Embed Size (px)
Citation preview

Stoichiometry and Energy Cost of CO2 Assimilation
• Fixation of three CO2 molecules yields one
glyceraldehyde 3-phosphate
• Nine ATP molecules and six NADPH molecules are consumed


Photosynthesis: From Light and CO2 to Glyceraldehyde 3-
phosphate
• The photosynthesis of one molecule of glyceraldehyde 3-phosphate requires the capture of roughly 24 photons
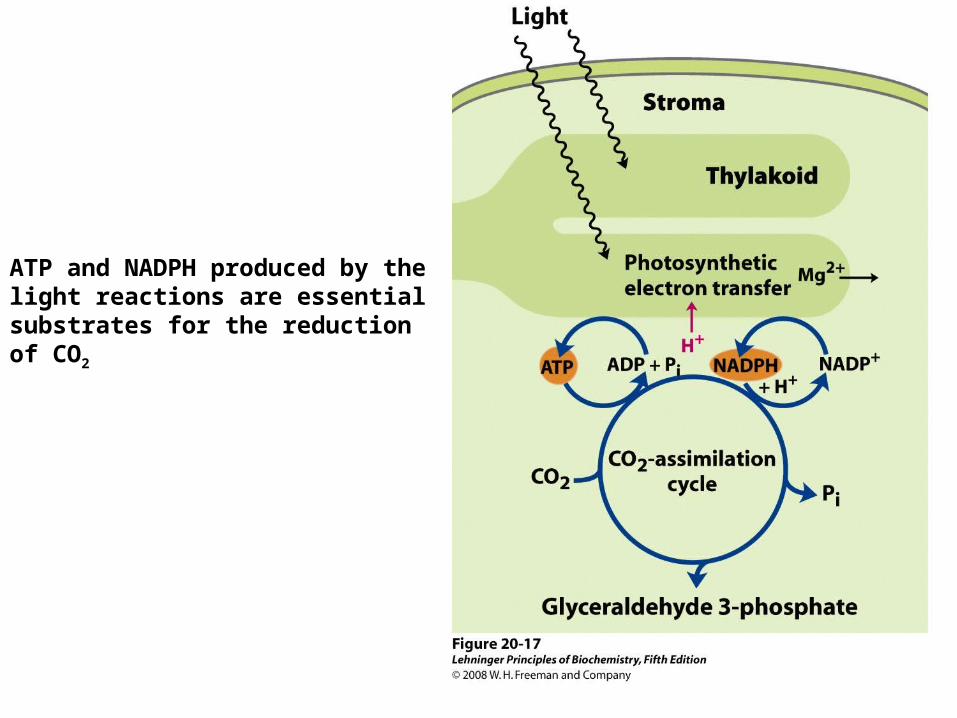
ATP and NADPH produced by the light reactions are essential substrates for the reduction of CO2

Enzymes in the Calvin Cycle are Regulated by Light
• Target enzymes are – ribulose 5-phosphate kinase, – fructose 1,6-bisphosphatase, – seduloheptose 1,7-bisphosphatase, and– glyceraldehyde 3-phosphate
dehydrogenase
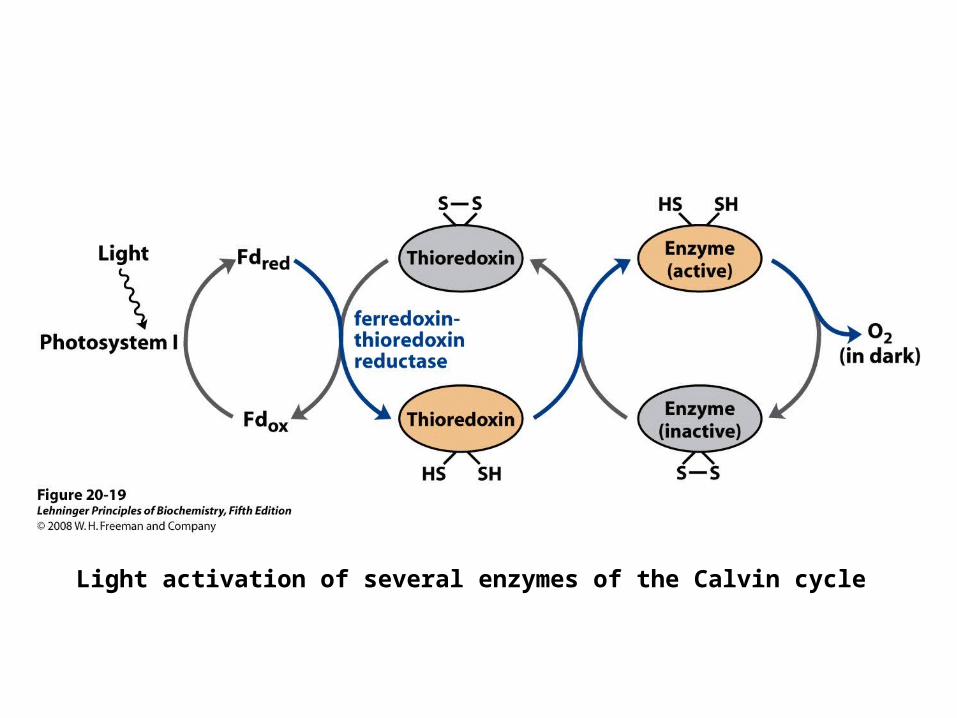
Light activation of several enzymes of the Calvin cycle

Photorespiration• So far, we saw that plants oxidize water to O2
and reduce CO2 to carbohydrates during the
photosynthesis
• Plants also have mitochondria where usual respiration with consumption of O2 occurs in the
dark
• In addition, a wasteful side reaction catalyzed by Rubisco occurs in mitochondria
• This reaction consumes oxygen and is called photorespiration; unlike mitochondrial respiration, this process does not yield energy

Oxygenase Activity of Rubisco
• The reactive nucleophile in the Rubisco reaction is the electron-rich enediol form of ribulose 1,5-bisphosphate
• The active site meant for CO2 also
accommodates O2
• Mg++ also stabilizes the hydroperoxy anion that forms by electron transfer from the enediol to oxygen


Salvage of 2-Phosphoglycerate
• Complex ATP-consuming process for the recovery of C2 fragments from the
photorespiration
• Requires oxidation of glycolate with molecular oxygen in peroxisomes, and formation of H2O2
• Involves a loss of a carbon as CO2 by
mitochondrial decarboxylation of glycine
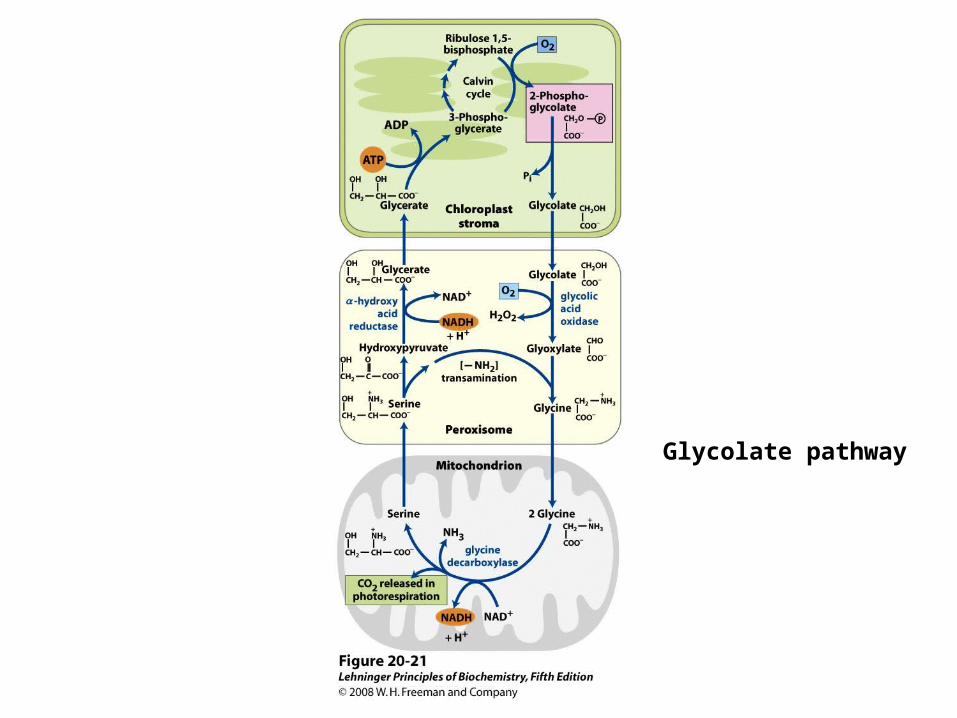
Glycolate pathway

Rubisco in C3 Plants Cannot
Avoid Oxygen
• Plants that assimilate dissolved CO2 in the
mesophyll of the leaf into three-carbon 3-phosphoglycerate are called the C3 plants
• Our atmosphere contains about 21% of oxygen and 0.038% of carbon dioxide
• The dissolved concentrations in pure water are about 260 M O2 and 11 M CO2 (at the
equilibrium and room temperature)
• The Km of Rubisco for oxygen is about 350 M

