Embed Size (px)
Citation preview

University of Groningen
Steroid transformation by Rhodococcus strains and bacterial cytochrome P450 enzymesDu Plessis-Rosloniec, Kamila Zofia
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2011
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Du Plessis-Rosloniec, K. Z. (2011). Steroid transformation by Rhodococcus strains and bacterialcytochrome P450 enzymes. Groningen: s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 09-04-2019

HOOFDSTUK 5
Samenvatting en algemene discussie
143

Bioactieve steroïden zijn biologisch en commercieel (de farmaceutische industrie)
belangrijke verbindingen. Voorlopers van deze steroïden worden voornamelijk gewonnen
uit planten (fytosterolen). Deze voorlopers kunnen zowel langs chemische als
biotechnologische weg worden omgezet in de gewenste bioactieve steroid producten.
Verschillende micro-organismen, die de gewenste enzymatische activiteiten bezitten,
worden hedendaags gebruikt voor de biotechnologische productie van commerciële
steroïden, voornamelijk soorten van de genera Mycobacterium, Rhodococcus,
Arthrobacter, Brevibacterium [23].
Rhodococci zijn in staat om een verscheidenheid aan natuurlijk voorkomende
verbindingen als koolstof- en energiebronnen te metaboliseren, waaronder ook
sterolen/steroïden. Om accumulatie van steroïden uit sterolen met behulp van
biotransformatie processen met Rhodococcus-stammen te realiseren, moeten specifieke
enzymreacties, voornamelijk de steroid ring-openbrekende enzymen, in de afbraak route
worden geblokkeerd [301] (Figuur 1).
Genetisch gemanipuleerde Rhodococcus stammen, geblokkeerd op het niveau van de
steroid ring afbraak, kunnen ook worden gebruikt als gastheer voor de expressie
van interessante steroid modificerende enzymen, zoal bijvoorbeeld cytochroom P450
monooxygenases (P450 of CYPs). Op die manier kan een verdere verbreding
van producten gesynthetiseerd met behulp van biotransformaties worden verkregen
(Figuur 1).
P450 enzymen zijn krachtige biokatalysatoren. Rhodococcus stammen coderen voor een
indrukwekkend aantal P450 enzymen, die ongetwijfeld bijdragen aan het feit dat deze
bacteriën katabole krachtpatsers zijn en in staat zijn om op een efficiënte manier een
diversiteit aan verbindingen af te breken [189].
144
Het is bekend dat vele P450 enzymen uit eukaryote organismen steroïden op
verschillende manieren kunnen omzetten. Bacteriële CYPs betrokken bij sterol afbraak
waren echter nog niet geïdentificeerd of gekarakteriseerd.
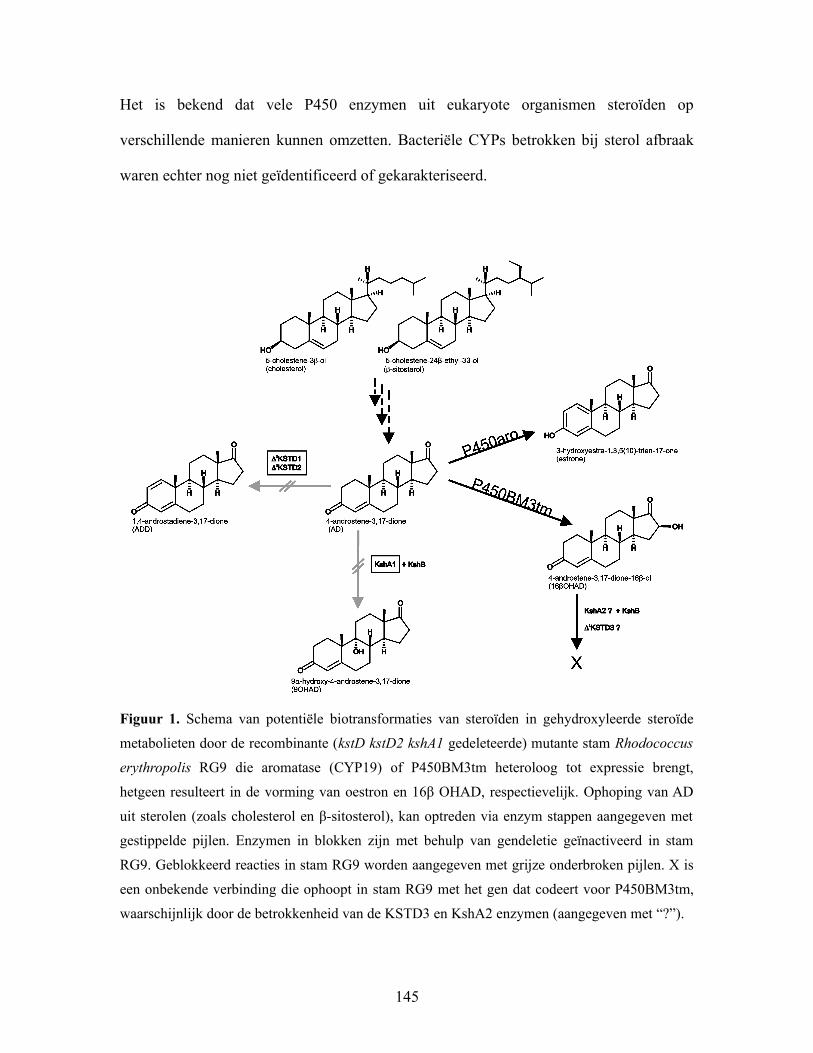
Figuur 1. Schema van potentiële biotransformaties van steroïden in gehydroxyleerde steroïde
metabolieten door de recombinante (kstD kstD2 kshA1 gedeleteerde) mutante stam Rhodococcus
erythropolis RG9 die aromatase (CYP19) of P450BM3tm heteroloog tot expressie brengt,
hetgeen resulteert in de vorming van oestron en 16β OHAD, respectievelijk. Ophoping van AD
uit sterolen (zoals cholesterol en β-sitosterol), kan optreden via enzym stappen aangegeven met
gestippelde pijlen. Enzymen in blokken zijn met behulp van gendeletie geïnactiveerd in stam
RG9. Geblokkeerd reacties in stam RG9 worden aangegeven met grijze onderbroken pijlen. X is
een onbekende verbinding die ophoopt in stam RG9 met het gen dat codeert voor P450BM3tm,
waarschijnlijk door de betrokkenheid van de KSTD3 en KshA2 enzymen (aangegeven met “?”).
145

