Embed Size (px)
Citation preview

Quorum sensing and biofilm formation inStreptococcal infections
Dennis G. Cvitkovitch, … , Yung-Hua Li, Richard P. Ellen
J Clin Invest. 2003;112(11):1626-1632. https://doi.org/10.1172/JCI20430.
Members of the bacterial genus Streptococcus are responsible for causing a wide variety ofinfections in humans. Many Streptococci use quorum-sensing systems to regulate severalphysiological properties, including the ability to incorporate foreign DNA, tolerate acid, formbiofilms, and become virulent. These quorum-sensing systems are primarily made of smallsoluble signal peptides that are detected by neighboring cells via a histidinekinase/response regulator pair.
Perspective Series
Find the latest version:
http://jci.me/20430-pdf

1626 The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11
Streptococcal infectionsMembers of the genus Streptococcus (i.e., streptococci) areubiquitous parasites of humans. Some are part of theindigenous microflora that are involved in opportunis-tic infections such as dental caries and others are exoge-nous pathogens that cause infections ranging frommild respiratory or skin diseases to life-threatening con-ditions such as pneumonia, septic shock, and necrotiz-ing fasciitis. Contemporary research has found thatmany species of streptococci and other Gram positivebacteria have evolved similar peptide pheromone quo-rum-sensing systems that probably help them adapt toand survive host-imposed fluctuations in their localenvironment and coincidently regulate the expressionof virulence factors that promote their pathogenicity.This review focuses on streptococcal peptide signalingpathways that are population density–dependent andthat impact on vital survival and virulence traits.
BiofilmsBiofilms are dense aggregates of surface-adherentmicroorganisms embedded in an exopolysaccharidematrix. The study of bacteria residing in biofilms as aninteractive community rather than free-living plank-tonic cells has recently gained a great deal of attention.This has arisen, in part, because of the estimate by theCenters for Disease Control and Prevention that 65%of human bacterial infections involve biofilms. Many
species of streptococci are known to form biofilms;however, the relationship between the pathogenic stateand the biofilm mode of growth has been most clearlyestablished with the oral streptococci, which are knownto initiate dental caries when the bacteria are living inthe biofilm environment of dental plaque.
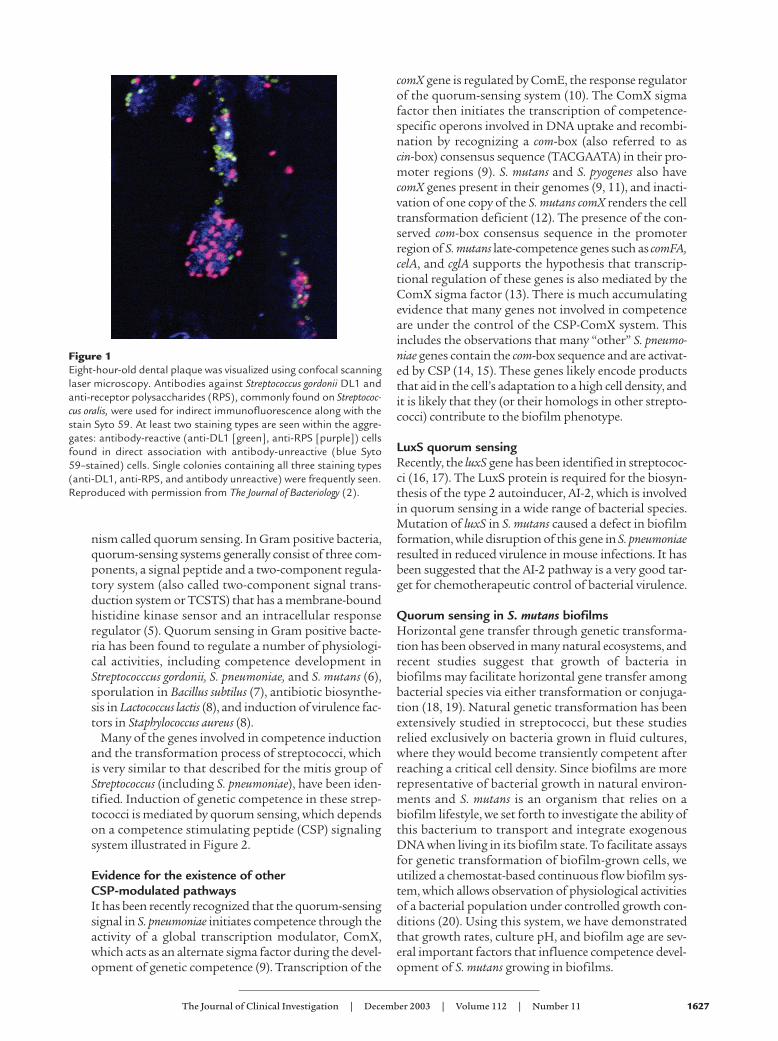
Dental plaqueThe human oral cavity is a complex ecosystem that sup-ports an extremely diverse microflora consisting of about500 species of microorganisms (1). Numerous physicaland nutritional interactions between oral bacteria con-tribute to a complex biofilm community (Figure 1) (2).Streptococci, including Streptococcus mutans, are ubiqui-tous in the oral microbiota of humans. S. mutans is con-sidered to be a principal etiological agent of dental caries,where it can cause dissolution of tooth enamel by acidend-products resulting from carbohydrate metabolism.The tooth surface is an indispensable natural habitat forS. mutans (3). S. mutans’ dental biofilm tropism most like-ly reflects its evolution of glucan synthesis and bindingfunctions as well as its relative aciduricity. Its competi-tiveness for this ecological niche may also relate to celldensity–dependent regulation of its acid toleranceresponse (ATR), natural genetic competence, and bacte-riocin activity. Since S. mutans has evolved to depend on abiofilm lifestyle for survival and persistence in the oralcavity combined with its role as an opportunisticpathogen, it has become the best-studied example of abiofilm-forming, disease-causing Streptococcus (4).Biofilm-like populations of pathogenic streptococci mayalso reach higher densities in confined areas like heartvalves, prosthetic devices, sinuses, tonsillar crypts, termi-nal respiratory passages, and in infectious skin lesions.