Separation of CO2 capture and the
Rubisco Reaction in C4 Plants
• Many tropical plants avoid wasteful photorespiration by a physical separation of CO2 capture and Rubisco
activity
• CO2 is captured into oxaloacetate (C4) in mesophyll
cells
• CO2 is transported to bundle-sheath cells where
Rubisco is located
• The local concentration of CO2 in bundle-sheath cells
is much higher than the concentration of O2

Carbon assimilation in C4 plants

Chapter 20: Summary
• ATP and NADPH from light reactions are needed in order to assimilate
CO2 into carbohydrates
• Assimilations of three CO2 molecules via the Calvin cycle leads to the
formation of one molecule of 3-phosphoglycerate
• 3-Phosphoglycerate is a precursor for the synthesis of larger
carbohydrates such as fructose and starch
• The key enzyme of the Calvin cycle, Rubisco, fixes carbon dioxide into
carbohydrates
• Low selectivity of Rubisco causes a wasteful incorporation of molecular
oxygen in C3 plants; this is avoided in C4 plants by increasing the
concentration of CO2 near Rubisco
In this chapter, we learned that:

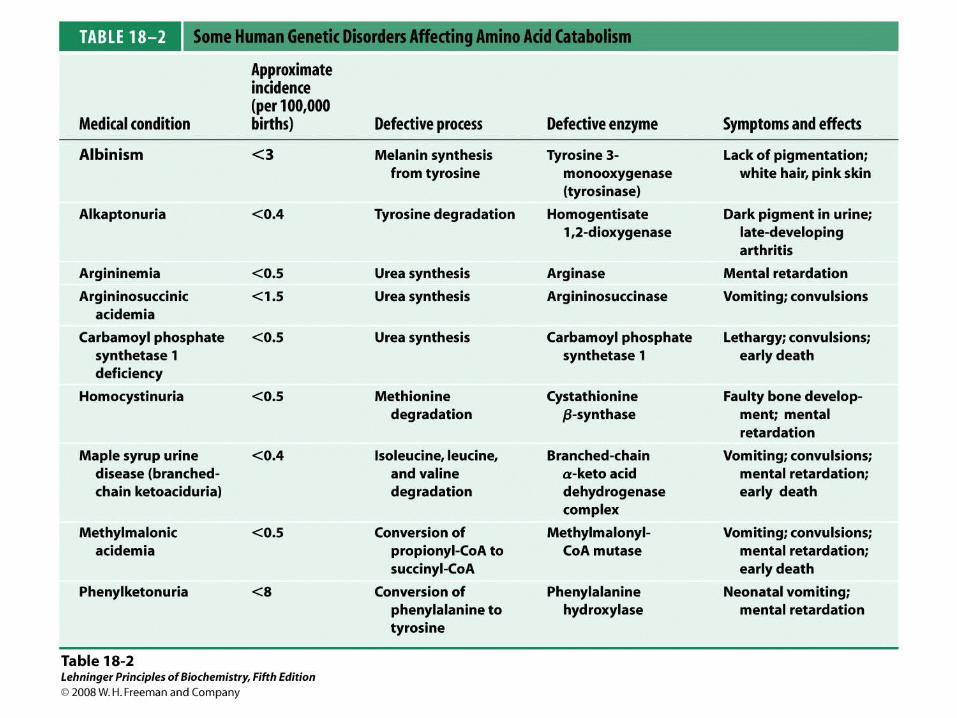
Lecture Connections18 | Amino Acid Oxidation Production of Urea
© 2009 W. H. Freeman and Company

CHAPTER 18 Amino Acid Oxidation
Production of Urea
– How proteins are digested in animals– How amino acids are degraded in animals– How urea is made in made and excreted
Key topics:

Oxidation of Amino Acids is a Significant Energy-Yielding Pathway
in Carnivores
• Not all organisms use amino acids as the source of energy
• About 90% of energy needs of carnivores can be met by amino acids immediately after a meal
• Only a small fraction of energy needs of herbivores are met by amino acids
• Microorganisms scavenge amino acids from their environment for fuel


Metabolic Circumstances of Amino Acid Oxidation
Amino acids undergo oxidative catabolism under three circumstances:
– Leftover amino acids from normal protein turnover are degraded
– Dietary amino acids that exceed body’s protein synthesis needs are degraded
– Proteins in the body are broken down to supply amino acids for catabolism when carbohydrates are in short supply (starvation, diabetes mellitus),

Dietary Proteins are Enzymatically Hydrolyzed
• Pepsin cuts protein into peptides in the stomach• Trypsin and chymotrypsin cut proteins and larger
peptides into smaller peptides in the small intestine
• Aminopeptidase and carboxypeptidases A and B degrade peptides into amino acids in the small intestine


Enzymatic Degradation of Dietary Proteins

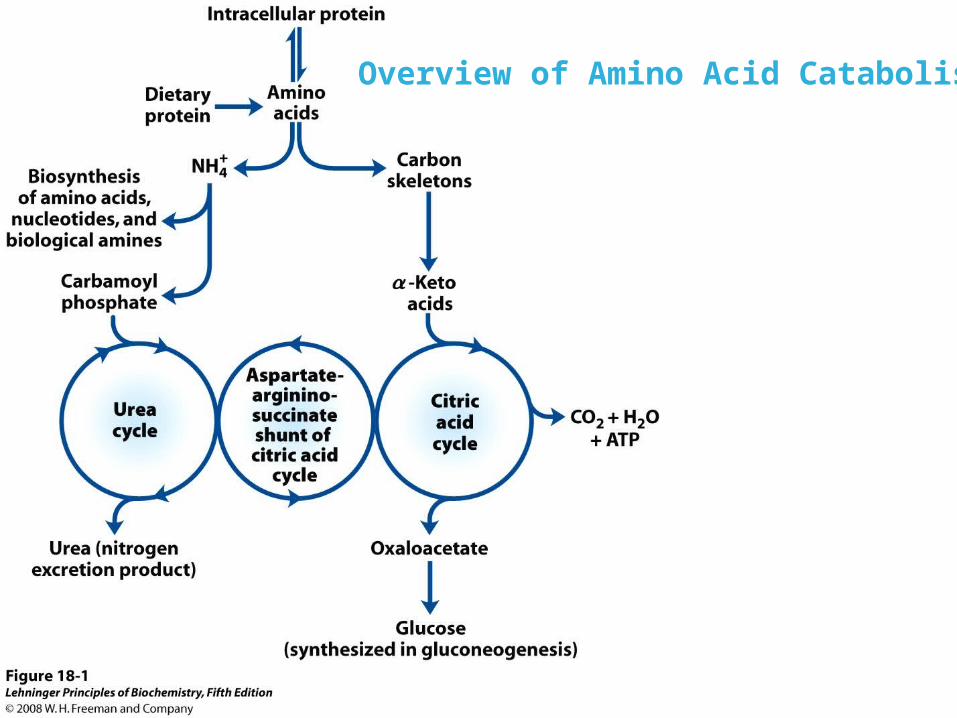
Overview of Amino Acid Catabolism

The Amino Group is Removed From All Amino Acids First

Fates of Nitrogen in Organisms• Plants conserve almost all the nitrogen• Many aquatic vertebrates release ammonia to their environment
– Passive diffusion from epithelial cells– Active transport via gills
• Many terrestrial vertebrates and sharks excrete nitrogen in the form of urea– Urea is far less toxic that ammonia– Urea has very high solubility
• Some animals, such as birds and reptiles excrete nitrogen as uric acid– Uric acid is rather insoluble– Excretion as paste allows to conserve water
• Humans and great apes excrete both urea (from amino acids) and uric acid (from purines)

Excretory Forms of Nitrogen

Enzymatic Transamination
• All aminotransferases rely on the pyridoxal phosphate cofactor
• Typically, -ketoglutarate accepts amino groups
• L-Glutamine acts as a temporary storage of nitrogen
• L-Glutamine can donate the amino group when needed for amino acid biosynthesis


Structure of Pyridoxal Phosphate and Pyridoxamine Phosphate
• Intermediate, enzyme-bound carrier of amino groups
• Aldehyde form can react reversibly with amino
groups
• Aminated form can react reversibly with carbonyl
groups


