Embed Size (px)
Citation preview

Fitriyana et al. Statistik Demografi Diaphania indica 105J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 105 – 113, September 2015
STATISTIK DEMOGRAFI DIAPHANIA INDICA SAUNDERS(LEPIDOPTERA: CRAMBIDAE)
Ita Fitriyana1, Damayanti Buchori1, Ali Nurmansyah1, Rosichon Ubaidillah2, & Akhmad Rizali3
1Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper, Kampus IPB Dramaga, Bogor 16680
2Bidang Zoologi Pusat Penelitian LIPI, BogorJl. Raya Jakarta - Bogor Km.46 Cibinong, Bogor 16911
3Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas BrawijayaJl. Veteran, Malang 65145
E-mail: [email protected]
ABSTRACT
Demographic statistic of Diaphania indica Saunders (Lepidoptera: Crambidae). Diaphania indica is a pest of Cucumissativus L. The information about the biology of D. indica is needed to determine appropriate pest management strategies. Theaim of this study was to study the biology and life cycle of D. indica. The study was conducted by observing life stagedevelopment of D. indica, such as life cycle, head width and body length. The life table was determined by calculatingmortality of D. indica individuals and fecundity at each age. Mortality and fecundity data of D. indica were then used tocalculate the parameters of population growth including gross reproduction rate (GRR), net reproductive rate (Ro), meangeneration time (T) and and intrinsic rate of increase (r ). The results show that the life cycle of D. indica is about 25.7 days,while egg stadium = 3.2 days, larval stadium = 13.5 days, pupal stadium = 7.9 days, imago stadium = 8.6 days, and preovipositionperiod = 3.9 days. GRR of D. indica is 263 individuals per generation. Ro = 51 individuals per female per generation. T is 31.3days and the innate capacity for increase (r) = 0.1 individual per female per day.
Key words: Diaphania indica, life cycle, life table
ABSTRAK
Statistik demografi Diaphania indica Saunders (Lepidoptera: Crambidae). Diaphania indica adalah hama pada tanamanmentimun Cucumis sativus L. Informasi mengenai biologi dan siklus hidup diperlukan untuk strategi pengendalian hama.Tujuan dari penelitian ini adalah untuk mengetahui biologi dan siklus hidup D. indica. Pengamatan biologi terkait siklus hidupD. indica dilakukan dengan mengamati setiap tahap perkembangan hidup D. indica, meliputi waktu yang diperlukan danukuran tubuh pada setiap stadium. Penelitian neraca kehidupan dilakukan dengan mencatat mortalitas individu D. indica dankeperidian setiap individu imago setiap harinya. Data mortalitas dan keperidian D. indica dimasukkan ke dalam tabel neracakehidupan yang kemudian digunakan untuk menghitung parameter pertumbuhan populasi D. indica, meliputi laju reproduksikotor (GRR), laju reproduksi bersih (Ro), rataan masa generasi (T), dan laju pertumbuhan instrinsik (r). Hasil penelitianmenunjukkan bahwa siklus hidup D. indica pada tanaman mentimun memerlukan waktu 26,6 hari, dengan lama stadium telur3,2 hari; stadium larva 13,5 hari, stadium pupa 7,9 hari, stadium imago 8,6 hari dan masa praoviposisi imago 3,9 hari. GRR D.indica adalah 263 individu/generasi, Ro sebesar 51 individu/induk/generasi, T selama 31,3 hari, dan r sebesar 0,1 individu/induk/hari.
Kata kunci: Diaphania indica, neraca kehidupan, siklus hidup
PENDAHULUAN
Pengetahuan mengenai aspek-aspek demografimerupakan salah satu langkah awal dalam mempelajariperkembangan suatu populasi serangga. Demografiadalah analisis kuantitatif karakteristik suatu populasi,terutama hubungannya dengan pola pertumbuhanpopulasi, hubungan ketahanan, dan pergerakan populasi.Hal ini sangat erat hubungannya dengan dinamika
populasi namun penekanannya agak berbeda, demografilebih memusatkan pada pola perkembangan, kelahiran,kematian, dan pergerakan, sementara itu sebab danakibat dari fenomena ini dipelajari dalam dinamikapopulasi. Aspek demografi suatu populasi terdapat dalamneraca kehidupan. Neraca kehidupan merupakan teknikmenghitung angka kelahiran dan kematian suatupopulasi. Neraca tersebut adalah ringkasan pernyataantentang kehidupan individu populasi atau kelompok. Dari

106 J. HPT Tropika Vol. 15 No. 2, 2015: 105 - 113
data yang dihasilkan dapat dihitung berapa lama harapanhidup yang masih tersisa suatu individu (Price, 1984).Ada dua tipe neraca kehidupan yaitu yang bersifatspesifik umur atau neraca kehidupan horisontal dan yangbersifat spesifik waktu atau neraca kehidupan vertikal(Bellows et al., 1992). Neraca kehidupan horisontalmencakup penghitungan yang berulang terhadap suatukelompok (kohort) tunggal yang terdiri dari individu yangsama umurnya sepanjang waktu. Sedangkan neracakehidupan vertikal adalah data diambil pada suatukejadian tunggal ketika diasumsikan bahwa semuagenerasinya sudah saling lingkup dengan sempurna olehkarena kelas umur secara simultan adalah sama. Neracakehidupan merupakan riwayat perkembangan kohortyang bersifat dinamis (Tarumingkeng, 1992). Carey(1993) mengungkapkan bahwa neraca kehidupan yangdirancang dengan baik dapat meningkatkan pemahamanmengenai proses-proses yang terjadi di dalam suatupopulasi yang dapat digunakan untuk memprediksistruktur populasi dan pengaruh interaksi antar spesiesterhadappopulasi tersebut. Informasi biologi dan neracakehidupan diperlukan dalam menentukan strategipengendalian hama.
Salah satu hama yang berpotensi menimbulkankerusakan pada tanaman adalah hama ulat daunDiaphania indica Saunders (Lepidoptera: Crambidae).MacLeod (2005) menyatakan bahwa D. indicamerupakan salah satu hama pada pertanaman mentimundi Asia dan Afrika. Hama ini juga ditemukan menyerangmentimun di Indonesia (Asikin, 2004). Larva D. indicamemakan daun, batang muda yang lunak dan menggerekbuah. Kerusakan yang paling merugikan adalah jikalarva menyerang buah mentimun. Pada buah yangterserang ditemukan lubang pada permukaan buah,menyebabkan buah menjadi cepat busuk sehingga tidaklayak untuk dikonsumsi dan dijual (CABI, 2005). Dalamupaya pengendalian hama D. indica dibutuhkaninformasi dasar seperti informasi biologi dan neracakehidupan dari serangga tersebut. Pengetahuanmengenai berbagai aspek biologi yang diperlukan antaralain meliputi perilaku, siklus hidup, perkembangan,fisiologi, dan reproduksi (DeBach, 1973). Penelitian yangberkaitan dengan biologi dan statistika demografi dariD. indica perlu dilakukan untuk menunjang keberhasilandalam pengendalian hama ulat daun D. indica. Tujuandari penelitian ini adalah untuk mempelajari biologi danstatistika demografi D. indica di laboratorium. Penelitianini diharapkan dapat memberikan informasi biologi D.indica yang dapat digunakan sebagai input dalammerancang dan mengambil keputusan yang tepat dalammengendalikan D. indica.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Pengendalian Hayati, DepartemenProteksi Tanaman, Fakultas Pertanian IPB, Bogor.Penelitian ini dilaksanakan dari bulan Februari sampaidengan Juni 2014.
Pemeliharaan Masal Diaphania indica. SeranggaD. indica dikumpulkan dari lahan pertanaman mentimunmilik petani di Desa Cihideung Udik, KecamatanCiampea, Kabupaten Bogor. Larva atau pupa yangdidapat dari lapangan selanjutnya dipelihara dilaboratorium untuk dikembangbiakkan. Larva dipeliharadalam wadah plastik berukuran 14 x 9 x 7 cm. Larvayang telah menjadi pupa kemudian ditempatkan padawadah plastik (diameter = 3 cm, tinggi = 18 cm) sampaimenjadi imago. Pakan untuk imago berupa larutan madu20%. Imago dipelihara hingga menghasilkan telur danmati. Telur yang dihasilkan dipindahkan ke dalam tabungplastik (diameter = 9 cm, tinggi = 12 cm). Larva yangmuncul dipelihara hingga menjadi imago.
Pengamatan Siklus Hidup D. indica di Lapangan.Untuk mempelajari siklus hidup D. indica di lapangan,sebanyak 100 larva dipilih secara acak pada tanamanmentimun. Larva instar I diberi tanda dan diamati setiaphari. Pengamatan dilakukan dengan mencatat masih adaatau tidaknya larva, masih hidup atau sudah mati. Larvayang masih hidup difoto dengan kamera digital CasioEX-ZS5. Larva difoto setiap hari untuk mengetahuiukuran panjang tubuh dan lebar kepala larva. Penentuanstadia larva dilakukan dengan mempelajari ukuran larvayang diperoleh dari foto yang kemudian diukurmenggunakan program TPS DIG version 2. Hasildigitasi kemudian dikonversi dengan Microsoft Excel2010 menjadi ukuran panjang tubuh dan lebar kepala.Untuk penentuan stadia instar larva digunakanpengukuran lebar kepala yang kemudian di analisisdengan distribusi frekuensi berdasarkan metode yangdigunakan Chen & Seybold (2013). Jumlah larva yanghidup, dan mati diamati dan dicatat perkembangannyasetiap hari hingga menjadi imago dan menghasilkan telur.Lama stadia larva, pupa dan imago, panjang dan lebarukuran pupa, serta jenis kelamin imago yang munculjuga dicatat.
Pengamatan Kohort D. indica. Kohort merupakankelompok individu yang lahir dalam interval waktu yanghampir sama (Begon et al., 2008). Pengamatan kohortD. indica dilakukan dalam empat tahap yang disesuaikan

Fitriyana et al. Statistik Demografi Diaphania indica 107
dengan fase perkembangan D. indica, yaitu fase telur,larva, pupa dan imago.
Pengamatan Stadia Telur D. indica. Telur D.indica yang digunakan sebagai populasi awal kohortberasal dari peletakan telur oleh imago betina hasilperbanyakan. Pengamatan dimulai denganmempersiapkan 100 telur yang berasal dari lima ekorbetina ke dalam toples plastik. Telur-telur diinkubasihingga menetas. Telur yang digunakan merupakan teluryang diletakkan pada hari yang sama. Pengamatandilakukan setiap hari dengan mencatat jumlah larva instarI yang muncul dari toples plastik. Larva instar I yangmuncul kemudian dipindahkan dan dipelihara untukpengamatan lama stadia larva. Pengamatan stadia telurmencakup lama stadia telur, jumlah telur yang diletakkan,jumlah telur yang menetas dan persentase penetasan.Persentase penetasan dihitung dengan menggunakanrumus:
Pengamatan Lama Stadia Larva D. indica.Larva instar I yang digunakan untuk pengujian inimerupakan larva yang muncul dari 100 telur yangdigunakan pada awal pengamatan. Larva yang munculdimasukkan kedalam wadah plastik berukuran 14 x 9 x7 cm yang berisi pakan daun mentimun. Pengamatanlama stadia larva dilakukan dengan mengamati jumlahlarva yang hidup dan yang mati. Perkembangan larvadicatat setiap hari sampai larva menjadi pupa.Pengamatan tahap ini memberikan data lama stadialarva, distribusi umur instar dan laju kesintasan larva.
Pengamatan Lama Stadia Pupa D. indica.Pengamatan lama stadia pupa dilakukan sejak pupaterbentuk sampai pupa menjadi imago. Pupa yangterbentuk dimasukkan ke dalam wadah plastik (diamater= 3 cm, tinggi = 18 cm). Pengamatan ini menghasilkandata lama stadia pupa.
Pengamatan Imago D. indica. Tahap terakhirdari pengamatan kohort adalah pengamatan imago.Imago yang muncul dari pupa pada pengamatansebelumnya dipelihara dalam wadah plastik yang berisikapas yang telah dibasahi dengan larutan madu 20%sebagai pakan D. indica dan daun mentimun sebagaimedia peletakan telur imago D. indica. Satu ekor imagobetina yang muncul kemudian dipaparkan dengan satuekor imago jantan. Pemaparan dilakukan agar imagojantan dan betina dapat berkopulasi sehinggamenghasilkan telur. Pemaparan dilakukan setiap hari dandihentikan ketika seluruh imago mati. Pengamatan inimenghasilkan data siklus dan lama hidup imago jantan
100%urjumlah telTotal
menetasyangurJumlah telPenetasan%
dan betina, periode praoviposisi, umur betina saat pertamakali meletakkan telur, periode oviposisi, dan fekunditas.
Analisis Data. Data mengenai kemampuan hidup danfekunditas disusun dalam bentuk tabel neraca kehidupan(life table). Parameter demografi yang dihitungmenurut Birch (1948) dalam Kurniawan (2007) meliputi:
1. Laju produksi kotor (GRR) =
2. Laju produksi bersih (Ro) =
3. Laju pertumbuhan instrinsik (rm) =
dengan rawal
=(ln Ro) / T
4. Rataan masa generasi (T) =
5. Populasi berlipat ganda (DT) = ln(2)/T
Statistik neraca kehidupan kemudian disusunmenggunakan metode Jackknife. Jackknife adalahmetode pengambilan contoh ulang (resampling) yangdigunakan untuk estimasi bias dan menduga standardeviasi (Quenouille, 1949). Prinsip dasar dari metodeJackknife terletak pada perhitungan suatu statistik secaraberulang dengan mengeluarkan satu atau lebihpengamatan dari suatu sampel (n) yang ditetapkan,sehingga menghasilkan sampel terpisah yang masing-masing memiliki besar ukuran n-1 atau n-d, (d < n).Pengambilan ulang sampel dalam penelitian ini dilakukansebanyak 50 kali.
HASIL DAN PEMBAHASAN
Siklus Hidup Diaphania indica. Siklus hidupD. indica terdiri dari empat fase yaitu telur, larva, pupadan imago. Larva D. indica terdiri dari lima instar (instarI, II, III, IV dan V). Stadia dan ukuran masing-masingfase perkembangan Diaphania indica dapat dilihatpada Tabel 1.
Telur D. indica diletakkan secara berkelompokdi bawah permukaan daun mentimun (Gambar 1). Telurmemiliki rata rata panjang 0,7 mm dan lebar 0,4 mm.Telur menetas setelah diletakkan 1 sampai 3 hari. Telurberbentuk bulat pipih. Pada hari pertama diletakkan telurberwarna kuning transparan, kemudian warna telursecara perlahan berubah menjadi kuning pekat yangmenandakan telur akan segera menetas.
Hasil analisis distribusi frekuensi ukuran lebarkepala menunjukkan bahwa larva D. indica mengalamilima instar yakni instar I hingga instar V. Tubuh larvaterdiri dari 3 ruas toraks dengan 2 pasang spirakel dan
xm
xmxl
1 rmxexmxl
xmxlxmxXl

108 J. HPT Tropika Vol. 15 No. 2, 2015: 105 - 113
Gambar 1.Telur Diaphania indica di bawah mikroskop (A) dan di bawah permukaan daun (B)
10 ruas abdomen dengan 8 pasang spirakel. Larvamemakan daun, batang yang bertekstur lunak dan buahmentimun. Tubuh larva instar I berwarna hijaukekuningan dengan kepala berwarna hijau muda(Gambar 2A). Ukuran rata-rata panjang tubuh 3,3 mmdan lebar kepala 0,3 mm. Larva instar I yang barumuncul tidak langsung aktif bergerak tetapi bersembunyiterlebih dahulu di antara tulang daun. Setelah dua harilarva instar I berubah warna menjadi kehijauan dan larvamulai aktif bergerak dan memakan bagian tengah menujubagian tepi daun. Lama stadia larva instar I adalah 2sampai 4 hari.
Tubuh larva instar II memiliki warna kehijauanyang lebih pekat dibandingkan dengan larva instar I(Gambar 2B). Pada larva instar II mulai muncul garis
putih pada bagian dorsal larva. Ukuran rata-rata panjangtubuh larva instar II adalah 5,7 mm dengan rata-ratalebar kepala 0,4 mm. Pada fase ini larva sudah lebihaktif bergerak dibandingkan dengan larva instar I . Lamastadia larva instar II adalah 2 sampai 5 hari.
Larva instar III dan IV berwarna hijau dengandua garis berwarna putih di sepanjang tubuh bagiandorsal, dan kepala berwarna hijau (Gambar 3A dan 3B).Larva instar III memiliki panjang 8,1 mm dengan lebarkepala 0,7 mm. Larva instar IV memiliki ukuran rata-rata panjang 10,5 mm dengan rata-rata lebar kepala 0,9mm. Pada kedua fase ini larva lebih aktif makan daundan batang lunak. Larva instar III dan IV memiliki lamastadia yang sama yaitu berkisar 2 sampai 4 hari.
Tabel 1. Rata-rata lama stadia, panjang tubuh dan lebar kepala larva Diaphania indica
FaseLama stadia (hari)
Selang (n) Rata-rata ± galat
Telur 3 – 4 (100) 3,2 ± 0,9Larva Instar 1 2 – 5 (100) 3,4 ± 0,7Larva Instar 2 2 – 4 (93) 2,5 ± 0,7Larva Instar 3 2 – 4 (79) 2,7 ± 0,6Larva Instar 4 2 – 5 (68) 2,4 ± 1,3Larva Instar 5 1 – 5 (68) 2,6 ± 1,4Pupa 6 – 12(62) 7,8 ± 1,4Jantan :
Lama hidup 5 – 13 (27) 8,6 ± 2,3Betina :
Lama hidup 8 – 14 (29) 10,6 ± 1,8Siklus hidup 23 – 32 (29) 25,7 ± 3,1Umur pertama meletakkan telur 26 – 35 (29) 29,6 ± 3,0Masa praoviposisi 2 – 6 (29) 3,9 ± 0,8Masa oviposisi 4 – 10 (29) 6,5 ± 1,6Fekunditas 37 – 309 (29) 103,3 ± 67,0

Fitriyana et al. Statistik Demografi Diaphania indica 109
A B
Larva instar V berwarna hijau dengan dua garisputih sepanjang tubuh yang semakin jelas dan kepalaberwarna hijau kekuningan. Panjang tubuh dan lebarkepala larva instar V adalah 12,0 mm dan 1,4 mm. Padafase ini larva mulai memasuki masa prapupa yangberlangsung 1 sampai 2 hari. Menjelang masa prapupatubuh larva berubah warna menjadi lebih pucat. Masaprapupa ditandai dengan menurunnya aktivitas dan
kemampuan makan. Larva mulai menyelubungitubuhnya dengan menjalin benang-benang yangdihasilkan dari mulutnya (Gambar 4). Biasanya larvatersebut melipat daun dan kemudian bersembunyi didalam lipatan daun tersebut. Saat fase prapupa larvaakan mengalami penyusutan ukuran tubuh sehinggaterlihat lebar dan gemuk. Lama stadia larva instar Vadalah 1 sampai 5 hari.
Gambar 4. Larva Diaphania indica instar V yang akan memasuki periode prapupa
Gambar 2. Larva Diaphania indica instar I (A) dan instar II (B)
A B
Gambar 3. Larva Diaphania indica instar III (A) dan instar IV (B)
A B

110 J. HPT Tropika Vol. 15 No. 2, 2015: 105 - 113
Pupa berwarna coklat mengkilap. Tipe pupa D.indica adalah obtekta. Bagian bakal antena, alat mulut,bakal sayap dan bakal tungkai dapat dilihat dengan jelas.Rata-rata panjang tubuh pupa 11,3 mm dan lebar tubuhpupa 2,5 mm (Gambar 5). Perbedaan pupa jantan danbetina dapat dilihat dari karakteristik morfologinya. Pupajantan memiliki bentuk ujung abdomen kerucut danmemiliki pola warna gelap terang yang jelas antar ruasabdomen, sedangkan pupa betina memiliki bentuk ujungabdomen yang lebih ramping serta pola warna gelapterang antar ruas abdomen yang kurang jelas (Gambar5). Selain itu, pada pupa betina terdapat celah padasternum ruas abdomen terakhir. Pupa yang akan menjadiimago ditandai dengan perubahan warna dari coklatmenjadi kehitaman. Stadia pupa berkisar antara 6 sampai12 hari dan biasanya terdapat dalam gulungan daun.
Imago yang baru terbentuk berwarna putihkekuningan (krem). Sayap berwarna putih dengan pitaberwarna coklat gelap di sepanjang pinggiran sayap.Warna sayap antara jantan dan betina hampir sama.
Gambar 5. Pupa Diaphania indica jantan (A) dan betina (B)
Imago jantan dan betina dapat dibedakan melaluimorfologi abdomennya. Pada abdomen imago betinaterdapat rumbai (bulu-bulu halus) berwarna kuningdengan jumlah yang lebih banyak, dan ukuran abdomenyang lebih besar dibandingkan dengan imago jantan(Gambar 6). Panjang larva umumnya berkaitan denganjenis kelamin dari imago D. indica. Larva D. indicadengan panjang 12,8–15,0 mm umumnya menjadi imagobetina, sedangkan larva dengan panjang 11,0–12,7 mmmenjadi imago jantan. Ukuran lebar rentang sayap jantanadalah 12,6 mm dan sayap betina 13,3 mm. Lama stadiajantan 6 sampai 13 hari, sedangkan lama stadia betina 8sampai 14 hari.
Masa Praoviposisi, Oviposisi dan FekunditasD. indica. Masa praoviposisi betina D. indica berkisarantara 2 sampai 6 hari. Telur diletakkan berkelompok disepanjang permukaan bawah tulang daun. Banyaknyatelur dalam satu helai daun dapat mencapai 71 butir telur/hari. Imago dapat bertelur hingga 6 hari. Selama
A B
Gambar 6. Imago Diaphania indica betina (A) dan jantan (B)
A B
Fitriyana et al. Statistik Demografi Diaphania indica 111
hidupnya imago betina mampu menghasilkan 309 butirtelur.
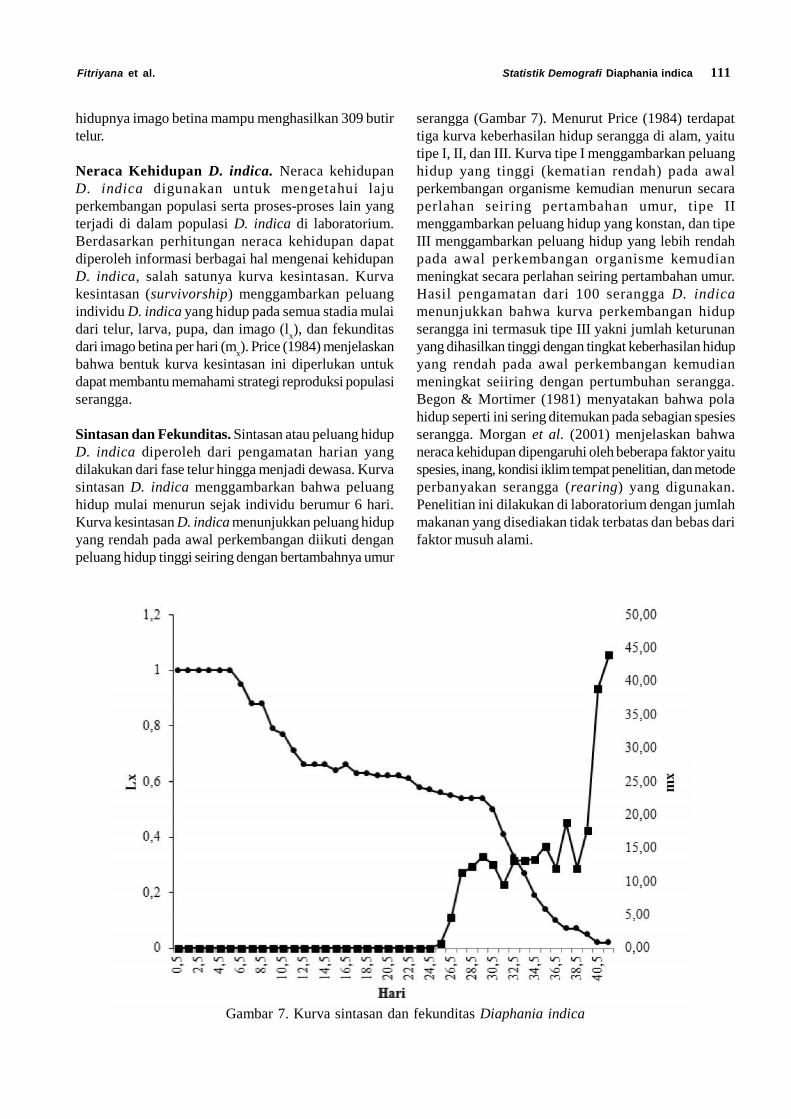
Neraca Kehidupan D. indica. Neraca kehidupanD. indica digunakan untuk mengetahui lajuperkembangan populasi serta proses-proses lain yangterjadi di dalam populasi D. indica di laboratorium.Berdasarkan perhitungan neraca kehidupan dapatdiperoleh informasi berbagai hal mengenai kehidupanD. indica, salah satunya kurva kesintasan. Kurvakesintasan (survivorship) menggambarkan peluangindividu D. indica yang hidup pada semua stadia mulaidari telur, larva, pupa, dan imago (l
x), dan fekunditas
dari imago betina per hari (mx). Price (1984) menjelaskan
bahwa bentuk kurva kesintasan ini diperlukan untukdapat membantu memahami strategi reproduksi populasiserangga.
Sintasan dan Fekunditas. Sintasan atau peluang hidupD. indica diperoleh dari pengamatan harian yangdilakukan dari fase telur hingga menjadi dewasa. Kurvasintasan D. indica menggambarkan bahwa peluanghidup mulai menurun sejak individu berumur 6 hari.Kurva kesintasan D. indica menunjukkan peluang hidupyang rendah pada awal perkembangan diikuti denganpeluang hidup tinggi seiring dengan bertambahnya umur
Gambar 7. Kurva sintasan dan fekunditas Diaphania indica
serangga (Gambar 7). Menurut Price (1984) terdapattiga kurva keberhasilan hidup serangga di alam, yaitutipe I, II, dan III. Kurva tipe I menggambarkan peluanghidup yang tinggi (kematian rendah) pada awalperkembangan organisme kemudian menurun secaraperlahan seiring pertambahan umur, tipe IImenggambarkan peluang hidup yang konstan, dan tipeIII menggambarkan peluang hidup yang lebih rendahpada awal perkembangan organisme kemudianmeningkat secara perlahan seiring pertambahan umur.Hasil pengamatan dari 100 serangga D. indicamenunjukkan bahwa kurva perkembangan hidupserangga ini termasuk tipe III yakni jumlah keturunanyang dihasilkan tinggi dengan tingkat keberhasilan hidupyang rendah pada awal perkembangan kemudianmeningkat seiiring dengan pertumbuhan serangga.Begon & Mortimer (1981) menyatakan bahwa polahidup seperti ini sering ditemukan pada sebagian spesiesserangga. Morgan et al. (2001) menjelaskan bahwaneraca kehidupan dipengaruhi oleh beberapa faktor yaituspesies, inang, kondisi iklim tempat penelitian, dan metodeperbanyakan serangga (rearing) yang digunakan.Penelitian ini dilakukan di laboratorium dengan jumlahmakanan yang disediakan tidak terbatas dan bebas darifaktor musuh alami.

112 J. HPT Tropika Vol. 15 No. 2, 2015: 105 - 113
Fekunditas imago betina dapat diketahui dengancara menghitung banyaknya telur yang diletakkan setiapharinya. Nilai mx menunjukkan banyaknya telur yangdihasilkan oleh imago betina yang berumur x hari setelahmemperhitungkan nisbah kelamin. Dari kurva juga dapatdilihat bahwa peneluran terjadi setelah hari ke 26 atau 3hari setelah D. indica menjadi imago. Velasco & Walter(1993) menyatakan bahwa keberhasilan perkembanganserangga dan fase reproduktif serangga sangatdipengaruhi oleh kualitas makanan.
Parameter demografi D. indica yang meliputi lajureproduksi kotor (GRR), laju reproduksi bersih (Ro), lajupertambahan instrinsik (r
m), rataan lama generasi (T),
dan waktu berlipat ganda (doubling time) (DT) dapatdilihat pada Tabel 2.
Laju reproduksi kotor (GRR) menggambarkanrataan jumlah keturunan betina per induk yang dihasilkanoleh individu D. indica yang hidupnya mencapai umurmaksimal. Nilai GRR D. indica adalah 263menunjukkan bahwa D. indica mampu menghasilkanketurunan sebesar 263 individu/induk/generasi.
Laju reproduksi bersih (Ro) D. indicamenunjukkan bahwa rataan banyaknya keturunan betinayang dihasilkan oleh seekor induk betina adalah 51,8betina/induk/generasi atau dapat dikatakan bahwapopulasi D. indica dapat berlipat sebanyak 52 kali dalamsetiap generasinya. Menurut Price (1997) suatu populasiakan bertambah jika memiliki nilai Ro > 1 dan stabil jikaRo = 1. Tingginya angka pertumbuhan serangga apabilaberada pada kondisi lingkungan yang optimum dapatmenyebabkan serangga tersebut dapat berkembangdengan cepat menjadi hama yang menyebabkankerugian secara ekonomi, seperti kehilangan hasil dankematian tanaman. Nilai Ro dan GRR menunjukkantingkat kesesuaian antara serangga dengan tanamaninang, semakin tinggi nilai Ro dan GRR menunjukkansemakin tinggi tingkat kesesuaian serangga dan tanamaninang, sebaliknya nilai Ro dan GRR yang rendahmenunjukkan tingkat kesesuaian serangga dan tanamaninang yang rendah pula.
Nilai rm
menggambarkan laju pertambahaninstrinsik pada keadaan sumber daya yang tidak terbatasdan dihitung dengan asumsi bahwa populasi memiliki
nilai lx dan m
x yang tetap serta kematian hanya terjadi
oleh faktor fisiologi (Price 1997). Nilai rm yang tinggi
dapat diartikan bahwa individu akan banyak mengalamikematian pada populasi dilapangan. Nilai r
mD. indica
adalah sebesar 0,1. Brewer (1979) menjelaskan bahwatinggi rendahnya nilai r dipengaruhi oleh jumlah keturunanper periode perkembangan, jumlah yang bertahan hidupdan selama masa reproduktif, usia saat reproduktifdimulai dan lama usia reproduktif.
Nilai rata-rata masa generasi (T) menggambarkanwaktu yang dibutuhkan sejak telur diletakkan sampaisaat imago betina yang berasal dari telur tersebutmenghasilkan keturunannya. Nilai T yang semakin kecilmenunjukkan semakin cepat suatu organisme untukberkembang biak. Nilai T dari D. indica sebesar 31,3menunjukkan bahwa dalam waktu 31,3 hari individubetina D. indica mampu menghasilkan keturunankembali. Berdasarkan nilai tersebut dapat diperkirakanbahwa dalam 1 tahun D. indica mampu menghasilkan12 generasi semakin kecil nilai T maka semakin cepatwaktu suatu individu untuk berkembang biak.
Doubling time atau waktu yang dibutuhkan untukpopulasi D.indica menjadi berlipat ganda (DT) adalah5,5 hari. Nilai DT yang tinggi dapat menyebabkanmeningkatnya laju reproduksi kotor (GRR) dan nilai lajureproduksi bersih (Ro) dalam satuan waktu tertentu.Menurut Birch (1948) dalam Kurniawan (2007), nilaiberlipat ganda yang tinggi pada suatu populasi, dapatmenyebabkan penurunan sumber daya lingkungan danmempengaruhi nilai laju pertambahan intrinsik (r
m).
Harcout (1969) menyatakan bahwa neracakehidupan bukanlah nilai akhir dari analisis dinamikapopulasi, tetapi sekedar penampilan sistematik dari dataketahanan hidup, mortalitas dan fekunditas dalampopulasi. Fungsi utama dari data ini adalah untukmemberikan informasi tentang gambaran kemampuanhidup suatu serangga pada kondisi lingkungan tertentu.Keseluruhan hasil pengamatan demografi dari D. indicamengungkapkan bahwa D. indica merupakan seranggadengan kemampuan perkembangan populasi yang tinggidan laju pertumbuhan yang cepat, hal ini dapat dilihatdari tingginya nilai laju reproduksi, doubling time, rataangenerasi serta pola kesintasan dari D. indica.
Tabel 2. Statistik demografi Diaphania indica di laboratorium
No. Parameter Rataan ± galat
1. Laju reproduksi kotor (GRR) 263,3 ± 2,52. Laju reproduksi bersih (Ro) 51,8 ± 0,93. Laju pertumbuhan intrinsik (rm) 0,1 ± 0,04. Rataan lama generasi (T) (hari) 31,3 ± 0,15. Doubling time (DT) (hari) 5,5 ± 0,0

Fitriyana et al. Statistik Demografi Diaphania indica 113
SIMPULAN
Siklus hidup Diaphania indica pada tanamanmentimun memerlukan waktu 25,7 hari, dengan kurvakeberhasilan hidup tipe III dimana pada pola ini jumlahketurunan yang dihasilkan tinggi dengan tingkatkeberhasilan hidup yang rendah pada awalperkembangan kemudian meningkat seiring denganpertumbuhan serangga. Neraca kehidupan D. indicamenunjukkan bahwa D. indica merupakan seranggayang memiliki tingkat perkembangan yang cepat, tingkatbertahan yang tinggi dan kapasitas reproduksi yangsangat besar, sehingga D. indica dapat berpotensimenjadi hama primer dan menyebabkan kerusakan yangsignifikan dalam jumlah besar pada tanaman ekonomipenting, apabila tidak dilakukan penanganan secara tepatyang dilaksanakan sesuai waktunya.
SANWACANA
Penelitian ini dibiayai oleh Direktorat PendidikanTinggi (Dikti) melalui Hibah Kompetensi, InstitutPertanian Bogor tahun 2013-2014. Terima kasih kepadaAmanda Mawan, S.P., M.Si. dan Ibu Adha Sari, S.P.atas bantuan analisis dan teknis, serta kepada BapakDarmawan atas bantuan identifikasi serangga parasitoidsehingga penelitian ini dapat berjalan dengan lancar.
DAFTAR PUSTAKA
Asikin S. 2004. Alternatif pengendalian hama seranggasayuran ramah lingkungan di lahan lebak.Laporan Tahunan Balai Penelitian PertanianLahan Rawa (Balitra). Banjarbaru.
Begon M & Montimer M. 1981. Population Ecology:a Unified Study of Animals and Plants .Sunderland Sinauer Associated, Massachussetts.
Begon M, Townsend CR, & Harper JL. 2008. Ecology:From Individuals to Ecosystems. 4th edition.Blackwell Publishing, Oxford.
Bellows Jr TS, Van Driesche RG, & Elkinton JS. 1992.Life-table construction in the evaluation of naturalenemies. Annu. Rev. Entomol. 37: 587–612.
Brewer R. 1979. Principles of Ecology. W.B SoundersCo, Philadelphia.
[CABI] Centre for Agriculture and BioscienceInternational. 2005. Corp ProtectionCompendium 2005 [CD-ROM]. CABI.Wallingford.
Carey JR. 1993. Applied Demography for Biologistwith Special Emphasis on Insect. pp. 11–41.Oxford University Press, New York.
Chen Y & Seybold SJ. 2013. Application of a frequencydistribution method for determining instars of thebeet armyworm (Lepidoptera: Noctuidae) fromwidths of cast head capsules. J. Econ. Entomol.106(2): 800–806.
DeBach P. 1973. The scope biological control. In:DeBach P (Ed). Biological Control of InsectPest and Weeds. pp. 3–20. Chapman and HallLtd., London.
Harcourt DG. 1969. The development and use of lifetables in the study of natural insect population.Annu. Rev. Entomol. 14: 175–196.
Kurniawan HA. 2007. Neraca kehidupan kutukebul,Bemisia tabaci Gennadius (Hemiptera:Aleyrodidae) biotipe-B dan non-B pada tanamanmentimun (Cucumis sativus L.) dan cabai(Capsicum annuum L.). Tesis. Sekolah PascaSarjana, Institut Pertanian Bogor, Bogor.
MacLeod A. 2005. Pest Risk Analysis for Diaphaniaindica. Central Science Laboratory, Sand HuttonYork.
Morgan D, Walters KFA, & Aegerter JN. 2001. Effectof temperature and cultivar on pea aphid,Acyrthosiphon pisum (Hemiptera: Aphididae)life history. Bull. Entomol. Res. 91(1): 47–52.
Price PW. 1997. Insect Ecology. 3th ed. John Wiley &Sons, New York.
Price PW. 1984. Insect Ecology. 2nded. John Wiley &Sons, New York.
Quenouille MH. 1949. Approximate tests of correlationin time series. J. R. Stat. Soc. B11(1): 68–84.
Tarumingkeng RC. 1992. Dinamika PertumbuhanPopulasi Serangga. Institut Pertanian Bogor,Bogor.
Velasco LRI & Walter GH. 1993. Potential of hostswitching in Nezara viridula (Hemiptera:Pentatomidae) to enhance survival andreproduction. Environ. Entomol. 22(2): 326–333.

