Embed Size (px)
Citation preview

This article was downloaded by: [University of Birmingham]On: 03 October 2014, At: 02:37Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Caryologia: International Journal of Cytology,Cytosystematics and CytogeneticsPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tcar20
Structural and cytochemical analysis of femalereproductive development in Ornithogalumsigmoideum Freyn & SintIsmailoglu Isil a & Meral Unal aa Department of Biology , Marmara University, Faculty of Science and Letter ,Goztepe , 34722 , Istanbul , TurkeyPublished online: 10 Feb 2014.
To cite this article: Ismailoglu Isil & Meral Unal (2011) Structural and cytochemical analysis of female reproductivedevelopment in Ornithogalum sigmoideum Freyn & Sint, Caryologia: International Journal of Cytology, Cytosystematicsand Cytogenetics, 64:1, 99-109, DOI: 10.1080/00087114.2011.10589769
To link to this article: http://dx.doi.org/10.1080/00087114.2011.10589769
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitability for anypurpose of the Content. Any opinions and views expressed in this publication are the opinions and viewsof the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sources of information. Taylorand Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses,damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connectionwith, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

CARYOLOGIA
*Corresponding author: Phone: +090 216 348 77 59-1177; fax: +090 216 347 87 83; e-mail: [email protected]
Vol. 64, no. 1: 99-109, 2011
Structural and cytochemical analysis of female reproductive devel-opment in Ornithogalum sigmoideum Freyn & Sint
Ismailoglu* Isil and Meral Unal
Marmara University, Faculty of Science and Letter, Department of Biology, Goztepe 34722 Istanbul-Turkey
Abstract — In the present study, after the determination of karyological characteristics of Ornithogalum sig-moideum, the development of female gametophyte, embryo and endosperm was investigated, and cytochemical features were identifi ed.The diploid chromosome number is 16 in O. sigmoideum. The ovule is anatropous, bitegmic and crassinucellate. The ovular structures such as obturator and nucellar epidermis secrete insoluble polysaccharides, and play a role as the guidance and growth of the pollen tubes. The tissue differentiation called hypostase is located at the chalazal part of the ovule. This tissue is stained deeply with PAS, and it stores a large quantity of starch grains.The embryo sac development is Polygonum type. In the mature embryo sac, the egg cell and synergids are sur-rounded by a complete cell wall. The synergids show a similar polarization to the egg cell, and their cell walls form a prominent fi liform apparatus at the base of cells. Cytoplasm of the egg cell is not rich in insoluble and acidic polysaccharides, proteins, and lipids whereas cy-toplasm of the synergids and antipodals are rich in insoluble polysaccharides, proteins but is not rich acidic polysaccharides and lipids. The starch grains are peripherally located in the central cell. The embryo develop-ment is Asterad type. The endosperm is helobial.
Key words: Cytochemistry, embryo, endosperm, female gametophyte, karyotype, Ornithogalum sigmoideum.
INTRODUCTION
There are 25 species of Ornithogalum genus, which contain bulb and perennial grasses plants in Turkey. Chromosome numbers in this genus differentiate among and within species. In the study conducted by CZAPIK (1965) over 19 dif-ferent populations in O. umbellatum, and 7 dif-ferent populations in O. gussonei, he noted that the chromosome number of O. umbellatum was 2n: 18, 19, 27, 28, 36, while the chromosome number of O. gussonei was 2n: 18, 19, 20, 20+f, 27, 28, 29. It was revealed that the diploid chro-mosome number of O. sigmoideum genus was 2n: 12, 16, 17, 19, 20 (DAVIS 1984).
The differences in the number of the chromo-some numbers are effective in the evaluation of
the taxonomic position of a plant. Besides, em-bryological studies contribute to the taxonomic evaluation in such cases when the evaluation is diffi cult to make. Since there might be differ-ences in chromosomes between various popula-tions, the chromosome samples collected from the same population were analysed, and it was made certain that our embriological outcomes were not effected by this difference.
Embryological studies on several species of Ornithogalum L. have been conducted by SCHNARFF (1928), DESOLE (1947), PIENAAR (1963), CZAPIK (1972), ZABINSKA (1972), TILTON (1980, 1981a, b) and TILTON and LERSTEN (1981a, b). Detailed light and electron microscope stud-ies on the development of the ovule of O. cauda-tum were carried out by TILTON (1981a, b) and TILTON and LERSTEN (1981a, b).
The hypostase in O. caudatum starts to differ-entiate during megasporogenesis and starch ac-cumulates in the cell cytoplasms (TILTON 1980a). There is a nucellar epidermis forming a nucellar cap, which has the ability to attract the pollen
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4

ISMAILOGLU and UNAL100
tubes by producing secretion near the micropyle of this structure (TILTON 1980a, b; TILTON and LERSTEN 1981a). The ovule of O. caudatum has a secretory obturator originating from the base of the funiculus (TILTON and HORNER 1980).
Aposporic embryo sac development was ob-served in some species of the genus Ornithogalum. ZABINSKA (1972) detected an embryo sac with unre-duced number of chromosomes in O. gussonei, at-tributing it to apospory (apomixis), a type of sexual reproduction. Furthermore, in a study on different species of Ornithogalum, CZAPIK (1972) observed a twin embryo sac. DESOLE (1947) did not mention apomictic reproduction in O. umbellatum.
All cells in the embryo sac of O. caudatum (TILTON 1981b) and Scilla sibirica (BHANDARI and SACHDEVA 1983) are structurally so organ-ized that the egg cell is covered by a complete cell wall. In O. umbellatum, the antipodals and synergids degenerate just after fertilization (CZAPIK 1966, 1972; TILTON and LERSTEN 1981b; VAN RENSBURG and ROBBERTSE 1988), while they live longer in O. gussonei (ZABINSKA 1972). In O. umbelatum, DESOLE (1947) demonstrated the presence of fi liform apparatus in synergids; whereas ZABINSKA (1972) did not record it in the synergids of O. gussonei. In O. nutans and O. na-num species, polyploidy was noted in the antipo-dal cells (KARAGOZAVA and VAN KHANKH 1972).
