Embed Size (px)
Citation preview

NOAA Technical Memorandum NMFS-AFSC-212
Status Review of the Ringed Seal (Phoca hispida)
by B. P. Kelly, J. L. Bengtson, P. L. Boveng, M. F. Cameron, S. P. Dahle, J. K. Jansen, E. A. Logerwell, J. E. Overland, C. L. Sabine, G. T. Waring, and J. M. Wilder
U.S. DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Administration
National Marine Fisheries Service Alaska Fisheries Science Center
December 2010

NOAA Technical Memorandum NMFS
The National Marine Fisheries Service's Alaska Fisheries Science Center uses the NOAA Technical Memorandum series to issue informal scientific and technical publications when complete formal review and editorial processing are not appropriate or feasible. Documents within this series reflect sound professional work and may be referenced in the formal scientific and technical literature.
The NMFS-AFSC Technical Memorandum series of the Alaska Fisheries Science Center continues the NMFS-F/NWC series established in 1970 by the Northwest Fisheries Center. The NMFS-NWFSC series is currently used by the Northwest Fisheries Science Center.
This document should be cited as follows:
B. P. Kelly, J. L. Bengtson, P. L. Boveng, M. F. Cameron, S. P. Dahle, J. K. Jansen, E. A. Logerwell, J. E. Overland, C. L. Sabine, G. T. Waring, and J. M. Wilder 2010. Status review of the ringed seal (Phoca hispida). U.S. Dep. Commer., NOAA Tech. Memo. NMFS-AFSC-212, 250 p.
Reference in this document to trade names does not imply endorsement by the National Marine Fisheries Service, NOAA.

December 2010
NOAA Technical Memorandum NMFS-AFSC-212
by
Status Review of the Ringed Seal(Phoca hispida)
U.S. DEPARTMENT OF COMMERCEGary F. Locke, Secretary
National Oceanic and Atmospheric Administration Jane Lubchenco, Under Secretary and Administrator
National Marine Fisheries ServiceEric. C. Schwaab, Assistant Administrator for Fisheries
B. P. Kelly1, J. L. Bengtson1, P. L. Boveng1, M. F. Cameron1,S. P. Dahle1, J. K. Jansen1, E. A. Logerwell1, J. E. Overland2,
C. L. Sabine2, G. T. Waring3, and J. M. Wilder4
1 Alaska Fisheries Science Center7600 Sand Point Way NE
Seattle, WA 98115www.afsc.noaa.gov
2 Pacific Marine Environmental LaboratoryOffice of Oceanic and Atmospheric Research
7600 Sand Point Way NESeattle, WA 98115
3 Northeast Fisheries Science Center166 Water Street
Woods Hole, MA 02543
4 U.S. Fish and Wildlife Service, Alaska Region, Marine Mammals Management1011 East Tudor RoadAnchorage, AK 99503

This document is available to the public through:
National Technical Information Service U.S. Department of Commerce 5285 Port Royal Road Springfield, VA 22161
www.ntis.gov

STATUS REVIEW OF THE RINGED SEAL (Phoca hispida)
Prepared and Edited by:
The 2010 Ringed Seal Biological Review Team Brendan P. Kelly1 (Chair), John L. Bengtson1, Peter L. Boveng1, Michael F. Cameron1,
Shawn P. Dahle1, John K. Jansen1, Elizabeth A. Logerwell1, James E. Overland2, Christopher L. Sabine2, Gordon T. Waring3, and James M. Wilder4
With Contributions by:
Josh M. London1, Erin E. Moreland1, Tamara L. Olson5, Duane E. Stevenson1, and Muyin Wang6,2
1 Alaska Fisheries Science Center, National Marine Fisheries Service, 7600 Sand Point Way NE, Seattle, WA 98115
2 Pacific Marine Environmental Laboratory, Office of Oceanic and Atmospheric Research, 7600 Sand Point Way NE, Seattle, WA 98115
3 Northeast Fisheries Science Center, National Marine Fisheries Service, 166 Water Street, Woods Hole, MA 02543
4 U.S. Fish and Wildlife Service, Alaska Region, Marine Mammals Management, 1011 East Tudor Road, Anchorage, AK 99503
5 Protected Resources Division, Alaska Region, National Marine Fisheries Service, 222 West 7th Ave., Anchorage, AK 99513
6 Joint Institute for the Study of the Atmosphere and Ocean, University of Washington, Box 355672, Seattle, WA 98195
December 2010
Photo by Brendan P. Kelly


vii
CONTENTS
Contents ...................................................................................................................................................... vii
Executive Summary ...................................................................................................................................... xi
1 Introduction .......................................................................................................................................... 1
2 Species Background .............................................................................................................................. 3
2.1 Taxonomy and Phylogeny ................................................................................................................... 3
2.2 Species Description ............................................................................................................................. 4
2.3 Behavior .............................................................................................................................................. 5
2.4 Seasonal Distribution, Habitat‐use, and Movements ......................................................................... 8
2.4.1 Open‐water period..................................................................................................................... 10
2.4.2 Subnivean period ....................................................................................................................... 12
2.4.3 Basking period ............................................................................................................................ 14
2.5 Reproduction and Molting ................................................................................................................ 14
2.6 Vital Parameters ................................................................................................................................ 16
2.7 Foraging and Diet .............................................................................................................................. 18
2.7.1 Marine environment .................................................................................................................. 18
2.7.2 Freshwater environment ........................................................................................................... 26
2.8 Historic and Current Abundance and Trends .................................................................................... 26
2.8.1 Arctic ringed seals ...................................................................................................................... 27
2.8.1.1 Greenland Sea and Baffin Bay ............................................................................................. 27
2.8.1.2 Hudson Bay ......................................................................................................................... 28
2.8.1.3 Beaufort and Chukchi Seas ................................................................................................. 29
2.8.1.4 White, Barents, Kara, and East Siberian Seas ..................................................................... 30
2.8.2 Okhotsk ringed seals .................................................................................................................. 30
2.8.3 Baltic ringed seals ...................................................................................................................... 31
2.8.4 Ladoga ringed seals .................................................................................................................... 32
2.8.5 Saimaa ringed seals .................................................................................................................... 32
3 Species Delineation ............................................................................................................................. 35
3.1 Evaluation of Discreteness ................................................................................................................ 35
3.1.1 Separation by Physical, Physiological, Ecological, or Behavioral Factors .................................. 35

viii
3.1.2 Delimitation by International Differences in Management and Regulatory Mechanisms ........ 38
3.2 Evaluation of Significance ................................................................................................................. 39
3.3 Determination of Distinct Population Segments .............................................................................. 39
4 Extinction Risk Assessment ................................................................................................................. 41
4.1 Time Frame: The Foreseeable Future ............................................................................................... 41
4.1.1 Factors in the Foreseeability of Threats to Ringed Seals ........................................................... 41
4.1.2 Factors in the Foreseeability of Ringed Seal Responses to Threats ........................................... 43
4.1.3 Lack of a Single Time Frame for the Foreseeable Future ........................................................... 44
4.2 Analysis of Factors Listed Under Section 4(a)(1) of the Endangered Species Act ............................ 44
4.2.1 Present or Threatened Destruction, Modification, or Curtailment of the Species’ Habitat or Range .................................................................................................................................................. 44
4.2.1.1 Global climate change ......................................................................................................... 44
4.2.1.2 Impacts of global climate change on ringed seals ............................................................ 104
4.2.2 Overutilization for Commercial, Recreational, Scientific, or Educational Purposes ................ 113
4.2.2.1 Commercial, subsistence, and illegal harvest ................................................................... 113
4.2.2.2 Scientific and educational utilization ................................................................................ 120
4.2.3 Diseases, Parasites, and Predation .......................................................................................... 121
4.2.3.1 Diseases ............................................................................................................................. 121
4.2.3.2 Parasites ............................................................................................................................ 124
4.2.3.3 Predation ........................................................................................................................... 126
4.2.4 Inadequacy of Existing Regulatory Mechanisms ...................................................................... 129
4.2.4.1 Existing conservation efforts ............................................................................................. 129
4.2.5 Other Natural or Human Factors Affecting the Species’ Continued Existence ........................ 140
4.2.5.1 Pollution and contaminants .............................................................................................. 140
4.2.5.2 Oil and gas exploration, development, and production ................................................... 147
4.2.5.3 Wind farms ........................................................................................................................ 161
4.2.5.4 Commercial fisheries interactions and bycatch ................................................................ 161
4.2.5.5 Shipping and transportation ............................................................................................. 164
4.2.6 Threats Assessment ................................................................................................................. 174
4.3 Analysis of Demographic Risks ........................................................................................................ 180
4.3.1 Abundance ............................................................................................................................... 180
4.3.2 Productivity .............................................................................................................................. 183

ix
4.3.3 Spatial Structure ....................................................................................................................... 185
4.3.4 Diversity ................................................................................................................................... 187
4.3.5 Relevant Modifying or Mitigating Factors ............................................................................... 189
4.3.5.1 Life‐history characteristics ................................................................................................ 189
4.3.5.2 Population characteristics ................................................................................................. 190
4.3.5.3 Habitat characteristics ...................................................................................................... 190
4.3.6 Demographic Risks Assessment ............................................................................................... 191
4.4 Conclusions of the Extinction Risk Assessment .............................................................................. 193
4.4.1 Arctic ringed seals .................................................................................................................... 193
4.4.2 Okhotsk ringed seals ................................................................................................................ 193
4.4.3 Baltic ringed seals .................................................................................................................... 194
4.4.4 Ladoga ringed seals .................................................................................................................. 194
4.4.5 Saimaa ringed seals .................................................................................................................. 194
5 Acknowledgments ............................................................................................................................. 195
6 Citations ............................................................................................................................................ 197
7 Appendix: Glossary of Abbreviations ................................................................................................ 249


xi
EXECUTIVE SUMMARY
A Biological Review Team (BRT) convened by the National Marine Fisheries Service assessed the best available information concerning the status of ringed seals (Phoca hispida) and past, present, and future threats to the species in response to a petition filed by the Center for Biological Diversity. The petition seeks to list the ringed seal as threatened or endangered under the U.S. Endangered Species Act (ESA), primarily due to concern about threats to the species’ habitat from climate warming and diminishing ice and snow cover.
The BRT’s review included delineating population structure within the species and assessing the risk of extinction at present and in the foreseeable future. The review is intended to inform the Secretary of Commerce’s decision whether to list the species as endangered or threatened in all or part of its range. The ESA defines an endangered species as “any species which is in danger of extinction throughout all or a significant portion of its range.” Threatened species is defined as “any species which is likely to become an endangered species within the foreseeable future throughout all or a significant portion of its range.” The foreseeable future was not considered to be fixed but, rather, threat specific. Ice and snow habitats are affected by climate which is forecasted to continue changing directionally at least until the end of the century in response to greenhouse gas (GHG) forcing. Variability in the magnitude—but not the directionality—of climate change increases over time and is reflected in inter‐model variability and in levels of certainty assigned to the BRT’s assessments of threats and demographic risks.
Species Background: Ringed seals are one of the smallest true seals (Phocidae), a group of marine carnivores descended from terrestrial mammals. The distribution of phocids is biased toward polar seas reflecting evolutionary adaptations to ice‐covered waters. Ringed seals are the most strongly ice‐associated seal, coming out of the water exclusively on sea ice except in marginal seas and freshwater lakes where ice disappears seasonally. Their global distribution has expanded and contracted with changing sea‐ice cover, and today they inhabit all the seasonally ice‐covered seas of the Northern Hemisphere as well as Lake Saimaa (Finland) and Lake Ladoga (Russia). After reaching sexual maturity, typically at 4‐8 years—and when environmental conditions are favorable—female ringed seals produce a single pup each year. The pups are born in subnivean lairs (snow caves) excavated above breathing holes in the ice, where they are nursed for 5‐9 weeks. Males typically become sexually mature at 5‐7 years. Survival rates are not well known, but ringed seals can live in excess of 40 years. Ringed seals eat a wide variety of prey but tend to prefer small, schooling species that form dense aggregations, such as cods, smelts, and crustaceans. Population estimates are highly uncertain in most areas and range from the low millions in the Arctic to less than 300 in Lake Saimaa.
Species Delineation: Five subspecies of ringed seals inhabit the Arctic Ocean, Sea of Okhotsk, Baltic Sea, Lake Ladoga, and Lake Saimaa and are discrete breeding populations. The Arctic Ocean subspecies may be comprised of multiple discrete subpopulations, but population structure is not fully resolved.
Extinction Risk Assessment: For each subspecies, the BRT evaluated 17‐18 threats grouped by the ESA Section 4(a)(1) factors:

xii
• the present or threatened destruction, modification, or curtailment of its habitat or range,
• overutilization for commercial, recreational, scientific, or educational purposes,
• disease or predation,
• the inadequacy of existing regulatory mechanisms, or
• other natural or manmade factors affecting its continued existence
The BRT also assessed the risks to population persistence posed by those threats in demographic terms (abundance, productivity, spatial structure, and diversity). The Arctic Ocean, Sea of Okhotsk, Baltic Sea, and Lake Ladoga subspecies all number several thousand or more seals and are not believed to be currently at risk from the effects of demographic stochasticity, inbreeding, loss of genetic diversity, or depensation. The Saimaa seal, however, numbers fewer than 300 individuals and shows substantially lower genetic diversity than do the other subspecies. Saimaa ringed seals have been listed as endangered under the ESA since 1993.
Present or threatened destruction, modification, or curtailment of the species’ habitat or range: Diminishing ice and snow cover are the greatest challenges to persistence of all of the ringed seal subspecies. Climate models consistently project overall diminishing ice and snow cover at least through the current century with regional variation in the timing and severity of those losses. Increasing atmospheric concentrations of GHGs, including carbon dioxide (CO2), will drive climate warming and increase acidification of the ringed seal’s ocean and lake habitats. Acidification threatens changes in prey communities on which ringed seals depend.
Ice loss will be greatest in the summer and fall months when the ringed seal’s use of ice as a resting platform is at a minimum. In those months, however, ice remains important to prey populations such as Arctic cod, and ringed seal populations will be affected by diminished prey populations. Increased competition with northward‐expanding, subarctic species may also affect prey densities. The greatest impacts to ringed seals of diminished ice cover will be mediated through diminished snow accumulation. While winter precipitation is forecasted to increase in a warming Arctic, the duration of ice cover will be substantially reduced, and the net affect will be lower snow accumulation on the ice. Model forecasts indicate that throughout the range of ringed seals, there will be substantial reductions in on‐ice snow cover. Snow depth limits the formation of subnivean lairs, and birth lairs require depths of at least 50‐65 cm. Such depths typically are found only where 20‐30 cm or more of snow has accumulated on flat ice and drifted along pressure ridges or ice hummocks. Within the century, snow cover is forecasted to be inadequate for the formation and occupation of birth lairs over most of the species’ range. Without the protection of the lairs, ringed seals—especially newborn—are vulnerable to freezing and predation. As populations decline, the significance of currently low‐level threats—including ocean acidification, increased human activity, and changes in populations of prey, predators, competitors, and parasites—may increase.
Overutilization for commercial, subsistence, recreational, scientific, or educational purposes: Subsistence and commercial harvests of Arctic ringed seals have been large in the past, but there is no

xiii
evidence that they have contributed to large‐scale population declines. Commercial harvests in the Sea of Okhotsk and predator‐control harvests in the Baltic Sea, Lake Ladoga, and Lake Saimaa caused population declines in the past but have since been restricted. Current harvest levels appear to be low and sustainable. Recreational, scientific, and educational uses are minimal and not projected to increase significantly in the foreseeable future for any of the subspecies.
Diseases, parasites, and predation: Ringed seals have co‐evolved with numerous parasites and diseases, and those relationships are presumed to be stable. Evidence of distemper virus, for example, has been reported in Arctic ringed seals, but there is no evidence of impacts to ringed seal population size or productivity. Abiotic and biotic changes to ringed seals’ habitat potentially could lead to exposure to new pathogens or new levels of virulence, but the BRT considered the potential threats to ringed seals as low.
Ringed seals are commonly preyed upon by polar bears and Arctic foxes, and less commonly by other terrestrial carnivores, sharks, and killer whales. Predation on newborn pups by gulls and ravens is typically prevented by the pups’ concealment in subnivean lairs. When the pups are prematurely exposed, however, predation by birds—as well as terrestrial carnivores—can be substantial.
Inadequacy of existing regulatory mechanisms: Harvests and incidental takes by fisheries and commercial activities are reasonably well regulated throughout the range of ringed seals. Currently, however, there are no effective mechanisms to regulate the global GHG emissions that are driving—via climate warming—destruction of ringed seal habitat. The BRT implicitly considered impacts of inadequate regulation of GHG emissions by way of the emissions scenarios used in forecast models; the scenarios were all “non‐mitigated”, meaning that they assumed no globally‐significant framework for regulating or reducing emissions would be implemented.
Other natural or human factors affecting the species’ continued existence: Drowning of seals in fishing nets and disturbance by human activities remain conservation concerns in Lake Saimaa and Lake Ladoga. Reduced productivity in the Baltic Sea subspecies in recent decades resulted from pollutants impairing fertility. Petroleum development, commercial fisheries, increased ship traffic, and pollutants pose moderate risks to the Arctic, Okhotsk, and Baltic subspecies. Their significance would increase, however, for any populations diminished by the effects of climate change or other threats.
Status of the ringed seal subspecies: The BRT reviewed published data and consulted with other experts to evaluate the specific threats and demographic risks to population persistence for each subspecies of ringed seals. Threats and demographic risks were scored quantitatively, and the level of certainty in scores was also recorded. The BRT concluded:
Arctic ringed seals: Persistence of the Arctic subspecies likely will be challenged as decreases in ice and, especially, snow cover lead to increased juvenile mortality from premature weaning, hypothermia, and predation. The depth and duration of snow cover are forecasted to decline substantially throughout the range of Arctic ringed seals. Risks to abundance, productivity, spatial structure, and diversity currently are low. In the foreseeable future, however, it is expected that abundance and productivity will decline and spatial structure will be disrupted by rapid loss of habitat. Initially, impacts may be somewhat

xiv
ameliorated if the subspecies’ range retracts northward with sea‐ice habitats. By 2100, however, average snow depths will fail to meet the 20‐30 cm minimum needed for successful formation and maintenance of birth lairs in a substantial portion of the subspecies’ range. Thus, within the foreseeable future, it is likely that the number of Arctic ringed seals will decline substantially, and they will no longer persist in substantial portions of their range.
Okhotsk ringed seals: Before the end of the current century, ice is forecasted to be limited to the northernmost regions of the Sea of Okhotsk during the whelping and nursing periods. On‐ice measurements of snow cover are not available for the Sea of Okhotsk, but model results indicate inadequate (< 20 cm average depth) snow for birth lairs throughout the subspecies’ range in the most recent decade. Okhotsk ringed seals apparently depend on sheltering in the lee of ice hummocks as snow cover is inadequate for lair construction in much of the habitat. In the foreseeable future, the diversity of the subspecies will be at moderate risk while its abundance, productivity, and spatial structure will be highly at risk. Okhotsk ringed seals likely will decline to levels that threaten their persistence as a consequence of a decrease in sea‐ice habitat suitable for whelping, nursing, and molting. The range of Okhotsk ringed seals is bounded by land to the north, and the opportunity to retract its range with the ice is limited accordingly.
Baltic ringed seals: Substantial reductions in sea‐ice extent by mid‐century, coupled with deteriorating snow conditions, will substantially alter the habitat of Baltic ringed seals and lead to decreased survival of pups. In the foreseeable future, risks to demographic attributes are expected to be moderate (diversity) to high (abundance, productivity, and spatial structure). The range of Baltic ringed seals is bounded by land to the north, and the opportunity to retract their range with the ice is limited accordingly. Degradation of ice and snow habitats is likely to cause substantial population declines and threaten the Baltic ringed seal’s persistence within the foreseeable future.
Ladoga ringed seals: Persistence of Ladoga ringed seals will be challenged by decreased ice habitat suitable for whelping and nursing as well as increased pup mortality from hypothermia and predation due to insufficient depth or duration of snow cover. At present, there is a moderate risk to the subspecies’ persistence since, as a landlocked population, Ladoga ringed seals cannot disperse to new habitats. Within the foreseeable future, persistence will be challenged by moderate risks to diversity and high risks to abundance, productivity, and spatial structure. Degradation of ice and snow habitats is likely to cause substantial population declines and threaten the Ladoga ringed seal’s persistence within the foreseeable future.
Saimaa ringed seals: Saimaa seals currently are listed as endangered under the ESA. The population remains low (< 300 seals) and pup mortality remains high. Forecasts of decreasing snow depth and seasonal duration likely will increase pup mortality through hypothermia and predation. Declining ice cover represents a further loss of habitat. The present risk to population persistence is moderate to high in terms of productivity and diversity and high to very high in terms of abundance and spatial structure. In the foreseeable future, the risks are expected to be high to very high for all of the demographic attributes. Degradation of ice and snow habitats is likely to cause substantial population declines and further threaten the Saimaa ringed seal’s persistence within the foreseeable future.

1
1 INTRODUCTION
On 28 March 2008, the National Marine Fisheries Service (NMFS) initiated conservation status reviews of the ringed seal (Phoca hispida), bearded seal (Erignathus barbatus), and spotted seal (Phoca largha) (National Marine Fisheries Service 2008b). On 28 May 2008, the Center for Biological Diversity (CBD) filed a petition with the Secretary of Commerce (Secretary) and NMFS to list these three species as threatened or endangered, and to designate critical habitat for these species pursuant to the U.S. Endangered Species Act (ESA) of 1973, as amended (16 U.S.C. 1531 et seq.) (Center for Biological Diversity 2008).
Section 4(b)(3)(A) of the ESA requires the Secretary to determine, to the maximal extent practicable, within 90 days of receiving a petition to list a species under the ESA, whether the petition presents substantial scientific or commercial information indicating that the petitioned action may be warranted. This finding is to be promptly published in the Federal Register. On 4 September 2008, NMFS published a positive 90‐day finding stating that the CBD’s petition presented substantial scientific or commercial information indicating that the petitioned action may be warranted (National Marine Fisheries Service 2008c). To assist in determining whether listing the ringed seal under the ESA is warranted, NMFS convened an expert panel (the 2010 Ringed Seal Biological Review Team, or BRT) to conduct an ESA status review for this species. The BRT was composed of 8 marine mammal biologists, 1 climate scientist, 1 ocean chemist, and 1 fishery biologist. A status review (Boveng et al. 2009) and 12‐month finding (National Marine Fisheries Service 2009) for spotted seals were published in October 2009, and a status review of bearded seals was conducted concurrent with this one.
There are two key tasks associated with conducting an ESA status review. The first task is to delineate the taxonomic group under consideration. To be considered for listing under the ESA, a group of organisms must constitute a “species”, which according to the ESA includes “any subspecies of fish or wildlife or plants, and any distinct population segment of any species of vertebrate fish or wildlife which interbreeds when mature.” The BRT applied the joint U.S. Fish and Wildlife Service (USFWS)‐NMFS Policy Regarding the Recognition of Distinct Population Segments Under the Endangered Species Act (U.S. Fish and Wildlife Service and National Marine Fisheries Service 1996) to determine whether the ringed seal or any of its subspecies merits delineation into distinct population segments (DPSs). That analysis can be found in Section 3 of the review.
The second key task of a status review is to conduct an extinction risk assessment to serve as the scientific basis for determining whether the petitioned species is threatened or endangered. The ESA defines the term endangered species as “any species which is in danger of extinction throughout all or a significant portion of its range.” The term threatened species is defined as “any species which is likely to become an endangered species within the foreseeable future throughout all or a significant portion of its range.” To make this assessment, the BRT evaluated the time frames over which future events can be reasonably said to be “foreseeable”, and assessed the risks based on specific demographic factors of the

2
species, such as abundance, productivity, spatial structure, and diversity, as well as specific threats faced by the species, as outlined in Section 4(a)(1) of the ESA:
• the present or threatened destruction, modification, or curtailment of its habitat or range,
• overutilization for commercial, recreational, scientific, or educational purposes,
• disease or predation,
• the inadequacy of existing regulatory mechanisms, or
• other natural or manmade factors affecting its continued existence
That analysis can be found in Section 4 of the review.
This document is a compilation of the best available scientific and commercial data describing the past, present, and likely future threats to the ringed seal. It does not represent a decision by NMFS on whether this taxon should be proposed for listing as threatened or endangered under the ESA. That decision will be made by NMFS after reviewing this document, other relevant biological and threat information not included herein, efforts being made to protect the species, and all relevant laws, regulations, and policies. The result of the decision will be posted on the NMFS website (refer to: http://www.nmfs.noaa.gov/pr/species/) and announced in the Federal Register.

3
2 SPECIES BACKGROUND
2.1 Taxonomy and Phylogeny
Pinnipeds, a monophyletic group of aquatic carnivores, diverged from their closest terrestrial relatives (mustelids or ursids) approximately 40 million years ago (Higdon et al. 2007, Rybczynski et al. 2009, Fulton and Strobeck 2010a). They likely originated in the Arctic (Davies 1958), feeding first in freshwater lakes and later moving to marine foraging (Rybczynski et al. 2009). Arctic sea ice has been implicated in the subsequent divergence of pinnipeds in to three families: the Phocidae (true seals), Otariidae (sea lions and fur seals), and Odobenidae (walruses) 20‐25 million years ago (Fulton and Strobeck 2010a). The Phocidae comprise two subfamilies: the Monachinae (“southern seals”) and Phocinae (“northern seals”) (Berta 2009). The Phocinae, including ringed seals, are believed to have adapted to breeding on sea ice 13‐17 million years ago based on their shared white natal coat (Árnason et al. 2006, Higdon et al. 2007). Biogeographic evidence suggests that the Phocina arose in the Greenland‐Barents Sea region and radiated in response to global changes in sea level and advances and contractions of sea ice (Árnason et al. 2006, Higdon et al. 2007, Fulton and Strobeck 2010a).
Three primitive members of the Phocinae—ringed, Baikal, and Caspian seals—are grouped together as Pusa, recognized by some as a subgenus of Phoca (Chapskii 1955, McLaren 1960, Burns and Fay 1970, Grigorescu 1976, Ray 1976, Repenning et al. 1979, Árnason et al. 1995, Sasaki et al. 2003, Árnason et al. 2006, Higdon et al. 2007), but given generic status by others (Allen 1880, Scheffer 1958, King 1964, De Muizon 1982, Rice 1998, Demere et al. 2003). The most comprehensive molecular analysis of phocid relationships to date (15 nuclear and 13 mitochondrial genes) emphasized the importance of including multiple individuals from each species (Fulton and Strobeck 2010b). Genetic distances were no greater between than within Pusa, Phoca, and Halichoerus (gray seal), consistent with assigning all three to subgenera within Phoca as suggested by Árnason et al. (1995, 2006). Noting the morphological distinctness of Halichoerus, however, Fulton and Strobeck (2010b) considered the generic distinctions to be unresolved. We follow Burns and Fay (1970) and Árnason et al. (2006) and classify ringed seals as Phoca hispida with the recognition that molecular and morphological analyses remain incompletely resolved.
We provisionally recognize five subspecies of ringed seals—the Arctic ringed seal (Phoca hispida hispida); the Baltic ringed seal (Phoca hispida botnica); the Okhotsk ringed seal (Phoca hispida ochotensis); the Ladoga ringed seal (Phoca hispida ladogensis); and the Saimaa ringed seal (Phoca hispida saimensis)—as “species” for the purposes of the ESA. The latter four subspecies are well supported; genetic structuring within the Arctic subspecies, however, has yet to be thoroughly investigated, and we caution that it may prove to be composed of multiple distinct populations.
As many as 10 subspecies of ringed seals have been proposed based on distribution and morphological variations (Allen 1880, Anderson 1934, Ognev 1935, Chapskii 1955, Scheffer 1958, King 1983), however, there remain several unresolved issues with ringed seal systematics (Amano et al. 2002, Palo 2003, Sell

4
2008, Kelly et al. 2009). Rice (1998) suggested that available information was consistent with five subspecies, but he noted that additional subspecies might be recognized with additional data.
During the Last Glacial Maximum, ringed seals were displaced southward from the Arctic Ocean by heavy sea ice and land bridges (Harington and Sergeant 1972, Palo 2003), and they were found as far south as southeastern Alaska and Portugal (Helle 1992 cited in Reeves 1998, Heaton and Grady 2003). The species’ range advanced northward again as the ice retreated, but refugial populations remained in the Baltic Sea region and the Sea of Okhotsk. Seals in the former basin were separated from Arctic ringed seals 11,000 years ago when an ice barrier formed the Baltic Ice Lake (Forstén and Alhonen 1975, Hyvärinen and Nieminen 1990). Seals confined to that freshwater lake were further fragmented when Oulujärvi, Saimaa, Ladoga, and other lakes were isolated 9,000 and 8,000 years ago (Müller‐Wille 1969). Ringed seal populations subsequently went extinct in the freshwater lakes with the exception of Lakes Saimaa and Ladoga (Forstén and Alhonen 1975, Ukkonen 1993, Ukkonen 2002). The Sea of Okhotsk provided year‐round ice to seals from 21,000 to 11,000 years ago, but only seasonal ice cover for the past 8,000 years (Shiga and Koizumi 1999).
Over the past 8,000‐9,000 years, ringed seals in Lake Ladoga and Lake Saimaa have accumulated sufficient differences in skull morphology and coat color to warrant subspecific status according to Hyvärinen and Nieminen (1990) and Amano et al. (2002). Craniological analyses did not warrant subspecific recognition for ringed seals in the Baltic Sea, but Amano et al. (2002) argued to retain subspecific rank based on the isolation from Arctic ringed seals and darker pelage. The light morph is rare or absent in the Baltic Sea subspecies. Like Arctic ringed seals, the lake‐inhabiting subspecies rest on ice in winter and spring, but they rest on islets or shorelines in the ice‐free seasons. Whereas the marine subspecies excavate subnivean lairs in snow drifts formed along pressure ridges and other deformities in the ice, the lake inhabiting subspecies excavate subnivean lairs along shorelines.
Amano et al. (2002) believed that their sample size (N = 12) was insufficient to reach a firm conclusion on the subspecific status of ringed seals in the Sea of Okhotsk. Fedoseev (1984), however, considered Sea of Okhotsk ringed seals to be “well differentiated by morphological and ecological characters” from Bering Sea ringed seals. Fedoseev and Nazarenko (1970) found little morphological differences between ringed seals from the Bering and Barents Seas, but they pointed out that resolving population structure within the Arctic Basin would require samples from more areas.
2.2 Species Description
Ringed seals are small phocids with typical adult body sizes of 1.5 m in length and 70 kg in weight. The species is dimorphic in pelage, having both dark and light phases (Murdoch 1885, Ognev 1935, McLaren 1966, Kelly 1981) similar to those described for harbor seals (Phoca vitulina) (Stutz 1967, Shaughnessy and Fay 1977, Kelly 1981). The light phase consists of a dark grey saddle with superimposed light rings and lightly‐colored lateral and ventral surfaces with or without darker spots, while the dark phase has a dark background with light rings overall. Body size and the ratio of light to dark phase seals vary widely in response to influences not well understood.

5
At birth, ringed seal are approximately 60‐65 cm in length and weigh 4.5‐5.0 kg with regional variation (Chapskii 1940, McLaren 1958a, Tikhomirov 1968, Fedoseev 1975, Smith and Stirling 1975, Lydersen et al. 1992). The pups are born with a white natal coat (lanugo) which is shed after 4‐6 weeks (Figure 1). They nurse for as long as 2 months in stable, shorefast or lake ice (Chapskii 1940, McLaren 1958a) and for as little as 3‐6 weeks in moving ice (Burns 1970, Fedoseev 1975). At weaning, pups are four times their birth weights (Hammill et al. 1991, Lydersen et al. 1992) and lose weight for several months after weaning (Smith 1987). In a sample of 24 one‐year‐old seals from the Canadian Arctic, the average lengths of males and females were 103 cm and 94 cm, respectively (McLaren 1958a). In a sample of unspecified size from the Bering and Chukchi Seas, the average length of one‐year‐old seals was 86.3 cm (Frost and Lowry 1981).
Figure 1. ‐‐ Recently molted Arctic ringed seal pup.
2.3 Behavior
Time spent in and out of the water by ringed seals varies regionally, although some general patterns are apparent. Except during the spring molt, Arctic ringed seals spend most of their time under the ice or concealed in subnivean lairs on top of the ice (Smith and Stirling 1975, Kelly et al. 2010). In the Beaufort and Chukchi Seas along the coast of Alaska, Kelly et al. (2010) found that ringed seals spent 10% or less of their time on the ice during August to November. Time out of the water increased—but remained less than 20%—during December to March and increased to an average of 55% when the seals basked on
NOAA photo by Shawn Dahle

6
the ice while molting in May and June. In northern Baffin Bay, Teilmann et al. (1999) reported that ringed seals were out of the water for an average of 2.3% of the time during late August to late December for an average of about 3 hours, and Born et al. (2002) found that percent time out of the water per month ranged between 4‐16% during November to January. In the Baltic Sea, ringed seals showed an unusual pattern of increasing time out of the water during the late summer and winter, spending up to 40‐50% of their time on the ice during August‐September and 60% in January, while their diving activity peaked during June‐July (Harkonen et al. 2008). Arctic ringed seals use sea ice as a platform for resting throughout the year, although they have been observed on offshore islands and sand bars in the White Sea during summer months (Lukin et al. 2006). In the Sea of Okhotsk (Krylov et al. 1964), Baltic Sea (Harkonen et al. 2008), Lake Saimaa (Hyvärinen et al. 1995), and Lake Ladoga (Sipilä et al. 1996), ringed seals also rest on rocky reefs, islands, or shorelines during the ice‐free periods of the year.
Ringed seals are uniquely adapted to remaining in heavily ice‐covered areas throughout the fall, winter, and spring by using the stout claws on their foreflippers to maintain breathing holes in the ice. As the spring pupping season approaches, ringed seals excavate lairs in the snow cover over their breathing holes, and they rest on the ice within those lairs. Adult females also whelp and nurse their young in subnivean lairs. In Lake Saimaa, ringed seals typically make their lairs where the ice meets the shorelines of small islands, the only place where snow accumulates deep enough to build lairs (Helle et al. 1984, Sipilä 1990). In Lake Ladoga, ringed seals make their lairs on both the shorelines of small islands and on the drifting ice near the middle of the lake (Sipilä et al. 1996, Kunnasranta et al. 2001). In April to mid‐May, Arctic ringed seals rested in lairs mostly at night for a median duration of 6 hours, and the median interval between on‐ice bouts was 27 hours (Kelly et al. 2010). Ringed seal pups are more aquatic than other ice‐associated northern phocid pups, spending about 50% of their time in the water during the nursing period, diving for up to 12 minutes and as deep as 89 m (Lydersen and Hammill 1993b). The average duration of the pups’ on‐ice bouts was 6.3 hours and the time between on‐ice bouts averaged 8.2 hours (Lydersen and Hammill 1993b). The pups’ large proportion of time spent in the water, early development of diving skills, use of multiple breathing holes and nursing/resting lairs, and prolonged lanugo stage were interpreted as adaptive responses to strong predation pressure, mainly by polar bears (Ursus maritimus) and Arctic foxes (Alopex lagopus) (Smith et al. 1991, Lydersen and Hammill 1993b). Ringed seal mothers have a strong bond with their pups and may physically move their pups from the birth lair to an alternate lair to avoid predation, sometimes risking their lives to defend their pups from polar bears or human hunters (Smith 1987).
The breeding system of ringed seals is not well known, however, males are often described as territorial and the breeding system as polygynous. During the spring rut, adult males produce a strong scent, reminiscent of gasoline, and such males are referred to as tigak by Canadian Inuit. The odor clings to the snow at their breathing holes and lairs and is believed to be used in territorial marking (Smith 1987, Hardy et al. 1991, Ryg et al. 1992). Observations of segregated age classes and disparate sex ratios (2.1 to 2.4 females per male) in fast‐ice breeding areas (Smith 1987, Krafft et al. 2007), aggressive behavior and bite wounds on the bodies of primarily adult and subadult males (Smith and Hammill 1981, Smith 1987), and increased underwater vocalizations during the breeding season (Stirling et al. 1983, Rautio et

7
al. 2009) have been cited as evidence of territorial behavior and limited polygyny. Telemetric tracking of ringed seals in the vicinity of Resolute Bay, Northwest Territories, Canada (Kelly and Wartzok 1996) and north of the Mackenzie River delta (L. Harwood, Department of Fisheries and Oceans [DFO], Canada, pers. comm., 9 July 2010) showed that adult males did not dive as deeply as adult females and subadult males during the breeding season suggesting that the breeding males were guarding territories or mates near the undersurface of the ice. Kelly et al. (2010) reported that the above and under‐ice ranges of females were larger than those of males during the breeding season and suggested that the restricted movements of the males indicates that they were guarding the primary breathing hole of one female until she was receptive, yielding a monogamous or perhaps mixed breeding system rather than polygyny. Krafft et al. (2007) found that adult males with the most adult female neighbors were significantly older but not larger than adult males with fewer female neighbors and suggested that experience (age) likely plays a strong role in achieving reproductive success. The mating act has not been observed in the wild, but presumably occurs under the ice in the vicinity of the pupping lair (Kelly 1988).
Numbers of ringed seals visible on the surface of the ice typically begin to increase during spring as temperatures begin to warm and the snow covering the seals’ lairs melts. In the Sea of Okhotsk, ringed seals are first seen on the ice at the end of March or early April (Tikhomirov 1961, Heptner et al. 1976). In Lakes Saimaa and Ladoga, ringed seals begin appearing on the ice in late April, but most of the lake seals molt on rocks and island shorelines in May to mid‐June (Kunnasranta et al. 2002). In the Beaufort and Chukchi Seas, time spent on the ice increased from 12% in March to 43% in early June, and most seals began basking on the ice in late May or June (Kelly and Quakenbush 1990). Peak numbers of basking seals are typically observed in June in the Arctic during the seals’ annual molt (Smith and Hammill 1981, Smith 1987, Born et al. 2002, Moulton et al. 2002, Carlens et al. 2006, Harwood et al. 2007, Kelly et al. 2010). Factors affecting time out of the water during this period include stage of snow melt, time of day, air temperature, and wind speed with peak numbers of seals out of the water around mid‐day on warm, calm days (Finley 1979, Smith and Hammill 1981, Smith 1987, Moulton et al. 2002, Carlens et al. 2006, Kelly et al. 2006). Carlens et al. (2006) reported that the duration of bouts out of the water varied by age class during the molting period, with pups coming out for short but frequent bouts and older seals out for longer periods, sometimes lasting more than 24 hours (Max. = 141 hours). Born et al. (2002) found that time out of the water increased in June from about 25% to about 57%, and Kelly et al. (2010) reported that ringed seals spent more than 60% of their time on the ice in June, but that dropped to 30% in July. The median duration of bouts out of the water was 9 hours during the molting period and the intervals between bouts were about 14 hours (Kelly et al. 2010). While basking on the ice, ringed seals are vigilant and typically are out singly or in small groups at the edge of a hole or in the center of large ice floes, often facing downwind towards the hole and oriented broadside to the sun (Finley 1979, Kingsley and Stirling 1991). As the sea ice breaks up during the summer and molting nears completion, ringed seals spend much of their time in the water feeding again.
Tagging studies revealed that ringed seals are capable of diving for at least 39 minutes (Teilmann et al. 1999) and to depths of over 500 m (Born et al. 2004), however, most dives reportedly lasted less than 10 minutes and dive depths were highly variable and were often limited by the relative shallowness of the areas in which the studies took place (Lydersen 1991, Kelly and Wartzok 1996, Teilmann et al. 1999,

8
Gjertz et al. 2000, Kunnasranta et al. 2002, Harkonen et al. 2008). Diving capacity has been shown to be related to body mass, with larger individuals being capable of diving deeper and longer (Kelly and Wartzok 1996, Kelly 1997, Teilmann et al. 1999, Kunnasranta et al. 2002). Based on three‐dimensional tracking, Simpkins et al. (2001) categorized ringed seal dives as either travel, exploratory, or foraging/social dives. Hyvärinen et al. (1995) found that ringed seals in Lake Saimaa made long duration diving bouts during the summer and fall, which they suggested were resting dives used to conserve energy and add more weight during the seals’ intensive feeding period. Ringed seals tend to come out of the water during the daytime and dive at night during the spring to early summer breeding and molting periods, while the inverse tended to be true during the late summer, fall, and winter (Kelly and Quakenbush 1990, Lydersen 1991, Teilmann et al. 1999, Kunnasranta et al. 2002, Carlens et al. 2006, Kelly et al. 2010). Captive diving experiments conducted by Elsner et al. (1989) indicated that ringed seals primarily use vision to locate breathing holes from under the ice, followed by their auditory and vibrissal senses for short‐range pilotage. Hyvärinen (1989) suggested that Lake Saimaa ringed seals may use a simple form of echolocation along with a highly developed vibrissal sense for orientation and feeding in the lake’s dark, murky waters. The vibrissae likely are important in detecting prey by sensing their turbulent wakes as demonstrated experimentally for harbor seals (Dehnhardt et al. 1998).
2.4 Seasonal Distribution, Habitatuse, and Movements
Ringed seals are circumpolar (Figure 2) and are found in all seasonally ice‐covered seas of the Northern Hemisphere as well as in certain freshwater lakes (King 1983) . They range southward to Newfoundland and the Baltic Sea in the North Atlantic Ocean and are found in the southern Bering Sea, the Sea of Okhotsk, and the Sea of Japan in the western North Pacific Ocean. They are found in Nettilling Lake on Baffin Island (Mansfield 1967), and landlocked populations inhabit Lakes Saimaa and Ladoga east of the Baltic Sea (Ognev 1935).
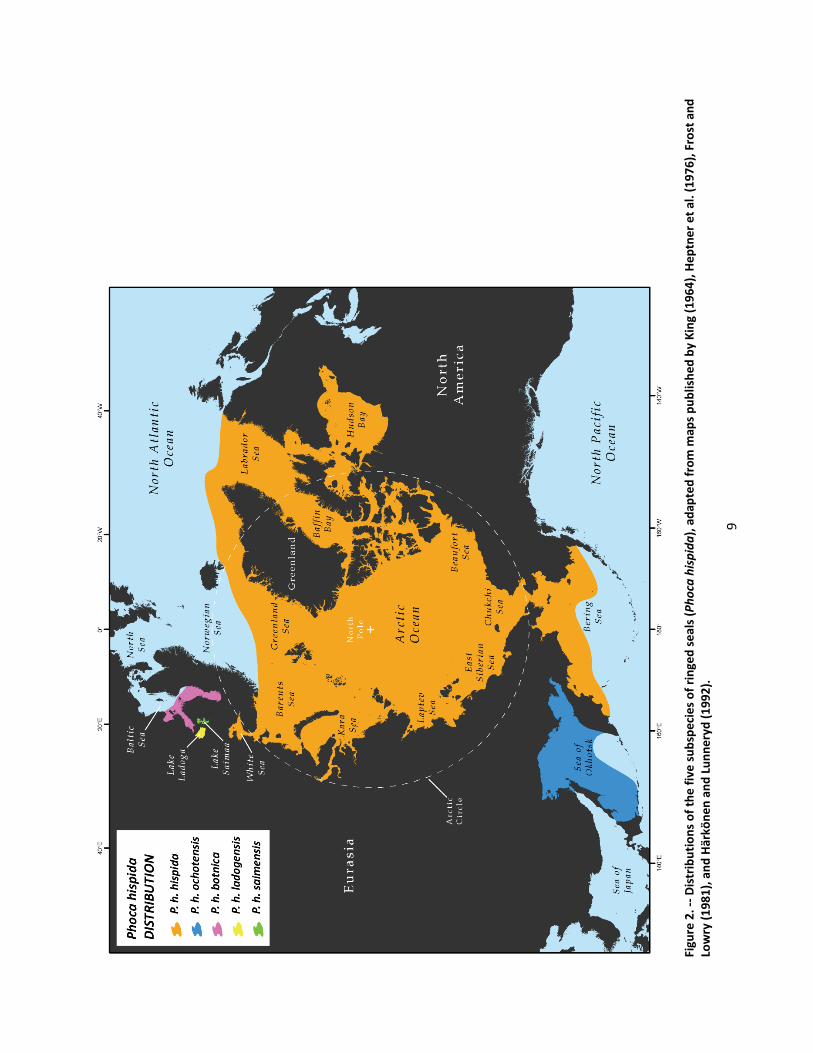
9
Figure 2. ‐‐ D
istributions of the
five sub
species of ringed seals (Pho
ca hispida
), ad
apted from
map
s pu
blishe
d by
King (1964), H
eptner et a
l. (1976), Frost and
Lowry (1
981), and
Härkö
nen an
d Lunn
eryd
(1992).

10
In the Arctic Ocean, marginal seas, and lakes, ringed seals have an affinity for ice‐covered waters and are able to occupy areas of even continuous ice cover by abrading breathing holes in that ice (Hall 1865, Bailey and Hendee 1926, Chapskii 1940, McLaren 1958a). Throughout most of its range, the Arctic subspecies does not come ashore and uses sea ice as a substrate for resting, pupping, and molting (Kelly 1988, Kelly et al. 2010; T. G. Smith, Eco Marine Corporation, pers. comm., 9 January 2010). In more southerly regions including the White Sea, the Sea of Okhotsk, and the Baltic Sea, ringed seals occasionally rest on island shores or offshore reefs in the ice‐free season (Ognev 1935, Härkönen et al. 1998, Trukhin 2000, Lukin et al. 2006). In Lakes Ladoga and Saimaa, ringed seals typically rest on rocks and island shores during the ice‐free season (Kunnasranta 2001).
Throughout most of the range, ringed seals give birth in late winter‐early spring in subnivean lairs on the sea ice (Arctic Ocean and Baltic Sea subspecies) or where the ice meets the shoreline (Lakes Saimaa and Ladoga). The Sea of Okhotsk subspecies is exceptional in that pups apparently are born primarily in moving pack ice both within subnivean lairs and in the open, typically in the lee of ice hummocks (Ognev 1935, Krylov et al. 1964, Fedoseev 1965b, Fedoseev 1975, Heptner et al. 1976). Based on hunters’ reports, Krylov et al. (1964) suggested that only those Okhotsk ringed seals breeding in shorefast ice used subnivean lairs. Observations of seals in the moving ice of the Okhotsk Sea have been limited mainly to observations from ice breaking ships, and Fedoseev (1965b) noted that it would be difficult to detect lairs from shipboard. Of more than 900 Okhotsk ringed seals that he observed from a ship during the pupping season, only three were pups. Heptner et al. (1976) pointed out that lairs “can be detected only with the help of a dog.” We are not aware of any attempts to locate subnivean lairs using dogs in the Sea of Okhotsk, and the extent to which ringed seals there rely on lairs is not known.
Today, the Baltic Sea is brackish and low salinity strongly influences its ecology (von Storch and Omstedt 2008). Whelping, nursing, and molting takes place on the sea ice, but during the ice‐free season, ringed seals do rest—especially in the southern portion of their Baltic Sea range—on offshore islets (Härkönen et al. 1998). Similarly, in the southern Sea of Okhotsk, ringed seals have recently been observed coming ashore on islands (Trukhin 2000).
The seasonality of ice cover strongly influences ringed seal movements, foraging, reproductive behavior, and vulnerability to predation. Born et al. (2004) recognized three “ecological seasons” as important to ringed seals off northwestern Greenland: the “open‐water season” in August‐October; the ice‐covered “winter” in November‐March; and “spring” in April‐July, when the seals breed and molt on the ice surface. Similarly, tracking seals in Alaska and the western Canadian Arctic, Kelly et al. (2010) referred to the open‐water period when ringed seals forage most intensively as the “foraging period”, early winter through late May to early June when seals rested primarily in subnivean lairs on the ice as the “subnivean period”, and the period between abandonment of the lairs (May or June) and ice break‐up (typically June or July) as the “basking period.”
2.4.1 Openwater period
Short and long distance movements by ringed seals in the open‐water period were suggested by observations of annual movements through Bering Strait (Bailey and Hendee 1926) and past Herschel

11
Island in the eastern Beaufort Sea (Smith et al. 1973) and by recoveries of tagged seals. Two seals tagged as juveniles in the Herschel Island area were subsequently collected in the same vicinity while 10 others tagged in the Canadian Beaufort Sea including at Herschel Island were tracked to (and in some cases recovered in) the Chukchi Sea (Smith 1976, Smith and Stirling 1978, Harwood and Smith 2003). A yearling ringed seal tagged in Resolute Passage in the Canadian Arctic Archipelago was harvested near the coast of southwestern Greenland (Kelly and Wartzok 1996, Kapel et al. 1998).
Satellite‐linked transmitters glued to the pelage have allowed ringed seals to be tracked for several months between tagging and the subsequent annual molt. One adult and three juvenile ringed seals were tracked for periods ranging from 2 weeks to 6 months off northwestern Greenland (Heide‐Jørgensen et al. 1992b) in summer and autumn. They remained within the fjord where they had been tagged until the ice broke up and then moved a few to over 200 km away. Similar transmitters were glued to seven adult ringed seals at Svalbard in early July, after the annual molt (Gjertz et al. 2000). Over the next 3‐11 months, those seals mostly made “local movements” near the Svalbard coast, but two ranged 400 km north to the pack ice before returning to the fjord where they were captured.
Seven juveniles and one adult ringed seal were tracked using satellite‐linked transmitters in the vicinity of the North Water polynya in Baffin Bay (Teilmann et al. 1999). The seals were tagged in fjords of northwestern Greenland in August and September and tracked for 1 to almost 5 months. During that period, the seals were mainly associated with shorefast ice where prey densities apparently were highest. Born et al. (2004) extended the observations of Teilmann et al. (1999) to include an additional 15 ringed seals tracked by satellite in the North Water polynya for up to 11 months. They confirmed the observations that the seals were concentrated in shallower waters, spending 90% of their time in water less than 100 m deep and that ringed seals preferentially exploited areas of lighter ice within the polynya. They recorded home ranges of 10,300‐18,500 km2 in the open‐water season. Eight ringed seals tagged in Canada’s Amundsen Gulf used similar sized open‐water home ranges as those tagged in the North Water study, ranging from 9,000‐18,000 km2 (L. Harwood, DFO, Canada, pers. comm., 9 July 2010).
Harkonen et al. (2008) tracked 19 adult ringed seals in the Baltic Sea for periods of 2 to 9 months. Those seals all remained within a few hundred kilometers of their capture sites and were clustered in three regions—Bothnian Bay, Gulf of Finland, and Gulf of Riga—taken by the authors to reflect distinct stocks.
Freitas et al. (2008) used satellite tracking to quantify at‐sea habitat selection and emphasized variation among individuals in their autumn and winter movements. They tagged 22 seals (17 juvenile and 4 mature) at Svalbard in July and reported on their movements for up to 9 months but with an emphasis on the period August‐October. They documented two main foraging strategies in which seals either moved away from their winter areas to the sea‐ice edge or remained close to winter areas at glacier fronts. Those that associated with sea ice showed a preference for ice concentrations of 40‐80% indicative of the ice edge. The authors suggested that both strategies—frequenting the sea‐ice edge or glacier fronts—provided access to food‐rich waters as well as to on‐ice resting sites. They speculated that the value of resting on the ice outside of the breeding or molting periods may relate to reducing thermal stress and minimizing predation, perhaps from Greenland sharks (Somniosus microcephalus).

12
Kelly et al. (2010) attached satellite‐linked transmitters to the hindflippers of seals (instead of gluing to the pelage) in order to extend tracking beyond the subsequent molt. They captured 25 ringed seals at four sites in the shorefast ice of the Chukchi and Beaufort Seas. The seals were captured in March to early June and tracked for up to 14 months. After the ice broke up in July, the seals moved offshore to moving ice. Nine seals were tracked throughout the year and—in July through December—6 of those moved to pack ice within 200 km of their tagging sites and 3 to pack ice 800 km or more from their tagging sites (including one that ranged almost 1,800 km). By the subsequent January, 8 of the 9 seals returned to within 55 km of the sites at which they had been captured during the previous breeding season. The ninth seal, an adult male tagged on shorefast ice in May, moved to a pack‐ice site 1,000 km to the west in August, returned to his tagging site in October, traveled 800 km east in November, and was back at his shorefast ice tagging site the following June.
Overall, the record from satellite tracking indicates that ringed seals breeding in shorefast ice practice one of two strategies during the open‐water foraging period (Freitas et al. 2008). Some forage within 100 km of their shorefast ice breeding habitat while others make extensive movements of 100s or 1,000s of kilometers to forage in highly productive areas (e.g., Viscount Melville Sound) and along the pack‐ice edge. Movements during the foraging period by ringed seals that breed in the pack ice are unknown. At the end of the foraging period, adult Arctic ringed seals return to the same sites used during the previous subnivean period (Smith and Hammill 1981, Krafft et al. 2007, Kelly et al. 2010). Saimaa and Ladoga ringed seals show similar site fidelity (Sipilä et al. 1996, Kunnasranta 2001, Koskela et al. 2002).
Just prior to freeze up, large groups of ringed seals frequently feed on dense schools of cod in near shore areas of Amundsen Gulf and Prince Albert Sound, Beaufort Sea (Smith 1987). In offshore areas of the Beaufort Sea and Amundsen Gulf, large, loose feeding aggregations of ringed seals have also been documented in the late summer and early fall (Harwood and Stirling 1992). High quality, abundant food is important to the annual energy budgets of ringed seals. Fall and early winter periods, prior to the occupation of breeding sites, are important in allowing ringed seals to accumulate enough fat stores to support estrus and lactation.
2.4.2 Subnivean period
Movements during the subnivean period typically are quite limited, especially where ice cover is extensive (Kelly and Quakenbush 1990). Harwood et al. (2007) tracked 20 ringed seals by satellite in the fast ice off the Mackenzie estuary/Yukon North Slope of Canada during the breeding seasons of 2005 and 2006. The seals’ movements were limited, and 95% of their surface locations were within areas averaging 14 km2. Similar patterns were found in Alaskan studies, where seals were tracked in the Beaufort Sea. Based on VHF radio tracking (58 seals) in the shorefast ice of the Alaskan Beaufort Sea and ultrasonic tracking (9 seals) beneath the ice in Barrow Strait, Canada, Kelly et al. (2010) measured smaller breeding home ranges. Ranges averaged less than 1 km2 above the ice and less than 0.15 km3 below the ice. Ringed seals with access to a large polynya (North Water), on the other hand, ranged 2,500‐7,000 km2 in winter and 800‐2,100 km2 in spring (Born et al. 2004). Similar to the North Water study, adult seals tracked in Canada’s Amundsen Gulf used winter home ranges from 3,400‐4,600 km2.

13
Despite this extensive winter home range, one adult female’s range was limited to 167 km2 for the month of February—presumably during lair construction and prior to parturition (L. Harwood, DFO, Canada, pers. comm., 9 July 2010).
At freeze up in fall, ringed seals surface to breathe in the remaining open water of cracks and leads. As these openings freeze over, the seals push through the ice to breathe until it is too thick (Lukin and Potelov 1978). They then open breathing holes by abrading the ice with the claws of their foreflippers (Bailey and Hendee 1926, Smith and Stirling 1975). As the ice thickens, the seals continue to maintain the breathing holes by scratching at the walls. The breathing holes can be maintained in ice 2 m or greater in thickness but often are concentrated in the thinner ice of refrozen cracks (Hammill and Smith 1989). The holes generally are cone‐shaped (Ognev 1935, Smith and Stirling 1975, Lukin and Potelov 1978) and approximately 30 cm in diameter at the upper ice surface and 76 cm at the lower surface (Kelly 1996).
As snow accumulates and buries the breathing hole, the seals breathe through the snow layer. Ringed seals excavate lairs in the snow above breathing holes where snow depth is sufficient (Chapskii 1940, McLaren 1958a, Smith and Stirling 1975). Subnivean lairs are occupied for resting, whelping, and nursing young in shorefast ice (McLaren 1958a, Burns 1970) and in pack ice (Finley et al. 1983, Wiig et al. 1999, Bengtson et al. 2005). Useable lairs are excavated in snow depths greater than or equal to 45 cm, which are rare except where snow has drifted next to a pressure ridge or ice hummock (Smith and Stirling 1975, Lydersen and Gjertz 1986, Kelly 1988, Furgal et al. 1996, Lydersen 1998). A general lack of such ridges or hummocks in Lakes Ladoga and Saimaa limits snow drifts to shorelines, where most lairs in Lake Ladoga and virtually all in Lake Saimaa are found (Sipilä and Hyvärinen 1998). Active lairs vary greatly in size and complexity with internal heights ranging from less than 30 to 70 cm and lengths typically ranging from 1 to 2 m but occasionally up to 5 m (Smith and Stirling 1975, Burns and Kelly 1982, Lydersen and Gjertz 1984, Furgal et al. 1996). Subnivean lairs protect ringed seals from extreme cold and from predators. Unoccupied lairs are heated by the water below (Lukin 1980), and—in a study in the Chukchi Sea—the air temperature in unoccupied subnivean lairs ranged between ‐3° and ‐10°C while effective outside air temperatures (including wind chill effects) ranged from ‐7° to ‐61°C (Kelly 1988). When occupied by a seal, the air temperature in the same lairs ranged between ‐5° and +5°C (Avg. = +0.7°C, SD = 2.19). When dry, ringed seal pups in lanugo can maintain thermal neutrality at temperatures above ‐25°C and, when wet, can regain thermal neutrality at temperatures above ‐8°C (Taugbøl 1982, Smith et al. 1991). Thus, subnivean lairs provide refuge from air temperatures too low for survival of ringed seal pups.
Lairs also conceal ringed seals from predators (see Section 4.2.3.3 ), an advantage especially important to the small pups that start life with minimal tolerance for immersion in cold water (Smith et al. 1991). When forced to flee in to the water to avoid predators, the pups depend on the subnivean lair to subsequently warm themselves. Offshore of southeastern Baffin Island, low snow depths were associated with increased predation attempts (y = ‐0.613, p = 0.02, N = 14) and successful predation (y = ‐0.896, p = 0.04, N = 5) on ringed seal pups by polar bears (Hammill and Smith 1991). When warm temperatures and rain at southeastern Baffin Island caused lairs to collapse, the success rate of bear

14
attacks on ringed seals tripled from 11.3% to 33.5%. Similarly, a decrease from 23 cm to 10 cm in average snow depth in Barrow Strait, Canada, in the mid‐1980s coincided with an increase in the predation rate by polar bears from 0.1 to 0.4 seals/km2 (Hammill and Smith 1991).
Occasionally, lairs are not excavated in snow but consist of natural cavities occurring where ice has buckled and rafted under pressure. Lairs in such ice cavities may afford better protection from predators yet present less insulation and greater danger because of the instability of such ice (Chapskii 1940, McLaren 1958a).
2.4.3 Basking period
The warmer temperatures and long hours of sunlight in late spring and early summer rapidly melt the snow cover, but the ice remains largely intact and serves as a resting substrate for the molting seals that spend many hours basking in the sun (Smith 1973, Finley 1979). Seals of the Arctic subspecies spend over 60% of their time on the ice in June when they are actively regenerating epidermal tissues (skin and hair) (Kelly et al. 2010). As seals complete that phase of the annual pelage cycle, they spend increasing amounts of time in the water. In July, the proportion of time on the ice in the Alaskan Beaufort Sea dropped to 30% (Kelly et al. 2010). Activity among polar bears increases during that time as the bears hunt seals visible on the ice surface (Stirling 1974, Messier et al. 1992).
2.5 Reproduction and Molting
Ringed seals breed annually, and males in the Arctic populations rut from late March to mid‐May, occasionally to mid‐June, and rarely even later (McLaren 1958a). Timing varies regionally, however, and the rut peaks in February in the Baltic Sea (Bakulina 1989). Arctic females ovulate in May and early June (Johnson et al. 1966, Smith 1973, Smith 1987). Mating is thought to take place under the ice in the vicinity of the pupping lair. The mating system is debated. Stirling (1977) inferred a monogamous or nearly monogamous breeding system with territoriality from an even adult sex ratio, predation pressure, evidence of inter‐male aggression, and presumed aggression among females. Smith and Hammill (1981) suggested a weakly polygynous system based on an odiferous secretion by males during the rut (Ognev 1935, Chapskii 1940, Hardy et al. 1991), an inferred under‐ice territoriality, increased vocalization rates during the breeding season (Stirling 1973, Stirling et al. 1983, Rautio et al. 2009), and evidence of site fidelity. Krafft et al. (2007) suggested a territorial system with weak polygny based on the locations of seals when they were killed and a sex ratio skewed slightly toward females. Tracking the movements of adults seals above and below the ice throughout the breeding season, however, showed that female ranges were larger than those of males, and Kelly et al. (2010) suggested that considerable overlap in areas and holes used by females as well as limited movements and diving by males were more consistent with a mate guarding strategy than with territoriality.
Mating takes place typically in May (Arctic subspecies) shortly after parturition while mature females are still lactating (Figure 3). Fertilization is followed by 3 to 3.5 months of arrested development before the blastocyst implants (McLaren 1958a, Fedoseev 1975, Smith 1987). Following implantation, active

15
gestation lasts approximately 240 days. A single pup (rarely twins) is born in a subnivean lair on either shorefast ice (McLaren 1958a) or pack ice (Finley et al. 1983, Kelly 1988). In much of the Arctic, whelping occurs in late March through April—when snow depth is maximal (Weeks 2010)—but the timing varies with latitude (Freuchen 1935, Chapskii 1940, McLaren 1958a, Smith 1987, Lydersen 1998, Sinisalo et al. 2008). Lukin and Potelov (1978) give February as the birthing season in the White Sea, but more recently, Lukin et al. (2006) stated that “most females give birth during the first ten days of March.” Belikov and Boltunov (1998), however, give the birthing season as mid‐March through mid‐April or later in the White Sea and the southern Barents Sea. Heptner et al. (1976) gave the pupping season in the Kara Sea as April. In the Baltic Sea, Lake Saimaa, and Lake Ladoga pups are born in February‐March (Fedoseev 1972, Helle 1979a, Sipilä and Hyvärinen 1998, Kunnasranta et al. 2001, Sinisalo et al. 2008, Rautio et al. 2009). Pupping in the Sea of Okhotsk takes place in March and April based on observations of pups in lanugo (Tikhomirov 1961, Fedoseev 1965b).
The pups are nursed in the lairs for an average of 39 days (Hammill et al. 1991) but with considerable variation. Fedoseev (1975) reported nursing periods of “little more than three week” for the Sea of Okhotsk and up to 6 weeks in the Bering Sea, a difference he attributed to breeding in pack ice in the Sea of Okhotsk and in shorefast ice in the Bering Sea. Saimaa ringed seals nurse for 7‐9 weeks (Käkelä and Hyvärinen 1993). Lydersen and Hammill (1993b) reported a nursing period of almost 8 weeks for one seal at Svalbard. At weaning the pups weigh about 20 kg (Lydersen et al. 1992, Lydersen and Hammill 1993a).
At or shortly after weaning, ringed seal pups shed their natal pelage (Kelly 1988, Lydersen and Hammill 1993b), the first of what becomes an annual molt. A ringed seal pup still in lanugo and photographed on 10 July 1968 in the northern Bering Sea (R. O. Stephenson, Alaska Department of Fish and Game (ADFG), pers. comm., 20 April 2010) presumably was born near the end of the birthing season. Adults generally molt from mid‐May to mid‐July (McLaren 1958a, Kunnasranta et al. 2002), although there seems to be regional variation. Ringed seals come out of the water for long periods of time during the molt (Smith 1973, Smith and Hammill 1981, Kelly and Quakenbush 1990, Kelly et al. 2010), a behavior ascribed to the need to maintain elevated skin temperatures. The cell divisions during epidermal regeneration require temperatures higher than can be achieved in the water (Feltz and Fay 1966). There is limited evidence that some ringed seals do not molt until autumn (Bychkov 1965), and Inuit on Baffin Island suggest that such seals were deprived of suitable ice in the usual spring molting season (McLaren 1958a). Feeding is reduced and the seal’s metabolism declines by an average 19% during the molt (Ashwell‐Erickson et al. 1986).

16
Figure 3. ‐‐ Approximate annual timing of reproduction and molting for the five subspecies of ringed seals. Yellow bars indicate the “normal” range over which each event is reported to occur and orange bars indicate the “peak” timing of each event.
2.6 Vital Parameters
At birth, the sex‐ratio among ringed seals typically is 1:1 (McLaren 1958a, Smith 1973, Fedoseev 1975, Helle 1980a, Frost and Lowry 1981, Lydersen and Gjertz 1987, Sipilä et al. 1990, Holst et al. 1999, Sipilä et al. 1999), which is purportedly maintained across all age groups (Frost and Lowry 1981).
Ringed seals generally attain sexual maturity at ages 4‐8 for females and at 5‐7 for males but with geographic and temporal variability depending on animal condition and population structure (McLaren
A. Arctic ringed seals
Adults
Pups
B. Okhotsk ringed seals
Adults
Pups
C. Baltic, Ladoga, and Saimaa ringed seals
Adults
Pups
July
Moltin
g
Breeding
Nursing
Whelping
Month February March April May June
July
Moltin
g
Breeding
Nursing
Whelping
Month February March April May June
July
Moltin
g
Breeding
Nursing
Whelping
Month February March April May June

17
1958a, Tikhomirov 1968, Smith 1973, Lydersen and Gjertz 1987, Kelly 1988, Reeves 1998, Sipilä and Hyvärinen 1998, Sipilä et al. 1999). Historical information on the age of sexual maturation (defined as the “age at first pregnancy”) was summarized by Frost and Lowry (1981, Table 2). Females mature at 6‐10 years and males at 5‐7 years (McLaren 1958a, Mansfield 1967, Burns and Fay 1970, Smith and Stirling 1975) in the Chukchi Sea and Arctic Ocean (Frost and Lowry 1981). Corresponding ages are 5‐7 and 6‐7 years in the Sea of Okhotsk (Fedoseev 1975) and 5 and 3‐4 years in the Baltic Sea (Curry‐Lindahl 1975). There is some doubt, however, regarding male maturation at 3‐4 years in the Baltic (T. Härkönen, Swedish Museum of Natural History, pers. comm., 8 March 2010). Holst et al. (1999) reported the average age of sexual maturation for females in Hudson Bay was 5.7 years, which is within the range reported for other areas. In Lake Saimaa, females reach sexual maturity at 4‐5 years and males at 4+ years (McLaren 1958a, Sipilä and Hyvärinen 1998). Similar values were reported by Popov (1979) for Lake Ladoga (i.e., ages 4‐5 for females and 6‐7 for males).
Reeves (1998) summarized published reproductive rates (defined as the “proportion of mature females that are either pregnant or have a corpus luteum of ovulation in any given year”) which ranged between 0.45 in Barrow Strait (Hammill 1987), 0.54 in Amundsen Gulf, 0.62 in Home Bay, Baffin Island (Smith 1987), 0.63 in the Choska Inlet (Nazarenko 1965), 0.78 near southern Baffin Island (McLaren's data cited in Hammill 1987), 0.81 in the Barrow Strait region (Smith et al. 1979), and 0.86 near Cape Thompson, Alaska (Johnson et al. 1966). A very low value of 0.28 for Bothnian Bay in the Baltic Sea (Helle 1980a) was related to uterine pathologies that have not been reported in other ringed seal populations (Reeves 1998). Hammill (1987) standardized several of these estimates by excluding animals less than 8 years old. The revised estimates were 0.63 for Barrow Strait, 0.75 for Amundsen Gulf, 0.77 for Home Bay, Baffin Island, and 0.81 for southern Baffin Island (Reeves 1998). Fedoseev (1975) reported a reproductive rate of 0.21 for populations in the Sea of Okhotsk, Bering Sea, and Chukchi Sea, but Reeves (1998) noted that the estimate is questionable due to lack of details on the sample sizes and methods. Sipilä et al. (1990) reported a pregnancy rate of ca. 0.70 in the Lake Saimaa population during the early 1980s, and Sipilä (2003) reported a range between 0.75 and 0.83 in the 1990s. Age at maturation and reproductive rates are dynamic due to environmental, ecological, and anthropogenic factors (Lydersen and Gjertz 1987, Kelly 1988, Holst et al. 1999, Sipilä 2003, Kovacs 2007). Generally, reproductive senescence has not been described for ringed seals, although McLaren (1958a) described a 35+ year‐old female as senile.
The average life span of ringed seals is about 15‐28 years (Frost and Lowry 1981, Sipilä and Hyvärinen 1998, Holst et al. 1999, Sipilä et al. 1999), and the maximal lifespan is in excess of 43 and as old as 48 years (McLaren 1958a, Lydersen and Gjertz 1987). Holst et al. (1999) reported that the maximal ages for male and female ringed seals in Hudson Bay (35 and 32, respectively) was similar to Smith’s (1973) data for Davis Strait. In contrast, Helle (1980a) noted that the oldest male and female seals in Bothnian Bay were 40 and 37 years old, respectively, whereas in Svalbard (Lydersen and Gjertz 1987), they were 30 and 45 years old, respectively. Lydersen and Gjertz (1987) noted that little information was available on the mean age of ringed seal males and females in different areas.

18
Age composition of Russian harvests in the northern part of the Sea of Okhotsk (1960‐1965) indicate that all age classes (0‐25) were killed, but seals less than 10 years old comprised the majority of the kills (Fedoseev 1968). In the region of the Shantar Islands, seals 7‐14 years old were dominant in 1960 and 1961 catches, whereas in 1965, most of the seals were less than 10 years old (Fedoseev 1968).
Kelly (1988) summarized age‐specific mortality rates from published life tables (McLaren 1958b, Smith 1973, Lowry et al. 1982) derived from harvest data. Mortality rates for most age classes were taken from the smoothed age‐frequency distribution, but data for the youngest and oldest seals were interpreted subjectively, since the youngest seals were overrepresented in the harvest and samples of the oldest seals were quite small. Estimated mortality was 30‐41% in pups, dropped to about 10% by sexual maturity, and increased slowly after about 15 years of age (Kelly 1988).
2.7 Foraging and Diet
2.7.1 Marine environment
Ringed seals eat a wide variety of prey spanning several trophic levels (Table 1). Their diet has been well documented, especially in the marine environment (e.g., Chapskii 1940, Dunbar 1941, Pikharev 1946, McLaren 1958a, Fedoseev 1965c, Johnson et al. 1966, Nazarenko 1967, Söderberg 1971, Tormosov and Rezvov 1978, Lowry et al. 1980, Bradstreet and Cross 1982, Bradstreet and Finley 1983, Gjertz and Lydersen 1986, Smith 1987, Węsławski et al. 1994, Siegstad et al. 1998, Wathne et al. 2000, Holst et al. 2001, Stenman and Pöyhönen 2005, Dehn et al. 2007, Labansen et al. 2007). Most ringed seal prey is small and preferred prey tend to be schooling species that form dense aggregations (Kovacs 2007). The most common prey size is 5‐10 cm for fish and 2‐6 cm for crustaceans, with a maximal size typically of about 20 cm (Węsławski et al. 1994). Estimates of prey size may be biased, however, if ringed seals do not consume the hard and identifiable parts (e.g., bones and otoliths) of larger prey (Smith 1977). Smith (1977) reported two Inuit hunters finding a partially‐eaten, 127 cm long wolffish (Anarhichas sp.) in the breathing hole of a 121 cm long ringed seal in northern Canada, suggesting that large prey may be taken on occasion. Ringed seals rarely prey upon more than 10‐15 species in any one area, and not more than 2‐4 species are considered as important prey (Węsławski et al. 1994). Several studies found no sex‐related differences in the diet of ringed seals (Johnson et al. 1966, Lowry et al. 1980, Bradstreet and Finley 1983, Gjertz and Lydersen 1986, Holst et al. 2001). Labansen et al. (2007), however, reported that adult females ate smaller cod more often than adult males or juveniles. McLaren (1958a) and Gjertz and Lydersen (1986) detected no age‐related differences in prey composition, but several studies found that young ringed seals ate more crustaceans and/or less fish when compared with adults (Fedoseev 1965c, Lowry et al. 1980, Bradstreet and Cross 1982, Bradstreet and Finley 1983, Smith 1987, Siegstad et al. 1998). Regional differences in diet probably result from differences in prey availability, prey preferences, and oceanographic conditions such as water depth, productivity level, and presence of sea ice.
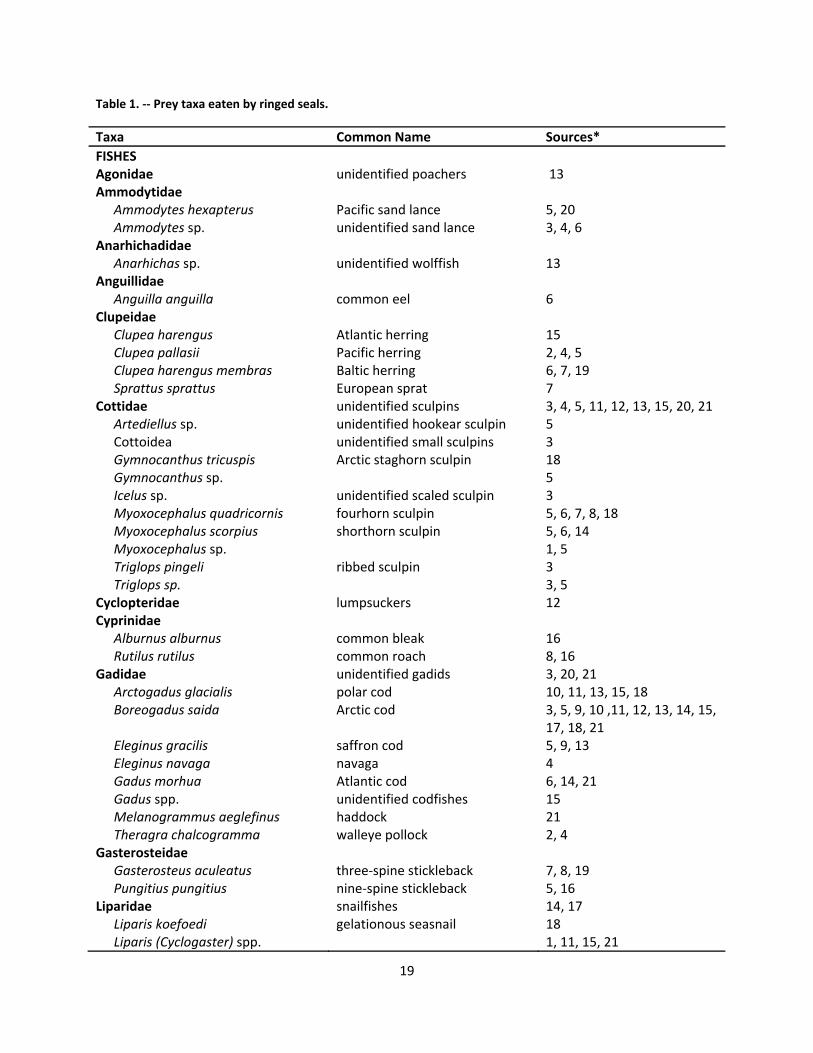
19
Table 1. ‐‐ Prey taxa eaten by ringed seals.
Taxa Common Name Sources* FISHES Agonidae unidentified poachers 13 Ammodytidae
Ammodytes hexapterus Pacific sand lance 5, 20 Ammodytes sp. unidentified sand lance 3, 4, 6
Anarhichadidae Anarhichas sp. unidentified wolffish 13
Anguillidae Anguilla anguilla common eel 6
Clupeidae Clupea harengus Atlantic herring 15 Clupea pallasii Pacific herring 2, 4, 5 Clupea harengus membras Baltic herring 6, 7, 19 Sprattus sprattus European sprat 7
Cottidae unidentified sculpins 3, 4, 5, 11, 12, 13, 15, 20, 21 Artediellus sp. unidentified hookear sculpin 5 Cottoidea unidentified small sculpins 3 Gymnocanthus tricuspis Arctic staghorn sculpin 18 Gymnocanthus sp. 5 Icelus sp. unidentified scaled sculpin 3 Myoxocephalus quadricornis fourhorn sculpin 5, 6, 7, 8, 18 Myoxocephalus scorpius shorthorn sculpin 5, 6, 14 Myoxocephalus sp. 1, 5 Triglops pingeli ribbed sculpin 3 Triglops sp. 3, 5
Cyclopteridae lumpsuckers 12 Cyprinidae
Alburnus alburnus common bleak 16 Rutilus rutilus common roach 8, 16
Gadidae unidentified gadids 3, 20, 21 Arctogadus glacialis polar cod 10, 11, 13, 15, 18 Boreogadus saida Arctic cod 3, 5, 9, 10 ,11, 12, 13, 14, 15,
17, 18, 21 Eleginus gracilis saffron cod 5, 9, 13 Eleginus navaga navaga 4 Gadus morhua Atlantic cod 6, 14, 21 Gadus spp. unidentified codfishes 15 Melanogrammus aeglefinus haddock 21 Theragra chalcogramma walleye pollock 2, 4
Gasterosteidae Gasterosteus aculeatus three‐spine stickleback 7, 8, 19 Pungitius pungitius nine‐spine stickleback 5, 16
Liparidae snailfishes 14, 17 Liparis koefoedi gelationous seasnail 18 Liparis (Cyclogaster) spp. 1, 11, 15, 21
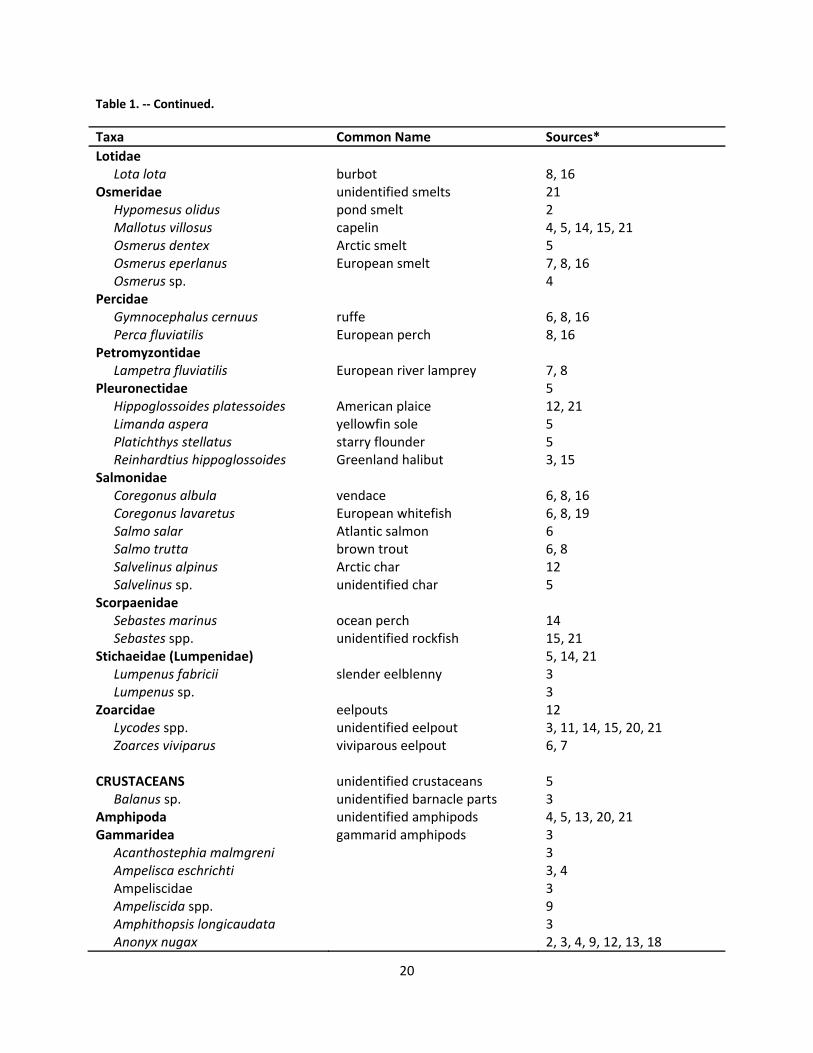
20
Table 1. ‐‐ Continued.
Taxa Common Name Sources* Lotidae
Lota lota burbot 8, 16 Osmeridae unidentified smelts 21
Hypomesus olidus pond smelt 2 Mallotus villosus capelin 4, 5, 14, 15, 21 Osmerus dentex Arctic smelt 5 Osmerus eperlanus European smelt 7, 8, 16 Osmerus sp. 4
Percidae Gymnocephalus cernuus ruffe 6, 8, 16 Perca fluviatilis European perch 8, 16
Petromyzontidae Lampetra fluviatilis European river lamprey 7, 8
Pleuronectidae 5 Hippoglossoides platessoides American plaice 12, 21 Limanda aspera yellowfin sole 5 Platichthys stellatus starry flounder 5 Reinhardtius hippoglossoides Greenland halibut 3, 15
Salmonidae Coregonus albula vendace 6, 8, 16 Coregonus lavaretus European whitefish 6, 8, 19 Salmo salar Atlantic salmon 6 Salmo trutta brown trout 6, 8 Salvelinus alpinus Arctic char 12 Salvelinus sp. unidentified char 5
Scorpaenidae Sebastes marinus ocean perch 14 Sebastes spp. unidentified rockfish 15, 21
Stichaeidae (Lumpenidae) 5, 14, 21 Lumpenus fabricii slender eelblenny 3 Lumpenus sp. 3
Zoarcidae eelpouts 12 Lycodes spp. unidentified eelpout 3, 11, 14, 15, 20, 21 Zoarces viviparus viviparous eelpout 6, 7
CRUSTACEANS unidentified crustaceans 5
Balanus sp. unidentified barnacle parts 3 Amphipoda unidentified amphipods 4, 5, 13, 20, 21 Gammaridea gammarid amphipods 3
Acanthostephia malmgreni 3 Ampelisca eschrichti 3, 4 Ampeliscidae 3 Ampeliscida spp. 9 Amphithopsis longicaudata 3 Anonyx nugax 2, 3, 4, 9, 12, 13, 18

21
Table 1. ‐‐ Continued.
Taxa Common Name Sources* Anonyx sarsi 13 Anonyx spp. 11, 21 Apherusa glacialis 3, 13 Atylus carinatus 3, 18 Calliopius laeviusculus 3 Eusirus cuspidatus 15 Gammaracanthus loricatus 1, 3, 11, 13, 15, 18 Gammarellus homari 3 Gammaridae 3 Gammarus locusta 1, 12 Gammarus oceanicus 3, 13 Gammarus schmidtii 2 Gammarus setosus 3, 14, 18 Gammarus wilkitskii 3, 11, 12, 13, 14, 15, 21 Gammarus spp. 1, 3, 9, 10, 11, 12, 21 Halirages nilssoni 3 Haploops setosa 3 Ischyrocerus anguipes 3 Ischyrocerus sp. 13, 14 Lysianassidae 3, 21 Melita sp. 14 Onisimus edwardsi 12, 18 Onisimus glacialis 1, 3, 11, 13, 21 Onisimus littoralis 3, 11, 13, 14, 18 Onisimus nanseni 1, 3 Onisimus (Pseudalibrotus) spp. 3, 10, 11 Orchomenella pinguis 3 Pardalisca cuspidata 3 Pontogeneia inermis 3 Rhachotropis aculeata 3, 12 Socarnes bidenticulatus 3 Stegocephalus inflatus 3, 12, 13, 21 Weyprechtia pinguis 3, 13
Hyperiidea hyperiid amphipods Hyperia galba 3, 18 Hyperia spinigera 1 Hyperia spp. 11 Parathemisto abyssorum 13 Parathemisto libellula 9, 11, 12, 13, 21 Parathemisto spp. 10, 11, 15 Themisto compressa 2 Themisto libellula 1, 3, 4, 14, 17, 18
Cumacea unidentified hooded shrimp 5 Decapoda unidentified decapods 1, 3, 4, 5, 11, 21 Natantia unidentified shrimp 3
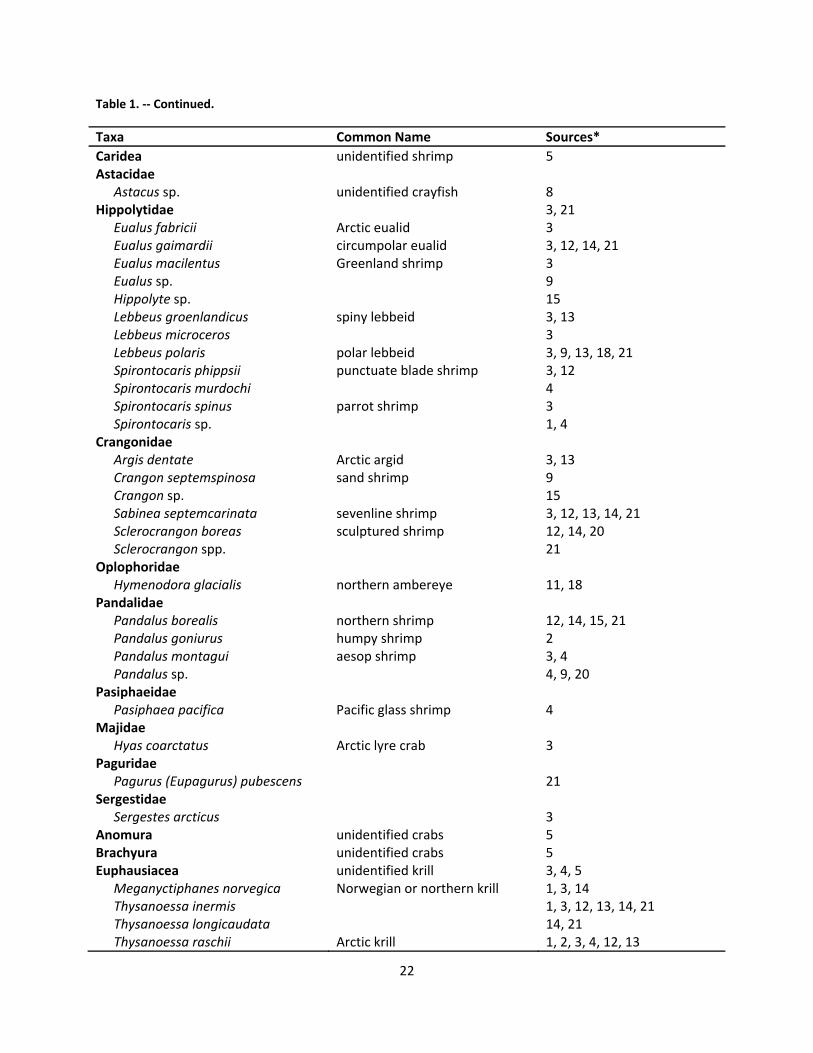
22
Table 1. ‐‐ Continued.
Taxa Common Name Sources* Caridea unidentified shrimp 5 Astacidae
Astacus sp. unidentified crayfish 8 Hippolytidae 3, 21
Eualus fabricii Arctic eualid 3 Eualus gaimardii circumpolar eualid 3, 12, 14, 21 Eualus macilentus Greenland shrimp 3 Eualus sp. 9 Hippolyte sp. 15 Lebbeus groenlandicus spiny lebbeid 3, 13 Lebbeus microceros 3 Lebbeus polaris polar lebbeid 3, 9, 13, 18, 21 Spirontocaris phippsii punctuate blade shrimp 3, 12 Spirontocaris murdochi 4 Spirontocaris spinus parrot shrimp 3 Spirontocaris sp. 1, 4
Crangonidae Argis dentate Arctic argid 3, 13 Crangon septemspinosa sand shrimp 9 Crangon sp. 15 Sabinea septemcarinata sevenline shrimp 3, 12, 13, 14, 21 Sclerocrangon boreas sculptured shrimp 12, 14, 20 Sclerocrangon spp. 21
Oplophoridae Hymenodora glacialis northern ambereye 11, 18
Pandalidae Pandalus borealis northern shrimp 12, 14, 15, 21 Pandalus goniurus humpy shrimp 2 Pandalus montagui aesop shrimp 3, 4 Pandalus sp. 4, 9, 20
Pasiphaeidae Pasiphaea pacifica Pacific glass shrimp 4
Majidae Hyas coarctatus Arctic lyre crab 3
Paguridae Pagurus (Eupagurus) pubescens 21
Sergestidae Sergestes arcticus 3
Anomura unidentified crabs 5 Brachyura unidentified crabs 5 Euphausiacea unidentified krill 3, 4, 5
Meganyctiphanes norvegica Norwegian or northern krill 1, 3, 14 Thysanoessa inermis 1, 3, 12, 13, 14, 21 Thysanoessa longicaudata 14, 21 Thysanoessa raschii Arctic krill 1, 2, 3, 4, 12, 13

23
Table 1. ‐‐ Continued.
Taxa Common Name Sources* Thysanoessa sp. 3, 9, 15, 17, 20
Isopoda unidentified isopods 4, 5 Idotea ochotensis 4 Saduria (Mesidothea) entomon 4, 6, 7, 13 Saduria (Mesidothea) sp. 2, 20 Synidotea nodulosa 21
Mysida unidentified mysids 4, 5 Mysidae 15 Mysis litoralis 11 Mysis oculata 1, 3, 11, 12, 13, 14, 18, 21 Mysis mixta 1, 3 Mysis relicta 6, 16 Mysis spp. 3, 9, 11, 20 Neomysis rayii 9 Neomysis spp. 20
Copepoda unidentified copepods 4, 13 Calanoida 3 Calanus finmarchicus 1, 3 Calanus hyperboreas 3, 13 Euchaeta glacialis 13 Metridia longa 1, 13 Paraeuchaeta norvegica 3
Ostracoda Philomedes globosus 13
MOLLUSCS unidentified molluscs 11 Bivalvia (Pelecypoda) unidentified bivalves 3, 4, 5, 18
Astarte elliptica elliptical astarte 3 Astarte sp. 3 Mytilus edulis blue mussel 6 Nucula tenuis smooth nutclam 3 Portlandia arctica Arctic nutclam 3
Cephalopoda unidentified squid beaks 3 Teuthida 20 Gonatidae 12, 14 Gonatus fabricii boreoatlantic armhook squid 15, 21 Rossia moelleri 18 Rossia sp. 13, 15 Octopoda unidentified octopuses 5 Bathypolypus arcticus spoonarm octopus 18 Octopus spp. 20
Gastropoda unidentified gastropods 3, 4, 5, 11, 13, 18 Buccinum sp. 3 Colus sp. 18 Fusus sp. 3
24
Table 1. ‐‐ Continued.
Taxa Common Name Sources* Limacina helicina helicid pteropod 1, 3, 4 Margarita helicina 3, 4 Margarita sp. 18 Turritellidae unidentified tower shells 3, 4
OTHER TAXA Echinodermata echinoderms 5
Ophiura robusta 11 Tunicata sea squirts 5, 11, 12 Porifera sponges 5 Anthozoa anemones and corals 5 Annelida segmented worms 5 Echiura burrow worms 20 Echiuroidea spoon worms 5 Polychaeta bristle worms 3, 13, 14, 21 Maldanidae bamboo worms 3, 4 Polynoidae scale worms 3 Nemertea (Nemertinea) proboscis worms 13 Pycnogonida sea spiders 5
Nymphon (Chaetonymphon) hirtripes
3
* Sources: (1) Dunbar 1941, (2) Pikharev 1946, (3) McLaren 1958, (4) Fedoseev 1965, (5) Johnson et al. 1966, (6) Söderberg 1971, (7) Tormosov and Rezvov 1978, (8) Tormosov and Filatov 1979, (9) Lowry et al. 1980, (10) Bradstreet and Cross 1982, (11) Bradstreet and Finley 1983, (12) Gjertz and Lydersen 1986, (13) Smith 1987, (14) Weslawski et al. 1994, (15) Siegstad et al. 1998, (16) Kunnasranta et al. 1999, (17) Wathne et al. 2000, (18) Holst et al. 2001, (19) Stenman and Pöyhönen 2005, (20) Dehn et al. 2007, (21) Labansen et al. 2007
Despite regional and seasonal variations in the diet of ringed seals, gadid fishes tend to dominate the diet from late autumn through early spring in many areas (Kovacs 2007). Arctic cod1 (Boreogadus saida) is often reported to be among the most important prey species, especially during the ice‐covered periods of the year (Chapskii 1940, McLaren 1958a, Lowry et al. 1980, Bradstreet and Finley 1983, Gjertz and Lydersen 1986, Smith 1987, Węsławski et al. 1994, Belikov and Boltunov 1998, Siegstad et al. 1998, Wathne et al. 2000, Holst et al. 2001, Labansen et al. 2007). Other members of the cod family, including polar cod1 (Arctogadus glacialis), saffron cod (Eleginus gracilis), and navaga (Eleginus nawaga), are also seasonally important to ringed seals in some areas (Fedoseev 1965c, Johnson et al. 1966, Lowry et al. 1980, Bradstreet and Finley 1983, Siegstad et al. 1998, Holst et al. 2001). Other fishes reported to be locally important to ringed seals include smelt (Osmerus sp.) and herring (Clupea sp.) in the Sea of Okhotsk (Fedoseev 1965c); sculpins (Cottidae) in the Chukchi Sea (Johnson et al. 1966); capelin
1 The common names for Arctic cod and polar cod are not internationally consistent. In the United States and in this document, Arctic cod refers to Boreogadus saida and polar cod refers to Arctogadus glacialis, but in Europe, these common names are reversed.

25
(Mallotus villosus), redfish (Sebastes sp.), and snailfish (Liparis sp.) around Greenland (Siegstad et al. 1998); redfish and pricklebacks (Stichaeidae) around Svalbard (Węsławski et al. 1994, Labansen et al. 2007); sand lance (Ammodytes sp.) in Hudson Bay (Vincent‐Chambellant 2010); and herring, smelt, fourhorn sculpins (Myoxocephalus quadricornis), Atlantic salmon (Salmo salar), three‐spine sticklebacks (Gasterosteus aculeatus), and ruffe (Gymnocephalus cernuus) in the Baltic Sea (Söderberg 1971, Tormosov and Rezvov 1978, Sinisalo et al. 2006, Sinisalo et al. 2008).
Invertebrates appear to become more important to ringed seals in many areas during the open‐water season, and are often found to dominate the diets of young seals (Dunbar 1941, Fedoseev 1965c, Johnson et al. 1966, Lowry et al. 1980, Bradstreet and Finley 1983, Smith 1987, Węsławski et al. 1994, Siegstad et al. 1998, Holst et al. 2001). Large amphipods (e.g., Themisto libellula), mysids (e.g., Mysis oculata), euphausiids (e.g., Thysanoessa spp.), shrimps (e.g., Eualus, Lebbeus, and Pandalus spp.), and squid (e.g., Gonatus sp.) are all commonly found in the diet of ringed seals and can be very important in some regions, at least seasonally (Chapskii 1940, Dunbar 1941, McLaren 1958a, Fedoseev 1965c, Söderberg 1971, Lowry et al. 1980, Bradstreet and Finley 1983, Gjertz and Lydersen 1986, Smith 1987, Węsławski et al. 1994, Siegstad et al. 1998, Holst et al. 2001, Dehn et al. 2007). Smith (1987) and Kelly (1988) reported that ringed seals often eat small fish by sucking them into their mouth and swallowing them whole, and McLaren (1958a) and Fedoseev (1965c) suggested that ringed seals consume large quantities of small crustaceans by filtering them with their teeth.
Ringed seals experience a distinct annual cycle in body condition that is related to their seasonal behavior and feeding levels (Johnson et al. 1966, Kelly 1988, Harkonen et al. 2008). Adult female ringed seals suppress ovulation in years when they have not accumulated sufficient blubber reserves (Harwood et al. 2000). Adequate pre‐reproductive season resources apparently are necessary for adult females to gain sufficient reserves to support reproduction.
Changes in body condition index (McLaren 1958a, Smith 1987), weight (Fedoseev 1965c, Lowry et al. 1980), blubber content (Ryg et al. 1990), and proportion of killed seals lost due to sinking during hunting (Smith 1987, Ryg et al. 1990) indicated that ringed seals typically lose a significant proportion of their blubber mass during the spring to early summer. During the spring, adult males are busy defending underwater territories and/or access to females, which increases their energy demands and decreases their time available to forage. Adult females are whelping and nursing their young during the spring, and while mothers continue to feed or perhaps even increase feeding during this time (Ryg and Øritsland 1991), it is not enough to offset the high energy demands of lactation (Ryg et al. 1990, Hammill et al. 1991, Lydersen and Hammill 1993a, Lydersen 1995, Lydersen and Kovacs 1999). Immediately following the reproductive season, adults go through their annual molting period, during which they spend much of their time basking on the ice and reduce, but do not cease, their foraging (Pikharev 1946, McLaren 1958a, Fedoseev 1965c, Johnson et al. 1966, Lowry et al. 1980, Smith 1987, Ryg et al. 1990). In one study conducted in Svalbard, adult females lost about 19% of their blubber content between March and June while adult males lost about 12% (Ryg et al. 1990). Sexually immature seals of both sexes gained more core body mass than blubber mass during spring and early summer, resulting in a slight decrease in percent blubber content (Ryg et al. 1990). Ringed seals replenish their blubber reserves by increasing

26
feeding during the late summer, fall, and winter. Harkonen et al. (2008) reported that female Baltic Sea ringed seals doubled their weight between the end of the molt in May and the following December.
Overlap in the composition of prey taken by ringed seals and other marine predators (e.g., Arctic cod, seabirds, and other marine mammals) suggests that competition for food may occur in some areas (Frost and Lowry 1984), however, few studies have explored the extent of actual competition. Lowry et al. (1978) suggested that ringed seals may compete with bowhead whales (Balaena mysticetus) for euphausiids and gammarid amphipods in parts of the Arctic, but it is not clear that those prey limit the seals or the whales. In the Barents Sea, Wathne et al. (2000) found an almost complete overlap in diet between ringed seals and harp seals (Phoca groenlandicus). The harp seals, however, preyed on significantly larger Arctic cod that were distributed in deeper water; thus, the two seal species may have been exploiting different fractions of the same resource. Kovacs (2007) suggested that harbor, bearded, and ringed seals were likely competing for Arctic cod, pricklebacks, and sculpins in a small area on the west coast of Svalbard, based on studies of their diets.
2.7.2 Freshwater environment
Kunnasranta et al. (1999) reported that small, schooling fishes such as European perch (Perca fluviatilis), common roach (Rutilus rutilus), vendace (Coregonus albula), European smelt (Osmerus eperlanus), and ruffe were the most important prey species in Lake Saimaa, and that crustaceans were of minor importance due their low densities in the lake. Tormosov and Filatov (1979, cited in Sipilä and Hyvärinen 1998) reported that Lake Ladoga ringed seals mainly eat smelt, vendace, ruffe, burbot (Lota lota), three‐spined stickleback, perch, roach, European whitefish (Coregonus lavaretus), brown trout (Salmo trutta), fourhorn sculpin, and occasionally river lamprey (Lampetra fluviatilis) and crayfish (Astacus sp.). Studies of captive ringed seals from Lake Saimaa revealed a seasonal feeding cycle similar to wild populations from marine environments (Kunnasranta et al. 1999).
2.8 Historic and Current Abundance and Trends
Several factors make it difficult to accurately assess ringed seal population size or trends. The remoteness and dynamic nature of their sea‐ice habitat, time spent below the surface, along with their broad distribution and seasonal movements makes surveying ringed seals expensive and logistically challenging. Additionally, the species’ range crosses political boundaries, and there has been limited international cooperation to conduct range‐wide surveys. Details of survey methods and data are often limited or have not been published, making it difficult to judge the reliability of the reported numbers, especially some produced by Soviet‐era government institutions. Some studies have relied on surveys of seal holes and then estimated the number of seals based on various assumptions of the ratio of seals to holes. Most surveys are conducted during the basking period, and the numbers of seals on ice is multiplied by some factor to estimate population size or determine a population index. While a few recent studies have recorded and modeled time out of the water to develop correction factors for seals submerged and unseen, many studies only present estimates for seals visible on ice (i.e., the ‘basking
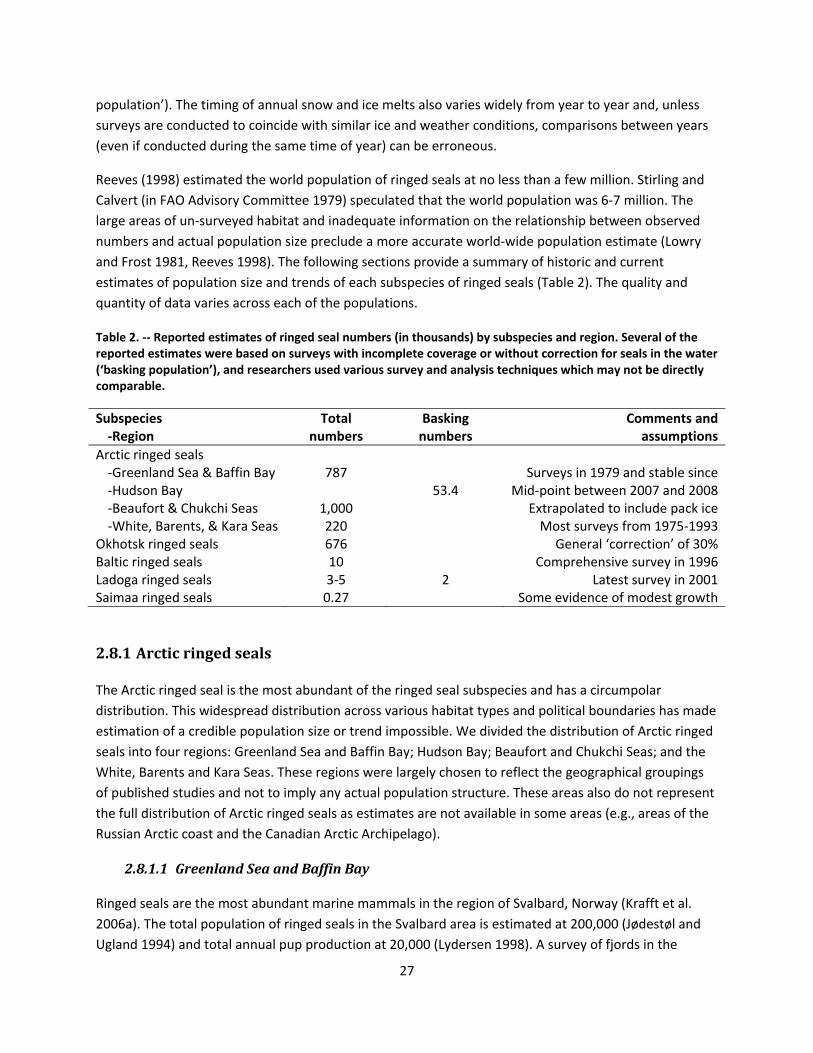
27
population’). The timing of annual snow and ice melts also varies widely from year to year and, unless surveys are conducted to coincide with similar ice and weather conditions, comparisons between years (even if conducted during the same time of year) can be erroneous.
Reeves (1998) estimated the world population of ringed seals at no less than a few million. Stirling and Calvert (in FAO Advisory Committee 1979) speculated that the world population was 6‐7 million. The large areas of un‐surveyed habitat and inadequate information on the relationship between observed numbers and actual population size preclude a more accurate world‐wide population estimate (Lowry and Frost 1981, Reeves 1998). The following sections provide a summary of historic and current estimates of population size and trends of each subspecies of ringed seals (Table 2). The quality and quantity of data varies across each of the populations.
Table 2. ‐‐ Reported estimates of ringed seal numbers (in thousands) by subspecies and region. Several of the reported estimates were based on surveys with incomplete coverage or without correction for seals in the water (‘basking population’), and researchers used various survey and analysis techniques which may not be directly comparable.
Subspecies ‐Region
Total numbers
Basking numbers
Comments andassumptions
Arctic ringed seals ‐Greenland Sea & Baffin Bay 787 Surveys in 1979 and stable since‐Hudson Bay 53.4 Mid‐point between 2007 and 2008‐Beaufort & Chukchi Seas 1,000 Extrapolated to include pack ice‐White, Barents, & Kara Seas 220 Most surveys from 1975‐1993
Okhotsk ringed seals 676 General ‘correction’ of 30%Baltic ringed seals 10 Comprehensive survey in 1996Ladoga ringed seals 3‐5 2 Latest survey in 2001Saimaa ringed seals 0.27 Some evidence of modest growth
2.8.1 Arctic ringed seals
The Arctic ringed seal is the most abundant of the ringed seal subspecies and has a circumpolar distribution. This widespread distribution across various habitat types and political boundaries has made estimation of a credible population size or trend impossible. We divided the distribution of Arctic ringed seals into four regions: Greenland Sea and Baffin Bay; Hudson Bay; Beaufort and Chukchi Seas; and the White, Barents and Kara Seas. These regions were largely chosen to reflect the geographical groupings of published studies and not to imply any actual population structure. These areas also do not represent the full distribution of Arctic ringed seals as estimates are not available in some areas (e.g., areas of the Russian Arctic coast and the Canadian Arctic Archipelago).
2.8.1.1 Greenland Sea and Baffin Bay
Ringed seals are the most abundant marine mammals in the region of Svalbard, Norway (Krafft et al. 2006a). The total population of ringed seals in the Svalbard area is estimated at 200,000 (Jødestøl and Ugland 1994) and total annual pup production at 20,000 (Lydersen 1998). A survey of fjords in the

28
Spitsbergen area during the peak molting period of 2002 relied on aerial, digital photographic surveys combined with a model of time out of the water (derived from VHF tagging data). The total estimate of ringed seals in the survey area was 7,585 (95% CI: 6,332‐9,085) (Krafft et al. 2006a).
Aerial strip surveys for ringed seals were conducted in Kong Oscars Fjord, Scoresby Sund, and adjacent areas in 1984 (Born et al. 1998). The aggregate estimate of ringed seals (unadjusted for seals in the water) was 28,882. Finley et al. (1983) reported estimates from surveys in 1979 for the northeastern Baffin Island shorefast ice and Baffin Bay pack‐ice habitats. More than 67,000 ringed seals were estimated for the shorefast ice near northeast Baffin Island and more than 417,000 within the pack‐ice habitat of Baffin Bay. Finley et al. (1983) further proposed the population of seals from Canada and Greenland inhabiting pack ice would be 787,000 once seals missed during the survey were accounted for. Miller et al. (1982) provide estimates for the West Greenland shorefast ice, also from surveys in 1979. The estimate for this region of eastern Baffin Bay was 97,800 ringed seals. Miller et al. (1982) also estimated an additional 15,500 seals were within in the shorefast ice habitats of Devon Island’s east coast and north to 80˚ latitude. Harvest records for Greenland in 1993 and 1994 indicated a total harvest of 75,644 and 64,661 seals and these harvest levels were within the annual harvest range estimated for 1971 to 1985 (59,000‐97,000) (Kapel and Rosing‐Asvid 1996). The estimate of 787,000 proposed by Finley et al. remains the only available, comprehensive estimate for the region, and the consistency in harvest records over time lends some confidence the population has not changed significantly.
2.8.1.2 Hudson Bay
Aerial surveys were conducted by Smith (1975) in the mid‐1970s throughout much of western Hudson Bay. The flight tracks were categorized by ice type and densities from these surveys were extrapolated to the entire region based on the distribution of those ice types. The result was an estimate (to the nearest thousand) of 455,000 seals in western Hudson Bay. This estimate is much larger than the 218,300 reported in the 1950s (McLaren 1958b). Smith pointed out the earlier studies did not account for seals using pack‐ice habitats and that this may explain the differences. In June 1995, Lunn et al. (1997) surveyed western Hudson Bay and estimated there were 140,880 (SE = 8,100) ringed seals on ice. They also concluded this estimate could equate to a total population of approximately 280,000 ringed seals when missed seals were accounted for. Ferguson and Secretariat (2009) conducted aerial surveys of seals on ice in western Hudson Bay for 2007 and 2008. They reported a density of 0.92 ± 0.07 seals/km2 and corresponding population estimate of 73,170 ± 5,440 (95% CI: 63,260‐84,633) in 2007. For 2008, the estimates for western Hudson Bay were lower. The reported density was only 0.44 ± 0.05 seals/km2 and the population estimate was 33,701 ± 3,704 (95% CI: 27,188‐41,776). The wide interannual variability in density and population estimates has been previously noted in western Hudson Bay, and the authors cautioned that the decline in 2008 may be more related to availability of seals to be counted than any actual decline. The mid‐point between the two years is 53,436 and may represent reasonable estimate of the population of seals on ice.