114 J. HPT Tropika Vol. 15 No. 2, 2015: 114 - 121J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 114 – 121, September 2015
EKSPRESI GEN PROTEIN SELUBUNG TOMATO INFECTIOUSCHLOROSIS VIRUS PADA ESCHERICHIA COLI
Fitrianingrum Kurniawati, Gede Suastika, & Giyanto
Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper, Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Expression of tomato infectious chlorosis virus coat protein gene on Escherichia coli. Tomato infectious chlorosis virus(TICV) is the causal agent of chlorotic disease of tomato. Detection of TICV can be carried out by RT-PCR and serological test.Titer of TICV in plant tissue is very low because TICV is limited to phloem. Serological detection of TICV requires antiserumwhich is not available in Indonesia. Producing antibody through cloning and coat protein gene (TICV CP gene) expression isa promising approach in producing antiserum. The objective of this study was to express TICV CP gene as antigen forantiserum production. TICV CP gene was amplified using RT-PCR from total RNA extracted from TICV infected leaves collectedfrom Cipanas, Cianjur, West Java. The amplified CP gene was then sequenced and sub-cloned into pET 21b expression vector,transformed into Escherichia coli strain BL21 DE3(pLysS) and induced expression using IPTG 1 mM overnight at 37 °C. CPthat contains 6xhistag was purified using NiNTA spin column and then confirmed by SDS-PAGE. The size of TICV CP genewas 750 bp and the gene was expressed on pET 21 b vector and SDS-PAGE showed a 29 kDa band.
Key words: coat protein gene, expression, tomato infectious chlorosis virus (TICV)
ABSTRAK
Ekspresi gen protein selubung tomato infectious chlorosis virus pada Escherichia coli. Penyakit klorosis pada tanamantomat disebabkan oleh Tomato infectious chlorosis virus (TICV). Deteksi TICV dapat dilakukan dengan transkripsi balik (RT-PCR) dan serologi. Titer TICV dalam jaringan tanaman sangat rendah karena terbatas pada jaringan floem. Deteksi serologimembutuhkan antiserum. Di Indonesia antiserum TICV ini belum tersedia. Usaha penyediaan antigen melalui kloning danekspresi gen protein selubung (CP) merupakan salah satu cara yang menjanjikan dalam produksi antiserum. Penelitian inibertujuan untuk mengekspresikan gen CP TICV sebagai antigen dalam produksi antiserum. Gen CP diamplifikasi dengantranskripsi balik (RT-PCR) dari RNA total yang diekstraksi dari daun yang terinfeksi yang berasal dari Cipanas, KabupatenCianjur, Jawa Barat. Gen CP hasil amplifikasi kemudian di sekuensing, disubkloning ke dalam vektor ekspresi pET 21b,ditransformasi ke dalam bakteri Escherichia coli strain BL21 DE3(pLysS) dan diekspresikan dengan diinduksi menggunakanIPTG 1 mM selama semalaman pada suhu 37 °C. Purifikasi CP yang mengandung 6xhistag dengan menggunakan NiNTA spincolumn dan hasilnya dikonfirmasi dengan sodium deodecyl sulphate polyacrilamide gel electrophoresis (SDS-PAGE). Hasilamplifikasi gen CP berhasil memperoleh fragmen gen CP yang berukuran 750 pb dan diekspresikan pada vektor pET 21b.Purifikasi CP yang mengandung 6xhistag menggunakan NiNTA spin column berhasil memperoleh pita CP yang berukuransekitar 29 kDa.
Kata kunci: ekspresi, gen protein selubung, tomato infectious chlorosis virus (TICV)
PENDAHULUAN
Tomato infectious chlorosis virus (TICV)merupakan salah satu virus yang menyebabkan penyakitklorosis pada tanaman tomat. Di Indonesia, penyakit initelah ditemukan di beberapa sentra produksi tomat,seperti di Garut, Cipanas-Cianjur, Bogor, Magelang, danYogyakarta (Sa’adah, 2013; Fitriasari, 2010; Hartono& Wijonarko, 2007). TICV merupakan anggota dari
genus Crinivirus (famili Closteroviridae) (Jacquemondet al., 2009).
Gejala serangan TICV pada tanaman tomatditunjukkan oleh klorosis pada bagian antara tulang daun.Jika gejala klorosis sangat parah, daun akan mengalaminekrotik (kematian jaringan) dan menjadi rapuh, sertaukuran buah menjadi lebih kecil, mudah gugur dan prosespemasakan terganggu sehingga hasil panen menurun(Wisler et al., 1998). TICV ditularkan dari satu tanaman

Kurniawati et al. Ekspresi Gen Protein Selubung 115
ke tanaman lainnya oleh serangga vektor Trialeurodesvaporariorum (Hemiptera: Aleyrodidae) yang dikenaldengan nama kutu kebul rumah kaca secarasemipersisten (Duffus et al., 1996).
Deteksi TICV dapat dilakukan dengan reversetranscription–polymerase chain reaction (RT-PCR)dan hasilnya sangat cepat dan akurat, tetapi deteksitersebut memerlukan bahan-bahan dan peralatan yangmahal. Teknik deteksi selain RT-PCR adalah deteksidengan pendekatan serologi yaitu enzyme linkedimmunosorbent assay (ELISA), tissue blotimmunosorbent assay (TBIA), dan dot blotimmunosorbent assay (DIBA). Deteksi dengan metodeini cepat dan murah. Deteksi secara serologi inimemerlukan antiserum. Antiserum TICV belum tersediadi Indonesia, sehingga menyulitkan dalam mendeteksivirus ini. Usaha ke arah penyediaan antiserum akansangat bermanfaat sebagai sarana dalam mendeteksiTICV. Titer TICV pada tanaman sangat rendah karenavirus ini hanya ditemukan pada jaringan floem.
Ekspresi gen coat protein (CP) TICV padaEscherichia coli menjanjikan tersedianya antigen dalamjumlah yang cukup untuk produksi antiserum.Keunggulan penyediaan antiserum dengan metode iniantara lain: protein yang dihasilkan bersifat spesifiksehingga tidak bereaksi terhadap protein tanaman,antigen dapat tersedia dalam jumlah yang mencukupisetiap saat apabila diperlukan untuk produksi antiserum(Cotillon et al., 2005). Melihat keunggulan di atas makadiperlukan suatu metode untuk dapat mengekspresikangen CP-TICV yang akan digunakan sebagai antigendalam produksi antiserum. Penelitian ini bertujuan untukmengekspresikan gen protein selubung (CP) TICV padaE. coli. Manfaat dari penelitian ini berupa proteinselubung TICV murni yang dapat digunakan sebagaiantigen pada tubuh mamalia (kelinci) sehingga terbentukantibodi terhadap TICV yang kemudian dapat digunakansebagai sarana deteksi virus yang bersangkutan.
METODE PENELITIAN
Tempat dan Waktu. Penelitian ini dilakukan diLaboratorium Virologi Tumbuhan, Departemen ProteksiTanaman, Fakultas Pertanian IPB dan LaboratoriumBiokimia Balai Besar Bioteknologi dan SumberdayaGenetik Pertanian dari bulan Mei 2012 sampai denganAgustus 2013.
Tanaman Sumber Virus. Untuk mendapatkan TICVisolat Indonesia dilakukan pengumpulan tanamanbergejala klorosis dari sentra produksi tomat di daerah
Cipanas, Kabupaten Cianjur, Provinsi Jawa Barat (1225m dpl). Daun tanaman yang terinfeksi TICV diamatidan difoto gejalanya, kemudian diambil untukdiidentifikasi dengan menggunakan transkripsi balik (RT-PCR).
Amplifikasi Gen CP TICVEkstraksi RNA Total. Ekstraksi RNA total
dilakukan dengan menggunakan Qiagen RNAeasy PlantMini Kit dan dikerjakan sesuai dengan protokol yangdiberikan (Qiagen, 2003).
Sintesis complementary (c) DNA. RNA hasilekstraksi digunakan sebagai cetakan dalam sintesiscDNA melalui reaksi RT mengikuti metode Fitriasari(2010). Siapan cDNA hasil RT digunakan sebagaicetakan dalam reaksi PCR.
PCR. PCR dilakukan dalam volume 25 µl yangterdiri dari 12,5 µl Go Tag Green PCR Master mix(Thermo Scientific, US), masing-masing 1 µl primer(TICV–CP F-Bam no ATG (5’-A A T T A A G G A T C C G A A A ACTTATCTGGTAATGCAAAC-3’ dan TICV–CP R-Hind 5’-AATTAAAAGCTTTTAGCATGGGTGTTTCATATC AGCC-3’), 8,5 µl ddH
2O dan 2 µl cDNA.
PCR dilakukan pada Automated Thermal cycler (GeneAmp PCR System 9700; PE Applied Biosystem, USA).Proses ini didahului dengan denaturasi awal pada 94 ºCselama 4 menit, dilanjutkan dengan 30 siklus yang terdiridari denaturasi pada 94 ºC selama 1 menit, penempelanprimer (annealing) pada 55 ºC selama 1 menit, danpemanjangan (extension) pada 72 ºC selama 2 menit,dan diikuti pemanjangan akhir pada 72 ºC selama 10menit.
Elusi Produk PCR. Produk PCRdielektroforesis dalam gel agarose 1%. Produk PCRdan penanda DNA 1kb (Thermo Scientific, US), masing-masing 10 µl dan 5 µl dimasukkan ke dalam sumuranyang telah disiapkan pada gel agarose. Elektroforesisdilakukan selama 30 menit pada 100 V. Produk PCRyang telah dielektroforesis kemudian divisualisasi denganUV transluminator. Bagian gel tepat pada produk PCRdipotong dan dimasukkan ke tabung mikro yang sudahditambahkan 500 µl buffer DF (HiYield, China), kemudiandivortex, dan diinkubasi pada suhu 55 ºC sampai gel cair.Siapan kemudian dimasukkan ke DF kolom dandisentrifugasi dengan kecepatan 8000 rpm selama 1menit. Selanjutnya, 500 µl wash buffer (HiYield, China)dimasukkan dalam DF kolom dan disentrifugasi dengankecepatan 8000 rpm selama 1 menit. Sebanyak 20 µlbuffer elusi (HiYield, China) ditambahkan pada tengahkolom dan disentrifugasi pada kecepatan 8000 rpm

116 J. HPT Tropika Vol. 15 No. 2, 2015: 114 - 121
selama 1 menit. Hasil elusi (yang merupakan gen CP-TICV) disimpan pada suhu -20 ºC sampai akandigunakan.
Penyisipan Gen CP TICV pada Plasmid VektorpET-21b
Pemotongan Plasmid Vektor pET-21b.Plasmid pET-21b dipotong dengan enzim restriksiBamH1 dan HindIII. Rektan untuk pemotongan plasmiddilakukan dengan total volume 20 µl yang terdiri dari 1µl plasmid pET-21b, 1 µl enzim BamH1, 1 µl enzimHindIII, 2 µ l buffer, dan 15 µ l ddH2O. Reaksipemotongan diinkubasi pada suhu 37 ºC selama satumalam.
Ligasi Plasmid dan Insert (Gen CP TICV).Gen CP TICV disisipkan ke dalam plasmid pET-21bmelalui reaksi ligasi mengikuti prosedur (Qiagen, US).Campuran ligasi dengan total volume 10 µl terdiri dari 2µl plasmid pET-21b, 2 µl hasil elusi gen CP TICV, 1 µlenzim T4 ligase, 1 µl buffer ligasi, dan 4 µl ddH
2O. Reaksi
ligasi dilakukan pada suhu 4 ºC selama 16 jam. Hasilligasi akan menghasilkan plasmid pET-21b yangmempunyai insert gen CP TICV, yang selanjutnyadisebut plasmid pET-CP TICV.
Persiapan Kompeten Sel E. coli strainBL21(DE3)pLysS. Stok E. coli strain BL21(DE3)pLysS dalam gliserol digores pada media LB agar yangmengandung antibiotik ampisilin 50 µ g/ml dankloramfenikol 20 µg/ml, kemudian diinkubasi pada suhu37 °C selama satu malam. Satu kultur biakan E. coliselama satu malam dipindahkan ke dalam 5 ml LuriaBertani (LB) broth yang mengandung antibiotikkemudian diinkubasi pada suhu 37 °C selama satumalam. Sekitar 2 ml kultur selama satu malam tersebutdiambil dan dipindahkan ke dalam 40 ml media A (LB,MgSO
47H
2O 10 mM, gukosa 0,2%), lalu diinkubasi
selama 2 jam. Setelah 2 jam kultur bakteri dipindahkanke tabung falcon steril dan diinkubasi dalam es batuselama 10 menit. Biakan yang berada pada tabung falcondisentrifugasi dengan kecepatan 4000 rpm, selama 15menit pada suhu 4 °C. Supernatan dibuang dan diambilpeletnya. Pelet diresuspensi dengan 2,5 ml media B (LB,glyserol 36%, PEG 7000 12%, dan MgSO
47H
2O 12
mM). Siapan bakteri tersebut kemudiandipindahpisahkan masing-masing 100 µl ke tabungeppendorf 1,5 ml dan disimpan di freezer -80 °C sampaidigunakan untuk transformasi (Nishimura et al., 2003).Transformasi. Transformasi dilakukan denganmencampur 10 µl hasil ligasi dengan 100 µl sel kompetenBL21(DE3)pLySs. Terhadap siapan ini berturut-turutdilakukan diinkubasi dalam es batu selama 20 menit,
heat shock pada suhu 42 °C selama 1 menit, laludipindahkan ke dalam es batu selama 2 menit,ditambahkan 500 µl LB cair, dan diinkubasi dengandishaker pada suhu 37 °C selama 4 jam, kemudiandisentrifus 12.000 rpm 1 menit, supernatan dibuang, peletdan LB yang masih tersisa sampai 100 µl dihomogenasi,dan ditumbuhkan dalam media LB agar 20 ml yang telahdiberi ampisilin 50 µg/ml dan kloramfenikol 20 µg/ml.Biakan tersebut diinkubasi pada suhu 37 °C selama satumalam (Sambrook & Russel, 2001).
Konfirmasi Transforman. Transforman yangmembawa plasmid pET-CP TICV terlebih dahuluditumbuhkan pada 5 ml media LB cair yang mengandung50 mg/ml ampisilin dan 20 mg/ml kloramfenikol padasuhu 37 °C selama satu malam, kemudian plasmiddiisolasi dengan metode alkalin lisis (Sambrook &Russel, 2001). Konfirmasi dilakukan dengan memotongplasmid pET-CP TICV dan koloni PCR.
Isolasi Plasmid dengan Metode Alkalin Lisis. Satukoloni bakteri diinokulasi ke dalam 10 ml LB dandiinkubasi selama satu malam pada suhu 37 °C, kemudiandipindahkan ke tabung eppendorf 1,5 ml dandisentrifugasi 12.000 rpm pada suhu 4 °C selama 2menit. Setelah itu supernatan dibuang. Resuspensi peletdengan 100 µl larutan I (50 mM glukosa, 10 mM EDTA,25 mM Tris HCl pH 8, 2 mg/ml lisozyme (ditambahkansaat akan digunakan)), kemudian divortex, ditambahkan200 µl larutan II (0,2 M NaOH, 1% SDS dibuat saatakan digunakan), divortex, ditambahkan 150 µl larutanIII (3 M NaOAc pH 4,8), tabungnya dibolak-balik agarbercampur, disentrifus 12.000 rpm selama 5 menit.Supernatan dipindahkan ke tabung eppendorf baru,ditambahkan 300 µl phenol kloroform, divortex, dandisentrifus 12.000 rpm 5 menit. Larutan yang paling atasdipindahkan ke tabung yang baru, kemudian ditambahkanetanol absolut 2 x volume, sentrifus 12.000 rpm selama20 menit, supernatan dibuang, dicuci dengan 800 µletanol 70%, vortex, disentrifus 12.000 rpm 5 menit,supernatan dibuang, pelet dikeringanginkan, kemudiandiresuspensi dengan 30 µl buffer TE atau ddH
2O
(Sambrook & Russel, 2001).
Pemotongan pET-CP TICV dengan EnzimRestriksi. Plasmid pET-CP TICV dipotong denganenzim restriksi BamH1 dan HindIII prosedur sepertipada restriksi yang telah diuraikan sebelumnya. Hasilrestriksi dielektroforesis pada gel agarose 1% selama30 menit dengan tegangan 100 V. Hasil elektroforesisdivisualisasikan dengan transilluminator ultraviolet dandidokumentasikan dengan kamera digital.

Kurniawati et al. Ekspresi Gen Protein Selubung 117
Mengkulturkan E. coli Transforman untukPurifikasi Protein CP-TICV. E. coli strain BL21(DE3)pLysS yang membawa plasmid rekombinanpET21-CP diinokulasi ke dalam 3 ml media LB yangmengandung 50 mg/ml ampisilin dan 20 mg/mlkloramfenikol. Biakan diinkubasi di dalam orbitalshaker (75 rpm) pada suhu 37 oC selama satu malam.Kemudian sebanyak 100 ml biakan diinokulasikan kedalam 10 ml media LB yang mengandung antibiotik dandiinkubasikan di dalam orbital shaker (75 rpm) padasuhu 37 oC. Setelah pertumbuhan bakteri mencapaiOD
600 0,5 (kira-kira 5-6 jam), biakan diinduksi dengan 1
mM isopropil-thio-D-galaktoside (IPTG) dandiinkubasikan semalaman. Sel bakteri yangmengekpresikan protein rekombinan dipanen dengansentrifugasi (12.000 rpm, 4 oC, 15 menit) dan dilisisdengan buffer B-7M urea 300 µl dan turbonuclease(Nacalai, Japan) 3 unit/ml kultur (0,4 µl). Sel hasil lisisdisentrifugasi dengan kecepatan 12.000 rpm pada suhu4 oC selama 15 menit. Protein rekombinan yang adadalam supernatan (soluble protein) dan pelet (insolubleprotein) dianalisis dengan analisis SDS-PAGE.
Purifikasi CP-TICV. Purifikasi CP-TICV dilakukandengan Qiagen NiNTA spin column pada kondisidenaturasi dan dikerjakan sesuai dengan protokol yangdiberikan (Qiagen, 2008). Setelah diperoleh proteinmurni, dilakukan analisis SDS-PAGE (Laemmli, 1970).Elektroforesis dilakukan dengan Biorad power pac 300selama 2 jam dengan voltase 150 V. Kemudian gel hasilelektroforesis dilepas dari cetakannya dan dimasukkanke dalam larutan staining (coomassie brilliant blue R-250, metanol, asam asetat glasial) dan dishaker selamasatu malam, setelah itu dicuci dengan larutan distaining(metanol, asam asetat glasial, aquadest) sampai gelterlihat bening dan pita protein terlihat berwarna biruselanjutnya hasilnya difoto.
HASIL DAN PEMBAHASAN
TICV Isolat Indonesia. Pengumpulan tanaman yangbergejala klorosis yang dilakukan di daerah Cipanas,Kabupaten Cianjur, Provinsi Jawa Barat berhasilmendapatkan tanaman tomat yang terinfeksi TICV(Gambar 1). Gejala penyakit klorosis pada tanamantomat ditunjukkan oleh klorosis pada bagian tulang daun(interveinal yellowing) yang dimulai pada daunterbawah, kemudian berkembang cepat secara meratake daun-daun bagian atas. Serangan klorosis yang parahakan menyebabkan nekrotik pada daun (Gambar 1B).Hal ini menyebabkan proses fotosintesis terganggu danmengakibatkan penurunan produksi (Wisler et al., 1998).
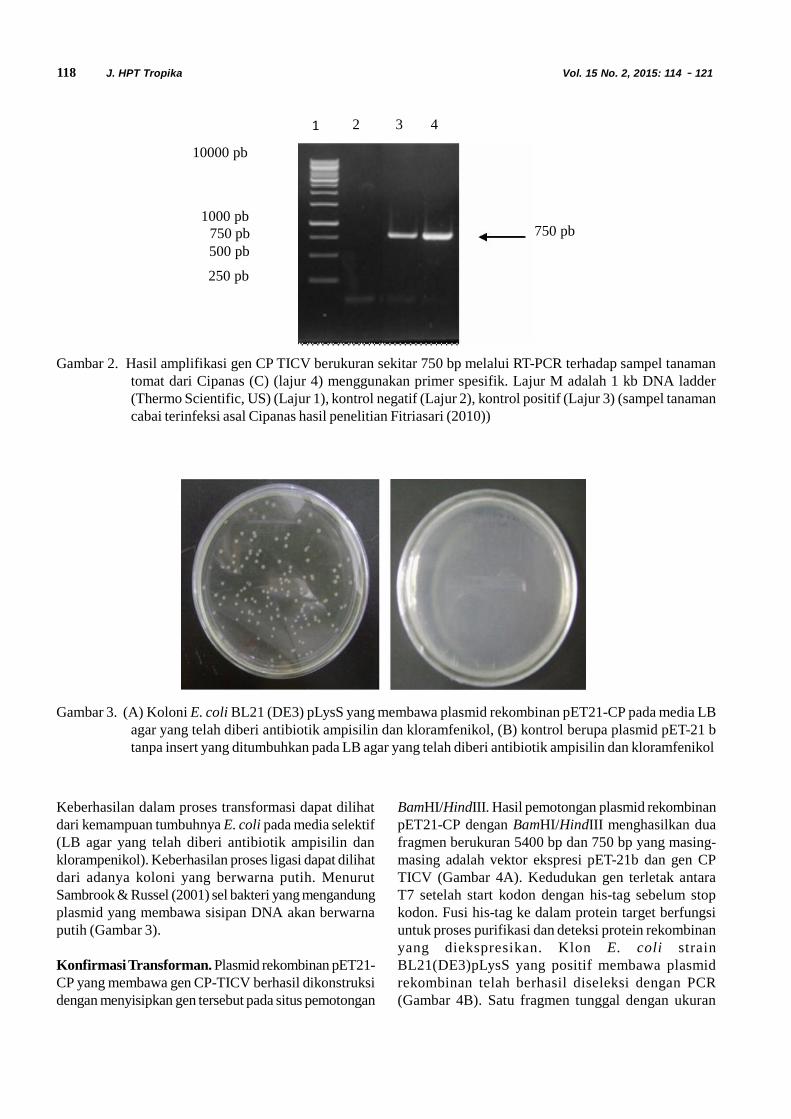
Amplifikasi Gen CP TICV. Gen CP berhasildiamplifikasi menggunakan sepasang primer spesifik(TICV–CP F-Bam no ATG (5’AATTAAGGATCCGAAAACTTATCTGGTAATGCAAAC-3’ dan TICV–CP R-Hind 5’AATTAAAAGCTTTTAGCATGGGTGTTTCATATCAGCC-3’). Produk PCRberukuran sekitar 750 pb yang disajikan dalam Gambar2, sesuai dengan hasil penelitian Fitriasari (2010).Fragmen gen CP hasil amplifikasi ini telah disekuensing(data tidak ditampilkan) dan mempunyai kemiripandengan isolat dari negara lain sebesar 99,4–100%.Fragmen gen CP ini selanjutnya digunakan pada tahapberikutnya yaitu kloning gen CP TICV.
Kloning Gen CP TICV. Proses ligasi dan transformasimerupakan tahapan utama dalam kloning gen.Penggabungan fragmen gen CP TICV ke dalam vektorekspresi pET-21 b merupakan proses ligasi. Hasil ligasiantara vektor ekspresi (pET-21b) dengan fragmen genCP TICV diintroduksikan ke dalam bakteri E. colisebagai inangnya yang kemudian ditumbuhkan ke dalammedia LB agar yang telah diberi antibiotik tertentu.
Gambar 1. Gejala penyakit klorosis pada tanaman tomat di lapangan, (A) interveinal yellowing, (B) nekrotik(Kurniawati et al., 2015)
A B
118 J. HPT Tropika Vol. 15 No. 2, 2015: 114 - 121
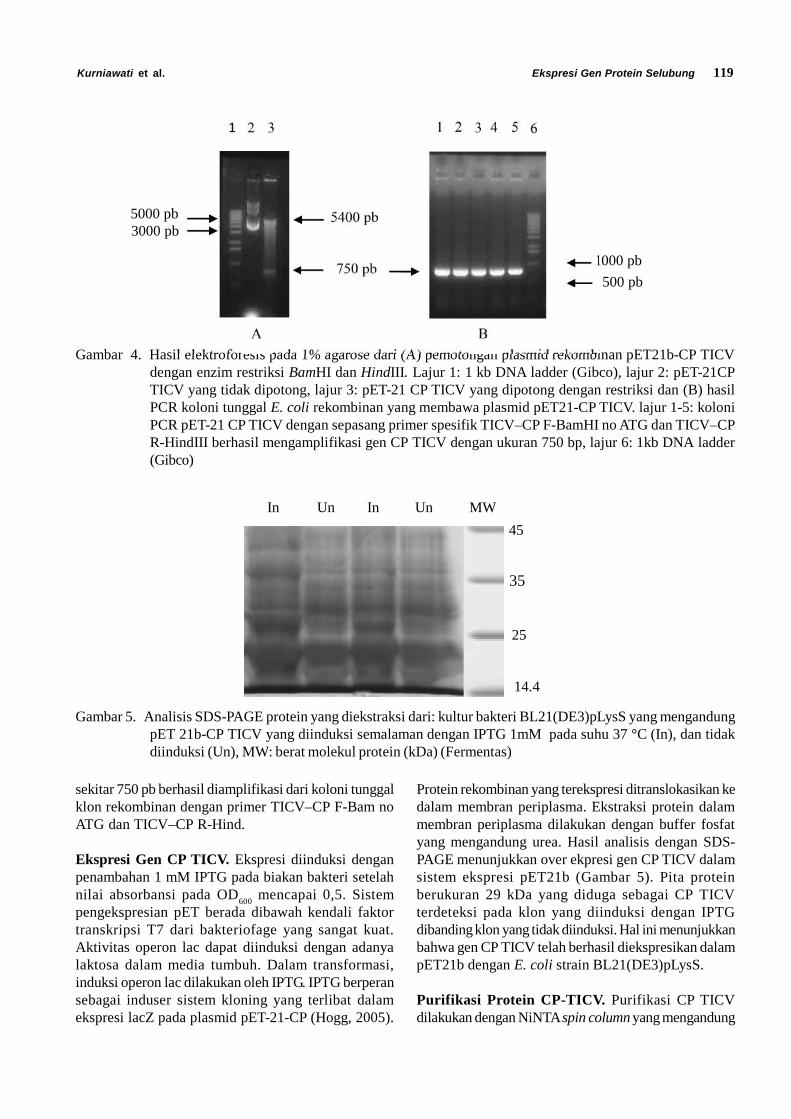
Keberhasilan dalam proses transformasi dapat dilihatdari kemampuan tumbuhnya E. coli pada media selektif(LB agar yang telah diberi antibiotik ampisilin danklorampenikol). Keberhasilan proses ligasi dapat dilihatdari adanya koloni yang berwarna putih. MenurutSambrook & Russel (2001) sel bakteri yang mengandungplasmid yang membawa sisipan DNA akan berwarnaputih (Gambar 3).
Konfirmasi Transforman. Plasmid rekombinan pET21-CP yang membawa gen CP-TICV berhasil dikonstruksidengan menyisipkan gen tersebut pada situs pemotongan
BamHI/HindIII. Hasil pemotongan plasmid rekombinanpET21-CP dengan BamHI/HindIII menghasilkan duafragmen berukuran 5400 bp dan 750 bp yang masing-masing adalah vektor ekspresi pET-21b dan gen CPTICV (Gambar 4A). Kedudukan gen terletak antaraT7 setelah start kodon dengan his-tag sebelum stopkodon. Fusi his-tag ke dalam protein target berfungsiuntuk proses purifikasi dan deteksi protein rekombinanyang diekspresikan. Klon E. coli strainBL21(DE3)pLysS yang positif membawa plasmidrekombinan telah berhasil diseleksi dengan PCR(Gambar 4B). Satu fragmen tunggal dengan ukuran
A B
Gambar 2. Hasil amplifikasi gen CP TICV berukuran sekitar 750 bp melalui RT-PCR terhadap sampel tanamantomat dari Cipanas (C) (lajur 4) menggunakan primer spesifik. Lajur M adalah 1 kb DNA ladder(Thermo Scientific, US) (Lajur 1), kontrol negatif (Lajur 2), kontrol positif (Lajur 3) (sampel tanamancabai terinfeksi asal Cipanas hasil penelitian Fitriasari (2010))
500 pb
1 2 3 4
750 pb
250 pb
1000 pb750 pb
10000 pb
Gambar 3. (A) Koloni E. coli BL21 (DE3) pLysS yang membawa plasmid rekombinan pET21-CP pada media LBagar yang telah diberi antibiotik ampisilin dan kloramfenikol, (B) kontrol berupa plasmid pET-21 btanpa insert yang ditumbuhkan pada LB agar yang telah diberi antibiotik ampisilin dan kloramfenikol
Kurniawati et al. Ekspresi Gen Protein Selubung 119
sekitar 750 pb berhasil diamplifikasi dari koloni tunggalklon rekombinan dengan primer TICV–CP F-Bam noATG dan TICV–CP R-Hind.
Ekspresi Gen CP TICV. Ekspresi diinduksi denganpenambahan 1 mM IPTG pada biakan bakteri setelahnilai absorbansi pada OD
600 mencapai 0,5. Sistem
pengekspresian pET berada dibawah kendali faktortranskripsi T7 dari bakteriofage yang sangat kuat.Aktivitas operon lac dapat diinduksi dengan adanyalaktosa dalam media tumbuh. Dalam transformasi,induksi operon lac dilakukan oleh IPTG. IPTG berperansebagai induser sistem kloning yang terlibat dalamekspresi lacZ pada plasmid pET-21-CP (Hogg, 2005).
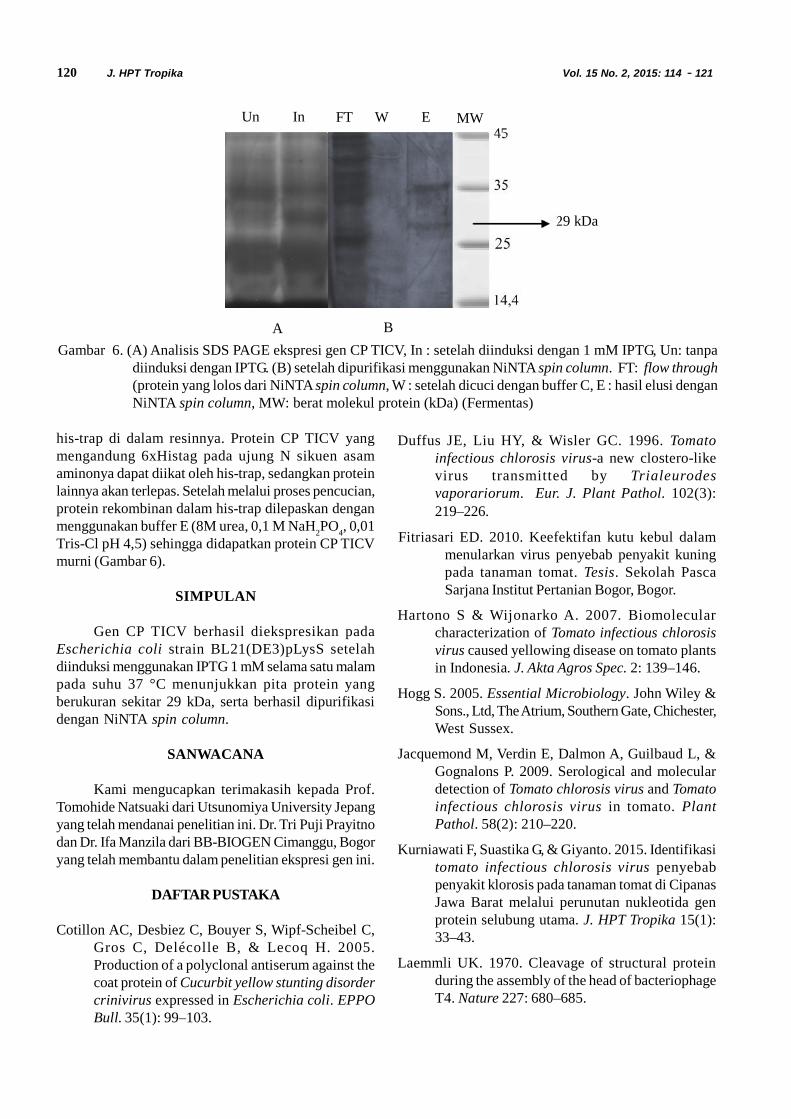
Protein rekombinan yang terekspresi ditranslokasikan kedalam membran periplasma. Ekstraksi protein dalammembran periplasma dilakukan dengan buffer fosfatyang mengandung urea. Hasil analisis dengan SDS-PAGE menunjukkan over ekpresi gen CP TICV dalamsistem ekspresi pET21b (Gambar 5). Pita proteinberukuran 29 kDa yang diduga sebagai CP TICVterdeteksi pada klon yang diinduksi dengan IPTGdibanding klon yang tidak diinduksi. Hal ini menunjukkanbahwa gen CP TICV telah berhasil diekspresikan dalampET21b dengan E. coli strain BL21(DE3)pLysS.
Purifikasi Protein CP-TICV. Purifikasi CP TICVdilakukan dengan NiNTAspin column yang mengandung
Gambar 4. Hasil elektroforesis pada 1% agarose dari (A) pemotongan plasmid rekombinan pET21b-CP TICVdengan enzim restriksi BamHI dan HindIII. Lajur 1: 1 kb DNA ladder (Gibco), lajur 2: pET-21CPTICV yang tidak dipotong, lajur 3: pET-21 CP TICV yang dipotong dengan restriksi dan (B) hasilPCR koloni tunggal E. coli rekombinan yang membawa plasmid pET21-CP TICV. lajur 1-5: koloniPCR pET-21 CP TICV dengan sepasang primer spesifik TICV–CP F-BamHI no ATG dan TICV–CPR-HindIII berhasil mengamplifikasi gen CP TICV dengan ukuran 750 bp, lajur 6: 1kb DNA ladder(Gibco)
3
A
1 32
3000 pb5000 pb 5400 pb
750 pb
B
1 4 652
1000 pb
500 pb
Gambar 5. Analisis SDS-PAGE protein yang diekstraksi dari: kultur bakteri BL21(DE3)pLysS yang mengandungpET 21b-CP TICV yang diinduksi semalaman dengan IPTG 1mM pada suhu 37 °C (In), dan tidakdiinduksi (Un), MW: berat molekul protein (kDa) (Fermentas)
14.4
45
35
25
In Un In Un MW
120 J. HPT Tropika Vol. 15 No. 2, 2015: 114 - 121
his-trap di dalam resinnya. Protein CP TICV yangmengandung 6xHistag pada ujung N sikuen asamaminonya dapat diikat oleh his-trap, sedangkan proteinlainnya akan terlepas. Setelah melalui proses pencucian,protein rekombinan dalam his-trap dilepaskan denganmenggunakan buffer E (8M urea, 0,1 M NaH
2PO
4, 0,01
Tris-Cl pH 4,5) sehingga didapatkan protein CP TICVmurni (Gambar 6).
SIMPULAN
Gen CP TICV berhasil diekspresikan padaEscherichia coli strain BL21(DE3)pLysS setelahdiinduksi menggunakan IPTG 1 mM selama satu malampada suhu 37 °C menunjukkan pita protein yangberukuran sekitar 29 kDa, serta berhasil dipurifikasidengan NiNTA spin column.
SANWACANA
Kami mengucapkan terimakasih kepada Prof.Tomohide Natsuaki dari Utsunomiya University Jepangyang telah mendanai penelitian ini. Dr. Tri Puji Prayitnodan Dr. Ifa Manzila dari BB-BIOGEN Cimanggu, Bogoryang telah membantu dalam penelitian ekspresi gen ini.
DAFTAR PUSTAKA
Cotillon AC, Desbiez C, Bouyer S, Wipf-Scheibel C,Gros C, Delécolle B, & Lecoq H. 2005.Production of a polyclonal antiserum against thecoat protein of Cucurbit yellow stunting disordercrinivirus expressed in Escherichia coli. EPPOBull. 35(1): 99–103.
Duffus JE, Liu HY, & Wisler GC. 1996. Tomatoinfectious chlorosis virus-a new clostero-likevirus transmitted by Trialeurodesvaporariorum. Eur. J. Plant Pathol. 102(3):219–226.
Fitriasari ED. 2010. Keefektifan kutu kebul dalammenularkan virus penyebab penyakit kuningpada tanaman tomat. Tesis. Sekolah PascaSarjana Institut Pertanian Bogor, Bogor.
Hartono S & Wijonarko A. 2007. Biomolecularcharacterization of Tomato infectious chlorosisvirus caused yellowing disease on tomato plantsin Indonesia. J. Akta Agros Spec. 2: 139–146.
Hogg S. 2005. Essential Microbiology. John Wiley &Sons., Ltd, The Atrium, Southern Gate, Chichester,West Sussex.
Jacquemond M, Verdin E, Dalmon A, Guilbaud L, &Gognalons P. 2009. Serological and moleculardetection of Tomato chlorosis virus and Tomatoinfectious chlorosis virus in tomato. PlantPathol. 58(2): 210–220.
Kurniawati F, Suastika G, & Giyanto. 2015. Identifikasitomato infectious chlorosis virus penyebabpenyakit klorosis pada tanaman tomat di CipanasJawa Barat melalui perunutan nukleotida genprotein selubung utama. J. HPT Tropika 15(1):33–43.
Laemmli UK. 1970. Cleavage of structural proteinduring the assembly of the head of bacteriophageT4. Nature 227: 680–685.
Gambar 6. (A) Analisis SDS PAGE ekspresi gen CP TICV, In : setelah diinduksi dengan 1 mM IPTG, Un: tanpadiinduksi dengan IPTG. (B) setelah dipurifikasi menggunakan NiNTA spin column. FT: flow through(protein yang lolos dari NiNTA spin column, W : setelah dicuci dengan buffer C, E : hasil elusi denganNiNTA spin column, MW: berat molekul protein (kDa) (Fermentas)
A B
Un In FT W E MW45
35
25
14,4
29 kDa

Kurniawati et al. Ekspresi Gen Protein Selubung 121
Nishimura M, Park G, & Xu JR. 2003. The G-betasubunit MGB1 is involved in regulating multiplesteps of infection-related morphogenesis inMagnaporthe grisea. Mol. Microbiol. 50(1): 231–243.
Qiagen. 2003. The Qiaexpressionist: a Handbook forHigh Level Expression and Purification of 6xHis-tagged Protein. Qiagen Inc., Valencia.
Qiagen. 2008. Ni-NTA Spin Kit Handbook. Ed ke-2.Qiagen Inc. Valencia.
Sa’adah L. 2013. Uji serologi diferensial dan simultan untukTomato chlorosis virus (ToCV) dan Tomatoinfectious chlorosis virus (TICV) pada tanamantomat. Skripsi. Institut Pertanian Bogor, Bogor.
Sambrook J & Russel DW. 2001. Molecular Cloning:a Laboratory Manual. Cold Spring HarborLaboratory, New York.
Wisler GC, Duffus JE, Liu HY, & Li RH. 1998. Ecologyand epidemiology of whitefly-transmittedclosteroviruses. Plant Dis. 82(3): 270–280.

132 J. HPT Tropika Vol. 15 No. 2, 2015: 132 - 140J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 132 – 140, September 2015
IDENTIFIKASI MOLEKULER BEAN COMMON MOSAIC VIRUS YANGBERASOSIASI DENGAN PENYAKIT MOSAIK KUNING KACANG PANJANG
Melinda1, Tri Asmira Damayanti2, & Sri Hendrastuti Hidayat2
1Fakultas Pertanian, Universitas Negeri Papua, ManokwariJl. Gunung Salju, Amban Manokwari, Papua Barat
2Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper, Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Molecular identification of bean common mosaic virus associated with yellow mosaic disease on yard long bean. Beancommon mosaic virus (BCMV) has been reported as one of the causal agents of yellow mosaic disease on yard long bean inWest Java and Central Java. Infected plants showed mosaic, yellowing, and mixture of yellow mosaic. The research wasconducted to identify the diversity of BCMV associated with yellow mosaic disease based on coat protein (CP) gene sequences.Symptomatic leaf samples were collected from yard long bean growing areas in several districts in West Java (Bogor, Cirebon,Subang, and Indramayu), and several districts in Central Java (Tegal, Klaten, Solo, Yogjakarta, Sleman, and Magelang).Molecular detection using RT-PCR method was carried out by using specific primer to BCMV which will amplify the CP gene.DNA fragment, + 860 bp in size, was successfully amplified from 8 out of 13 leaf samples, i.e samples from three villages inBogor District (Cangkurawok, Bubulak, Bojong), and five samples from District of Cirebon, Subang, Solo, Sleman, and Tegal.Sequence analysis of those DNA fragment showed that 4 isolates (Bogor-Cangkurawok, Subang, Solo and Sleman) had thehighest homology to BCMV-BlC from Taiwan, whereas 2 isolates (Cirebon and Tegal) had the highest homology to BCMV-NL1 from England. Further, phyllogenetic analysis revealed that those of 4 isolates were closely related to BCMV-BlC fromTaiwan based on nucleotide as well as amino acid sequences; while those other 2 isolates were closely related to BCMV-NL1from England based on nucleotide sequences but closely related to BCMV-BlC Y from China based on amino acid sequences.Phyllogenetic analysis showed that those of 6 BCMV isolates separated in two different clusters; 4 isolates (Bogor-Cangkurawok, Subang, Solo, and Sleman) in cluster 1 together with BCMV-BlC from Taiwan, while other 2 isolates (Cirebonand Tegal) in cluster 2 together with BCMV-NL1.
Key words: bean common mosaic virus, coat protein, diversity, yard long bean
ABSTRAK
Identifikasi molekuler bean common mosaic virus yang berasosiasi dengan penyakit mosaik kuning kacang panjang.Bean common mosaic virus (BCMV) telah dilaporkan sebagai salah satu penyebab penyakit mosaik kuning di pertanamankacang panjang di Jawa Barat dan Jawa Tengah. Tanaman terinfeksi menunjukkan gejala mosaik, menguning, dan campuranmosaik kuning. Penelitian ini dilakukan untuk mengidentifikasi keragaman BCMV yang berasosiasi dengan penyakit mosaikkuning berdasarkan urutan gen protein selubung (CP). Sampel yang bergejala dikumpulkan dari pertanaman kacang panjangdi beberapa kabupaten di Jawa Barat (Bogor, Cirebon, Subang, dan Indramayu), dan beberapa kabupaten di Jawa Tengah(Tegal, Klaten, Solo, Yogjakarta, Sleman, dan Magelang). Deteksi molekuler menggunakan metode RT-PCR, dilakukan denganmenggunakan primer spesifik BCMV yang menyandi gen CP. Fragmen DNA berukuran + 860 pb berhasil diamplifikasi 8 dari 13sampel daun, yaitu sampel dari 3 desa di Kabupaten Bogor (Cangkurawok, Bubulak, Bojong) dan 5 sampel dari KabupatenCirebon, Subang, Solo, Sleman, dan Tegal. Analisis urutan fragmen DNA menunjukkan bahwa 4 isolat (Bogor-Cangkurawok,Subang, Solo dan Sleman) memiliki homologi tertinggi dengan BCMV-BlC dari Taiwan, sedangkan 2 isolat (Cirebon dan Tegal)memiliki homologi tertinggi dengan BCMV-NL1 dari Inggris. Analisis filogenetika lebih lanjut menunjukkan bahwa 4 isolat(Bogor-Cangkurawok, Subang, Solo, dan Sleman) memiliki kekerabatan yang dekat dengan BCMV-BlC dari Taiwan berdasarkanurutan nukleotida serta urutan asam amino, sedangkan 2 isolat (Cirebon dan Tegal), memiliki kekerabatan yang dekat denganBCMV-NL1 dari Inggris berdasarkan urutan nukleotida dan memiliki kekerabatan yang dekat dengan BCMV-BlC Y dari Cinaberdasarkan urutan asam amino. Analisis filogenetika menunjukkan 6 isolat BCMV terpisah menjadi 2 kluster; 4 isolat (Bogor-Cangkurawok, Subang, Solo, dan Sleman) pada kluster 1 bersama BCMV-BlC dari Taiwan dan 2 isolat (Cirebon dan Tegal) padakluster 2 bersama BCMV-NL1.
Kata kunci: bean common mosaic virus, kacang panjang, keragaman, protein selubung

Melinda et al. Identifikasi Molekuler BCMV 133
PENDAHULUAN
Tanaman kacang panjang (Vigna sinensis ssp.sesquipedalis) merupakan tanaman hortikultura yangmemiliki peluang sebagai komoditas strategis untukpangan masa depan serta sebagai alternatif sumberpertumbuhan ekonomi (Pitojo, 2006). Berdasarkan dataBadan Pusat Statistik (2011), produksi kacang panjangdi Indonesia pada tahun 2010 mencapai 489.449 ton danpada tahun 2011 menurun menjadi 458.307 ton. Salahsatu faktor pembatas produksi kacang panjang adalahgangguan hama dan penyakit tanaman. Hama yangumum ditemukan pada pertanaman kacang panjangadalah kutu daun Aphis craccivora Koch, penggerekpolong (Maruca testulalis Geyer), kutu kebul (Bemisiatabaci Genn), ulat jengkal (Plusia chalcites Esper), lalatkacang (Agromyza phaseoli Cog), tungau merah(Tetranychus cinnabarinus Boisd.) dan kepik polong(Riptortus linearis F.). Sedangkan penyakit yang seringditemui diantaranya adalah layu (Fusarium oxysporumf.sp. phaseoli), antraknosa (Colletotrichumlindemuthianum (Sacc et Magn), mosaik yangdisebabkan oleh Bean common mosaic virus(BCMV), Bean yellow mosaic virus (BYMV) danCowpea aphid-borne mosaic virus (CABMV), daunkecil kacang panjang (Cowpea witches-broom virus)dan puru akar (Meloidogyne sp.) (Anwar et al., 2005;Haryanto et al., 2010; Lazuardi, 2005; Siregar, 1996).
Pada tahun 2008–2009, Damayanti et al. (2009)melaporkan insidensi penyakit mosaik kuning kacangpanjang yang meluas di beberapa daerah pantai utaraJawa Barat. Tanaman yang terinfeksi menunjukkangejala mosaik kuning, penghambatan pertumbuhan danmalformasi daun. Infeksi pada tanaman yang masihmuda menyebabkan tidak terbentuknya polong dan jikainfeksinya sangat parah dapat mengakibatkan kematian.Salah satu virus yang terdeteksi pada tanaman kacangpanjang dengan gejala mosaik kuning dari daerahBubulak, Dramaga Bogor dan Sidorejo, KaranganyarPekalongan adalah BCMV strain Blackeye (BCMV-BlC).
BCMV umumnya menginfeksi tanaman kacang-kacangan seperti pada spesies Phaseolus vulgaris,Vigna unguiculata, dan V. radiata. Tanaman yangterinfeksi menunjukkan gejala utama mosaik dannekrosis, tulang daun berwarna hijau tua sedangkandaerah interveinal menjadi hijau muda. Adanyaperubahan warna daun biasanya diikuti denganmalformasi daun berupa kerutan dan menggulung. Gejalapada tanaman umumnya muncul 7 sampai 10 harisetelah inokulasi. Tipe gejala yang ditimbulkan ditentukanoleh strain virus, suhu lingkungan pertanaman dan
genotipe inang. Terdapat 10 strain BCMV yang telahdiketahui yaitu Blackeye, US1, US5, NL2, NL3, NL4,NL5, NL6, NL7 dan NL8 (Morales, 1988). Sebelumnyadilaporkan infeksi BCMV dan BCMNV (Bean commonmosaic necrosis virus) menyebabkan gejala yangmosaik yang berbeda dari gejala khas pada tanamanPhaseolus lunatus (lima bean) di Peru (Melgarejo etal., 2007). Selain itu beberapa isolat BCMV-BlC jugadilaporkan menginfeksi vanili di India (Bhadramurthy& Bhat, 2009) dan menjadi masalah serius yangmengancam produksi kedelai di Cina (Zhou et al., 2014).
Mengingat sedikitnya informasi mengenai strainBCMV atau keragaman BCMV yang menginfeksitanaman kacang panjang di Indonesia, maka perludilakukan identifikasi secara molekuler terhadap BCMVyang menyebabkan penyakit mosaik kuning padatanaman kacang panjang. Insidensi penyakit mosaikkuning di lapangan dilaporkan masih cukup tinggiterutama di Jawa Barat dan Jawa Tengah. Oleh karenaitu penelitian ini bertujuan untuk mengidentifikasikeragaman BCMV dari tanaman kacang panjang yangberasal dari beberapa daerah di Jawa Barat dan JawaTengah.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Virologi Tumbuhan, Departemen ProteksiTanaman, Fakultas Pertanian, Institut Pertanian Bogormulai bulan Juni sampai dengan Desember 2012.
Pengamatan Gejala di Lapangan sertaPengambilan Sampel Tanaman. Pengamatan gejaladan pengambilan sampel dilakukan di pertanamankacang panjang di daerah Dramaga, Kabupaten BogorPropinsi Jawa Barat yaitu di daerah Cangkurawok,Bubulak, Situgede dan Bojong. Sebanyak 20 sampel daunyang menunjukkan gejala mosaik kuning diambil darimasing-masing lokasi. Sampel daun dengan gejala kuningjuga diperoleh dari koleksi Laboratorium VirologiTumbuhan IPB yaitu sampel yang berasal dari beberapaKabupaten di Jawa Barat (Bogor, Cirebon, Subang, danIndramayu) dan beberapa kabupaten di Jawa Tengah(Tegal, Solo, Yogjakarta, Sleman, Klaten, dan Magelang).
Deteksi BCMV dengan Teknik ReverseTranscription-Polymerase Chain Reaction (RT-PCR)
Ekstraksi RNA Total. RNA total diekstraksi darijaringan daun tanaman bergejala sebanyak 0,1 gmenggunakan NucleoSpin RNA Plant (Macherey-Nagel Inc., USA) atau Xprep Plant RNA Mini Kit