The development of endosperm is helobial type and embryo development conforms to the Asterad type in the species of Ornithogalum (CZAPIK 1966, 1972; ZABINSKA 1972; VAN RENS-BURG and ROBBERTSE 1988).
This study aims to explore the structural and cytochemical features of the embryo sac, embryo and endosperm development in O. sigmoideum.
MATERIALS AND METHODS
The karyological analysis of O. sigmoideum was performed on the mitotic metaphases. The root tips were fi xed in acetic-alcohol (1:3, v/v) after pretreatment with bromonaphthalene and squash preparations were made in %2 aceto-or-sein. Chromosome measurements including long arm, short arm, total length of chromosome set, arm ratio index and relative chromosome length were made from 5 enlarged well-spread met-aphase cells of each population. Chromosomes were classifi ed according to the nomenclature of LEVAN et al (1964). The homologous chromo-somes were paired by their similarity in size and shape. The best 5 metaphase plates were used to
construct the respective ideogram according to the relative chromosome length.
Flower buds were collected at different de-velopmental stages in Basibuyuk region (Istan-bul, Turkey). The material at different stages was fi xed in acetic-alcohol (1:3, v/v) at 4°C for 24 hours, and then dehydrated in an ascending series of ethanol. Thereafter the material was embedded in paraffi n. The 4-12 µm thick micro-tome sections (Leica RM 2125 RT) of 4-12 were stained with Regaud’s hematoxylin.
For cytochemical observations, fl ower buds were fi xed in 2% glutaraldehyde in phosphate buffer (pH 8.0) for 1 h at room temperature, and then in 3% fresh glutaraldehyde (same buffer) at 4°C overnight. Following dehydration in an ascending series of ethanol, the material was embedded in epon. Semi-thin sections (1-2 µm) were cut with glass knives and used to localize different cytochemical components.
Total insoluble polysaccharides were identi-fi ed with periodic acid-Schiff (PAS) reagent; pec-tinaceous material and acidic polysaccharides with 1% Alcian blue in 3% acetic acid; proteins with Coomassie brilliant blue in a mixture of water, methanol and acetic acid (v:v:v, 87:10:3); and lipoidal material with Sudan black B in 70% ethanol (HESLOP-HARRISON and SHIVANNA 1977; O’BRIEN et al. 1964; FEDER and O’BRIEN 1968; HESLOP-HARRISON 1979, HESLOP-HARRISON et al. 1973; PEARSE 1961).
The preparations were analyzed with Im-age-Pro Express Version 6.0 scientifi c image processing and analysis software, photographed by using an Evolution LC color camera and an Olympus BH-2 microscope.
RESULTS
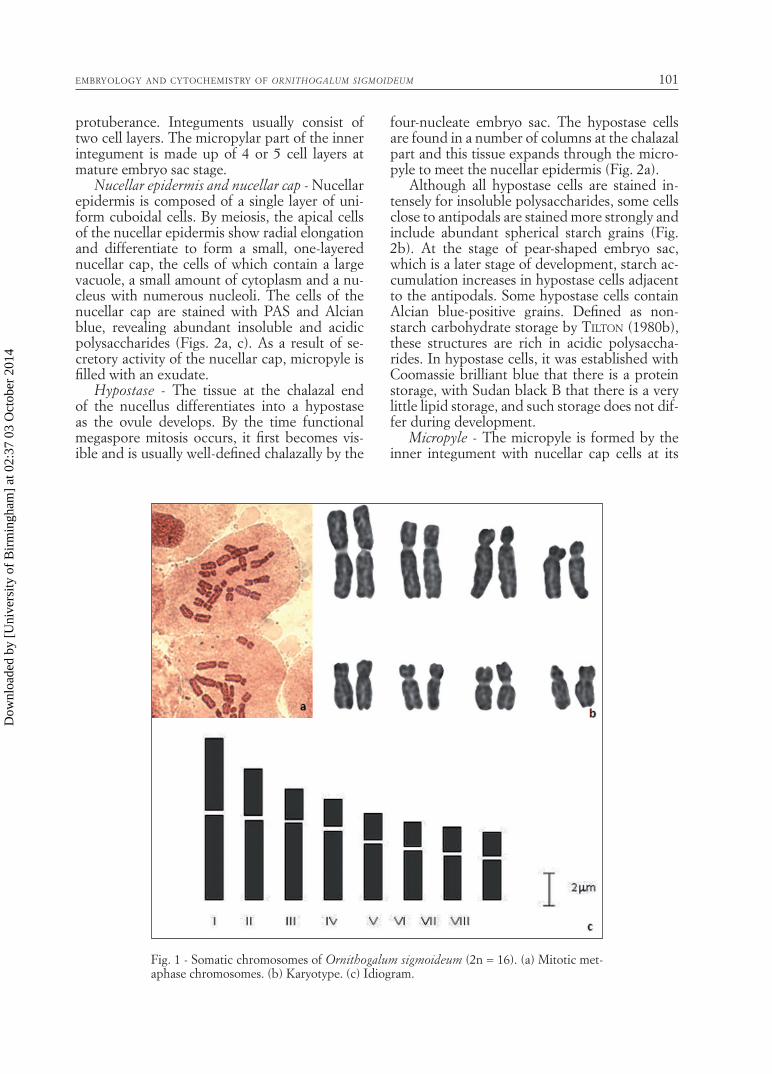
The karyotype - To avoid a high level of karyo-logical differentiation in different populations of O. sigmoideum, all the materials were collected from the same area and karyotype analysis was carried out. The diploid chromosome number is 16, consisting of a pair of metasentric and 7 pairs of submetasentric chromosomes (Fig. 1.a). The chromosome lengths vary between 3.9805 and 9.8107 µm. The chromosomes have no satellites. The results of the karyological analysis are pre-sented in Table 1 and Fig. 1.b, c.
Ovular structures - In O. sigmoideum, the mature ovule is anatropous, bitegmic and crassi-nucellate. The inner integument initiates fi rst and then outer integuments develop into a small
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4
EMBRYOLOGY AND CYTOCHEMISTRY OF ORNITHOGALUM SIGMOIDEUM 101
protuberance. Integuments usually consist of two cell layers. The micropylar part of the inner integument is made up of 4 or 5 cell layers at mature embryo sac stage.
