated and organotypic cultures (see Ganguly et al., 2001), we used chronic treatment on hippocampal cultures with the action-potential blocker TTX (10μM); NMDA and non-NMDA antagonist, APV (40μM) and NBQX
(10μM); as well as the GABAA receptor blocker PTX (50μM). Measurements of KCC2 immunostaining intensity did not reveal any signifi cant alteration in KCC2 developmental up-regulation in cultures treated with various combinations of the blockers (II; Fig. 5). The results were further confi rmed by western blot analysis. These data show that neither neuronal spiking nor ionotropic glutamatergic and GABAergic transmission are required for the developmental expression of KCC2 in mouse hippocampal neurons in vitro.
The data presented do not rule out the possibility that expression of KCC2 could be modulated by trophic factors, such as brain-derived neurotrophic factor (Aguado et al., 2003) or insulin-like growth factor1 (Kelsch et al., 2001). Indeed, when studying the expression of KCC2 during plasticity, we observed down-regulation of KCC2 mRNA and protein levels by spontaneous activity. In these experiments, hippocampal slices were exposed to a solution devoid of Mg 2+ (0- Mg 2+). After 1-3 hours, RT-PCR from the slices showed that there is a progressive fall of KCC2 mRNA that starts within 30min exposure. Hippocampal slices in Mg 2+ -free solution usually generate continuous interictal-like spontaneous activity (Anderson et al., 1986; Mody et al., 1987), which commences at ~10-15 min and shows a progressive increase in frequency during the 1-3hr recording sessions. Western blot analyses showed a pronounced decrease of KCC2 protein during 0- Mg 2+ exposure for 1-3 hr (IV; Fig. 1). The down-regulation of KCC2 expression was not caused by the withdrawal of Mg 2+ per se but by the consequent discharge activity, because pharmacologically blocking the activity with NBQX plus AP-5 retained KCC2 mRNA as well as protein at a level that was indistinguishable from parallel controls.
Results and Discussion

42
This activity-induced down-regulation displayed an area-specifi c pattern shown by KCC2 immunofluorescent staining in the slices. The decrease was most prominent in the CA1 region than other regions (IV; Fig. 2).
BDNF-TrkB-mediated signaling has been found to be a major molecular cascade responsible for changes in protein expression required for plasticity such as LTP and epileptogenesis. To investigate the molecular mechanism for activity-induced downregulation of KCC2, we superfused acute hippocampal slices with 0-Mg2+ solution for 3 hr in the presence or absence of the tyrosine kinase inhibitor K252a, which prevents TrkB activation by inhibiting autophosphorylation of the receptors. KCC2 mRNA and protein levels remained at control levels in the presence of K252a (IV; Fig. 6). Control experiments showed that K252a had no effect on 0-Mg2+ induced interictal-like spontaneous activity. Similar results were obtained when TrkB receptor bodies (TrkB-Fc) were applied to slices to scavenge endogenously released BDNF. These results demonstrated that TrkB activation by endogenous BDNF is required for the activity-induced down-regulation of KCC2 .
BDNF-TrkB intracellular signaling is activated through the autophosphorylation of tyrosine residues of the intracellular domain of TrkB. There are two tyrosine phosphorylation residues creating docking sites for recruiting signaling adaptor proteins linked to PLCγ (Tyr 816), and Shc/FRS-2 (Tyr 515) (Bibel and Barde, 2000; Minichiello et al., 2002). These two docking sites mediate two different signaling cascades: PLCγ activates the CaM kinase/IP3 kinase signal pathway, and Shc/FRS-2 the Ras/MAPK pathways. To further examine which signaling pathway,
downstream of TrkB, is involved in the downregulation of KCC2, we analyzed two different transgenic mice, one with a targeted mutation in the PLCγ docking site of TrkB (TrkBPLC/PLC), and the other with a targeted mutation in the Shc binding site (TrkBSHC/SHC). Interestingly, KCC2 protein levels were not significantly affected in TrkBSHC/SHC mutant slices, indicating that the activation of Shc pathway is necessary for KCC2 down-regulation. Strikingly, KCC2 protein levels in the TrkBPLC/PLC mutant slices were elevated by the exposure to BDNF, indicating that in the absence of PLCγ cascades, the Shc pathway is able to upregulate KCC2 (IV; Fig. 8).
Similar results were obtained from experiments in which TrkB mutant slices were exposed to the 0-Mg2+ solution for 3 hr. In addition, immunostaining for pCREB demonstrated that CREB activation (Finkbeiner et al., 1997; Bibel and Barde, 2000; Minichiello et al., 2002) may be involved in the mechanism for KCC2 downregulation in 0-Mg2+ treated slices. Scavenging endogenous BDNF also showed suppressed CREB activation in slices exposed to the 0-Mg2+ solution. The robust increase in CREB phosphorylation observed in the CA1 region after 1 hr coincides with the down-regulation of KCC2 in the same region. These data support the involvement of CREB in regulating KCC2 transcription under spontaneous activity.
These results show that the expression of KCC2 is regulated by neuronal activity. Consequently, neuronal hyperactivity leads to compromised Cl- extrusion which is required for GABAergic postsynaptic hyperpolarization. This prediction was demonstrated experimentally by measuring the reversal potential of GABAA-mediated IPSPs with a somatic
Results and Discussion

43
chloride load. The driving forces of IPSPs between control slices and 0-Mg2+ treated slices were signifi cantly different. In treated slices depolarizing reversal potentials was measured as compared to control. Our results provide evidence for the fast qualitative change in GABAergic transmission from hyperpolarizing to depolarizing under epileptic-like conditions (Kapur et al., 1995; Nabekura et al., 2002; Payne et al., 2003; see also Kaila et al., 1997). Interestingly, a recent report suggests that a Ca2+-dependent decrease in K-Cl cotransport activity is associated with coincident presynaptic and postsynaptic spiking (Woodin et al., 2003). This effect may share the same mechanism as indicated in our studies.
Our results revealed a direct link between BDNF-TrkB signaling and downregulation of KCC2 and the underlying intracellular signaling
cascades. We found that both the PLCγ and Shc/FRS-2 activated signaling cascades are required for KCC2 down-regulation. Experiments from transgenic TrkBPLC/PLC and TrkBSHC/SHC mice suggested that the Shc pathway is crucial for both the down-regulation and up-regulation of KCC2. Down-regulation of KCC2 appears to take place if the Shc pathway is activated in conjunction with the PLCγ cascades, whereas an up-regulation is triggered by the Shc pathway acting in the absence of the PLCγ cascades. These may partly explain the different effects of BDNF on KCC2 mRNA expression in immature and mature neurons (Aguado et al., 2003; Rivera et al., 2002). We speculate that similar mechanisms for long-term plasticity are important under normal and pathophysiological conditions as well as in the maturation of functional neuronal networks.
Results and Discussion

44
Conclusions
1. The spatiotemporal expression patterns of distinct CCCs during embryonic development suggests that Cl- regulatory mechanisms are critically involved in the control of neuronal development.
2. Developmental expression of KCC2 in vivo and in vitro follows neuronal maturation and is strongly correlated with synaptogenesis, but neither neuronal spiking nor ionotropic glutamatergic and GABAergic transmission are required for the developmental up-regulation of KCC2 expression in mouse cultured cortical neurons.
3. KCC2-/- cortical neurons have abnormal morphology of dendritic protrusions and the number of functional excitatory synapses is reduced in KCC2-/- neurons. The interaction between KCC2 and dendritic cytoskeleton protein, 4.1N, is required for spine development.
4. The expression of KCC2 is regulated in diverse manifestations of neuronal plasticity. We show that interictal-like activity down-regulates KCC2 mRNA and protein expression in hippocampal slices, which leads to a reduced capacity for neuronal Cl- extrusion. This effect is mediated by endogenous BDNF acting on TrkB, with down-stream cascades involving both Shc/FRS-2 and PLCγ cAMP response element-binding protein signaling.
Conclusion