Pyridoxal Phosphate is Covalently Linked to the Enzyme
In the Resting Enzyme • The linkage is made via an nucleophilic attack of
the amino group an active-site lysine side chain
• After dehydration, a Schiff base linkage is formed
• The covalent complex is called internal aldimine because the Schiff base connects PLP to the enzyme
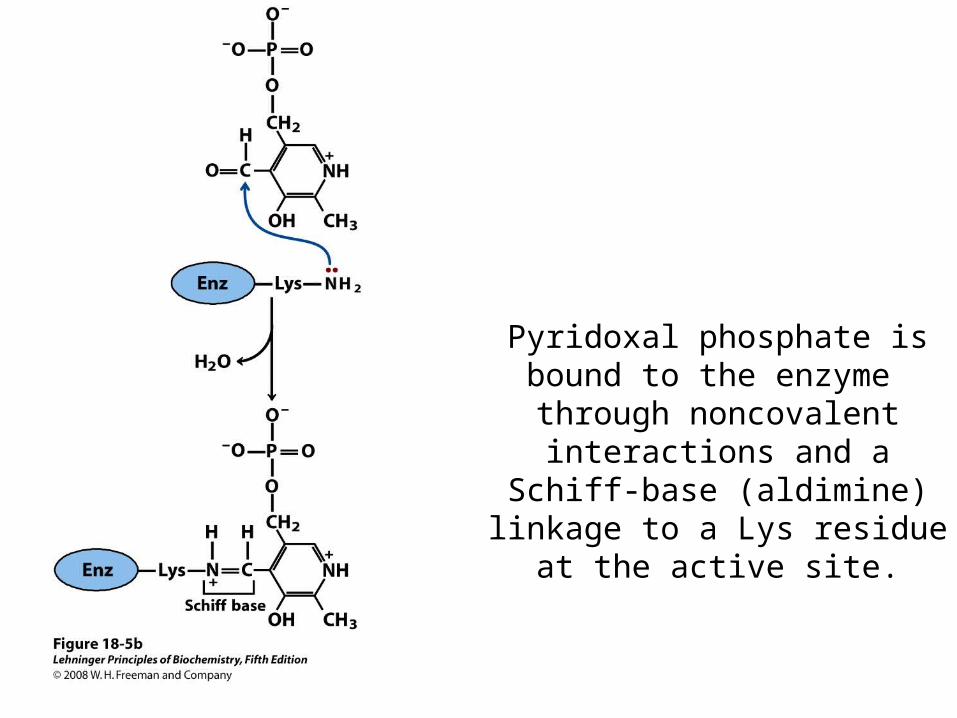
Pyridoxal phosphate is bound to the enzyme through noncovalent
interactions and a Schiff-base (aldimine) linkage to a
Lys residue at the active site.

PLP (red) bound to one of the two active sites of
the dimeric enzyme aspartate aminotransferase, a typical aminotransferase

PLP (red, with yellow phosphorus) in aldimine linkage with the side chain of
Lys258 (purple)

Chemistry of the Amino Group Removal by the Internal
Aldimine
• The external aldimine of PLP is a good electron sink, allowing removal of -hydrogen
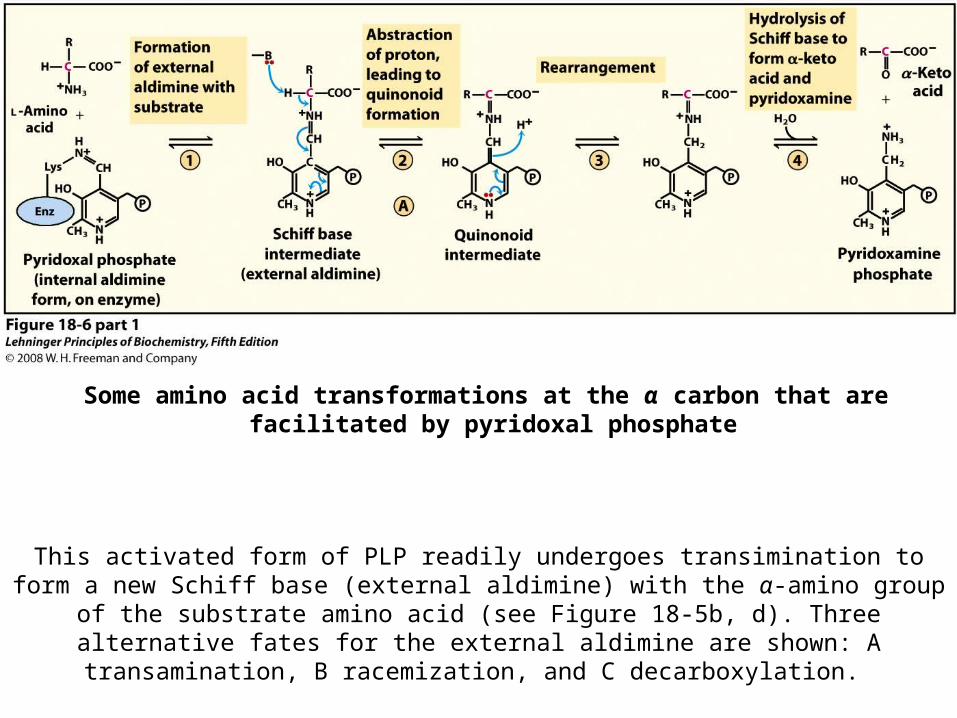
Some amino acid transformations at the α carbon that are facilitated by pyridoxal phosphate
This activated form of PLP readily undergoes transimination to form a new Schiff base (external aldimine) with the α-amino group of the substrate amino acid (see Figure 18-5b, d). Three alternative fates for the external aldimine are shown: A transamination, B
racemization, and C decarboxylation.

PLP Also Catalyzes Racemization of Amino Acids
• The external aldimine of PLP is a good electron sink, allowing removal of -hydrogen
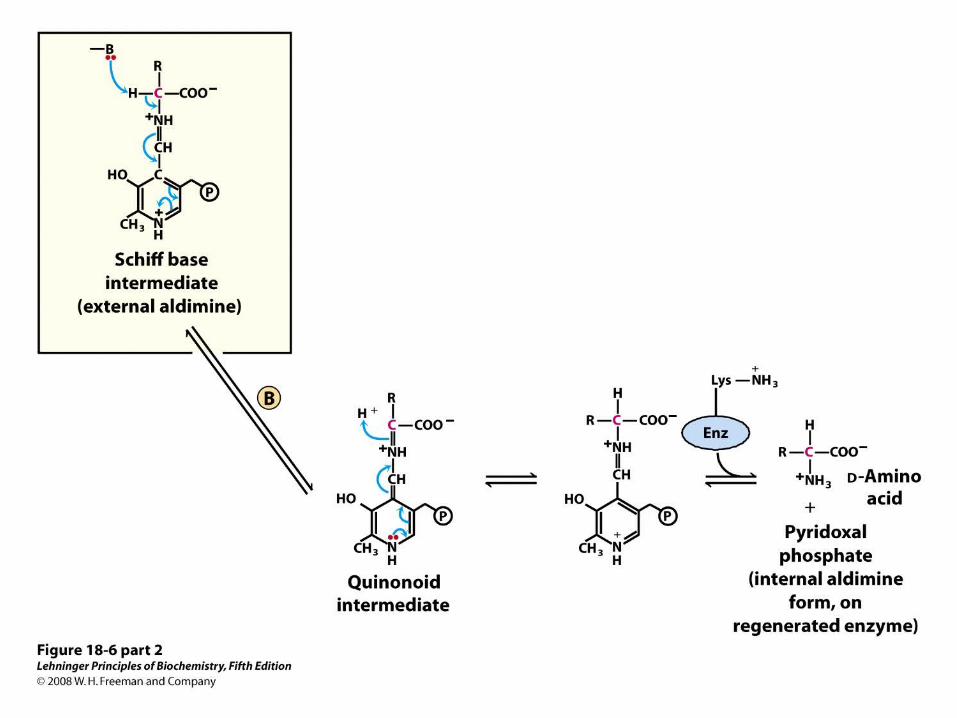

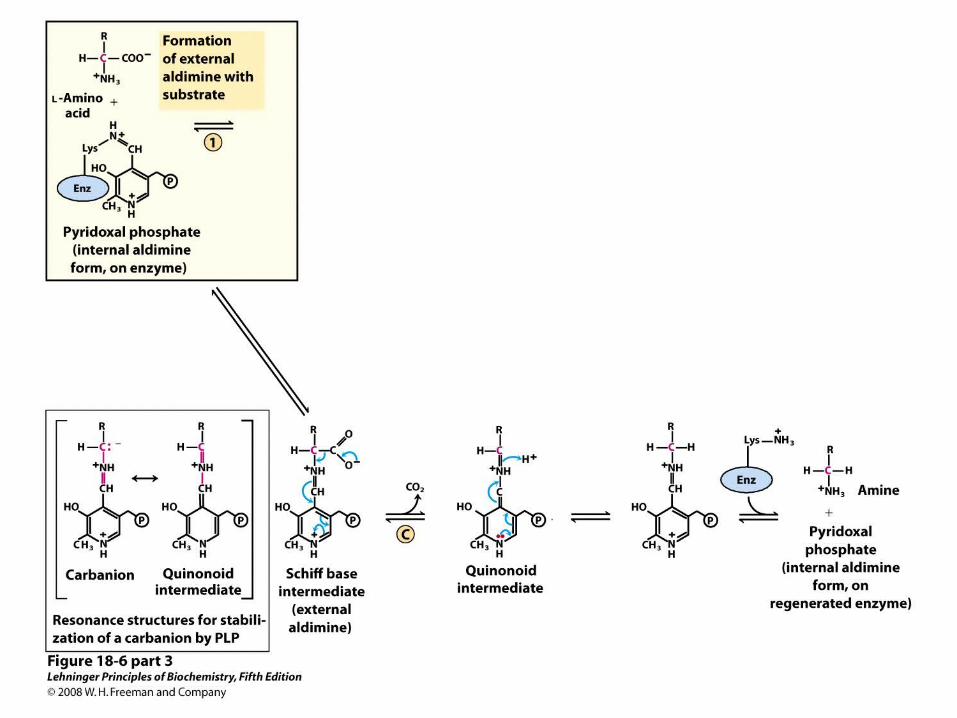
PLP Also Catalyzes Decarboxylation of Amino Acids
• The external aldimine of PLP is a good electron sink, allowing removal of -carboxylate