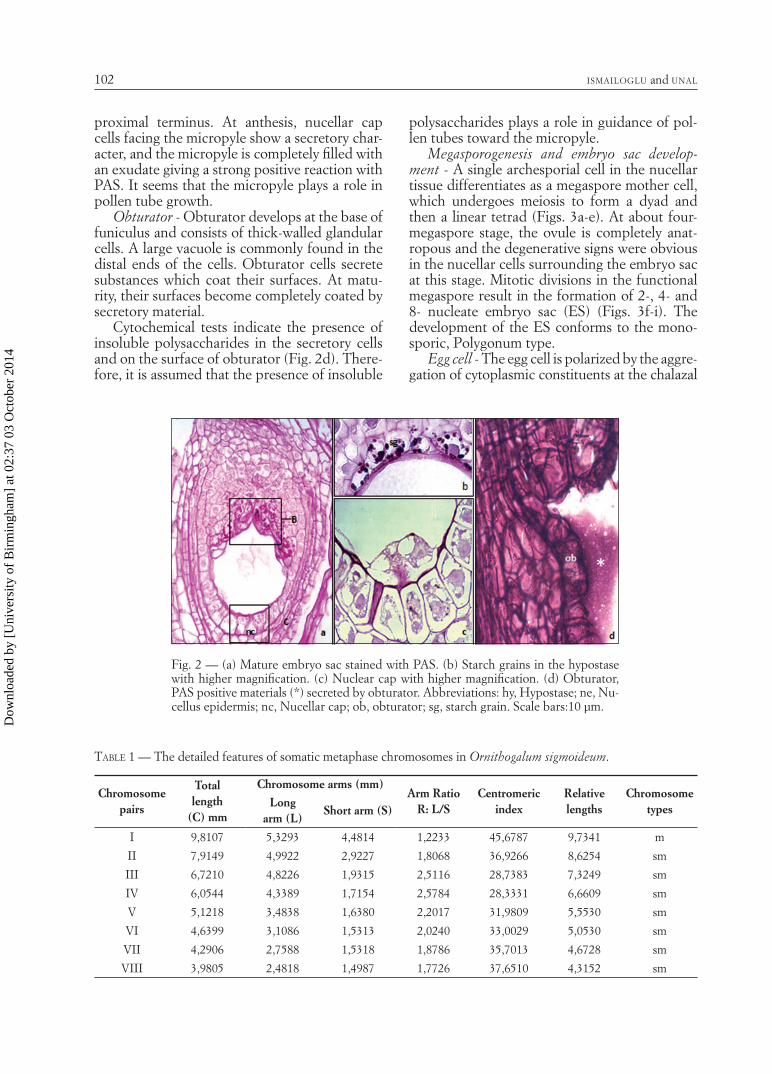
Nucellar epidermis and nucellar cap - Nucellar epidermis is composed of a single layer of uni-form cuboidal cells. By meiosis, the apical cells of the nucellar epidermis show radial elongation and differentiate to form a small, one-layered nucellar cap, the cells of which contain a large vacuole, a small amount of cytoplasm and a nu-cleus with numerous nucleoli. The cells of the nucellar cap are stained with PAS and Alcian blue, revealing abundant insoluble and acidic polysaccharides (Figs. 2a, c). As a result of se-cretory activity of the nucellar cap, micropyle is fi lled with an exudate.
Hypostase - The tissue at the chalazal end of the nucellus differentiates into a hypostase as the ovule develops. By the time functional megaspore mitosis occurs, it fi rst becomes vis-ible and is usually well-defi ned chalazally by the
four-nucleate embryo sac. The hypostase cells are found in a number of columns at the chalazal part and this tissue expands through the micro-pyle to meet the nucellar epidermis (Fig. 2a).
Although all hypostase cells are stained in-tensely for insoluble polysaccharides, some cells close to antipodals are stained more strongly and include abundant spherical starch grains (Fig. 2b). At the stage of pear-shaped embryo sac, which is a later stage of development, starch ac-cumulation increases in hypostase cells adjacent to the antipodals. Some hypostase cells contain Alcian blue-positive grains. Defi ned as non-starch carbohydrate storage by TILTON (1980b), these structures are rich in acidic polysaccha-rides. In hypostase cells, it was established with Coomassie brilliant blue that there is a protein storage, with Sudan black B that there is a very little lipid storage, and such storage does not dif-fer during development.
Micropyle - The micropyle is formed by the inner integument with nucellar cap cells at its
Fig. 1 - Somatic chromosomes of Ornithogalum sigmoideum (2n = 16). (a) Mitotic met-aphase chromosomes. (b) Karyotype. (c) Idiogram.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4
ISMAILOGLU and UNAL102
proximal terminus. At anthesis, nucellar cap cells facing the micropyle show a secretory char-acter, and the micropyle is completely fi lled with an exudate giving a strong positive reaction with PAS. It seems that the micropyle plays a role in pollen tube growth.
Obturator - Obturator develops at the base of funiculus and consists of thick-walled glandular cells. A large vacuole is commonly found in the distal ends of the cells. Obturator cells secrete substances which coat their surfaces. At matu-rity, their surfaces become completely coated by secretory material.
Cytochemical tests indicate the presence of insoluble polysaccharides in the secretory cells and on the surface of obturator (Fig. 2d). There-fore, it is assumed that the presence of insoluble
polysaccharides plays a role in guidance of pol-len tubes toward the micropyle.