45
Acknowledgments
This study was carried out at the Institute of Biotechnology and Department of Environmental and Biological Sciences under the supervision of Dr. Claudio Rivera and Professor Kai Kaila during 2000-2007.
First of all, I would like to express my deepest thanks to Claudio who took me into his group. Claudio has endless kindness and patience in helping me deal with every detail of my experiments among these years. More importantly, he has given me the freedom and opportunities to think and test different aspects of the subject. Along my Ph.D study, his continued encouragement accelerated my enthusiasm in science. Without his help, I would have not been able to complete my Ph.D studies.
I want to thank Kai who gave me the opportunity to be his student and led me into the interesting fi eld of neuroscience. It is enjoyable to be his student because of his patience and scientifi c advices. Without his guidance and encouragement, I would have not been able to fi nish my Ph.D study.
I would also give my special thank to professor Mart Saarma, the Director of the Institute of Biotechnology, who also supervised my Ph.D study. Mart encouraged me continuously and provided valuable advices and suggestions to improve my thesis work. In addition, I am grateful to him for providing excellent facilities and resources for my research work.
I warmly thank Professor Dan Lindholm and Professor Kid Tornquist for reviewing my thesis and providing constructive criticisms to the thesis book.
Special thanks to Miika Palviainen and Marjo Heikura, for their technical supports, in particular for their pleasant help, all the fun in the lab and long-term friendship. Warmly thanks to Sergey for teaching me electrophysiology method. Thanks to Anastasia Ludwig, Anastasia Shulga, Hannele Lahtinen, Judith Thomas-Crusells, for relax work atmosphere and collaborations.
I am grateful to Docent Matti Airaksinen for collaborations and giving me valuable advices and suggestions in my thesis work. Thanks Janne Tornberg for pleasant collaboration and help.
I would like to thank Stanislav Khirug and Peter Blaesse for collaboration. All the members in the group have been a great help. Professor Juha Voipio, Eva Ruusuvuori, Junko Yamada.
Deeply thanks to Katri Wegelius for providing kind help with my study and living in Finland.
Many Thanks to Leonard Khiroug, Julia Kolikova, Ramil Afzalov, Kari Keinanen, Sarah. Coleman, Sari Lauri for pleasant collaborations.
Many thanks to the Neurogroup members, both former and present, have been a great help, Liying Yu, Jianmin Yang, Urmas Arumae, Congyun Zheng, Maili Jakobson, Maria Lindahl, Satu Akerber, Eila Kujamaki, Pavel Uvarov, Jari Rossi, Paivi Lindfors, Yunfu Sun, Xinqun Liang, Maxim Bespalov, Mikhail Paveliev, Paivi Lindholm, Pia Runeberg-Roos, Maria Lumen, Liina Lonka.
Outside of the lab, I would like to thank Li Tian, for her friendship and help during these years.
Finally, I would like to thank my husband, Chunlin, for his love, collaboration and support; and our son, Yanzhe, for bringing me endless happiness.
Hong Li, November 2007, Helsinki
Acknowledgements

46
Reference List
Adragna NC, Fulvio MD, Lauf PK (2004) Regulation of K-Cl cotransport: from function to genes. J Membr Biol 201: 109-137.
Aguado F, Carmona MA, Pozas E, Aguilo A, Martinez-Guijarro FJ, Alcantara S, Borrell V, Yuste R, Ibanez CF, Soriano E (2003) BDNF regulates spontaneous correlated activity at early developmental stages by increasing synaptogenesis and expression of the K+/Cl- co-transporter KCC2. Development 130: 1267-1280.
Alberts, B (1994) Molecular biology of the cell, 3rd edition. Garland.
Altman J & Bayer SA (1995) Atlas of Prenatal Rat Brain Development CRC Press, London, UK
Anderson WW, Lewis DV, Swartzwelder HS, Wilson WA (1986) Magnesium-free medium activates seizure-like events in the rat hippocampal slice. Brain Res 398: 215-219.
Aoki C, Miko I, Oviedo H, Mikeladze-Dvali T, Alexandre L, Sweeney N, Bredt DS (2001) Electron microscopic immunocytochemical detection of PSD-95, PSD-93, SAP-102, and SAP-97 at postsynaptic, presynaptic, and nonsynaptic sites of adult and neonatal rat visual cortex. Synapse 40: 239-257.
Bamji SX (2005) Cadherins: actin with the cytoskeleton to form synapses. Neuron 47: 175-178.
Barnard EA, Skolnick P, Olsen RW, Mohler H, Sieghart W, Biggio G, Braestrup C, Bateson AN, Langer SZ (1998) International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classifi cation on the basis of subunit structure and receptor function. Pharmacol Rev 50: 291-313.
Baumann SW, Baur R, Sigel E (2001) Subunit arrangement of gamma-aminobutyric acid type A receptors. J Biol Chem 276: 36275-36280.
Becker CM, Hoch W, Betz H (1988) Glycine receptor heterogeneity in rat spinal cord during postnatal development. EMBO J 7: 3717-3726.
Ben Ari Y (2001) Developing networks play a similar melody. Trends Neurosci 24: 353-360.
Ben Ari Y (2002) Excitatory actions of gaba during development: the nature of the nurture. Nat Rev Neurosci 3: 728-739.
Ben Ari Y, Cherubini E, Corradetti R, Gaiarsa JL (1989) Giant synaptic potentials in immature rat CA3 hippocampal neurones. J Physiol 416: 303-325.
Beretta F, Sala C, Saglietti L, Hirling H, Sheng M, Passafaro M (2005) NSF interaction is important for direct insertion of GluR2 at synaptic sites. Mol Cell Neurosci 28: 650-660.
Berridge MJ (1998) Neuronal calcium signaling. Neuron 21: 13-26.
Betz H, Laube B (2006) Glycine receptors: recent insights into their structural organization and functional diversity. J Neurochem 97: 1600-1610.
Betz H, Kuhse J, Schmieden V, Malosio ML, Langosch D, Prior P, Schmitt B, Kirsch J (1991) How to build a glycinergic postsynaptic membrane. J Cell Sci Suppl 15: 23-25.
Bibel M, Barde YA (2000) Neurotrophins: key regulators of cell fate and cell shape in the vertebrate nervous system. Genes Dev 14: 2919-2937.
Binder DK, Croll SD, Gall CM, Scharfman HE (2001) BDNF and epilepsy: too much of a good thing? Trends Neurosci 24: 47-53.
Blaesse P, Guillemin I, Schindler J, Schweizer M, Delpire E, Khiroug L, Friauf E, Nothwang HG 2006Oligomerization of KCC2 correlates with development of inhibitory neurotransmission. J Neurosci 26(41):10407-19
Bliss TV., Collingridge GL. (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 361(6407):31-9
Boettger T, Rust MB, Maier H, Seidenbecher T, Schweizer M, Keating DJ, Faulhaber J, Ehmke H, Pfeffer C, Scheel O, Lemcke B, Horst J, Leuwer R, Pape HC, Volkl H, Hubner CA, Jentsch TJ (2003) Loss of K-Cl co-transporter KCC3 causes deafness, neurodegeneration and reduced seizure threshold. EMBO J 22: 5422-5434.
Reference List

47
Boettger T, Hubner CA, Maier H, Rust MB, Beck FX, Jentsch TJ (2002) Deafness and renal tubular acidosis in mice lacking the K-Cl co-transporter Kcc4. Nature 416: 874-878.
Bolea S, Avignone E, Berretta N, Sanchez-Andres JV, Cherubini E (1999) Glutamate controls the induction of GABA-mediated giant depolarizing potentials through AMPA receptors in neonatal rat hippocampal slices. J Neurophysiol 81: 2095-2102.
Bormann J (2000) The 'ABC' of GABA receptors. Trends Pharmacol Sci 21: 16-19.
Borodinsky LN, Root CM, Cronin JA, Sann SB, Gu X, Spitzer NC (2004) Activity-dependent homeostatic specification of transmitter expression in embryonic neurons. Nature 429: 523-530.
Brickley SG, Cull-Candy SG, Farrant M (1996) Development of a tonic form of synaptic inhibition in rat cerebellar granule cells resulting from persistent activation of GABAA receptors. J Physiol 497 ( Pt 3): 753-759.
Bureau M, Olsen RW (1990) Multiple distinct subunits of the gamma-aminobutyric acid-A receptor protein show different ligand-binding affi nities. Mol Pharmacol 37: 497-502.
Cai C, Li H, Rivera C, Keinanen K (2006) Interaction between SAP97 and PSD-95, two Maguk proteins involved in synaptic traffi cking of AMPA receptors. J Biol Chem 281: 4267-4273.
Cai C, Coleman SK, Niemi K, Keinanen K (2002) Selective binding of synapse-associated protein 97 to GluR-A alpha-amino-5-hydroxy-3-methyl-4-isoxazole propionate receptor subunit is determined by a novel sequence motif. J Biol Chem 277: 31484-31490.
Caron L, Rousseau F, Gagnon E, Isenring P (2000) Cloning and funct ional characterization of a cation-Cl- cotransporter-interacting protein. J Biol Chem 275: 32027-32036.
Casula S, Shmukler BE, Wilhelm S, Stuart-Tilley AK, Su W, Chernova MN, Brugnara C, Alper SL (2001) A dominant negative mutant of the KCC1 K-Cl cotransporter:
both N- and C-terminal cytoplasmic domains are required for K-Cl cotransport activity. J Biol Chem 276: 41870-41878.
Chen G, Trombley PQ, van den Pol AN (1996) Excitatory actions of GABA in developing rat hypothalamic neurones. J Physiol 494 ( Pt 2): 451-464.
Chen L, Chetkovich DM, Petralia RS, Sweeney NT, Kawasaki Y, Wenthold RJ, Bredt DS, Nicoll RA (2000) Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 408: 936-943.
Cherubini E, Gaiarsa JL, Ben Ari Y (1991) GABA: an excitatory transmitter in early postnatal life. Trends Neurosci 14: 515-519.
Chetkovich DM, Bunn RC, Kuo SH, Kawasaki Y, Kohwi M, Bredt DS (2002) Postsynaptic targeting of alternative postsynaptic density-95 isoforms by distinct mechanisms. J Neurosci 22: 6415-6425.
Chih B, Engelman H, Scheiffele P (2005) Control of excitatory and inhibitory synapse formation by neuroligins. Science 307: 1324-1328.
Cho KO, Hunt CA, Kennedy MB (1992) The rat brain postsynaptic density fraction contains a homolog of the Drosophila discs-large tumor suppressor protein. Neuron 9: 929-942.
Clayton GH, Owens GC, Wolff JS, Smith RL (1998) Ontogeny of cation-Cl- cotransporter expression in rat neocortex. Brain Res Dev Brain Res 109: 281-292.
Cline H (2005) Synaptogenesis: a balancing act between excitation and inhibition. Curr Biol 15: R203-R205.
Coleman SK, Cai C, Mottershead DG, Haapalahti JP, Keinanen K (2003) Surface expression of GluR-D AMPA receptor is dependent on an interaction between its C-terminal domain and a 4.1 protein. J Neurosci 23: 798-806.
Colman DR (1997) Neurites, synapses, and cadherins reconciled. Mol Cell Neurosci 10: 1-6.
Connolly CN, Krishek BJ, McDonald BJ, Smart TG, Moss SJ (1996) Assembly and cell surface expression of heteromeric and homomeric gamma-aminobutyric acid type A receptors. J Biol Chem 271: 89-96.
Reference List