In dit onderzoek hebben we ons gericht op een gedetailleerde karakterisatie van steroid
induceerbare enzymen, voornamelijk een aantal specifiek geselecteerde P450 enzymen
van Rhodococcus jostii RHA1, een stam waarvan de volledige genoomsequentie bekend
is. Een dergelijke aanpak is belangrijk voor het verkrijgen van meer inzicht in bacteriële
katabole routes van steroïden en opent mogelijkheden voor verdere manipulatie van deze
afbraakroutes. P450s zijn rijk vertegenwoordigd in genomen van rhodococci. Het
genoom van R. jostii RHA1 (9,7 Mb; www.rhodococcus.ca; GenBank CP000431) codeert
voor 26 CYP-enzymen. Rhodococcus erythropolis stam PR4 (6,9 Mb; GenBank
AP008957) codeert voor 16 CYPs [10], het vermoedelijke aantal CYPs in Rhodococcus
opacus stam B4 (8,8 Mb, GenBank AP011115; NC_012523, NC_006969, NC_006970,
NC_012520, NC_012521) is 27; het genoom van R. erythropolis stam SK121 (6,8 Mb,
NZ_ACNO00000000) telt 14 CYPs, terwijl er in Rhodococcus equi 103S (5,1 Mb;
http://www.sanger.ac.uk/Projects/R_equi/) 6 CYPs worden gevonden (gebasseerd op een
BLAST homologie vergelijking met gebruik van de CYP125(A14) eiwitsequentie van
stam RHA1 [250], Hoofdstuk 2). Hoewel er enkele CYP-enzymen uit Rhodococcus zijn
gekarakteriseerd [337], was er aan het begin van dit promotieonderzoek nog niets bekend
over P450 enzymen uit Rhodococcus die activiteit vertonen met sterolen of steroïden.
In dit proefschrift worden een aantal P450-enzymen uit R. jostii stam RHA1 beschreven.
Hierbij bleek dat CYP125(A14) een essentiële rol heeft in de katabole route van sterolen
(Hoofdstuk 2), en dat CYP257A1 vermoedelijk een rol heeft in sterol metabolisme
(Hoofdstuk 3). CYP257A1 en CYP125(A14) werden voor een nadere karakterisatie
geselecteerd, op basis van een analyse van het transcriptoom van R. jostii RHA1 cellen
gegroeid op cholesterol [309] (Tabel 1). De transcriptie van de CYP257A1 en
146

CYP125(A14) genen bleek het hoogst te zijn van alle CYP genen in RHA1 cellen
gegroeid op cholesterol, in vergelijking met RHA1 cellen gegroeid op pyruvaat [309].
Tabel 1. Overzicht van de 26 CYP eiwitten en hun mogelijke redox donoren, in de geannoteerde
genoomsequentie van R. jostii stam RHA1 (www.rhodococcus.ca). Toewijzing aan een CYP
familie is volgens Nelson [211] en de bijbehorende gennamen zijn overgenomen uit
(www.rhodococcus.ca). Het CYP125(A14)-enzym dat hier weergegeven wordt is het produkt van
een geherannoteerd ro04679 gen zoals beschreven in [250], Hoofdstuk 2. De verhoogde gen
expressie van de CYP genen in cellen van stam RHA1 gegroeid op cholesterol versus pyruvaat
wordt hier weergegeven als “Opregulatie op cholesterol”; "na" en "ni" staan respectievelijk voor
niet beschikbaar en niet geïdentificeerd. CYPs die in vet gedrukt worden weergegeven zijn >2-
voudig opgereguleerd op cholesterol [309]. De vier leden van de CYP125 familie zijn
aangegeven met arcering.
Het onderzoek heeft geleid tot de ontdekking van een essentiële rol voor CYP125(A14)
in de sterol afbraak (Hoofdstuk 2). CYP125(A14) blijkt een belangrijke rol te spelen als
een steroid 26-hydroxylase in de initiatie van de steroid zijketen afbraak in R. jostii
RHA1 (Hoofdstuk 2). In Hoofdstuk 3 wordt een mogelijke rol van CYP257A1 in de
afbraak van fytosterolen besproken. CYP257A1 is een vertegenwoordiger van een
147