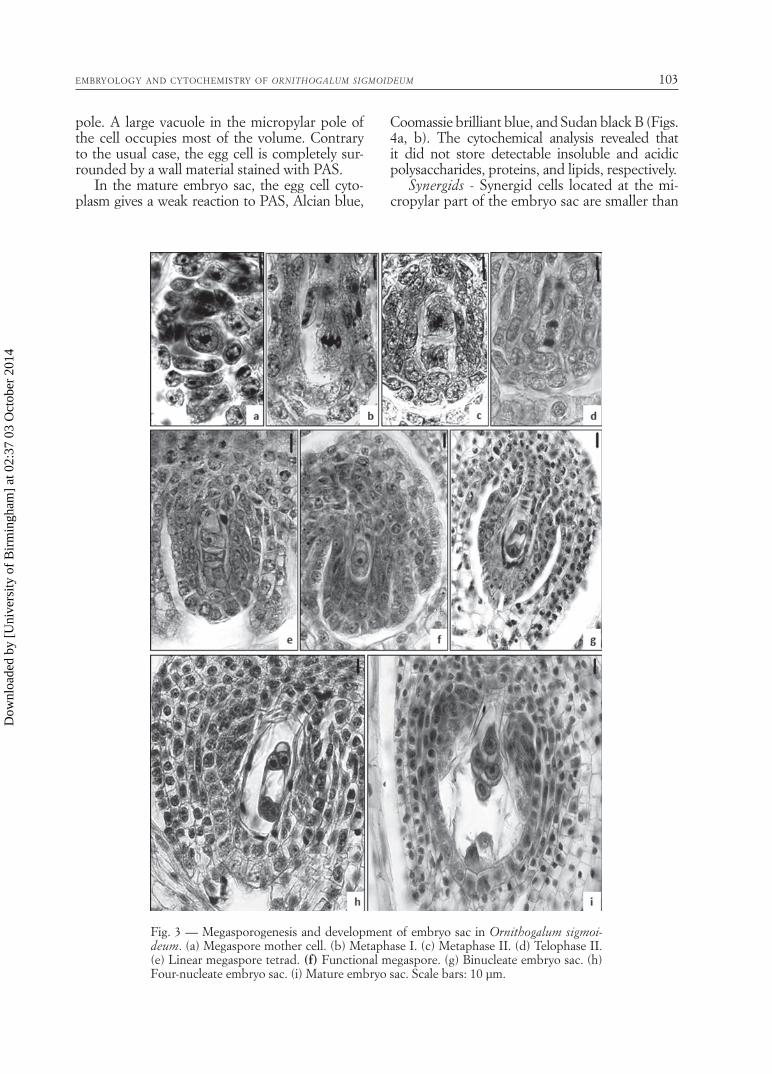
Megasporogenesis and embryo sac develop-ment - A single archesporial cell in the nucellar tissue differentiates as a megaspore mother cell, which undergoes meiosis to form a dyad and then a linear tetrad (Figs. 3a-e). At about four-megaspore stage, the ovule is completely anat-ropous and the degenerative signs were obvious in the nucellar cells surrounding the embryo sac at this stage. Mitotic divisions in the functional megaspore result in the formation of 2-, 4- and 8- nucleate embryo sac (ES) (Figs. 3f-i). The development of the ES conforms to the mono-sporic, Polygonum type.
Egg cell - The egg cell is polarized by the aggre-gation of cytoplasmic constituents at the chalazal
TABLE 1 — The detailed features of somatic metaphase chromosomes in Ornithogalum sigmoideum.
Chromosome pairs
Total length
(C) mm
Chromosome arms (mm)Arm Ratio
R: L/SCentromeric
indexRelativelengths
Chromosome typesLong
arm (L)Short arm (S)
I 9,8107 5,3293 4,4814 1,2233 45,6787 9,7341 m
II 7,9149 4,9922 2,9227 1,8068 36,9266 8,6254 sm
III 6,7210 4,8226 1,9315 2,5116 28,7383 7,3249 sm
IV 6,0544 4,3389 1,7154 2,5784 28,3331 6,6609 sm
V 5,1218 3,4838 1,6380 2,2017 31,9809 5,5530 sm
VI 4,6399 3,1086 1,5313 2,0240 33,0029 5,0530 sm
VII 4,2906 2,7588 1,5318 1,8786 35,7013 4,6728 sm
VIII 3,9805 2,4818 1,4987 1,7726 37,6510 4,3152 sm
Fig. 2 — (a) Mature embryo sac stained with PAS. (b) Starch grains in the hypostase with higher magnifi cation. (c) Nuclear cap with higher magnifi cation. (d) Obturator, PAS positive materials (*) secreted by obturator. Abbreviations: hy, Hypostase; ne, Nu-cellus epidermis; nc, Nucellar cap; ob, obturator; sg, starch grain. Scale bars:10 µm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4
EMBRYOLOGY AND CYTOCHEMISTRY OF ORNITHOGALUM SIGMOIDEUM 103
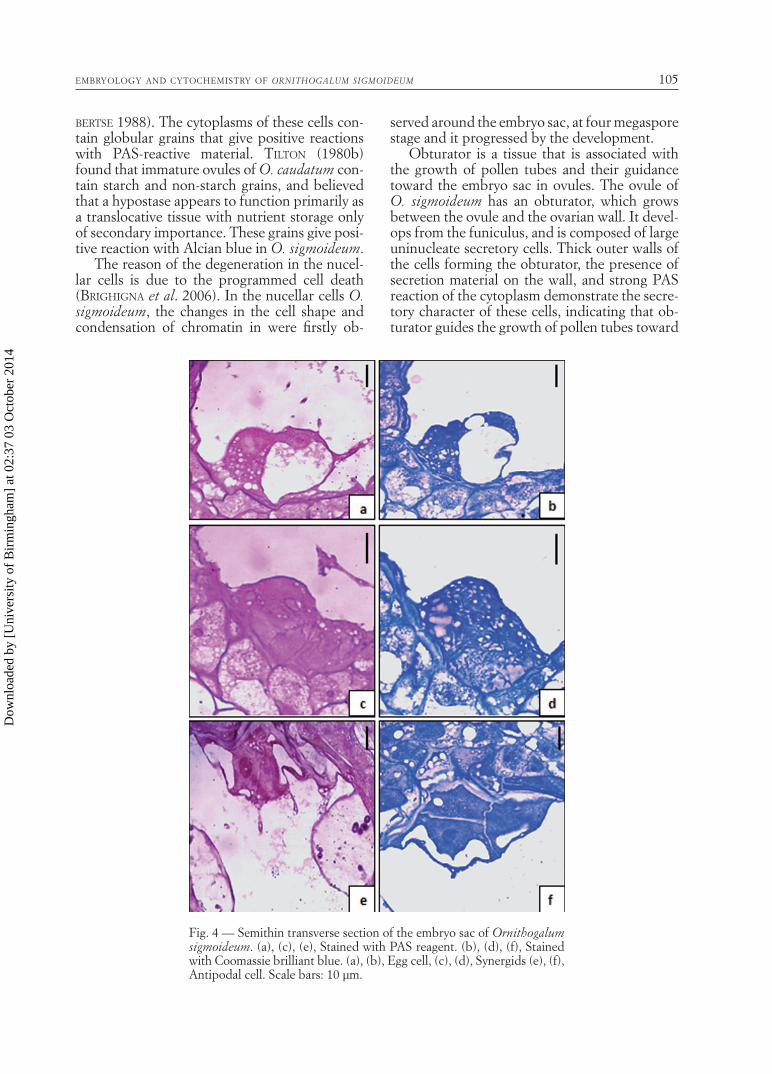
pole. A large vacuole in the micropylar pole of the cell occupies most of the volume. Contrary to the usual case, the egg cell is completely sur-rounded by a wall material stained with PAS.