134 J. HPT Tropika Vol. 15 No. 2, 2015: 132 - 140
(Philakorea Technology) dilakukan sesuai protokol yangdianjurkan pembuatnya.
Sintesis cDNA. RNA total hasil ekstraksiselanjutnya digunakan untuk proses transkripsi balik(reverse transcription-RT) menjadi cDNA(complementary DNA) dengan menggunakan enzimReverse Transcriptase. Reaksi transkripsi balik terdiridari: 4 µl RNA total, 4 µl 5x bufer RT, 2 µl DTT(dithiothreitol) 50 mM, 2 µl dNTP 10 mM, 1 µl M-MuLV(200 U/µl), 1 µl RNase inhibitor (40 U/µl), 2 µl oligod(T)
18 10 µM dan air bebas nuklease sampai volume
total 20 µl. Reaksi RT dilakukan dalam mesin AutomatedThermal Cycler (Gene Amp PCR System 9700; PEApplied Biosystem, USA) yang diprogram untuk satusiklus pada suhu 25 oC selama 5 menit, 42 oC selama 60menit, dan 70 oC selama 15 menit. Siapan cDNA hasilRT ini digunakan sebagai DNA cetakan dalam reaksiamplifikasi.
Amplifikasi DNA dengan PCR. AmplifikasiDNA virus dilakukan dengan metode Polymerase ChainReaction (PCR) dengan menggunakan pasangan primerspesifik gen coat protein BCMV yaitu primer forwardBCMV-F dan primer reverse BCMV-R (Anggraini &Hidayat, 2014) dengan hasil amplifikasi berukuran + 860pb (pasang basa). Reaksi PCR terdiri dari: 1 µl cDNA,12,5 µl premix Go Taq Green (Promega Jepang),masing-masing 1 µl primer BCMV-F dan BCMV-R (10µM/µl), dan air bebas nuklease sampai volume total 25µl. Amplifikasi dilakukan pada Automated ThermalCycler (Gene Amp PCR System 9700; PE AppliedBiosystem, USA). Amplifikasi didahului dengandenaturasi awal pada suhu 94 oC selama 5 menit,dilanjutkan dengan 35 siklus yang terdiri dari denaturasipada suhu 94 oC selama 1 menit, penempelan primerpada suhu 68 oC selama 1 menit dan ekstensi pada suhu72 oC selama 1 menit. Siklus terakhir ditambahkan 7menit pada suhu 72 oC untuk tahapan sintesis dan siklusberakhir pada suhu 4 oC.
Visualisasi DNA. Gel agarosa konsentrasi 1% dibuatdari 0,3 g agarosa yang dilarutkan dalam bufer TBE0,5x sebanyak 30 ml dengan bantuan microwave.Sebelum digunakan, larutan gel agarosa didinginkanhingga suhu 50 oC selama kurang lebih 15 menit laluditambahkan 3 µl ethidium bromide (0,5 µg/10 ml).Elektroforesis dilakukan pada tegangan 100 V selama20 menit. Hasil elektroforesis selanjutnya divisualisasidibawah transilluminator ultraviolet dan didokumentasidengan kamera digital.
Perunutan dan penyejajaran DNA. Fragmen DNAhasil amplifikasi PCR dikirim ke First Base GeneticaScience (Singapura) untuk proses perunutan nukleotida.Hasil perunutan nukleotida digunakan untuk melihattingkat kesamaan dengan runutan nukleotida yangterdapat di GenBank menggunakan program BLAST(Basic Local Alignment Search Tool) (http://www.ncbi.nlm.nih.gov/BLAST) dan identitas matrikshomologi menggunakan perangkat lunak BioEdit (Hall,1999), serta MEGA 5.05. untuk konstruksi pohonfilogenetika.
HASIL DAN PEMBAHASAN
Gejala Mosaik Kuning pada Tanaman KacangPanjang di Lapangan. Sampel-sampel tanaman kacangpanjang sakit yang diambil dari lapangan menunjukkangejala mosaik kuning. Gejala mosaik kuning di lapangandapat dibedakan menjadi dua kelompok berdasarkanwarna dan perubahan bentuk pada daun. Kelompokpertama menunjukkan gejala mosaik kuning dengan polamosaik berwarna hijau muda sampai tua kekuningandengan warna terang, terjadi penebalan tulang daun (veinbanding), malformasi daun (daun menggulung sepertikrupuk, mengerut sepanjang tulang daun), dan terdapatlepuhan. Bila gejala berlanjut tanaman menjadi kerdildan terhambat pertumbuhannya. Kelompok keduamenunjukkan gejala dengan pola mosaik berwarna hijaumuda sampai tua dengan batas kuning yang kurangtegas, terjadi penebalan tulang daun (vein banding),dan malformasi daun (Tabel 1).
Gejala yang ditunjukkan tanaman sampel padakelompok pertama mengindikasikan tanaman terinfeksiBean common mosaic virus (BCMV). Tanaman yangterinfeksi BCMV menunjukkan gejala utama mosaik dannekrosis, tulang daun berwarna hijau tua sedangkandaerah interveinal menjadi hijau muda diikuti denganperubahan warna daun dan malformasi daun berupakerutan dan menggulung (CABI, 2007). Menurut Agrios(2005) gejala awal daun yang terinfeksi BCMV adalahdaun menjadi bergelombang kemudian warna daunmenjadi berubah dan tidak merata, seiring denganberjalannya waktu daun melengkung ke bawah dan keatas. Daun selanjutnya terlihat mengerut dan terjadimosaik, malformasi daun dan penebalan di sekitarpertulangan daun berwarna hijau tua.
Berdasarkan deskripsi yang diuraikan di atas, adaindikasi bahwa gejala mosaik kuning pada kacangpanjang dapat disebabkan oleh infeksi beberapa jenisvirus. Kepastian adanya infeksi BCMV dapat ditentukanberdasarkan metode deteksi secara molekuler.

Melinda et al. Identifikasi Molekuler BCMV 135
*Mkr = Mosaik kuning ringan; Mks = Mosaik kuning sedang; Mkb = Mosaik kuning berat; Md = Malformasi daun;Vb = Vein banding; K = Kerdil; + = BCMV; - = bukan BCMV.
Deteksi Virus dengan Metode RT-PCR danPerunutan Basa Nukleotida. Pita DNA berukuran+ 860 pb berhasil teramplifikasi dari sampel asalKabupaten Bogor (Desa Cangkurawok, Bubulak,Bojong), Cirebon, Subang, Solo, Sleman, dan Tegal.BCMV tidak teramplifikasi dari sampel asal Bogor-Situgede, Indramayu, Yogjakarta, Klaten dan Magelang(Gambar 1). Hasil deteksi ini menunjukkan bahwapengamatan gejala penyakit saja tidak cukup untukmendeteksi dan mengidentifikasi virus pada tanamansehingga diperlukan diagnosis secara molekuler.Damayanti et al. (2009) berhasil mendeteksi BCMVdan Cucumber mosaic virus (CMV) dari tanamankacang panjang yang menunjukkan gejala mosaik kuning.Oleh karena itu gejala mosaik yang tampak pada sampel-
sampel daun pada penelitian ini, kemungkinan jugadisebabkan oleh infeksi virus selain BCMV. Nurulita(2014) melaporkan PCR DNA menggunakan primeruniversal Begomovirus dari gejala mosaik kuning kacangpanjang positif Begomovirus.
Pita DNA hasil amplifikasi digunakan untukperunutan basa nukleotida dan diperoleh hasil perunutanberkisar 779 sampai 861 pb (Tabel 2). Setelah dilakukananalisis homologi terhadap sekuen yang ada di Genbankmenggunakan program BLAST (http://www.ncbi.nlm.nih.gov/BLAST) diketahui bahwa isolat-isolat Bogor-Cangkurawok, Subang, Solo dan Slemanmemiliki 99% sampai 100% homologi dengan BCMV-BlC asal Taiwan (AY575773.1), sedangkan isolat–isolatCirebon dan Tegal memiliki 90% homologi dengan
Gambar 1. Pita DNA hasil amplifikasi dengan primer spesifik gen CP BCMV (BCMV-F/BCMV-R) pada gelagarosa 1%. M = Penanda DNA 100 pb, DNA sampel asal; 1. Situgede, 2. Cangkurawok, 3.Bubulak.Bojong, 5. Subang, 6. Indramayu, 7. Cirebon, 8. Tegal, 9. Klaten, 10. Solo, 11. Magelang, 12.Yogjakarta, 13. Sleman; K- = Kontrol negatif (air), K+ = Kontrol positif (isolat BCMV)
Tabel 1. Tipe gejala dan hasil deteksi BCMV menggunakan metode RT-PCR
Asal sampel Tipe gejala*) Hasil PCR
Bogor-Cangkurawok Mkb, Md, Vb, K +-Bubulak Mks, Vb +-Situgede Mks, Vb --Bojong Mks, Vb +
Cirebon Mkb, Md, Vb +Subang Mkb, Md, Vb, K +Indramayu Mks, Vb -Klaten Mks, Vb -Solo Mkb, Md, Vb, K +Yogjakarta Mks, Vb -Sleman Mks, Md, Vb +Tegal Mkb, Md, Vb +Magelang Mkr, Vb -
900 pb
M 1 2 3 4 5 6 7 8 9 10 11 12 13 K- K+
600 pb
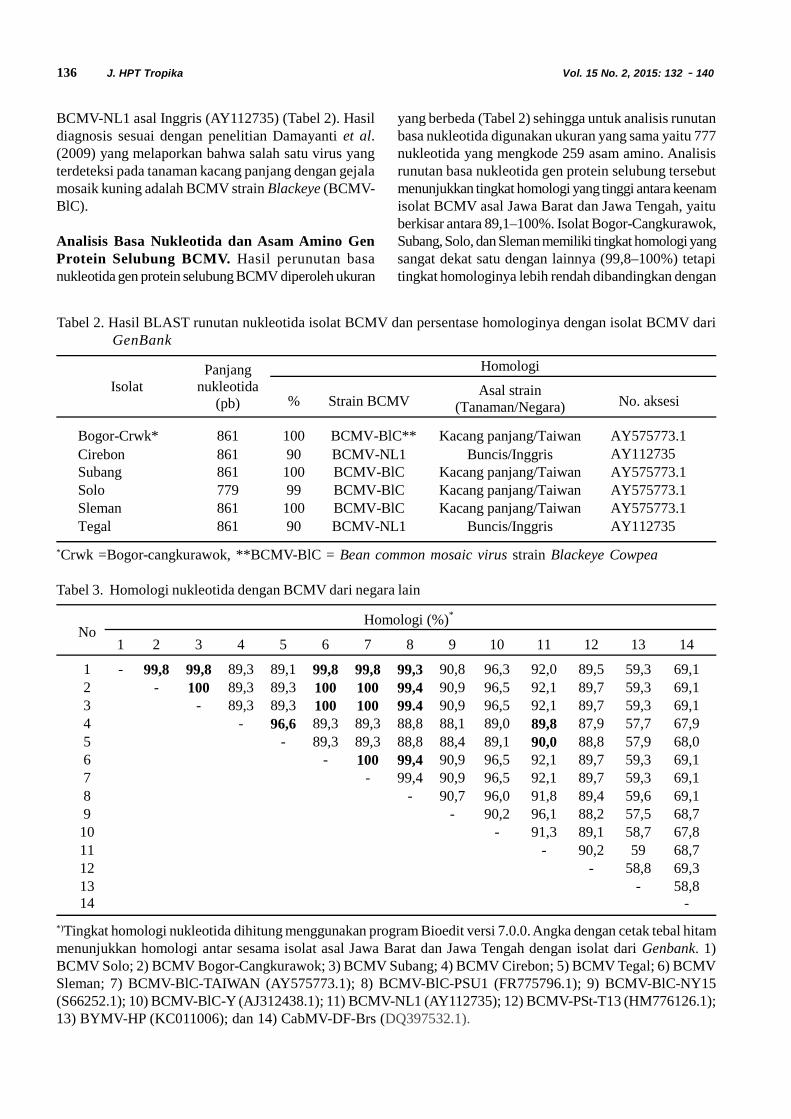
136 J. HPT Tropika Vol. 15 No. 2, 2015: 132 - 140
BCMV-NL1 asal Inggris (AY112735) (Tabel 2). Hasildiagnosis sesuai dengan penelitian Damayanti et al.(2009) yang melaporkan bahwa salah satu virus yangterdeteksi pada tanaman kacang panjang dengan gejalamosaik kuning adalah BCMV strain Blackeye (BCMV-BlC).
Analisis Basa Nukleotida dan Asam Amino GenProtein Selubung BCMV. Hasil perunutan basanukleotida gen protein selubung BCMV diperoleh ukuran
yang berbeda (Tabel 2) sehingga untuk analisis runutanbasa nukleotida digunakan ukuran yang sama yaitu 777nukleotida yang mengkode 259 asam amino. Analisisrunutan basa nukleotida gen protein selubung tersebutmenunjukkan tingkat homologi yang tinggi antara keenamisolat BCMV asal Jawa Barat dan Jawa Tengah, yaituberkisar antara 89,1–100%. Isolat Bogor-Cangkurawok,Subang, Solo, dan Sleman memiliki tingkat homologi yangsangat dekat satu dengan lainnya (99,8–100%) tetapitingkat homologinya lebih rendah dibandingkan dengan
Tabel 2. Hasil BLAST runutan nukleotida isolat BCMV dan persentase homologinya dengan isolat BCMV dariGenBank
*Crwk =Bogor-cangkurawok, **BCMV-BlC = Bean common mosaic virus strain Blackeye Cowpea
Tabel 3. Homologi nukleotida dengan BCMV dari negara lain
NoHomologi (%)*
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 - 99,8 99,8 89,3 89,1 99,8 99,8 99,3 90,8 96,3 92,0 89,5 59,3 69,12 - 100 89,3 89,3 100 100 99,4 90,9 96,5 92,1 89,7 59,3 69,13 - 89,3 89,3 100 100 99.4 90,9 96,5 92,1 89,7 59,3 69,14 - 96,6 89,3 89,3 88,8 88,1 89,0 89,8 87,9 57,7 67,95 - 89,3 89,3 88,8 88,4 89,1 90,0 88,8 57,9 68,06 - 100 99,4 90,9 96,5 92,1 89,7 59,3 69,17 - 99,4 90,9 96,5 92,1 89,7 59,3 69,18 - 90,7 96,0 91,8 89,4 59,6 69,19 - 90,2 96,1 88,2 57,5 68,710 - 91,3 89,1 58,7 67,811 - 90,2 59 68,712 - 58,8 69,313 - 58,814 -
*)Tingkat homologi nukleotida dihitung menggunakan program Bioedit versi 7.0.0. Angka dengan cetak tebal hitammenunjukkan homologi antar sesama isolat asal Jawa Barat dan Jawa Tengah dengan isolat dari Genbank. 1)BCMV Solo; 2) BCMV Bogor-Cangkurawok; 3) BCMV Subang; 4) BCMV Cirebon; 5) BCMV Tegal; 6) BCMVSleman; 7) BCMV-BlC-TAIWAN (AY575773.1); 8) BCMV-BlC-PSU1 (FR775796.1); 9) BCMV-BlC-NY15(S66252.1); 10) BCMV-BlC-Y (AJ312438.1); 11) BCMV-NL1 (AY112735); 12) BCMV-PSt-T13 (HM776126.1);13) BYMV-HP (KC011006); dan 14) CabMV-DF-Brs (DQ397532.1).
IsolatPanjang
nukleotida(pb)
Homologi
% Strain BCMVAsal strain
(Tanaman/Negara) No. aksesi
Bogor-Crwk* 861 100 BCMV-BlC** Kacang panjang/Taiwan AY575773.1Cirebon 861 90 BCMV-NL1 Buncis/Inggris AY112735Subang 861 100 BCMV-BlC Kacang panjang/Taiwan AY575773.1Solo 779 99 BCMV-BlC Kacang panjang/Taiwan AY575773.1Sleman 861 100 BCMV-BlC Kacang panjang/Taiwan AY575773.1Tegal 861 90 BCMV-NL1 Buncis/Inggris AY112735

Melinda et al. Identifikasi Molekuler BCMV 137
Tabel 4. Homologi asam amino dengan BCMV asal negara lain
N0.Homologi (%)
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 - 99,6 99,6 93,0 93,0 99,6 99,6 99,2 92,2 99,2 93,0 92,2 57,3 66,42 - 100 93,4 93,4 100 100 99,6 92,6 99,6 93,4 92,6 57,3 66,43 - 93,4 93,4 100 100 99,6 92,6 99,6 93,4 92,6 57,3 66,44 - 98,8 93,4 93,4 93,0 90,7 93,8 91,8 93,0 57,3 67,55 - 93,4 93,4 93,0 91,1 93,8 92,2 93,0 57,3 67,56 - 100 99,6 92,6 99,6 93,4 92,6 57,3 66,47 - 99,6 92,6 99,6 93,4 92,6 57,3 66,48 - 93,0 992 93,8 93,0 57,3 66,79 - 93,0 94,9 90,7 57,3 66,7
10 - 93,8 93,0 57,3 66,411 - 92,6 58,4 66,712 - 58 68,313 - 54,914 -
*)Tingkat homologi nukleotida dihitung menggunakan program BioEdit versi 7.0.0. Angka dengan cetak tebal hitammenunjukkan homologi antar sesama isolat asal Jawa Barat dan Jawa Tengah dengan isolat dari Genbank. 1)BCMV Solo; 2) BCMV Cangkurawok; 3) BCMV Subang; 4) BCMV Cirebon; 5) BCMV Tegal; 6) BCMVSleman; 7) BCMV-BlC-TAIWAN (AY575773.1); 8) BCMV-BlC-PSU1 (FR775796.1); 9) BCMV-BlC-NY15(S66252.1); 10) BCMV-BlC-Y (AJ312438.1); 11) BCMV-NL1 (AY112735); 12) BCMV-PSt-T13 (HM776126.1);13) BYMV-HP (KC011006); dan 14) CabMV-DF-Brs (DQ397532.1).
isolat Cirebon dan Tegal (89,1–89,3%). Dua isolatBCMV asal Cirebon dan Tegal memiliki tingkat homologiyang sangat dekat satu dengan lainnya (96,6%) tetapitidak terlalu dekat dengan keempat isolat lainnya (Tabel3).
Analisis homologi dengan melibatkan beberapaisolat lain dari Genbank menunjukkan bahwa 4 isolatBCMV (isolat-isolat Bogor-Cangkurawok, Subang, Solodan Sleman) memiliki hubungan homologi yang dekatdengan BCMV-BlC dari Taiwan (AY575773.1) danBCMV-BlC PSU1 dari Thailand (FR775796.1) dengantingkat homologi berkisar 99,3–100%. Dua isolat BCMVasal Cirebon dan Tegal lebih dekat hubungankekerabatannya dengan BCMV-NL1 (AY112735) dariInggris dengan tingkat homologi 89,8–90,0%dibandingkan hubungannya dengan BCMV-BlC asalTaiwan ataupun BCMV-BlC PSU1 asal Thailand.Perbedaan tingkat homologi tersebut memberikanindikasi adanya keragaman diantara isolat-isolat BCMVdari daerah Jawa Barat dan Jawa Tengah.
Analisis runutan asam amino gen protein selubungmenunjukkan tingkat homologi yang tinggi antara keenamisolat BCMV asal Jawa Barat dan Jawa Tengah, yaituberkisar antara 93,0–100%. Isolat Bogor-Cangkurawok,
Subang, Solo, dan Sleman memiliki tingkat homologi yangsangat dekat satu dengan lainnya (99,6–100%) tetapitingkat homologinya tidak terlalu dekat dengan isolatCirebon dan Tegal (93,0–93,4%). Dua isolat BCMVasal Cirebon dan Tegal memiliki tingkat kekerabatanyang sangat dekat satu dengan lainnya (98,8%) tetapitidak terlalu dekat dengan keempat isolat lainnya (Tabel4).
Analisis kekerabatan dengan melibatkan beberapaisolat lain dari Genbank menunjukkan bahwa 4 isolatBCMV dari Bogor-Cangkurawok, Subang, Solo danSleman memiliki hubungan kekerabatan yang dekatdengan BCMV-BlC dari Taiwan (AY575773.1) dengantingkat kesamaan berkisar 99,6–100%. Dua isolatBCMV asal Cirebon dan Tegal lebih dekat hubungankekerabatannya dengan BCMV-BlC Y asal Cina(AJ312438.1) dengan tingkat kesamaan 93,8%dibandingkan hubungannya dengan BCMV-BlC asalTaiwan.
Menurut Adams et al. (2005), sekuen demarkasigen protein selubung dalam Potyvirus disebut spesiesyang sama jika homologi nukleotidanya berkisar 76–77%dan homologi runutan asam aminonya 82%. Homologinukleotida dan asam amino 6 isolat BCMV yang diisolasi
138 J. HPT Tropika Vol. 15 No. 2, 2015: 132 - 140
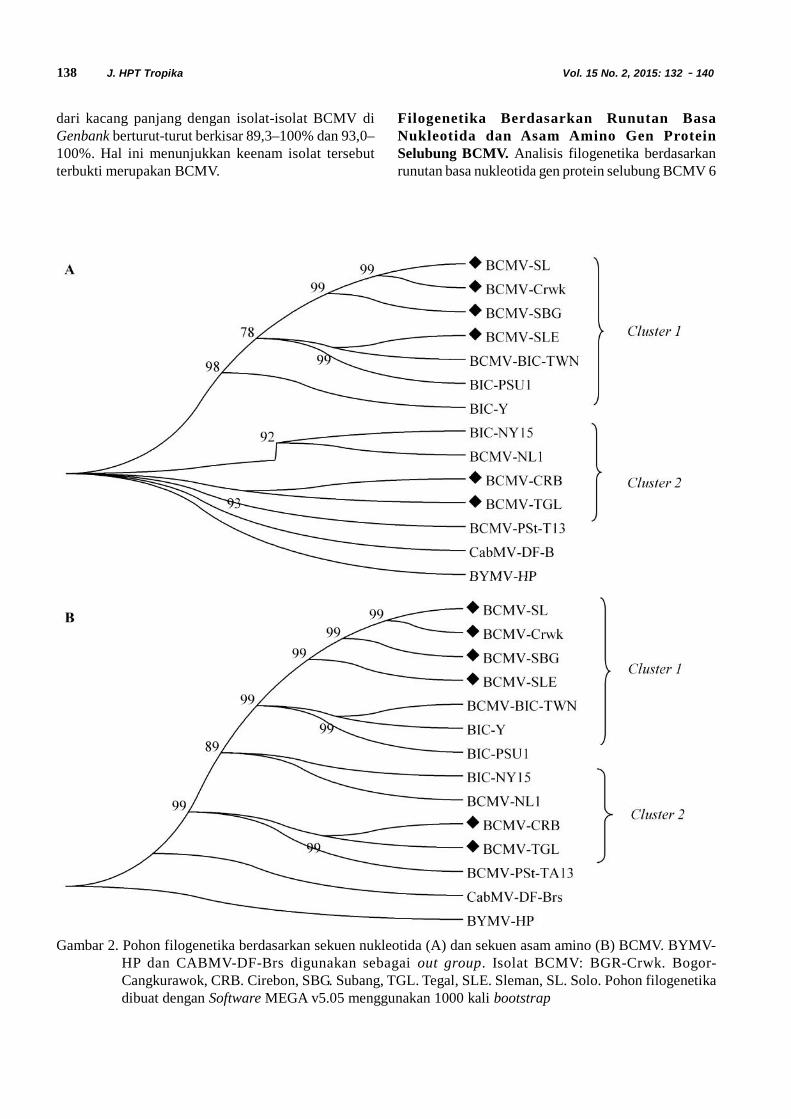
Gambar 2. Pohon filogenetika berdasarkan sekuen nukleotida (A) dan sekuen asam amino (B) BCMV. BYMV-HP dan CABMV-DF-Brs digunakan sebagai out group. Isolat BCMV: BGR-Crwk. Bogor-Cangkurawok, CRB. Cirebon, SBG. Subang, TGL. Tegal, SLE. Sleman, SL. Solo. Pohon filogenetikadibuat dengan Software MEGA v5.05 menggunakan 1000 kali bootstrap
dari kacang panjang dengan isolat-isolat BCMV diGenbank berturut-turut berkisar 89,3–100% dan 93,0–100%. Hal ini menunjukkan keenam isolat tersebutterbukti merupakan BCMV.
Filogenetika Berdasarkan Runutan BasaNukleotida dan Asam Amino Gen ProteinSelubung BCMV. Analisis filogenetika berdasarkanrunutan basa nukleotida gen protein selubung BCMV 6
A
B
BCMV-SL
BCMV-Crwk
BCMV-SBG
BCMV-SLE
BCMV-BIC-TWN
BIC-PSU1
BIC-Y
BIC-NY15
BCMV-NL1
BCMV-CRB
BCMV-TGL
BCMV-PSt-T13
CabMV-DF-B
BYMV-HP
93
92
98
78
99
9999
Cluster 1
Cluster 2
BCMV-SL
BCMV-Crwk
BCMV-SBG
BCMV-SLE
BCMV-BIC-TWN
BIC-Y
BIC-PSU1
BIC-NY15
BCMV-NL1
BCMV-CRB
BCMV-TGL
BCMV-PSt-TA13
CabMV-DF-Brs
BYMV-HP
99
99
89
99
99
99
9999
Cluster 2
Cluster 1

Melinda et al. Identifikasi Molekuler BCMV 139
isolat asal Jawa Barat dan Jawa Tengah terhadap isolatdari negara lain menunjukkan terbentuknya 2 kelompok/cluster. Keenam isolat tersebut terbagi kedalam clusteryang berbeda. Cluster pertama terdiri dari isolat-isolatBogor-Cangkurawok, Subang, Solo dan Sleman, beradadalam satu cluster yang dekat dengan isolat BCMV-BlC Taiwan, BCMV-BlC Y dan BCMV-BlC PSU1.Cluster kedua terdiri dari isolat-isolat Cirebon dan Tegal,berada dalam satu cluster yang dekat dengan isolatBCMV-BlC NY15, BCMV-NL1 dan BCMV-PSt-TA13(Gambar 2a). Hal ini menunjukkan adanya keragamanisolat BCMV ditemukan diantara enam isolat. Sebagaipembanding di luar cluster (outgroup) digunakan Beanyellow mosaic virus-HP (BYMV-HP) dan Cowpeaaphid-borne mosaic virus-DF-Brs (CabMV-DF-Brs).
Analisis filogenetika berdasarkan sekuen asamamino menunjukkan terbentuknya 2 cluster yang samadengan runutan basa nukleotida. Walaupun membentuk2 cluster, namun kekerabatan antar isolat masih sangatdekat dengan isolat BCMV-BlC dari Taiwan danBCMV-BlC Y dari Cina (Gambar 2b). Isolat BCMV-BlC asal Taiwan dan BCMV-BlC asal Cina sebelumnyadilaporkan menyebabkan penyakit yang parah dengangejala daun kuning pada kacang panjang di Taiwan danCina (Zheng et al., 2002; Wang & Fang, 2004).
SIMPULAN
Gejala mosaik kuning pada tanaman kacangpanjang di lapangan sangat bervariasi. Deteksi molekulermenggunakan primer spesifik CP-BCMV berhasilmengamplifikasi pita DNA dari sampel tanaman asalJawa Barat (Bogor; Cangkurawok, Bubulak, Bojong),Subang, dan Cirebon dan asal Jawa Tengah (Solo,Sleman, dan Tegal). Gejala mosaik kuning pada tanamankacang panjang tidak hanya terinfeksi oleh BCMV tetapikemungkinan disebabkan oleh infeksi virus lain. IsolatBCMV asal Bogor-Cangkurawok, Subang, Solo danSleman memiliki homologi dan kekerabatan yang dekatdengan BCMV-BlC dari Taiwan berdasarkan runutanbasa nukleotida dan asam amino. Isolat BCMV asalCirebon dan Tegal memiliki homologi dan kekerabatanyang dekat dengan BCMV-NL1 dari Inggrisberdasarkan runutan basa nukleotida, namunberdasarkan runutan asam amino lebih dekat ke BCMV-BlC Y dari Cina.
SANWACANA
Penelitian terlaksana atas dana pendidikan danpenelitian dari NUFFIC AGRI-4 UNIPA untuk penulisutama dan secara parsial dari dana penelitian IPVDNtahun 2012.
DAFTAR PUSTAKA
Adams MJ, Antoniw JF, & Fauquet CM. 2005.Molecular criteria for genus and speciesdiscrimination within the family Potyviridae. ArchVirol. 150(3): 459–479.
Agrios GN. 2005. Plant Pathology. 5th eds. AcademicPress. New York.
Anggraini S & Hidayat SH. 2014. Sensitivitas metodeserologi dan polymerase chain reaction untukmendeteksi Bean Common Mosaic Potyviruspada kacang panjang. J. Fitopatol. Indones.10(1): 17-22.
Anwar A, Sudarsono, & Ilyas S. 2005. Perbenihansayuran di Indonesia: kondisi terkini dan prospekbisnis benih sayuran. Bul Agron. 33(1): 38–47.
Badan Pusat Statistik. 2011. Produksi Sayuran diIndonesia. [internet]. Jakarta (ID). http://www.bps.go.id/tab_sub/view.php. Diakses 19November 2012.
Bhadramurthy V & Bhat AI. 2009. Biological andmolecular characterization of Bean commonmosaic virus associated with vanilla in India.Indian J. Virol. 20(2): 70–77.
[CABI] Centre for Agriculture and BiosciencesInternational. 2007. Crop ProtectionCompendium. (Serial Online). CABInternational. Wallingford.
Damayanti TA, Alabi OJ, Naidu RA, & Rauf A. 2009.Severe outbreak of a yellow mosaic disease onthe yard long bean in Bogor, West Java. HayatiJ. Biosci. 16(2): 78-82.
Haryanto E, Suhartini T, & Rahayu E. 2010. Budi DayaKacang Panjang. Penebar Swadaya, Jakarta.

140 J. HPT Tropika Vol. 15 No. 2, 2015: 132 - 140
Hall TA. 1999. Bioedit: a user-friendly biologicalsequence alignment editor and analysis programfor Windows 95/98/NT. Nucl. Acids Symp. Ser.41: 95–98.
Lazuardi F. 2005. Perkembangan beberapa penyakitpenting pada kacang panjang (Vigna sinensis L.)di lahan petani sekitar kampus IPB Dramaga,Kabupaten Bogor. Skripsi. Institut PertanianBogor, Bogor.
Melgarejo TA, Lehtonen MT, Fribourg CE, Rannali M,& Valkonen JPT. 2007. Strains of BCMV andBCMNV characterized from lima bean plantsaffected by deforming mosaic disease in Peru.Arch. Virol. 152(10): 1941–1949.
Morales FJ. 1988. Bean common mosaic virus.www.dpvweb.net/dpv/showdpv.php?dpvno=337.Diakses 12 Oktober 2012.
Nurulita S. 2014. Molecular characterization ofBegomovirus infecting yard long bean (Vignaunguiculata subsp. sesquipedalis L.) andconstruction of its specific primers. Tesis. InstitutPertanian Bogor, Bogor.
Pitojo S. 2006. Benih Kacang Panjang. Kanisius,Yogyakarta.
Siregar S. 1996. Pengamatan hama dan penyakit pentingtanaman kacang panjang (Vigna sinensis L.) diKecamatan Ciampea, Kabupaten Bogor JawaBarat. Skripsi. Institut Pertanian Bogor, Bogor.
Wang HL & Fang CC. 2004. Molecular sequencing andanalysis of the viral genomic regions of Blackeyecowpea mosaic virus Taiwan strain. PlantPathol. Bull. 13: 117–126.
Zheng H, Chen J, Chen J, Adams MJ, & Hou M. 2002.Bean common mosaic virus isolates causingdifferent symptoms in asparagus bean in Chinadiffer greatly in the 5’-parts of their genomes.Arch. Virol. 147(6): 1257–1262.
Zhou GC, Wu XY, Zhang YM, Wu P, Wu XZ, Liu LW,Wang Q, Hang YY, Yang JY, Shao ZQ, Wang B,& Chen JQ. 2014. A genomic survey of thirtysoybean-infecting Bean common mosaic virus(BCMV) isolates from China pointed BCMV asa potential threat to soybean production. VirusRes. 191: 125–133.

152 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 152 – 163, September 2015
KETAHANAN BEBERAPA GENOTIPE PADI TERHADAP PENYAKITHAWAR DAUN BAKTERI (XANTHOMONAS ORYZAE PV. ORYZAE)
Wira Hadianto1,2, Lukman Hakim1, & Bakhtiar1
1Program Studi Magister Agroekoteknologi, Program Pasca Sarjana Universitas Syiah KualaJl. Tgk. Hasan Krueng Kalee, No. 3. Kopelma Darussalam, Banda Aceh 23111
2Program Studi Agroteknologi Universitas Teuku Umar Jl. Alue Penyareng, Meulaboh 23615
E-mail: [email protected]
ABSTRACT
The resistance rice genotypes against bacterial leaf blight (Xanthomonas oryzae pv. oryzae). The research was conductedto analyze genotypes of rice that were resistant to bacterial leaf blight (BLB) disease. The experiment was conducted at thescreen house of Faculty of Agriculture, Syiah Kuala University, Banda Aceh from February to October 2014. This experimentwas arranged in randomized complete block design with 73 treatments and 3 replications. IR-BB27 and IR-64 were used ascontrol of resistant and susceptible varieties to BLB, respectively. The result showed that the bacterial isolates tested werevirulent. The longest incubation period found in Limboto, Inpari 10, Situ Patenggang, Sirendeh Semantuk Wayla, Tamboen,Sepasie, Bontok and Arias. Genotypes Inpari 1, Limboto, Tuwoti, Inpari 10, Lekat Rambot Linuet, Rom Mokot, Paki Gajah,Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok, Sirendeh Semantuk Wayla and Sambei showed resistance reaction to bacterialleaf blight disease.
Key words: bacterial leaf blight, rice genotypes, Xanthomonas oryzae pv. oryzae
ABSTRAK
Ketahanan beberapa genotipe padi terhadap penyakit hawar daun bakteri (Xanthomonas oryzae pv. oryzae). Penelitan inibertujuan untuk mendapatkan genotipe padi yang tahan terhadap penyakit hawar daun bakteri (HDB). Penelitian dilaksanakandi screen house Fakultas Pertanian Universitas Syiah Kuala, Banda Aceh, pada bulan Februari sampai Oktober 2014. Rancanganyang digunakan adalah Rancangan Acak Kelompok dengan 73 perlakuan dan 3 ulangan. IR-BB27 digunakan sebagaipembanding tahan dan IR-64 sebagai pembanding rentan terhadap HDB. Hasil penelitian menunjukkan isolat bakteri yangdiuji virulen. Masa inkubasi paling lama dijumpai pada genotipe Limboto, Inpari 10, Situ Patenggang, Sirendeh SemantukWayla, Tamboen, Sepasie, Bontok dan Arias. Padi genotipe IR-BB27, Inpari 1, Limboto, Tuwoti, Inpari 10, Lekat RambotLinuet, Rom Mokot, Paki Gajah, Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok, Sirendeh Semantuk Wayla dan Sambei yangmenunjukan reaksi tahan terhadap HDB.
Kata kunci: genotipe padi, hawar daun bakteri, Xanthomonas oryzae pv. oryzae
PENDAHULUAN
Penyakit hawar daun bakteri (HDB) merupakansalah satu faktor pembatas dalam peningkatan produksipadi. Penyakit HDB disebabkan oleh Xanthomonasoryzae pv. oryzae (Xoo), yang dapat menurunkan hasilpadi 30–40% dan menurunkan mutu beras yangdihasilkan. Penyakit ini menginfeksi pada fase vegetatifhingga fase generatif (Herlina & Silitonga, 2011).Xanthomonas oryzae pv. oryzae (Xoo) menginfeksitanaman dengan cara masuk ke dalam jaringan tanamanmelalui lubang alami seperti hidatoda dan benih yangterkontaminasi (Ou, 1985). Gejala dari serangan Xoomenyebabkan daun padi berubah menjadi kuning pucat,layu, dan kemudian mati (Wahyudi et al., 2011). Salah
satu upaya pengendalian penyakit HDB denganpenggunaan varietas tahan. Varietas tahan lebih amandigunakan karena tidak mencemari lingkungandibandingkan dengan penggunaan pestisida yangmeninggalkan residu. Varietas yang tahan dapatdiperoleh melalui perakitan varietas denganmenggabungkan gen ketahanan pada tetua yang telahberadaptasi dan berdaya hasil tinggi.
Perakitan varietas tahan diawali denganidentifikasi genotipe sebagai tetua untuk pembentukanpopulasi dasar. Penggunaan gen-gen tahan terhadapberbagai cekaman yang dimiliki varietas lokal dalampemuliaan tanaman dapat meningkatkan keunggulanvarietas unggul yang akan dihasilkan (Berthaud et al.,2000). Padi lokal dapat dijadikan sebagai tetua untuk

Hadianto et al. Ketahanan Beberapa Genotipe Padi 153
memperoleh varietas unggul yang mempunyai sifatmorfologi dan fisiologi yang lebih baik, seperti gabahhampa lebih sedikit dan lebih tahan terhadap hama danpenyakit utama (Abdullah, 2009). Dengan demikian,varietas lokal yang dibudidayakan secara turun temurunoleh petani merupakan salah satu sumber daya genetikbagi perakitan varietas.
Provinsi Aceh memiliki banyak varietas lokal yangmasih dibudidayakan petani terutama di daerah AcehBarat-Selatan. Bakhtiar et al. (2011) melaporkan bahwavarietas lokal padi Aceh sangat beragam. Namundemikian, potensi varietas tersebut sebagai sumber genketahanan terhadap HDB belum dievaluasi.
Berdasarkan permasalahan di atas diperlukansuatu penelitian untuk mengevaluasi ketahanan beberapagenotipe padi terhadap penyakit hawar daun bakteri.Tujuan penelitian adalah untuk mengetahui danmendapatkan genotipe padi yang tahan terhadappenyakit HDB.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan di kebunpercobaan Fakultas Pertanian, Universitas Syiah Kuala,Banda Aceh, bulan Februari sampai dengan Oktober2014. Rancangan yang digunakan adalah rancanganacak kelompok (RAK) dengan 73 genotipe padi sebagaiperlakuan dan 3 ulangan. Genotipe IR-BB27 digunakansebagai pembanding tahan dan genotipe IR-64 sebagaipembanding rentan terhadap penyakit HDB.
Bahan Tanam. Bahan tanam yang digunakan terdiridari 71 genotipe tanaman padi, yang terdiri atas varietasunggul nasional sebanyak 16 genotipe, varietas lokalAceh tipe ketan sebanyak 11 genotipe dan varietas lokalAceh tipe bukan ketan sebanyak 44 genotipe.
Persiapan Benih. Benih terlebih dahulu direndamdengan air bersih selama satu malam untuk prosesimbibisi, kemudian benih dikecambahkan selama dua haridengan cara dibalut dengan kain lembab. Setelahberkecambah benih tersebut dipindahkan ke wadahpersemaian yang telah disiapkan. Media tanam adalahtanah alluvial yang dimasukkan dalam bak plastik dandiberikan air, selanjutnya diaduk sampai merata.
Persiapan Inokulum. Daun padi varietas Ciherangyang terinfeksi HDB dikumpulkan dari Gampong BlangKecamatan Blang Bintang Kabupaten Aceh Besar, padamusim tanam 2013/2014. Persiapan inokulum mengikutiprosedur yang dilakukan Shehzad et al. (2012), daunterinfeksi HDB dibilas dengan air, kemudian dipotong-
potong 5–10 mm dan disterilisasi dengan etanol 70%selama 10 detik dan disimpan pada cawan Petri selama20 menit supaya bacterial ooze-keluar. Media NutrientAgar (NA) digunakan untuk mengisolasi inokulum yangdiinkubasikan selama 3 hari sampai muncul koloni kuning.
Biakan murni bakteri disimpan pada media agarmiring pada suhu 20–25 oC agar terhindar darikontaminasi. Pada saat akan digunakan, koloni bakteridiukur konsentrasinya setara dengan 106 sel hidup/ml.Inokulum yang disiapkan dalam media NA 20 g/1akuades, selanjutnya dimasukkan ke dalam Erlenmeyeryang dibalut dengan aluminium foil, kemudian siapdiinokulasi pada daun padi.
Uji Hipersensitivitas. Uji hipersensitivitas isolat bakteriterhadap tanaman tembakau dilakukan menurut Zou etal. (2006). Koloni hawar daun bakteri hasil isolasidengan kerapatan ±106 sel/ml dalam kultur cairdisuntikkan ke tulang daun tanaman tembakau. Sebagaikontrol, pada tulang daun tembakau lainnya disuntikdengan akuades. Pengamatan gejala penyakit dilakukanhingga 48 jam setelah penyuntikan dan difoto.
Penanaman dan Inokulasi Bakteri. Semua bahantanam ditanam masing-masing sebanyak 5 tanamandalam 1 ulangan. Penanaman dilakukan dalam bakplastik dan ditempatkan dalam screen house FakultasPertanian Unsyiah, Banda Aceh. Pengendalian hamamenggunakan insektisida karbofuran 3G dengan takaran20 kg formulasi/ha. Untuk mendorong perkembanganpenyakit HDB, tanaman diberi pupuk N dengan takaran250 kg urea/ha. Agar bakteri tidak dihadapkan pada suhuyang terlalu panas, inokulasi dilakukan menjelang sore,antara pukul 16.00-18.00. Inokulasi dilakukan dengancara pengguntingan daun padi untuk pelukaan sebagaijalan masuk bagi infeksi bakteri. Pengguntingandilakukan 3–5 cm dari ujung daun, menggunakan guntingyang telah dicelupkan ke dalam Erlenmeyer berisisuspensi isolat bakteri dengan kerapatan 106 sel hidup/ml. Inokulasi tanaman dengan isolat bakteriXanthomonas oryzae pv. oryzae dilakukan satu kali,yaitu pada saat tanaman berumur 42 HST atau memasukifase vegetatif aktif.
Suhu dan Kelembaban. Diamati setiap hari mulaiinokulasi penyakit sampai 14 hari setelah inokulasi.
Masa inkubasi. Masa inkubasi dihitung dari saatinokulasi sampai munculnya gejala penyakit yangdilakukan setiap hari hingga 14 hari setelah inokulasidalam satuan hari.