nieuwe cytochroom P450 familie. Een fysiologische rol van CYP257A1 in sterol
metabolisme is waarschijnlijk, maar hiervoor is geen sluitend bewijs gevonden
(Hoofdstuk 3).
In Hoofdstuk 4 wordt ook de heterologe expressie van het steroid modificerend P450-
enzym P450BM3tm in R. erythropolis RG9, een kstD1 kstD2 kshA1 deletie mutant
volledig geblokkeerd in steroid ring afbraak, beschreven [312; 306] (Figuur 1).
P450BM3tm is een recombinant cytochroom P450BM3 (CYP102A1) enzym uit Bacillus
megaterium, waarin drie puntmutaties zijn aangebracht [174]. In Hoofdstuk 4 tonen we
aan dat P450BM3tm de16β-hydroxylatie van 4-androstene-3,17-dione (AD) katalyseert.
Heterologe expressie van eukaryote steroide-modificerende P450-enzymen is een
uitdaging
In het begin van het onderzoek hebben we ons gericht op de expressie van functioneel
humaan cytochroom P450 (aromatase, CYP19, P450aro) [97] in een gemuteerde
Rhodococcus stam die volledig geblokkeerd is in steroide ring afbraak (Figuur 1). CYP19
zet androgenen om in oestrogenen, in de aanwezigheid van elektronen afkomstig van
NADPH via NADPH-cytochroom P450-reductase (CPR). Interactie tussen aromatase en
CPR is van essentieel belang voor katalyse, maar het mechanisme hiervan wordt nog niet
volledig begrepen [102]. Incubatie van humaan aromatase met alternatieve redoxdonor-
eiwitten uit runderen, adrenodoxin en adrenodoxin reductase, resulteerde niet in vorming
van een product [231]. Rhodococci coderen voor meerdere redox-systemen ter
ondersteuning van P450-activiteit (zie Tabel 1 voor een overzicht van R. jostii RHA1).
Expressie van functioneel aromatase in rhodococci zou ook kunnen worden ondersteund
door een dergelijk endogeen redoxsysteem. Het gebruik van een endogeen Rhodococcus
148

redoxsysteem voor de activiteit van humaan CYP19 in Rhodococcus cellen is een
aantrekkelijke optie maar wellicht moeilijk te realiseren. Tevens hebben we getracht de
aromatase ondersteunende reductase (CPR) en het reductase domein van B. megaterium
P450BM3, dat homoloog is aan het eukaryote cytochroom P450-reductase, doch
oplosbaar, te gebruiken (zie sectie flavocytochrome P450BM3 [CYP102A1] in
Hoofdstuk 1).
De beoogde R. erythropolis gastheer stam voor CYP19 expressie, geblokkeerd in steroide
ring afbraak, bleek in bioconversies met hele cellen niet in staat oestron af te breken, het
product van de 4-androstene-3,17-dione (AD) transformatie door aromatase. Succesvolle
expressie van CYP19 in een Rhodococcus stam die AD accumuleert uit fytosterolen zou
het mogelijk maken oestron uit steroïden en sterolen te produceren (Figuur 1). Stammen
van R. erythropolis die AD van fytosterolen accumuleren zijn echter nog niet
beschikbaar. Verschillende strategieën zijn toegepast voor expressie van het humane
aromatase gen in Rhodococcus; expressie van (1) het complete cDNA van humaan
aromatase, (2) een recombinant aromatase waarin het N-terminale deel dat in het
celmembraan steekt is gedeleteerd (volgens Zhang et al. [335]), (3) een recombinant
aromatase, met een gedeleteerde N-terminus, hetgeen een stabiel enzym oplevert als het
wordt geproduceerd in E. coli [127], en (4) een codon-geoptimaliseerd aromatase gen met
en zonder deletie van het N-terminale deel dat in het celmembraan steekt.
Voor heterologe genexpressie van aromatase in Rhodoccocus hebben we gebruik gemaakt
van de induceerbare expressievector, pRESX [304]. Ook werd het aromatase gen stabiel
in het Rhodococcus genoom geplaats achter de sterke induceerbare kstD promoter [305].
Uit experimenten met expressie van een β-lactamase reporter gen bleek echter dat het
plasmide-afgeleide pRESX expressie systeem een efficiëntere methode voor heterologe
149