48
Constantine-Paton M, Cline HT (1998) LTP and activity-dependent synaptogenesis: the more alike they are, the more different they become. Curr Opin Neurobiol 8: 139-148.
Coull JAM, Boudreau D, Bachand K, Prescott SA, Nault F, Sik A, De Koninck P, De Koninck Y (2003) Trans-synaptic shift in anion gradient in spinal lamina I neurons as a mechanism of neuropathic pain. Nature 424: 938-942.
Crair MC, Malenka RC (1995) A critical period for long-term potentiation at thalamocortical synapses. Nature 375: 325-328.
Craven SE, Bredt DS (1998) PDZ proteins organize synaptic signaling pathways. Cell 93: 495-498.
Dailey ME, Smith SJ (1996) The dynamics of dendritic structure in developing hippocampal slices. J Neurosci 16: 2983-2994.
Dammerman RS, Flint AC, Noctor S, Kriegstein AR (2000) An excitatory GABAergic plexus in developing neocortical layer 1. J Neurophysiol 84: 428-434.
Darman RB, Flemmer A, Forbush B (2001) Modulation of ion transport by direct targeting of protein phosphatase type 1 to the Na-K-Cl cotransporter. J Biol Chem 276: 34359-34362.
Davies P, Anderton B, Kirsch J, Konnerth A, Nitsch R, Sheetz M (1998) First one in, last one out: the role of gabaergic transmission in generation and degeneration. Prog Neurobiol 55: 651-658.
DeFazio RA, Keros S, Quick MW, Hablitz JJ (2000) Potassium-coupled chloride cotransport controls intracellular chloride in rat neocortical pyramidal neurons. J Neurosci 20: 8069-8076.
Delpire E, Mount DB (2002) Human and murine phenotypes associated with defects in cation-chloride cotransport. Annu Rev Physiol 64: 803-843.
Delpire E, Lu J, England R, Dull C, Thorne T (1999) Deafness and imbalance associated with inactivation of the secretory Na-K-2Cl co-transporter. Nat Genet 22: 192-195.
Demarque M, Represa A, Becq H, Khalilov I, Ben Ari Y, Aniksztejn L (2002) Paracrine intercellular communication by a Ca2+-
and SNARE-independent release of GABA and glutamate prior to synapse formation. Neuron 36: 1051-1061.
Dev KK, Nishimune A, Henley JM, Nakanishi S (1999) The protein kinase C alpha binding protein PICK1 interacts with short but not long form alternative splice variants of AMPA receptor subunits. Neuropharmacology 38: 635-644.
Dingledine R, Borges K, Bowie D, Traynelis SF (1999) The glutamate receptor ion channels. Pharmacol Rev 51: 7-61.
Dong H, Zhang P, Liao D, Huganir RL (1999) Characterization, expression, and distribution of GRIP protein. Ann N Y Acad Sci 868: 535-540.
Dong H, O'Brien RJ, Fung ET, Lanahan AA, Worley PF, Huganir RL (1997) GRIP: a synaptic PDZ domain-containing protein that interacts with AMPA receptors. Nature 386: 279-284.
Durand GM, Kovalchuk Y, Konnerth A (1996) Long-term potentiation and functional synapse induction in developing hippocampus. Nature 381: 71-75.
Dzhala VI, Talos DM, Sdrulla DA, Brumback AC, Mathews GC, Benke TA, Delpire E, Jensen FE, Staley KJ (2005) NKCC1 transporter facilitates seizures in the developing brain. Nat Med 11: 1205-1213.
El Husseini AE, Schnell E, Chetkovich DM, Nicoll RA, Bredt DS (2000) PSD-95 involvement in maturation of excitatory synapses. Science 290: 1364-1368.
Esteban JA (2003) AMPA receptor traffi cking: a road map for synaptic plasticity. Mol Interv 3: 375-385.
Ethell IM, Pasquale EB (2005) Molecular mechanisms of dendritic spine development and remodeling. Prog Neurobiol 75: 161-205.
Farrant M., Kaila K., (2007) The cellular, molecular and ionic basis of GABA(A) receptor signaling. Prog Brain Res.;160:59-87.
Farrar SJ, Whiting PJ, Bonnert TP, McKernan RM (1999) Stoichiometry of a ligand-gated ion channel determined by fluorescence energy transfer. J Biol Chem 274: 10100-10104.
Reference List

49
Feigenspan A, Bormann J (1998) GABA-gated Cl- channels in the rat retina. Prog Retin Eye Res 17: 99-126.
Feller MB (1999) Spontaneous correlated activity in developing neural circuits. Neuron 22: 653-656.
Feng W, Long JF, Fan JS, Suetake T, Zhang M (2004) The tetrameric L27 domain complex as an organization platform for supramolecular assemblies. Nat Struct Mol Biol 11: 475-480.
Flagella M, Clarke LL, Miller ML, Erway LC, Giannella RA, Andringa A, Gawenis LR, Kramer J, Duffy JJ, Doetschman T, Lorenz JN, Yamoah EN, Cardell EL, Shull GE (1999) Mice lacking the basolateral Na-K-2Cl cotransporter have impaired epithelial chloride secretion and are profoundly deaf. J Biol Chem 274: 26946-26955.
Finkbeiner S, Tavazoie SF, Maloratsky A, Jacobs KM, Harris KM, Greenberg ME (1997) CREB: a major mediator of neuronal neurotrophin responses. Neuron 19: 1031-1047
Flint AC, Liu X, Kriegstein AR (1998) Nonsynaptic glycine receptor activation during early neocortical development. Neuron 20: 43-53.
Fritschy JM, Brunig I (2003) Formation and plasticity of GABAergic synapses: p h y s i o l o g i c a l m e c h a n i s m s a n d pathophysiological implications. Pharmacol Ther 98: 299-323.
Fritschy JM, Mohler H (1995) GABAA-receptor heterogeneity in the adult rat brain: differential regional and cellular distribution of seven major subunits. J Comp Neurol 359: 154-194.
Fujita A, Kurachi Y (2000) SAP family proteins. Biochem Biophys Res Commun 269: 1-6.
Gagnon KB, England R, Delpire E (2006) Volume sensitivity of cation-Cl- cotransporters is modulated by the interaction of two kinases: Ste20-related proline-alanine-rich kinase and WNK4. Am J Physiol Cell Physiol 290: C134-C142.
Gamba G, Miyanoshita A, Lombardi M, Lytton J, Lee WS, Hediger MA, Hebert SC (1994) Molecular cloning, primary structure,
and characterization of two members of the mammalian electroneutral sodium-(potassium)-chloride cotransporter family expressed in kidney. J Biol Chem 269: 17713-17722.
Ganguly K, Schinder AF, Wong ST, Poo M (2001) GABA itself promotes the developmental switch of neuronal GABAergic responses from excitation to inhibition. Cell 105: 521-532.
Gao FB, Raff M (1997) Cell size control and a cell-intrinsic maturation program in proliferating oligodendrocyte precursor cells. J Cell Biol 138: 1367-1377.
Garaschuk O, Hanse E, Konnerth A (1998) Developmental profi le and synaptic origin of early network oscillations in the CA1 region of rat neonatal hippocampus. J Physiol 507 ( Pt 1): 219-236.
Gardner SM, Takamiya K, Xia J, Suh JG, Johnson R, Yu S, Huganir RL (2005) Calcium-permeable AMPA receptor plasticity is mediated by subunit-specific interactions with PICK1 and NSF. Neuron 45: 903-915.
Gimenez I, Isenring P, Forbush B (2002) Spatially distributed alternative splice variants of the renal Na-K-Cl cotransporter exhibit dramatically different affi nities for the transported ions. J Biol Chem 277: 8767-8770.
Goda Y (2002) Cadherins communicate structural plasticity of presynaptic and postsynaptic terminals. Neuron 35: 1-3.
Govek EE, Newey SE, Akerman CJ, Cross JR, Van d, V, Van Aelst L (2004) The X-linked mental retardation protein oligophrenin-1 is required for dendritic spine morphogenesis. Nat Neurosci 7: 364-372.
Graf ER, Zhang X, Jin SX, Linhoff MW, Craig AM (2004) Neurexins induce differentiation of GABA and glutamate postsynaptic specializations via neuroligins. Cell 119: 1013-1026.
Grichtchenko II, Choi I, Zhong X, Bray-Ward P, Russell JM, Boron WF (2001) Cloning, characterization, and chromosomal mapping of a human electroneutral Na(+)-driven Cl-HCO3 exchanger. J Biol Chem 276: 8358-8363.
Reference List