154 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163
Panjang lesio. Panjang lesio diamati 14 hari setelahinokulasi dengan cara mengukur panjang lesio dari limadaun per tanaman, kemudian dihitung rata-ratanya dalamsatuan cm.
HASIL DAN PEMBAHASAN
Uji Hipersensitivitas. Uji hipersensitivitas dilakukanuntuk mengetahui patogenesitas patogen ataukemampuan patogen dalam menyebabkan penyakit.Tanaman tembakau digunakan sebagai tanamanindikator untuk uji hipersensitif (Suswanto et al., 1996).Bakteri Xanthomonas oryzae pv. oryzae (Xoo) yangdisuntik pada tulang daun tanaman tembakau mampumenginduksi reaksi hipersensitif. Daun tembakau menjadikecoklatan pada area masuknya bakteri. Reaksi ini palingjelas teramati 48 jam setelah penyuntikan. Sebagaikontrol negatif yang disuntik dengan akuades tidakmenunjukkan reaksi serupa (Gambar 1).
Respon hipersensitif diartikan sebagai reaksipertahanan yang cepat dari tanaman menghadapipatogen yang tidak kompatibel disertai dengan kematiansel yang cepat pada jaringan di daerah yang diinjeksisuspensi bakteri (Klement et al., 1990). Dengandemikian bakteri yang diisolasi dari lapangan tersebutdapat digunakan sebagai inokulum untuk evaluasiketahanan terhadap HDB dari 73 genotipe padi yangakan diuji.
Suhu. Pada penelitian ini, suhu udara dicatat setiap harisejak inokulasi sampai pengamatan gejala penyakitterakhir, yaitu 14 HSI. Suhu minimum 22 oC danmaksimum 34 oC dengan rata-rata 27,04 oC selamapenelitian. Menurut Devadath (1989) perkembangangejala HDB akan lebih cepat pada suhu tinggi dibandingsuhu rendah dimana suhu optimal berkisar antara 24,3-34 oC. Selanjutnya, Wiyono (2007) menyebutkan bakteripenyebab penyakit kresek pada padi menghendaki suhu
optimum pada 30ºC. Hal ini menunjukkan bahwa suhuselama percobaan mendukung perkembangan penyakitHDB.
Kelembaban. Kelembaban udara yang cukup tinggiakan meningkatkan pertumbuhan penyakit hawar daunbakteri. Pada penelitian ini, kelembaban udara diamatisetiap hari sejak inokulasi sampai pengamatan gejalapenyakit terakhir pada 14 HSI. Kelembaban udaraselama pengamatan gejala penyakit berada pada kisaran43,60–68,23% dengan rata-rata 53,18%. Menurut Sudiret al. (2012), kelembaban yang tinggi dapatmempercepat perkembangan penyakit ini. Hal inimenunjukkan bahwa kondisi kelembaban pada penelitianini dapat meningkatkan infeksi penyakit HDB yangdiinokulasi pada daun padi.
Masa Inkubasi. Hasil pengamatan terhadap masainkubasi bakteri Xoo pada beberapa genotipe varietasunggul nasional disajikan pada Tabel 1. Gejala penyakithawar daun pada varietas yang diuji ditandai denganmunculnya gejala hijau kelabu selanjutnya menguningberdekatan dengan ujung daun yang dipotong. Gejalapenyakit tersebut terus memanjang di sepanjang tepi daun(Gambar 2).
Masa inkubasi bakteri hawar daun bakteri pada16 genotipe varietas unggul nasional yang diuji berkisar4 sampai 10,67 hari setelah inokulasi. Gejala penyakitpaling cepat muncul dijumpai pada varietas Inpari 7 danIpari 16, yaitu 4,67 hari setelah inokulasi. Gejala penyakitpada IR-64 sebagai pembanding rentan mulai munculpada 8,67 hari setelah inokulasi. Gejala penyakit palinglama muncul pada genotipe unggul dijumpai padaLimboto, Inpari 10, dan Situ Patenggang yang tidak jauhberbeda dibandingkan masa inkubasi pada pembandingtahan yaitu galur IR-BB27, yaitu 12 hari setelah inokulasi(Tabel 1).
A B
Gambar 1. Hasil uji hipersensitivitas setelah disuntik membentuk lesio pada daun tembakau yang terinfeksi bakteriXoo (A), kontrol negatif (B)

Hadianto et al. Ketahanan Beberapa Genotipe Padi 155
Hasil pengamatan terhadap masa inkubasi bakteriXoo pada beberapa genotipe padi lokal Aceh tipe ketandisajikan pada Tabel 2. Masa inkubasi bakteri hawardaun bakteri pada 11 genotipe tipe ketan yang diujiberkisar 3 sampai 9 hari setelah inokulasi. Gejalapenyakit paling cepat muncul dijumpai pada genotipePulut Merah, yaitu 3,33 hari setelah inokulasi. Gejalapenyakit paling lama dijumpai pada genotipe LekatRambot Lineut yaitu muncul 9,33 hari setelah inokulasitetapi lebih cepat dibandingkan gejala penyakit pada IR-BB27 sebagai pembanding tahan (Tabel 2).
Hasil pengamatan terhadap periode inkubasibakteri Xoo pada beberapa genotipe padi lokal Acehtipe bukan ketan disajikan pada Tabel 3. Periode inkubasibakteri hawar daun bakteri pada 44 genotipe padi lokalbukan ketan yang diuji berkisar 3 sampai 11 hari setelah
inokulasi. Gejala penyakit paling cepat muncul dijumpaipada genotipe Pade Merah Lamtuba, yaitu 3,67 harisetelah inokulasi. Gejala penyakit paling lama munculdijumpai pada Sirendeh Semantuk Wayla yaitu 11 harisetelah inokulasi hampir sama dengan gejala penyakitpada IR-BB27 sebagai pembanding tahan (Tabel 3).
Hasil penelitian terhadap masa inkubasi bakteriXoo pada 71 genotipe yang diuji penyakit HDB terlihatbahwa genotipe varietas unggul nasional dengan masainkubasi tercepat dijumpai pada varietas Inpari 16 danInpari 7 (4,67 hari). Genotipe padi lokal Aceh tipe ketandengan masa inkubasi tercepat dijumpai pada genotipePulut Merah (3,33 hari) dan genotipe padi lokal Acehtipe bukan ketan dengan masa inkubasi tercepat padagenotipe Pade Merah Lamtuba (3,67 hari). Gejalapenyakit pada IR-64 sebagai pembanding rentan mulai
Gambar 2. Gejala HDB pada daun padi yang diinokulasi dengan Xoo (kiri) dan tidak memperlihatkan gejala padadaun padi tanpa Xoo (kanan)
Tabel 1. Masa inkubasi pada varietas unggul nasional
No Genotipe unggul Masa inkubasi (hari)
1 IR-BB27 (kontrol tahan) 12,00 ± 1,002 Limboto 10,67 ± 0,583 Inpari 10 10,33 ± 2,084 Situ Patenggang 10,00 ± 1,735 IPB 4 S 9,67 ± 1,536 Situ Bagendit 9,67 ± 1,537 Ciherang 9,67 ± 2,528 Inpari 1 9,33 ± 1,539 Inpari 19 9,33 ± 2,0810 Cirata 8,67 ± 1,1511 IPB 3S 8,67 ± 1,1512 Tuwoti 8,67 ± 0,5813 Inpago 6 7,67 ± 0,5814 Danau Gaung 6,33 ± 1,5315 Kencana Bali 5,00 ± 1,7316 Inpari 16 4,67 ± 1,5317 Inpari 7 4,67 ± 1,1518 IR-64 (kontrol rentan) 8,67 ± 1,53

156 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163
muncul pada 8.67 hari setelah inokulasi. Hal inimenunjukkan bahwa genotipe tersebut rentan terhadappenyakit HDB yang terlihat dari cepat terinfeksi olehpenyakit yang diinokulasi dengan Xoo pada daun padi.
Masa inkubasi bakteri Xoo terlama dijumpai padavarietas pembanding IR-BB27 yaitu 12 hari setelahinokulasi, sedangkan pada varietas unggul nasionaldijumpai pada Limboto, Inpari 10, dan Situ Patenggang.Genotipe padi lokal Aceh tipe ketan dengan masainkubasi terlama dijumpai pada genotipe Lekat RambotLineut (9,33 hsi) dan genotipe padi lokal Aceh tipe bukanketan dengan masa inkubasi terlama pada genotipeSirendeh Semantuk Wayla (11 hsi). Hal ini menunjukkanbahwa genotipe tersebut memiliki ketahanan terhadapinfeksi penyakit HDB dapat dilihat lamanya munculgejala serangan. Masa inkubasi penyakit menjadi salahsatu faktor yang menentukan virulensi suatu patogen,demikan pula sebaliknya masa inkubasi penyakit yanglama menandakan suatu varietas tahan terhadap suatupatogen (Rahim et al., 2012). Genotipe tahan memilikigen ketahanan terhadap ras/patotipe Xoo yangdikendalikan oleh gen R mayor (Nino-Liu et al., 2006).
Masa inkubasi bakteri Xoo pada fase vegetatifdari 71 genotipe yang diuji dengan dibagi tiga kriteriayaitu lama, sedang sampai cepat (Tabel 4). IR-64tergolong kedalam genotipe dengan masa inkubasipenyakit HDB sedang, dengan demikian, varietas IR-64 cocok digunakan sebagai kontrol rentan. IR-BB27tergolong ke dalam genotipe dengan masa inkubasipenyakit HDB lama sehingga IR-BB27 cocok digunakansebagai kontrol tahan berdasarkan muncul gejalapenyakit terhadap HDB.
Panjang Lesio. Hasil pengamatan terhadap panjanglesio bakteri Xoo pada beberapa genotipe varietas unggul
nasional disajikan pada Tabel 5. Panjang lesio yaitu reaksigejala layu pada ujung daun dan muncul bercak kelabupada tepi daun yang kemudian mulai berkerut seluruhbagian daun mulai berwarna putih kecoklatan dan kuningpucat menuju pangkal daun. Panjang lesio bakteri hawardaun bakteri pada 17 genotipe varietas unggul nasionalyang diuji berkisar 1,42 sampai 19,09 cm. Panjang lesioterpendek dijumpai pada pembanding tahan galur IR-BB27 dan varietas Inpari 1, yaitu 1,42 cm. Panjang lesioterpanjang dijumpai pada varietas Inpari 16, yaitu 19,09cm (Tabel 5). Panjang lesio pada varietas IR-64 sebagaipembanding rentan yaitu 7,43 cm.
Panjang lesio pada varietas Inpari 1, Limboto,Tuwoti, dan Inpari 10 berbeda tidak nyata dibandingkanpanjang lesio pada genotipe pembanding IR-BB27 (Tabel5). Hal ini menunjukkan bahwa varietas tersebut tahanterhadap HDB. Panjang lesio pada varietas DanauGaung, IPB 4 S, Situ Bagendit, Inpago, Cirata dan IPB3S berbeda nyata dibandingkan panjang lesio padagenotipe pembanding IR-BB27 (Tabel 5). Hal inimenunjukkan bahwa varietas tersebut moderat tahanterhadap HDB. Panjang lesio pada varietas Ciherang,Situ Patenggang, Kencana Bali, Inpari 19, Inpari 7 danInpari 16 berbeda sangat nyata dibandingkan panjanglesio pada genotipe pembanding IR-BB27 (Tabel 5). Halini menunjukkan bahwa varietas tersebut rentan terhadapHDB.
Panjang lesio bakteri Xoo pada padi lokal Acehtipe ketan disajikan pada Tabel 6. Panjang lesio bakterihawar daun bakteri pada 11 genotipe tipe ketan yangdiuji berkisar 3,06 sampai 19,39 cm. Panjang lesioterpendek dijumpai pada genotipe Lekat Rambot Lineut,yaitu 3,6 cm. Panjang lesio terpanjang dijumpai padagenotipe Lekat Tuleng, yaitu 19,93 cm. Panjang lesio
Tabel 2. Masa inkubasi pada genotipe padi lokal Aceh tipe ketan
No Genotipe ketan Masa inkubasi (hari)
1 IR-BB27 12,00 ± 1,002 Leukat Rambot Lineut 9,33 ± 0,583 Pulut Hitam 9,00 ± 1,004 Ketan Putih 8,67 ± 1,535 Lekat Singke 8,00 ± 0,006 Lekat jerajak Lango 8,00 ± 1,007 Lekat Adang 8,00 ± 1,008 Lekat Alahu 7,67 ± 2,319 Pulut Simanik 6,33 ± 0,5810 Lekat Tuleng 4,67 ± 2,0811 Lekat Kumbob 4,33 ± 0,5812 Pulut Merah 3,33 ± 0,58
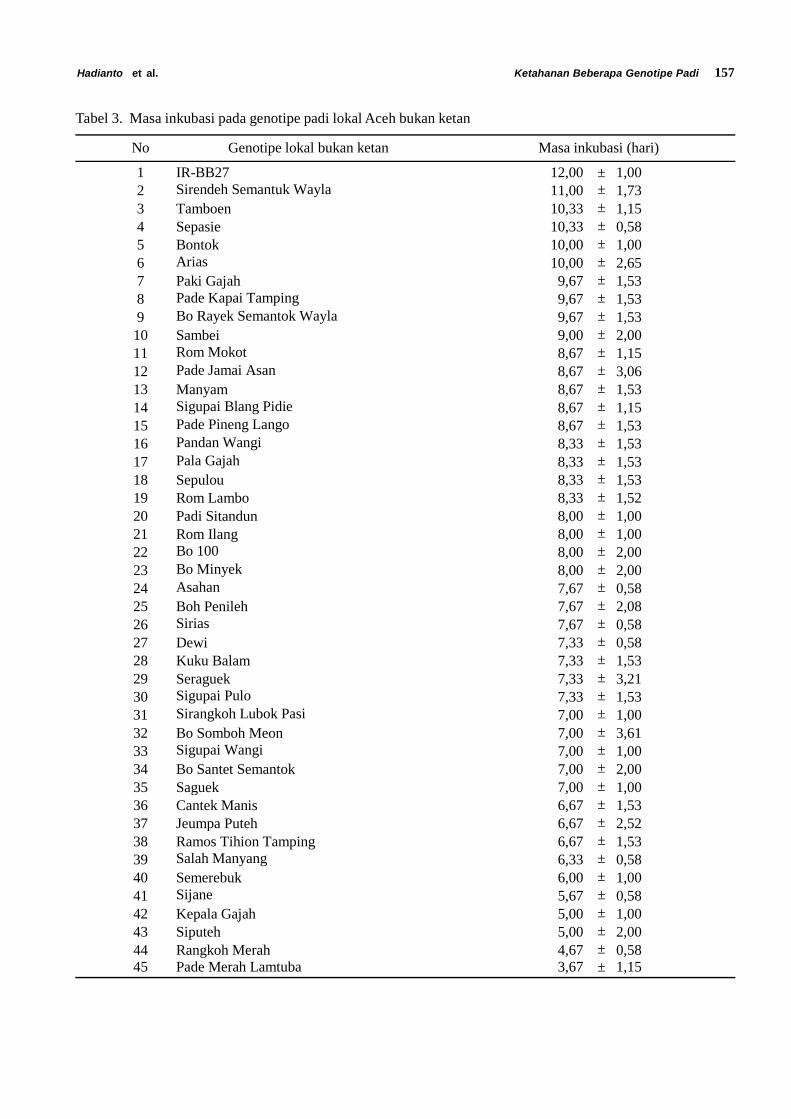
Hadianto et al. Ketahanan Beberapa Genotipe Padi 157
Tabel 3. Masa inkubasi pada genotipe padi lokal Aceh bukan ketan
No Genotipe lokal bukan ketan Masa inkubasi (hari)
1 IR-BB27 12,00 ± 1,002 Sirendeh Semantuk Wayla 11,00 ± 1,733 Tamboen 10,33 ± 1,154 Sepasie 10,33 ± 0,585 Bontok 10,00 ± 1,006 Arias 10,00 ± 2,657 Paki Gajah 9,67 ± 1,538 Pade Kapai Tamping 9,67 ± 1,539 Bo Rayek Semantok Wayla 9,67 ± 1,5310 Sambei 9,00 ± 2,0011 Rom Mokot 8,67 ± 1,1512 Pade Jamai Asan 8,67 ± 3,0613 Manyam 8,67 ± 1,5314 Sigupai Blang Pidie 8,67 ± 1,1515 Pade Pineng Lango 8,67 ± 1,5316 Pandan Wangi 8,33 ± 1,5317 Pala Gajah 8,33 ± 1,5318 Sepulou 8,33 ± 1,5319 Rom Lambo 8,33 ± 1,5220 Padi Sitandun 8,00 ± 1,0021 Rom Ilang 8,00 ± 1,0022 Bo 100 8,00 ± 2,0023 Bo Minyek 8,00 ± 2,0024 Asahan 7,67 ± 0,5825 Boh Penileh 7,67 ± 2,0826 Sirias 7,67 ± 0,5827 Dewi 7,33 ± 0,5828 Kuku Balam 7,33 ± 1,5329 Seraguek 7,33 ± 3,2130 Sigupai Pulo 7,33 ± 1,5331 Sirangkoh Lubok Pasi 7,00 ± 1,0032 Bo Somboh Meon 7,00 ± 3,6133 Sigupai Wangi 7,00 ± 1,0034 Bo Santet Semantok 7,00 ± 2,0035 Saguek 7,00 ± 1,0036 Cantek Manis 6,67 ± 1,5337 Jeumpa Puteh 6,67 ± 2,5238 Ramos Tihion Tamping 6,67 ± 1,5339 Salah Manyang 6,33 ± 0,5840 Semerebuk 6,00 ± 1,0041 Sijane 5,67 ± 0,5842 Kepala Gajah 5,00 ± 1,0043 Siputeh 5,00 ± 2,0044 Rangkoh Merah 4,67 ± 0,5845 Pade Merah Lamtuba 3,67 ± 1,15
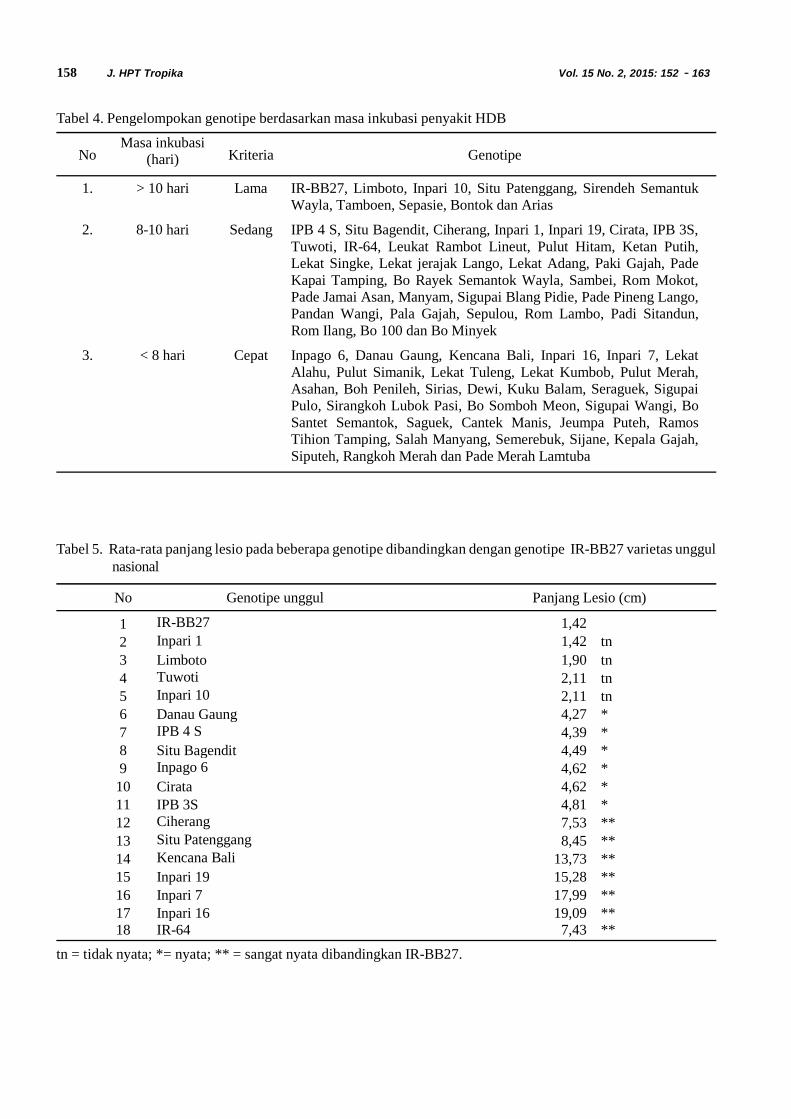
158 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163
Tabel 4. Pengelompokan genotipe berdasarkan masa inkubasi penyakit HDB
NoMasa inkubasi
(hari) Kriteria Genotipe
1. > 10 hari Lama IR-BB27, Limboto, Inpari 10, Situ Patenggang, Sirendeh SemantukWayla, Tamboen, Sepasie, Bontok dan Arias
2. 8-10 hari Sedang IPB 4 S, Situ Bagendit, Ciherang, Inpari 1, Inpari 19, Cirata, IPB 3S,Tuwoti, IR-64, Leukat Rambot Lineut, Pulut Hitam, Ketan Putih,Lekat Singke, Lekat jerajak Lango, Lekat Adang, Paki Gajah, PadeKapai Tamping, Bo Rayek Semantok Wayla, Sambei, Rom Mokot,Pade Jamai Asan, Manyam, Sigupai Blang Pidie, Pade Pineng Lango,Pandan Wangi, Pala Gajah, Sepulou, Rom Lambo, Padi Sitandun,Rom Ilang, Bo 100 dan Bo Minyek
3. < 8 hari Cepat Inpago 6, Danau Gaung, Kencana Bali, Inpari 16, Inpari 7, LekatAlahu, Pulut Simanik, Lekat Tuleng, Lekat Kumbob, Pulut Merah,Asahan, Boh Penileh, Sirias, Dewi, Kuku Balam, Seraguek, SigupaiPulo, Sirangkoh Lubok Pasi, Bo Somboh Meon, Sigupai Wangi, BoSantet Semantok, Saguek, Cantek Manis, Jeumpa Puteh, RamosTihion Tamping, Salah Manyang, Semerebuk, Sijane, Kepala Gajah,Siputeh, Rangkoh Merah dan Pade Merah Lamtuba
Tabel 5. Rata-rata panjang lesio pada beberapa genotipe dibandingkan dengan genotipe IR-BB27 varietas unggulnasional
No Genotipe unggul Panjang Lesio (cm)
1 IR-BB27 1,422 Inpari 1 1,42 tn3 Limboto 1,90 tn4 Tuwoti 2,11 tn5 Inpari 10 2,11 tn6 Danau Gaung 4,27 *7 IPB 4 S 4,39 *8 Situ Bagendit 4,49 *9 Inpago 6 4,62 *10 Cirata 4,62 *11 IPB 3S 4,81 *12 Ciherang 7,53 **13 Situ Patenggang 8,45 **14 Kencana Bali 13,73 **15 Inpari 19 15,28 **16 Inpari 7 17,99 **17 Inpari 16 19,09 **18 IR-64 7,43 **
tn = tidak nyata; *= nyata; ** = sangat nyata dibandingkan IR-BB27.

Hadianto et al. Ketahanan Beberapa Genotipe Padi 159
pada galur IR-BB27 sebagai pembanding tahan yaitu1,42 cm (Tabel 6).
Panjang lesio pada genotipe padi lokal Aceh tipeketan Leukat Rambot Lineut tidak berbeda nyatadibandingkan panjang lesio pada genotipe pembandingIR-BB27 (Tabel 6). Hal ini menunjukkan bahwa genotipetersebut tahan terhadap HDB. Panjang lesio padagenotipe padi lokal Aceh tipe ketan Lekat Adang, Lekatjerajak Lango, Pulut Hitam, Lekat Singke, Lekat Alahu,Ketan Putih, Pulut Merah, Pulut Simanik, Lekat Kumbobdan Lekat Tuleng berbeda sangat nyata dibandingkanpanjang lesio pada genotipe pembanding IR-BB27 (Tabel6). Hal ini menunjukkan bahwa genotipe tersebut rentanterhadap HDB.
Hasil pengamatan terhadap panjang lesio bakteriXoo pada beberapa genotipe padi lokal Aceh tipe bukanketan disajikan pada Tabel 7. Panjang lesio bakteri hawardaun bakteri pada 44 genotipe padi lokal bukan ketanyang diuji berkisar 1,91 sampai 14,74 cm. Panjang lesioterpendek dijumpai pada genotipe Rom Mokot, yaitu 1,91cm. Lesio terpanjang dijumpai pada Siputeh, yaitu 14,74cm (Tabel 7). Panjang lesio pada galur IR-BB27 sebagaipembanding tahan yaitu 1,42 cm.
Panjang lesio pada genotipe Rom Mokot, PakiGajah, Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok,Sirendeh Semantuk Wayla dan Sambei berbeda tidaknyata dibandingkan panjang lesio pada genotipepembanding IR-BB27 (Tabel 7). Hal ini menunjukkanbahwa genotipe tersebut tahan terhadap HDB. Panjanglesio pada genotipe Pade Jamai Asan, Salah Manyang,Arias, Pandan Wangi dan Pade Merah Lamtuba berbedanyata dibandingkan panjang lesio pada genotipe
pembanding IR-BB27 (Tabel 7). Hal ini menunjukkanbahwa genotipe tersebut moderat tahan terhadap HDB.
Panjang lesio pada genotipe Bo Somboh Meon,Sigupai Wangi, Kuku Balam, Kepala Gajah, Asahan,Manyam, Boh Penileh, Saguek, Dewi, Jeumpa Puteh,Sigupai Blang Pidie, Pala Gajah, Rangkoh Merah, PadeKapai Tamping, Sitandun, Sirangkoh Lubok Pasi,Sepulou, Pade Pineng Lango, Seraguek, Semerebuk,Ramos Tihion Tamping, Sijane, Rom Lambo, Sirias,Sigupai Pulo, Bo Santet Semantok, Bo Rayek SemantokWayla, Cantek Manis, Rom Ilang dan Siputeh berbedasangat nyata dibandingkan panjang lesio pada genotipepembanding IR-BB27 (Tabel 7). Hal ini menunjukkanbahwa genotipe tersebut rentan terhadap HDB.
Reaksi 71 genotipe yang diuji dengan bakteriXanthomonas oryzae pv. oryzae pada fase vegetatifsangat bervariasi mulai dari tahan, moderat tahan sampairentan (Tabel 8). IR-64 menunjukkan reaksi rentan,sedangkan IR-BB27 menunjukkan reaksi tahanterhadap penyakit HDB. Dengan demikian, varietas IR-64 cocok digunakan sebagai kontrol rentan dan IR-BB27cocok digunakan sebagai kontrol tahan terhadap HDB.
Pada penelitian ini, ditemukan empat belasgenotipe yang tahan terhadap bakteri Xoo berdasarkanpanjang bercak (Ogawa et al., 1988). Bercak pada daundari genotipe tersebut tidak berkembang dan hanyakering berwarna coklat tua pada ujung daun denganpanjang lesio kurang dari 3 cm. Genotipe yang tahanterhadap HDB adalah Inpari 1, Limboto, Tuwoti, Inpari10, Lekat Rambot Linuet, Rom Mokot, Paki Gajah,Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok, SirendehSemantuk Wayla dan Sambei (Tabel 8). Genotipe
Tabel 6. Rata-rata panjang lesio pada beberapa genotipe dibandingkan dengan genotipe IR-BB27 pada genotipepadi lokal Aceh tipe ketan
No Genotipe ketan Panjang lesio (cm)
1 IR-BB27 1,422 Leukat Rambot Lineut 3,06 tn3 Lekat Adang 5,39 **4 Lekat jerajak Lango 6,45 **5 Pulut Hitam 6,75 **6 Lekat Singke 8,40 **7 Lekat Alahu 8,78 **8 Ketan Putih 9,10 **9 Pulut Merah 10,13 **10 Pulut Simanik 10,82 **11 Lekat Kumbob 17,00 **12 Lekat Tuleng 19,93 **
tn = tidak nyata; ** = sangat nyata dibandingkan IR-BB27

160 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163
Tabel 7. Rata-rata panjang lesio pada beberapa genotipe dibandingkan dengan genotipe IR-BB27 padi lokal bukanketan
No Genotipe lokal bukan ketan Panjang lesio (cm)
1 IR-BB27 1,422 Rom Mokot 1,91 tn3 Paki Gajah 2,20 tn4 Tamboen 2,26 tn5 Bo 100 2,36 tn6 Sepasie 2,56 tn7 Bo Minyek 2,94 tn8 Bontok 2,99 tn9 Sirendeh Semantuk Wayla 3,03 tn10 Sambei 3,20 tn11 Pade Jamai Asan 4,00 *12 Salah Manyang 4,37 *13 Arias 4,53 *14 Pandan Wangi 4,68 *15 Pade Merah Lamtuba 4,80 *16 Bo Somboh Meon 5,41 **17 Sigupai Wangi 5,45 **18 Kuku Balam 5,96 **19 Kepala Gajah 6,22 **20 Asahan 6,37 **21 Manyam 6,58 **22 Boh Penileh 6,71 **23 Saguek 6,71 **24 Dewi 6,72 **25 Jeumpa Puteh 6,90 **26 Sigupai Blang Pidie 6,99 **27 Pala Gajah 7,05 **28 Rangkoh Merah 7,14 **29 Pade Kapai Tamping 7,15 **30 Padi Sitandun 7,16 **31 Sirangkoh Lubok Pasi 7,37 **32 Sepulou 7,60 **33 Pade Pineng Lango 7,63 **34 Seraguek 7,87 **35 Semerebuk 8,29 **36 Ramos Tihion Tamping 8,70 **37 Sijane 8,89 **38 Rom Lambo 8,89 **39 Sirias 8,92 **40 Sigupai Pulo 8,97 **41 Bo Santet Semantok 9,01 **42 Bo Rayek Semantok Wayla 9,25 **43 Cantek Manis 9,25 **44 Rom Ilang 9,36 **45 Siputeh 14,74 **
tn = tidak nyata; * = nyata; ** = sangat nyata dibandingkan IR-BB27.

Hadianto et al. Ketahanan Beberapa Genotipe Padi 161
Tabel 8. Pengelompokan ketahanan genotipe padi terhadap Xanthomonas oryzae pv. oryzae pada fase vegetatifberdasarkan panjang lesio
No Kriteria Genotipe
1. Tahan IR-BB27, Inpari 1, Limboto, Tuwoti, Inpari 10, Lekat Rambot Linuet, Rom Mokot, PakiGajah, Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok, Sirendeh Semantuk Wayla danSambei
2. ModeratTahan
Danau Gaung, IPB 4S, Situ Bagedit, Inpago, Cirata, IPB 3S, Pade Jamai Asan, SalahManyang, Arias, Pandan Wangi dan Pade Merah Lamtuba.
3. Rentan Inapri 16, Ciherang, Situ Patenggang, Kencana Bali, Inpari 19, Inpari 7, IR-64, LekatAlahu, Lekat Singke, Pulut Hitam, Pulut Simanik, Pulut Merah, Lekat Tuleng, LekatKumbob, Lekat jerajak Lango, Lekat Adang, Ketan Putih, Bo Somboh Meon, SigupaiWangi, Kuku Balam, Kepala Gajah, Asahan, Manyam, Boh Penileh, Saguek, Dewi,Jeumpa Puteh, Sigupai Blang Pidie, Pala Gajah, Rangkoh Merah, Pade Kapai Tamping,Padi Sitandun, Sirangkoh Lubok Pasi, Sepulou, Pade Pineng Lango, Seraguek,Semerebuk, Ramos Tihion Tamping, Sijane, Rom Lambo, Sirias, Sigupai Pulo, BoSantet Semantok, Bo Rayek Semantok Wayla, Cantek Manis, Rom Ilang dan Siputeh
tersebut memiliki gen-gen ketahanan terhadap penyakitHDB dapat digunakan untuk membentuk varietas ungguldan tahan penyakit HDB melalui pemuliaan tanaman.Sifat unggul spesifik yang dimiliki padi lokal perludiinkorporasikan ke dalam genom varietas unggul agarmemiliki sifat unggul yang unik. Varietas lokal dengansifat-sifat unggul perlu dilestarikan sebagai aset sumberdaya genetik nasional dan dimanfaatkan dalam programpemuliaan (Sitaresmi et al., 2013).
Pada penelitian ini, dijumpai 11 genotipe yangmoderat tahan menunjukkan infeksi dan gejala layu padaujung daun dan warna putih kecoklatan dengan panjanglesio mencapai 3-6 cm. Diperoleh juga 48 genotipe yangrentan terhadap penyakit HDB. Reaksi gejala layu padaujung daun dan bercak kelabu pada tepi daun genotipetersebut sudah dimulai 3 hari setelah inokulasi, kemudiandaun mulai berkerut pada 14 hari setelah inokulasiseluruh bagian daun mulai berwarna putih kecoklatandan kuning pucat menuju pangkal daun. Pada genotipeyang menunjukkan gejala moderat tahan dan rentanterhadap HDB, patogen sangat cepat berkembangterutama pada keadaan lembab. Penyakit HDB inimampu menghasilkan ras baru, sehingga seringmenyulitkan pengendalian dengan varietas tahan.Gnanamanickam et al. (1999) menyatakan bahwapenyakit HDB merupakan penyakit yang menginfeksisecara sistemik dengan gejala berupa bercak berwarnaabu abu putih di sepanjang tulang daun.
Patogen masuk melalui hidatoda. Luka pada dauntanaman juga merupakan jalan masuk yang potensial
bagi patogen. Kebanyakan infeksi yang berhasil terjadimelalui luka dibandingkan dengan yang melalui hidatoda,dan luka yang masih baru lebih kondusif bagi patogendibandingkan dengan luka yang sudah lama. Panjanglesio pada tanaman padi dipengaruhi oleh sinar mataharidan suhu selama perkembangan penyakit, latar belakanggenetik dari galur yang digunakan, konsentrasi inokulum,dan virulensi (Ogawa et al., 1988).
SIMPULAN
Masa inkubasi paling lama terdapat pada genotipeIR-BB27, Limboto, Inpari 10, Situ Patenggang, SirendehSemantuk Wayla, Tamboen, Sepasie, Bontok dan Arias.Sedangkan masa inkubasi tercepat terdapat padagenotipe Inpago 6, Danau Gaung, Kencana Bali, Inpari16, Inpari 7, Lekat Alahu, Pulut Simanik, Lekat Tuleng,Lekat Kumbob, Pulut Merah, Asahan, Boh Penileh,Sirias, Dewi, Kuku Balam, Seraguek, Sigupai Pulo,Sirangkoh Lubok Pasi, Bo Somboh Meon, SigupaiWangi, Bo Santet Semantok, Saguek, Cantek Manis,Jeumpa Puteh, Ramos Tihion Tamping, Salah Manyang,Semerebuk, Sijane, Kepala Gajah, Siputeh, RangkohMerah dan Pade Merah Lamtuba. Genotipe yang tahanterhadap HDB dijumpai pada Inpari 1, Limboto, Tuwoti,Inpari 10, Lekat Rambot Linuet, Rom Mokot, Paki Gajah,Tamboen, Bo 100, Sipasie, Bo Minyek, Bontok, SirendehSemantuk Wayla dan Sambei. Genotipe yangmenunjukkan reaksi moderat tahan dijumpai pada DanauGaung, IPB 4S, Situ Bagedit, Inpago, Cirata, IPB 3S,

162 J. HPT Tropika Vol. 15 No. 2, 2015: 152 - 163
Pade Jamai Asan, Salah Manyang, Arias, Pandan Wangidan Pade Merah Lamtuba. Namun genotipe yangmenunjukkan reaksi rentan terhadap HDB dijumpai padaInapri 16, Ciherang, Situ Patenggang, Kencana Bali,Inpari 19, Inpari 7, IR-64, Lekat Alahu, Lekat Singke,Pulut Hitam, Pulut Simanik, Pulut Merah, Lekat Tuleng,Lekat Kumbob, Lekat jerajak Lango, Lekat Adang,Ketan Putih, Bo Somboh Meon, Sigupai Wangi, KukuBalam, Kepala Gajah, Asahan, Manyam, Boh Penileh,Saguek, Dewi, Jeumpa Puteh, Sigupai Blang Pidie, PalaGajah, Rangkoh Merah, Pade Kapai Tamping, PadiSitandun, Sirangkoh Lubok Pasi, Sepulou, Pade PinengLango, Seraguek, Semerebuk, Ramos Tihion Tamping,Sijane, Rom Lambo, Sirias, Sigupai Pulo, Bo SantetSemantok, Bo Rayek Semantok Wayla, Cantek Manis,Rom Ilang dan Siputeh.
SANWACANA
Penulis mengucapkan terimakasih kepadaUniversitas Syiah Kuala, Kementerian Pendidikan danKebudayaan, yang telah membiayai sebagian danapenelitian ini melalui Penelitian Unggulan PerguruanTinggi Tahun Anggaran 2014.
DAFTAR PUSTAKA
Abdullah B. 2009. Progress of rice improvement throughrecurrent selection. J. Agron. Indones. 37(3):188–193.
Bakhtiar, Kesumawati E, Hidayat T, & Rahmawati M.2011. Karakterisasi plasma nutfah padi lokal Acehuntuk perakitan varietas adaptif pada tanahmasam. Agrista 15(3): 79–86.
Berthaud J, Clement JC, Emperaire L, Louette D, PintonF, Sanou J, & Second G. 2000. The role of local-level geneflow in enhancing and maintaininggenetic diversity. In: Cooper HD, Spillene C, &Hodgken (Eds.). Broadening the Genetic Baseof Crops Production. pp. 81–104. IGRI, FAO,CABI Publishing, Wallingford.
Devadath S. 1989. Chemical control of bacterial blightof rice. In: Bacterial Blight of Rice. pp. 89–98.IRRI, Manila. Philippines.
Gnanamanickam SS, Priyadarisini VB, Narayanan NN,Vasudevan P, & Kavitha S. 1999. An overviewof bacterial blight disease of rice and strategiesfor its management. Curr. Sci. 77(11): 1435–1443.
Herlina L & Silitonga TS. 2011. Seleksi lapang ketahananbeberapa varietas padi terhadap infeksi hawardaun bakteri strain IV dan VIII. Bul. PlasmaNutfah 17(2): 80–87.
Klement Z, Stall RE, Novacky A, Ersek T, Fett T W,Huang J, & Beckman C. 1990. Mechanism ofresistance. In: Klement Z, Rudolph K, & SandsDC (Eds.). Methods in Phytobacteriology. pp.469–473. Budapest, Hungary.
Nino-Liu DO, Ronald PC, & Bogdanove AJ. 2006.Xanthomonas oryzae pathovars: modelpathogens of a model crop. Mol. Plant Pathol.7(5): 303–324.
Ogawa T, Busto GA, Tabien RE, & Khush GS. 1988.Further study of Xa-4b gene for resistance tobacterial blight of rice. Rice Genetics Newsletter5: 104–105.
Ou SH. 1985. Rice Diseases . Second edition.Commowealth Mycological Institute. TheCambrian News, Aberystwyth.
Rahim A, Khaeruni A, & Taufik M. 2012. Reaksiketahanan beberapa varietas padi komersialterhadap patotipe Xanthomonas oryzae pv.oryzae isolat Sulawesi Tenggara. BerkalaPenelitian Agronomi 1(2): 132–138.
Shehzad FD, Farhatullah, Iqbal N, Shah SMA, & AhmadM. 2012. Screening of local rice germplasmagainst bacterial leaf blight caused byXanthomonas oryzae pv. oryzae. Sarhad J.Agric. 28(4): 565–569.
Sitaresmi T, Wening RH, Rakhmi AT, Yunani N, &Susanto U. 2013. Pemanfaatan plasma nutfahpadi varietas lokal dalam perakitan varietasunggul. Iptek Tan. Pangan 8(1): 22–30.
Sudir, Nuryanto B, & Kadir TS. 2012. Epidemiologi,patotipe, dan strategi pengendalian penyakit hawardaun bakteri pada tanaman padi. Iptek TanamanPangan 7(2): 79–87.
Suswanto A, Friska H, & Sudirman I. 1996. KarakterisasiPseudomonas fluorescens B29 dan B39; profilDNA genom, uji hipersensitivitas, dan asaisenyawa bioaktif. Hayati 3(1): 15–20.
Wahyudi AT, Meliah S, & Nawangsih AA. 2011.Xanthomonas oryzae pv. oryzae bakteripenyebab hawar daun pada padi: isolasi,karakterisasi, dan telaah mutagenesis dengantransposon. Makara Sains 15(1): 89–96.