Quorum sensing in streptococciMany bacteria, including streptococci, are known to reg-ulate diverse physiological processes through a mecha-
Quorum sensing and biofilm formation in Streptococcal infections
Dennis G. Cvitkovitch, Yung-Hua Li, and Richard P. Ellen
Dental Research Institute, University of Toronto, Toronto, Ontario, Canada
Members of the bacterial genus Streptococcus are responsible for causing a wide variety of infections inhumans. Many Streptococci use quorum-sensing systems to regulate several physiological properties,including the ability to incorporate foreign DNA, tolerate acid, form biofilms, and become virulent. Thesequorum-sensing systems are primarily made of small soluble signal peptides that are detected by neigh-boring cells via a histidine kinase/response regulator pair.
J. Clin. Invest. 112:1626–1632 (2003). doi:10.1172/JCI200320430.
PERSPECTIVE SERIESQuorum sensing | E. Peter Greenberg, Series Editor
Address correspondence to: Dennis Cvitkovitch, Room 449A,Dental Research Institute, University of Toronto, 124 EdwardStreet, Toronto, Ontario M5G 1G6, Canada. Phone: (416) 979-4917 ext. 4592; Fax: (416) 979-4936; E-mail: [email protected] of interest: The authors have declared that no conflict ofinterest exists.Nonstandard abbreviations used: acid tolerance response (ATR);two-component signal transduction system (TCSTS);competence-stimulating peptide (CSP); extracellular inductioncomponent (EIC).
The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11 1627
nism called quorum sensing. In Gram positive bacteria,quorum-sensing systems generally consist of three com-ponents, a signal peptide and a two-component regula-tory system (also called two-component signal trans-duction system or TCSTS) that has a membrane-boundhistidine kinase sensor and an intracellular responseregulator (5). Quorum sensing in Gram positive bacte-ria has been found to regulate a number of physiologi-cal activities, including competence development inStreptococccus gordonii, S. pneumoniae, and S. mutans (6),sporulation in Bacillus subtilus (7), antibiotic biosynthe-sis in Lactococcus lactis (8), and induction of virulence fac-tors in Staphylococcus aureus (8).
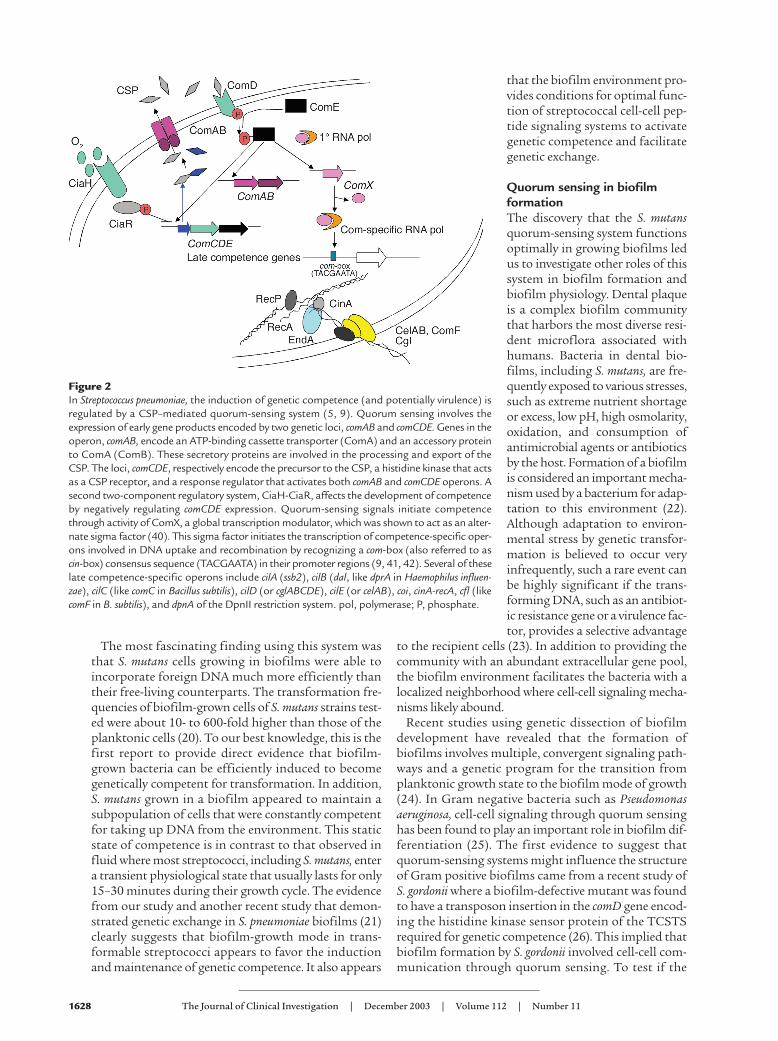
Many of the genes involved in competence inductionand the transformation process of streptococci, whichis very similar to that described for the mitis group ofStreptococcus (including S. pneumoniae), have been iden-tified. Induction of genetic competence in these strep-tococci is mediated by quorum sensing, which dependson a competence stimulating peptide (CSP) signalingsystem illustrated in Figure 2.