50
Groc L, Gustafsson B, Hanse E (2006) AMPA signalling in nascent glutamatergic synapses: there and not there! Trends Neurosci 29: 132-139.
Gulyas AI, Sik A, Payne JA, Kaila K, Freund TF (2001) The KCl cotransporter, KCC2, is highly expressed in the vicinity of excitatory synapses in the rat hippocampus. Eur J Neurosci 13: 2205-2217.
Halpain S (2000) Actin and the agile spine: how and why do dendritic spines dance? Trends Neurosci 23: 141-146.
Harvey RJ, Schmieden V, Von Holst A, Laube B, Rohrer H, Betz H (2000) Glycine receptors containing the alpha4 subunit in the embryonic sympathetic nervous system, spinal cord and male genital ridge. Eur J Neurosci 12: 994-1001.
Hering H, Sheng M (2001) Dendritic spines: structure, dynamics and regulation. Nat Rev Neurosci 2: 880-888.
Hiki K, D'Andrea RJ, Furze J, Crawford J, Woollatt E, Sutherland GR, Vadas MA, Gamble JR (1999) Cloning, characterization, and chromosomal location of a novel human K+-Cl- cotransporter. J Biol Chem 274: 10661-10667.
Hollmann M, Heinemann S (1994) Cloned glutamate receptors. Annu Rev Neurosci 17: 31-108.
Hollrigel GS, Ross ST, Soltesz I (1998) Temporal patterns and depolarizing actions of spontaneous GABAA receptor activation in granule cells of the early postnatal dentate gyrus. J Neurophysiol 80: 2340-2351.
Hoover KB, Bryant PJ (2000) The genetics of the protein 4.1 family: organizers of the membrane and cytoskeleton. Curr Opin Cell Biol 12: 229-234.
Howard HC, Mount DB, Rochefort D, Byun N, Dupre N, Lu J, Fan X, Song L, Riviere JB, Prevost C, Horst J, Simonati A, Lemcke B, Welch R, England R, Zhan FQ, Mercado A, Siesser WB, George AL, Jr., McDonald MP, Bouchard JP, Mathieu J, Delpire E, Rouleau GA (2002) The K-Cl cotransporter KCC3 is mutant in a severe peripheral neuropathy associated with agenesis of the corpus callosum. Nat Genet 32: 384-392.
Huang K, El Husseini A (2005) Modulation of neuronal protein traffi cking and function by palmitoylation. Curr Opin Neurobiol 15: 527-535.
Huang Y, Man HY, Sekine-Aizawa Y, Han Y, Juluri K, Luo H, Cheah J, Lowenstein C, Huganir RL, Snyder SH (2005) S-nitrosylation of N-ethylmaleimide sensitive factor mediates surface expression of AMPA receptors. Neuron 46: 533-540.
Hubner CA, Lorke DE, Hermans-Borgmeyer I (2001a) Expression of the Na-K-2Cl-cotransporter NKCC1 during mouse development. Mech Dev 102: 267-269.
Hubner CA, Stein V, Hermans-Borgmeyer I, Meyer T, Ballanyi K, Jentsch TJ (2001b) Disruption of KCC2 reveals an essential role of K-Cl cotransport already in early synaptic inhibition. Neuron 30: 515-524.
Hussain NK, Sheng M (2005) Neuroscience. Making synapses: a balancing act. Science 307: 1207-1208.
Inoue K, Ueno S, Fukuda A (2004) Interaction of neuron-specific K+-Cl- cotransporter, KCC2, with brain-type creatine kinase. FEBS Lett 564: 131-135.
Irie F, Yamaguchi Y (2002) EphB receptors regulate dendritic spine development via intersectin, Cdc42 and N-WASP. Nat Neurosci 5: 1117-1118.
Isaac JT, Crair MC, Nicoll RA, Malenka RC (1997) Silent synapses during development of thalamocortical inputs. Neuron 18: 269-280.
Jacob TC, Bogdanov YD, Magnus C, Saliba RS, Kittler JT, Haydon PG, Moss SJ (2005) Gephyrin regulates the cell surface dynamics of synaptic GABAA receptors. J Neurosci 25: 10469-10478.
Ji Y, Pang PT, Feng L, Lu B (2005) Cyclic AMP controls BDNF-induced TrkB phosphorylation and dendritic spine formation in mature hippocampal neurons. Nat Neurosci 8: 164-172.
Johnson RC, Penzes P, Eipper BA, Mains RE (2000) Isoforms of kalirin, a neuronal Dbl family member, generated through use of different 5'- and 3'-ends along with an internal translational initiation site. J Biol Chem 275: 19324-19333.
Reference List

51
Johnston GA (1996) GABAc receptors: relatively simple transmitter -gated ion channels? Trends Pharmacol Sci 17: 319-323.
Kahle KT, Wilson FH, Lifton RP (2005) Regulation of diverse ion transport pathways by WNK4 kinase: a novel molecular switch. Trends Endocrinol Metab 16: 98-103.
Kaila K., Ransom B. (eds) (1998) pH and Brain Function. Wiley-Liss, New York.
Kaila K, Lamsa K, Smirnov S, Taira T, Voipio J (1997) Long-lasting GABA-mediated depolarization evoked by high-frequency stimulation in pyramidal neurons of rat hippocampal slice is attributable to a network-driven, bicarbonate-dependent K+ transient. J Neurosci 17: 7662-7672.
Kaila K (1994) Ionic basis of GABAA receptor channel function in the nervous system. Prog Neurobiol 42: 489-537.
Kaila K, Voipio J (1987) Postsynaptic fall in intracellular pH induced by GABA-activated bicarbonate conductance. Nature 330: 163-165.
Kakazu Y, Akaike N, Komiyama S, Nabekura J (1999) Regulation of intracellular chloride by cotransporters in developing lateral superior olive neurons. J Neurosci 19: 2843-2851.
Kanaka C, Ohno K, Okabe A, Kuriyama K, Itoh T, Fukuda A, Sato K (2001) The differential expression patterns of messenger RNAs encoding K-Cl cotransporters (KCC1,2) and Na-K-2Cl cotransporter (NKCC1) in the rat nervous system. Neuroscience 104: 933-946.
Kaneda M, Farrant M, Cull-Candy SG (1995) Whole-cell and single-channel currents activated by GABA and glycine in granule cells of the rat cerebellum. J Physiol 485 ( Pt 2): 419-435.
Kandel ER, Shwartz JH, Jessell TM (2000) Principles of Neural Science 4th ed, McGraw-Hill Medical.
Kapur J, Pillai A, Henry TR (1995) Psychogenic elaboration of simple partial seizures. Epilepsia 36: 1126-1130.
Karadsheh MF, Byun N, Mount DB, Delpire E (2004) Localization of the KCC4 potassium-
chloride cotransporter in the nervous system. Neuroscience 123: 381-391.
Karadsheh MF, Delpire E (2001) Neuronal restrictive silencing element is found in the KCC2 gene: molecular basis for KCC2-specific expression in neurons. J Neurophysiol 85: 995-997.
Karnak D, Lee S, Margolis B (2002) Identifi cation of multiple binding partners for the amino-terminal domain of synapse-associated protein 97. J Biol Chem 277: 46730-46735.
Kasyanov AM, Safiulina VF, Voronin LL, Cherubini E (2004) GABA-mediated giant depolarizing potentials as coincidence detectors for enhancing synaptic efficacy in the developing hippocampus. Proc Natl Acad Sci U S A 101: 3967-3972.
Katz LC, Crowley JC (2002) Development of cortical circuits: lessons from ocular dominance columns. Nat Rev Neurosci 3: 34-42.
Kaufmann WA, Humpel C, Alheid GF, Marksteiner J (2003) Compartmentation of alpha 1 and alpha 2 GABA(A) receptor subunits within rat extended amygdala: implications for benzodiazepine action. Brain Res 964: 91-99.
Kelsch W, Hormuzdi S, Straube E, Lewen A, Monyer H, Misgeld U (2001) Insulin-like growth factor 1 and a cytosolic tyrosine kinase activate chloride outward transport during maturation of hippocampal neurons. J Neurosci 21: 8339-8347.
Khalilov I, Dzhala V, Ben Ari Y, Khazipov R (1999) Dual role of GABA in the neonatal rat hippocampus. Dev Neurosci 21: 310-319.
Khazipov R, Esclapez M, Caillard O, Bernard C, Khalilov I, Tyzio R, Hirsch J, Dzhala V, Berger B, Ben Ari Y (2001) Early development of neuronal activity in the primate hippocampus in utero. J Neurosci 21: 9770-9781.
Khazipov R, Leinekugel X, Khalilov I, Gaiarsa JL, Ben Ari Y (1997) Synchronization of GABAergic interneuronal network in CA3 subfi eld of neonatal rat hippocampal slices. J Physiol 498 ( Pt 3): 763-772.
Reference List