genexpressie in Rhodococcus was dan expressie vanaf het genoom (data niet
weergegeven). Expressie van aromatase in Rhodococcus werd echter niet gevonden. RT-
PCR en Northern blot hybridisatie-analyse gaven aan dat er geen mRNA werd
geproduceerd, hetgeen waarschijnlijk te wijten is aan een te zwak expressie systeem. Met
de hulp van dr. Barbara van Vugt-Lussenburg, Vrije Universiteit Amsterdam, hebben we
vervolgens geprobeerd expressie te realiseren van eukaryote P4501A2 (CYP1A2), een
humaan cytochroom P450 dat de oxidatie van oestron en 17β-estradiol, alsook dat van
bepaalde geneesmiddelen, katalyseert [66, 163]. P4501A2 werd eerst in actieve vorm
geproduceerd in E. coli samen met de bijbehorende reductase (blijkend uit CO-verschil
spectra en een cytochroom c reductie analyse, respectievelijk). Hierbij werd gebruik
gemaakt van een bi-cistronisch expressie plasmide (een gift van P. Guengerich,
Vanderbilt University School of Medicine, USA), een goed-gedocumenteerde en
efficiente methode voor het verkrijgen van het actieve en functionele P450 enzym [82,
227]. Met behulp van dit modelsysteem hebben we vervolgens geprobeerd essentiële
omstandigheden voor P450-expressie te vinden die we vervolgens konden toepassen op
ons werk met rhodococci. Daarna vervingen we het CYP1A2 gen op dit expressie-
plasmide door humaan CYP19, met als doel om actief aromatase-enzym te produceren op
dezelfde wijze als voor CYP1A2, namenlijk eerst met E. coli. Helaas werd er geen actief
aromatase waargenomen. Duidelijk is dat elk P450-enzym individueel moet worden
geoptimaliseerd voor over-expressie [334]. Ondanks uitgebreid onderzoek zijn helaas ook
al onze pogingen om actief CYP19 of CYP1A2 tot expressie te brengen in recombinante
R. erythropolis stammen mislukt, ook wanneer we gebruik maakten van pTip-QC1 [205],
een sterker expressie plasmide dan pRESX.
150

De negatieve resultaten met de heterologe expressie van humane P450 eiwitten
in R. erythropolis worden niet in beschreven in de experimentele hoofdstukken van dit
proefschrift. Vervolgens hebben we ons onderzoek gericht op bacteriële P450BM3tm
productie in mutante Rhodococcus stammen.
Heterologe expressie van bacterieel cytochroom P450BM3tm in Rhodococcus
Hoofdstuk 4 beschrijft een analyse van de stereo- en regio-selectiviteit van P450BM3tm
op AD ende heterologe expressie van P450BM3tm in de R. erythropolis RG9 mutant
geblokkeerd in AD afbraak (Figuur 1). Het P450BM3tm enzym (aanwezig in E. coli-cel-
vrije extracten) zette AD om in 4-androstene-3,17-dione-16β-ol (16β-hydroxy-AD). Ook
de biotransformatie van AD met cellen van R. erythropolis stam RG9BM3tm was zeer
efficiënt. AD conversie vond niet plaats met stam RG9 die alleen de lege expressie vector
bevatte, dus zonder het gen coderend voor P450BM3tm. Verrassend genoeg werd er geen
16β-hydroxy-AD geproduceerd door celkweken van RG9BM3tm tijdens AD
bioconversie, maar was er een tijdelijke ophoping te zien van een onbekende steroïde.
Het feit dat geen 16β-hydroxy-AD accumulatie waargenomen werd in stam RG9BM3tm
weerspiegelt waarschijnlijk een snel metabolisme van deze verbinding door de katabole
enzymen die nog aanwezig zijn, bv. 3-ketosteroid Δ1-dehydrogenase KSTD3 [141] en 3-
ketosteroid 9α-hydroxylase KshA2 [304] (Fig. 1). In tegenstelling tot AD induceert 16β-
hydroxy-AD blijkbaar deze katabole enzymen in stam RG9BM3tm. Verdere metabole
engineering van stam RG9BM3tm op moleculair niveau, voornamelijk gericht op het
inactiveren van KSTD3 en KshA2 activiteiten, zal zeer waarschijnlijk resulteren in een
gemuteerde stam die in staat zal zijn 16β-hydroxy-AD te accumuleren uit AD.
151

Rhodococcus en Escherichia coli stammen als gastheer voor expressie van CYP
genen
E. coli is een goed bestudeerde bacterie en een bewezen goede gastheer voor eiwit
expressie in het algemeen en ook voor P450 enzymes in het bijzonder [332, 78, 325,
334]. E. coli-stammen coderen niet voor P450-genen, waardoor ze als zeer nuttige
gastheren worden beschouwd voor de heterologe productie en analyse van CYP eiwitten,
en de activiteiten van CYP eiwitten (bv. Arisawa et al. [10]). De afwezigheid van P450
genen in E. coli stammen zorgt dus voor een schone achtergrond, maar heeft als nadeel
dat medium additieven nodig zijn, zoals δ-aminolevuline zuur, voor productie van actieve
P450 eiwitten. Rhodococcus stammen zijn echter natuurlijke producenten van een
verscheidenheid aan P450 enzymen. Het gebruik van rhodococci als gastheer voor de
expressie van CYPs heeft als voordeel dat geen medium additieven nodig zijn. Recent
zijn er echter ook recombinante E. coli stammen geconstrueerd voor de heterologe
productie van actieve P450 enzymen die geen δ-aminolevuline zuur in het medium nodig
hebben [90].
Uit onze resultaten blijkt dat Rhodococcus stammen, zoals R. erythropolis, goede
gastheren zijn voor de (heterologe) productie van actieve cytochroom P450-enzymen,
zoals de R. jostii enzymen CYP125(A14) [250] (Hoofdstuk 2, expressie niveaus van 5
μM, zonder media supplementatie), CYP257A1 (0.5 and 5 μM, respectievelijk zonder en
met media supplementatie, Hoofdstuk 3) en B. megaterium P450BM3tm (0.2 μM, zonder
media supplementatie, Hoofdstuk 4). In het geval van CYP257A1 productie door R.
erythropolis, maar niet van P450BM3tm-productie, was de enzymopbrengst tien keer
hoger na medium supplementatie met δ-aminolevuline zuur, thiamine, ijzer (III) chloride
en sporenelementen. De P450-opbrengst in rhodococci bleek over het algemeen lager dan
152