Evidence for the existence of other CSP-modulated pathwaysIt has been recently recognized that the quorum-sensingsignal in S. pneumoniae initiates competence through theactivity of a global transcription modulator, ComX,which acts as an alternate sigma factor during the devel-opment of genetic competence (9). Transcription of the
comX gene is regulated by ComE, the response regulatorof the quorum-sensing system (10). The ComX sigmafactor then initiates the transcription of competence-specific operons involved in DNA uptake and recombi-nation by recognizing a com-box (also referred to as cin-box) consensus sequence (TACGAATA) in their pro-moter regions (9). S. mutans and S. pyogenes also havecomX genes present in their genomes (9, 11), and inacti-vation of one copy of the S. mutans comX renders the celltransformation deficient (12). The presence of the con-served com-box consensus sequence in the promoterregion of S. mutans late-competence genes such as comFA,celA, and cglA supports the hypothesis that transcrip-tional regulation of these genes is also mediated by theComX sigma factor (13). There is much accumulatingevidence that many genes not involved in competenceare under the control of the CSP-ComX system. Thisincludes the observations that many “other” S. pneumo-niae genes contain the com-box sequence and are activat-ed by CSP (14, 15). These genes likely encode productsthat aid in the cell’s adaptation to a high cell density, andit is likely that they (or their homologs in other strepto-cocci) contribute to the biofilm phenotype.
LuxS quorum sensingRecently, the luxS gene has been identified in streptococ-ci (16, 17). The LuxS protein is required for the biosyn-thesis of the type 2 autoinducer, AI-2, which is involvedin quorum sensing in a wide range of bacterial species.Mutation of luxS in S. mutans caused a defect in biofilmformation, while disruption of this gene in S. pneumoniaeresulted in reduced virulence in mouse infections. It hasbeen suggested that the AI-2 pathway is a very good tar-get for chemotherapeutic control of bacterial virulence.
Quorum sensing in S. mutans biofilmsHorizontal gene transfer through genetic transforma-tion has been observed in many natural ecosystems, andrecent studies suggest that growth of bacteria inbiofilms may facilitate horizontal gene transfer amongbacterial species via either transformation or conjuga-tion (18, 19). Natural genetic transformation has beenextensively studied in streptococci, but these studiesrelied exclusively on bacteria grown in fluid cultures,where they would become transiently competent afterreaching a critical cell density. Since biofilms are morerepresentative of bacterial growth in natural environ-ments and S. mutans is an organism that relies on abiofilm lifestyle, we set forth to investigate the ability ofthis bacterium to transport and integrate exogenousDNA when living in its biofilm state. To facilitate assaysfor genetic transformation of biofilm-grown cells, weutilized a chemostat-based continuous flow biofilm sys-tem, which allows observation of physiological activitiesof a bacterial population under controlled growth con-ditions (20). Using this system, we have demonstratedthat growth rates, culture pH, and biofilm age are sev-eral important factors that influence competence devel-opment of S. mutans growing in biofilms.
Figure 1Eight-hour-old dental plaque was visualized using confocal scanninglaser microscopy. Antibodies against Streptococcus gordonii DL1 andanti-receptor polysaccharides (RPS), commonly found on Streptococ-cus oralis, were used for indirect immunofluorescence along with thestain Syto 59. At least two staining types are seen within the aggre-gates: antibody-reactive (anti-DL1 [green], anti-RPS [purple]) cellsfound in direct association with antibody-unreactive (blue Syto59–stained) cells. Single colonies containing all three staining types(anti-DL1, anti-RPS, and antibody unreactive) were frequently seen.Reproduced with permission from The Journal of Bacteriology (2).
1628 The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11
The most fascinating finding using this system wasthat S. mutans cells growing in biofilms were able toincorporate foreign DNA much more efficiently thantheir free-living counterparts. The transformation fre-quencies of biofilm-grown cells of S. mutans strains test-ed were about 10- to 600-fold higher than those of theplanktonic cells (20). To our best knowledge, this is thefirst report to provide direct evidence that biofilm-grown bacteria can be efficiently induced to becomegenetically competent for transformation. In addition,S. mutans grown in a biofilm appeared to maintain asubpopulation of cells that were constantly competentfor taking up DNA from the environment. This staticstate of competence is in contrast to that observed influid where most streptococci, including S. mutans, entera transient physiological state that usually lasts for only15–30 minutes during their growth cycle. The evidencefrom our study and another recent study that demon-strated genetic exchange in S. pneumoniae biofilms (21)clearly suggests that biofilm-growth mode in trans-formable streptococci appears to favor the inductionand maintenance of genetic competence. It also appears
that the biofilm environment pro-vides conditions for optimal func-tion of streptococcal cell-cell pep-tide signaling systems to activategenetic competence and facilitategenetic exchange.