52
Khirug S, Huttu K, Ludwig A, Smirnov S, Voipio J, Rivera C, Kaila K, Khiroug L (2005) Distinct properties of functional KCC2 expression in immature mouse hippocampal neurons in culture and in acute slices. Eur J Neurosci. 21(4):899-904
Kim E, Sheng M (2004) PDZ domain proteins of synapses. Nat Rev Neurosci 5: 771-781.
Klausberger T, Sarto I, Ehya N, Fuchs K, Furtmuller R, Mayer B, Huck S, Sieghart W (2001) Alternate use of distinct intersubunit contacts controls GABAA receptor assembly and stoichiometry. J Neurosci 21: 9124-9133.
Kneussel M, Betz H (2000) Receptors, gephyrin and gephyrin-associated proteins: novel insights into the assembly of inhibitory postsynaptic membrane specializations. J Physiol 525 Pt 1: 1-9.
Knight AR, Stephenson FA, Tallman JF, Ramabahdran TV (2000) Monospecific antibodies as probes for the stoichiometry of recombinant GABA(A) receptors. Receptors Channels 7: 213-226.
Kornau HC, Seeburg PH (1997) Partner selection by PDZ domains. Nat Biotechnol 15: 319.
Kullmann DM, Asztely F, Walker MC., (2000) The role of mammalian ionotropic receptors in synaptic plasticity: LTP, LTD and epilepsy. Cell Mol Life Sci. 57(11):1551-61.
Kullmann DM, Ruiz A, Rusakov DM, Scott R, Semyanov A, Walker MC (2005) Presynaptic, extrasynaptic and axonal GABAA receptors in the CNS: where and why? Prog Biophys Mol Biol 87: 33-46.
Lamsa K, Palva JM, Ruusuvuori E, Kaila K, Taira T (2000) Synaptic GABA(A) activation inhibits AMPA-kainate receptor-mediated bursting in the newborn (P0-P2) rat hippocampus. J Neurophysiol 83: 359-366.
Langosch D, Hoch W, Betz H (1992) The 93 kDa protein gephyrin and tubulin associated with the inhibitory glycine receptor are phosphorylated by an endogenous protein kinase. FEBS Lett 298: 113-117.
Larramendi EM, Victor T (1967) Synapses on the Purkinje cell spines in the mouse. An
electronmicroscopic study. Brain Res 5: 15-30.
Lauf PK, Adragna NC (2000) K-Cl cotransport: properties and molecular mechanism. Cell Physiol Biochem 10: 341-354.
Le Rouzic P, Ivanov TR, Stanley PJ, Baudoin FM, Chan F, Pinteaux E, Brown PD, Luckman SM (2006) KCC3 and KCC4 expression in rat adult forebrain. Brain Res 1110: 39-45.
Lee H, Chen CX, Liu YJ, Aizenman E, Kandler K (2005) KCC2 expression in immature rat cortical neurons is suffi cient to switch the polarity of GABA responses. Eur J Neurosci 21: 2593-2599.
Lee S, Fan S, Makarova O, Straight S, Margolis B (2002) A novel and conserved protein-protein interaction domain of mammalian Lin-2/CASK binds and recruits SAP97 to the lateral surface of epithelia. Mol Cell Biol 22: 1778-1791.
Lee SY, Maniak PJ, Rhodes R, Ingbar DH, O’Grady SM. (2003) Basolateral Cl- transport is stimulated by terbutaline in adult rat alveolar epithelial cells. J Membr Biol. 191(2):133-9.
Leinekugel X, Medina I, Khalilov I, Ben Ari Y, Khazipov R (1997) Ca2+ oscillations mediated by the synergistic excitatory actions of GABA(A) and NMDA receptors in the neonatal hippocampus. Neuron 18: 243-255.
Leinekugel X, Tseeb V, Ben Ari Y, Bregestovski P (1995) Synaptic GABAA activation induces Ca2+ rise in pyramidal cells and interneurons from rat neonatal hippocampal slices. J Physiol 487 ( Pt 2): 319-329.
Leonard AS, Davare MA, Horne MC, Garner CC, Hell JW (1998) SAP97 is associated with the alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor GluR1 subunit. J Biol Chem 273: 19518-19524.
Levinson JN, Chery N, Huang K, Wong TP, Gerrow K, Kang R, Prange O, Wang YT, El Husseini A (2005) Neuroligins mediate excitatory and inhibitory synapse formation: involvement of PSD-95 and neurexin-1beta
Reference List

53
in neuroligin-induced synaptic specifi city. J Biol Chem 280: 17312-17319.
Li RW, Yu W, Christie S, Miralles CP, Bai J, Loturco JJ, De Blas AL (2005) Disruption of postsynaptic GABA receptor clusters leads to decreased GABAergic innervation of pyramidal neurons. J Neurochem 95: 756-770.
Liao D, Malinow R (1996) Deficiency in induction but not expression of LTP in hippocampal slices from young rats. Learn Mem 3: 138-149.
Liao D, Zhang X, O’Brien R, Ehlers MD, Huganir RL (1999) Regulation of morphological postsynaptic silent synapses in developing hippocampal neurons. Nat Neurosci 2: 37-43.
Lim IA, Merrill MA, Chen Y, Hell JW (2003) Disruption of the NMDA receptor-PSD-95 interaction in hippocampal neurons with no obvious physiological short-term effect. Neuropharmacology 45: 738-754.
Lise MF, El Husseini A (2006) The neuroligin and neurexin families: from structure to function at the synapse. Cell Mol Life Sci 63: 1833-1849.
LoTurco JJ, Owens DF, Heath MJ, Davis MB, Kriegstein AR (1995) GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron 15: 1287-1298.
Lu J, Karadsheh M, Delpire E (1999) Developmental regulation of the neuronal-specifi c isoform of K-Cl cotransporter KCC2 in postnatal rat brains. J Neurobiol 39: 558-568.
Lynch JW (2004) Molecular structure and function of the glycine receptor chloride channel. Physiol Rev 84: 1051-1095.
Lytle C, McManus T (2002) Coordinate modulation of Na-K-2Cl cotransport and K-Cl cotransport by cell volume and chloride. Am J Physiol Cell Physiol 283: C1422-C1431.
Lytle C (1998) A volume-sensitive protein kinase regulates the Na-K-2Cl cotransporter in duck red blood cells. Am J Physiol 274: C1002-C1010.
Ma XM, Johnson RC, Mains RE, Eipper BA (2001) Expression of kalirin, a neuronal
GDP/GTP exchange factor of the trio family, in the central nervous system of the adult rat. J Comp Neurol 429: 388-402.
MacVicar BA, Feighan D, Brown A, Ransom B (2002) Intrinsic optical signals in the rat optic nerve: role for K(+) uptake via NKCC1 and swelling of astrocytes. Glia 37: 114-123.
Maletic-Savatic M, Malinow R, Svoboda K (1999) Rapid dendritic morphogenesis in CA1 hippocampal dendrites induced by synaptic activity. Science 283: 1923-1927.
Malinow R and Malenka RC (2002) AMPA receptor traffi cking and synaptic plasticity. Annu Rev Neurosci. 25:103-26.
Malosio ML, Grenningloh G, Kuhse J, Schmieden V, Schmitt B, Prior P, Betz H (1991) Alternative splicing generates two variants of the alpha 1 subunit of the inhibitory glycine receptor. J Biol Chem 266: 2048-2053.
Mantyh PW, Hunt SP (2004) Setting the tone: superfi cial dorsal horn projection neurons regulate pain sensitivity. Trends Neurosci 27: 582-584.
Marrs GS, Green SH, Dailey ME (2001) Rapid formation and remodeling of postsynaptic densities in developing dendrites. Nat Neurosci 4: 1006-1013.
Marty S, Wehrle R, Alvarez-Leefmans FJ, Gasnier B, Sotelo C (2002) Postnatal maturation of Na+, K+, 2Cl- cotransporter expression and inhibitory synaptogenesis in the rat hippocampus: an immunocytochemical analysis. Eur J Neurosci 15: 233-245.
Marty S, Berninger B, Carroll P, Thoenen H (1996) GABAergic stimulation regulates the phenotype of hippocampal interneurons through the regulation of brain-derived neurotrophic factor. Neuron 16: 565-570.
Matskevich I, Hegney KL, Flatman PW (2005) Regulation of erythrocyte Na-K-2Cl cotransport by threonine phosphorylation. Biochim Biophys Acta 1714: 25-34.
Matus A (2000) Actin-based plasticity in dendritic spines. Science 290: 754-758.
Mauceri D, Cattabeni F, Di Luca M, Gardoni F (2004) Calcium/calmodulin-dependent protein kinase II phosphorylation drives synapse-associated protein 97 into spines. J Biol Chem 279: 23813-23821.
Reference List