29
2.8.1.3 Beaufort and Chukchi Seas
The population assessments of ringed seals in the Beaufort and Chukchi Seas mostly have been confined to the U.S. and Canadian waters. In 1970, the number of ringed seals visible on shorefast ice along the North Slope of Alaska was estimated to be at least 11,612 (Burns and Harbo 1972). Frost and Lowry (1984) accumulated data from aerial surveys between 1970 and 1976 and calculated a mean density on ice of 0.40 seals/km2. With a correction for availability bias of a factor of 2, their total density estimate was 0.80 seals/km2. They extrapolated to the entire Alaskan Beaufort Sea and estimated 40,000 ringed seals for the winter and spring months. The estimate for the summer months was 80,000 based on an assumption the population would double as seals migrate from the west and south. Aerial surveys in the late 1990s (Frost et al. 2004) resulted in density estimates ranging from 0.57 to 1.14 seals/km2 although these surveys were limited to within 40 km of shore.
Bengtson et al. (2005) conducted surveys along the Alaskan Chukchi Sea coast in 1999‐2000. While surveys were focused on the coastal zone (within 37 km of shore), some survey lines were flown between 148 and 185 km from shore. The density and population estimates for ringed seals were derived from both survey results and a model of time out of the water developed from satellite data loggers deployed on six seals in Kotzebue Sound and Prudhoe Bay, Alaska. The average density of ringed seals was estimated to be 1.91 seals/km2 and 1.62 seals/km2 in 1999 and 2000, respectively. The highest densities were south of Kivalina, Alaska and near Kotzebue Sound. Population estimates for the entire survey area were 252,488 (SE = 47,204) in 1999 and 208,857 (SE = 25,502) in 2000.
Estimates have also been derived for all Alaskan shorefast ice habitats in both the Chukchi and Beaufort Seas (Frost 1985) based on aerial surveys in 1985‐1987. The estimates were 250,000 ringed seals in the shorefast ice and 1‐1.5 million including seals in the pack‐ice habitat. Kingsley and Lunn (1983) conducted aerial surveys in the eastern Beaufort and Amundsen Gulf regions of Canada in 1981‐1982. They estimated the number of ringed seals on ice in the eastern Beaufort at between 5,400 and 5,500. The estimate from their surveys in the Amundsen Gulf was 30,900 in 1981 and 70,500 in 1982. The wide interannual variability is similar to other regions and underscores the importance of understanding the relationships between seal behavior, snow and ice conditions, and survey methods. Using the more recent estimates from surveys by Bengtson et al. (2005) and Frost et al. (2004) in the late 1990s, the total population in the Alaskan Chukchi and Beaufort Sea regions is at least 300,000 seals. That number is likely an underestimate as Frost et al.’s surveys were limited to within 40 km of shore (mostly shorefast ice habitat) for the Beaufort. Frost (1985) proposed the total could be as much as 1.5 million once seals in pack ice were taken in to account. While the density of seals in the pack ice is lower, the large area covered by pack‐ice habitat results in a substantial increase in the population estimates. Considering these factors and at least an additional 50,000 seals from the eastern Beaufort and Amundsen Gulf, a reasonable estimate for the total population of ringed seals in the Chukchi and Beaufort Seas is 1 million seals.

30
2.8.1.4 White, Barents, Kara, and East Siberian Seas
The White, Barents, Kara, and East Siberian Seas encompass at least half of the worldwide distribution of ringed seals. Heptner et al. (1976) reported a preliminary estimate of the number of ringed seals from the eastern Barents Sea to the Bering Strait of between 2.0 and 2.5 million seals. Additional estimates have been derived for these seas, and the methods used for the assessments varied widely. Ognetov (2002) estimated between 35,000 and 50,000 ringed seals in the Barents Sea based on observations of seals on ice and the area of shorefast ice in June 1988 and May 1994. He estimated between 24,000 and 30,000 ringed seals in the White Sea in the 1970s and 1980s. Additional estimates—based on shipboard observations in the White Sea from 1975‐1993—ranged from 9,700 to 31,000. The Kara Sea appears to contain the largest number of ringed seals in this region of the Arctic. Ognetov (2002) extrapolated estimates from several regions of the Kara Sea to yield a total of 90,000 to 150,000 ringed seals in the Kara Sea. Ognetov suggested the estimate of 150,000 ringed seals is more likely. Thus, the total population of ringed seals across the White, Barents, and Kara Seas may be as many as 220,000 seals.
2.8.2 Okhotsk ringed seals
Fedoseev (1970, 1971b) conducted aerial surveys in the Sea of Okhotsk during 1968‐1969 and estimated a population of 800,000 ringed seals with approximately 130,000 around Sakhalin Island. Fedoseev (1971b) did not make adjustments for seals in the water since the estimated survey error (± 20%) was greater than the assumed correction coefficient (16‐17%). This was the same as the estimate previously back‐calculated from catch data in 1966 when a population decline due to hunting was identified (Fedoseev 1968, 1971a). These calculations also suggested that ringed seals in the Sea of Okhotsk had been declining steadily since 1955 when the population was estimated to exceed 1 million. Shustov (1972) also reported an estimate of 800,000 ringed seals in the Sea of Okhotsk based on aerial surveys during 1969 and 1970 but suggested that seal numbers had stabilized at low levels after years of intensive harvesting. Popov (1976) repeated Fedoseev’s (1971a, 1971b) estimates of 800,000 ringed seals and indicated that the population may have declined “in recent years”, but provided no further information. Aerial surveys were conducted over large portions of the western, northern, and eastern coasts of the Sea of Okhotsk during July‐October 1986, and total counts of 7,490 ringed were reported, of which 105 were on land (Lagarev 1988). Lagarev (1988) also reported coastal survey results from the southern Sea of Okhotsk, around Sakhalin Island and in the Tartar Strait, however, counts were not reported by species.
Based on aerial surveys, Fedoseev (2000) estimated 676,000 to 855,000 ringed seals in the Sea of Okhotsk between 1968 and 1990. Those estimates included increments to account for 30% of the population that was assumed to be unseen in the water. Fluctuations in population estimates since catch limits were initiated in 1968 were suspected to be natural. Regional estimates within the Sea of Okhotsk were reported at 110,000 in the Shelikhov Gulf, 485,000 from Tauyskaya Bay to the Shantarskiye Islands, and 130,000 off the coast of Sakhalin Island (Fedoseev 2000). Based on these surveys, a conservative estimate of ringed seals in the Sea of Okhotsk would be 676,000.

31
2.8.3 Baltic ringed seals
Based on bounty records, Kokko et al. (1999) back‐calculated an estimate of 50,000 to 450,000 Baltic ringed seals in 1900. A more thorough compilation that considered age classes in the harvest and a broader geographic coverage concluded that the minimum population must have been about 180,000 ringed seals at that time (Harding and Härkönen 1999). The large range in the estimates reflects uncertainty in the hunting dynamics and whether the populations were historically subject to density dependence. By the 1940s, the population had been reduced to 25,000, in large part due to Swedish and Finnish extermination efforts (Harkonen et al. 2008). The population appears to have been stable at that level until 1965, after which it declined to less than 5,000 in the late 1970s (Harding and Härkönen 1999) as a consequence of sterility caused by organochlorines (Helle 1980b, Bergman and Olsson 1985).
Ringed seals in the Baltic are found in three general regions: the Bothnian Bay, Gulf of Finland, and Gulf of Riga plus the Estonian west coast. Low numbers of ringed seals are also present in the Bothnian Sea and the southwestern region of Finland (Härkönen et al. 1998). Härkönen et al. (1998) compiled a table of Baltic ringed seal surveys between 1970 and 1996. The surveys varied in methods and levels of effort.
The greatest concentration of Baltic ringed seals is in the Bothnian Bay. Surveys in the Bothnian Bay in the 1970s estimated at least 2,000 to 3,000 animals (Härkönen and Lunneryd 1992). Surveys using the same aerial methods in 1984 and 1987 indicated a decline in numbers to the mid‐1980s (Helle 1990). Surveys of the Bothnian Bay in 1989‐1990 yield an estimated basking population at 2,470 ± 454 in 1989 and 2,497 ± 367 in 1990 (Härkönen and Lunneryd 1992), implying no statistically significant trend from 1975 to 1990. A time series of surveys carried out with compatible methods for the period 1988‐2009 showed that the Bothnian Bay population has increased by 4.5% annually and that estimated numbers on ice were about 6,100 (Karlsson et al. 2008). About 75% of the Baltic ringed seal population is found in the Bothnian Bay population.
Estimates of the ringed seal population in the Gulf of Finland prior to the 1980s are available from Soviet‐era studies (Tormosov and Rezvov 1978). Little information is available regarding the methods employed in these surveys, and the reported numbers, which ranged from 2,100 to 8,200 seals, are likely gross overestimates (Härkönen et al. 1998). The current best estimates of seals in the Gulf of Finland are derived from surveys in 1994‐1996 and range from 149 to 173 (Härkönen et al. 1998). Later annual surveys confirmed that only some hundreds of ringed seals are found in the area and that the population shows a growth rate close to zero (T. Härkönen, Swedish Museum of Natural History, pers. comm., 1 September 2010). Little historical information is available on the number of ringed seals in the Gulf of Riga. Härkönen et al. (1998) reported the first surveys in 1994 and an additional survey of the entire region in 1996. The estimate in 1996 was 1,407 ± 590 seals on the ice.
Härkönen et al. (1998) provided the first comprehensive estimate of ringed seals out of the water in the Baltic Sea at 5,510 (± 42%) in 1996. Harkonen et al. (2008) corrected this to a “true” population estimate of approximately 10,000 seals in the Baltic based on an estimated 57% of seals out of the water during the molt. The Baltic population, as a whole, appears to be increasing since that time although this is

32
entirely reflective of the Bothnian Bay population, and potential declines of the southern regions bear consideration (Harkonen et al. 2008, Karlsson et al. 2008).
2.8.4 Ladoga ringed seals
The current population of ringed seals in Lake Ladoga is thought to number between 3,000 and 5,000 seals, a decrease from numbers reported for the early 1900s. Zheglov and Chapskii (1974) reported estimates of 20,000 seals in the 1930s and estimates of 5,000 to 10,000 seals in the 1960s. Citing Russian reports from the 1970s and 1980s, Sipilä and Hyvärinen (1998) reported that earlier population estimates ranged broadly from 3,000 to 20,000 seals. Zheglov and Chapskii (1974) conducted a test aerial survey in Lake Ladoga. They conducted an initial flight in March and found holes of ringed seals 5 to 30 km from the eastern and western shores. During a subsequent flight in April, they encountered both seal holes and seals. They reported the density of seals in the northern part of the lake on what they referred to as ‘landfast ice’ at 0.028 seals/km2 and the density of holes at 0.084 seals/km2. In other parts of the lake, Zheglov and Chapskii (1974) described the ice as ‘breaking‐up landfast ice derived from drifted‐in ice’ and reported those densities much higher at 0.19/km2 for seals and 0.36/km2 for observed holes. They reported encountering 2,200 seals but believed this to be an underestimate because, for several years prior to the survey, seals were harvested at a rate of 500 per year. Additionally, Zheglov and Chapskii (1974) estimated the number of holes encountered to be 5,100. They assumed the number of holes per seal was approximately 1 and reported the population estimate at 5,000. Sipilä and Hyvärinen (1998) also provided results from another Russian aerial survey in the 1970s that estimated the population of ringed seals in Lake Ladoga to be 3,500 to 4,700 animals. An aerial survey in 2001 determined the population of seals basking on the ice to be 2,000 (± 70), and Agafonova et al. (2007) concluded that the total population is between 3,000 and 5,000 seals.
2.8.5 Saimaa ringed seals
The population of ringed seals in Lake Saimaa was estimated to have been between 4,000 and 6,000 animals approximately 5,000 years ago (Sipilä and Hyvärinen 1998, Sipilä 2006). Using a back‐casting process based on reported bounty statistics, however, Kokko et al. (1999) estimated the population in 1893 to be between 100 and 1,300 seals. Sipilä et al. (1990) reported a marked decrease from approximately 250 seals in 1971 to an estimated 140 seals remaining in 1984. The more recent surveys for seals in Lake Saimaa have employed both aerial surveys and lair mapping.
Lake Saimaa is a complex body of water, and the population trends and estimates for Saimaa ringed seals have differed across the various regions. The southernmost and northern regions of the lake experienced drastic decreases from the mid‐1960s to the 1980s. In 1966 in the northern region of the lake, near Pyhaselka, 30 seals were seen basking on the ice. In 1977, only 13 seals were seen at the corresponding time. The central regions of the lake, near Linnansaari National Park, remained stable over the same period of time.

33
In 1993, the Saimaa seal was listed as endangered under the ESA (National Marine Fisheries Service 1993) and as depleted under the U.S. Marine Mammal Protection Act (MMPA) of 1972, as amended (16 U.S.C. 1361 et seq.) (National Marine Fisheries Service 2010).
The current population estimate is less than 300 seals (Sipilä 2006; M. Kunnasranta, Finnish Game and Fisheries Research Institute, pers. comm., 9 July 2010), and the mean growth rate from 1990 to 2004 was 1.026. As was noted in previous studies, the growth rates vary across the different regions of Lake Saimaa and range from 0.919 to 1.056 with some of the higher growth rates occurring in the central regions of the lake and lower numbers in the northernmost and southernmost regions. Sipilä (2006) projected that the population of Saimaa ringed seals may reach 400 by 2015, but cautioned that the seals may no longer be present in some regions of the lake.


35
3 SPECIES DELINEATION
The ESA defines a “species” as “any subspecies of fish or wildlife or plants, and any distinct population segment of any species of vertebrate fish or wildlife which interbreeds when mature.” The NMFS and the USFWS also recognize DPSs under the ESA. The agencies’ Policy Regarding the Recognition of Distinct Vertebrate Population Segments Under the Endangered Species Act calls for recognizing a segment of a population as distinct if it is both “discrete” and “significant.” A population segment may be considered discrete if:
1. It is markedly separated from other populations of the same taxon as a consequence of physical, physiological, ecological, or behavioral factors. Quantitative measures of genetic or morphological discontinuity may provide evidence of this separation; or
2. It is delimited by international governmental boundaries within which differences in control of exploitation, management of habitat, conservation status, or regulatory mechanisms exist that are significant in light of section 4(a)(1)(D) of the Act.
If a population segment is considered discrete under one or more of the above conditions, its biological and ecological significance will then be considered in light of Congressional guidance (see Senate Report 151, 96th Congress, 1st Session) to list DPSs ‘‘… sparingly’’ while encouraging the conservation of genetic diversity. In determining the significance of a discrete population segment to its species, the NMFS considers:
1. Persistence of the discrete population segment in an ecological setting unusual or unique for the taxon,
2. Evidence that loss of the discrete population segment would result in a significant gap in the range of a taxon,
3. Evidence that the discrete population segment represents the only surviving natural occurrence of a taxon that may be more abundant elsewhere as an introduced population outside its historic range, or
4. Evidence that the discrete population segment differs markedly from other populations of the species in its genetic characteristics.
3.1 Evaluation of Discreteness
3.1.1 Separation by Physical, Physiological, Ecological, or Behavioral Factors
Suggestions that ecological factors created discrete populations within the Arctic and Okhotsk subspecies of ringed seals have not been fully resolved. Variations in body size and growth rates have

36
been taken as evidence of different breeding populations of ringed seals in shorefast versus pack ice. Fedoseev (1975) contrasted body size and growth rates of ringed seals collected in the Sea of Okhotsk, Bering Sea, and Chukchi Sea, and reported that seals breeding in pack ice were smaller and matured at earlier ages than those breeding in shorefast ice. He apparently assumed, however, that all seals collected in the Sea of Okhotsk and the Chukchi Sea bred in pack ice and that all seals collected in the Bering Sea bred in shorefast ice; the validity of those assumptions is uncertain. Consistent with Fedoseev’s (1975) suggestion of pack ice and shorefast ice “ecotypes,” however, are observations by Native hunters of two types of ringed seals that differ markedly in adult body size. Large shorefast ice seals and smaller pack‐ice seals are reported by hunters in the northern Bering Sea (C. Noongwook, Savoonga, Alaska, pers. comm., March 1980), northern Baffin Island (Finley et al. 1983), southern Baffin Island (McLaren 1958a), and northwestern Greenland (Vibe 1950). Finley et al. (1983) compared body size, craniology, diet, and parasite loads of ringed seals collected in shorefast ice and pack ice of Baffin Bay and concluded that seals in those two habitats were reproductively isolated.
Physical factors are believed to separate Baltic and Okhotsk ringed seals from Arctic ringed seals. Throughout their range, the seals are adapted to ice environments, and large expanses of ice‐free waters in the North Atlantic and North Pacific oceans limit movements between the Arctic Ocean and the Baltic Sea or the Sea of Okhotsk. Movements between the Baltic Sea or the Sea of Okhotsk and the Arctic Ocean are presumed to be rare, but the actual frequency is unknown. Based on movements of 19 seals over periods of 9 months or less, Harkonen et al. (2008) suggested that Baltic Sea ringed seals may be comprised of three stocks in the Bothnian Bay, Gulf of Finland, and Gulf of Riga. Genetic analyses, however, do not seem to support that conclusion (Palo et al. 2001). Many sections of Lake Saimaa are connected only by small straits, but there is no evidence that the Saimaa or Ladoga subspecies are subdivided in to multiple discrete populations.
Physiological factors that could influence population discreteness most typically involve genetic variation. Genetic variation among ringed seal populations is higher than among closely related species. For example, the expected heterozygosity in microsatellite loci was 0.77 in the gray seal (Halichoerus grypus) and 0.56 in the harbor seal but 0.81 in the ringed seal (Palo et al. 2001). In two recent studies of genetic diversity among ringed seal populations, the number of alleles per microsatellite locus averaged 12.4 (Palo et al. 2001) and 11.7 (Sell 2008, Kelly et al. 2009). The greater genetic diversity among ringed seals likely reflects historical population sizes of several million ringed seals in contrast to populations numbering considerably less than 1 million for gray and harbor seals (Palo et al. 2001). Whereas populations of less ice‐adapted species were presumed to have declined during the last glaciations, ringed seals likely thrived with the expanded ice habitats (Palo 2003).
Palo et al. (2001) examined the genetic structure of ringed seals within the Baltic Sea, contrasting microsatellite DNA diversity among 149 Baltic Sea ringed seals with 39 ringed seals sampled at Svalbard and considered representative of the Arctic subspecies. Their analysis of eight microsatellite loci showed low allele frequency differentiation considering the putative subspecific rank of Baltic and Arctic ringed seals. They examined the likelihood that the observed genetic variation was consistent with post glacial isolation or recurrent gene flow between the Arctic and Baltic populations, and they concluded that the

37
latter was more likely. Palo et al. (2001) noted that the Baltic Sea subspecies has recently been fragmented in to three breeding segments—Gulf of Riga, Gulf of Finland, and Gulf of Bothnia—but that genetic evidence of the separation is not yet evident.
Genetic diversity among ringed seals in the Arctic Ocean, Baltic Sea, Lake Ladoga, and Lake Saimaa was evaluated using mitochondrial DNA (mtDNA) and microsatellite markers by Palo (2003). Haplotype and nucleotide diversity were examined for a 416 base pair segment of the mtDNA control region for ringed seals collected in the Bering and Beaufort Seas off Alaska (N = 7), near Svalbard (N = 39), the White Sea (N = 34), the Baltic Sea (N = 38), Lake Ladoga (N = 8), and Lake Saimaa (N = 25). Eight microsatellite loci were examined for seals sampled at Svalbard (N = 39), the Baltic Sea (N = 149), and Lake Saimaa (N = 80). Both the microsatellite and mtDNA markers showed high levels of diversity among Arctic ringed seals, and the mtDNA showed no significant differentiation between Alaskan and Svalbard seals. Palo (2003) found lower—but still substantial—genetic diversity in the Baltic Sea and Lake Ladoga populations relative to the Arctic Ocean population. Lake Saimaa ringed seals showed 68% less microsatellite diversity and 87% less mtDNA diversity than did ringed seals sampled at Svalbard.
Davis et al. (2008) examined genetic variability in 11 microsatellite loci from over 300 ringed seals collected at eight Arctic Ocean locations. They reported that seals collected in the White Sea differed significantly in allele frequencies from all other populations and showed modest overall differentiation. Comparisons between the other sample locations, however, revealed no significant differences in allele frequencies. Genetic distance was significantly correlated with geographic distance, but a Bayesian analysis suggested that the most likely number of populations within the Arctic was one.
Samples analyzed by Palo (2003) and Palo et al. (2001) were collected opportunistically during research projects or from animals found dead. Similarly, those reported by Davis et al. (2008) came from seals harvested in subsistence hunts or handled in scientific studies (Davis 2004). Many of the samples in those studies likely were collected outside of the breeding season. Ringed seals fitted with satellite‐linked transmitters in Alaska made extensive movements outside of the breeding season but returned to the same breeding sites in successive years (Kelly et al. 2010). Thus, seals from multiple populations may have been intermixed in Palo et al.’s (2001) and Davis et al.’s (2008) investigations. Ringed seals tracked by satellite in the Baltic Sea showed fidelity to three areas with no overlap in range (Harkonen et al. 2008).
Kelly et al. (2009) examined population structure by analyzing mitochondrial and nuclear DNA of 358 ringed seals collected at breeding sites (Swanson et al. 2006). Analyses of 9 microsatellite loci and a 359 base pair sequence of the Cytochrome Oxidase I mtDNA locus region were consistent with ongoing gene flow between Arctic Ocean and Baltic Sea breeding sites, although not with the Lake Saimaa population. They pointed to the history of large effective population sizes among ringed seals, and could not rule out the existence of multiple, genetically isolated populations in which genetic drift has been weak. Isolation among Arctic, Baltic, and Saimaa regions was supported by an analysis in which 98% of the seals sampled in those regions were genetically assigned to their capture sites (Sell 2008, Kelly et al. 2009).

38
Behavioral factors that might influence population subdivision are poorly known. Evidence of fidelity to breeding sites, however, suggests the potential for multiple discrete populations within the Arctic subspecies, at least (Kelly et al. 2010). Further investigation is needed to determine whether, in fact, there are multiple, discrete populations within the Arctic subspecies.
3.1.2 Delimitation by International Differences in Management and Regulatory Mechanisms
The normal range of ringed seals includes 10‐15 countries deploying a variety of management and regulatory mechanisms.
Japan – The southern portion of the Okhotsk ringed seal’s range encompasses the northern shores of Hokkaido, waters they share with several other species of pinnipeds. Until recently, Japan had no regulations or laws for the conservation or management of pinnipeds, in part, due to opposition to conservation measures from commercial fishing interests (Wada et al. 1991). Since 2002, however, seal harvests in Japan have been controlled under “Wildlife Protection and Appropriate Hunting Law” (Ministry of Environment and Hokkaido Prefectural Government 2007; Y. Mitani, Hokkaido University, pers. comm., 14 September 2009). Apparently, few ringed seals have been harvested around Hokkaido in recent years.
Russia – Arctic and Okhotsk ringed seals are harvested by permit for subsistence (indigenous people) and for commercial purposes in Russia. Sport hunting is not allowed. The Pacific Research Fisheries Center (TINRO) sets annual total allowable catches (TACs) based on regional biological reviews. Permitees are required to record harvests. Enforcement is lacking in remote regions, and reporting is probably incomplete. Nonetheless, it is believed that the overall take is small (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 4 May 2010).
Canada – In Canada, the DFO has jurisdiction over marine mammals, and management of Arctic ringed seals is regulated by the Fisheries Act and the Marine Mammal Regulations, which identify regional Sealing Areas in Arctic and Atlantic waters (P. A. Hall, DFO, Canada, pers. comm., 6 May 2010). Harvesting requires a license except that licenses are not required for Canadian aboriginals and land claim beneficiaries to harvest seals for food/social/ceremonial purposes or for non‐aboriginal residents harvesting for food only in Sealing Areas 1 to 4. Ringed seals are not harvested commercially in Canada, and subsistence harvests are managed regionally. Where aboriginal land claims have been settled, the DFO co‐manages marine mammal resources with the Fisheries Joint Management Committee in the western Arctic, the Nunavut Wildlife Management Board in Nunavut, the Nunavik Marine Region Wildlife Board in Nunavik (northern Quebec), and the Labrador Inuit Association in Labrador. No specific management objectives have been identified for ringed in the Canadian Arctic (Sealing Areas 1‐3).
Elsewhere, the DFO manages ringed seal harvests. In Labrador and Newfoundland (Sealing Areas 4‐33), ringed seals are included in the Atlantic Seal Management Plan although they are not harvested commercially.

39
Greenland – Arctic ringed seal harvests are not regulated in Greenland. Subsistence hunters regulate their own harvest by avoiding certain fjords, and the government is considering regulations for ringed seal harvests (A. Rosing‐Asvid, Greenland Institute of Natural Resources, pers. comm., 22 April 2010).
Norway – In the waters around Svalbard, a few hundred Arctic ringed seals are harvested throughout the year except for a 2‐month closed period during the breeding season (K. Kovacs, Norwegian Polar Institute, pers. comm., 22 April 2010). Off of the mainland coast of northern Norway, where ringed seals are rare, small numbers are harvested by sport hunters regulated by a quota system.
United States – In Alaska, Arctic ringed seals are harvested by indigenous hunters under an exception to the MMPA. The MMPA permits harvests for subsistence purposes by coastal dwelling Alaska Natives and establishes mechanisms for co‐management with Alaska Natives. The Ice Seal Committee, an Alaska Native organization, has an agreement with the NMFS to co‐manage ringed seals and other ice‐associated seals in Alaska. At present, there are no limitations on the subsistence take.
Baltic Sea countries – Denmark, Estonia, Finland, Germany, Latvia, Lithuania, Poland, Russia, and Sweden banned harvest of Baltic ringed seals based on a recommendation of the Helsinki Commission (HELCOM). Limited take for scientific research is allowed under the agreement, and additional harvests may be permitted if the populations show growth and exceed a “Limit Reference Level.”
The Saimaa ringed seal occurs solely in waters governed by Finland; the Ladoga ringed seal occurs exclusively in Russian waters, and the Okhotsk ringed seal is found primarily in Russian waters but also in Japanese waters. Multiple countries have jurisdiction over Arctic ringed seals (Russia, United States, Canada, Greenland, and Norway) and Baltic ringed seals (Denmark, Estonia, Finland, Germany, Latvia, Lithuania, Poland, Russia, and Sweden), although most of the Baltic population is in the Swedish and Finnish waters of the Bothnian Bay.
Conservation and management by the various nations within the ringed seals’ range are largely consistent, and cooperation is good. Additional population subdivision along international boundaries would not enhance conservation efforts for any of the five subspecies.
3.2 Evaluation of Significance
No discrete population segments were identified within any of the five subspecies of ringed seals. Therefore, it was not necessary to evaluate the significance of population subdivision.
3.3 Determination of Distinct Population Segments
Five recognized subspecies of ringed seals in the Sea of Okhotsk, Arctic Ocean (including the Bering Sea), Baltic Sea, Lake Saimaa, and Lake Ladoga are distinct based on morphological and genetic differentiation and/or the uniqueness of their ecological settings. At present, it is not possible to conclude whether the Arctic Ocean subspecies is made up of a single, panmictic population or multiple discrete populations.

40
With the exception of the freshwater populations, genetic differentiation is low among subspecies of ringed seals and may suggest on‐going gene flow. Alternatively, a history of large population sizes—and apparently rapid population growth following recolonizations of post‐glacial habitats—may have limited the loss of alleles despite limited gene flow. The small remnant population in Lake Saimaa is a notable exception; that population experienced an extreme bottle neck dipping as low as 100 animals. Fidelity to breeding sites and results of genetic assignment tests support philopatry and suggest that there may, in fact, be more distinct population units.
The BRT recognizes five distinct populations of ringed seals corresponding to the five subspecies and emphasizes that further investigation may well discern additional distinct units, especially within the Arctic subspecies. The geographic distributions of the five subspecies are shown in Figure 2.

41
4 EXTINCTION RISK ASSESSMENT
4.1 Time Frame: The Foreseeable Future
The BRT assessed the risk of extinction to ringed seals as background for the Secretary’s decision whether any of the subspecies should be listed as threatened or endangered under the ESA. The ESA defines endangered species as “any species which is in danger of extinction throughout all or a significant portion of its range.” A threatened species is “any species which is likely to become an endangered species within the foreseeable future throughout all or a significant portion of its range.” Determining whether a species is threatened, therefore, requires consideration of the time frame over which the population status can be said to be “foreseeable”, in the sense of a credible prediction of the likely outcome. To be credible, a prediction must have a substantial element of rigor that derives from factors such as relevant data, consideration of uncertainty (to assess whether an outcome is more likely than not to occur), and concurrence of relevant subject‐matter experts. Speculation alone does not constitute credible prediction or foreseeability.
The foreseeability of a species’ future status depends upon both the foreseeability of threats to the species and foreseeability of the species’ response to those threats. When a species is exposed to a variety of threats, each threat may be foreseeable on a different time frame. For example, a threat stemming from well‐established, observed trends in a global physical process may be foreseeable on a much longer time horizon than a threat stemming from a potential episodic process such as an outbreak of disease that may never have been observed to occur in the species.
4.1.1 Factors in the Foreseeability of Threats to Ringed Seals
The petition by the CBD to list ringed seals cited global warming as the foremost concern, and others have speculated similarly that ringed seals and other ice‐associated marine mammals are at risk from loss of sea‐ice habitat in a warming climate (Tynan and DeMaster 1997, Lowry 2000, Kelly 2001, Learmonth et al. 2006, Simmonds and Isaac 2007, Kovacs and Lydersen 2008, Laidre et al. 2008, Moore and Huntington 2008). Other potential threats, such as modification of ringed seals’ prey community by ocean acidification may be related to warming by the common driver of greenhouse gas (GHG) emissions, the root cause of the largest portion of observed and projected climate change. Therefore, the predictability of GHG emissions is of primary consideration in the foreseeability of climate‐related threats to ringed seals.
The analysis and synthesis of information presented by the Intergovernmental Panel on Climate Change (IPCC) in its Fourth Assessment Report (AR4) (IPCC 2007a) represents the scientific consensus view on the causes and future of climate change (but see Oppenheimer et al. 2007 for a reminder that achieving consensus should be balanced with consideration of the full range of uncertainty and plausible outcomes). The IPCC AR4 is the most recent comprehensive summary of observations, analyses, and models that collectively have been found to be compelling by all major scientific bodies in the United

42
States with directly relevant expertise (Oreskes 2004). The IPCC AR4 used a range of future GHG emissions produced under six “marker” scenarios from the Special Report on Emissions Scenarios (SRES; IPCC 2000) to project plausible outcomes under clearly stated assumptions about socioeconomic factors that will influence the emissions. Conditional on each scenario, the best estimate and likely range of emissions were projected through the end of the 21st century.
The factors that distinguish the SRES marker scenarios include economic and human population growth rates, technological development, and the mix of energy sources used to meet global needs. The policy of the IPCC is to consider all six SRES marker scenarios equally likely. The differences in emissions under these scenarios, however, contribute to the variation in predicted future conditions, especially in the latter half of the 21st century.
Conditions such as surface air temperature (SAT) and sea‐ice area are linked in the IPCC climate models to GHG emissions by the physics of radiation processes. When carbon dioxide (CO2) is added to the atmosphere, it has a long residence time and is only slowly removed by ocean absorption and other processes. Based on IPCC AR4 climate models, expected global warming—defined as the change in global mean SAT—by the year 2100 depends strongly on the assumed emissions of CO2 and other GHGs. By contrast, warming out to about 2040‐2050 will be primarily due to emissions that have already occurred and those that will occur over the next decade. Thus, conditions projected to mid‐century are less sensitive to assumed future emissions scenarios (Figure SPM.5 in IPCC 2007c). Uncertainty in the amount of warming out to mid‐century is primarily a function of model‐to‐model differences in the way that the physical processes are incorporated, and this uncertainty can be addressed in predicting ecological responses by incorporating the range in projections from different models.
The IPCC AR4 emphasized the importance of this consideration for evaluating its 21st‐century projections (Meehl et al. 2007):
“There is close agreement of globally averaged SAT multi‐model mean warming for the early 21st century for concentrations derived from the three non‐mitigated IPCC Special Report on Emission Scenarios (SRES: B1, A1B and A2) scenarios (including only anthropogenic forcing). . . this warming rate is affected little by different scenario assumptions or different model sensitivities, and is consistent with that observed for the past few decades. . . Possible future variations in natural forcings (e.g., a large volcanic eruption) could change those values somewhat, but about half of the early 21st‐century warming is committed in the sense that it would occur even if atmospheric concentrations were held fixed at year 2000 values. By mid‐century (2046‐2065), the choice of scenario becomes more important for the magnitude of multi‐model globally averaged SAT warming. . . About a third of that warming is projected to be due to climate change that is already committed. By late century (2090‐2099), differences between scenarios are large, and only about 20% of that warming arises from climate change that is already committed.”
The USFWS used this guidance from the IPCC AR4 to define the horizon of the foreseeable future as the year 2050 in its decision to list the polar bear as threatened (U.S. Fish and Wildlife Service 2008). The

43
NMFS used the same guidance and time horizon in its decision not to propose any listing for the ribbon seal (National Marine Fisheries Service 2008a).
In this review of ringed seal population status, the BRT recognized that the physical basis for some of the primary threats faced by the species have been projected, under certain assumptions, through the end of the 21st century, and that these projections currently form the most widely accepted version of the best available information about future conditions. Therefore, in the risk assessment that follows, the BRT used the full 21st‐century projections as the basis for the foreseeability of threats stemming from climate change.
Because the current consensus is to treat all SRES scenarios as equally likely, one option for representing the full range of variability in potential outcomes would be to project from any model under all six scenarios. This may be impractical in many situations, so the typical procedure for projecting impacts is to use an intermediate scenario, such as A1B or B2 to predict trends, or one intermediate and one extreme scenario (e.g., A1B and A2) to represent a significant range of variability or uncertainty.
It is important to note that the SRES scenarios are non‐mitigated; that is, they do not contain explicit assumptions about implementation of agreements or protocols on emission limits beyond current mitigation policies and related sustainable development practices. Recent studies have begun to explore the projected outcomes of emissions mitigation scenarios (Van Vuuren et al. 2008, Strassmann et al. 2009). The mitigated scenarios produce substantially less 21st century warming than the SRES scenarios, though even the most stringent mitigation scenarios result in an average of about 1.4°C warming (range of 0.5°‐2.8°C) above 1990 levels (Van Vuuren et al. 2008). Incorporating the likelihood and effectiveness of further mitigation policies into an assessment of risks to ringed seals, however, is beyond the purview and capabilities of the BRT. It is left as a policy choice as to whether anticipation of climate mitigation measures should be a factor in the decision of whether to list ringed seals under the ESA.
Not all potential threats to ringed seals are climate related, and therefore not all can be regarded as foreseeable through the 21st century. As a simple example, morbillivirus infections have caused mass mortality in European harbor seals, and Arctic ringed seals have tested positive for the virus. Thus, distemper may be considered a threat to ringed seals, but the foreseeability of an inherently episodic and novel threat is difficult or impossible to establish.
4.1.2 Factors in the Foreseeability of Ringed Seal Responses to Threats
A threat to a species and the species’ response to that threat are not, in general, equally foreseeable. The demographic, ecological, and evolutionary responses of ringed seals to threats from a warming climate are, in most cases, difficult to predict, even though future warming is highly likely to continue. The difficulty stems both from limited knowledge of the species’ current status (i.e., population density, trends, and vital rates) and its resilience to the effects of climate change. As discussed in more detail in Sections 4.2 and 4.3, the data on size and trends of most of the populations are imprecise, especially in the Arctic and Okhotsk subspecies, limiting quantitative predictions of the impact of projected environmental conditions on ringed seal survival or reproduction. The range of uncertainty in forward

44
projections of ringed seal population size is bounded above by the species maximal growth rate, approximately 12% annually. There is, of course, no lower bound on the rate of population change, as any population could conceivably go extinct instantly from a sufficiently severe perturbation. These extreme scenarios of hypothetical population responses, however, are not very helpful in the practical matter of judging whether ringed seals are likely to reach some threshold conservation status within a particular period of time. We must therefore rely upon the judgment of experts familiar with the natural history of the species and its evolutionary origins.
4.1.3 Lack of a Single Time Frame for the Foreseeable Future
Many of the anticipated effects of GHGs have been projected through the end of the 21st century, and a broad consensus has formed around various outcomes in those projections, subject to certain inputs and assumptions. These nearly century‐long projections should be considered in the assessment of the outlook for ringed seals, yet there is no single period of time that is appropriate for consideration of the risks from all the apparent threats faced by the species, and the species’ responses to the threats. In assessing whether ringed seals are threatened or endanger, the foreseeability of each threat and the species’ responses should be considered separately. For a species like the ringed seal, composed of multiple subspecies, these assessments should be made separately for each subspecies.
4.2 Analysis of Factors Listed Under Section 4(a)(1) of the Endangered Species Act
Section 4(a)(1) of the ESA requires the determination of whether a species is endangered or threatened because of any of the following factors:
(A) the present or threatened destruction, modification, or curtailment of its habitat or range;
(B) overutilization for commercial, recreational, scientific, or educational purposes;
(C) disease or predation;
(D) the inadequacy of existing regulatory mechanisms; or
(E) other natural or manmade factors affecting its continued existence.
4.2.1 Present or Threatened Destruction, Modification, or Curtailment of the Species’ Habitat or Range
4.2.1.1 Global climate change
Research, monitoring, and modeling of global climate change have progressed rapidly during the past several decades, yielding a vast body of information on causes of climate change, effects, and ways to mitigate the problems. In 1988, the World Meteorological Organization and the United Nations

45
Environmental Programme established the IPCC to provide an objective source of information about this complex issue (IPCC 2008). The IPCC has produced four assessment reports that represent syntheses of the best available and most comprehensive scientific information on climate change to date. The following two excerpts from the IPCC’s Climate Change 2007: Synthesis Report, Summary for Policymakers (IPCC 2007b), highlight some of the observed and projected changes in climate and their anticipated effects/impacts:
“Observed changes in climate and their effects:
• Warming of the climate system is unequivocal, as is now evident from observations of increases in global average air and ocean temperatures, widespread melting of snow and ice and rising global average sea level.
• Observational evidence from all continents and most oceans shows that many natural systems are being affected by regional climate changes, particularly temperature increases.
• There is medium confidence that other effects of regional climate change on natural and human environments are emerging, although many are difficult to discern due to adaptation and non‐climatic drivers.”
“Projected climate change and its impacts: • There is high agreement and much evidence that with current climate change mitigation
policies and related sustainable development practices, GHG emissions will continue to grow over the next few decades.
• Continued GHG emissions at or above current rates would cause further warming and induce many changes in the global climate system during the 21st century that would very likely be larger than those observed during the 20th century.
• There is now higher confidence than in the TAR [Third Assessment Report] in projected patterns of warming and other regional‐scale features, including changes in wind patterns, precipitation and some aspects of extremes and sea ice.
• Studies since the TAR have enabled more systematic understanding of the timing and magnitude of impacts related to differing amounts and rates of climate change.
• Altered frequencies and intensities of extreme weather, together with sea level rise, are expected to have mostly adverse effects on natural and human systems.
• Anthropogenic warming and sea level rise would continue for centuries due to the time scales associated with climate processes and feedbacks, even if GHG concentrations were to be stabilized
• Anthropogenic warming could lead to some impacts that are abrupt or irreversible, depending upon the rate and magnitude of the climate change.”
Both the observed and the projected effects of a warming global climate are most extreme in northern high‐latitude regions (ACIA 2005, Meehl et al. 2007; Fig. 10.6c), in large part due to the ice‐albedo feedback mechanism in which melting of snow and sea ice lowers reflectivity and, thereby, further increases surface warming by absorption of solar radiation (e.g., Weatherly et al. 1991). A vast and

46
rapidly growing body of information documenting this and other Arctic climate processes, and projecting future changes, has been comprehensively reviewed and assessed in widely available formats (ACIA 2005, IPCC 2007c, Walsh 2008), notwithstanding the very recent findings, which seem to be published on an almost weekly basis (e.g., Stroeve et al. 2008, Kwok and Rothrock 2009, Wang and Overland 2009, Zhang 2010).
Our focus in this section is to assess the observed and projected changes with significant potential to impact the ringed seal’s range and habitat, including both the physical and biological components of habitat. We address changes in sea ice, snow cover, ocean temperature, ocean pH (acidity), and associated changes in ringed seal predator and prey species.
4.2.1.1.1 Effects of climate change on the annual formation of ringed seals’ ice and snow habitat
Ringed seals depend on ice as a platform for resting, whelping, nursing, and molting, and they depend on snow cover to provide protection from cold and predators. Ice and snow cover are changing and will continue to do so as the climate warms.
Sea ice in the Northern Hemisphere can be divided into first‐year sea ice that formed in the most recent autumn‐winter period, and multi‐year sea ice that has survived at least one summer melt season. Multi‐year sea ice is generally confined to the Arctic Ocean. More southerly regions, such as the Bering Sea, Barents Sea, Baffin Bay, the Baltic Sea, Hudson Bay, and the Sea of Okhotsk are known as seasonal ice zones, where sea ice is renewed every winter. Similarly, freshwater ice in Lake Ladoga and Lake Saimaa forms and melts annually.
Sea‐ice extent at the end of summer (September) 2007 in the Arctic Ocean was a record low (4.3 million km2), nearly 40% below the long‐term average and 23% below the previous record set in 2005 (5.6 million km2) (Stroeve et al. 2008). Sea‐ice extent in September 2008 and 2009 was greater than in 2007 but still more than two standard deviations below the 1979‐2000 average. Most of the loss of sea ice was on the Pacific side of the Arctic. Of even greater long‐term significance was the loss of over 40% of Arctic multi‐year sea ice over the last 5 years (Kwok et al. 2009) (Figure 4). While the annual minimum of sea‐ice extent is often taken as an index of the state of Arctic sea ice, the recent reductions of the area of multi‐year sea ice and the reduction of sea‐ice thickness are of greater physical importance. It would take many years to restore the ice thickness through annual growth, and the loss of multi‐year sea ice makes it unlikely that the Arctic will return to previous climatological conditions. Continued loss of sea ice will be a major driver of changes across the Arctic over the next decades, especially in late summer and autumn.
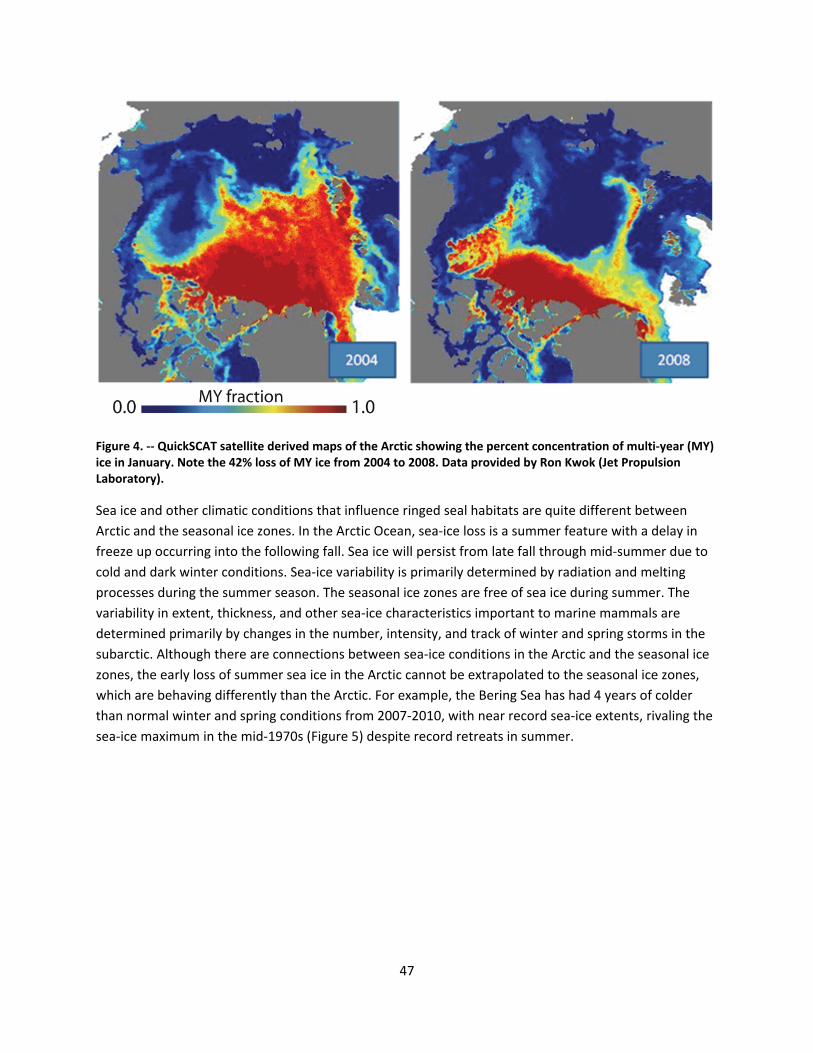
47
Figure 4. ‐‐ QuickSCAT satellite derived maps of the Arctic showing the percent concentration of multi‐year (MY) ice in January. Note the 42% loss of MY ice from 2004 to 2008. Data provided by Ron Kwok (Jet Propulsion Laboratory).
Sea ice and other climatic conditions that influence ringed seal habitats are quite different between Arctic and the seasonal ice zones. In the Arctic Ocean, sea‐ice loss is a summer feature with a delay in freeze up occurring into the following fall. Sea ice will persist from late fall through mid‐summer due to cold and dark winter conditions. Sea‐ice variability is primarily determined by radiation and melting processes during the summer season. The seasonal ice zones are free of sea ice during summer. The variability in extent, thickness, and other sea‐ice characteristics important to marine mammals are determined primarily by changes in the number, intensity, and track of winter and spring storms in the subarctic. Although there are connections between sea‐ice conditions in the Arctic and the seasonal ice zones, the early loss of summer sea ice in the Arctic cannot be extrapolated to the seasonal ice zones, which are behaving differently than the Arctic. For example, the Bering Sea has had 4 years of colder than normal winter and spring conditions from 2007‐2010, with near record sea‐ice extents, rivaling the sea‐ice maximum in the mid‐1970s (Figure 5) despite record retreats in summer.
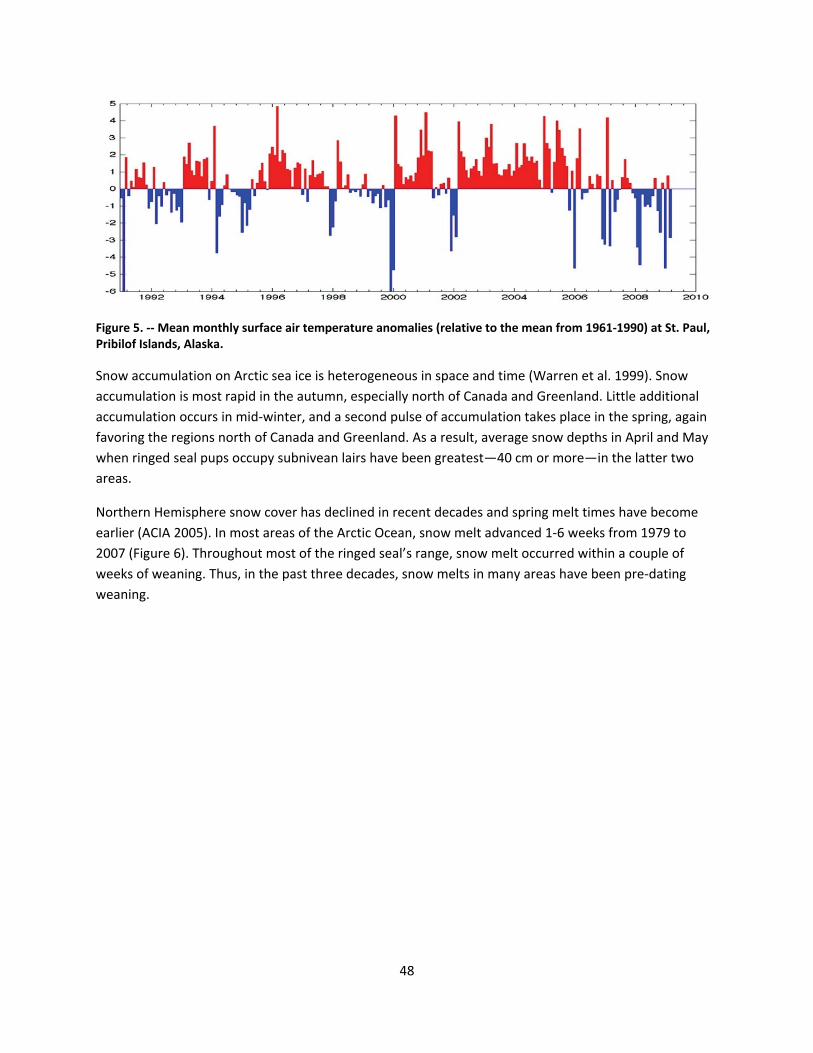
48
Figure 5. ‐‐ Mean monthly surface air temperature anomalies (relative to the mean from 1961‐1990) at St. Paul, Pribilof Islands, Alaska.
Snow accumulation on Arctic sea ice is heterogeneous in space and time (Warren et al. 1999). Snow accumulation is most rapid in the autumn, especially north of Canada and Greenland. Little additional accumulation occurs in mid‐winter, and a second pulse of accumulation takes place in the spring, again favoring the regions north of Canada and Greenland. As a result, average snow depths in April and May when ringed seal pups occupy subnivean lairs have been greatest—40 cm or more—in the latter two areas.
Northern Hemisphere snow cover has declined in recent decades and spring melt times have become earlier (ACIA 2005). In most areas of the Arctic Ocean, snow melt advanced 1‐6 weeks from 1979 to 2007 (Figure 6). Throughout most of the ringed seal’s range, snow melt occurred within a couple of weeks of weaning. Thus, in the past three decades, snow melts in many areas have been pre‐dating weaning.

49
Figure 6. ‐‐ Trends in the annual onset of snow melt on Arctic sea ice from 1979 to 2007 (Source: Drobot and Anderson 2001, updated 2009).
4.2.1.1.1.1 IPCC model projections
Comprehensive Atmosphere‐Ocean General Circulation Models (AOGCMs) are the major objective tools that scientists use to understand the complex interaction of processes that determine future climate change. The IPCC used the simulations from as many as two dozen AOGCMs developed by 17 international modeling centers as the basis for the AR4 (IPCC 2007a). Regional projections from these models are also being used by management agencies to assess and plan for future ecological and societal impacts (e.g., Scenarios Network for Alaska Planning: http://www.snap.uaf.edu/home). The AOGCM results are archived as part of the Coupled Model Intercomparison Project‐Phase 3 (CMIP3) at the Program for Climate Model Diagnosis and Intercomparison.
The IPCC’s AR4 emphasizes that current generation AOGCMs provide credible quantitative estimates of future climate change at continental scales and above (IPCC 2007a). The CMIP3 models are improved over the models used for the Third Assessment Report (Randall et al. 2007, Reichler and Kim 2008). Climate researchers are also making use of the AR4 AOGCM simulations on regional scales (e.g., Walsh et al. 2008). The Program for Climate Model Diagnosis and Intercomparison shows over 1,100 projects and over 500 publications using CMIP3, most based on regional studies. The CMIP3 AOGCMs provide reliable projections, because they are built on well‐known dynamical and physical principles, and they simulate quite well many large scale aspects of present‐day conditions (Randall et al. 2007, Knutti et al.

50
2008). Further, some biases in simulated climate by different models can be unsystematic (Räisänen 2007, Jun et al. 2008). The coarse resolution of most current climate models certainly dictates careful application on small scales in heterogeneous regions such as along coastlines or rugged orography. Our experience and that of other groups conducting model evaluations indicates the importance of multiple, complementary approaches.
There are three main contributors to the divergence in AOGCM climate projections: large natural variations, the range in emissions scenarios, and across‐model differences (Hawkins and Sutton 2009). First, it is known that if climate models are run several times with slightly different initial conditions, the trajectory of day‐to‐day and indeed year‐to‐year evolution will have different timing of events, even though the underlying statistical‐spectral character of the model climate tends to be similar for each run. This variability is a feature of the real climate system, and consumers of climate projections must recognize its importance. Natural variability is a source of ambiguity in the comparison of models with each other and with observational data. This variability can affect decadal or even longer means, so it is relevant to the use of model‐derived climate projections. The variability can be incorporated by averaging the projections over decades or, preferably, by forming ensemble averages from several runs of the same model.
A second source of variation in projections arises from the range in plausible emissions scenarios. Emissions scenarios have been developed based on assumptions for future development of humankind (Nakicenovic and Swart 2000); they are converted into GHG and aerosol concentrations, which are then used to drive the AOGCMs in the form of radiative forcing specified in the CMIP3 models and summarized in the IPCC AR4. For this study, we used emissions scenario A1B, a middle range case which is also the closest to the CO2 concentrations observed in the last decade, and A2, a high emissions scenario (IPCC 2000). Because of the residence time of carbon in the atmosphere and the thermal inertia of the climate system, climate projections are often somewhat insensitive to the precise details of which future emissions scenarios are used over the next few decades. The impacts of the scenarios are rather similar before mid‐21st century (Hawkins and Sutton 2009). For the second half of the 21st century, however, and especially by 2100, the choice of the emissions scenario becomes the major source of variation among climate projections and dominates over natural variability and model‐to‐model differences (IPCC 2007a). If 2030‐2050 is a timescale of interest, climatologists will often use the A1B scenario—or A1B and A2 together—to increase the number of potential ensemble members as their CO2 trajectories are similar before 2050.
The third source of variability is termed across‐model uncertainty2 (Knutti et al. 2008). Different numerical approximations of the model equations, spatial resolution, and other model development factors introduce structural uncertainty between different models. The potential for model bias introduces uncertainty. Because this bias is rarely systemic, however, this uncertainty can be addressed and mitigated in part by using the ensemble means from multiple models (Overland et al. In review).
2 Note that here we use “uncertainty” to refer to statistical measures of variation that should not be confused with the more general use of the term to refer to something that is unreliable or untrustworthy.

51
There is no universal method for combining AOGCMs for climate projections (Gleckler et al. 2008, Räisänen et al. 2010), and there is no one best model. Our objective was to reduce the impact of models with large hindcast error. We culled the poor performing models—rather than the selecting the best models—and retained several models as a measure of model variability.
4.2.1.1.1.2 Northern Hemisphere
4.2.1.1.1.2.1 Data and analytical methods
The CMIP3 model simulations used in our analyses were obtained from the Program for Climate Model Diagnosis and Intercomparison online (PCMDI 2010). There are several sources of sea‐ice data available; we chose to use the gridded sea‐ice concentration analysis from the Hadley Centre (MET Office Hadley Data Centre 2008) as the observed values in this study. Conventionally, sea‐ice extent was then defined as the area where the ice concentration is more than 15% in a grid box.
Confidence that climate models provide credible quantitative projections of future climate is built upon their demonstrated ability to reproduce observed features of recent climate (Gerdes and Köberle 2007). It is therefore important to apply an observational constraint on the CMIP3 models, and cull poor performing models from further consideration. Inspired by Knutti et al. (2006), Wang and Overland (2009) required that models simulate the seasonal cycle and the mean of September sea‐ice extent to within 20% of the Hadley Centre sea‐ice and sea surface temperature data set (HadISST) analysis for the period of 1980‐1999. Wang and Overland (2009) provided the basic model for sea‐ice projection analysis used in this report. Reproducing the correct magnitude of the seasonal cycle of sea‐ice extent is one way of demonstrating the models’ sensitivity to changes in external forcing (e.g., solar insolation). The September mean sea‐ice extent is an efficient constraint to eliminate models with systematic biases. Our constraints are based on comparisons to the HadISST sea‐ice concentration analysis, which was made more homogeneous by compensating satellite microwave‐based sea‐ice concentrations for the impact of surface melt effects in the Arctic (Rayner et al. 2003). The combination of the seasonal cycle and mean conditions is an improved constraint relative to previous studies (Overland and Wang 2007, Stroeve et al. 2007). The culling process not only reduces the range of variability in the future model projections, but it also shows that models with a reasonable seasonal cycle relative to the observations project a faster decline of future September sea‐ice extent (Figure 7).
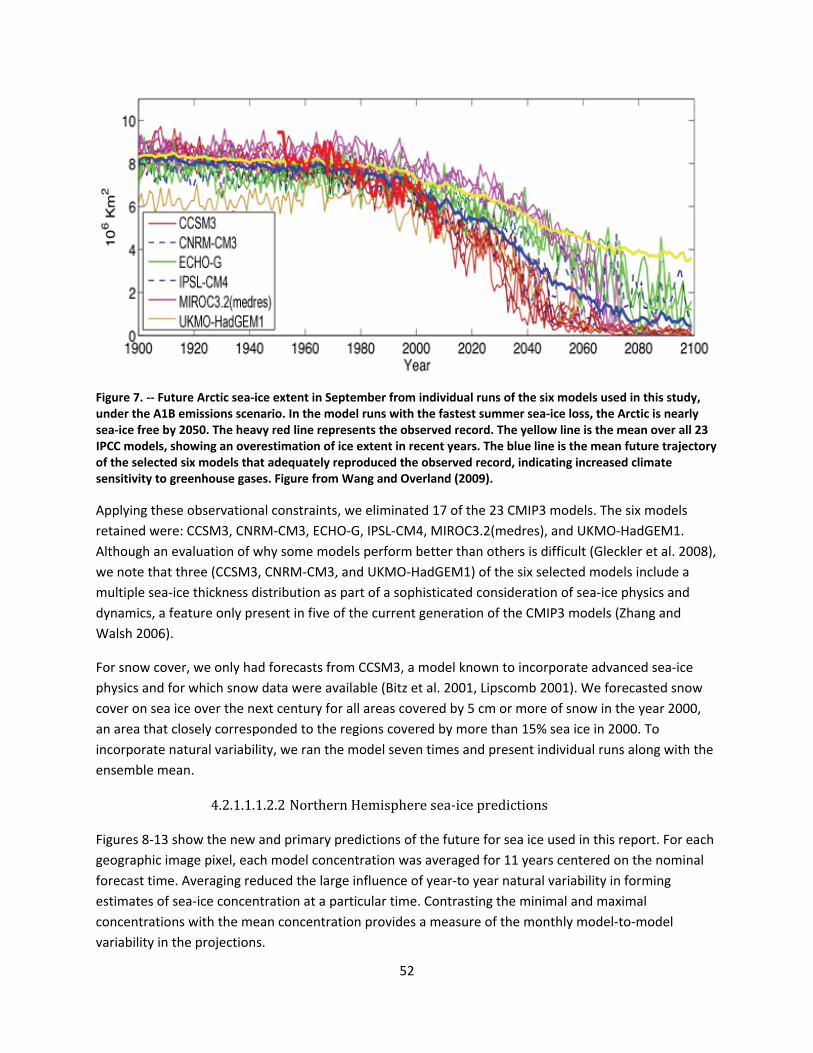
52
Figure 7. ‐‐ Future Arctic sea‐ice extent in September from individual runs of the six models used in this study, under the A1B emissions scenario. In the model runs with the fastest summer sea‐ice loss, the Arctic is nearly sea‐ice free by 2050. The heavy red line represents the observed record. The yellow line is the mean over all 23 IPCC models, showing an overestimation of ice extent in recent years. The blue line is the mean future trajectory of the selected six models that adequately reproduced the observed record, indicating increased climate sensitivity to greenhouse gases. Figure from Wang and Overland (2009).
Applying these observational constraints, we eliminated 17 of the 23 CMIP3 models. The six models retained were: CCSM3, CNRM‐CM3, ECHO‐G, IPSL‐CM4, MIROC3.2(medres), and UKMO‐HadGEM1. Although an evaluation of why some models perform better than others is difficult (Gleckler et al. 2008), we note that three (CCSM3, CNRM‐CM3, and UKMO‐HadGEM1) of the six selected models include a multiple sea‐ice thickness distribution as part of a sophisticated consideration of sea‐ice physics and dynamics, a feature only present in five of the current generation of the CMIP3 models (Zhang and Walsh 2006).
For snow cover, we only had forecasts from CCSM3, a model known to incorporate advanced sea‐ice physics and for which snow data were available (Bitz et al. 2001, Lipscomb 2001). We forecasted snow cover on sea ice over the next century for all areas covered by 5 cm or more of snow in the year 2000, an area that closely corresponded to the regions covered by more than 15% sea ice in 2000. To incorporate natural variability, we ran the model seven times and present individual runs along with the ensemble mean.
4.2.1.1.1.2.2 Northern Hemisphere sea‐ice predictions
Figures 8‐13 show the new and primary predictions of the future for sea ice used in this report. For each geographic image pixel, each model concentration was averaged for 11 years centered on the nominal forecast time. Averaging reduced the large influence of year‐to year natural variability in forming estimates of sea‐ice concentration at a particular time. Contrasting the minimal and maximal concentrations with the mean concentration provides a measure of the monthly model‐to‐model variability in the projections.
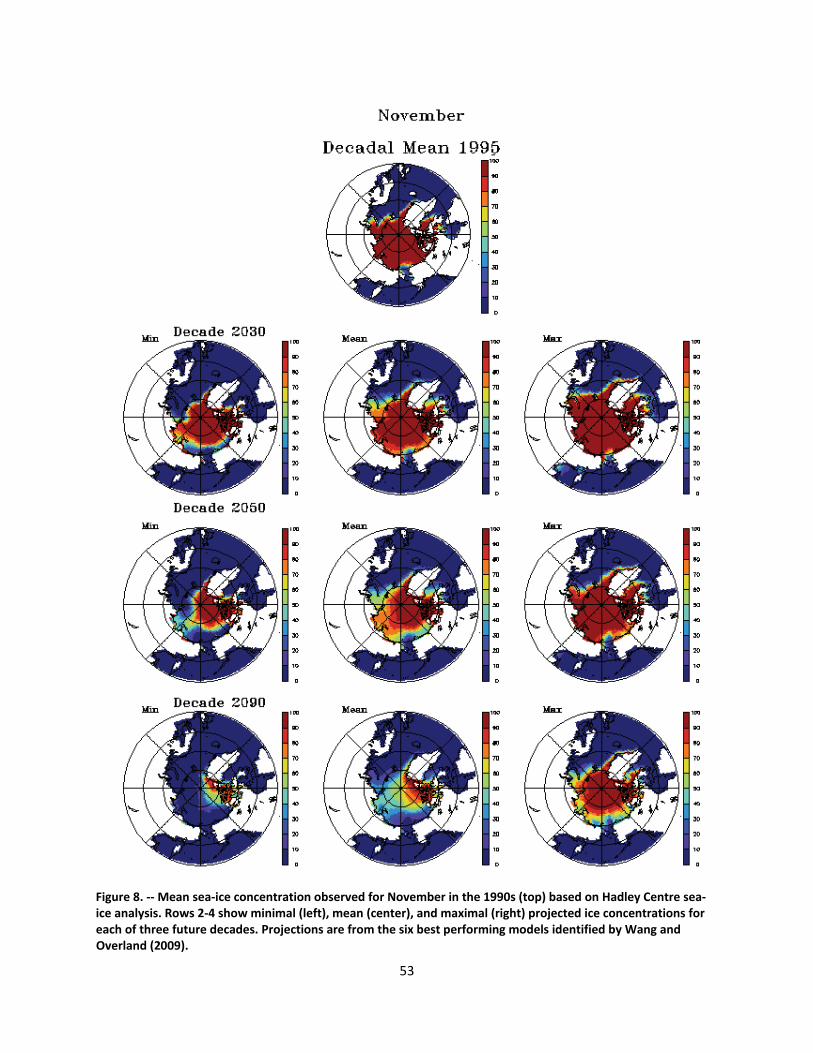
53
Figure 8. ‐‐ Mean sea‐ice concentration observed for November in the 1990s (top) based on Hadley Centre sea‐ ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).
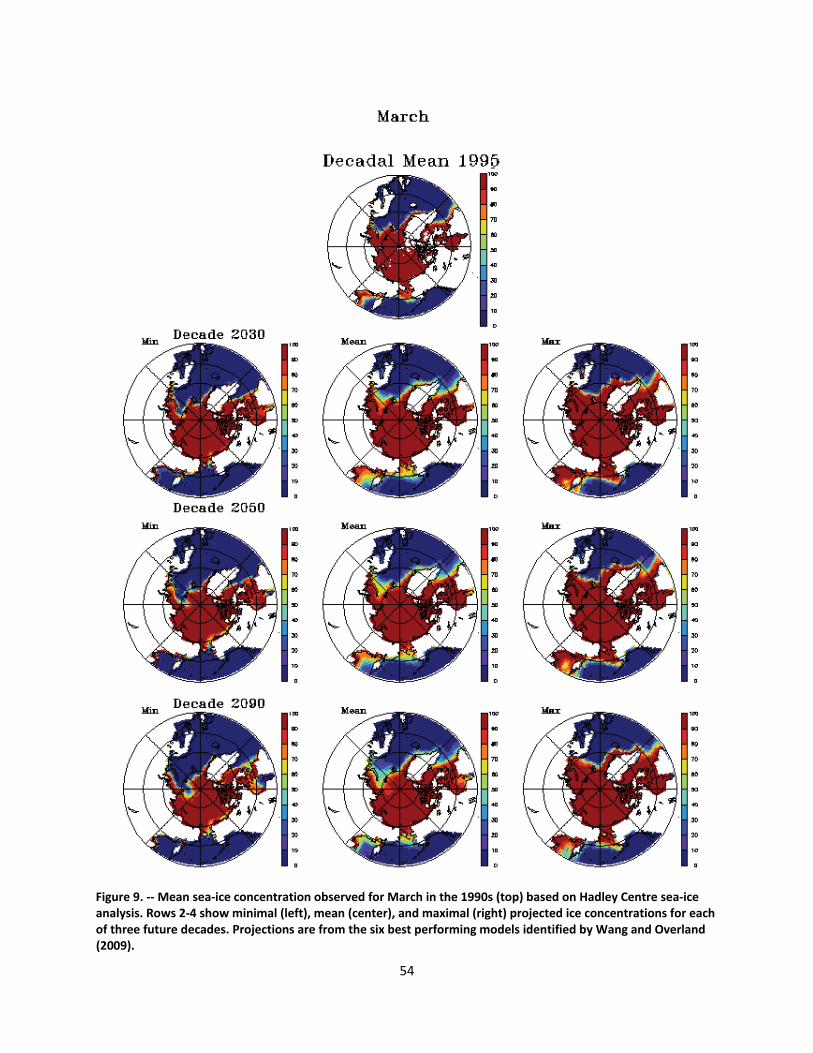
54
Figure 9. ‐‐ Mean sea‐ice concentration observed for March in the 1990s (top) based on Hadley Centre sea‐ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).
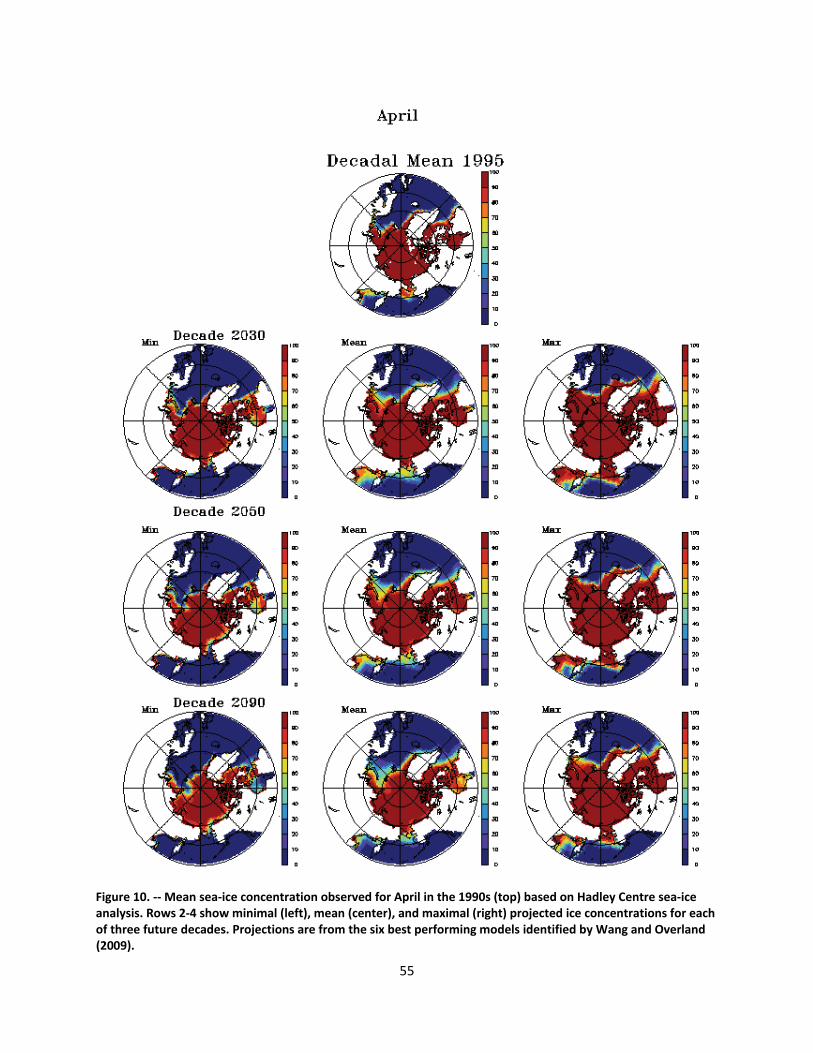
55
Figure 10. ‐‐ Mean sea‐ice concentration observed for April in the 1990s (top) based on Hadley Centre sea‐ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).
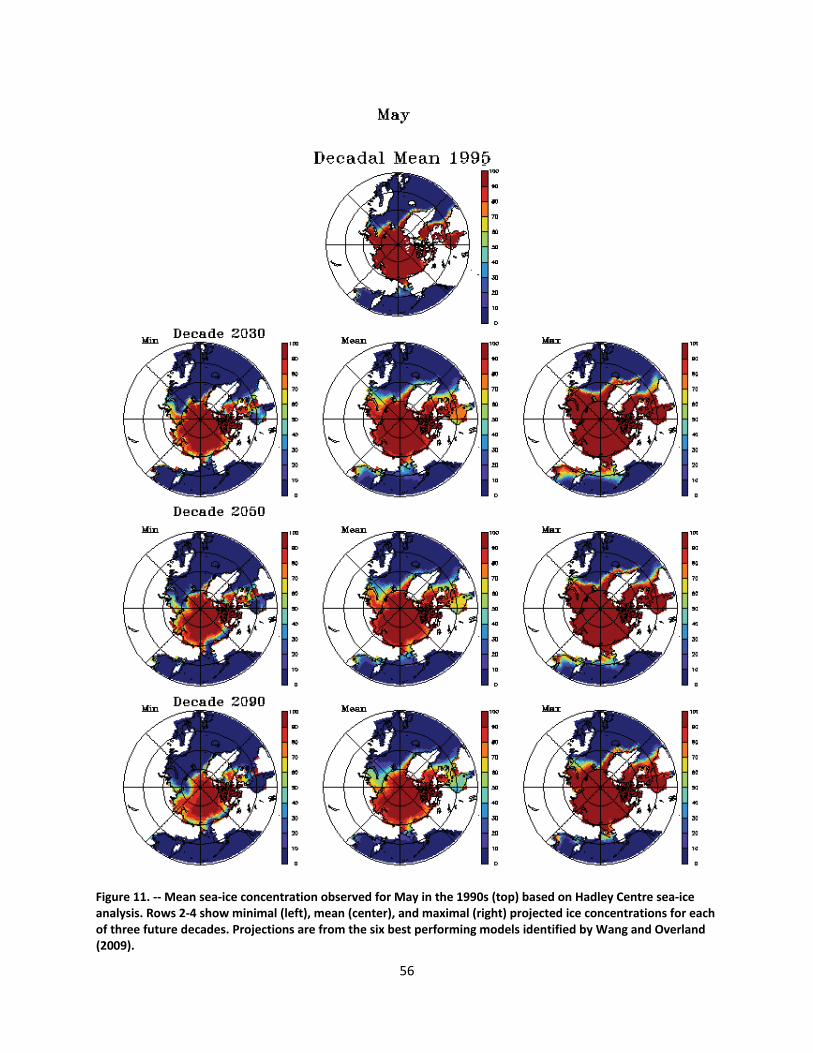
56
Figure 11. ‐‐ Mean sea‐ice concentration observed for May in the 1990s (top) based on Hadley Centre sea‐ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).
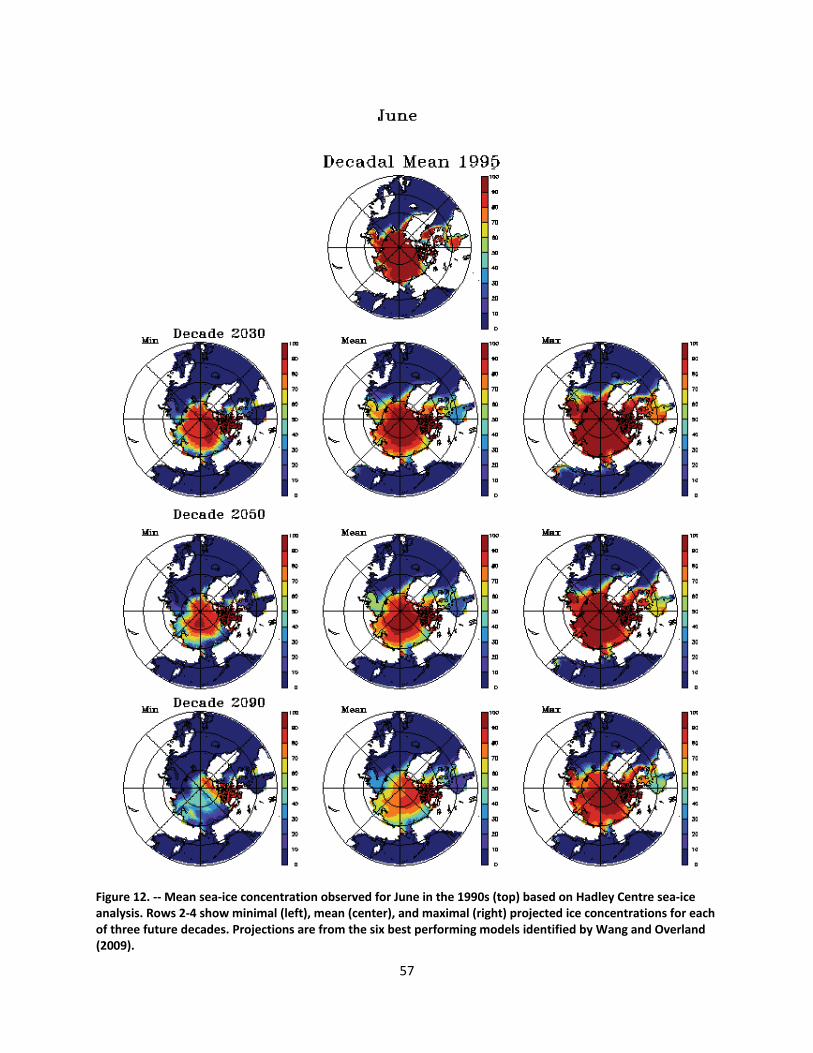
57
Figure 12. ‐‐ Mean sea‐ice concentration observed for June in the 1990s (top) based on Hadley Centre sea‐ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).
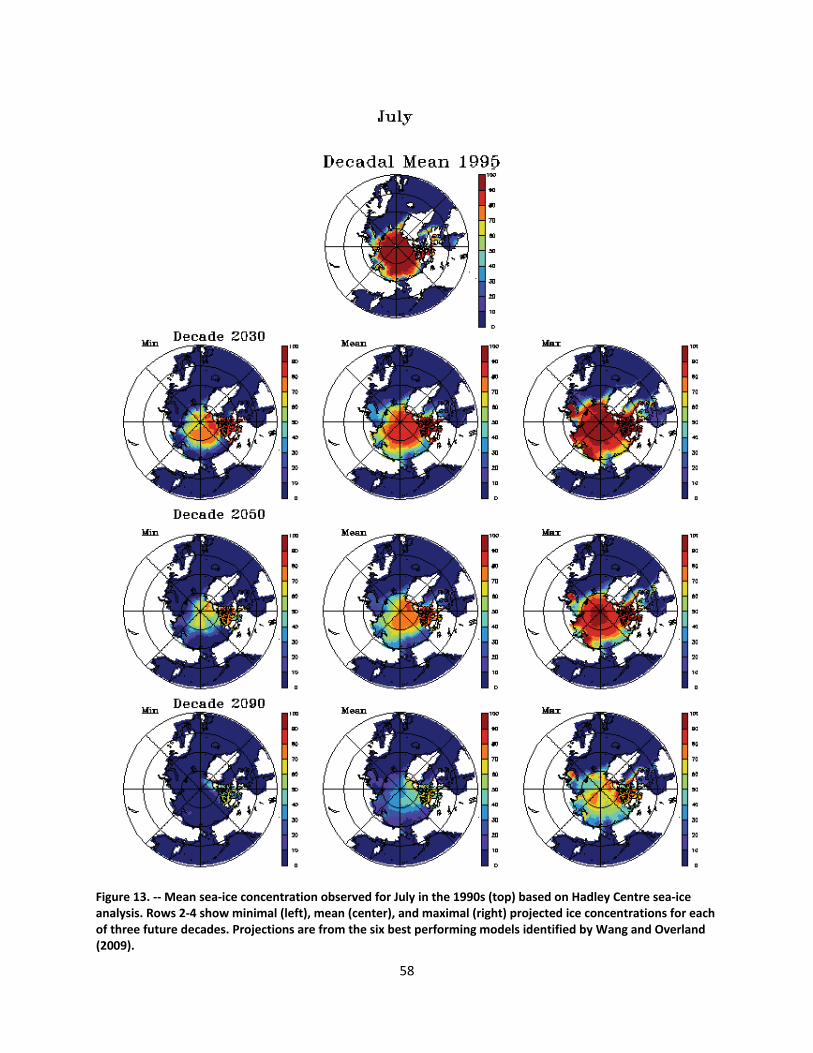
58
Figure 13. ‐‐ Mean sea‐ice concentration observed for July in the 1990s (top) based on Hadley Centre sea‐ice analysis. Rows 2‐4 show minimal (left), mean (center), and maximal (right) projected ice concentrations for each of three future decades. Projections are from the six best performing models identified by Wang and Overland (2009).

59
The projections for November (Figure 8) indicate a major delay in fall freeze‐up by 2050 north of Alaska and in the Barents Sea. By 2090, the average sea‐ice concentration is below 50% in the Russian Arctic and some models show a nearly ice free Arctic, except for the region of the Canadian Arctic Archipelago. In March and April (Figures 9 and 10, respectively), generally winter type conditions persist out to 2090. There is some reduction of sea ice by 2050 in the outer portions of the seasonal ice zones, but the sea ice south of Bering Strait, eastern Barents Sea, Baffin Bay, and the Kara and Laptev Seas remains substantial. May (Figure 11) is similar to April but with diminishing sea‐ice cover at 2050 and 2090 in the Barents Sea and Sea of Okhotsk. The month of June (Figure 12) begins to show substantial changes as the century progresses. Current conditions occasionally exhibit a lack of sea ice near the Bering Strait by mid‐June. By 2050, however, this sea‐ice loss becomes a major feature with open water continuing along the northern Alaskan coast in most models. Open water in June spreads to the East Siberian Shelf by 2090. The eastern Barents Sea experiences a reduction in sea ice between 2030 and 2050. The models indicate that sea ice in Baffin Bay will be affected very little until the end of the century. In July (Figure 13), the Arctic Ocean shows a marked effect of global warming, with the sea ice retreating to a central core as the century progresses. The loss of multi‐year sea ice over the last 5 years has provided independent evidence for this conclusion. By 2050, the continental shelves of the Beaufort, Chukchi, and East Siberian Seas are nearly ice free in July with ice concentrations less than 20% in the ensemble mean projections. The Kara and Laptev Seas also show a reduction of sea ice in coastal regions by mid‐century in most but not all models. The Canadian Arctic Archipelago and the adjacent Arctic Ocean north of Canada and Greenland, however, are predicted to become a refuge for sea ice through the end of the century. This conclusion is supported by typical Arctic wind patterns which tend to blow onshore in this region. Indeed, this refuge region is why sea‐ice scientists use the phrase: “a nearly sea ice free summer Arctic by mid‐century.”
4.2.1.1.1.3 Sub‐regions
4.2.1.1.1.3.1 Regional boundaries and overview of methods
Based on the six models identified by Wang and Overland (2009), the decadal mean sea‐ice concentration shown in Figures 8‐13 clearly indicate that sea ice declines at faster rate in the fall (November) and early summer (July) than in late winter (March) or spring (April, May, and June). Climate models generally perform better on continental or larger scales, but because habitat changes are not uniform throughout the hemisphere, we further evaluated these six models independently on their performance at 14 different sub‐regions throughout the ringed seals’ range: Baffin Bay (50‐80°N, 70‐45°W), Baltic Sea (53‐65°N, 12‐33°E), Barents Sea (65‐80°N, 12‐60°E), Beaufort Sea (70‐80°N, 157‐112°W), eastern Bering Sea (54‐66°N, 175‐157°W), western Bering Sea (54‐66°N, 165°E‐175°W), Canadian Arctic Archipelago (62‐84°N, 112‐70°W), central Arctic (80‐90°N, 180°W‐180°E), Chukchi Sea (66‐80°N, 180‐157°W), East Siberian Sea (70‐80°N, 145‐180°E), Greenland Sea (60‐84°N, 45°W‐12°E), Hudson Bay (50‐62°N, 95‐70°W), Kara‐Laptev Seas (70‐80°N, 60‐145°E), and the Sea of Okhotsk (44‐62°N, and 137‐162°E). The boundaries of each selected region are shown in Figure 14.

60
All six of the models met the performance criteria for predicting sea ice in the Chukchi Sea, East Siberian Sea, and the central Arctic. To forecast sea‐ice extent and concentration in the other regions, we used a subset of models, results from previous analyses, or other analytical methods appropriate for the region:
• Beaufort Sea – Four of the six models (CCSM3, ECHO‐G, IPSL‐CM4, and UKMO‐HadGEM1) simulated both the mean and seasonal cycle of Beaufort Sea ice extent with reasonable agreement with observations, so the projections are based on these four models.
• Eastern Bering Sea – Four of the six models (CCSM3, CNRM‐CM3, ECHO‐G, and MIROC3.2(medres)) met the criteria, which still allowed projections to be made from a basis that includes model‐to‐model variation and sufficient numbers of available model runs.
• Western Bering and the Barents Seas – Only one of the six models (CCSM3) met the performance criteria in these regions and most of the other models tended to overestimate the observed ice extent year‐round. We used the single model as the basis for projecting sea ice with caveats about the reliability described below.
• None of the models performed satisfactorily in the remaining regions and so we employed other methods to predict sea ice:
o Existing analyses for the Baltic Sea (Jylhä et al. 2008) and Hudson Bay (Joly et al. 2010) were used to infer ice conditions in those areas.
o For the Sea of Okhotsk, we investigated the use of SAT for projecting future ocean and ice conditions.
o For Baffin Bay, the Canadian Arctic Archipelago, and the Greenland, Kara, and Laptev Seas, we performed a qualitative analysis of seasonal and decadal changes using results of the hemispheric predictions with caveats about the reliability described below.
Below we present details of these analyses for each region. Whenever the relevant model outputs were available, we analyzed the conditions projected under the “medium” A1B and “high” A2 emissions scenarios (IPCC 2000). By including both the A1B and A2 emissions scenario, we doubled the number of ensemble members and represented much of the range of variability contained in the SRES scenarios. For regions where ice projections could not be obtained, we present results of the other regionally appropriate analyses.
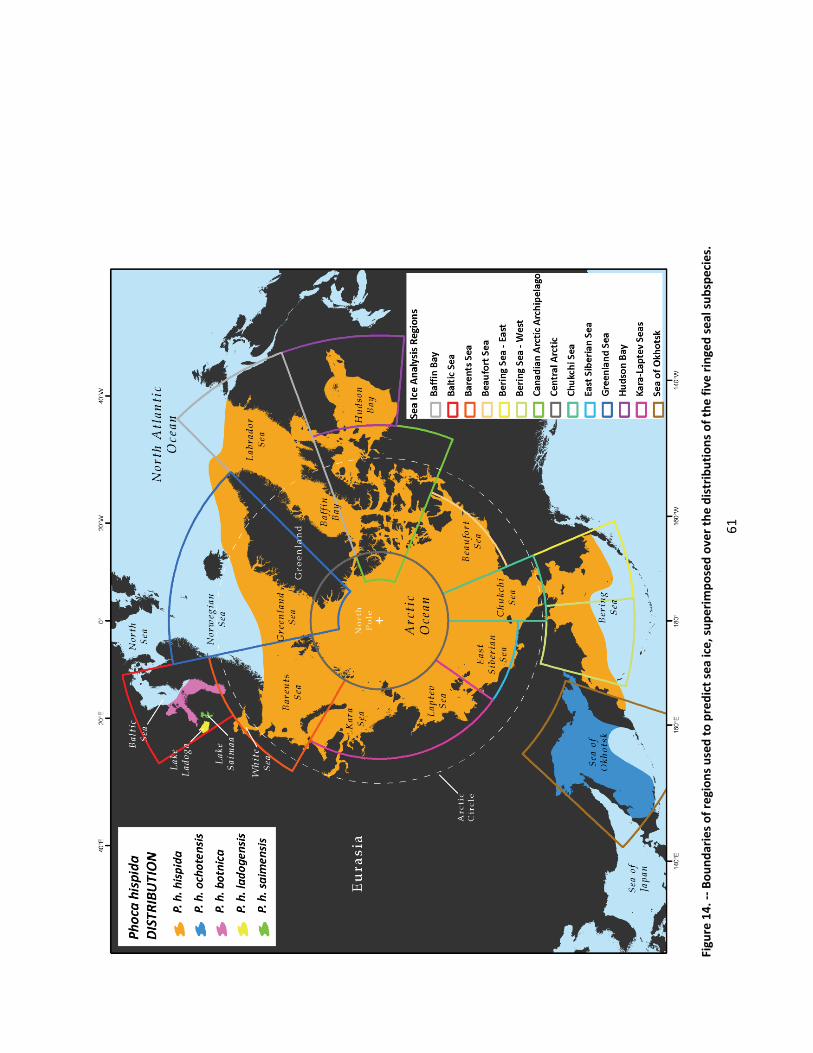
61
Figure 14. ‐‐ Bou
ndaries of regions used to predict sea
ice, sup
erim
posed over th
e distribu
tion
s of th
e fiv
e ringed
seal sub
species.

62
4.2.1.1.1.3.2 Regional sea‐ice predictions
4.2.1.1.1.3.2.1 Chukchi Sea
The Chukchi Sea is located north of the Arctic Circle, with its northern boundary adjoining the Arctic Ocean. Sea ice starts to retreat in late May or later, and part of the region will be covered by ice during summer. We found no significant ice reduction projected for winter and early spring (January to May). This is in contrast to sharp declining trends near the end of the 21st century for the autumn and early summer (i.e., November and July (Figure 15)). The downward trends are especially apparent after 2070 in the projections for November and July, and only significant by the end of the century for June, but by then the difference between the emissions scenarios (blue for A1B, and magenta for A2; Figure 15) becomes a major contributor to the trends. It is also obvious from the figures that the model projections are more variable after the mid‐century, which is shown by a wider spread of the area covered by the grey lines.
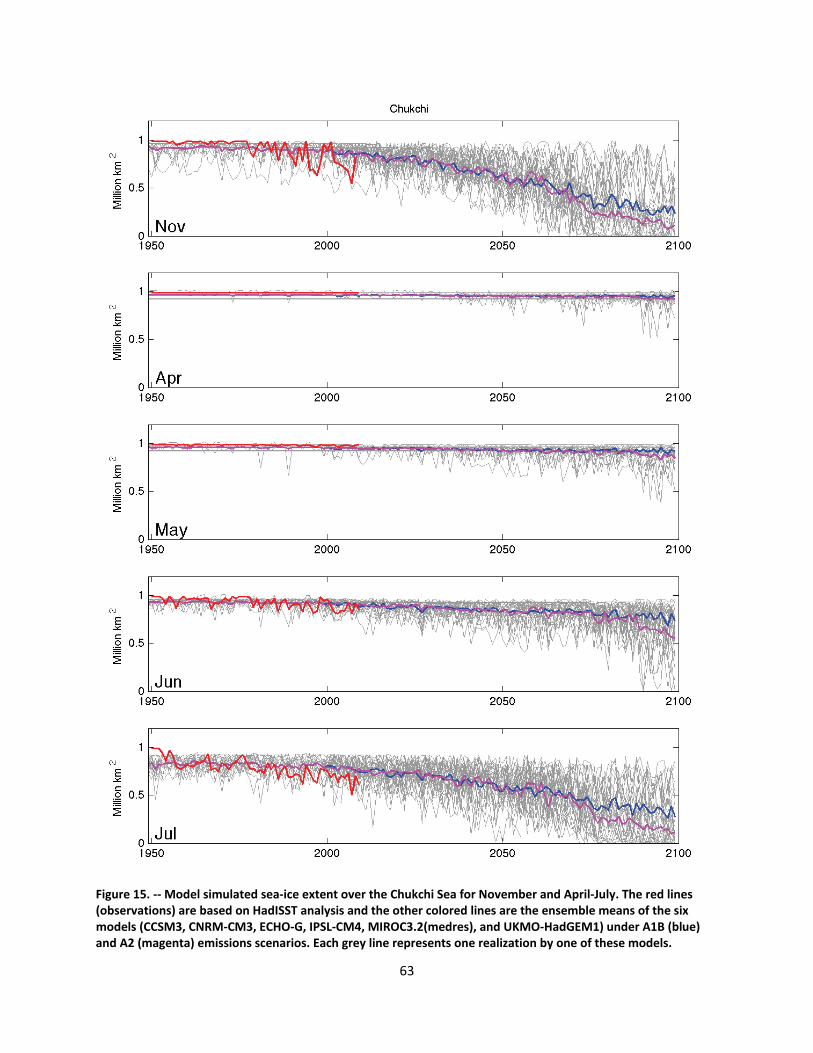
63
Figure 15. ‐‐ Model simulated sea‐ice extent over the Chukchi Sea for November and April‐July. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the six models (CCSM3, CNRM‐CM3, ECHO‐G, IPSL‐CM4, MIROC3.2(medres), and UKMO‐HadGEM1) under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by one of these models.

64
4.2.1.1.1.3.2.2 East Siberian Sea
All six models simulated the seasonal cycle of the sea‐ice extent over the East Siberian Sea in reasonable agreement with observations. The projection from these six models shows that the decline of sea ice will be obvious only after 2050 for November and July (Figure 16). The change in winter and spring ice condition is minimal.
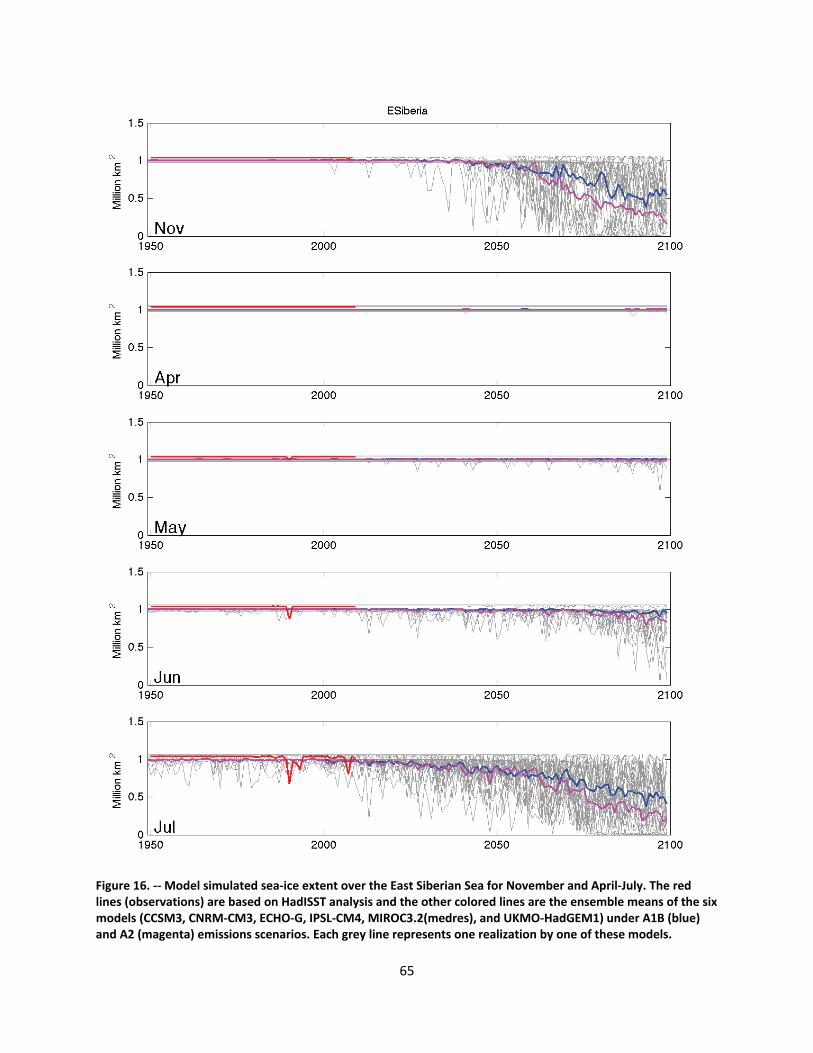
65
Figure 16. ‐‐ Model simulated sea‐ice extent over the East Siberian Sea for November and April‐July. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the six models (CCSM3, CNRM‐CM3, ECHO‐G, IPSL‐CM4, MIROC3.2(medres), and UKMO‐HadGEM1) under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by one of these models.