Quorum sensing in biofilmformationThe discovery that the S. mutansquorum-sensing system functionsoptimally in growing biofilms ledus to investigate other roles of thissystem in biofilm formation andbiofilm physiology. Dental plaqueis a complex biofilm communitythat harbors the most diverse resi-dent microflora associated withhumans. Bacteria in dental bio-films, including S. mutans, are fre-quently exposed to various stresses,such as extreme nutrient shortageor excess, low pH, high osmolarity,oxidation, and consumption ofantimicrobial agents or antibioticsby the host. Formation of a biofilmis considered an important mecha-nism used by a bacterium for adap-tation to this environment (22).Although adaptation to environ-mental stress by genetic transfor-mation is believed to occur veryinfrequently, such a rare event canbe highly significant if the trans-forming DNA, such as an antibiot-ic resistance gene or a virulence fac-tor, provides a selective advantage
to the recipient cells (23). In addition to providing thecommunity with an abundant extracellular gene pool,the biofilm environment facilitates the bacteria with alocalized neighborhood where cell-cell signaling mecha-nisms likely abound.
Recent studies using genetic dissection of biofilmdevelopment have revealed that the formation ofbiofilms involves multiple, convergent signaling path-ways and a genetic program for the transition fromplanktonic growth state to the biofilm mode of growth(24). In Gram negative bacteria such as Pseudomonasaeruginosa, cell-cell signaling through quorum sensinghas been found to play an important role in biofilm dif-ferentiation (25). The first evidence to suggest thatquorum-sensing systems might influence the structureof Gram positive biofilms came from a recent study ofS. gordonii where a biofilm-defective mutant was foundto have a transposon insertion in the comD gene encod-ing the histidine kinase sensor protein of the TCSTSrequired for genetic competence (26). This implied thatbiofilm formation by S. gordonii involved cell-cell com-munication through quorum sensing. To test if the
Figure 2In Streptococcus pneumoniae, the induction of genetic competence (and potentially virulence) isregulated by a CSP–mediated quorum-sensing system (5, 9). Quorum sensing involves theexpression of early gene products encoded by two genetic loci, comAB and comCDE. Genes in theoperon, comAB, encode an ATP-binding cassette transporter (ComA) and an accessory proteinto ComA (ComB). These secretory proteins are involved in the processing and export of theCSP. The loci, comCDE, respectively encode the precursor to the CSP, a histidine kinase that actsas a CSP receptor, and a response regulator that activates both comAB and comCDE operons. Asecond two-component regulatory system, CiaH-CiaR, affects the development of competenceby negatively regulating comCDE expression. Quorum-sensing signals initiate competencethrough activity of ComX, a global transcription modulator, which was shown to act as an alter-nate sigma factor (40). This sigma factor initiates the transcription of competence-specific oper-ons involved in DNA uptake and recombination by recognizing a com-box (also referred to ascin-box) consensus sequence (TACGAATA) in their promoter regions (9, 41, 42). Several of theselate competence-specific operons include cilA (ssb2), cilB (dal, like dprA in Haemophilus influen-zae), cilC (like comC in Bacillus subtilis), cilD (or cglABCDE), cilE (or celAB), coi, cinA-recA, cfl (likecomF in B. subtilis), and dpnA of the DpnII restriction system. pol, polymerase; P, phosphate.

The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11 1629
S. mutans ComCDE quorum-sensing system wasinvolved in biofilm formation, we examined the abilityof S. mutans mutants defective in various componentsof the system to form biofilms. We found that inacti-vation of any one of the genes encoding the compo-nents of the quorum-sensing signaling system resultsin the formation of an abnormal biofilm (Figure 3)(12). Particularly, the comC mutant (unable to produceor secrete the signal peptide) formed a biofilm thatlacked the wild-type architecture, whereas the comDand comE mutants defective in sensing and respondingto the signal peptide formed biofilms with a reducedbiomass. The architectural change in the comC mutantbiofilms may be associated with a defect in cell separa-tion with mutations in this gene resulting in the for-mation of large aggregates or “weblike” biofilms thatwere easily removed from the surface. The observationthat the mutants unable to produce or secrete the CSPformed biofilms that differed from those formed by themutant defective in the comD or comE genes suggestedthat S. mutans has multiple CSP receptors. Although wehave clearly demonstrated that the ComCDE quorum-sensing system is directly connected to the ability of S.mutans to form biofilms, the molecular and biochemi-cal mechanisms involved in expression of the “wild-type biofilm phenotype” remain to be investigated.
CSP modulation of the ATRThe pH levels in dental biofilms are highly variable andfrequently shift from above pH 7.0 in the resting pHstate to as low as pH 3.0 during the ingestion of dietarycarbohydrates by the host. Thus, pH exerts a significantecological pressure on S. mutans, and its ability to toler-ate and grow in low pH environments is crucial to its sur-vival and eventual dominance in dental plaque, leadingto caries (27). Considerable evidence has shown that S.mutans has evolved a number of sophisticated mecha-nisms to survive these pH changes including inductionof an ATR in which exposure of S. mutans cells to a mild
or moderately acidic pH (5.0–6.0) results inenhanced survival of a significant proportionof the cell population in a lower pH of3.0–3.5 (28). This ATR involves a number ofde novo proteins that appear to be importantfor adaptation to an acidic environment (29).Although many of the molecular mecha-nisms of the ATR in S. mutans remainunclear, this “signal pH” that results in syn-thesis of protective proteins appears to beimportant for induction of the ATR.