54
McLean HA, Caillard O, Khazipov R, Ben Ari Y, Gaiarsa JL (1996) Spontaneous release of GABA activates GABAB receptors and controls network activity in the neonatal rat hippocampus. J Neurophysiol 76: 1036-1046.
Mehta AK, Ticku MK (1999) An update on GABAA receptors. Brain Res Brain Res Rev 29: 196-217.
Menendez dlP, Bolea S, Sanchez-Andres JV (1998) Origin of the synchronized network activity in the rabbit developing hippocampus. Eur J Neurosci 10: 899-906.
Menezes, J.R. & Luskin, M.B. (1994) Expression of neuron-specifi c tubulin defi nes a novel population in the proliferative layers of the developing telencephalon. J. Neurosci., 4, 5399-5416.
Menezes, J.R., Smith, C.M., Nelson, K.C. & Luskin, M.B. (1995) The division of neuronal progenitor cells during migration in the neonatal mammalian forebrain. Mol. Cell. Neurosci., 6, 496-508.
Mercado A, Broumand V, Zandi-Nejad K, Enck AH, Mount DB (2006) A C-terminal domain in KCC2 confers constitutive K+-Cl- cotransport. J Biol Chem 281: 1016-1026.
Mercado A, Vazquez N, Song L, Cortes R, Enck AH, Welch R, Delpire E, Gamba G, Mount DB (2005) NH2-terminal heterogeneity in the KCC3 K+-Cl- cotransporter. Am J Physiol Renal Physiol 289: F1246-F1261.
Mercado A, Mount DB, Gamba G (2004) Electroneutral cation-chloride cotransporters in the central nervous system. Neurochem Res 29: 17-25.
Meyer G, Kirsch J, Betz H, Langosch D (1995) Identifi cation of a gephyrin binding motif on the glycine receptor beta subunit. Neuron 15: 563-572.
Mikawa S, Wang C, Shu F, Wang T, Fukuda A, Sato K (2002) Developmental changes in KCC1, KCC2 and NKCC1 mRNAs in the rat cerebellum. Brain Res Dev Brain Res 136: 93-100.
Miller M, Peters A (1981) Maturation of rat visual cortex. II. A combined Golgi-electron microscope study of pyramidal neurons. J Comp Neurol 203: 555-573.
Minichiello L, Calella AM, Medina DL, Bonhoeffer T, Klein R, Korte M (2002) Mechanism of TrkB-mediated hippocampal long-term potentiation. Neuron 36: 121-137.
Misgeld U, Deisz RA, Dodt HU, Lux HD (1986) The role of chloride transport in postsynaptic inhibition of hippocampal neurons. Science 232: 1413-1415.
Mody I, Lambert JD, Heinemann U (1987) Low extracellular magnesium induces epileptiform activity and spreading depression in rat hippocampal slices. J Neurophysiol 57: 869-888.
Mohler H, Luscher B, Fritschy JM, Benke D, Benson J, Rudolph U (1998) GABA(A)-receptor assembly in vivo: lessons from subunit mutant mice. Life Sci 62: 1611-1615.
Mok H, Shin H, Kim S, Lee JR, Yoon J, Kim E (2002) Association of the kinesin superfamily motor protein KIF1Balpha with postsynaptic density-95 (PSD-95), synapse-associated protein-97, and synaptic scaffolding molecule PSD-95/discs large/zona occludens-1 proteins. J Neurosci 22: 5253-5258.
Mount DB, Mercado A, Song L, Xu J, George AL, Jr., Delpire E, Gamba G (1999) Cloning and characterization of KCC3 and KCC4, new members of the cation-chloride cotransporter gene family. J Biol Chem 274: 16355-16362.
Murphy DD, Cole NB, Segal M (1998) Brain-derived neurotrophic factor mediates estradiol-induced dendritic spine formation in hippocampal neurons. Proc Natl Acad Sci U S A 95: 11412-11417.
Nabekura J, Ueno T, Okabe A, Furuta A, Iwaki T, Shimizu-Okabe C, Fukuda A, Akaike N (2002) Reduction of KCC2 expression and GABAA receptor-mediated excitation after in vivo axonal injury. J Neurosci 22: 4412-4417.
Naisbitt S, Kim E, Tu JC, Xiao B, Sala C, Valtschanoff J, Weinberg RJ, Worley PF, Sheng M (1999) Shank, a novel family of postsynaptic density proteins that binds to the NMDA receptor/PSD-95/GKAP complex and cortactin. Neuron 23: 569-582.
Reference List

55
Nakayama AY, Harms MB, Luo L (2000) Small GTPases Rac and Rho in the maintenance of dendritic spines and branches in hippocampal pyramidal neurons. J Neurosci 20: 5329-5338.
Nicoll RA, Malenka RC (1999) Expression mechanisms underlying NMDA receptor-dependent long-term potentiation. Ann N Y Acad Sci 868: 515-525.
Nishimune A, Isaac JT, Molnar E, Noel J, Nash SR, Tagaya M, Collingridge GL, Nakanishi S, Henley JM (1998) NSF binding to GluR2 regulates synaptic transmission. Neuron 21: 87-97.
Nusser Z, Mody I (2002) Selective modulation of tonic and phasic inhibitions in dentate gyrus granule cells. J Neurophysiol 87: 2624-2628.
O’Brien RJ, Lau LF, Huganir RL (1998) Molecular mechanisms of glutamate receptor clustering at excitatory synapses. Curr Opin Neurobiol 8: 364-369.
Obrietan K, van den Pol AN (1995) GABA neurotransmission in the hypothalamus: developmental reversal from Ca2+ elevating to depressing. J Neurosci 15: 5065-5077.
Okabe A, Yokokura M, Toyoda H, Shimizu-Okabe C, Ohno K, Sato K, Fukuda A (2003) Changes in chloride homeostasis-regulating gene expressions in the rat hippocampus following amygdala kindling. Brain Res 990: 221-226.
Okabe S, Miwa A, Okado H (2001) Spine formation and correlated assembly of presynaptic and postsynaptic molecules. J Neurosci 21: 6105-6114.
Owens DF, Kriegstein AR (2002) Is there more to GABA than synaptic inhibition? Nat Rev Neurosci 3: 715-727.
Owens DF, Boyce LH, Davis MB, Kriegstein AR (1996) Excitatory GABA responses in embryonic and neonatal cortical slices demonstrated by gramicidin perforated-patch recordings and calcium imaging. J Neurosci 16: 6414-6423.
Owens DF, Kriegstein AR (1998) Patterns of intracellular calcium fl uctuation in precursor cells of the neocortical ventricular zone. J Neurosci. 18: 5374-88.
Paarmann I, Spangenberg O, Lavie A, Konrad M (2002) Formation of complexes between Ca2+.calmodulin and the synapse-associated protein SAP97 requires the SH3 domain-guanylate kinase domain-connecting HOOK region. J Biol Chem 277: 40832-40838.
Pace AJ, Lee E, Athirakul K, Coffman TM, O’Brien DA, Koller BH (2000) Failure of spermatogenesis in mouse lines defi cient in the Na(+)-K(+)-2Cl(-) cotransporter. J Clin Invest 105: 441-450.
Palva JM, Lamsa K, Lauri SE, Rauvala H, Kaila K, Taira T (2000) Fast network oscillations in the newborn rat hippocampus in vitro. J Neurosci 20: 1170-1178.
Park E, Na M, Choi J, Kim S, Lee JR, Yoon J, Park D, Sheng M, Kim E (2003) The Shank family of postsynaptic density proteins interacts with and promotes synaptic accumulation of the beta PIX guanine nucleotide exchange factor for Rac1 and Cdc42. J Biol Chem 278: 19220-19229.
Payne JA, Rivera C, Voipio J, Kaila K (2003) Cation-chloride co-transporters in neuronal communication, development and trauma. Trends Neurosci 26: 199-206.
Payne JA (1997) Functional characterization of the neuronal-specifi c K-Cl cotransporter: implications for [K+]o regulation. Am J Physiol 273: C1516-C1525.
Payne JA, Stevenson TJ, Donaldson LF (1996) Molecular characterization of a putative K-Cl cotransporter in rat brain. A neuronal-specifi c isoform. J Biol Chem 271: 16245-16252.
Pearson MM, Lu J, Mount DB, Delpire E (2001) Localization of the K(+)-Cl(-) cotransporter, KCC3, in the central and peripheral nervous systems: expression in the choroid plexus, large neurons and white matter tracts. Neuroscience 103: 481-491.
Penn AA, Shatz CJ (1999) Brain waves and brain wiring: the role of endogenous and sensory-driven neural activity in development. Pediatr Res 45: 447-458.
Penn AA, Riquelme PA, Feller MB, Shatz CJ (1998) Competition in retinogeniculate patterning driven by spontaneous activity. Science 279: 2108-2112.
Reference List