66
4.2.1.1.1.3.2.3 Central Arctic
The central Arctic is defined as the regions north of 80°N. In the past, this area has been completely covered by sea ice throughout the entire year (Figures 8‐13, top row). This pattern changed in the late summer of 2007, however, when for the first time, satellites observed a reduction in sea‐ice extent to less than 5 million km2. This reduction moved the ice edge north as far as 85°N, leaving a large area with open water in the eastern central Arctic. Although the ice extent had grown again by the end of summer in 2008 and 2009, these extents were still 2 standard deviations below the 1979‐2000 average. More importantly, in the past five years multi‐year sea ice in the Arctic has declined over 40% (Kwok et al. 2009, see Fig. 1). The first‐year sea ice which has replaced the multi‐year ice is thinner, more fragile, easier to melt, and more readily exported by wind. As a consequence, we can expect increasing rates of sea‐ice decline. The increased rate of sea‐ice loss is most obvious in the fall (November) and early summer (July) after 2050 (Figure 17).
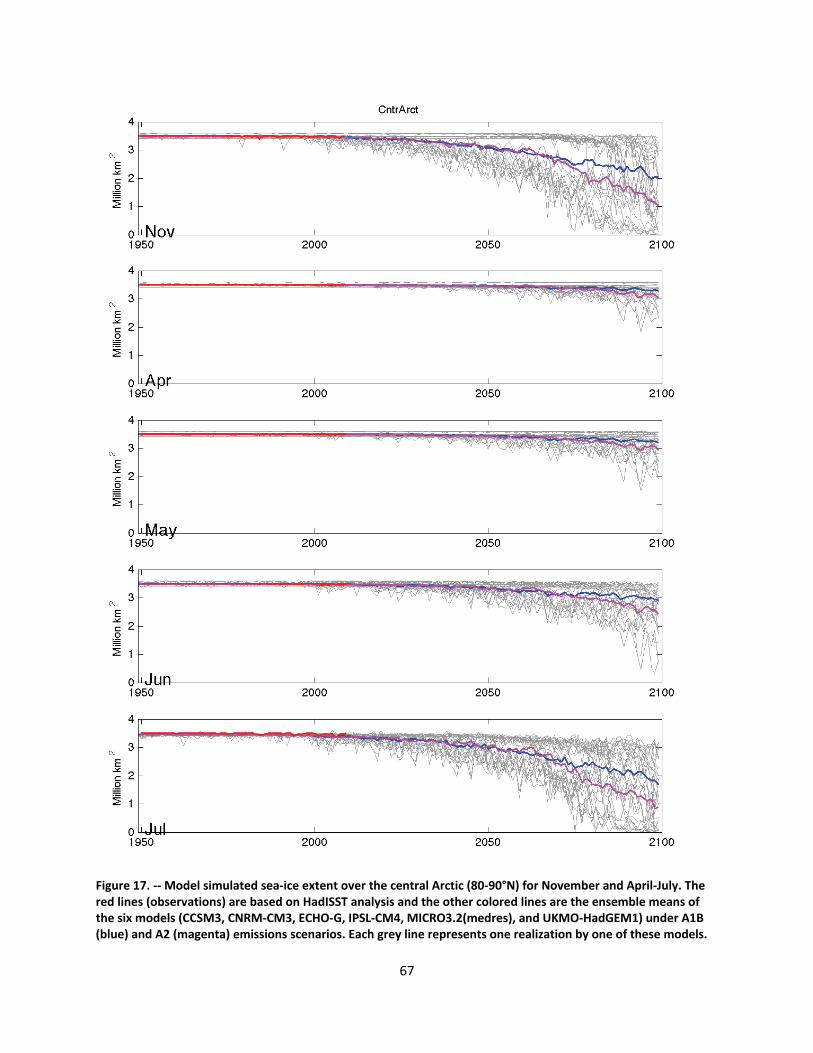
67
Figure 17. ‐‐ Model simulated sea‐ice extent over the central Arctic (80‐90°N) for November and April‐July. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the six models (CCSM3, CNRM‐CM3, ECHO‐G, IPSL‐CM4, MICRO3.2(medres), and UKMO‐HadGEM1) under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by one of these models.

68
4.2.1.1.1.3.2.4 Beaufort Sea
All six models simulated the seasonal cycle of sea‐ice extent over the Beaufort Sea in reasonable agreement with observations, although two of them underestimated the summer sea‐ice extent. The projections for the future are therefore made from four models (CCSM3, ECHO‐G, IPSL‐CM4, and UKMO‐HadGEM1). Sea ice is forecasted to be present in the Beaufort Sea in the winter and spring, even until the end of 21st century. In early summer (July) and fall (November), we see an accelerated decline after 2050 (Figure 18). The shortened period of sea‐ice cover in the Beaufort Sea suggests that coastal areas might be impacted the most.
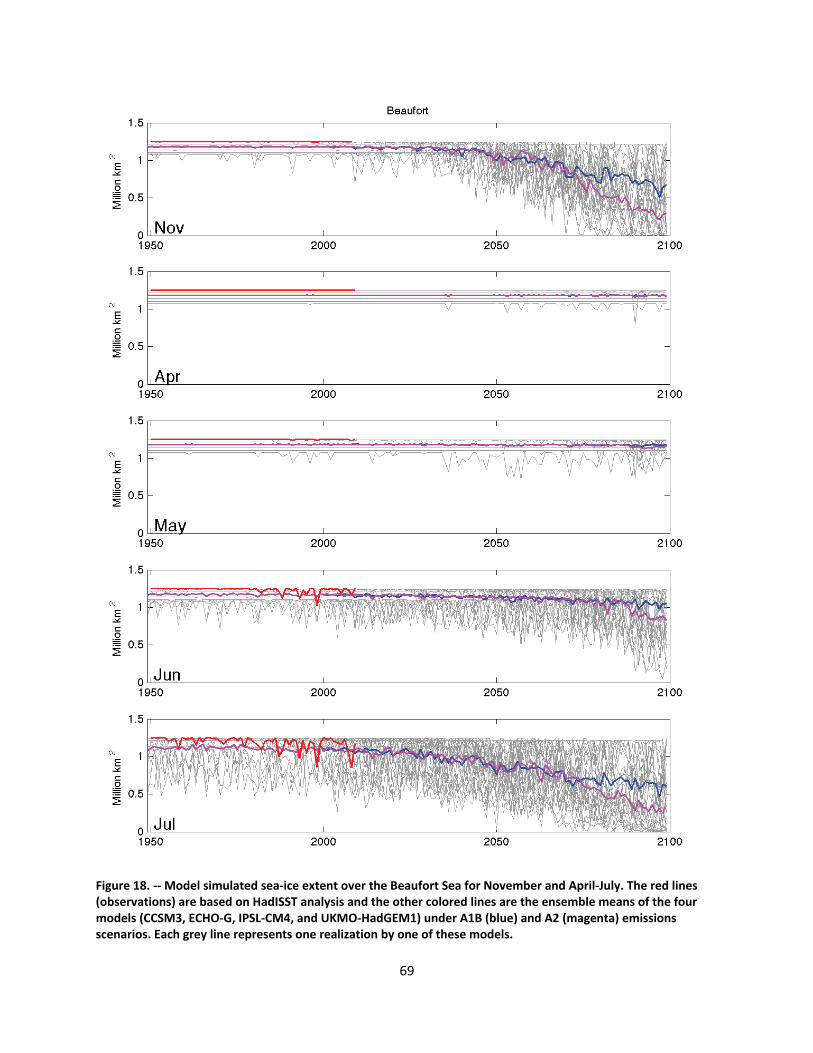
69
Figure 18. ‐‐ Model simulated sea‐ice extent over the Beaufort Sea for November and April‐July. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the four models (CCSM3, ECHO‐G, IPSL‐CM4, and UKMO‐HadGEM1) under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by one of these models.

70
4.2.1.1.1.3.2.5 Eastern Bering Sea
Sea ice begins to cover the eastern Bering Sea in November, and gradually advances south to reach its maximum in March. Figure 19 shows the projections of sea‐ice extent over the eastern Bering Sea for March, April, May, and June. From March to May, the interannual variability of sea‐ice extent is large, and the overall downward trend is small, but visually obvious. Very little ice has remained in the eastern Bering in June since the mid‐1970s. At the scale of these models, very little change in extent is predicted to the end of the century. The largest decline in sea‐ice extent is projected to occur in the late autumn months of November and December (not shown). By 2050, the averaged sea‐ice extent in late autumn would be 28% of the 1980‐1999 mean, whereas the average spring sea‐ice extent (average of March to May) would be at 58% of the present value. By 2075, the average spring sea‐ice extent would decline to 37% of present day value, and the autumn average extent (not shown) would be at only 12% of present day value.
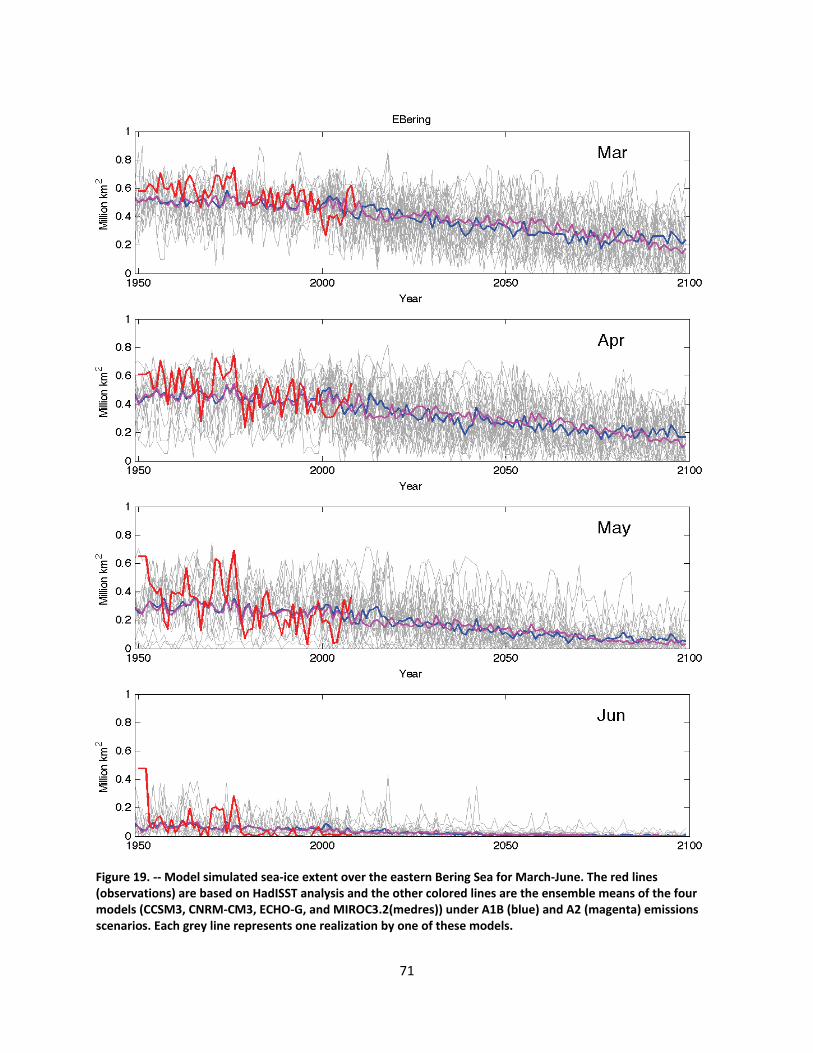
71
Figure 19. ‐‐ Model simulated sea‐ice extent over the eastern Bering Sea for March‐June. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the four models (CCSM3, CNRM‐CM3, ECHO‐G, and MIROC3.2(medres)) under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by one of these models.

72
4.2.1.1.1.3.2.6 Western Bering Sea
As noted above, we discovered that the majority of models tended to overestimate the sea‐ice coverage for the western Bering Sea in winter, with one model underestimating the ice conditions, and only one model (CCSM3) passing the selection criteria. For reference, we provide this single model’s output and caution that the results must be interpreted in the context of possible bias and lack of model‐to‐model variation. Projections from a single model may fail to represent the full range of variability or may be subject to biases of a particular model formulation, perhaps reducing confidence in the output for this region. The western Bering Sea projections are shown in Figure 20 for March to June. Compared with historical observations, this model overestimated sea‐ice extent in both March and April, but performed reasonably well for May and June. It projected a rapid decline in sea‐ice extent over the first half of the 21st century, then a stable rate of decline to the end of the century (top 3 panels of Figure 20). The mean linear trends estimated from the CCSM3 model were 28 x 103 km2/decade (8%/decade) and 17 x 103 km2/decade (9%/decade) for April and May, respectively, during the 21st century. Under these scenarios, the western Bering Sea is projected to have ice in March and April through nearly the end of the 21st century, though the average extent in the latter half‐century would be approximately 25% of the present‐day extent. The projection for May indicates that there will commonly be years with little or no sea ice beyond mid‐century.
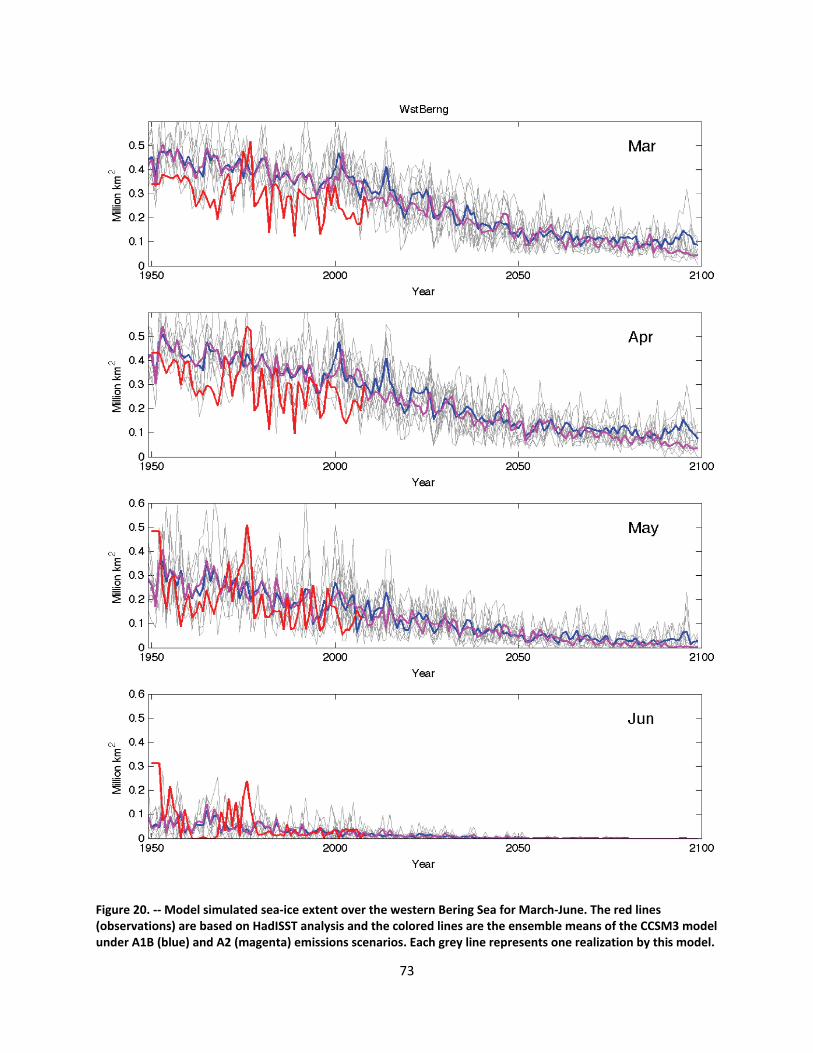
73
Figure 20. ‐‐ Model simulated sea‐ice extent over the western Bering Sea for March‐June. The red lines (observations) are based on HadISST analysis and the colored lines are the ensemble means of the CCSM3 model under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by this model.

74
To put the seasonal time series of sea‐ice extent for both the eastern and western Bering Sea into a geographic perspective, an example of the ice concentration projected by the CCSM3 model for May and November during the coming decades is shown in Figure 21. This clearly shows a spatial view of how the average November ice extent is projected to decrease faster than the average May ice extent.
Figure 21. ‐‐ The shaded area shows an example of projected sea‐ice concentration under the A1B emissions scenario from one run of the CCSM3 model for May (left) and November (right). The colored lines indicate the average position of the ice edge, which is defined as 15% ice concentration in a grid box, in the recent past (1980‐1999, black), in 2011‐2019 (red), in 2031‐2039 (green), and in 2051‐2059 (blue).
4.2.1.1.1.3.2.7 Barents Sea
Previous studies have shown that most models tend to have a cold bias in their SAT simulations and excessive sea‐ice cover over the Barents Sea region (Overland et al. In review). Indeed only one of the six models (CCSM3) simulates the seasonal cycles of the sea‐ice extent over the Barents Sea region satisfactorily. Figure 22 shows the sea‐ice extent projected by CCSM3 for November and from April through July. As this projection is from only one model, caution must be used when considering the future state of the Barents Sea sea‐ice extent. The model simulates the April and May ice conditions in reasonable agreement with observations and projects that ice will continue to decline in the region at constant linear rate throughout the 21st century. The model underestimates the ice conditions in the fall (November), however, and for the months of June and July, it predicts that ice will disappear rapidly in the coming decades.
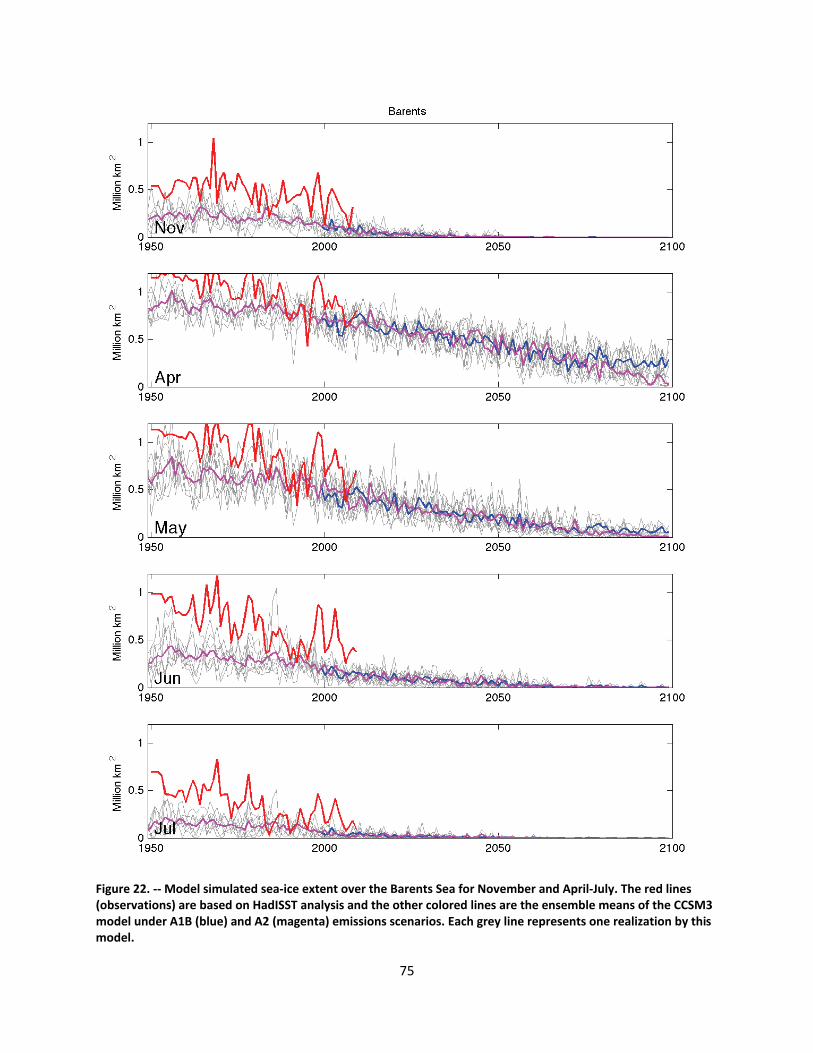
75
Figure 22. ‐‐ Model simulated sea‐ice extent over the Barents Sea for November and April‐July. The red lines (observations) are based on HadISST analysis and the other colored lines are the ensemble means of the CCSM3 model under A1B (blue) and A2 (magenta) emissions scenarios. Each grey line represents one realization by this model.
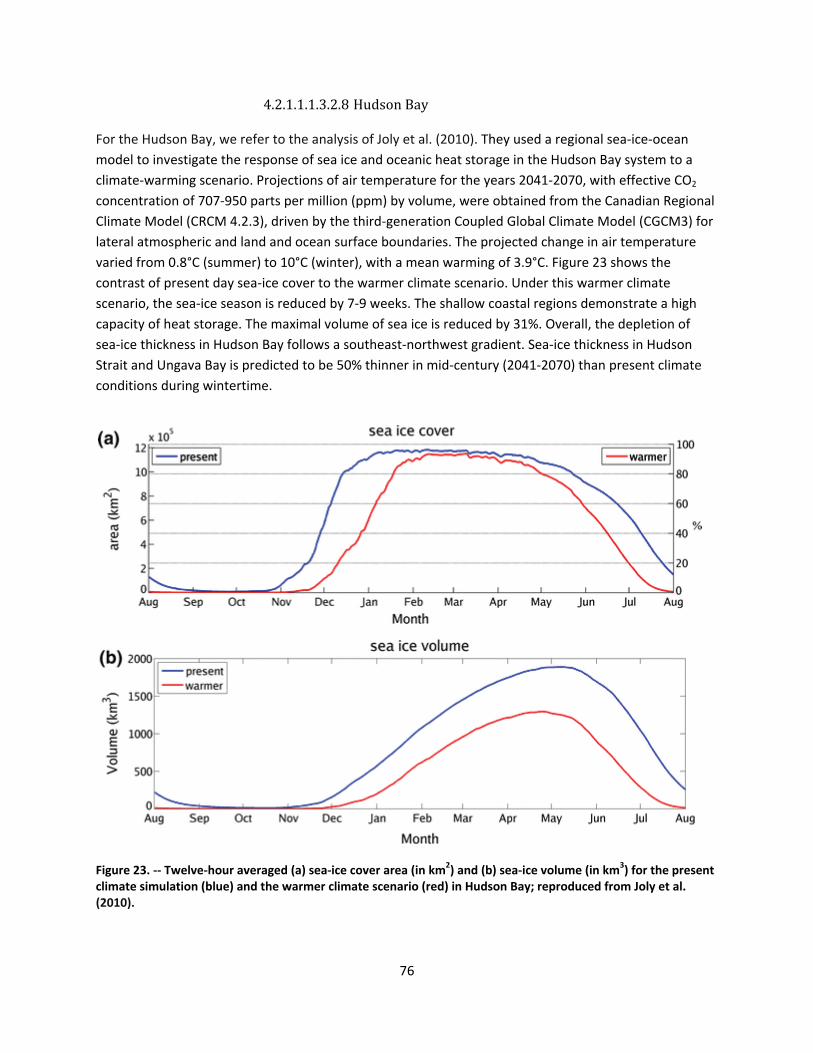
76
4.2.1.1.1.3.2.8 Hudson Bay
For the Hudson Bay, we refer to the analysis of Joly et al. (2010). They used a regional sea‐ice‐ocean model to investigate the response of sea ice and oceanic heat storage in the Hudson Bay system to a climate‐warming scenario. Projections of air temperature for the years 2041‐2070, with effective CO2 concentration of 707‐950 parts per million (ppm) by volume, were obtained from the Canadian Regional Climate Model (CRCM 4.2.3), driven by the third‐generation Coupled Global Climate Model (CGCM3) for lateral atmospheric and land and ocean surface boundaries. The projected change in air temperature varied from 0.8°C (summer) to 10°C (winter), with a mean warming of 3.9°C. Figure 23 shows the contrast of present day sea‐ice cover to the warmer climate scenario. Under this warmer climate scenario, the sea‐ice season is reduced by 7‐9 weeks. The shallow coastal regions demonstrate a high capacity of heat storage. The maximal volume of sea ice is reduced by 31%. Overall, the depletion of sea‐ice thickness in Hudson Bay follows a southeast‐northwest gradient. Sea‐ice thickness in Hudson Strait and Ungava Bay is predicted to be 50% thinner in mid‐century (2041‐2070) than present climate conditions during wintertime.
Figure 23. ‐‐ Twelve‐hour averaged (a) sea‐ice cover area (in km2) and (b) sea‐ice volume (in km3) for the present climate simulation (blue) and the warmer climate scenario (red) in Hudson Bay; reproduced from Joly et al. (2010).

77
4.2.1.1.1.3.2.9 Baltic Sea
For the Baltic Sea, we reviewed the analysis of Jylha et al. (2008). They used seven regional climate models and found good agreement with observations for the 1902‐2000 comparison period. For the forecast period 2071‐2100, one model predicted a change to mostly mild conditions (49‐58% of average ice area), while the remaining models predicted unprecedentedly mild conditions (19‐37% of average ice area). They noted that their estimates for a warming climate were in agreement with the findings of Tinz (1996) and Meier et al. (2004), who found unprecedentedly mild ice extent conditions in the majority of years after about 2030.
4.2.1.1.1.3.2.10 Sea of Okhotsk
The sea‐ice forecasts for the Sea of Okhotsk are not sufficiently reliable for the ringed seal assessment due to model deficiencies and the small size of the region compared to the spatial resolution of the climate models. Instead, we examined the model forecasts of SAT from the IPCC‐CMIP3 models relative to the current climate conditions. If future monthly mean temperatures during spring approach the melting point of sea ice, ~0°C, sea ice presumably will not persist.
The Sea of Okhotsk lies to the southwest of the Bering Sea and, thus, can be expected to have earlier radiative heating in spring. The region is dominated in winter and spring, however, by cold continental air masses and offshore flow (Wang et al. 2007). During winter and spring, typical air temperatures in the northern Sea of Okhotsk are colder than in the northern Bering Sea (Wang et al. 2007). Sea ice is formed rapidly and is generally advected southward (Sasaki et al. 2007). As the region is dominated by cold air masses for much of the winter and spring, we would expect the present seasonal cycle of first‐year sea ice to continue to dominate the future habitat of the Sea of Okhotsk, similar to the Bering Sea. The Sea of Okhotsk in winter and spring lies between the extreme cold region of Siberia to the west and a storm track that brings warm air northward from Japan to the east. Figure 24 shows the average SATs for the months of March, April, and May. March is dominated by the cold continental air mass with temperatures below ‐6°C for most of the Sea of Okhotsk. During April, there is a large gradient in SATs between the northern and southern portions of the sea. There are ‐4°C temperatures to the north and 0°C to the south. During May, SATs are warmer to the west, and the Sea of Okhotsk region has warmed to the melting point of sea ice throughout the region.
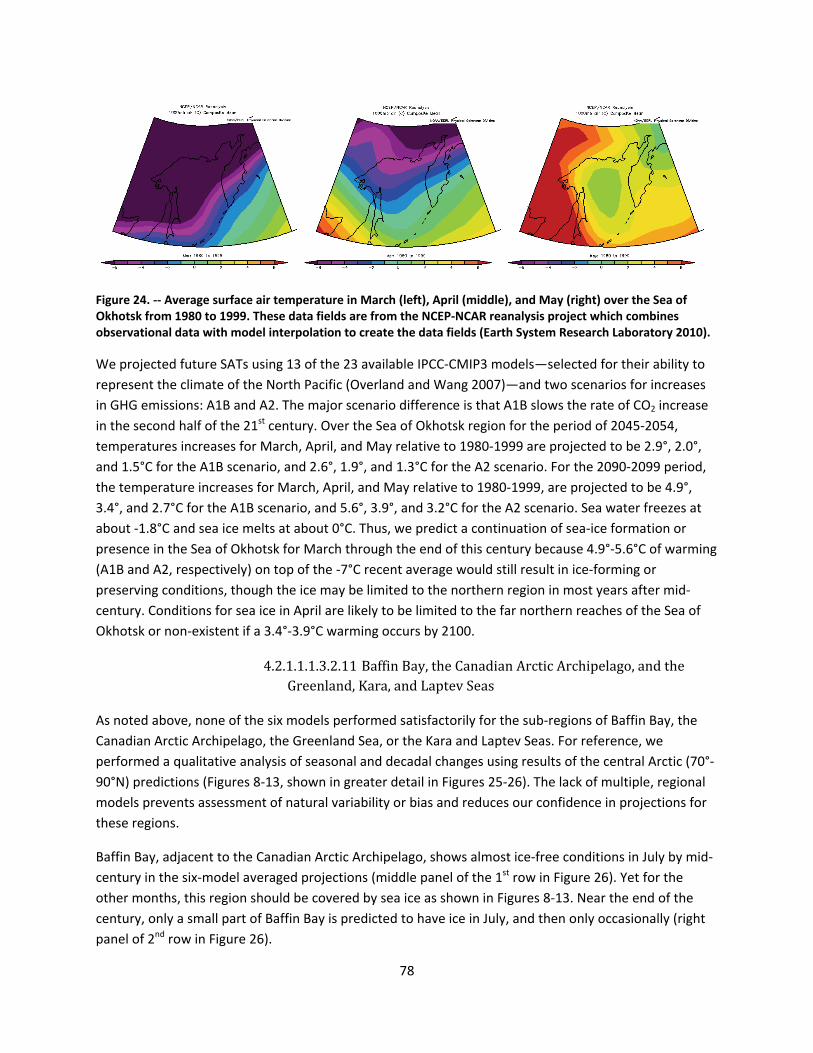
78
Figure 24. ‐‐ Average surface air temperature in March (left), April (middle), and May (right) over the Sea of Okhotsk from 1980 to 1999. These data fields are from the NCEP‐NCAR reanalysis project which combines observational data with model interpolation to create the data fields (Earth System Research Laboratory 2010).
We projected future SATs using 13 of the 23 available IPCC‐CMIP3 models—selected for their ability to represent the climate of the North Pacific (Overland and Wang 2007)—and two scenarios for increases in GHG emissions: A1B and A2. The major scenario difference is that A1B slows the rate of CO2 increase in the second half of the 21st century. Over the Sea of Okhotsk region for the period of 2045‐2054, temperatures increases for March, April, and May relative to 1980‐1999 are projected to be 2.9°, 2.0°, and 1.5°C for the A1B scenario, and 2.6°, 1.9°, and 1.3°C for the A2 scenario. For the 2090‐2099 period, the temperature increases for March, April, and May relative to 1980‐1999, are projected to be 4.9°, 3.4°, and 2.7°C for the A1B scenario, and 5.6°, 3.9°, and 3.2°C for the A2 scenario. Sea water freezes at about ‐1.8°C and sea ice melts at about 0°C. Thus, we predict a continuation of sea‐ice formation or presence in the Sea of Okhotsk for March through the end of this century because 4.9°‐5.6°C of warming (A1B and A2, respectively) on top of the ‐7°C recent average would still result in ice‐forming or preserving conditions, though the ice may be limited to the northern region in most years after mid‐century. Conditions for sea ice in April are likely to be limited to the far northern reaches of the Sea of Okhotsk or non‐existent if a 3.4°‐3.9°C warming occurs by 2100.
4.2.1.1.1.3.2.11 Baffin Bay, the Canadian Arctic Archipelago, and the Greenland, Kara, and Laptev Seas
As noted above, none of the six models performed satisfactorily for the sub‐regions of Baffin Bay, the Canadian Arctic Archipelago, the Greenland Sea, or the Kara and Laptev Seas. For reference, we performed a qualitative analysis of seasonal and decadal changes using results of the central Arctic (70°‐90°N) predictions (Figures 8‐13, shown in greater detail in Figures 25‐26). The lack of multiple, regional models prevents assessment of natural variability or bias and reduces our confidence in projections for these regions.
Baffin Bay, adjacent to the Canadian Arctic Archipelago, shows almost ice‐free conditions in July by mid‐century in the six‐model averaged projections (middle panel of the 1st row in Figure 26). Yet for the other months, this region should be covered by sea ice as shown in Figures 8‐13. Near the end of the century, only a small part of Baffin Bay is predicted to have ice in July, and then only occasionally (right panel of 2nd row in Figure 26).

79
As mentioned earlier, even once the Arctic reaches near ice‐free conditions, sea ice will continue to persist in the Canadian Arctic Archipelago region, the ice “refuge.” According to the models (Figure 12), a reduction in ice concentration appears possible near the end of 21st century in June along the eastern portion of the archipelago, but the channels in the archipelago would still contain sea ice. Even in July, the eastern archipelago may contain ice in concentrations of more than 20% by 2090. These archipelagos are packed with ice in winter, a situation that does not change much throughout the century (Figures 25‐26). In the northern latitudes of the western archipelago, ice concentrations of over 60% are predicted. Around 2050, the archipelago will be packed with ice in the fall and, to a lesser extent, in the early summer as well. Ice grows very fast in the fall, and there will be an average ice concentration of more than 80% over most of the archipelago in November, even by 2090 (Figure 26, last row).
Most of the ice covering the Greenland Sea is near the east coast of Greenland. This spatial pattern is consistent throughout the year (Figures 8‐13) and continues in the future. The one obvious change is in the early summer (July) when, by the end of the 21st century, ice will only cover the northern tip of the Greenland, leaving most of the east coast of Greenland ice free.
Although sea ice has covered most of the Kara and Laptev Seas throughout most of the year in the past, models show that this may change in early summer (July) by the middle of 21st century (Figure 26). The delayed freeze‐up (in November) in the Kara‐Laptev Sea region becomes prominent around 2090 with a majority of the area having ice concentrations less than 40%, which is about half of present day’s value (Figures 8, 12, and 26). The changes in winter and early spring ice conditions, however, are less significant (Figures 8‐10).
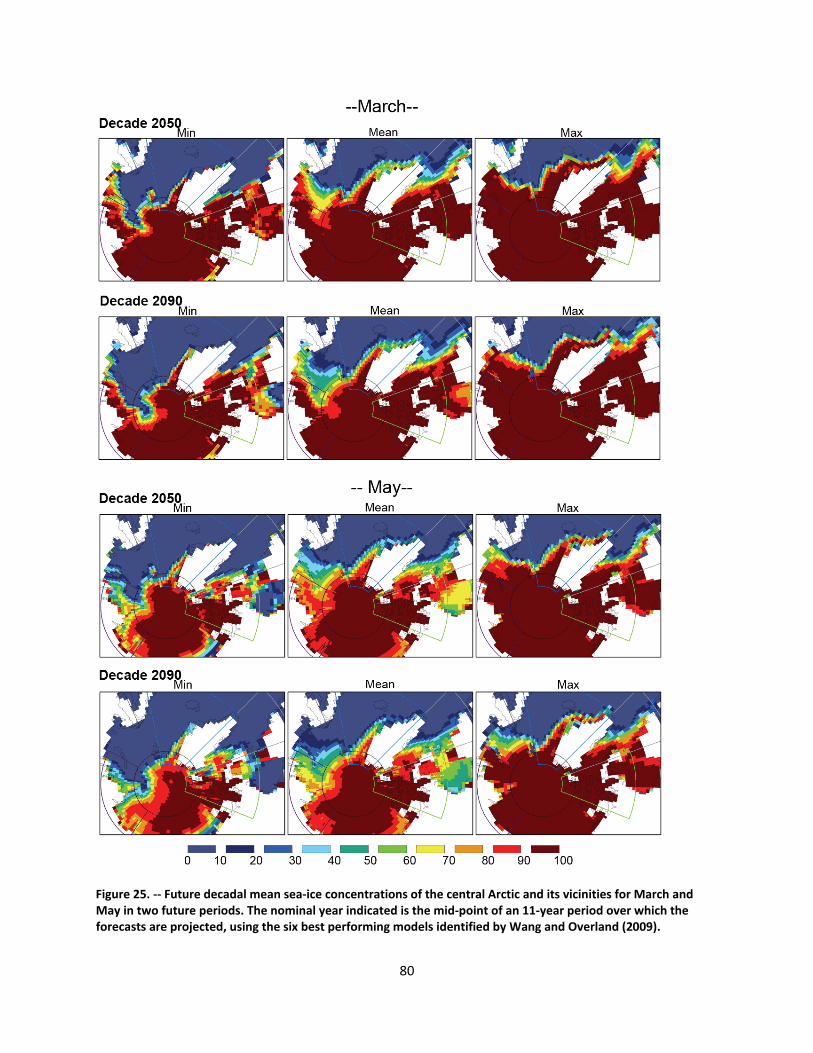
80
Figure 25. ‐‐ Future decadal mean sea‐ice concentrations of the central Arctic and its vicinities for March and May in two future periods. The nominal year indicated is the mid‐point of an 11‐year period over which the forecasts are projected, using the six best performing models identified by Wang and Overland (2009).
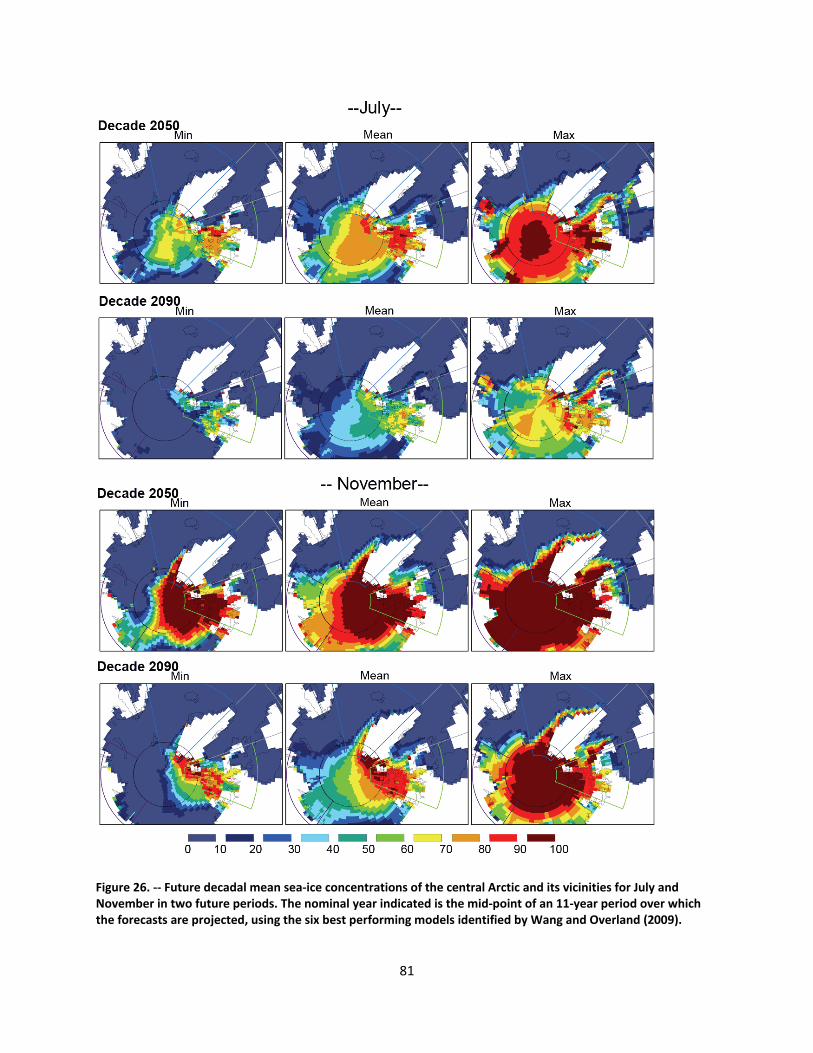
81
Figure 26. ‐‐ Future decadal mean sea‐ice concentrations of the central Arctic and its vicinities for July and November in two future periods. The nominal year indicated is the mid‐point of an 11‐year period over which the forecasts are projected, using the six best performing models identified by Wang and Overland (2009).
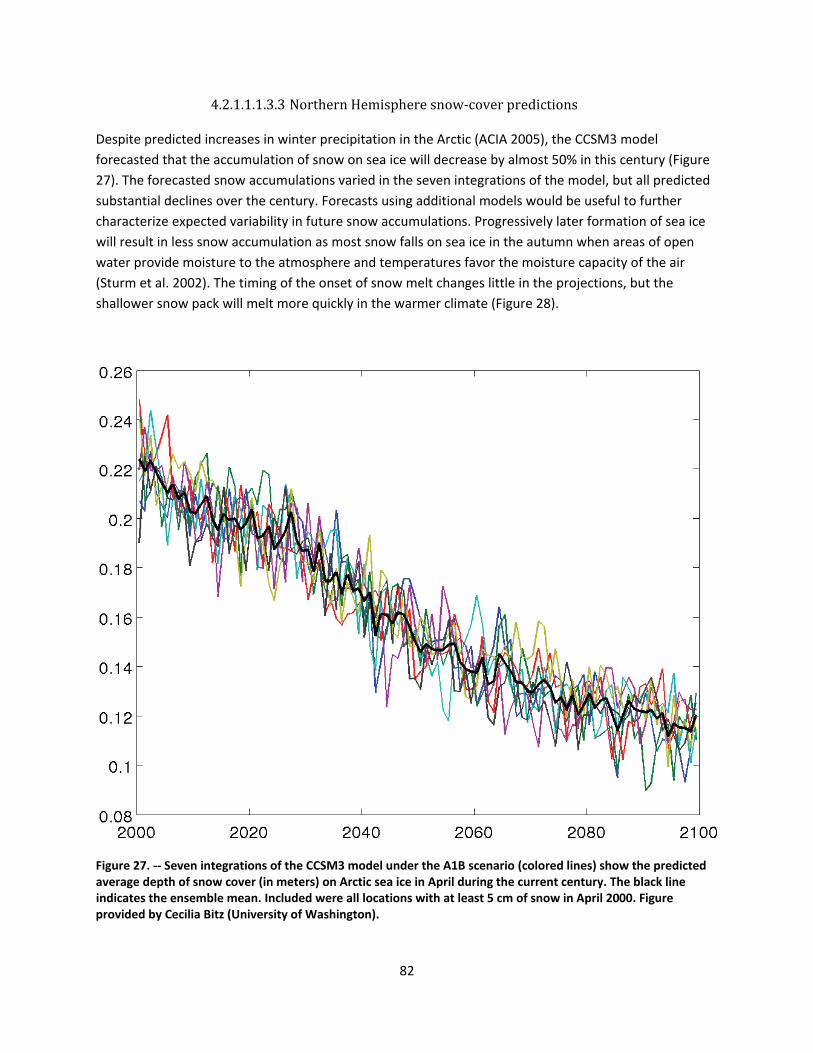
82
4.2.1.1.1.3.3 Northern Hemisphere snow‐cover predictions
Despite predicted increases in winter precipitation in the Arctic (ACIA 2005), the CCSM3 model forecasted that the accumulation of snow on sea ice will decrease by almost 50% in this century (Figure 27). The forecasted snow accumulations varied in the seven integrations of the model, but all predicted substantial declines over the century. Forecasts using additional models would be useful to further characterize expected variability in future snow accumulations. Progressively later formation of sea ice will result in less snow accumulation as most snow falls on sea ice in the autumn when areas of open water provide moisture to the atmosphere and temperatures favor the moisture capacity of the air (Sturm et al. 2002). The timing of the onset of snow melt changes little in the projections, but the shallower snow pack will melt more quickly in the warmer climate (Figure 28).
Figure 27. ‐‐ Seven integrations of the CCSM3 model under the A1B scenario (colored lines) show the predicted average depth of snow cover (in meters) on Arctic sea ice in April during the current century. The black line indicates the ensemble mean. Included were all locations with at least 5 cm of snow in April 2000. Figure provided by Cecilia Bitz (University of Washington).
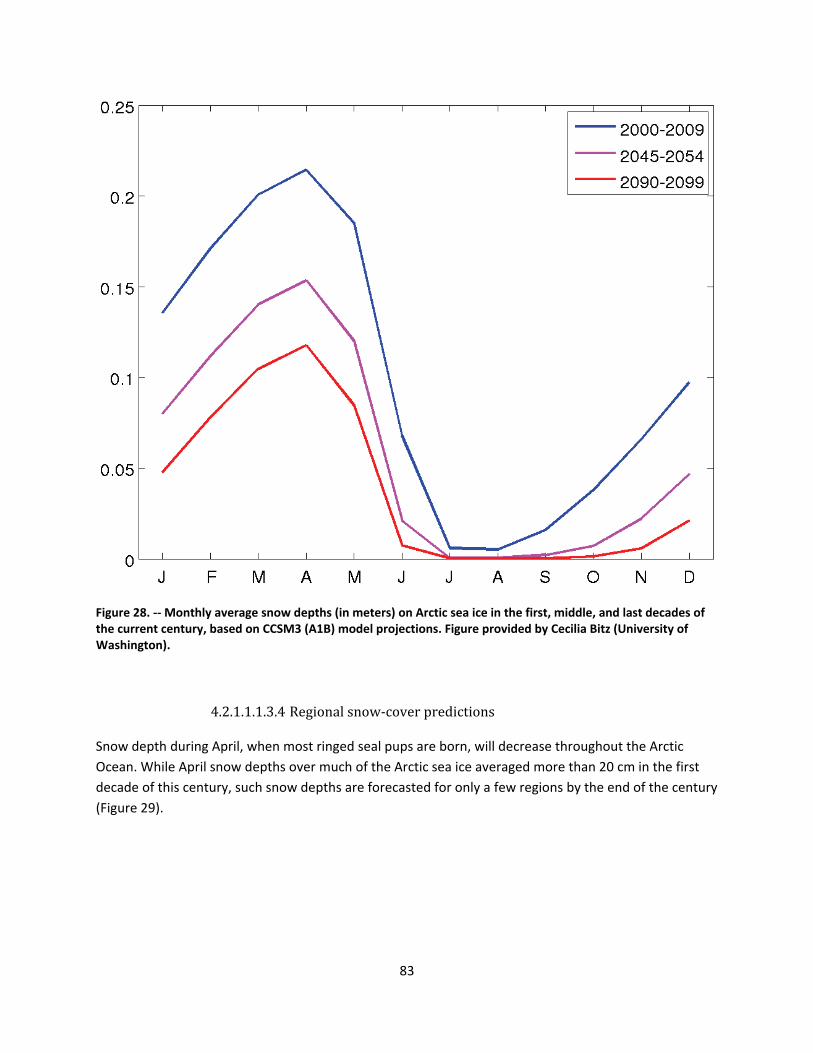
83
Figure 28. ‐‐ Monthly average snow depths (in meters) on Arctic sea ice in the first, middle, and last decades of the current century, based on CCSM3 (A1B) model projections. Figure provided by Cecilia Bitz (University of Washington).
4.2.1.1.1.3.4 Regional snow‐cover predictions
Snow depth during April, when most ringed seal pups are born, will decrease throughout the Arctic Ocean. While April snow depths over much of the Arctic sea ice averaged more than 20 cm in the first decade of this century, such snow depths are forecasted for only a few regions by the end of the century (Figure 29).
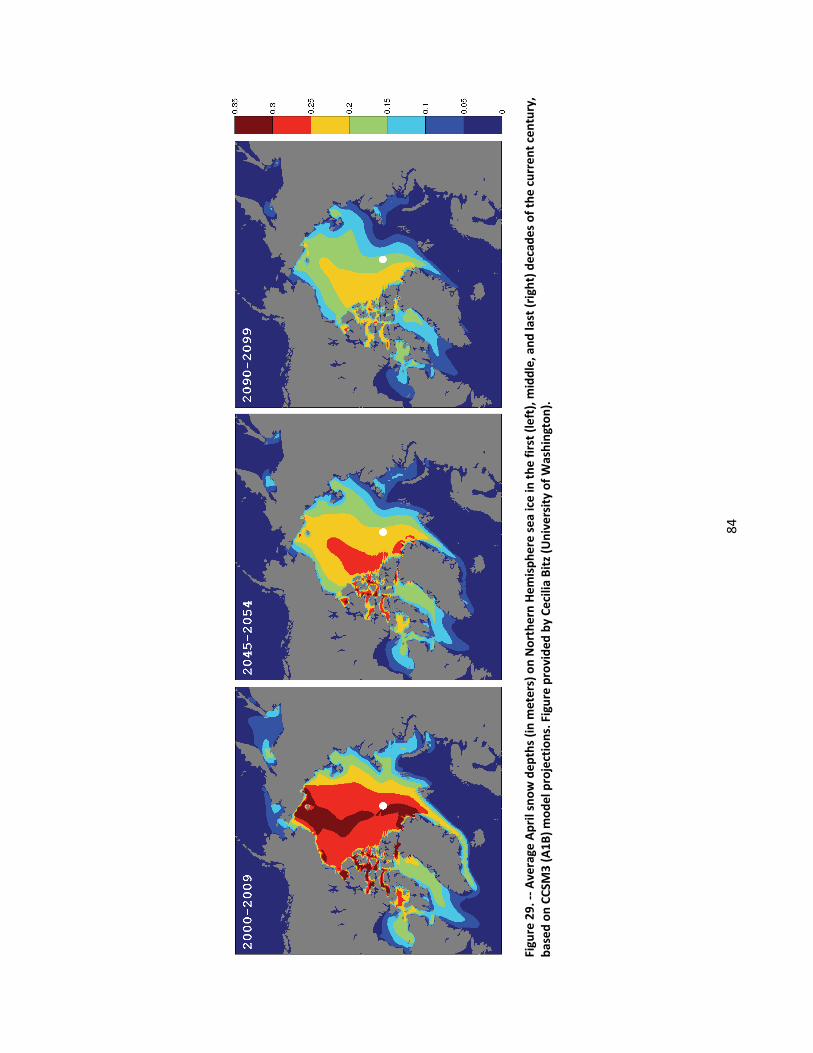
84
Figure 29. ‐‐ Average April snow
dep
ths (in
meters) on Northern Hem
isph
ere sea ice in th
e first (left), m
iddle, and
last (right) d
ecad
es of the
current cen
tury,
based on
CCSM3 (A1B
) mod
el projections. Figure provided
by Ce
cilia
Bitz (University of W
ashington).

85
4.2.1.1.1.3.4.1 Chukchi Sea
In the first 10 years of this century, snow depths over most of the Chukchi Sea exceeded 25 cm in April. By mid‐century, virtually none of the ice in the Chukchi Sea will be covered by that much snow and, by the end of the century, snow depths will typically be less than 20 cm throughout the Chukchi Sea.
4.2.1.1.1.3.4.2 East Siberian Sea
Snow cover in the East Siberian Sea is forecasted to follow a similar course as that in the Chukchi Sea. Snow depths of over 25 cm in most of the sea are forecasted to decline to 15‐20 cm over the century.
4.2.1.1.1.3.4.3 Central Arctic
April snow depths of 25‐35 cm were common on sea ice of the central Arctic Ocean in the first decade of the century and are forecasted to decline heterogeneously thereafter. By mid‐century, the eastern portion of the central Arctic is projected to have 20‐25 cm of snow, while a region of approximately 25‐30 cm snow cover will extend in to the western portion from the coast of North America. By the end of the century, April snow cover will have declined to 20‐25 cm in the western portion and 15‐20 cm in the eastern portion.
4.2.1.1.1.3.4.4 Beaufort Sea
In the Beaufort Sea, average snow depths in April are forecasted to decline from 20‐35 cm in the first decade of the century, to 10‐20 cm by mid‐century, and 5‐15 cm by the end of the century.
4.2.1.1.1.3.4.5 Bering Sea
Average snow depths exceeded 10 cm only in the northern portions of the Bering Sea in April during the first decade of this century. By mid‐century and beyond, April snow depths are forecasted to be less than 5 cm throughout the Bering Sea.
4.2.1.1.1.3.4.6 Barents Sea
April snow depths averaged 10‐15 cm in the vicinity of Svalbard and to the south of Novaya Zemlya in the first decade of the century and are forecasted to decline to 10 cm or less by mid‐century and to less than 5 cm by the end of the century.
4.2.1.1.1.3.4.7 Hudson Bay
Weather station data indicated that average snow depth in Hudson Bay decreased from 45 cm to 18 cm between 1962 and 2000 (Ferguson et al. 2005). Consistent with those measurements, the CCSM3 model forecasted average snow depths of 15‐20 cm only in northern Hudson Bay in the first decade of the century. Snow depths will diminish substantially over the century and, by the end of the century, average snow depths are forecasted to be less than 15 cm throughout Hudson Bay.

86
4.2.1.1.1.3.4.8 Baltic Sea
The global model did not provide adequate resolution for the Baltic Sea. The seven regional climate models analyzed by Jylhä et al. (2008), however, forecasted decreases of 45 to 60 days in duration of snow cover by the end of the century in the northern Baltic Sea region. The shortened seasonal snow cover would result primarily from earlier spring melts but also from delayed onset of snow cover. Depth of snow is forecasted to decrease 50‐70% in the region over the same period. The depth of snow also will be decreased my mid‐winter thaws and rain events. Simulations of the snow cover indicated that an increasing proportion of the snow pack will consist of icy or wet snow (Rasmus et al. 2004).
4.2.1.1.1.3.4.9 Sea of Okhotsk
The CCSM3 projection showed snow depths of 15‐20 cm only in small areas of the northern Sea of Okhotsk in the past ten years and no snow deeper than 15 cm by mid‐century. By the end of the century average snow depths are projected to be 15 cm or less—and mostly less than 10 cm—on the ice that remains in the Sea of Okhotsk.
4.2.1.1.1.3.4.10 Baffin Bay
In the first decade of the century, April snow depths on the ice averaged 15‐20 cm for most of Baffin Bay and 20‐25 cm in a small portion of the northern bay. A modest decline is forecasted by mid‐century. By the end of the century, however, depths greater than 20 cm can be expected only in a small portion of northeastern Baffin Bay.
4.2.1.1.1.3.4.11 Canadian Arctic Archipelago
In the Canadian Arctic Archipelago, April snow depths mostly exceeded 25 cm from 2000‐2009. By mid‐century, such depths are forecasted for a smaller portion of the archipelago, and by the end of the century, depths are expected to be 20‐25 cm over most of the region with pockets of 15‐20 cm and 25‐30 cm.
4.2.1.1.1.3.4.12 Greenland Sea
During the first decade of the century, April snow depths on the ice of the Greenland Sea were largely between 20 and 30 cm, exceeding 30 cm in the northwestern‐most areas. The depths are forecasted to decrease throughout the century. By the end of the century, depths will be less than 20 cm over most of the Greenland Sea and 20 to 25 cm only in small areas off of northeastern Greenland.
4.2.1.1.1.3.4.13 Kara and Laptev Seas
Snow depths along the coasts of the Kara and Laptev Seas in April averaged less than 20 cm in the first decade of the century and increased moving poleward to 20‐25 cm in the northernmost areas. April snow depths will decrease throughout the century with depths averaging less than 10 cm in the Kara Sea and less than 15 cm in the Laptev Sea by the end of the century.

87
4.2.1.1.1.3.4.14 Lake Ladoga and Lake Saimaa
Snow cover in Fennoscandia is projected to decrease 10‐30% before mid‐century and 50‐90% by the end of the century (Saelthun et al. 1998 cited in Kuusisto 2005). High rates of mortality of Saimaa seal pups in birth lairs during 2005‐2007 (more than double that in 1980‐2000) were attributed to insufficient snow for lair formation and occupation (Sipilä and Kokkonen 2008 cited in ICES 2008).
4.2.1.1.2 Effects of climate change on the quality of ringed seals’ habitat
The petition to list ringed seals as threatened or endangered described threats to the species from reduction of sea ice, changes in seasonal snow cover, and ocean acidification (Center for Biological Diversity 2008).
Ice‐associated seals, including the ringed seal, are vulnerable to habitat loss from changes in the extent or concentration of Arctic ice, because they depend on sea‐ice habitat for pupping, foraging, molting, and resting (Tynan and DeMaster 1997, Derocher et al. 2004). The southern edge of the ringed seal’s range may shift north, because ringed seals stay with the ice as it annually advances and retreats (Tynan and DeMaster 1997). Whether ringed seals will continue to move north with retreating ice over the deeper, less productive Arctic Basin waters and whether forage fishes that they prey on will also move north is uncertain.
Sea‐ice extent in September 2007 in the central Arctic Ocean was a record low, nearly 40% below the long‐term average and 23% below the previous record set in 2005 (Stroeve et al. 2008). Most of this loss was on the Pacific side of the Arctic. Arctic Ocean ice extent remained low in the summers of 2008, 2009, and 2010 (National Snow and Ice Data Center 2008, Wang and Overland 2009). Sea‐ice projections at the end of summer for the years 2045‐2054 from the IPCC AR4, combined with the recent result that Arctic sea ice is on a faster track for loss compared to these projections, provided support for the recent listing of polar bears as threatened under the ESA (U.S. Fish and Wildlife Service 2008). The range of ringed seals largely overlaps with that of polar bears, and ringed seals depend on ice throughout their range for resting, whelping, nursing, and molting. Ice cover strongly influences community structure in ice‐covered ecosystems by limiting access for some species and providing habitat for specialist species. Changes in the phenology and extent of ice extent will alter community composition, presenting ringed seals with new competitors, predators, and prey (Grebmeier et al. 2006b).
Sea ice changes in response to environmental factors may have consequences for the distribution and productivity of ringed seals. In the absence of solid data, it has generally been assumed that seal populations occur at high numbers and are stable. Unexpectedly low pregnancy rates and proportions of young‐of‐the‐year, however, were reported in a sample from western Hudson Bay in 1991‐1992 (Holst et al. 1999, Ferguson et al. 2005). A follow‐up study spanning 1998‐2000 also found a lower than expected pregnancy rate and proportion of young‐of‐the‐year. Thus, ringed seal recruitment appears to be declining in Hudson Bay and can be expected to lead to a population decline (Stirling 2002).
In the southern Beaufort Sea, anomalous heavy sea‐ice conditions in the mid‐1970s and mid‐1980s caused significant declines in productivity of ringed seals (Stirling 2002). Each event lasted

88
approximately 3 years and caused similar declines in the birth rate of polar bears and survival of subadults, after which reproductive success and survival of both species increased again.
Ringed seals create birth lairs in areas of accumulated snow on stable ice including the shorefast ice over continental shelves along Arctic coasts, bays, and inter‐island channels (Smith and Hammill 1981). While some authors suggest that landfast ice is the preferred pupping habitat of ringed seals due to its stability throughout the pupping and nursing period (McLaren 1958a, Burns 1970), others have documented ringed seal pupping on drifting pack ice both nearshore and offshore (Burns 1970, Finley et al. 1983, Smith 1987, Wiig et al. 1999, Lydersen et al. 2004). Either of these habitats can be affected by earlier warming and break‐up in the spring, which shortens the length of time pups have to grow and mature (Kelly 2001, Smith and Harwood 2001). Harwood et al. (2000) reported that an early spring break‐up negatively impacted the growth, condition, and apparent survival of nursing ringed seal pups. Early break‐up was believed to have interrupted lactation in adult females, which in turn, negatively affected the condition and growth of pups. Earlier ice break‐ups similar to those documented by Harwood et al. (2000) and Ferguson et al. (2005) are predicted to occur more frequently with warming temperatures and result in a predicted decrease in productivity and numbers of ringed seals (Kelly 2001, Ferguson et al. 2005). Additionally, high fidelity to birthing sites exhibited by ringed seals makes them more susceptible to localized degradation of snow cover (Kelly et al. 2010).
Unusually heavy ice has also resulted in markedly lower productivity of ringed seals and reduced polar bear productivity (Stirling 2002). While reduced ice thickness associated with warming in some areas could be expected to improve seal productivity, the transitory and localized benefits of reduced ice thickness on ringed seals are expected to be outweighed by the negative effects of increased vulnerability of seals to predation and thermoregulatory costs (Hammill and Smith 1989, Smith et al. 1991, Harwood et al. 2000, Derocher et al. 2004, Stirling and Smith 2004, Ferguson et al. 2005). For example, high pup mortality in the White Sea in 1977 was attributed to warm conditions resulting in early snow melts that destroyed subnivean lairs and early break‐up of unusually thin ice and the subsequent premature separation of nursing pups from their mothers (Lukin et al. 2006).
Ringed seals—especially newborns—depend also on snow cover for protection from cold temperatures and predators. Occupation of subnivean lairs is especially critical when pups are nursed in late March to June. Ferguson et al. (2005) showed that ringed seal recruitment in western Hudson Bay dropped sharply to near zero when the average snow depth in April and May was less than 32 cm. Reduced snowfall results in less snow accumulation next to pressure ridges, and pups in lairs with thin snow cover are more vulnerable to predation than pups in lairs with thick cover (Hammill and Smith 1989, Ferguson et al. 2005). Warming temperatures that melt snow‐covered birth lairs can result in pups being exposed to ambient conditions and suffering from hypothermia (Stirling and Smith 2004). Others have noted that when lack of snow cover has forced birthing to occur in the open, nearly 100% of pups died from predation (Kumlien 1879, Lydersen et al. 1987, Lydersen and Smith 1989, Smith et al. 1991, Smith and Lydersen 1991). Polar bears and Arctic foxes—important predators of Arctic ringed seals—are absent from the Sea of Okhotsk, however, and most Okhotsk ringed seals apparently give birth on pack ice in the lee of ice hummocks (Krylov et al. 1964, Heptner et al. 1976). Use of subnivean lairs in the Sea of

89
Okhotsk may be restricted to the limited areas of shorefast ice (Krylov et al. 1964), although there are some reports of lairs in the pack ice as well (Fedoseev 1965b). More recently, telemetric monitoring of Arctic ringed seals and satellite measurements of snow melt—using passive microwave emissions—showed that the seals’ emergence from lairs was related to structural failure of the snow pack (Kelly et al. 2006). Warmer temperatures will continue to have negative effects on ringed seal pup survival.
Increased rain‐on‐snow events during the late winter also will negatively impact ringed seal recruitment by damaging or eliminating snow‐covered pupping lairs, increasing exposure and the risk of hypothermia, and facilitating predation by polar bears, Arctic foxes, gulls, ravens, and other predators (Stirling and Smith 2004). Stirling and Smith (2004) documented the collapse of subnivean lairs during unseasonal rains near southeastern Baffin Island and the subsequent exposure of ringed seals to hypothermia. They surmised that most of the pups that survived exposure to cold were eventually killed by polar bears (Stirling and Archibald 1977), Arctic foxes (Smith 1976), or gulls (Lydersen and Smith 1989). Stirling and Smith (2004) postulated that should early season rain become regular and widespread in the future, mortality of ringed seal pups will increase, especially in more southerly parts of their range.
4.2.1.1.3 Effects of climate change on ocean conditions
4.2.1.1.3.1 Ocean warming
The western Beaufort Sea, the Chukchi Sea, and northern Bering Sea surface waters have warmed by as much as 3.5°C since 1990 (Steele et al. 2008). Summer temperatures of the southeastern Bering Sea warmed 2°C from 1995‐2003 (Overland and Stabeno 2004). Ocean temperatures warmed in summer 2007 and 2008 by as much as 4°C in the East Siberian Sea, 3°C in the Chukchi Sea, 2°C in the Kara Sea, and 1.5°C in parts of the Laptev Sea (Richter‐Menge and Overland 2009). In contrast, the Barents, Greenland, and parts of the Laptev Seas experienced occasional years of mild (≤ 1.5°C) cooling over the same period. Warming of sea surface temperatures would likely have little direct impact on ringed seals, although the efficiency of molting and thermoregulation might be somewhat improved (Boily 1995, Harding et al. 2005). Indirect effects of ocean warming, however, could be substantial, particularly through altered distributions of prey, pathogenic vectors, and predators. These effects are considered in Sections 4.2.1.1.3.3.2.1, 4.2.3.1 , and 4.2.3.3 respectively.
4.2.1.1.3.2 Ocean acidification
Since the beginning of the industrial revolution in the mid‐18th century, the release of GHGs including CO2 from human activities, commonly referred to as “anthropogenic CO2”, has resulted in an increase in atmospheric CO2 concentrations from approximately 280 to nearly 390 ppm, with 30% of the increase occurring in the last three decades. The atmospheric concentration of CO2 is now higher than experienced on Earth for more than 800,000 years (Lüthi et al. 2008). Over the industrial era, the ocean has absorbed about one‐third of anthropogenic carbon emissions (Canadell et al. 2007). When anthropogenic CO2 is absorbed by seawater, chemical reactions occur that reduce both seawater pH and the concentration of carbonate ions in a process known as “ocean acidification.”

90
Results from global ocean CO2 surveys over the past two decades have shown that ocean acidification is a predictable consequence of rising atmospheric CO2 levels. Seawater carbonate chemistry is governed by a series of abiotic chemical reactions (CO2 dissolution, acid/base chemistry, and calcium carbonate dissolution) and biologically mediated reactions (photosynthesis, respiration, and calcium carbonate precipitation). The pH of ocean surface waters has already decreased (i.e., become less alkaline) by about 0.1 units since the beginning of the industrial revolution (Caldeira and Wickett 2003, Caldeira and Wickett 2005), with a decrease of ~0.0018 yr
‐1 observed over the last quarter century at several open
ocean sites (Bates 2007, Bates and Peters 2007, Santana‐Casiano et al. 2007). By the middle of this century, atmospheric CO2 levels could reach more than 500 ppm and over 800 ppm by the end of the century (Orr et al. 2005). The result would be an additional decrease in surface water pH of approximately 0.3 pH units by 2100. Acidity in the ocean would increase by about 150% relative to the beginning of the industrial era.
As the pH of the ocean decreases, the equilibrium between calcium carbonate (CaCO3) and its dissolution products (Ca2+ and CO3
2‐) favors dissolution. Ocean acidification reduces the calcium carbonate saturation levels, which stresses calcifying organisms by making calcification more difficult. Significant reductions in calcium carbonate saturation have been observed in the ocean since the industrial revolution (Feely et al. 2004). The carbonate saturation horizon is shoaling (becoming shallower), shrinking the layer of carbonate‐saturated surface waters in which calcification by organisms can occur. The observed and expected future shoaling of the saturation depth in the North Pacific are greater than in most of the other oceans due to respiration processes as ocean water circulates along the deep conveyor belt from the Atlantic to the Indian and Pacific Oceans (Feely et al. 2004). Shoaling of undersaturated waters has been observed in the high latitude North Pacific (Feely et al. 1988) and the Chukchi Sea (Bates and Mathis 2009) where the seasonal undersaturation of carbonate minerals in subsurface waters is enhanced by the sinking organic matter. Local upwelling can exacerbate the ocean acidification impacts by bringing deep, CO2 rich waters to the surface exposing organisms to strongly corrosive conditions (Feely et al. 2008). Two recent studies suggested that aragonite undersaturation due to ocean acidification will develop in Arctic surface waters by or before mid‐century under the B1 and A2 IPCC emissions scenarios (Orr et al. 2009, Steinacher et al. 2009).
The process of ocean acidification has long been recognized (Broecker and Takahashi 1966, Broecker et al. 1971, Bacastow and Keeling 1973, Feely and Chen 1982, Feely et al. 1988), but the ecological implications of such chemical changes have only recently begun to be appreciated. Although initial concerns about the effects of acidification focused on the negative effect that decreased calcium carbonate saturation state has on the ability of organisms to produce calcium carbonate shells, it is becoming increasingly clear that changes in CO2 and pH per se can affect the growth, survival, and behavior for a range of marine organisms (Pörtner 2008). Many non‐calcareous species are affected by acidification, and the ability of calcareous species to produce shells can be affected by acidification factors other than just saturation state (Pörtner 2008). In addition to the effects of changes in CO2, pH, and saturation state, secondary chemical reactions can change other components of seawater, such as the concentration of various forms of trace elements and nutrients, which in turn can affect species growth and survival (Doney et al. 2009). Ocean acidification also affects sound absorption in the ocean.

91
Brewer and Hester (2009), for example, suggest that changes in borate and carbonate ion concentrations associated with ocean acidification is reducing the absorption of low frequency sound (~300 Hz‐10 kHz) important to some marine mammals. Potential direct effects of ocean acidification include:
• Reduced calcification rates – The reduced saturation state affects the ability to produce calcium carbonate shells, makes the process more physiologically costly, or leads to dissolution of existing calcium carbonate structures.
• Altered survival and reproduction from reduced pH – Organisms generally require energy to maintain appropriate inter‐cellular pH balance. Altering the external pH of seawater can overwhelm pH control mechanisms, affecting survival or reproduction.
• Reduced olfaction in fish – Increased CO2 in seawater can affect the ability of fishes to detect critical olfactory cues.
• Increased photosynthesis – Because CO2 is required for photosynthesis, some photosynthetic organisms, especially those without effective carbon concentrating mechanisms, may have increased photosynthetic rates with increased CO2.
• Hypercapnia – Increased CO2 in internal fluids, especially in highly energetic species like squid, can affect survival or reproduction. The ability of organisms to decrease CO2 titers in internal fluids by transferring CO2 across membranes to seawater is reduced when seawater CO2 concentrations are high.
• Acoustic disruption from noisier ocean – Changes in ocean pH will alter the acoustic properties of the ocean, increasing transmission of low frequency sounds, which may affect species relying on acoustic information.
• Response to altered metals, nutrient or toxics speciation – Acidification will alter speciation (ionic from) of various metals, nutrients, and toxins in a way which might affect species survival and reproduction.
The waters of the Arctic and adjacent seas are among the most vulnerable to ocean acidification (Orr et al. 2005, Gehlen et al. 2007, Cao and Caldeira 2008). Modeling studies have suggested that the polar seas will experience aragonite undersaturation by the middle of the century. Recent modeling studies (Feely et al. 2009, Steinacher et al. 2009) argue that the surface waters of the Arctic Ocean will start experiencing localized aragonite undersaturation within the next decade.
Seasonal undersaturation has already been documented (Bates et al. 2009, Yamamoto‐Kawai et al. 2009). Bates et al. (2009) measured saturation states for aragonite and calcite for the Chukchi Sea shelf and Canada Basin from 2002 to 2004 and found that aragonite undersaturation is occurring in the subsurface waters due to ocean acidification combined with subsurface remineralization of particulate organic carbon. Patches of surface water undersaturated in aragonite were also found in the Canada Basin, which resulted from significant sea‐ice melt contributions. In a separate study, Yamamoto‐Kawai et al. (2009) also found that the synergistic effects of increasing CO2 and ice melt resulted in surface waters in the Canada Basin that were undersaturated with respect to aragonite.

92
Even though the Atlantic Ocean generally has higher saturation states than the Pacific, carbonate chemistry measurements in the Baltic Sea show that the central basin becomes undersaturated (or nearly so) in winter, with respect to both aragonite and calcite mineral forms of calcium carbonate; undersaturation is even more severe in Bothnian Bay (Tyrrell et al. 2008). Thus, the Baltic Sea is equally as vulnerable to increasing CO2 levels as the Arctic. European researchers are actively studying ocean acidification in the Baltic.
We are not aware of specific acidification studies in the Lakes Ladoga and Saimaa. Freshwater systems, however, are much less buffered than ocean waters and are likely to experience even larger changes in acidification levels than marine systems. Aquatic ecosystem acidification can have serious deleterious impacts on aquatic organisms, as shown by an increasing number of laboratory studies, as well as historical studies on poorly buffered, soft water lakes that experienced acidification and ecosystem impacts during the 20th century due to the deposition of acidic nitrogen (N) and sulfur (S) compounds (e.g., Water, Air, and Soil Pollution (vols. 31[3‐4]: 1986 and 35 [1‐2]: 1987), Ambio (vol. 22[5]: 1993), and Journal of Paleolimnology (vol. 3[3]: 1990).
Although no scientific studies have directly addressed the impacts of ocean acidification on ringed seals, the effects would likely be through their ability to find food. Most pinniped species are high trophic predators that live in regions with high productivity at least seasonally (e.g., Bowen and Siniff 1999). Ringed seals consume most of their annual energy in a period from late summer through to early winter (Ryg and Øritsland 1991), focusing on lipid rich, large zooplankton, Arctic cod, and polar cod. Climate warming, however, has been credited with global declines in phytoplankton concentrations (Boyce et al. 2010) and shifts in community organization and productivity in the Bering Sea, Aleutian Islands, and Gulf of Alaska (Anderson and Piatt 1999, Ciannelli et al. 2005, Grebmeier et al. 2006b, Litzow et al. 2006, Litzow and Ciannelli 2007, Mueter and Litzow 2008). Ocean acidification is likely to have increasingly profound impacts on the ecosystem structure in the ringed seal habitats. The exact nature of these impacts cannot be predicted, and some likely will amplify more than others. For example, populations of upper trophic level pelagic species’ may decline if their early life stages consume prey items (e.g., pterapods; Comeau et al. 2009) that cannot survive the added stress of ocean acidification. Pteropods are important food sources for larval and juvenile walleye pollock (Theragra chalcogramma), Pacific herring (Clupea pallasii), and cod. The ringed seals depend on cod, particularly juvenile cod that are less than 20 cm in length (Lowry et al. 1980). The loss of calcifying species like pteropods from the ecosystem could have a cascading effect on the ringed seals.
There are a number of potential direct impacts of highly elevated CO2 on embryo and larval fish that could affect the availability of cod and other fish as a food source in the future (Ishimatsu et al. 2005). Likewise, Yamada and Ikeda (1999) found that exposure to very high CO2 concentrations (> 2,000 ppm) does reduce survival of six different copepod species, but levels expected over the next century do not seem to have a significant impact (Kurihara and Ishimatsu 2008).

93
4.2.1.1.3.3 Ringed seal prey communities
4.2.1.1.3.3.1 Current status and trends of prey
The breadth of the ringed seal’s diet reflects its diverse range of habitats including the marine Arctic Ocean and adjacent seas, the brackish Baltic Sea, and freshwater lakes. Ringed seals are shallow‐water pelagic predators, and most of the primary prey taxa are small schooling species that form dense aggregations in the water column (Kovacs 2007). Fish are generally more important than invertebrates, and gadid species are most prevalent in the diets of ringed seals, but preferred prey items vary by region and by season. Key demersal fish species include Arctic cod, polar cod, saffron cod, navaga, rockfish, sculpins, pricklebacks, and snailfish. Common pelagic fish prey include capelin, smelt, and herring. Pelagic invertebrate prey include amphipods (Gammaridea and Hyperiidea), mysids (Mysidae), euphausiids (Thysanoessa spp.), isopods (Isopoda) and squid (Gonatidae). Shrimp are also preyed upon by ringed seals. Ringed seals in the freshwater Lake Ladoga and Lake Saimaa consume a variety of pelagic and benthic fish including smelt, ruffe, perch, roach, and vendace.
4.2.1.1.3.3.1.1 The Arctic
Ringed seals in the Arctic feed primarily on fish. Most prominent among these are gadids, including Arctic cod, polar cod, saffron cod, and navaga. The Arctic cod is distributed throughout the Arctic, and extends south into the Labrador Sea in the North Atlantic Basin as well as Hudson Bay and the northern Bering Sea (Cohen et al. 1990). Thus, it is available throughout the range of Arctic ringed seals. It is primarily a coastal species, and is usually found either in nearshore waters or offshore associated with sea ice. Arctic cod may be the most abundant fish species in some Arctic areas (Barber et al. 1997, Mecklenberg et al. 2002) and have been fished heavily in the Barents Sea and northwest Atlantic. Its short life span and high fecundity, however, have protected the species from notable depletion (Cohen et al. 1990). The polar cod has a disjunct but broad distribution in the Arctic and has been reported from both sides of Greenland as well as the Canadian High Arctic and the Laptev Sea (Cohen et al. 1990). It is a pelagic species primarily found in offshore waters and associated with sea ice, and its biology is poorly known. Two species of Eleginus, saffron cod and navaga, are prominent in ringed seal diets. The saffron cod is found throughout the North Pacific and Bering Sea and extends into the Arctic in the East Siberian, Chukchi, and Beaufort Seas (Cohen et al. 1990). Navaga are found in the Barents, Kara, and White Seas. Both are primarily demersal species found in shallow coastal waters.
In addition to gadids, several species of pelagic forage fishes, such as smelts and herring, are regularly consumed by ringed seals. The smelt species most commonly found in ringed seal diets is capelin, which is found throughout the Arctic, extending south into the North Atlantic and North Pacific. This pelagic species migrates onshore in large schools to spawn and, therefore, large numbers are accessible to ringed seals in shallow water during some times of the year (Eschmeyer and Herald 1983, Mecklenberg et al. 2002). Other species of smelts consumed by ringed seals (e.g., Osmerus spp.) occupy large portions of the Arctic and are anadromous species generally found in shallow coastal waters. Similarly, Atlantic herring (Clupea harengus) and Pacific herring are abundant, pelagic, schooling fishes found primarily in coastal waters. The Atlantic herring is primarily restricted to the subarctic portions of the North Atlantic,

94
Labrador, and North Seas, but extends into the Arctic in the Barents and Greenland Seas. The Pacific herring is found throughout the North Pacific and Bering Sea, but also extends into the Chukchi and Beaufort Seas, and disjunct populations in the Kara and Barents Seas are sympatric with Atlantic herring (Whitehead 1985). Both Atlantic and Pacific stocks have been depleted by decades of fishing efforts, but those efforts have been concentrated in the subarctic portions of the species range. Arctic populations of herring are fished less than their subarctic counterparts, and are poorly known (Hay et al. 2001).
Several species of bathypelagic and demersal fishes are also found regularly in ringed seal diets, including Atlantic redfish (Sebastes spp.) as well as sculpins, pricklebacks, snailfishes, and eelpouts. The Atlantic redfish complex includes two or more species of Sebastes (S. marinus, S. mentella, S. fasciatus) that extend from the North Atlantic into the Labrador, Greenland, and Barents Seas. These are bathypelagic species generally found in deep waters, but they migrate vertically up into the water column at night (Gauthier and Rose 2005). Other demersal species found in ringed seal diets generally reflect the most abundant and diverse groups in the region. Sculpins (Cottidae) are represented throughout the Arctic by several genera, including Artediellus, Icelus, Gymnocanthus, Myoxocephalus, and Triglops (Walters 1955, Mecklenberg et al. 2002, Mecklenburg et al. 2007, Møller et al. 2010). Eelpouts are particularly diverse in Arctic waters, with 26 species known from Greenland alone (Møller et al. 2010).
A suite of environmental variables from the North Pacific changed abruptly in 1976‐1977 (Ebbesmeyer et al. 1991, Trenberth and Hurrell 1995, Hare and Mantua 2000) in what has now become known as a major ocean climate “regime shift.” The regime shift was associated with warmer ocean temperatures and decreased winter ice cover in the Bering Sea (Niebauer 1988, Hollowed and Wooster 1995). This reduction in the extent of winter ice reduced the southerly extent of very cold (< 2°C) bottom water on the Bering Sea shelf during summer (a.k.a. the “cold pool”) with subsequent expansion of walleye pollock to the south and contraction of Arctic cod to the north (Wyllie‐Echeverria and Wooster 1998). Further research has shown that the northward shift of the summer cold pool has apparently resulted in the northward shift in the center of distribution of multiple taxa of demersal fish and invertebrates (Mueter and Litzow 2008). Subarctic taxa increasingly occupy the area formerly covered by the cold pool resulting in a shift from a less species diversity Arctic community devoid of groundfish to a more diverse subarctic community rich in groundfish. Continued reductions in winter sea‐ice cover appear to be causing a change from Arctic to subarctic conditions in the northern Bering Sea and a northward shift of the pelagic‐dominated marine ecosystem that was previously limited to the southeastern Bering Sea (Grebmeier et al. 2006b). The regime shift had mixed impacts on production of groundfish. Recruits per spawner decreased after the regime shift for flatfish (e.g., yellowfin sole [Limanda aspera]) and increased for rockfish (e.g., Pacific ocean perch [Sebastes alutus]). Although production of groundfish (e.g., walleye pollock and Pacific cod [Gadus macrocephalus]) was not impacted by the regime shift (Low 2008), there was a shift in the importance of pollock to the ecosystem, from near 10% of the energy flow in the 1950s to 1960s to over 50% in the 1980s (PICES 2005). Biomass trends in the southeastern Bering Sea do show trends coherent with the regime shift. The density of demersal fish and non‐crab benthic invertebrates increased from the late 1970s to the early 1990s and then decreased somewhat in the late 1990s (Conners et al. 2002).

95
The warm period occurring after the 1976‐1977 regime shift was followed by a weak cold period from 1989‐1998 with a subtle but evident increase in winter ice extent in the Bering Sea (Schumacher et al. 2003). The Bering Sea then experienced another suite of unusual physical and biological conditions in the late 1990s associated with unusually warm sea surface temperatures: the first recorded major coccolithophore blooms, a large die‐off of shearwaters, salmon returns far below predictions and reduced onshore transport of slope water (Schumacher et al. 2003). Production (recruits‐per‐spawner) decreased for several species of flatfish after 1988‐89 (e.g., arrowtooth flounder [Atheresthes stomias], rock sole [Lepidopsetta bilineata], and flathead sole [Hippoglossoides elassodon]) and increased for rockfish (e.g., Pacific ocean perch) (Low 2008). Cold water species such as Greenland turbot (Reinhardtius hippoglossoides), Arctic cod, and snow crab (Chionoecetes opilio) are no longer found in abundance in the southeast Bering Sea (PICES 2005). Adult walleye pollock biomass in the eastern Bering Sea has ranged from 3.2 to 12.6 million metric tons since 1979 and is currently low due to the lack of strong recruitment in recent years (Ianelli et al. 2009). The numbers of smaller, juvenile pollock in the eastern Bering Sea is highly variable due to high interannual variation in year‐class strength (Ianelli 2005).
Capelin stocks in the Barents Sea have fluctuated dramatically since the early 1970s. Stock size was high in the 1970s and early 1980s. The stock collapsed around 1984, and the commercial fishery was closed from 1987‐1990. The stock apparently recovered for a brief period after 1988 and then stock collapsed again, and the fishery was closed in 1994 (Gjøsæter 1998). The declines in capelin stock size are thought to be related to decreases in plankton biomass, increases in predation by herring and cod, and increases in fishing pressure (Skjoldal et al. 1992, Gjøsæter 1998).
In addition to fishes, ringed seals in the Arctic feed on a variety of invertebrates, including several species of crustaceans, euphausiids, amphipods, mysids, and mollusks. Crustaceans favored by ringed seals include virtually all species of shrimps known from the Arctic as well as majid and pagurid crabs. Some of the shrimp, such as Pandalus borealis, are abundant enough to be commercially fished (Hop et al. 2002). At least four species of the epipelagic euphausiid genus Thysanoessa are found in Arctic waters: T. inermis, T. longicaudata, T. longipes, and T. raschii (Brinton 1962). T. longipes is found only in the North Pacific and adjacent Arctic waters, T. longicaudata is found only in the North Atlantic and adjacent Arctic waters, and T. inermis and T. raschii are found on both sides of the Arctic. Whether their populations are continuous across the Arctic is unknown (Brinton 1962). Arctic amphipods consumed by ringed seals include pelagic species, such as Themisto libellula, as well as sympagic species inhabiting the underside of the sea ice, such as Gammarus wilkitzkii, which is the dominant amphipod of the Arctic sympagic community (Gulliksen and Lonne 1989, Hop et al. 2002).
The Bering Sea regime shift of 1976/77 and subsequent ocean warming resulted in a decrease in zooplankton crustaceans and an increase in gelatinous zooplankton (jellyfish) whose diet of juvenile pollock, euphausiids, and amphipods potentially overlaps with that of ringed seals (Schumacher et al. 2003, Bluhm and Gradinger 2008). In addition, rebounding populations of planktivorous whales may have put further pressure on zooplankton food resources used by fish and ringed seals (Schumacher et al. 2003). Similarly, zooplankton biomass and chlorophyll concentrations in the central and western

96
subarctic Pacific decreased after the early 1970s (Sugimoto and Tadakoro 1997). Interannual variability in the Barents Sea zooplankton community is thought to be related to the amount of inflow of North Atlantic water (Skjoldal and Rey 1989). For example, the population size of Arctic amphipod species, such as Themisto libellula, has been shown to be correlated with the amount of Arctic water in the Barents Sea, such that the population was low in years when Atlantic inflow was high (Dalpadado 2002). During the last decades (since the late 1980s) the influx of Atlantic water into the Arctic Ocean has increased (Morison et al. 2000). In the Canada Basin there have been changes in the ice‐associated phytoplankton and zooplankton communities since the 1970s‐1980s that are thought to be a result of ice melt and subsequent warming and freshening of the ocean water. Specifically observed were changes in the species composition of phytoplankton, a decrease in viable populations of copepods and amphipods within the ice interior, and a decrease in the number of species and overall population size of zooplankton associated with the under surface of the ice (Melnikov et al. 2002).
4.2.1.1.3.3.1.2 Sea of Okhotsk
In the Sea of Okhotsk, ringed seals feed primarily on fish and euphausiids (Fedoseev 1965c). Fish prey include saffron cod, Arctic smelt (Osmerus dentex), and Pacific herring, all of which are abundant in the Sea of Okhotsk (Sheiko and Federov 2000). The Arctic cod, an important component of the diet of ringed seals in other areas, is not found in the Sea of Okhotsk, but capelin are abundant in the region (Cohen et al. 1990, Sheiko and Federov 2000). Euphausiids also represent a significant portion of the diet of ringed seals in the Sea of Okhotsk, and at least two species of the epipelagic euphausiid genus Thysanoessa are found there: T. inermis and T. raschii (Brinton 1962).
Pacific herring stocks in the Sea of Okhotsk declined severely in the latter part of the 20th century due to intense fishing pressure, and commercial herring fishing in the region has been prohibited (Hay et al. 2001). A recent cooling trend in the Sea of Okhotsk, beginning in 1997, resulted in a decrease in walleye pollock, cod, flatfish, and sardine and an increase in herring, capelin, anchovy, and squid (Dulepova and Klyashtorin 2008). Despite dramatic declines, pollock remain the most abundant epipelagic fish in the Sea of Okhotsk (PICES 2005). Total biomass of epipelagic fish is currently below levels of the 1970s‐1990s (PICES 2005). In addition, there has been a significant decline in the biomass of all major demersal fish species in the Sea of Okhotsk since 1997, excluding sculpins (PICES 2005).
4.2.1.1.3.3.1.3 Baltic Sea
The salinity, particularly in the surface layers, of the Baltic Sea is much lower than in the Arctic seas or the Sea of Okhotsk, and the Baltic Sea prey community includes a mixture of marine and freshwater species. The coastal fish communities of the Baltic Sea, particularly in the northern portion and Gulf of Bothnia, are dominated by freshwater species (Ådjers et al. 2006), such as perch, roach, and ruffe, all of which have been reported in ringed seal diets. Marine fishes important in the diet of ringed seals in the Baltic Sea include European smelt, Baltic herring (Clupea harengus membras), three‐spined stickleback, and fourhorn sculpin. European smelt are common throughout the coastal waters of the Baltic and are particularly abundant in the gulfs and bays of the northern and eastern Baltic, where several distinct populations are recognized (Shpilev et al. 2005). Baltic herring are found throughout the Baltic Sea and

97
are heavily fished in a mixed fishery along with Baltic sprat (Sprattus sprattus balticus). Baltic herring stocks declined from at least the mid‐1970s until about 2000 while being fished at high levels. Fishing pressure has declined since 2000, and the stock is growing again (Hay et al. 2001). The fourhorn sculpin is a circumpolar marine and brackish water species with a relict distribution in the Baltic Sea. It is common in the northern Baltic Sea and has recently been recorded for the first time from the southern Baltic (Bacevičius 2009).
Invertebrates important in the diet of ringed seals in the Baltic Sea include the isopod Saduria entomon and the mysid Mysis relicta. Saduria entomon is a large benthic isopod with a circumpolar distribution, and glacial relict populations exist in the Baltic Sea as well as several freshwater lakes (Haahtela 1990). In most of its Arctic range, it prefers shallow estuarine waters (Percy 1983), but in the Baltic Sea, its range is generally deeper and specimens have been found as deep as 290 m (Haahtela 1990). Mysis relicta is a nectobenthic mysid crustacean abundant in the shallow waters of the northern Baltic Sea, while a similar species, M. mixta, is more common in the high salinity waters of the southern Baltic (Salemaa et al. 1990).
The oceanography of the Baltic Sea is dominated by epidsodic water flows from the North Sea which bring salt, fish larvae, plankton, and oxygen to the often stagnant deeper sea depths. Inflows are highly variable in volume—after the longest stagnation period on record, starting in 1977, a major inflow of salty and oxygen‐rich North Sea waters in 1992 oxygenated the bottom waters. The salinity of the bottom water has decreased since then due to decreased frequency and volume of inflows (Jansson 2003). The dynamics of North Sea inflow are important for the fish community—the successful spawning of cod is partly regulated by oxygenation periods in deep water (Jansson 2003). During the 1977‐1992 period of stagnation and high salinities, the weight‐at‐age of herring decreased by 50%, presumably a result in the decline of their copepod prey (Flinkman et al. 1998, Vuorinen et al. 1999). The benthic communities of the Baltic are also impacted by fluctuations from non‐anoxic to anoxic conditions. The benthic area of intermittent oxygen deficiency has more than tripled since the 1940s due to long and frequent stagnation periods (Elmgren 1989, Jonsson et al. 1990).
The Baltic salmon population has suffered from the damming of rivers, disease, and overfishing. Artificial rearing and release of hatchery smolts has compensated somewhat for the decline in wild stocks but has resulted in a decrease in genetic diversity and an increase in vulnerability to environmental stress. The wild population has decreased to 10‐15% of the total standing stock, the rest being artificially reared (Jansson 2003).
4.2.1.1.3.3.1.4 Lake Ladoga and Lake Saimaa
The diet of ringed seals in Lake Ladoga and Lake Saimaa consists primarily of pelagic schooling fishes, including European perch, roach, ruff, vendace, and smelt. Other more benthic species, such as burbot and fourhorn sculpin, however, have also been found in the diets of ringed seals in the lakes. Most of the fish species preyed upon by ringed seals in these lakes are broadly distributed throughout Eurasia, and some (e.g., burbot and fourhorn sculpin) are circum‐Arctic.

98
4.2.1.1.3.3.2 Projected changes in prey
Using evidence from recent past warming episodes to forecast climate change effects on ecosystems may not always be appropriate. (Bakun 1990) cautions about performing such extrapolations because of the different causal mechanisms at work in controlling marine ecosystem dynamics. His caution, however, is most appropriate for very complex oceanographic systems such as upwelling zones.
4.2.1.1.3.3.2.1 Impacts of ocean warming on prey
In general, marine populations exhibit a wide array of distribution patterns, reflecting their relative sensitivities to the seasonal temperature cycle (Taylor et al. 1957, Colton 1972, Scott 1982, Murawski and Finn 1988). Warming of the oceans is predicted to drive species ranges toward higher latitudes (Parmesan and Yohe 2003). Climate change can strongly influence fish distribution and population size (Wood and McDonald 1997) through changes in growth, survival, reproduction, and spawning distribution (Sundby and Nakken 2008). Rapid warming might exceed the ability of local forage species to adapt, thereby causing a major restructuring of regional ecosystems as was observed in the North and Baltic Sea ecosystems (MacKenzie and Schiedek 2007). It can be predicted with some certainty that further shifts in spatial distribution and northward range extensions are inevitable and that the species composition of the plankton and fish communities will continue to change under a warming climate (Mueter et al. 2009).
The demersal fish prey of ringed seals are likely to be impacted by ocean warming. For example, Arctic cod is primarily an Arctic species associated with cold waters. The location and extent of cold bottom water less than 2°C in the summer (a.k.a. “the cold pool”) in the Bering Sea is linked to the extent of ice cover during the previous winter such that the cold pool is more extensive and expands to the south and east after high ice years. Arctic cod have been found primarily in these cold pool waters such that their summer distribution is also more extensive and further to the south and east after high ice winters. Conversely, Arctic cod distribution is contracted towards the north and west in summers following low winter ice cover (Wyllie‐Echeverria and Wooster 1998). Thus, continued ocean warming and decreased winter ice extent in the future could potentially reduce the southerly and easterly extent of the distribution of Arctic cod in the Bering Sea. Arctic cod in the Beaufort Sea are similarly associated with cold waters. A survey of marine fish of the Beaufort Sea conducted in summer 2008 showed that adult Arctic cod in both benthic and pelagic habitats apparently prefer the very cold winter water that emanates from the Chukchi Sea (Logerwell et al. 2010). This is the region of the cold halocline that characterized Chukchi Sea outflow through Barrow canyon, and fish distributions and model results suggest that the fish prefer these low temperatures. The Chukchi Sea outflow of cold, winter‐formed waters are also rich in dissolved and particulate organic carbon, with this water carried offshore into the halocline (Pickart et al. 2005, Mathis et al. 2007). Previous studies in the marine coastal habitat of the Beaufort Sea have similarly shown that Arctic cod prefer waters that are cold (‐1° to 3°C) and of high salinity (27‐32 ppt; Craig 1984). Future warming of Chukchi and Beaufort Seas potentially will bring about changes in the distribution and/or numbers of Arctic cod.

99
Ocean warming will not only impact gadids such as Arctic cod. Community‐wide responses involving a number of demersal fish and invertebrate species to warming conditions in the Bering Sea have been observed, for example: northward distribution shifts, changes in community metrics such as total biomass, species richness, and average trophic level. Those observations suggest the potential for highly disruptive responses of the benthic community to continued climate change (Mueter and Litzow 2008). Demersal fish in the Atlantic Arctic have also demonstrated sensitivity to ocean warming. Nearly two‐thirds of exploited and non‐exploited demersal fish species in the North Sea showed a northward shift (average shift was 172.3 km/°C or 12 km per decade; Perry et al. 2005) in response to recent (post‐1980s) rapidly warming water temperatures (Perry et al. 2005). The rapid warming led to the northward migration of southern species and the ecosystem changed from one dominated by cold‐water species to the one dominated by warm‐water species. In the northwest Atlantic Ocean, centroids of mean catches of fish from bottom trawl surveys also shifted north by 0.5‐0.8 degrees of latitude for each 1°C increase in average water temperature (Murawski 1993).
Ocean warming may also impact the distribution of pelagic fish that are prey for ringed seals. There have been range extensions of pelagic fish such as pollock and Pacific salmon in the northern Bering Sea associated with increased temperatures and decreased ice cover, and the evidence is for a continued trend towards more subarctic conditions in that area (Grebmeier et al. 2006b). In the Barents Sea, higher zooplankton biomass has been observed during recent years with warm ocean temperatures and increased inflow of North Atlantic water (1991‐2000) compared to previous cold years (1984‐1990) (Dalpadado et al. 2003). Thus, warm years would be expected to be favorable for pelagic zooplanktivorous fish such as capelin and herring. In addition, the distribution of capelin has been shown to be linked to ocean temperatures, being shifted to the north and east during warm ocean years (Loeng 1989). In the Russian North Pacific, model predictions indicate cooling until approximately 2020 with decreases in “warm water” pelagic species such as salmon, pollock, and sardines and then increases after 2020. Herring and other “cold water” species will increase until 2020 and then decline (Dulepova and Klyashtorin 2008). The pelagic zooplankton prey of ringed seals may also be impacted by future ocean warming. Observations that warming associated with the 1976/77 regime shift led to decreases in zooplankton crustaceans in the Bering Sea and subarctic Pacific support this hypothesis (Sugimoto and Tadakoro 1997, Schumacher et al. 2003, Bluhm and Gradinger 2008).
In addition to shifts in the distribution of species ranges within Arctic Ocean regions, it is reasonable to expect trans‐Arctic invasions of the Atlantic by North Pacific species concurrent with a decrease in sea ice and ocean warming. These invasions last occurred during the warm mid‐Pliocene epoch (Vermeij and Roopnarine 2008). Given that marine invasions rarely lead to extinctions, the expectation is that the trans‐Arctic invasions will result in an increase in species diversity in the Atlantic. Few Atlantic to Pacific invasions are predicted because Pacific‐derived species are larger‐bodied than Atlantic and would thus be capable of excluding Atlantic invaders.
4.2.1.1.3.3.2.2 Impacts of changes in oceanographic processes on prey
Several of the dominant zooplankton taxa that are either preyed upon directly by seals or are food of pelagic fish upon which seals prey are linked to the Arctic oceanographic regime. Thus, fluctuations or

100
long‐term changes in large‐scale oceanographic processes could alter the species composition of zooplankton on Arctic shelves (Bluhm and Gradinger 2008). For example, greater inflow of North Atlantic water into the Barents Sea is associated with warm ocean conditions and higher zooplankton biomass overall (Dalpadado et al. 2003). Greater inflow of Atlantic water, however, is associated with lower biomass of Arctic amphipod species such as Themista libullula (Dalpadado 2002).
The following scenario for the Bering Sea has been formulated by experts in their fields, assuming that the next decade resembles the unusual conditions of the late 1990s (Schumacher et al. 2003). The key process would be a reduction in on‐shelf transport (observed in 1997). Such a reduction would result in a decrease in flux of nutrients to the shelf, weaker stratification, reduced influence of sea ice, and warmer ocean temperatures. The expected biological response would be a decline in annual primary production and a spring phytoplankton bloom of lower biomass but longer duration, favoring pelagic rather than benthic production.
Increased river runoff is another predicted consequence of a warming climate, with associated increases in inorganic nutrients, increased turbidity, and decreased salinity in coastal waters (ACIA 2004). Higher biomasses of phytoplankton and zooplankton have been observed in marine waters compared to estuarine waters associated with river runoff such that a future with more freshwater input could mean lower prey biomass in the coastal pelagic ecosystem of the Arctic (Bluhm and Gradinger 2008). In the Baltic Sea modeling, the effects of global warming and increased precipitation indicate a gradual reduction in salinity with possible shifts in the species composition of the biota (Omstedt et al. 2000).
4.2.1.1.3.3.2.3 Impacts of changes in distribution and abundance of sea ice on prey
Ice algae contribute 4‐26% of the production in seasonally ice‐covered waters and greater than 50% in the permanently covered central Arctic (Gosselin et al. 1997). Spring algal blooms in the bottom layer of sea ice can be substantial, above 70 mg Chl a m‐2 (Smith et al. 1993). The ice community is important to the Arctic ecosystem not only because it contributes to primary production but also because it extends the grazing season of herbivores (ice algae blooms are available before the open water spring bloom). In addition, high densities of algae on the 2‐dimensional undersurface of the ice my increase the grazing efficiency of zooplankton compared to the 3‐dimensional open‐water habitat (Bradstreet and Cross 1982). The association of Arctic cod with sea ice is well documented in coastal and offshore Arctic locations across the globe (reviewed in Bradstreet et al. 1986). Arctic cod appear to use sea ice for both feeding and for protection from predators by hiding in water wedges along the edges of ice floes (Gradinger and Bluhm 2004). For example, several species of Gammaridean amphipods are endemic consumers of algal ice production in many different areas of the Arctic and in turn are important prey for Arctic cod (Bradstreet and Cross 1982, Lønne and Gulliksen 1989, Lønne and Gulliksen 1991a, Lønne and Gulliksen 1991b, Carey 1992, Polterman et al. 2000, Werner and Gradinger 2002). It is expected that thinning of Arctic sea ice and the resulting increased meltwater and change in morphology of floes will have a negative effect on these under‐ice amphipods (Werner and Gradinger 2002). Feces of ringed seals collected near the ice edge have been shown to be dominated by Arctic cod and amphipods (Bradstreet and Cross 1982).