It is widely accepted that bacteria living inbiofilms are more resistant to mechanical,physical, and chemical stresses. SinceS. mutans normally resides in high cell densi-ty biofilms, the ability to withstand acid inthis physiological state is likely an importantadaptive response. We therefore addressedthe question of whether acid adaptationinvolved cell density–dependent events or
cell-cell signaling in biofilms. Changes in external pHcan significantly influence many physiological param-eters, such as energy coupling, ion transport, protonmovement, and export of metabolic products, therebytriggering numerous secondary signals. During growthat pH 5.0, Escherichia coli can signal stress tolerance toother unadapted cells by secreting a proteinlike mole-cule, termed extracellular induction component (EIC)(30). Although the signal molecule remains unidenti-fied, induction of stress (including acid) adaptation inE. coli presumably involves cell-cell communication.Since S. mutans normally encounters acid while living indense biofilm communities, we proposed that quorumsensing at high cell densities of S. mutans might facili-tate its survival against low pH challenges. Testing thishypothesis has led us to find that the ATR interfaceswith the density-dependent signaling pathway that alsoinitiates genetic competence. We have demonstratedthat mutants defective in the comC, D, or E genes have adiminished log-phase ATR and the neutralized culturefiltrates prepared from acid-adapted wild-type cells alsoinduce a partial log-phase ATR in cells that have neverencountered the signal pH (31). S. mutans grown at highcell density established adaptation to the signal pHmore rapidly than cells grown at low density. Similarly,S. mutans cells grown in a high cell-density biofilm weremore resistant to the killing pH than planktonic-phasecells. In fact, S. mutans cells grown in biofilms not onlysurvived better than the planktonic cells but also werecapable of growth at the lower pH following a glucosepulse. Based on the evidence obtained from this work,we propose that S. mutans, upon exposure to low pH ina growing culture, releases extracellular signal mole-cules, one of which is the CSP, to enhance induction ofacid adaptation in the population. It is likely that opti-mal induction of acid adaptation in a population of S.mutans requires a coordinated activity through mecha-nisms involving both low pH induction and cell density–dependent intercellular signals.
Figure 3Scanning confocal laser microscope images of 16 h S. mutans biofilms. Panels a–cshow the X-Y planes (top view), while the panels d–f show the X-Z planes (sideview). The panels illustrate (a, d) a normal biofilm formed by the parent strainNG8 and (b, e) an aberrant biofilm formed by the comC– mutant defective in mak-ing a CSP. The wild-type appearance of the biofilm is restored by addition of syn-thetic CSP (c, f). Reproduced with permission from The Journal of Bacteriology (12).

1630 The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11
Biofilms likely provide bacterial cells with a uniqueenvironment to fully express their adaptive survivalmechanisms. Because of three-dimensional structures,high cell density, and diffusion barriers, bacterial cells atdifferent locations within a biofilm may not sense thesame degree of extracellular stress simultaneously. Thecells that first sense a pH stress may rapidly process theinformation and pass their “secondary signal” to theother members of the population through cell-cell sig-naling systems to initiate a coordinated protectiveresponse against potentially lethal forces, like acid. Unlikesome planktonic cells that need to reach a critical con-centration of signal molecules and cell density, biofilmscan allow signal molecules to accumulate rapidly in thelocal environment to initiate coordinated activities farmore quickly (25). In addition, physiological states of bac-terial cells living in a biofilm, in terms of growth rate,growth phase, or metabolic activities, are heterogeneous;this allows the cells to respond to stress in different ways.Apparently, biofilm populations have several advantagesover their free-living counterparts since the cells havemore time, a sufficient concentration of signal molecules,and high population density to adapt to stress relative toplanktonic cells. The high cell density biofilms may pro-vide a unique environment for induction of acid adapta-tion via quorum sensing in S. mutans. It is likely that theS. mutans quorum-sensing signaling pathway is signifi-cant for the ATR to intersect with the regulatory net-works initiating genetic competence as well as a switch tothe “biofilm phenotype.”
The ComC signal peptide may activate more than one signal transduction pathwayOur previous study revealed that an S. mutans comCmutant unable to produce CSP formed a biofilm thatdiffered from that formed by the mutant defective inthe comD or comE genes suggested that there might be asecond receptor that also responded to the CSP but acti-vated a different pathway in order to invoke the pheno-type (12, 20). Based on the available information, wehave proposed a “two-receptor” cell-cell signaling modelto illustrate how the quorum-sensing system in S.mutans functions to regulate genetic competence,biofilm formation, and the ATR (12). The principle ofthis model is that the signal peptide (CSP) encoded bycomC can simultaneously interact with two cognatereceptors, one encoded by comD and another encodedby an unknown gene. These receptors likely transfer theinput signal through two different pathways. Althoughthe genes encoding the components involved in the sec-ond transduction pathway remain unknown, the workin our lab has recently characterized another TCSTS,named HK/RR11, which is also demonstrated to involvebiofilm formation and acid resistance in S. mutans (32).