In the mature embryo sac, the egg cell cyto-plasm gives a weak reaction to PAS, Alcian blue,
Coomassie brilliant blue, and Sudan black B (Figs. 4a, b). The cytochemical analysis revealed that it did not store detectable insoluble and acidic polysaccharides, proteins, and lipids, respectively.
Synergids - Synergid cells located at the mi-cropylar part of the embryo sac are smaller than
Fig. 3 — Megasporogenesis and development of embryo sac in Ornithogalum sigmoi-deum. (a) Megaspore mother cell. (b) Metaphase I. (c) Metaphase II. (d) Telophase II. (e) Linear megaspore tetrad. (f) Functional megaspore. (g) Binucleate embryo sac. (h) Four-nucleate embryo sac. (i) Mature embryo sac. Scale bars: 10 µm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4

ISMAILOGLU and UNAL104
the egg cell. Contrary to many species, they show unusual polarization; their nuclei and the bulk of the cytoplasm are located at the chalazal pole and a single, large vacuole is found at the chala-zal pole. They are ephemeral structures, and de-generate simultaneously soon after fertilization. The synergids are completely surrounded by a wall material giving PAS-positive reaction. The fi liform apparatus is the most conspicuous struc-ture in the synergids, and it extends from the wall in the proximal portion of the cell toward the center (Fig. 2c). It is a structure of the wall with fi nger-like projections into the cytoplasm and stains intensely for insoluble polysaccharides.
In the mature embryo sac, cytoplasm of syn-ergid cells gives a strong PAS-positive reaction; however, PAS reaction in the fi liform apparatus is much stronger (Fig. 4c). The cytoplasm also stains intensely with Commasie brillant blue (Fig. 4d). These reactions indicate that synergid cytoplasm is very rich in insoluble polysaccha-rides and proteins. It was established in staining with Alcian blue and Sudan black B that syn-ergid cells do not contain acidic polysaccharides and lipids.
Antipodal cells - The three antipodals are persistent uninucleate cells with dense, darkly stained cytoplasm. They become more darkly stained and fl at at later stages of development. Their remnants can be observed until the fi nal stages of development.
Antipodal cells in female gametophyte give a darker reaction than other embryo sac cells with PAS and Commassie brillant blue, and give a negative reaction with Alcian blue and Sudan black B (Fig. 4e, f). These results indicate that antipodal cells are rich in insoluble polysac-charides and proteins but do not contain acidic polysaccharides and lipids.
Central cell - A central cell is the largest cell in female gametophyte, and is occupied by a central vacuole. Most of the cell cytoplasm is peripherally located with narrow cytoplasmic strands extending toward the center. The polar nuclei of the central cell suspended in the center fuse before fertilization to form the secondary nucleus, which is located near antipodals.
Starch grains accumulate at the periphery of the female gametophyte during maturation, while they are rarely found after pollination (Fig. 4, 5). In the central cell, cytoplasmic nar-row strands are observed to give positive reac-tion with Coomassie brilliant blue.
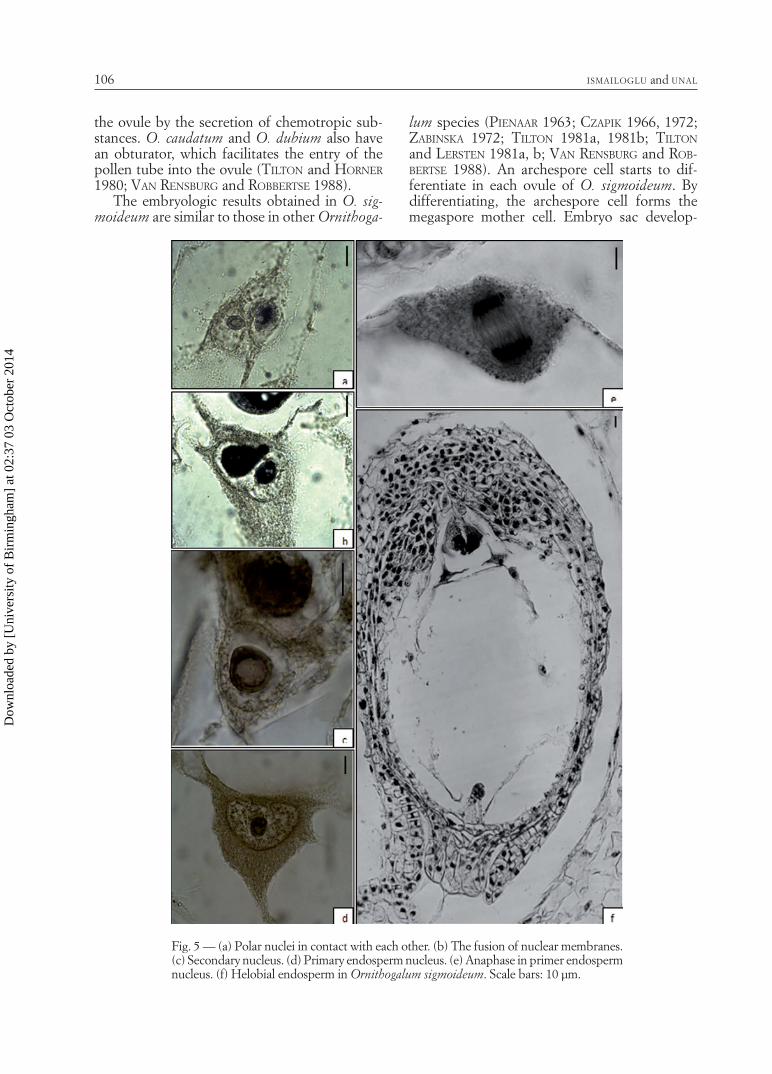
Endosperm development - Endosperm is helo-bial in O. sigmoideum. The primary endosperm
nucleus is formed by the fusion of a sperm with the secondary nucleus and divides soon thereaf-ter. The nucleus of primary endosperm divides at the level of antipodals to form a large micropy-lar and a small chalazal chamber. Many nuclear divisions take place in the micropylar chamber before it undergoes cellularization (Fig. 5a-f).