101
Loss of summer sea ice due to ocean warming could result in a decrease in the availability of this ice‐based food web for foraging seals (Bluhm and Gradinger 2008). On the other hand, increased summer ice melt could result in increased primary productivity as more of the ocean surface is exposed to sunlight (Anderson and Kaltin 2001). Nutrients, however, could limit the extent of increases in production. Scenarios based on contemporary nutrient concentrations suggest a doubling of production in the central Arctic with loss of summer sea ice, but no increase in coastal and shelf regions (Bluhm and Gradinger 2008). Increased sea‐ice melting is also expected to lead to more brackish conditions (i.e., warmer and fresher water) with potential impacts on the ice‐associated food web. For example, since the 1970s‐1980s, there have been changes in the ice‐associated phytoplankton and zooplankton communities of the Canada Basin attributed to ice melt and subsequent warming and freshening. Specifically observed were changes in the species composition of phytoplankton, a decrease in viable populations of copepods and amphipods within the ice interior, and a decrease in the number of species and overall biomass of zooplankton associated with the under surface of the ice (Melnikov et al. 2002).
Ocean warming and loss of winter sea ice may alter the coupling between pelagic and benthic production, further impacting Arctic food webs. In the northern Bering Sea, late retreat of winter ice cover is thought to result in an ice‐associated phytoplankton bloom early in the spring. Herbivorous zooplankton are in an early phase of their annual ontogenetic cycle at this time and cannot graze down the phytoplankton bloom with the result that much of the production sinks to the bottom and fuels a benthic food web (Hunt et al. 2002, Grebmeier et al. 2006a). In contrast, early retreat of winter ice cover associated with warmer ocean waters is hypothesized to result in a later spring phytoplankton bloom in open water. In this situation, herbivorous zooplankton would be further along in their ontogenetic cycle and able to graze the phytoplankton bloom such that less production goes to the benthos. By a similar mechanism, zooplankton in the Barents Sea are able to crop more of the primary production in years when water temperatures are high (Rey et al. 1987, Skjoldal et al. 1987). Thus, warming and decrease in ice extent could increase pelagic productivity in favor of ringed seals (Bluhm and Gradinger 2008).
4.2.1.1.3.3.2.4 Impacts of ocean acidification on prey
Ocean acidification is expected to hamper the ability of phytoplankton such as foraminifera and coccolithophorids to maintain their calcareous structures, with negative consequences for the rest of the marine food web (Feely et al. 2004, Orr et al. 2005). Laboratory experiments on the coccolithophore, Emiliania huxleyi, have resulted in inconsistent results (Riebesell et al. 2000, Iglesias‐Rodriguez et al. 2008). Further research on species‐specific responses of phytoplankton to increased ambient CO2 is needed. Ocean acidification also could have severe consequences for calcifying zooplankton, especially shelled pteropods which are prey for carnivorous zooplankton and fish such as salmon, herring, and cod (Lalli and Gilmer 1989, Willette et al. 2001, Boldt and Haldorson 2003, Orr et al. 2005). Experiments and in vitro measurements show that pteropods are unable to maintain their shells in waters that are undersaturated with calcium carbonate, a consequence of ocean acidification, with expected negative impacts on their survival (Byrne et al. 1984, Feely et al. 2004, Orr et al. 2005).
In addition to interfering with calcification of organisms at lower trophic levels, changes in ocean chemistry can have direct effects on the physiology of marine invertebrates and fish. In general, the

102
expectation is that an increase in CO2 will cause changes in the mode of metabolism of marine organisms, with possible reductions in growth and reproduction. The hypothesis, however, has not been tested in long‐term field experiments (Pörtner et al. 2004). CO2 impacts animal physiology predominantly through its acidifying effect on acid‐base balances. Changes in acid‐base balance will impact membrane‐bound ion regulation, as observed for both fishes and crabs. Compensation for increased acidity causes potentially unfavorable changes in the ionic composition of plasma and other body fluids (Pörtner et al. 2004). These processes are not likely to be life‐threatening but are expected to hamper growth, reproduction, and survival (Pörtner et al. 2005). For example, studies have demonstrated detrimental effects of low pH on the growth of shrimp and sturgeon (Wickins 1984, Crocker and Cech 1996), and on reproductive success of perch (Vinogradov and Komov 1985). Another common adaptive strategy to increased CO2 is to suppress aerobic energy rates, or “metabolic depression” (reviewed in Guppy and Withers 1999). Comparing patterns seen in fish with those found in invertebrates indicates that the effect of metabolic depression occurs with smaller increases in CO2 in invertebrates. Vertebrates apparently have an enhanced capacity to avoid early metabolic depression in the face of long‐term elevated CO2 levels. The processes that control vertebrate metabolism under conditions of increased CO2, however, reflect changes in energy use and allocation with potential consequences for long‐term survival (Pörtner et al. 2005).
Squid may be particularly intolerant of increases in CO2 due to their unusually high metabolic rate and their unique respiratory physiology. Compared to squid, fish are expected to be more tolerant of increases in CO2, a result of their lower metabolic rate and venous oxygen reserve. Deep‐sea fishes are expected to be the least tolerant group of fishes because of their adaptations to respiration in low‐oxygen environments (Pörtner et al. 2004). A laboratory experiment on a Mediterranean sea bream (Sparus aurata) indicated that prolonged exposure to moderate increases in CO2 caused shifts in metabolic pathways from aerobic to anaerobic metabolism (Michaelidis et al. 2007). Other studies have shown an alteration in protein biosysthesis or patterns of gene expression (Langenbuch and Pörtner 2003). It is not known whether these processes are adaptive to long‐term increases in ocean CO2
concentrations or whether they would contribute to decreased growth and survival (Pörtner et al. 2005).
Additional concerns about ocean acidification are related to the fact that future climate change scenarios will involve not only increases in oceanic CO2 concentrations but also increased warming and decreased dissolved oxygen due to eutrophication. Because CO2 affects several physiological mechanisms that are also affected by thermal extremes and oxygen deficiency, it has been suggested that current trends of warming, CO2 increase, and oxygen reduction in marine waters may exert harmful synergistic effects on marine organisms (Pörtner et al. 2005). For example, physiological responses to increased CO2 will likely result in a narrowing of the thermal tolerance of marine organisms, which would be most detrimental for animals living at the edges of their distributional range. This interaction could cause shifts in geographical distribution as well as alteration of ecosystem composition and functioning (Pörtner et al. 2005).

103
Current rates of change in ocean acidity are more than 100 times any changes in the past 100,000 years (Raven et al. 2005). The high rate of increase in CO2 concentration means that organisms are being exposed to changes that are faster than they have encountered in their recent evolutionary history. The capacity of calcifying marine organisms to adapt to progressively acidified oceans is not known, but will depend, in part, on the species’ generation time. Long‐lived species—such as benthic molluscs—will be less able to respond adaptively. Shorter generation times may afford increased opportunities for micro‐evolutionary adaptation (Raven et al. 2005). A recent National Academies report concluded that “the chemistry of the ocean is changing at an unprecedented rate and magnitude due to anthropogenic CO2 emissions; the rate of change exceeds any known to have occurred for at least the past hundreds of thousands of years” (National Research Council 2010). The report went on to point out that while the ultimate consequences of ocean acidification are still unknown, both the magnitude and rate of change present a risk of ecosystem changes that threaten coral reefs, fisheries, protected species, and other natural resources of value to society.
4.2.1.1.3.3.2.5 Other anthropogenic impacts on prey
The drainage area of the Baltic Sea is home to a human population of around 85 million in an area four times larger than the sea itself, thus the impact of anthropogenic nutrients and toxins—such as metals, polychlorinated biphenyls (PCBs), and dichloro‐diphenyl‐trichloroethane (DDT)—is unusually high (Jansson 2003). The HELCOM developed a Joint Comprehensive Action Program in the early 1990s to develop strategies to minimize the environmental effects of future expansions of human activities in the Baltic Sea region. A target reduction in nitrogen and phosphorous loads as well as heavy metals was identified. As of the early 2000s, however, only Poland and Russia had come close to the 50% reduction in nutrient loads, mostly due to economic recession and the resulting decline in the use of fertilizer in the 1990s. Success in achieving the target reduction in heavy metals seems unlikely in the near future as 50,000 to 100,000 different substances are in use and thousands of new ones are introduced annually in industries and households. Although target reductions have not been reached, many countries report improvements in environmental conditions, such as the hygienic condition of beaches resulting in the improvement of sewage treatment facilities (Jansson 2003).
Concentrations of nitrogen and phosphorous have increased by four and eight times, respectively, since 1900 (Larsson et al. 1985) but have leveled off after the 1980s (Jansson 2003). The result of this nutrient input has been an increase in primary production by 30‐70%, and an increase in zooplankton of about 25% (Elmgren 1989). Sediment loads of metals (mercury, copper, lead, zinc and cadmium) started to rise in the 1950s, peaked during the 1960s and 1970s and decreased after the 1980s. Mercury has been a major threat to top predators such as birds of prey and seals (Jansson 2003). PCBs and DDT were widely used during the 1960s and 1970s with negative impacts on eagles, seabirds, seals that prey on herring. After these toxins were banned in the 1970s, the concentrations in tissues of herring declined and threatened populations are recovering, although PCB emissions still occur through dumping and incineration of old materials containing PCBs. Cadmium concentrations in the tissues of marine organisms in the central Baltic are increasing in spite of restrictions on its use (Jansson 2003).

104
4.2.1.2 Impacts of global climate change on ringed seals
Ocean warming and acidification will impact ringed seals primarily through changes in community composition. The identity and impacts of new parasites, competitors, prey, and predators can be predicted only in general terms. As ice cover diminishes, the ringed seal’s ability to inhabit ice‐covered seas year round will no longer be a competitive advantage, nor will it provide refuge from predators such as killer whales (Orcinus orca). The most substantial impacts to ringed seals in a changing climate are likely to issue from changes in ice and snow cover.
4.2.1.2.1 Impacts related to changes in ice and snow cover
The number of breeding species among pinnipeds peaks in the seasonally ice‐covered seas of the Northern and Southern Hemispheres (Kelly 2001). Contributing to the reversal of the usual pattern of increasing species richness with increasing latitude is the fact that pinnipeds divide their time between foraging in the water and reproducing and molting out of the water. They are especially vulnerable to predation out of the water, since their limbs and overall body shape are adapted for aquatic locomotion with a consequent decrease in their agility on land (Allen 1880). In the tropical and temperate zones, islands provide havens from predation and are the main whelping and molting sites, but islands compose only 1,400,000 km2 or less than 1% of the earth’s land area. In the polar regions, however, sea ice provides a more extensive substrate—approximately 36,000,000 km2 in recent decades—and is used regularly as a whelping and molting platform by 13 of the 19 extant species of phocids and by walruses (Odobenus rosmarus).
Seasonal sea ice in the Arctic began to form as long as 46 million years ago (Moran et al. 2006), predating the divergence of seals from their terrestrial ancestors (Higdon et al. 2007, Rybczynski et al. 2009, Fulton and Strobeck 2010a). Perennial ice apparently formed in the Arctic Ocean about 14 million years ago (Polyak et al. 2010) consistent with the suggestion that pinnipeds adapted to breeding on Arctic sea ice 13‐17 million years ago (Árnason et al. 2006, Higdon et al. 2007). Episodes of seasonally ice‐free conditions may have occurred in the Arctic Ocean prior to the Quaternary glaciations, and ice extent may have been below 20th century observations during some interglacial periods, but additional data and analyses are needed to determine the actual extent of ice cover in those periods (Polyak et al. 2010). Ringed seals survived those interglacials as a species, but we do not know whether all population segments survived or how low their populations might have become. Declines in sea‐ice cover in recent decades are more extensive and rapid than any known for at least the last few thousand years (Polyak et al. 2010).
Indigenous hunters have long recognized the importance of sea‐ice dynamics to marine mammal populations (Shapiro et al. 1979, Noongwook 2000, Pungowiyi 2000). Vibe (1950, 1967) noted that climate influenced seal and walrus distributions by its strong effects on sea ice. Fay (1974) reviewed the role of ice in the ecology of marine mammals in the Bering Sea, and pointed out that the timing of reproduction and molting among ringed seals and other ice‐associated pinnipeds was adapted to seasonal ice conditions. Burns et al. (1981) and Braham et al. (1981) expanded on Fay’s descriptions

105
drawing on extensive surveys in the Bering Sea and on remote sensing records of sea‐ice distribution and phenology.
Stirling (1997) reviewed the importance to ringed seals of highly productive waters adjacent to sea ice. Tynan and DeMaster (1997) suggested that ringed seals might be especially sensitive to loss of sea ice and reduced snow cover, and they also suggested that plasticity—evident in some populations breeding in shorefast ice and others in pack ice—might favor adaptation to changing environments.
The ringed seal’s sensitivity to ice phenology was evident in a population monitored in the Canadian Arctic in the 1970s and 1990s. Reduced food availability, poor body condition, and low ovulation rates were attributed to unusually heavy winter ice conditions in 1974/1975 (Harwood et al. 2000, Smith and Harwood 2001). In 1998, however, sea ice in the same area broke up 6 weeks earlier than the average for the previous eight years. Food availability and ovulation rates in 1998 were high, but nursing was interrupted by the premature ice break‐up, and pup survival was reduced. Body condition, growth, and survival of the unweaned pups were significantly reduced. At the same time, however, the early break‐up in 1998 resulted in an apparently greater amount of food and increased body condition in the other age classes of seals.
In Amundsen Gulf, the body condition of over 2,000 harvested seals varied widely among years from 1971‐1979 and from 1992‐2009 (L. Harwood, DFO, Canada, pers. comm., 9 July 2010). The extremes in body condition matched the timing of break‐up; seals sampled in years of late break‐up had the lowest body condition indices (e.g., 1974 and 2005), and the highest body condition was found in years with an early break‐up (e.g., 1998 and 2008). In the two late break‐up years, the seals were in significantly poorer body condition, and there were major declines in reproductive output. Break‐up date influences the amount of time there is to feed and perhaps the absolute quantity of food as well as the energetic quality of the prey items available during June when seals increase feeding after lactation and the annual molt (McLaren 1958a, Smith 1973, Smith 1987).
Sea‐ice phenology will change substantially over the next 90 years and probably beyond. Changes will be rapid relative to ringed seal generation time and, thereby, will limit adaptive responses. The changes will be most severe in the marginal ice zones, and suitable ice regimes for ringed seal reproduction and molting likely will be substantially reduced or lost in the Sea of Okhotsk, the Baltic Sea, Lake Ladoga, and Lake Saimaa by the end of the century (Meier et al. 2004). Suitable ice is likely to persist in the Arctic Ocean, albeit to a greatly reduced extent.
Ringed seals require snow cover to provide shelter from extreme cold and from predators, and they very likely will be negatively impacted by shorter periods of seasonal snow cover as the climate warms (ACIA 2005). As early as the reports of the Howgate Polar Expedition (1877‐1878), it has been recognized that ringed seals born without benefit of a subnivean lair “almost always fall a prey to foxes and ravens” and other predators (Kumlien 1879). McLaren (1958a) pointed out that the southern extent of the ringed seal’s range corresponds to the latitudes to which snow cover—sufficient to form and maintain subnivean lairs—extends. When snow cover is insufficient, pups freeze in their lairs as documented in 1974 when roofs of lairs in the White Sea were only 5‐10 cm thick (Lukin and Potelov 1978). Similarly,

106
pup mortality from freezing and polar bear predation increased when unusually warm spring temperatures caused early melting near Baffin Island in the late 1970s (Smith and Hammill 1980b, Stirling and Smith 2004). Prematurely exposed pups also are vulnerable to predation by wolves and foxes, as documented during an early snow melt in the White Sea in 1977 (Lukin 1980); by gulls and ravens, as documented in the Barents Sea (Gjertz and Lydersen 1983, Lydersen and Gjertz 1987, Lydersen et al. 1987, Lydersen and Smith 1989, Lydersen and Ryg 1990, Lydersen 1998); and possibly by snowy owls (Bubo scandiacus) (Lydersen and Ryg 1991). Avian predation is facilitated not only by lack of sufficient snow cover but also by conditions favoring influxes of birds. Lydersen and Smith (1989) noted that the proximity of open water at Svalbard exposed unconcealed ringed seal pups to predation by glaucous gulls (Larus hyperboreus). Gulls and ravens have been increasing along the Arctic coast of Alaska as a result of habitat changes associated with industrial activities (Johnson and Herter 1989, Truett et al. 1997, Johnson and Noel 2005). Lydersen and Smith (1989) pointed out that the small size of newborn ringed seals, coupled with their prolonged nursing period, makes them vulnerable to predation by birds and likely sets a southern limit to their distribution. That is, the southern limit corresponds to the latitudes at which snow conditions are suitable for excavating and occupying subnivean lairs. Similarly, unseasonal melting of snow dens is believed to threaten polar bears, especially in the southern portions of their range (Clarkson and Irish 1991, Stirling and Derocher 1993).
Over most of the Arctic Ocean, snow accumulation is sufficient for lair formation only where pressure ridges or hummocks in the ice cause the snow to form drifts (Smith and Hammill 1980a). Lydersen et al. (1990) linked low densities of ringed seals in Van Mijen Fiord, Svalbard to a lack of lack of snow drifts in especially flat ice. Ringed seals maintained breathing holes but almost no lairs in fjords with average snow depths less than 30 cm (Lydersen et al. 1990, Lydersen and Ryg 1991). Smith and Lydersen (1991) contrasted quality of sea ice as ringed seal habitat in the Canadian Arctic Archipelago to that at Svalbard. They considered the amount of time prior to the whelping season that stable ice accumulated snow to be “the primary factor influencing the quality of ringed seal breeding habitat.” They also pointed out that sufficient snow depths for lair formation only occurred where wind conditions and ice deformation favored drifting. In the Canadian Arctic Archipelago, they found large areas of deformed ice with snow drifts deep enough for ringed seal lairs consistent with reports that snow accumulation on Arctic sea ice is maximal in the Canadian sector (Radionov et al. 1997). In contrast, only small areas around glacier fronts in Svalbard accumulated sufficient snow for lairs. Other areas, such as Van Mijenfjorden, accumulate little snow as a consequence of late ice formation and minimal deformation to cause drifts. At Van Mijenfjorden, Lydersen et al. (1987) observed evidence of high pup mortality during a year of especially low snow accumulation. They observed six pups born outside of subnivean lairs, and all six were killed by foxes or birds. They also reported that 10 of 11 mature females that they collected that year had lost pups.
Based on studies in the White Sea, Lukin et al. (2006) concluded that the depth of snow cover and the timing of ice break‐up were “the key factors affecting” reproduction and population trajectories of ringed seals. They reported that the survival rates of ringed seal pups in the White Sea decreased since 1978 with a nearly doubling of the frequency of mild winters accompanied by shorter periods of ice cover and lower snow accumulation.

107
The Saimaa ringed seal’s habit of excavating lairs along shorelines is attributed to the fact that—in the absence of deformed ice—the only drifts of sufficient depth occur along the shores (Helle et al. 1984). The majority of lairs in Lake Saimaa are found on the northern and eastern sides of islands where snow drifts persist longest in the spring (Helle et al. 1984). Pressure ridges are limited to only some portions of Lake Ladoga ice, and snow accumulation at those ridges is sufficient for lair formation and occupation. Most of the lairs—including all birth lairs—in Lake Ladoga are found, however, in deeper drifts along shorelines (Kunnasranta et al. 2001).
Tynan and DeMaster (1997) noted that climate models (e.g., Manabe et al. 1992, IPCC 1996) predict increased precipitation in the Arctic Ocean and marginal seas with potentially beneficial effects for lair occupation by ringed seals. The climate models, however, also forecast a decreased duration of ice cover, and the net result is that overall snow accumulation may decrease (Sturm et al. 2002), and the date of snow melts will advance leading to premature exposure of pups to wet and cold conditions as well as to predation. Support for those predictions was reported in a multi‐decadal study of recruitment of ringed seals in Hudson Bay (Ferguson et al. 2005) and a 5‐year study of factors influencing the emergence of ringed seals from lairs in the Alaskan Beaufort Sea (Kelly et al. 2006).
Ferguson et al. (2005) examined over 600 ringed seals harvested by Native hunters in Hudson Bay and found correlations between survival of young ringed seals and sea‐ice and snow conditions. In the 1970s and 1980s, they observed that variation in survival corresponded to variations in ice break‐up. Survival was enhanced by early break‐up of the ice, an effect attributed to better access to prey. In the 1990s, however, they found that the greatest proportion of variance in pup survival was explained by snow depth. Overall, pup survival was low in the 1990s and correlated with shallow snow cover and warmer temperatures during the spring months when pups are most dependent on subnivean lairs. Ferguson et al. (2005) also reported that break‐up in Hudson Bay advanced by just over 3 weeks in the past 3 decades. Snow depth at western Hudson Bay weather stations was less than 20 cm during 7 of 8 recent years and, combined with high spring air temperatures, likely caused early failure of many subnivean lairs.
Ice conditions in Hudson Bay are not conducive to on‐ice field investigations, and Ferguson et al.’s (2005) study was necessarily correlative. Shorefast ice along the Alaskan Beaufort Sea coast is accessible by surface travel, and Kelly et al. (2006) reported direct measurements of snow conditions and the responses of radio‐tagged ringed seals during the whelping seasons of 1999‐2003. They showed that ringed seals abandoned subnivean lairs when the snow became saturated with melt water, an event readily detected by temperature sensors in the snow and by satellite‐borne sensors measuring microwave emissions from the snow surface. An algorithm developed by Belchansky et al. (2004) determined the date of snow melt from passive microwave emissions, and those snow melt dates were strong predictors of the dates of final lair abandonment by ringed seals (r2 = 0.982, p = 0.001). Applying that algorithm to the satellite microwave record of snow on the sea ice near Prudhoe Bay, Alaska between 1979 and 2007 showed that the dates on which snow melt would have forced ringed seals from their lairs varied considerably from year to year (Figure 30). Overall, however, the snow melt at that site has advanced by 7 days in the past 30 years.
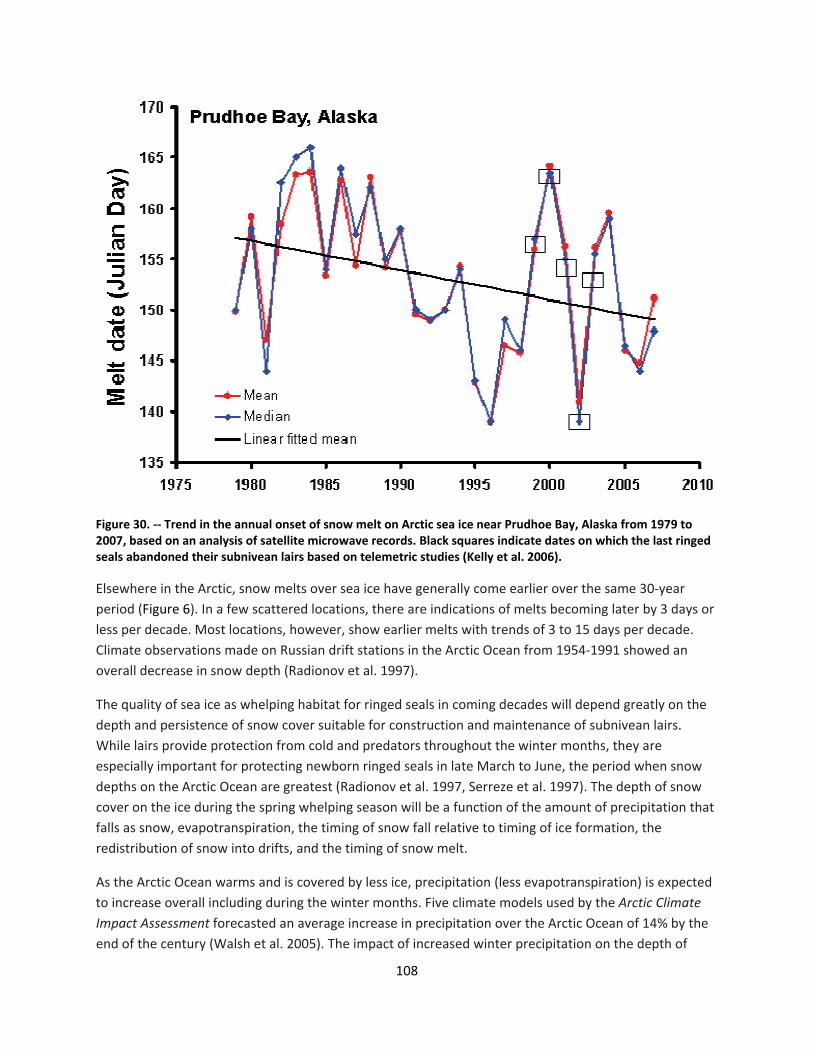
108
Figure 30. ‐‐ Trend in the annual onset of snow melt on Arctic sea ice near Prudhoe Bay, Alaska from 1979 to 2007, based on an analysis of satellite microwave records. Black squares indicate dates on which the last ringed seals abandoned their subnivean lairs based on telemetric studies (Kelly et al. 2006).
Elsewhere in the Arctic, snow melts over sea ice have generally come earlier over the same 30‐year period (Figure 6). In a few scattered locations, there are indications of melts becoming later by 3 days or less per decade. Most locations, however, show earlier melts with trends of 3 to 15 days per decade. Climate observations made on Russian drift stations in the Arctic Ocean from 1954‐1991 showed an overall decrease in snow depth (Radionov et al. 1997).
The quality of sea ice as whelping habitat for ringed seals in coming decades will depend greatly on the depth and persistence of snow cover suitable for construction and maintenance of subnivean lairs. While lairs provide protection from cold and predators throughout the winter months, they are especially important for protecting newborn ringed seals in late March to June, the period when snow depths on the Arctic Ocean are greatest (Radionov et al. 1997, Serreze et al. 1997). The depth of snow cover on the ice during the spring whelping season will be a function of the amount of precipitation that falls as snow, evapotranspiration, the timing of snow fall relative to timing of ice formation, the redistribution of snow into drifts, and the timing of snow melt.
As the Arctic Ocean warms and is covered by less ice, precipitation (less evapotranspiration) is expected to increase overall including during the winter months. Five climate models used by the Arctic Climate Impact Assessment forecasted an average increase in precipitation over the Arctic Ocean of 14% by the end of the century (Walsh et al. 2005). The impact of increased winter precipitation on the depth of

109
snow on sea ice, however, will be counteracted by later formation of sea ice. Over most of the Arctic Ocean, snow cover reaches its maximal depth in May, but most of that accumulation takes place in the autumn (Sturm et al. 2002). Snow depths reach 50% of the annual maximum by the end of October and 67% of the maximum by the end of November (Radionov et al. 1997). Thus, delays of 1‐2 months in the date of ice formation will result in substantial decreases in spring snow depths despite the potential for increased winter precipitation. Thinner ice, however, will be more susceptible to deforming and producing pressure ridges and ice hummocks favoring snow drifts where depths exceed those on flat ice (Iacozza and Barber 1999, Sturm et al. 2006). As spring air temperatures continue to warm, snow melt will continue to come earlier in the year. Our model projections are for a reduction in snow cover on Arctic sea ice of approximately 50% by the end of the century (Figures 27‐28). A regional climate model likewise predicts decreases in the duration of seasonal ice cover of 1‐3 months for the Baltic Sea (Meier et al. 2004), shortening the duration of snow catchment. In the Norwegian Arctic, air temperatures have increased by 1°C in the past century and are projected in regional models to increase at a similar rate in the current century (Hanssen‐Bauer 2002). Between 1975 and 2001, the fraction of precipitation falling as snow in that region diminished by almost 50% (Førland and Hanssen‐Bauer 2003).
Anomalies in the seasonal cycles of ice and snow cover occur from year to year and can affect the whelping, breeding, and molting of ringed seals. Unusually early and heavy ice has been implicated in shifting distribution, high winter mortality, and reduced productivity (Stirling et al. 1975, Burns and Eley 1978, Smith and Stirling 1978). Stirling et al. (1982) suggested that heavy ice affects ringed seals by increasing the difficulty of maintaining breathing holes and by decreasing light penetration through the ice and thus decreasing primary production. Lukin (1980) attributed the apparent freezing death of pups in the White Sea to insufficient snow cover. Kelly et al. (1986) attributed low densities of pupping lairs along much of the Alaskan coast in 1983 to minimal snow accumulation. Rapid melting of snow along the Beaufort Sea coast of Alaska that same year resulted in reduced areas suitable for molting, with unknown consequences (Kelly et al. 1986).
The combination of late freeze‐ups and early snow melts will deprive ringed seals of sufficient snow cover for subnivean lair occupation, especially during the spring months when young are born and nursed in lairs. Ringed seal pups born on the ice will be subject to high rates of mortality through hypothermia and predation.
Persistence of ringed seals will depend on the amount and phenology of ice and snow cover, habitat features forecasted to change heterogeneously over the next century. As pointed out previously, the period over which snow accumulates on ice is “the primary factor influencing the quality of ringed seal breeding habitat” (Smith and Lydersen 1991). Snow drifted to 45 cm or more is needed for excavation and maintenance of simple lairs, and birth lairs require depths of 50 cm (Lukin et al. 2006) to 65 cm or more (Smith and Stirling 1975, Lydersen and Gjertz 1986, Kelly 1988, Furgal et al. 1996, Lydersen 1998). Such drifts typically occur only where average snow depths (on flat ice) are at least 20‐30 cm and where drifting has taken place along pressure ridges or ice hummocks (Lydersen et al. 1990, Hammill and Smith 1991, Lydersen and Ryg 1991, Smith and Lydersen 1991). Ferguson et al. (2005) observed evidence that pup survival dropped sharply when snow depths were less than 32 cm. They found that ringed seal

110
recruitment increased with increasing snow depth (r2 = 0.51, p = 0.021), and suggested that reduced recruitment in recent years resulted from low snow fall yielding lairs excavated in drifts too shallow to protect against predators.
We take 20 cm to be the minimal average snow depth needed for the formation of lairs, and average depths of 30 cm or more appear to be necessary for lairs of sufficient depth to adequately protect against predation. We summarize the regional impacts to ringed seals of changing ice and snow cover as follows.
4.2.1.2.1.1 Chukchi Sea
The model forecasts suggest that sea ice in the Chukchi Sea will provide an adequate substrate for resting, whelping, and nursing young throughout the current century, although ice suitable for resting will decline by the century’s end during June and July when ringed seals are molting. By the end of the century, ice formation in the Chukchi Sea will be delayed as evidenced by forecasts of significantly reduced ice cover in November. The delayed freeze‐up will contribute to a decline in snow accumulation, forecasted to average less than 20 cm by the end of the century. By then, snow cover will be inadequate to form and maintain subnivean lairs.
4.2.1.2.1.2 East Siberian Sea
Model forecasts for the East Siberian Sea also suggest adequate substrate for resting, whelping, and nursing young throughout the current century and some loss of ice during the molting season by the end of the century. Snow depths, however, will decline to be less than 20 cm—below the threshold needed for subnivean lairs—over most of the East Siberian Sea by the end of the century.
4.2.1.2.1.3 Central Arctic Ocean
By mid‐century, sea ice in the central Arctic Ocean is forecasted to decrease in extent, especially in early summer and fall. Ice loss in summer will impact molting and resting ringed seals. The reduction in fall ice will negatively impact snow accumulation. The model forecasts indicate that snow cover will be inadequate for lair occupation in much of the central Arctic Ocean by the end of the century. Snow depths are forecasted to be less than 25 cm throughout and less than 20 cm in half or more of the region.
4.2.1.2.1.4 Beaufort Sea
The models project adequate sea‐ice cover in the Beaufort Sea in the winter and spring through the end of the current century. In the second half of the century, however, sea‐ice extent in the Beaufort Sea will decline during the early summer molting period and in the fall, especially in the coastal areas preferred by ringed seals. Snow cover on the Beaufort Sea ice will be inadequate for lair use by mid‐century (averaging 15‐20 cm) and continue to decline at least until the end of the century (averaging 10‐15 cm).
4.2.1.2.1.5 Bering Sea
Spring sea ice (whelping habitat) declines minimally in model projections for the eastern Bering Sea throughout the century. Larger declines are projected for late autumn with decreases to 28% of current

111
extent by mid‐century and 12% of current extent by 2075. The model used for the western Bering Sea projected sea ice to decline by 75% in the spring whelping and nursing period by the end of the century. In the second half of the century, years with no ice cover in May—when ringed seals are nursing and beginning to molt—will be common. April snow depths in the Bering Sea are projected to average less than 5 cm, far below threshold values for lair formation, by mid‐century.
4.2.1.2.1.6 Barents Sea
By the end of the century, the extent of sea ice in the Barents Sea during the spring whelping and nursing periods is projected to be less than half that of present. Virtually no ice is projected to exist in the Barents Sea during the molting period before the end of the century. April snow depths averaged 10‐15 cm in the vicinity of Svalbard and to the east of Novaya Zemlya in the first decade of the century and are forecasted to decline to 10 cm or less by mid‐century and to less than 5 cm by the end of the century. Thus, snow depths likely will be insufficient for lair formation by mid‐century and almost certainly by the end of the century.
4.2.1.2.1.7 Hudson Bay
The duration of sea‐ice cover in Hudson Bay is forecasted to decline by 7‐9 weeks, and the volume of ice is expected to be reduced by 31% by mid‐century (2041‐2070). Average snow depth on Hudson Bay ice diminished by more than 50% in the past four decades, and the CCSM3 model forecasts continuing declines. Average snow depths already appear to be below levels necessary for lair formation in much of Hudson Bay and are forecasted to inadequate throughout Hudson Bay before the end of the century.
4.2.1.2.1.8 Baltic Sea
Regional climate models forecast reduced sea‐ice extent in the Baltic Sea before mid‐century with drastic decreases by the end of the century. By the end of the century, the depth and insulating properties of snow cover on Baltic Sea ice are forecasted to decrease due to late ice formation, mid‐winter thaws, rain‐on‐snow events, and early snow melts. Such shallow, wet snow will provide poor insulation and—combined with earlier melt seasons—likely will decrease pup survival.
4.2.1.2.1.9 Sea of Okhotsk
Based on temperature proxies, we expect sea ice to persist in the Sea of Okhotsk through the onset of pupping in March through the end of this century. Ice suitable for whelping and nursing likely will be limited to the northern most portions of the sea as ice is likely to be limited to that region in April by the end of the century. Modeling, however, suggested that average snow depths already are below 20 cm—consistent with observation of seals sheltering not under the snow but in the lee of ice hummocks—and snow depths are projected to decrease further through the current century.
4.2.1.2.1.10 Baffin Bay
Sea ice in Baffin Bay is projected to be available to ringed seals for whelping, nursing, and molting, although molting may be truncated by ice loss in July. Snow depths in April, however, are projected to be inadequate for lair use except in a small portion of northeastern Baffin Bay by mid‐century.

112
4.2.1.2.1.11 Canadian Arctic Archipelago
The models project persistence of sea ice in the Canadian Arctic Archipelago throughout most of the year with some reduction in summer ice by the end of the century. The amount of ice covered with adequate snow for lair formation is projected to decline in the Canadian Arctic Archipelago, and by the end of the century, average snow depths will mostly be 20‐25 cm with up to 30 cm in a few isolated locations.
4.2.1.2.1.12 Greenland Sea
The area of sea ice in the Greenland Sea available to ringed seals during the later part of the molt season will decline over the century. Adequate snow depths for lair formation will disappear from all but a small portion of the northwestern Greenland Sea by the end of the century.
4.2.1.2.1.13 Kara and Laptev Seas
Ice extent during the late molt season also is expected to decline in the Kara and Laptev Seas by mid‐century. Freeze‐up in the region will be substantially delayed by the end of the century with impacts on snow accumulation. April snow depths in recent years have been marginal in most areas of Kara and Laptev Seas (averaging 10‐20 cm), and are projected to decline throughout the century.
4.2.1.2.1.14 Lake Ladoga and Lake Saimaa
Simulations of future climate in northern Europe suggest warming winters with reduced ice and snow cover for Lakes Saimaa and Ladoga (Jylhä et al. 2008). Snow cover in the region is projected to decrease 10‐30% before the middle of the century and 50‐90% by the end of the century (Saelthun et al. 1998 cited in Kuusisto 2005). Mortality of Saimaa and Ladoga seal pups likely will increase as the snow cover declines in depth, duration, and insulating quality.
4.2.1.2.2 Impacts related to changes in ocean conditions
The most likely impact of ocean acidification on ringed seals will be through lower tropic levels on which the species’ prey depends. Cascading effects are likely both in the marine and freshwater environments. Our limited understanding of planktonic and benthic calcifiers in the Arctic (e.g., even their baseline geographical distributions) means that future changes will be difficult to detect and evaluate (Orr et al. 2009).
Warming water temperatures and decreasing ice likely will result in a contraction in the range of Arctic cod, a primary prey of ringed seals. The same changes will lead to colonization of the Arctic Ocean by more southerly species, including potential prey, predators, and competitors. The outcome of new competitive interactions cannot be specified, but as sea‐ice specialists, ringed seals may be at a disadvantage in competition with generalists in an ice‐diminished Arctic. Prey biomass may be reduced as a consequence of increased freshwater input and loss of sea‐ice habitat for amphipods and copepods. On the other hand, overall pelagic productivity may increase.

113
4.2.2 Overutilization for Commercial, Recreational, Scientific, or Educational Purposes
4.2.2.1 Commercial, subsistence, and illegal harvest
Ringed seals have been hunted by humans for millennia and remain a fundamental subsistence resource for many northern coastal communities today (Krupnik 1988, ACIA 2005, Kovacs 2007, Hovelsrud et al. 2008). Ringed seals were also harvested commercially in large numbers during the 20th century, which lead to the depletion of their stocks in many parts of their range. Ringed seal meat has been used as food for people, sled dogs, and livestock; their skins sold for cash, traded for goods, or used for clothing, crafts, or other household items; their blubber rendered into oil for food and fuel; and their flippers, bones, and viscera used for many household, industrial, or medicinal purposes (Krylov et al. 1964, Frost 1985, Stewart et al. 1986, Belikov and Boltunov 1998). The following sections summarize the past and present commercial, subsistence, and illegal harvests of ringed seals by region, and ends with a brief discussion of the potential threat of overutilization in the future.
4.2.2.1.1 The Arctic
4.2.2.1.1.1 Alaska
Ringed seals have been an important subsistence resource for many Alaska Native communities along the coasts of the northern Bering, Chukchi, and Beaufort Seas for centuries, however, their harvest levels decreased during the 1970s, likely due to changes in the Natives’ lifestyle and the enactment of the MMPA in 1972 (Frost 1985). The annual harvest in Alaska dropped from 7,000‐15,000 ringed seals per year during 1962‐1972 to 3,000‐6,000 during 1973‐1977, and to 2,000‐3,000 by 1979 (Frost 1985). Based on limited data from two villages on St. Lawrence Island, Kelly (1988) suggested that the annual take in Alaska likely exceeded 3,000 ringed seals during the mid‐1980s.
The ADFG’s Division of Subsistence maintains a database that provides information on the subsistence harvest of seals in different regions of Alaska. Data were compiled for 129 villages from various subsistence harvest reports (e.g., Coffing et al. 1998, Georgette et al. 1998, Wolfe and Hutchinson‐Scarbrough 1999). Data were lacking for 22 villages, so their harvests were estimated using the annual per capita harvest rates from nearby villages. For 16 of these villages, harvest levels were estimated from data collected during the 1980s; otherwise, data from 1990 to 1998 were used. As of August 2000, an estimated 9,500 ringed seals were harvested for subsistence use in Alaska per year (Allen and Angliss 2010). Measures of error were not available for this estimate. Currently, there is no comprehensive effort to quantify harvest levels of seals in Alaska (Allen and Angliss 2010).
Commercial harvesting of marine mammals has been prohibited in U.S. waters since 1972 by the MMPA. Refer to Section 4.2.4 for more information about existing regulatory mechanisms related to ringed seal harvest (and other threats) in all parts of the species’ range.

114
4.2.2.1.1.2 Canada
Ringed seals are by far the most important seal species for human consumption and utilization in the Canadian Arctic (ACIA 2005). Reeves et al. (1998) reviewed the catch history of ringed seals in Canada and concluded that harvest levels were probably highest (likely exceeding 100,000 ringed seals per year) during the 1960s and 1970s when both the value of sealskins and the local demand for seal products were particularly high. Ringed seals may have been locally depleted within the vicinity of some communities where exploitation was most intensive (Mansfield 1970 cited in Reeves et al. 1998). The Royal Commission on Seals and the Sealing Industry in Canada cautioned that “resumption of catching at the 1970s level might see the collapse of some of these stocks” (Malouf 1986 cited in Reeves et al. 1998). Catches of ringed seals declined substantially during the 1980s following a European ban on pup skins and the subsequent decline in sealskin prices (Reeves et al. 1998). Reeves et al. (1998) estimated that the total catch in Canada ranged between about 50,000 and 65,000 ringed seals per year during the 1980s and early 1990s, with the total kill (accounting for hunting losses) ranging between about 60,000 and 80,000 ringed seals per year.
Ringed seals are primarily hunted throughout the Canadian Arctic for subsistence uses (Department of Fisheries and Oceans 2008), however, formal monitoring or reporting is lacking, and harvest studies have only done on a regional basis (Stewart et al. 1986, Reeves et al. 1998). For example, Priest and Usher (2004) estimated that about 25,000 ringed seals were harvested annually by Inuit hunters in Nunavut during 1996‐2001, and approximately 1,500‐2,000 ringed seals were reportedly taken each year off the coast of Labrador during 1997‐2003, although the latter hunting statistics are known to be incomplete (Department of Fisheries and Oceans 2008). TACs or allocations are not set for ringed seals in Canada, but any commercial harvests are regulated by licenses and permits (Department of Fisheries and Oceans 2008). The estimates made by Reeves et al. (1998) of 60,000‐80,000 killed are likely the most recent available for Canada’s annual ringed seal take.
4.2.2.1.1.3 Greenland
Seal hunting is a vital component of everyday life and culture in Greenland and provides a significant amount of nutrition and income to families living in remote coastal communities (Greenland Home Rule 2009). Teilmann and Kapel (1998) provided a thorough review of the exploitation of ringed seals in Greenland. Combining data from all regions (excluding North Greenland during the 1950s, for which no catch records exist), they determined that the catch of ringed seals was about 43,000 per year during the 1950s, fluctuated between about 45,000 and 75,000 per year during the 1960s, steadily increased during the 1970s to a peak of almost 100,000 in 1979, and then decreased to a nearly constant level of about 70,000 per year during the 1980s and early 1990s (Teilmann and Kapel 1998). The authors noted that ringed seals were widely distributed in low densities around Greenland and that hunters mainly took the youngest age‐classes while only catching adult breeders in small numbers (Teilmann and Kapel 1998). They concluded that as long as existing hunting practices and methods were maintained, the existing level of exploitation was unlikely to pose a threat to the Greenland ringed seal population as a whole, but that intensive hunting activity could affect distribution and density on a local scale (Teilmann and Kapel 1998).

115
The reported annual catch of ringed seals in Greenland averaged 82,421 (SD = 4,697) during 1995‐2006 (Greenland Home Rule 2009). About half of the total number of sealskins taken from harp, hooded, and ringed seals in Greenland during 2000‐2006 were sold to tanneries each year, with the other half presumably being used for private purposes. Thus, the seal hunt in Greenland is both subsistence‐oriented and a commercial activity (Greenland Home Rule 2009). Currently, there are no national laws providing protection for ringed seals in Greenland or catch quotas regulating their harvest, however, sealing is regulated in a wildlife sanctuary in Melville Bay and a national park in northeast Greenland, and most municipalities restrict the area, season, or method of seal hunting through bylaws (Greenland Home Rule 2009). Greenlandic citizens may hunt seals year‐round provided they have either a commercial or leisure time hunting permit and report their catch to the government each year (Greenland Home Rule 2009).
4.2.2.1.1.4 Svalbard and Norway
Large‐scale commercial harvesting of marine mammals began in Svalbard shortly after its official discovery in 1596, however, ringed seals did not appear to play an important role in these early harvests since hunters preferred larger prey, such as bearded seals and walruses (Lydersen 1998). Since 1946, Norwegian hunting records indicated that ringed seal catches varied between 0 (several years) and a maximum of 745 per year (in 1973), Those records included kills from areas outside of Svalbard, however, and likely overestimate the harvests for Svalbard alone (Lydersen 1998). Currently, ringed seals are protected in Svalbard during the breeding season (March 20 to May 20), but licensed hunters can take seals at other times of the year outside of the national parks and nature reserves (Kovacs 2007). Systematic harvest records are not kept in Svalbard, but Lydersen (1998) estimated that “some few hundred seals are taken annually, mainly for dog food.” In mainland Norway, ringed seals are hunted for sport, and the harvest is regulated by quotas and licensing with only a few hundred seals taken each year (Kovacs 2007).
4.2.2.1.1.5 Russian Federation
Ringed seals were harvested in the Russian Arctic as long as 3,000‐5,000 years ago (Krupnik 1993), but they do not appear to have been subjected to intensive sealing in the region as were harp seals and Pacific walruses (Chapskii 1940, Belikov and Boltunov 1998). Hunting of ringed seals in the western Russian Arctic was thoroughly reviewed by Belikov and Boltunov (1998), based mainly on catch data presented by Nazarenko (1969), Timoshenko (1984), and an anonymous (1994) author. According to Nazarenko (1969), about 3,200 ringed seals were harvested in the Barents Sea in 1894 and 4,512 ringed seals were harvested in the White Sea in 1903. The annual harvest of ringed seals in the White Sea averaged about 7,500 during 1905‐1909; 6,200 during 1910‐1915; and 6,400 during 1928‐1931 (Nazarenko 1969). The combined catches of ringed seals from the White and Barents Seas was 12,600 in 1933 and averaged about 5,600 per year during 1954‐1965, while the harvest of ringed seals in the Kara Sea was 13,200 in 1933 and averaged about 2,900 per year during 1960‐1962 (Nazarenko 1969). Those numbers only reflect the commercial harvest conducted by the fishing industry and procurement companies, which Nazarenko (1969) believed could be increased by 10‐15% to account for data missing from areas that did not respond to his inquiries. Nazarenko (1969) did not assess the subsistence

116
harvest by local people, but he stated that individual hunters may have taken up to 200 seals per year for their own needs.
Timoshenko (1984 cited in Belikov and Boltunov 1998) compared data from the White, Barents, and Kara Seas during the 1960s and 1970s and found a decrease in ringed seal harvests in all areas. The average annual harvest of ringed seals declined from about 3,150 during 1960‐1975 to about 1,350 during 1976‐1978 in the White Sea, and declined from about 4,600 during the 1960s to 2,000 during 1970‐1978 for the combined Barents and Kara Sea catches. The declines were believed to result from reduced effort by the sealing industry associated with falling prices rather than a decline in seal numbers (Timoshenko 1984 cited in Belikov and Boltunov 1998). Popov (1982) believed that most stocks of ringed seals in the Russian Arctic were “underexploited” and stated that regular, intensive sealing only occurred in the White Sea, where an annual harvest quota of 3,500 ringed seals was established. The average annual catch of ringed seals dropped to about 800 in the White Sea during 1985‐1994 and dropped to less than 400 in the Barents and Kara Seas combined during 1985‐1992 (Anonymous 1994 cited in Belikov and Boltunov 1998). The declines were believed to have resulted from a closure of most of the fish‐processing plants and procurement companies that made up the sealing industry, due to deteriorating economic conditions in the northern regions of Russia as well as an underestimate of the true seal catch (Belikov and Boltunov 1998). The reported harvests only reflect commercial takes and do not include subsistence takes.
The Soviet sealing fleet conducted large‐scale commercial harvests in the Bering Sea during 1961‐1969, and about 5,100 ringed seals (or 6,400 accounting for hunting losses) were killed each year (Fedoseev 2000). Large‐scale ship‐based harvests of ringed and bearded seals have been prohibited in the Bering Sea since 1969 (Popov 1982, Fedoseev 2000), but small‐scale ship‐based harvests of these species continued. Fedoseev (2000) stated that the annual ship‐based harvest did not exceed 3,300 ringed seals (4,100 accounting for losses) during 1970‐1990, while Mineev (1981, 1984) reported that this harvest ranged between 287 and 766 ringed seals per year, with an average of 471 during 1979‐1983.
Historically, the largest harvest of ringed seals from the Russian portions of the Bering and Chukchi Seas came from shore‐based hunting by native peoples. Heptner et al. (1976, citing P.G. Nikulin as their source) reported that the annual harvest of ringed seals by Russian Natives living along the coast of the Chukchi Sea averaged 25,000 during the late 1930s; 23,500 during the 1940s; and 15,500 during the late 1950s. Popov (1982) stated that the shore‐based catch in the Bering Sea was between 30,000 and 35,000 ringed seals per year during the “postwar years” but that the harvest decreased to 10,000‐12,000 ringed seals annually “in later years,” mainly because there were fewer hunters. Fedoseev (1984) reported an annual shore‐based harvest of 40,000 ringed seals along the coasts of the Bering, Chukchi, and East Siberian Seas during 1940‐1954. The shore‐based harvest in the Bering Sea has been restricted to low levels (i.e., 2,000‐3,000 ringed seals per year) since 1970 (Popov 1982). Fedoseev (1984) stated that the shore‐based harvest from the Bering and Chukchi Seas was not more than 3,000‐4,000 ringed seals per year in the early 1980s, and Mineev (1981, 1984) reported that this harvest ranged between 991 and 3,607 ringed seals per year with an average of 2,405 during 1979‐1983. Fedoseev (2000) also stated that the shore‐based harvest in the Bering and Chukchi Seas did not exceed 7,000‐8,000 ringed

117
seals per year during 1970‐2000. The decline in the shore‐based harvest during the mid‐20th century was attributed to the movement of native peoples from small coastal camps to larger settlements and a subsequent change in their social traditions (Fedoseev 1984).
Currently, the Russian Federation manages marine mammal harvests in its waters using a system of TACs. Scientific institutions review biological data every year and establish a TAC quota for each species by region (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 4 May 2010). Local fisheries management agencies issue permits to hunters (for both commercial and subsistence takes) which restrict the species, season, location, and method of hunting, and hunters are required to keep a record of their harvest. Ringed seal TACs in the western Bering Sea and along the eastern coast of the Kamchatka Peninsula ranged between 5,900 and 6,500 per year during 2002‐2005 (Marine Mammal Council 2008) and were set at 1,900 in 2008, the most recent year for which TACs are available. Ringed seal TACs were set at 200 in the Laptev and East Siberian Seas and 5,200 in the Chukchi Sea and Chukotka Zone in 2008. The current ringed seal harvest in Russia is conducted mostly along the coast of the Arctic Ocean. It is not regulated well, and hunters in rural areas often ignore permit requirements, take seals as needed, and do not report their catches, so the official catch records do not reflect the actual harvest levels well. The current harvest level, however, is believed to be “quite low” and probably does not affect the population (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 4 May 2010). Hovelsrud et al. (2008) stated that ringed seals are the most commonly hunted seal in the Chukotka region of northeastern Russia and considered the marine mammal harvest there to be “small‐scale” and “sustainable.”
Current harvest levels of ringed seals in the western Russian Arctic are not available but, based on recent TACs, are likely low. During 2002‐2005, TACs for ringed seals ranged from 1,000 to 2,500 in the Barents Sea and from 1,000 to 1,300 in the White Sea; TACs were apparently not established for ringed seals in the Kara Sea during 2002‐2004, but a quota of 1,500 was set there in 2005 (Marine Mammal Council 2008). In 2008, ringed seal TACs were set at 100 in the Barents and Kara Seas and 500 in the White Sea (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 10 May 2010).
4.2.2.1.2 Sea of Okhotsk
Soviet sealing statistics reported from the Sea of Okhotsk vary considerably from source to source, but by all accounts, ringed seals were overexploited there during the mid‐20th century. Seals have been hunted by native peoples along the coasts of the Sea of Okhotsk for centuries (Krylov et al. 1964), and prior to the 1950s, the subsistence hunters’ annual take did not exceed 25,000‐35,000 seals of which about 50% were ringed seals (Fedoseev 2000). Popov (1982) reported that not more than 25,000‐30,000 ringed seals were taken each year during the same period and that the harvest did not reduce the population. Ship‐based sealing expeditions were first conducted in the Sea of Okhotsk in 1932 (Krylov et al. 1964), and during the years following World War II to the early 1950s, an average of 6,000 ringed seals were hunted annually, apparently a sustainable harvest (Heptner et al. 1976). The ship‐based commercial harvest increased substantially in 1954 as the sealing fleet grew in size, skill, and intensity (Krylov et al. 1964, Heptner et al. 1976), and hunting was unregulated until 1969 (Fedoseev 2000). The

118
commercial harvest in the Sea of Okhotsk ranged between 38,938 and 81,710 (Avg. = 60,413) ringed seals per year during 1955‐1968 (Fedoseev (1984, Table 1). Later, Fedoseev (2000) put the average catch during the same period at 78,500 ringed seals per year, and Popov (1982) put this figure at 72,000 ringed seals per year. Seals that were mortally wounded but escaped into the water or that sank before they could be retrieved were not accounted for, and Fedoseev (2000) estimated that an additional 25‐30% of the numbers harvested were killed but lost. The large numbers killed reduced the ringed seal population in the Sea of Okhotsk and altered the natural age structure of the populations (Krylov et al. 1964, Fedoseev 1984).
An annual harvest limit of 32,000 ringed seals was imposed on sealing ships in the Sea of Okhotsk in 1969 and reduced to 25,000 in 1972 and 18,000 in 1975 (Popov 1982). As a result, the harvest of ringed seals decreased from 32,742 in 1969 to 25,000 in 1974; no data were presented for 1975 (Fedoseev 1984, Table 1). It is not known whether harvest limits were further reduced, but the reported harvest dropped to an average of 4,834 ringed seals per year during 1976‐1980 (Fedoseev 1984, Table 1). A catch restriction of 7,000 ringed seals per year was also imposed on the shore‐based harvest in the Sea of Okhotsk in 1975 (Popov 1982). Harvest limits or levels during the 1980s are unknown, but Fedoseev (2000) stated that the commercial catch was “strongly limited” in those years and minimally impacted the population. Grachev (2006) reported that an average of 12,664 ringed seals were harvested by ships in the Sea of Okhotsk during 1990‐1994 and proposed that 12,000 ringed seals could be safely harvested each year and their organs used to develop new medical and pharmaceutical industries in Russia. That proposal does not appear to have been implemented.
Large‐scale ship‐based harvests ended in the Sea of Okhotsk in 1994 (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 20 August 2008). TACs set by the Russian Federation allowed for 20,100‐22,600 ringed seals to be harvested annually from the Sea of Okhotsk during 2002‐2005 (Marine Mammal Council 2008), however, the actual harvest (including four seal species) was less than 1,000 seals per year during 1995‐2005 (Grachev 2006). The ringed seal TAC in the Sea of Okhotsk was lowered to 3,500 in 2008 (the most recent year for which TACs are available), and the current harvest level is believed to be much lower than 1,000 ringed seals per year (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 9 May 2010).
Little information is available regarding ringed seal hunting near Hokkaido, Japan in the southern Sea of Okhotsk. Yoshikazu (2006) reported that the Japanese conducted “industrial exploitation” of seals following World War II that continued to 1977. The highest annual catch was estimated to exceed 10,000 seals in 1950, but harvest numbers were not reported by species or location. Naito (1971) reported that a small‐scale seal hunt was conducted around Hokkaido by fishermen during their off‐season. Based on data collected from fur factories, Naito (1971) estimated that about 190 ringed seals were harvested in 1968, which represented less than 10% of the total seal harvest. The catch was thought to be similar in 1969 and 1970 (Naito 1971). Seal hunting was unregulated in Japan until 2002, when revision of the “Wildlife Protection and Appropriate Hunting Law” restricted the capture or killing of seals without proper authorization (Ministry of Environment and Hokkaido Prefectural Government

119
2007; Y. Mitani, Hokkaido University, pers. comm., 14 September 2009). Current harvest levels of ringed seals around Hokkaido are unknown but likely are low.
4.2.2.1.3 Baltic Sea
Bounty hunters reduced Baltic ringed seals numbers from an estimated few hundred thousand in 1900 to about 6,000‐10,000 by the 1980s (Almkvist 1982, Helle 1986). The population reduction was accompanied by a decrease in the average age of sexual maturity from 4.7 to 2.7 years (Söderberg 1978). During the early 1900s, more than 20,000 ringed seals were killed annually in Finnish and Swedish waters (Helle and Stenman 1990 cited in Reijnders et al. 1993). The kill dropped to about 6,000 ringed seals per year during 1924‐1939 and less than 2,000 during 1956‐1975 (Bergman 1956 cited in Härkönen et al. 1998, Söderberg 1975 cited in Härkönen et al. 1998, Helle 1979b). During 1933‐1970, about 650 ringed seals were taken each year in Estonian waters (Tormosov and Rezvov 1978). Several seal hunting regulations were enacted in the Baltic region beginning in the 1970s (Söderberg 1973, Stenman 1977, Popov 1980, Ministry of Agriculture and Forestry 2007), and all killing of Baltic ringed seals was banned in the Soviet Union in 1980, in Sweden in 1986, and in Finland in 1988 (Härkönen et al. 1998). Gray seal hunting was restarted in Finland in 1998 and in Sweden in 2001 but is strictly regulated in both countries (NAMMCO 2004, Ministry of Agriculture and Forestry 2007). Beginning in 2010, Finland is permitting harvest of a small number of ringed seals (initially 30) in the Bothnian Bay. Illegal killing of gray, ringed, and harbor seals still occurs in parts of the Baltic, but reliable estimates of that take are not available (M. Kunnasranta, Finnish Game and Fisheries Research Institute, pers. comm., 9 July 2010). Poaching was not thought to constitute a significant threat to seal populations in Finland in recent years (Ministry of Agriculture and Forestry 2007).
4.2.2.1.4 Lake Ladoga
Ringed seals experienced significant hunting pressure in Lake Ladoga during the early part of the 20th century which likely caused their numbers to dramatically decline (Sipilä and Hyvärinen 1998). During 1909‐1918, 436 to 1,278 ringed seals were harvested annually in Lake Ladoga (Heptner et al. 1976), and about 950 per year were killed in the northern part of the lake during 1924‐1939 (Jääskeläinen 1942 cited in Sipilä and Hyvärinen 1998). In the 1970s, about 100 seals were hunted annually in the southern part of the lake (Tormosov and Filatov 1979 cited in Sipilä et al. 2002). Bounty statistics indicated that 500 to 1,000 ringed seals were harvested annually during 1950‐1974, with an estimated total of 10,000‐20,000 seals taken during that period (Verevkin et al. 2006, Agafonova et al. 2007). Hunting of Lake Ladoga ringed seals was prohibited by law in either 1975 (Filatov 1990 cited in Sipilä et al. 2002, Verevkin et al. 2006) or 1980 (Sipilä and Hyvärinen 1998). Fishermen reportedly still kill some seals every year (Sipilä and Hyvärinen 1998) but exact numbers are unknown.
4.2.2.1.5 Lake Saimaa
Ringed seals in Lake Saimaa were considered pests during the early part of the 20th century due to their perceived impact on the fishery, and a bounty was paid for killing them from 1882 to 1948 (Metsähallitus 2009). According to bounty statistics, 246 ringed seals were killed during 1893‐1905 (Kokko et al. 1998), 140 seals were shot in the middle part of the lake during 1909‐1918 (Kilkki and

120
Marttinen 1984 cited in Sipilä and Hyvärinen 1998), and about 278 were killed during 1928‐1947 (Kokko et al. 1998). During the 1950s, ringed seals were eradicated from some parts of the lake by fishermen (Sipilä and Hyvärinen 1998). In 1955, Lake Saimaa ringed seals were protected from hunting by law because their numbers had become so small (Sipilä and Hyvärinen 1998, Wilson et al. 2001). Since that time, only two cases of poaching in the early 1980s have been reported (Sipilä 2003).
4.2.2.1.6 Future threat due to commercial, subsistence, or illegal harvest
Climate change is likely to alter existing subsistence harvest patterns of marine mammals by changing species’ local densities or distributions in relation to hunting communities (ACIA 2005, Hovelsrud et al. 2008). For example, if changes in sea ice lead to decreased densities near hunting communities, seal populations might increase, but if seal densities increased around hunting communities, local populations might be depleted (Mansfield 1970 cited in Reeves et al. 1998, Teilmann and Kapel 1998). Vibe (1967) correlated temperature records with numbers of ringed seals harvested in Greenland from the early 1800s to the mid‐1960s. When the local climate warmed and sea ice was reduced in West Greenland (e.g., before 1810 and around 1930), ringed seal catches were low, but when the climate cooled (e.g., 1810‐1860) and ice stagnated in the northern regions of West Greenland, ringed seal harvests rose considerably. The impacts of climate change on subsistence hunting will be complicated by the complexity of interacting variables. Nonetheless, many of the consequences of climate change are likely to be negative for marine mammals as well as for marine mammal hunters (Hovelsrud et al. 2008).
Huntington (2009) qualitatively assessed the threats facing Arctic marine mammals and concluded that hunting poses only a modest threat because it “is well understood, is likely to have a low impact, and is amenable to effective conservation through hunting management, which can respond to change relatively quickly”, making it likely “the most readily managed human activity affecting marine mammals in the Arctic.” He also suggested that co‐management regimes, in which hunters and wildlife managers together take into account the population biology of the species and the cultural and economic dimensions of the hunters, are the most effective means of mitigating the risks posed by harvesting (Huntington 2009). Accurate information on both harvest levels and the species’ population size and trends are needed, however, in order to assess the impacts of hunting as well as to respond appropriately to potential future changes in population levels due to climate change (ACIA 2005, Hovelsrud et al. 2008, Huntington 2009).
4.2.2.2 Scientific and educational utilization
Ringed seals have occasionally been collected for zoos and aquariums (e.g., one is currently held at SeaWorld San Diego and two are at the Okhotsk Tokkari Center, Japan; C. Reichmuth, University of California Santa Cruz, pers. comm., 22 June 2010) or killed for scientific research (e.g., Krafft et al. 2006b). Total numbers of ringed seals used for scientific or educational purposes are small and likely have no impact on the populations of any of the subspecies.

121
4.2.3 Diseases, Parasites, and Predation
4.2.3.1 Diseases
4.2.3.1.1 Protozoa
Neospora caninum, Sarcocystic neurona, and Toxoplasma gondii are protozoan parasites that can cause encephalitis and can be fatal in several species of domestic and wild animals (Van Pelt and Dieterich 1973, Migaki et al. 1977, Holshuh et al. 1985, Lapointe et al. 1998, Cole et al. 2000, Miller et al. 2001, Dubey et al. 2003). Serum antibody prevalences of those protozoans have been examined in several species of marine mammals (Lapointe et al. 1998, Dubey et al. 2003, Fujii et al. 2007). Although 48 ringed seals tested negative for T. gondii antibodies in the North Atlantic (Oksanen et al. 1998), 5 of 32 ringed seals tested in Alaska showed evidence of T. gondii exposure and four were positive for antibodies to N. caninum (Dubey et al. 2003). More recently, 49 of 262 ringed seals tested in Svalbard between 1992 and 2008 tested positive for antibodies to T. gondii (Jensen et al. 2010). The prevalence of T. gondii in ringed seals increased significantly over the past decade.
Jensen et al. (2010) suggested that seals are exposed to protozoan oocysts primarily through their prey, especially filter‐feeding species. Ringed seals from Svalbard are known to ingest filter feeders (Labansen et al. 2007). The two main sources of postnatal T. gondii infection are ingestion of oocysts in contaminated food or water and ingestion of T. gondii‐infected tissues (Dubey et al. 2003, Fujii et al. 2007). Felids and dogs are the only known hosts that can excrete oocysts of T. gondii and N. caninum, respectively (Dubey et al. 2003, Fujii et al. 2007). Oocysts from both species may be washed into the ocean in runoff contaminated by excrement (Fujii et al. 2007). Areas of high freshwater outflow along the California coast have been associated with T. gondii positive sea otters (Miller et al. 2002). T. gondii oocysts are extremely resistant to environmental influences and are viable even after several months of exposure to seawater temperatures as low as 4°C (Dubey et al. 2003, Lindsay et al. 2003). Jensen et al. (2010) suggested a number of possible explanations for the increased prevalence of T. gondii antibodies in ringed seals off of Svalbard. A significant increase in ship‐traffic over the past decade may be introducing vectors through ballast water discharge and oocysts through dumping of biodegradable waste. Oocysts also may be transported from southern latitudes by the North Atlantic Current. A recent increase in sea water temperatures has brought temperate species into the area which may serve as vectors to a potential influx of oocysts. It is unknown how the increase in sea temperature may affect the survivability of transported oocysts.
4.2.3.1.2 Viruses
Phocine distemper virus (PDV), and canine distemper virus (CDV), both of the genus morbillivirus, have caused large die‐offs of Baikal seals (Phoca sibirica) in 1987‐1988 (Grachev et al. 1989), harbor seals and gray seals in the North and Baltic Seas in 1988 (Kennedy et al. 1988, Osterhaus et al. 1990), and Caspian seals (Phoca caspica) in 1997 and 2000 (Kennedy et al. 2000, Barrett et al. 2003). The 1988 die off in northern Europe was repeated on a greater scale in 2002 when about 50% of the harbor seals along the European continent died, whereas harbor seals around the British Isles were less effected (Härkönen et al. 2006). In both events, mortalities were more frequent among harbor seals than gray seals (Barrett et

122
al. 2003). Seals infected with morbillivirus show lesions in the lungs, central nervous system, lymphatic tissue, intestine, and liver resulting in pneumonia, encephalitis, compromised immune function, reduced coordination, and muscle tremors (reviewed in Heide‐Jørgensen et al. 1992a, Kennedy 1998, Kennedy‐Stoskopf 2001). Many infected seals had additional bacterial infections, were exposed to biotoxins, or were in areas with high pollutant levels, contributing to immunosupression and complicating the interpretation of clinical signs (Ross et al. 1996, Kennedy‐Stoskopf 2001, Troisi et al. 2001). Few ringed seals in Canada and Greenland tested positive for PDV neutralizing antibodies between 1972 and 1988 (reviewed in Heide‐Jørgensen et al. 1992a). Forty‐one percent of ringed seals sampled between 1992 and 1994 from seven locations in the Canadian Arctic tested positive for exposure to PDV and CDV (Duignan et al. 1997). Duignan et al. (1997) also showed PDV antibody prevalence was significantly higher in ringed seals whose range overlapped with harp seals. Harp seals have a high prevalence (83%) of PDV antibodies and may have been a vector for the northern European die‐offs of 1988 and 2002 (Heide‐Jørgensen et al. 1992a, Duignan et al. 1997, Barrett et al. 2003). Terrestrial and marine Arctic species (e.g., polar bears and Arctic foxes) show serological evidence of PVD and CVD exposure, and predators and scavengers may contribute to the spread of morbillivirus (Cattet et al. 2004, Tryland et al. 2005a). Aggregations of ringed seals sharing breathing holes may facilitate contact with other species and opportunities to transfer infection, which could then be dispersed as far west as the Bering Sea (Duignan et al. 1997). No evidence of morbillivirus was detected in 60 ringed seals tested in the Bering Sea in the 1980s (Osterhaus et al. 1988), and serologic surveys prior to 2000 indicated that Pacific marine mammals had not been exposed to PDV. Recently, however, PVD was detected in Northern sea otters (Enhydra lutris kenyoni) off of Alaska (Goldstein et al. 2009). Sequence analysis confirmed the viral fragment was identical to the 2002 phocine distemper that caused the outbreak in northern Europe, raising the possibility that the virus was transmitted through the Arctic to the North Pacific after the 2002 European epidemic. Ranges of circumpolar seal species—such as bearded and ringed seals—overlap with subarctic species such as ribbon and spotted seals. Although PDV vector species are largely unknown, Arctic and subarctic migrating seals have been suggested to be carriers of PDV in the Atlantic. Reduced sea‐ice cover may alter seal migration patterns and contact between species further spreading the virus. Thus, ringed seals may be both susceptible to and vectors for PDV in northern seas.
Herpesvirus infections have been found in several marine mammal species from the Northern Hemisphere (Kennedy‐Stoskopf et al. 1986, Harris et al. 1990, Zarnke et al. 1997). Herpesviruses have been associated with both fatal and nonfatal infections of harbor seals from the north Pacific, central and northern California, and the Netherlands (Borst et al. 1986, Spraker et al. 1994, Gulland et al. 1997). Exposures to two phocid herpesviruses have been detected in phocid seals in Alaska. Phocid herpesvirus‐1 (PhHV‐1) is an alpha herpesvirus, related to both canine herpesvirus and felid herpesvirus, and phocid herpesvirus‐2 (PhHV‐2) is a gamma herpesvirus (Osterhaus et al. 1985, Harder et al. 1996, Zarnke et al. 1997). PhHV‐1 caused the death of 11 harbor seal pups in a nursery in the Netherlands and has caused disease in other pinnipeds with clinical signs that include pneumonia, adrenocortical necrosis, and hepatic necrosis (Osterhaus et al. 1985, Borst et al. 1986, Kennedy‐Stoskopf et al. 1986, Gulland et al. 1997). PhHV‐1 is highly contagious and transmitted through aerosols or direct contact (Zarnke et al. 1997). It is unknown how PhHV‐2, the gamma herpesvirus, is transmitted, and there is no evidence that PhHV‐2 causes clinical disease in pinnipeds (Zarnke et al. 1997). Zarnke et al. (1997) tested

123
marine mammals from Alaska and Russia for antibodies to PhHV‐1 and PhHV‐2. They examined walrus, two species of otariids, and five species of phocids, including four ringed seals. In ringed seals, serum antibody prevalence for PhHV‐1 and PhHV‐2 were both 50%, and antibody prevalence for neither virus was 25%. Antibody prevalence for PhHV‐1 was higher than for PhHV‐2 in most of the species examined, and the highest prevalence of antibodies to PhHV‐1 was found in phocid seals. Zarnke et al. (1997) suggested that serum antibody prevalences found in this study indicate that marine mammals off the coasts of Alaska and Russia are regularly exposed to PhHV‐1 and PhHV‐2 and possibly to other related herpesviruses.
A combination of lesions found in intestine, kidneys, adrenals, bone, blood vessels, and uteruses of Baltic gray and ringed seals have been described as the Baltic seal disease complex (Bergman and Olsson 1985). Similar lesions were not found in Arctic ringed seals sampled in Svalbard, and there is strong evidence that the disease complex is associated with local levels of PCBs and related contaminants (Olsson et al. 1994, Bergman et al. 2001). The uterine lesions cause infertility, abortion, and fetal resorption. The subsequent reduction in reproductive rates in the Gulf of Bothnia may have also been linked to contaminants (Helle et al. 1976a, Helle et al. 1976b). Many of the pathological changes have occurred less frequently after PCB and DDT contaminant levels began declining in the 1970s (Bergman 1999, Jüssi et al. 2005).
Some caliciviruses are known to have a marine origin and are able to spread and cause disease in both marine and terrestrial species, including humans (Barlough et al. 1987, Smith et al. 1998). Well‐known examples include vesicular exanthema of swine and San Miguel sea lion virus (Smith et al. 1998). Barlough et al. (1987) sought to determine whether Tillamook calicivirus (TCV), which infects bovines, has a marine origin. Blood samples from several species of marine mammals from Pacific populations, including 1 ringed seal, were tested for the presence of serum neutralizing antibodies to TCV (Barlough et al. 1987). Spotted, ribbon, and bearded seals, a few non‐ice associated phocids, Northern fur seals (Callorhinus ursinus), Pacific walruses, and a few cetaceans were all negative for antibodies to TCV (Barlough et al. 1987). California sea lions (Zalophus californianus), Steller sea lions (Eumetopias jubatus), Northern fur seals, and walruses have all tested positive for antibodies, consistent with the observation that most caliciviruses isolated from marine mammals have been from the subfamily Otariinae (Barlough et al. 1987, Smith et al. 1998).
4.2.3.1.3 Bacteria
Brucella is a bacterium known to cause reproductive failure in domestic animals (Godfroid 2002) and suspected of causing abortions in marine mammals (Ewalt et al. 1994, Miller et al. 1999, Rhyan et al. 2001). The first evidence of Brucella exposure in ringed seals was reported in 1996 (Nielsen et al. 1996) when anti‐Brucella antibodies were detected in 4% of 248 ringed seals sampled from the Canadian Arctic between 1992 and 1994. Evidence of Brucella exposure was also detected in 10% of apparently healthy ringed seals collected from Svalbard and Franz Joseph Land between 1992 and 1995 (Tryland et al. 1999). Brucellosis in ringed seals was documented in ringed seals near Baffin Island infected with a novel Brucella sp. (Forbes et al. 2000). More recent collections from the Svalbard area failed to find

124
Brucella sp. in ringed seals (Tryland et al. 2005b). No evidence of reproductive failure in ringed seals exposed to Brucella has been reported.
Infections with the bacterium, Leptrospira, are known to cause renal disease and death in harbor seals (Stamper et al. 1998, Kik et al. 2006), California sea lions (Gulland et al. 1996), and Northern fur seals (Smith et al. 1977). There is one report of a bearded seal testing positive for exposure to Leptrospira in the Bering Sea (Calle et al. 2008), but no evidence of exposure in ringed seals.
4.2.3.2 Parasites
Helminth parasites—including cestodes, trematodes, nematodes, and acanthocephalan worms—have been found in ringed seals from all parts of their range (Table 3) (Popov 1975, Heptner et al. 1976, Shults 1977, Fay et al. 1979, Eley 1981, Shults 1982, Delyamure et al. 1984, Takahashi 1999).
Dipetalonema spirocauda, a filaroid nematode that infects the cardiovascular system in pinnipeds, has been identified in ringed seals (Popov 1975, Delyamure et al. 1980, Measures et al. 1997). Eley (1981) examined hearts and pulmonary arteries from marine mammals in Alaskan waters to determine the prevalence and distribution of D. spirocauda. The parasite was identified in spotted, ringed, ribbon, bearded, and harbor seals and most often in the right ventrical and also the pulmonary artery. The parasite was more common in harbor seals (17%), which are not associated with sea ice, than in any of the four species of ice‐associated seals, where occurrence was 4‐8% (Eley 1981). Measures et al. (1997) also found these roundworms in 14 of 221 ringed seals sampled from six sites in the Canadian Arctic, with the majority of infected seals less than 1 year old. D. spirocauda were not found in eight other Arctic marine mammal species (Eley 1981). Delyamure et al. (1980) reported D. spirocauda infections in four emaciated seals of Lake Ladoga.
Parafilaroides articus and P. krascheninnikovi were discovered in the lung of ringed seals in the Chukchi Sea (Delyamure and Alekseev 1966, Yurakhno and Skryabin 1971). P. krascheninnikovi were found in the bronchi, bronchiole, and alveoli of ringed seals in the Chukchi (16 of 60 seals) and Bering (15 of 118) Seas (Yurakhno and Skryabin 1971). Lungworms found in Baltic ringed seals were associated with local hemorrhaging where P. gymnurus were embedded in lung tissue (Delyamure et al. 1980). Otostrongylus circumlitus has been described in ringed seals from Amundsen Gulf and the Canadian Arctic (Onderka 1989, Bergeron et al. 1997). P. gymnurus causes damage to airways and inflammation and, because infection appears to be limited to young‐of‐the‐year, the parasite could cause population level effects (Onderka 1989, Bergeron et al. 1997). Parafilaroides hispidus was more prevalent (53%) in all age classes, but no significant lesions were observed in the alveoli (Onderka 1989). Although Bergeron et al. (1997) found O. circumlitus to be prevalent (48% of 190 seals) in the eastern Canadian Arctic, Gosselin et al. (1998) did not find it in 31 seals from the Northwest Territories. The ringed seal was a new host for Parafilaroides gymnurus, and gravid female worms (Parafilaroides spp.) were more prevalent in ringed seals than in other ice‐associated seals. Infected seals were not in poor body condition (Gosselin et al. 1998). Parafilaroides hispidus was described from a ringed seal from the Beaufort Sea (Kennedy 1986) and identified in two ringed seals from the Canadian Arctic (Gosselin et al. 1998).
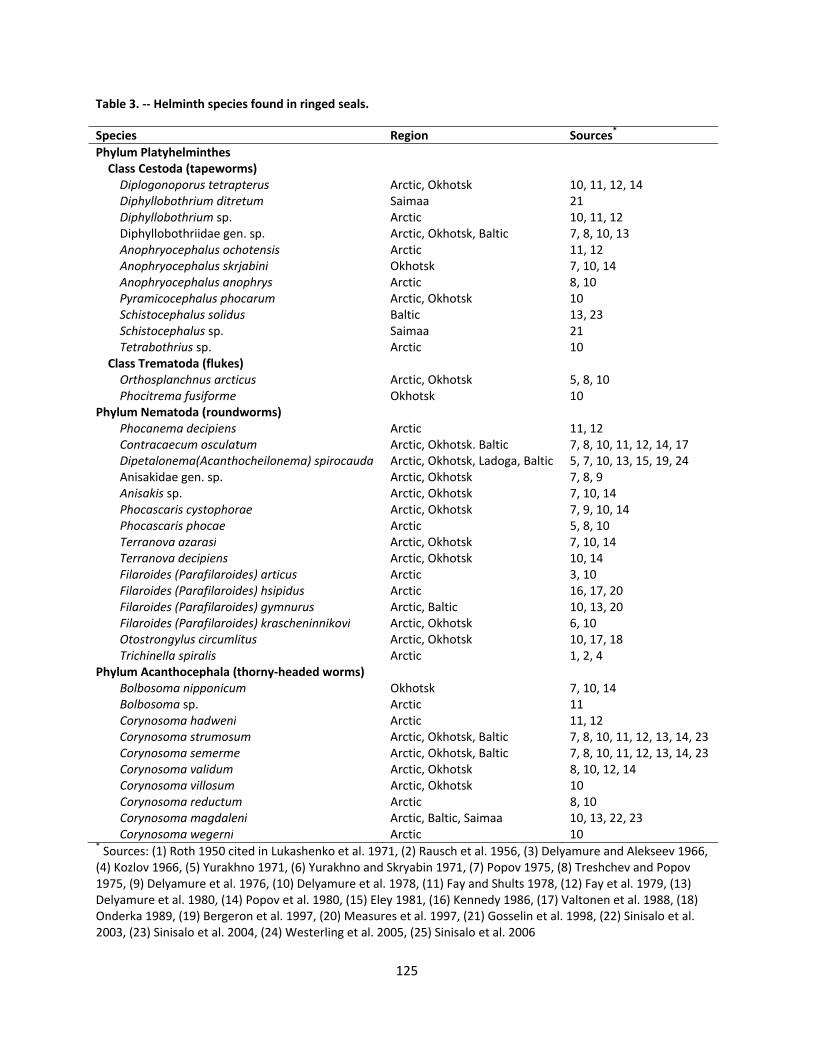
125
Table 3. ‐‐ Helminth species found in ringed seals.
Species Region Sources* Phylum Platyhelminthes Class Cestoda (tapeworms)
Diplogonoporus tetrapterus Arctic, Okhotsk 10, 11, 12, 14 Diphyllobothrium ditretum Saimaa 21 Diphyllobothrium sp. Arctic 10, 11, 12 Diphyllobothriidae gen. sp. Arctic, Okhotsk, Baltic 7, 8, 10, 13 Anophryocephalus ochotensis Arctic 11, 12 Anophryocephalus skrjabini Okhotsk 7, 10, 14 Anophryocephalus anophrys Arctic 8, 10 Pyramicocephalus phocarum Arctic, Okhotsk 10 Schistocephalus solidus Baltic 13, 23 Schistocephalus sp. Saimaa 21 Tetrabothrius sp. Arctic 10
Class Trematoda (flukes) Orthosplanchnus arcticus Arctic, Okhotsk 5, 8, 10 Phocitrema fusiforme Okhotsk 10
Phylum Nematoda (roundworms) Phocanema decipiens Arctic 11, 12 Contracaecum osculatum Arctic, Okhotsk. Baltic 7, 8, 10, 11, 12, 14, 17Dipetalonema(Acanthocheilonema) spirocauda Arctic, Okhotsk, Ladoga, Baltic 5, 7, 10, 13, 15, 19, 24Anisakidae gen. sp. Arctic, Okhotsk 7, 8, 9 Anisakis sp. Arctic, Okhotsk 7, 10, 14 Phocascaris cystophorae Arctic, Okhotsk 7, 9, 10, 14 Phocascaris phocae Arctic 5, 8, 10 Terranova azarasi Arctic, Okhotsk 7, 10, 14 Terranova decipiens Arctic, Okhotsk 10, 14 Filaroides (Parafilaroides) articus Arctic 3, 10 Filaroides (Parafilaroides) hsipidus Arctic 16, 17, 20 Filaroides (Parafilaroides) gymnurus Arctic, Baltic 10, 13, 20 Filaroides (Parafilaroides) krascheninnikovi Arctic, Okhotsk 6, 10 Otostrongylus circumlitus Arctic, Okhotsk 10, 17, 18 Trichinella spiralis Arctic 1, 2, 4
Phylum Acanthocephala (thorny‐headed worms)Bolbosoma nipponicum Okhotsk 7, 10, 14 Bolbosoma sp. Arctic 11 Corynosoma hadweni Arctic 11, 12 Corynosoma strumosum Arctic, Okhotsk, Baltic 7, 8, 10, 11, 12, 13, 14, 23Corynosoma semerme Arctic, Okhotsk, Baltic 7, 8, 10, 11, 12, 13, 14, 23Corynosoma validum Arctic, Okhotsk 8, 10, 12, 14 Corynosoma villosum Arctic, Okhotsk 10 Corynosoma reductum Arctic 8, 10 Corynosoma magdaleni Arctic, Baltic, Saimaa 10, 13, 22, 23 Corynosoma wegerni Arctic 10
* Sources: (1) Roth 1950 cited in Lukashenko et al. 1971, (2) Rausch et al. 1956, (3) Delyamure and Alekseev 1966, (4) Kozlov 1966, (5) Yurakhno 1971, (6) Yurakhno and Skryabin 1971, (7) Popov 1975, (8) Treshchev and Popov 1975, (9) Delyamure et al. 1976, (10) Delyamure et al. 1978, (11) Fay and Shults 1978, (12) Fay et al. 1979, (13) Delyamure et al. 1980, (14) Popov et al. 1980, (15) Eley 1981, (16) Kennedy 1986, (17) Valtonen et al. 1988, (18) Onderka 1989, (19) Bergeron et al. 1997, (20) Measures et al. 1997, (21) Gosselin et al. 1998, (22) Sinisalo et al. 2003, (23) Sinisalo et al. 2004, (24) Westerling et al. 2005, (25) Sinisalo et al. 2006

126
Intestinal parasites have been found in all subspecies of ringed seals (Popov 1975, Treshchev and Popov 1975, Delyamure et al. 1978, Fay et al. 1979, Popov et al. 1980, Olson et al. 1997, Sinisalo et al. 2003, Sinisalo et al. 2004). Helminth species have also been found in the liver, gall bladder, and pancreas (Treshchev and Popov 1975, Fay and Shults 1978). Baltic, Saimaa, and Ladoga ringed seals are host to a much more restrictive helminth fauna (Delyamure et al. 1980). Corynosoma magdaleni is the only acanthocephalan species found in Saimaa seals (Sinisalo et al. 2003). The intensity of C. magdaleni infection was postively correlated with age of the seal, possibly due to food items available only to the deeper diving older animals (Sinisalo et al. 2003). A positive relationship between nematode load and the age of ice‐associated seals has been observed elsewhere as well (Delyamure et al. 1980). C. magdaleni was also found in Baltic ringed seals but not in 18 Ladoga seals (Delyamure et al. 1980). Two species of cestodes (tapeworms of the family Diphyllobothriidae) were found in Saimaa and Ladoga seals (Delyamure et al. 1980, Sinisalo et al. 2003). In the Barents Sea, 154 ringed seals all carried intestinal parasites with the greatest numbers in the small intestines (Treshchev and Popov 1975).
Focal hepatitis and hepatic abscesses have been found in ringed seals infested with Orthosplanchnus fraterculus off the Alaska coast (Fay and Shults 1978). O. arcticus is encountered more often than other trematodes (Heptner et al. 1976) and is found in the liver, gall bladder, and—most frequently—pancreas of ringed seals in the Barents Sea (Treshchev and Popov 1975). Larval Trichinella spiralis has been reported in ringed seal muscle tissue collected off Alaska (Rausch et al. 1956), Greenland (Roth 1950), and in the Chukchi Sea (Kozlov 1966), although it was not found in 178 ringed seals from the Bering and Chukchi Seas (Lukashenko et al. 1971) or in 252 seals from Svalbard (Larsen and Kjos‐Hanssen 1983).
Heptner et al. (1976) stated that only one species of lice, Echinophthirius horridus, has been documented infesting ringed seals. Fay et al. (1979) determined that E. horridus were only found rarely on bearded, ringed, and ribbon seals (1‐5 lice per seal). Nasal mites (Acarina: Halarachnidae) are parasites that inhabit the respiratory tract—primarily the nasal passages—of mammals infrequently including bearded, ringed, ribbon, and spotted seals (Fay et al. 1979, Fay and Furman 1982). Fay and Furman (1982) examined nasal passages from marine mammals collected in the eastern Bering, eastern Chukchi, and western Beaufort Seas. They did not find nasal mites in the 43 ringed seals examined but suggested that the species is rarely infected. (Fay et al. 1979) reported that 4 of 202 seals—including ringed, ribbon, bearded, and spotted seals—were infected with nasal mites, and all four were spotted seals.
4.2.3.3 Predation
Ringed seal predators include polar bears, brown bears (Ursus arctos), Arctic foxes, red foxes (Vulpes vulpes), gray wolves (Canis lupus), lynx (Lynx lynx), European mink (Mustela lutreola), walruses, killer whales, Greenland sharks, common ravens (Corvus corax), and glaucous gulls (Burns and Eley 1976, Heptner et al. 1976, Fay et al. 1990, Sipilä 2003, Melnikov and Zagrebin 2005). Ringed seals and bearded seals are the primary prey of polar bears (Heptner et al. 1976, Derocher et al. 2004).
Polar bears prey heavily on ringed seals but with regional and temporal variation. In the Beaufort Sea, ringed seals comprised 98% of the polar bears’ diet. In Western Hudson Bay, ringed seals accounted for

127
80% of the polar bears’ diet in the early 1990s, but later in that decade, as ice break‐up came earlier, the proportion declined while the proportion of bearded and harbor seals in the diet increased (Stirling and Parkinson 2006). Adult polar bears need to consume approximately 2 kg (4.4 lbs) of fat per day to survive (Best 1985), and Stirling and Øritsland (1995) calculated that a hypothetical polar bear population containing 1,800 bears would need approximately 77,400‐80,293 ringed seals per year. Kingsley (1998) estimated that the polar bears in Baffin Bay and associated waters (N = ca. 4,000) would need to eat 120,000 to 160,000 ringed seals per year to sustain themselves.
Polar bear predation on ringed seals is most successful in moving, offshore ice (Stirling et al. 1975, Stirling and Archibald 1977), often along floe edges and rarely in ice‐free waters (Furnell and Oolooyuk 1980). Polar bears also successfully hunt ringed seals on stable shorefast ice by catching animals when they surface to breathe and by pouncing on lairs and pinning their prey (Stirling and Latour 1978, Smith 1980). The snow cover at birth lairs typically is deeper than at lairs used only by adult seals (Smith 1980, Lydersen and Gjertz 1984, Kelly et al. 1986, Lydersen 1998), and bears often cannot collapse the roofs of birth lairs. In those instances, bears may excavate a hole in the snow above the lair and wait with their body pressed into the hole for a seal to surface (Smith 1980). Smith (1980) proposed that multiple lairs—thought to be maintained by pupping females—and the distribution of lairs might be adaptive responses to lessen the likelihood of polar bear predation. The distribution of lairs also is limited by snow depth, and the distribution of lairs reflects the distribution of pressure ridges and ice hummocks that form snow drifts (Kelly and Quakenbush 1990, Sipilä and Hyvärinen 1998). Hammill and Smith (1991) found that polar bear predation increased significantly with increased density of lairs and that bear predation on pups tripled when pups were prematurely exposed as a consequence of unseasonably warm conditions. Overall, they found that bears removed between 8 and 44% of the annual pup production. Hammill and Smith (1991) further noted that polar bear predation on ringed seal pups increased four‐fold when average snow depths in their study area decreased from 23 cm to 10 cm. They concluded that while a high proportion of pups born each year are lost to predation, “without the protection provided by the subnivean lair, pup mortality would be much higher.”
The distribution of Arctic foxes broadly overlaps with that of Arctic ringed seals (Hersteinsson and MacDonald 1992), and some foxes spend over 40% of the year on the ice (Pamperin et al. 2008). Arctic foxes prey on newborn seals by tunneling in to the birth lairs (Kumlien 1879, Degerbøl and Freuchen 1935). Smith (1976) reported that Arctic foxes were the most frequent cause of death among ringed seals in the first year of life in nearshore areas of the western Canadian Arctic with 9‐40% of the pups killed by foxes. Elsewhere, the percentages of lairs entered by Arctic foxes were 21% (Svalbard) and 13% (Alaskan Beaufort Sea), and the percentages of pups killed when birth lairs were entered in those locations were 38% and 25%, respectively (Lydersen and Gjertz 1984, Kelly et al. 1986, Kelly and Quakenbush 1990). Smith (1976) suggested that patterns of lair use by ringed seals were adaptations, in part, to fox predation. The range of Arctic foxes does not coincide with that of the Okhotsk, Baltic, Saimaa, or Ladoga subspecies of ringed seals. Red foxes, however, do overlap with those subspecies and, on rare occasions, prey on newborn ringed seals in lairs (Andriashek and Spencer 1989).

128
Terrestrial predators can readily access shorefast sea‐ice habitats, and other carnivores also are occasional predators. Sipilä (2003) noted that the brown bear is a potential predator of the Lake Saimaa ringed seal, though no specific records were identified. In Lake Ladoga, ringed seal lairs have been marked and penetrated by wolves, dogs, and red foxes (Kunnasranta 2001). In Lake Saimaa, ringed seal pups have been killed in their lairs by canids—either dogs or foxes (Sipilä 2003). The wolf, lynx, and mink are also described as potential land predators of the Lake Saimaa ringed seal (Kunnasranta 2001, Sipilä 2003). Tunnels thought to have been made by mink penetrated seal lairs, and a mink has been observed eating a pup in a lair (Kunnasranta 2001). Okhotsk ringed seals are largely restricted to pack ice where terrestrial predators are rare. The scarcity of predators probably is important in the Okhotsk ringed seal’s ability to raise young on ice with insufficient snow cover for lair formation.
Marine predation is only partially limited by the ringed seal’s occupation of heavy ice cover. Lowry and Fay (1984) presented evidence of walrus predation on ringed, spotted, and bearded seals in the Bering and Chukchi Seas. They found ringed seal remains in 25 of 364 walrus stomachs from the Bering Sea and in 5 of 44 walrus stomachs from the Chukchi Sea. In the Gulf of Anadyr, hunters reported that ringed seal numbers declined with the arrival of walruses each year, but specific observations of predation were not noted (Fedoseev 1965a). Killer whales likely prey on ringed seals in open water or along ice margins, but the whales are limited by their inability to penetrate far in to the ice. Greenland sharks are not limited by ice cover and are known to prey on ringed seals, but the frequency of such predation is unknown. The Greenland shark and killer whale are considered important predators of the ringed seal in the Barents Sea (Heptner et al. 1976).
Gulls, ravens, and possibly snowy owls prey on ringed seal pups when the latter are forced out of subnivean lairs prematurely because of low snow accumulation and/or early melts (Kumlien 1879, Gjertz and Lydersen 1983, Lydersen and Gjertz 1987, Lydersen et al. 1987, Lydersen and Smith 1989, Lydersen and Ryg 1990, Lydersen 1998). Avian predation is facilitated not only by lack of sufficient snow cover but also by conditions favoring influxes of birds. Lydersen and Smith (1989) noted that the proximity of open water at Svalbard exposed unconcealed ringed seal pups to predation by glaucous gulls. Gulls and ravens have been increasing along the Arctic coast of Alaska as a result of habitat changes associated with industrial activities (Johnson and Herter 1989, Truett et al. 1997, Johnson and Noel 2005). Lydersen and Smith (1989) pointed out that the small size of newborn ringed seals, coupled with their prolonged nursing period, makes them vulnerable to predation by birds and likely sets a southern limit to their distribution. That is, the southern limit corresponds to the latitudes at which snow conditions are suitable for excavating and occupying subnivean lairs. Similarly, unseasonal melting of snow dens is believed to threaten polar bears, especially in the southern portions of their range (Clarkson and Irish 1991, Stirling and Derocher 1993).
4.2.3.3.1 Future considerations: climate change
Snow cover provides protection from cold and predators for ringed seals. The small pups (about 4 kg) are protected from predation and cold exposure by occupying subnivean lairs when born through the first 6‐9 weeks of their lives. High rates of predation by polar bears, Arctic foxes, gulls, ravens, and other predators have been associated with anomalous weather events that caused the lairs to collapse or melt

129
before ringed seal pups were weaned. Ringed seals abandon lairs as the snow warms in the spring. In the Beaufort Sea, lair abandonment began progressively earlier each year from 1999 to 2003 (Kelly et al. 2003). The snow cover failed especially early in 2002, and by mid‐May of that year, all of the seals had abandoned lairs. Increased juvenile mortality among ringed seals likely will result if the snow continues to melt early.
4.2.4 Inadequacy of Existing Regulatory Mechanisms
The ESA requires that a listing determination be based solely on the best scientific and commercial data available after conducting a review of the status of the species and after taking into account any efforts being made to protect the species. The consideration of protective efforts requires an evaluation of the probability that those efforts will be implemented and effective. Because such an evaluation requires inherently non‐scientific, sociological, and political considerations—such as the likelihood of future funding and future local agency involvement—the BRT did not include threats stemming from inadequacy or lack of existing regulatory or protective efforts in its threats assessment. The human activities that are the subject of any such efforts, however, are included in the suite of threats considered under the other four ESA Section 4(a)(1) factors (Sections 4.2.1, 4.2.2, 4.2.3, and 4.2.5). Therefore, a summary of regulatory mechanisms relevant to ringed seal conservation is presented below for perspective on the related threats.
4.2.4.1 Existing conservation efforts
4.2.4.1.1 International agreements and oversight
4.2.4.1.1.1 The Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) treaty
The CITES treaty is aimed at protecting species at risk from unregulated international trade. CITES regulates international trade in animals and plants by listing species in one of three appendices. The level of monitoring and control to which an animal or plant species is subject depends on the appendix in which the species is listed. Appendix I includes species threatened with extinction which are—or may be—affected by trade. Trade of Appendix I species is only allowed in exceptional circumstances. Appendix II includes species not necessarily threatened with extinction presently, but for which trade must be regulated in order to avoid utilization incompatible with their survival. Appendix III includes species that are subject to regulation in at least one country and for which that country has asked other CITES Party countries for assistance in controlling and monitoring international trade in that species. Ringed seals have no special status under CITES.
4.2.4.1.1.2 The International Union for Conservation the of Nature and Natural Resources (IUCN) Red List of Threatened Species
The IUCN Red List of Threatened Species identifies and documents those species most in need of conservation attention if global extinction rates are to be reduced and is widely recognized as the most comprehensive, apolitical, global approach for evaluating the conservation status of plant and animal species. In order to produce Red Lists of threatened species worldwide, the IUCN Species Survival

130
Commission draws on a network of scientists and partner organizations, which use a scientifically standardized approach to determine species’ risks of extinction. The ringed seal is currently classified as a species of “Least Concern” on the IUCN Red List, due to the large population size and broad distribution of the Arctic subspecies (Kovacs et al. 2008). Kovacs et al. (2008) noted that given the risks posed to the ringed seal by climate change, the conservation status of all ringed seal subspecies should be reassessed within a decade. The ringed seal was chosen by the IUCN Species Survival Commission as one of 10 climate change flagship species to illustrate the impacts of climate change on polar habitats, including the effects of ice loss on ice‐adapted species (IUCN Species Survival Commission 2009).
The European Red list compiles assessments of the conservation status of European species according to IUCN regional red listing guidelines. The Saimaa seal is currently classified as an “Endangered” species and the Ladoga seal is classified as a “Vulnerable” species on the list (European Mammal Assessment Team 2007). The Baltic ringed seal is classified as a species of “Least Concern” on the European Red List with the qualifications that population numbers remain low and that there are significant conservation concerns regarding ringed seals in some parts of the Baltic (European Mammal Assessment Team 2007).
4.2.4.1.1.3 The Council of Europe’s Bern Convention on the Conservation of European Wildlife and Habitats
The Bern Convention is a regional European treaty on conservation of wild flora and fauna and their natural habitats. Current parties to the Bern Convention within the range of the ringed seal include Norway, Sweden, Finland, Estonia, and Latvia (the Russian Federation participates as an observer). The agreement calls for signatories to provide special protection for fauna species listed in Appendix II (species to be strictly protected) and Appendix III to the convention (species for which any exploitation is to be regulated). The Saimaa seal and Ladoga seal are listed under Appendix II, and other ringed seals fall under Appendix III.
4.2.4.1.1.4 Council of the European Union (EU) Directive 92/43/EEC on the Conservation of Natural Habitats and of Wild Fauna and Flora (EU Habitats Directive)
The provisions of the EU Habitats Directive are intended to promote the conservation of biodiversity in EU member countries. EU members meet the habitat conservation requirements of the directive by designating qualified sites for inclusion in a special conservation areas network known as Natura 2000. Current members of the EU within the range of the ringed seal include Sweden, Finland, and Estonia. Annex II to the Habitats Directive lists species whose conservation is to be specifically considered in designating special conservation areas, Annex IV identifies species determined to be in need of strict protection, and Annex V identifies species whose exploitation may require specific management measures to maintain favorable conservation status. The Saimaa seal is listed in Annex II (as a priority species) and IV, the Baltic ringed seal is listed in Annex II and V, and the Arctic ringed seal is listed in Annex V.