56
Penzes P, Johnson RC, Alam MR, Kambampati V, Mains RE, Eipper BA (2000) An isoform of kalirin, a brain-specific GDP/GTP exchange factor, is enriched in the postsynaptic density fraction. J Biol Chem 275: 6395-6403.
Petralia RS, Esteban JA, Wang YX, Partridge JG, Zhao HM, Wenthold RJ, Malinow R (1999) Selective acquisition of AMPA receptors over postnatal development suggests a molecular basis for silent synapses. Nat Neurosci 2: 31-36.
Petralia RS, Wenthold RJ (1992) Light and electron immunocytochemical localization of AMPA-selective glutamate receptors in the rat brain. J Comp Neurol 318: 329-354.
Piechotta K, Lu J, Delpire E (2002) Cation chloride cotransporters interact with the stress-related kinases Ste20-related proline-alanine-rich kinase (SPAK) and oxidative stress response 1 (OSR1). J Biol Chem 277: 50812-50819.
Plata C, Meade P, Vazquez N, Hebert SC, Gamba G (2002) Functional properties of the apical Na+-K+-2Cl- cotransporter isoforms. J Biol Chem 277: 11004-11012.
Plata C, Meade P, Hall A, Welch RC, Vazquez N, Hebert SC, Gamba G (2001) Alternatively spliced isoform of apical Na(+)-K(+)-Cl(-) cotransporter gene encodes a furosemide-sensitive Na(+)-Cl(-)cotransporter. Am J Physiol Renal Physiol 280: F574-F582.
Polenzani L, Woodward RM, Miledi R (1991) Expression of mammalian gamma-aminobutyric acid receptors with distinct pharmacology in Xenopus oocytes. Proc Natl Acad Sci U S A 88: 4318-4322.
Prange O, Wong TP, Gerrow K, Wang YT, El Husseini A (2004) A balance between excitatory and inhibitory synapses is controlled by PSD-95 and neuroligin. Proc Natl Acad Sci U S A 101: 13915-13920.
Prior P, Schmitt B, Grenningloh G, Pribilla I, Multhaup G, Beyreuther K, Maulet Y, Werner P, Langosch D, Kirsch J, . (1992) Primary structure and alternative splice variants of gephyrin, a putative glycine receptor-tubulin linker protein. Neuron 8: 1161-1170.
Qualmann B, Boeckers TM, Jeromin M, Gundelfinger ED, Kessels MM (2004) Linkage of the actin cytoskeleton to the postsynaptic density via direct interactions of Abp1 with the ProSAP/Shank family. J Neurosci 24: 2481-2495.
Race JE, Makhlouf FN, Logue PJ, Wilson FH, Dunham PB, Holtzman EJ (1999) Molecular cloning and functional characterization of KCC3, a new K-Cl cotransporter. Am J Physiol 277: C1210-C1219.
Rao A, Craig AM (2000) Signaling between the actin cytoskeleton and the postsynaptic density of dendritic spines. Hippocampus 10: 527-541.
Rao VR and Finkbeiner S, (2007) NMDA and AMPA receptors: old channels, new tricks. Trends Neurosci. 6:284-91.
Rivera C, Li H, Thomas-Crusells J, Lahtinen H, Viitanen T, Nanobashvili A, Kokaia Z, Airaksinen MS, Voipio J, Kaila K, Saarma M (2002) BDNF-induced TrkB activation down-regulates the K+-Cl- cotransporter KCC2 and impairs neuronal Cl- extrusion. Journal of Cell Biology 159: 747-752.
Rivera C, Voipio J, Payne JA, Ruusuvuori E, Lahtinen H, Lamsa K, Pirvola U, Saarma M, Kaila K (1999) The K+/Cl- co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature 397: 251-255.
Rohrbough J, Spitzer NC (1996) Regulation of intracellular Cl- levels by Na(+)-dependent Cl- cotransport distinguishes depolarizing from hyperpolarizing GABAA receptor-mediated responses in spinal neurons. J Neurosci 16: 82-91.
Romero MF, Hediger MA, Boulpaep EL, Boron WF (1997) Expression cloning and characterization of a renal electrogenic Na+/HCO3- cotransporter. Nature 387: 409-413.
Rubio ME, Wenthold RJ (1999) Differential distribution of intracellular glutamate receptors in dendrites. J Neurosci 19: 5549-5562.
Russell JM (2000) Sodium-potassium-chloride cotransport. Physiol Rev 80: 211-276.
Sala C, Piech V, Wilson NR, Passafaro M, Liu G, Sheng M (2001) Regulation of dendritic
Reference List

57
spine morphology and synaptic function by Shank and Homer. Neuron 31: 115-130.
Salinas PC, Price SR (2005) Cadherins and catenins in synapse development. Curr Opin Neurobiol 15: 73-80.
Sangan P, Brill SR, Sangan S, Forbush B 3rd, Binder HJ (2000) Basolateral K-Cl cotransporter regulates colonic potassium absorption in potassium depletion.J Biol Chem. 275(40):30813-6.
Sato K, Kiyama H, Tohyama M (1992) Regional distribution of cells expressing glycine receptor alpha 2 subunit mRNA in the rat brain. Brain Res 590: 95-108.
Schmidt A, Hall A (2002) Guanine nucleotide exchange factors for Rho GTPases: turning on the switch. Genes Dev 16: 1587-1609.
Schomberg SL, Bauer J, Kintner DB, Su G, Flemmer A, Forbush B, Sun DD (2003) Cross talk between the GABA(A) receptor and the Na-K-Cl cotransporter is mediated by intracellular Cl-. Journal of Neurophysiology 89: 159-167.
Scott EK, Reuter JE, Luo L (2003) Small GTPase Cdc42 is required for multiple aspects of dendritic morphogenesis. J Neurosci 23: 3118-3123.
Segal M, Andersen P (2000) Dendritic spines shaped by synaptic activity. Curr Opin Neurobiol 10: 582-586.
Semyanov A, Walker MC, Kullmann DM, Silver RA (2004) Tonically active GABA A receptors: modulating gain and maintaining the tone. Trends Neurosci 27: 262-269.
Semyanov A, Walker MC, Kullmann DM (2003) GABA uptake regulates cortical excitability via cell type-specific tonic inhibition. Nat Neurosci 6: 484-490.
Serafini R, Valeyev AY, Barker JL, Poulter MO (1995) Depolarizing GABA-activated Cl- channels in embryonic rat spinal and olfactory bulb cells. J Physiol 488 ( Pt 2): 371-386.
Shen L, Liang F, Walensky LD, Huganir RL (2000) Regulation of AMPA receptor GluR1 subunit surface expression by a 4. 1N-linked actin cytoskeletal association. J Neurosci 20: 7932-7940.
Sheng M (2001) Molecular organization of the postsynaptic specialization. Proc Natl Acad Sci U S A 98: 7058-7061.
Sheng M, Sala C (2001) PDZ domains and the organization of supramolecular complexes. Annu Rev Neurosci 24: 1-29.
Shi SH, Hayashi Y, Petralia RS, Zaman SH, Wenthold RJ, Svoboda K, Malinow R (1999) Rapid spine delivery and redistribution of AMPA receptors after synaptic NMDA receptor activation. Science 284: 1811-1816.
Shillingford JM, Miyoshi K, Flagella M, Shull GE, Hennighausen L (2002) Mouse mammary epithelial cells express the Na-K-Cl cotransporter, NKCC1: characterization, localization, and involvement in ductal development and morphogenesis. Mol Endocrinol 16: 1309-1321.
Sieghart W (1995) Structure and pharmacology of gamma-aminobutyric acidA receptor subtypes. Pharmacol Rev 47: 181-234.
Sieghart W (2000) Unraveling the function of GABA(A) receptor subtypes. Trends Pharmacol Sci 21: 411-413.
Sipilä ST, Kaila K (2007) GABAergic Control of CA3-driven network events in the developing hippocampus. In: Inhibitory Regulation of Excitatory Neurotransmission, Ed. Mark Darlison, Springer, Heidelberg (in press)
Sipila ST, Schuchmann S, Voipio J, Yamada J, Kaila K. (2006b) The cation-chloride cotransporter NKCC1 promotes sharp waves in the neonatal rat hippocampus. J Physiol. 573:765-773.
Sipila ST, Huttu K, Voipio J, Kaila K (2006a) Intrinsic bursting of immature CA3 pyramidal neurons and consequent giant depolarizing potentials are driven by a persistent Na+ current and terminated by a slow Ca2+ -activated K+ current. Eur J Neurosci 23: 2330-2338.
Sipila ST, Huttu K, Soltesz I, Voipio J, Kaila K (2005) Depolarizing GABA acts on intrinsically bursting pyramidal neurons to drive giant depolarizing potentials in the immature hippocampus. J Neurosci 25: 5280-5289.
Reference List