Ammonia in Transported in the Bloodstream Safely as
Glutamate
• Un-needed glutamine is processed in intestines, kidneys and liver
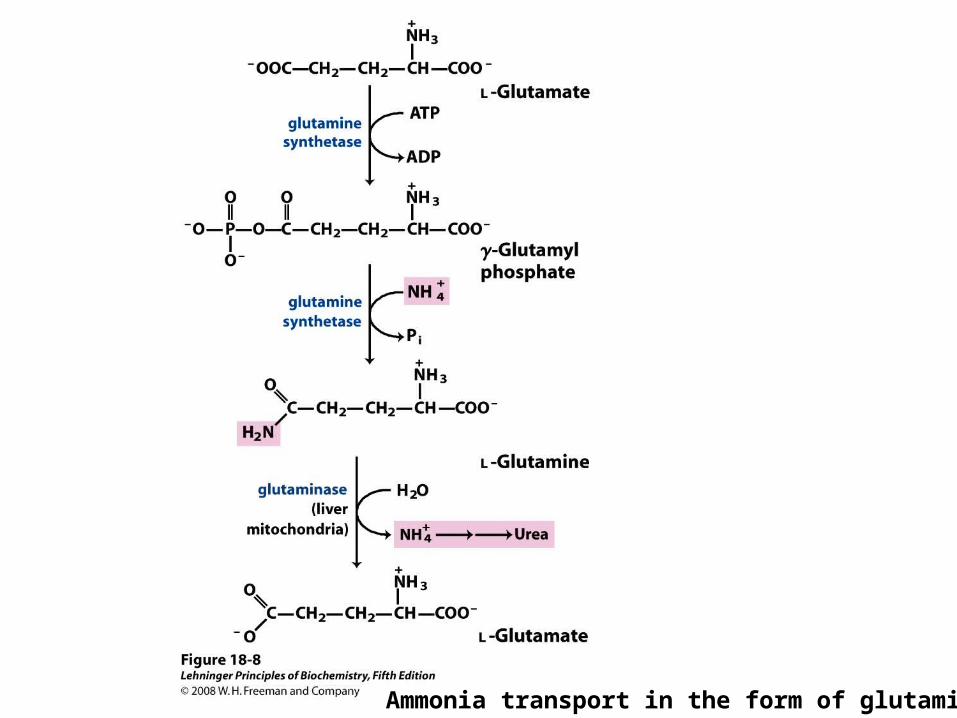
Ammonia transport in the form of glutamine.

Glutamate can Donate Ammonia to Pyruvate to Make Alanine
• Vigorously working muscles operate nearly anaerobically and rely on glycolysis for energy
• Glycolysis yields pyruvate that muscles cannot metabolize aerobically; if not eliminated lactic acid will build up
• This pyruvate can be converted to alanine for transport into liver
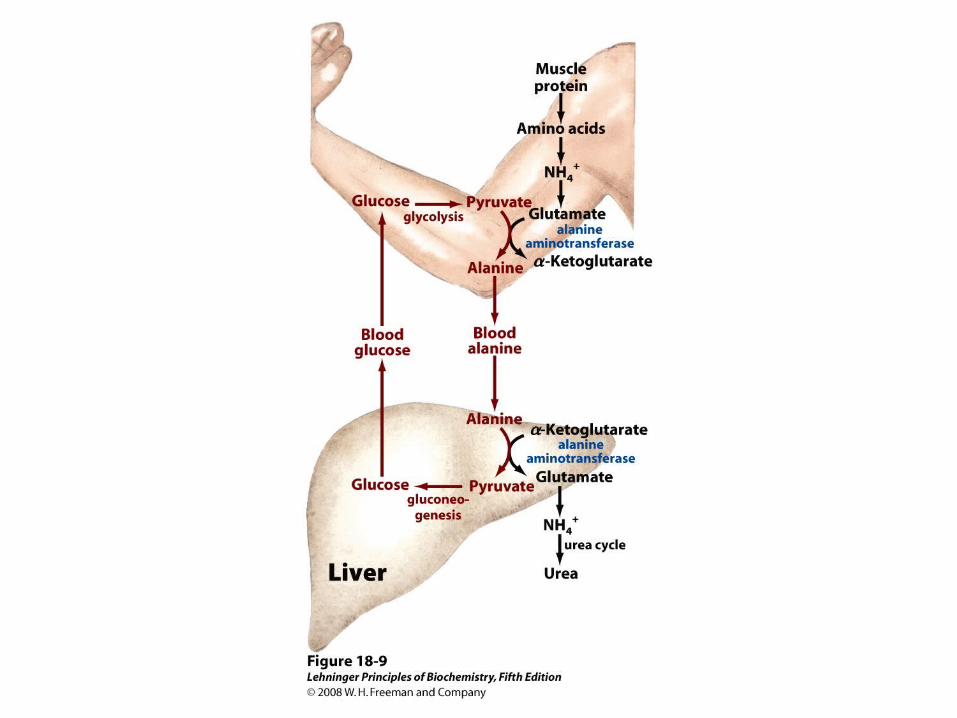

Excess Glutamate is Metabolized in the Mitochondria
of Hepatocytes

Reactions that feed amino groups into the urea cycle

The Glutamate Dehydrogenase Reaction
• Two-electron oxidation of glutamate followed by hydrolysis
• Net process is oxidative deamination of glutamate
• Occurs in mitochondrial matrix in mammals
• Can use either NAD+ or NADP+ as electron acceptor

Reaction catalyzed by glutamate dehydrogenase. The glutamate
dehydrogenase of mammalian liver has the unusual capacity to use either NAD+ or
NADP+ as cofactor. The glutamate DHs of plants and microorganisms are generally
specific for one or the other. The mammalian enzyme is allosterically regulated by GTP
and ADP.

Ammonia is Re-captured via Synthesis of Carbamoyl Phosphate
• This is the first nitrogen-acquiring reaction

Nitrogen-acquiring reactions in the synthesis of urea.
This reaction has two activation steps (1 and 3).
Nitrogen from Carbamoyl Phosphate Enters the Urea Cycle

Urea cycle and reactions that feed amino groups into the cycle
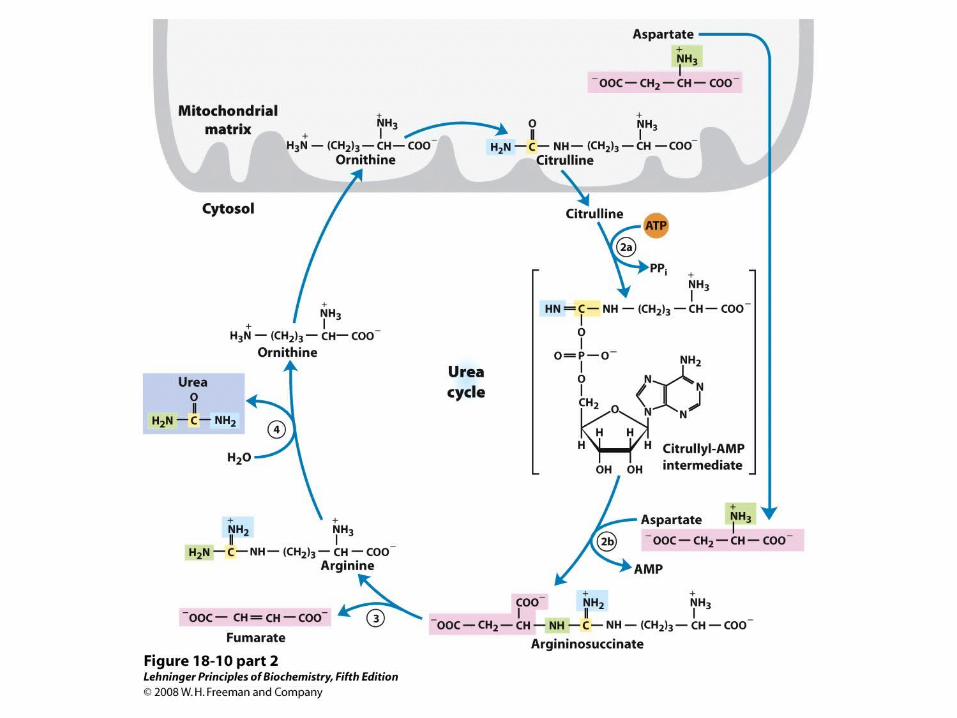