in E. coli; de verdere verbetering van gen expressie systemen in Rhodococcus-stammen
blijft dus een uitdagend doel.
Rhodococcus stammen zijn van nature in staat om een breed scala van hydrofobe
substraten af te breken, waaronder sterolen/steroïden, en bezitten actieve systemen voor
hun opname in de cel [194]. E. coli stammen die P450 eiwitten exporteren in de
periplasmatische ruimte zijn beschreven, waardoor de biotransformatie van hydrofobe
substraten in celkweken mogelijk wordt [126]. Beide eigenschappen, endogene δ-
aminolevuline zuur synthese voor productie van actieve CYP eiwitten, en het transport
van hydrofobe verbindingen, zijn van nature reeds aanwezig in rhodococci. Dit moet
echter nog gerealiseerd worden in E coli.
E. coli stammen zijn o.h.a. snelle groeiers maar bezitten geen katabole activiteiten op
sterol of steroid substraten. Het ontbreekt E. coli aan alle enzymen voor sterol en steroid
conversie. Daarom is het een grote uitdaging om E. coli stammen te construeren die
sterolen kunnen omzetten in bioactieve steroïden. Het is waarschijnlijk gemakkelijker om
recombinante Rhodococcus stammen te construeren voor de productie van interessante
steroïden. Deze laatste benadering is echter ook uitdagend, omdat steroid modificerende
enzymen van interessante Rhodococcus stammen en hun regulatie voor de expressie van
genen nog grondig moet worden geanalyseerd. In ons onderzoek aan Rhodococcus
stammen worden echter snelle vorderingen gemaakt, resulterend in ontrafeling van hun
complexe repertoire van steroïde modificerende enzymen en de regulatie systemen
betrokken bij hun steroid katabole routes [142, 194, 233, 250, 309]. Uiteindelijk hangt de
keuze van de optimale bacteriële gastheer voor CYP expressie van vele factoren af,
bijvoorbeeld de specifieke CYP toepassing, de gewenste opbrengst van CYP eiwit, totale
proces kosten, gebruiksgemak en productopbrengsten.
153

CYP125(A14) is een steroid 26-hydroxylase
Hoofdstuk 2 beschrijft de kenmerken van CYP125(A14) uit R. jostii stam RHA1
(CYP125RHA1), een microbieël enzym verantwoordelijk voor het initiëren van de sterol
zijketen afbraak. Stam RHA1Δcyp125, met een gedeleteerd CYP125RHA1 gene, was niet
in staat om te groeien op 3-hydroxy-sterolen met een relatief lange alifatische zijketen,
bijvoorbeeld cholesterol. Stam RHA1Δcyp125 groeide nog wel op 5α-cholestane-3β-ol
en 3-oxo-steroiden, zoals bijvoorbeeld 4-cholestene-3-one, vergelijkbaar aan de RHA1
stam. Interessant is dat het gezuiverde CYP125RHA1 eiwit niet alleen cholesterol bindt,
maar ook 5α-cholestane-3β-ol and 4-cholestene-3-one, op een wijze die typisch is voor
P450-substraten. Een cyp125 disruptie mutant van R. rhodochrous RG32
(RG32Ωcyp125) is ook volledig geblokkeerd in cholesterol-zijketen afbraak, maar niet in
de afbraak van 5-cholestene-26-carboxylzuur-3β-ol. 5-Cholestene-26-carboxylzuur-3β-ol
is een verwacht intermediair van de cholesterol-zijketen afbraak. Verwacht wordt dat
CYP125RG32 ook de steroid 26-hydroxyleringsreactie katalyseert. Terwijl RHA1Δcyp125
volledig geblokkeerd was in de cholesterol afbraak, was stam RG32Ωcyp125 nog in staat
om cholesterol om te zetten in 3-keto-Δ4 en 3-keto-Δ1,4 intermediairen, hetgeen aangeeft
dat ring-oxidatie in deze stam niet volledig geblokkeerd is. De sterol afbraak routes van
R. rhodochrous RG32 en R. jostii RHA1 zijn duidelijk verschillend. De resultaten tonen
aan dat CYP125RHA1 essentieel is voor de eerste stap in het cholesterol katabolisme in R.
jostii RHA1, waaruit blijkt dat cholesterol afbraak wordt geïnitieerd door een aanval van
CYP125RHA1 op de sterol zijketen. CYP125RG32 initieert echter niet de cholesterol afbraak
in R. rhodochrous RG32; andere enzymen hebben blijkbaar deze functie. In lijn met deze
bevindingen werd in stam RHA1 geen 3β-hydroxysteroid dehydrogenase (3βHSD)
activiteit met cholesterol waargenomen. 3βHSD activiteit werd wel waargenomen in stam
154