Hadianto et al. Ketahanan Beberapa Genotipe Padi 163
Wiyono S. 2007. Perubahan iklim dan ledakan hamadan penyakit tanaman. Makalah SeminarSehari tentang Keanekaragaman Hayati ditengah Perubahan Iklim: Tantangan MasaDepan Indonesia. pp.1–10. Jakarta. 18 Juni2008.
Zou LF, Wang XP, Xiang Y, Zhang B, Li YR, Xiao YL,Wang JS, Walmsley AR, & Chen GY. 2006.Elucidation of the hrp clusters of Xanthomonasoryzae pv. oryzicola that control thehypersensitive response in nonhost tobacco andpathogenicity in susceptible host rice. Appl.Environ. Microbiol. 72(9): 6212–6224.

164 J. HPT Tropika Vol. 15 No. 2, 2015: 164 - 169J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 164 – 169, September 2015
EVALUASI LIMA JENIS INNER CARRIER DAN FORMULASIBACILLUS SUBTILIS UNTUK PENGENDALIAN HAWAR
PELEPAH JAGUNG (RHIZOCTONIA SOLANI KUHN)
Amran Muis, Nurasiah Djaenuddin, & Nurnina Nonci
Balai Penelitian Tanaman SerealiaJl. Dr. Ratulangi No. 274, Maros 90514
E-mail: [email protected]
ABSTRACT
Evaluation of five inner carriers and Bacillus subtilis formulation to control banded leaf and sheath blight (Rhizoctoniasolani Kuhn). One alternative control method against plant pathogens is the use of antagonistic microorganisms, such asBacillus subtilis. The use of the antagonistic bacteria on corn especially in Indonesia is still lack. The objective of thisresearch was to evaluate some inner carrier and to make formulated antagonistic B. subtilis to be used as biological controlagents on corn diseases. This research consists of laboratory and greenhouse activities. The laboratory activities consist ofB. subtilis biomass production, formulation of B. subtilis, and evaluation of five types of inner carrier. In the greenhouse,testing the formulation B. subtilis with talc as an inner carrier, which is compared with the treatment solution of B. subtilis,nordox, metalaxyl fungicides. The data collected in this study were percentage of germination, damping off due to pathogenR. solani, plant height, plant fresh weight, and percentages of R. solani incidence on 14 DAP. The results showed that talcpowder and corn flour were the best inner carrier to be used in sorage formulation of antagonistic Bacillus. FormulatedBacillus subtilis TM4 showed no negative affect on seed germination and able to suppress the development of R. solani ingreenhouse.
Key words: Bacillus subtilis, formulation, inner carrier, Rhizoctonia solani
ABSTRAK
Evaluasi lima jenis inner carrier dan formulasi Bacillus subtilis untuk pengendalian hawar pelepah jagung (Rhizoctoniasolani Kuhn). Salah satu alternatif pengendalian patogen tanaman yaitu pemanfaatan mikroorganisme antagonis sepertiBacillus subtilis. Penggunaan bakteri antagonis tersebut pada tanaman jagung di Indonesia masih sangat kurang. Penelitianini bertujuan mengevaluasi beberapa inner carrier dan membuat formulasi bakteri antagonis B. subtilis untuk digunakansebagai agensia pengendali hayati penyakit pada tanaman jagung. Kegiatan penelitian ini terdiri dari kegiatan laboratoriumdan rumah kaca. Di laboratorium dilakukan produksi biomass B. subtilis, pembuatan formulasi B. subtilis, dan evaluasiformulasi dari lima jenis inner carrier. Di rumah kaca, dilakukan pengujian formulasi B. subtilis dengan talc sebagai innercarrier yang dibandingkan dengan perlakuan larutan B. subtilis, fungisida nordox, dan fungisida metalaxyl. Pengamatandilakukan terhadap daya kecambah, damping off akibat patogen R. solani, tinggi tanaman, berat segar tanaman, dan persentaseserangan R. solani pada 14 HST. Hasil penelitian menunjukkan bahwa talc dan tepung jagung merupakan inner carrier yangterbaik untuk digunakan dalam pembuatan formulasi bakteri antagonis B. subtilis. Formulasi B. subtilis TM4 tidak berpengaruhnegatif terhadap daya kecambah benih serta mampu menekan perkembangan R. solani di rumah kaca.
Kata kunci: Bacillus subtilis, formulasi, inner carrier, Rhizoctonia solani
PENDAHULUAN
Semua bagian dari tanaman jagung adalah rentanterhadap serangan berbagai jenis penyakit yang dapatmenurunkan baik kualitas maupun kuantitas. Selainpenyakit bulai, sejumlah penyakit penting pada tanamanjagung seringkali berpengaruh secara ekonomi,diantaranya adalah penyakit hawar pelepah yangdisebabkan oleh cendawan Rhizoctonia solani dan
penyakit bercak daun yang disebabkan oleh cendawanBipolaris maydis.
Cendawan R. solani adalah patogen tular tanahdan memiliki kisaran inang yang luas, sehingga sulitdikendalikan (Ogoshi, 1987). Pengendalian terhadappenyakit tersebut selama ini masih mengandalkan padapenggunaan pestisida kimiawi, sedangkan penggunaanvarietas tahan masih sangat terbatas jumlahnya.

Muis et al. Evaluasi Lima jenis Inner Carrier 165
Beberapa tahun belakangan ini, penelitian tentangpemanfaatan mikroorganisme antagonis untukmengendalikan penyakit-penyakit tersebut banyakdilakukan. Salah satu mikroorganisme antagonis yangmenunjukkan hasil yang baik dalam pengendalianpatogen penyebab penyakit adalah bakteri antagonisBacillus subtilis yang keefektifannya dalammengendalikan penyakit-penyakit pada berbagaikomoditas telah banyak menunjukkan hasil yang baik(Kilian et al., 2000; Schisler et al., 2002; EPA, 2003).Agensia pengendali hayati menunjukkan kemampuannyadalam menghambat perkembangan penyakit tanamanserta meningkatkan pertumbuhan tanaman denganmekanisme yang khas (Keel & Defago, 1997). Rustamet al. (2011) telah memperoleh beberapa bakteriantagonis yang potensial sebagai agensia pengendalihayati penyakit hawar pelepah padi. Namun penggunaanbakteri antagonis sebagai agensia hayati tersebut padatanaman jagung masih sangat kurang terutama diIndonesia.
Untuk memudahkan aplikasi bakteri antagonisuntuk mengendalikan penyakit tanaman, dibutuhkansuatu formulasi dengan bahan inner carrier yang tepatsebagai sumber karbon. Berdasarkan hal tersebut di ataspenelitian ini dilakukan untuk mengevaluasi beberapainner carrier untuk digunakan dalam pembuatanformulasi bakteri antagonis Bacillus.
METODE PENELITIAN
Tempat dan Waktu. Penelitian ini dilaksanakan diLaboratorium Penyakit Tanaman dan rumah kaca BalaiPenelitian Tanaman Serealia (Balitsereal) di MarosSulawesi Selatan, pada bulan Februari sampai denganMei 2013.
Produksi Biomass B. subtilis. Isolat B. subtilis (3isolat terbaik dari penelitian sebelumnya) diperbanyakpada media Potato Dextrose Broth (PDB) (250 gkentang + 20 g dextrose + 1000 ml akuades steril).Sebanyak 180 ml larutan PDB tersebut dimasukkan kedalam Erlenmeyer berukuran 1000 ml yang diulang 5kali, kemudian disterilkan pada tekanan 15 psi selama15 menit. Setelah sterilisasi, setiap erlenmeyerdiinokulasi dengan B. subtilis yang berumur 36 jam danterlebih dahulu dilarutkan ke dalam akuades steril. Kulturtersebut kemudian dikocok selama 36 jam. Biomass yangdihasilkan dipanen dengan menggunakan alatsentrifugasi pada kecepatan 5000 rpm selama 6 menit.Sel bakteri hasil sentrifugasi inilah yang digunakan didalam formulasi.
Evaluasi beberapa Inner carrier sebagai BahanFormulasi B. subtilis. Inner carrier yang akandievaluasi dan bahan additive yang akan digunakandalam formulasi adalah:A. Talc + 0,25% yeast extract + 1% CMCB. Tepung beras + 0,25% yeast extract + 1% CMCC. Tepung tapioka + 0,25% yeast extract + 1% CMCD. Tepung jagung + 0,25% yeast extract + 1% CMCE. Tepung beras pecah + 0,25 yeast extract + 1% CMC
Dua puluh lima g inner carrier, 0,25% yeastextract, dan 1% Carboxymethyl Cellulose (CMC)dicampur di dalam wadah aluminium steril. Bahancampuran tersebut lalu dimasukkan ke dalam erlenmeyerberukuran 500 ml, kemudian disterilkan pada tekanan15 psi selama 30 menit. Setelah disterilkan dandidinginkan, dimasukkan 20 ml sel bakteri yang telahdisiapkan sebelumnya ke dalam masing-masingcampuran inner carrier tadi, kemudian dicampurkansecara merata dan dikeringanginkan di dalam laminarflow selama 48 jam. Setelah kering, bahan formulasitersebut dimasukkan ke dalam kantong plastik sterilkemudian disimpan di dalam suhu ruangan untukdigunakan selanjutnya.
Bahan inner carrier yang paling baik adalah yangmengandung sel bakteri paling banyak. Untukmengetahui kandungan sel bakteri pada setiap bahaninner carrier, dilakukan pengujian sebagai berikut: satug formulasi bakteri dimasukkan ke dalam kantong plastiksteril yang berisi 100 g biji jagung dan telah ditambahkan1 ml akuades steril, lalu dikocok hingga formulasi tersebutmembungkus biji jagung secara merata. Kemudiandiambil tiga biji jagung tadi secara acak lalu dimasukkanke dalam tabung reaksi yang berisi 10 ml akuades steril,lalu diencerkan hingga 10-2. Dari larutan yang telahdiencerkan tadi, diambil 0,1 ml menggunakan pipet sterildan diletakkan pada cawan Petri berisi Potato dextrosepeptone agar (PDPA) dengan komposisi 200 g kentang,20 g dekstrose, 20 g agar, 15 g peptone dan 1000 mlakuades, lalu disebar dengan menggunakan glass rodhockey. Cawan Petri tersebut kemudian diberi selotipdan diinkubasi dengan posisi terbalik. Setelah 24 jam,koloni bakteri yang terbentuk pada setiap cawan Petridihitung.
Pengujian Formulasi B. subtilis TM4 terhadapRhizoctonia solani di Rumah Kaca. Sebelumdilakukan pengujian terlebih dahulu disiapkan formulasiB. subtilis TM4 dengan mencampurkan 20 ml suspensibakteri yang diambil dari kultur bakteri + talc yang sudahdisterilkan + 0,25% yeast extract dan 1% CMC lalu

166 J. HPT Tropika Vol. 15 No. 2, 2015: 164 - 169
dikeringanginkan di dalam laminar flow chamberselama dua hari.
Inokulum patogen R. solani disiapkan denganmembiakkannya pada media campuran beras dan sekampadi yang dimasukkan di dalam plastik berukuran 1 L.Sebelum dibiakkan, media tersebut disterilkan selama 2jam pada tekanan 15 psi. Setelah sterilisasi diambilpotongan biakan R. solani dari cawan Petri (3 mm2)lalu dimasukkan ke dalam media steril tadi. Biakantersebut disimpan di dalam suhu kamar selama 2 minggu.Setelah biakan R. solani siap, inokulum tersebutkemudian dicampurkan ke dalam baki plastik yang telahdiisi tanah steril dan dibiarkan selama 14 hari. Sebagaikontrol, digunakan tanah pada baki plastik yang tidakdiinokulasi dengan R. solani. Setelah inokulum siap didalam baki tadi, ke dalam baki-baki tersebut ditanambiji jagung yang telah diberi perlakuan benih dengan cara10 g formulasi bakteri dicampurkan dengan 1 kg benihjagung di dalam kantong plastik dan dikocok hingga rata.Perlakuan disusun dalam rancangan acak lengkap yangterdiri dari 7 perlakuan dan 3 ulangan sebagai berikut:
Pengamatan dilakukan terhadap daya kecambahpada 7 Hari Setelah Tanam (HST), tinggi tanaman,damping off, berat segar keseluruhan tanaman danpersentase serangan R. solani pada 14 HST.Persentase serangan R. solani dihitung denganmenggunakan rumus sebagai berikut:
100%b
aP
dengan:P = Persentase serangan R. solania = Jumlah tanaman terserang R. solanib = Jumlah tanaman yang diamati
Analisis Data. Data hasil lima jenis inner carrier invitro dan hasil pengujian in vivo di rumah kaca dianalisisdengan program komputer SAS ver. 9.1.3 (SAS InstituteInc., 2006).
HASIL DAN PEMBAHASAN
Hasil pengamatan terhadap jumlah kolonibakteri antagonis pada lima inner carrier menunjukkanbahwa talc adalah inner carrier terbaik diantara innercarrier lainnya yang diuji diikuti dengan tepung jagung(Tabel 1). Hal ini diduga karena ukuran partikel daritalc paling kecil diantara inner carrier lain, ini ditandaidengan serbuk talc lebih halus dari inner carrier laindan sifatnya mudah menyebar sehingga jika disebarkandalam air terbentuk suspensi yang kental dan akan mudahberikatan dengan larutan bakteri. Menurut Anonim(2014), talc merupakan garam anhidrat yang bersifatcepat terhidrasi dengan mengikat air atau senyawa-senyawa yang dapat mengikat air melalui pengikatan dipermukaan (surface adhesion) tanpa menjadi basahdan menggumpal.
Pada Tabel 1 tampak bahwa dari tiga isolat B.subtilis yang diuji, isolat TM4 menunjukkan populasibakteri yang terbanyak yakni rata-rata 99 x 105 cfu/ml.Selain talc, tepung jagung juga memperlihatkan hasilyang baik sebagai inner carrier, terutama bagi isolatBNT5. Pada Tabel 1 nampak bahwa jumlah kolonibakteri antagonis dengan formulasi tepung jagung
Tabel 1. Jumlah koloni 3 isolat B. subtilis pada 5 formulasi di laboratorium
Inner carrierJumlah koloni B. subtilis (x 105 cfu/ml)
Isolat TLB1 (57) Isolat TM4 (87) Isolat BNt5 (109)
Talc 73,33 a 99,00 a 96,67 aTepung jagung 29,00 b 32,33 b 88,00 aTepung beras pecah 13,00 c 9,67 c 24,33 bTepung beras 11,33 c 13,00 c 12,33 bTepung tapioka 5,67 c 4,33 c 15,33 b
KK (%) 29,22 15,09 13,79
Angka dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji BNT 5%.
P1 = Seed treatment dengan larutan B. subtilis TM4P2 = Seed treatment dengan formulasi B. subtilis
TM4+ 0,5% gum ArabicP3 = Seed treatment dengan fungisida nordoxP4 = Seed treatment dengan fungisida metalaxylP5 = Seed treatment dengan fungisida nordox +
metalaxylP6 = Kontrol (diinokulasi dengan R. solani)P7 = Kontrol (tanpa inokulasi dengan R. solani)
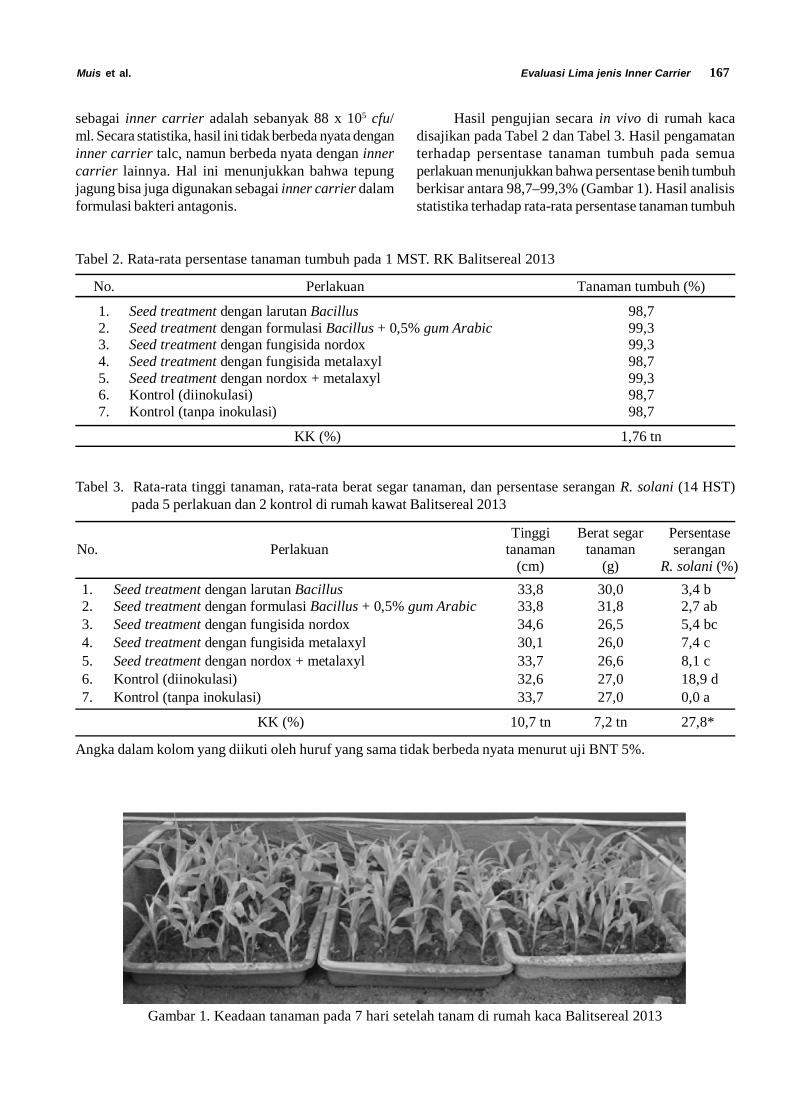
Muis et al. Evaluasi Lima jenis Inner Carrier 167
sebagai inner carrier adalah sebanyak 88 x 105 cfu/ml. Secara statistika, hasil ini tidak berbeda nyata denganinner carrier talc, namun berbeda nyata dengan innercarrier lainnya. Hal ini menunjukkan bahwa tepungjagung bisa juga digunakan sebagai inner carrier dalamformulasi bakteri antagonis.
Hasil pengujian secara in vivo di rumah kacadisajikan pada Tabel 2 dan Tabel 3. Hasil pengamatanterhadap persentase tanaman tumbuh pada semuaperlakuan menunjukkan bahwa persentase benih tumbuhberkisar antara 98,7–99,3% (Gambar 1). Hasil analisisstatistika terhadap rata-rata persentase tanaman tumbuh
Gambar 1. Keadaan tanaman pada 7 hari setelah tanam di rumah kaca Balitsereal 2013
Tabel 2. Rata-rata persentase tanaman tumbuh pada 1 MST. RK Balitsereal 2013
No. Perlakuan Tanaman tumbuh (%)
1. Seed treatment dengan larutan Bacillus 98,72. Seed treatment dengan formulasi Bacillus + 0,5% gum Arabic 99,33. Seed treatment dengan fungisida nordox 99,34. Seed treatment dengan fungisida metalaxyl 98,75. Seed treatment dengan nordox + metalaxyl 99,36. Kontrol (diinokulasi) 98,77. Kontrol (tanpa inokulasi) 98,7
KK (%) 1,76 tn
Angka dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji BNT 5%.
Tabel 3. Rata-rata tinggi tanaman, rata-rata berat segar tanaman, dan persentase serangan R. solani (14 HST)pada 5 perlakuan dan 2 kontrol di rumah kawat Balitsereal 2013
No. PerlakuanTinggi
tanaman(cm)
Berat segartanaman
(g)
Persentaseserangan
R. solani (%)
1. Seed treatment dengan larutan Bacillus 33,8 30,0 3,4 b2. Seed treatment dengan formulasi Bacillus + 0,5% gum Arabic 33,8 31,8 2,7 ab3. Seed treatment dengan fungisida nordox 34,6 26,5 5,4 bc4. Seed treatment dengan fungisida metalaxyl 30,1 26,0 7,4 c5. Seed treatment dengan nordox + metalaxyl 33,7 26,6 8,1 c6. Kontrol (diinokulasi) 32,6 27,0 18,9 d7. Kontrol (tanpa inokulasi) 33,7 27,0 0,0 a
KK (%) 10,7 tn 7,2 tn 27,8*

168 J. HPT Tropika Vol. 15 No. 2, 2015: 164 - 169
pada 1 minggu setelah tanam (MST) menunjukkan tidakada perbedaan yang nyata antar perlakuan. Hal inimenunjukkan bahwa seed treatment dengan formulasibakteri antagonis B. subtilis TM4 + yeast extract +CMC + gum Arabic tidak berpengaruh negatif terhadapdaya kecambah benih. Kilian et al. (2000) melaporkanbahwa kolonisasi akar dan rizosfer dipengaruhi olehsejumlah faktor termasuk teknik aplikasi. SelanjutnyaCanaday & Ownley (1999) mengemukakan bahwaproporsi populasi B. subtilis pada akar tergantung padajumlah B. subtilis yang diaplikasikan.
Hasil pengamatan terhadap tinggi tanaman pada14 HST menunjukkan bahwa hanya perlakuan P4 yangagak pendek dibanding dengan perlakuan lainnya (Tabel3). Hasil analisis statistika terhadap rata-rata tinggitanaman menunjukkan tidak ada perbedaan yang nyataantar perlakuan. Pada Tabel 2 tampak bahwa perlakuanbenih dengan B. subtilis baik yang tanpa formulasi (P1)maupun dengan formulasi (P2) menunjukkan tinggitanaman yang hampir sama dengan kontrol tanpainokulasi R. solani.
Hasil pengamatan terhadap berat segar 10tanaman pada 14 HST disajikan pada Tabel 2. PadaTabel 2 terlihat bahwa walaupun rata-rata tinggi tanamanpada semua perlakuan hampir sama, namun perlakuanbenih dengan formulasi B. subtilis menunjukkan rata-rata berat segar tanaman yang lebih tinggi dibandingperlakuan lainnya. Hal ini menunjukkan bahwa B.subtilis disamping sebagai antagonis bagi cendawan,juga berfungsi sebagai pemacu daya tumbuh tanaman,seperti yang dikemukakan oleh Van Loon et al. (1998),perlakuan benih dengan plant growth promotingrhizobacterium (PGPR) seperti Bacillus spp.meningkatkan persentase kecambah benih, vigortanaman, pertumbuhan akar, biomas tanaman, dan lain-lain. Selanjutnya Prathibha & Siddalingeshwara (2013)mengemukakan bahwa PGPR seperti B. subtilis efektifmeningkatkan kualitas benih seperti daya kecambah,indeks vigor, dan kualitas nutrisi seperti kandunganprotein dan karbohidrat.
Mishra et al. (2010) mengemukakan bahwaperlakuan benih dengan B. subtilis meningkatkanproduksi pada tanaman Pelagronium gravelen.Selanjutnya Mohamed & Gomaa (2012) melaporkanbahwa benih lobak yang diinokulasi dengan B. subtilissebelum tanam, meningkatkan secara nyata biomassegar dan kering akar dan daun. Selain itu bakteritersebut juga meningkatkan kandungan fitohormon (IAAdan GA) serta kandungan N, P, K, Ca, dan Mg. Baugh(2014) juga mengemukakan bahwa B. subtilis
menghasilkan auksin yang bersifat meningkatkan dayatumbuh tanaman.
Hasil pengamatan terhadap persentase seranganR. solani pada 14 HST disajikan pada Tabel 3. Hasilanalisis statistika terhadap persentase serangancendawan R. solani menunjukkan bahwa perlakuanSeed treatment dengan formulasi B. subtilis TM4 +0,5% gum Arabic berbeda nyata dengan perlakuan Seedtreatment dengan fungisida metalaxyl dan seedtreatment dengan fungisida nordox + metalaxyl sertadengan perlakuan kontrol (diinokulasi dengan R. solani).Perlakuan Seed treatment dengan formulasi B. subtilisTM4 + 0,5% gum Arabic tersebut tidak berbeda nyatadengan perlakuan seed treatment dengan larutan B.subtilis TM4, seed treatment dengan fungisida nordox,dan kontrol (tanpa inokulasi dengan R. solani). Namundemikian, persentase serangan cendawan R. solaniditemukan terendah pada perlakuan Seed treatmentdengan formulasi B. subtilis TM4 + 0,5% gum Arabicdengan persentase serangan 2,7%. Hal ini didugatanaman memberikan reaksi ketahanannya terhadapinfeksi patogen akibat adanya bakteri antagonis yangjuga berperan dalam meningkatkan mekanismeketahanan jaringan tanaman. Menurut Van Loon et al.(1998), mekanisme ketahanan tanaman yangdipengaruhi oleh bakteri antagonis ini dikenal denganistilah ketahanan terinduksi secara sistemik (inducedsystemic resistance). Dan persentase serangan tertinggiditunjukkan oleh perlakuan kontrol (tanpa inokulasidengan R. solani).
Hasil penelitian ini menunjukkan bahwa formulasibakteri antagonis B. subtilis TM4 dapat menekanperkembangan serangan cendawan R. solani.Kemampuan ini mungkin disebabkan oleh antibiotik yangdihasilkan oleh bakteri tersebut. Seperti yangdikemukakan oleh Islam et al. (2012), bahwa komposisinutrisi (karbon dan nitrogen) dan kondisi lingkungan (pHdan temperatur) mempengaruhi produksi antibiotik daribakteri antagonis. EPA (2003) melaporkan bahwa B.subtilis strain QST 713 dapat mencegah perkembangansejumlah jenis cendawan tertentu yang kemungkinandengan cara bersaing dalam hal kebutuhan hara,lingkungan pertumbuhan tanaman, dan secara langsungmengkolonisasi cendawan patogen. Niu et al. (2011)melaporkan bahwa Bacillus cereus AR156 merupakanPGPR yang menginduksi ketahanan tanaman terhadapberbagai jenis patogen termasuk Pseudomonassyringae pv. tomato DC3000. Bakteri ini meningkatkanbiomas tanaman dan menekan perkembangan penyakitdi daun.

Muis et al. Evaluasi Lima jenis Inner Carrier 169
SIMPULAN
Talc dan tepung jagung merupakan inner carrieryang terbaik untuk digunakan dalam pembuatanformulasi bakteri antagonis Bacillus subtilis. Formulasibakteri antagonis Bacillus subtilis TM4 tidakberpengaruh negatif terhadap daya kecambah benih.Formulasi bakteri antagonis Bacillus isolat TM4 mampumenekan perkembangan cendawan patogen R. solanidi rumah kaca.
DAFTAR PUSTAKA
Anonim. 2014. Bahan Tambahan Pangan. Buku PutihKementerian Kesehatan Republik Indonesia,Jakarta.
Baugh B. 2014. Bacillus subtilis Plant GrowthPromoter. http:/living-soils.com/bacillus-subtilis-plant-growth-promoter/. Diakses 3 April 2014.
Canaday CH & Ownley BH. 1999. Effects of SeedTreatment Chemicals and Bacillus subtilis onSnap Bean Seedling Diseases, Growth, andYield. The University of Tennessee, Knoxville.
EPA (U.S. Environmental Protection Agency). 2003.Bacillus subtilis QST713 (006479). http://www.epa.gov. Diakses 9 Mei 2005.
Islam MdR, Jeong YT, Lee YS, & Song CH. 2012.Isolation and identification of antifungalcompounds from Bacillus subtilis C9 inhibitingthe growth of plant pathogenic fungi.Mycobiology 40(1): 59–66.
Keel C & Defago G. 1997. Interactions betweenbeneficial soil bacteria and root pathogens:mechanisms and ecological impact. In: Gange AC& Brown VK (Eds.). 36th Symposium of TheBritish Ecological Society MultitrophicInteraction in terrestrial system. pp. 27–47.Blackwell Science Oxford.
Kilian M, Steiner U, Krebs B, Junge H, SchmiedeknechtG, & Hain R. 2000. FZB24® Bacillus subtilis –mode of action of a microbial agent enhancingplant vitality. Pflanzenschutz-NachrichtenBayer 1/00 1: 72–93.
Mishra RK, Prakash Om, Alam M, & Dikshit A. 2010.Influence of plant growth promoting rhizobacteria(PGPR) on the productivity of Pelargoniumgraveolens L. Herit. Recent Res. Sci. Technol.2(5): 53–57.
Mohamed HI & Gomaa EZ. 2012. Effect of plantgrowth promoting Bacillus subtilis andPseudomonas fluorescens on growth andpigment composition of radish plants (Raphanussativus) under NaCl stress. Photosynthetica50(2): 263–272.
Niu DD, Liu HX, Jiang CH, Wang YP, Wang QY, JinHL, & Guo JH. 2011.The plant growth–promotingrhizobacterium Bacillus cereus AR156 inducessystemic resistance in Arabidopsis thaliana bysimultaneously activating salicylate-andjasmonate/ethylene-dependent signalingpathways. Mol. Plant-Microbe Interact. 24(5):533–542.
Ogoshi A. 1987. Ecology and pathogenicity ofanastomosis and intraspecific group ofRhizoctonia solani KÜhn. Annu. Rev.Phytopathol. 25: 125–143.
Prathibha KS & Siddalingeshwara KG. 2013. Effect ofplant growth promoting Bacillus subtilis andPseudomonas fluorescence as Rhizobacteria onseed quality of sorghum. Int. J. Curr. Microbiol.App. Sci. 2(3): 11–18.
Rustam, Giyanto, Wiyono S, Santosa DA, & Susanto S.2011. Seleksi dan identifikasi bakteri antagonissebagai agens pengendali hayati penyakit hawarpelepah padi. Penelitian Pertanian TanamanPangan 30(3): 164–171.
SAS Institute Inc. 2006. Base SAS ® 9.1.3 ProceduresGuide. Second Edition. Volumes 1, 2, 3, and 4.Cary.
Schisler DA, Khan NI, Boehm MJ, & Slininger PJ. 2002.Greenhouse and field evaluation of biologicalcontrol of Fusarium head blight on durum wheat.Plant Dis. 86(12): 1350–1356.
van Loon LC, Bakker PAHM, & Pieterse CMJ. 1998.Systemic resistance induced by rhizospherebacteria. Annu. Rev. Phytopathol. 36: 453–483.

170 J. HPT Tropika Vol. 15 No. 2, 2015: 170 - 179J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 170 – 179, September 2015
EKSPLORASI DAN UJI SENYAWA BIOAKTIF BAKTERI AGENSIA HAYATIUNTUK PENGENDALIAN PENYAKIT KRESEK PADA PADI
Sri Kurniawati1, Kikin Hamzah Mutaqin2, & Giyanto2
1 BPTP Banten, Badan Penelitian dan Pengembangan Kementerian PertanianJl. Raya Ciptayasa Km 01 Ciruas, Serang-Banten
2 Laboratorium Bakteriologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Insitut Pertanian BogorJl. Kamper, Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Exploration of bacterial biocontrol agent and its potential bioactive compound to control rice bacterial leaf blight. Theresearch aims were to obtain bacterial isolates which were potential as biological control agent of X. oryzae pv. oryzae, thecausal agent of rice bacterial blight and to assess the effectiveness of their bioactive compounds, and to identify of thepotential isolates. The research steps included bacterial isolation, screening based on antibiosis activity and pathogenicitytest, characterization based on chitinolytic enzyme production, siderophores, and phosphate dissolution test, effectivenesstest of bioactive compounds and molecular identification of potential isolates. Out of 156 bacterial isolates from rice croptested, 11 isolates showed to be non plant pathogenic and to have activity as biological agents against X. oryzae pv. oryzaepathotype III, IV and VIII. Further characterization of 11 isolates resulted in 2 isolates that showrd ability to produce chitinase(isolates T5-1118 and R7-1018), phosphatase (isolates T5-1105 and T6-1109), and siderophores (isolates T5-1118 and T6-1109). The test of bioactive compound effectiveness of 4 isolates to the growth of X. oryzae pv. oryzae showed thatT5-1118,T5-1105, T6-1109 and R7-1018 have ability to inhibit X. oryzae pv. oryzae at 48 hours after inoculation of 66,61%, 62,4%,23,97% and 12,40%, respectively. Identification of 4 bacterial biocontrol isolates with partial sequencing of 16S rRNA geneshowed that those bacteria are close to Bacillus nealsonii strain F22 (R7-1018), Chromobacterium sp. MWU328 (T5-1118),Streptomyces sp. Antag 1 (T5-1105) and Kitasatospora nipponensis strains H2-4 (T6-1109).
Key words: Bacillus, characterization, Chromobacterium, Kitasatospora, Streptomyces
ABSTRAK
Eksplorasi dan uji senyawa bioaktif bakteri agensia hayati untuk pengendalian penyakit kresek pada padi. Penelitian inibertujuan untuk memperoleh isolat bakteri yang berpotensi sebagai agensia hayati untuk menghambat pertumbuhan X.oryzae pv. oryzae, menguji keefektifan senyawa bioaktif yang dihasilkannya dan mengidentifikasi isolat bakteri yang terujiberpotensi sebagai agensia hayati tersebut. Metode penelitian meliputi isolasi, penapisan melalui uji antibiosis dan patogenisitas,karakterisasi dengan uji produksi enzim kitinolitik, siderofor dan pelarutan fosfat. Isolat yang terpilih diuji potensi senyawabioaktifnya dan identifikasi secara molekuler melalui sekuensing gen 16S rRNA. Sebanyak 156 isolat bakteri telah diperolehdari pertanaman padi, 11 isolat bakteri diantaranya menunjukkan potensi sebagai agensia hayati yang ditunjukkan dengankemampuan menghambat X. oryzae pv. oryzae patotipe III, IV dan VIII dan tidak bersifat patogenik terhadap tanaman.Karakterisasi terhadap11 isolat tersebut menghasilkan 4 isolat yang memiliki keunggulan berupa kemampuan aktivitas kitinolitik(T5-1118 dan R7-1018), melarutkan fosfat ( T5-1105 dan T6-1109), dan produksi siderofor (T5-1118 dan T6-1109). Pengujianpotensi senyawa bioaktif dari keempat isolat secara in vitro menunjukkan bahwa isolat T5-1118, T5-1105, T6-1109 dan R7-1018dapat menghambat pertumbuhan X. oryzae pv. oryzae pada 48 jam setelah inokulasi masing-masing sebesar 66,61; 62,4; 23,97dan 12,40%. Identifikasi berdasarkan sekuensing parsial gen pengkode 16S rRNA menunjukkan bahwa keempat isolat tersebutadalah Bacillus nealsonii strain F22 (R7-1018), Chromobacterium sp. MWU328 (T5-1118), Streptomyces sp. Antag1 (T5-1105) dan Kitasatospora nipponensis strain H2-4 (T6-1109).
Kata kunci: Bacillus, Chromobacterium, karakterisasi, Kitasatospora, Streptomyces
PENDAHULUAN
Pemerintah Republik Indonesia menargetkanproduksi padi tahun 2013 sebesar 72,06 juta ton gabah
kering giling (GKG) dan surplus beras 10 juta ton padatahun 2015. Namun, produksi padi yang dicapai tahun2013 adalah 71,23 juta ton GKG dengan realisasi luaspanen 13,84 juta ha (BPS, 2014). Fakta tersebut

Kurniawati et al. Eksplorasi dan Uji Senyawa Bioaktif Bakteri 171
menunjukkan terdapat kesenjangan antara target danrealisasi produksi padi. Salah satu faktor pembatas dalamproduksi padi di Indonesia adalah adanya penyakit kresekatau hawar daun bakteri yang disebabkan olehXanthomonas oryzae pv. oryzae. Penyakit ini dapatmenyebabkan kehilangan hasil sebesar 35,8%(Suparyono & Sudir, 1992). Bakteri patogen inidilaporkan memiliki beberapa patotipe dan yang dominandi Indonesia adalah patotipe III, IV dan VIII (Suparyonoet al., 2004).
Sejauh ini pengendalian penyakit kresek adalahmenggunakan varietas unggul baru (VUB) tahan kresekseperti Conde dan Angke. Namun demikian, penggunaanbenih VUB ini memiliki beberapa kelemahan diantaranyabenih sulit diperoleh di pasaran dan kurang diminati olehpetani maupun konsumen. Selain itu, kelemahan lainnyadalam penggunaan varietas tahan adalah ketahananterhadap suatu patotipe X. oryzae pv. oryzae mudahdipatahkan jika ditanam dalam pola tanam monokulturyang luas dan dalam waktu yang lama. Teknikpengendalian lainnya adalah dengan penggunaanbakterisida sintetik yang memiliki keefektifan yang tinggidalam mengendalikan bakteri patogen ini, namunpenggunaan bakterisida secara terus menerus dapatmemicu terjadinya resistensi patogen, matinya organismebukan sasaran, pencemaran lingkungan, residu padaproduk yang membahayakan bagi kesehatan manusia.
Penggunaan agensia hayati sebagai komponenpengendalian dalam mengendalikan X.oryzae pv. oryzaememiliki prospek yang baik karena diyakini bersifatefektif, kompatibel atau sinergi dengan teknikpengendalian lain dan aman bagi lingkungan. Penelitian-penelitian tentang pengembangan agensia hayatiterhadap penyakit kresek pada padi telah banyakdilaporkan. Velusamy et al. (2006) berhasil mengisolasiPseudomonas flourescens yang menghasilkansenyawa anti mikroba yang dikenal sebagai 2,4-diacetylphloroglucinol (DAPG). Selanjutnya, Hoa et al. (2012)melakukan skrining agensia antagonis dan memperolehbeberapa strain Actinomycetes termasuk Streptomycesvirginiae yang mampu menghambat 10 patotipe X.oryzae pv. oryzae. Hastuti et al. (2012) melaporkanbahwa Streptomyces spp. dapat menurunkan keparahanpenyakit kresek dalam percobaan di rumah kaca maupundi lapangan yang setara dengan perlakuan bakterisida.Demikian pula penggunaan bakteri agensia hayatiPseudomonas diminuta, P. aeruginusa dan Bacillussubtilis dapat mengendalikan bakteri penyebab penyakitkresek ini dan meningkatkan pertumbuhan tanaman padi(Agustiansyah et al., 2013).
Tujuan penelitian ini adalah untuk memperolehisolat bakteri agensia hayati dari pertanaman padi yang
berpotensi menghambat pertumbuhan bakteri penyebabpenyakit kresek X. oryzae pv. oryzae patotipe III, IVdan VIII, menguji keefektifan senyawa bioaktif yangdihasilkannya dan mengidentifikasi isolat bakteripenghasil senyawa bioaktif yang mampu menghambatpertumbuhan bakteri patogen tersebut.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilakukan diLaboratorium Bakteri Patogen Tumbuhan, DepartemenProteksi Tanaman, Institut Pertanian Bogor dari bulanApril sampai dengan Desember 2013.
Isolasi Bakteri Agensia Hayati dari Tanah,Rizosfer dan Bakteri Endofit. Contoh tanaman adalahtanaman padi sehat yang diambil di antara tanaman yangterserang penyakit kresek. Lokasi pengambilan contohdari lahan sawah tadah hujan dilakukan di DesaWidodomartani, Kecamatan Ngemplak dan DesaHarjobinangun, Kecamatan Tratas, Kabupaten Sleman,Yogyakarta (Contoh 1-6), contoh dari lahan sawah irigasidi Desa Gempol Sari, Kecamatan Patok Beusi,Kabupaten Subang, Jawa Barat (Contoh 7), dan contohdari lahan rawa tipe A di Desa Karang Indah,Kecamatan Mandastana, Kabupaten Barito Kuala,Kalimantan Selatan pada agroekosistem lahan rawa tipeA (Contoh 8).
Isolasi Bakteri dari Tanah dan RizosferPertanaman Padi. Bakteri kandidat agensia hayatidiisolasi dari rizosfer tanaman padi dengan metodepengenceran bertingkat dan pencawanan (serialdilution and plating). Sebanyak 10 g tanah dimasukkanke dalam 90 ml larutan fisiologis (0,85% NaCl) dandibuat seri pengenceran 100–10-6. Suspensi tersebutdihomogenisasikan dalam rotary shaker pada kecepatan150 rpm selama 30 menit. Suspensi diambil sebanyak0,1 ml dan disebar pada medium tryptic soy agar/TSA(pancreatic digest of casein 17 g, NaCl 5 g, papaicdigest of soybean meal 3 g, K
2HPO
4 2,5 g, glukosa
2,5 g dan agar 15 g dalam 1000 ml akuades) 100% dan5%, King’s B (pepton protease 20 g, K
2HPO
4 1,5 g,
MgSO4. 7H
2O 1,5 g, agar 15 g dan gliserol 15 ml dalam
1000 ml akuades), water-yeast extract-agar/WYE(yeast extract 0,25 g, K
2HPO
4 0,5 g dan agar 18 g,
dalam 1000 ml akuades), casamino acid-yeast extract-glocose-agar/YCED (yeast extract 0,3 g, casaminoacid 0,3 g, D-glucose 0,3 g, K
2HPO
4 2 g, dan agar 18
g dalam 1000 ml akuades) dan nutrient agar/NA (Beefextract 3 g, pepton 5 g dan agar 15 g dalam 1000 ml