Entry of Aspartate into the Urea Cycle
• This is the second nitrogen-acquiring reaction
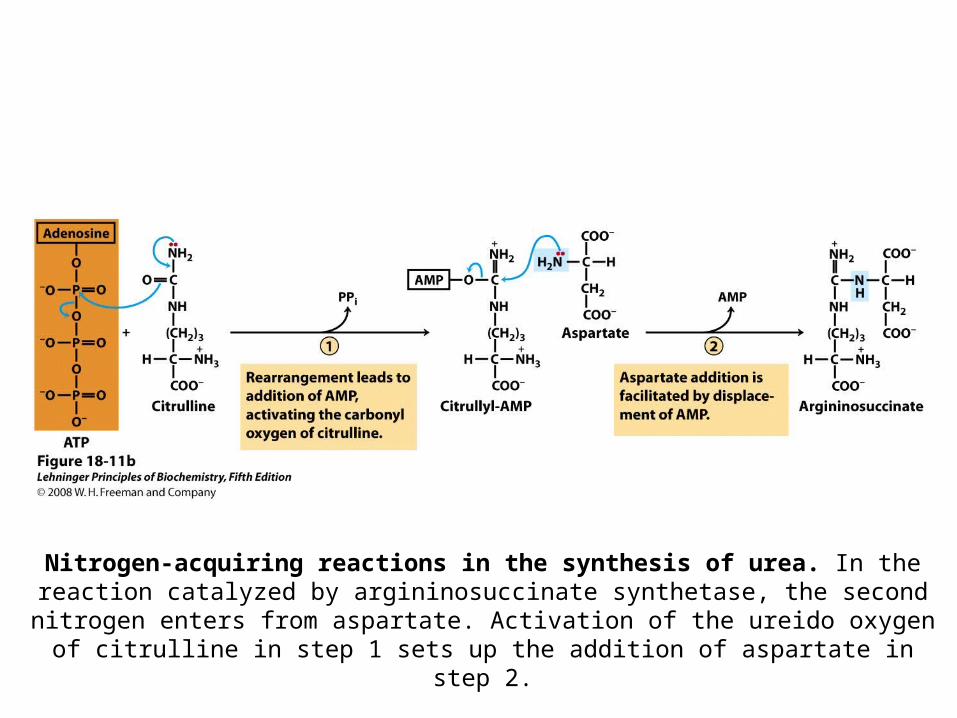
Nitrogen-acquiring reactions in the synthesis of urea. In the reaction catalyzed by argininosuccinate synthetase, the second nitrogen enters from aspartate. Activation of
the ureido oxygen of citrulline in step 1 sets up the addition of aspartate in step 2.
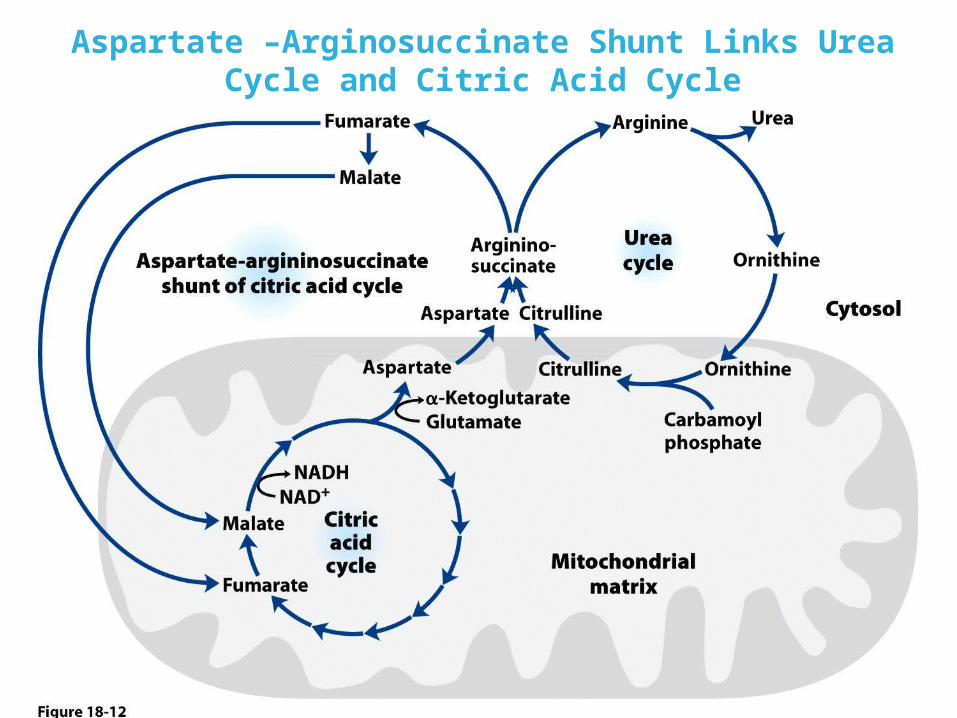
Aspartate –Arginosuccinate Shunt Links Urea Cycle and Citric Acid Cycle

Not All Amino Acids can be Synthesized in Humans
• These amino acids must be obtained as dietary protein
• Consumption of a variety of foods (including vegetarian only diets) well supplies all the essential amino acids
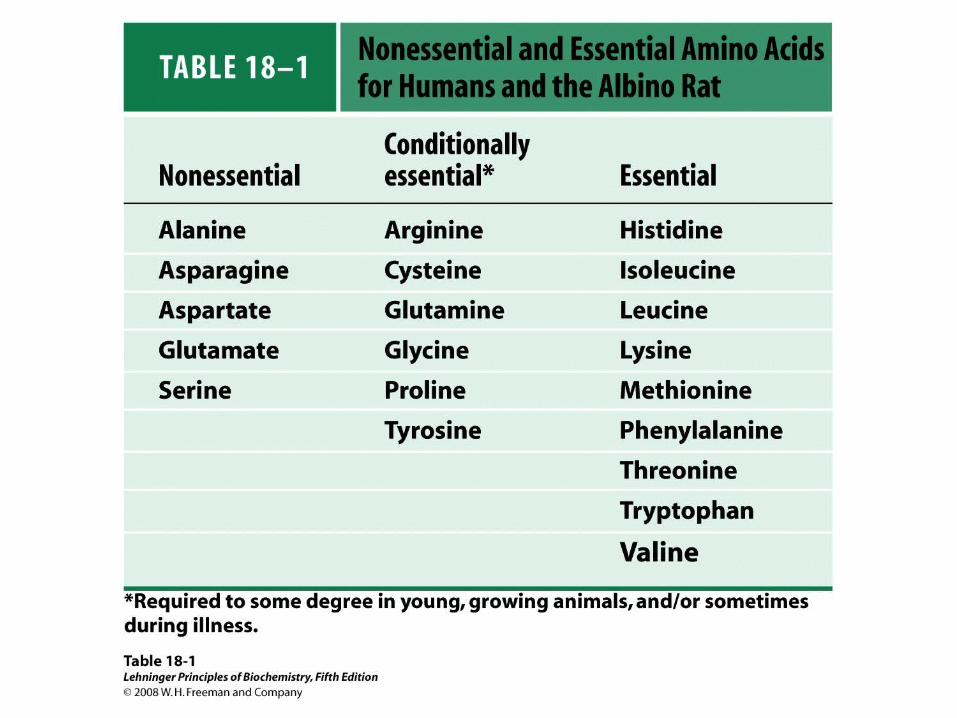