One of the defects observed in HK/RR11 mutantbiofilms was the development of a spongelike architec-ture composed of cells in very long chains, a feature thatwe previously observed with the biofilm formed by thecomC mutant unable to produce CSP. Since there is no
putative substrate, signal or function assigned for theHK/RR11 transduction system, we suspected that theTCSTS encoded by hk/rr11 may activate a second path-way to respond to the CSP. To test this hypothesis, weadded CSP to the HK/RR11 mutant cultures to assessthe effect on chain formation by the hk11 and rr11mutant biofilm cells. The results revealed that additionof CSP to the mutant cultures had no observableimpact on the length of cell chains comprising themutant biofilms. This result was consistent with HK11acting as a CSP receptor but provided no direct evidenceto conclusively assign a role to HK11 as a CSP receptor.A study for characterizing the relationship between thecomC-encoded signal peptide and the HK/RR11 signaltransduction pathway is now underway.
Evaluation of CSP analogs to act as inhibitors of biofilm formationAnalogs of quorum-sensing peptides can competitive-ly inhibit the activity of the peptide-mediated pheno-type. It has been demonstrated that analogous signalpeptides from Staphylococcus epidermidis can inhibit theagr signaling system of Staphylococcus aureus, therebymodulating S. aureus virulence (33). Since peptide sig-naling–sensing systems are very similar among Grampositive bacteria (including the S. mutans CSP system)analogs of the S. mutans CSP may be able to interferewith the quorum-sensing process, leading to inhibitionof induction of the “biofilm phenotype.”
Mucosal pathogensQuorum sensing among mucosal pathogens may notrequire high cell density. In contrast to the very densedental plaques colonized by S. mutans and several otheroral streptococci, pathogenic streptococci that infectmucosal tissues colonize environments where bacterialmicrocolonies are usually less dense due to bathingeffects of secretions and desquamation of epithelium.Yet, several of these streptococcal and other Gram pos-itive species have evolved peptide pheromone signalingpathways that have autoregulating functions analogousto the quorum-sensing systems that we described abovefor S. mutans. The finding that initiation of genetic com-petence in S. pneumoniae is sensitive to relatively low lev-els of CSP at about 107 bacteria/ml (34) suggests thatpathogenic streptococci of mucosal surfaces mayrespond to gradients of cell densities that occur fre-quently during natural infections, raising the questionof whether quorum sensing affects pathways by whichpathogenic streptococci grow to predominance overnormally protective indigenous species. It has beenrecently established that mutants of S. pneumoniae defec-tive in the quorum-sensing competence induction path-way have diminished virulence in a murine model rela-tive to the parent strain (35). Differential fluorescenceinduction also showed that several Com genes wereinduced during infection in mice (36). Clearly, the roleof quorum sensing during infection by S. pneumoniaewarrants further examination.

The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11 1631
BacteriocinsBacteriocins are antimicrobial peptides that are generat-ed by some bacteria and target others that are sensitive.The signaling networks that regulate bacteriocin pro-duction and transport as well as immunity to bacteri-ocins involve peptide pheromone sensing pathways thatare very similar to those involved in genetic competence(8). Operons encoding bacteriocin peptide precursors,cognate peptide processing transporters, and two-com-ponent regulatory sensors and response regulators areknown to be under the control of cell density. For exam-ple, the virulence-related blp and com regulons of S. pneu-moniae have many analogous features including highlysimilar signal peptides and peptide secretion systems(15). Moreover, activation of the competence and bacte-riocin peptide pheromone response regulators may, inturn, activate downstream pathways that intersect in acommon regulatory gene. One such example is the pre-viously mentioned comX, a transcriptional regulator ofso-called “late” competence genes (9). The genome of M1S. pyogenes strain SF370 contains comX and a completebacteriocin salA1 locus; yet, it lacks the genes comABC thatencode the CSP and its secretion apparatus (11). Thisimplies that S. pyogenes comX (and downstream pathways)may be activated in response to environmental signalsother than CSPs. S. pyogenes is a pathogen with a hostrange limited to humans and a unique set of virulenceproperties not shared by other streptococci. It is possiblethat it has evolved a sophisticated bacteriocin peptide reg-ulatory system to help its population competitivelyemerge on mucosal surfaces, yet protect its genome fromcontamination with foreign DNA by deleting (or neverevolving) genes for the cell density–dependent CSP.Recently, Jenkinson’s lab reported that the sensory andimmunity pathways for the pheromone antibiotics SalA1of S. pyogenes and SalA of the indigenous oral species Strep-tococcus salivarius are conserved, closely related, and cross-sensitive (37). Such novel findings suggest that determi-nants of population dynamics on bathed mucosalsurfaces probably encompass subtle combinations ofcomplex environmental sensing systems that are not lim-ited to cell density, bacteriocin production, and compe-tence stimulation. It is also possible that S. pyogenes uti-lizes a bacteriocin-like hemolysin as a quorum-sensingmolecule. Recently, Enterococcus faecalis was shown to reg-ulate gene expression by such a mechanism involving itscytolysin, a molecule with both antimicrobial andhemolytic activity (38). S. pyogenes has a hemolysin that isgenetically very similar to quorum-sensing autoinductionoperons found in streptococci (39).
Fluctuations in total bacterial burden and populationdensity also determine the pathogenicity of bacterialmicrocolonies and biofilms. For example, the ability tosuppress competing streptococci through bacteriocinactivity may be considered a virulence property if it allowsthe pathogenic species to emerge sufficiently to damagethe host. Evidence is growing that cell density–dependentgene regulation also affects the expression of many othervirulence-related proteins of pathogenic streptococci. For
example, virulence factor expression and regulation are,in part, determined by growth phase and activation oftwo-component regulatory sensors of environmental sig-nals, which are characteristics of quorum sensing inGram positive bacteria (38). Thus, the extracellular con-centration of autoregulating peptides appears to serve asone of several environmental conditions that regulate vir-ulence genes through signal transduction pathways thatare initiated via two-component regulatory systems. In S.pyogenes, in addition to sagA, there appear to be severalcoordinated regulatory loci such as mga, csrRS, fasBCA,and rgg that affect the expression of genes for its numer-ous and diverse virulence factors.