58
Song I, Kamboj S, Xia J, Dong H, Liao D, Huganir RL (1998) Interaction of the N-ethylmaleimide-sensitive factor with AMPA receptors. Neuron 21: 393-400.
Song L, Mercado A, Vazquez N, Xie Q, Desai R, George AL, Jr., Gamba G, Mount DB (2002) Molecular, functional, and genomic characterization of human KCC2, the neuronal K-Cl cotransporter. Brain Res Mol Brain Res 103: 91-105.
Sotelo C (1978) Purkinje cell ontogeny: formation and maintenance of spines. Prog Brain Res 48: 149-170.
Sotelo C (1990) Cerebellar synaptogenesis: what we can learn from mutant mice. J Exp Biol. 153:225-49
Spitzer NC, Root CM, Borodinsky LN (2004) Orchestrating neuronal differentiation: patterns of Ca2+ spikes specify transmitter choice. Trends Neurosci 27: 415-421.
Steiger JL, Russek SJ (2004) GABAA receptors: building the bridge between subunit mRNAs, their promoters, and cognate transcription factors. Pharmacol Ther 101: 259-281.
Stell BM, Mody I (2002) Receptors with different affi nities mediate phasic and tonic GABA(A) conductances in hippocampal neurons. J Neurosci 22: RC223.
Strange K, Singer TD, Morrison R, Delpire E (2000) Dependence of KCC2 K-Cl cotransporter activity on a conserved carboxy terminus tyrosine residue. Am J Physiol Cell Physiol 279: C860-C867.
Strata F, Atzori M, Molnar M, Ugolini G, Tempia F, Cherubini E (1997) A pacemaker current in dye-coupled hilar interneurons contributes to the generation of giant GABAergic potentials in developing hippocampus. J Neurosci 17: 1435-1446.
Studer R, von Boehmer L, Haenggi T, Schweizer C, Benke D, Rudolph U, Fritschy JM (2006) Alteration of GABAergic synapses and gephyrin clusters in the thalamic reticular nucleus of GABA(A) receptor alpha3 subunit-null mice. Eur J Neurosci 24: 1307-1315.
Sung KW, Kirby M, McDonald MP, Lovinger DM, Delpire E (2000) Abnormal GABAA
receptor-mediated currents in dorsal root ganglion neurons isolated from Na-K-2Cl cotransporter null mice. J Neurosci 20: 7531-7538.
Sutor B, Luhmann HJ (1995) Development of excitatory and inhibitory postsynaptic potentials in the rat neocortex. Perspect Dev Neurobiol 2: 409-419.
Tashiro A, Minden A, Yuste R (2000) Regulation of dendritic spine morphology by the rho family of small GTPases: antagonistic roles of Rac and Rho. Cereb Cortex 10: 927-938.
Thomas U, Phannavong B, Muller B, Garner CC, Gundelfinger ED (1997) Functional expression of rat synapse-associated proteins SAP97 and SAP102 in Drosophila dlg-1 mutants: effects on tumor suppression and synaptic bouton structure. Mech Dev 62: 161-174.
Thompson SM, Gahwiler BH (1989) Activity-dependent disinhibition. I. Repetitive stimulation reduces IPSP driving force and conductance in the hippocampus in vitro. J Neurophysiol 61: 501-511.
Titz S, Hans M, Kelsch W, Lewen A, Swandulla D, Misgeld U (2003) Hyperpolarizing inhibition develops without trophic support by GABA in cultured rat midbrain neurons. J Physiol 550: 719-730.
Tomita S, Adesnik H, Sekiguchi M, Zhang W, Wada K, Howe JR, Nicoll RA, Bredt DS (2005) Stargazin modulates AMPA receptor gating and traffi cking by distinct domains. Nature 435: 1052-1058.
Tornberg J, Voikar V, Savilahti H, Rauvala H, Airaksinen MS (2005) Behavioural phenotypes of hypomorphic KCC2-defi cient mice. Eur J Neurosci 21: 1327-1337.
Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K (2002) Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature 420: 788-794.
Tu JC, Xiao B, Yuan JP, Lanahan AA, Leoffert K, Li M, Linden DJ, Worley PF (1998) Homer binds a novel proline-rich motif and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron 21: 717-726.
Reference List

59
Uvarov P, Pruunsild P, Timmusk T, Airaksinen MS (2005) Neuronal K+/Cl- co-transporter (KCC2) transgenes lacking neurone restrictive silencer element recapitulate CNS neurone-specific expression and developmental up-regulation of endogenous KCC2 gene. J Neurochem 95: 1144-1155.
Vandenberghe W, Nicoll RA, Bredt DS (2005) Stargazin is an AMPA receptor auxiliary subunit. Proc Natl Acad Sci U S A 102: 485-490.
Walensky LD, Blackshaw S, Liao D, Watkins CC, Weier HU, Parra M, Huganir RL, Conboy JG, Mohandas N, Snyder SH (1999) A novel neuron-enriched homolog of the erythrocyte membrane cytoskeletal protein 4.1. J Neurosci 19: 6457-6467.
Wall MJ, Usowicz MM (1997) Development of action potential-dependent and independent spontaneous GABAA receptor-mediated currents in granule cells of postnatal rat cerebellum. Eur J Neurosci 9: 533-548.
Walsh MJ, Kuruc N (1992) The postsynaptic density: constituent and associated proteins characterized by electrophoresis, immunoblotting, and peptide sequencing. J Neurochem 59: 667-678.
Wang C, Shimizu-Okabe C, Watanabe K, Okabe A, Matsuzaki H, Ogawa T, Mori N, Fukuda A, Sato K. (2002) Developmental changes in KCC1, KCC2, and NKCC1 mRNA expressions in the rat brain. Brain Res Dev Brain Res. 139(1):59-66
Williams JR, Sharp JW, Kumari VG, Wilson M, Payne JA (1999) The neuron-specific K-Cl cotransporter, KCC2. Antibody development and initial characterization of the protein. J Biol Chem 274: 12656-12664.
Wisden W, Seeburg PH (1992) GABAA receptor channels: from subunits to functional entities. Curr Opin Neurobiol 2: 263-269.
Woo NS, Lu J, England R, McClellan R, Dufour S, Mount DB, Deutch AY, Lovinger
DM, Delpire E (2002) Hyperexcitability and epilepsy associated with disruption of the mouse neuronal-specifi c K-Cl cotransporter gene. Hippocampus 12: 258-268.
Woodin MA, Ganguly K, Poo MM (2003) Coincident pre- and postsynaptic activity modif ies GABAergic synapses by postsynaptic changes in Cl- transporter activity. Neuron 39: 807-820.
Wu G, Malinow R, Cline HT (1996) Maturation of a central glutamatergic synapse. Science 274: 972-976.
Xia J, Zhang X, Staudinger J, Huganir RL (1999) Clustering of AMPA receptors by the synaptic PDZ domain-containing protein PICK1. Neuron 22: 179-187.
Xu JC, Lytle C, Zhu TT, Payne JA, Benz E Jr, Forbush B, III (1994) Molecular cloning and functional expression of the bumetanide-sensitive Na-K-Cl cotransporter. Proc Natl Acad Sci U S A 91: 2201-2205.
Yamada Y, Fukuda A, Tanaka M, Shimano Y, Nishino H, Muramatsu K, Togari H, Wada Y (2001) Optical imaging reveals cation--Cl(-) cotransporter-mediated transient rapid decrease in intracellular Cl(-) concentration induced by oxygen--glucose deprivation in rat neocortical slices. Neurosci Res 39: 269-280.
Yuste R, Bonhoeffer T (2004) Genesis of dendritic spines: insights from ultrastructural and imaging studies. Nat Rev Neurosci 5: 24-34.
Zhang H, Webb DJ, Asmussen H, Horwitz AF (2003) Synapse formation is regulated by the signaling adaptor GIT1. J Cell Biol 161: 131-142.
Zhang SJ, Jackson MB (1995) GABAA receptor activation and the excitability of nerve terminals in the rat posterior pituitary. J Physiol 483 ( Pt 3): 583-595.
Ziv NE, Smith SJ (1996) Evidence for a role of dendritic fi lopodia in synaptogenesis and spine formation. Neuron 17: 91-102.
Reference List























![KCC2-dependent Steady-state Intracellular Chloride ...€¦ · To gain insight into the factors affecting neuronal intracellular steady-state Cl- concentration, we compared [Cl-]](https://img.pdfslide.us/doc/110x75/5eaed7f64037310648663520/kcc2-dependent-steady-state-intracellular-chloride-to-gain-insight-into-the.jpg)