Fate of Individual Amino Acids• Seven to acetyl-CoA
– Leu, Ile, Thr, Lys, Phe, Tyr, Trp
• Six to pyruvate– Ala, Cys, Gly, Ser, Thr, Trp
• Five to -ketoglutarate– Arg, Glu, Gln, His, Pro
• Four to succinyl-CoA– Ile, Met, Thr, Val
• Two to fumarate– Phe, Tyr
• Two to oxaloacetate – Asp, Asn
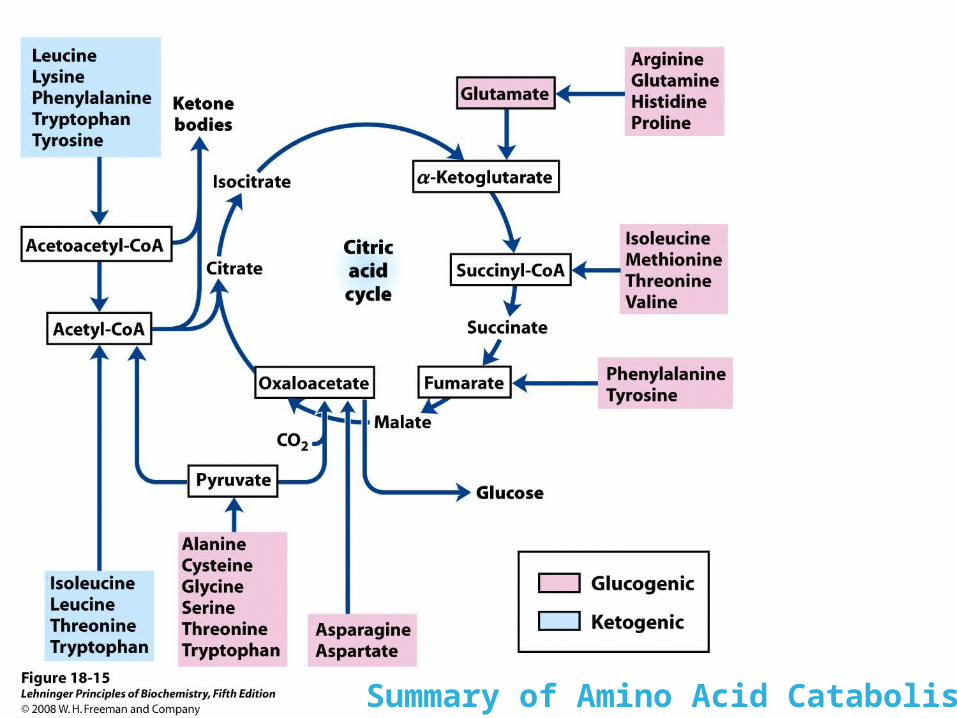
Summary of Amino Acid Catabolism

Some enzyme cofactors important in one-carbon transfer reactions

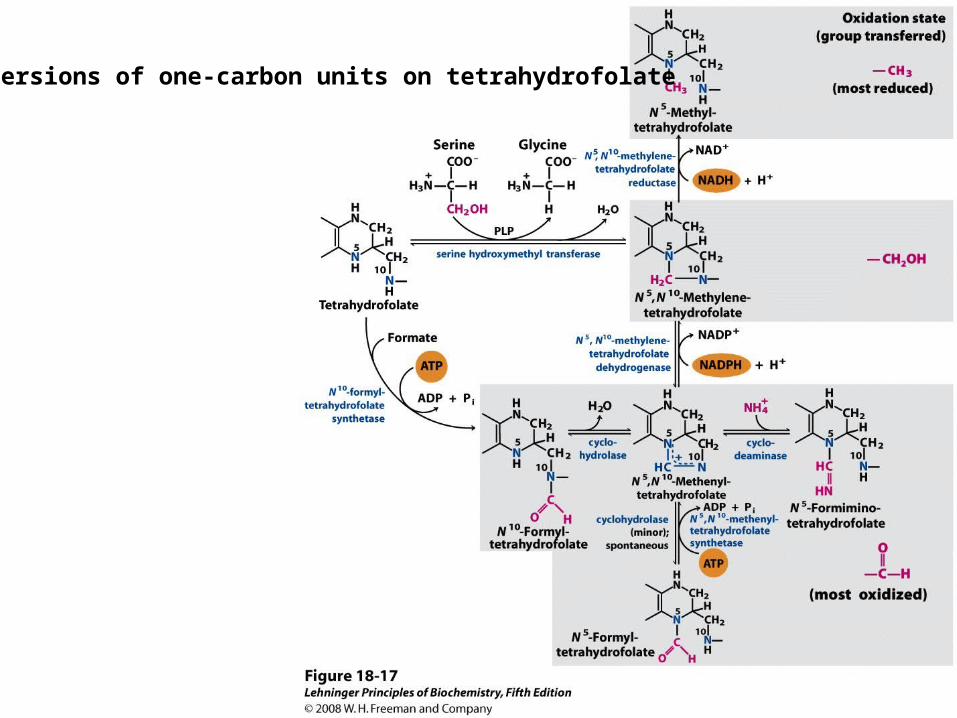
Conversions of one-carbon units on tetrahydrofolate
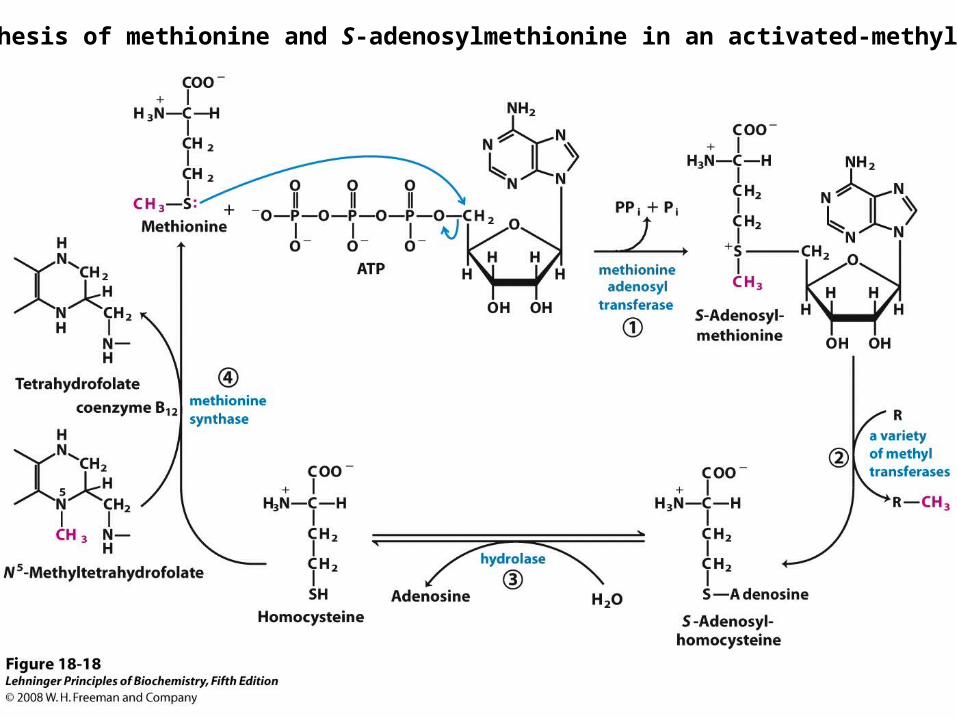
Synthesis of methionine and S-adenosylmethionine in an activated-methyl cycle
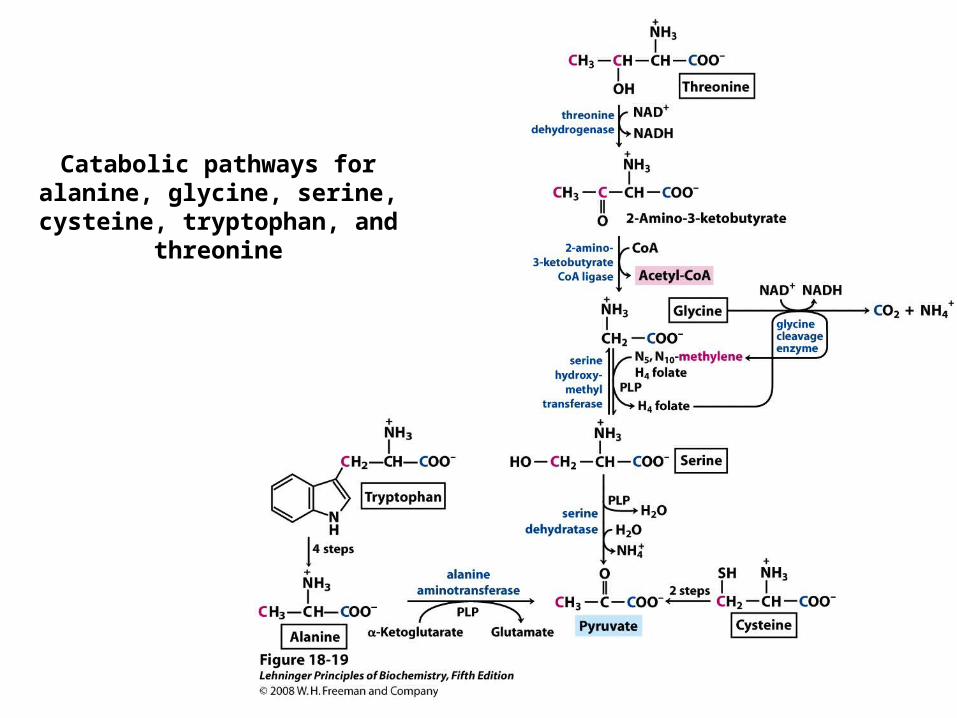
Catabolic pathways for alanine, glycine, serine, cysteine,
tryptophan, and threonine
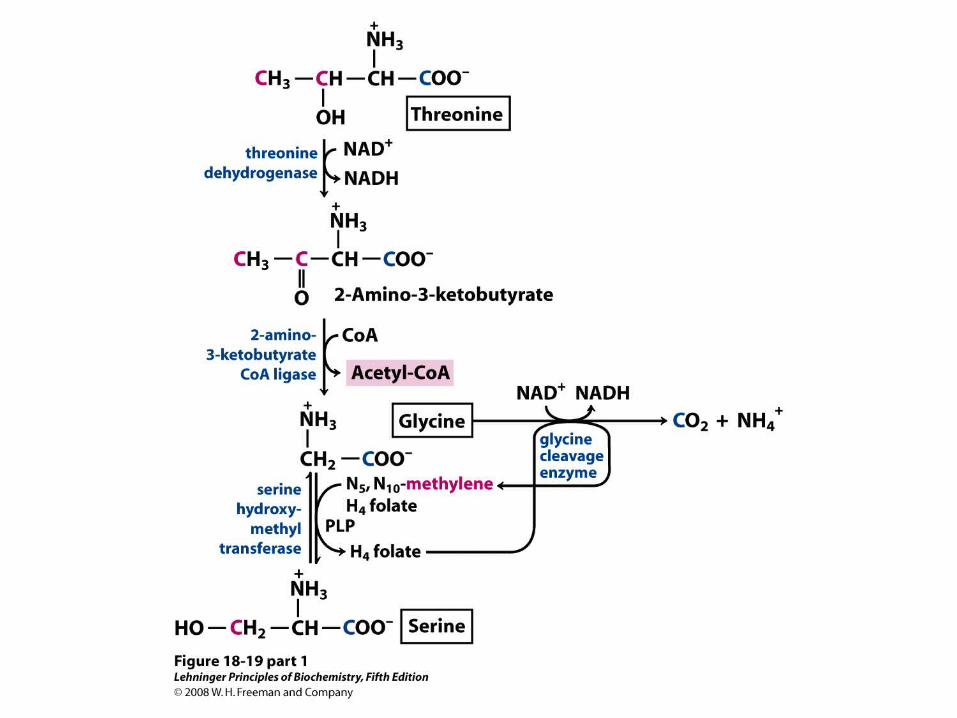