172 J. HPT Tropika Vol. 15 No. 2, 2015: 170 - 179
akuades), selanjutnya diinkubasikan selama 2–14 haripada suhu kamar.
Isolasi Bakteri Endofit Tanaman Padi. Isolasi bakteriendofit pada bagian akar, batang dan daun padimengikuti metode Hallmann et al. (1997) dan Munif etal. (2012).
Penyediaan Isolat Bakteri X. oryzae pv. oryzae.Isolat X. oryzae pv. oryzae patotipe III, IV dan VIIIdiperoleh dari koleksi Balai Besar Penelitian TanamanPadi di Sukamandi, Subang, Jawa Barat. Isolatdiremajakan dalam media Wakimoto Agar (WA) (bactopepton 7 g, sukrosa 17 g, Ca(NO
3)
2.4H
20 0,5 g,
Na2HPO
4.6H
2O 0,5 g, agar 18 g, kaldu dari kentang
300 g dalam 1000 ml akuades) sebelum digunakan untukpengujian.
Uji Kemampuan Antibiosis. Seleksi bakteri kandidatagensia hayati melalui pengujian antibiosis terhadap X.oryzae pv. oryzae patotipe III, IV dan VIII. Pengujianyang digunakan adalah dengan menggunakan metodedouble layer (Lisboa et al., 2006) dan cross cross-streak (Madigan et al., 1997) yang telah dimodifikasi.
Metode Double Layer. Isolat bakteri dibiakkandalam media NB (Nutrient Broth) selama 48 jam diatas inkubator bergoyang (100 rpm). Selanjutnya,suspensi bakteri dengan kerapatan 106–109 cfu/ml(OD
600 = 0,16-1,2) digunakan dalam pengujian. Bakteri
X. oryzae pv. oryzae dibiakkan pada media WakimotoBroth 48 jam di atas inkubator bergoyang (100 rpm).Sebanyak 100 µ l suspensi X. oryzae pv. oryzaedisebarkan pada permukaan media WA secara meratamenggunakan glass beads dan dikeringanginkan.Selanjutnya, 5 potongan kertas steril dengan diameter 5mm diletakan secara teratur pada permukaan media.Sebanyak 4 potongan kertas saring masing-masingditetesi 5 µl suspensi isolat bakteri yang berbeda dan 1potongan kertas saring ditetesi 5 µl media biakan yangtidak mengandung bakteri sebagai kontrol. Pengamatandilakukan terhadap lebar zone bening di sekitar kertassaring yang merupakan reaksi penghambatan dari bakteriagensia hayati terhadap X. oryzae pv. oryzae.
Metode Cross-Streak. Metode ini digunakanuntuk pengujian kelompok bakteri aktinomiset padamedia WA. Isolat aktinomiset digores pada satu sisicawan seluas sepertiga cawan. Aktinomiset diinkubasiselama 5 hari untuk memberi kesempatan tumbuh danmenghasilkan senyawa bioaktif yang akan berdifusi kemedia agar. Setelah 5 hari, bakteri X. oryzae pv. oryzaedigoreskan pada sisi cawan yang kosong sepanjang 4,5cm dengan arah tegak lurus terhadap goresan isolat
aktinomiset dan diinkubasi hingga tumbuh. Isolataktinomiset yang berpotensi antagonis akan ditunjukkandengan terbentuknya zona hambatan di mana X. oryzaepv. oryzae yang digores tidak tumbuh.
Karakterisasi Bakteri Agens Hayati. Karakterisasibakteri agens hayati terdiri dari pengujian reaksihipersensitif, aktivitas kitinolitik, pelarutan posfat danproduksi siderofor.
Pengujian Reaksi Hipersensitif. Bakterikandidat agensia hayati yang menghasilkan senyawabioaktif, diuji potensi patogenisitasnya denganmenggunakan uji hipersensitif pada tembakau varietasWhite Burley. Bakteri yang telah dikulturkan dalammedia NB selama 24 jam di atas inkubator bergoyangdengan kecepatan 100 rpm pada kerapatan 108–109 cfu/ml (OD
600= 0,16-1,2) kemudian diinfiltrasikan
menggunakan jarum suntik (syringe) ke dalam jaringandaun tembakau pada permukaan bawah daun. Respontanaman berupa gejala hipersenstif diamati pada 1–2hari setelah inokulasi.
Uji Aktivitas Kitinolitik. Pengujian dilakukanpada media agar koloidal kitin 0,2% (Lingappa &Lockwood, 1962). Pengujian aktivitas kitinolitik dilakukandengan meneteskan suspensi bakteri pada kertas saringsebanyak 5 µ l di atas media koloidal kitin dandiinkubasikan selama 4–7 hari pada suhu ruangan.Aktivitas kitinolitik ditunjukkan dengan adanya zonabening disekitar biakan bakteri.
Uji Produksi Siderofor. Produksi siderofor daribakteri agensia hayati dideteksi menggunakan mediaChroma Azurol Sulfonate (CAS) agar (Gross, 1990).Suspensi bakteri sebanyak 5 µl diteteskan pada kertassaring di atas media CAS dan diinkubasikan selama 4-7hari pada suhu ruangan. Produksi siderofor diindikasikandengan adanya warna orange di sekeliling koloni bakteri.
Uji Pelarutan Fosfat. Kemampuan bakteri dalammelarutkan fosfat diuji dengan menggunakan mediaPikovskaya Agar (Rao & Sinha, 1962). Suspensi bakterisebanyak 5 µl diteteskan pada kertas saring di atas mediaPikovskaya dan diinkubasikan selama 4-7 hari pada suhuruangan. Kemampuan bakteri dalam melarutkan fosfatdiindikasikan dengan adanya zona bening di sekelilingkoloni bakteri.
Uji Potensi Senyawa Bioaktif untukMenekan Pertumbuhan X. oryzae pv. oryzae.Potensi senyawa bioaktif bakteri agensia hayatidilakukan dengan menggunakan metode peracunanmedia (poisoned food technique) berdasarkan metodeSingh et al. (2005) yang dimodifikasi. Bakteri agensiahayati dibiakkan pada media NB, selama 5 hari untukkelompok bakteri aktinomiset dan 2 hari untuk kelompok

Kurniawati et al. Eksplorasi dan Uji Senyawa Bioaktif Bakteri 173
bakteri lainnya pada inkubator bergoyang 100 rpm.Selanjutnya bahan bioaktif yang dihasilkan bakteritersebut dipisahkan menggunakan filter (Corning NY14831 Germany) ukuran 0,45 µl. Media LB (Tryptone10 g, NaCl 5 g, yeast extract 5 g, akuades l000 ml)sebanyak 90 ml disiapkan untuk masing-masingperlakuan isolat bakteri dan dicampur dengan senyawabioaktif sebanyak 10 ml. Suspensi X. oryzae pv. oryzaepatotipe IV sebanyak 100 µl yang telah dibiakkan padamedia Wakimoto cair selama 24 jam (OD
600 0,610)
diinokulasikan pada media yang mengandung senyawabioaktif selanjutnya diinkubasikan pada inkubatorbergoyang dan diamati secara berkala nilai Absorbance(OD
600) sampai dengan 48 jam. Percobaan ini
menggunakan rancangan acak lengkap (RAL) dengan3 ulangan. Data dianalisis menggunakan Analysis ofVariance (ANOVA) dan dilanjutkan dengan uji Duncanpada taraf nyata 5%, menggunakan piranti lunakStatistical Analysis System (SAS) versi 9.1.3 untukWindows.
Identifikasi Isolat Bakteri dengan SekuensingParsial Gen Pengkode 16S rRNA. Identifikasi secaramolekuler dilakukan pada isolat bakteri yang mampumenghambat pertumbuhan bakteri dan memilikikarakteristik unggul (kemampuan kitinolitik, melarutkanfosfat atau memproduksi siderofor). Persiapan yangdilakukan sebelum melakukan proses sekuensing,terlebih dahulu dilakukan ekstraksi DNA yang mengacupada protokol kit ekstraksi DNA (Gene JET GenomicDNA Purification Kit # K0722), selanjutnya amplifikasigen 16S rRNA dengan Polymerase Chain Reaction(PCR) dan dilanjutkan dengan sekuensing. Data sekuengen 16S-rRNA yang diperoleh kemudian dimasukkandalam program BLASTN untuk dianalisis tingkathomologi atau kemiripannya dengan sekuen gen bakterilain yang ada pada data base.
Amplifikasi DNA kromosom bakteri dilakukandengan menggunakan sepasang primer general untuk
kelompok prokariotik (bakteri), yaitu 27F (5’-AGA GTTTGA TCC TGG CTC AG-3’) dan 1492R (5’-GGT TACCTT ACG ACT T-3’) (Lane, 1991). Reaksi PCRdilakukan pada volume total 50 µ l (Dream TaqGreenPCR Master Mix 2X1(Fermentas) 25 µl, Primer27 F 20 pmol, Primer 1492 20 pmol, ddH
2O 17 µl,
template 4-6 µl konsentrasi DNA 0,45-21,44 ng/µl).Proses PCR (menggunakan mesin Gene Amp PCRSystem 9700) terdiri dari 35 siklus dengan pemanasanawal pada suhu 95 oC selama 5 menit, denaturasi padasuhu 95 oC selama 1 menit, kemudian annealing padasuhu 55 oC selama 1 menit, ekstensi pada suhu 72 oCselama 1 menit dan ekstensi akhir pada suhu 72 oC selama10 menit. DNA hasil PCR kemudian dielektroforesisgel agarose 1% dalam 2X TAE buffer, pada 75 voltselama 35 menit. Hasil elektroforesis divisualisasi padatransiluminator ultraviolet untuk mengamati pita DNAyang terbentuk.
HASIL DAN PEMBAHASAN
Isolasi Bakteri Kandidat Agensia Hayati. Hasilisolasi bakteri dari bagian tanah, rizosfer dan endofitakar, batang dan daun padi pada penelitian ini diperoleh1145 isolat. Dari sejumlah isolat tersebut dipilih 156 isolatyang memiliki karakteristik koloni bakteri yangmenunjukkan kelompok bakteri yang berpotensi sebagaiagensia hayati seperti aktinomiset, Pseudomonaskelompok flourescens, Bacillus dan Chromobacter.Selanjutnya, pengujian aktivitas antibiosis dilakukan pada156 isolat bakteri tersebut terhadap X. oryzae pv. oryzaepatotipe III, IV dan VIII. Hasil pengujian antibiosisditunjukkan oleh terbentuknya zona bening antara bakteriagensia hayati dengan X. oryzae pv. oryzae (Gambar1) dan terdapat keragaman kemampuan dari masing-masing bakteri dalam membentuk zona bening (Tabel1). Pengujian lebih lanjut diperoleh 11 isolat bakteri yangdapat menekan X. oryzae pv. oryzae patotipe III, IV
Gambar 1. Uji antibiosis dengan menggunakan metode cross-streak. (A) isolat T5-1105, (B) kontrol, (C) uji antibiosisdengan metode double layer

174 J. HPT Tropika Vol. 15 No. 2, 2015: 170 - 179
dan VIII dan tidak memiliki potensi sebagai patogentumbuhan (Tabel 1).
Uji Kemampuan Antibiosis. Isolasi agensia hayatipada pertanaman padi telah banyak dilakukan,diantaranya adalah isolasi bakteri endofit dari padi gogoyang memiliki potensi antibiosis terhadap Rhizoctoniasolani dan Pyricularia grisea dan dapat memacupertumbuhan tanaman padi (Munif et al., 2012).Selanjutnya, Santosa et al. (2003), berhasil mengisolasibakteri yang berasal dari filosfer padi IR64 yang memilikipotensi sebagai agensia pemacu pertumbuhan. Adapunisolasi agensia hayati dari kelompok cendawan endofittelah dilakukan oleh Suada et al. (2012) yang dapatmenghambat pertumbuhan P. oryzae.
Patogenesitas. Pengujian reaksi hipersensitif(hypersensitive response/HR) pada tanaman tembakaudilakukan untuk mengetahui apakah suatu isolat bakteribersifat patogenik atau tidak. Jika suatu isolatdiinfiltrasikan pada daun tembakau dan menunjukkangejala nekrosis dalam waktu 24 jam, maka isolat tersebutmemiliki potensi patogenik. Nekrosis tidak terjadi padabakteri yang tidak memiliki potensi patogenik. Kematiansel yang cepat di dalam dan di sekitar sel yang terinfeksioleh patogen merupakan reaksi hipersensitif dan hal iniberasosiasi dengan respon ketahanan tanaman (Garcionet al., 2007).
Karakteristik Bakteri Agensia Hayati.Karakterisasi lebih lanjut 11 isolat bakteri agensia hayatimeliputi pengujian aktivitas kitinolitik, pelarutan fosfatdan produksi siderofor. Dari hasil pengujian tersebut
diperoleh empat isolat bakteri yang berpotensi memilikisalah satu atau lebih karakteristik unggulan (aktivitaskitinolitik, pelarutan fosfat dan produksi siderofor)sebagai agensia hayati (Tabel 2).
Aktivitas kitinolitik dimiliki oleh mikroba yangmenghasilkan ezim kitinase. Enzim ini salah satu yangberperan penting dalam mekanisme antibiosis yaitumenghancurkan dinding sel beberapa cendawan patogenseperti Sclerotium rolfsii, Fusarium oxysporum (Pal& Gardener, 2006) dan Rhizoctonia solani (Wahyudiet al., 2011). Bakteri kitinolotik telah banyak diketahuiberasal dari kelompok Bacillus, Serratia (Pal &Gardener, 2006), Aeromonas, Enterobacter,Paenibacillus, Streptomyces dan Chromobacterium(Park et al., 2005).
Adapun aktivitas pelarutan fosfat dan produksisiderofor merupakan mekanisme kompetisi nutrisi yangdilakukan oleh agensia hayati dengan patogen. Fosfatdi dalam tanah seringkali berada dalam bentuk yangterikat dengan kalsium pada tanah basa, sedangkan padatanah masam terikat dengan aluminum dan besi. Bakteripelarut fosfat seperti Bacillus, Pseudomonas danRhizobium dapat meningkatkan ketersediaan fosfat didalam tanah (Goldstein, 1995) bagi tanaman dan mikrobatanah lainnya. Hal ini berkaitan dengan dihasilkannyaasam organik seperti formic, acetic, propionic, lactic,glycolic, fumaric dan succinic yang dapatmeningkatkan pH tanah dan menghasilkan H+ denganmenggantikan Ca2+ kemudian melepaskan HPO
42- ke
dalam tanah (Yasmin et al., 2009).Demikian pula dengan ketersediaan besi yang
dapat diserap oleh organisme sangat rendah yaitu sekitar
Tabel 1. Hasil pengujian kemampuan antibiosis dan reaksi hipersensitif bakteri agensia hayati
Kode isolat
Zona hambat (mm) Asal isolat
Patotipe X. oryzae pv. oryzaeHabitat Kabupaten Agroekosistem
III IV VIII
T5-1118 3 5 3 Tanah Sleman Sawah tadah hujanR7-1018 3 3 2 Rhizosfer Subang Sawah irigasiEA4-1130 7 4 2 Endofit akar Sleman Sawah tadah hujanR7-1024 2 3 2 Rhizosfer Subang Sawah IrigasiT6-1112* 30 25 34 Tanah Sleman Sawah tadah hujanEA8-910 2 5 5 Endofit akar Barito Kuala RawaT6-1109* 45 40 39 Tanah Sleman Sawah tadah hujanR1-1095* 14 24 28 Rhizosfer Sleman Sawah tadah hujanT5-1105* 33 23 25 Tanah Sleman Sawah tadah hujanEB7-1032 3 5 2 Endofit batang Subang Sawah irigasiR7-1013 3 8 6 Rhizosfer Subang Sawah irigasi
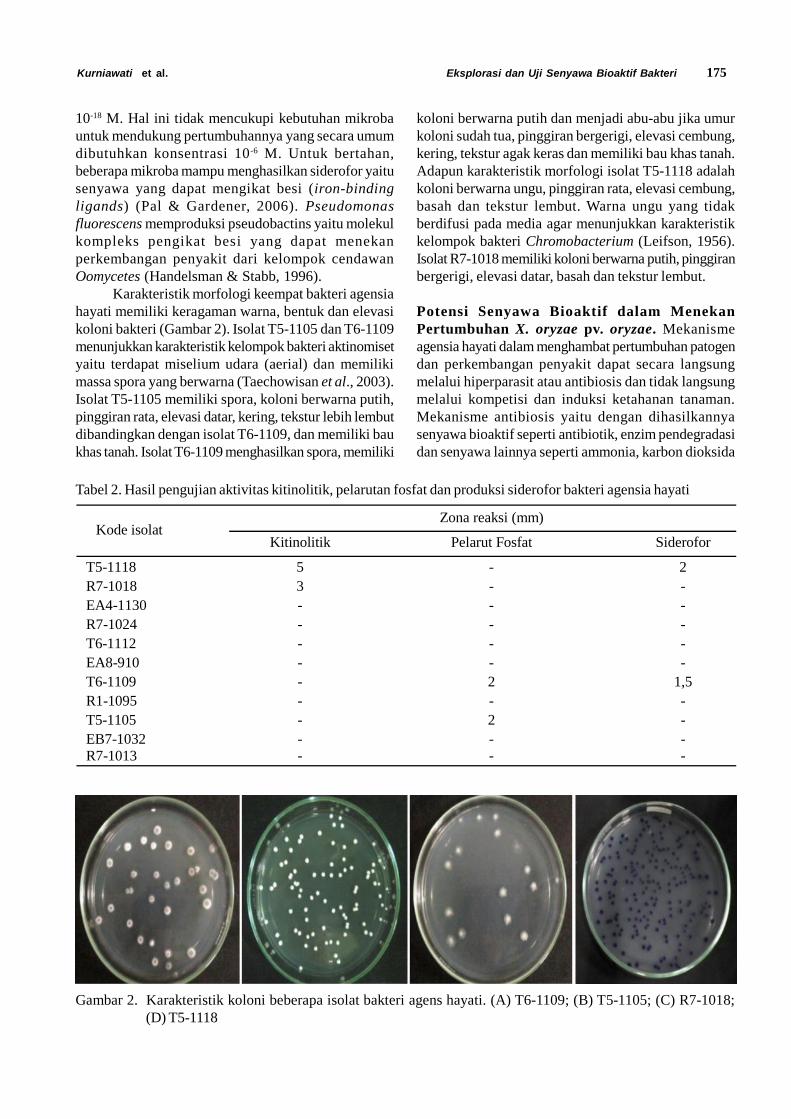
Kurniawati et al. Eksplorasi dan Uji Senyawa Bioaktif Bakteri 175
10-18 M. Hal ini tidak mencukupi kebutuhan mikrobauntuk mendukung pertumbuhannya yang secara umumdibutuhkan konsentrasi 10-6 M. Untuk bertahan,beberapa mikroba mampu menghasilkan siderofor yaitusenyawa yang dapat mengikat besi (iron-bindingligands) (Pal & Gardener, 2006). Pseudomonasfluorescens memproduksi pseudobactins yaitu molekulkompleks pengikat besi yang dapat menekanperkembangan penyakit dari kelompok cendawanOomycetes (Handelsman & Stabb, 1996).
Karakteristik morfologi keempat bakteri agensiahayati memiliki keragaman warna, bentuk dan elevasikoloni bakteri (Gambar 2). Isolat T5-1105 dan T6-1109menunjukkan karakteristik kelompok bakteri aktinomisetyaitu terdapat miselium udara (aerial) dan memilikimassa spora yang berwarna (Taechowisan et al., 2003).Isolat T5-1105 memiliki spora, koloni berwarna putih,pinggiran rata, elevasi datar, kering, tekstur lebih lembutdibandingkan dengan isolat T6-1109, dan memiliki baukhas tanah. Isolat T6-1109 menghasilkan spora, memiliki
koloni berwarna putih dan menjadi abu-abu jika umurkoloni sudah tua, pinggiran bergerigi, elevasi cembung,kering, tekstur agak keras dan memiliki bau khas tanah.Adapun karakteristik morfologi isolat T5-1118 adalahkoloni berwarna ungu, pinggiran rata, elevasi cembung,basah dan tekstur lembut. Warna ungu yang tidakberdifusi pada media agar menunjukkan karakteristikkelompok bakteri Chromobacterium (Leifson, 1956).Isolat R7-1018 memiliki koloni berwarna putih, pinggiranbergerigi, elevasi datar, basah dan tekstur lembut.
Potensi Senyawa Bioaktif dalam MenekanPertumbuhan X. oryzae pv. oryzae. Mekanismeagensia hayati dalam menghambat pertumbuhan patogendan perkembangan penyakit dapat secara langsungmelalui hiperparasit atau antibiosis dan tidak langsungmelalui kompetisi dan induksi ketahanan tanaman.Mekanisme antibiosis yaitu dengan dihasilkannyasenyawa bioaktif seperti antibiotik, enzim pendegradasidan senyawa lainnya seperti ammonia, karbon dioksida
Gambar 2. Karakteristik koloni beberapa isolat bakteri agens hayati. (A) T6-1109; (B) T5-1105; (C) R7-1018;(D) T5-1118
A B C D
Tabel 2. Hasil pengujian aktivitas kitinolitik, pelarutan fosfat dan produksi siderofor bakteri agensia hayati
Kode isolatZona reaksi (mm)
Kitinolitik Pelarut Fosfat Siderofor
T5-1118 5 - 2R7-1018 3 - -EA4-1130 - - -R7-1024 - - -T6-1112 - - -EA8-910 - - -T6-1109 - 2 1,5R1-1095 - - -T5-1105 - 2 -EB7-1032 - - -R7-1013 - - -

176 J. HPT Tropika Vol. 15 No. 2, 2015: 170 - 179
dan hidrogen sianida yang dapat membunuh ataumenghambat pertumbuhan mikroorganisme lain (Pal &Gardener, 2006). Antibiotik dapat menghambat sintesadinding sel, sintesa protein, sintesa DNA/RNA dansintesa koenzim folat (Walsh, 2003). Selain itu, senyawaantibiotik dapat berperan sebagai agensia penginduksi(elicitor) ketahanan tanaman terhadap penyakit (Lyon,2007).
Pengujian senyawa bioaktif dilakukan pada empatisolat yang memiliki karakteristik unggulan. Senyawabioaktif yang dihasilkan oleh isolat T6-1109, T5-1105,R7-1018 dan isolat T5-1118 menunjukkan potensimenghambat pertumbuhan X. oryzae pv. oryzaepatotipe IV secara nyata berdasarkan uji Duncan padataraf kepercayaan 95% (Gambar 3). Pertumbuhan X.oryzae pv. oryzae terendah dihasilkan oleh perlakuansenyawa bioaktif asal isolat T5-1118 dengan keefektifanpenekanan sebesar 66,61%. Selanjutnya diikuti berturut-turut isolat T5-1105, T6-1109 dan R7-1018 memilikiefektivitas penekanan sebesar 62,4; 23,97 dan 12,40%.
Pertumbuhan X. oryzae pv. oryzae dapatdihambat oleh senyawa bioaktif yang dihasilkan oleh keempat isolat bakteri agensia hayati diduga dapat melaluiberbagai mekanisme, diantaranya produksi senyawaantibiotik. Antibiotik kasugamycin yang dihasilkan olehStreptomyces kasugaensis bersifat bakterisida danfungisida. Antibiotik ini berperan sebagai penghambat(inhibitor) dalam sintesa protein pada mikroorganisme.Senyawa antibiotik ini telah banyak digunakan untukmengendalikan penyakit blast pada padi dan beberapapenyakit yang disebabkan oleh bakteri patogen kelompokPseudomonas (Doumbou et al., 2001). Antibiotiklainnya yang dihasilkan oleh aktinomiset adalahvancomycin, teicoplanin, nocardicin, dan thienamycindapat menghambat sintesa dinding sel (peptidoglycan),erythromycin, kanamycin dan oleandomycin dapatmengikat ribosom dan rifamycin dapat menghambatsintesa RNA (Walsh, 2003).
Mekanisme senyawa bioaktif dalam menghambatpertumbuhan patogen lainnya adalah produksi enzimpendegradasi senyawa polimerik seperti protein, kitin,selulosa, hemiselulosa dan DNA. Senyawa volatil yangdapat menghambat pertumbuhan mikroba adalahhidrogen sianida (HCN) dan amonia. HCN dapatmenghambat proses oksidasi cytochrome dan sangattoksik terhadap mikroorganisme yang bersifat aerob.Salah satu penghasil HCN adalah Pseudomonasfluorescens yang dapat menghambat perkembanganpenyakit black root pada tembakau. Adapun amoniaadalah senyawa yang bersifat volatil, salah satunyadihasilkan oleh Enterobacter cloacae dapat menekanperkembangan penyakit rebah kecambah pada kapas
yang disebabkan oleh Pythium ultimum (Pal &Gardener, 2006).
Identifikasi Isolat Bakteri dengan SekuensingParsial Gen Pengkode 16S rRNA. Keempat isolatbakteri yang menunjukkan potensi menghambatpertumbuhan X. oryzae pv. oryzae berdasarkan ujisenyawa bioaktif selanjutnya diidentifikasi secaragenotifik untuk memastikan identitas bakteri. Informasiini diperlukan untuk pengembangan lebih lanjut daribakteri agensia hayati tersebut. Hasil PCR DNA genpengkode 16S rRNA keempat isolat selanjutnyadianalisis dengan sekuensing secara parsial. Hasilanalisis sekuen menggunakan program BLASTN dipusat data Gen Bank menunjukkan isolat T5-1105 danT6-1109 dari kelompok bakteri Aktinomiset, isolat T5-1118 adalah Chromobacterium dan isolat R7-1018adalah Bacillus (Tabel 3).
Sekuensing 16S rRNA merupakan teknik yangsangat baik dan paling umum digunakan untukmengidentifikasi bakteri dengan membandingkanseberapa besar persamaan urutan nukleotidanya. Halini dikarenakan pada beberapa bagian 16 S rRNAmerupakan gen yang sangat konservatif. Oleh karenanya,penggunaan pasangan primer pada PCR dapatmengenali bagian gen yang konservatif tersebut danmengamplifikasinya. Produk PCR selanjutnyadisekuensing dan diperoleh urutan nukleotidanyakemudian dapat dibandingkan dengan data bakteri yangtelah diketahui dan dideterminasi (Dale & Park, 2010).
Kelompok bakteri aktinomiset telah banyakdiketahui sebagai penghasil beberapa senyawa antibiotikseperti polyketides, β-lactams dan peptida yangberfungsi sebagai anti fungal, anti tumor dan immunosuppresive (Behal, 2000). Genus Kitasatosporamerupakan kelompok bakteri aktinomiset yaitu dari familiStreptomycetaceae (Groth et al., 2004).
Hasil identifikasi genotif selanjutnya terhadapisolat T5-1118 (Chromobacterium sp.) dan isolat R7-1018 (Bacillus nealsonii), dapat menjelaskan bahwabakteri tersebut dapat menekan pertumbuhan X. oryzaepv. oryzae berdasarkan hasil-hasil penelitiansebelumnya. Kim et al. (2014) melaporkan bahwaChromobacterium sp. strain C61 menghasilkanantibiotik chromobactomycin yang dapat menghambatpertumbuhan beberapa cendawan patogen dan menekanperkembangan beberapa penyakit tumbuhan. Demikianhalnya dengan kelompok bakteri Bacillus juga telahbanyak diketahui menghasilkan antibiotik seperti, iturinA, surfactins (Beric et al., 2012), bacillomycin D,mycosubtilin dan zwittermicin A (Pal & Gardener, 2006).
Kurniawati et al. Eksplorasi dan Uji Senyawa Bioaktif Bakteri 177
Berdasarkan potensi senyawa bioaktif yangdihasilkan oleh keempat isolat tersebut maka, isolat iniberpotensi untuk dikembangkan sebagai agensia hayatidalam menekan penyakit kresek yang disebabkan olehX. oryzae pv. oryzae. Namun demikian perlu dilakukanpengujian lanjutan secara inplanta baik di rumah kacamaupun di lapangan untuk menguji keefektifannya dalammengendalikan penyakit kresek.
SIMPULAN
Dari seleksi bakteri dari pertanaman padidiperoleh 11 isolat bakteri agensia hayati yang berpotensidapat menghambat X. oryzae pv. oryzae patotipe III,IV dan VIII dan tidak bersifat patogenik terhadaptanaman. Karakterisasi terhadap 11 isolat tersebutmenunjukkan bahwa isolat-isolat tersebut memilikikemampuan kitinolitik (isolat T5-1118 dan R7-1018),melarutkan fosfat (T5-1105 dan T6-1109), dan
memproduksi siderofor (isolat T5-1118 dan T6-1109).Pengujian senyawa bioaktif dari masing-masing 4 isolattersebut terhadap pertumbuhan X. oryzae pv. oryzaesecara in vitro menunjukkan bahwa isolat T5-1118, T5-1105, T6-1109 dan R7-1018 dapat menghambatpertumbuhan X. oryzae pv. oryzae pada 48 jam setelahinokulasi masing-masing sebesar 66,61; 62,4; 23,97 dan12,40%. Selanjutnya hasil identifikasi dengan sekuensingparsial gen pengkode 16S rRNA terhadap keempat isolattersebut adalah Chromobacterium sp. MWU328 (T5-1118), Streptomyces sp. Antag 1 (T5-1105),Kitasatospora nipponensis strain H2-4 (T6-1109) danBacillus nealsonii strain F22 (R7-1018).
SANWACANA
Penelitian ini merupakan salah satu bagian darihasil penelitian KKP3N 2013 kerjasama dengan BadanPenelitian dan Pengembangan Pertanian, Kementrian
Tabel 3. Hasil analisis sekuen parsial gen 16S rRNA isolat bakteri dengan sekuen gen 16S rRNA di pusat dataGenBank
Kode isolat Spesies yang homolog Kemiripan (%) No aksesi
T5-1105 Streptomyces sp. Antag1 98 (JQ417268.1)T6-1109 Kitasatospora nipponensis H2-4 94 (HQ857768.1)R7-1018 Bacillus nealsonii strain F22 99 (JQ579625.1)T5-1118 Chromobacterium sp.MWU328 99 (JN653466.1)
Gambar 3. Pengaruh senyawa bioaktif bakteri agensia hayati terhadap pertumbuhan populasi X. oryzae pv.oryzae patotipe IV. Huruf yang sama menunjukkan tidak berpengaruh nyata secara statistikberdasarkan uji Duncan dengan taraf nyata 5% terhadap kontrol
Kontrol E. coli
Kontrol Xoo

178 J. HPT Tropika Vol. 15 No. 2, 2015: 170 - 179
Pertanian RI dengan judul: “Aplikasi Teknik Metagenomdalam Eksplorasi Agens Hayati dan Induksi Resistensiterhadap Penyakit Kresek yang Disebabkan olehXanthomonas oryzae pv. oryzae pada Tanaman Padi”,No. Kontrak : 691/LB.620/I.1/2/2013 tanggal 25 Februari2013.
DAFTAR PUSTAKA
Agustiansyah, Ilyas S, Sudarsono, & Machmud M. 2013.Karakterisasi rizobakteri yang berpotensimengendalikan bakteri Xanthomonas oryzae pv.oryzae dan meningkatkan pertumbuhan tanamanpadi. J. HPT Tropika 13(1): 42–51.
[BPS] Badan Pusat Statistik. 2014. Produksi padi, jagungdan kedelai (angka tetap tahun 2013). BeritaResmi Statistik. 50(7): 1–10.
Behal V. 2000. Bioactive products from Streptomyces.Adv. Appl. Microbiol. 47: 113–156.
Berić T, Kojić M, Stancović S, Topisirović L, DegrassiG, Myers M, Venturi V, & Fira D. 2012. Antimi-crobial activity of Bacillus sp. natural isolates andtheir potential use in the biocontrol of phytopatho-genic bacteria. Food Technol. Biotech. 50(1):25–31.
Dale JW & Park SF. 2010. Molecular Genetics ofBacteria. 5thEd.Wiley-Blackwell, Oxford.
Doumbou CL, Salove MKH, Crawford DL, & BeaulieuC. 2001. Actinomycetes, promising tools to controlplant diseases and promote plant growth.Phytoprotection 82(3): 85–102.
Garcion C, Lamotte O, & Métraux JP. 2007.Mechanisms of defence to pathogens:biochemistry and physiology. In: Walters D,Newton A, & Lyon G (Eds.). Induced Resistancefor Plant Defense: Sustainable Approach toCrop Protection. pp. 109–132. BlackwellPublishing, Oxford.
Goldstein AH. 1995. Recent progress in understandingthe molecular genetics and biochemistry ofcalcium phosphate solubilization by gram negativebacteria. Biol. Agric. Hort. 12(2): 185–193.
Gross M. 1990. Siderophores and fluorescent pigments.In: Klement Z, Rudolph K, & Sand DC (Eds.).Methods in Phytobacteriology. pp. 434–438.Budapest, Hungary.
Groth I, Rodriguez C, Schütze B, Schmitz P, LeistnerE, & Goodfellow M. 2004. Five novelKitasatospora species from soil: Kitasatosporaarboriphila sp. nov., K. gansuensis sp. nov., K.nipponensis sp. nov., K. paranensis sp. nov. andK. terrestris sp. nov. Int. J. Syst. Evol.Microbiol. 54: 2121–2129.
Hallmann J, Quadt-Hallmann A, Mahaffee WF, &Kloepper JW. 1997. Bacterial endophytes inagricultural crops. Can. J. Microbiol. 43(10):895–914.
Handelsman J & Stabb EV. 1996. Biocontrol of soilborneplant pathogens. Plant Cell. 8: 1855–1869.
Hastuti RD, Lestari Y, Saraswati R, Suwanto A, &Chaerani. 2012. Capability of Streptomyces spp.in controlling bacterial leaf blight disease in riceplants. Am. J. Agri. Biol. Sci. 7(2): 217–223.
Hoa PTP, Quang ND, Sakiyama Y, Hop DV, Hang DT,Ha TH, Van NT, Quy NTK, & Dao NTA. 2012.Screening for Actinomyces isolated from soil withthe ability to inhibit Xanthomonas oryzae pv.oryzae causing rice bacterial blight disease inVietnam. Afr. J. Biotechnol. 11(80): 14586–14594.
Kim HJ, Choi HS, Yang SY, Kim IS, Yamaguchi T, SohngJK, Park SK, Kim JC, Lee CH, Garderner BM,& Kim YC. 2014. Both extracellular chitinaseand new cyclic lipopeptide, chromobactomycin,contribute to the biocontrol activity ofChromobacterium sp. C61. Mol. PlantPathol.15(2): 122–132.
Lane DJ. 1991. 16S/23S rRNA sequencing. In:Stackebrandt E & Goodfellow M (Eds.). Nucleicacid Techniques in Bacterial Systematics. pp.115–175. John Wiley and Sons, Chichester, NewYork.
Leifson E. 1956. Morphological and physiologicalcharacteristics of the genus Chromobacterium.J. Bacteriol. 71(4): 393–400.
Lingappa Y & Lockwood JL. 1962. Chitin media forselective isolation and culture of actinomycetes.Phytopathology 52: 317–323.
Lisboa MP, Bonatto D, Bizani D, Henriques JAP, &Brandelli A. 2006. Characterization of abakteriosin-like substance produced by Bacillusamyloliquefaciens isolated from the BrazilianAtlantic forest. Int. Microbiol. 9: 111–118.

Kurniawati et al. Eksplorasi dan Uji Senyawa Bioaktif Bakteri 179
Lyon G. 2007. Agens that can elicit induced resistance.In: Walters D, Newton A, & Lyon G (Eds.).Induced Resistance for Plant Defense:Sustainable Approach to Crop Protection. pp.9–29. Blackwell Publishing, Oxford.
Madigan MT, Martinko JM, & Parker J. 1997. Brock’sBiology of Microorganisms. Ed ke-8. : Prentice-Hall, Inc New Jersey.
Munif A, Wiyono S, & Suwarno. 2012. Isolasi bakteriendofit asal padi gogo dan potensinya sebagaiagens biokontrol dan pemacu pertumbuhan. J.Fitopatol. Indones. 8(3): 57–64.
Pal KK & Gardener BM. 2006. Biological control ofplant pathogens. The Plant Health Instructor.DOI: 10.1094/PHI-A-2006-1117-02. APSnet 25p.
Park SK, Lee MC, & Harman GE. 2005. The biocontrolactivity of Chromobacterium sp. strain C-61against Rhizoctonia solani depends on theproductive ability of chitinase. Plant Pathol. J.21(3): 275–282.
Rao SWCB & Sinha MK. 1962. Phosphate dissolvingmicroorganism in the soil and rhizosphere. Ind.J. Sci. 23: 272–278.
Santosa DA, Handayani N, & Iswandi A. 2003. Isolasidan seleksi bakteri filosfer pemicu tumbuh daridaun padi (Oryza sativa L.) varietas IR64. J.Tanah dan Lingkungan 5(1): 7–12.
Singh G, Maurya S, deLampasona MP, & Catalan C.2005. Chemical constituents, antimicrobialinvestigations and antioxidative potentials ofAnethumgraveolens L. essential oil and acetoneextract part 52. J. Food Sci. 70(4): 208–215.
Suada IK, Suhartini DMWY, Sunariasih NPL, WirawanIGP, Chun KW, Cha JY, & Ohga S. 2012. Abilityof endophytic fungi isolated from rice to inhibitPyricularia oryzae-induced rice blast inIndonesia. J. Fac. Agr. 57(1): 51–53.
Suparyono, Sudir, & Suprihanto. 2004. Pathotype profileof Xanthomonas oryzae pv. oryzae isolates fromthe rice ecosystem in Java. Indones. J. Agric.Sci. 5(2): 63–69.
Suparyono & Sudir. 1992. Perkembangan penyakitbakterihawar daun pada stadia tumbuh yangberbeda dan pengaruhnya terhadap hasil padi.Media Penelitian Sukamandi 12: 6–9.
Taechowisan T, Peberdy JF, & Lumyong S. 2003.Isolation of endophytic actinomycetes fromselected plants and their antifungal activity. WorldJ. Microbiol. Biotechnol. 19(4): 381–385.
Velusamy P, Immanuel JE, Gnanamanickam SS, &Thomashow L. 2006. Biological control of ricebacterial blight by plant-associated bacteriaproducing 2,4-diacetylphloroglucinol. Can. J.Microbiol. 52: 56–65.
Wahyudi AT, Astuti RI, & Giyanto. 2011. Screening ofPseudomonas sp. isolated from rhizosphere ofsoybean plant as plant growth promoter andbiocontrol agent. Am. J. Agric. Biol. Sci. 6(1):134–141.
Walsh C. 2003. Antibiotics: Action, Origins,Resistance. 1st Edition. ASM Press, Washington.
Yasmin F, Othman R, Sijam K, & Saad MS. 2009.Characterization of beneficial properties of plantgrowth-promoting rhizobacteria isolated fromsweet potato rhizosphere. Afr. J. Microbiol. Res.3(11): 815–821.