131
4.2.4.1.1.5 The Convention on the Protection of the Marine Environment of the Baltic Sea Area (Helsinki Convention)
The Helsinki Convention is a regional agreement, which was signed by the contracting parties in 1974 (entered force in 1980) and revised in 1992, that is aimed at controlling and limiting severe pollution affecting the Baltic Sea area. The Baltic Marine Environment Protection Commission (also known as HELCOM) is the governing body of the Helsinki Convention. Current parties to the convention include all of the countries bordering the Baltic Sea, along with the EU. As an international advisory body to Baltic Sea area countries, HELCOM provides information about environmental trends and the status of the Baltic Sea ecosystem and, based on that information, develops objectives and recommendations aimed at ecosystem restoration and protection. Countries that are parties to the convention are expected to implement HELCOM recommendations through national legislation and environmental programs.
HELCOM has adopted numerous recommendations that are aimed at controlling and minimizing land‐based pollution in the Baltic Sea area. Although appreciable decreases have been documented over the past 20 to 30 years in annual emissions of some hazardous substances, concentrations of some heavy metals and persistent organic pollutants (POPs) remain markedly higher in the Baltic Sea than in the North Atlantic (HELCOM 2007a). Contaminant levels in ringed seals have decreased somewhat since the 1970s but remain high (Ministry of Agriculture and Forestry 2007).
In 2007, HELCOM adopted the Baltic Sea Action Plan, which aims to reduce pollution and eutrophication in the Baltic Sea and to restore and conserve natural resources based on an ecosystem approach (HELCOM 2007a). As with other plans and recommendations adopted by HELCOM, effectiveness will depend on follow‐up actions by all HELCOM member countries. In 2009, the European Commission also adopted a strategy and related action plan to coordinate efforts among EU countries and other interested parties to address a number of Baltic Sea issues, including reducing pollution and eutrophication (Commission of the European Communities 2009).
Current joint international spill response plans for the Baltic Sea are detailed in the HELCOM Response Manual, and HELCOM member countries participate in annual multi‐country spill response exercises (HELCOM 2009). In addition, to address increasing risks of oil and hazardous materials spills due to increasing maritime traffic in the Baltic Sea, a 3‐year project was begun in 2009 to assess sub‐regional response capacities and to develop plans to address any deficiencies identified (HELCOM 2009).
HELCOM has designated a network of Baltic Sea Protected Areas (BSPAs) in member countries, some of which include Baltic ringed seal habitat (HELCOM 2007c). Most of the BSPAs receive some level of protection from national legislation; a number of these BSPAs are also designated as Natura 2000 or Emerald Network sites (an extension of Natura 2000 to include non‐member countries).
In 2006, HELCOM adopted Recommendation 27‐28/2 that calls for development and implementation of national management plans for Baltic seals, implementation of effective measures to reduce illegal killing and incidental bycatch by 2012, and organization of a HELCOM seal experts group. The seal experts group, which first convened in 2006, was tasked with coordinating development of national management plans for seals, identifying healthy seal population levels, developing and coordinating seal

132
monitoring programs, and identifying and establishing sites within the HELCOM BSPA Network that include important actual and potential seal habitats (HELCOM 2006b). In regions outside of Finnish parts of the Gulf of Bothnia and Aland Sea, the Baltic seal has been listed by HELCOM as a threatened and/or declining species in immediate need of protective measures (HELCOM 2007b).
4.2.4.1.1.6 Agreement on Cooperation in Research, Conservation, and Management of Marine Mammals in the North Atlantic (North Atlantic Marine Mammal Commission [NAMMCO])
The NAMMCO was established in 1992 by a regional agreement among the governments of Greenland, Iceland, Norway, and the Faroe Islands to cooperatively conserve and manage marine mammals in the North Atlantic. Canada and the Russian Federation participate in the NAMMCO as observers.
4.2.4.1.1.7 Regulation of the European Parliament and the Council of the EU on Trade in Seal Products (Regulation 1007/2009)
An EU regulation banning marketing and importation of commercial seal products was adopted in September 2009 by the European Parliament and the Council of the European Union in response to animal welfare concerns. Further measures are to be developed within 9 months to allow limited exceptions for products derived from subsistence harvest by indigenous communities. The governments of Canada and Norway have initiated the World Trade Organization’s dispute resolution process concerning the regulation (Lester 2010), and a lawsuit to overturn the regulation has been filed in the European Union General Court by a number of Inuit organizations (Inuit Tapiriit Kanatami 2010).
4.2.4.1.1.8 Mechanisms to limit sea‐ice reduction and ocean acidification by regulation of GHG emissions
There are no known regulatory mechanisms that effectively address changes to ringed seal habitat from reductions in sea ice or from ocean acidification. The primary international regulatory mechanisms addressing GHG emissions and global warming are the United Nations Framework Convention on Climate Change and the Kyoto Protocol. The Kyoto Protocol’s first commitment period, however, only sets targets for action through 2012. There are no regulatory mechanisms governing GHG emissions in the years beyond 2012. The United States is a signatory to the Kyoto Protocol but has not ratified it; therefore, the Kyoto Protocol is nonbinding on the United States.
4.2.4.1.1.9 Other agreements on contaminants
The Stockholm Convention on Persistent Organic Pollutants (SCPOP), which went into effect in 2004, is a global treaty to protect human health and the environment from POPs. Most of the countries within the range of the ringed seal have signed, but not all have ratified, the agreement. Twelve POPs were identified under the SCPOP to be banned or restricted, and recent amendments to the SCPOP added 9 other POPs. In addition, the Convention on Long‐Range Transboundary Air Pollution, which went into effect in 1983, has produced internationally legally binding protocols to reduce and control major air pollutants in signatory countries. Most European countries, the Russian Federation, Japan, Canada, and the United States have signed, but not all have ratified, the agreement and its related protocols. Levels of many of the 12 POPs originally targeted by the Stockholm Convention have declined in the Arctic

133
(AMAP 2009). Concentrations of some of these POPs, however, have remained high in some Arctic marine mammals and seabirds (AMAP 2009). In addition, there is evidence of Arctic accumulations of a number of compounds with POP characteristics that are currently in commercial use but are not yet subject to any international agreements.
In 2005, the International Maritime Organization (IMO) designated the Baltic Sea area outside of Russian territorial waters as a Particularly Sensitive Sea Area (PSSA), which provides a framework under the IMO’s International Convention for the Prevention of Pollution from Ships (MARPOL 73/78) for developing internationally agreed upon measures to reduce risks posed from maritime shipping activities. To date, a maritime traffic separation scheme is the sole protective measure associated with the Baltic Sea PSSA. Expansion of Russian oil terminals is contributing to a marked increase in oil transport in the Baltic Sea, however, the Russian Federation has declined to support the Baltic Sea PSSA designation (Kotiranta 2008).
4.2.4.1.2 Domestic regulatory mechanisms
4.2.4.1.2.1 United States
4.2.4.1.2.1.1 Endangered Species Act of 1973, as amended (ESA)
The ESA provides for the conservation of species that are endangered or threatened throughout all or a significant portion of their range and for the conservation of the ecosystems on which they depend. A "species" is considered endangered if it is in danger of extinction throughout all or a significant portion of its range and is considered threatened if it is likely to become an endangered species within the foreseeable future. The Saimaa seal was listed as endangered under the ESA in 1993 (National Marine Fisheries Service 1993).
4.2.4.1.2.1.2 Marine Mammal Protection Act of 1972, as amended (MMPA)
Ringed seals are protected in U.S. waters and on the high seas by the MMPA (16 U.S.C. 1361 et seq.). The MMPA was enacted in response to growing concerns among scientists and the general public that certain species and populations of marine mammals were in danger of extinction or depletion as a result of human activities. The MMPA set forth a national policy to prevent marine mammal species or population stocks from diminishing to the point where they are no longer a significant functioning element of the ecosystems.
The MMPA places an emphasis on habitat and ecosystem protection. The habitat and ecosystem goals set forth include: (1) management of marine mammals to ensure they do not cease to be a significant element of the ecosystem to which they are a part, (2) protection of essential habitats, including rookeries, mating grounds, and areas of similar significance “from the adverse effects of man's action”, (3) recognition that marine mammals “affect the balance of marine ecosystems in a manner that is important to other animals and animal products” and that marine mammals and their habitats should therefore be protected and conserved, and (4) directing that the primary objective of marine mammal management is to maintain “the health and stability of the marine ecosystem.” Congressional intent to protect marine mammal habitat is also reflected in the definitions section of the MMPA. The terms

134
“conservation” and “management” of marine mammals are specifically defined to include habitat acquisition and improvement.
The MMPA includes a general moratorium on the taking and importing of marine mammals, which is subject to a number of exceptions. Some of these exceptions include take for scientific purposes, public display, subsistence use by Alaska Natives, and unintentional incidental take coincident with conducting lawful activities. Take is defined in the MMPA to include the “harassment” of marine mammals. “Harassment” includes any act of pursuit, torment, or annoyance which “has the potential to injure a marine mammal or marine mammal stock in the wild” (Level A harassment), or “has the potential to disturb a marine mammal or marine mammal stock in the wild by causing disruption of behavioral patterns, including but not limited to, migration, breathing, nursing, breeding, feeding, or sheltering” (Level B harassment).
The Secretaries of Commerce and of the Interior have primary responsibility for implementing the MMPA. The Department of Commerce, through the National Oceanic and Atmospheric Administration (NOAA), has authority with respect to whales, porpoises, seals, and sea lions. The remaining marine mammals, including polar bears, walruses, sea otters, and manatees are managed by the Department of the Interior through the USFWS. Both agencies are responsible for the promulgation of regulations, the issuance of permits, the conduct of scientific research, and enforcement as necessary to carry out the purposes of the MMPA.
U.S. citizens who engage in a specified activity other than commercial fishing (which is specifically and separately addressed under the MMPA) within a specified geographical region may petition the Secretaries to authorize the incidental, but not intentional, taking of small numbers of marine mammals within that region for a period of not more than five consecutive years (16 U.S.C. 1371(a)(5)(A)). The Secretary “shall allow” the incidental taking if the Secretary finds that “the total of such taking during each 5 year (or less) period concerned will have a negligible impact on such species or stock and will not have an immitigable adverse impact on the availability of such species or stock for taking for subsistence uses.” If the Secretary makes the required findings, the Secretary also prescribes regulations that specify (1) permissible methods of taking, (2) means of affecting the least practicable adverse impact on the species and their habitat, and (3) requirements for monitoring and reporting. The regulatory process does not authorize the activities themselves, but authorizes the incidental take of the marine mammals in conjunction with otherwise legal activities described within the regulations.
Similar to promulgation of incidental take regulations, the MMPA also established a process by which citizens of the United States can apply for an authorization to incidentally take small numbers of marine mammals where the take will be limited to harassment (16 U.S.C. 1371(a)(5)(D)). These authorizations are limited to one‐year and, as with incidental take regulations, the Secretary must find that the total of such taking during the period will have a negligible impact on such species or stock and will not have an immitigable adverse impact on the availability of such species or stock for taking for subsistence uses. The Service refers to these authorizations as Incidental Harassment Authorizations.

135
Certain exceptions from the prohibitions on taking are provided. The MMPA exempts coastal‐dwelling Alaska Natives from the prohibitions on the taking of marine mammals, including ringed seals, for subsistence purposes. Sections 101(b)(3) and 103 of the MMPA provide for subsistence harvest regulations for marine mammal stocks designated as depleted under that Act, after notice and administrative hearings as prescribed by the MMPA. Section 119 of the MMPA allows the Secretary of Commerce to enter into cooperative agreements with Alaska Native organizations to conserve marine mammals and provide co‐management of subsistence uses.
4.2.4.1.2.1.3 National Environmental Policy Act (NEPA)
The NEPA requires federal agencies to consider the environmental impacts of their proposed actions and reasonable alternatives to those actions. To meet this requirement, federal agencies conduct environmental reviews, including environmental impact statements and environmental assessments. The NEPA does not itself regulate ringed seals, but it does require full evaluation and disclosure of information regarding the effects of contemplated federal actions on ringed seals and their habitat.
4.2.4.1.2.1.4 Outer Continental Shelf Lands Act (OCSLA)
The OCSLA (43 U.S.C. 331 et seq.) established federal jurisdiction over submerged lands on the outer continental shelf (OCS) seaward of the state boundaries (3‐mile limit) in order to expedite exploration and development of oil and gas resources on the OCS. Implementation of OCSLA is delegated to the Minerals Management Service (MMS, now known as the Bureau of Ocean Energy Management, Regulation and Enforcement) of the Department of the Interior. OCS projects that could adversely impact the coastal zone are subject to federal consistency requirements under terms of the Coastal Zone Management Act, as noted below. OCSLA also mandates that orderly development of OCS energy resources be balanced with protection of human, marine, and coastal environments. The OCSLA does not itself regulate the take of ringed seals, although through consistency determinations it helps to ensure that OCS projects do not adversely impact ringed seals or their habitats.
4.2.4.1.2.1.5 Coastal Zone Management Act (CZMA)
The CZMA (16 U.S.C. 1451 et seq.) was enacted to “preserve, protect, develop, and where possible, to restore or enhance the resources of the Nation's coastal zone.” The CZMA is a state program subject to federal approval. The CZMA requires that federal actions be conducted in a manner consistent with the state's coastal zone management plan to the maximal extent practicable. Federal agencies planning or authorizing an activity that affects any land or water use or natural resource of the coastal zone must provide a consistency determination to the appropriate state agency. The CZMA applies to ringed seal habitats of Alaska, although the CZMA does not itself regulate the take of ringed seals.
4.2.4.1.2.1.6 Marine Protection, Research and Sanctuaries Act (MPRSA)
The MPRSA (33 U.S.C. 1401 et seq.) was enacted in part to “prevent or strictly limit the dumping into ocean waters of any material that would adversely affect human health, welfare, or amenities, or the marine environment, ecological systems, or economic potentialities.” The MPRSA does not itself

136
regulate the take of ringed seals, although it operates to protect the quality of marine habitats that ringed seals rely upon.
4.2.4.1.2.1.7 Mechanisms to limit sea‐ice reduction and ocean acidification by regulation of GHG emissions
In 2003, the U.S. Environmental Protection Agency (EPA) rejected a petition urging it to regulate GHG emissions from automobiles under the Clean Air Act. In 2007, the U.S. Supreme Court overturned the EPA’s refusal to regulate these emissions and remanded the matter to the agency for further consideration (Supreme Court of the United States 2007). On 15 December 2009, the EPA published a final finding that six GHGs taken in combination endanger the public health and welfare of current and future generations, and that emissions of these GHGs from new vehicles contribute to the total GHG air pollution, and thus to this endangerment (U.S. Environmental Protection Agency 2009). The finding does not impose any emission reduction requirements but allowed the EPA to finalize GHG emissions standards for new light‐duty vehicles as part of joint rulemaking with the Department of Transportation (U.S. Environmental Protection Agency and U.S. Department of Transportation 2010).
Finalizing these new vehicle emission standards also had implications for regulation of GHGs from stationary sources. The EPA’s interpretation is that a newly regulated pollutant becomes subject to permit requirements of the Prevention of Significant Deterioration (PSD) and title V programs at the time regulatory requirements to controls emissions of that pollutant take effect (U.S. Environmental Protection Agency 2010a). Thus, GHG emissions from stationary sources become subject to the permitting requirements of these programs when the new vehicle emission standards take effect. On 13 May 2010, the EPA issued a final rule that limits the applicability of PSD and title V permitting requirements for GHG emissions to the largest emitters and excludes smaller stationary emission sources from these permitting requirements until at least 30 April 2016 (U.S. Environmental Protection Agency 2010b).
The BRT did not attempt to separate the risk posed by the lack of a regulatory mechanism for GHG emissions from the risks posed by the effects of those emissions. In Section 4.2.1, the risks posed by increased GHG emissions, via potential destruction or modification of ringed seal habitat, were assessed by evaluating the best available projections of future conditions under scenarios of no regulation of GHGs (the projections were based on “non‐mitigated” scenarios for future emissions). Therefore, the implications of the current lack of regulations are already included in the evaluation of risks to ringed seal habitat. In other words, while there are no regulatory mechanisms that effectively address reductions in sea‐ice habitat or ocean acidification, we do not expect this shortcoming to result in population‐level impacts beyond those already identified in the section on present or threatened destruction of habitat.
4.2.4.1.2.2 Canada
Hunting of seals in Canada is managed at the federal level by the DFO according to the Marine Mammal Regulations established under the Fisheries Act. In regions affected by native land claims agreements, the DFO co‐manages seal hunting with regional renewable resource boards in accordance with those

137
agreements (Department of Fisheries and Oceans 2007). Native subsistence hunting of ringed seals for food, social, or ceremonial purposes does not require a license (Department of Fisheries and Oceans 2008). A portion of seal skins harvested surplus to annual native subsistence needs is sold in international markets (Standing Committee on Fisheries and Oceans 2007). Hunting of seals by other residents for personal consumption is also permitted without a license north of 53°N latitude; a personal use sealing license, allowing the harvest of up to six seals (not species‐specific) annually for personal consumption, is available to non‐native coastal residents in Newfoundland, Labrador, and Quebec south of this latitude (Department of Fisheries and Oceans 2008). All other uses require a sealing license. Hunting of ringed seals is allowed year‐round outside of an approximately 5‐month annual closure period in Labrador (Department of Fisheries and Oceans 2008). Ringed seals are not currently harvested commercially in Canada (P. Hall, DFO, Canada, pers. comm., 6 May 2010). The Atlantic Seal Management Plan (Department of Fisheries and Oceans 2008) indicates that no TACs or allocations are set for ringed seals and that any commercial harvest of the species would be controlled through issuance of licenses and permits. Ringed seals are classified as “not at risk” under the Canadian Wildlife Species at Risk Act (Committee on the Status of Endangered Wildlife in Canada 2009).
4.2.4.1.2.3 Greenland
Harp and ringed seals are the primary species hunted for subsistence, recreational, and commercial purposes in Greenland. Licensed hunting of ringed seals is allowed year‐round with no established harvest quotas outside of Melville Bay Nature Sanctuary, Northeast Greenland National Park, and some municipalities (Greenland Home Rule 2009). Ringed seals are identified as a species of “Least Concern” on the Greenland Red List (Boertmann 2008).
4.2.4.1.2.4 Norway
Along the coast of Norway, where ringed seals are rare, harvest of ringed seals is managed under the Marine Resources Act (Directorate of Fisheries 2010; D. Vongraven, Norwegian Polar Institute, pers. comm., 2 July 2010). Small numbers of ringed seals are harvested there annually by licensed hunters without set quotas. In Svalbard territorial waters, harvest of ringed seals is managed under the Svalbard Environmental Protection Act within the framework that “productivity and diversity of species and their habitats are maintained and Svalbard’s natural wilderness is protected.” Small numbers of ringed seals are harvested annually by licensed hunters outside the major national parks and nature reserves that include about 86% of Svalbard’s territorial waters (Lier et al. 2010; I. Gjertz, Research Council of Norway, pers. comm., 23 June 2010).There are no set harvest quotas throughout the year except for a closed period during the breeding season (K. Kovacs, Norwegian Polar Institute, pers. comm., 22 April 2010; D. Vongraven, Norwegian Polar Institute, pers. comm., 2 July 2010). The ringed seal is not included on the Norwegian Red List (Kålås et al. 2006).
4.2.4.1.2.5 Russian Federation
Both subsistence and commercial hunting of ringed seals occurs in Russia. The Russian Federation government approves TACs that are assigned by species and geographical region (COWI 2008, Marine Mammal Council 2008). During 2008, annual TACs for the Western Bering Sea and Eastern Kamchatka

138
zone; Sea of Okhotsk; White, Barents, Kara, and Laptev Seas; and the Eastern Siberian Sea, Chukchi Sea, and Chukotka zone were set at 1,900, 3,500, 1,500, and 5,400 ringed seals, respectively (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 10 May 2010). Hunters are licensed; but, there are few restrictions on time or place of harvest outside of state nature reserves and refuges (Belikov and Boltunov 1998).
Harvest of ringed seals declined markedly across most of Russia after the mid‐1980s as commercial sealing became less economically viable, and large‐scale commercial harvest of ringed seals ended in the early 1990s (Belikov and Boltunov 1998, Grachev 2006, COWI 2008). Organized commercial harvest of seals has resumed on a small‐scale in some areas in the past few years (COWI 2008). Annual harvest of seals in the Sea of Okhotsk was reported as well below the TAC between 1995 and 2005 (Grachev 2006) and harvest levels are believed to have remained well below TACs in recent years (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 9 May 2010). It should be noted that harvest reporting and enforcement of catch quotas are difficult to manage in particular in remote areas of Russia (Kovacs et al. 2008). Arctic ringed seals are not listed in the Red Book of the Russian Federation (Danilov‐Danilyan 2001).
4.2.4.1.2.6 Japan
In Japan, licensing of hunters is overseen by local prefectures and allows for application to hunt wildlife for the purpose of damage prevention (Ministry of the Environment and Hokkaido Prefectural Government 2007, Ministry of the Environment 2009). The ringed seal is not included on the Red List of Threatened Mammals of Japan (Ministry of the Environment 2000).
4.2.4.1.2.7 Baltic States
In accordance with HELCOM Recommendation 9/1, hunting of Baltic ringed seals was suspended in all countries within the range of the seal in 1988, and harvest was discontinued before this time in several countries (Wilson et al. 2001). In Finland, the Baltic ringed seal is considered a game species that may be taken by licensed hunters during a designated hunting season, however, no hunting licenses have been issued since 1988 (Ministry of Agriculture and Forestry 2007). Seals found in fishing gear in the Baltic Sea of Finland can be kept by the owner without any required reporting (Ministry of Agriculture and Forestry 2007). There has been continuing pressure from the fishing industry to reestablish a ringed seal hunt in the Gulf of Bothnia (Ministry of Agriculture and Forestry 2007). Finland and Sweden have reported annual takes of a limited number of Baltic ringed seals for scientific sampling purposes, as well as removal of individual seals in response to fishery conflicts (ICES 2003). These countries also reported that a small number of Baltic ringed seals were mistakenly taken under permits issued for gray seals (ICES 2003).
Some protected areas designated in Sweden, Finland, the Russian Federation, and Estonia include Baltic ringed seal habitat (HELCOM 2001, HELCOM 2007c). The Baltic ringed seal is included in the Red Book of the Russian Federation as “category 2” (decreasing abundance), is classified as “endangered” in the Red Data Book of Estonia, and is listed as “near threatened” on the Finnish and Swedish Red Lists (Danilov‐

139
Danilyan 2001, Rassi et al. 2001, Jüssi et al. 2005, Commission for Nature Conservation 2008, ArtDatabanken 2010).
Hunting of ringed seals in Lake Ladoga has been prohibited since 1980 (Sipilä and Hyvärinen 1998). The number of seals lost due to entanglement in fishing nets is unknown but is believed to be a primary source of mortality for Ladoga seals (Verevkin et al. 2006). There are currently no fishing restrictions aimed at limiting seal mortalities from fishery interactions (Shibaeva 2009). Valaam Archipelago Nature Park, encompassing over 50 islands in the northwest portion of Lake Ladoga, includes terrestrial resting sites used by seals during open‐water periods. Increasing disturbance of resting seals and construction on remote islands within the park, however, have been reported (Shibaeva 2007, 2009). In May 2009, Ladoga Skerries National Park was added to the Russian Federation’s list of protected areas to be established (Shibaeva 2009). Ladoga Skerries National Park will encompass northern and northwest Lake Ladoga and will include seal pupping, molting, and resting areas. The Ladoga seal is included in the Red Data Books of the Russian Federation (as “category 3” [rare]) in the Leningrad and Karelia regions (Danilov‐Danilyan 2001, Jüssi et al. 2005).
The Saimaa seal is classified as a non‐game species and has been protected from hunting under Finnish law since 1955 (Wilson et al. 2001). The seal is also designated as an “Endangered” species on the Finnish Red List (Rassi et al. 2001). To conserve seal breeding areas, new construction is not permitted within designated shoreline conservation areas (water bodies excluded), some of which are located within two national parks (Wilson et al. 2001, Sipilä 2003). Sipilä (2003) estimated that 34% of birth lair sites occur within the national parks, and 70% of seal lair sites (birth and resting) are included in the shoreline conservation areas. New construction outside of designated shoreline conservation areas has been regulated since 1999 to limit the density of new buildings (Wilson et al. 2001), however, lakeshore development has still increased substantially (Sipilä 2003). To reduce pup mortalities that have occurred when lairs collapsed due to unstable ice conditions, artificial lowering of the water level of Lake Saimaa has been prohibited during the seal’s pupping season since 1991, and effects of natural reductions in water level have been reduced by artificial regulation of water in lakes that flow into Lake Saimaa (Sipilä and Hyvärinen 1998).
To reduce Saimaa seal mortalities in fisheries, restrictions have been placed on certain types of fishing gear within breeding areas since 1999 (Bell et al. 2008). Gill net fishing also has been closed from mid‐April to June annually through voluntary agreements between the Finnish government and local statutory fishing associations (Bell et al. 2008). Survival rates of weaned pups were about 10% higher in areas with seasonal gill net fishing closures than in areas without restriction (Sipilä 2003). Despite seasonal closure agreements with numerous fishing associations, seals—especially juveniles—continue to drown in fishing nets (ICES 2008). A single contiguous protected area within which all net fishing would be seasonally prohibited is under consideration along with other fishing restrictions (Ministry of Agriculture and Forestry 2010a). The Finnish Ministry of Agriculture and Forestry (2010b) has signed agreements with most of the Saimaa Lake fishing associations and is continuing to negotiate agreements with others. In May 2010, however, the European Commission put Finland on notice that it has failed to

140
implement adequate measures to protect the Saimaa seal and that better targeted measures are needed (European Commission 2010).
4.2.5 Other Natural or Human Factors Affecting the Species’ Continued Existence
4.2.5.1 Pollution and contaminants
Contaminant loads in ringed seals have been investigated in most parts of the species’ range (although not in the Sea of Okhotsk), reflecting the ringed seal’s importance in the diets of polar bears and coastal people (Dietz et al. 1995, Zhu et al. 1995, Muir et al. 1999, Fisk et al. 2001, Fisk et al. 2005, Kovacs 2007, Dietz 2008).
Pollutants such as organochlorine (OC) compounds and heavy metals have been found in all ringed seal populations (Kostareva et al. 1970, Smith and Armstrong 1978, Johansen et al. 1980, Helle 1981, Sipilä et al. 1996, Cameron et al. 1997, Dietz et al. 1998, Hyvärinen et al. 1998, Addison et al. 2000, Becker 2000, Bang et al. 2001, Dehn et al. 2005, Rigét et al. 2006, Kovacs 2007, Ashley et al. 2008, Dietz 2008, Quakenbush and Citta 2008, Routti et al. 2009, Sonne et al. 2009). The variety, sources, and transport mechanisms of the contaminants vary across the seal’s range (Addison et al. 2009). Muir et al. (1999) schematically described the pathways of transport, deposition, redistribution, and accumulation of persistent organics and metals to Arctic marine ecosystems. Many compounds are imbedded in the Arctic marine food chain (Wiberg et al. 2000, Kovacs 2007, Addison et al. 2009). Borgå et al. (2005) noted that OC contamination is greater in the European Arctic than in the Canadian or U.S. Arctic.
4.2.5.1.1 Heavy metals
Heavy metals such as mercury, cadmium, lead, selenium, arsenic, and nickel accumulate in ringed seal vital organs, including the liver and kidneys, as well as in their central nervous system (Koeman et al. 1975, Smith and Armstrong 1978, Wagemann 1985, Wagemann 1989, Wagemann et al. 1996, Atwell et al. 1998, Hyvärinen et al. 1998, Rigét et al. 2005, Quakenbush and Sheffield 2007, Gaden et al. 2009, Sonne et al. 2009).
4.2.5.1.1.1 The Arctic
Heavy metal burdens in Arctic ringed seals have been reported over the last several decades (Koeman et al. 1975, Smith and Armstrong 1978, Wagemann 1985, Wagemann 1989, Wagemann et al. 1996, Atwell et al. 1998, Dietz et al. 1998, Hyvärinen et al. 1998, Rigét et al. 2005, Kovacs 2007, Quakenbush and Sheffield 2007, Gaden et al. 2009, Sonne et al. 2009). Smith and Armstrong (1978) examined mercury and selenium levels in ringed seal tissues from seven locations across the Canadian Arctic. Their findings confirmed prior reports of high total mercury (THg) levels in liver and muscle, and they found no significant regional differences. Mercury and selenium were positively correlated with age, and the ratio of the two elements was linear (1:1). The authors did not detect any toxic effects of these naturally‐occurring elements and suggested that marine mammals have developed mechanisms (Koeman et al. 1975) to cope with the high level of mercury in their diet. Rigét et al. (2005) compared mercury and cadmium concentrations in ringed seal liver and kidneys from 11 locations across the Arctic, including

141
Alaska, Canada, Greenland, Svalbard, and the White Sea. They found that concentrations differed significantly among the studied locations. Ringed seals in the western Canadian Arctic had the highest concentrations of mercury in liver, while cadmium in liver was highest in the eastern Canadian Arctic and West Greenland. Concentrations in liver and kidney were also significantly higher in adult ringed seals as compared to subadults, and the circumpolar patterns were most pronounced in adult ringed seals. The authors suggested that the distribution of mercury and cadmium in ringed seals reflected natural and geological differences in the distribution of the metals. They noted that mercury and cadmium in the environment are derived from both natural and anthropogenic sources. Gaden et al. (2009) examined mercury levels in ringed seals from the western Canadian Arctic from 1973 to 2007. They detected no temporal trends in THg in muscle tissue, but a curvilinear relationship existed with the length of the ice‐free periods. THg levels were higher in both short (2‐month) and long (5‐month) ice‐free seasons. They suggested that during ice‐free periods the seals will eat more Arctic cod (and mercury). The authors also found that THg levels increased with age for both sexes, which is similar to Dehn et al.’s (2005) findings near Barrow, Alaska.
Dietz et al. (1998) reported on cadmium, mercury, zinc, and selenium levels in ringed seals from Greenland and Svalbard. Cadmium and mercury levels were associated with age and sampling locations. Mercury accumulated throughout life, whereas cadmium levels peaked in the 5‐ to 10‐year age groups, then declined significantly. Zinc concentrations in liver and kidney tissues showed some differences among sampling areas, but there was no correlation with age. Selenium levels exhibited the same patterns as cadmium and mercury. As reported in other studies, selenium and mercury levels were strongly correlated in the liver (1:1). Further, cadmium and mercury were correlated in all tissues, whereas cadmium and selenium and mercury and zinc were only correlated in kidneys and liver. High cadmium levels in adults from some regions were linked to their prey (e.g., Arctic cod). The authors also noted that average cadmium levels from Greenland were similar to levels reported by Wagemann et al. (1996) for the eastern Canadian Arctic; levels from western Greenland were much higher than levels reported for the Gulf of Bothnia in the Baltic Sea. Dietz et al. (1998) also summarized previous findings of mercury levels in Arctic ringed seals. They cited Zeisler et al.’s (1993) findings of high mercury levels in 1 and 2‐year old seals in Alaskan waters and noted that those levels corresponded to concentrations reported from western Canadian Arctic and eastern Greenland samples. Further, Dietz et al. (1998) noted that mercury levels in samples analyzed from the eastern Canadian Arctic, West Greenland, Svalbard, and northern Norway were three times lower (see Smith and Armstrong 1975, Smith and Armstrong 1978, Johansen et al. 1980, Carlberg and Bøler 1985, Wagemann 1989, Skaare et al. 1994, Wagemann et al. 1996). Sonne‐Hansen et al. (2002) investigated the effect on the skeletal system on the high cadmium concentrations in the kidneys of ringed seals from northwestern Greenland. Despite the high levels of cadmium, none of the seals exhibited signs of cadmium‐induced nephropathy or osteodystrophy. They noted that the ringed seals’ diet contains high levels of vitamin D, calcium, phosphorus, zinc, selenium, and protein, which would counteract the effect of cadmium.
4.2.5.1.1.2 Baltic Sea
Helle (1981) conducted the first study on heavy metal (mercury, cadmium, selenium, zinc, and chromium) levels in Baltic (Gulf of Bothnia) ringed seals. Mercury was the only element that differed

142
between sexes, with higher levels observed in females. Except for zinc, no trend with age was detected, but the author noted that a lack of samples from pups and subadults may have obscured age effects. Fant et al. (2001) compared mercury, cadmium, lead, and selenium levels in ringed seals from the Baltic and Svalbard. Mercury and selenium levels were significantly higher in the Baltic, whereas cadmium levels were lower. There was no geographic difference in lead levels. Compared to prior Baltic studies, metal levels have remained at 1980s levels. Similar to studies in other regions, mercury, cadmium, and selenium levels accumulated with age. Further, the authors did not detect any pathological changes that were associated with metal contamination. They also noted that information on the toxicological effects of metals was scarce.
4.2.5.1.1.3 Lake Saimaa
Hyvärinen and Sipilä (1984) compared nickel, mercury, cadmium, chromium, and lead concentrations in ringed seals across all age groups (still‐born pups to adults). Nickel levels were significantly higher in still‐born pups as compared to other age groups. Mercury concentration was significantly higher in still‐born pups, as compared to first‐year adults. No differences were detected in levels of cadmium, lead, and chromium between still‐born seals and older age groups. The authors noted that the high level of nickel in the hair of still‐born pups suggests it may have played a role in the high pup mortality seen in the region. Hyvärinen et al. (1998) monitored mercury concentrations in ringed seals from 1980 to 1995. They reported that mercury concentrations in livers of very young pups did not change significantly during the study period, whereas, mercury levels in weaned pups were significantly higher in 1981‐1983 than after 1984. Likewise, mercury levels in kidney and muscle tissues were significantly higher in 1981‐1983 than in 1990‐1995. In adult seals, however, there were no clear changes in mercury concentration over the time period. As in other regions, mercury levels increased with age. The levels detected in muscle and liver tissue were higher than those reported for the Baltic Sea and Arctic Ocean. Hyvärinen et al. (1998) suggested that the high levels may have contributed to the population decline in the 1960s and 1970s. They noted that the high level of mercury in the seals’ prey and shortage of selenium would reduce the seals capacity for metabolic detoxification. Sipilä (2003) noted that the major source of mercury in Lake Saimaa is from the pulp industry.
4.2.5.1.1.4 Lake Ladoga
Sipilä et al. (1996) could not attribute increases in cadmium and lead levels in Ladoga seals collected in the early 1990s to anthropogenic activities. Mercury concentration in kidney and liver tissues, however, were elevated and comparable to Saimaa seals. Heavy metal levels were determined for ringed seal tissues collected from 1990‐1993 in Lake Ladoga and the White Sea (Medvedev et al. 1997, Note: the findings of this study were also published by Ivanter et al. 1998). Mercury levels in Lake Ladoga samples were significantly higher than in the White Sea, and mercury levels in hair increased with age in both regions. Cadmium levels in kidney and liver tissues were significantly higher in Lake Ladoga samples, whereas, levels in hair and muscle were significantly higher in White Sea seals. Lead levels in liver and kidney tissues were similar in both regions, but levels in muscle and hair were significantly higher in the White Sea. Copper concentrations were similar in both regions. Nickel and zinc levels were significantly higher in Lake Ladoga seals. Medvedev et al. (1997) concluded that the level of heavy metals in Lake

143
Ladoga ringed seals was considerable and that the low levels observed in the White Sea could not be attributed to anthropogenic sources.
4.2.5.1.1.5 Sea of Okhotsk
There are no published records of heavy metal burdens in ringed seals in the Sea of Okhotsk. Based on the level of industrial activity in coastal areas, however, it seems likely that ringed seals in the region also carry high levels of toxic metals.
4.2.5.1.2 Organochlorine pollutants
OC pollutants, including compounds like polybrominated diphenyl ethers (PBDEs), DDT, and PCBs have been reported in ringed seals (Helle et al. 1976b, Olsson et al. 1986, Becker 2000, Nyman et al. 2002, Quakenbush and Sheffield 2007, Quakenbush 2007). OC contaminants are of particular concern because they are lipophilic compounds that have potential detrimental effects on health and reproduction (Helle et al. 1976a, O'Shea 1999, Aguilar et al. 2002, Gregory and Cyr 2003, Kovacs 2007). O’Shea (1999) summarized findings on the concentrations of DDT, PCB, and dichloro‐diphenyl‐dichloroethylene (DDE) in male and female ringed seals by geographic region (also see Addison and Smith 1974). In all regions, concentrations increased with age in males, whereas females reduced their contaminant levels via transfer to nursing pups. Senescent females do not show the same reduction in concentrations (see Fisk et al. 2002). Similar findings have been reported in subsequent reviews and studies (e.g., Cameron et al. 1997, Muir et al. 1999, Kucklick et al. 2002).
Less is known about the toxicity of flame retardants, but they are widely used in plastics, textiles, electronic equipment, and other materials (Rigét et al. 2006). PBDEs are ubiquitous in the environment. They are found in air, water, fish, birds, marine mammals, and humans, and levels have increased exponentially over the past 30 years (reviewed in Hites 2004). PBDEs adversely affect thyroid function and neurodevelopment in mammals (Darnerud 2003, Viberg et al. 2004).
4.2.5.1.2.1 The Arctic
Addison and Smith (1974) analyzed OC residues in blubber lipids from ringed seals collected in the Canadian Arctic (Northwest Territories). The levels of DDT, DDE, and PCBs were similar to previous Arctic studies, but well below levels found in Baltic ringed seals. Ringed seals collected from the Kara Sea in the Russian Arctic during 1995 (Nakata et al. 1998) contained higher levels of DDTs and PCBs than did seals from the Canadian Arctic and Norwegian Arctic. Nakata et al. (1998) noted that scant information was available on OC contamination in higher trophic animals in the Russian Arctic. Local areas were deemed to be the likely sources of the high contaminant levels. PCB levels in ringed seals from Svalbard were five times higher than in seals from the Canadian Arctic, consistent with the higher contaminant levels in the European Arctic (Wolkers et al. (1998a). Bang et al. (2001) noted, however, that OC levels in blood samples from Svalbard were similar to the North American Arctic, but lower than previous values reported for blubber. Kosmato et al. (2000a) compared DDT and PCB levels in tissues collected in Lake Saimaa, Lake Ladoga, and the Arctic (White Sea) and reported that the lowest levels were in the Arctic samples. Kucklick et al. (2002) measured concentrations of POPs in ringed seals from Barrow, Alaska and reported that they were similar to levels in the western Canadian Arctic. Addison et al. (2005) reported

144
that concentrations of some OCs in Arctic ringed seals have not changed between 1981 and 2000. Ashley et al. (2008) found a significant relationship between OC levels and sea‐ice trends in western Canadian Arctic ringed seals. During longer ice‐free seasons, seals are fatter because they have access to more prey, thus consuming more contaminants. Kovacs (2007) noted that the presence of POPs and other contaminants was inconsistent with the perception of the Arctic as one of the last pristine wilderness areas.
Rigét et al. (2006) examined temporal trends in PBDEs and PCBs in ringed seals from East Greenland. The total PBDE concentrations ranged from 21.8 (1986) to 39.3 (1999) ng g‐1 lipid weight. The levels observed in East Greenland were higher than reported from the Canadian Arctic (six locations), the Russian Arctic (three locations), Svalbard (one location), and West Greenland (one location) (Braune et al. 2005, de Wit et al. 2006). Sources of PBDEs in the Arctic include Western Europe, eastern North America, highly populated local areas, and southern regions through long‐range atmospheric transport (de Wit et al. 2006).
4.2.5.1.2.2 Baltic Sea
High levels of DDT and PCBs were found in Baltic (Bothnian Bay) ringed seals in the 1960s and 1970s (Helle et al. 1976a, Helle et al. 1976b), and PCB levels were correlated with reproductive failure. DDT and PCB levels were higher in females exhibiting pathological uterine occlusions. Tissue samples collected from age 1 and older seals in the early 1980s indicated that neither DDT nor PCB levels changed, but mean levels declined from the 1970s (Helle and Stenman 1984). Nyman et al. (2002) conducted a comparison of DDT and PCB levels reported by Helle et al. (1976b) and detected a 72‐85% reduction in total DDT levels, particularly in healthy females, followed by occluded females and males. Total PCB levels, however, only exhibited a minor decrease (25%) in females and no reduction in males.
4.2.5.1.2.3 Lake Saimaa
Helle et al. (1983) reported on OC levels (PCBs and DDT) in blubber samples collected in 1977‐1981. OC compound levels were higher than those reported in oceanic regions (see Wagemann and Muir 1981) but considerably lower than in the Baltic Sea. While DDT levels declined during the 1980s and 1990s (Helle et al. 1983, Sipilä and Hyvärinen 1998), PCB levels did not show a similar trend. Kosmato et al. (2000a) compared tissues collected in Lake Saimaa, Lake Ladoga, and the White Sea and reported that the highest concentrations of total PCBs and DDT were found in Lake Saimaa ringed seals. They also noted that while concentrations in Lake Saimaa seals were high, they were well below late 1960s and early 1970s levels.
4.2.5.1.2.4 Lake Ladoga
Kosmato et al. (2000a) reported that the mean concentrations of total PCBs and total DDT in ringed seals sampled in the early 1990s had not changed since findings published by Olsson et al. (1986) and were lower than those of Baltic and Lake Saimaa populations (Olsson et al. 1986, Kostamo et al. 2000a, Kostamo et al. 2000b, Sipilä et al. 2002).

145
4.2.5.1.2.5 Sea of Okhotsk
There are no published reports on OC contaminants in ringed seals from the Sea of Okhotsk. Based on the level of industrial activity in coastal areas, however, it seems likely that ringed seals in the region also carry high levels of OC contaminants.
4.2.5.1.3 Perfluorinated contaminants
Perfluorinated contaminants (PFCs)—such as perfluorooctane sulfonate (PFOS) and related synthetic compounds—are global pollutants (Bossi et al. 2005a, Quakenbush and Citta 2008) and are found in remote areas of the Arctic (Bossi et al. 2005b). PFCs are used in many industrial products (e.g., fire‐fighting foams, insecticide and herbicide production, lubricants, adhesives, paints; Kannan et al. 2001b). In marine mammals they bind to proteins rather than lipids and accumulate in the liver, kidney, and spleen with negative effects on cellular function (Van de Vijver et al. 2005).
4.2.5.1.3.1 The Arctic
PFCs and related synthetic compounds have been detected in ringed seals in the Alaskan Bering and Chukchi Seas (Quakenbush and Citta 2008), Canada (Martin et al. 2004, Butt et al. 2007), and Greenland (Bossi et al. 2005a). Bossi et al. (2005a) examined archived liver samples collected in East and West Greenland. They reported that PFOS was the major contributor to the burden of PFCs, followed by perfluoroundecanoic acid (PFUnA). Perfluorononanoic acid (PFNA) and perfluorodecanoic acid (PFDA) were also detected in most samples. Perfluorohexane sulfonate (PFHxS) and perfluorooctane sulfonamide (PFOSA) were only found sporadically. Perfluorooctanoic acid was not found in detectable concentrations in any sample. Concentrations of PFOS, PFDA, and PFUnA at both locations were increasing significantly. Quakenbush and Citta (2008) found lower levels of PFOS but higher levels of PFNA in the Alaskan Bering and Chukchi Seas, as compared to studies in Canada and Greenland. They also determined that PFCs do not appear to bioaccumulate with age in either male or female seals. Finally, levels of PFOS and PFNA appeared to be higher at lower latitude, possibly indicating that sources of PFOS and PFNA are closer to the Bering Sea than the Chukchi Sea. Kannan et al. (2001b) detected concentrations of PFOS in all blood samples collected in 1996 and 1998 from ringed seals in Svalbard. Concentrations of PFOS collected in 1998 were greater than those collected in 1996.
4.2.5.1.3.2 Baltic Sea
Kannan et al. (2001b) detected concentrations of PFOS in all blood samples collected in the northern Baltic (Bothnian Bay) from 1996 to 1998. PFOS levels were 15 times greater than those from the Arctic. Blood from a 4‐year‐old male ringed seal collected in 1998 contained the highest PFOS concentration (475 ng/ml). Kannan et al. (2002) also reported on PFOS levels in ringed seal livers. Concentrations were similar in males and females and did not increase with age. Levels were significantly greater than in samples from Baltic Sea gray seals.

146
4.2.5.1.3.3 Lake Saimaa, Lake Ladoga, and the Sea of Okhotsk
There are no published records of PFCs in ringed seals in Lake Saimaa, Lake Ladoga, or the Sea of Okhotsk. Based on the global distribution of these contaminants (Bossi et al. 2005a, Quakenbush and Citta 2008), however, it seems likely that ringed seals in these regions are burdened with PFCs.
4.2.5.1.4 Other contaminants
4.2.5.1.4.1 The Arctic
Butyltin (BT) compounds are used as antifouling agents in ship bottom paints and aquaculture nets (Iwata et al. 1997). They are retained in all tissues and largely in the liver rather than the blubber where PCBs and DDT accumulate (Iwata et al. 1997). BT compounds have been detected in marine mammal species in North Pacific, Asian, and California coastal waters (Iwata et al. 1997, Tanabe et al. 1998, Kannan et al. 2001a). Tanabe et al. (1998) reported that “BT accumulation in pinnipeds was lower than in cetaceans, confirming earlier notion that pinnipeds have greater capacity to degrade TBT [tributyltin] in the liver and excrete BTs through molting.” BT compounds have been found to suppress immune responses in rats (Snoeij et al. 1988). Berge et al. (2004) note that few studies on BT compounds have been conducted in Arctic waters. Based on analysis of liver, muscle, and blubber, they found low‐levels of BT contamination in ringed seals from northern Norwegian territories (Spitsbergen, Bear Island).
Cytochrome P450s (CYPs) are a class of hemoproteins induced by exposure to contaminants and pharmaceuticals. They are used as biomarkers for exposure to certain contaminants, including OCs (Assunção and Ross 2001, Fujita et al. 2001, Nyman et al. 2003). Elevated levels of CYP1A and CYP2A have been identified in Canadian Arctic, Baltic, and Svalbard ringed seals (Mattson et al. 1998, Wolkers et al. 1998a, Muir et al. 1999, Nyman et al. 2001, Nyman et al. 2003). Ringed seals off Svalbard demonstrated a clear negative relationship between enzymes involved in steroid breakdown (CYP3A) and PCB burdens (Wolkers et al. 1998b, Kovacs 2007).
Radiocaesium levels in Arctic seals have been evaluated in the Barents Sea, Svalbard, and North Greenland Sea (Carroll et al. 2002, Andersen et al. 2006). The authors note that in those regions seals are continuously exposed to low level radioactive contamination. This contaminant accumulates in soft tissues, particularly muscle, and it cannot be broken down in the body. Andersen et al. (2006) noted that their findings are consistent with previous studies in the Barents Sea, Svalbard, and Alaska (Cooper et al. 2000, Carroll et al. 2002) indicating low levels of radiocaesium in Arctic seals.
4.2.5.1.4.2 Baltic Sea, Lake Saimaa, Lake Ladoga, and the Sea of Okhotsk
There are no published records concerning BT contaminants, cytochrome P450s, or radiocaesium levels in ringed seals in the Baltic Sea, Lake Saimaa, Lake Ladoga, or the Sea of Okhotsk.
4.2.5.1.5 Future considerations
De Wit et al. (2006) predicted that OC levels surrounding the Arctic will continue to rise. Tynan and DeMaster (1997) noted that climate change has the potential to increase the transport of pollutants from lower latitudes to the Arctic, highlighting the importance of continued monitoring of ringed seal

147
contaminant levels. Kovacs (2007), noted that ringed seals are an important species to use in the study of global dispersal patterns of pollutants because of their broad circumpolar distribution, large populations, and high trophic status.
4.2.5.2 Oil and gas exploration, development, and production
4.2.5.2.1 Overview
4.2.5.2.1.1 The Arctic
Commercial oil and gas activities began in the Arctic in the 1920s in Canada’s Northwest Territories, and greatly expanded during the 1970s, particularly in Alaska, Russia, and Norway (AMAP 2007). By the 1980s and early 1990s, oil and gas activities had extended farther into the Arctic with several fields being explored and developed offshore. To date, cumulative production from the Arctic is in the billions of cubic meters for both oil and gas, and the Arctic is currently producing about a tenth of the world’s oil and a quarter of its gas (AMAP 2007). In 2008, the U.S. Geological Survey (USGS) completed an assessment of undiscovered conventional oil and gas resources in all areas north of the Arctic Circle. Using a geology‐based probabilistic method, the USGS estimated that 90 billion barrels of oil, 1,669 trillion cubic feet of natural gas, and 44 billion barrels of natural gas liquids may remain to be found in the Arctic, of which approximately 84% is expected to occur in offshore areas (U.S. Geological Survey 2008). These reserves may represent as much as a quarter of the world’s undiscovered oil and gas (AMAP 2007). Such extensive reserves coupled with rising global demand make it very likely that oil and gas activity will increase in the Arctic in the future. According to the Arctic Oil and Gas 2007 assessment (AMAP 2007):
“More activity is expected in the next two decades, however projections father into the future become increasingly speculative since the pace of activity is affected by a number of factors including economic conditions, societal considerations, regulatory processes, and technological advances. Global climate change may introduce additional factors that need to be taken into account.”
Climate change is expected to enhance marine access to offshore oil and gas reserves by reducing sea‐ice extent, thickness, and seasonal duration, thereby improving ship accessibility to these resources around the margins of the Arctic Basin (ACIA 2004, AMAP 2007). Reduced sea‐ice extent and thickness is also likely to allow construction and operation of more economical offshore platforms since the ice forces these structures must withstand may also be reduced (ACIA 2005, AMAP 2007). Conversely, land‐based infrastructure such as ice roads, pipelines, artificial islands, and coastal installations will likely be negatively impacted by climate change due to thawing of the permafrost and increased storm surges and coastal erosion (ACIA 2005, AMAP 2007). These effects may make future development on land more complicated and expensive, providing further incentive for the development of marine‐based production and transportation infrastructure (AMAP 2007).

148
4.2.5.2.1.1.1 United States (Alaska)
The United States schedules oil and gas lease sales on its OCS regions in 5‐year programs that indicate the size, timing, and location of proposed leasing activity for each period. In June 2007, the Secretary of the Interior approved the 2007‐2012 OCS Oil and Gas Leasing Program. Lease sales were planned in the Chukchi Sea in 2008, 2010, and 2012; in the Beaufort Sea in 2009 and 2011; and in the North Aleutian Basin of the southeastern Bering Sea in 2011 (Minerals Management Service 2007b). Two lawsuits were filed against the lease program in 2007, and in April 2009, the U.S. Court of Appeals for the District of Columbia vacated and remanded the program and required the Department of the Interior to “conduct a more complete comparative analysis of the environmental sensitivity of different areas” (Minerals Management Service 2010a). In March 2010, the MMS announced that the Preliminary Revised 2007‐2012 OCS Oil and Gas Leasing Program retains the Chukchi Sea lease sale from 2008 and allows proposed exploratory activities on leases in the Chukchi and Beaufort Seas to proceed, but removes the five remaining lease sales in the North Aleutian Basin and Beaufort and Chukchi Seas from the program (Minerals Management Service 2010c). A final 2007‐2012 leasing program will be approved after the Secretary of the Interior considers public comments on the revised plan (Minerals Management Service 2010a).
On 16 January 2009, the MMS announced the release of a 5‐year Draft Proposed Program (DPP) for offshore oil and gas lease sales for 2010‐2015 (Minerals Management Service 2009b), 2 years earlier than the usual schedule. The DPP calls for lease sales in the Chukchi Sea in 2010, 2012, and 2014; in the Beaufort Sea in 2013 and 2015; and in the North Aleutian Basin in 2011 and 2014. On 10 February 2009, the Secretary of the Interior announced that he was extending the comment period on the DPP by 180 days to provide additional time for states, stakeholders, and affected communities to provide input on the plan (Minerals Management Service 2009c). In the meantime, the MMS has initiated the process to develop the new 2012‐2017 OCS Oil and Gas Leasing Program, which will supersede the 2010‐2015 DPP once it’s completed. Currently, this process includes the Chukchi and Beaufort Seas as potential leasing locations for the 2012‐2017 program but does not include the North Aleutian Basin (Minerals Management Service 2010b).
The 20 April 2010 explosion of the Deepwater Horizon drilling platform in the Gulf of Mexico and the catastrophic oil blowout that resulted (estimated to be 4.9 million barrels total, Deepwater Horizon Response 2010) likely will impact future oil and gas activities in the United States, including Alaska. For example, on 27 May 2010, the Secretary of the Interior announced that Shell would not be permitted to drill five exploratory wells in their Beaufort and Chukchi Sea leases (scheduled for the summer of 2010) until at least 2011, pending further review by the MMS (U.S. Department of Interior 2010).
The State of Alaska also conducts lease sales and licensing for the exploration of oil and gas on state lands and in state waters. At least 20 lease sales have been made in state waters of the Beaufort Sea since 1979, and new lease sales are scheduled to occur annually according to the 5‐year leasing program for 2010‐2014 (Alaska Department of Natural Resources 2010b). The petroleum potential in these coastal waters is considered to be moderate to high.

149
4.2.5.2.1.1.1.1 Beaufort and Chukchi Seas
Oil and gas activities have been conducted off the coast of Alaska since the 1970s, with most of the activity occurring in the Beaufort Sea. Between 1976 and 2003, 31 offshore exploratory wells were drilled in the Beaufort Sea, 30 were drilled in the Bering Sea (including 6 deep stratigraphic test wells), and 5 were drilled in the Chukchi Sea (Minerals Management Service 2009a). The MMS has conducted 10 offshore lease sales in federal waters of the Beaufort Sea since 1979, and seven oil companies had active leases there as of January 2009 (Figure 31). The Alaska Department of Natural Resources has conducted an additional 20 lease sales in state waters of the Beaufort Sea since 1979 (Alaska Department of Natural Resources 2009), and many exploration and development activities are ongoing there (Alaska Department of Natural Resources 2010a). Three oil fields are currently in production in the Beaufort Sea where drilling is being conducted from artificial islands in state waters: Endicott field has been in production since October 1987, Northstar field since October 2001, and Oooguruk field since June 2008. By 2009, these three fields had produced nearly 600 million barrels of oil (Alaska Department of Natural Resources 2009). Development of the offshore Liberty oil field began in 2008 with production expected to begin in 2011 (Lee 2008). Production at the Nikaitchuq oil field is scheduled to begin from on onshore drilling pad in December 2010 and from an offshore drilling island in 2011 (Anchorage Daily News 2010). Several other oil fields on Alaska’s North Slope (e.g., Point Thompson, Badami, Dewline, and Beechey Point) have had development wells drilled near the coast of the Beaufort Sea in recent years (Alaska Department of Natural Resources 2010a). Operations or accidents at these facilities could potentially affect ringed seals in areas near shore.
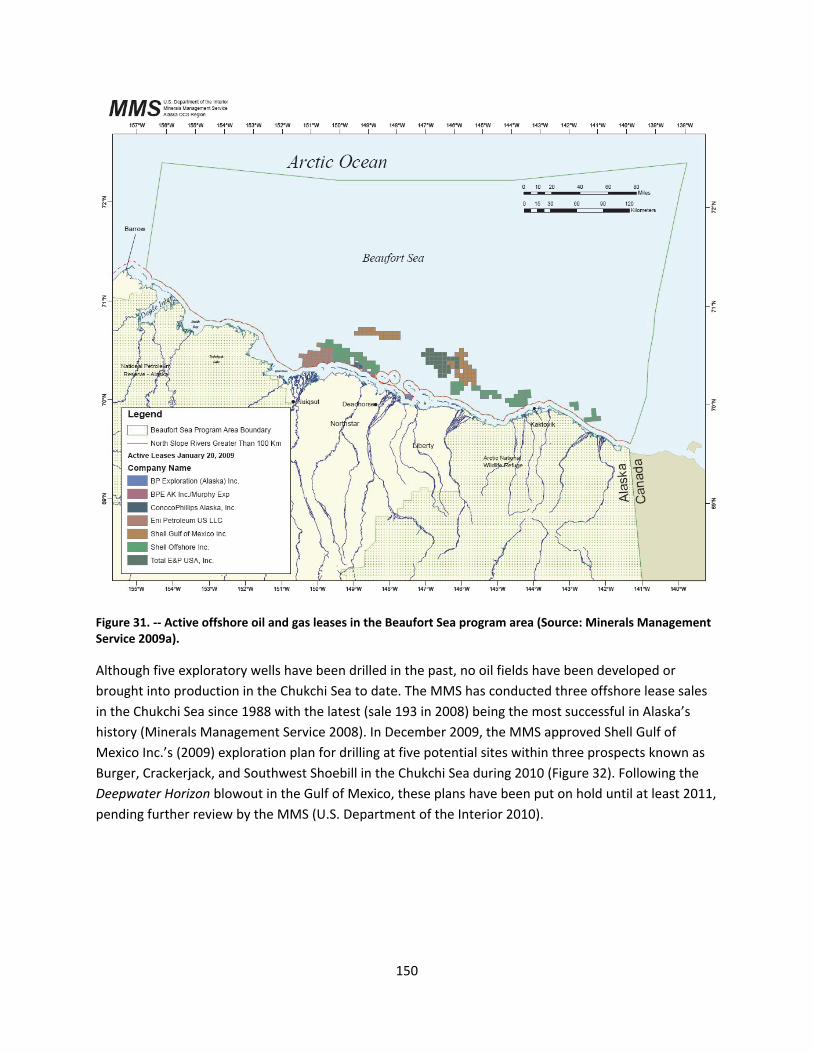
150
Figure 31. ‐‐ Active offshore oil and gas leases in the Beaufort Sea program area (Source: Minerals Management Service 2009a).
Although five exploratory wells have been drilled in the past, no oil fields have been developed or brought into production in the Chukchi Sea to date. The MMS has conducted three offshore lease sales in the Chukchi Sea since 1988 with the latest (sale 193 in 2008) being the most successful in Alaska’s history (Minerals Management Service 2008). In December 2009, the MMS approved Shell Gulf of Mexico Inc.’s (2009) exploration plan for drilling at five potential sites within three prospects known as Burger, Crackerjack, and Southwest Shoebill in the Chukchi Sea during 2010 (Figure 32). Following the Deepwater Horizon blowout in the Gulf of Mexico, these plans have been put on hold until at least 2011, pending further review by the MMS (U.S. Department of the Interior 2010).

151
Figure 32. ‐‐ Locations of three prospects (Burger, Crackerjack, and Southwest Shoebill) identified as potential drilling sites by Shell in their 2010 exploration plan (Source: Shell Gulf of Mexico Inc. 2009).
There are no offshore oil or gas fields in development or production in the Bering Sea, and all previous lease sales have expired (Minerals Management Service 2009a). Lease sales are not expected to resume in this region until 2012 at the earliest (Minerals Management Service 2010b).
4.2.5.2.1.1.2 Canada
From 1969 through the late 1980s, more than 400 exploratory wells were drilled in the Canadian Arctic (Geological Survey of Canada 2007). That exploration resulted in the discovery of 20 oil and gas fields in the Arctic Archipelago (Cranstone 2002) and 47 in Mackenzie Delta and Beaufort Sea regions (Geological Survey of Canada 2007). In the Arctic Archipelago, two of the gas fields are some of the largest in Canada, and total estimates across all fields of “proven, probable, and possible reserves” is 509 billion cubic meters of gas, 9.7 million cubic meters of natural gas liquids, and 200‐500 million barrels of oil (Cranstone 2002). In general, the economic costs and logistical challenges of extracting and transporting these petroleum resources south have been too high. One field (Bent Horn) in the Arctic Archipelago was put into production in the early 1980s, and approximately 100‐200 thousand barrels of oil were shipped annually to Montreal via an ice‐breaking tanker. By the time this practice ended in 1996 (due to a decline in crude prices), 2.8 million barrels of oil had been shipped from Bent Horn (Cranstone 2002).

152
Over the years, construction of natural gas pipelines from the Arctic Archipelago region have been proposed but have yet to be proven economically viable.
Substantial reserves of oil and gas are present within the Mackenzie Delta and the Canadian Beaufort Sea. Estimates of recoverable resources are as much as 1.8 trillion cubic meters of natural gas and 6.7 billion barrels of oil (Cranstone 2002). The level of future oil and gas activities in Canada probably depends on factors such as the price of oil or the development of new pipelines. For example, construction of a proposed Mackenzie Valley gas pipeline would likely increase exploration and development activities within the Mackenzie Delta and Beaufort Sea region (AMAP 2007) and may also open the door for a pipeline from the Arctic Archipelago. There has been historical opposition to these pipelines from Native peoples in the region, however, and even if the economics of pipeline construction were favorable, construction would likely not proceed without support from these communities. There are currently no active drilling operations within the Canadian Beaufort Sea (CAPP 2010).
4.2.5.2.1.1.3 Greenland
Oil and gas exploration was initiated in areas offshore West Greenland in the early 1970s with almost 21,000 km of seismic surveys (GHEXIS 2005). Five exploratory wells were drilled in Davis Strait during 1976‐1977, but no oil was found. Exploration activities moved to the East Greenland shelf in 1978 where over 63,000 km of aeromagnetic surveys and nearly 7,800 km of marine seismic surveys were conducted between 1979 and 1982 (GHEXIS 2005). No exploratory wells have been reported in the area.
Previous seismic data from West Greenland was re‐interpreted in the early 1990s, and it was determined that many of the earlier wells had been abandoned prematurely. At least 47,500 km of additional seismic surveys were conducted offshore West Greenland (including the northern regions of Baffin Bay) between 1990 and 2004, and nearly 7,000 km were conducted offshore Northeast and East Greenland between 1990 and 1996 (GHEXIS 2005). While exploration activities have yet to yield major oil or gas discoveries in these areas, they are being conducted at moderate levels, and efforts to date have produced results warranting further evaluation (AMAP 2007). The government of Greenland has recently adopted more favorable policies to increase oil and gas industry interest in Greenland’s shelves, and exploration activities are on‐going (AMAP 2007).
4.2.5.2.1.1.4 Norway and Svalbard
The Norwegian portion of the Barents Sea holds an estimated 2.2 billion barrels of oil and 30 trillion cubic feet of gas, of which about 90% is yet to be discovered (WWF 2010). Exploration for offshore oil and gas began in the Norwegian portion of the Barents Sea in the 1980s, which resulted in the discovery of a gas field in 1984 and an oil field in 2000 (AMAP 2007). Production began at the gas field in 2007 (AMAP 2007), and the oil field is currently under development (Fouche and Moskwa 2010). Norway is exploring the Barents Sea intensively and is planning continued oil and gas development along with associated offshore pipeline and tanker transport infrastructure in the future (AMAP 2007). In April 2010, Norway and Russia settled a 40‐year maritime border dispute in the Barents Sea by dividing the

153
disputed area into two equally significant parts, thereby, paving the way for the area to be opened for oil and gas exploration as early as 2012‐2013 (Fouche and Moskwa 2010). The entire disputed area holds and estimated 10 billion barrels of petroleum (Fouche and Moskwa 2010).
Oil and gas exploration activities occurred around Svalbard in the period between 1960 and 1979 with at least two exploratory wells being drilled offshore (AMAP 2007). Recently, several nations and oil companies have expressed interest in exploring for oil and gas around Svalbard (Adam 2007), and Russia announced plans to conduct surveys in the waters surrounding the archipelago in 2009 despite protests from the Norwegian government (BarentsObserver 2008b). Norway claims the right to establish an economic zone around Svalbard in which all commercial activities would be subject to its regulations, but the claims are being disputed by several nations (BarentsObserver 2008b).
4.2.5.2.1.1.5 Russian Federation
About 80% of the oil and 99% of the gas produced in the Arctic today comes from the Russian Arctic (AMAP 2007). With over 75% of known Arctic oil, over 90% of known Arctic gas, and vast estimates of undiscovered oil and gas reserves, Russia will continue to be the dominant producer of Arctic oil and gas in the future (AMAP 2007). According to Patin (1999), most estimates suggest that oil and gas reserves may be found on about 90% of all Russian shelves and that 90‐100 billion tons of oil equivalent (of which 80% is natural gas) may be potentially recoverable. Two huge oil and gas basins located on the shelves of the Barents and Kara Seas contain numerous oil and gas fields (Figure 33) with potential resources of at least 50‐60 billion tons of oil equivalent (Patin 1999). Just two of these fields located in the Kara Sea shelf hold an estimated 5 trillion cubic meters of natural gas, which in 1999 was equivalent to 2.5 times the worldwide gas production (Patin 1999). A gas condensate field located on the Barents Sea shelf is probably the world’s largest known offshore gas field, with reserves of about 3 trillion cubic meters of gas and more than 20 million tons of gas condensate (Patin 1999). Another field on the Barents Sea shelf was estimated to hold more than 200 million tons of oil (Patin 1999). Oil and gas developments in the Kara and Barents Seas began in 1992, and large‐scale production activities were initiated during 1998‐2000 (Patin 1999). Oil and gas production activities are expected to grow in the western Siberian provinces and Kara and Barents Seas in the future and will likely include the construction of several new pipelines and marine terminals and subsequent increases in tanker traffic to transport oil and gas to world markets (AMAP 2007).

154
Figure 33. ‐‐ Locations of oil (pink) and gas (yellow and red) fields in the Barents and Kara Seas and Yamal region of Russia (Source: BarentsObserver 2008a).
The shelves of Russia’s East Siberian, Chukchi, and Bering Seas have excellent prospects for large‐scale and long‐term developments of offshore oil and gas with potential recoverable reserves estimated in the billions of tons of conventional fuel (Patin 1999). Arctic waters offshore eastern Russia are thought to contain more than one‐third of the Russian Arctic’s recoverable offshore oil reserves and more than 10% of its recoverable gas reserves (Frantzen 2007). Recently, there has been renewed interest in the Russian Chukchi Sea as new evidence emerges to support the notion that the region may contain world‐class oil and gas reserves. Global oil and gas exploration companies are becoming more interested in the Russian Arctic offshore, because it is one of the world’s few remaining petroleum frontiers with significant resource potential. In 2006, 3,700 km of seismic surveys were conducted in Russia’s North and South Chukchi Basins to explore for economically viable oil and gas reserves. Preliminary results were described as “very encouraging” (Frantzen 2007).
4.2.5.2.1.2 Sea of Okhotsk
In the Sea of Okhotsk, oil and natural gas operations are active off the northeastern coast of Sakhalin Island, and future developments are planned in the Magadan and western Kamchatka regions (Figure 34). The project referred to as Salkhain‐1 consists of three deposits with extraction capability up to 10 billion cubic meters per year, and Salkhalin‐2 is one of the world’s largest integrated oil and gas projects with annual production of liquefied gas at nearly 20 billion cubic meters (Chernenko 2007). Oil extraction from these projects has already started with associated construction of a platform, a terminal for oil shipment, and a floating oil tank with a capacity of one million barrels (Lapko and Radchenko 2000). Initial work and seismic exploration for the projects Salkahin‐3,4,5 and 6 has also begun
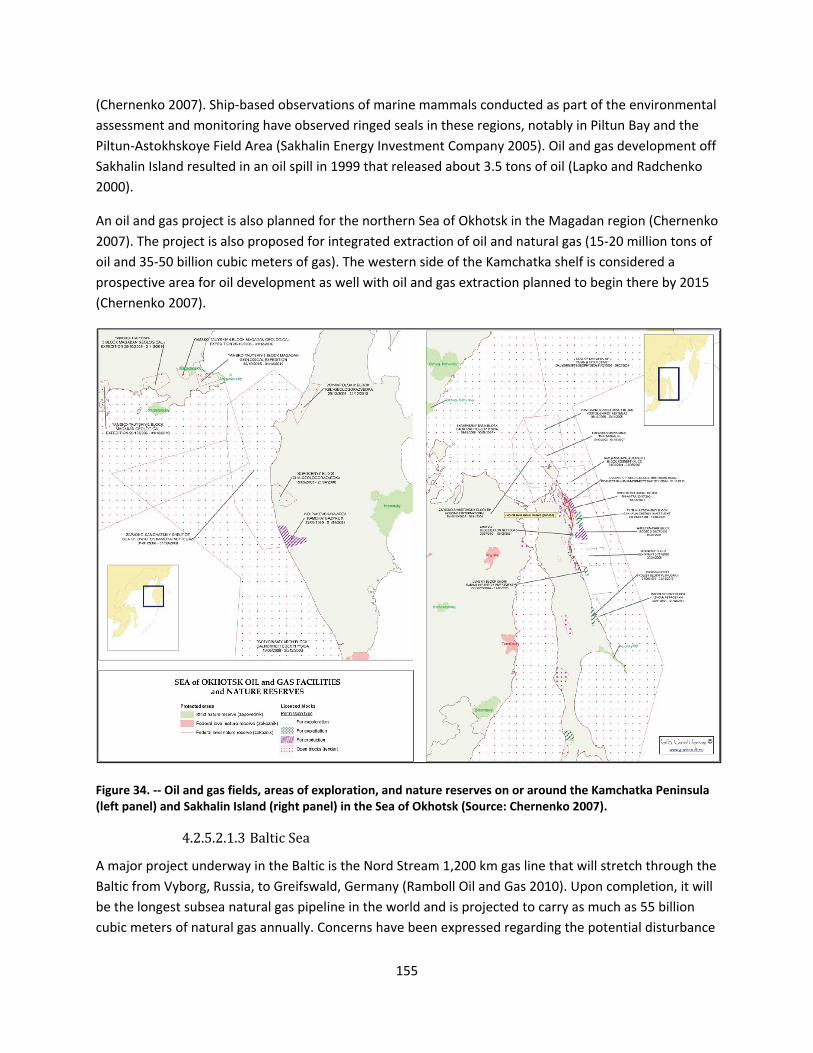
155
(Chernenko 2007). Ship‐based observations of marine mammals conducted as part of the environmental assessment and monitoring have observed ringed seals in these regions, notably in Piltun Bay and the Piltun‐Astokhskoye Field Area (Sakhalin Energy Investment Company 2005). Oil and gas development off Sakhalin Island resulted in an oil spill in 1999 that released about 3.5 tons of oil (Lapko and Radchenko 2000).
An oil and gas project is also planned for the northern Sea of Okhotsk in the Magadan region (Chernenko 2007). The project is also proposed for integrated extraction of oil and natural gas (15‐20 million tons of oil and 35‐50 billion cubic meters of gas). The western side of the Kamchatka shelf is considered a prospective area for oil development as well with oil and gas extraction planned to begin there by 2015 (Chernenko 2007).
Figure 34. ‐‐ Oil and gas fields, areas of exploration, and nature reserves on or around the Kamchatka Peninsula (left panel) and Sakhalin Island (right panel) in the Sea of Okhotsk (Source: Chernenko 2007).
4.2.5.2.1.3 Baltic Sea
A major project underway in the Baltic is the Nord Stream 1,200 km gas line that will stretch through the Baltic from Vyborg, Russia, to Greifswald, Germany (Ramboll Oil and Gas 2010). Upon completion, it will be the longest subsea natural gas pipeline in the world and is projected to carry as much as 55 billion cubic meters of natural gas annually. Concerns have been expressed regarding the potential disturbance

156
of World War II landmines and chemical toxins in the sediment during construction. Once the pipeline is completed and operational, there are additional concerns about potential leaks and spills from the pipeline and the impacts on the Baltic Sea marine environment. Circulation of waters in the Baltic Sea is limited and any contaminants may not be flushed efficiently.
Exploration of the Baltic Sea off the Latvian coast began in 1976 and, through 1991, 24,500 km of seismic surveys were conducted to identify potential deposits. While several prospective areas were identified, only two locations were drilled. Additional seismic exploration was conducted in the early 1990s near the Swedish and Lithuanian borders. Seismic exploration by the Latvian government has been limited since the early 1990s due to insufficient funding. In 2001, Latvia initiated open offshore licenses for oil exploration and production off the southern coast along the Lithuanian border. Three oil companies have been awarded licenses. These licenses were awarded for up to 30 years and include exploration activity for up to 5 years.
4.2.5.2.1.4 Lakes Ladoga and Saimaa
There are no oil or gas exploration or development projects in Lakes Ladoga or Saimaa.
4.2.5.2.2 Potential threats to ringed seals from offshore oil and gas exploration, development, and production
Potentially harmful oil and gas exploration, development, and production activities include seismic surveys; exploratory, delineation, and production drilling; construction of artificial‐islands, causeways, ice roads, shore‐based facilities, and pipelines; and vessel and aircraft operations. These activities have the potential to impact ringed seals, primarily through the noise, physical disturbance, and pollution they cause, particularly in the event of a large oil spill or blowout.
4.2.5.2.2.1 Noise and physical disturbance
Noise and physical disturbances associated with oil and gas exploration, drilling, construction, and support activities could potentially interfere with ringed seals’ ability to function normally and impact their health. The ‘noisiest’ and ‘busiest’ period of offshore oil and gas operations occurs during exploration and site establishment (Richardson et al. 1995). Conversely, production activities are generally quieter and require fewer support operations. To varying degrees, drilling operations produce low‐frequency sounds with strong tonal components.
Moulton et al. (2005) reported that during spring aerial surveys for ringed seals, there was no evidence that construction, drilling, and production activities at BP’s Northstar oil development in the Beaufort Sea affected local ringed seal distribution and numbers. Drilling and production sounds from Northstar likely were audible to ringed seals, at least intermittently, out to about 1.5 km in water and 5 km in air (Blackwell et al. 2004). Underwater sounds from construction, drilling, and production reached background values at 2‐4 km (Richardson and Williams 2004), while underwater sound from vessels often were detectable as far as 30 km offshore. Likewise, Richardson and Williams (2004) concluded that there was little effect from the low to moderate level, low‐frequency industrial sounds emanating from the Northstar facility on ringed seals during the open‐water period and that the overall effects of the

157
construction and operation of the facility were minor, short‐term, and localized, with no consequences to seal populations as a whole. Because the Northstar facility is on a manmade artificial island, it is not known whether these results are applicable to other types of drilling and production facilities.
Harwood et al. (2007) evaluated the potential impacts of offshore exploratory drilling on ringed seals in the near shore Canadian Beaufort Sea, during February to June 2003‐2006. The first 3 years of the study (2003‐2005) were conducted prior to industry activity in the area, while a fourth year of study (2006) was conducted during the latter part of a single exploratory drilling season. Seal breathing holes and lairs were not significantly different in distance from industrial activities during the non‐industry (2003 and 2004) and industry (2006) years. The movements, behavior, and home range size of 10 seals tagged in 2006 also did not vary statistically between the 19 days when industry was active (20 March to 8 April) and the following 19 days when industry operations were completed. The density of basking seals was not significantly different among the different study years and was comparable to densities found in this same area during surveys conducted in 1974‐1979. No detectable effect on ringed seals could be discerned in the one season of drilling in the study area (Harwood et al. 2007). The effects of longer exposures to industrial activity, or exposure to multiple industrial sources, remain unknown.
Air and vessel traffic are often needed to support offshore oil and gas exploration and development. Low‐flying aircraft may cause resting ringed seals to enter the water (Burns and Harbo 1972), however, most of these disturbances will likely be minor, highly variable, and brief in nature. The effects of noise and physical disturbance from ships are discussed in Section 4.2.5.5.1.3.2.
4.2.5.2.2.1.1 Seismic surveys
Seismic surveys are a particularly intense source of noise and, thus, warrant further consideration as a potential threat. Pinnipeds use the acoustic properties of sea water to aid in navigation, social communication, and possibly predator avoidance. There is considerable variability in the vocalizations of seals, and many of the ice‐associated species vocalize underwater in association with territorial and mating behaviors. Most phocid seals spend greater than 80% of their time submerged in the water (Gordon et al. 2003); consequently, they will be exposed to sounds from seismic surveys that occur in their vicinity. Phocids have good low‐frequency hearing; thus, it is expected that they will be more susceptible to masking of biologically significant signals by low frequency sounds, such as those from seismic surveys (Gordon et al. 2003). Masking of biologically significant sounds by anthropogenic noise is equivalent to a temporary loss of hearing acuity. Brief, small‐scale masking episodes might, in themselves, have few long‐term consequences for individuals or groups of ringed seals. The consequences might be more serious in areas where many surveys are occurring simultaneously. Underwater audiograms for phocids suggest that they have very little hearing sensitivity below 1 kHz, though they can hear underwater sounds at frequencies up to 60 kHz and make calls between 90 Hz and 16 kHz (Richardson et al. 1995). A more recent review suggests that the auditory bandwidth for pinnipeds in water should be considered to be 75 Hz to 75 kHz (Southall et al. 2007). While seismic surveys can contain energy up to 1 kHz, most of the emitted energy is less than 200 Hz.

158
There is no specific evidence that exposure to pulses of airgun sound can cause permanent threshold shifts to the hearing of any marine mammal, even with large arrays of airguns. Nevertheless, direct impacts causing injury from seismic surveys would likely occur only if animals entered the zone immediately surrounding the sound source. Southall et al. (2007) proposed that auditory injury would occur to pinnipeds exposed to a single pulse of sound at 218 dB re: 1 micropascal in water and 143 dB re: 20 micropascals in air. Although it is unlikely that airgun operations during most seismic surveys would cause permanent threshold shifts in ringed seals, caution is warranted given the limited knowledge about noise‐induced hearing damage in this species.
Reported seal responses to seismic surveys have been variable and often contradictory, although they do suggest that pinnipeds frequently do not avoid the area within a few hundred meters of operating airgun arrays (Brueggeman et al. 1991, Harris et al. 2001, Miller and Davis 2002). Kelly et al. (1988), however, found that the frequency of abandonment of breathing holes by ringed seals was significantly greater within 150 m of on‐ice seismic exploration than at greater distances. They concluded that the population level effects would be minor, in part, due to an assumption that ringed seals could readily move to other areas under the ice. Subsequent evidence of strong fidelity to under‐ice home ranges, however, suggested that such displacement may be costly to the fitness of ringed seals (Kelly et al. 2010).
Telemetry work by Thompson et al. (1998) indicated that harbor seals and gray seals exhibit strong avoidance behavior of small seismic airgun arrays, including swimming rapidly away from seismic sources, ceasing feeding activities, and exiting the water, possibly to avoid underwater noise. The behavior of most of the seals reportedly returned to normal within 2 hours of the seismic array falling silent. The authors suggested that responses to more powerful commercial arrays might be more dramatic and occur at greater ranges. Based on tagging studies conducted in 2001, Cott et al. (2003) reported that Canadian marine seismic surveys in the Beaufort Sea did not appear to affect the timing or route of the western fall migration of ringed seals. A comparison of ringed seal densities in areas of shorefast ice with seismic activity (using explosive charges) vs. undisturbed areas indicated that ringed seals were not appreciably displaced by even intensive seismic activity, however, the study took place immediately offshore of Prudhoe Bay where ringed seals were said to “normally occur at very low densities” (Burns and Harbo 1972).
4.2.5.2.2.2 Oil spills
Large oil spills or blowouts are generally considered to be the greatest threat of oil and gas activities in marine environments (AMAP 2007). In contrast to spills on land, large spills at sea are difficult to contain and may spread over hundreds or thousands of kilometers. Responding to a spill in the Arctic environment would be particularly challenging. Many oil and gas activities occur in locations far from population centers, where spill response resources are typically located. Reaching a spill site and responding effectively would be especially difficult in winter, when weather can be severe and daylight extremely limited. Oil spills under ice or in ice‐covered waters are the most challenging to deal with, simply because they cannot be contained or recovered effectively with current technology. Current techniques, such as use of dispersants and in‐situ burning, may be attempted, however, both techniques

159
have drawbacks that may be made worse in Arctic conditions (AMAP 2007). New techniques, such as the use of ice rather than booms to contain and concentrate oil until it can be recovered by skimmers, have shown some promise, but most response techniques require rapid action to be effective, which may not be possible in remote areas without prior staging of equipment and personnel (AMAP 2007). The difficulties experienced in stopping and containing the oil blowout at the Deepwater Horizon well in the Gulf of Mexico, where environmental conditions and response preparedness are comparatively good, point towards even greater challenges of attempting a similar feat in a much more severe and remote Arctic location.
Although planning, management, and use of best practices can help reduce risks and impacts, the history of oil and gas activities, including recent events, indicates that accidents cannot be eliminated (AMAP 2007). Tanker spills, pipeline leaks, and oil blowouts are likely to occur in the future, even under the most stringent regulatory and safety systems. According to a figure in AMAP’s (2007) report, one oil tanker spill of 35,000 cubic meters or greater is estimated to occur each year, and an oil blowout of 2,000 cubic meters or greater is estimated to occur every 10 years. And according to the MMS (2007a), the probability of one or more large oil spills (≥ 1,000 barrels) occurring over the production life of a new development in the Chukchi Sea was estimated to be between 35‐40%. To date, there have been no large spills in the Arctic marine environment from oil and gas activities, however, based on experiences of spills in subarctic regions (e.g., Prince William Sound, Alaska; the North Sea in Europe; and the Gulf of St. Lawrence, Canada), the potential impacts of a spill in the Arctic are likely to be severe for Arctic species and ecosystems (AMAP 2007).
Ringed seals could be impacted by oil spills in several ways. The greatest impacts would likely result from an oil spill during the pupping season (Smith 1987, St. Aubin 1990). Researchers have suggested that pups may be particularly vulnerable to fouling because of their dense lanugo coat (Johnson 1983, St. Aubin 1990, Jenssen 1996). Fouled pelage of neonates would have a lower insulative value, putting them at greater risk of low‐temperature stress when out of the water (Kooyman et al. 1977, St. Aubin 1990), lower mass at weaning (Davis and Anderson 1976), and lower survival (Harding et al. 2005). Ringed seals pups rely on their lanugo for insulation and typically retain this coat for 4‐6 weeks after birth. During this time, they would be particularly prone to the physical impacts of contacting oil. Energetic costs may be compounded if mothers attempted to move their pups out of the affected area. The increased time spent swimming in water could lead to hypothermia and increased mortality in pups (Smith 1987). Adults, juveniles, and weaned young‐of‐the‐year rely on blubber for insulation, so effects on their thermoregulation are expected to be minimal (Jenssen 1996).
Other acute effects of oil exposure, which have been shown to reduce seals’ health and possibly survival, include: skin irritation, disorientation, lethargy, conjunctivitis, corneal ulcers, and liver legions (Geraci and Smith 1976a, St. Aubin 1988). Many of these effects are thought to be largely reversible, but others such as brain legions and nerve damage may be fatal (Engelhardt 1983, Frost and Lowry 1994, Frost et al. 1994b, Lowry et al. 1994, Spraker et al. 1994, Salazar 2003). Six captive ringed seals that were experimentally dosed with crude oil for 24‐hr in the field exhibited profuse tearing, behavioral changes, corneal erosions and ulcers, severe conjunctivitis, and swollen nictitating membranes (Geraci and Smith

160
1976b). All seals recovered quickly after being placed in clean water, and within 3‐4 days, showed no signs of irritation. Conversely, three captive ringed seals that were dosed with crude oil in a laboratory died within 21‐71 minutes of the exposure. The researchers attributed their deaths to severe stress caused by the presence of the oil, rather than any toxic effects of the oil (Geraci and Smith 1976b).
Direct ingestion of oil, ingestion of contaminated prey, or inhalation of volatile hydrocarbons transfers toxins to body fluids, muscle, liver, and blubber, causing effects that may lead to death, as suspected in dead gray and harbor seals found with oil in their stomachs (Engelhardt et al. 1977, Engelhardt 1982, St. Aubin 1990, Frost et al. 1994b, Lowry et al. 1994, Spraker et al. 1994, Jenssen 1996). Furthermore, ingestion of hydrocarbons irritates and destroys epithelial cells in the stomach and intestine, affecting motility, digestion, and absorption, which can result in death or reproductive failure (St. Aubin 1990). Engelhardt et al. (1977) reported results from immersion and ingestion studies on captive ringed seals and noted that although petroleum hydrocarbons were rapidly absorbed and distributed throughout body tissues and fluids, there were no major pathological effects.
Freshly spilled oil contains high levels of toxic aromatic compounds that, if inhaled, could cause serious health effects or death in ringed seals, as occurred with an estimated 300 harbor seals following the Exxon Valdez oil spill in Prince William Sound, Alaska (Frost et al. 1994a, Frost et al. 1994b, Lowry et al. 1994, Spraker et al. 1994). Oil that disperses from a spill site may still have high levels of toxic aromatic compounds, depending on the temperature and whether the oil becomes frozen into ice (St. Aubin 1990). Pinnipeds stressed by parasitism or other metabolic disorders may be susceptible to injury or death from even brief exposure to low concentrations of hydrocarbon vapors (St. Aubin 1990). For example, parasitized lungs—common in pinnipeds—can exacerbate the effects of even mild irritation of respiratory tissues (St. Aubin 1990). Toxicity of oil is generally greater in younger animals so exposure to oil contamination during the breeding season would likely cause higher mortality among pups (Jenssen 1996, Jenssen et al. 1996).
4.2.5.2.3 Oil and gas activity threat assessment
Though the probability of an oil spill affecting a significant portion of a ringed seal population is likely low, the potential impacts from such a spill could be significant, particularly if subsequent clean‐up efforts were ineffective. Because toxicity varies widely across oil types and with exposure, it is difficult to generalize about the toxicological impacts on ringed seals that come in contact with oil. Based on the documented exposures of ringed seals and other phocid species to oil, however, significant effects on health and survival would be expected for any seal that is immersed or coated in oil during the days and weeks following a spill (St. Aubin 1990). The spatial extent, persistence, and proximity of oil to areas of higher density of ringed seals (e.g., in geographically confined regions)—and the effectiveness of the spill response—are less predictable factors that would greatly modify exposures and, thus, the biological impacts of a large release of oil into the ocean.
It is important to evaluate the effects of anthropogenic perturbations, such as oil spills, in the context of historical data. Without historical data on distribution and population size, it is difficult to predict the impacts of an oil spill on ringed seals. Population monitoring studies need to be implemented in areas

161
where significant industrial activities are likely to occur, so that it will be possible to compare future impacts with historical patterns and determine the magnitude of potential effects (Frost et al. 2004).
4.2.5.3 Wind farms
As many as 2,000 wind generators are planned to be erected in the Bothnian Bay, the main breeding area for Baltic ringed seals (M. Kunnasranta, Finnish Game and Fisheries Research Institute, pers. comm., 9 July 2010). Impacts of disturbance to ringed seals associated with the construction and operation of wind farms are unknown.
4.2.5.4 Commercial fisheries interactions and bycatch
4.2.5.4.1 Bycatch
4.2.5.4.1.1 The Arctic
Commercial fisheries may impact ringed seals through direct interactions (i.e., incidental take or bycatch) and indirectly through competition for prey resources and impacts on size structure, genetics, and/or life history of prey populations. Direct interactions between commercial fishing and marine mammals in Alaska federal waters are monitored by way of shipboard observers (who also monitor the fishery catch). During the 1990s, three commercial fisheries were monitored: the BSAI groundfish trawl fishery, longline fisheries, and pot fisheries. Observer coverage during this period ranged from 9.7‐74% of tons caught. Two incidental mortalities of ringed seals in the BSAI trawl fishery were observed in 1992; no mortalities were observed after 1992, and the average annual mortality of ringed seals for the decade was zero (Angliss and Lodge 2002). Beginning in 2003, the three fisheries categories were further divided into 12, reflecting not a change in fishing effort, but an improvement in the information about which components of the fisheries might be responsible for incidental catch of marine mammals (Angliss and Allen 2009). During 2000‐2004, incidental injuries and mortalities of ringed seals only occurred in the BSAI pollock trawl fishery. Observer coverage during this period ranged from 76.2‐82.2%. One mortality was observed in 2000 and one in 2001 for a mean annual mortality during this period of 0.71 (Angliss and Allen 2009). During 2002‐2006, incidental injuries and mortalities of ringed seals only occurred in the BSAI flatfish trawl survey. Observer coverage ranged from 58.4‐67.8%. One mortality was observed in 2005 and one in 2006 for mean annual mortality of 0.46 (Allen and Angliss 2010).
Estimates of ringed seal bycatch levels in other parts of the Arctic are not available. In an assessment of the status and threats of ringed seals in the North Atlantic region, NAMMCO (2002) stated that ringed seals are seldom accidentally caught in fishing gear because their distribution does not coincide with intensive fisheries in most areas.
4.2.5.4.1.2 Sea of Okhotsk
The frequency of ringed seal bycatch in the Sea of Okhotsk is unknown. Given the intensity of commercial fishing in the Sea of Okhotsk (Lapko and Radchenko 2000, Dulepova and Radchencko 2004), however, some bycatch seems likely.

162
4.2.5.4.1.3 Baltic Sea
Drowning in fishing gear is one of the most significant mortality factors for seals in the Baltic Sea, especially for young seals which are prone to getting trapped in fishing nets (Ministry of Agriculture and Forestry 2007). There are no reliable estimates of seal bycatch in the Baltic Sea, and available estimates for many areas are believed to under represent actual takes (Ministry of Agriculture and Forestry 2007). Approximately 5% of the commercial fishing effort is monitored, and it was estimated that 80 ringed seals were caught in Swedish coastal fisheries in 2004 (Lunneryd et al. 2005). In Latvia, 200‐400 seals are reported as bycatch each year, however, the majority of these are young gray seals (Urtans et al. 2005). There are no estimates of seal bycatch from Lithuanian, Estonian, or Russian waters of the Baltic. The incidence of seal bycatch was expected to increase along with increasing seal numbers in the Baltic Sea, however, decreases in the use of the most harmful types of nets (i.e., gillnets and unprotected trap nets) along with the development of seal‐proof of fishing gear were thought to have resulted in a decline in the share of bycatch (Ministry of Agriculture and Forestry 2007).
4.2.5.4.1.4 Lake Ladoga
It was estimated that 200‐400 Lake Ladoga ringed seals died annually in fishing gear during the late 1980s and early 1990s (Sipilä et al. 2002). Fishing patterns, however, have changed since then due to changes in the economic market, and decreases in total catch have been observed since 1990. Thus, as of the late 1990s, fishing was not regarded to be a threat to Ladoga ringed seal populations but could again be a threat if market conditions improve (Sipilä and Hyvärinen 1998). Sipilä et al. (2002) recommended several restrictions related to fishing gear, seasons, and areas to minimize bycatch of newly‐weaned pups and adult ringed seals in Lake Ladoga and also noted that bycatch data should be measured more carefully in the future. Based on interviews with fishermen in Lake Ladoga, at least 483 ringed seals were killed in fishing gear in 2003, even though official records only recorded 60 cases of bycatch (Verevkin et al. 2006).
4.2.5.4.1.5 Lake Saimaa
Small‐scale fishing was thought to be the most serious threat to ringed seals in Lake Saimaa (Sipilä and Hyvärinen 1998). Of 182 carcasses that were examined for the period 1977 to 2000, 97 (53%) were determined to have died from drowning in fishing gear, making this the most common cause of death for ringed seals in Lake Saimaa (Sipilä 2003). In an effort to protect newly‐weaned pups, major breeding areas were identified, and people were encouraged to avoid those areas when fishing. In addition, the Finnish government restricted certain types of fishing gear (strong gill nets, certain fyke nets, and baited hooks) year round in major breeding areas. During 1982‐1984, 39% of pups born in areas with fishing restrictions were captured in nets in their first 2 years of life, while 50% were captured in areas without restrictions (Sipilä and Hyvärinen 1998). The restricted areas were later expanded, and captures fell to 13% in restricted areas and 21% in unrestricted areas by the late 1990s (Sipilä 2003). During the late 1990s, 1‐3 adult ringed seals were lost annually to nets, hooks, and traps (Sipilä and Hyvärinen 1998). Sipilä (2003) believed this threat to the population was likely to increase in the future due to trends in fishing methods and concluded that the Saimaa ringed seal was in great danger of extinction without fishing restrictions.

163
4.2.5.4.2 Indirect interactions
Commercial fisheries target a number of ringed seal prey species, such as walleye pollock, Pacific cod, herring, and capelin. Those fisheries may indirectly impact ringed seal populations through reductions in prey biomass. The U.S. fisheries in the North Pacific are managed to prevent overfishing of individual stocks. As such, strict limits on catch and bycatch are placed on all groundfish species or species groups. Even well‐managed fisheries, however, will result in reduced levels of biomass relative to theoretical mean unfished levels. The extent that reduced numbers in these individual stocks affect the viability of ringed seal populations is unknown. In the U.S. exclusive economic zone (EEZ), overall biomass levels of all groundfish species have remained stable between 15 and 20 million metric tons of biomass after showing substantial increases since the 1970s (Mueter and Megrey 2006). Ringed seals were not believed to be significantly competing with or affected by commercial fisheries in the waters of Alaska (Frost 1985, Kelly 1988), Canada (Malouf 1986 cited in Reeves 1998), Russia (Heptner et al. 1976), the Baltic Sea (Ministry of Agriculture and Forestry 2007), Lake Saimaa (Auvinen et al. 2005), or Lake Ladoga (Sipilä et al. 2002). Reeves (1998) suggested that ringed seals may be in competition with the shrimp fishery around Svalbard and the shrimp and Greenland halibut fisheries around Greenland.
Commercial fishing can also have indirect effects on marine mammals through changes in genetics, reproductive capacity, and life history characteristics of their prey. Fisheries generally select particular individuals (usually larger and older fish) and focus on particular locations (such as spawning or feeding grounds) such that fishing is non‐random with respect to fish characteristics (or phenotypes). If there is a genetic component to differences in phenotypes between fish, then fishing will cause evolutionary change. The argument that fishing could cause phenotypic evolution is widely known in general terms. There are numerous examples of changes in life history characteristics of commercially exploited stocks over time, such as weight‐at‐age, length‐at‐age, length‐at‐maturation, and age‐at‐maturation (reviewed in Law 2000). Current patterns of fishing for Northeast Arctic cod are selecting for fish that allocate resources to reproduction rather than growth. The sustainable yield associated with this life history could be less than half the yield potentially available (Law and Grey 1989).
Fishing can impact reproductive capacity through changes in fish size. Reductions in fish size can result from long‐term size‐selective fishing (e.g., Zwanenburg 2000). The decrease in the proportion of large fish might have negative impacts on reproductive capacity of the population if smaller, first‐time spawners are less successful in producing viable eggs than are larger, more experienced spawners (Trippel et al. 1997). In addition, some temperate demersal fishes, such as cod and other gadids, are thought to rely on the longevity and size of mature individuals to bridge the gaps between years of strong recruitment (Longhurst 1999). A reduction in the average size of prey species also could reduce the per capita energy content and may increase the foraging effort exerted by ringed seals. Conversely, older fish may be more cryptic, harder to catch, and less numerous. Groundfish stocks are known to have a high degree of interannual variability in recruitment and it is likely that such fluctuations occurred prior to fishing. As such, ringed seals dependence on different size composition for groundfish species would seem to be fairly adaptable.