Embryo development - Embryo develop-ment is Asterad type (Figs. 6.a-l). The zygote is smaller than the egg cell, which is resulted from the shrinking of vacuole. The fi rst division of zy-gote is transversal, giving rise to an apical cell and a basal cell. The apical cell divides verti-cally to form two juxtaposed cells. The basal cell undergoes a transverse division to produce two superposed cells. Thus, a four-celled proembryo is formed, whose cells are divided in various planes to form a globular embryo. The globular embryo stage appears as prolonged, even at the early stages of seed development.
Cleavage polyembryony - In O. sigmoideum, polyembryony is identifi ed, which occurs by the cleavage of the proembryo. This rare phenom-enon is detected only in a single sac.
DISCUSSION
DAVIS (1984) recorded in Flora of Turkey that diploid chromosome number is 12, 16, 17, 19 and 20 in O. sigmoideum. In the present study, we identifi ed the diploid chromosome number of the material collected from the same popula-tion as 2n:16. In order to obtain our embryologi-cal and cytochemical results from the plants with the same chromosome number, the materials were collected from the same population.
In Ornithogalum species, the ovule is anatro-pous, bitegmic and crassinucellate (DAVIS 1966). TILTON and LERSTEN (1981a) demonstrated that in O. caudatum, the embryo sac is surrounded by the nucellar epidermis, which forms a secretory nucellar cap to attract the pollen tubes. Similar-ly, it was established in O. sigmoideum that nu-cellar epidermal cells and nucellar cap cells give positive reactions for insoluble polysaccharides and acidic polysaccharides; however, these cells do not contain starch grains. PAS-positive sub-stances released by the nucellar cap and stored in the micropylar opening may play a role in at-tracting the pollen tubes.
Similar to O. caudatum and O. dubium, in O. sigmoideum, hypostase cells near the embryo sac and the antipodals, in particular, vary in their content (TILTON 1980b; VAN RENSBURG and ROB-
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4
EMBRYOLOGY AND CYTOCHEMISTRY OF ORNITHOGALUM SIGMOIDEUM 105
BERTSE 1988). The cytoplasms of these cells con-tain globular grains that give positive reactions with PAS-reactive material. TILTON (1980b) found that immature ovules of O. caudatum con-tain starch and non-starch grains, and believed that a hypostase appears to function primarily as a translocative tissue with nutrient storage only of secondary importance. These grains give posi-tive reaction with Alcian blue in O. sigmoideum.
The reason of the degeneration in the nucel-lar cells is due to the programmed cell death (BRIGHIGNA et al. 2006). In the nucellar cells O. sigmoideum, the changes in the cell shape and condensation of chromatin in were fi rstly ob-
served around the embryo sac, at four megaspore stage and it progressed by the development.
Obturator is a tissue that is associated with the growth of pollen tubes and their guidance toward the embryo sac in ovules. The ovule of O. sigmoideum has an obturator, which grows between the ovule and the ovarian wall. It devel-ops from the funiculus, and is composed of large uninucleate secretory cells. Thick outer walls of the cells forming the obturator, the presence of secretion material on the wall, and strong PAS reaction of the cytoplasm demonstrate the secre-tory character of these cells, indicating that ob-turator guides the growth of pollen tubes toward
Fig. 4 — Semithin transverse section of the embryo sac of Ornithogalum sigmoideum. (a), (c), (e), Stained with PAS reagent. (b), (d), (f), Stained with Coomassie brilliant blue. (a), (b), Egg cell, (c), (d), Synergids (e), (f), Antipodal cell. Scale bars: 10 µm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4
ISMAILOGLU and UNAL106
the ovule by the secretion of chemotropic sub-stances. O. caudatum and O. dubium also have an obturator, which facilitates the entry of the pollen tube into the ovule (TILTON and HORNER 1980; VAN RENSBURG and ROBBERTSE 1988).
The embryologic results obtained in O. sig-moideum are similar to those in other Ornithoga-
lum species (PIENAAR 1963; CZAPIK 1966, 1972; ZABINSKA 1972; TILTON 1981a, 1981b; TILTON and LERSTEN 1981a, b; VAN RENSBURG and ROB-BERTSE 1988). An archespore cell starts to dif-ferentiate in each ovule of O. sigmoideum. By differentiating, the archespore cell forms the megaspore mother cell. Embryo sac develop-
Fig. 5 — (a) Polar nuclei in contact with each other. (b) The fusion of nuclear membranes. (c) Secondary nucleus. (d) Primary endosperm nucleus. (e) Anaphase in primer endosperm nucleus. (f) Helobial endosperm in Ornithogalum sigmoideum. Scale bars: 10 µm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4

EMBRYOLOGY AND CYTOCHEMISTRY OF ORNITHOGALUM SIGMOIDEUM 107
ment is monosporic Polygonum type. Following cellularization of the embryo sac, the micropy-lar part is round, while the chalazal part is pear-shaped.