180 J. HPT Tropika Vol. 15 No. 2, 2015: 180 - 187J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 180 – 187, September 2015
IDENTIFIKASI FENOTIPIK GALUR-GALUR KEDELAITERHADAP KETAHANAN SERANGAN HAMA ULAT GRAYAK
(SPODOPTERA LITURA F.)
Gatut Wahyu Anggoro Susanto & Moh. Muchlish Adie
Balai Penelitian Tanaman Aneka Kacang dan Umbi, Malang, Jawa TimurJl. Raya Kendalpayak Km 8, Desa Kendalpayak, Kecamatan Kedungkandang, Kabupaten Malang
E-mail: [email protected]
ABSTRACT
Phenotypic identification of soybean lines against armyworm pest resistance (Spodoptera litura F). This research consistedof two parts which aimed to find out the intensity of damage to the leaves and the influence of soybean on the biologicalaspect of armyworms. The materials tested were Shr/W-C-60, Aochi/Wil-60, 9837/K-D-8-185, 9837/K-D-3-185-95, W/9837-D-6-220, 9837/K-D-3-185-82, 9837/W-D-5-211, GI, G100H breeding lines and Wilis varieties. The research was conducted at aBalitkabi screenhouse in February 2011, using randomized block design and each treatment was replicated three times. Theplanting media were plastic pots (diameter 18 cm) filled with earth, two seeds per pot were planted and intensively raised.When the plants were 27 days old after planting, at each replication consisting of 10 materials, they were covered with gauzecages (2 x 2 x 2 m). Then the plants in each pot was infested with 10 instar I armyworm larvae. The other part of research wascarried out in Balitkabi Breeding Improvement Laboratory. For testing purposes, there was a need for a third nodal leaf ofsoybean aged 27 days after planting in each tested material. The research used a completely randomized design each treatmentwas replicated three times. One nodal leaf in each material was placed in a petri dish 15 cm in diameter, which was coated withmoist filter paper, and this was later called treatment. Each treatment was infested with one instar I armyworm larva. The resultof research indicated that an assessment of soybean resistance to armyworms could be made from the density of the trichomeand/or the length of the trichome on the leaves. The G100H breeding line was found to be resilient with its characteristics ofhaving dense trichome (25/4 mm2) and long trichome (1.1 mm).
Key words: caterpillar army worm, soybean, trichoma
ABSTRAK
Identifikasi fenotipik galur-galur kedelai terhadap ketahanan serangan hama ulat grayak (Spodoptera litura F). Penelitianterdiri dua bagian yang bertujuan untuk mengetahui intensitas kerusakan daun dan pengaruh kedelai terhadap aspek biologiulat grayak. Materi yang diuji adalah galur Shr/W-C-60, Aochi/Wil-60, 9837/K-D-8-185, 9837/K-D-3-185-95, W/9837-D-6-220,9837/K-D-3-185-82, 9837/W-D-5-211, GI, G100H dan varietas Wilis. Penelitian dilakukan di rumah kasa Balitkabi pada Februari2011, dirancang menggunakan acak kelompok setiap perlakuan diulang tiga kali. Media tanam pot plastik (diameter 18 cm)yang diisi tanah, ditanam dua biji/pot dan dipelihara secara intensif. Saat tanaman berumur 27 hari setelah tanam (HST), padasetiap ulangan yang terdiri dari 10 materi disungkup dengan kurungan kasa (2 x 2 x 2 m). Selanjutnya tanaman pada masing-masing pot diinfestasi dengan larva ulat grayak instar I sebanyak 10 ekor. Penelitian lainnya dilakukan di LaboratoriumPemuliaan Balitkabi. Untuk pengujiannya dibutuhkan daun buku ketiga kedelai yang berumur 27 HST pada masing-masingmateri yang diuji. Penelitian dirancang menggunakan acak lengkap yang diulang tiga kali. Satu daun buku ketiga pada setiapmateri ditempatkan pada cawan Petri berdiameter 15 cm yang telah dilapisi kertas saring yang basah air (lembab), dan inikemudian disebut sebagai perlakuan. Setiap perlakuan diinfestasi larva ulat grayak instar I sebanyak satu ekor. Hasil penelitianmenunjukkan penilaian ketahanan kedelai terhadap ulat grayak dapat ditilik dari kerapatan trikoma dan atau panjang trikomadaun. Galur G100H dinilai tahan dengan karakteristik trikoma padat (25/4 mm2) dan panjang (1,1 mm).
Kata kunci: kedelai, trikoma, ulat grayak
PENDAHULUAN
Serangan hama merupakan salah satu masalahpenting budidaya kedelai. Ulat grayak (Spodopteralitura F.) merupakan serangga hama yang bersifat
polifag (Ravishankar & Venkatesha, 2010) sehinggahama tersebut memiliki penyebaran dan kisaran inangyang cukup luas. Di Indonesia ulat grayak merupakanhama pemakan daun utama pada tanaman kedelai(Arifin & Koswanudin, 2010; Suharsono, 2011),

Susanto & Adie Identifikasi Fenotipik Galur-Galur Kedelai 181
penurunan hasil kedelai akibat serangan hama ulatgrayak dapat mencapai 90%, tergantung pada fasepertumbuhan dan waktu serangan (Suharsono et al.,2007; Marwoto & Suharsono, 2008) maupun varietasyang digunakan (Adie et al., 2012). Hal tersebutmengindikasikan bahwa ulat grayak merupakan hamapotensial merugikan yang perlu mendapatkanpenanganan secara bijaksana.
Tingkat ketahanan suatu varietas terhadap hamatertentu adalah bersifat relatif, karenanya pendekatanpengukuran ketahanan dapat didasarkan pada tingkatkepekaan dari suatu varietas unggul yang digunakansebagai standar perbaikan. Identifikasi yang dilakukanoleh Igita et al. (1996) tidak satupun tanaman kedelaiasal Indonesia yang bereaksi tahan terhadap ulat grayak,termasuk Wilis merupakan varietas yang berdaya hasiltinggi dan memiliki penyebaran yang cukup luas(Heriyanto, 2012). Berdasarkan tingkat kepekaan yangterdapat pada varietas Wilis dapat dijadikan sebagaikriteria perbaikan ketahanan suatu varietas terhadap ulatgrayak. Galur-galur yang merupakan hasil persilanganantara genotipe tahan dengan varietas unggul diharapkanmenghasilkan galur yang memiliki ketahanan sekaligusberproduksi tinggi. Tersedianya varietas kedelai tahanterhadap hama utama diharapkan mampumempertahankan stabilitas hasil kedelai.
Beberapa hasil penelitian melaporkan bahwa hasilkajiannya tentang penggunaan pupuk N sebesar 112,5kg urea/ha dapat meningkatkan perkembangan larva S.litura; durasi larva, terutama pada instar IV, VI, prepupadan durasi seluruh larva dibandingkan pemberian pupuk37,5 kg/ha pada tanaman kedelai (Hartati, 2009). Artinya,pemberian pupuk yang berbeda berpengaruh nyataterhadap perkembangan ulat grayak pada stadia tertentu.
Mekanisme ketahanan tanaman terhadap hamadapat disebabkan karena faktor antibiosis (Komatsu etal., 2004). Ketahanan antibiosis merupakan mekanismeketahanan tanaman yang disebabkan oleh adanya zatkimia tertentu yang mampu menghambat bahkanmematikan serangga hama. Menurut Singh (1986)terganggunya proses metabolisme fisiologis larva akibatadanya faktor antibiosis dapat berupa kematian larvaterutama pada instar awal, pengurangan ukuran dan beratlarva, kematian sebelum mencapai fase dewasa atauterganggunya siklus hidup dari larva itu sendiri. Dengandemikian pengukuran uji antibiosis dapat berdasar padaperubahan yang terjadi pada larva akibat mengkonsumsidaun dari setiap galur kedelai yang akan diuji. Penelitianbertujuan untuk mengetahui tingkat toleransi galurkedelai terhadap serangan ulat grayak.
METODE PENELITIAN
Tempat dan Waktu. Penelitian ini dilakukan di rumahkasa dan di Laboratorium Pemuliaan Balitkabi Malangpada bulan Februari 2011.
Pengujian. Penelitian terdiri dari dua bagian kegiatanyang bertujuan untuk mengetahui intensitas kerusakandaun dan pengaruh daun kedelai terhadap aspek biologiulat grayak. Materi yang diuji kedua penelitian ini adalahgalur kedelai hasil persilangan tunggal yang telahmencapai homosigot secara fenotipik yaitu Shr/W-C-60, Aochi/Wil-60, 9837/K-D-8-185, 9837/K-D-3-185-95,W/9837-D-6-220, 9837/K-D-3-185-82, 9837/W-D-5-211, GI, G100H dan varietas Wilis.
Penelitian di rumah kasa dirancang menggunakanacak kelompok yang diulang sebanyak tiga kali, ujianalisis beda nyata terkecil (BNT) dilakukan jika hasilanalisi uji F nyata > 95%. Media tanam yangdipergunakan adalah pot plastik (Ø 18 cm) yang telahterisi tanah, ditanam dua biji per pot dan dipelihara secaraintensif. Saat tanaman berumur 27 hari, tanamandisungkup dengan kurungan kasa (2 x 2 x 2 m). Satusungkup merupakan satu ulangan yang berisi 10 pot(materi yang di uji), sehingga seluruhnya terdapat tigakurungan kasa. Penyusunan tanaman pada pot diatursedemikian rupa supaya daun antar tanaman kedelaisaling bersentuhan sehingga memungkinkan bagi larvauntuk memilih daun tanaman kedelai yang disukai secarabebas. Selanjutnya tanaman pada pot yang berisi duatanaman diinvestasikan larva ulat grayak instar I masing-masing sebanyak 5 ekor larva, sehingga dalam satu potberjumlah 10 ekor larva. Bahan larva ulat grayak yangdinvestasi ke tanaman merupakan hasil pembiakanmelalui telur ulat grayak yang diperoleh dari pertanamankedelai di lapang. Telur-telur ulat grayak ditempatkanpada kotak kurungan dipelihara hingga menjadi menjadiimago. Selama periode pembiakan diberi makan daunjarak dan dijaga kehidupannya secara intensif.Selanjutnya, telur imago yang telah menetas menjadistadia larva instar I digunakan sebagai bahan investasidalam kegiatan penelitian.
Variabel yang diamati adalah intensitas kerusakandaun tanaman (%) pada umur 4, 8, 12 dan 16 hari setelahinvestasi (HSI). Intensitas kerusakan daun dihitungmengikuti cara yang dilakukan oleh Rusdy (2009),Leatemia & Rumthe (2011), Sembiring et al. (2013),dan Luhukay et al. (2013) dengan rumus sebagai berikut:
100%NZ
v)(nI
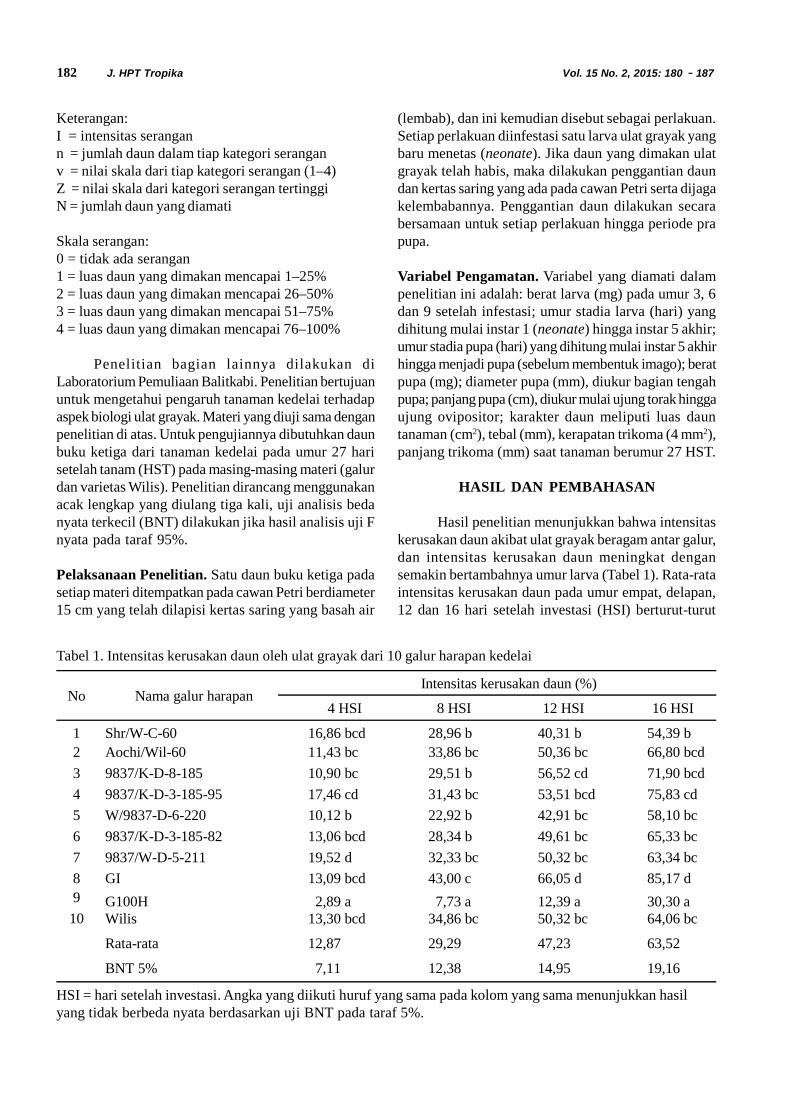
182 J. HPT Tropika Vol. 15 No. 2, 2015: 180 - 187
Keterangan:I = intensitas serangann = jumlah daun dalam tiap kategori seranganv = nilai skala dari tiap kategori serangan (1–4)Z = nilai skala dari kategori serangan tertinggiN = jumlah daun yang diamati
Skala serangan:0 = tidak ada serangan1 = luas daun yang dimakan mencapai 1–25%2 = luas daun yang dimakan mencapai 26–50%3 = luas daun yang dimakan mencapai 51–75%4 = luas daun yang dimakan mencapai 76–100%
Penelitian bagian lainnya dilakukan diLaboratorium Pemuliaan Balitkabi. Penelitian bertujuanuntuk mengetahui pengaruh tanaman kedelai terhadapaspek biologi ulat grayak. Materi yang diuji sama denganpenelitian di atas. Untuk pengujiannya dibutuhkan daunbuku ketiga dari tanaman kedelai pada umur 27 harisetelah tanam (HST) pada masing-masing materi (galurdan varietas Wilis). Penelitian dirancang menggunakanacak lengkap yang diulang tiga kali, uji analisis bedanyata terkecil (BNT) dilakukan jika hasil analisis uji Fnyata pada taraf 95%.
Pelaksanaan Penelitian. Satu daun buku ketiga padasetiap materi ditempatkan pada cawan Petri berdiameter15 cm yang telah dilapisi kertas saring yang basah air
(lembab), dan ini kemudian disebut sebagai perlakuan.Setiap perlakuan diinfestasi satu larva ulat grayak yangbaru menetas (neonate). Jika daun yang dimakan ulatgrayak telah habis, maka dilakukan penggantian daundan kertas saring yang ada pada cawan Petri serta dijagakelembabannya. Penggantian daun dilakukan secarabersamaan untuk setiap perlakuan hingga periode prapupa.
Variabel Pengamatan. Variabel yang diamati dalampenelitian ini adalah: berat larva (mg) pada umur 3, 6dan 9 setelah infestasi; umur stadia larva (hari) yangdihitung mulai instar 1 (neonate) hingga instar 5 akhir;umur stadia pupa (hari) yang dihitung mulai instar 5 akhirhingga menjadi pupa (sebelum membentuk imago); beratpupa (mg); diameter pupa (mm), diukur bagian tengahpupa; panjang pupa (cm), diukur mulai ujung torak hinggaujung ovipositor; karakter daun meliputi luas dauntanaman (cm2), tebal (mm), kerapatan trikoma (4 mm2),panjang trikoma (mm) saat tanaman berumur 27 HST.
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa intensitaskerusakan daun akibat ulat grayak beragam antar galur,dan intensitas kerusakan daun meningkat dengansemakin bertambahnya umur larva (Tabel 1). Rata-rataintensitas kerusakan daun pada umur empat, delapan,12 dan 16 hari setelah investasi (HSI) berturut-turut
No Nama galur harapanIntensitas kerusakan daun (%)
4 HSI 8 HSI 12 HSI 16 HSI
1 Shr/W-C-60 16,86 bcd 28,96 b 40,31 b 54,39 b2 Aochi/Wil-60 11,43 bc 33,86 bc 50,36 bc 66,80 bcd
3 9837/K-D-8-185 10,90 bc 29,51 b 56,52 cd 71,90 bcd
4 9837/K-D-3-185-95 17,46 cd 31,43 bc 53,51 bcd 75,83 cd
5 W/9837-D-6-220 10,12 b 22,92 b 42,91 bc 58,10 bc
6 9837/K-D-3-185-82 13,06 bcd 28,34 b 49,61 bc 65,33 bc
7 9837/W-D-5-211 19,52 d 32,33 bc 50,32 bc 63,34 bc
8 GI 13,09 bcd 43,00 c 66,05 d 85,17 d9 G100H 2,89 a 7,73 a 12,39 a 30,30 a10 Wilis 13,30 bcd 34,86 bc 50,32 bc 64,06 bc
Rata-rata 12,87 29,29 47,23 63,52
BNT 5% 7,11 12,38 14,95 19,16
Tabel 1. Intensitas kerusakan daun oleh ulat grayak dari 10 galur harapan kedelai
HSI = hari setelah investasi. Angka yang diikuti huruf yang sama pada kolom yang sama menunjukkan hasilyang tidak berbeda nyata berdasarkan uji BNT pada taraf 5%.
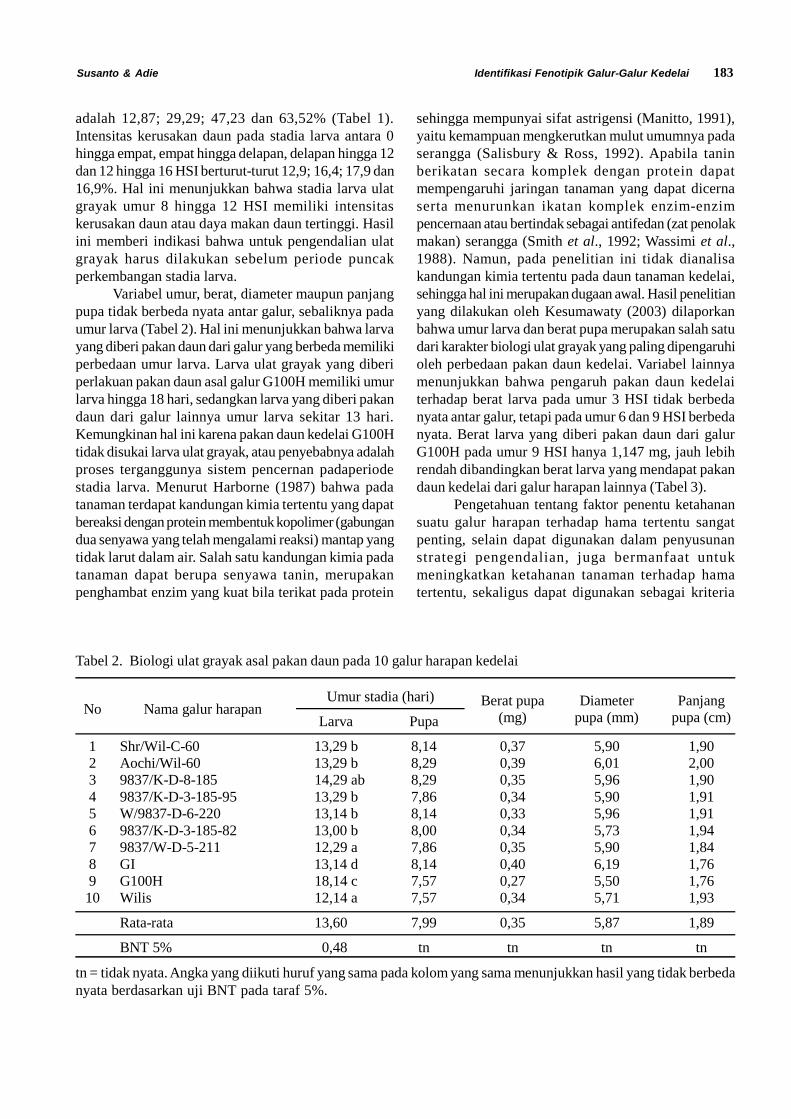
Susanto & Adie Identifikasi Fenotipik Galur-Galur Kedelai 183
adalah 12,87; 29,29; 47,23 dan 63,52% (Tabel 1).Intensitas kerusakan daun pada stadia larva antara 0hingga empat, empat hingga delapan, delapan hingga 12dan 12 hingga 16 HSI berturut-turut 12,9; 16,4; 17,9 dan16,9%. Hal ini menunjukkan bahwa stadia larva ulatgrayak umur 8 hingga 12 HSI memiliki intensitaskerusakan daun atau daya makan daun tertinggi. Hasilini memberi indikasi bahwa untuk pengendalian ulatgrayak harus dilakukan sebelum periode puncakperkembangan stadia larva.
Variabel umur, berat, diameter maupun panjangpupa tidak berbeda nyata antar galur, sebaliknya padaumur larva (Tabel 2). Hal ini menunjukkan bahwa larvayang diberi pakan daun dari galur yang berbeda memilikiperbedaan umur larva. Larva ulat grayak yang diberiperlakuan pakan daun asal galur G100H memiliki umurlarva hingga 18 hari, sedangkan larva yang diberi pakandaun dari galur lainnya umur larva sekitar 13 hari.Kemungkinan hal ini karena pakan daun kedelai G100Htidak disukai larva ulat grayak, atau penyebabnya adalahproses terganggunya sistem pencernan padaperiodestadia larva. Menurut Harborne (1987) bahwa padatanaman terdapat kandungan kimia tertentu yang dapatbereaksi dengan protein membentuk kopolimer (gabungandua senyawa yang telah mengalami reaksi) mantap yangtidak larut dalam air. Salah satu kandungan kimia padatanaman dapat berupa senyawa tanin, merupakanpenghambat enzim yang kuat bila terikat pada protein
sehingga mempunyai sifat astrigensi (Manitto, 1991),yaitu kemampuan mengkerutkan mulut umumnya padaserangga (Salisbury & Ross, 1992). Apabila taninberikatan secara komplek dengan protein dapatmempengaruhi jaringan tanaman yang dapat dicernaserta menurunkan ikatan komplek enzim-enzimpencernaan atau bertindak sebagai antifedan (zat penolakmakan) serangga (Smith et al., 1992; Wassimi et al.,1988). Namun, pada penelitian ini tidak dianalisakandungan kimia tertentu pada daun tanaman kedelai,sehingga hal ini merupakan dugaan awal. Hasil penelitianyang dilakukan oleh Kesumawaty (2003) dilaporkanbahwa umur larva dan berat pupa merupakan salah satudari karakter biologi ulat grayak yang paling dipengaruhioleh perbedaan pakan daun kedelai. Variabel lainnyamenunjukkan bahwa pengaruh pakan daun kedelaiterhadap berat larva pada umur 3 HSI tidak berbedanyata antar galur, tetapi pada umur 6 dan 9 HSI berbedanyata. Berat larva yang diberi pakan daun dari galurG100H pada umur 9 HSI hanya 1,147 mg, jauh lebihrendah dibandingkan berat larva yang mendapat pakandaun kedelai dari galur harapan lainnya (Tabel 3).
Pengetahuan tentang faktor penentu ketahanansuatu galur harapan terhadap hama tertentu sangatpenting, selain dapat digunakan dalam penyusunanstrategi pengendalian, juga bermanfaat untukmeningkatkan ketahanan tanaman terhadap hamatertentu, sekaligus dapat digunakan sebagai kriteria
tn = tidak nyata. Angka yang diikuti huruf yang sama pada kolom yang sama menunjukkan hasil yang tidak berbedanyata berdasarkan uji BNT pada taraf 5%.
No Nama galur harapanUmur stadia (hari) Berat pupa
(mg)Diameter
pupa (mm)Panjang
pupa (cm)Larva Pupa
1 Shr/Wil-C-60 13,29 b 8,14 0,37 5,90 1,902 Aochi/Wil-60 13,29 b 8,29 0,39 6,01 2,003 9837/K-D-8-185 14,29 ab 8,29 0,35 5,96 1,904 9837/K-D-3-185-95 13,29 b 7,86 0,34 5,90 1,915 W/9837-D-6-220 13,14 b 8,14 0,33 5,96 1,916 9837/K-D-3-185-82 13,00 b 8,00 0,34 5,73 1,947 9837/W-D-5-211 12,29 a 7,86 0,35 5,90 1,848 GI 13,14 d 8,14 0,40 6,19 1,769 G100H 18,14 c 7,57 0,27 5,50 1,7610 Wilis 12,14 a 7,57 0,34 5,71 1,93
Rata-rata 13,60 7,99 0,35 5,87 1,89
BNT 5% 0,48 tn tn tn tn
Tabel 2. Biologi ulat grayak asal pakan daun pada 10 galur harapan kedelai
184 J. HPT Tropika Vol. 15 No. 2, 2015: 180 - 187
No Nama galur harapanBerat larva (mg/ekor)
3 HSI 6 HSI 9 HSI
1 Shr/W-C-60 0,022 0,463 bcde 1,818 ab2 Aochi/Wil-60 0,026 0,549 de 2,455 c3 9837/K-D-8-185 0,027 0,503 cde 2,340 c4 9837/K-D-3-185-95 0,025 0,487 cde 2,088 bc5 W/9837-D-6-220 0,027 0,410 bc 1,499 ab6 9837/K-D-3-185-82 0,026 0,445 bcd 2,015 bc7 9837/W-D-5-211 0,024 0,354 ab 1,607 ab8 GI 0,028 0,550 de 1,501 ab9 G100H 0,022 0,323 a 1,147 a10 Wilis 0,027 0,536 cde 1,993 bc
Rata-rata 0,026 0,462 1,860
BNT 5% tn 0,11 0,72
Tabel 3. Berat larva ulat grayak asal pakan daun pada 10 galur harapan kedelai
tn = tidak nyata. Angka yang diikuti huruf yang sama pada kolom yang sama menunjukkan hasil yang tidak berbedanyata berdasarkan uji BNT pada taraf 5%.
seleksi tak langsung. Penentuan dan pencarian kriteriaseleksi tak langsung sangat penting, karena akanmeningkatkan efisiensi program pemuliaan.
Di antara galur-galur yang diuji maka G100Hmemiliki daun lebih tebal, trikoma yang lebih rapat, sertatrikoma yang panjang (Tabel 4). Berkaitan dengankarakter-karakter tersebut menunjukkan bahwa galurG100H memiliki intensitas kerusakan daun yangterendah, berbeda dengan galur lainnya maupun denganvarietas cek Wilis. Norris & Kogan (1980)mengemukakan bahwa karakter trikoma sebagai faktorpertahanan fisik terhadap ulat grayak ditentukan olehkerapatan dan panjang trikoma. IAC-100 merupakangalur yang bertrikoma padat konsisten tahan terhadapserangan hama pemakan daun, terutama ulat grayak(Suharsono et al., 2008; Suharsono & Adie, 2010),maupun G 100 H (Suharsono & Suntono, 2007) hal inimembuktikan bahwa trikoma berperan dalampertahanan terhadap serangan hama ulat grayak.
Karakter morfologi penentu ketahanan kedelaiterhadap ulat grayak dikaji dengan analisis korelasi daribeberapa karakter morfologi daun dengan intensitaskerusakan daun maksimum yang dicapai setelah infestasilarva selama 16 hari (Tabel 5). Terdapat dua karaktermorfologi daun yang berperan sebagai penentuketahanan kedelai terhadap ulat grayak yakni kerapatantrikoma pada permukaan daun bagian atas dan panjangtrikoma daun pada permukaan bawah daun, masing-masing dengan nilai r = -0,753 dan r = -0,689. Menilih
hasil ini maka galur G100H memiliki ketahatan terhadapulat grayak dengan karakteristik bertrikoma padat danpanjang dibandingkan dengan galur lainnya maupunvarietas cek Wilis. Adapun peran karakter ketebalandan luas daun terhadap ketahanan kedelai terhadap ulatgrayak relatif kecil.
Struktur daun kedelai nampaknya belum terlalukuat menahan larva ulat grayak yang bertipe mulutmenggigit dan mengunyah. Intensitas serangan padadaun tergantung pada populasi ulat grayak. Makin tinggipopulasi ulat grayak makin tinggi intensitas seranganakibat aktivitas ulat mengkonsumsi daun kedelai sebagaisumber pakan (Suharsono, 2011). Kajian untuk tanamanserealia, Blum (1982) mengemukakan bahwa varietassorgum yang tahan terhadap lalat bibit Atherigonavaria, ditentukan oleh adanya daun-daun muda yangmempunyai dinding sel tebal, sehingga berpengaruhterhadap proses penggangguan sistem pencernaan. Polayang sama juga ditemukan pada hama belalang hijauMelanoplus confusus yang tidak mampu mencernadaun rumput-rumputan (Panicum virgatu,Andropogon gerardi dan Schizachyrium scoparum).
Kerapatan trikoma daun permukaan atas danbawah berkorelasi nyata positif, pola yang samaditemukan juga pada karakter panjang trikoma. Faktaini sekaligus mengindikasikan bahwa penilaian ketahanankedelai terhadap ulat grayak dapat ditilik dari kerapatantrikoma daun dan atau panjang trikoma. Seperti hasilpenelitian Hendrival et al. (2013) yang menyimpulkan
Susanto & Adie Identifikasi Fenotipik Galur-Galur Kedelai 185
No Nama galur harapanKetebalandaun (mm)
Kerapatan trikoma/4 mm2 Panjang trikoma (mm) Luasdaun/
tanaman(cm2)
Bagianpermukaan
atas
Bagianpermukaan
bawah
Bagianpermukaan
atas
Bagianpermukaan
bawah
1 Shr/W-C-60 0,80 cdef 15,33 a 11,00 ab 0,52 b 0,52 b 32,04 cd2 Aochi/Wil-60 0,70 abcd 15,67 a 10,00 a 1,00 c 0,87 c 59,37 e3 9837/K-D-8-185 0,67 abc 15,33 a 16,00 c 0,57 b 0,57 b 25,44 bc4 9837/K-D-3-185-95 0,67 abc 14,67 a 15,67 bc 0,63 b 0,60 b 51,65 e5 W/9837-D-6-220 0,63 ab 18,67 a 16,33 c 0,57 b 0,80 c 36,95 d6 9837/K-D-3-185-82 0,87 def 14,67 a 15,67 bc 0,60 b 0,45 b 35,85 d7 9837/W-D-5-211 0,90 ef 15,67 a 15,67 bc 1,40 d 1,10 d 36,04 d8 GI 0,77 bcde 15,00 a 10,00 a 0,23 a 0,10 a 20,72 a9 G100H 0,97 f 25,00 b 29,00 e 1,07 c 1,10 d 55,23 e10 Wilis 0,57 a 16,00 a 22,67 d 0,47 b 0,50 b 29,96 cd
Rata-rata 0,75 16,60 16,20 0,70 0,66 38,32
BNT 5% 0,17 5,82 4,29 0,23 0,16 8,48
Tabel 4. Karakter morfologi daun dari 10 galur harapan kedelai
BNT = beda nyata terkecil. Angka sekolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut ujiBNT 5%.
SifatKerusakan
daun 16 HSITBDN KR-A KR-B PJ-A PJ-B
TBDN 0,101 1,000KR-A -0,753** -0,473 1,000KR-B -0,629 -0,335 0,744* 1,000PJ-A -0,561 -0,073 0,528 0,045 1,000PJ-B -0,689* -0,310 0,783** 0,291 0,910** 1,000LD 0,451 -0,159 0,082 -0,361 0,129 0,151
Tabel 5.Korelasi antara intensitas kerusakan daun 16 HSI dengan karakter morfologi daun
*nyata pada p = 0,05 ; ** = nyata pada p = 0,01; TBDN = tebal daun; KR-A = kerapatan trikoma daun bagianpermukaan atas; KR-B = kerapatan trikoma daun bagian permukaan bawah; PJ-A = panjang trikoma daun bagianpermukaan atas; PJ-B = panjang trikoma daun bagian permukaan bawah; LD = luas daun/tanaman.
bahwa trikoma pada daun kedelai berperan terhadapketahahanan morfologis terhadap hama ulat grayak.Berdasarkan aspek kepraktisan mengukur kepadatantrikoma lebih mudah dibandingkan dengan panjangtrikoma, karenanya karakter kepadatan trikoma, baikpada daun pada permukaan atas maupun bawah, dapatdipertimbangkan digunakan sebagai kriteria seleksiketahanan kedelai terhadap hama ulat grayak.Berdasarkan serangkaian penelitian ini, maka galurG100H memiliki trikoma padat di daun dan berkriteriatahan serangan ular grayak. Artinya kewaspadaan
diperlukan untuk mengantisipasi infestasi ulat grayak,agar kerugian hasil dapat dihindari.
SIMPULAN
Untuk menilai tingkat ketahanan tanaman kedelaiterhadap ulat grayak dapat ditilik dari kerapatan trikomadan atau panjang trikoma daun. Galur G100H berkriteriatahan serangan ulat grayak dibandingkan dengan galur-galur yang diuji maupun varietas cek Wilis.

186 J. HPT Tropika Vol. 15 No. 2, 2015: 180 - 187
SANWACANA
Terima kasih kepada DIPA Badan LitbangPertanian yang membiayai penelitian dan kepada Toni(teknisi pemuliaan Balitkabi Malang) yang membantuserangkaian penelitian.
DAFTAR PUSTAKA
Adie MM, Krisnawati A, & Mufidah AZ. 2012. Derajatketahanan genotipe kedelai terhadap hama ulatgrayak. Dalam: Rahmianna AA, Yusnawan E,Taufiq A, Sholihin, Suharsono, Sundari T, &Hermanto (Eds.). Prosiding Seminar HasilPenelitian Tanaman Aneka Kacang dan Umbi.Peningkatan Daya Saing dan ImplementasiPengembangan Komoditas Kacang dan UmbiMendukung Pencapaian Empat SuksesPembangunan Pertanian. pp. 29–36. PusatPenelitian dan Pengembangan Tanaman Pangan,Malang. 5 Juli 2012.
Arifin M & Koswanudin D. 2010. Alternatif teknologipengendalian ulat grayak pada kedelai denganberbagai jenis insektisida biorasional. Dalam:Kardinan A, Laba IW, Kartohardjono A, &Harnoto (Eds.). Prosiding Seminar Nasional VIPEI. Peranan Entomologi dalam MendukungPengembangan Pertanian Ramah Lingkungandan Kesehatan Masyarakat. pp. 419–434.Bogor. 24 Juni 2010.
Blum A. 1982. Evidence for genetic variability in droughtresistance and its applications in plant breeding.In: IRRI. Drought Resistance in Crops withEmphasis on Rice. pp. 56–68. Publisher IRRI,Los Banos.
Harborne JB. 1987. Metode Fitokimia (PenuntunCara Modern Menganalisis Tumbuhan). 2.ITB. Bandung. pp. 102–109.
Hartati S. 2009. Biologi Spodoptera litura F.(Lepidoptera: Noctuidae) pada Tanaman Kedelaidengan Dosis Pupuk Nitrogen yang Berbeda.http://uripsantoso.wordpress.com/2009/07/24/biologi-spodoptera-litura-f-lepidoptera-noctuidae-pada-tanaman-kedelai-dengan-dosis-pupuk-nitrogen-yang-berbeda/. Diakses 6 Desember2011.
Hendrival, Latifah, & Hayu R. 2013. PerkembanganSpodoptera litura F. (Lepidoptera: Noctuidae)pada kedelai. J. Floratek 8: 88–100.
Heriyanto. 2012. Upaya percepatan respon petani dalampeningkatan kontribusi varietas unggul kedelaiterhadap pendapatan daerah Jawa Timur. J.Cakrawala 6(2): 114–128.
Igita K, Adie MM, Suharsono, & Tridjaka. 1996. Secondbrief report: Method of cultivation of soybeanin cropping systems with low input (pesticide)in Indonesia. RILET-JIRCAS, Malang.
Kesumawaty R. 2003. Hubungan morfologi daunbeberapa genotipe kedelai dengan ketahanan ulatgrayak. Tesis. Universitas Brawijaya, Malang.
Komatsu K, Okuda S, Takashi M, & Madsunaga R.2004. Antibiotic effect and insect-resistantsoybean on common armyworm (Spodopteralitura) and its inheritance. Breed. Sci. 54: 27–32.
Leatemia JA & Rumthe RY. 2011. Studi kerusakan akibatserangan hama pada tanaman pangan diKecamatan Bula, Kabupaten Seram BagianTimur, Propinsi Maluku. J. Agroforestri 6(1): 53–56.
Luhukay JN, Uluputty MR, & Rumthe RY. 2013.Respons lima varietas kubis (Brassica oleraceal.) terhadap serangan hama pemakan daunPlutella xylostella (Lepidoptera: Plutellidae).Agrologia: J. Ilmu Budidaya Tan. 2(2): 164–169.
Manitto P. 1991. Biosintesa Produk Alami .Diterjemahkan oleh Sammes PG. John Wiley &Sons, New York.
Marwoto & Suharsono. 2008. Strategi dan komponenteknologi pengendalian ulat grayak (Spodopteralitura Fabricius) pada tanaman kedelai. J.Litbang Pertanian 27(4): 131–136.
Norris DM & Kogan M. 1980. Biochemical andmorphological bases of resistance. In: MaxwellFG & Jennings PR (Eds.). Breeding plantsResistant to Insects. pp. 23–60. John Wiley andSons, New York.

Susanto & Adie Identifikasi Fenotipik Galur-Galur Kedelai 187
Ravishankar BS & Venkatesha MG. 2010.Effectiveness of slnpv of Spodoptera litura(Fab.) (Lepidoptera: Noctuidae) on different hostplants. J. Biopestisida 3: 168–171.
Rusdy A. 2009. Efektivitas ekstrak daun mimba dalampengendalian ulat grayak (Spodoptera litura F.)pada tanaman selada. J. Floratek 4: 41–54.
Salisbury FB & Ross CW. 1992. Plant Physiology.4th, Wadsworth Publishing Co., Adivision ofWadsworth, Inc.
Sembiring N, Tarigan MN, & Lisnawita. 2013. Tingkatserangan ulat kantong Metisa plana Walker(Lepidoptera: Psychidae) terhadap umur tanamankelapa sawit (Elaeis guineensis Jacq.) di kebunMatapao PT. Socfin Indonesia. J.Agroekoteknologi 1(4): 135-143
Singh DP. 1986. Breeding for resistance to diseaseand insect pests. Crop Protection MonographsSpringer-Verlag, Berlin.
Smith WC, McCarty Jr JC, Altamarino TP, Lege KE,Schuster MF, Phillips JR, & Lopez JD.1992.Condensed tannins in cotton and bollwormbudworm (Lepidoptera: Noctuidae) resistance. J.Econ. Entomol. 85(6): 2211–2217.
Suharsono. 2011. Kepekaan galur kedelai toleran jenuhair terhadap ulat grayak Spodoptera litura F.Suara Perlindungan Tanaman 1(3): 13–22.
Suharsono & Adie MM. 2010. Identifikasi sumberketahanan aksesi plasma nutfah kedelai untuk ulatg r a y a k Spodoptera litura F. Buletin PlasmaNutfah 16(1): 29–37.
Suharsono & Suntono. 2007. Efektivitas beberapa jenisinsektisida kimia dan galur tahan untukmengendalikan hama perusak daun. HasilPenelitian Tahun 2015. Balitkabi, Malang.
Suharsono, Rahayu M, Hardaningsih S, Tengkano W,Indiati SW, Marwoto, Bedjo, & Baliadi Y. 2007.Perbaikan dan evaluasi komponen teknologipengendalian hama dan penyakit terpadu (PHT)pada tanaman kedelai. Laporan Akhir HasilPenelitian Tahun 2007. Pusat Penelitian danPengembangan Tanaman Pangan, Bogor.
Suharsono, Rahayu M, Hardaningsih S, Tengkano W,Indiati SW, Marwoto, Bedjo, & Baliadi Y. 2008.Perbaikan dan evaluasi komponen teknologipengendalian hama dan penyakit terpadu (PHT)pada tanaman kedelai. Laporan Akhir HasilPenelitian Tahun 2008. Pusat Penelitian danPengembangan Tanaman Pangan, Bogor
Wassimi NN, Hosfield GL, & Uebersax MA. 1988.Combining ability of tanin content and proteincharacteristics of row and cooked dry beans.Crop Sci. 28(3): 452–458.