RG32. Ook komen de genen die coderen voor cholesterol oxidases in stam RHA1 niet tot
expressie bij groei op cholesterol [309]. 3βHSD oxidase en cholesterol zijn de enzymen
die zeer waarschijnlijk verantwoordelijk zijn voor het initiëren van de cholesterol afbraak
in stam RG32 (zie ook Hoofdstuk 1, Fig. 2).
Onze suggestie dat CYP125RHA1, CYP125RG32 en de orthologe CYP125A van
Mycobacterium tuberculosis H37Rv (CYP125H37Rv) een vergelijkbare rol in sterol zijketen
afbraak hebben (Hoofdstuk 2), is inmiddels bevestigd door Capyk et al. [27]. Zij toonden
aan dat CYP125H37Rv cholesterol hydroxyleert op koolstofatoom C26, C27, of beide.
Recent werd hydroxylatie van cholesterol op koolstofatoom C27 door CYP125H37Rv
aangetoond [188], ondersteund door gegevens van een kristalstructuur van dit eiwit met
gebonden cholesterol. CYP125RHA1 en CYP125H37Rv zijn waarschijnlijk orthologen gezien
hun overeenkomsten in aminozuursequentie (62% identiteit) [309]. Daarom is het
waarschijnlijk dat ook CYP125RHA1 steroid 27-hydroxylering kan katalyseren. Ter
bevestiging van de exacte activiteit van CYP125RHA1 zouden de E. coli redox-systemen
[188], of het KshB enzym uit stam RHA1 gebruikt kunnen worden analoog aan
experimenten van Capyk et al. [27].
CYP125A enzymen lijken op grote schaal aanwezig te zijn binnen de familie van de
actinobacteria [188] en deze eiwitten zijn daarom interessante targets voor evolutionaire
studies (zie Lamb et al. [155]). De eiwitsequentie-identiteit van alle CYP125A die
voorkomen in Rhodococcus stammen waarvan de complete genoom sequentie bekend is
(zie hierboven), is tot 97%, hetgeen het belang van dit cytochroom P450 enzym aangeeft.
CYP125A van M. tuberculosis H37Rv bleek, samen met vijf naburige genen (het igr
operon), tevens een rol te spelen in de pathogeniciteit van deze bacterie. De igr genen
spelen een rol in de intracellulaire groei van M. tuberculosis H37Rv in zowel macrofagen
155

als in muizen [29]. CYP125H37Rv is bestand tegen reactieve stikstofmonoxide, hetgeen al
aangeeft dat de enzym activiteit zal overleven in de macrofaag-achtige omgeving [223].
Samenvattend, de geconserveerde CYP125A enzymen spelen een rol in het sterol
metabolisme en in de pathogeniciteit van M. tuberculosis. Het vaststellen van de rol van
CYP125A in R. equi, een andere ziekteverwekker uit de familie van de actinobacteria,
zou kunnen bijdragen aan het verkrijgen van meer begrip van de virulentie van deze
pathogene bacterie.
CYP257A1 en zijn mogelijke rol in de β-sitosterol afbraak
CYP257A1 uit R. jostii RHA1 katalyseert de N-demethylering van dextromethorfan
(DXM), blijkbaar met steun van donor redox-systemen van E. coli (Hoofdstuk 3). De
fysiologische rol van CYP257A1 in R. jostii RHA1 is nog onduidelijk. Aangezien de
expressie van het CYP257A1 gen sterk verhoogd is in cellen gekweekt op cholesterol
(Tabel 1), wordt verondersteld dat CYP257A1 een katabole enzym codeert dat betrokken
is bij het cholesterol metabolisme. Er werd echter geen CYP257A1 activiteit
waargenomen met cholesterol, of met tussenproducten van de cholesterol afbraakroute als
substraat. Bindingsstudies toonden de complexe aard aan van de affiniteit van
CYP257A1 voor cholesterol, lanosterol, β-sitosterol en DXM. β-Sitosterol induceerde
spectrale veranderingen in het CYP257A1 enzyme die karakteristiek zijn voor een
substraat. Activiteitsbepalingen en de karakterisering van een CYP257A1 disruptie
mutant van R. jostii RHA1 gaven echter geen nader bewijs dat β-sitosterol een substraat
is voor CYP257A1, noch dat CYP257A1 betrokken is bij β-sitosterol afbraak. Meer
analyses zullen nodig zijn voor het begrijpen van de interactie van β-sitosterol met
CYP257A1. Hierbij zou men kunnen denken aan incubaties van het enzym met gelabeld
156

β-sitosterol gevolgd door product analyse met een methode gekoppeld aan massa
spectrometrie om te zien of het eiwit inderdaad sterolen omzet. We postuleren dat
CYP257A1 een mogelijke fysiologische rol heeft in sterol afbraak en een nog niet
eïdentificeerde demethylering stap in de afbraak route van het C24 vertakte sterol β-
sitosterol katalyseert.
Sequentie analyse van CYP257A1 suggereerde dat het enzym een brede specificiteit zou
kunnen hebben, weerspiegeld door zijn activiteit met DXM, een onnatuurlijk substraat.
CYP257A1 van R. jostii RHA1 is uniek en heeft, voor zover bekend uit genoom
sequenties, geen orthologen in andere Rhodococcus stammen, of in andere actinobacteria.
CYP257A1 vertegenwoordigt een nieuwe familie van cytochroom P450-enzymen
(Hoofdstuk 3).
Andere CYPs uit R. jostii stam RHA1 die mogelijk steroiden omzetten
Kennis over steroid modificerende enzymen, ook die van de CYP-enzymen, vergroot ons
begrip van het steroïd metabolisme in Rhodococcus en maakt engineering van
Rhodococcus stammen voor toepassing in steroïde synthese processen mogelijk. Het
genoom van R. jostii stam RHA1 codeert voor 26 CYPs (Tabel 1) (www.rhodococcus.ca).
CYP125(A14), die betrokken is bij de sterol zijketen afbraak, en 20 andere CYPs, liggen
op het chromosoom (7,8 Mb [189]), terwijl vijf andere CYPs plasmide-gecodeerd zijn
(Tabel 1). Hiervan zijn er drie gelegen op pRHL3 (0,33 Mb), inclusief CYP257A1. Twee
CYPs zijn te vinden op pRHL1 (1,1 Mb) (Tabel 1). Plasmide pRHL2 (0.44Mb) bevat
geen CYP genen. Vier van deze 26 CYPs waren meer dan 2-voudig opgereguleerd in
RHA1 cellen gekweekt op cholesterol [309], te weten CY257A1, CYP125(A14),
CYP116C2 en CYP102B3 (Tabel 1). Deze spelen dus mogelijk een rol in de afbraak van
157