In O. sigmoideum, the egg cell is surrounded by a complete wall. Observed in the form of a partial wall in many species, the wall of the egg cell has been detected in some plants; i.e., Capsella (SCHULZ and JENSEN 1968), Epidendrum
(COCUCCI and JENSEN 1969), Plumbago capensis (CASS 1972), Papaver (OLSON and CASS 1981), O. caudatum (TILTON 1981b), Scilla sibirica (BHANDARI and SACHDEVA 1983), S. autumnalis (COSKUN and UNAL 2010) and Genlisea aurea (PŁACHNO, 2010). In an electron microscope study on O. caudatum, TILTON (1981b) noted that the egg cell is surrounded by a complete wall which is thick and near the micropylar part
Fig. 6 — Embryo development in Ornithogalum sigmoideum (a) Zygote. (b) Two-celled proembryo. (c), (d) Four-celled proembryo. (e) Transverse cell division in basal cell. (f) Four-celled proembryo and two-celled suspensor. (g-l), The formation of globular embryo by the periclinal and anticlinal divisions in proembryo. Scale bars: 10 µm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4

ISMAILOGLU and UNAL108
of the cell, showing thinning or rupture toward the chalazal part. We observed in our study with PAS reaction that the cell is surrounded by a complete wall; yet, further EM studies will help confi rm our fi ndings.
Atypically, in O. sigmoideum, nuclei of syn-ergid cells are located at the chalazal part of the cell, which is different from other Ornithoga-lum species (ZABINSKA 1972; CZAPIK 1966, 1972; TILTON 1981b; TILTON and LERSTEN 1981a; VAN RENSBURG and ROBBERTSE 1988).
In O. sigmoideum, synergid cells are also surrounded by a complete wall which forms an indented structure into the cytoplasm at the micropylar part. This structure called the fi li-form apparatus gives a strong positive reaction with PAS, implying that the fi liform apparatus is rich in insoluble polysaccharides. The fact that synergids contain fi liform apparatus was demonstrated by DESOLE (1947) in O. umbel-latum and by TILTON (1981b) in O. caudatum; however, ZABINSKA (1972) did not identify the fi liform apparatus in O. gussonei. In O. gussonei, at least one synergid remains undegenerated af-ter fertilization, while synergids in O. umbella-tum degenerate at fertilization (ZABINSKA 1972; CZAPIK 1966). Similarly, in O. sigmoideum, both synergids degenerate just after fertilization. Synergid cytoplasm is rich in insoluble polysac-charides and proteins but poor in lipids and acidic polysaccharides. In Capsella bursa-pas-toris (SCHULZ and JENSEN 1968a), Nicotiana rus-tica (SEHGAL and GIFFORD 1979), Scilla sibirica (BHANDARI and SACHDEVA 1983), and O. cauda-tum (TILTON 1981b), synergid cytoplasms gave positive reactions for protein.
Antipodals remain undegenerated for a long time in O. gussonei as well; whereas they start to degenerate at an early stage in O. umbellatum (CZAPIK 1966; ZABINSKA 1972). The antipodal cytoplasms in O. sigmoideum give positive reac-tion for insoluble polysaccharides and proteins. These data make clear that the antipodal cells are metabolically very active.
Secondary nucleus is situated near the an-tipodals in O. sigmoideum. In O. dubium (VAN RENSBURG and ROBBERTSE 1988) and O. gussonei (ZABINSKA 1972), secondary nucleus is close to the antipodals, while it is located in the centre in O. caudatum (TILTON and LERSTEN 1981a).
Starch grains are located at the periphery of mature embryo sac, while the central cell rarely contains starch grains at later stages of devel-opment. Similarly, starch grains were observed in the central cells of O. caudatum (TILTON and
LERSTEN 1981a), O. dubium (VAN RENSBURG and ROBBERTSE 1988), and Scilla sibirica (BHANDARI and SACHDEVA 1983); however, no starch grains were found in O. gussonei (ZABINSKA 1972) and O. umbellatum (CZAPIK 1966; ZABINSKA 1972).
Endosperm development is helobial type in O. sigmoideum. Helobial endosperm devel-opment is observed in many species of the ge-nus Ornithogalum exemplifi ed with O. nutans (SCHNARF 1928), O. caudatum (CHIAPPINI 1962), O. umbellatum (CZAPIK 1966), and O. gussonei (ZABINSKA 1972).
In O. sigmoideum, the embryo development is Asterad type. The fi rst division of zygote is transversal. VAN RENSBURG and ROBBERTSE (1988) noted similar cases in O. dubium, stating that the suspensor is multicellular at the globular stage. In O. sigmoideum, the suspensor is four-celled at the globular embryo stage.
Acknowledgements — This work was supported by the Research Foundation of Marmara Univer-sity (Bapko), (No. FEN-DKR-250405-0114). We are thankful to Prof. Dr. Romana Czapik for her valuable support and advice. We dedicated this paper to the memory of Prof. Dr. Romana Czapik.
REFERENCES
BHANDARI, N.N., SACHDEVA, A., 1983 — Some aspects of organization and histochemistry of the embryo sac of Scilla sibirica Sato. Protoplasma, 116: 170-178.
BRIGHIGNA, L., PAPINI, A., MILOCANI, E., VESPRINI, J.L, 2006 — Programmed cell death in the nucellus of Tillandsia (Bromeliaceae). Caryologia, 59: (4) 334-339.
CASS, D.D., 1972 — Occurance and development of a fi liform apparatus in the egg of Plumbago capensis. American Journal of Botany, 59: 279-283.
CHIAPPINI. M., 1962 — Osservazioni carioembriolog-iche in Ornithogalum di Sardegna. Giornale Bo-tanica Italiano, 69: 91-102.
COCUCCI A., JENSEN W.A., 1969 — Orchid embryol-ogy: megagametophyte of Epidendrum scutella fol-lowing fertilization. American Journal of Botany, 56: 629-640.
COSKUN, Z.M., UNAL, M., 2010 — Embryological and Cytochemical features of Scilla autumnalis L. Turk-ish Journal of Botany, 34: 291-301.
CZAPIK, R., 1972 — Cyctoembryology of experimental hybrids between two related species of Ornithoga-lum L.. Acta Biologica Cracoviensia, XV: 165- 177.