164
Some fisheries may be expected to expand or shift northward in response to an increased length of the ice‐free, open‐water season in the future. If such shifts occur, the likelihood of both direct and indirect fisheries interactions with ringed seals may increase. The North Pacific Fishery Management Council has established an Arctic Fisheries Management Plan that would place a moratorium on development of fisheries in federally managed waters in the U.S. EEZ north of Bering Strait (North Pacific Fishery Management Council 2009). Several Russian companies recently sent longline vessels to explore fishing prospects in the Chukchi Sea, with unknown results, and the fishing season in the western Bering Sea has reportedly been increasing due to reduced sea‐ice conditions (V. Burkanov, Kamchatka Branch of the Pacific Institute of Geography, pers. comm., 19 September 2008).
4.2.5.5 Shipping and transportation
4.2.5.5.1 The Arctic
The substantial reduction in Arctic sea ice that has occurred in recent years has renewed interest in the Arctic Ocean as a waterway for coastal, regional, and trans‐Arctic marine operations (Brigham and Ellis 2004). Declines in sea‐ice extent and thickness have provided greater access to marine navigation routes, especially along the margins of the Arctic Basin, which historically have been ice‐covered for most or all of the year (ACIA 2004). Climate models predict that the warming trend in the Arctic will accelerate, causing the sea ice to begin melting earlier in the spring, retreat farther away from most Arctic landmasses, get thinner during the summer, and resume freezing later in the fall, resulting in an expansion of potential shipping routes and lengthening the potential navigation season (ACIA 2004, Khon et al. 2010). The reduction in sea ice “is very likely to increase marine transport and access to resources” in the Arctic during this century (ACIA 2004). A comprehensive review and analysis of current (2004) and future (2020) marine shipping activities in the Arctic was presented in the Arctic Marine Shipping Assessment (AMSA) 2009 Report (Arctic Council 2009). Much of the following information was incorporated from this report.
According to the AMSA report (Arctic Council 2009), the term “shipping” refers to the various uses of all types of ships (except naval vessels), including tankers, bulk carriers, offshore supply vessels, passenger ships, tug‐barge combinations, fishing vessels, ferries, research vessels, and icebreakers. These ships may travel to or from destinations within the Arctic (destinational shipping) or may use the Arctic Ocean as a marine link between the Atlantic and Pacific Oceans (trans‐Arctic shipping). At present, the two main navigation routes crossing the Arctic are the Northwest Passage (NWP) and Northern Sea Route (NSR). A proposed new route termed the Central Arctic Ocean Route (CAOR), which would cross a significant portion of the Arctic Basin, could be navigable at least intermittently and be economically feasible by mid‐century (Holland et al. 2006, Ellis 2008). Compared to the NSR, the CAOR would reduce the distance between Russian ports by 10‐15%.
The NSR, which is actually the central portion of a longer trans‐Arctic route called the Northeast Passage, traverses the Russian Arctic along the northern coast of Eurasia from the Barents Sea in the west to the Bering Sea in the east (Figure 35). For ships travelling between northern Europe and Far East Asia or Alaska, the NSR represents a savings of 35‐60% in distance when compared to the normal

165
shipping routes through the Suez or Panama Canals (Arctic Council 2009). This shallow, seasonally ice‐covered route has been maintained year‐round in its western portion by Russian icebreakers since 1979 and has been open to international marine traffic since 1991 (ACIA 2004).
Figure 35. ‐‐ Major shipping routes and bathymetry of the Arctic Ocean (Source: Arctic Council 2009; Map 2.1).
The NWP traverses the Arctic along the northern coast of North America from the Labrador Sea in the east, through the Canadian Arctic Archipelago, to the Bering Sea in the west (Figure 35). The operating season along the NWP is short—generally from late July to mid‐October—and depends on the ice conditions in any given year (Arctic Council 2009). This perennially ice‐choked passage was ice‐free for the first time in recorded history during the summer of 2007 (National Snow and Ice Data Center 2007). Studies by the Canadian Ice Service indicated that sea‐ice conditions in this region during the past three decades have been characterized by high interannual variability, making prospects for regular marine transportation less predictable (ACIA 2004). Additional studies by Canada’s Institute of Ocean Sciences

166
suggested that increasing amounts of multi‐year sea ice and glacial ice bergs could be flushed through the channels and straits of the NWP more frequently as continued melting weakens the perennial blockages or “ice bridges” that have controlled ice movements in the past (ACIA 2004). These larger, denser ice features could present serious navigational hazards along this route.
4.2.5.5.1.1 Current shipping activity in the Arctic
A comprehensive survey of all shipping activity in the Arctic was conducted as part of the AMSA study, using 2004 as the baseline year (Arctic Council 2009). Responses to the survey varied greatly between Arctic states, with several states unable to provide complete data for 2004; therefore, the AMSA survey likely underestimated the levels of shipping activity during the reporting year. The survey estimated that approximately 6,000 individual vessels operated in the Arctic during 2004, of which about half were vessels travelling along the Great Circle Route through the Aleutian Islands in the North Pacific Ocean/southern Bering Sea. Excluding vessels using the Great Circle Route, the most vessels by category were fishing vessels at about 1,600 or slightly less than 50% of the remaining total, followed by bulk carriers at about 20%. Shipping activity took place throughout the Arctic in 2004, including eight icebreaker expeditions to the North Pole during the summer, however, most operations occurred along the periphery of the Arctic Ocean in coastal waters that were either ice‐free year‐round or ice‐covered only seasonally (Figure 36). In the seasonally ice‐covered areas, nearly all of the vessel activity in 2004 occurred during or after the ice melt, when icebreakers were not required for access (Arctic Council 2009).
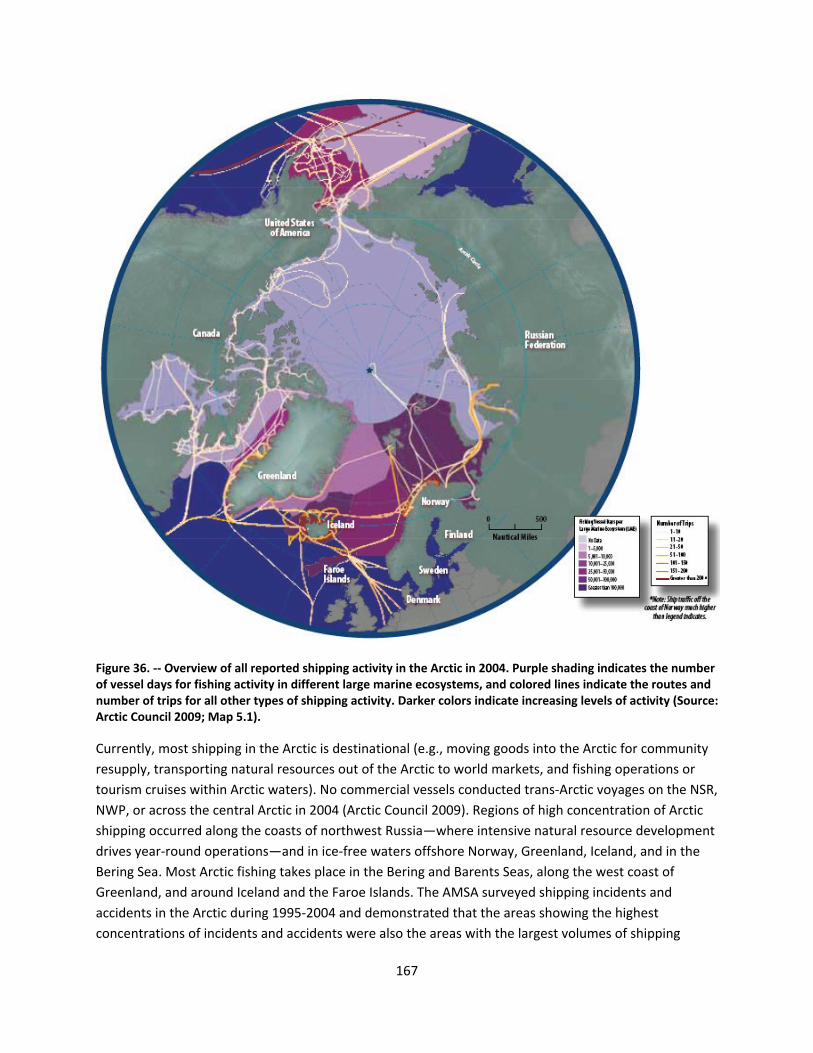
167
Figure 36. ‐‐ Overview of all reported shipping activity in the Arctic in 2004. Purple shading indicates the number of vessel days for fishing activity in different large marine ecosystems, and colored lines indicate the routes and number of trips for all other types of shipping activity. Darker colors indicate increasing levels of activity (Source: Arctic Council 2009; Map 5.1).
Currently, most shipping in the Arctic is destinational (e.g., moving goods into the Arctic for community resupply, transporting natural resources out of the Arctic to world markets, and fishing operations or tourism cruises within Arctic waters). No commercial vessels conducted trans‐Arctic voyages on the NSR, NWP, or across the central Arctic in 2004 (Arctic Council 2009). Regions of high concentration of Arctic shipping occurred along the coasts of northwest Russia—where intensive natural resource development drives year‐round operations—and in ice‐free waters offshore Norway, Greenland, Iceland, and in the Bering Sea. Most Arctic fishing takes place in the Bering and Barents Seas, along the west coast of Greenland, and around Iceland and the Faroe Islands. The AMSA surveyed shipping incidents and accidents in the Arctic during 1995‐2004 and demonstrated that the areas showing the highest concentrations of incidents and accidents were also the areas with the largest volumes of shipping

168
activity. In the few years since the AMSA survey, shipping activity in the Arctic has increased significantly. For example, cruise ship traffic around Greenland has increased exponentially in recent years, and several new icebreaking tankers are providing a continuous supply of oil and gas for export out of northwest Russia and northern Norway. At the current rate of increase, it will not take long to double or triple the 2004 shipping activity levels in the Arctic (Arctic Council 2009).
4.2.5.5.1.2 Future shipping activity in the Arctic
Plausible and diverse future scenarios were developed as part of the AMSA study to identify the major sources of uncertainty that would likely shape the future of Arctic shipping activities to the years 2020 and 2050 (Arctic Council 2009). Table 4 describes the four future scenarios that were chosen as being the most plausible, the most relevant to the Arctic and maritime affairs, and as including the appropriate level of external factors. The greatest uncertainties included laws and governance of marine use of the Arctic Ocean, changes in global commodity prices and trade dynamics, degree of Arctic state cooperation, multiple‐use conflicts, climate change severity and variability, role of the marine insurance industry, an Arctic maritime disaster, new resource discoveries, and advances in marine technologies. Although climate change and reductions in sea ice were believed to be important in providing improved marine access and potentially longer navigation seasons, economic factors such as natural resource development and regional trade were viewed as being the primary drivers of increased Arctic shipping activity in the future.
In their summary of findings, the AMSA authors noted that the future of Arctic shipping remains largely uncertain and quantitative predictions of future shipping activity are unavailable. Still, their findings indicated that increases in nearly all types of Arctic shipping—including fishing operations, transport of goods into and resources out of the Arctic, commercial icebreaking, scientific explorations, and marine tourism—were either “plausible” or “anticipated” in the future (Arctic Council 2009). Arctic shipping was expected to remain overwhelmingly destinational (for regional trade) through 2020, with trans‐Arctic voyages only being plausible on an experimental basis during some summers. The report stated that recent offshore leases in the Beaufort and Chukchi Seas may lead to increased marine traffic in the Bering Strait region, which may require formally established vessel routing measures. The NWP is not expected to become a viable trans‐Arctic route through 2020, but destinational shipping is anticipated to increase in the Canadian Arctic. The annual navigation season for light ice‐class ships was projected to increase from 2 to 4 months for the NWP and from 3 to 6 months for the NSR by 2100 (Khon et al. 2010). Compared to transit through the Suez Canal, use of the NSR was projected to result in economic savings of 15% per year in shipping costs by the end of the century (Khon et al. 2010). Large investments in resource development and transport made in the Barents Sea may stimulate decadal increases in shipping along the western portion of the NSR. A lack of major ports and other maritime infrastructure were seen as being significant limiting factors to future Arctic marine operations. Finally, it was believed that future socio‐economic responses to climate change (e.g., mandatory emission controls) would very likely impact all elements of future Arctic shipping activity (Arctic Council 2009).
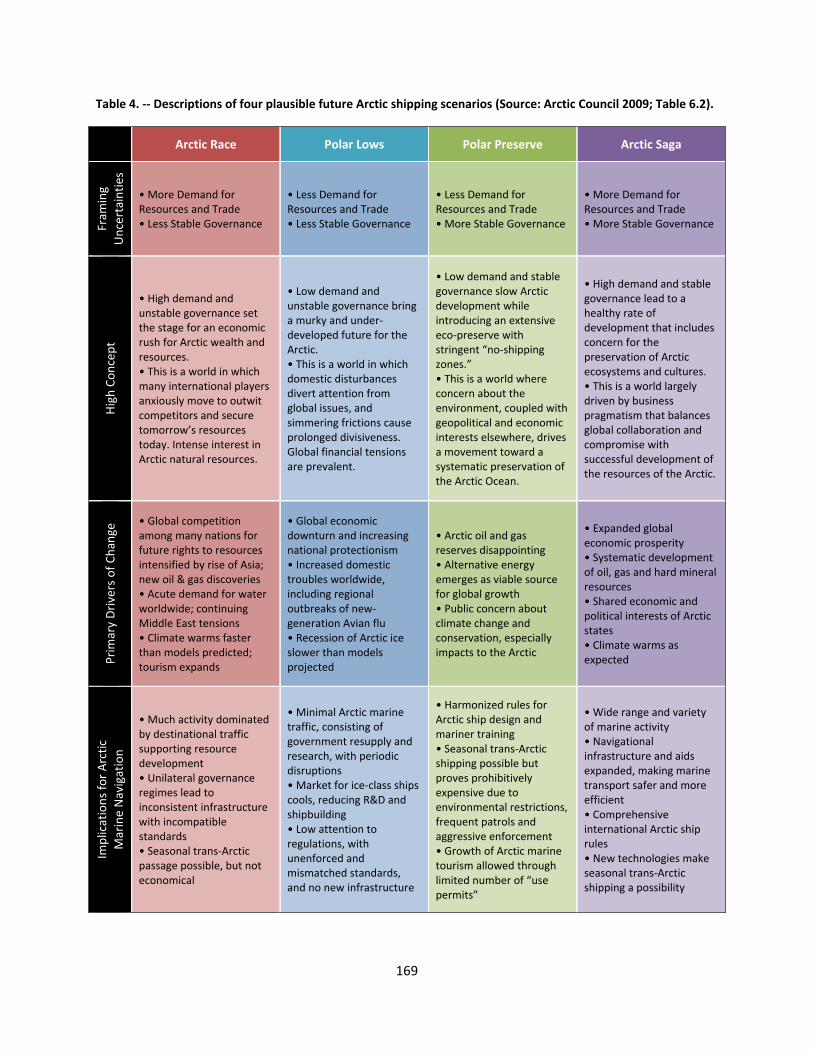
169
Table 4. ‐‐ Descriptions of four plausible future Arctic shipping scenarios (Source: Arctic Council 2009; Table 6.2). Arctic Race Polar Lows Polar Preserve Arctic Saga
Fram
ing
Uncertaintie
s
• More Demand for Resources and Trade • Less Stable Governance
• Less Demand for Resources and Trade • Less Stable Governance
• Less Demand for Resources and Trade • More Stable Governance
• More Demand for Resources and Trade • More Stable Governance
High Co
ncep
t
• High demand and unstable governance set the stage for an economic rush for Arctic wealth and resources. • This is a world in which many international players anxiously move to outwit competitors and secure tomorrow’s resources today. Intense interest in Arctic natural resources.
• Low demand and unstable governance bring a murky and under‐developed future for the Arctic. • This is a world in which domestic disturbances divert attention from global issues, and simmering frictions cause prolonged divisiveness. Global financial tensions are prevalent.
• Low demand and stable governance slow Arctic development while introducing an extensive eco‐preserve with stringent “no‐shipping zones.” • This is a world where concern about the environment, coupled with geopolitical and economic interests elsewhere, drives a movement toward a systematic preservation of the Arctic Ocean.
• High demand and stable governance lead to a healthy rate of development that includes concern for the preservation of Arctic ecosystems and cultures. • This is a world largely driven by business pragmatism that balances global collaboration and compromise with successful development of the resources of the Arctic.
Prim
ary Drivers of C
hange • Global competition
among many nations for future rights to resources intensified by rise of Asia; new oil & gas discoveries • Acute demand for water worldwide; continuing Middle East tensions • Climate warms faster than models predicted; tourism expands
• Global economic downturn and increasing national protectionism • Increased domestic troubles worldwide, including regional outbreaks of new‐ generation Avian flu • Recession of Arctic ice slower than models projected
• Arctic oil and gas reserves disappointing • Alternative energy emerges as viable source for global growth • Public concern about climate change and conservation, especially impacts to the Arctic
• Expanded global economic prosperity • Systematic development of oil, gas and hard mineral resources • Shared economic and political interests of Arctic states • Climate warms as expected
Implications fo
r Arctic
Marine Navigation
• Much activity dominated by destinational traffic supporting resource development • Unilateral governance regimes lead to inconsistent infrastructure with incompatible standards • Seasonal trans‐Arctic passage possible, but not economical
• Minimal Arctic marine traffic, consisting of government resupply and research, with periodic disruptions • Market for ice‐class ships cools, reducing R&D and shipbuilding • Low attention to regulations, with unenforced and mismatched standards, and no new infrastructure
• Harmonized rules for Arctic ship design and mariner training • Seasonal trans‐Arctic shipping possible but proves prohibitively expensive due to environmental restrictions, frequent patrols and aggressive enforcement • Growth of Arctic marine tourism allowed through limited number of “use permits”
• Wide range and variety of marine activity • Navigational infrastructure and aids expanded, making marine transport safer and more efficient • Comprehensive international Arctic ship rules • New technologies make seasonal trans‐Arctic shipping a possibility

170
4.2.5.5.1.3 Potential threats to ringed seals from shipping activities
4.2.5.5.1.3.1 Oil spills
The most significant threat posed by shipping activities to ringed seals in the Arctic is the accidental or illegal discharge of oil or other toxic substances carried by ships due to their immediate and potentially long‐term effects on individual animals, populations, food webs, and the environment (Arctic Council 2009). This threat is magnified by the lack of accident response resources in the Arctic, the lack of effective techniques for containing or cleaning up spilled oil under ice or in broken ice, and the challenges of conducting a rapid, effective spill response in a region where weather is often severe, daylight may be limited, and accidents may happen in very remote locations (AMAP 2007). The potential threat of oil spills to ringed seals was previously reviewed in the section on oil and gas exploration, development, and production (Section 4.2.5.2.2.2). The regular and legal discharge of oily sludge, garbage, and other debris from ships also poses significant risks to ringed seals since these types of pollution are more common and widespread than accidents or illegal discharges and can have similar immediate and long‐term impacts on individuals, communities, and the environment (Arctic Council 2009).
4.2.5.5.1.3.2 Noise and physical disturbance
All vessels produce sound during operation, which when propagated at certain frequencies and intensities can alter the normal behavior of marine mammals, mask their underwater communications and other uses of sound, cause them to avoid noisy areas, and in extreme cases (e.g., high‐powered sonar), damage their auditory systems and even cause death (Marine Mammal Commission 2007, Arctic Council 2009, OSPAR Commission 2009). The mere presence and movements of ships in the vicinity of seals can also cause disturbance of their normal behaviors (Jansen et al. 2010) and potentially cause ringed seals to abandon their preferred breeding habitats in areas with high traffic (Smiley and Milne 1979, Mansfield 1983). Vessel strikes are probably not a threat to ringed seals except in the case of icebreakers, which could potentially crush individuals (particularly young pups and perhaps their protective mothers) while they occupy their subnivean lairs in the springtime (Smiley and Milne 1979, Ministry of Agriculture and Forestry 2007). Pups have a greater potential for heat loss than adults, and thus, would be more prone to incur energetic costs of increased time in the water if vessel disturbance became a frequent event. Ringed seals pups may also be at risk of abandonment by their mothers as a result of disturbance by icebreakers (Smiley and Milne 1979). The opening of channels through the ice by icebreaking vessels could also impact ringed seals by altering the natural dynamics of the ice (Smith 1987) or by providing additional access to open‐water leads, if only temporarily, which could have both positive and negative effects on ringed seals (Smiley and Milne 1979, Mansfield 1983).
4.2.5.5.1.3.3 Indirect effects
Ships contribute significantly to global climate change and air pollution through their emissions of GHGs, aerosols, nitrogen and sulfur oxides, carbon monoxide, and particulate matter. The release and deposition of black carbon in the Arctic is of particular concern because of the effect it has on reducing

171
the albedo of sea ice and snow cover, which in turn increases the rate of melting significantly (ACIA 2005, Arctic Council 2009). The introduction of exotic species into the Arctic by ships could also affect ringed seals, although likely in a more indirect manner, such as through changes in the lower trophic levels of their food webs (Arctic Council 2009).
4.2.5.5.1.4 Shipping threat assessment
Current and future shipping activities in the Arctic pose varying levels of threat to ringed seals depending on the type and intensity of the shipping activity and its degree of spatial and temporal overlap with ringed seals. These factors are inherently difficult to know or predict. There is no one scenario of global supply and demand that seems most likely (ECON 2007), and knowledge about present impacts is even limited because few are witnessed, documented, or studied. Nonetheless, based on our knowledge of ringed seal populations and shipping activity in the Arctic, some general assessments can be made.
Ringed seal densities vary with many factors (reviewed by Reeves 1998), however, they are often reported to be widely distributed in low densities (averaging about 1‐2 seals/km2 in “good” habitats (Kovacs 2007)) and rarely congregate in large numbers as many other pinniped species do. The dispersed distribution may help mitigate the risks of localized shipping threats (e.g., oil spills or physical disturbance), since the impacts from such events would be less likely to affect large numbers of seals. The fact that nearly all shipping activity in the Arctic (with the exception of icebreaking) purposefully avoids areas of ice and primarily occurs during the ice‐free or low‐ice seasons also helps to mitigate the risks of shipping to ringed seals since this species is closely associated with ice at nearly all times of the year and especially during the whelping, breeding, and molting periods when the seals (especially young pups) may be most vulnerable to shipping impacts (Smith 1987).
Icebreakers pose special risks to ringed seals since they are capable of operating year‐round in all but the heaviest ice conditions and are often used to escort other types of vessels (e.g., tankers and bulk carriers) through ice‐covered areas. Furthermore, new classes of ships are being designed that serve the dual roles of both tanker/carrier and icebreaker (Arctic Council 2009). Therefore, if icebreaking activities increase in the Arctic in the future as expected, the likelihood of negative impacts (e.g., oil spills, pollution, noise, disturbance, and habitat alteration) occurring in ice‐covered areas where ringed seals reside will likely also increase.
Spatially, ringed seals may be at the greatest risk from shipping threats in areas of the Arctic where geographic constriction concentrates seals and shipping activity into confined areas, such as the Bering Strait, Hudson Strait, Lancaster Sound, Pechora Sea, and Kara Port (Arctic Council 2009). Effective governance and regulations will be necessary to mitigate the threats of shipping to ringed seals and other marine species in the Arctic, especially in areas and during periods of high risk (Arctic Council 2009). In a preliminary assessment of future threats to Arctic marine mammals, Huntington (2009) considered shipping to be a low level threat with modest impacts that should be amenable to effective regulation. Indeed, shipping impacts alone may comprise a low risk to entire populations, but when

172
combined with the effects related to diminishing ice cover, such as increasingly denser aggregations, the impacts may be magnified and may play an important role in affecting the future health of populations.
The following three sections review the types and levels of shipping activity in the Sea of Okhotsk, Baltic Sea, and Lakes Ladoga and Saimaa, as well as any specific shipping threat information that could be found for these regions. The potential threats in these regions are largely the same as in the Arctic, so will not be repeated. Threats specific to these regions are summarized below. Two obvious but important distinctions are that these bodies of water (particularly the gulfs of the Baltic Sea and Lakes Ladoga and Saimaa) are geographically smaller and more confined than many areas where the Arctic subspecies lives and contain much smaller populations of ringed seals. Therefore, shipping impacts and ringed seals are more likely to overlap spatially in these regions, and a single accident (e.g., a large oil spill) could potentially impact these smaller populations severely.
4.2.5.5.2 Sea of Okhotsk
Though few details are available regarding actual shipping levels in the Sea of Okhotsk, resource development over the last decade stands out as a likely significant contributor (Reeves et al. 2005). It is clear that considerable ship traffic is needed to support present oil and gas operations, primarily off the northeastern coast of Sakhalin Island and the western coast of the Kamchatka Peninsula (Lapko and Radchenko 2000, Dulepova and Radchencko 2004), with future developments pointing to an ever‐growing shipping industry to support the area’s energy and minerals commerce (United Nations Environment Programme 2006). Large‐scale commercial fishing occurs in many parts of the sea, accounting for half of the total Russian fish harvest from the Pacific Ocean (Dulepova and Radchencko 2004). Recent analysis of satellite imagery revealed fairly regular, large offshore oil slicks in the Sea of Okhotsk (Ivanov and Zatyagalova 2008). Sources of the oil could not be identified, but many of the largest slicks were linear in form and believed to have been left by passing ships. Fairly regular dumping of engine room and bilge water by fishing and factory processor ships is also believed to be a significant source of pollution (Ivanov and Zatyagalova 2008).
The Russian government and international companies have focused primarily on developing oil and gas fields near Sakhalin and less on developing the resources and technical expertise to respond adequately to large or remote oil spills. For example, many oil tankers operating in the region do not have double hulls (Lawn et al. 1999, United Nations Environment Programme 2006). Although Russian waters are covered by the MARPOL treaty, inadequate port reception facilities, high costs of waste disposal, inaction against violators, and an inability to detect illegal dumping are believed to result in significant waste discharges at sea (Schei and Brubaker 2006, Ivanov and Zatyagalova 2008). An accident at an oil production complex resulted in a large (3.5 ton) spill in 1999 (Lapko and Radchenko 2000). In winter 2009, an unknown quantity of oil associated with a tanker fouled 3 km of coastline and hundreds of birds in Aniva Bay (Associated Foreign Press 2009). Navigation in this sea is reported to be difficult or impossible during winter due to heavy ice conditions, yet shipping still occurs year‐round at the largest ports with the help of icebreakers (Kommersant 2004). Winter shipping activities in the southern Sea of Okhotsk are expected to increase considerably as oil and gas production pushes the development and

173
use of new classes of icebreaking ships, thereby increasing the potential for shipping accidents and spills in the ice‐covered regions of this sea (Shumei et al. 2001, Choi 2008).
4.2.5.5.3 Baltic Sea
The Baltic Sea is one of the most heavily trafficked shipping areas in the world, with more than 2,000 large ships (including about 200 oil tankers) sailing on its waters on an average day (Arctic Council 2009). Additionally, some of the world’s biggest ferries transport goods and people between Sweden and Finland every day, and several other ferry lines connect the other states of the Baltic proper. Up to 2,000 fishing boats can be at sea on an average day during most times of the year, and during summer, large numbers of cruise ships enter the Baltic Sea to visit the many cultural centers along its shores (Arctic Council 2009). Both the number and size of ships (especially oil tankers) have grown in recent years, and the amount of oil transported in the Baltic (especially from the Gulf of Finland) has increased significantly since 2000 (HELCOM 2006a). Oil transportation is expected to increase by 40%, and cargo and container traffic is expected to triple by 2017 (Baltic Sea 2020 2009).
There were 374 ship accidents reported in the Baltic during 2000‐2004, mostly from groundings and collisions, and 8% of these resulted in some type of pollution (HELCOM 2006a). Shipping traffic, accidents, and oil spills appeared to be highest in the southwestern Baltic Sea (around Denmark), followed by the Gulf of Finland, and appeared to be low in the Gulfs of Bothnia and Riga (HELCOM 2006a). The risk of oil exposure for seals living in the Baltic Sea was considered to be greatest in the Gulf of Finland, where oil shipping routes pass through ringed seal pupping areas as well as close to rocks and islets where seals come out of the water (Ministry of Agriculture and Forestry 2007). Icebreaking during the winter was considered to be the most significant marine traffic factor for seals in the Baltic Sea, especially in Bothnia Bay where the largest harbors have been kept open through winter since the 1980s (Ministry of Agriculture and Forestry 2007). Heavy icebreaking activity reportedly breaks up the normally firm ice cover in Bothnia Bay, creating artificial drift‐ice conditions. Icebreakers may destroy ringed seal lairs and kill pups which lie on or near their paths, however, the incidence is believed to be low since ringed seal mothers may avoid shipping lanes when choosing locations to build their birthing lairs (Sipilä 1990, Ministry of Agriculture and Forestry 2007). During the ice‐free season, the dredging and channeling of shipping lanes was thought to have the greatest impact on the marine environment by churning up the sediments and mixing nutrients and contaminants back into the water column. Yachting and other marine recreational activities (such as fishing and snowmobiling on the shorefast ice during winter) have been increasing in recent years, especially in the Baltic Sea’s archipelagos and coastal areas. Those activities are not thought to have a large impact on Baltic seal populations in general, but may cause significant disturbance on a local level (Ministry of Agriculture and Forestry 2007).
4.2.5.5.4 Lakes Ladoga and Saimaa
Lakes Ladoga and Saimaa are connected to the Baltic Sea and other bodies of water via a network of rivers and canals and are used as waterways to transport people, resources, and cargo throughout the Baltic region. Additional boat traffic is associated with commercial and subsistence fishing operations on these lakes (Sipilä and Hyvärinen 1998).

174
At least two large fuel spills from ships occurred on Lake Ladoga in 1997, and pollution and disturbance from increasing vessel traffic have been mentioned as concerns there (Seal Conservation Society 2001). Boating activity was reported to cause mass disturbance events at large, on‐shore resting sites in the Valaam Archipelago of Lake Ladoga (Agafonova et al. 2007). In Lake Saimaa, icebreaking and the regular use of shipping lanes during winter was thought to reduce the availability of suitable breeding habitat for ringed seals and cause some level of disturbance, but there were no observations that winter shipping had increased the mortality of young pups (Sipilä 2003). Shipping‐related activities (other than accidental bycatch in fishing gear; see Section 4.2.5.4.1) were not considered to pose significant risks to the conservation status of ringed seals in either Lake Ladoga or Saimaa (Sipilä and Hyvärinen 1998, Sipilä et al. 2002, Sipilä 2003).
4.2.6 Threats Assessment
The BRT members’ assessments of the significance of the threats to each ringed seal subspecies’ persistence were summarized in numerical scores. The primary threats were grouped by ESA Section 4(a)(1) factors (excluding Inadequacy of Existing Regulatory Mechanisms), and each individual threat was scored—in consideration of their severity, geographic scope, and likelihood of occurrence—as follows: 1 = low or zero significance, 2 = moderate significance, 3 = high significance, and 4 = very high significance. For each threat, the level of certainty (i.e., amount and quality of available information) in the threat assessment was also scored on the same scale. Then, each BRT member assigned an overall score for each ESA Section 4(a)(1) factor, in recognition that multiple threats may have cumulative effects that sum or magnify the overall threat for the factor. The average score and the range of scores among BRT members are presented for each threat and ESA Section 4(a)(1) factor in Tables 5‐9.
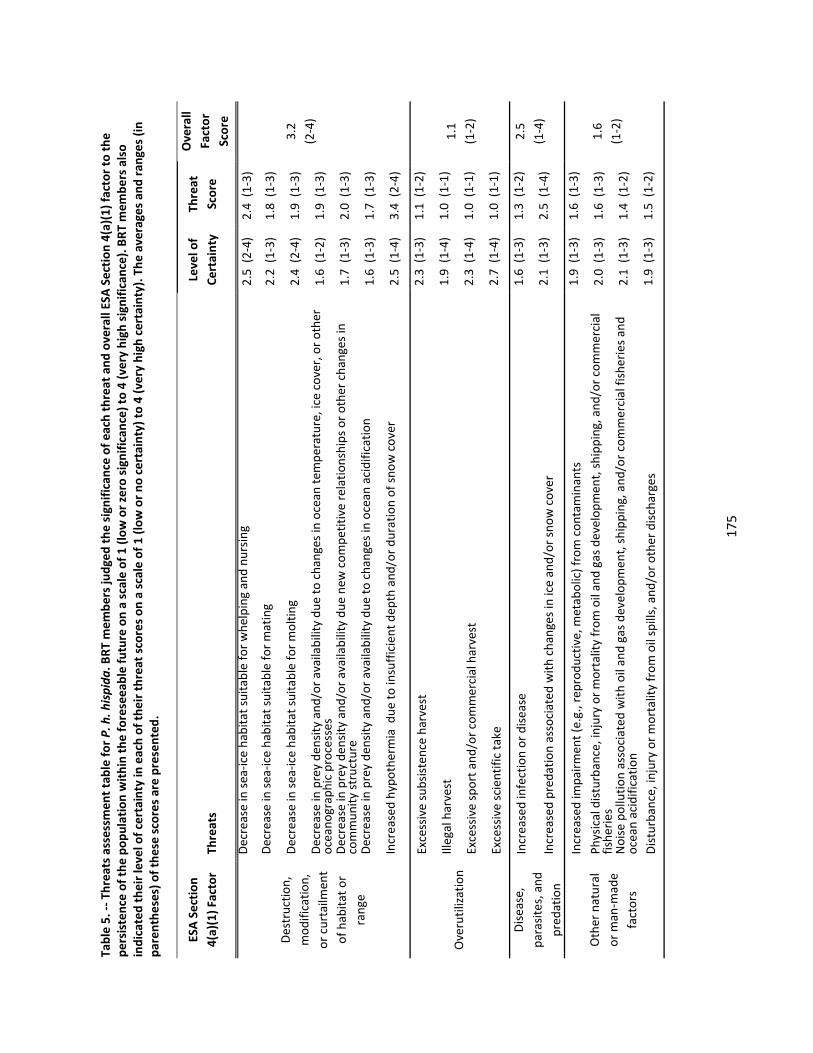
175
Table 5. ‐‐ Threats assessm
ent table for P. h. h
ispida
. BRT
mem
bers ju
dged
the significance of each threat and
overall ESA Section
4(a)(1) fa
ctor to the
persistence of th
e po
pulation
within the foreseeable future on a scale of 1 (low
or zero significan
ce) to 4 (very high
significan
ce). BRT
mem
bers also
indicated their level of certainty in
each of th
eir threat scores on
a scale of 1
(low
or no
certainty) to 4 (very high
certainty). Th
e averages and
ran
ges (in
pa
renthe
ses) of the
se scores are presen
ted.
ESA Section
4(a)(1) Factor
Threats
Level of
Certainty
Threat
Score
Overall
Factor
Score
Destructio
n,
mod
ificatio
n,
or curtailm
ent
of habita
t or
range
Decrease in sea‐ice habitat suitable for w
helping and nu
rsing
2.5 (2
‐4)
2.4 (1
‐3)
3.2
(2‐4)
Decrease in sea‐ice habitat suitable for m
ating
2.2 (1
‐3)
1.8 (1
‐3)
Decrease in sea‐ice habitat suitable for m
oltin
g2.4 (2
‐4)
1.9 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
tempe
rature, ice cover, or othe
r oceano
graphic processes
1.6 (1
‐2)
1.9 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e ne
w com
petitive relatio
nships or othe
r changes in
commun
ity structure
1.7 (1
‐3)
2.0 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
acidification
1.6 (1
‐3)
1.7 (1
‐3)
Increased hypo
thermia due
to insufficient dep
th and
/or d
uration of sno
w cover
2.5 (1
‐4)
3.4 (2
‐4)
Overutilization
Excessive subsistence harvest
2.3 (1
‐3)
1.1 (1
‐2)
1.1
(1‐2)
Illegal harvest
1.9 (1
‐4)
1.0(1‐1)
Excessive sport a
nd/or com
mercial harvest
2.3 (1
‐4)
1.0 (1
‐1)
Excessive scientific take
2.7 (1
‐4)
1.0 (1
‐1)
Disease,
parasites, and
pred
ation
Increased infection or disease
1.6 (1
‐3)
1.3 (1
‐2)
2.5
(1‐4)
Increased predation associated
with
changes in
ice and/or sno
w cover
2.1 (1
‐3)
2.5 (1
‐4)
Other natural
or m
an‐m
ade
factors
Increased im
pairmen
t (e.g., rep
rodu
ctive, m
etabolic) from con
taminants
1.9 (1
‐3)
1.6 (1
‐3)
1.6
(1‐2)
Physical disturbance, injury or m
ortality from
oil and gas de
velopm
ent, shipp
ing, and
/or commercial
fishe
ries
2.0 (1
‐3)
1.6 (1
‐3)
Noise pollutio
n associated
with
oil and gas de
velopm
ent, shipp
ing, and
/or commercial fisheries and
ocean acidificatio
n 2.1 (1
‐3)
1.4 (1
‐2)
Disturbance, injury or m
ortality from
oil spills, and
/or o
ther discharges
1.9 (1
‐3)
1.5 (1
‐2)
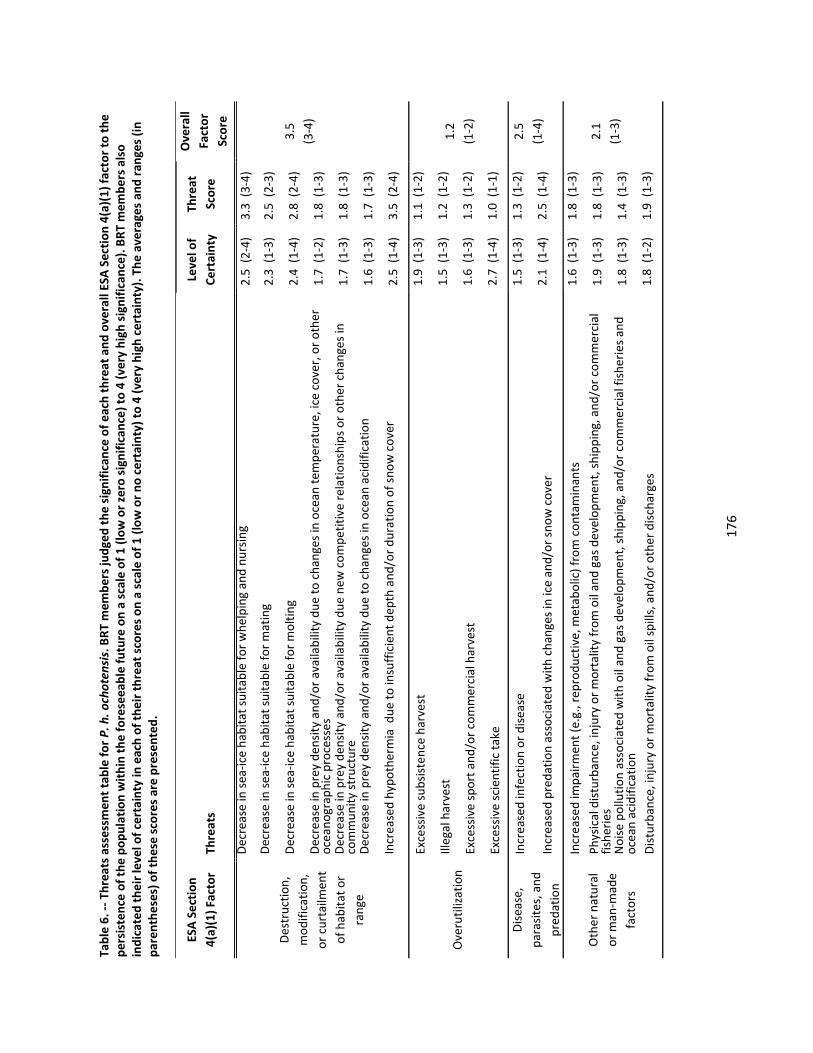
176
Table 6. ‐‐ Threats assessm
ent table for P. h. o
choten
sis. BRT
mem
bers judged
the
significan
ce of e
ach threat and
overall ESA Section
4(a)(1) fa
ctor to
the
persistence of th
e po
pulation
within the foreseeable future on a scale of 1 (low
or zero significan
ce) to 4 (very high
significan
ce). BRT
mem
bers also
indicated their level of certainty in
each of th
eir threat scores on
a scale of 1
(low
or no
certainty) to 4 (very high
certainty). Th
e averages and
ran
ges (in
pa
renthe
ses) of the
se scores are presen
ted.
ESA Section
4(a)(1) Factor
Threats
Level of
Certainty
Threat
Score
Overall
Factor
Score
Destructio
n,
mod
ificatio
n,
or curtailm
ent
of habita
t or
range
Decrease in sea‐ice habitat suitable for w
helping and nu
rsing
2.5 (2
‐4)
3.3 (3
‐4)
3.5
(3‐4)
Decrease in sea‐ice habitat suitable for m
ating
2.3 (1
‐3)
2.5 (2
‐3)
Decrease in sea‐ice habitat suitable for m
oltin
g2.4 (1
‐4)
2.8 (2
‐4)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
tempe
rature, ice cover, or othe
r oceano
graphic processes
1.7 (1
‐2)
1.8 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e ne
w com
petitive relatio
nships or othe
r changes in
commun
ity structure
1.7 (1
‐3)
1.8 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
acidification
1.6 (1
‐3)
1.7 (1
‐3)
Increased hypo
thermia due
to insufficient dep
th and
/or d
uration of sno
w cover
2.5 (1
‐4)
3.5 (2
‐4)
Overutilization
Excessive subsistence harvest
1.9 (1
‐3)
1.1 (1
‐2)
1.2
(1‐2)
Illegal harvest
1.5 (1
‐3)
1.2 (1
‐2)
Excessive sport a
nd/or com
mercial harvest
1.6 (1
‐3)
1.3 (1
‐2)
Excessive scientific take
2.7 (1
‐4)
1.0 (1
‐1)
Disease,
parasites, and
pred
ation
Increased infection or disease
1.5 (1
‐3)
1.3 (1
‐2)
2.5
(1‐4)
Increased predation associated
with
changes in
ice and/or sno
w cover
2.1 (1
‐4)
2.5 (1
‐4)
Other natural
or m
an‐m
ade
factors
Increased im
pairmen
t (e.g., rep
rodu
ctive, m
etabolic) from con
taminants
1.6 (1
‐3)
1.8 (1
‐3)
2.1
(1‐3)
Physical disturbance, injury or m
ortality from
oil and gas de
velopm
ent, shipp
ing, and
/or commercial
fishe
ries
1.9 (1
‐3)
1.8 (1
‐3)
Noise pollutio
n associated
with
oil and gas de
velopm
ent, shipp
ing, and
/or commercial fisheries and
ocean acidificatio
n 1.8 (1
‐3)
1.4 (1
‐3)
Disturbance, injury or m
ortality from
oil spills, and
/or o
ther discharges
1.8 (1
‐2)
1.9 (1
‐3)
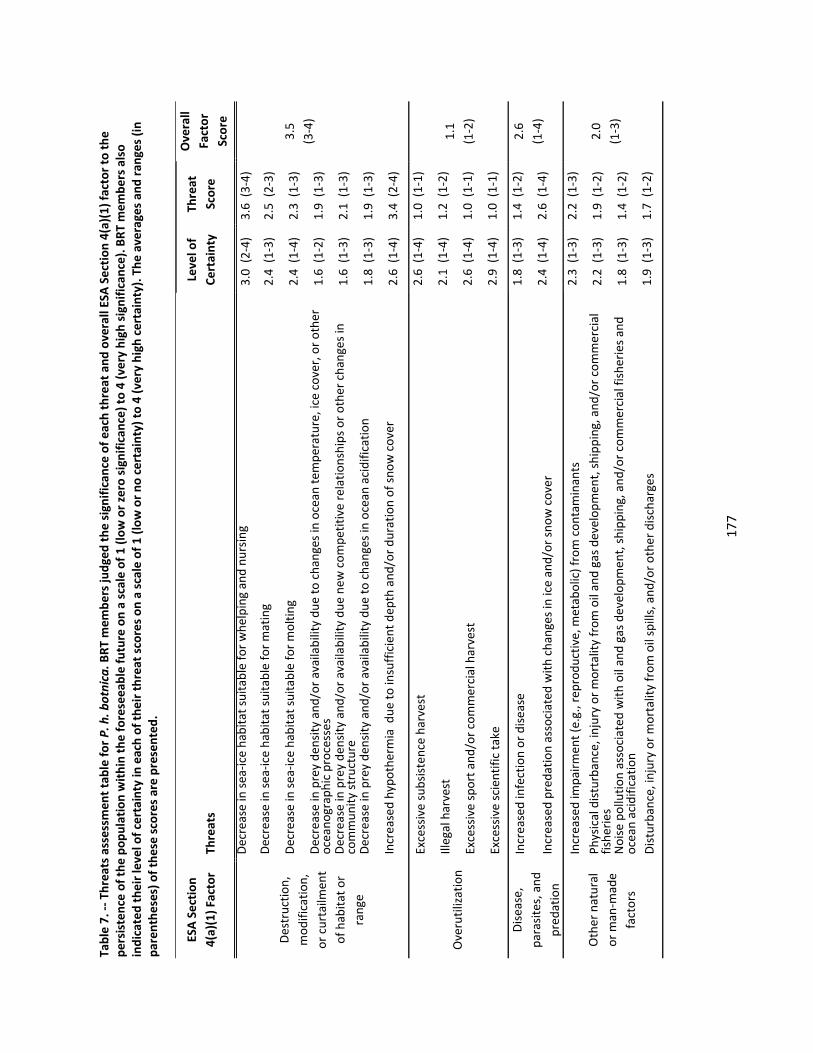
177
Table 7. ‐‐ Threats assessm
ent table for P. h. b
otnica. B
RT m
embe
rs ju
dged
the
significan
ce of e
ach threat and
overall ESA Section
4(a)(1) fa
ctor to the
persistence of th
e po
pulation
within the foreseeable future on a scale of 1 (low
or zero significan
ce) to 4 (very high
significan
ce). BRT
mem
bers also
indicated their level of certainty in
each of th
eir threat scores on
a scale of 1
(low
or no
certainty) to 4 (very high
certainty). Th
e averages and
ran
ges (in
pa
renthe
ses) of the
se scores are presen
ted.
ESA Section
4(a)(1) Factor
Threats
Level of
Certainty
Threat
Score
Overall
Factor
Score
Destructio
n,
mod
ificatio
n,
or curtailm
ent
of habita
t or
range
Decrease in sea‐ice habitat suitable for w
helping and nu
rsing
3.0(2‐4)
3.6 (3
‐4)
3.5
(3‐4)
Decrease in sea‐ice habitat suitable for m
ating
2.4 (1
‐3)
2.5 (2
‐3)
Decrease in sea‐ice habitat suitable for m
oltin
g2.4 (1
‐4)
2.3 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
tempe
rature, ice cover, or othe
r oceano
graphic processes
1.6 (1
‐2)
1.9 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e ne
w com
petitive relatio
nships or othe
r changes in
commun
ity structure
1.6 (1
‐3)
2.1 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
ocean
acidification
1.8 (1‐3)
1.9 (1
‐3)
Increased hypo
thermia due
to insufficient dep
th and
/or d
uration of sno
w cover
2.6 (1
‐4)
3.4 (2
‐4)
Overutilization
Excessive subsistence harvest
2.6 (1
‐4)
1.0 (1
‐1)
1.1
(1‐2)
Illegal harvest
2.1 (1
‐4)
1.2 (1
‐2)
Excessive sport a
nd/or com
mercial harvest
2.6 (1
‐4)
1.0 (1
‐1)
Excessive scientific take
2.9 (1
‐4)
1.0 (1
‐1)
Disease,
parasites, and
pred
ation
Increased infection or disease
1.8 (1
‐3)
1.4 (1
‐2)
2.6
(1‐4)
Increased predation associated
with
changes in
ice and/or sno
w cover
2.4 (1
‐4)
2.6 (1
‐4)
Other natural
or m
an‐m
ade
factors
Increased im
pairmen
t (e.g., rep
rodu
ctive, m
etabolic) from con
taminants
2.3 (1
‐3)
2.2 (1
‐3)
2.0
(1‐3)
Physical disturbance, injury or m
ortality from
oil and gas de
velopm
ent,shipping, and
/or commercial
fishe
ries
2.2 (1
‐3)
1.9 (1
‐2)
Noise pollutio
n associated
with
oil and gas de
velopm
ent, shipp
ing, and
/or commercial fisheries and
ocean acidificatio
n 1.8 (1
‐3)
1.4 (1
‐2)
Disturbance, injury or m
ortality from
oil spills, and
/or o
ther discharges
1.9 (1
‐3)
1.7 (1
‐2)
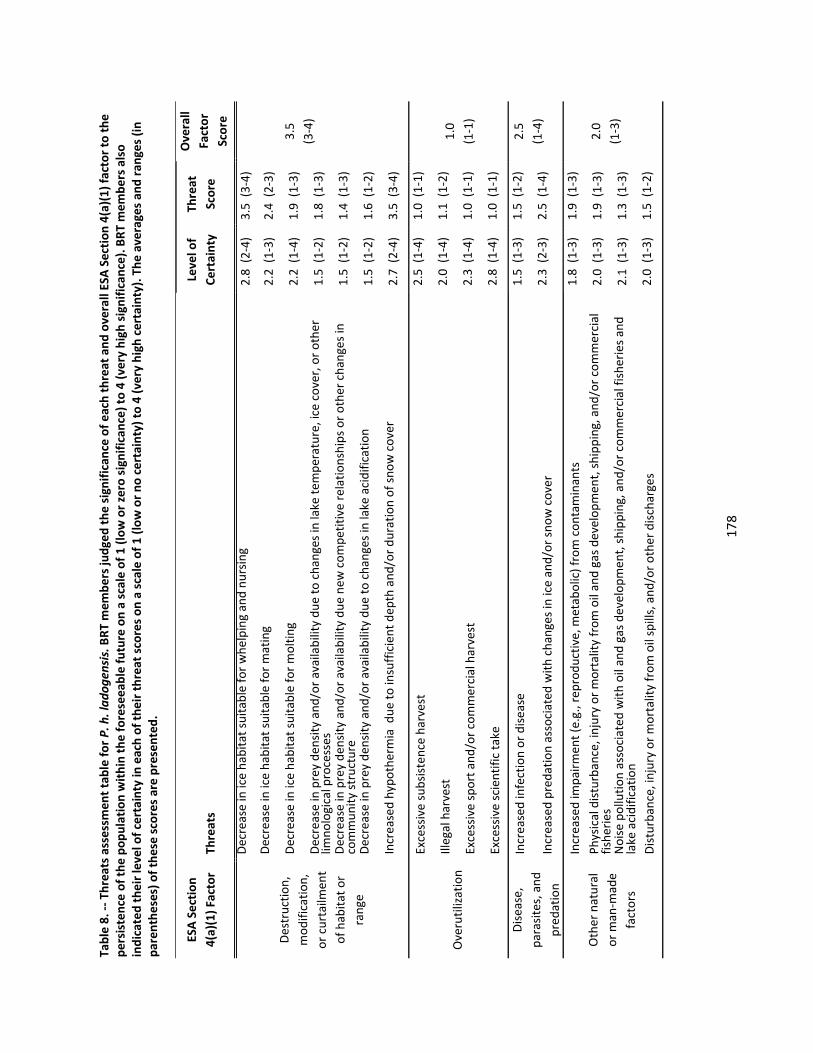
178
Table 8. ‐‐ Threats assessm
ent table for P. h. lad
ogensis. BRT
mem
bers judged
the
significan
ce of e
ach threat and
overall ESA Section
4(a)(1) fa
ctor to
the
persistence of th
e po
pulation
within the foreseeable future on a scale of 1 (low
or zero significan
ce) to 4 (very high
significan
ce). BRT
mem
bers also
indicated their level of certainty in
each of th
eir threat scores on
a scale of 1
(low
or no
certainty) to 4 (very high
certainty). Th
e averages and
ran
ges (in
pa
renthe
ses) of the
se scores are presen
ted.
ESA Section
4(a)(1) Factor
Threats
Level of
Certainty
Threat
Score
Overall
Factor
Score
Destructio
n,
mod
ificatio
n,
or curtailm
ent
of habita
t or
range
Decrease in ice habitat suitable for whe
lping and nu
rsing
2.8 (2
‐4)
3.5 (3
‐4)
3.5
(3‐4)
Decrease in ice habitat suitable for mating
2.2 (1
‐3)
2.4 (2
‐3)
Decrease in ice habitat suitable for moltin
g2.2 (1
‐4)
1.9 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
lake
tempe
rature, ice cover, or othe
r lim
nological processes
1.5 (1
‐2)
1.8 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e ne
w com
petitive relatio
nships or othe
r changes in
commun
ity structure
1.5 (1
‐2)
1.4 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
lake
acidificatio
n 1.5 (1
‐2)
1.6 (1
‐2)
Increased hypo
thermia due
to insufficient dep
th and
/or d
uration of sno
w cover
2.7 (2
‐4)
3.5 (3
‐4)
Overutilization
Excessive subsistence harvest
2.5 (1
‐4)
1.0 (1
‐1)
1.0
(1‐1)
Illegal harvest
2.0 (1
‐4)
1.1 (1
‐2)
Excessive sport a
nd/or com
mercial harvest
2.3 (1
‐4)
1.0 (1
‐1)
Excessive scientific take
2.8 (1
‐4)
1.0 (1
‐1)
Disease,
parasites, and
pred
ation
Increased infection or disease
1.5 (1
‐3)
1.5 (1
‐2)
2.5
(1‐4)
Increased predation associated
with
changes in
ice and/or sno
w cover
2.3 (2
‐3)
2.5 (1
‐4)
Other natural
or m
an‐m
ade
factors
Increased im
pairmen
t (e.g., rep
rodu
ctive, m
etabolic) from con
taminants
1.8 (1
‐3)
1.9 (1
‐3)
2.0
(1‐3)
Physical disturbance, injury or m
ortality from
oil and gas de
velopm
ent, shipp
ing, and
/or commercial
fishe
ries
2.0 (1
‐3)
1.9 (1
‐3)
Noise pollutio
n associated
with
oil and gas de
velopm
ent, shipp
ing, and
/or commercial fisheries and
lake acidification
2.1 (1
‐3)
1.3 (1
‐3)
Disturbance, injury or m
ortality from
oil spills, and
/or o
ther discharges
2.0 (1
‐3)
1.5 (1
‐2)
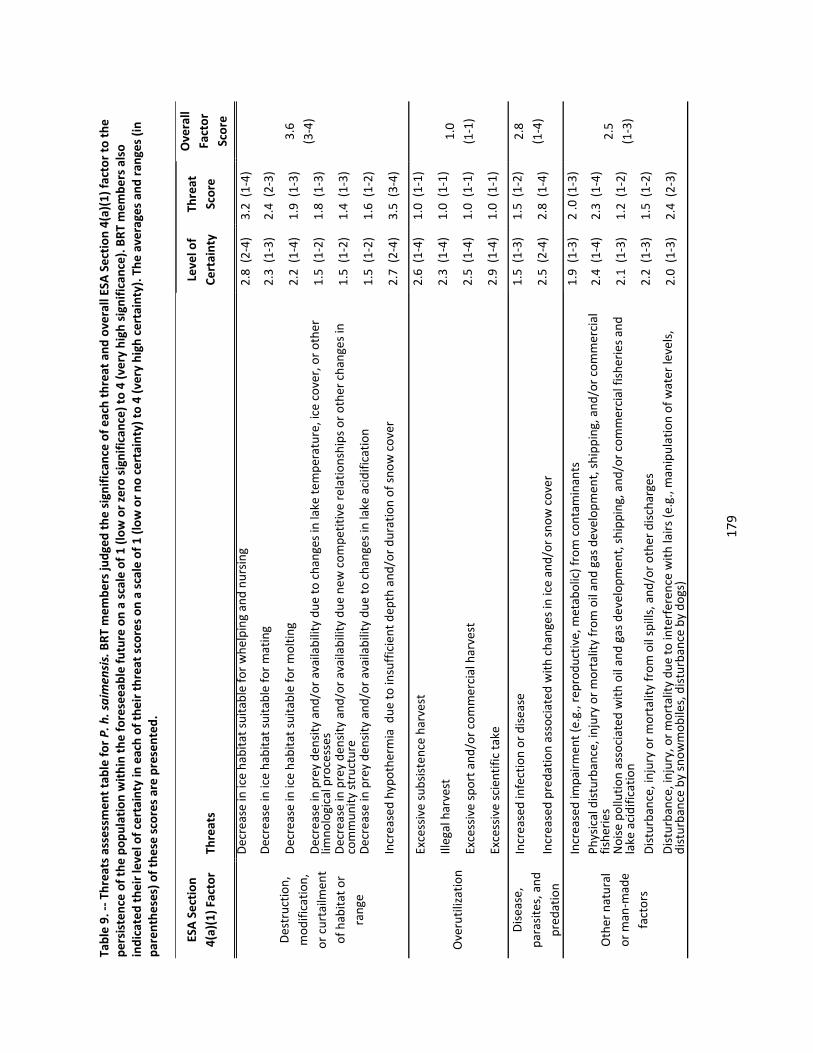
179
Table 9. ‐‐ Threats assessm
ent table for P. h. saimen
sis. BRT
mem
bers judged
the significance of each threat and
overall ESA Section
4(a)(1) fa
ctor to the
persistence of th
e po
pulation
within the foreseeable future on a scale of 1 (low
or zero significan
ce) to 4 (very high
significan
ce). BRT
mem
bers also
indicated their level of certainty in
each of th
eir threat scores on
a scale of 1
(low
or no
certainty) to 4 (very high
certainty). Th
e averages and
ran
ges (in
pa
renthe
ses) of the
se scores are presen
ted.
ESA Section
4(a)(1) Factor
Threats
Level of
Certainty
Threat
Score
Overall
Factor
Score
Destructio
n,
mod
ificatio
n,
or curtailm
ent
of habita
t or
range
Decrease in ice habitat suitable for whe
lping and nu
rsing
2.8 (2
‐4)
3.2 (1
‐4)
3.6
(3‐4)
Decrease in ice habitat suitable for mating
2.3 (1
‐3)
2.4 (2
‐3)
Decrease in ice habitat suitable for moltin
g2.2 (1
‐4)
1.9 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
lake
tempe
rature, ice cover, or othe
r lim
nological processes
1.5 (1
‐2)
1.8 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e ne
w com
petitive relatio
nships or othe
r changes in
commun
ity structure
1.5 (1
‐2)
1.4 (1
‐3)
Decrease in prey de
nsity
and
/or availability du
e to changes in
lake
acidificatio
n 1.5 (1
‐2)
1.6 (1
‐2)
Increased hypo
thermia due
to insufficient dep
th and
/or d
uration of sno
w cover
2.7 (2
‐4)
3.5 (3
‐4)
Overutilization
Excessive subsistence harvest
2.6 (1
‐4)
1.0 (1
‐1)
1.0
(1‐1)
Illegal harvest
2.3 (1
‐4)
1.0 (1
‐1)
Excessive sport a
nd/or com
mercial harvest
2.5 (1
‐4)
1.0 (1
‐1)
Excessive scientific take
2.9 (1
‐4)
1.0 (1
‐1)
Disease,
parasites, and
pred
ation
Increased infection or disease
1.5 (1
‐3)
1.5 (1
‐2)
2.8
(1‐4)
Increased predation associated
with
changes in
ice and/or sno
w cover
2.5 (2
‐4)
2.8 (1
‐4)
Other natural
or m
an‐m
ade
factors
Increased im
pairmen
t (e.g., rep
rodu
ctive, m
etabolic) from con
taminants
1.9 (1
‐3)
2 .0(1‐3)
2.5
(1‐3)
Physical disturbance, injury or m
ortality from
oil and gas de
velopm
ent, shipp
ing, and
/or commercial
fishe
ries
2.4 (1
‐4)
2.3 (1
‐4)
Noise pollutio
n associated
with
oil and gas de
velopm
ent, shipp
ing, and
/or commercial fisheries and
lake acidification
2.1 (1
‐3)
1.2 (1
‐2)
Disturbance, injury or m
ortality from
oil spills, and
/or o
ther discharges
2.2 (1
‐3)
1.5 (1
‐2)
Disturbance, injury, or mortality du
e to interferen
ce with
lairs (e
.g., manipulation of water levels,
disturbance by sno
wmob
iles, disturbance by do
gs)
2.0 (1
‐3)
2.4 (2
‐3)

180
4.3 Analysis of Demographic Risks
Threats to a species’ long‐term persistence, such as those evaluated in Section 4.2 are manifested demographically as risks to its abundance; productivity; spatial structure and connectivity; and genetic and ecological diversity. These demographic risks thus provide the most direct indices or proxies of extinction risk. In this section, the current status of each of these risks is assessed in turn by responding to a set of questions adapted from McElhany et al. (2000) and incorporated into a draft NMFS interim protocol for conducting ESA status reviews (Nammack et al. 2007). Although the interim protocol itself has not yet been officially adopted as a whole, these questions are based on general conservation biology principles applicable to a wide variety of species. The BRT used these questions to guide their considerations of the broader demographic risk categories of abundance, productivity, spatial structure, and diversity.
4.3.1 Abundance
Is the species’ abundance so low that it is at imminent risk of extinction due to environmental variation or anthropogenic perturbations (of the patterns and magnitudes observed in the past and expected in the foreseeable future)?
Saimaa ringed seal – The current population of the Saimaa ringed seals—less than 300 individuals—is only 4‐7% of the estimated population size 5,000 years ago. Since 1993, the subspecies has been listed as endangered under the ESA. The subspecies no longer occurs in some portions of the lake, but is showing modest growth in other portions. On‐going losses due to entanglement in fishing nets, “lair deaths” due to human activities, and pollutants pose continuing risks of extinction (Sipilä 2003).
Ladoga ringed seal – The best current estimates of the number of Ladoga ringed seals range from 3,000‐5,000, down from a historic high of, perhaps, 20,000. Hunting has been banned since 1980, but drowning in fishing nets remains an important source of mortality. The current population is not so low as to put the subspecies at imminent risk of extinction due to the kind of environmental variation or perturbations experienced in the past. Strongly directional changes in ice and snow cover in its habitat, however, are likely to decrease the population to levels where it is at imminent risk of extinction. The managers who work most closely with the subspecies protect it as “rare” (Red Data Books of the Russian Federation) and “vulnerable” (European Mammal Assessment Team 2007).
Baltic ringed seal – The best estimate of current population size of the Baltic subspecies (10,000) indicates that it is not at imminent risk of extinction due to environmental variation or anthropogenic perturbations. Strongly directional changes in ice and snow cover in the Baltic Sea are likely to decrease the population to levels where it is at imminent risk of extinction.
Arctic ringed seal – There are no credible estimates of population size for the Arctic subspecies, but most experts would postulate that the population numbers in the millions. At present, there is no evidence that the population is so low that it is at imminent risk of extinction due to environmental

181
variation or anthropogenic perturbations. Strongly directional changes in ice and snow cover in the Arctic are likely to decrease the population, and, within the foreseeable future, it is likely that they will no longer persist in substantial portions of their range.
Okhotsk ringed seal – The best estimate of current population size of the Okhotsk subspecies (676,000) indicates that it presently is not at imminent risk of extinction due to environmental variation or anthropogenic perturbations. Low snow cover on sea ice and strongly directional changes in ice and snow cover in the Sea of Okhotsk are likely to decrease the population to levels where it is at imminent risk of extinction.
Is the species’ abundance so low, or variability in abundance so high, that it is at imminent risk of extinction due to depensatory processes?
Depensatory processes, in which per‐capita growth rate declines with decreasing abundance (opposite of density dependence), are associated with very low abundance levels and include breakdown of social structures or mating systems, failure to muster foraging groups, and failure of group defenses against predators.
Saimaa ringed seal – The very small population size of the Saimaa subspecies puts it at risk of depensatory processes.
Ladoga ringed seal – If the current, estimated population size of Ladoga ringed seals is accurate, it is large enough to likely avoid depensation. Experts familiar with the estimates, however, consider them unreliable (M. Kunnasranta, Finnish Game and Fisheries Research Institute, pers. comm., 9 July 2010). Combined with long life spans and slow reproduction, a population of the purported size is not likely to experience extreme fluctuations that could lead to depensation. The threshold for depensation in seals, however, is unknown.
Baltic ringed seal – The population of Baltic ringed seals currently is large enough to likely avoid depensation. Combined with long life spans and slow reproduction, a population of this size is not likely to experience extreme fluctuations that could lead to depensation. The threshold for depensation in seals, however, is unknown.
Arctic ringed seal – The current population of Arctic ringed seals presumably is large enough to avoid depensation. Combined with long life spans and slow reproduction, a population of this size is not likely to experience extreme fluctuations that could lead to depensation. The threshold for depensation in seals, however, is unknown.
Okhotsk ringed seal – At present, the population of Okhotsk ringed seals is large enough to likely avoid depensation. Combined with long life spans and slow reproduction, a population of this size is not likely to experience extreme fluctuations that could lead to depensation. The threshold for depensation in seals, however, is unknown.

182
Is the species’ abundance so low that its genetic diversity is at imminent risk due to inbreeding depression, loss of genetic variants, or fixation of deleterious mutations?
Saimaa ringed seal – Assuming that the subspecies does not experience further declines or become separated in to multiple breeding populations by habitat fragmentation, it should be safe from genetic effects associated with low population size. Experts on the population are not certain that the subspecies is safe from such effects, and a population genetics study is underway (M. Kunnasranta, Finnish Game and Fisheries Research Institute, pers. comm., 9 July 2010).
Ladoga ringed seal – Despite its long isolation, the Ladoga ringed seal population retains substantial genetic diversity, although somewhat lower than in the Arctic subspecies. At current population levels, it is not likely at imminent risk due to inbreeding depression, loss of genetic variants, or fixation of deleterious mutations.
Baltic ringed seal – If, as suggested by Palo (2003), there is on‐going genetic exchange with the Arctic subspecies, then the Baltic subspecies currently is not at imminent risk due to inbreeding depression, loss of genetic variants, or fixation of deleterious mutations. On the other hand, the Baltic subspecies may prove to be isolated and composed of three discrete populations (Harkonen et al. 2008). In that case, the smaller of those populations could be at risk.
Arctic ringed seal – The Arctic subspecies exhibits substantial genetic diversity. It presently is not likely at imminent risk due to inbreeding depression, loss of genetic variants, or fixation of deleterious mutations.
Okhotsk ringed seal – Genetic diversity in the Okhotsk subspecies has not been investigated. The apparently large population size suggests that presently it is not likely at imminent risk due to inbreeding depression, loss of genetic variants, or fixation of deleterious mutations.
Is the species’ abundance so low that it is at imminent risk of extinction due to its inability to provide important ecological functions throughout its lifecycle?
Species that depend on critical numbers or density for modification of their or other organisms’ physical or biological environments potentially are at risk at low population size. The life cycle of ringed seals does not include such a dependence on a certain density.
Is the species’ abundance so low that it is at imminent risk due to demographic stochasticity?
Saimaa ringed seal – An estimate of the demographic effective population size of Saimaa ringed seals indicated that low population size is exacerbated by habitat fragmentation and that the subspecies is “vulnerable to extinction due to demographic stochasticity alone” (Kokko et al. 1998).
Ladoga ringed seal – We are not aware of any estimates of the demographic effective population size of the Ladoga subspecies. Estimates of current population size, however, are an order of magnitude greater than the numbers typically associated with populations at imminent risk due to demographic stochasticity.

183
Baltic ringed seal – Palo et al. (2001) calculated an effective population size of 20,000 Baltic ringed seals at the end of the last glaciations. Subsequently, the population declined by about 75%. Estimates of current population size, however, are an order of magnitude greater than the numbers typically associated with populations at imminent risk due to demographic stochasticity. Note, however, the possibility of population subdivision described above.
Arctic ringed seal – We are not aware of any estimation of the demographic effective population size of the Arctic subspecies. Current—albeit crude—population estimates are many times larger than the numbers typically associated with populations at imminent risk due to demographic stochasticity. Note, however, that the population structure of Arctic ringed seals is not resolved, and this subspecies may, in fact, be comprised of multiple breeding populations.
Okhotsk ringed seal – We are not aware of any estimates of the demographic effective population size of the Okhotsk subspecies. Current population estimates are many times larger than the numbers typically associated with populations at imminent risk due to demographic stochasticity. Note, however, that the population structure of Okhotsk ringed seals has not been investigated.
4.3.2 Productivity
Is the species’ average productivity below replacement and such that it is at imminent risk of satisfying the abundance conditions described above?
Saimaa ringed seal – The Saimaa subspecies population is growing slowly under active management by the Finnish government.
Ladoga ringed seal – The population of Ladoga ringed seals is believed to have been stable for the past three decades (Sipilä and Hyvärinen 1998) suggesting that the average productivity is not below replacement.
Baltic ringed seal – Overall, the Baltic ringed seal population is stable or increasing, but the data are dominated by the larger, Bothnian Bay population. Productivity in the smaller—possibly discrete—southern populations may be below replacement.
Arctic ringed seal – The population trend is unknown for Arctic ringed seals. If the subspecies is comprised of multiple populations, it is possible that one or more are at risk from inadequate productivity.
Okhotsk ringed seal – The population of Okhotsk ringed seals is believed to be stable, but there are no firm data for that conclusion.

184
Is the species’ average productivity below replacement and such that it is unable to exploit requisite habitats/niches/etc. or at imminent risk due to depensatory processes during any life‐history stage?
Saimaa ringed seal – The Saimaa subspecies’ current average productivity is not known to be below replacement, but the subspecies no longer occupies portions of it former range in Lake Saimaa and, therefore, is no longer able to exploit some habitats as a consequence of population decline.
Ladoga ringed seal – Productivity of the Ladoga ringed seal is not well known.
Baltic ringed seal – The Baltic subspecies average productivity does not appear to be below replacement, but see cautions above about possible population subdivision.
Arctic ringed seal – Average productivity is unknown for the Arctic subspecies.
Okhotsk ringed seal – Average productivity is unknown for the Okhotsk subspecies.
Does the species exhibit trends or shifts in demographic or reproductive traits that portend declines in per capita growth rate which pose imminent risk of satisfying any of the preceding conditions?
Saimaa ringed seal – Changes in demographic or reproductive traits of Saimaa ringed seals are not known to pose risks.
Ladoga ringed seal – Changes in demographic or reproductive traits of Ladoga ringed seals are not known to pose risks.
Baltic ringed seal – Changes in demographic or reproductive traits of Baltic ringed seals are not known to pose risks.
Arctic ringed seal – Changes in demographic or reproductive traits of Arctic ringed seals are not known to pose risks.
Okhotsk ringed seal – Changes in demographic or reproductive traits of Arctic ringed seals are not known to pose risks.
Species status evaluations should take into account uncertainty in estimates of growth rate and productivity‐related parameters.
Saimaa ringed seal – Population size and trends are well established for Saimaa ringed seals.
Ladoga ringed seal – There are no reliable estimates of population growth rate or productivity for Ladoga ringed seals.
Baltic ringed seal – Population size and trends are well established for Baltic ringed seals.

185
Arctic ringed seal – There are no good estimates of growth rate or productivity‐related parameters for Arctic ringed seals.
Okhotsk ringed seal – There are no good estimates of growth rate or productivity‐related parameters for Okhotsk ringed seals.
4.3.3 Spatial Structure
Are habitat patches being destroyed faster than they are naturally created such that the species is at imminent risk of extinction due to environmental and anthropogenic perturbations or catastrophic events?
Saimaa ringed seal – The diminishing quantity and quality of ice and snow cover represent significant changes in the habitats available to Saimaa ringed seals. Reduction of ice as a platform for pupping and resting, as well as reduction in the duration and quality of snow cover, will substantially impact ringed seal habitat. As that habitat deteriorates, Saimaa ringed seals will not be able to emigrate.
Ladoga ringed seal – Ice cover has diminished about 12% over the past 50 years in Lake Ladoga (Karetnikov and Naumenko 2008), but we are not aware of any forecasts specific to the lake for future ice conditions. Loss of snow cover, likely will be a substantial and negative impact on habitat. As the habitat in the lake deteriorates, Ladoga ringed seals will not be able to respond by shifting their range.
Baltic ringed seal – The diminishing quantity and quality of sea ice and snow cover represent significant changes in the habitats available to Baltic ringed seals. Reduction of ice as a platform for pupping, molting, and resting, as well as reduction in the duration and quality of snow cover, will substantially impact ringed seal habitat. As that habitat deteriorates, Baltic ringed seals will not be able to shift their range northward.
Arctic ringed seal – The diminishing quantity and quality of ice and snow cover represent significant changes in the habitats available to Arctic ringed seals. Reduction of ice as a platform for pupping, molting, and resting, as well as reduction in the duration and quality of snow cover, will substantially impact ringed seal habitat. As that habitat deteriorates, Arctic ringed seals initially may be able to compensate by emigrating northward. Within the century, however, snow cover likely will be inadequate for lair use over substantial portions of the subspecies’ range, including the highest latitudes.
Okhotsk ringed seal – The diminishing quantity and quality of sea ice and snow cover represent significant changes in the habitats available to Okhotsk ringed seals. Reduction of ice as a platform for pupping, molting, and resting, as well as further reduction in the duration and quality of snow cover, may have already substantially impacted ringed seal habitat. As that habitat continues to deteriorate, Okhotsk ringed seals will be limited in their ability to shift their range northward. Some Okhotsk ringed seals have been reported on terrestrial resting sites, but these provide inferior whelping and nursing habitat.

186
Are natural rates of dispersal among populations, metapopulations, or habitat patches so low that the species is at imminent risk of extinction due to insufficient genetic exchange among populations, or an inability to find or exploit available resource patches?
Saimaa ringed seal – Dispersal patterns among Saimaa ringed seals are currently under investigation. Movements of adult seals apparently are quite limited, but pups seem to move throughout the lake (Koskela et al. 2002). Population subdivision, however, is considered a significant threat (Kokko et al. 1998, Sipilä 2003). Furthermore, the subspecies is landlocked so dispersal is limited to within the lake.
Ladoga ringed seal – The Ladoga ringed seal has been landlocked for the past 8,000‐9,000 years. Dispersal from the lake is not possible, but its long persistence in the lake suggest that lack of dispersal opportunities does not put it at imminent risk of extinction due to insufficient genetic exchange among populations, or an inability to find or exploit available resource patches.
Baltic ringed seal – Tracking studies suggest that there is no or limited dispersal between the three breeding areas in the Baltic Sea. Genetic studies, however, suggest at least intermittent gene flow (Palo et al. 2001). Resolving the patterns of gene flow among those and other areas of the ringed seal’s range will be critical to assessing the likelihood of extinctions due to insufficient genetic exchange among populations, or an inability to find or exploit available resource patches.
Arctic ringed seal – Rates of dispersal are unknown for the Arctic subspecies. High genetic diversity has been interpreted as evidence of considerable dispersal. High site fidelity, on the other hand, suggests that dispersal may be low. Low rates of genetic drift provide an alternative explanation for the observed levels of genetic diversity. If dispersal is low and the subspecies is composed of numerous discrete populations, the risks of extinctions would be increased.
Okhotsk ringed seal – Neither the structure of the population nor the rates of dispersal have been investigated for the Okhotsk subspecies.
Is the species at imminent risk of extinction due to the loss of critical source populations, subpopulations, or habitat patches?
Saimaa ringed seal – Loss of adequate snow cover for the construction and maintenance of subnivean lairs will constitute loss of crucial habitat patches.
Ladoga ringed seal – Loss of adequate snow cover for the construction and maintenance of subnivean lairs will constitute loss of crucial habitat patches.
Baltic ringed seal – Loss of adequate snow cover for the construction and maintenance of subnivean lairs will constitute loss of crucial habitat patches.
Arctic ringed seal – Loss of adequate snow cover for the construction and maintenance of subnivean lairs will constitute loss of crucial habitat patches.

187
Okhotsk ringed seal – Snow cover is inadequate for the construction and maintenance of subnivean lairs in most areas of the Sea of Okhotsk.
Analyses of species’ spatial processes should take uncertainty into account.
Saimaa ringed seal – The distribution of Saimaa ringed seal breeding areas is well known, and their movements are fairly well understood.
Ladoga ringed seal – The distribution of breeding areas is reasonably well known for Ladoga ringed seals.
Baltic ringed seal – Spatial patterns of habitat use are reasonably well known for Baltic ringed seals.
Arctic ringed seal – Important aspects of the Arctic subspecies’ spatial processes need to be resolved.
Okhotsk ringed seal – Important aspects of the Okhotsk subspecies’ spatial processes need to be investigated.
4.3.4 Diversity
Is the species at risk due to a substantial change or loss of variation in life‐history traits, population demography, morphology, behavior, or genetic characteristics?
Saimaa ringed seal – The Saimaa subspecies has substantially less variability in microsatellite and mitochondrial DNA markers than do Arctic ringed seals. The genetic variability presumably was reduced due to genetic drift in the small, isolated population. It is unknown whether genetic variability is sufficient to permit adaptation to environmental changes such as diminishing ice and snow cover.
Ladoga ringed seal – The Ladoga subspecies is intermediate between Saimaa and Arctic ringed subspecies in terms of genetic diversity. Genetic variability presumably was reduced due to genetic drift in the isolated population. It is unknown whether genetic variability is sufficient to permit adaptation to environmental changes such as diminishing ice and snow cover.
Baltic ringed seal – There is no evidence that at current population levels the Baltic Sea ringed seal is at risk due to a substantial change or loss of variation in life‐history traits, population demography, morphology, behavior, or genetic characteristics.
Arctic ringed seal – There is no evidence that at current population levels the Arctic ringed seal is at risk due to a substantial change or loss of variation in life‐history traits, population demography, morphology, behavior, or genetic characteristics.
Okhotsk ringed seal – There is no evidence that at current population levels the Okhotsk ringed seal is at risk due to a substantial change or loss of variation in life‐history traits, population demography, morphology, behavior, or genetic characteristics.

188
Is the species at risk because natural processes of dispersal and gene flow among populations have been significantly altered?
Saimaa ringed seal – Isolation in Lake Saimaa 8,000‐9,000 years ago disrupted gene flow with other populations. Population subdivision in the large and complex lake potentially could limit dispersal and maintenance of diversity.
Ladoga ringed seal – Isolation in Lake Ladoga 8,000‐9,000 years ago disrupted gene flow with other populations. Natural processes of dispersal and gene flow have not been altered since that initial isolation and do not appear to pose a risk to the subspecies at present.
Baltic ringed seal – Harkonen et al. (2008) suggested that gene flow among Baltic Sea ringed seals has been disrupted, although genetic data suggest any such disruption must be quite recent. Such a change would add to the subspecies’ risk.
Arctic ringed seal – Natural processes of dispersal and gene flow among Arctic ringed seals are inadequately known, and we lack the means to assess whether they have been significantly altered.
Okhotsk ringed seal – Natural processes of dispersal and gene flow among Okhotsk ringed seals are inadequately known, and we lack the means to assess whether they have been significantly altered.
Is the species at risk because natural processes that cause ecological variation have been significantly altered?
Saimaa ringed seal – The expected reductions in the duration of ice and snow cover will represent significantly reduced ecological variation. The Saimaa ringed seal’s whelping and nursing seasons are adapted to the phenology of ice and snow. The subspecies’ persistence will be greatly challenged by significantly altered patterns of ecological variation.
Ladoga ringed seal – The expected reductions in the duration of ice and snow cover will represent significantly reduced ecological variation. The Ladoga ringed seal’s whelping and nursing seasons are adapted to the phenology of ice and snow. The subspecies’ persistence will be greatly challenged by significantly altered patterns of ecological variation.
Baltic ringed seal – The expected reductions in the duration of ice and snow cover will represent significantly reduced ecological variation. The Baltic ringed seal’s whelping and nursing seasons are adapted to the phenology of ice and snow. The subspecies’ persistence will be greatly challenged by significantly altered patterns of ecological variation.
Arctic ringed seal – The expected reductions in the duration of ice and snow cover will represent significantly reduced ecological variation. The Arctic ringed seal’s whelping and nursing seasons are adapted to the phenology of ice and snow. The subspecies’ persistence will be greatly challenged by significantly altered patterns of ecological variation.

189
Okhotsk ringed seal – The expected continuing reductions in the duration of ice and snow cover will represent significantly reduced ecological variation. The Okhotsk ringed seal’s whelping and nursing seasons are adapted to the phenology of ice and snow. The subspecies’ persistence will be greatly challenged by significantly altered patterns of ecological variation.
Species status evaluations should take uncertainty about requisite levels of diversity into account.
Saimaa ringed seal – We do not know whether the lower genetic diversity among Saimaa ringed seals poses a significant risk. Given the drastic and rapid environmental changes facing the subspecies, however, higher levels of diversity would increase the likelihood of persistence.
Ladoga ringed seal – It is unlikely that observed levels of genetic diversity among Ladoga ringed seals pose a significant risk. We cannot say whether that level of genetic diversity will be adequate in the face large environmental changes facing the subspecies.
Baltic ringed seal – The possibility of population subdivision among Baltic ringed seals raises the distinct possibility that one or more of the three breeding areas lacks for genetic diversity and, therefore, may be more vulnerable to environmental change.
Arctic ringed seal – It is unlikely that current levels of genetic diversity among Arctic ringed seals pose a significant risk. We cannot say whether that level of genetic diversity will be adequate in the face large environmental changes facing the subspecies. Important aspects of population structure among the Arctic need to be resolved.
Okhotsk ringed seal – Genetic diversity has not been investigated among Okhotsk ringed seals. We cannot say whether that level of genetic diversity will be adequate in the face large environmental changes facing the subspecies.
4.3.5 Relevant Modifying or Mitigating Factors
4.3.5.1 Lifehistory characteristics
The wide distribution of ringed seals in fresh, brackish, and salt water environments; from marginal ice zones to the High Arctic; and in shorefast and pack‐ice habitats suggest adaptability to changing environments. The ringed seal’s broad distribution, ability to undertake long movements, diverse diet, and association with widely varying ice conditions suggest resilience in the face of environmental variability (Laidre et al. 2008). Ringed seals—with the exception of the Saimaa subspecies—also exhibit high levels of genetic diversity that might predispose them to adaptive responses to climate change (Palo et al. 2001, Palo et al. 2003). Low effective population size contributes to continued risk of extinction for the Saimaa ringed seal.
The long generation time and ability to produce only a single pup each year may limit the ringed seal’s ability to respond to environmental challenges such as diminishing ice and snow cover that will be dramatic in a matter of decades. Ringed seals apparently thrived during glacial maxima and survived

190
warm interglacial periods. How they survived the latter periods or in what numbers is not known. The rates of environmental change facing ringed seals in the coming century will be rapid and may outpace possible adaptive responses.
The evolution of pinnipeds involved a trade‐off between mobility out of the water—where young are born and nursed—and mobility in the water—where prey are captured (Bartholomew 1970). The trade‐off favored aquatic mobility with the result that pinnipeds move slowly and are vulnerable to predation out of the water (Allen 1880). Predators abound on continental shorelines, but are largely absent from small islands. The latter, however, offer very limited area to whelping pinnipeds. Seasonal sea ice, in contrast, is continental in area, and predators are absent (Antarctica) or sparse (Arctic), and pinniped diversity is greatest in those habitats (Kelly 2001). Ringed seals are among the smallest pinnipeds, and they must nurse their young for 1‐2 months. The small pups are vulnerable to cold temperatures and avian and mammalian predators on the ice, especially during the nursing period. Thus, a specific habitat requirement for ringed seals is adequate snow cover for the occupation of subnivean lairs, especially in spring when pups are born and nursed.
4.3.5.2 Population characteristics
The low density of the Saimaa ringed seal population coupled with limited dispersal opportunities and depensatory effects continue to put the subspecies at risk. Present population levels of the other ringed seal subspecies favor resilience. Dispersal opportunities are also limited for the Okhotsk, Baltic, and Ladoga ringed seal subspecies.
4.3.5.3 Habitat characteristics
The key factor limiting the viability of all five ringed seal subspecies is the forecasted reduction in spring snow cover. Early snow melts already are evident in much of the species’ range. Increasingly late ice formation in autumn contributes to expectations of substantial decreases in snow accumulation. The Arctic sea ice and its snow cover are especially vulnerable to climate warming because loss of snow and ice has a positive feedback on warming.
The ringed seal’s specific requirement for habitats with adequate spring snow cover is manifested in the pups’ low tolerance for exposure to wet, cold conditions and their vulnerability to predation. Premature failure of the snow cover has caused high mortality due to freezing and predation. Climate warming will result in increasingly early snow melts exposing vulnerable ringed seal pups to predators. The warming also will increase immigration of avian and other predators in to ringed seal habitat. The shifts in ice and snow habitats are directional, and we have high confidence that the climate will warm, reducing ice and snow cover for at least the next century and probably beyond.

191
4.3.6 Demographic Risks Assessment
The BRT members’ assessment of the severity of the demographic risks to the persistence of each ringed seal subspecies was formalized using a numerical scoring system. Scoring was modeled on similar approaches used in other ESA status reviews (e.g., Atlantic Wolffish Biological Review Team 2009, Butler et al. 2009). The system was designed to elicit expert judgment about the likelihood that the known and potential threats will impact a subspecies’ persistence. Specifically, each BRT member considered the risk that the subspecies may be placed in danger of extinction by demographic problems with abundance, productivity, spatial structure, or diversity, and then assigned a score to each of these demographic categories as follows: 1 = very low or zero risk, 2 = low risk, 3 = moderate risk, 4 = high risk, and 5 = very high risk. The questions posed in sections 4.3.1‐4.3.4 and used to guide consideration of each demographic category (McElhany et al. 2000) emphasized the present risks of extinction. To assess future risks of extinction, the BRT members also scored the risks that problems associated with those demographic categories will, in the foreseeable future, place the subspecies in danger of extinction. The average score and the range of scores among BRT members are presented in Table 10.

192
Table 10. ‐‐ B
RT m
embe
rs ju
dged
the
risks th
at th
reats to the
persisten
ce of e
ach ringed
seal sub
species wou
ld be man
ifested
as de
mograph
ic problem
s associated
with ab
unda
nce, produ
ctivity, spa
tial structure, o
r diversity. The
severity of the
risk that th
e subspe
cies m
ay be placed
in dan
ger of extinction by
these de
mograph
ic problem
s—bo
th presently and
in th
e foreseeable future—was given
a score of 1
(very low or zero risk), 2
(low
risk), 3
(mod
erate risk), 4
(high risk), or 5 (very high
risk). The
averages an
d ranges (in pa
renthe
ses) of the
se scores are presen
ted.
Dem
ograph
ic
Risks
P. h. h
ispida
P. h. o
chotensis
P. h. b
otnica
P. h. lad
ogensis
P. h. saimen
sis
Presen
t Foreseeable
Future
Presen
t Foreseeable
Future
Presen
t Foreseeable
Future
Presen
t Foreseeable
Future
Presen
t Foreseeable
Future
Abu
ndan
ce
1.1
2.9
1.3
3.8
2.0
4.0
2.8
4.3
4.3
4.8
(1‐2)
(2‐4)
(1‐2)
(3‐5)
(1‐3)
(3‐5)
(2‐4)
(3‐5)
(2‐5)
(3‐5)
Prod
uctivity
1.4
3.7
1.8
4.3
2.1
4.1
2.6
4.6
3.8
4.6
(1‐2)
(2‐5)
(1‐2)
(3‐5)
(2‐3)
(3‐5)
(1‐4)
(3‐5)
(2‐5)
(3‐5)
Spatial
Structure
2.1
4.0
2.2
4.2
2.9
4.3
3.4
4.4
4.1
4.8
(1‐3)
(2‐5)
(1‐3)
(3‐5)
(2‐3)
(4‐5)
(2‐4)
(3‐5)
(3‐5)
(4‐5)
Diversity
1.7
2.8
1.8
3.2
2.1
3.3
2.7
3.4
3.6
4.2
(1‐3)
(1‐3)
(1‐3)
(2‐4)
(1‐3)
(2‐4)
(2‐3)
(3‐4)
(3‐5)
(3‐5)

193
4.4 Conclusions of the Extinction Risk Assessment
Warming—driven by GHG emissions—is accelerated in the Arctic by positive feedbacks including reduced albedo. Recent reductions in the areal extent of sea ice and the seasonal duration of snow on sea ice have contributed strongly to the reduction in albedo, meaning more heat is retained by the ocean and earth’s surface. Current atmospheric levels of GHGs are sufficient to continue warming the climate and diminishing ice and snow cover throughout the century. The changes to the ice and snow habitats of ringed seals are forecasted to be rapid relative to generation time, challenging the species’ ability to respond adaptively.
Ringed seal populations will be impacted indirectly through changes in biological community composition as consequences of ocean and lake warming and acidification. Direct effects will result from diminishing ice and snow cover. The BRT considered 17‐18 threats to each subspecies of ringed seals and assessed how those threats are likely to manifest demographically (presently and in the foreseeable future) as risks to abundance, productivity, spatial structure, and diversity.
4.4.1 Arctic ringed seals
The BRT judged the greatest threat to the Arctic ringed seal to be increased hypothermia due to decreasing depth and duration of snow cover. The threat considered the second highest to this subspecies was increased predation, also associated with diminishing snow cover. The BRT considered current risks to be low for all four demographic attributes, but they rated risks higher in the foreseeable future for all attributes. Overall, risks to productivity and spatial structure were rated high in the foreseeable future. Persistence of the Arctic subspecies likely will be challenged as decreases in ice and, especially, snow cover lead to increased juvenile mortality from hypothermia and predation. Spatial structure likely will be disrupted by rapid loss of habitat patches, and the subspecies likely will disappear from a substantial portion of its range.
4.4.2 Okhotsk ringed seals
The greatest threat to Okhotsk ringed seals was judged by the BRT to be increased hypothermia due to decreasing depth and duration of snow cover. The threat judged second highest for the Okhotsk ringed seal was a decrease in sea‐ice habitat suitable for whelping and nursing, reflecting forecasts that April ice will be limited to the northernmost Sea of Okhotsk and that snow cover already appears to be inadequate for birth lairs throughout the range. The BRT judged the current risks to persistence to be low for ringed seals in the Sea of Okhotsk but judged risks in the foreseeable future to be moderate (diversity) or high (abundance, productivity, and spatial structure). By the end of the century, reduced extent of sea ice and inadequate snow accumulation will challenge the persistence of ringed seals throughout their range in the Sea of Okhotsk.

194
4.4.3 Baltic ringed seals
The BRT judged decreased sea‐ice habitat suitable for whelping and nursing, and increased hypothermia due to insufficient depth or duration of snow cover, to be the greatest threats to the persistence of the Baltic ringed seal. The threats were judged currently to pose low to moderate risks for each of the four demographic attributes. In the foreseeable future, however, the BRT judged the risks to be moderate (diversity) to high (abundance, productivity, and spatial structure). Forecasts of substantial reductions in sea‐ice extent by mid‐century, coupled with deteriorating snow conditions, represent drastic habitat modifications expected to decrease survival of ringed seal pups throughout the Baltic Sea.
4.4.4 Ladoga ringed seals
Decreased ice habitat suitable for whelping and nursing, and increased hypothermia due to insufficient depth or duration of snow cover, were judged by the BRT to be the greatest threats to the persistence of the Ladoga ringed seals. The present risk to population persistence was judged to be moderate in terms of abundance, productivity, spatial structure, and diversity. The concern about spatial structure reflected the fact that the subspecies is landlocked and cannot respond to habitat loss by dispersing to new habitat. Risks in the foreseeable future were judged to be moderate (diversity) or high to very high (abundance, productivity, and spatial structure). Drastic reductions in snow cover forecasted for the region can be expected to result in increased pup mortality throughout Lake Ladoga.
4.4.5 Saimaa ringed seals
The greatest threats to Saimaa seals, in the judgment of the BRT, were increased hypothermia due to insufficient depth or duration of snow cover and decreased ice habitat suitable for whelping and nursing. Given the low population size (< 300 seals), the present risk to population persistence was judged to be high for all of the demographic attributes. In the foreseeable future, the risks were judged by the BRT to be high to very high for all of the demographic attributes. Drastic reductions in snow cover forecasted for the region can be expected to result in increased pup mortality throughout Lake Saimaa.

195
5 ACKNOWLEDGMENTS
Ringed seals are broadly distributed in the Northern Hemisphere, and we depended on our U.S. and international colleagues for data and reviews. We extend our appreciation to Aqqalu Rosing‐Asvid, Micaela Martinez‐Bakker, Cecilia Bitz, Yuri Bukhtiarov, Vladimir Burkanov, Sheldon Drobot, Tracey Goldstein, Patt Hall, Tero Härkönen, Lois Harwood, Kit Kovacs, Mervi Kunnasranta, Bruce Marcot, Jim Maslanik, Melissa Neuman, Donald Perovich, Jeff Short, Tero Sipilä, Thomas G. Smith, David Tallmon, Barbara Taylor, Alexy Trukhin, Dag Vongraven, and John Walsh. Elizabeth Morgan, Erin Richmond, Luciana Santos, and Kym Yano maintained the electronic bibliographic library that was used while writing this document and Jessica Tingg provided technical support for the BRT’s organizational website. We acknowledge the Program for Climate Model Diagnosis and Intercomparison and the World Climate Research Programme's Working Group on Coupled Modeling for their roles in making available the CMIP3 multi‐model dataset. Support of this dataset is provided by the Office of Science, U.S. Department of Energy.


197
6 CITATIONS
ACIA. 2004. Impacts of a warming Arctic: Arctic climate impact assessment. Cambridge University Press, New York. 144 p. Accessed at http://www.acia.uaf.edu.
ACIA. 2005. Arctic Climate Impact Assessment. Cambridge University Press, Cambridge, UK. 1042 p. Accessed September 2008 at http://www.acia.uaf.edu.
Adam, D. 2007. Research boom in Arctic village as oil reserves draw big powers. The Guardian, London, England. Accessed June 2010 at http://www.guardian.co.uk/environment/2007/aug/22/energy.oilandpetrol.
Addison, R. F., M. G. Ikonomou, and T. G. Smith. 2000. Residues of PCDD, PCDF, NO‐CB, and MO‐CB in ringed seals (Phoca hispida) from Holman, NWT, in 1981, 1991, and 1996. Department of Fisheries and Oceans, Science Branch, Canadian Data Report of Fisheries and Aquatic Sciences. 2000 p.
Addison, R. F., M. G. Ikonomou, and T. G. Smith. 2005. PCDD/F and PCB in harbour seals (Phoca vitulina) from British Columbia: response to exposure to pulp mill effluents. Marine Environmental Research 59:165‐176.
Addison, R. F., D. C. G. Muir, M. G. Ikonomou, L. Harwood, and T. G. Smith. 2009. Hexachlorocyclohexanes (HCH) in ringed seal (Phoca hispida) from Ulukhaktok (Holman), NT: trends from 1978 to 2006. Science of the Total Environment 407:5139‐5146.
Addison, R. F., and T. G. Smith. 1974. Organochlorine residue levels in Arctic ringed seals: Variation with age and sex. Oikos 25:335‐377.
Ådjers, K., M. Appelberg, R. Eschbaum, A. Lappalainen, A. Minde, R. Repečka, and G. Thoresson. 2006. Trends in coastal fish stocks of the Baltic Sea. Boreal Environment Research 11:13‐25.
Agafonova, E. V., M. V. Verevkin, R. A. Sagitov, T. Sipilä, M. V. Sokolovskay, and V. U. Shahnazarova. 2007. The ringed seal in Lake Ladoga and the Valaam Archipelago. Baltic Fund for Nature of Saint Petersburg Naturalists Society. 60 p. (In Russian, English Summary).
Aguilar, A., A. Borrell, and P. Reijnders. 2002. Geographical and temporal variation in levels of organochlorine contaminants in marine mammals. Marine environmental research 53:425‐452.
Alaska Department of Natural Resources. 2009. Alaska oil and gas report November 2009. Alaska DNR, Division of Oil and Gas, Anchorage, AK. 63 p. Accessed at http://www.dog.dnr.state.ak.us/oil/products/publications/annual/2009_annual_report/updated_2009_annual_report/Annual%20Report%202009%20Updated%205‐18‐10.pdf.
Alaska Department of Natural Resources. 2010a. Historic oil and gas activity maps. Alaska DNR, Division of Oil and Gas, Anchorage, AK. Accessed June 2010 at http://www.dog.dnr.state.ak.us/oil/products/maps/historic_activity_maps.htm.
Alaska Department of Natural Resources. 2010b. State of Alaska five‐year program of proposed oil and gas lease sales. Alaska DNR, Division of Oil and Gas, Anchorage, AK. 56 p. Accessed at http://www.dog.dnr.alaska.gov/oil/products/publications/otherreports/5_year_reports/5_year_report.html.
Allen, B. M., and R. P. Angliss. 2010. Alaska Marine Mammal Stock Assessments, 2009. U.S. Department of Commerce, NOAA Technical Memorandum NMFS‐AFSC‐206. 276 p.
Allen, J. A. 1880. History of North American pinnipeds: a monograph of the walruses, sea‐lions, sea‐bears and seals of North America. U.S. Department of the Interior, U.S. Government Printing Office, Washington, D.C. 785 p.
Almkvist, L. 1982. Baltic marine mammals ‐‐ a status report. International Council for the Exploration of the Sea, C.M. 1982/N:16. 19 p.

198
Amano, M., A. Hayano, and N. Miyazaki. 2002. Geographic variation in the skull of the ringed seal, Pusa hispida. Journal of Mammalogy 83:370‐380.
AMAP. 2007. Arctic oil and gas 2007. Arctic Monitoring and Assessment Programme, Oslo, Norway. 70 p. Accessed at http://www.amap.no/oga/.
AMAP. 2009. Arctic Pollution 2009. Arctic Monitoring and Assessment Programme, Oslo, Norway. 83 p. Accessed at http://www.amap.no/.
Anchorage Daily News. 2010. Eni Petroleum set to produce Nikaitchuq oil. Anchorage Daily News, Anchorage, AK. Accessed June 2010 at http://www.adn.com/2010/02/16/1142601/eni‐petroleum‐set‐to‐produce‐nikaitchuq.html.
Andersen, M., J. P. Gwynn, M. Dowdall, K. M. Kovacs, and C. Lydersen. 2006. Radiocaesium (137Cs) in marine mammals from Svalbard, the Barents Sea and the North Greenland sea. Science of the Total Environment 363:87‐94.
Anderson, L. G., and S. Kaltin. 2001. Carbon fluxes in the Arctic Ocean ‐‐ potential impact by climate change. Polar Research 20:225‐232.
Anderson, P. J., and J. F. Piatt. 1999. Community reorganization in the Gulf of Alaska following ocean climate regime shift. Marine Ecology Progress Series 189:117‐123.
Anderson, R. M. 1934. Mammals of the eastern Arctic and Hudson Bay. Pages 67‐108 in W. C. Bethune, editor. Canada's Eastern Arctic: Its History, Resources, Population and Administration. U.S. Department of Interior, Ottawa, Canada.
Andriashek, D., and C. Spencer. 1989. Predation on a ringed seal, Phoca hispida, pup by a red fox, Vulpes vulpes. Canadian Field‐Naturalist 103:600.
Angliss, R. P., and B. M. Allen. 2009. Alaska marine mammal stock assessments, 2008. U.S. Department of Commerce, NOAA Technical Memorandum NMFS‐AFSC‐193. 258 p.
Angliss, R. P., and K. L. Lodge. 2002. Alaska marine mammal stock assessments, 2002. U.S. Department of Commerce, NOAA Technical Memorandum NMFS‐AFSC‐133. 224 p.
Anonymous. 1994. Otchet o deyatelnosti Sevrybvod [Official report on the activity of Sevrybvod]. Arkhangelsk. (In Russian).
Arctic Council. 2009. Arctic marine shipping assessment 2009 report. Arctic Council, Tromsø, Norway. 187 p. Accessed at http://www.pame.is/amsa/amsa‐2009‐report.
Árnason, U., K. Bodin, A. Gullberg, C. Ledje, and S. Mouchaty. 1995. A molecular view of pinniped relationships with particular emphasis on the true seals. Journal of Molecular Evolution 40:78‐85.
Árnason, U., A. Gullberg, A. Janke, M. Kullberg, N. Lehman, E. A. Petrov, and R. Väinölä. 2006. Pinniped phylogeny and a new hypothesis for their origin and dispersal. Molecular Phylogenetics and Evolution 41:345‐354.
ArtDatabanken. 2010. The 2010 Red List of Swedish Species. ArtDatabanken, Uppsala, Sweden. Accessed May 2010 at http://snotra.artdata.slu.se/artfakta/GetSpecies.aspx?SearchType=Advanced.
Ashley, G., L. Harwood, H. Melling, G. Stern, and S. Ferguson. 2008. Organochlorine contaminants in ringed seals (Phoca hispida) in the western Canadian Arctic: Trends with sea ice. Organohalogen Compounds 70:001028‐001031.
Ashwell‐Erickson, S., F. H. Fay, and R. Elsner. 1986. Metabolic and hormonal correlates of molting and regeneration of pelage in Alaskan harbor and spotted seals (Phoca vitulina and Phoca largha). Canadian Journal of Zoology 64:1086‐1094.
Associated Foreign Press. 2009. Oil spill in Russian Far East kills hundreds of birds: reports. Associated Foreign Press, Moscow, Russia. Accessed June 2010 at http://www.google.com/hostednews/afp/article/ALeqM5g1LeTJjXj2LGoogMI331NPFt5oQA.
Assunção, M., and P. Ross. 2001. Cytochrome P450 1A enzymes as non‐invasive biomarkers of contaminant exposure in skin from harbour seals (Phoca vitulina). T. Droscher, editor.

199
Proceedings of the 2001 Puget Sound Research Conference, Olympia, WA. Puget Sound Action Team.
Atlantic Wolffish Biological Review Team. 2009. Status review of Atlantic wolffish (Anarhichas lupus). Report to National Marine Fisheries Service, Northeast Regional Office. September 30, 2009. 149 p.
Atwell, L., K. A. Hobson, and H. E. Welch. 1998. Biomagnification and bioaccumulation of mercury in an arctic marine food web: insights from stable nitrogen isotope analysis. Canadian Journal of Fisheries and Aquatic Sciences 55:1114‐1121.
Auvinen, H., J. Jurvelius, J. Koskela, and T. Sipilä. 2005. Comparative use of vendace by humans and Saimaa ringed sea in Lake Pihlajavesi, Finland. Biological Conservation 125:381‐389.
Bacastow, R., and C. D. Keeling. 1973. Atmospheric carbon‐dioxide and radiocarbon in the natural carbon cycle: II. Changes from A.D. 1700 to 2070 as deduced from a geochemical model. Brookhaven Symposia in Biology 30:86‐135.
Bacevičius, E. 2009. The first record of the fourhorn sculpin ((Triglopsis quadricornis Linnaeus, 1758): Actinopterygii: Scorpaeniformes: Cottidae) in the southeastern part of the Baltic Sea (Lithuanian coastal zone). Acta Zoologica Lituanica 19:324‐329.
Bailey, A. M., and R. W. Hendee. 1926. Notes on the mammals of northwestern Alaska. Journal of Mammalogy 7:9‐28.
Bakulina, E. D. 1989. Some features of spermatogenesis of the Baltic ringed seal (Pusa hispida botnica). Pages 68‐73 in A. V. Yablokov and M. Olsson, editors. Influence of Human Activities on the Baltic Ecosystem. Proceedings of the Soviet‐Swedish Symposium. Effects of Toxic Substances on Dynamics of Seal Populations, Moscow, USSR. Leningrad Gidrometeoizdat.
Bakun, A. 1990. Global climate change and intensification of coastal ocean upwelling. Science 247:198‐201.
Baltic Sea 2020. 2009. Baltic Sea challenge: shipping. Baltic Sea 2020, Stockholm, Sweden. 1 p. Accessed at http://www.balticsea2020.org/attachments/128_Challenges_ship.pdf.
Bang, K., B. M. Jenssen, C. Lydersen, and J. U. Skaare. 2001. Organochlorine burdens in blood of ringed and bearded seals from north‐western Svalbard. Chemosphere 44:193‐203.
Barber, W. E., R. L. Smith, M. Vallarino, and R. M. Meyer. 1997. Demersal fish assemblages of the northeastern Chukchi Sea, Alaska. Fishery Bulletin 95:195‐209.
BarentsObserver. 2008a. Gazprom: less for Shtokman, more for Yamal. BarentsObserver, Kirkenes, Norway. Accessed June 2010 at http://www.barentsobserver.com/gazprom‐less‐for‐shtokman‐more‐for‐yamal.4500148‐16149.html.
BarentsObserver. 2008b. Norway challenged by Russian plans in Spitsbergen waters. BarentsObserver, Kirkenes, Norway. Accessed June 2010 at http://www.barentsobserver.com/norway‐challenged‐by‐russian‐plans‐in‐spitsbergen‐waters.4500045‐28235.html.
Barlough, J. E., E. S. Berry, A. W. Smith, and D. E. Skilling. 1987. Prevalence and distribution of serum neutralizing antibodies to Tillamook (bovine) calicivirus in selected populations of marine mammals. Journal of Wildlife Diseases 23:45‐51.
Barrett, T., P. Sahoo, and P. D. Jepson. 2003. Seal distempter outbreak 2002. Microbiology Today 30:162‐164.
Bartholomew, G. A. 1970. A model for the evolution of pinniped polygyny. Evolution 24:546‐559. Bates, N. R. 2007. Interannual variability of the oceanic CO2 sink in the subtropical gyre of the North
Atlantic Ocean over the last 2 decades. Journal of Geophysical Research 112:C09013. Bates, N. R., and J. T. Mathis. 2009. The Arctic Ocean marine carbon cycle: evaluation of air‐sea CO2
exchanges, ocean acidification impacts and potential feedbacks. Biogeosciences Discussions 6:6695‐6747.

200
Bates, N. R., J. T. Mathis, and L. W. Cooper. 2009. Ocean acidification and biologically induced seasonality of carbonate mineral saturation states in the western Arctic Ocean. Journal of Geophysical Research 114:C11007.
Bates, N. R., and A. J. Peters. 2007. The contribution of atmospheric acid deposition to ocean acidification in the subtropical North Atlantic Ocean. Marine Chemistry 107:547‐558.
Becker, P. R. 2000. Concentration of chlorinated hydrocarbons and heavy metals in Alaska Arctic marine mammals. Marine Pollution Bulletin 40:819‐829.
Belchansky, G. I., D. C. Douglas, I. N. Mordvintsev, and N. G. Platonov. 2004. Estimating the time of melt onset and freeze onset over Arctic sea‐ice area using active and passive microwave data. Remote Sensing of Environment 92:21‐39.
Belikov, S. E., and A. N. Boltunov. 1998. The ringed seal (Phoca hispida) in the western Russian Arctic. Pages 63‐82 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Bell, S., K. Hampshire, and M. Tonder. 2008. Person, place, and knowledge in the conservation of the Saimaa ringed seal. Society and Natural Resources 21:277‐293.
Bengtson, J. L., L. M. Hiruki‐Raring, M. A. Simpkins, and P. L. Boveng. 2005. Ringed and bearded seal densities in the eastern Chukchi Sea, 1999‐2000. Polar Biology 28:833‐845.
Berge, J. A., E. M. Brevik, A. Bjørge, N. Følsvik, G. W. Gabrielsen, and H. Wolkers. 2004. Organotins in marine mammals and seabirds from Norwegian territory. Journal of Environmental Monitoring 6:108‐112.
Bergeron, E., L. N. Measures, and J. Huot. 1997. Lungworm (Otostrongylus circumlitus) infections in ringed seals (Phoca hispida) from eastern Arctic Canada. Canadian Journal of Fisheries and Aquatic Sciences 54:2443‐2448.
Bergman, A. 1999. Health condition of the Baltic grey seal (Halichoerus grypus) during two decades ‐‐ Gynaecological health improvement but increased prevalence of colonic ulcers. APMIS 107:270‐282.
Bergman, A., A. Bergstrand, and A. Bignert. 2001. Renal lesions in Baltic grey seals (Halichoerus grypus) and ringed seals (Phoca hispida botnica). Ambio 30:397‐409.
Bergman, A., and M. Olsson. 1985. Pathology of Baltic grey seal and ringed seal females with special reference to adrenocortical hyperplasia: is environmental pollution the cause of a widely distributed disease syndrome? Finnish Game Research 44:47‐62.
Bergman, G. 1956. Sälbeståndet vid våra kuster [The seal population in our coastal waters]. Nordenskiöld‐samfundets tidskrift:49‐65. (In Swedish).
Berta, A. 2009. Pinnipedia, Overview. Pages 878‐885 in P. W.F., B. Wursig, and J. G. M. Thewissen, editors. Encyclopedia of Marine Mammals. Academic Press, San Diego, CA.
Best, R. C. 1985. Digestibility of ringed seals by the polar bear. Canadian Journal of Zoology 63:1033‐1036.
Bitz, C. M., M. M. Holland, A. J. Weaver, and M. Eby. 2001. Simulating the ice‐thickness distribution in a coupled climate model. Journal of Geophysical Research‐Oceans 106:2441‐2463.
Blackwell, S. B., C. R. Greene, Jr., and W. J. Richardson. 2004. Drilling and operational sounds from an oil production island in the ice‐covered Beaufort Sea. Journal of the Acoustical Society of America 116:3199‐3211.
Bluhm, B. A., and R. Gradinger. 2008. Regional variability in food availability for Arctic marine mammals. Ecological Applications 18:S77‐S96.
Boertmann, D. 2008. Greenland Red List 2007. National Environmental Research Institute, Aarhus University, and Directorate for Environment and Nature, Nuuk, Greenland. 156 p. Accessed May 2010 at http://www2.dmu.dk/Pub/Groenlands_Roedliste_2007_DK.pdf.