sterolen en steroïden, zoals hieronder wordt besproken. R. jostii RHA1 bevat meerdere
CYP genen die coderen voor eiwitten die gelijkenis vertonen met gekarakteriseerde
enzymen van andere bacteriën met activiteiten op steroïden.
De karakterisatie van CYP125(A14) en CYP257A1, de hoogst opgereguleerde CYPs in
stam RHA1 gegroeid op cholesterol, zijn beschreven in respectievelijk de Hoofdstukken
2 en 3 van dit proefschrift. CYP125(A14) is het enige lid van de CYP125 familie in stam
RHA1 die tot expressie komt bij groei op cholesterol (Tabel 1).
De fysiologische rol van de drie andere CYP125 familieleden moeten nog worden
geïdentificeerd. Opvallend is dat CYP125B1 nabij CYP123A5 en CYP51B1 ligt op het
RHA1 genoom (Tabel 1), hetgeen wijst op een mogelijke functie van deze drie CYPs in
dezelfde, maar tot nog toe nog onbekende route. Van een lid van de CYP123 familie,
afkomstig van R. erythropolis PR4, werd reeds activiteit op testosteron aangetoond [3]
(zie ook Fig. 5 in Hoofdstuk 1). De eiwit sequentie van CYP51B1 van stam RHA1
(CYP51B1RHA1) is voor 74% identiek aan die van CYP51 van M. tuberculosis H37Rv, die
actief is met lanosterol, 24,25-dihydrolanosterol, 24-methylenedihydrolanosterol,
obtusifoliol en 4β-desmethyllanosterol (norlanosterol) [167]. Het is dus mogelijk dat
CYP51B1RHA1 ook activiteit op sterolen vertoont, te meer omdat alle tot nu toe
gekarakteriseerde CYP51 enzymen een vergelijkbare functie hebben, namelijk 14α-
demethylatie van Δ8-sterols. Eukaryote CYP51 enzymen katalyseren sterol
transformaties, meestal als onderdeel van sterol biosynthese routes. In bacteriën is de
fysiologische rol van CYP51 echter grotendeels onduidelijk gebleven [167].
Met uitzondering van M. smegmatis [157] zijn er geen actinomyceten bekend die sterolen
synthetiseren. De rol van CYP51 in actinomyceten en andere bacteriën, die geen sterolen
synthetiseren, lijkt dus anders te zijn. Voor sommige CYP51s uit zoogdieren, parasieten
158

en planten is een regulerende rol in de biosynthese van cholesterol aangetoond en
mogelijk ook in de meiose, apoptose of spermatogenese [48, 72, 167, 336]. Daarnaast
speelt CYP51 een rol in de synthese van antimicrobiële verbindingen: het plantengeslacht
Avena synthetiseert avenacins, hetgeen triterpeenglycosiden (saponinen) zijn [242]. De
functie van CYP51 in Rhodococcus stammen moet nog verder worden onderzocht.
Samengevat, CYP125B1, CYP123A5 en CYP51B1 van stam RHA1 zijn mogelikerwijs
allen betrokken bij steroid modificaties.
Expressie van het CYP116C2 gen is sterk verhoogd in stam RHA1 cellen gegroeid op
cholesterol (6.8 maal; Tabel 1). CYP116C1 is een isovorm van CYP116C2. CYP116C1
ligt gecodeerd op een ander plasmide en komt niet tot expressie bij groei op cholesterol
(Tabel 1). De functies van CYP116C1 en CYP116C2 zijn echter onbekend. Gezien de
verhoogde expressie van CYP116C2 op cholesterol is dit enzym mogelijkerwijs actief
met sterolen, alhoewel geen van de gekarakteriseerde CYP116 familieleden sterolen of
steroïde verbindingen bleken om te kunnen zetten (bv. Liu et al. [172], Munro et al.
[198]).
Ook het CYP102B3 gen kwam verhoogd tot expressie in stam RHA1 cellen gekweekt op
cholesterol (2.3 voudig; Tabel 1). Microbiële cytochroom P450 systemen van familie
CYP102 en subfamilie A zijn zeer interessant gezien de beschikbare kennis van
CYP102A1 (P450BM3) van B. megaterium stam ATCC 14581, een zelfvoorzienend CYP
[196]. Geen van de CYPs geïdentificeerd in stam RHA1 zijn echter zelfvoorzienend.
Ondanks dat er verschillende enzymen van CYP102 uit de subfamilie A uit verschillende
stammen Bacillus werden gekarakteriseerd, samengevat door Furuya et al. [69], is slechts
van een CYP102 enzym (CYP102A1 van B. megaterium) aangetoond dat het de
hydroxylering van testosteron op 7β positie kan katalyseren [3] (zie ook Fig. 5 in
159