188 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198J. HPT Tropika. ISSN 1411-7525Vol. 15, No. 2: 188 – 199, September 2015
IDENTIFIKASI MOLEKULER BROAD BEAN WILT VIRUS 2 (BBWV2)DAN CYMBIDIUM MOSAIC VIRUS (CYMMV)
ASAL TANAMAN NILAM (POGOSTEMON CABLIN BENTH.)
Miftakhurohmah1, Gede Suastika2, Tri Asmira Damayanti2, & Rita Noveriza1
1Balai Penelitian Tanaman Rempah dan ObatJl. Tentara Pelajar No. 3, Bogor 16111
2Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian BogorJl. Kamper, Kampus IPB Dramaga, Bogor 16680
E-mail: [email protected]
ABSTRACT
Molecular identification Broad Bean Wilt Virus 2 (BBWV2) and Cymbidium Mosaic Virus (CymMV) from patchouliplant (Pogostemon cablin Benth.). Several viruses have been reported to be associated with mosaic disease on patchouliplant in Indonesia. This study aims to identify the two viruses in patchouli cultivation in West Java by studying the molecularcharacterization. Mosaic symptomatic leaf samples taken from patchouli cultivation in Manoko (Bandung Barat District, WestJava Province). RNA extraction was performed using Xprep Plant RNA mini kit. RNA amplification with RT-PCR techniqueusing primers for the cp gene region of BBWV2 and CymMV. The PCR product was sent to PT. Science Genetics Indonesia todo sequencing, then analyzed nucleotide sequences. Results of RT-PCR were performed successfully obtained DNA bandswith size accordance with the predictions of the primer design for BBWV2 and CymMV cp region. Further, based on nucleotideand amino acid sequence analyses, the two virus isolates were confirmed as BBWV2 and CymMV respectively. Phylogeneticanalyses revealed that BBWV2 Manoko clustered with BBWV2 from Singapore (original host of Brazilian red-cloak), China(pepper) and South Korea (chili). Whereas, CymMV Manoko become one cluster with CymMV from India (Phaius sp.),Indonesia (Dendrobium), China (vanilla), Thailand (Oncidium), Hawai (Dendrobium) and South Korea (Cymbidium).
Key words: nucleotide sequence, virus, patchouli, BBWV2, CymMV
ABSTRAK
Identifikasi molekuler Broad Bean Wilt Virus 2 (BBWV2) dan Cymbidium Mosaic Virus (CymMV) asal tanaman nilam(Pogostemon cablin Benth.). Beberapa virus dilaporkan berasosiasi dengan penyakit mosaik pada tanaman nilam di Indonesia.Penelitian ini bertujuan mengidentifikasi dua virus pada pertanaman nilam di Jawa Barat dengan mempelajari karakterisasimolekulnya. Sampel daun bergejala mosaik diambil dari pertanaman nilam di Manoko (Kabupaten Bandung Barat, ProvinsiJawa Barat). Ekstraksi RNA dilakukan menggunakan Xprep Plant RNA mini kit. Amplifikasi RNA dengan teknik RT-PCRmenggunakan primer untuk daerah gen cp BBWV2 dan CymMV. Produk PCR yang dihasilkan dikirimkan ke PT. GenetikaScience Indonesia untuk dilakukan perunutan nukleotida, dan selanjutnya dianalisis runutan nukleotidanya. Hasil RT-PCRyang dilakukan berhasil didapatkan pita DNA dengan ukuran sesuai prediksi dari desain primer untuk bagian cp BBWV2 danCymMV. Berdasarkan analisis runutan nukleotida dan asam amino, dua virus yang ditemukan merupakan BBWV2 dan CymMV.Analisis homologi dan filogenetik menunjukkan bahwa BBWV2 Manoko mengelompok dengan BBWV2 dari Singapura(Brazilian red-cloak), Cina (lada) dan Korea Selatan (cabai). Sedangkan CymMV Manoko berada dalam satu kelompokdengan CymMV dari India (Phaius sp.), Indonesia (Dendrobium), China (vanili), Thailand (Oncidium), Hawai (Dendrobium)dan Korea Selatan (Cymbidium).
Kata kunci: sekuen nukleotida, virus, nilam, BBWV2, CymMV
PENDAHULUAN
Infeksi virus pada tanaman nilam di Indonesiatelah ditemukan sejak tahun 1996 oleh Sumardiyono etal. (1996), yang berdasarkan uji serologi dengan antibodipatchouli mottle virus (PatMoV), menunjukkan reaksi
positif. Pada pertanaman nilam di Cianjur dan Bogor,ditemukan infeksi campuran antara cucumber mosaicvirus (CMV) dan genus Potyvirus (Sukamto et al.,2007). Gejala mosaik ditemukan pada beberapapertanaman nilam di Jawa Tengah, Jawa Barat,Sumatera Utara dan Sumatera Barat. Berdasarkan hasil

Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 189
deteksi dan identifikasi, penyakit berasosiasi dengantelosma mosaic virus (TeMV) yang tergolong ke dalamgenus Potyvirus (Noveriza et al., 2012).
Selain genus Potyvirus dan CMV, juga ditemukaninfeksi virus lain pada pertanaman nilam di Manoko(Kabupaten Bandung Barat, Provinsi Jawa Barat) yangberdasarkan deteksi secara serologi merupakanBBWV2 dan CymMV. Kedua virus tersebut merupakanvirus pada tanaman nilam yang tergolong patogen A1,dalam arti belum pernah ada laporan serangan virus inipada tanaman di Indonesia (Badan Karantina Pertanian,2011). Berdasarkan pengamatan, saat ini sebagian besarbibit nilam hasil perbanyakan vegetatif sudah terserangvirus dengan berbagai gejala sebelum ditanam dilapangan.
Berdasarkan hal tersebut diatas, perlu dilakukanidentifikasi lanjut BBWV2 dan CymMV yangmenginfeksi pertanaman nilam. Deteksi secara serologimemiliki kelemahan, yaitu tidak bisa membedakan virusyang sekerabat, terutama bila yang digunakan antiserumuniversal genus. Selain itu, beberapa antigenic sitedimiliki oleh virus yang tidak sekerabat, sehinggamemungkinkan terjadinya reaksi silang (Putnam, 1995).Oleh karena itu, hasil deteksi secara serologi perludilanjutkan dengan identifikasi berdasarkan asam nukleat.Hasil identifikasi digunakan sebagai salah satu dasardalam pengambilan tindakan pengendalian.
Identifikasi berdasarkan asam nukleat dilakukansecara molekuler, dengan teknik Polymerase chainreaction (PCR). Untuk diagnosis virus tanaman yangmemiliki asam nukleat RNA, RNA target dikonversikanmenjadi complementary DNA (cDNA) yang dikopidengan transkripsi balik sebelum PCR dimulai (Naidu& Hughes, 2003).
Dari hasil RT-PCR, selanjutnya dilakukanperunutan nukleotidanya, yang dapat digunakan untukmengklasifikasikan virus berdasarkan genomnya.Penelitian terdahulu menunjukkan bahwa analisisrunutan nukleotida coat protein (cp) sangat bergunadalam kegiatan identifikasi BBWV1 dan BBWV2(Kobayashi et al., 2005; Kondo et al., 2005). Beberapasoftware telah tersedia untuk mengklasifikasikan virusberdasarkan genomnya, diantaranya BioEdit, MEGA,dan GeneDoc (Tamura et al., 2007; Hall, 1999).
Penelitian bertujuan mengidentifikasi BBWV2dan CymMV pada tanaman nilam di Jawa Barat denganmempelajari karakterisasi molekulnya.
METODE PENELITIAN
Tempat dan Waktu. Penelitian dilaksanakan diLaboratorium Virologi Tumbuhan, Fakultas Pertanian,
Institut Pertanian Bogor, dari bulan Februari sampaidengan Agustus 2012.
Sumber Virus. Virus diambil dari daun bergejala mosaikpada pertanaman nilam di Kebun Percobaan (KP)Manoko (Kabupaten Bandung Barat). Gejala mosaikyang ditemukan bervariasi, dari mosaik lemah, mosaikkuning hijau, mosaik hijau muda hijau tua, mosaik denganpenebalan dan mosaik dengan perubahan bentuk daun(malformasi).
Reverse transcription polymerase chain reaction(RT-PCR)
Ekstraksi RNA. Ekstraksi RNA dilakukandengan Xprep Plant RNA mini kit (PKT-PhilekoreaTechnology). Bufer XPRB disiapkan denganmenambahkan 1% mercaptoethanol (ME). Sampeldaun nilam sebanyak 0,1 g digerus menggunakan nitrogencair pada mortar, sampai menjadi serbuk. Serbuk sampelditambah bufer XPRB yang telah ditambah ME,dimasukkan ke dalam kolom filter dan disentrifugasiselama dua menit pada kecepatan 13.000 rpm.Supernatan dipindahkan ke tabung eppendorf baru danetanol absolut ditambahkan sebanyak 0,5 kali volumesupernatan, dan dicampur dengan cara dipipet ataudibolak-balik. Kemudian supernatan dalam alkoholdimasukkan ke dalam kolom XPPLR mini, dandisentrifugasi selama satu menit pada kecepatan 13.000rpm. Setelah supernatan dibuang, kolom diberi washbuffer 1 (WB1) sebanyak 500 µl, disentrifugasi selamasatu menit, dan supernatan dibuang. Wash buffer 2(WB2) sebanyak 750 µl ditambahkan ke dalam kolom,disentrifugasi selama satu menit, dan supernatandibuang. Selanjutnya, untuk mengeringkan kolom,dilakukan sentrifugasi kolom selama tiga menit padakecepatan 13.000 rpm. RNA total dikoleksi dengan caramemberikan air bebas nuklease sebanyak 50 µl ke pusatmembran kolom XPPLR yang diletakkan pada tabungeppendorf baru, dibiarkan selama satu menit, laludisentrifugasi selama satu menit. RNA yang telahdiperoleh disimpan di freezer -80 oC, sampai digunakan.
Sintesis complementary DNA (c-DNA ). Reaksireverse transcription (RT) dilakukan untuk membuatcDNA dengan menggunakan enzim reversetranscriptase. Pereaksi yang digunakan terdiri atas airbebas nuklease (3,7 µl), bufer RT 5x (2 µl), DTT 50mM (0,35 µl), dNTP 10 mM (0,5 µl), M-MuLV Rev(Fermentas) (0,35 µl), RNase Inhibitor (0,35 µl), Oligod(T) 10 µM (0,75 µl) dan RNA templat (2 µl) dengantotal volume sebanyak 10 µl. Setiap komponen reaksidipipet satu persatu, dimasukkan ke dalam tabung

190 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198
eppendorf berukuran 0,5 µl, kemudian dimasukkan kedalam mesin PCR. Reaksi RT dilakukan selama satujam pada suhu 42 oC.
Amplifikasi cDNA. Untuk amplifikasi DNA,digunakan sepasang primer degenerate untuk BBWV(Kondo et al., 2005) dan satu pasang primer spesifikCymMV (Lee & Chang, 2008) dengan urutannukletotida yang dapat dilihat pada Tabel 1. cDNA yangdihasilkan dari hasil RT, diamplifikasi dengan teknik PCRpada volume 25 µl yang terdiri atas: air bebas nuklease9,5 µl, PCR mix (Go Green Taq-Promega) 12,5 µl, primerforward 10 µM dan reverse 10 µM, masing–masing 1µl dan DNA templat 1 µl. Setiap komponen reaksi dipipetsatu persatu, dimasukkan ke dalam tabung eppendorfberukuran 0,5 µl, kemudian dimasukkan dalam mesinPCR. Program PCR diatur berbeda, tergantungpasangan primer yang digunakan (Tabel 2).
Visualisasi DNA. DNA hasil amplifikasi dariPCR diseparasi pada gel agarose 1,5% yang telahditambah etidium bromida (0,5 µl/10 ml TBE). Sampeldimasukkan ke dalam sumuran gel agarose, kemudiandielektroforesis pada 50 V selama 60 menit. Adanyapita DNA pada gel agarose dilihat di bawahtransilluminator ultraviolet dan didokumentasi dengankamera digital.
Analisis Runutan Nukleotida cp BBWV2, CymMV,dan Asam Amino CP BBWV2 dan CymMV
Perunutan Nukleotida. Perunutan nukleotidamenggunakan mesin sequencer ABI-Prism 3100-AvantGenetic Analyzer di Laboratorium Research andDevelopment Centre PT. Genetika Science. Indonesia.
Hasil runutan dianalisis menggunakan software Blast(www. NCBI. Nml. Niv.gov) dan software Wu-Blast(www.ebi.ac.uk). Contiq hasil perunutan DNAdilakukan dengan bantuan program Sequenche r4.8 dansoftware Complementor (www.justbio.com).Selanjutnya, hasil runutan yang sudah diolah, ditranslasimenjadi urutan protein (asam amino), dengan bantuansoftware Translate (www.expasy.org.tools).
Analisis Identitas Matriks dan Filogenetika.Runutan nukleotida yang diperoleh dibandingkan dengandata runutan nukleotida yang ada di GenBank. Matriksidentitas nukleotida dan asam amino diperoleh denganmenggunakan software BioEdit versi 7.0. Selanjutnya,gambar pensejajaran runutan nukleotida didapatkandengan menggunakan program GeneDoc versi 2.7.000.Pohon filogenetika dikonstruksi dengan menggunakansoftware MEGA 4.0 (Molecular EvolutionaryGenetic Analysis software) (Tamura et al., 2007),dengan metode neighbour-joining menggunakanbootstrap 1000 kali ulangan.
HASIL DAN PEMBAHASAN
Broad Bean Wilt Virus 2RT-PCR. Primer yang digunakan yaitu BBWV25
dan BBWV3487M merupakan primer degenerate untukmengamplifikasi gen coat protein (cp) BBWV baikBBWV1 maupun BBWV2, sehingga produk yangdihasilkan merupakan gen cp BBWV. Pasangan primerini telah berhasil digunakan untuk mengamplifikasi gencp BBWV2 dari tanaman Chinese yam (Dioscoreaopposita Thunb.) di Ojima, Jepang (Kondo et al., 2005).
Tabel 1. Primer-primer yang digunakan untuk deteksi virus mosaik nilam
No Primer Urutan basa Ukuran DNA Referensi
1 BBWV25 5’-AATGARRTKGTNCTCAAYTA-3’2000 pb Kondo et al. ( 2005)
2 BBW3487M 5’-AMAMAGGTCATGGAACCCA-3’.3 CyCP-F1 5-ATGGGAGAGYCCACTCCARCYCCAGC-3'
679 pb Lee & Chang (2008)4 CyCP-R1 5'-ATCGCTCGAGTTCAGTAGGGGGTGCAGGCA-3'
Tabel 2. Program PCR untuk setiap primer yang digunakan untuk kegiatan PCR
No Primer Program PCR Referensi
1 BBWV25 Denaturasi awal pada suhu 95 oC ( 5 menit); 35 siklus: 99 oC (1 menit),55 oC (2 menit) dan 72 oC (3 menit); diakhiri dengan 72 oC (5 menit)
Kondo et al. (2005)2 BBW3487M3 CyCP-F1 Denaturasi awal pada suhu 96 oC (5 menit); 30 siklus: 96 oC (30 detik),
52 oC (30 detik) dan 72 oC (30 detik); diakhiri dengan 72 oC (7 menit) Lee & Chang (2008)4 CyCP-R1
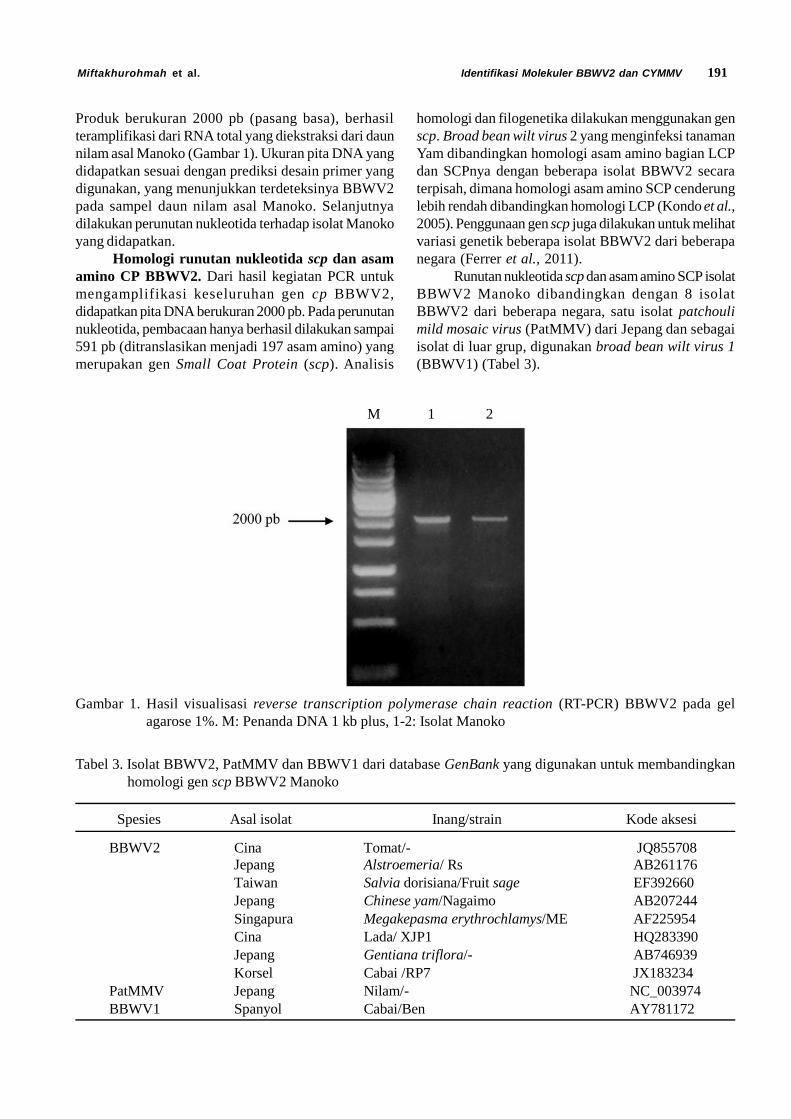
Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 191
Produk berukuran 2000 pb (pasang basa), berhasilteramplifikasi dari RNA total yang diekstraksi dari daunnilam asal Manoko (Gambar 1). Ukuran pita DNA yangdidapatkan sesuai dengan prediksi desain primer yangdigunakan, yang menunjukkan terdeteksinya BBWV2pada sampel daun nilam asal Manoko. Selanjutnyadilakukan perunutan nukleotida terhadap isolat Manokoyang didapatkan.
Homologi runutan nukleotida scp dan asamamino CP BBWV2. Dari hasil kegiatan PCR untukmengamplifikasi keseluruhan gen cp BBWV2,didapatkan pita DNA berukuran 2000 pb. Pada perunutannukleotida, pembacaan hanya berhasil dilakukan sampai591 pb (ditranslasikan menjadi 197 asam amino) yangmerupakan gen Small Coat Protein (scp). Analisis
homologi dan filogenetika dilakukan menggunakan genscp. Broad bean wilt virus 2 yang menginfeksi tanamanYam dibandingkan homologi asam amino bagian LCPdan SCPnya dengan beberapa isolat BBWV2 secaraterpisah, dimana homologi asam amino SCP cenderunglebih rendah dibandingkan homologi LCP (Kondo et al.,2005). Penggunaan gen scp juga dilakukan untuk melihatvariasi genetik beberapa isolat BBWV2 dari beberapanegara (Ferrer et al., 2011).
Runutan nukleotida scp dan asam amino SCP isolatBBWV2 Manoko dibandingkan dengan 8 isolatBBWV2 dari beberapa negara, satu isolat patchoulimild mosaic virus (PatMMV) dari Jepang dan sebagaiisolat di luar grup, digunakan broad bean wilt virus 1(BBWV1) (Tabel 3).
Tabel 3. Isolat BBWV2, PatMMV dan BBWV1 dari database GenBank yang digunakan untuk membandingkanhomologi gen scp BBWV2 Manoko
Spesies Asal isolat Inang/strain Kode aksesi
BBWV2 Cina Tomat/- JQ855708Jepang Alstroemeria/ Rs AB261176Taiwan Salvia dorisiana/Fruit sage EF392660Jepang Chinese yam/Nagaimo AB207244Singapura Megakepasma erythrochlamys/ME AF225954Cina Lada/ XJP1 HQ283390Jepang Gentiana triflora/- AB746939Korsel Cabai /RP7 JX183234
PatMMV Jepang Nilam/- NC_003974BBWV1 Spanyol Cabai/Ben AY781172
Gambar 1. Hasil visualisasi reverse transcription polymerase chain reaction (RT-PCR) BBWV2 pada gelagarose 1%. M: Penanda DNA 1 kb plus, 1-2: Isolat Manoko
M 1 2
2000 pb

192 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198
Tabel 4. Homologi runutan nukleotida dan asam amino gen scp BBWV2 Manoko dengan beberapa anggota genusFabavirus
Hasil analisis homologi runutan nukleotida danasam amino menunjukkan bahwa gen scp isolat asalManoko menunjukkan homologi tertinggi sebesar 93,0%dan 93,9% dengan BBWV2 asal Singapura. Sedangkandengan isolat BBWV2 lain dan PatMMV, homologiberdasarkan runutan nukleotida berkisar antara 78,8–90,0%, dan berdasarkan asam amino berkisar antara88,8–93,4%. Homologi isolat asal Manoko denganBBWV1, hanya sebesar 63,6% berdasarkan runutannukleotida, dan sebesar 58,8% berdasarkan asam amino(Tabel 4). Hasil ini menunjukkan bahwa virus yangditemukan merupakan salah satu spesies BBWV2.Dalam genus Fabavirus, virus diklasifikasikan ke dalamspesies yang sama bila homologi gen cp lebih dari 75%(Fauquet et al., 2005).
Genus Fabavirus nilam di Jepang diidentifikasisebagai PatMMV. PatMMV memiliki kemiripanmorfologi partikel, berat molekul CP dan hubunganserologi dengan isolat BBWV dan lamium mild mosaicvirus (LMMV). Namun, PatMMV menunjukkanperbedaan kisaran inang dan gejalanya pada Vicia fabadan Nicotiana tabacum, sehingga digolongkan sebagaispesies baru dalam genus Fabavirus (Natsuaki et al.,1994). Genus Fabavirus yang menginfeksi pertanamannilam di Indonesia, menunjukkan homologi nukleotidadan asam amino lebih tinggi terhadap BBWV2 asalSingapura, dibandingkan dengan PatMMV. Dengandemikian, genus Fabavirus yang menginfeksi tanamannilam merupakan BBWV2. Selain itu, PatMMVakhirnya juga diklasifikasikan sebagai salah satu strainBBWV2, karena perbedaan gen cp-nya hanya berkisarantara 3–21% dengan BBWV2 (Fauquet et al., 2005).
BBWV2 memiliki kisaran inang yang luas,menginfeksi tanaman holtikultura dan hias (Ferrer et al.,
2011), namun belum pernah dilaporkan menginfeksinilam. Secara serologi BBWV2 terdeteksi menginfeksinilam (Noveriza et al., 2012). Penelitian ini berhasilmengidentifikasi BBWV2 secara molekuler melaluiteknik RT-PCR menggunakan primer dengan target gencp serta analisis runutan nukleotida dan asam aminonya.
Pohon Filogenetika Runutan Nukleotida danAsam Amino Gen scp BBWV2. Hasil analisisfilogenetika berdasarkan runutan nukleotida gen scpmenunjukkan terbentuknya dua kelompok/cluster.BBWV2 Manoko berada dalam satu kelompok denganBBWV2 Singapura (yang menginfeksi Megakepasmaerythroclamys/ME), Cina (lada) dan Korea Selatan(cabai). Isolat PatMMV (nilam) berada pada kelompoklain dengan BBWV2 Taiwan (S. dorisiana), Jepang(Gentiana, yam, Alstroemeria) dan Cina (tomat)(Gambar 2). Hasil analisa filogenetik ini mendukunghasil analisa homologi runutan nukleotida dan asamamino, dimana BBWV2 Manoko memiliki homologitertinggi dengan BBWV2 Singapura, dan pada pohonfilogeni berada dalam satu kelompok.
Hasil analisis filogenetika menunjukkan bahwapengelompokan isolat-isolat BBWV2 tidak berdasarkanasal isolat. Hal ini terlihat pada kedua isolat Cina, beradapada kelompok yang berbeda. Hasil yang sama jugaterjadi pada analisis filogenetika isolat-isolat BBWV2dari beberapa negara dan dari inang yang berbedaberdasarkan gen scp, terlihat bahwa pengelompokkantidak berdasarkan inang dan asal isolat (Ferrer et al.,2011).
Cymbidium Mosaic VirusRT-PCR. Primer yang digunakan untuk
mengamplifikasi gen cp CymMV adalah primer
Spesies Asal isolat Inang/strainHomologi (%)
Nukleotida Asam amino
BBWV2 Cina Tomat/- 80,5 91,3
Jepang Alatroemeria/Rs 78,8 88,8Taiwan Salvia dorisiana/Fruit Sage 80,0 88,0Singapura Megakepasma erythroclamys/ME 93,0 94,9Cina Lada/XJP1 90,0 93,4Jepang Gentiana triflora/- 80,3 89,3Korsel Cabai/RP7 88,1 92,8Jepang Chinese yam/Nagaimo 80,7 86,8
PatMMV Jepang Nilam/- 79,6 89,8BBWV1 Spanyol Cabai/Ben 63,6 58,8
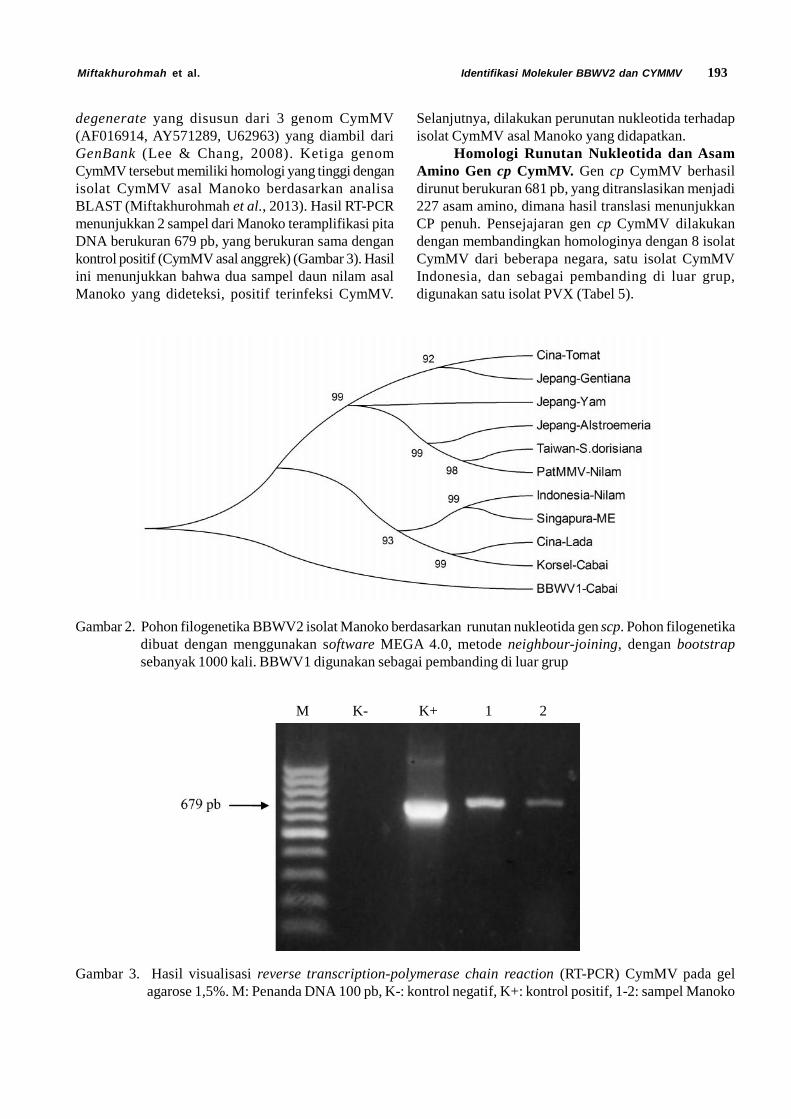
Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 193
Cina-Tomat
Jepang-Gentiana
Jepang-Yam
Jepang-Alstroemeria
Taiwan-S.dorisiana
PatMMV-Nilam
Indonesia-Nilam
Singapura-ME
Cina-Lada
Korsel-Cabai
BBWV1-Cabai
99
99
93
9899
92
99
Gambar 2. Pohon filogenetika BBWV2 isolat Manoko berdasarkan runutan nukleotida gen scp. Pohon filogenetikadibuat dengan menggunakan software MEGA 4.0, metode neighbour-joining, dengan bootstrapsebanyak 1000 kali. BBWV1 digunakan sebagai pembanding di luar grup
Gambar 3. Hasil visualisasi reverse transcription-polymerase chain reaction (RT-PCR) CymMV pada gelagarose 1,5%. M: Penanda DNA 100 pb, K-: kontrol negatif, K+: kontrol positif, 1-2: sampel Manoko
679 pb
M K- K+ 1 2
degenerate yang disusun dari 3 genom CymMV(AF016914, AY571289, U62963) yang diambil dariGenBank (Lee & Chang, 2008). Ketiga genomCymMV tersebut memiliki homologi yang tinggi denganisolat CymMV asal Manoko berdasarkan analisaBLAST (Miftakhurohmah et al., 2013). Hasil RT-PCRmenunjukkan 2 sampel dari Manoko teramplifikasi pitaDNA berukuran 679 pb, yang berukuran sama dengankontrol positif (CymMV asal anggrek) (Gambar 3). Hasilini menunjukkan bahwa dua sampel daun nilam asalManoko yang dideteksi, positif terinfeksi CymMV.
Selanjutnya, dilakukan perunutan nukleotida terhadapisolat CymMV asal Manoko yang didapatkan.
Homologi Runutan Nukleotida dan AsamAmino Gen cp CymMV. Gen cp CymMV berhasildirunut berukuran 681 pb, yang ditranslasikan menjadi227 asam amino, dimana hasil translasi menunjukkanCP penuh. Pensejajaran gen cp CymMV dilakukandengan membandingkan homologinya dengan 8 isolatCymMV dari beberapa negara, satu isolat CymMVIndonesia, dan sebagai pembanding di luar grup,digunakan satu isolat PVX (Tabel 5).
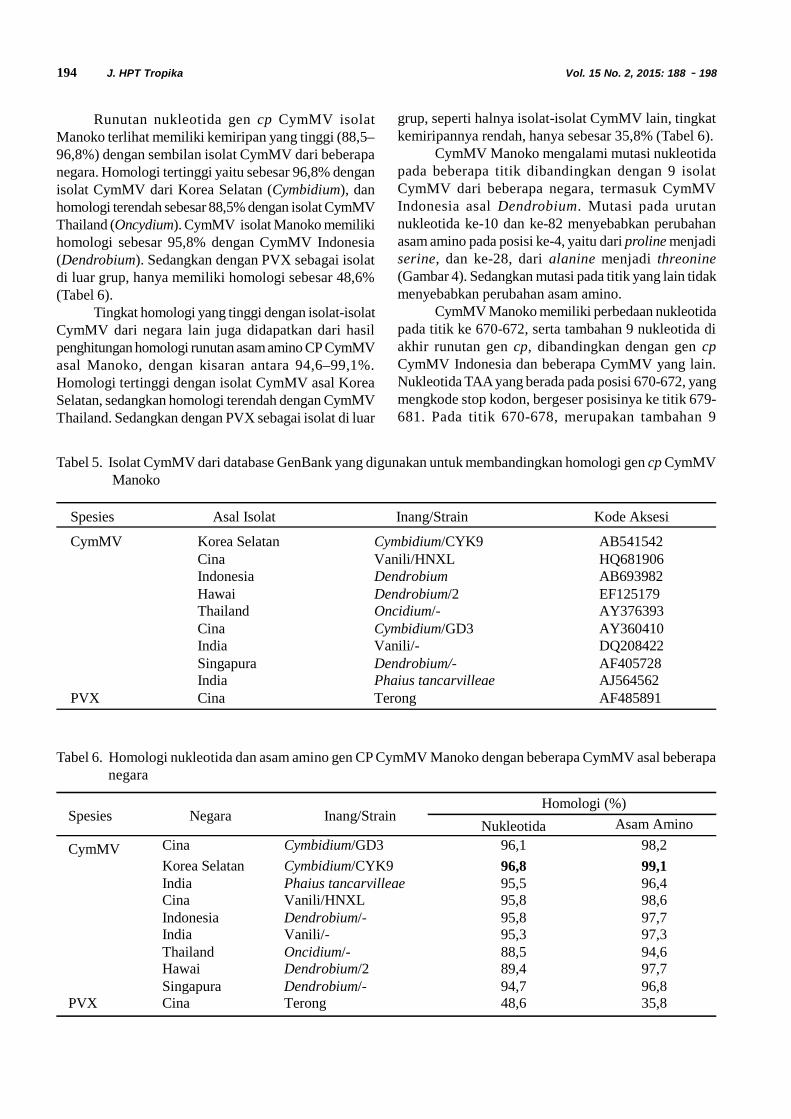
194 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198
Tabel 6. Homologi nukleotida dan asam amino gen CP CymMV Manoko dengan beberapa CymMV asal beberapanegara
Spesies Negara Inang/StrainHomologi (%)
Nukleotida Asam Amino
CymMV Cina Cymbidium/GD3 96,1 98,2
Korea Selatan Cymbidium/CYK9 96,8 99,1India Phaius tancarvilleae 95,5 96,4Cina Vanili/HNXL 95,8 98,6Indonesia Dendrobium/- 95,8 97,7India Vanili/- 95,3 97,3Thailand Oncidium/- 88,5 94,6Hawai Dendrobium/2 89,4 97,7Singapura Dendrobium/- 94,7 96,8
PVX Cina Terong 48,6 35,8
Runutan nukleotida gen cp CymMV isolatManoko terlihat memiliki kemiripan yang tinggi (88,5–96,8%) dengan sembilan isolat CymMV dari beberapanegara. Homologi tertinggi yaitu sebesar 96,8% denganisolat CymMV dari Korea Selatan (Cymbidium), danhomologi terendah sebesar 88,5% dengan isolat CymMVThailand (Oncydium). CymMV isolat Manoko memilikihomologi sebesar 95,8% dengan CymMV Indonesia(Dendrobium). Sedangkan dengan PVX sebagai isolatdi luar grup, hanya memiliki homologi sebesar 48,6%(Tabel 6).
Tingkat homologi yang tinggi dengan isolat-isolatCymMV dari negara lain juga didapatkan dari hasilpenghitungan homologi runutan asam amino CP CymMVasal Manoko, dengan kisaran antara 94,6–99,1%.Homologi tertinggi dengan isolat CymMV asal KoreaSelatan, sedangkan homologi terendah dengan CymMVThailand. Sedangkan dengan PVX sebagai isolat di luar
grup, seperti halnya isolat-isolat CymMV lain, tingkatkemiripannya rendah, hanya sebesar 35,8% (Tabel 6).
CymMV Manoko mengalami mutasi nukleotidapada beberapa titik dibandingkan dengan 9 isolatCymMV dari beberapa negara, termasuk CymMVIndonesia asal Dendrobium. Mutasi pada urutannukleotida ke-10 dan ke-82 menyebabkan perubahanasam amino pada posisi ke-4, yaitu dari proline menjadiserine, dan ke-28, dari alanine menjadi threonine(Gambar 4). Sedangkan mutasi pada titik yang lain tidakmenyebabkan perubahan asam amino.
CymMV Manoko memiliki perbedaan nukleotidapada titik ke 670-672, serta tambahan 9 nukleotida diakhir runutan gen cp, dibandingkan dengan gen cpCymMV Indonesia dan beberapa CymMV yang lain.Nukleotida TAA yang berada pada posisi 670-672, yangmengkode stop kodon, bergeser posisinya ke titik 679-681. Pada titik 670-678, merupakan tambahan 9
Tabel 5. Isolat CymMV dari database GenBank yang digunakan untuk membandingkan homologi gen cp CymMVManoko
Spesies Asal Isolat Inang/Strain Kode Aksesi
CymMV Korea Selatan Cymbidium/CYK9 AB541542Cina Vanili/HNXL HQ681906Indonesia Dendrobium AB693982Hawai Dendrobium/2 EF125179Thailand Oncidium/- AY376393Cina Cymbidium/GD3 AY360410India Vanili/- DQ208422Singapura Dendrobium/- AF405728India Phaius tancarvilleae AJ564562
PVX Cina Terong AF485891

Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 195
nukleotida, yang ditranslasikan menjadi asam aminoleucine, glutamic acid dan arginine (Gambar 5).
Perubahan dan tambahan beberapa asam aminoCP CymMV Manoko ini diduga mempengaruhiperbedaan inang CymMV Manoko dan isolat CymMVlain. CymMV dilaporkan hanya menginfeksi tanamandalam famili Orchidaceae, sedangkan CymMV Manokomenginfeksi tanaman nilam yang tergolong ke dalamfamili Lamiaceae. Hal ini terjadi karena gen cp genusPotexvirus selain berperan sebagai pembentuk selubungprotein, juga berperan dalam pergerakan virus antar sel(sebagai tambahan), yang mempengaruhi penyebaranvirus (Scholthof, 2005). Gen cp CymMV M1 berperanpenting dalam perpindahan sel ke sel. Perbedaan empatasam amino pada CP dua strain CymMV asal anggrek
menyebabkan CymMV M1 mampu berpindah keseluruh bagian tanaman tembakau (Nicotiana tabacum)yang menyebabkan gejala sistemik, sedangkan CymMVM2 hanya menyebabkan gejala lokal (Lu et al., 2009).
Genus Potexvirus pada nilam di Brazildiidentifikasi sebagai virus baru, yang dinamakan denganPatVX (Filho et al., 2002). Namun demikian, penelitidari Brazil tersebut hanya melakukan pembuatanantiserum, uji serologi baik dengan antiserum dan denganuji protein, tanpa melakukan sekuensing. Dengandemikian, runutan nukleotida dan asam amino isolatCymMV yang didapatkan dari nilam tidak bisadibandingkan homologinya dengan genom PatVX yangditemukan di Brazil.
Gambar 4. Hasil alignment asam amino antara genom CymMV isolat Manoko dengan nukleotida genom- genomCymMV yang didapatkan dari database GeneBank; keterangan: latar belakang warna hitammenunjukkan kesamaan runutan nukleotida antar isolat, sedangkan warna abu-abu menunjukkanketidaksamaan. Pensejajaran dilakukan dengan program Bioedit. dilanjutkan dengan program GeneDogVer. 2.7.000 (www.psc.edu/biomed/genedoc)

196 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198

Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 197
Gambar 5. Hasil alignment nukleotida antara genom CymMV isolat Manoko dengan nukleotida genom-genomCymMV yang didapatkan dari database GeneBank; keterangan: latar belakang warna hitammenunjukkan kesamaan runutan nukleotida antar isolat, sedangkan warna abu-abu menunjukkanketidaksamaan. Pensejajaran dilakukan dengan program Bioedit. dilanjutkan dengan program GeneDogVer. 2.7.000 (www.psc.edu/biomed/genedoc)
CymMV telah dilaporkan menginfeksi tanamananggrek di beberapa negara. Selain tanaman anggrek,CymMV juga dilaporkan menginfeksi tanaman vanili(Grisoni et al., 2004). Di Indonesia, CymMV telahdilaporkan menginfeksi tanaman anggrek (Lakani et al.,2010). Namun demikian, infeksi CymMV pada tanamannilam belum pernah dilaporkan. Dengan demikian,penelitian ini merupakan laporan pertama infeksiCymMV pada tanaman nilam.
Pohon Filogenetika Runutan Nukleotida cp danAsam Amino CP CymMV. Hasil analisis filogenetikarunutan nukleotida gen cp membentuk 2 kelompok/
cluster. Isolat Manoko berada dalam satu kelompokdengan CymMV Indonesia (Dendrobium), Cina(Cymbidium), Korea Selatan (Cymbidium), India(Phaius sp.), Thailand (Oncidium) dan Hawai(Dendrobium). Isolat Singapura (Dendrobium) danIndia (vanili) membentuk kelompok terpisah (Gambar6).Hasil analisa homologi asam amino dan nukleotida sertafilogenetik menunjukkan bahwa isolat Manoko yangditemukan merupakan CymMV. Virus diklasifikasikanke dalam spesies yang sama bila gennya memilikihomologi nukleotida lebih dari 72% atau asam aminolebih dari 80% (Fauquet et al., 2005).

198 J. HPT Tropika Vol. 15 No. 2, 2015: 188 - 198
Mnk(Ind)-nilam
Indonesia-Dendrobium
Cina-Cymbidium
Cina-Vanili
Korsel-Cymbidium
India-Phaius sp
Thailand-Oncidium
Hawai-Dendrobium
Singapura-Dendrobium
India-Vanili
PVX-Terong
99
9167
65
Gambar 6. Pohon filogenetika CymMV isolat Manoko (Lembang) berdasarkan runutan nukleotida gen cp. Pohonfilogenetika dibuat menggunakan software MEGA 4.0, metode neighbour-joining, dengan bootstrapsebanyak 1000 kali. PVX digunakan sebagai pembanding di luar grup
SIMPULAN
Virus yang berasosiasi dengan gejala mosaik padatanaman nilam adalah BBWV2 dan CymMV. Hasilanalisis homologi dan pohon filogenetika menunjukkanbahwa BBWV2 asal Manoko mengelompok denganBBWV2 dari Singapura asal tanaman hiasMegakepasma erythroclamys/ME), Cina asal tanamanlada dan Korea Selatan asal tanaman cabai. SedangkanCymMV asal Manoko berada dalam satu kelompokdengan CymMV dari Indonesia, Cina, Korea Selatan,India, Thailand, dan Hawai yang berasal dari beberapajenis anggrek. Isolat CymMV yang ditemukanmerupakan laporan pertama teridentifikasi pada nilamdi Indonesia.
SANWACANA
Penulis mengucapkan terima kasih kepada Dr.Irwan Lakani yang telah membantu kegiatan analisisrunutan nukleotida gen cp BBWV2 dan CymMV sertaasam amino CP BBWV2 dan CymMV.
DAFTAR PUSTAKA
Badan Karantina Pertanian. 2011. Basis DataOrganisme Pengganggu TumbuhanKarantina. Badan Karantina Pertanian.Departemen Pertanian Republik Indonesia.http://karantina.deptan.go.id/optk/index.php.Diakses tanggal 14 April 2011.
Fauquet CM, Mayo MA, Maniloff J, Desselberger U,& Ball LA. 2005. Virus Taxonomy.Classification and Nomenclature of Viruses.Elsevier Academic Press. New York.
Ferrer RM, Ferrior I, Moreno P, Guerri J, & Rubio L.2011. Genetic variation and evolutionary analysisof Broad bean wilt virus 2. Arch. Virol. 156(8):1445–1450.
Filho PEM, Resende RO, Lima MI, & Kitajima EW.2002. Pantchouli virus x, a new potexvirus fromPogostemon clabin. Annu. Appl. Biol. 141: 267–274.
Grisoni M, Davidson F, Hyrondelle C, Farreyrol K,Caruana ML, & Pearson M. 2004. Nature,incidence and symptomatology of viruses infectingVanilla tahitensis in French Polysenia. Plant Dis.88(2): 119–124.
Hall TA. 1999. BioEdit: a user-friendly biologicalsequence alignment editor and analysis programfor Windows 95/98/NT. Nucleic Acid Symp. Ser.41: 95–98.
Kondo T, Fuji S, Yamashita K, Kang DK, & Chang MU.2005. Broad bean wilt virus 2 in yams. J. Gen.Plant Pathol. 71: 441–443.
Kobayashi YO, Kobayashi A, Hagiwara K, Uga H,Mikoshiba Y, Naito T, Honda Y, & Omura T. 2005.Gentian mosaic virus: a new spesies in the genusFabavirus. Phytopathology 95(2): 192–197.

Miftakhurohmah et al. Identifikasi Molekuler BBWV2 dan CYMMV 199
Lakani I, Suastika G, Mattjik N, & Damayanti TA. 2010.Identification and molecular characterization ofOdontoglossum ringspot virus (ORSV) fromBogor, Indonesia. Hayati J. Biosci. 17(2): 101–104.
Lee SC & Chang YC. 2008. Performances andapplication of antisera produced by recombinantcapsid proteins of Cymbidium mosaic virus andOdontoglossum ringspot virus. Eur. J. PlantPathol. 122: 297–306.
Lu HC, Chen CE, Tsai MH, Wang H, Su HJ, & YehHH. 2009. Cymbidium mosaic potexvirusisolate-dependent host movement systems revealtwo movement control determinants and the coatprotein is the dominant. Virology 388(1): 147–159.
Miftakhurohmah, Suastika G, & Damayanti TA. 2013.Deteksi secara serologi dan PCR beberapa jenisvirus yang berasosiasi dengan penyakit mosaikpada tanaman nilam (Pogostemon cablin Benth).J. Littri. 19(3): 130–138.
Naidu RA & Hughes JDA. 2003. Methods for thedetection of plant viral diseases of plant viraldiseases in plant virology in sub-Saharan Africa.Proceedings of Plant Virology. pp. 233–260.Nigeria. http://old.iita.org/cms/details/virology/pdf_files /233-260.pdf. Accessed on 2011 June16.
Natsuaki KT, Tomaru K, Ushiku S, Ichikawa Y,Sugimura Y, Natsuaki T, Okuda S, & TeranakaM. 1994. Characterization of two viruses isolatedfrom patchouli in Japan. Plant Dis. 78(1): 1094–1097.
Noveriza R, Suastika G, Hidayat SH, & KartosuwondoU. 2012. Potyvirus associated with mosaicdisease on patchouli (Pogostemon cablin(Blanco) Benth.) plants in Indonesia. J. ISSAAS.18(1): 131–146.
Putnam ML. 1995. Evaluation of selected methods ofplant diagnosis. Crop Prot. 14(6): 517–525.
Scholthof HB. 2005. Plant virus transport: motions offunctional equivalence. Trends Plant Sci. 10(8):376–382.
Sukamto, Rahardjo IB, & Sulyo Y. 2007. Detection ofPotyvirus on patchouli plant (Pogostemon cablinBENTH) from Indonesia. Proceeding ofInternational Seminar on Essential Oil. pp. 72–77. Jakarta. 7–9 November 2007.
Sumardiyono YB, Sulandari S, & Hartono S. 1996.Penyakit mosaik kuning pada nilam (Pogostemoncablin). Risalah Konggres Nasional XII danSeminar Ilmiah Perhimpunan FitopatologiIndonesia. pp. 912–916. Yogyakarta. 6-8September 1996.
Tamura K, Dudley J, Nei M, & Kumar S. 2007. MEGA4: molecular evolutionary genetic analysis(MEGA) software version 4.0. Mol. Biol. Evol.24(8): 1596 – 1599.





