201
Boily, P. 1995. Theoretical heat flux in water and habitat selection of phocid seals and beluga whales during the annual molt. Journal of Theoretical Biology 172:235‐244.
Boldt, J. L., and L. J. Haldorson. 2003. Seasonal and geographical variation in juvenile pink salmon diets in the Northern Gulf of Alaska and Prince William Sound. Transactions of the American Fisheries Society 132:1035‐1052.
Borgå, K., G. W. Gabrielsen, J. U. Skaare, L. Kleivane, R. J. Norstrom, and A. T. Fisk. 2005. Why do organochlorine differences between arctic regions vary among trophic levels? Environmental Science & Technology 39:4343‐4352.
Born, E. W., J. Teilmann, M. Acquarone, and F. F. Rigét. 2004. Habitat use of ringed seals (Phoca hispida) in the North Water Area (North Baffin Bay). Arctic 57:129‐142.
Born, E. W., J. Teilmann, and F. Rigét. 1998. Abundance of ringed seals (Phoca hispida) in the Kong Oscars Fjord, Scoresby Sund and adjacent areas in eastern Greenland. Pages 152‐166 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Born, E. W., J. Teilmann, and F. Rigét. 2002. Haul‐out activity of ringed seals (Phoca hispida) determined from satellite telemetry. Marine Mammal Science 18:167‐181.
Borst, G. H. A., H. C. Walvoort, P. J. H. Reijnders, J. S. van der Kamp, and A. Osterhaus. 1986. An outbreak of a herpesvirus infection in harbor seals (Phoca vitulina). Journal of Wildlife Diseases 22:1‐6.
Bossi, R., F. F. Rigét, and R. Dietz. 2005a. Temporal and spatial trends of perfluorinated compounds in ringed seal (Phoca hispida) from Greenland. Environmental Science & Technology 39:7416‐7422.
Bossi, R., F. F. Rigét, R. Dietz, C. Sonne, P. Fauser, M. Dam, and K. Vorkamp. 2005b. Preliminary screening of perfluorooctane sulfonate (PFOS) and other fluorochemicals in fish, birds and marine mammals from Greenland and the Faroe Islands. Environmental Pollution 136:323‐329.
Boveng, P. L., J. L. Bengtson, T. W. Buckley, M. F. Cameron, S. P. Dahle, B. P. Kelly, B. A. Megrey, J. E. Overland, and N. J. Williamson. 2009. Status review of the spotted seal (Phoca largha). U.S. Department of Commerce, NOAA Technical Memorandum NMFS‐AFSC‐200. 153 p.
Bowen, W. D., and D. B. Siniff. 1999. Distribution, population biology, and feeding ecology of marine mammals. Pages 423‐484 in J. E. I. Reynolds and S. A. Rommel, editors. Biology of Marine Mammals. Smithsonian Institution Press, Washington, D. C.
Boyce, D. G., M. R. Lewis, and B. Worm. 2010. Global phytoplankton decline over the past century. Nature 466:591‐596.
Bradstreet, M. S. W., and W. E. Cross. 1982. Trophic relationships at high Arctic ice edges. Arctic 35:1‐12. Bradstreet, M. S. W., and K. J. Finley. 1983. Diet of ringed seals (Phoca hispida) in the Canadian High
Arctic. LGL Limited Environmental Research Associates, Prepared for Petro‐Canada Exploration Inc. by LGL Limited Environmental Research Associates. 36 p.
Bradstreet, M. S. W., K. J. Finley, A. D. Sekerak, W. B. Griffiths, C. R. Evans, M. F. Fabijan, and H. E. Stallard. 1986. Aspects of the biology of arctic cod (Boreogadus saida) and its importance in arctic marine food chains. Department of Fisheries and Oceans, Canadian Technical Report of Fisheries and Aquatic Sciences, No. 1491. 193 p.
Braham, H. W., J. Burns, J., G. A. Fedoseev, and B. D. Krogman. 1981. Distribution and density of ice‐associated pinnipeds in the Bering Sea. NOAA, National Marine Fisheries Service, Northwest and Alaska Fisheries Center, National Marine Mammal Laboratory. 27 p.
Braune, B. M., P. M. Outridge, A. T. Fisk, D. C. G. Muir, P. A. Helm, K. Hobbs, P. F. Hoekstra, Z. A. Kuzyk, M. Kwan, R. J. Letcher, W. L. Lockhart, R. J. Norstrom, G. A. Stern, and I. Stirling. 2005. Persistent organic pollutants and mercury in marine biota of the Canadian Arctic: an overview of spatial and temporal trends. Science of the Total Environment 351:4‐56.
Brewer, P. G., and K. Hester. 2009. Ocean acidification and the increasing transparency of the ocean to low‐frequency sound. Oceanography 22:86‐93.

202
Brigham, L., and B. Ellis, editors. 2004. Arctic marine transport workshop. Final Report, 28‐30 September 2004. Institute of the North, U.S. Arctic Research Commission, and International Arctic Science Committee, Cambridge, UK. 56 p. Accessed September 2008 at http://www.institutenorth.org/servlet/content/reports.html.
Brinton, E. 1962. The distribution of Pacific euphausiids. Bulletin of the Scripps Institution of Oceanography 8:21‐270.
Broecker, W. S., Y. H. Li, and T. H. Peng. 1971. Carbon dioxide ‐‐ man's unseen artifact. Pages 287‐324 in D. W. Hood, editor. Impingement of man on the oceans. John Wiley and Sons, Inc., New York, NY.
Broecker, W. S., and T. Takahashi. 1966. Calcium carbonate precipitation on Bahama Banks. Journal of Geophysical Research 71:1575‐1602.
Brueggeman, J. J., D. P. Volsen, R. A. Grotefendt, G. A. Green, J. J. Burns, and D. K. Ljungblad. 1991. 1990 walrus monitoring program: the popcorn, burger, and crackerjack prospects in the Chukchi Sea. Report from EBASCO Environmental, Bellevue, WA, for Shell Western E&P Inc. and Chevron USA Inc. Shell Western E&P Inc., Houston, TX. 53 p.
Burns, J. J. 1970. Remarks on the distribution and natural history of pagophilic pinnipeds in the Bering and Chukchi Seas. Journal of Mammalogy 51:445‐454.
Burns, J. J., and T. J. Eley. 1976. The natural history and ecology of the bearded seal (Erignathus barbatus) and the ringed seal (Phoca (Pusa) hispida). Pages 263‐294 in Environmental Assessment of the Alaskan Continental Shelf. Annual Reports from Principal Investigators. April 1976. Volume 1 Marine Mammals. U.S. Department of Commerce, NOAA, Boulder, CO.
Burns, J. J., and T. J. Eley. 1978. The natural history and ecology of the bearded seal (Erignathus barbatus) and the ringed seal (Phoca hispida). Pages 99‐160 in Environmental Assessment of the Alaskan Continental Shelf. Annual Reports of Principal Investigators for the year ending March 1978. Volume 1. Receptors‐‐Mammals‐Birds. U.S. Department of Commerce, NOAA and U.S. Department of the Interior, Bureau of Land Management, Boulder, CO.
Burns, J. J., and F. H. Fay. 1970. Comparative morphology of the skull of the ribbon seal, Histriophoca fasciata, with remarks on systematics of Phocidae. Journal of Zoology 161:363‐394.
Burns, J. J., and S. J. Harbo. 1972. An aerial census of ringed seals, northern coast of Alaska. Arctic 25:179‐290.
Burns, J. J., and B. P. Kelly. 1982. Studies of ringed seals in the Alaskan Beaufort Sea during winter: impacts of seismic exploration. Alaska Department of Fish and Game, Annual Report. 57 p.
Burns, J. J., L. H. Shapiro, and F. H. Fay. 1981. Ice as marine mammal habitat in the Bering Sea. Pages 781‐797 in D. W. Hood and J. A. Calder, editors. The Eastern Bering Sea Shelf: Oceanography and Resources. Volume Two. U.S. Department of Commerce, NOAA, U.S. Department of Interior, Office of Marine Pollution Assessment, and Bureau of Land Management, Washington, D.C.
Butler, J., A. DeVogelaere, R. Gustafson, C. Mobley, M. Neuman, D. Richards, S. Rumsey, B. Taylor, and G. VanBlaricom. 2009. Status review report for black abalone. National Marine Fisheries Service, Southwest Region, Long Beach, CA. 135 p.
Butt, C. M., D. C. G. Muir, I. Stirling, M. Kwan, and S. A. Mabury. 2007. Rapid response of Arctic ringed seals to changes in perfluoroalkyl production. Environmental Science & Technology 41:42‐49.
Bychkov, V. A. 1965. Autumn molting in seals. Bulletin of the Moscow Society of Naturalists, Biological Division 70:113‐118.
Byrne, R. H., J. G. Acker, P. R. Betzer, R. A. Feely, and M. H. Cates. 1984. Water column dissolution of aragonite in the Pacific Ocean. Nature 312:321‐326.
Caldeira, K., and M. E. Wickett. 2003. Anthropogenic carbon and ocean pH. Nature 425:365. Caldeira, K., and M. E. Wickett. 2005. Ocean model predictions of chemistry changes from carbon
dioxide emissions to the atmosphere and ocean. Journal of Geophysical Research 110:C09S04.

203
Calle, P. P., D. J. Seagars, C. McClave, D. Senne, C. House, and J. A. House. 2008. Viral and bacterial serology of six free‐ranging bearded seals Erignathus barbatus. Diseases of Aquatic Organisms 81:77‐80.
Cameron, M. E., T. L. Metcalfe, C. D. Metcalfe, and C. R. Macdonald. 1997. Persistent organochlorine compounds in the blubber of ringed seals (Phoca hispida) from the Belcher Islands, Northwest Territories, Canada. Marine Environmental Research 43:99‐116.
Canadell, J. G., C. Le Quere, M. R. Raupach, C. B. Field, E. T. Buitenhuis, P. Ciais, T. J. Conway, N. P. Gillett, R. A. Houghton, and G. Marland. 2007. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proceedings of the National Academy of Sciences of the United States of America 104:18866‐18870.
Cao, L., and K. Caldeira. 2008. Atmospheric CO2 stabilization and ocean acidification. Geophysical Research Letters 35:L19609.
CAPP. 2010. Offshore drilling. Canadian Association of Petroleum Producers, Calgary, Canada. Accessed June 2010 at http://www.capp.ca/canadaIndustry/oil/offshore/Pages/default.aspx#T869miWhlfg8.
Carey, A. G. 1992. The ice fauna in the shallow southwestern Beaufort Sea, Arctic Ocean. Journal of Marine Systems 3:225‐236.
Carlberg, G. E., and J. B. Bøler. 1985. Determination of persistent chlorinated hydrocarbons and inorganic elements in samples from Svalbard. Pages 1‐18 in Report No. 8311 01‐1. Center for Industrial Research, Oslo, Norway.
Carlens, H., C. Lydersen, B. A. Krafft, and K. M. Kovacs. 2006. Spring haul‐out behavior of ringed seals (Pusa hispida) in Kongsfjorden, Svalbard. Marine Mammal Science 22:379‐393.
Carroll, J., H. Wolkers, M. Andersen, and K. Rissanen. 2002. Bioaccumulation of radiocaesium in Arctic seals. Marine Pollution Bulletin 44:1366‐1371.
Cattet, M. R. L., P. J. Duignan, C. A. House, and D. J. St Aubin. 2004. Antibodies to canine distemper and phocine distemper viruses in polar bears from the Canadian Arctic. Journal of Wildlife Diseases 40:338‐342.
Center for Biological Diversity. 2008. Petition to list three seal species under the Endangered Species Act: ringed seal (Pusa hispida), bearded seal (Erignathus barbatus), and spotted seal (Phoca largha). 139 p. Center for Biological Diversity, San Francisco, CA.
Chapskii, K. K. 1940. The ringed seal of western seas of the Soviet Arctic (The morphological characteristic, biology and hunting production). Page 147 in N. A. Smirnov, editor. Proceedings of the Arctic Scientific Research Institute, Chief Administration of the Northern Sea Route. Izd. Glavsevmorputi, Leningrad, Moscow. (Translated from Russian by the Fisheries Research Board of Canada, Ottawa, Canada, Translation Series No. 1665, 147 p.).
Chapskii, K. K. 1955. An attempt at revision of the systematics and diagnostics of seals in the subfamily Phocinae. Trudy Zoologicheskovo Instituta Akademii Nauk SSSR 17:160‐199. (Translated from Russian by T.F. Jeletzky, Fisheries Research Board of Canada, Montreal, Canada, Translation Series No. 114, 57 p.).
Chernenko, T. 2007. Appendix 1. The status of the yellow‐billed loon in Russia: a summary of Russian language information. Comments submitted by Center for Biological Diversity, Pacific Environmental, and Natural Resources Defence Council on August 6, 2007, to the U.S. Fish and Wildlife Service on the 90‐day Finding and Status Review of the Yellow‐billed Loon. 16 p.
Choi, K. 2008. Recent trend in design parameters of ice‐transiting vessels. International Journal of Offshore and Polar Engineering 18:282‐287. (Abstract online; Accessed May 2010 at http://www.isope.org/publications/journals/ijope‐18‐4/abst‐18‐4‐p282‐RF‐40‐Choi.pdf).

204
Ciannelli, L., K. M. Bailey, K. S. Chan, A. Belgrano, and N. C. Stenseth. 2005. Climate change causing phase transitions of walleye pollock (Theragra chalcogramma) recruitment dynamics. Proceedings of the Royal Society B 272:1735‐1743.
Clarkson, P. L., and D. Irish. 1991. Den collapse kills female polar bear and two newborn cubs. Arctic 44:83‐84.
Coffing, M., C. L. Scott, and C. J. Utermohle. 1998. The subsistence harvest of seals and seal lions by Alaska Natives in three communities of the Yukon‐Kuskokwim Delta, Alaska, 1997‐98. Alaska Department of Fish and Game, Division of Subsistence, Technical Paper No. 255. 56 p.
Cohen, D. M., T. Inada, T. Iwamoto, and N. Scialabba. 1990. FAO species catalogue. Vol. 10. Gadiform fishes of the world (Order Gadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers and other gadiform fishes known to date. Food and Agriculture Organization of the United Nations, FAO Fisheries Synopsis No. 125. 442 p.
Cole, R. A., D. S. Lindsay, D. K. Howe, C. L. Roderick, J. P. Dubey, N. J. Thomas, and L. A. Baeten. 2000. Biological and molecular characterizations of Toxoplasma gondii strains obtained from southern sea otters (Enhydra lutris nereis). Journal of Parasitology 86:526‐530.
Colton, J. B., Jr. 1972. Temperature trends and distribution of groundfish in continental shelf waters, Nova Scotia to Long Island. Fishery Bulletin 70:637‐657.
Comeau, S., G. Gorsky, R. Jeffree, J. L. Teyssié, and J. P. Gattuso. 2009. Key Arctic pelagic mollusc (Limacina helicina) threatened by ocean acidification. Biogeosciences Discussions 6:2523‐2537.
Commission for Nature Conservation. 2008. Estonian Red List of Threatened Species. Commission for Nature Conservation, Estonian Academy of Sciences, Tartu, Estonia. Accessed February 2010 at http://elurikkus.ut.ee/prmt.php?lang=eng.
Commission of the European Communities. 2009. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions Concerning the European Union Strategy for the Baltic Sea Region. Action Plan. Commission of the European Communities, Com(2009) 248 Final. 11 p.
Committee on the Status of Endangered Wildlife in Canada. 2009. Canadian wildlife species at risk. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ontario, Canada. 92 p.
Conners, M. E., A. B. Hollowed, and E. Brown. 2002. Retrospective analysis of Bering Sea bottom trawl surveys: regime shift and ocean reorganization. Progress in Oceanography 55:209‐222.
Cooper, L. W., I. L. Larsen, T. M. O'Hara, S. Dovlin, V. Woshner, and G. F. Cota. 2000. Radionuclide contaminant burdens in arctic marine mammals harvested during subsistence hunting. Arctic 53:174‐182.
Cott, P. A., B. W. Hanna, and J. A. Dahl. 2003. Discussion on seismic exploration in the Northwest Territories 2000‐2003. Canadian Manuscript Report for Fisheries and Aquatic Sciences 2648. 36 p.
COWI. 2008. Assessment of the potential impact of a ban of products derived from seal species. European Commission Direcorate‐General Environment. 45 p.
Craig, P. C. 1984. Fish use of coastal waters of the Alaska Beaufort Sea: a review. Transactions of the American Fisheries Society 113:265‐282.
Cranstone, D. A. 2002. A history of mining and mineral exploration in Canada and outlook for the future. Natural Resources Canada, Ottawa, Canada. 44 p. Accessed at http://www.nrcan‐rncan.gc.ca/mms‐smm/busi‐indu/cmy‐amc/arch‐arch/hist‐eng.pdf.
Crocker, C. E., and J. J. Cech. 1996. The effects of hypercapnia on the growth of juvenile white sturgeon, Acipenser transmontanus. Aquaculture 147:293‐299.
Curry‐Lindahl, K. 1975. Ecology and conservation of the grey seal Halichoerus grypus, common seal Phoca vitulina, and ringed seal, Pusa hispida, in the Baltic Sea. Biology of the Seal. Proceedings

205
of a Symposium held in Guelph 14‐17 August 1972. Rapports et Proces‐verbaux des Réunions. Conseil International pour l'Éxploration de la Mer. 169:527‐532.
Dalpadado, P. 2002. Inter‐specific variations in distribution, abundance and possible life‐cycle patterns of Themisto spp. (Amphipoda) in the Barents Sea. Polar Biology 25:656‐666.
Dalpadado, P., R. Ingvaldsen, and A. Hassel. 2003. Zooplankton biomass variation in relation to climatic conditions in the Barents Sea. Polar Biology 36:233‐241.
Danilov‐Danilyan, V. I. 2001. Red book of the Russian Federation. 1 p. Accessed May 2010 at http://www.sevin.ru/redbook/index.html.
Darnerud, P. O. 2003. Toxic effects of brominated flame retardants in man and in wildlife. Environment International 29:841‐853.
Davies, J. L. 1958. The Pinnipedia: an essay in zoogeography. Geographical Review 48:474‐493. Davis, C. S. 2004. Phylogenetic relationships of the Phocidae and populations genetics of ice breeding
seals. Ph. D. Dissertation. University of Alberta Edmonton, Canada. 166 p. Davis, C. S., I. Stirling, C. Strobeck, and D. W. Coltman. 2008. Population structure of ice‐breeding seals.
Molecular Biology 17:3078‐3094. Davis, J. E., and S. S. Anderson. 1976. Effects of oil pollution on breeding grey seals. Marine Pollution
Bulletin 7:115‐118. De Muizon, C. 1982. Phocid phylogeny and dispersal. Annals of the South African Museum 89:175‐213. de Wit, C. A., M. Alaee, and D. C. G. Muir. 2006. Levels and trends of brominated flame retardants in the
Arctic. Chemosphere 64:209‐233. Deepwater Horizon Response. 2010. August 2, 2010 news release: U.S. scientific teams refine estimates
of oil flow from BP’s well prior to capping. Unified Command, Joint Information Center, Accessed September 2010 at http://www.restorethegulf.gov/release/2010/08/02/us‐scientific‐teams‐refine‐estimates‐oil‐flow‐bps‐well‐prior‐capping.
Degerbøl, M., and P. Freuchen. 1935. Report of the mammals collected by the Fifth Thule Expedition to the Arctic. North America: Part I. Systematic Notes. Report of the Fifth Thule Expedition 2:1‐67.
Dehn, L.‐A., G. Sheffield, E. Follmann, L. Duffy, D. Thomas, and T. O’Hara. 2007. Feeding ecology of phocid seals and some walrus in the Alaskan and Canadian Arctic as determined by stomach contents and stable isotope analysis. Polar Biology 30:167‐181.
Dehn, L.‐A., G. G. Sheffield, E. H. Follmann, L. K. Duffy, D. L. Thomas, G. R. Bratton, R. J. Taylor, and T. M. O'Hara. 2005. Trace elements in tissues of phocid seals harvested in the Alaskan and Canadian Arctic: influence of age and feeding ecology. Canadian Journal of Zoology 83:726‐746.
Dehnhardt, G., B. Mauck, and H. Bleckmann. 1998. Seal whiskers detect water movements. Nature 394:235‐236.
Delyamure, S. L., and E. V. Alekseev. 1966. Parafilaroides arcticus n. sp. ‐ a parasite of the Chukchee Sea ringed seal. Problemy Parazitologii 6:11‐15. (Translated from Russian by the Canadian Translation of Fisheries and Aquatic Sciences, No. 5564, 7 p.).
Delyamure, S. L., V. N. Popov, and A. N. Trashchenkov. 1980. Study of the helminth fauna of the seals of the Baltic Sea and Lake Ladoga. Doklady Vysshei Shkoly, Biologicheskie Nauki (University Scientific Reports, Biological Sciences) 7:43‐45. (Translated from Russian by the Canada Institute for Scientific and Technical Information, Ottawa, Ontario, Canada, Translation Series No. 5112, 7 p.).
Delyamure, S. L., M. V. Yurakhno, and V. N. Popov. 1976. On the helminth fauna of pinnipeds from the Karaginsk Gulf (the Bering Sea). Parazitologiya 10:325‐332. (In Russian, English Summary).
Delyamure, S. L., M. V. Yurakhno, V. N. Popov, L. M. Shults, and F. H. Fay. 1984. Helminthological comparison of subpopulations of Bering Sea spotted seals, Phoca largha Pallas. Pages 61‐66 in F. H. Fay and G. A. Fedoseev, editors. Soviet‐American Cooperative Research on Marine Mammals. Volume 1 ‐ Pinnipeds. Under Project V.6. Marine Mammals, of the US‐USSR Agreement on

206
Cooperation in the Field of Environmental Protection. U.S. Department of Commerce. NOAA Technical Report NMFS 12, Washington, D.C.
Delyamure, S. L., M. V. Yurakhno, V. N. Popov, and V. V. Treschev. 1978. Helminth fauna of the ringed seal in various parts of its range. Pages 104‐106 in Marine Mammals. Abstracts of papers 7th All‐Union Conference, Moscow, Russia. Ichthilogical Commission, Ministry of Fisheries.
Demere, T. A., A. Berta, and P. J. Adam. 2003. Pinnipedimorph evolutionary biogeography. Bulletin of the American Museum of Natural History 279:32‐76.
Department of Fisheries and Oceans. 2007. An integrated Aboriginal policy framework 2006‐2010. Fisheries and Oceans Canada, Ottawa, Ontario, Canada. 33 p. Accessed May 2010 at http://www.dfo‐mpo.gc.ca/fm‐gp/aboriginal‐autochtones/IAPF‐CIPA‐eng.pdf.
Department of Fisheries and Oceans. 2008. Overview of the Atlantic seal hunt 2006‐2010. Fisheries and Oceans Canada, Ottawa, Canada. Accessed April 2010 at http://www.dfo‐mpo.gc.ca/fm‐gp/seal‐phoque/reports‐rapports/mgtplan‐plangest0610/mgtplan‐plangest0610‐eng.htm.
Derocher, A. E., N. J. Lunn, and I. Stirling. 2004. Polar bears in a warming climate. Integrative and Comparative Biology 44:163‐176.
Dietz, R. 2008. Contaminants in marine mammals in Greenland ‐‐ with linkages to trophic levels, effects, diseases and distribution. DSc Thesis. University of Aarhus, Denmark, Copenhagen, Denmark. 120 p.
Dietz, R., E. W. Born, C. T. Agger, and C. O. Nielsen. 1995. Zinc, cadmium, mercury and selenium in polar bears (Ursus maritimus) from Central East Greenland. Polar Biology 15:175‐185.
Dietz, R., P. Paludan‐Müller, C. T. Agger, and C. O. Nielsen. 1998. Cadmium, mercury, zinc and selenium in ringed seals (Phoca hispida) from Greenland and Svalbard. Pages 242‐273 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Directorate of Fisheries. 2010. Regulations on quotas in the hunting of coastal seals 2010. Directorate of Fisheries, Bergen, Norway. Accessed May 2010 at http://www.fiskeridir.no/fiske‐og‐fangst/j‐meldinger/gjeldende‐j‐meldinger/j‐225‐2009. (In Norwegian).
Doney, S. C., V. J. Fabry, R. A. Feely, and J. A. Kleypas. 2009. Ocean acidification: the other CO2 problem. Annual Review of Marine Science 1:169‐192.
Drobot, S., and M. Anderson. 2001, updated 2009. Snow melt onset over Arctic sea ice from SMMR and SSM/I brightness temperatures. National Snow and Ice Data Center, Boulder, CO. Accessed June 2010 at http://nsidc.org/data/docs/daac/nsidc0105_arctic_snowmelt_onset_dates.gd.html.
Dubey, J. P., R. Zarnke, N. J. Thomas, S. K. Wong, W. Van Bonn, M. Briggs, J. W. Davis, R. Ewing, M. Mense, O. C. H. Kwok, S. Romand, and P. Thulliez. 2003. Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis‐like infections in marine mammals. Veterinary Parasitology 116:275‐296.
Duignan, P. J., O. Nielsen, C. House, K. M. Kovacs, N. Duffy, G. Early, S. Sadove, D. J. St. Aubin, B. K. Rima, and J. R. Geraci. 1997. Epizootiology of morbillivirus infection in harp, hooded, and ringed seals from the Canadian Arctic and western Atlantic. Journal of Wildlife Diseases 33:7‐19.
Dulepova, E., and V. Radchencko. 2004. Okhotsk Sea. Pages 95‐111 in PICES Special Publication: Marine Ecosystems of the North Pacific. North Pacific Marine Science Organization, Sidney, Canada.
Dulepova, E. P., and L. B. Klyashtorin. 2008. Russia. Pages 137‐162 in R. J. Beamish, editor. Impacts of climate and climate change on the key species in the fisheries in the North Pacific. PICES Scientific Report, No. 35. North Pacific Marine Science Organization, Sidney, Canada.
Dunbar, M. J. 1941. On the food of seals in the Canadian eastern Arctic. Canadian Journal of Research 19:150‐155.

207
Earth System Research Laboratory. 2010. Physical sciences division (Online database). U.S. Department of Commerce, NOAA, Washington, D.C. Accessed June 2010 at http://www.esrl.noaa.gov/psd/cgi‐bin/data/composites/comp.pl.
Ebbesmeyer, C. C., D. R. Cayan, D. R. McLain, F. H. Nichols, D. H. Peterson, and K. T. Redmond. 1991. 1976 step in the Pacific climate: forty environmental changes between 1968‐1975 and 1977‐1984. Pages 115‐126 in J. L. Betancourt and V. L. Tharp, editors. Proceedings of the Seventh Annual Pacific Climate (PACLIM) Workshop, April 1990. California Department of Water Resources. Interagency Ecological Studies Program Technical Report 26, Asilomar, CA.
ECON. 2007. Arctic shipping 2030: from Russia with oil, stormy passage, or Arctic great game? Norshipping, ECON Report No. 2007‐070, Project No. 50450. 45 p.
Eley, T. J. 1981. Dipetalonema spirocauda in Alaskan marine mammals. Journal of Wildlife Diseases 17:65‐67.
Ellis, B. 2008. Arctic transport today and tomorrow: implications of a changing marine environment at the top of the world (Presentation). Institute of the North, Moscow, Russia. Accessed at http://www.institutenorth.org/servlet/content/presentations.html.
Elmgren, R. 1989. Man’s impact on the ecosystem of the Baltic Sea: energy flows today and at the turn of the century. Ambio 18:326‐332.
Elsner, R., D. Wartzok, N. B. Sonafrank, and B. P. Kelly. 1989. Behavioral and physiological reactions of Arctic seals during under‐ice pilotage. Canadian Journal of Zoology 67:2506‐2513.
Engelhardt, F. R. 1982. Hydrocarbon metabolism and cortisol balance in oil‐exposed ringed seals, Phoca hispida. Comparative Biochemistry and Physiology C 72:133‐136.
Engelhardt, F. R. 1983. Petroleum effects on marine mammals. Aquatic Toxicology 4:199‐217. Engelhardt, F. R., J. R. Geraci, and T. G. Smith. 1977. Uptake and clearance of petroleum hydrocarbons in
the ringed seal, Phoca hispida. Journal of the Fisheries Research Board of Canada 34:1143‐1147. Eschmeyer, W. M., and E. S. Herald. 1983. A field guide to Pacific coast fishes of North America from the
Gulf of Alaska to Baja California. Houghton Mifflin, Boston, MA. European Commission. 2010. May 5, 2010 news release: Environment: Commission asks Finland to
protect its endangered native seals. European Commission, Brussels, Germany. Accessed May 2010 at http://europa.eu/rapid/pressReleasesAction.do?reference=IP/10/523&type=HhTML.
European Mammal Assessment Team. 2007. Pusa hispida, IUCN Red List of Threatened Species. Version 2010.1. International Union for Conservation of Nature and Natural Resources (IUCN), Cambridge, UK. Accessed March 2010 at http://www.iucnredlist.org/apps/redlist/details/41672/1/full.
Ewalt, D. R., J. B. Payeur, B. M. Martin, D. R. Cummins, and W. G. Miller. 1994. Characteristics of a Brucella species from a bottlenose dolphin (Tursiops truncatus). Journal of Veteterinary Investigations 6:448‐452.
Fant, M. L., M. Nyman, E. Helle, and E. Rudback. 2001. Mercury, cadmium, lead and selenium in ringed seals (Phoca hispida) from the Baltic Sea and from Svalbard. Environmental Pollution 111:493‐501.
FAO Advisory Committee. 1979. Mammals in the seas. Food and Agriculture Organization of the United Nations, Rome, Italy. 151 p.
Fay, F. H. 1974. The role of ice in the ecology of marine mammals of the Bering Sea. Pages 383‐399 in D. W. Hood and E. J. Kelley, editors. Oceanography of the Bering Sea. Institute of Marine Science, Hakodate, Japan.
Fay, F. H., R. A. Dieterich, L. M. Shults, N. K. Murray, A. Hoover, and B. P. Kelly. 1979. Morbidity and mortality of marine mammals. Pages 1‐34 in Environmental Assessment of the Alaskan Continental Shelf. Annual Reports of Principal Investigators for the Year Ending March 1979.

208
Volume 1. Receptors‐‐Mammals‐Birds. U.S. Department of Commerce, NOAA and U.S. Department of the Interior, Bureau of Land Management, Boulder, CO.
Fay, F. H., and D. P. Furman. 1982. Nasal mites (Acari: Halarachnidae) in the spotted seal, Phoca largha Pallas, and other pinnipeds of Alaskan waters. Journal of Wildlife Diseases 18:63‐68.
Fay, F. H., J. L. Sease, and R. L. Merrick. 1990. Predation on a ringed seal, Phoca hispida , and a black guillemot, Cepphus grylle , by a Pacific walrus, Odobenus rosmarus divergens. Marine Mammal Science 6:348‐350.
Fay, F. H., and L. M. Shults. 1978. An amendment to RU 232 trophic relationships among ice inhabiting phocid seals, and RU 230 natural history of ringed and bearded seals. U.S. National Oceanic and Atmospheric Administration, Boulder, CO. Environmental Research Laboratories, Environmental Assessment of the Alaskan Continental Shelf. Annual Reports of Principal Investigators for the Year Ending March 1978. Volume 1. Mammals‐‐Birds. 366‐372 p.
Fedoseev, G. A. 1965a. Comparative characteristics of ringed seal populations in the Chukotskii peninsula coastal waters. Izvestiya TINRO 59:194‐212. (Translated from Russian by the Fisheries Marine Service, Quebec, Canada, Translation Series No. 3428, 38 p.).
Fedoseev, G. A. 1965b. The ecology of the reproduction of seals on the northern part of the Sea of Okhotsk. Izvestiya TINRO 65:212‐216. (Translated from Russian by the Fisheries and Marine Service, Quebec, Canada, Translation Series No. 3369, 8 p.).
Fedoseev, G. A. 1965c. Food of the ringed seal. U.S. National Oceanic and Atmospheric Administration, Boulder, CO. Environmental Research Laboratories, Environmental Assessment of the Alaskan Continental Shelf. Annual Reports for Principal Investigators for the Year Ending March 1977. Volume 1. Receptors‐‐Mammals. 373‐383 p. (Translated from Russian by Meg L. Poppino, 11 p.).
Fedoseev, G. A. 1968. Determination of abundance and grounds for establishing the catch quota for ringed seals in the Sea of Okhotsk. Pages 180‐188 in A. V. A. and K. I. Panin, editors. Pinnipeds of the North Pacific Israel Program for Scientific Translations, Jerusalem, Israel.
Fedoseev, G. A. 1970. Distribution and numerical strength of seals off Sakhalin Island. Izvestiya TINRO 71:319‐324. (Translated from Russian by Department of the Environment, Fisheries Research Board of Canada, Translation Series No. 2400, 15 p.).
Fedoseev, G. A. 1971a. Determining numbers and basis for establishing the catch quota of ringed seals in the Sea of Okhotsk Pages 175‐183 in V. A. Arsen'ev and K. I. Panin, editors. Pinnipeds of the North Pacific. Israel Program for Scientific Translations, Jerusalem, Israel.
Fedoseev, G. A. 1971b. The distribution and numbers of seals on whelping and moulting patches in the Sea of Okhotsk. Pages 87‐99 in K. K. Chapskii and E. S. Mil'chenko, editors. Research on Marine Mammals. Atlantic Research Institute of Marine Fisheries and Oceanography (AtlantNIRO), Kaliningrad, Russia. (Translated from Russian by Fisheries and Marine Service, Canada, 24 p.).
Fedoseev, G. A. 1972. Morphoecological types of the populations of ringed seal (Pusa hispida Schreber, 1777) (Abstract). International Council for the Exploration of the Sea, Symposium on the Biology of the Seal. 14‐17 August 1972, University of Guelph, Ontario, Canada. 3 p.
Fedoseev, G. A. 1975. Ecotypes of the ringed seal (Pusa hispida Schreber, 1777) and their reproductive capabilities. Biology of the Seal. Proceedings of a Symposium held in Guelph 14‐17 August 1972. Rapports et Proces‐verbaux des Réunions. Conseil International pour l'Éxploration de la Mer. 169:156‐160.
Fedoseev, G. A. 1984. Population structure, current status, and perspective for utilization of the ice‐inhabiting forms of pinnipeds in the northern part of the Pacific Ocean Pages 130‐146 in A. V. Yablokov, editor. Marine Mammals. Nauka, Moscow, Russia. (Translated from Russian by F. H. Fay and B. A. Fay, 17 p.).

209
Fedoseev, G. A. 2000. Population biology of ice‐associated forms of seals and their role in the northern Pacific ecosystems. Center for Russian Environmental Policy, Russian Marine Mammal Council, Moscow, Russia. 271 p. (Translated from Russian by I. E. Sidorova, 271 p.).
Fedoseev, G. A., and Y. I. Nazarenko. 1970. On intraspecific structure of ringed seals in the Arctic. Izvestiya TINRO 71:301‐307. (Translated from Russian by the Department of the Environment, Fisheries Research Board of Canada, Quebec, Canada, Translation Series No. 2411, 11 p.).
Feely, R. A., R. H. Byrne, J. G. Acker, P. R. Betzer, C. T. A. Chen, J. F. Gendron, and M. F. Lamb. 1988. Winter‐summer variations of calcite and aragonite saturation in the Northeast Pacific. Marine Chemistry 25:227‐241.
Feely, R. A., and C. T. A. Chen. 1982. The effect of excess CO2 on the calculated calcite and aragonite saturation horizons in the Northeast Pacific. Geophysical Research Letters 9:1294‐1297.
Feely, R. A., S. C. Doney, and S. R. Cooley. 2009. Ocean acidification: present conditions and future changes in a high‐CO2 world. Oceanography 22:36‐47.
Feely, R. A., C. L. Sabine, J. M. Hernandez‐Ayon, D. Ianson, and B. Hales. 2008. Evidence for upwelling of corrosive "acidified" water onto the continental shelf. Science 320:1490‐1492.
Feely, R. A., C. L. Sabine, K. Lee, W. Berelson, J. Kleypas, V. J. Fabry, and F. J. Millero. 2004. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 305:362‐366.
Feltz, E. T., and F. H. Fay. 1966. Thermal requirements in vitro of epidermal cells from seals. Cryobiology 3:261‐264.
Ferguson, S., and C. S. A. Secretariat. 2009. Review of aerial survey estimates for ringed seals (Phoca hispida) in western Hudson Bay. Center for Science Advice, Central and Arctic Region, Fisheries and Oceans Canada, Science Advisory Report 2009/004. 5 p.
Ferguson, S. H., I. Stirling, and P. McLoughlin. 2005. Climate change and ringed seal (Phoca hispida) recruitment in western Hudson Bay. Marine Mammal Science 21:121‐135.
Filatov, I. E. 1990. Ladozhskaja kolchataja nerpa. Pages 57‐64 in V. E. Sokolov and A. L. Janshin, editors. Redkie ischezaushcije vidy mlekopitajushikh SSSR. Nauka, Moscow, Russia. (In Russian).
Finley, K. J. 1979. Haul‐out behaviour and densities of ringed seals (Phoca hispida) in the Barrow Strait area, N.W.T. Canadian Journal of Zoology 57:1985‐1997.
Finley, K. J., G. W. Miller, R. A. Davis, and W. R. Koski. 1983. A distinctive large breeding population of ringed seals (Phoca hispida) inhabiting the Baffin Bay pack ice. Arctic 36:162‐173.
Fisk, A. T., C. A. de Wit, M. Wayland, Z. Z. Kuzyk, N. Burgess, R. Robert, B. Braune, R. Norstrom, S. P. Blum, C. Sandau, E. Lie, H. J. S. Larsen, J. U. Skaare, and D. C. G. Muir. 2005. An assessment of the toxicological significance of anthropogenic contaminants in Canadian arctic wildlife. Science of the Total Environment 351:57‐93.
Fisk, A. T., K. A. Hobson, and R. J. Norstrom. 2001. Influence of chemical and biological factors on trophic transfer of persistent organic pollutants in the northwater polynya marine food web. Environmental Science & Technology 35:732‐738.
Fisk, A. T., M. Holst, K. A. Hobson, J. Duffe, J. Moisey, and R. J. Norstrom. 2002. Persistent organochlorine contaminants and enantiomeric signatures of chiral pollutants in ringed seals (Phoca hispida) collected on the east and west side of the northwater polynya, Canadian Arctic. Archives of Environmental Contamination and Toxicology 42:118‐126.
Flinkman, J., E. Aro, I. Vuorinen, and M. Viitasalo. 1998. Changes in northern Baltic zooplankton and herring nutrition from 1980s to 1990s: top‐down and bottom‐up processes at work. Marine Ecology Progress Series 165:127‐136.
Forbes, L. B., O. Nielsen, L. Measures, and D. R. Ewalt. 2000. Brucellosis in ringed seals and harp seals from Canada. Journal of Wildlife Diseases 36:595‐598.
Førland, E. J., and I. Hanssen‐Bauer. 2003. Past and future climate variations in the Norwegian Arctic: overview and novel analyses. Polar Research 22:113‐124.

210
Forstén, A., and P. Alhonen. 1975. The subfossil seals of Finland and their relation to the history of the Baltic Sea. Boreas 4:143‐155.
Fouche, G., and W. Moskwa. 2010. Arctic border deal may extend Norway's oil boom. Commodities Now, London, England. Accessed June 2010 at http://www.commodities‐now.com/news/power‐and‐energy/2502‐arctic‐border‐deal‐may‐extend‐norways‐oil‐boom.html.
Frantzen, E. 2007. From Russia with love. Hart Energy Publishing, LP, Houston, TX. Accessed May 2010 at http://www.epmag.com/archives/features/405.htm.
Freitas, C., K. M. Kovacs, R. A. Ims, M. A. Fedak, and C. Lydersen. 2008. Ringed seal post‐moulting movement tactics and habitat selection. Oecologia 155:193‐204.
Freuchen, P. 1935. Mammals. Part II. Field notes and biological observations. Pages 66‐218 in M. Degerbol and P. Freuchen, editors. Mammals. Gyldendalske Boghandel, Nordisk Forlag, Copenhagen, Denmark.
Frost, K. J. 1985. The ringed seal (Phoca hispida). Pages 79‐87 in J. J. Burns, K. J. Frost, and L. F. Lowry, editors. Marine Mammals Species Accounts. Alaska Department Fish and Game, Juneau, AK.
Frost, K. J., and L. Lowry. 1981. Ringed, Baikal and Caspian seals ‐‐ Phoca hispida Schrebner, 1775 Phoca sibrinica Gmelin, 1758 and Phoca caspica Gmelin, 1788. Pages 29‐53 in S. H. Ridgeway and R. J. Harrison, editors. Handbook of Marine Mammals Volume 2: Seals. Academic Press, New York.
Frost, K. J., and L. F. Lowry. 1984. Trophic relationships of vertebrate consumers in the Alaskan Beaufort Sea. Pages 381‐401 in P. W. Barnes, D. M. Schell, and E. Reimnitz, editors. The Alaskan Beaufort Sea ‐‐ Ecosystems and Environments. Academic Press, Inc., New York, NY.
Frost, K. J., and L. F. Lowry. 1994. Assessment injury to harbor seals in Prince William Sound, Alaska, and adjacent areas following the Exxon Valdez oil spill. Alaska Department of Fish and Game, Wildlife Conservation Division, Exxon Valdez Oil Spill. State/Federal Natural Resource Damage Assessment Final Report. 157 p.
Frost, K. J., L. F. Lowry, G. Pendleton, and H. R. Nute. 2004. Factors affecting the observed densities of ringed seals, Phoca hispida, in the Alaskan Beaufort Sea, 1996‐99. Arctic 57:115‐128.
Frost, K. J., L. F. Lowry, E. H. Sinclair, J. Ver Hoef, and D. C. McAllister. 1994a. Impacts on distribution, abundance, and productivity of harbor seals. Pages 97‐118 in T. R. Loughlin, editor. Marine Mammals and the Exxon Valdez. Academic Press, Inc., San Diego, CA.
Frost, K. J., C. A. Manen, and T. L. Wade. 1994b. Petroleum hydrocarbons in tissues of harbor seals from Prince William Sound and the Gulf of Alaska. Pages 331‐358 in T. R. Loughlin, editor. Marine Mammals and the Exxon Valdez. Academic Press, Inc., San Diego, CA.
Fujii, K., C. Kakumoto, M. Kobayashi, S. Saito, T. Kariya, Y. Watanabe, X. N. Xuan, I. Igarashi, and M. Suzuki. 2007. Seroepidemiology of Toxoplasma gondii and Neospora caninum in seals around Hokkaido, Japan. Journal of Veterinary Medical Science 69:393‐398.
Fujita, S., I. Chiba, M. Ishizuka, H. Hoshi, H. Iwata, A. Sakakibara, S. Tanabe, A. Kazusaka, M. Masuda, Y. Masuda, and H. Nakagawa. 2001. P450 in wild animals as a biomarker of environmental impact. Biomarkers 6:19‐25.
Fulton, T. L., and C. Strobeck. 2010a. Multiple fossil calibrations, nuclear loci and mitochondrial genomes provide new insight into biogeography and divergence timing for true seals (Phocidae, Pinnipedia). Journal of Biogeography 37:814‐829.
Fulton, T. L., and C. Strobeck. 2010b. Multiple markers and multiple individuals refine true seal phylogeny and bring molecules and morphology back in line. Proceedings of the Royal Society B 277:1065‐1070.
Furgal, C. M., S. Innes, and K. M. Kovacs. 1996. Characteristics of ringed seal, Phoca hispida, subnivean structures and breeding habitat and their effects on predation. Canadian Journal of Zoology 74:858‐874.

211
Furnell, D. J., and D. Oolooyuk. 1980. Polar bear predation on ringed seals in ice‐free water. Canadian Field‐Naturalist 94:88‐89.
Gaden, A., S. H. Ferguson, L. Harwood, H. Melling, and G. A. Stern. 2009. Mercury trends in ringed seals (Phoca hispida) from the western Canadian Arctic since 1973: associations with length of ice‐free season. Environmental Science & Technology 43:3646‐3651.
Gauthier, S., and G. A. Rose. 2005. Diel vertical migration and shoaling heterogeneity in Atlantic redfish: effects on acoustic and bottom‐trawl surveys. ICES Journal of Marine Science 62:75‐85.
Gehlen, M., R. Gangsto, B. Schneider, L. Bopp, O. Aumont, and C. Ethe. 2007. The fate of pelagic CaCO3 production in a high CO2 ocean: a model study. Biogeosciences 4:505‐519.
Geological Survey of Canada. 2007. Arctic oil and gas. Geological Association of Canada, St. John's, Newfoundland. 2 p. Accessed at http://www.gac.ca/PopularGeoscience/factsheets/ArcticOilandGas_e.pdf.
Georgette, S., M. Coffing, C. Scott, and C. Utermohle. 1998. The subsistence harvest of seals and sea lions by Alaska Natives in the Norton Sound‐Bering Strait region, Alaska, 1996‐97. Alaska Department of Fish and Game, Division of Subsistence, Technical Paper No. 242. 88 p.
Geraci, J. R., and T. G. Smith. 1976a. Behavior and pathophysiology of seals exposed to crude oil. Pages 447‐462 in Proceedings of the Symposium American University: Sources, Effects, and Sinks of Hydrocarbons in the Aquatic Environment, Washinton, D.C. The American Institute of Biological Sciences.
Geraci, J. R., and T. G. Smith. 1976b. Direct and indirect effects of oil on ringed seals (Phoca hispida) of the Beaufort Sea. Journal of the Fisheries Research Board of Canada 33:1976‐1984.
Gerdes, R., and C. Köberle. 2007. Comparison of Arctic sea ice thickness variability in IPCC Climate of the 20th Century experiments and in ocean ‐‐ sea ice hindcasts. Journal of Geophysical Research 112:C04S13.
GHEXIS. 2005. Exploration history. Greenland Hydrocarbon Exploration Information Service (GHEXIS). Geological Survey of Denmark and Greenland (GEUS), Copenhagen, Denmark. Accessed May 2010 at http://www.geus.dk/ghexis/expl‐his.htm.
Gjertz, I., K. M. Kovacs, C. Lydersen, and Ø. Wiig. 2000. Movements and diving of adult ringed seals (Phoca hispida) in Svalbard. Polar Biology 23:651‐656.
Gjertz, I., and C. Lydersen. 1983. Ringed seal Phoca hispida pupping in the Svalbard area. Fauna 36:65‐66.
Gjertz, I., and C. Lydersen. 1986. The ringed seal (Phoca hispida) spring diet in northwestern Spitsbergen, Svalbard. Polar Research 4:53‐56.
Gjøsæter, H. 1998. The population biology and exploitation of capelin (Mallotus villosus) in the Barents Sea. Sarsia 83:453‐496.
Gleckler, P. J., K. E. Taylor, and C. Doutriaux. 2008. Performance metrics for climate models. Journal of Geophysical Research 113:D06104.
Godfroid, J. 2002. Brucellosis in wildlife. Revue Scientifique Et Technique De L Office International Des Epizooties 21:277‐286.
Goldstein, T., J. A. K. Mazet, V. A. Gill, A. M. Doroff, K. A. Burek, and J. A. Hammond. 2009. Phocine distemper virus in northern sea otters in the Pacific Ocean, Alaska, USA. Emerging Infectious Diseases 15:925‐927.
Gordon, J., D. Gillespie, J. Potter, A. Frantzis, M. P. Simmonds, R. Swift, and D. Thompson. 2003. A review of the effects of seismic surveys on marine mammals. Marine Technology Society Journal 37:16‐34.
Gosselin, J.‐F., L. N. Measures, and J. Huot. 1998. Lungworm (Nematoda: Metastrongyloidea) infections in Canadian phocids. Canadian Journal of Fisheries and Aquatic Sciences 55:825‐834.

212
Gosselin, M., M. Lavasseur, P. A. Wheeler, R. A. Horner, and B. C. Booth. 1997. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep‐Sea Research II 44:1623‐1644.
Grachev, A. I. 2006. Analysis of present‐day harvest of true seals in the Sea of Okhotsk and its prospects. Pages 155‐159 in V. M. Belkovich, editor. Marine Mammals of the Holarctic. 2006. Collection of Scientific Papers, St. Petersburg, Russia. Marine Mammal Council.
Grachev, M. A., V. P. Kumarev, L. V. Mamaev, V. L. Zorin, L. V. Baranova, N. N. Denikina, S. I. Belikov, E. A. Petrov, V. S. Kolesnik, R. S. Kolesnik, V. M. Dorofeev, A. M. Beim, V. N. Kudelin, F. G. Nagieva, and V. N. Sidorov. 1989. Distemper virus in Baikal seals. Nature 338:209‐209.
Gradinger, R. R., and B. A. Bluhm. 2004. In‐situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the High Arctic Canada Basin. Polar Biology 27:595‐603.
Grebmeier, J. M., L. W. Cooper, H. M. Feder, and B. I. Sirenko. 2006a. Ecosystem dynamics of the Pacific‐influenced Northern Bering and Chukchi Seas in the Amerasian Arctic. Progress in Oceanography 71:331‐361.
Grebmeier, J. M., J. E. Overland, S. E. Moore, E. V. Farley, E. C. Carmack, L. W. Cooper, K. E. Frey, J. H. Helle, F. A. McLaughlin, and S. L. McNutt. 2006b. A major ecosystem shift in the northern Bering Sea. Science 311:1461‐1464.
Greenland Home Rule. 2009. Management and utilization of seals in Greenland. Department of Fisheries, Hunting and Agriculture. 24 p.
Gregory, M., and D. G. Cyr. 2003. Effects of environmental contaminants on the endocrine system of marine mammals. Pages 67‐81 in J. G. Vos, G. D. Bossart, M. Fournier, and T. J. O'Shea, editors. Toxicology of Marine Mammals. Taylor and Francis, London, UK.
Grigorescu, D. 1976. Paratethyan seals. Systematic Zoology 25:407‐419. Gulland, F. M. D., M. Koski, L. J. Lowenstine, A. Colagross, L. Morgan, and T. Spraker. 1996. Leptospirosis
in California sea lions (Zalophus californianus) stranded along the the central California coast, 1981‐1994. Journal of Wildlife Diseases 32:572‐580.
Gulland, F. M. D., L. J. Lowenstine, J. M. Lapointe, T. Spraker, and D. P. King. 1997. Herpesvirus infection in stranded Pacific harbor seals of coastal California. Journal of Wildlife Diseases 33:450‐458.
Gulliksen, B., and O. J. Lonne. 1989. Distribution, abundance, and ecological importance of marine sympagic fauna in the Arctic. Rapports et Proces‐verbaux des Réunions. Conseil International pour l'Éxploration de la Mer 188:133‐138.
Guppy, M., and P. Withers. 1999. Metabolic depression in animals: physiological perspectives and biochemical generalizations. Biological Review 74:1‐40.
Haahtela, I. 1990. What do Baltic studies tell us about the isopod Saduria entomon (L.)? Annales Zoologici Fennici 27:269‐278.
Hall, C. F. 1865. Arctic researchers, and life among the Esquimaux: being the narrative of an expedition in search of Sir John Franklin, in the years 1860, 1861, and 1862. Harper and Brothers, New York. 595 p.
Hammill, M. O. 1987. Ecology of the ringed seal (Phoca hispida Schreber) in the fast‐ice of Barrow Strait, Northwest Territories. Ph.D. Dissertation. Macdonald College of McGill University, Montreal, Quebec, Canada. 108 p.
Hammill, M. O., C. Lydersen, M. Ryg, and T. G. Smith. 1991. Lactation in the ringed seal (Phoca hispida). Canadian Journal of Fisheries and Aquatic Sciences 48:2471‐2476.
Hammill, M. O., and T. G. Smith. 1989. Factors affecting the distribution and abundance of ringed seal structures in Barrow Strait, Northwest Territories. Canadian Journal of Zoology 67:2212‐2219.
Hammill, M. O., and T. G. Smith. 1991. The role of predation in the ecology of the ringed seal in Barrow Strait, Northwest Territories, Canada. Marine Mammal Science 7:123‐135.

213
Hanssen‐Bauer, I. 2002. Temperature and precipitation in Svalbard 1912 – 2050: measurement and scenarios. Polar Record 38:225‐232.
Harder, T. C., M. Harder, H. Vos, K. Kulonen, S. Kennedy‐Stoskopf, B. Liess, M. J. G. Appel, and A. D. M. E. Osterhaus. 1996. Characterization of phocid herpesvirus‐1 and ‐2 as putative alpha‐ and gammaherpesviruses of North American and European pinnipeds. Journal of General Virology 77:27‐35.
Harding, K. C., M. Fujiwara, Y. Axberg, and T. Härkönen. 2005. Mass‐dependent energetics and survival in harbour seal pups. Functional Ecology 19:129‐135.
Harding, K. C., and T. J. Härkönen. 1999. Development in the Baltic grey seal (Halichoerus grypus) and ringed seal (Phoca hispida) populations during the 20th century. Ambio 28:619‐627.
Hardy, M. H., E. Roff, T. G. Smith, and M. Ryg. 1991. Facial skin glands of ringed and grey seals, and their possible function as odoriferous organs. Canadian Journal of Zoology 69:189‐200.
Hare, S. R., and N. J. Mantua. 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Progress in Oceanography 47:103‐145.
Harington, C. R., and D. E. Sergeant. 1972. Pleistocene ringed seal skeleton from Champlain Sea deposits near Hull, Quebec ‐‐ a reidentification. Canadian Journal of Earth Sciences 9:1039‐1051.
Härkönen, T., R. Dietz, P. Reijnders, J. Teilmann, K. Harding, A. Hall, S. Brasseur, U. Siebert, S. J. Goodman, P. D. Jepson, T. D. Rasmussen, and P. Thompson. 2006. A review of the 1988 and 2002 phocine distemper virus epidemics in European harbour seals. Diseases of Aquatic Organisms 68:115‐130.
Harkonen, T., M. Jüssi, I. Jüssi, M. Verevkin, L. Dmitrieva, E. Helle, R. Sagitov, and K. C. Harding. 2008. Seasonal activity budget of adult Baltic ringed seals. PLoS ONE 3:e2006.
Härkönen, T., and S. G. Lunneryd. 1992. Estimating abundance of ringed seals in the Bothnian Bay. Ambio 21:497‐503.
Härkönen, T., O. Stenman, M. Jüssi, I. Jüssi, R. Sagitov, and M. Verevkin. 1998. Population size and distribution of the Baltic ringed seal (Phoca hispida botnica). Pages 167‐180 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Harris, R. E., G. W. Miller, and W. J. Richardson. 2001. Seal responses to airgun sounds during summer seismic surveys in the Alaskan Beaufort Sea. Marine Mammal Science 17:795‐812.
Harris, R. K., R. B. Moeller, T. P. Lipscomb, J. M. Pletcher, R. J. Haebler, P. A. Tuomi, C. R. McCormick, A. R. Degange, D. Mulcahy, and T. D. Williams. 1990. Identification of a herpes‐like virus in sea otters during rehabilitation after the T/V Exxon Valez oil spill. Pages 366‐368 in K. Bayha and J. Kennedy, editors. Sea otter symposium: Proceedings of a symposium to evaluate the response effort on behalf of sea otters after the T/V Exxon Valdez oil spill into Prince William Sound, Anchorage, Alaska, 17‐19 April 1990. U.S. Fish and Wildlife Service, Anchorage, AK.
Harwood, L., T. G. Smith, and H. Melling. 2007. Assessing the potential effects of near shore hydrocarbon exploration on ringed seals in the Beaufort Sea region 2003‐2006. Environmental Research Studies Funds Report No. 162. 103 p.
Harwood, L. A., and T. G. Smith. 2003. Movements and diving of ringed seals in the Beaufort and Chukchi Seas, 1999‐2003 (Poster presentation). Abstracts of the 15th Biennial Conference on the Biology of Marine Mammals, 14‐19 December 2003, Greensboro, NC.
Harwood, L. A., T. G. Smith, and H. Melling. 2000. Variation in reproduction and body condition of the ringed seal (Phoca hispida) in Western Prince Albert Sound, NT, Canada, as assessed through a harvest‐based sampling program. Arctic 53:422‐431.
Harwood, L. A., and I. Stirling. 1992. Distribution of ringed seals in the southeastern Beaufort Sea during late summer. Canadian Journal of Zoology 70:891‐900.

214
Hawkins, E., and R. Sutton. 2009. The potential to narrow uncertainty in regional climate predictions. Bulletin of the American Meteorological Society 90:1095‐1107.
Hay, D. E., R. Toresen, R. Stephenson, M. Thompson, F. Funk, E. Ivshina, J. Jakobsson, T. Kobayashi, I. McQuinn, G. Melvin, J. Molloy, N. Naumenko, K. T. Oda, R. Parmanne, M. Power, V. Radchenko, J. Schweigert, J. Simmonds, K. Stevenson, R. Tanasichuk, Q. Tang, D. L. Watters, and J. Wheeler. 2001. Taking stock: an inventory and review of world herring stocks in 2000. Pages 381‐455 in F. Funk, J. Blackburn, D. Hay, A. J. Paul, R. Stephenson, R. Toresen, and D. Witherell, editors. Herring: Expectations for a New Millennium. University of Alaska Sea Grant, Fairbanks, AK.
Heaton, T. H., and F. Grady. 2003. The late Wisconsin vertebrate history of Prince of Wales Island, southeast Alaska. Pages 17‐53 in B. W. Schubert, J. I. Mead, and R. W. Graham, editors. Ice Age Cave Faunas of North America. Indiana University Press, Bloomington, IN.
Heide‐Jørgensen, M. P., T. Härkönen, R. Dietz, and P. M. Thompson. 1992a. Retrospective of the 1988 European seal epizootic. Diseases of Aquatic Organisms 13:37‐62.
Heide‐Jørgensen, M. P., B. S. Stewart, and S. Leatherwood. 1992b. Satellite tracking of ringed seals Phoca hispida off Northwest Greenland. Ecography 15:56‐61.
HELCOM. 2001. Conservation and management of seal populations in the Baltic. Action plan for the implementation of the HELCOM project on seals. Ministry of Environment and Energy, Danish Forest and Nature Agency, Helsinki, Finland. 77 p. Accessed at http://meeting.helcom.fi/c/document_library/get_file?p_l_id=83489&folderId=73284&name=DLFE‐29324.pdf.
HELCOM. 2006a. Maritime transportation in the Baltic Sea. Draft HELCOM Thematic Assessment in 2006. Helsinki Commission (HELCOM), Helsinki, Finland. 24 p. Accessed at http://helcom.navigo.fi/stc/files/BSAP/FINAL%20Maritime.pdf.
HELCOM. 2006b. Minutes of the first meeting of ad hoc HELCOM seal expert group. Helsinki Commission (HELCOM), Helsinki, Finland. Accessed February 2010 at http://meeting.helcom.fi/c/document_library/get_file?p_l_id=16352&folderId=83903&name=DLFE‐33407.pdf.
HELCOM. 2007a. HELCOM Baltic Sea Action Plan. Helsinki Commission (HELCOM), HELCOM Ministerial Meeting. 101 p.
HELCOM. 2007b. HELCOM list of threatened and/or declining species of the Baltic Sea area. Pages 10‐13 in D. Boedeker and H. Von Nordheim, editors. Baltic Sea Environment Proceedings, No. 113. Helsinki Commission (HELCOM), Helsinki, Finland.
HELCOM. 2007c. Pearls of the Baltic Sea. Helsinki Commission (HELCOM), Networking for Life: Special Nature in a Special Sea. 198 p.
HELCOM. 2009. Reinforcing oil spill response capacity in the Baltic. Helsinki Commission (HELCOM). 15 p. Helle, E. 1979a. Growth and size of the ringed seal Phoca (Pusa) hispida Schreber in the Bothnian Bay,
Baltic. International Journal of Mammalian Biology 44:208‐220. Helle, E. 1979b. Structure and numbers of seal populations in the northern Baltic Sea: a study based on
Finnish bounty statistics, 1956‐1975. Aquilo Ser. Zool. 19:65‐71. Helle, E. 1980a. Age structure and sex ratio of the ringed seal Phoca (Pusa) hispida Schreber population
in the Bothnian Bay, northern Baltic Sea. Zeitschrift Fur Saugetierkunde‐International Journal of Mammalian Biology 45:310‐317.
Helle, E. 1980b. Lowered reproductive capacity in female ringed seals (Pusa hispida) in the Bothnian Bay, northern Baltic Sea, with special reference to uterine occlusions. Annales Zoologici Fennici 17:147‐158.
Helle, E. 1981. Reproductive trends and occurrence of organochlorines and heavy metals in the Baltic seal populations. International Council for the Exploration of the Sea, C.M. 1981/E:37. 13 p.

215
Helle, E. 1986. The decrease in the ringed seal population of the Gulf of Bothnia in 1975‐84. Finnish Game Research 44:28‐32.
Helle, E. 1990. The state of the ringed seal population in the Bothenian Bay in the 1980s. Suomen Riista 36:31‐36.
Helle, E. 1992. Phoca hispida Schreber, 1775 ‐‐ Ringelrobbe. Pages 138‐161 in R. Duguy and D. Robineau, editors. Handbuch der Säugetiere Europas, Band 6: Meeressäuger, Teil II: Robben ‐ Pinnipedia. AULA‐Verlag, Wiesbaden, Germany. (In German).
Helle, E., H. Hyvärinen, H. Pyysalo, and K. Wickström. 1983. Levels of organochlorine compounds in an inland seal population in eastern Finland. Marine Pollution Bulletin 14:256‐260.
Helle, E., H. Hyvärinen, and T. Sipilä. 1984. Breeding habitat and lair structure of the Saimaa ringed seal Phoca hispida saimensis Nordq. in Finland. Acta Zoologica Fennica 172:125‐127.
Helle, E., M. Olsson, and S. Jensen. 1976a. DDT and PCB levels and reproduction in ringed seal from the Bothnian Bay. Ambio 5:188‐189.
Helle, E., M. Olsson, and S. Jensen. 1976b. PCB levels correlated with pathological changes in seal uteri. Ambio 5:261‐263.
Helle, E., and O. Stenman. 1984. Recent trends in levels of PCB and DDT compounds in seals from the Finnish waters of the Baltic Sea. International Council for the Exploration of the Sea, C.M. 1984/E:43. 8 p.
Helle, E., and O. Stenman. 1990. Baltic seal populations. Maailman Luonnon Sätiŏn WWF, Suomen Rahaston Raportteja Nro 1. 76 p. (In Finnish, English Summary).
Heptner, L. V. G., K. K. Chapskii, V. A. Arsen'ev, and V. T. Sokolov. 1976. Ringed seal. Phoca (Pusa) hispida Schreber, 1775. Pages 218‐260 in L. V. G. Heptner, N. P. Naumov, and J. Mead, editors. Mammals of the Soviet Union. Volume II, Part 3‐‐Pinnipeds and Toothed Whales, Pinnipedia and Odontoceti. Vysshaya Shkola Publishers, Moscow, Russia. (Translated from Russian by P. M. Rao, 1996, Science Publishers, Inc., Lebanon, NH).
Hersteinsson, P., and D. W. MacDonald. 1992. Interspecific competition and the geographical distribution of red and Arctic foxes Vulpes Vulpes and Alopex lagopus. Oikos 64:505‐515.
Higdon, J. W., O. R. P. Bininda‐Emonds, R. M. D. Beck, and S. H. Ferguson. 2007. Phylogeny and divergence of the pinnipeds (Carnivora: Mammalia) assessed using a multigene dataset. BMC Evolutionary Biology 7:216.
Hites, R. A. 2004. Polybrominated diphenyl ethers in the environment and in people: a meta‐analysis of concentrations. Environmental Science & Technology 38:945‐956.
Holland, M. M., C. M. Bitz, and B. Tremblay. 2006. Future abrupt reductions in the summer Arctic sea ice. Geophysical Research Letters 33:L23503.
Hollowed, A. B., and W. S. Wooster. 1995. Decadal‐scale variations in the eastern subarctic Pacific: II. Response of northeast Pacific fish stocks. Pages 373‐385 in R. J. Beamish, editor. Climate Change and Northern Fish Populations. Canadian Journal of Fisheries and Aquatic Sciences 121.
Holshuh, H. J., A. E. Sherrod, C. R. Taylor, B. F. Andrews, and E. B. Howard. 1985. Toxoplasmosis in a feral northern fur seal. Journal of the American Veterinary Medical Association 187:1229‐1230.
Holst, M., I. Stirling, and W. Calvert. 1999. Age structure and reproductive rates of ringed seals (Phoca hispida) on the northwestern coast of Hudson Bay in 1991 and 1992. Marine Mammal Science 15:1357‐1364.
Holst, M., I. Stirling, and K. A. Hobson. 2001. Diet of ringed seals (Phoca hispida) on the east and west sides of the North Water Polynya, northern Baffin Bay. Marine Mammal Science 17:888‐908.
Hop, H., T. Pearson, E. N. Hegseth, K. M. Kovacs, C. Wiencke, S. Kwaśniewski, K. Eiane, F. Mehlum, B. Gulliksen, M. Wlodarska‐Kowalezuk, C. Lydersen, J. M. Węsławski, S. Cochrane, G. W. Gabrielsen, R. J. G. Leakey, O. J. Lonne, M. Zajaczkowski, S. Falk‐Petersen, M. Kendall, S. A. Wängberg, K. Bischof, A. Y. Voronkov, N. A. Kovaltchouk, J. Wiktor, M. Poltermann, G. di Prisco,

216
C. Papucci, and S. Gerland. 2002. The marine ecosystem of Kongsfjorden, Svalbard. Polar Research 21:167‐208.
Hovelsrud, G. K., M. McKenna, and H. P. Huntington. 2008. Marine mammal harvests and other interactions with humans. Ecological Applications 18:S135‐S147.
Hunt, G. L., Jr., P. Stabeno, G. Walters, E. Sinclair, R. D. Brodeur, J. M. Napp, and N. A. Bond. 2002. Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep‐Sea Research II 49:5821‐5853.
Huntington, H. P. 2009. A preliminary assessment of threats to arctic marine mammals and their conservation in the coming decades. Marine Policy 33:77‐82.
Hyvärinen, H. 1989. Diving in darkness: whiskers as sense‐organs of the Ringed Seal (Phoca hispida saimensis). Journal of Zoology 218:663‐678.
Hyvärinen, H., E. Hämäläinen, and M. Kunnasranta. 1995. Diving behavior of the Saimaa ringed seal (Phoca hispida saimensis Nordq.). Marine Mammal Science 11:324‐334.
Hyvärinen, H., and M. Nieminen. 1990. Differentiation of the ringed seal in the Baltic Sea, Lake Ladoga and Lake Saimaa. Finnish Game Research 47:21‐27.
Hyvärinen, H., and T. Sipilä. 1984. Heavy metals and high pup mortality in the Saimaa ringed seal population in eastern Finland. Marine Pollution Bulletin 15:335‐337.
Hyvärinen, H., T. Sipilä, M. Kunnasranta, and J. T. Koskela. 1998. Mercury pollution and the Saimaa ringed seal (Phoca hispida saimensis). Marine Pollution Bulletin 36:76‐81.
Iacozza, J., and D. G. Barber. 1999. An examination of the distribution of snow on sea‐ice. Atmosphere‐Ocean 37:21‐51.
Ianelli, J. 2005. Assessment and fisheries management of eastern Bering Sea walleye pollock: is sustainability luck? Bulletin of Marine Science 76:321‐335.
Ianelli, J. N., S. Barbeaux, T. Honkalehto, S. Kotwicki, K. Aydin, and N. Williamson. 2009. Assessment of walleye pollock stock in the Eastern Bering Sea. Pages 49‐148 in Stock Assessment and Fishery Evaluation Report for the Groundfish Resources of the Bering Sea/Aleutian Islands Regions. North Pacific Fishery Management Council, Seattle, WA.
ICES. 2003. Report of the working group on marine mammal ecology (WGMME), 25‐29 March 2003, Hel, Poland. International Council for the Exploration of the Sea, C.M. 2003/ACE:03. 75 p.
ICES. 2008. Report of the working group on marine mammal ecology (WGMME), 25‐29 February 2008, St. Andrews, UK. International Council for the Exploration of the Sea, C.M. 2008/ACOM:44. 83 p.
Iglesias‐Rodriguez, M. D., P. R. Halloran, R. E. M. Rickaby, I. R. Hall, E. Colmenero‐Hidalgo, J. R. Gittins, D. R. H. Green, T. Tyrrell, S. J. Gibbs, P. von Dassow, E. Rehm, E. V. Armbrust, and K. P. Boessenkool. 2008. Phytoplankton calcification in a high‐CO2 world. Science 320:336‐340.
Inuit Tapiriit Kanatami. 2010. Inuit sue European Union (EU) to overturn seal product import ban, defending Inuit rights and upholding the rule of law. Inuit Tapiriit Kanatami, Ottowa, Canada. Accessed February 2010 at http://www.itk.ca/media‐centre/media‐releases/inuit‐sue‐european‐union‐eu‐overturn‐seal‐product‐import‐ban‐defending‐i.
IPCC. 1996. Climate change 1995: Impacts, adaptations and mitigation of climate change: Scientific‐technical analyses. Contribution of Working Group II to the Second Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. 878 p.
IPCC. 2000. Summary for policymakers. Pages 1‐20 in Emissions Scenarios. A Special Report of the Intergovernmental Panel on Climate Change Working Group III. IPCC, Geneva, Switzerland.
IPCC. 2007a. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland. 104 p.
IPCC. 2007b. Summary for policymakers. Pages 1‐22 in Core Writing Team, R. K. Pachauri, and A. Reisinger, editors. Climate Change 2007: Synthesis Report. IPCC, Geneva, Switzerland.

217
IPCC. 2007c. Summary for policymakers. Pages 1‐18 in S. Solomon, D. Qin, M. Manning, M. Marquis, K. Averyt, M. M. B. Tignor, H. L. Miller, Jr., and Z. Chen, editors. Climate Change 2007: The Physical Science Basis. Cambridge University Press, Cambridge, UK and New York, NY.
IPCC. 2008. About IPCC (Website). Intergovernmental Panel on Climate Change, Geneva, Switzerland. Accessed August 22, 2008 at http://www.ipcc.ch/about/index.htm.
Ishimatsu, A., M. Hayashi, K. S. Lee, T. Kikkawa, and J. Kita. 2005. Physiological effects on fishes in a high‐CO2 world. Journal of Geophysical Research 110:C09209.
IUCN Species Survival Commission. 2009. Species and climate change: more than just the polar bear. International Union for Conservation of Nature and Natural Resources (IUCN). 46 p.
Ivanov, A. Y., and V. V. Zatyagalova. 2008. A GIS approach to mapping oil spills in a marine environment. International Journal of Remote Sensing 29:6297‐6313.
Ivanter, E. V., N. V. Medvedev, and N. A. Panichev. 1998. Heavy metal content in the Ladoga ringed seal, Phoca hispida ladogiensis. Russian Journal of Ecology 29:98‐102.
Iwata, H., S. Tanabe, T. Mizuno, and R. Tatsukawa. 1997. Bioaccumulation of butyltin compounds in marine mammals: the specific tissue distribution and composition. Applied Organometallic Chemistry 11:257‐264.
Jääskeläinen, V. 1942. Laatokka kalavetenä [Lake Ladoga and fishing]. Pages 38‐45 in V. Näsi and E. Ovaska, editors. Laatokan mainingit, Laatokan ja sen rannikon elämää sanoin ja kuvin. Otava, Helsinki, Finland. (In Finnish).
Jansen, J. K., P. L. Boveng, S. P. Dahle, and J. L. Bengtson. 2010. Reaction of harbor seals to cruise ships. Journal of Wildlife Management 74:1186‐1194.
Jansson, B.‐O. 2003. The Baltic Sea. Pages 145‐170 in G. Hempel and K. Sherman, editors. Large Marine Ecosystems of the World: Trends in Exploitation, Protection and Research. Elsevier, Amsterdam.
Jensen, S. K., J. Aars, C. Lydersen, K. M. Kovacs, and K. Asbakk. 2010. The prevalence of Toxoplasma gondii in polar bears and their marine mammal prey: evidence for a marine transmission pathway? Polar Biology 33:599‐606.
Jenssen, B. M. 1996. An overview of exposure to, and effects of, petroleum oil and organochlorine pollution in grey seals (Halichoerus grypus). Science of the Total Environment 186:109‐118.
Jenssen, B. M., J. U. Skaare, M. Ekker, D. Vongraven, and S. H. Lorentsen. 1996. Organochlorine compounds in blubber, liver and brain in neonatal grey seal pups. Chemosphere 32:2115‐2125.
Jødestøl, K., and K. I. Ugland. 1994. Sårbarhetsanalyse for ringsel og grønlandsel i Barentshavet nord. Det Norske Veritas. Technical Report No. 93‐3740. 59 p. (In Norwegian, English Summary).
Johansen, P., F. O. Kapel, and I. Kraul. 1980. Heavy metals and organochlorines in marine mammals from Greenland. International Council for the Exploration of the Sea, C.M. 1980/E:32. 12 p.
Johnson, L. 1983. Assessment of the effects of oil on Arctic marine fish and marine mammals. Canadian Technical Report of Fisheries and Aquatic Sciences No. 1200. 15 p.
Johnson, M. L., C. H. Fiscus, B. T. Ostenson, and M. L. Barbour. 1966. Marine mammals. Pages 877‐924 in N. J. Wilimovsky and J. N. Wolfe, editors. Environment of the Cape Thompson Region, Alaska. U.S. Atomic Energy Commission, Oak Ridge, TN.
Johnson, S. R., and D. R. Herter. 1989. The birds of the Beaufort Sea. British Petroleum Exploration Inc., Anchorage, AK. 372 p.
Johnson, S. R., and L. E. Noel. 2005. Temperature and predation effects on abundance and distribution of lesser snow geese in the Sagavanirktok River Delta, Alaska. Waterbirds 28:292‐300.
Joly, S., S. Senneville, D. Caya, and F. Saucier. 2010. Sensitivity of Hudson Bay sea ice and ocean climate to atmospheric temperature forcing. Climate Dynamics. DOI: 10.1007/s00382‐009‐0731‐4. (Published online January 21, 2010).
Jonsson, P., R. Carman, and F. Wulff. 1990. Laminated sediments in the Baltic –‐ a tool for evaluating mass balances. Ambio 19:152‐158.

218
Jun, M., R. Knutti, and D. W. Nychka. 2008. Spatial analysis to quantify numerical model bias and dependence: how many climate models are there? Journal of the American Statistical Association 103:934‐947.
Jüssi, I., E. Helle, O. Stenman, T. Blomberg, T. Sipilä, H. Benke, W. Dinter, K. Harder, U. Siebert, V. Pilats, A. Grudas, P. Kulikov, I. Kuklik, K. E. Skóra, V. I. Chernook, V. V. Melentiev, R. Sagitov, N. Medvedev, M. V. Verevkin, B.‐M. Bäcklin, B. Helander, O. Karlsson, S. G. Lunneryd, and I. Sundvisson. 2005. Management of the seal populations in the Baltic area. Pages 80‐84 in E. Helle, O. Stenman, and M. Wikman, editors. Symposium on Biology and Management of Seals in the Baltic Area, Helsinki, Finland. Finnish Game and Fisheries Research Institute.
Jylhä, K., S. Fronzek, H. Tuomenvirta, T. Carter, and K. Ruosteenoja. 2008. Changes in frost, snow and Baltic sea ice by the end of the twenty‐first century based on climate model projections for Europe. Climatic Change 86:441‐462.
Käkelä, R., and H. Hyvärinen. 1993. Fatty acid composition of fats around the mystacial and superciliary vibrissae differs from that of blubber in the Saimaa ringed seal (Phoca hispida saimensis). Comparative Biochemistry and Physiology B 105B:547‐552.
Kålås, J. A., A. Viken, and T. Bakken, editors. 2006. 2006 Norwegian red list. ArtsDatabanken, Trondheim, Norway. 416 p.
Kannan, K., S. Corsolini, J. Falandysz, G. Oehme, S. Focardi, and J. P. Giesy. 2002. Perfluorooctanesulfonate and related fluorinated hydrocarbons in marine mammals, fishes, and birds from coasts of the Baltic and the Mediterranean Seas. Environmental Science & Technology 36:3210‐3216.
Kannan, K., N. Kajiwara, S. Tanabe, and J. P. Giesy. 2001a. Organochlorines and butyltins in blubber and livers of stranded California Sea lions, elephant seals and harbor seals from Coastal California, USA. (Poster Presentation). Page PM157 in Society for Environmental Toxicology and Chemistry (SETAC) 22nd Annual Meeting. Changing Evironmental Awareness: Societal Concerns and Scientific Responses, Baltimore, MD.
Kannan, K., J. Koistinen, K. Beckmen, T. Evans, J. F. Gorzelany, K. J. Hansen, P. D. Jones, E. Helle, M. Nyman, and J. P. Giesy. 2001b. Accumulation of perfluorooctane sulfonate in marine mammals. Environmental Science & Technology 35:1593‐1598.
Kapel, F. O., J. Christiansen, M. P. Heide‐Jørgensen, T. Härkønen, E. W. Born, L. O. Knutsen, F. Rigét, and J. Teilmann. 1998. Netting and conventional tagging used to study movements of ringed seals (Phoca hispida) in Greenland. Pages 211‐228 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Kapel, O. F., and A. Rosing‐Asvid. 1996. Seal hunting statistics for Greenland 1993 and 1994, according to a new system of collecting information, compared to previous lists‐of‐game. Northwest Atlantic Fisheries Organization (NAFO) Scientific Council Studies 26:71‐86.
Karetnikov, S. G., and M. A. Naumenko. 2008. Recent trends in Lake Ladoga ice cover. Hydrobiologia 599:41‐48.
Karlsson, O., T. Härkönen, and B. M. Bäcklin. 2008. Growing populations. Pages 91‐92 in K. Viklund, editor. Havet 2008. Environmental Conditions in Swedish Coastal Waters. Swedish Environmental Protection Agency, Vallhallavägen, Stockholm. (In Swedish).
Kelly, B. 1997. Behavior of ringed seals diving under shore‐fast sea ice. Page 1073 in Dissertation Abstracts International. ProQuest, Ann Arbor, MI.
Kelly, B. P. 1981. Pelage polymorphism in Pacific harbor seals. Canadian Journal of Zoology 59:1212‐1219.
Kelly, B. P. 1988. Ringed seal, Phoca hispida. Pages 57‐75 in J. W. Lentifer, editor. Selected Marine Mammal Species of Alaska: Species Accounts with Research and Managment Recommendations. Marine Mammal Commission, Washington, D.C.

219
Kelly, B. P. 1996. Live capture of ringed seals in ice‐covered waters. Journal of Wildlife Management 60:678‐684.
Kelly, B. P. 2001. Climate change and ice breeding pinnipeds. Pages 43‐55 in G.‐R. Walther, C. A. Burga, and P. J. Edwards, editors. "Fingerprints" of Climate Change ‐‐ Adapted Behavior and Shifting Species Ranges. Kluwer Academic / Plenum Publishers, New York, NY.
Kelly, B. P., O. H. Badajos, M. Kunnasranta, and J. Moran. 2006. Timing and re‐interpretation of ringed seal surveys. Coastal Marine Institute University of Alaska Fairbanks, Final Report. 60 p.
Kelly, B. P., O. H. Badajos, M. Kunnasranta, J. R. Moran, M. Martinez‐Bakker, D. Wartzok, and P. Boveng. 2010. Seasonal home ranges and fidelity to breeding sites among ringed seals. Polar Biology 33:1095‐1109.
Kelly, B. P., J. J. Burns, and L. T. Quakenbush. 1988. Responses of ringed seals (Phoca hispida) to noise disturbance. Pages 27‐38 in W. M. Sackinger, M. O. Jeffries, J. L. Imm, and S. D. Treacy, editors. Port and Ocean Engineering Under Arctic Conditions, Volume II, Symposium on Noise and Marine Mammals, Fairbanks, Alaska.
Kelly, B. P., O. R. Harding, and M. Kunnasranta. 2003. Early onset of snow melts and increased vulnerability of an ice‐inhabiting seal. Page 206 in Extreme Events: Understanding Perturbations to the Physical and Biological Environment, Fairbanks, AK. American Association for the Advancement of Science (AAAS), Arctic Division.
Kelly, B. P., M. Ponce, D. A. Tallmon, B. J. Swanson, and S. K. Sell. 2009. Genetic diversity of ringed seals sampled at breeding sites; implications for population structure and sensitivity to sea ice loss. Univeristy of Alaska Southeast, North Pacific Research Board 631 Final Report. 28 p.
Kelly, B. P., and L. T. Quakenbush. 1990. Spatiotemporal use of lairs by ringed seals (Phoca hispida). Canadian Journal of Zoology 68:2503‐2512.
Kelly, B. P., L. T. Quakenbush, and J. R. Rose. 1986. Ringed seal winter ecology and effects of noise disturbance. Pages 447‐536 in Outer Continental Shelf Environmental Assessment Program. Final Reports of Principal Investigators, Volume 61. Minerals Management Service, Alaska Outer Continental Shelf Office, Anchorage, AK.
Kelly, B. P., and D. Wartzok. 1996. Ringed seal diving behavior in the breeding season. Canadian Journal of Zoology 74:1547‐1555.
Kennedy‐Stoskopf, S. 2001. Viral diseases. Pages 285‐308 in L. A. Dierauf and F. Gulland, editors. CRC Handbook of Marine Mammal Medicine. CRC Press LLC, Boca Raton, FL.
Kennedy‐Stoskopf, S., M. K. Stoskopf, M. A. Eckhaus, and J. D. Strandberg. 1986. Isolation of a retrovirus and a herpesvirus from a captive California sea lion. Journal of Wildlife Diseases 22:156‐164.
Kennedy, M. J. 1986. Filaroides (Parafilaroides) hispidus n.sp. (Nematoda: Metastrongyloidea) from the lungs of the ringed seal, Phoca hispida (Phocidae), from the Beaufort Sea, Canada. Canadian Journal of Zoology 64:1864‐1868.
Kennedy, S. 1998. Morbillivirus infections in aquatic mammals. Journal of Comparative Pathology 119:201‐225.
Kennedy, S., T. Kuiken, P. D. Jepson, R. Deaville, M. Forsyth, T. Barrett, M. W. G. van de Bildt, A. D. M. E. Osterhaus, T. Eybatov, C. Duck, A. Kydyrmanov, I. Mitrofanov, and S. Wilson. 2000. Mass die‐off of Caspian seals caused by canine distemper virus. Emerging Infectious Diseases 6:637‐639.
Kennedy, S., J. A. Smyth, S. J. McCullough, G. M. Allan, F. McNeilly, and S. McQuaid. 1988. Confirmation of cause of recent seal deaths. Nature 335:404.
Khon, V. C., I. I. Mokhov, M. Latif, V. A. Semenov, and W. Park. 2010. Perspectives of Northern Sea Route and Northwest Passage in the twenty‐first century. Climatic Change 100:757‐768.
Kik, M. J. L., M. G. Goris, J. H. Bos, R. A. Hartskeerl, and G. M. Dorrestein. 2006. An outbreak of leptospirosis in seals (Phoca vitulina) in captivity. Veterinary Quarterly 28:33‐39.

220
Kilkki, V., and K. Marttinen. 1984. Henkipatosta suojelukohteeksi‐ saimaannorpan ja ihmisen historiaa. [Saimaa seal and man, a historical review]. Pages 98‐102 in P. Becker, editor. Saimaanorppa. Suomen Luonnonsuojelun Tuki Oy, Helsinki, Finland. (In Finnish).
King, J. E. 1964. Seals of the world. 1st edition. British Museum (Natural History) London, UK. 154 p. King, J. E. 1983. Seals of the world. 2nd edition. British Museum (Natural History) and Oxford University
Press, London, UK. 240 p. Kingsley, M. C. S. 1998. The numbers of ringed seals (Phoca hispida) in Baffin Bay and associated waters.
Pages 181‐196 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Kingsley, M. C. S., and N. J. Lunn. 1983. The abundance of seals in the eastern Beaufort Sea, northern Amundsen Gulf and Prince Albert Sound, 1982. Canadian Wildlife Service, Prepared for Dome Petroleum Limited, Gulf Canada Resources, Inc. 16 p.
Kingsley, M. C. S., and I. Stirling. 1991. Haul‐out behavior of ringed and bearded seals in relation to defense against surface predators. Canadian Journal of Zoology 69:1857‐1861.
Knutti, R., M. R. Allen, P. Friedlingstein, J. M. Gregory, G. C. Hegerl, G. A. Meehl, M. Meinshausen, J. M. Murphy, G.‐K. Plattner, S. C. B. Raper, T. F. Stocker, P. A. Stott, H. Teng, and T. M. L. Wigley. 2008. A review of uncertainties in global temperature projections over the twenty‐first century. Journal of Climate 21:2651‐2663.
Knutti, R., G. A. Meehl, M. R. Allen, and D. A. Stainforth. 2006. Constraining climate sensitivity from the seasonal cycle in surface temperature. Journal of Climate 19:4224‐4233.
Koeman, J. H., W. S. M. van de Ven, J. J. M. Goeij, P. S. Tjioe, and J. L. van Haaften. 1975. Mercury and selenium in marine mammals and birds. Science of the Total Environment 3:279‐287.
Kokko, H., E. Helle, J. Lindström, E. Ranta, T. Sipilä, and F. Courchamp. 1999. Backcasting population sizes of ringed and grey seals in the Baltic and Lake Saimaa during the 20th century. Annales Zoologici Fennici 36:65‐73.
Kokko, H., J. Lindström, E. Ranta, T. Sipilä, and J. Koskela. 1998. Estimating the demographic effective population size of the Saimaa ringed seal (Phoca hispida saimensis Nordq.). Animal Conservation 1:47‐54.
Kommersant. 2004. Magadan Region (Online news article). Kommersant Publishing House, Moscow, Russia. Accessed May 2010 at http://www.kommersant.com/p‐52/r_396/Magadan_Region/.
Kooyman, G. L., R. W. Davis, and M. A. Castellini. 1977. Thermal conductance of immersed pinniped and sea otter pelts before and after oiling with Prudhoe Bay crude. Pages 151‐157 in D. A. Wolfe, editor. Fate and effects of petroleum hydrocarbons in marine ecosystems and organisms. Pergamon Press, New York, NY.
Koskela, J. T., M. Kunnasranta, E. Hämäläinen, and H. Hyvärinen. 2002. Movements and use of haul‐out sites of radio‐tagged Saimaa ringed seal (Phoca hispida saimensis Nordq.) during the open‐water season. Annales Zoologici Fennici 39:59‐67.
Kostamo, A., N. Medvedev, J. Pellinen, H. Hyvärinen, and J. V. K. Kukkonen. 2000a. Analysis of organochlorine compounds and extractable organic halogen in three subspecies of ringed seal from Northeast Europe. Environmental Toxicology and Chemistry 19:848‐854.
Kostamo, A., M. Viljanen, J. Pellinen, and J. Kukkonen. 2000b. EOX and organochlorine compounds in fish and ringed seal samples from Lake Ladoga, Russia. Chemosphere 41:1733‐1740.
Kostareva, G. V., A. A. Buchurin, and N. V. Sokolova. 1970. Strontium‐90, strontium and calcium in some aquatic organisms from the Sea of Okhotsk. Radiobiologiia 10:145‐147. (In Russian).
Kotiranta, E. 2008. Shipping of oil products in the Baltic Region. Pages 15‐16 in K. Liuhto, editor. Baltic Rim Economies – Bimonthly Review. Pan‐European Institute, Turku School of Economics, Turku, Finland.

221
Kovacs, K., L. Lowry, and T. Härkönen. 2008. Pusa hispida. IUCN Red List of Threatened Species. Version 2010.1. International Union for Conservation of Nature and Natural Resources (IUCN), Cambridge, UK. Accessed April 2010 at http://www.iucnredlist.org/apps/redlist/details/41672/0.
Kovacs, K. M. 2007. Background document for development of a circumpolar ringed seal (Phoca hispida) monitoring plan. Marine Mammal Commission, Workshop to Develop Monitoring Plans for Arctic Marine Mammals. 45 p.
Kovacs, K. M., and C. Lydersen. 2008. Climate change impacts on seals and whales in the North Atlantic Arctic and adjacent shelf seas. Science Progress 92:117‐150.
Kozlov, D. P. 1966. Trikhinellez u morzhei v Sovetskoi Arktike (Walrus trichinellosis in Soviet Arctic). Pages 131‐133 in Materialy k Nauchnym Konferentsiiam Vsesoiuznogo Obshchestva Gel'mintologov
Krafft, B. A., K. M. Kovacs, M. Andersen, J. Aars, C. Lydersen, T. Ergon, and T. Haug. 2006a. Abundance of ringed seals (Pusa hispida) in the fjords of Spitsbergen, Svalbard, during the peak molting period. Marine Mammal Science 22:394‐412.
Krafft, B. A., K. M. Kovacs, A. K. Frie, T. Haug, and C. Lydersen. 2006b. Growth and population parameters of ringed seals (Pusa hispida) from Svalbard, Norway, 2002‐2004. ICES Journal of Marine Science 63:1136‐1144.
Krafft, B. A., K. M. Kovacs, and C. Lydersen. 2007. Distribution of sex and age groups of ringed seals Pusa hispida in the fast‐ice breeding habitat of Kongsfjorden, Svalbard. Marine Ecology Progress Series 335:199‐206.
Krupnik, I. 1993. The evolution of maritime hunting. Pages 185‐215 in M. Levenson, editor. Arctic Adaptations, Native Whalers and Reindeer Herders of Northern Eurasia. University Press of New England, Hanover, MD and London, England. (Translated from Russian by M. Levenson, 355 p.).
Krupnik, I. I. 1988. Asiatic Eskimos and marine resources: a case of ecological pulsations or equilibrium? Arctic Anthropology 25:94‐106.
Krylov, V. I., G. A. Fedoseev, and A. P. Shustov. 1964. Pinnipeds of the Far East. Pischevaya Promyshlennost (Food Industry), Moscow, Russia. 59 p. (Translated from Russian by F. H. Fay and B. A. Fay, University of Alaska, Fairbanks, AK, 47 p.).
Kucklick, J. R., W. D. J. Struntz, P. R. Becker, G. W. York, T. M. O'Hara, and J. E. Bohonowych. 2002. Persistent organochlorine pollutants in ringed seals and polar bears collected from northern Alaska. Science of the Total Environment 287:45‐59.
Kumlien, L. 1879. Mammals. Pages 55‐61 in Contributions to the Natural History of Arctic America made in connection with the Howgate Polar Expedition 1877‐78. Government Printing Office, Washington, D.C.
Kunnasranta, M. 2001. Behavioural biology of two ringed seal (Phoca hispida) subspecies in the large European lakes Saimaa and Ladoga. University of Joensuu, Joensuu, Finland. 86 p.
Kunnasranta, M., H. Hyvärinen, J. Häkkinen, and J. T. Koskela. 2002. Dive types and circadian behaviour patterns of Saimaa ringed seals Phoca hispida saimensis during the open‐water season. Acta Theriologica 47:63‐72.
Kunnasranta, M., H. Hyvärinen, T. Sipilä, and J. T. Koskela. 1999. The diet of the Saimaa ringed seal Phoca hispida saimensis. Acta Theriologica 44:443‐450.
Kunnasranta, M., H. Hyvärinen, T. Sipilä, and N. Medvedev. 2001. Breeding habitat and lair structure of the ringed seal (Phoca hispida ladogensis) in northern Lake Ladoga in Russia. Polar Biology 24:171‐174.
Kurihara, H., and A. Ishimatsu. 2008. Effects of high CO2 seawater on the copepod (Acartia tsuensis) through all life stages and subsequent generations. Marine Pollution Bulletin 56:1086‐1090.

222
Kuusisto, E. 2005. Snow as a geographic element. Pages 159‐173 in M. Seppälä, editor. The Physical Geography of Fennoscandia. Oxford University Press, Oxford, UK.
Kwok, R., G. F. Cunningham, M. Wensnahan, I. Rigor, H. J. Zwally, and D. Yi. 2009. Thinning and volume loss of the Arctic Ocean sea ice cover: 2003‐2008. Journal of Geophysical Research 114:C07005.
Kwok, R., and D. A. Rothrock. 2009. Decline in Arctic sea ice thickness from submarine and ICESat records: 1958‐2008. Geophysical Research Letters 36:L15501.
Labansen, A. L., C. Lydersen, T. Haug, and K. M. Kovacs. 2007. Spring diet of ringed seals (Phoca hispida) from northwestern Spitsbergen, Norway. ICES Journal of Marine Science 64:1246‐1256.
Lagarev, S. I. 1988. Results of an aerial survey of coastal rookeries in the Sea of Okhotsk in 1986. Pages 68‐75 in N. S. Chernysheva, editor. Scientific Research on Sea Mammals of the Northern Part of the Pacific Ocean in 1986‐1987. All‐Union Scientific Research Institute of Sea Fisheries and Oceanography (VNIRO), Moscow, Russia. (Translated from Russian by Canada Institute for Scientific and Technical Information, National Research Council, Ottawa, Canada, 8 p.).
Laidre, K. L., I. Stirling, L. F. Lowry, Ø. Wiig, M. P. Heide‐Jørgensen, and S. H. Ferguson. 2008. Quantifying the sensitivity of Arctic marine mammals to climate‐induced habitat change. Ecological Applications 18:S97‐S125.
Lalli, C. M., and R. Gilmer. 1989. Pelagic snails: the biology of holoplanktonic gastropod mollusks. Stanford University Press, Stanford, CA. 276 p.
Langenbuch, M., and H. O. Pörtner. 2003. Energy budget of Antarctic fish hepatocytes (Pachycara brachycephalum and Lepidonotohen kempi) as a function of ambient CO2: pH dependent limitations of cellular protein biosynthesis? Journal of Experimental Biology 206:3895‐3903.
Lapko, V. V., and V. I. Radchenko. 2000. Sea of Okhotsk. Marine Pollution Bulletin 41:179‐187. Lapointe, J. M., P. J. Duignan, A. E. Marsh, F. M. Gulland, B. C. Barr, D. K. Naydan, D. P. King, C. A.
Farman, K. A. B. Huntingdon, and L. J. Lowenstine. 1998. Meningoencephalitis due to a Sarcocystis neurona‐like protozoan in Pacific harbor seals (Phoca vitulina richardsi). Journal of Parasitology 84:1184‐1189.
Larsen, T., and B. Kjos‐Hanssen. 1983. Trichinella sp. in polar bears from Svalbard, in relation to hide length and age. Polar Research 1:89‐96.
Larsson, U., R. Elmgren, and F. Wulff. 1985. Eutrophication and the Baltic Sea: causes and consequences. Ambio 14:9‐14.
Law, R. 2000. Fishing, selection, and phenotypic evolution. ICES Journal of Marine Science 57:659‐668. Law, R., and D. R. Grey. 1989. Evolution of yields from populations with age‐specific cropping.
Evolutionary Ecology 3:343‐359. Lawn, D., R. Steiner, and J. Wills. 1999. Sakhalin's oil: doing it right. Applying gold standards to public
participation, environmental monitoring, oil spill prevention and response and liability standards in the Sakhalin Oblast of the Russian Federation. Sakhalin Environment Watch and Pacific Environment and Resources Center. 46 p.
Learmonth, J. A., C. D. MacLeod, M. B. Santos, G. J. Pierce, H. Q. P. Crick, and R. A. Robinson. 2006. Potential effects of climate change on marine mammals. Oceanography and Marine Biology: An Annual Review 44:431‐464.
Lee, J. 2008. BP to begin developing Liberty oil field (Online news article). Anchorage Daily News, Anchorage, AK. Accessed June 2010 at http://www.adn.com/2008/07/14/464438/bp‐to‐begin‐developing‐liberty.html.
Lester, S. 2010. The WTO seal products dispute: a preview of the key legal issues. The American Society of International Law. 6 p.
Lier, M., S. Aarvik, K. Fossum, C. von Quillfeldt, O. Overrein, S. Barr, J.‐P. H. Hansen, and M. Ekker. 2010. Protected areas in Svalbard – securing internationally valuable cultural and natural heritage. Norwegian Directorate for Nature Management, Norwegian Polar Institute, Norwegian

223
Directorate for Cultural Heritage, and Governor of Svalbard, Trondheim, Norway. 19 p. Accessed at http://www.dirnat.no/attachment.ap?id=1043.
Lindsay, D. S., M. V. Collins, S. M. Mitchell, R. A. Cole, G. Flick, J., C. N. Wetch, A. Lindquist, and J. P. Dubey. 2003. Sporulation and survival of Toxoplasma gondii oocysts in seawater. Journal of Eukaryotic Microbiology 50:S687‐S688.
Lipscomb, W. H. 2001. Remapping the thickness distribution in sea ice models. Journal of Geophysical Research‐Oceans 106:13989‐14000.
Litzow, M. A., K. M. Bailey, F. G. Prahl, and R. Heintz. 2006. Climate regime shifts and reorganization of fish communities: the essential fatty acid limitation hypothesis. Marine Ecology Progress Series 315:1‐11.
Litzow, M. A., and L. Ciannelli. 2007. Oscillating trophic control induces community reorganization in a marine ecosystem. Ecology Letters 10:1124‐1134.
Loeng, H. 1989. The influence of temperature on some fish population parameters in the Barents Sea. Journal of Northwest Atlantic Fisheries Science 9:103‐113.
Logerwell, E. A., K. Rand, S. Parker‐Stetter, J. K. Horne, T. Weingartner, and B. A. Bluhm. 2010. Beaufort Sea marine fish monitoring 2008: pilot survey and test of hypotheses. Final Report. Minerals Management Service, Anchorage, AK.
Longhurst, A. R. 1999. Does the benthic paradox tell us something about surplus production models? Fisheries Research 867:1‐7.
Lønne, O. J., and B. Gulliksen. 1989. Size, age and diet of polar cod, Boreogadus saida (Lepechin, 1773), in ice covered waters. Polar Biology 9:187‐191.
Lønne, O. J., and B. Gulliksen. 1991a. On the distribution of sympagic macro‐fauna in the seasonally ice covered Barents Sea. Polar Biology 11:457‐469.
Lønne, O. J., and B. Gulliksen. 1991b. Sympagic macro‐fauna from multiyear sea‐ice near Svalbard. Polar Biology 11:471‐477.
Low, L. L. 2008. United States of America ‐‐ Alaska region. Pages 163‐205 in R. J. Beamish, editor. Impacts of Climate and Climate Change on the Key Species in the Fisheries in the North Pacific. PICES Scientific Report, No. 35. North Pacific Marine Science Organization (PICES), Sidney, Canada.
Lowry, L. F. 2000. Marine mammal‐sea ice relationships. Pages 91‐96 in H. P. Huntington, editor. Report of the Marine Mammal Commission Workshop: Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic, Girdwood, AK. Marine Mammal Commission.
Lowry, L. F., and F. H. Fay. 1984. Seal eating by walruses in the Bering and Chukchi Seas. Polar Biology 3:11‐18.
Lowry, L. F., and K. J. Frost. 1981. Feeding and trophic relationships of phocid seals and walruses in the eastern Bering Sea. Pages 813‐824 in D. W. Hood and J. A. Calder, editors. The Eastern Bering Shelf: Oceanography and Resources. U.S. Department of Commerce, NOAA, Office of Marine Pollution Assessment, Seattle, WA.
Lowry, L. F., K. J. Frost, and J. J. Burns. 1978. Food of ringed seals and bowhead whales near Point Barrow, Alaska. Canadian Field‐Naturalist 92:67‐70.
Lowry, L. F., K. J. Frost, and J. J. Burns. 1980. Variability in the diet of ringed seals, Phoca hispida, in Alaska. Canadian Journal of Fisheries and Aquatic Sciences 37:2254‐2261.
Lowry, L. F., K. J. Frost, D. Calkins, G. L. Swartzman, and S. Hills. 1982. Feeding habits, food requirements, and status of the Bering Sea marine mammals. Alaska Department of Fish and Game, Council Document No. 19. 292 p.
Lowry, L. F., K. J. Frost, and K. W. Pitcher. 1994. Observations of oiling of harbor seals in Prince William Sound. Pages 209‐225 in T. R. Loughlin, editor. Marine Mammals and the Exxon Valdez. Academic Press, Inc., San Diego, CA.

224
Lukashenko, N. P., A. G. Vol'fson, V. A. Istomin, and V. Chernov. 1971. Trichinellosis of animals in Chukotka, USSR: a general review. International Journal for Parasitology 1:287‐296.
Lukin, L. P., G. N. Ognetov, and N. S. Boiko. 2006. Ecology of the ringed seal in the White Sea. UrO RAN, Ekaterinburg, Russia. 165 p. (Translated from Russian by the Baltic Fund for Nature (BFN), State University of St. Petersburg, Russia).
Lukin, L. R. 1980. The habitat of the White Sea ringed seal during early postnatal development. Marine Biology 5:33‐37.
Lukin, L. R., and V. A. Potelov. 1978. Living conditions and distribution of ringed seal in the White Sea in the winter. Soviet Journal of Marine Biology 4:684‐690.
Lunn, N. J., I. Stirling, and S. N. Nowicki. 1997. Distribution and abundance of ringed (Phoca hispida) and bearded seals (Erignathus barbatus) in western Hudson Bay. Canadian Journal of Fisheries and Aquatic Sciences 54:914‐921.
Lunneryd, S. G., M. Hemmingsson, S. Tärnlund, and A. Fjälling. 2005. A voluntary logbook scheme as a method of monitoring the by‐catch of seals in Swedish coastal fisheries. International Council for the Exploration of the Sea, C.M. 2005/X:04. 10 p.
Lüthi, D., M. Le Floch, B. Bereiter, T. Blunier, J. M. Barnola, U. Siegenthaler, D. Raynaud, J. Jouzel, H. Fischer, K. Kawamura, and T. F. Stocker. 2008. High‐resolution carbon dioxide concentration record 650,000‐800,000 years before present. Nature 453:379‐382.
Lydersen, C. 1991. Monitoring ringed seal (Phoca hispida) activity by means of acoustic telemetry. Canadian Journal of Zoology 69:1178‐1182.
Lydersen, C. 1995. Energetics of pregnancy, lactation and neonatal development in ringed seals (Phoca hispida). Pages 319‐327 in A. S. Blix, L. Walløe, and Ø. Ulltang, editors. Whales, Seals, Fish and Man. Elsevier Science, Amsterdam, Netherlands.
Lydersen, C. 1998. Status and biology of ringed seals (Phoca hispida) in Svalbard. Pages 46‐62 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Lydersen, C., and I. Gjertz. 1984. Studies of the ringed seal (Phoca hispida Schreber 1775) in its breeding habitat in Kongsfjorden, Svalbard. Department of Marine Zoology and Marine Chemistry, University of Oslo, Norsk Polarinstitutt Rapportserie No. 19. 46 p.
Lydersen, C., and I. Gjertz. 1986. Studies of the ringed seal (Phoca hispida Schreber 1775) in its breeding habitat in Kongsfjorden, Svalbard. Polar Research 4:57‐63.
Lydersen, C., and I. Gjertz. 1987. Population parameters of ringed seals (Phoca hispida Schreber, 1775) in the Svalbard area. Canadian Journal of Zoology 65:1021‐1027.
Lydersen, C., and M. O. Hammill. 1993a. Activity, milk intake and energy‐consumption in free‐living ringed seal (Phoca hispida) pups. Journal of Comparative Physiology B 163:433‐438.
Lydersen, C., and M. O. Hammill. 1993b. Diving in ringed seal (Phoca hispida) pups during the nursing period. Canadian Journal of Zoology 71:991‐996.
Lydersen, C., M. O. Hammill, and M. S. Ryg. 1992. Water flux and mass gain during lactation in free‐living ringed seal (Phoca hispida) pups. Journal of Zoology 228:361‐369.
Lydersen, C., P. M. Jensen, and E. Lydersen. 1987. Studies of the ringed seal (Phoca hispida) population in the Van Mijen fiord, Svalbard, in the breeding period 1986. Norsk Polarinstitutt Rapportserie, No. 34. 89‐112 p.
Lydersen, C., P. M. Jensen, and E. Lydersen. 1990. A survey of the Van Mijen Fiord, Svalbard, as habitat for ringed seals, Phoca hispida. Holarctic Ecology 13:130‐133.
Lydersen, C., and K. M. Kovacs. 1999. Behaviour and energetics of ice‐breeding, North Atlantic phocid seals during the lactation period. Marine Ecology Progress Series 187:265‐281.