Interplay of the pyridoxal phosphate and tetrahydrofolate cofactors in serine and glycine metabolism

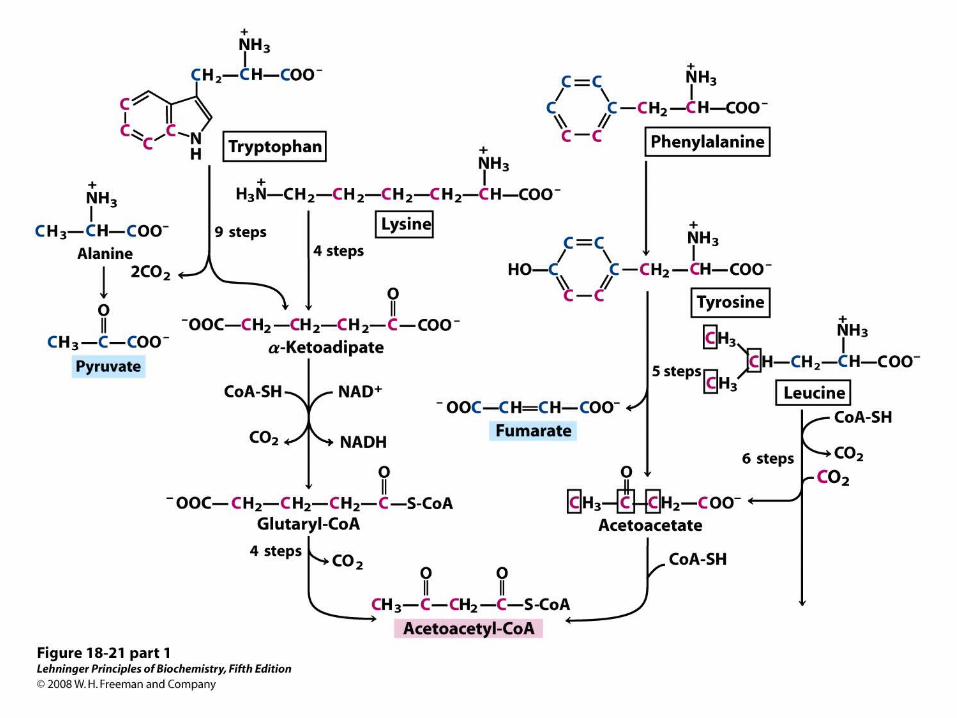
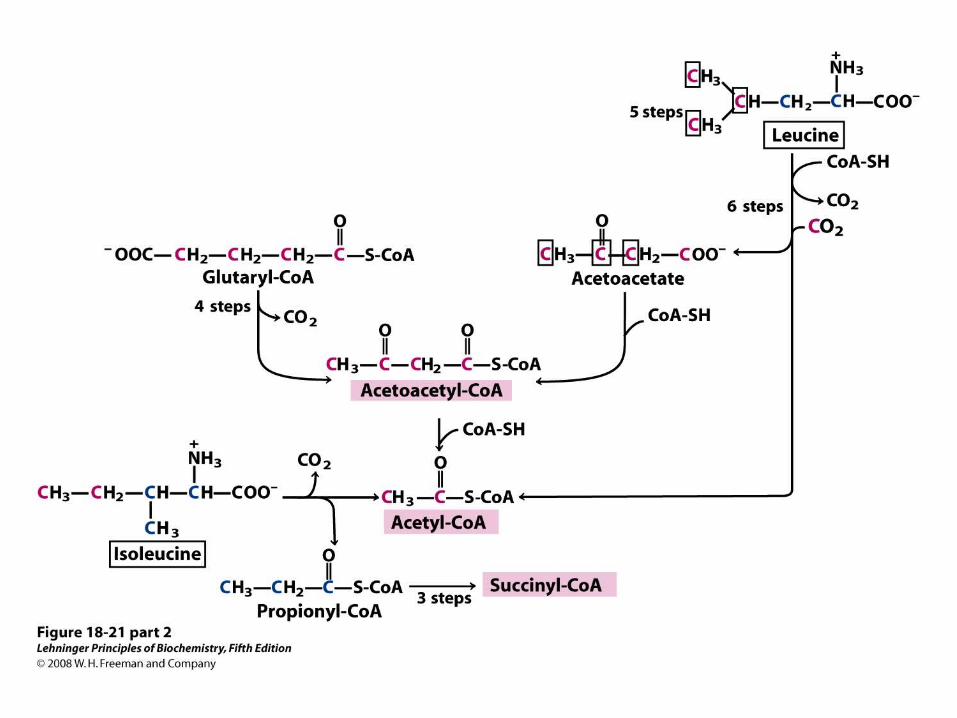
Catabolic pathways for tryptophan, lysine, phenylalanine, tyrosine, leucine, and isoleucine.
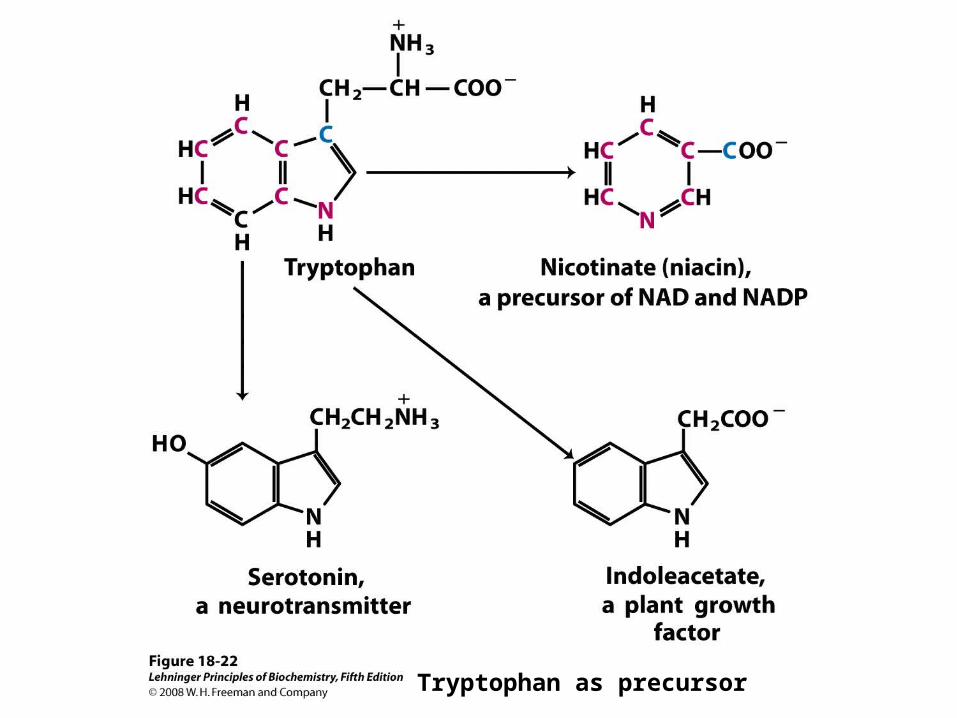
Tryptophan as precursor