CZAPIK, R., 1966 — The mechanism of cyctological dif-ferentiation in triploid populations of Ornithoga-lum umbellatum L.. Acta Biologica Cracoviensia, IX: 65- 89.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4

EMBRYOLOGY AND CYTOCHEMISTRY OF ORNITHOGALUM SIGMOIDEUM 109
CZAPIK, R., 1965 — Karyotype analysis of Ornithoga-lum umbellatum L and O. gussonei Ten.. Acta Bio-logica Cracoviensia, VIII: 21-36.
DAVIS, P. H., 1984- Flora of Turkey. Vol.VIII, Edin-burg.
DAVIS, G.L., 1966 — Systematic embryology of the an-giosperm. John Wiley, New York.
DESOLE, L., 1947 — Studio embriologico di Ornithog-alum umbellatum L.. Atti Società Toscana di Sci-enze Naturali - Memorie, 55: 3-8.
FEDER, N., O’BRIEN, T.P., 1968 — Plant microtech-nique: Some Principles and New Methods. Ameri-can Journal of Botany, 55: 123-142.
HESLOP-HARRISON, J., HESLOP-HARRISON, Y., KNOX, R.B., HOWLETT, B., 1973 — Pollen wall proteins: gametophytic and sporophytic fractions in the pol-len walls of the Malvaceae. Annals of Botany, 37: 403-412.
HESLOP-HARRISON, J., 1979 — Aspects of the structure, cytochemistry and germination of the polen of rye (Secale cereale L.). Annals of Botany, 44: 1-47.
HESLOP-HARRISON, Y., SHIVANNA, K.R., 1977 — The receptive surface of the angiosperm stigma. Annals of Botany, 41: 1233-1258.
KARAGOZOVA, M.D., KHANKH, F.V., 1972 — Basic charactristics and variations in the development of Ornithogalum nanum S.. Godoshnik na Sofi jskiya Universitet Biologicheski Facultet, Kniga 2. Bo-tanica, Microbiologiya, Fizyologiya i Biokhimiya na Rasteniyata, 64: 1-18.
LEVAN, A., FREDGA, K., SANDBERG, A.A., 1964 — No-menclature for centromeric position on chromo-some. Hereditas, 52: 201-220.
OLSON, A.R., CASS, D.D., 1981 — Changes in megag-ametophyte structure in Papaver nudicaule follow-ing in vitro placental pollination. American Jour-nal of Botany, 68: 1333-1341.
O’BRIEN, T.P., FEDER, N., MCCULLY, M.E., 1966 — Polychromatic staining of plant cell walls by Tolui-dine Blue O. Protoplasma, 59: 368-373.
PEARSE, A.G.E., 1961 — Histochemistry, Theoretical and Applied. 2nd end. Boston: Like Brown.
PIENAAR, R.D.E.V., 1963 — Citogenetiese studies in die genus Ornithogalum L.. inleidende oorsig. Journal of South African Botany, 29: 111-130.
PLACHNO, B.J., 2010 — Female germ unit in Genlisea and Utricularia, with remarks about the evolution of the extra-ovular female gametophyte in members of Lentibulariaceae. Protoplasma, DOI: 10.1007/s00709-010-0185-x.
SCHNARF, K., 1928 — Über die endospermentwicklung bei Ornithogalum. Oesterreichische Botanische Zeitschrift, 77: 173-177.
SEHGAL, C.B., GIFFORD, E.M.JR., 1979 — Development and Histochemical Studies of the Ovules of Nicotiana rustica L. Botanical Gazete, 140: 180-188.
SCHULZ, R., JENSEN, W.A., 1968a — Capsella Embryo-genesis: The Synergids Before and After Fertiliza-tion. American Journal of Botany, 55: 541-552.
TILTON, V.R., HORNER, H.T., 1980 — Stigma, style and obturator of Ornithogalum caudatum (Liliace-ae) and their function in the reproductive process. American Journal of Botany, 67: (7)1113-1131.
TILTON, V.R., LERSTEN, N.R., 1981a — Ovule develop-ment in Ornithogalum caudatum (Liliaceae) with a rewiew of selected papers on angiosperm reproduc-tion: I. Integuments, funikulus and vascular tissue. New Phytologist, 88: 439-457.
TILTON, V.R., LERSTEN, N.R., 1981b — Ovule develop-ment in Ornithogalum caudatum (Liliaceae) with a rewiew of selected papers on angiosperm repro-duction: III. Nucellus and megagametophyte. New Phytologist: 88: 477-504.
TILTON, V.R., 1980a — The nucellar epidermis and mi-cropyle of Ornithogalum caudatum (Liliaceae) with a review of these structures in other taxa. Canadian Journal of Botany, 58: 1872-1884.
TILTON, V.R., 1980b — Hypostase development in Or-nithogalum caudatum (Liliaceae) and notes on other types of modifi cations in the chalaza of angiosperm ovules. Canadian Journal of Botany, 58: 2059-2066.
TILTON, V.R., 1981a — Ovule development in Orni-thogalum caudatum (Liliaceae) with a rewiew of selected papers on angiosperm reproduction: II. Megaspoogenesis. New Phytologist, 88: 459-476.
TILTON, V.R., 1981b — Ovule development in Orni-thogalum caudatum (Liliaceae) with a rewiew of selected papers on angiosperm reproduction: IV. Egg apparatus structure and function. New Phy-tologist, 88: 505-531.
VAN RENSBURG, J.G.J., ROBBERTSE, P.J., 1988 — Seed development of Ornithogalum dubium, with spe-cial reference to fertilization and the egg apparatus. South African Journal of Botany, 54: (3)196-202.
ZABINSKA, D., 1972 — Embryo sac and endosperm de-velopment in some representatives of the group Or-nithogalum gussonei from Poland. Acta Biologica Cracoviensia, XV: 178-192.
Received July 14th 2010; accepted January 14th 2011
Dow
nloa
ded
by [
Uni
vers
ity o
f B
irm
ingh
am]
at 0
2:37
03
Oct
ober
201
4





