Hoofdstuk 1). CYP102A1 activiteit op testosteron werd daarintegen niet gevonden door
van Vugt-Lussenburg et al. [311]. Slechts een van de leden van de CYP102 subfamilie B
is tot nu toe nader bestudeerd, namelijk CYP102B1 uit Streptomyces coelicolor stam
A3(2) welke arachidonzuur metaboliseert [159]. CYP102B3 uit stam RHA1 moet nog
nader worden gekarakteriseerd.
Agematu et al. [3] heeft aangetoond dat de CYP gecodeerd door nfa33880 van Nocardia
farcinica stam IFM10152 (CYPnfa33880) testosteron oxidatie activiteit heeft voor de 12β en
17 posities (zie ook Fig. 5 in Hoofdstuk 1). CYP256A1 uit stam RHA1, gecodeerd door
ro08984, komt overeen met CYPnfa33880 (75% identiteit): mogelijkerwijs heeft ook dit
CYP256A1 enzym een steroid modificerende activiteit.
Van CYP105s uit actinomyceten werd aangetoond dat ze testosteron oxideren op de 2α,
2β, 15β en 17 posities [3] (zie ook Fig. 5 in Hoofdstuk 1). CYP105Y1 uit stam RHA1
(CYP105Y1RHA1) heeft mogelijkerwijs ook activiteit met testosteron. Kristalstructuren
van verschillende CYP105s enzymen van actinomyceten zijn bekend [117, 282, 326, 327,
331]. Gezien de snelle toename in kennis over de familie CYP105 enzymen zal de
activiteit van het CYP105Y1RHA1 enzym en de in vivo rol naar verwachting binnenkort
opgehelderd worden.
Outlook
Rhodococcus stammen zijn katabole krachtpatsers [189]. De verkenning van hun set van
enzymen, waaronder de talrijke geannoteerde CYPs in hun genoom sequenties, opent een
fascinerend gebied voor onderzoek, met interessante mogelijkheden voor industriele
toepassingen. Rhodococcus stammen zijn efficiënte afbrekers van plantaardige en
dierlijke sterolen/steroïden. In eerder werk hebben we gemuteerde stammen gemaakt die
160

geblokkeerd waren in de steroïd ring afbraak. Deze stammen kunnen ook dienen als
gastheer voor de efficiënte heterologe expressie van geselecteerde steroïd modificerende
enzymen (bv. CYP eiwitten), voor de biosynthese van bioactieve en farmaceutisch
interessante steroïden (Fig. 1).
In dit proefschrift hebben we afbraak waargenomen van sterolen door de mutante stam
RG32Ωcyp125 (Hoofdstuk 2) en een snelle omzetting van 16β-hydroxy-AD door de
mutante stam RG9 (Hoofdstuk 4), hetgeen vermoedelijk te wijten is aan (onbekende)
regulerende mechanismen, resulterend in de inductie van meerdere katabole enzymen
door steroïd verbindingen. In toekomstig werk is het daarom van belang om een grondige
kennis op te bouwen van de genen die coderen voor deze katabole enzymen en over de
regulering van hun genexpressie. Op basis van die kennis kunnen dan nieuwe
Rhodococcus stammen worden ontwikkeld met gemanipuleerde metabole routes die
efficiënt de gewenste steroïd moleculen accumuleren. Karakterisatie van de volledige set
van steroïd enzymen in Rhodococcus stammen zou een beter begrip van de katabole
routes in deze bacteriën geven en ons tevens in staat stellen om vergelijkende studies met
nauw verwante stammen van actinomyceten, bv. in (pathogene) Mycobacterium
stammen, uit te voeren. Diverse steroïd katabole genclusters lijken te zijn geconserveerd
in verschillende actinomyceten (bv. Knol et al. [142], van der Geize et al. [309]). Kennis
over de regulatie van cholesterol katabolisme in mycobacteriën neemt geleidelijk aan toe
[132].
Nadere analyses van de moleculaire mechanismen van natieve en recombinante CYPs zal
nodig zijn voor het ontwikkelen van efficiënte bioconversie processen met Rhodococcus
stammen. Recent is aangetoond dat het actief centrum van CYPs twee subsites`heeft die
een interactie met elkaar aangaan, een voor de binding van een substraat en de andere
161

voor de regulering van het enzym tijdens de katalyse [267]. Tevens zijn er vaak
transcriptionele regulator genen gelegen naast CYP genen in stam RHA1
(www.rhodococcus.ca), wijzend op de aanwezigheid van controlemechanismen in stam
RHA1 die nog nader moeten worden opgehelderd.
De aanwezigheid van CYP redox donoreiwitten en NAD(P)H zijn van cruciaal belang
voor het goed functioneren van de meeste P450 enzymen (Hoofdstuk 1, Tabel 2). Voor
realisatie van doeltreffende toepassingen van CYP-enzymen in bacteriële bioconversies is
ook een verdere optimalisatie nodig van de aanwezigheid van functionele CYP donor
redox-systemen en voldoende NAD(P)H.
162






