Such a pattern of global regulation is reminiscent ofthe induction of virulence genes via quorum-sensingpathways in biofilms of Pseudomonas aeruginosa. There-fore, it is likely that the genomes of streptococci haveevolved density-dependent regulons to control expres-sion of downstream genes that affect bacterial survivalin response to changing environmental conditions onmucosal and tooth surfaces, and the selective survivalof the pathogenic streptococcal species leads to clinicalinfection. Although the key molecules of the quorum-sensing pathways of pathogenic streptococci such as S.pneumoniae, S. pyogenes, and S. mutans are distinct fromthose first reported for Gram negative bacteria, theyseem to serve an analogous function, to modulatephysiologic homeostasis and adaptation to environ-mental conditions in response to fluctuations in pop-ulation density. The use of this information to exploitthese pathways to control streptococcal infections isnow being implemented and in the near future we maysee a more selective and targeted approach to control-ling persistent biofilm-dwelling bacteria.
AcknowledgmentsSpecial thanks to Paul Kolenbrander and Rob Palmerfrom the National Institute of Dental and CraniofacialResearch (NIDCR) for providing Figure 1. Work con-ducted by the authors was generously funded by theCanadian Institutes of Health Research, the NIDCR,and the Canada Foundation for Innovation.
1. Kroes, I., Lepp, P.W., and Relman, D.A. 1999. Bacterial diversity withinthe human subgingival crevice. Proc. Natl. Acad. Sci. U. S. A.96:14547–14552.
2. Palmer, R.J., Jr., et al. 2003. Coaggregation-mediated interactions ofstreptococci and actinomyces detected in initial human dental plaque.J. Bacteriol. 185:3400–3409.
3. Carlsson, J., Soderholm, G., and Almfeldt, I. 1969. Prevalence of Strepto-coccus sanguis and Streptococcus mutans in the mouth of persons wearingfull-dentures. Arch. Oral Biol. 14:243–249.
4. Burne, R.A. 1998. Oral streptococci . . . products of their environment.J. Dent. Res. 77:445–452.
5. Kleerebezem, M., et al. 1997. Quorum sensing by peptide pheromonesand two-component signal-transduction systems in Gram-positive bac-teria. Mol. Microbiol. 24:895–904.
6. Cvitkovitch, D.G. 2001. Genetic competence and transformation in oralstreptococci. Crit. Rev. Oral Biol. Med. 12:217–243.
7. Lazazzera, B.A. 2000. Quorum sensing and starvation: signals for entryinto stationary phase. Curr. Opin. Microbiol. 3:177–182.
8. Kleerebezem, M., and Quadri, L.E. 2001. Peptide pheromone-dependentregulation of antimicrobial peptide production in Gram-positive bacte-ria: a case of multicellular behavior. Peptides. 22:1579–1596.
9. Lee, M.S., and Morrison, D.A. 1999. Identification of a new regulator in

1632 The Journal of Clinical Investigation | December 2003 | Volume 112 | Number 11
Streptococcus pneumoniae linking quorum sensing to competence forgenetic transformation. J. Bacteriol. 1999. 181:5004–5016.
10. Ween, O., Gaustad, O., and Havarstein, L.S. 1999. Identification of DNAbinding sites for ComE, a key regulator of natural competence in Strep-tococcus pneumoniae. Mol. Microbiol. 33:817–827.
11. Ferretti, J.J., et al. 2001. Complete genome sequence of an M1 strain ofStreptococcus pyogenes. Proc. Natl. Acad. Sci. U. S. A. 98:4658–4663.
12. Li, Y.H., et al. 2002. A quorum-sensing signaling system essential forgenetic competence in Streptococcus mutans is involved in biofilm forma-tion. J. Bacteriol. 184:2699–2708.
13. Lee, J.H., et al. 2001. Genetic transformation in Streptococcus mutans: iden-tification of competence genes by functional genomic analysis. Abstractsof the 101st Annual General Meeting of the American Society for Microbiology.Orlando, Florida, USA. (Abstr. I-76)
14. Peterson, S., et al. 2000. Gene expression analysis of the Streptococcus pneu-moniae competence regulons by use of DNA microarrays. J. Bacteriol.182:6192–6202.
15. de Saizieu, A., et al. 2000. Microarray-based identification of a novelStreptococcus pneumoniae regulon controlled by an autoinduced peptide.J. Bacteriol. 182:4696–4703.
16. Merritt, J., et al. 2003. Mutation of luxS affects biofilm formation inStreptococcus mutans. Infect. Immun. 71:1972–1979.
17. Stroeher, U.H., et al. 2003. Mutation of luxS of Streptococcus pneumoniaeaffects virulence in a mouse model. Infect. Immun. 71:3206–3212.
18. Christensen, B.B., et al. 1998. Establishment of new genetic traits in amicrobial biofilm community. Appl. Environ. Microbiol. 64: 2247–2255.