225
Lydersen, C., O. A. Nøst, K. M. Kovacs, and M. A. Fedak. 2004. Temperature data from Norwegian and Russian waters of the northern Barents Sea collected by free‐living ringed seals. Journal of Marine Systems 46:99‐108.
Lydersen, C., and M. Ryg. 1991. Evaluating breeding habitat and populations of ringed seals Phoca hispida in Svalbard fjords. Polar Record 27:223‐228.
Lydersen, C., and M. S. Ryg. 1990. An evaluation of Tempelfjorden and Sassenfjorden as breeding habitat for ringed seals Phoca hispida. Pages 33‐40 in T. Severinsen and R. Hansson, editors. Environmental Atlas Gipsdalen, Svalbard. Vol. III: Reports on the Fauna of Gipsdalen. Norsk Polarinstitutt Rapportserie, No.66
Lydersen, C., and T. G. Smith. 1989. Avian predation on ringed seal Phoca hispida pups. Polar Biology 9:489‐490.
MacKenzie, B. R., and D. Schiedek. 2007. Daily ocean monitoring since the 1860s shows record warming of northern European seas. Global Change Biology 13:1335‐1347.
Malouf, A. H. 1986. Seals and sealing in Canada. Report of the Royal Commission. Minister of Supply and Services. 3 volumes; Vol. 1: 65 p.; Vol. 62: 622 p.; Vol. 623: 679 p.
Manabe, S., M. J. Spelman, and R. J. Stouffer. 1992. Transient responses of a coupled ocean‐atmosphere model to gradual changes of atmospheric CO2. Part II: seasonal response. Journal of Climate 5:105‐126.
Mansfield, A. W. 1967. Seals of arctic and eastern Canada. Bulletin Fisheries Research Board of Canada 137:1‐35.
Mansfield, A. W. 1970. Population dynamics and exploitation of some Arctic seals. Pages 429‐446 in M. W. Holdgate, editor. Antarctic Ecology. Academic Press, London, UK.
Mansfield, A. W. 1983. The effects of vessel traffic in the arctic on marine mammals and recommendations for future research. Government of Canada Fisheries and Oceans, Canadian Technical Reports in Fisheries and Aquatic Sciences No. 1186. 97 p.
Marine Mammal Commission. 2007. Marine mammals and noise. A sound approach to research and management. Marine Mammal Commission, Report to Congress. 370 p.
Marine Mammal Council. 2008. Russian Federation Government Decrees #1482‐r, #1644‐r, #1603‐r, and #1551‐r. Marine Mammal Council, Moscow, Russia. Accessed February 2010 at http://2mn.org/engl/directory_en.htm.
Martin, J. W., M. M. Smithwick, B. M. Braune, P. F. Hoekstra, D. C. G. Muir, and S. A. Mabury. 2004. Identification of long‐chain perfluorinated acids in biota from the Canadian Arctic. Environmental Science & Technology 38:373‐380.
Mathis, J. T., R. S. Pickart, D. A. Hansell, D. Kadko, and N. R. Bates. 2007. Eddy transport of organic carbon and nutrients from the Chukchi Shelf: impact on the upper halocline of the western Arctic Ocean. Journal of Geophysical Research 112:C05011.
Mattson, M., H. Raunio, O. Pelkonen, and E. Helle. 1998. Elevated levels of cytochrome P4501A (CYP1A) in ringed seals from the Baltic Sea. Aquatic Toxicology 43:41‐50.
McElhany, P., M. H. Ruckelshaus, M. J. Ford, T. C. Wainwright, and E. P. Bjorkstedt. 2000. Viable salmonid populations and the recovery of evolutionarily significant units. U.S. Department of Commerce, NOAA Technical Memorandum NMFS‐NWFSC‐42. 156 p.
McLaren, I. A. 1958a. The biology of the ringed seal (Phoca hispida Schreber) in the eastern Canadian Arctic. Bulletin Fisheries Research Board of Canada 118:97.
McLaren, I. A. 1958b. The economics of seals in the eastern Canadian Arctic. Fisheries Research Board of Canada, Circular No. 1. 94 p.
McLaren, I. A. 1960. On the origin of the Caspian and Baikal seals and the paleoclimatological implication. American Journal of Science 258:47‐65.

226
McLaren, I. A. 1966. Taxonomy of harbor seals of the western North Pacific and evolution of certain other hair seals. Journal of Mammalogy 47:466‐473.
Measures, L. N., J. F. Gosselin, and E. Bergeron. 1997. Heartworm, Acanthocheilonema spirocauda (Leidy, 1858), infections in Canadian phocid seals. Canadian Journal of Fisheries and Aquatic Sciences 54:842‐846.
Mecklenberg, C. W., T. A. Mecklenberg, and L. K. Thorsteinson. 2002. Fishes of Alaska. American Fisheries Society, Bethesda, MD. 1037 p.
Mecklenburg, C. A., D. L. Stein, B. A. Sheiko, N. V. Chernova, T. A. Mecklenburg, and B. A. Holladay. 2007. Russian‐American long‐term census of the Arctic: benthic fishes trawled in the Chukchi Sea and Bering Strait, August 2004. Northwestern Naturalist 88:168‐187.
Medvedev, N., N. Panichev, and H. Hyvärinen. 1997. Levels of heavy metals in seals of Lake Ladoga and the White Sea. Science of the Total Environment 206:95‐105.
Meehl, G. A., T. F. Stocker, W. D. Collins, P. Friedlingstein, A. T. Gaye, J. M. Gregory, A. Kitoh, R. Knutti, J. M. Murphy, A. Noda, S. C. B. Raper, I. G. Watterson, A. J. Weaver, and Z.‐C. Zhao. 2007. Global climate projections. Pages 747‐846 in S. Solomon, D. Qin, M. Manning, M. Marquis, K. Averyt, M. M. B. Tignor, J. Henry LeRoy Miller, and Z. Chen, editors. Climate Change 2007: The Physical Science Basis. Cambridge University Press, Cambridge, UK and New York, NY.
Meier, H. E. M., R. Döscher, and A. Halkka. 2004. Simulated distributions of Baltic Sea‐ice in warming climate and consequences for the winter habitat of the Baltic ringed seal. Ambio 33:249‐256.
Melnikov, I. A., E. G. Kolosova, H. E. Welch, and L. S. Zhitina. 2002. Sea ice biological communities and nutrient dynamics in the Canada Basin of the Arctic Ocean. Deep‐Sea Research I 49:1623‐1649.
Melnikov, V. V., and I. A. Zagrebin. 2005. Killer whale predation in coastal waters of the Chukotka Peninsula. Marine Mammal Science 21:550‐556.
Messier, F., M. K. Taylor, and M. A. Ramsay. 1992. Seasonal activity patterns of female polar bears (Ursus maritimus) in the Canadian Arctic as revealed by satellite telemetry. Journal of Zoology 226:219‐229.
MET Office Hadley Data Centre. 2008. Hadley Centre sea ice and sea surface temperature data sets (HadISST). MET Office Hadley Data Centre, Exeter, Devon UK. Accessed June 2010 at http://hadobs.metoffice.com/hadisst/.
Metsähallitus, editor. 2009. Saimaa ringed seal (Phoca hispida saimensis): the most endangered seal in the world? Metsähallitus, Natural Heritage Services, Savonlinna, Finland. 4 p.
Michaelidis, B., A. Spring, and H. O. Pörtner. 2007. Effects of long‐term acclimation to environmental hypercapnia on extracellular acid‐base status and metabolic capacity in Mediterranean fish Sparus aurata. Marine Biology 150:1417‐1429.
Migaki, G., J. F. Allen, and H. W. Casey. 1977. Toxoplasmosis in a California sea lion (Zalophus californianus). American Journal of Veterinary Research 38:135‐136.
Miller, G. W., and R. A. Davis. 2002. Marine mammal and acoustical monitoring of Anderson Exploration Limited’s open‐water seismic program in the southeastern Beaufort Sea, 2001. LGL Ecological Research Associates, Inc., LGL Report TA 2618‐1. 199 p.
Miller, G. W., R. A. Davis, and K. J. Finley. 1982. Ringed seals in the Baffin Bay region: habitat use. population dynamics and harvest levels. Report Arctic Pilot Project, 94 p. LGL Ltd., Toronto, Canada.
Miller, M. A., I. A. Gardner, C. Kreuder, D. M. Paradies, K. R. Worcester, D. A. Jessup, E. Dodd, M. D. Harris, J. A. Ames, A. E. Packham, and P. A. Conrad. 2002. Coastal freshwater runoff is a risk factor for Toxoplasma gondii infection of southern sea otters (Enhydra lutris nereis). International Journal for Parasitology 32:997‐1006.

227
Miller, M. A., K. Sverlow, P. R. Crosbie, B. C. Barr, L. J. Lowenstine, F. M. Gulland, A. Packham, and P. A. Conrad. 2001. Isolation and characterization of two parasitic protozoa from a Pacific harbor seal (Phoca vitulina richardsi) with meningoencephalomyelitis. Journal of Parasitology 87:816‐822.
Miller, W. G., L. G. Adams, T. A. Ficht, N. F. Cheville, J. P. Payeur, D. R. Harley, C. House, and S. H. Ridgway. 1999. Brucella‐induced abortions and infection in bottlenose dolphins (Tursiops truncatus). Journal of Zoo and Wildlife Medicine 30:100‐110.
Mineev, V. N. 1981. Protection and regulation of the harvest of marine mammals in the Bering and Chukchi seas. Pages 101‐102 in L. A. Popov, editor. Scientific Investigations of the Marine Mammals of the North Pacific Ocean in 1980/81. VNIRO, Moscow, Russia. (Translated from Russian, 3 p.).
Mineev, V. N. 1984. Protection and regulation of the harvest of marine mammals in the Bering and Chukchi seas. Pages 76‐78 in L. A. Popov, editor. Scientific Investigations of the Marine Mammals of the North Pacific Ocean in 1982/83. VNIRO, Moscow, Russia. (Translated from Russian by S. Pearson, 6 p.).
Minerals Management Service. 2007a. Chukchi Sea planning area: oil and gas lease sale 193 and seismic surveying activities in the Chukchi Sea. Final Environmental Impact Statement. U.S. Department of the Interior, Minerals Management Service, Anchorage, AK. 631 p.
Minerals Management Service. 2007b. Proposed final program Outer Continental Shelf oil and gas leasing program 2007‐2012, April 2007. U.S. Department of the Interior, Minerals Management Service, Anchorage, AK. 146 p.
Minerals Management Service. 2008. February 6, 2008 news release: MMS Chukchi Sea lease sale 193 breaks energy records with $2.6 billion in high bids. U.S. Department of the Interior, Minerals Management Service, Accessed August 2008 at http://www.mms.gov/ooc/press/2008/press0206.htm.
Minerals Management Service. 2009a. Alaska OCS Region (Website). Minerals Management Service, Anchorage, AK. Accessed June 2010 at http://www.mms.gov/alaska/fo/index.htm.
Minerals Management Service. 2009b. Draft proposed Outer Continental Shelf (OCS) oil and gas leasing program 2010‐2015. Minerals Management Service, Washington, D.C. 140 p. Accessed at http://www.mms.gov/5‐year/PDFs/2010‐2015/DPP%20FINAL%20(HQPrint%20with%20landscape%20maps).pdf.
Minerals Management Service. 2009c. February 10, 2009 news release: Secretary Salazar details strategy for comprehensive energy plan on U.S. Outer Continental Shelf ‐‐ provides more time for public comment; incorporates renewable energy. U.S. Department of the Interior, Minerals Management Service, Washington, D.C. Accessed May 2010 at http://www.doi.gov/archive/news/09_News_Releases/021009.html.
Minerals Management Service. 2010a. 2007‐2012 5‐year program. Minerals Management Service, Washington, D.C. Accessed June 2010 at http://www.mms.gov/5‐year/2007‐2012FiveYearProgram.htm.
Minerals Management Service. 2010b. Introduction ‐‐ 5‐year leasing program. Minerals Management Service, Washington, D.C. Accessed June 2010 at http://www.mms.gov/5‐year/.
Minerals Management Service. 2010c. Preliminary revised program Outer Continental Shelf oil and gas leasing program 2007‐2012. Minerals Management Service, Washington, D.C. 216 p. Accessed at http://www.mms.gov/5‐year/PDFs/PRP2007‐2012.pdf.
Ministry of Agriculture and Forestry. 2007. Management plan for the Finnish seal populations in the Baltic Sea. Ministry of Agriculture and Forestry. 95 p.
Ministry of Agriculture and Forestry. 2010a. January 2, 2010 news release: Follow‐up group for Saimaa ringed seal and fishing: a single protected area needed for the Saimaa seal. Valtioneuvosto

228
Finnish Government, Helsinki, Finland. Accessed April 2010 at http://www.vn.fi/ajankohtaista/tiedotteet/tiedote/sv.jsp.print?oid=286843&c=0&toid=2213.
Ministry of Agriculture and Forestry. 2010b. September 4, 2010 news release: Net fishing ban to protect Saimaa seal pups. Valtioneuvosto Finnish Government, Helsinki, Finland. Accessed April 2010 at http://www.government.fi/ajankohtaista/tiedotteet/tiedote/en.jsp.print?oid=292311.
Ministry of Environment and Hokkaido Prefectural Government. 2007. The multiple use marine management plan and explanatory material for Shiretoko World Natural Heritage Site. Ministry of Environment, Government of Japan, and Hokkaido Prefectural Government, Tokyo, Japan. Accessed May 2010 at http://www.env.go.jp/en/nature/npr/fcpn/index.html.
Ministry of the Environment. 2000. Red list of threatened mammals of Japan. Ministry of the Environment, Tokyo, Japan. Accessed May 2010 at http://www.biodic.go.jp/english/rdb/rdb_e.html#red6.
Ministry of the Environment. 2009. Protection and management of wildlife. Pages 15‐21 in For Coexistence of People and Nature. Nature Conservation Bureau, Tokyo, Japan.
Ministry of the Environment and Hokkaido Prefectural Government. 2007. The multiple use marine management plan and explanatory material for Shiretoko World Natural Heritage Site. Ministry of the Environment, Government of Japan, and Hokkaido Prefectural Government. 26 p.
Møller, P. R., J. G. Nielsen, S. W. Knudsen, J. Y. Poulsen, K. Sünksen, and O. A. Jørgensen. 2010. A checklist of the fish fauna of Greenland waters. Zootaxa 2378:1‐84.
Moore, S. E., and H. P. Huntington. 2008. Arctic marine mammals and climate change: impacts and resilience. Ecological Applications 18:S157‐S165.
Moran, K., J. Backman, H. Brinkhuis, S. C. Clemens, T. Cronin, G. R. Dickens, F. Eynaud, J. Gattacceca, M. Jakobsson, R. W. Jordan, M. Kaminski, J. King, N. Koc, A. Krylov, N. Martinez, J. Matthiessen, D. McInroy, T. C. Moore, J. Onodera, M. O'Regan, H. Palike, B. Rea, D. Rio, T. Sakamoto, D. C. Smith, R. Stein, K. St John, I. Suto, N. Suzuki, K. Takahashi, M. Watanabe, M. Yamamoto, J. Farrell, M. Frank, P. Kubik, W. Jokat, and Y. Kristoffersen. 2006. The Cenozoic palaeoenvironment of the Arctic Ocean. Nature 441:601‐605.
Morison, J., K. Aagaard, and M. Steele. 2000. Recent environmental changes in the Arctic: a review. Arctic 53:359‐371.
Moulton, V. D., W. J. Richardson, R. E. Elliott, T. L. McDonald, C. Nations, and M. T. Williams. 2005. Effects of an offshore oil development on local abundance and distribution of ringed seals (Phoca hispida) of the Alaskan Beaufort Sea. Marine Mammal Science 21:217‐242.
Moulton, V. D., W. J. Richardson, T. L. McDonald, R. E. Elliott, and M. T. Williams. 2002. Factors influencing local abundance and haulout behaviour of ringed seals (Phoca hispida) on landfast ice of the Alaskan Beaufort Sea. Canadian Journal of Zoology 80:1900‐1917.
Mueter, F. J., C. Broms, K. F. Drinkwater, K. D. Friedland, J. A. Hare, G. L. Hunt, Jr., W. Melle, and M. Taylor. 2009. Ecosystem response to recent oceanographic variability in high‐latitude Northern Hemisphere ecosystems. Progress in Oceanography 81:93‐110.
Mueter, F. J., and M. A. Litzow. 2008. Sea ice retreat alters the biogeography of the Bering Sea continental shelf. Ecological Applications 18:309‐320.
Mueter, F. J., and B. A. Megrey. 2006. Using multi‐species surplus production models to estimate ecosystem‐level maximum sustainable yields. Fisheries Research 81:189‐201.
Muir, D., B. Braune, B. DeMarch, R. Norstrom, R. Wagemann, L. Lockhart, B. Hargrave, D. Bright, R. Addison, J. Payne, and K. Reimer. 1999. Spatial and temporal trends and effects of contaminants in the Canadian Arctic marine ecosystem: a review. Science of the Total Environment 230:83‐144.

229
Müller‐Wille, L. L. 1969. Biometrical comparison of four populations of Phoca hispida Schreb. in the Baltic and White Seas and Lakes Ladoga and Saimaa. Commentationes Biologicae Societas Scientiarum Fennica 31:1‐12.
Murawski, S. A. 1993. Climate change and marine fish distributions: forecasting from historical analogy. Transactions of the American Fisheries Society 122:647‐658.
Murawski, S. A., and J. T. Finn. 1988. Biological bases for mixed‐species fisheries: species co‐distribution in relation to environmental and biotic variables. Canadian Journal of Fisheries and Aquatic Sciences 45:1720‐1735.
Murdoch, J. 1885. Part IV ‐‐ Natural History, I. ‐‐ Mammals. Pages 92‐103 in P. H. Ray, editor. Report of the International Polar Expedition to Point Barrow, Alaska. The Resolution of the House of Representatives of December 11, 1884. 48th Congress, 2nd Session, House Executive Document 44. Government Printing Office, Washington, D.C.
Naito, Y. 1971. Synopsis of the seal fishery. Scientific Reports of the Whales Research Institute 238:49‐52.
Nakata, H., S. Tanabe, R. Tatsukawa, Y. Koyama, N. Miyazaki, S. Belikov, and A. Boltunov. 1998. Persistent organochlorine contaminants in ringed seals (Phoca hispida) from the Kara Sea, Russian Arctic. Environmental Toxicology and Chemistry 17:1745‐1755.
Nakicenovic, N., and R. Swart, editors. 2000. Special report on emissions scenarios: a special report of Working Group III of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. 612 p.
Nammack, M., L. Manning, A. Mullen, and S. Rumsey. 2007. Interim protocol for conducting Endangered Species Act status reviews. National Marine Fisheries Service, November 2007 Draft. 43 p.
NAMMCO. 2002. Status of marine mammals in the North Atlantic; the ringed seal. North Atlantic Marine Mammal Commission (NAMMCO), Tromsø, Norway. 7 p. Accessed at http://www.nammco.no/webcronize/images/Nammco/653.pdf.
NAMMCO. 2004. Report of the NAMMCO workshop on hunting methods for seals and walrus. North Atlantic Marine Mammals Commission (NAMMCO), North Atlantic House Copenhagen, Denmark, 7–9 September 2004. 60 p.
National Marine Fisheries Service. 1993. Endangered and threatened species; Saimaa seal. Federal Register 58:86 (May 6, 1993) p. 26920‐26921.
National Marine Fisheries Service. 2008a. Notice of 12‐month finding on a petition to list the ribbon seal as a threatened or endangered species. Federal Register 73:250 (December 30, 2008) p. 79822‐79828.
National Marine Fisheries Service. 2008b. Notice of 90‐day finding on a petition to list the ribbon seal as a threatened or endangered species. Federal Register 73:61 (March 28, 2008) p. 16617‐16619.
National Marine Fisheries Service. 2008c. Notice of 90‐day finding on a petition to list the three ice seal species as a threatened or endangered species. Federal Register 73:172 (September 4, 2008) p. 51615‐51617.
National Marine Fisheries Service. 2009. Proposed threatened and not warranted status for distinct population segments of the spotted seal. Federal Register 74:201 (October 20, 2009) p. 53683‐53696.
National Marine Fisheries Service. 2010. Saimaa seal (Phoca hispida saimensis). NOAA Fisheries Office of Protected Resources, Silver Spring, MD. Accessed April 2010 at http://www.nmfs.noaa.gov/pr/species/mammals/pinnipeds/saimaaseal.htm.
National Research Council. 2010. Ocean acidification: a national strategy to meet the challenges of a changing ocean. National Academies Press, Washington, D.C. 175 p.
National Snow and Ice Data Center. 2007. Arctic sea ice shatters all previous record lows. Diminished summer sea ice leads to opening of the fabled Northwest Passage. NSIDC, Boulder, CO. Accessed

230
September 2008 at http://www.nsidc.org/news/press/2007_seaiceminimum/20071001_pressrelease.html.
National Snow and Ice Data Center. 2008. October 2, 2008 news release: Arctic sea ice down to second‐lowest extent; likely record‐low volume. Despite cooler temperatures and ice‐favoring conditions, long‐term decline continues. NSIDC, Boulder, CO. Accessed February 2009 at http://www.nsidc.org/news/press/20081002_seaice_pressrelease.html.
Nazarenko, Y. I. 1965. Contributions to the study of the reproduction of the ringed seal (Phoca hispida) of the Choska Inlet. Pages 171‐175 in Marine Mammals. Nauka Publishing House (Translated from Russian by the Fisheries Research Board of Canada, Quebec, Canada, Translation Series No. 1461, 10 p.).
Nazarenko, Y. I. 1967. Feeding of the ringed seal of the European north of the USSR. Canadian Translations of Fisheries and Aquatic Sciences 21:81‐85. (Translated from Russian by Canada Institiute for Scientific and Technical Information, Ottowa, Canada, Number 5166, 9 p.).
Nazarenko, Y. I. 1969. Data on the hunting of the northern ringed seal. Pages 234‐236 in Marine Mammals (Translated from Russian by the Research Fisheries Board of Canada, Quebec, Candada, Tranlsation Series No. 1462, 8 p.).
Niebauer, H. J. 1988. Effects of El Nino‐Southern Oscillation and North Pacific weather patterns on interannual variability in the subarctic Bering Sea. Journal of Geophyisical Research 93:5051‐5068.
Nielsen, O., K. Nielsen, and R. E. A. Stewart. 1996. Serologic evidence of Brucella spp. exposure in Atlantic walruses (Odobenus rosmarus rosmarus) and ringed seals (Phoca hispida) of Arctic Canada. Arctic 49:383‐386.
Noongwook, G. 2000. Native observations of local climate change around St. Lawrence Island. Pages 21‐24 in H. P. Huntington, editor. Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic. Marine Mammal Commission, Bethesda, MD.
North Pacific Fishery Management Council. 2009. Fishery management plan for fish resources of the Arctic management area. NPFMC, Anchorage, AK. 158 p. Accessed May 2010 at http://www.fakr.noaa.gov/npfmc/fmp/arctic/ArcticFMP.pdf.
Nyman, M., M. Bergknut, M. L. Fant, H. Raunio, M. Jestoi, C. Bengs, A. Murk, J. Koistinen, C. Backman, O. Pelkonen, M. Tysklind, T. Hirvi, and E. Helle. 2003. Contaminant exposure and effects in Baltic ringed and grey seals as assessed by biomarkers. Marine Environmental Research 55:73‐99.
Nyman, M., J. Koistinen, M. L. Fant, T. Vartiainen, and E. Helle. 2002. Current levels of DDT, PCB and trace elements in the Baltic ringed seals (Phoca hispida baltica) and grey seals (Halichoerus grypus). Environmental Pollution 119:399‐412.
Nyman, M., H. Raunio, P. Taavitsainen, and O. Pelkonen. 2001. Characterization of xenobiotic‐metabolizing cytochrome P450 (CYP) forms in ringed and grey seals from the Baltic Sea and reference sites. Comparative Biochemistry and Physiology C 128:99‐112.
O'Shea, T. J. 1999. Environmental contaminants and marine mammals. Pages 485‐563 in J. E. I. Reynolds and S. A. Rommel, editors. Biology of Marine Mammals. Smithsonian Institution Press, Washington, D. C.
Ognetov, G. N. 2002. Estimate of the ringed seal (Phoca hispida) abundance in the White, Barents, and Kara seas. Pages 209‐210 in V. M. Belkovich, editor. Marine Mammals of the Holarctic. 2002. Abstracts of Reports, Baikal, Russia. Marine Mammal Council.
Ognev, S. I. 1935. Mammals of U.S.S.R. and adjacent countries. Volume 3. Carnivora. Glavpushnina NKVT, Moscow, Russia. 641 p. (Translated from Russian by the Israel Program for Scientific Translations, Jerusalem 1962, 741 p.).

231
Oksanen, A., M. Tryland, K. Johnsen, and J. P. Dubey. 1998. Serosurvey of Toxoplasma gondii in North Atlantic marine mammals by the use of agglutination test employing whole tachyzoites and dithiothreitol. Comparative Immunology Microbiology and Infectious Diseases 21:107‐114.
Olson, M. E., P. D. Roach, M. Stabler, and W. Chan. 1997. Giardiasis in ringed seals from the western Arctic. Journal of Wildlife Diseases 33:646‐648.
Olsson, M., B. Karlsson, and E. Ahnland. 1994. Diseases and environmental contaminants in seals from the Baltic and the Swedish west coast. Science of the Total Environment 154:217‐227.
Olsson, M., L. Reutergardh, and A. V. Yablokov. 1986. DDT and PCB levels in ringed seal (Pusa hispida) from Lake Ladoga and the Gulf of Bothnia. Pages 117‐129 in A. V. Yablokov and M. Olsson, editors. Influence of Human Activities on the Baltic Ecosystem. Proceedings of the Soviet‐Swedish Symposium, Effects of Toxic Substances on Dynamics of Seal Populations, Moscow, USSR.
Omstedt, A. B., B. Gustafsson, J. Rodhe, and G. Walin. 2000. Use of Baltic Sea modelling to investigate the water cycle and the heat blance in GCM and regional climate models. Climate Research 15:95‐108.
Onderka, D. K. 1989. Prevalence and pathology of nematode infections in the lungs of ringed seals (Phoca hispida) of the western Arctic of Canada. Journal of Wildlife Diseases 25:218‐224.
Oppenheimer, M., B. C. O'Neill, M. Webster, and S. Agrawala. 2007. Climate change: the limits of consensus. Science 317:1505‐1506.
Oreskes, N. 2004. The scientific consensus on climate change. Science 306:1686. Orr, J. C., V. J. Fabry, O. Aumont, L. Bopp, S. C. Doney, R. A. Feely, A. Gnanadesikan, N. Gruber, A. Ishida,
F. Joos, R. M. Key, K. Lindsay, E. Maier‐Reimer, R. Matear, P. Monfray, A. Mouchet, R. G. Najjar, G.‐K. Plattner, K. B. Rodgers, C. L. Sabine, J. L. Sarmiento, R. Schlitzer, R. D. Slater, I. J. Totterdell, M.‐F. Weirig, Y. Yamanaka, and A. Yool. 2005. Anthropogenic ocean acidification over the twenty‐first century and its impact on calcifying organisms. Nature 437:681‐686.
Orr, J. C., S. Jutterström, L. Bopp, L. G. Anderson, P. Cadule, V. J. Fabry, T. Frölicher, E. P. Jones, F. Joos, A. Lenton, E. Maier‐Reimer, J. Segschneider, M. Steinacher, and D. Swingedouw. 2009. Amplified acidification of the Arctic Ocean (Abstract). IOP Conference Series: Earth and Environmental Science 6:462009.
OSPAR Commission. 2009. Overview of the impacts of anthropogenic underwater sound in the marine environment. OSPAR Commission, Biodiversity Series. 134 p.
Osterhaus, A., J. Groen, P. De Vries, F. Uytde Haag, B. Klingeborn, and R. Zarnke. 1988. Canine distemper virus in seals. Nature 335:403‐404.
Osterhaus, A., J. Groen, H. E. M. Spijkers, H. W. J. Broeders, F. UytdeHaag, P. de Vries, J. S. Teppema, and et al. 1990. Mass mortality in seals caused by a newly discovered virus‐like morbillivirus.
Osterhaus, A., H. Yang, H. E. M. Spijkers, J. Groen, J. S. Teppema, and G. Van Steenis. 1985. The isolation and partial characterization of a highly pathogenic herpesvirus from the harbor seal (Phoca vitulina). Archives of Virology 86:239‐251.
Overland, J. E., and P. J. Stabeno. 2004. Is the climate of the Bering Sea warming and affecting the ecosystem? EOS, Transactions of the American Geophysical Union 85:309‐316.
Overland, J. E., M. Wang, N. A. Bodn, J. E. Walsh, V. M. Kattsov, and W. L. Chapman. In review. Considerations in the selection of global climate models for regional climate projections: the Arctic as a case study. Journal of Climate.
Overland, J. E., and M. Y. Wang. 2007. Future regional Arctic sea ice declines. Geophysical Research Letters 34:L17705.
Palo, J. 2003. Genetic diversity and phylogeography of landlocked seals. Dissertation. Universiy of Helsinki, Helsinki, Finland. 29 p.

232
Palo, J. U., H. Hyvärinen, E. Helle, H. S. Mäkinen, and R. Väinölä. 2003. Postglacial loss of microsatellite variation in the landlocked Lake Saimaa ringed seal. Conservation Genetics 4:117‐128.
Palo, J. U., H. S. Mäkinen, E. Helle, O. Stenman, and R. Väinölä. 2001. Microsatellite variation in ringed seals (Phoca hispida): genetic structure and history of the Baltic Sea population. Heredity 86:609‐617.
Pamperin, N. J., E. H. Follmann, and B. T. Person. 2008. Sea‐ice use by arctic foxes in northern Alaska. Polar Biology 31:1421‐1426.
Parmesan, C., and G. Yohe. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37‐43.
Patin, S. 1999. Russian oil and gas offshore. EcoMonitor Publishing, East Northport, NY. Accessed April 29, 2010 at http://www.offshore‐environment.com/russianoil.html.
PCMDI. 2010. Welcome to PCMDI. Program for Climate Model Diagnosis and Intercomparison, Livermore, CA. Accessed June 2010 at http://www‐pcmdi.llnl.gov/.
Percy, J. A. 1983. Distribution of Arctic marine isopods of the Mesidotea (= Saduria) complex in relation to depth, temperature, and salinity in the southern Beaufort Sea. Arctic 36:3431‐3439.
Perry, A. L., P. J. Low, J. R. Ellis, and J. D. Reynolds. 2005. Climate change and distribution shifts in marine fishes. Science 308:1912‐1915.
PICES. 2005. Report of the study group on fisheries and ecosystem responses to recent regime shifts. North Pacific Marine Science Organization (PICES), PICES Scientific Report No. 28. 162 p.
Pickart, R. S., T. Weingartner, L. J. Pratt, S. T. Zimmerman, and D. J. Torres. 2005. Flow of winter‐transformed Pacific water into the western Arctic. Deep‐Sea Research II 52:3175‐3198.
Pikharev, G. A. 1946. The food of the seal Phoca hispida. Izvestia TNIRO 22:259‐261. (Translated from Russian by the Bureau for Translations, Ottawa, Canada, Translation Series No. 150, 4 p.).
Polterman, M., H. Hop, and S. Falk‐Petersen. 2000. Life under Arctic sea ice ‐‐ reproduction strategies of two sympagic (ice‐associated) amphipod species, Gammarus wilkitzkii and Apherusa glacialis. Marine Biology 136:913‐920.
Polyak, L., R. B. Alley, J. T. Andrews, J. Brigham‐Grette, T. M. Cronin, D. A. Darby, A. S. Dyke, J. J. Fitzpatrick, S. Funder, M. Holland, A. E. Jennings, G. H. Miller, M. O'Regan, J. Savelle, M. Serreze, K. St. John, J. W. C. White, and E. Wolff. 2010. History of sea ice in the Arctic. Quaternary Science Reviews 29:1757‐1778.
Popov, L. 1979. Ladoga seal. Pages 70‐71 in FAO Fisheries Series No. 5. Mammals in the Seas. Volume II: Pinniped Species Summaries and Report on Sirenians. Food and Agriculture Organization of the United Nations, Rome, Italy.
Popov, L. A. 1976. Status of main ice forms of seals inhabiting waters of the USSR and adjacent to the country marine areas. Pages 1‐17 in Scientific Consultation on Marine Mammals, Bergen, Norway. Food and Agriculture Organization of the United Nations.
Popov, L. A. 1980. Soviet investigations of the Baltic seals and legislation to protect their stocks. Research Institute of Marine Fisheries and Oceanography (VNIRO) 17:10‐11.
Popov, L. A. 1982. Status of the main ice‐living seals inhabiting inland waters and coastal marine areas of the USSR. Pages 361‐381 in FAO Fisheries Series No. 5. Mammals in the Seas. Volume IV‐‐Small Cetaceans, Seals, Sirenians and Otters. Food and Agriculture Organization of the United Nations, Rome, Italy.
Popov, V. N. 1975. Helminth fauna of Phocidae in the Sea of Okhotsk in Fall. Problemy Parazitologii 2:116‐118. (Translated from Russian by Canadian Translation of Fisheries and Aquatic Sciences, Ottawa, Ontario, Canada, No. 5608, 3 p.).
Popov, V. N., M. V. Jurakhno, and V. A. Skryabin. 1980. Helminth fauna of the Okhotsk ringed seal living in Patience Bay and Laperouse Strait. Parazitologiya 14:48‐54. (Translated from Russian by the

233
Canadian Translation of Fisheries and Aquatic Sciences, Ottawa, Ontario, Canada, No. 5590, 16 p.).
Pörtner, H. 2008. Ecosystem effects of ocean acidification in times of ocean warming: a physiologist's view. Marine Ecology Progress Series 373:203‐217.
Pörtner, H. O., M. Langenbuch, and B. Michaelidis. 2005. Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: from Earth history to global change. Journal of Geophysical Research 110:C09S10.
Pörtner, H. O., M. Langenbuch, and A. Reipschläger. 2004. Biological impact of elevated CO2 concentrations: lessons from animal physiology and earth history. Journal of Oceanography 60:705‐718.
Priest, H., and P. J. Usher. 2004. The Nunavut wildlife harvest study. Nunavut Wildlife Management Board, Final Report. 822 p.
Pungowiyi, C. 2000. Native observations of change in the marine environment of the Bering Strait Region. Page 98 in Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic, Girdwood, AK. Marine Mammal Commission.
Quakenbush, L., and G. Sheffield. 2007. Ice seal bio‐monitoring in the Bering‐Chukchi Sea region. North Pacific Research Board (NPRB) Project 312 Final Report, 47 p. Alaska Department of Fish and Game, Fairbanks, AK.
Quakenbush, L. T. 2007. Polybrominated diphenyl ether compounds in ringed, bearded, spotted, and ribbon seals from the Alaskan Bering Sea. Marine Pollution Bulletin 54:232‐236.
Quakenbush, L. T., and J. J. Citta. 2008. Perfluorinated contaminants in ringed, bearded, spotted, and ribbon seals from the Alaskan Bering and Chukchi Seas. Marine Pollution Bulletin 56:1809‐1814.
Radionov, V. F., N. N. Bryazgin, and E. I. Alexandrov. 1997. The snow cover of the Arctic Basin. Arctic and Antarctic Institute, Technical Report, APL‐UW TR 9701. 98 p.
Räisänen, J. 2007. How reliable are climate models? Tellus 59A:2‐29. Räisänen, J., L. Ruokolainen, and J. Ylhäisi. 2010. Weighting of model results for improving best
estimates of climate change. Climate Dynamics 35:407‐422. Ramboll Oil and Gas. 2010. Ramboll Oil and Gas. Ramboll Group, Esbjerg, Denmark. Accessed April 2010
at http://www.ramboll‐oilgas.com/. Randall, D. A., R. A. Wood, S. Bony, R. Colman, T. Fichefet, J. Fyfe, V. Kattsov, A. Pitman, J. Shukla, J.
Srinivasan, R. J. Stouffer, A. Sumi, and K. E. Taylor. 2007. Climate models and their evaluation. Pages 589‐662 in S. Solomon, D. Qin, M. Manning, M. Marquis, K. Averyt, M. M. B. Tignor, J. Henry LeRoy Miller, and Z. Chen, editors. Climate Change 2007: The Physical Science Basis. Cambridge University Press, Cambridge, UK and New York, NY.
Rasmus, S., J. Räisänen, and M. Lehning. 2004. Estimating snow conditions in Finland in the late 21st century using the SNOWPACK model with regional climate scenario data as input. Annals of Glaciology 38:238‐244.
Rassi, P., A. Alanen, T. Kanerva, and I. Mannerkoski. 2001. The red list of Finnish species. Ministry of the Environment and Finnish Environment Institute, Helsinki, Finland. Accessed February 2010 at http://www.environment.fi/print.asp?contentid=262131&lan=en&clan=en.
Rausch, R., B. B. Babero, R. V. Rausch, and E. L. Schiller. 1956. Studies on the helminth fauna of Alaska. XXVII. The occurrence of larvae of Trichinella spiralis in Alaskan mammals. Journal of Parasitology 42:259‐271.
Rautio, A., M. Niemi, M. Kunnasranta, I. J. Holopainen, and H. Hyvärinen. 2009. Vocal repertoire of the Saimaa ringed seal (Phoca hispida saimensis) during the breeding season. Marine Mammal Science 25:920‐930.
Raven, J., K. Caldeira, H. Elderfield, O. Hoegh‐Guldberg, P. S. Liss, U. Reisbell, J. Shepard, C. Turley, and A. J. Watson. 2005. Ocean acidification due to increasing atmospheric carbon dioxide. The Royal

234
Society, London, UK. 60 p. Accessed July 2010 at http://dge.stanford.edu/labs/caldeiralab/Caldeira%20downloads/RoyalSociety_OceanAcidification.pdf.
Ray, C. E. 1976. Geography of phocid evolution. Systematic Zoology 25:391‐406. Rayner, N. A., D. E. Parker, E. B. Horton, C. K. Folland, L. V. Alexander, D. P. Rowell, E. C. Kent, and A.
Kaplan. 2003. Global analyses of sea surface temperature, sea ice, and night marine air temperature since the late nineteenth century. Journal of Geophysical Research‐Atmospheres 108:4407.
Reeves, R., R. L. J. Brownell, A. M. Burdin, J. C. Cooke, J. D. Darling, G. P. Donovan, F. Gulland, S. E. Moore, D. P. Nowacek, T. J. Ragen, R. G. Steiner, G. R. VanBlaricom, A. Vedenev, and A. V. Yablakov. 2005. Report of the independent scientific review panel on the impacts of Sakhalin II phase 2 on western North Pacific gray whales and related biodiversity. International Union for the Conservation and Nature and Natural Resources (IUCN). 123 p.
Reeves, R. R. 1998. Distribution, abundance and biology of ringed seals (Phoca hispida): an overview. Pages 9‐45 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Reeves, R. R., G. W. Wenzel, and M. C. S. Kingsley. 1998. Catch history of ringed seals (Phoca hispida) in Canada. Pages 100‐129 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Reichler, T., and J. Kim. 2008. How well do coupled models simulate today's climate? Bulletin of the American Meteorological Society 89:303‐311.
Repenning, C. A., C. E. Ray, and D. Grjgorescu. 1979. Pinniped biogeography. Pages 357‐369 in J. Gray and A. J. Boucot, editors. Historical Biogeography, Plate Tectonics and the Changing Environment. Oregon State University Press, Corvallis, OR.
Rey, F., H. R. Skjoldal, and D. Slagstad. 1987. Primary production in relation to climatic changes in the Barents Sea. Pages 29‐46 in H. Loeng, editor. The Effect of Oceanographic Conditions on Distribution and Population Dynamics of Commercial Fish Stocks in the Barents Sea. Institute of Marine Research, Bergen, Norway.
Rhyan, J. C., T. Gidlewski, D. R. Ewalt, S. G. Hennager, D. M. Lambourne, and S. C. Olsen. 2001. Seroconversion and abortion in cattle experimentally infected with Brucella sp. isolated from a Pacific harbor seal (Phoca vitulina richardsi). Journal of Veterinary Diagnostic Investigation 13:379‐382.
Rice, D. W. 1998. Marine mammals of the world: systematics and distribution. Society for Marine Mammalogy, Lawrence, KS. 231 p.
Richardson, W. J., C. R. Greene, Jr., C. I. Malme, and D. H. Thomson. 1995. Marine mammals and noise. Academic Press, Inc., San Diego, CA. 576 p.
Richardson, W. J., and M. T. Williams. 2004. Monitoring of industrial sounds, seals, and bowhead whales near BP’s Northstar oil development, Alaskan Beaufort Sea, 1999‐2003. Annual and comprehensive report, Dec 2004. A report from LGL Ltd., Greenridge Sciences Inc., and WEST Inc. for BP Exploration (Alaska) Inc. LGL Report TA 4001, 297 p. BP Exploration (Alaska) Inc., Anchorage, AK.
Richter‐Menge, J., and J. E. Overland. 2009. Arctic report card 2009. 77 p. Accessed June 2010 at http://www.arctic.noaa.gov/reportcard.
Riebesell, U., I. Zondervan, B. Rost, P. D. Tortell, R. E. Zeebe, and F. M. M. Morel. 2000. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 407:364‐367.
Rigét, F., D. Muir, M. Kwan, T. Savinova, M. Nyman, V. Woshner, and T. O'Hara. 2005. Circumpolar pattern of mercury and cadmium in ringed seals. Science of the Total Environment 351:312‐322.

235
Rigét, F., K. Vorkamp, R. Dietz, and S. C. Rastogi. 2006. Temporal trend studies on polybrominated diphenyl ethers (PBDEs) and polychlorinated biphenyls (PCBs) in ringed seals from East Greenland. Journal of Environmental Monitoring 8:1000‐1005.
Ross, P., R. De Swart, R. Addison, H. Van Loveren, J. Vos, and A. Osterhaus. 1996. Contaminant‐induced immunotoxicity in harbour seals: Wildlife at risk? Toxicology 112:157‐169.
Roth, H. 1950. Nouvelles experiences sur la trichinose, avec considerations speciales sur son existence dans les regions arctiques. Bulletin de I'Office International des Etpizooties 34:197‐220.
Routti, H., B. van Bavel, R. J. Letcher, A. Arukwe, S. G. Chu, and G. W. Gabrielsen. 2009. Concentrations, patterns and metabolites of organochlorine pesticides in relation to xenobiotic phase I and II enzyme activities in ringed seals (Phoca hispida) from Svalbard and the Baltic Sea. Environmental Pollution 157:2428‐2434.
Rybczynski, N., M. R. Dawson, and R. H. Tedford. 2009. A semi‐aquatic Arctic mammalian carnivore from the Miocene epoch and origin of Pinnipedia. Nature 458:1021‐1024.
Ryg, M., and N. A. Øritsland. 1991. Estimates of energy expenditure and energy consumption of ringed seals (Phoca hispida) throughout the year. Polar Research 10:595‐602.
Ryg, M., T. G. Smith, and N. A. Øritsland. 1990. Seasonal changes in body mass and body composition of ringed seals (Phoca hispida) on Svalbard. Canadian Journal of Zoology 68:470‐475.
Ryg, M., Y. Solberg, C. Lydersen, and T. G. Smith. 1992. The scent of rutting male ringed seals (Phoca hispida). Journal of Zoology 226:681‐689.
Saelthun, N. R., P. Aittoniemi, S. Bergström, K. Einarsson, T. Jóhannesson, G. Lindström, P. E. Ohlsson, T. Thomsen, B. Vehviläinen, and K. O. Aamodt. 1998. Climate change impacts on runoff and hydropower in the Nordic countries. TemaNord 1998: 552, Nordic Council of Ministers, Final Report from the Project "Climate Change and Energy Production".
Sakhalin Energy Investment Company. 2005. Marine mammals (other than western gray whales). Environmental Impact Assessment Addendum, Chapter 5. Sakhalin Energy Investment Company, Yuzhno‐Sakhalinsk, Russia. 55 p. Accessed June 2010 at http://www.sakhalinenergy.ru/en/library.asp?p=lib_environment_shelf&l=lib_environment_additional.
Salazar, S. 2003. Impacts of the Jessica oil spill on sea lion (Zalophus wollebaeki) populations. Marine Pollution Bulletin 47:313‐318.
Salemaa, H., I. Vuorinen, and P. Välipakka. 1990. The distribution and abundance of Mysis populations in the Baltic Sea. Annales Zoologici Fennici 27:253‐257.
Santana‐Casiano, J. M., M. Gonzalez‐Davila, M. J. Rueda, O. Llinas, and E. F. Gonzalez‐Davila. 2007. The interannual variability of oceanic CO2 parameters in the northeast Atlantic subtropical gyre at the ESTOC site. Global Biogeochemical Cycles 21:GB1015.
Sasaki, H., K. Numachi, and M. A. Grachev. 2003. The origin and genetic relationships of the Baikal seal, Phoca sibirica, by restriction analysis of mitochondrial DNA. Zoological Science 20:1417‐1422.
Sasaki, Y. N., Y. Katagiri, S. Minobe, and I. G. Rigor. 2007. Autumn atmospheric preconditioning for interannual variability of wintertime sea‐ice in the Okhotsk Sea. Journal of Oceanography 63:255‐265.
Scheffer, V. B. 1958. Seals, sea lions and walruses: a review of the Pinnipedia. Stanford University Press, Palo Alto, CA. 179 p.
Schei, P. J., and R. D. Brubaker. 2006. Suggestions for a conceptual design of an Okhotsk Sea environmental regime ‐‐ comparison with the Barents Sea. Pages 259‐276 in H. Kitagawa, editor. New Era in Far East Russian and Asia. Ocean Policy Research Foundation, Tokyo, Japan.
Schumacher, J. D., N. A. Bond, R. Brodeur, P. Livingston, J. Napp, and P. Stabeno. 2003. Climate change in the southeastern Bering Sea and some consequences for habitat. Pages 17‐40 in G. Hempel

236
and K. Sherman, editors. Large Marine Ecosystems of the World: Trends in Exploitation, Protection and Research. Elsevier, Amsterdam, UK.
Scott, J. S. 1982. Depth, temperature and salinity preferences of common fishes of the Scotian Shelf. Journal of Northwest Atlantic Fishery Science 3:29‐39.
Seal Conservation Society. 2001. Ringed seal (Phoca hispida). Seal Conservation Society, Saintfield, Ireland. Accessed May 2010 at http://www.pinnipeds.org/species/ringed.htm.
Sell, S. K. 2008. Investigating population structure and philopatry in ringed seals (Phoca hispida). M.S. Thesis. Central Michigan University, Mount Pleasant, Michigan. 35 p.
Serreze, M. S., J. A. Maslanik, and J. R. Key. 1997. Atmospheric and sea ice characteristics of the Arctic Ocean and the SHEBA Field Region in the Beaufort Sea. National Snow and Ice Data Center, Special Report 4.
Shapiro, L. H., R. E. Metzner, and K. Toovak, Jr. 1979. Historical references to ice conditions along the Beaufort Sea coast of Alaska. Pages 632‐682 in Environmental Assessment of the Alaskan Continental Shelf. Annual Reports of Principal Invesitoators for the year ending March 1979. Volume 9. Hazards. U.S. Department of Commerce, NOAA and U.S. Department of Interior, Bureau of Land Management, Boulder, CO.
Shaughnessy, P. D., and F. H. Fay. 1977. A review of taxonomy and nomenclature of North Pacific harbor seals. Journal of Zoology 182:385‐419.
Sheiko, B. A., and V. V. Federov. 2000. Chapter 1. Class Cephalaspidomorphi ‐ lampreys. Class Chondrichthyes ‐ cartilaginous fishes. Class Holocephali ‐ chimaera. Class Osteichthyes ‐ bony fishes. Pages 7‐69 in R. S. Moiseev and A. M. Tokranov, editors. Catalog of Vertebrates of Kamchatka and Adjancent Waters. Kamchatsky Petchatniy Dvor, Petropavlovsk‐Kamchatsky, Russia.
Shell Gulf of Mexico Inc. 2009. Exploration plan; 2010 exploration drilling program; Posey blocks 6713, 6714, 6763, 6764, and 6912; Karo blocks 6864 and 7007; Burger, Crackerjack, and SW Shoebill prospects; OCS lease sale 193; Chukchi Sea, Alaska. Shell Gulf of Mexico Inc., Anchorage, AK. 1414 p. Accessed at http://www.mms.gov/alaska/ref/ProjectHistory/2009_Chukchi_Shell/2009_0811_Chukchi_EP_PublicCopy.pdf.
Shibaeva, T. 2007. Conservation of the Ladoga seal population. ECOS, Final Report for Second Rufford Small Grants for Nature Conservation. 27 p.
Shibaeva, T. 2009. Conservation of the Ladoga seal (Phoca hispida ladogensis) population, Russia. ECOS, Final Report for Second Rufford Small Grants for Nature Conservation, reporting period August 2008‐August 2009. 9 p.
Shiga, K., and I. Koizumi. 1999. Latest Quaternary oceanographic changes in the Okhotsk Sea based on diatom records. Marine Micropaleontology 38:91‐117.
Shpilev, H., E. Ojaveer, and A. Lankov. 2005. Smelt (Osmerus eperlanus L.) in the Baltic Sea. Biology and Ecology 54:230‐241.
Shults, L. M. 1977. Helminth parasites of ice‐inhabiting phocid seals in southeastern Bering Sea (Abstract). Page 82 in The Second Conference on the Biology of Marine Mammals, San Diego, CA. Society for Marine Mammalogy.
Shults, L. M. 1982. Helminths of the spotted seal, Phoca largha, from the Bering Sea. Journal of Wildlife Diseases 18:59‐62.
Shumei, N., K. Shigeru, I. Ko, U. Shotaro, T. Kenkichi, and T. Tadanori. 2001. A preliminary study for future shipping safety control regime in the ice‐infested waters of the Sea of Okhotsk. Papers of the Ship Research Institute 38:209‐225. (In Japanese, English Abstract).
Shustov, A. P. 1972. On the condition of the stocks and the distribution of true seals and walruses in the North Pacific (Abstract). Pages 146‐147 in V. A. Arsen'ev, editor. Fifth All‐Union Conference on

237
Studies of Marine Mammals, Makhachkala, USSR. Akad. Nauk. SSSR. (Translated from Russian by F.H. Fay, University of Alaska, Fairbanks, AK, 2 p.).
Siegstad, H., P. B. Neve, M. P. Heide‐Jørgensen, and T. Härkönen. 1998. Diet of the ringed seal (Phoca hispida) in Greenland. Pages 229‐241 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Simmonds, M. P., and S. J. Isaac. 2007. The impacts of climate change on marine mammals: early signs of significant problems. Oryx 41:19‐26.
Simpkins, M. A., B. P. Kelly, and D. Wartzok. 2001. Three‐dimensional diving behaviors of ringed seals (Phoca hispida). Marine Mammal Science 17:909‐925.
Sinisalo, T., R. I. Jones, E. Helle, and E. T. Valtonen. 2008. Changes in diets of individual Baltic ringed seals (Phoca hispida botnica) during their breeding season inferred from stable isotope analysis of multiple tissues. Marine Mammal Science 24:159‐170.
Sinisalo, T., M. Kunnasranta, and E. T. Valtonen. 2003. Intestinal helminths of a landlocked ringed seal (Phoca hispida saimensis) population in eastern Finland. Parasitology Research 91:40‐45.
Sinisalo, T., R. Poulin, H. Högmander, T. Juuti, and E. T. Valtonen. 2004. The impact of sexual selection on Corynosoma magdaleni (Acanthocephala) infrapopulations in Saimaa ringed seals (Phoca hispida saimensis). Parasitology 128:179‐185.
Sinisalo, T., E. T. Valtonen, E. Helle, and R. I. Jones. 2006. Combining stable isotopes and intestinal parasite information to evaluate dietary differences between individual ringed seals (Phoca hispida botnica). Canadian Journal of Zoology 84:823‐831.
Sipilä, T. 1990. Lair structure and breeding habitat of the Saimaa ringed seal (Phoca hispida saimensis Nordq.) in Finland. Finnish Game Research 47:11‐20.
Sipilä, T. 2003. Conservation biology of Saimaa ringed seal (Phoca hispida saimensis) with reference to other European seal populations. Ph.D. Dissertation. University of Helsinki, Helsinki, Finland. 40 p.
Sipilä, T. 2006. The past and future size of the Saimaa ringed seal population. Pages 158‐160 in Proceedings from the IVth International Symposium "Dynamics of game animal populations in Northern Europe", Petrozavodsk, Karelia, Russia.
Sipilä, T., E. Helle, and H. Hyvärinen. 1990. Distribution, population size and reproductivity of the Saimaa ringed seal (Phoca hispida saimensis Nordq.) in Finland, 1980‐84. Finnish Game Research 47:3‐10.
Sipilä, T., and H. Hyvärinen. 1998. Status and biology of Saimaa (Phoca hispida saimensis) and Ladoga (Phoca hispida ladogensis) ringed seals. Pages 83‐99 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.
Sipilä, T., and T. Kokkonen. 2008. Saimaannorppakannan tila vuonna 2008. 18 p. (In Finnish). Sipilä, T., J. Koskela, and H. Hyvärinen. 1999. Mortality of the Saimaa seal (Phoca hispida saimensis).
Page 174 in Thirteenth Biennial Conference on the Biology of Marine Mammals, Wailea, HI. Society for Marine Mammalogy.
Sipilä, T., N. Medvedev, M. Kunnasranta, V. Bogdanov, and H. Hyvärinen. 2002. Present status and recommended conservation actions for the Ladoga seal (Phoca hispida ladogensis) population. WWF Finland, Report No. 15. 30 p.
Sipilä, T., N. V. Medvedev, and H. Hyvärinen. 1996. The Ladoga seal (Phoca hispida ladogensis Nordq). Hydrobiologia 322:193‐198.
Skaare, J. U., E. Degre, P. E. Aspholm, and K. I. Ugland. 1994. Mercury and selenium in Arctic and coastal seals off the coast of Norway. Environmental Pollution 85:153‐160.
Skjoldal, H. R., H. Gjøsæter, and H. Loeng. 1992. The Barents Sea ecosystem in the 1980s: ocean climate, plankton, and capelin growth. ICES Marine Science Symposium 195:278‐290.

238
Skjoldal, H. R., A. Hassel, F. Rey, and H. Loeng. 1987. Spring phytoplankton development and zooplankton reproduction in the central Barents Sea in the period 1979‐1984. Pages 59‐89 in H. Loeng, editor. The Effect of Oceanographic Conditions on Distribution and Population Dynamics of Commercial Fish Stocks in the Barents Sea. Institute of Marine Research, Bergen, Norway.
Skjoldal, H. R., and F. Rey. 1989. Pelagic production and variability of the Barents Sea ecosystem. Pages 241‐186 in K. Sherman and L. M. Alexander, editors. Biomass Yields and Geography of Large Marine Ecosystems. American Association for the Advancement of Science, Boulder, CO.
Smiley, B. D., and A. R. Milne. 1979. LNG transport in Parry Channel: possible environmental hazards. Institute of Ocean Sciences. 47 p.
Smith, A. W., R. J. Brown, D. E. Skilling, H. L. Bray, and M. C. Keyes. 1977. Naturally‐occurring leptospirosis in northern fur seals (Callorhinus ursinus). Journal of Wildlife Diseases 13:144‐148.
Smith, A. W., D. E. Skilling, N. Cherry, J. H. Mead, and D. O. Matson. 1998. Calicivirus emergence from ocean reservoirs: zoonotic and interspecies movements. Emerging Infectious Diseases 4:13‐20.
Smith, R. E. H., J. F. Cavaletto, B. J. Eadie, and W. S. Gardner. 1993. Growth and lipid composition of high Arctic ice algae during the spring bloom at Resolute, Northwest Territories, Canada. Marine Ecology Progress Series 97:19‐29.
Smith, T. G. 1973. Population dynamics of the ringed seal in the Canadian eastern Arctic. Department of the Environment, Fisheries Research Board of Canada, Bulletin 181. 55 p.
Smith, T. G. 1975. Ringed seals in James Bay and Hudson Bay: population estimates and catch statistics. Arctic 28:170‐182.
Smith, T. G. 1976. Predation of ringed seal pups (Phoca hispida) by the Arctic fox (Alopex agopus). Canadian Journal of Zoology 54:1610‐1616.
Smith, T. G. 1977. The wolffish, cf. Anarhichas denticulatus, new to the Amundsen Gulf area, Northwest Territories, and a probable prey of the ringed seal. Canadian Field‐Naturalist 91:288.
Smith, T. G. 1980. Polar bear predation of ringed and bearded seals in the land‐fast sea ice habitat. Canadian Journal of Zoology 58:2201‐2209.
Smith, T. G. 1987. The ringed seal, Phoca hispida, of the Canadian western Arctic. Bulletin Fisheries Research Board of Canada. 81 p.
Smith, T. G., and F. A. J. Armstrong. 1975. Mercury in seals, terrestrial carnivores, and principal food items of the Inuit, from Holman, N.W.T. Journal of the Fisheries Research Board of Canada 32:795‐801.
Smith, T. G., and F. A. J. Armstrong. 1978. Mercury and selenium in ringed and bearded seal tissues from Arctic Canada. Arctic 31:75‐84.
Smith, T. G., B. Beck, and G. A. Sleno. 1973. Capture, handling, and branding of ringed seals. Journal of Wildlife Management 37:579‐583.
Smith, T. G., M. H. Hammill, D. W. Doidge, T. Cartier, and G. A. Selno. 1979. Marine mammal studies in southeastern Baffin Island. Final report to the Eastern Arctic Marine Environmental Studies (EAMES) project. Department of Fisheries and Ocean, Arctic Biological Station, Canadian Manuscript Report of Fisheries and Aquatic Sciences, No. 1552. 70 p.
Smith, T. G., and M. O. Hammill. 1980a. Ringed seal, Phoca hispida, breeding habitat survey of Bridport Inlet and adjacent coastal ice. Canadian Manuscript Report of Fisheries and Aquatic Sciences. 31 p.
Smith, T. G., and M. O. Hammill. 1980b. A survey of the breeding habitat of ringed seals and a study of their behavior during the spring haul‐out period in southeastern Baffin Island. Addendum to the Final Report to the Eastern Arctic Marine Environmental Studies (EAMES) project. Department of Fisheries and Oceans, Arctic Biological Station, Canadian Manuscript Report of Fisheries and Aquatic Sciences, No. 1561. 47 p.

239
Smith, T. G., and M. O. Hammill. 1981. Ecology of the ringed seal, Phoca hispida, in its fast ice breeding habitat. Canadian Journal of Zoology 59:966‐981.
Smith, T. G., M. O. Hammill, and G. Taugbøl. 1991. A review of the developmental, behavioural and physiological adaptations of the ringed seal, Phoca hispida, to life in the Arctic winter. Arctic 44:124‐131.
Smith, T. G., and L. A. Harwood. 2001. Observations of neonate ringed seals, Phoca hispida, after early break‐up of the sea ice in Prince Albert Sound, Northwest Territories, Canada, spring 1998. Polar Biology 24:215‐219.
Smith, T. G., and C. Lydersen. 1991. Availability of suitable land‐fast ice and predation as factors limiting ringed seal populations, Phoca hispida, in Svalbard. Polar Research 10:585‐594.
Smith, T. G., and I. Stirling. 1975. The breeding habitat of the ringed seal (Phoca hispida). The birth lair and associated structures. Canadian Journal of Zoology 53:1297‐1305.
Smith, T. G., and I. Stirling. 1978. Variation in the density of ringed seal (Phoca hispida) birth lairs in the Amundsen Gulf, Northwest Territories. Canadian Journal of Zoology 56:1066‐1070.
Snoeij, N., A. Penninks, and W. Seinen. 1988. Dibutyltin and tributyltin compounds induce thymus atrophy in rats due to a selective action on thymic lymphoblasts. International Journal of Immunopharmacology 10:891‐899.
Söderberg, S. 1971. Commercial damage and feeding habits of seals in the Baltic; a preliminary report. ICES, CM 1971/N:3. 6 p.
Söderberg, S. 1973. Seal observations in spring 1973. Fauna and Flora 5:182‐192. (Translated from Swedish by the Fisheries and Marine Service, Quebec, Canada, Translation Series Number 2999, 19 p.).
Söderberg, S. 1975. Sealhunting in Sweden. Pages 104‐116 in Proceedings from the Symposium on the Seal in the Baltic, June 4‐6, 1974, Lidingö, Sweden. National Swedish Environment Protection Board.
Söderberg, S. 1978. Falling age at sexual maturity in Baltic seals. Pages 27‐31 in T. Lampio and O. Stenman, editors. Proceedings from the Symposium on the Conservation of Baltic Seals April 26‐28, 1977, Haikko, Finland. Finnish Game and Fisheries Research Institute, Game Division.
Sonne‐Hansen, C., R. Dietz, P. Leifsson, L. Hyldstrup, and F. F. Rigét. 2002. Cadmium toxicity to ringed seals (Phoca hispida): an epidemiological study of possible cadmium‐induced nephropathy and osteodystrophy in ringed seals (Phoca hispida) from Qaanaaq in Northwest Greenland. Science of the Total Environment 295:167‐181.
Sonne, C., O. Aspholm, R. Dietz, S. Andersen, M. H. G. Berntssen, and K. Hylland. 2009. A study of metal concentrations and metallothionein binding capacity in liver, kidney and brain tissues of three Arctic seal species. Science of the Total Environment 407:6166‐6172.
Southall, B. L., A. E. Bowles, W. T. Ellison, J. J. Finneran, R. L. Gentry, C. R. Greene, Jr., D. Kastak, D. R. Ketten, J. H. Miller, P. E. Nachtigall, W. J. Richardson, J. A. Thomas, and P. L. Tyack. 2007. Marine mammal noise exposure criteria: initial scientific recommendations. Aquatic Mammals 33:411‐521.
Spraker, T. R., L. F. Lowry, and K. J. Frost. 1994. Gross necropsy and histopathological lesions found in harbor seals. Pages 281‐311 in T. R. Loughlin, editor. Marine Mammals and the Exxon Valdez. Academic Press, Inc., San Diego, CA.
St. Aubin, D. J. 1988. Physiological and toxicologic effects on pinnipeds. Pages 120‐142 in J. R. Geraci and D. J. St. Aubin, editors. Synthesis of Effects of Oil on Marine Mammals. U.S. Department of the Interior, Minerals Management Service, Atlantic OCS Region, New Orleans, LA.
St. Aubin, D. J. 1990. Physiological and toxic effects on pinnipeds. Pages 103‐127 in J. R. Geraci and D. J. St. Aubin, editors. Sea Mammals and Oil: Confronting the Risks. Academic Press, Inc., San Diego, CA.

240
Stamper, M. A., F. M. D. Gulland, and T. Spraker. 1998. Leptospirosis in rehabilitated Pacific harbor seals from California. Journal of Wildlife Diseases 34:407‐410.
Standing Committee on Fisheries and Oceans. 2007. Ensuring a sustainable and humane seal harvest. Parliamentary Information Research Service, Library of Parliament, Report of the Standing Committee on Fisheries and Oceans, 39th Parliament, 1st Session. 41 p.
Steele, M., W. Ermold, and J. L. Zhang. 2008. Arctic Ocean surface warming trends over the past 100 years. Geophysical Research Letters 35:L02614.
Steinacher, M., F. Joos, T. L. Frolicher, G. K. Plattner, and S. C. Doney. 2009. Imminent ocean acidification in the Arctic projected with the NCAR global coupled carbon cycle‐climate model. Biogeosciences 6:515‐533.
Stenman, O. 1977. Legislation, protection and research on seals in Finland. Finnish Game Research 37:8. Stenman, O., and O. Pöyhönen. 2005. Food remains in the alimentary tracts of the Baltic grey and ringed
seals (Abstract). Pages 51‐53 in E. Helle, O. Stenman, and M. Wikman, editors. Symposium on Biology and Management of Seals in the Baltic Area, 15‐18 February 2005, Helsinki, Finland. Finnish Game and Fisheries Research Institute.
Stewart, R. E. A., P. Richard, M. C. S. Kingsley, and J. J. Houston. 1986. Seals and sealing in Canada's Northern and Arctic regions. Western Region, Department of Fisheries and Oceans, Canadian Technical Report of Fisheries and Aquatic Sciences, No. 1463. 31 p.
Stirling, I. 1973. Vocalization in the ringed seal (Phoca hispida). Journal of the Fisheries Research Board of Canada 30:1592‐1594.
Stirling, I. 1974. Midsummer observations on the behavior of wild polar bears (Ursus maritimus). Canadian Journal of Zoology 52:1191‐1198.
Stirling, I. 1977. Adaptations of Weddell and ringed seals to exploit the polar fast ice habitat in the absence or presence of surface predators. Pages 741‐748 in Adaptations within Antarctic Ecosystems. Proceedings of the 3rd SCAR Symposium on Antarctic Biology. Smithsonian Institute, Washington, D.C.
Stirling, I. 1997. The importance of polynyas, ice edges, and leads to marine mammals and birds. Journal of Marine Systems 10:9‐21.
Stirling, I. 2002. Polar bears and seals in the eastern Beaufort Sea and Amundsen Gulf: a synthesis of population trends and ecological relationships over three decades. Arctic 55:59‐76.
Stirling, I., R. Archibald, and D. DeMaster. 1975. Distribution and abundance of seals in the eastern Beaufort Sea. Canadian Wildlife Service, Beaufort Sea Technical Report Number 1. 58 p.
Stirling, I., and W. R. Archibald. 1977. Aspects of predation of seals by polar bears. Journal of the Fisheries Research Board of Canada 34:1126‐1129.
Stirling, I., W. Calvert, and H. Cleator. 1983. Underwater vocalizations as a tool for studying the distribution and relative abundance of wintering pinnipeds in the High Arctic. Arctic 36:262‐274.
Stirling, I., and A. E. Derocher. 1993. Possible impacts of climatic warming on polar bears. Arctic 46:240‐245.
Stirling, I., M. Kingsley, and W. Calvert. 1982. The distribution and abundance of seals in the eastern Beaufort Sea, 1974‐79. Environment Canada, Canadian Wildlife Service, Edmonton, Canada. 25 p.
Stirling, I., and P. B. Latour. 1978. Comparative hunting abilities of polar bear cubs of different ages. Canadian Journal of Zoology 56:1768‐1772.
Stirling, I., and N. A. Øritsland. 1995. Relationships between estimates of ringed seal (Phoca hispida) and polar bear (Ursus maritimus) populations in the Canadian Arctic. Canadian Journal of Fisheries and Aquatic Sciences 52:2594‐2612.
Stirling, I., and C. L. Parkinson. 2006. Possible effects of climate warming on selected populations of polar bears (Ursus maritimus) in the Canadian Arctic. Arctic 59:261‐275.

241
Stirling, I., and T. G. Smith. 2004. Implications of warm temperatures, and an unusual rain event for the survival of ringed seals on the coast of southeastern Baffin Island. Arctic 57:59‐67.
Strassmann, K., G.‐K. Plattner, and F. Joos. 2009. CO2 and non‐CO2 radiative forcings in climate projections for twenty‐first century mitigation scenarios. Climate Dynamics 33:737‐749.
Stroeve, J., M. M. Holland, W. Meier, T. Scambos, and M. Serreze. 2007. Arctic sea ice decline: faster than forecast. Geophysical Research Letters 34:L09501.
Stroeve, J., M. Serreze, S. Drobot, S. Gearheard, M. M. Holland, J. Maslanik, W. Meier, and T. Scambos. 2008. Arctic sea ice extent plummets in 2007. EOS, Transactions of the American Geophysical Union 89:13‐14.
Sturm, M., J. Holmgren, and D. K. Perovich. 2002. Winter snow cover on the sea ice of the Arctic Ocean at the Surface Heat Budget of the Arctic Ocean (SHEBA): temporal evolution and spatial variability. Journal of Geophysical Research 107:8047.
Sturm, M., J. A. Maslanik, D. K. Perovich, J. C. Stroeve, J. Richter‐Menge, T. Markus, J. Holmgren, J. F. Heinrichs, and K. Tape. 2006. Snow depth and ice thickness measurements from the Beaufort and Chukchi seas collected during the AMSR‐Ice03 campaign. IEEE Transactions on Geoscience and Remote Sensing 44:3009‐3020.
Stutz, S. S. 1967. Pelage patterns and population distributions in the Pacific harbour seal (Phoca vitulina richardi). Journal of the Fisheries Research Board of Canada 24:451‐455.
Sugimoto, T., and K. Tadakoro. 1997. Interannual‐interdecadal variations in zooplankton biomass, chlorophyll concentration and physical environment in the subarctic Pacific and Bering Sea. Fisheries Oceanography 6:74‐93.
Sundby, S., and O. Nakken. 2008. Spatial shifts in spawning habitats of Arcto‐Norwegian cod related to multidecadal climate oscillations and climate change. ICES Journal of Marine Science 65:953‐962.
Supreme Court of the United States. 2007. Massachusetts v. Environmental Protection Agency (No. 05‐1120). Cornell University Law School, Accessed January 2010 at http://www.law.cornell.edu/supct/html/05‐1120.ZO.html.
Swanson, B. J., B. P. Kelly, C. K. Maddox, and J. R. Moran. 2006. Shed skin as a source of DNA for genotyping seals. Molecular Ecology Notes 6:1006‐1009.
Takahashi, C. 1999. Study on stomach nematodes (Anisakidae) among Steller sea lions, spotted seals and ribbon seals captured at northeast coast of Hokkaido, Japan. Japanese Journal of Veterinary Research 47:76‐77.
Tanabe, S., M. Prudente, T. Mizuno, J. Hasegawa, H. Iwata, and N. Miyazaki. 1998. Butyltin contamination in marine mammals from North Pacific and Asian coastal waters. Environmental Science & Technology 32:193‐198.
Taugbøl, G. 1982. Ringed seal thermoregulation, energy balance and development in early life, a study on Pusa hispida in Kongsfj., Svalbard. University of Oslo, Institute of Zoophysiology, Blindern, Norway. 102 p.
Taylor, C. C., H. B. Bigelow, and H. W. Graham. 1957. Climate trends and the distribution of marine animals in New England. Fishery Bulletin 57:293‐345.
Teilmann, J., E. W. Born, and M. Acquarone. 1999. Behaviour of ringed seals tagged with satellite transmitters in the North Water polynya during fast‐ice formation. Canadian Journal of Zoology 77:1934‐1946.
Teilmann, J., and F. O. Kapel. 1998. Exploitation of ringed seals (Phoca hispida) in Greenland. Pages 130‐151 in M. P. Heide‐Jørgensen and C. Lydersen, editors. Ringed Seals in the North Atlantic. NAMMCO Scientific Publications, Volume 1, Tromsø, Norway.

242
Thompson, D., C. D. Duck, and B. J. McConnell. 1998. Biology of seals of the north‐east Atlantic in relation to seismic surveys. Pages 4.1‐4.7 in M. L. Tasker and C. Weir, editors. Proceedings of the Seismic and Marine Mammals Workshop, London, UK.
Tikhomirov, E. A. 1961. Distribution and migration of seals in waters of the Far East. Pages 199‐210 in Conference on Pelagic Mammals, 1959, Moscow, Russia. Ichthyological Commission of the Academy of Sciences of the USSR. (Translated from Russian by L.V. Sagen, U.S. Fish and Wildlife Service, Marine Mammal Biological Laboratory, Seattle, WA, 26 p.).
Tikhomirov, E. A. 1968. Body growth and development of reproductive organs of the North Pacific phocids. Pages 213‐241 in V. A. Arsen'ev and K. I. Panin, editors. Pinnipeds of the North Pacific. Pischevaya Promyshlennost (Food Industry), Moscow, Russia. (Translated from Russian by J. Salkind, Israel Program for Science Translations, Jerusalem, Israel, 29 p.).
Timoshenko, Y. K. 1984. Sostoyaniye issledovaniy i perspectivy promysla kolchatoy nerpy v Belom, Barentsevom i Karskom moryakh [State research and perspectives of harvest of the ringed seal in the White, Barents and Kara seas]. Pages 104‐109 in Morskiye mlekopitayushiye. Biologicheskiye resursy gidrosfery i ikh ispolzovaniye [Marine Mammals. Biological resources of the hydrosphere and their uses]. M. Nauka Press, Moscow, Russia. (In Russian).
Tinz, B. 1996. On the relation between annual maximum extent of ice cover in the Baltic Sea and sea level pressure as well as air temperature field. Geophysica 32:319‐341.
Tormosov, D. D., and I. J. Filatov. 1979. Lodoga ringed seal ‐ the object of studies. Priroda 3:103‐107. (In Russian).
Tormosov, D. D., and G. V. Rezvov. 1978. Information on the distribution, number and feeding habits of ringed and grey seals in the Gulfs of Finland and Riga in the Baltic Sea. Pages 14‐17 in T. Lampio and O. Stenman, editors. Proceedings of the Symposium on the Conservation of Baltic Seals, 26‐28 April 1977, Haikko, Finland. Finnish Game and Fisheries Research Institute.
Trenberth, K. E., and J. W. Hurrell. 1995. Decadal coupled atmospheric‐ocean variations in the North Pacific Ocean. Pages 15‐24 in R. J. Beamish, editor. Climate Change and Northern Fish Populations. Canadian Special Publication in Fisheries and Aquatic Science 121.
Treshchev, V. V., and V. N. Popov. 1975. Helminths of the ringed seal in the southeastern part of the Barents Sea. Pages 131‐132 in G. B. Agarkov, editor. Proceedings of the Sixth All‐Union Conference, Kiev, USSR. Naukova Dumka. (Translated from Russian by the Canadian Translation of Fisheries and Aquatic Sciences, Ontario, Canada, No. 5600, 3 p. ).
Trippel, E. A., O. S. Kjesbu, and P. Solemdal. 1997. Effects of adult age and size structure on reproductive output in marine fishes. Pages 31‐63 in R. C. Chambers and E. A. Trippel, editors. Early Life History and Recruitment in Fish Populations. Chapman and Hall, New York, NY.
Troisi, G. M., K. Haraguchi, D. S. Kaydoo, M. Nyman, A. Aguilar, A. Borrell, U. Siebert, and C. F. Mason. 2001. Bioaccumulation of polychlorinated biphenyls (PCBs) and dichlorodiphenylethane (DDE) methyl sulfones in tissues of seal and dolphin morbillivirus epizootic victims. Journal of Toxicology and Environmental Health, Part A 62:1‐8.
Truett, J. C., M. E. Miller, and K. Kertell. 1997. Effects of Arctic Alaska oil development on brant and snow geese. Arctic 50:138‐146.
Trukhin, A. M. 2000. Ringed seal on the eastern coast of Sakhalin Island. Page 4 in V. M. Belkovich, A. N. Boltunov, and I. V. J. Smelova, editors. Marine Mammals of the Holarctic. 2000. Materials from the International Conference, Archangel, Russia. Pravda Severa. (Translated from Russian by Olga Romanenko, 4 p.).
Tryland, M., L. Kleivane, A. Alfredsson, M. Kjeld, A. Arnason, S. Stuen, and J. Godfroid. 1999. Evidence of Brucella infection in marine mammals in the North Atlantic Ocean. Veterinary Record 144:588‐592.

243
Tryland, M., E. Neuvonen, A. Huovilainen, H. Tapiovaara, A. Osterhaus, O. Wiig, and A. E. Derocher. 2005a. Serologic survey for selected virus infections in polar bears at Svalbard. Journal of Wildlife Diseases 41:310‐316.
Tryland, M., K. K. Sørensen, and J. Godfroid. 2005b. Prevalence of Brucella pinnipediae in healthy hooded seals (Cystophora cristata) from the North Atlantic Ocean and ringed seals (Phoca hispida) from Svalbard. Veterinary Microbiology 105:103‐111.
Tynan, C. T., and D. P. DeMaster. 1997. Observations and predictions of Arctic climatic change: potential effects on marine mammals. Arctic 50:308‐322.
Tyrrell, T., B. Schneider, A. Charalampopoulou, and U. Riebesell. 2008. Coccolithophores and calcite saturation state in the Baltic and Black Seas. Biogeosciences 5:485‐494.
U.S. Department of Interior. 2010. Fact sheet; A comprehensive, science‐based offshore energy plan. Department of Interior, Washington, D.C. 3 p. Accessed at http://www.doi.gov/deepwaterhorizon/loader.cfm?csModule=security/getfile&PageID=33566.
U.S. Department of the Interior. 2010. Fact sheet: A comprehensive, science‐based offshore energy plan. U.S. Department of Interior, Washington, D.C. 3 p. Accessed at http://www.doi.gov/deepwaterhorizon/loader.cfm?csModule=security/getfile&PageID=33566.
U.S. Environmental Protection Agency. 2009. Endangerment and cause or contribute findings for greenhouse gases under Section 202(a) of the Clean Air Act. Federal Register 74:239 (December 15, 2009) p. 66496‐66546.
U.S. Environmental Protection Agency. 2010a. Prevention of Significant Deterioration and title V greenhouse gas tailoring rule; Final Rule. Federal Register 75:106 (June 3, 2010) p. 31514‐31606.
U.S. Environmental Protection Agency. 2010b. Reconsideration of interpretation of regulations that determine pollutants covered by clean air act permitting programs; Final Action on Reconsideration of Interpretation. Federal Register 75:63 (April 2, 2010) p. 17004‐17023.
U.S. Environmental Protection Agency and U.S. Department of Transportation. 2010. Light‐duty vehicle greenhouse gas emission standards and corporate average fuel economy standards. Federal Register 75:88 (May 7, 2010) p. 25324‐25372.
U.S. Fish and Wildlife Service. 2008. Determination of threatened status for the polar bear (Ursus maritimus) throughout its range; Final Rule. Federal Register 73:95 (May 15, 2008) p. 28212‐28303.
U.S. Fish and Wildlife Service, and National Marine Fisheries Service. 1996. Policy regarding the recognition of distinct vertebrate population segments under the Endangered Species Act; Notice. Federal Register 61:26 (February 7, 1996) p. 4721‐4725.
U.S. Geological Survey. 2008. Circum‐Arctic resource appraisal: estimates of undiscovered oil and gas north of the Arctic Circle; Fact sheet 2008‐3049. U.S. Geological Survey, Menlo Park, CA. Accessed at http://pubs.usgs.gov/fs/2008/3049/fs2008‐3049.pdf.
Ukkonen, P. 1993. The post‐glacial history of the Finnish mammalian fauna. Annales Zoologici Fennici 30:249‐264.
Ukkonen, P. 2002. The early history of seals in the northern Baltic. Annales Zoologici Fennici 39:187‐207. United Nations Environment Programme. 2006. Sea of Okhotsk. Global International Waters Assessment
Regional Assessment 30, 53 p. University of Kalmar, Kalmar, Sweden. Urtans, E., N. Liskins, and V. Pilats. 2005. Seal monitoring in Latvia 1999‐2004 (Abstract). Page 69 in E.
Helle, O. Stenman, and M. Wikman, editors. Symposium on Biology and Management of Seals in the Baltic Area, 15‐18 February 2005 Helsinki, Finland. Finnish Game and Fisheries Research Institute.
Valtonen, E. T., H. P. Fagerholm, and E. Helle. 1988. Contracaecum osculatum (Nematoda: Anisakidae) in fish and seals in Bothnian Bay (northeastern Baltic Sea). International Journal for Parasitology 18:365‐370.

244
Van de Vijver, K. I., P. Hoff, K. Das, S. Brasseur, W. Van Dongen, E. Esmans, P. Reijnders, R. Blust, and W. De Coen. 2005. Tissue distribution of perfluorinated chemicals in harbor seals (Phoca vitulina) from the Dutch Wadden Sea. Environmental Science & Technology 39:6978‐6984.
Van Pelt, R. W., and R. A. Dieterich. 1973. Staphylococcal infection and toxoplasmosis in a young harbor seal. Journal of Wildlife Diseases 9:258‐261.
Van Vuuren, D. P., M. Meinshausen, G. K. Plattner, F. Joos, K. M. Strassmann, S. J. Smith, T. M. L. Wigley, S. C. B. Raper, K. Riahi, F. de la Chesnaye, M. G. J. den Elzen, J. Fujino, K. Jiang, N. Nakicenovic, S. Paltsev, and J. M. Reilly. 2008. Temperature increase of 21st century mitigation scenarios. Proceedings of the National Academy of Sciences 105:15258‐15262.
Verevkin, M. V., N. Medvedev, and T. Sipilä. 2006. Bycatch mortality of the Ladoga ringed seal. Bycatch Communication Network Newsletter:3‐4.
Vermeij, G. J., and P. D. Roopnarine. 2008. The coming Arctic invasion. Science 321:780‐781. Vibe, C. 1950. The marine mammals and the marine fauna in the Thule District (Northwest Greenland)
with observations on ice conditions in 1939‐41. Pages 1‐115 in Meddelelser om Grønland, Bind 150. C.A. Reitzels Forlag, Copenhagen, Denmark.
Vibe, C. 1967. Arctic animals in relation to climate fluctuations, the Danish zoogeographical investigations in Greenland. Meddelelser om Grønland, Bergen, Norway. 226 p.
Viberg, H., A. Fredriksson, and P. Eriksson. 2004. Investigations of strain and/or gender differences in developmental neurotoxic effects of polybrominated diphenyl ethers in mice. Toxicological Sciences 81:344‐353.
Vincent‐Chambellant, M. 2010. Ecology of ringed seals (Phoca hispida) in western Hudson Bay, Canada. Ph.D. Thesis. University of Manitoba, Winnipeg, Canada. 279 p.
Vinogradov, G. A., and V. T. Komov. 1985. Ion regulation in perch, Perca fluviatilis, in connection with the problem of acidification of water bodies. Journal of Ichthyology 25:63‐61.
von Storch, H., and A. Omstedt. 2008. Introduction and summary. Pages 1‐34 in H.‐J. Bolle, M. Menenti, and I. Rasool, editors. Assessment of Climate Change for the Baltic Sea Basin. Springer, Geesthacht, Germany.
Vuorinen, I., M. Hanninen, U. Helminen, and H. Kuosa. 1999. Proportion of copepod biomass declines together with increasing salinities of the Baltic Sea. ICES Journal of Marine Science 55:767‐774.
Wada, K., S. I. Hayama, T. Nakaoka, and H. Uno. 1991. Interactions between Kuril seals and salmon trap net fishery in the coastal waters of southeastern Hokkaido. Marine Mammal Science 7:75‐84.
Wagemann, R. 1985. Heavy metals in ringed seals from the Canadian Arctic. Department of Fisheries and Oceans, Prepared for the Department of Indian and Northern Development, Water Resources Division. 42 p.
Wagemann, R. 1989. Comparison of heavy metals in two groups of ringed seals (Phoca hispida) from the Canadian Arctic. Canadian Journal of Fisheries and Aquatic Sciences 46:1558‐1563.
Wagemann, R., S. Innes, and P. R. Richard. 1996. Overview and regional and temporal differences of heavy metals in Arctic whales and ringed seals in the Canadian Arctic. Science of the Total Environment 186:41‐66.
Wagemann, R., and D. C. G. Muir. 1981. Assessment of heavy metals and organochlorine concentrations in marine mammals of northern waters. International Council for the Exploration of the Sea, C.M. 1981/N:9. 102 p.
Walsh, J. E. 2008. Climate of the Arctic marine environment. Ecological Applications 18:S3‐S22. Walsh, J. E., O. Anisimov, J. O. M. Hagen, T. Jakobsson, J. Oerlemans, T. D. Prowse, V. Romanovsky, N.
Savelieva, M. Serreze, A. Shiklomanov, I. Shiklomanov, and S. Solomon. 2005. Section 6.2. Precipitation and evapotranspiration. Pages 184‐189 in Arctic Climate Impact Assessment. Cambridge University Press, Cambridge, UK.

245
Walsh, J. E., W. L. Chapman, V. Romanovsky, J. H. Christensen, and M. Stendel. 2008. Global climate model performance over Alaska and Greenland. Journal of Climate 21:6156‐6174.
Walters, V. 1955. Fishes of western Arctic America and eastern Arctic Siberia: taxonomy and zoogeography. Bulletin of the American Museum of Natural History 106:255‐368.
Wang, M., N. A. Bond, and J. E. Overland. 2007. Comparison of atmospheric forcing in four sub‐arctic seas. Deep‐Sea Research II 54:2543‐2559.
Wang, M. Y., and J. E. Overland. 2009. A sea ice free summer Arctic within 30 years? Geophysical Research Letters 36:1‐5.
Warren, S. G., I. G. Rigor, N. Untersteiner, V. F. Radionov, N. N. Bryazgin, Y. I. Aleksandrov, and R. Colony. 1999. Snow depth on Arctic sea ice. Journal of Climate 12:1814‐1829.
Wathne, J. A., T. Haug, and C. Lydersen. 2000. Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress Series 194:233‐239.
Weatherly, J. W., J. E. Walsh, and H. J. Zwally. 1991. Antarctic sea ice variations and seasonal air temperature relationships. Journal of Geophysical Research 96:15119‐15130.
Weeks, W. F. 2010. Snow. Pages 417‐426 in On Sea Ice. University of Alaska Press, Fairbanks, AK. Werner, I., and R. Gradinger. 2002. Under‐ice amphipods in the Greenland Sea and Fram Strait (Arctic):
environmental controls and seasonal patterns below the pack ice. Marine Biology 140:317‐326. Węsławski, J. M., M. Ryg, T. G. Smith, and N. A. Øritsland. 1994. Diet of ringed seals (Phoca hispida) in a
fjord of West Svalbard. Arctic 47:109‐114. Westerling, B., O. Stenman, and E. Rudbäck. 2005. Pathology of seals from the Finnish coastal waters,
Lake Saimaa and Lake Ladoga in the years 1982‐2004 (Abstract). Pages 76‐79 in E. Helle, O. Stenman, and M. Wikman, editors. Symposium on Biology and Management of Seals in the Baltic Area, 15‐18 February 2005, Helsinki, Finland. Finnish Game and Fisheries Research Institute.
Whitehead, P. J. P. 1985. FAO species catalogue. Vol. 7. Clupeoid fishes of the world (suborder Clupeoidei). An annotated and illustrated catalogue of the herrings, sardines, pilchards, sprats, shads, anchovies and wolf‐herrings. Part 1‐‐Chirocentridae, Clupeidae and Pristigasteridae. United Nations Development Programme, Food and Agriculture Organization of the United Nations, FAO Fisheries Synopsis No. 125. 303 p.
Wiberg, K., R. J. Letcher, C. D. Sandau, R. J. Norstrom, M. Tysklind, and T. F. Bidleman. 2000. The enantioselective bioaccumulation of chiral chlordane and alpha‐HCH contaminants in the polar bear food chain. Environmental Science & Technology 34:2668‐2674.
Wickins, J. F. 1984. The effect of hypercapnic sea water on growth and mineralization in penaeid prawns. Aquaculture 41:37‐48.
Wiig, Ø., A. E. Derocher, and S. E. Belikov. 1999. Ringed seal (Phoca hispida) breeding in the drifting pack ice of the Barents Sea. Marine Mammal Science 15:595‐598.
Willette, T. M., R. T. Cooney, V. Partrick, D. M. Mason, G. L. Thomas, and D. Scheel. 2001. Ecological processes influencing mortality of juvenile pink salmon (Oncorhynchus gorbuscha) in Prince William Sound, Alaska. Fisheries Oceanography 10:14‐41.
Wilson, S. C., G. Mo, and T. Sipilä. 2001. Legal protection for seals in small populations in European community and Mediterranean coastal waters. Mammalia 65:335‐348.
Wolfe, R., and L. B. Hutchinson‐Scarbrough. 1999. The subsistence harvest of harbor seal and sea lion by Alaska Natives in 1998. Alaska Department of Fish and Game, Division of Subsistence, Technical Paper No. 250.
Wolkers, J., I. C. Burkow, C. Lydersen, S. Dahle, M. Monshouwer, and R. F. Witkamp. 1998a. Congener specific PCB and polychlorinated camphene (toxaphene) levels in Svalbard ringed seals (Phoca

246
hispida) in relation to sex, age, condition and cytochrome P450 enzyme activity. Science of the Total Environment 216:1‐11.
Wolkers, J., I. C. Burkow, R. F. Witkamp, C. Lydersen, S. Dahle, and M. Monshouwer. 1998b. Cytochrome P450 mediated enzyme activities in relation to congener specific PCB accumulation in arctic seals. Organohalogen Compounds 37:421‐424.
Wood, C. M., and D. G. McDonald. 1997. Global warming: implications for freshwater and marine fish. Cambridge University Press, Cambridge, UK. 441 p.
WWF. 2010. Oil and gas in the Arctic: Barents Sea, Norway/Russia. World Wide Fund, Gland, Switzerland. Accessed June 2010 at http://wwf.panda.org/what_we_do/where_we_work/arctic/what_we_do/oil_gas/barents_oil_gas/.
Wyllie‐Echeverria, T., and W. S. Wooster. 1998. Year to‐year variations in Bering Sea ice cover and some consequences for fish distributions. Fisheries Oceanography 7:159‐170.
Yamada, Y., and T. Ikeda. 1999. Acute toxicity of lowered pH to some oceanic zooplankton. Plankton Biology and Ecology 46:62‐67.
Yamamoto‐Kawai, M., F. A. McLaughlin, E. C. Carmack, S. Nishino, and K. Shimada. 2009. Aragonite undersaturation in the Arctic Ocean: effects of ocean acidification and sea ice melt. Science 326:1098‐1100.
Yoshikazu, U. 2006. Review of catch records of marine mammals in Japanese waters (Poster Presentation). Pages 536‐538 in V. M. Belkovich, editor. Marine Mammals of the Holarctic 2006. Collection of Scientific Papers, Saint Petersburg, Russia. Marine Mammal Council.
Yurakhno, M. V. 1971. Helminths of commercial marine mammals of the Bering and Chukotsk seas. M.Sc. Dissertation. Crimean Pedagogical Institute, Simferopol, Crimea. 24 p. (Translated from Russian by M. Poppino, Davis, CA, 25 p.).
Yurakhno, M. V., and A. S. Skryabin. 1971. Parafilaroides krascheninnikovi sp.n., a parasite of the lungs of the ringed seal (Pusa hispida krascheninnikovi Naumov and Smirnov). Vestnik Zoologii 5:32‐36. (Translated from Russian by the Canadian Translations of Fisheries and Aquatic Sciences, Ottawa, Canada, No. 5563, 9 p.).
Zarnke, R. L., T. C. Harder, H. W. Vos, J. M. Ver Hoef, and A. D. M. E. Osterhaus. 1997. Serologic survey for phocid herpesvirus‐1 and ‐2 in marine mammals from Alaska and Russia. Journal of Wildlife Diseases 33:459‐465.
Zeisler, R., R. Demiralp, B. J. Koster, P. R. Becker, M. Burow, P. Ostapczuk, and S. A. Wise. 1993. Determination of inorganic constituents in marine mammal tissues. Science of the Total Environment 139/140:365‐386.
Zhang, X. D. 2010. Sensitivity of arctic summer sea ice coverage to global warming forcing: towards reducing uncertainty in arctic climate change projections. Tellus A 62:220‐227.
Zhang, X. D., and J. E. Walsh. 2006. Toward a seasonally ice‐covered Arctic Ocean: scenarios from the IPCC AR4 model simulations. Journal of Climate 19:1730‐1747.
Zheglov, V. A., and K. K. Chapskii. 1974. Test of an aerial survey of the ringed seal and the grey seal and of their holes in the gulfs of the Baltic Sea and on Lake Ladoga. Pages 323‐342 in K. K. Chapskii and E. S. Mil'chenko, editors. Research on Marine Mammals. Atlantic Research Institute of Marine Fisheries and Oceanography (AtlantNIRO), Russia. (Translated from Russian by Fisheries and Marine Service, Canada, Translation 3185, 36 p.).
Zhu, J. P., R. J. Norstrom, D. C. G. Muir, L. A. Ferron, L. P. Weber, and E. Dewailly. 1995. Persistent chlorinated cyclodiene compounds in ringed seal blubber, polar bear fat, and human plasma from Northern Quebec, Canada ‐ identification and concentrations of photoheptachlor. Environmental Science & Technology 29:267‐271.

247
Zwanenburg, K. C. T. 2000. The effects of fishing on demersal fish communities of the Scotian Shelf. ICES Journal of Marine Science 57:503‐509.

249
7 APPENDIX: GLOSSARY OF ABBREVIATIONS
Abbreviation Description ACIA Arctic Climate Impact Assessment ADFG Alaska Department of Fish and Game AMAP Arctic Monitoring and Assessment Programme AMSA Arctic Council's Arctic Marine Shipping Assessment AOGCM Atmosphere‐Ocean General Circulation Model AR4 IPCC's Fourth Assessment Report BRT biological review team BSAI Bering Sea‐Aleutian Islands BSPA Baltic Sea Protected Area BT butyltin CaCO3 calcium carbonate CAOR Central Arctic Ocean Route CBD Center for Biological Diversity CDV canine distemper virus CGCM3 Coupled Global Climate Model version 3 CI confidence interval CITES Convention on International Trade in Endangered Species of Wild Fauna and Flora CMIP3 Coupled Model Intercomparison Project Phase 3 CO2 carbon dioxide CRCM Canadian Regional Climate Model CYP cytochrome P450 CZMA U.S. Coastal Zone Management Act DDE dichloro‐diphenyl‐dichloroethylene DDT dichloro‐diphenyl‐trichloroethane DFO Canada’s Department of Fisheries and Oceans DPP Draft Proposed Program DPS distinct population segment EEZ exclusive economic zone EPA U.S. Environmental Protection Agency ESA U.S. Endangered Species Act EU European Union GHG greenhouse gas HadISST Hadley Centre sea ice and sea surface temperature data set HELCOM Helsinki Commission IMO International Maritime Organization IPCC Intergovernmental Panel on Climate Change IUCN International Union for Conservation the of Nature and Natural Resources MARPOL International Convention for the Prevention of Pollution from Ships MMPA U.S. Marine Mammal Protection Act MMS U.S. Minerals Management Service, now known as the Bureau of Ocean Energy
Management, Regulation, and Enforcement MPRSA U.S. Marine Protection, Research, and Sanctuaries Act mtDNA mitochondrial DNA

250
MY multi‐year NAMMCO North Atlantic Marine Mammal Commission NCAR National Center for Atmospheric Research NCEP NOAA’s National Centers for Environmental Prediction NEPA U.S. National Environmental Policy Act NMFS U.S. National Marine Fisheries Service NOAA U.S. National Oceanic and Atmospheric Administration NSR Northern Sea Route NWP Northwest Passage OC organochlorine OCS outer continental shelf OCSLA U.S. Outer Continental Shelf Lands Act PBDE polybrominated diphenyl ether PCB polychlorinated biphenyl PDV phocine distemper virus PFC perfluorinated contaminant PFDA perfluorodecanoic acid PFHxS perfluorohexane sulfonate PFNA perfluorononanoic acid PFOS perfluorooctane sulfonate PFOSA perfluorooctane sulfonamide PFUnA perfluoroundecanoic acid PhHV phocid herpesvirus POP persistent organic pollutant ppm parts per million PSD EPA’s Prevention of Significant Deterioration program PSSA Particularly Sensitive Sea Area SAT surface air temperature SCPOP Stockholm Convention on Persistent Organic Pollutants SD standard deviation SE standard error SRES IPCC's Special Report on Emissions Scenarios TAC total allowable catches TAR IPCC's Third Assessment Report TCV Tillamook calicivirus THg total mercury TINRO Russian Federation’s Pacific Research Fisheries Center USFWS U.S. Fish and Wildlife Service USGS U.S. Geological Survey

RECENT TECHNICAL MEMORANDUMS
Copies of this and other NOAA Technical Memorandums are available from the National Technical Information Service, 5285 Port Royal Road, Springfield, VA 22167 (web site: www.ntis.gov). Paper and electronic (.pdf) copies vary in price.
AFSC
211 CAMERON, M. F., J. L. BENGTSON, P. L. BOVENG, J. K. JANSEN, B. P. KELLY, S. P. DAHLE, E. A. LOGERWELL, J. E. OVERLAND, C. L. SABINE, G. T. WARING, and J. M. WILDER. 2010. Status review of the bearded seal (Erignathus barbatus), 246 p. NTIS number pending.
210 JOHNSON, S. W., J. F. THEDINGA, A. D. NEFF, and C. A. HOFFMAN. 2010. Fish fauna in nearshore waters of a barrier island in the western Beaufort Sea, Alaska, 28 p. NTIS No. PB2011102346. .
209 RONE, B. K., A. B. DOUGLAS, A. N. ZERBINI, L. MORSE, A. MARTINEZ, P. J. CLAPHAM, and J. CALAMBOKIDIS. 2010. Results from the April 2009 Gulf of Alaska line transect survey (GOALS) in the Navy training exercise area, 39 p. NTIS No. ADA522221.
208 VON SZALAY, P. G., N. W. RARING, F. R. SHAW, M. E. WILKINS, and M. H. MARTIN. 2010. Data Report: 2009 Gulf of Alaska bottom trawl survey, 245 p. NTIS No. PB2011102345.
207 PUNT, A. E., and P. R. WADE. 2010. Population status of the eastern North Pacific stock of gray whales in 2009, 43 p. NTIS No. PB2011102344.
206 ALLEN, B. M., and R. P. ANGLISS. 2010. Alaska marine mammal stock assessments, 2009, 276 p. NTIS No. PB2010-107408.
205 CAHALAN, J., J. MONDRAGON, and J. GASPER. 2010. Catch sampling and estimation in the Federal groundfish fisheries off Alaska, 42 p. NTIS No. PB2010-105918.
204 LAUTH, R. R. 2010. Results of the 2009 eastern Bering Sea continental shelf bottom trawl survey of groundfish and invertebrate resources, 228 p. NTIS No. PB2010-113178.
203 LAAKE, J., A. PUNT, R. HOBBS, M. FERGUSON, D. RUGH, and J. BREIWICK. 2009. Re-analysis of gray whale southbound migration surveys 1967-2006, 55 p. NTIS No. PB2010-103939.
202 FOWLER, C. W., and L. HOBBS. 2009. Are we asking the right questions in science and management?, 59 p. NTIS No. PB2010-105917.
201 CHILTON, E. A., C. E. ARMISTEAD, and R. J. FOY. 2009. The 2009 eastern Bering Sea continental shelf bottom trawl survey: Results for commercial crab species, 101 p. NTIS No. PB2010-103938.
200 BOVENG, P. L., J. L. BENGTSON, T. W. BUCKLEY, M. F. CAMERON, S. P. DAHLE, B. P. KELLY, B. A. MEGREY, J. E. OVERLAND, and N. J. WILLIAMSON. 2009. Status review of the spotted seal (Phoca largha), 153 p. NTIS No. PB2010-101436.
199 CONNERS, M. E., J. CAHALAN, S. GAICHAS, W. A. KARP, T. LOOMIS, and J. WATSON. 2009. Sampling for estimation of catch composition in Bering Sea trawl fisheries, 77 p. NTIS No. PB2010100895.
198 BARBEAUX, S. J., and D. FRASER. 2009. Aleutian Islands cooperative acoustic survey study for 2006, 90 p. NTIS No. PB2010-100894.
197 HOFF, G. R., and L. L. BRITT. 2009. Results of the 2008 eastern Bering Sea upper continental slope survey of groundfish and invertebrate resources, 294 p. NTIS No. PB2010-100893.
196 BUCKLEY, T. W., A. GREIG, and J. L. BOLDT. 2009. Describing summer pelagic habitat over the continental shelf in the eastern Bering Sea, 1982-2006, 49 p. NTIS No. PB2009-115399.





