Catabolic pathways for phenylalanine and tyrosine. In humans these amino acids are normally converted to acetoacetyl-CoA and fumarate. Genetic defects in many of
these enzymes cause inheritable human diseases.
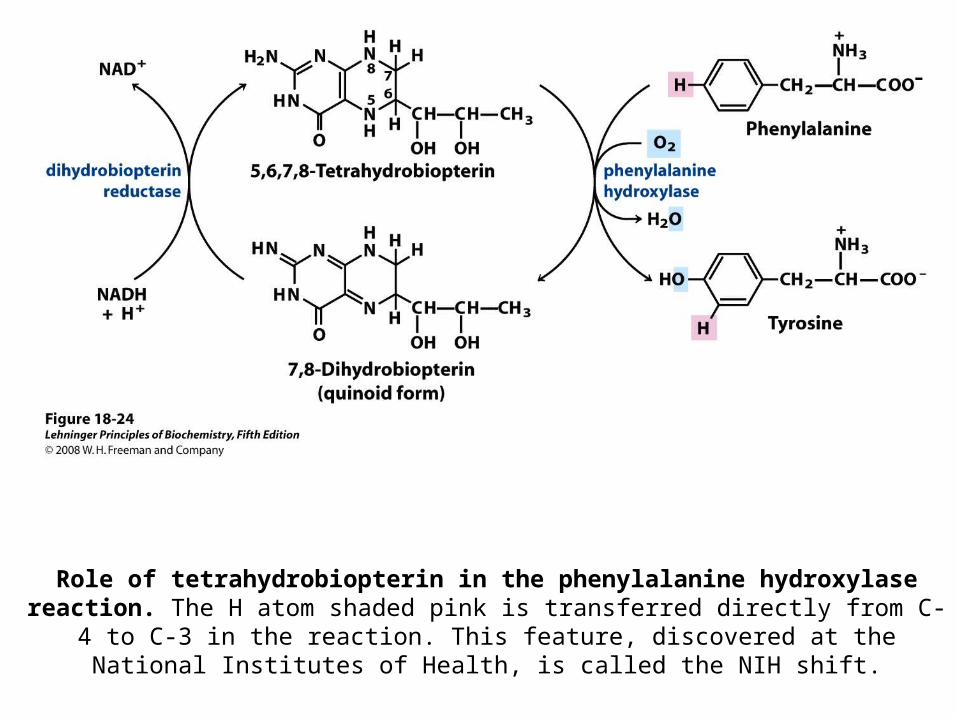
Role of tetrahydrobiopterin in the phenylalanine hydroxylase reaction. The H atom shaded pink is transferred directly from C-4 to C-3 in the reaction. This feature,
discovered at the National Institutes of Health, is called the NIH shift.

In PKU, phenylpyruvate accumulates in the
tissues, blood, and urine. The urine may also
contain phenylacetate and phenyllactate.

Catabolic pathways for arginine, histidine, glutamate, glutamine, and proline
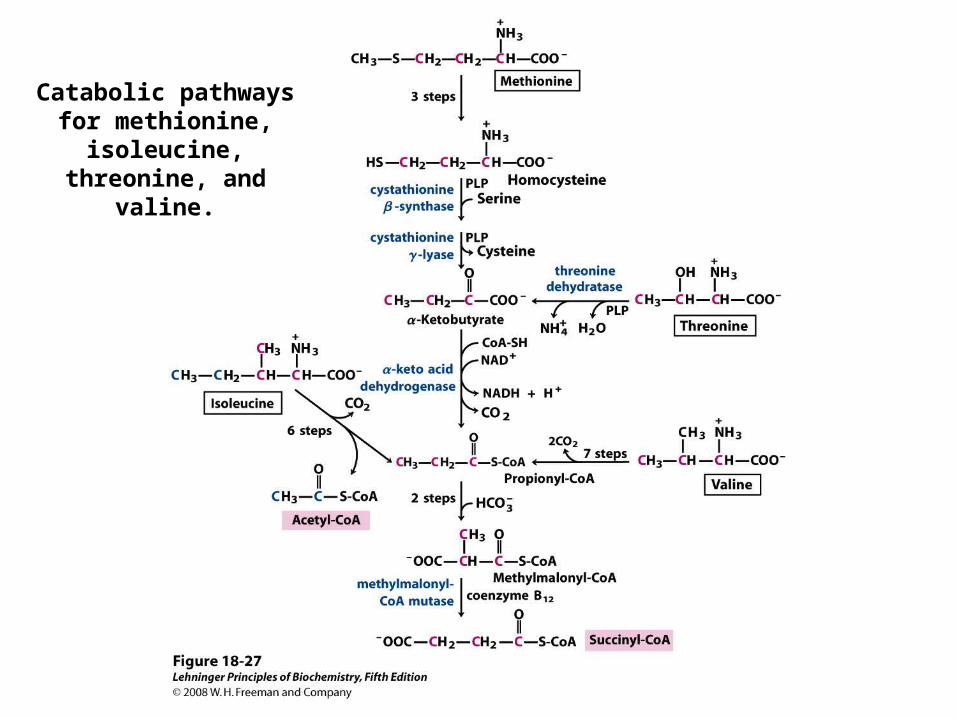
Catabolic pathways for methionine,
isoleucine, threonine, and valine.
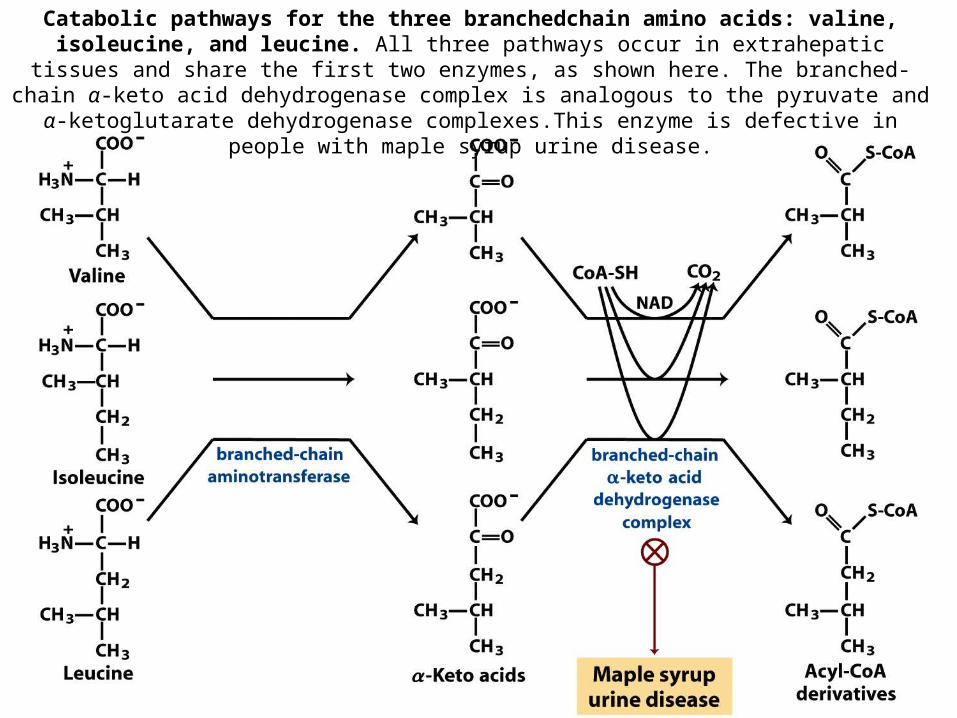
Catabolic pathways for the three branchedchain amino acids: valine, isoleucine, and leucine. All three pathways occur in extrahepatic tissues and share the first two enzymes, as
shown here. The branched-chain α-keto acid dehydrogenase complex is analogous to the pyruvate and α-ketoglutarate dehydrogenase complexes.This enzyme is defective in people with
maple syrup urine disease.

Catabolic pathway for asparagine and aspartate

Chapter 18: Summary
• Amino acids from protein are an important energy source
in carnivorous animals
• Catabolism of amino acids involves transfer of the amino
group via PLP-dependent aminotransferase to a donor
such as -ketoglutarate to yield L-glutamine
• L-glutamine can be used to synthesize new amino acids,
or it can dispose of excess nitrogen as ammonia
• In most mammals, toxic ammonia is quickly recaptured
into carbamoyl phosphate and passed into the urea cycle
In this chapter, we learned that:


![The Plastidial Glyceraldehyde-3-Phosphate Dehydrogenase Is … · The Plastidial Glyceraldehyde-3-Phosphate Dehydrogenase Is Critical for Viable Pollen Development in Arabidopsis1[W]](https://img.pdfslide.us/doc/110x75/600aa8912522092462533f3e/the-plastidial-glyceraldehyde-3-phosphate-dehydrogenase-is-the-plastidial-glyceraldehyde-3-phosphate.jpg)


