19. Hausner, M., and Wuertz, S. 1999. High rates of conjugation in bacteri-al biofilms as determined by quantitative in situ analysis. Appl. Environ.Microbiol. 65:3710–3713.
20. Li, Y.H., et al. 2001. Natural genetic transformation of Streptococcusmutans growing in biofilms. J. Bacteriol. 183:897–908.
21. Waite, R.D., Struthers, J.K., and Dowson, C.G. 2001. Spontaneoussequence duplication within an open reading frame of the pneumococ-cal type 3 capsule locus causes high-frequency phase variation. Mol.Microbiol. 45:1223–1232.
22. Donlan, R.M., and Costerton, J.W. 2002. Biofilms: survival mechanismsof clinically relevant microorganisms. Clin. Microbiol. Rev. 15:167–193.
23. Dowson, C.G., et al. 1997. Horizontal gene transfer and the evolution ofresistance and virulence determinants in Streptococcus. Soc. Appl. Bacteriol.Symp. Ser. 26:42S–51S.
24. O’Toole, G.A., and Kolter, R. 1998. Initiation of biofilm formation inPseudomonas fluorescens WCS365 proceeds via multiple, convergent sig-nalling pathways: a genetic analysis. Mol. Microbiol. 28:449–461.
25. Parsek, M.R., and Greenberg, E.P. 2000. Acyl-homoserine lactone quo-rum sensing in gram-negative bacteria: a signaling mechanism involvedin associations with higher organisms. Proc. Natl. Acad. Sci. U. S. A.97:8789–8793.
26. Loo, C.Y., Corliss, D.A., and Ganeshkumar, N. 2000. Streptococcus gordonii
biofilm formation: identification of genes that code for biofilm pheno-types. J. Bacteriol. 182:1374–1382.
27. Kuramitsu, H.K. 1993. Virulence factors of mutans streptococci: role ofmolecular genetics. Crit. Rev. Oral Biol. Med. 4:159–176.
28. Svensater, G., et al. 1997. Acid tolerance response and survival by oralbacteria. Oral Microbiol. Immunol. 12:266–273.
29. Wilkins, J.C., Homer, K.A., and Beighton, D. 2002. Analysis of Streptococ-cus mutans proteins modulated by culture under acidic conditions. Appl.Environ. Microbiol. 68:2382–2390.
30. Rowbury, R.J., and Goodson, M. 2001. Extracellular sensing and sig-nalling pheromones switch-on thermotolerance and other stressresponses in Escherichia coli. Sci. Prog. 84:205–233.
31. Li, Y.H., et al. 2001. Cell density modulates acid adaptation in Streptococ-cus mutans: implications for survival in biofilms. J. Bacteriol.183:6875–6884.
32. Li, Y.H., et al. 2002. A novel two-component regulatory system involvedin biofilm formation and acid resistance in Streptococcus mutans. J. Bacte-riol. 184:6333–6342.
33. Otto, M., et al. 1999. Inhibition of virulence factor expression in Staphy-lococcus aureus by the Staphylococcus epidermidis agr pheromone and deriv-atives. FEBS Lett. 450:257–262.
34. Steinmoen, H., Knutsen, E., and Havarstein, L.S. 2002. Induction of nat-ural competence in Streptococcus pneumoniae triggers lysis and DNArelease from a subfraction of the cell population. Proc. Natl. Acad. Sci. U. S. A. 99:7681–7686.
35. Lau, G.W., et al. 2001. A functional genomic analysis of type 3 Strepto-coccus pneumoniae virulence. Mol. Microbiol. 40:555–571.
36. Bartilson, M., et al. 2001. Differential fluorescence induction revealsStreptococcus pneumoniae loci regulated by competence stimulatory pep-tide. Mol. Microbiol. 39:126–135.
37. Upton, M., et al. 2001. Intra- and interspecies signaling between Strepto-coccus salivarius and Streptococcus pyogenes mediated by SalA and SalA1antibiotic peptides. J. Bacteriol. 183:3931–3938.
38. Haas, W., Shepard, B.D., and Gilmore, M.S. 2002. Two-component reg-ulator of Enterococcus faecalis cytolysin responds to quorum-sensingautoinduction. Nature. 415:84–87.
39. Nizet, V., et al. 2000. Genetic locus for streptolysin S production bygroup A streptococcus. Infect. Immun. 68:4245–4254.
40. Luo, P., and Morrison, D.A. 2003. Transient association of an alternativesigma factor, ComX, with RNA polymerase during the period of com-petence for genetic transformation in Streptococcus pneumoniae. J. Bacteri-ol. 185:349–358.
41. Pestova, E.V., and Morrison, D.A. 1998. Isolation and characterizationof three Streptococcus pneumoniae transformation-specific loci by use of alacZ reporter insertion vector. J. Bacteriol. 180:2701–2710.
42. Campbell, E.A., Choi, S.Y., and Masure, H.R. 1998. A competence regu-lon in Streptococcus pneumoniae revealed by genomic analysis. Mol. Micro-biol. 27:929–939.









![Natural Anti-Quorum Sensing agents against Pseudomonas ... · 2. Quorum Sensing: a Novel Target Vfr Quorum sensing (QS) is a population-dependent event [13]. The capability to sense](https://img.pdfslide.us/doc/110x75/5edbcc02ad6a402d66663060/natural-anti-quorum-sensing-agents-against-pseudomonas-2-quorum-sensing-a.jpg)



















