Embed Size (px)
Citation preview

STARCH PHOSPHORYLASE OF POTATO
BY D. E. GREEN* AND P. K. STUMPF
(From the DepartwLent of Biological Chemistry, Harvard Medical School, Boston)
(Received for publication, October 10, 1941)
Cori and his colleagues (l-6) discovered and elucidated the series of reversible chemical reactions by which glucose is trans- formed into glycogen in animal tissues. These reactions are catalyzed by a group of enzymes, the most fundamental of which is phosphorylase, the enzyme which catalyzes the condensation of glucose-l-phosphate (Cori ester) to starch or glycogen. The properties of phosphorylase from animal sources have been system- atically investigated by Cori et al. (l-5), while Hanes (7, 8) dis- covered a similar enzyme in extracts from pea and potato, and made a thorough study of the crude potato enzyme. The present investigation deals with the preparation and some of the character- istics of purified potato phosphorylase.
Method of Testing and Preparation
1 unit of phosphorylase activity has been defined as the amount of enzyme which catalyzes the liberation of 0.1 mg. of inorganic phosphate from glucose-l-phosphate in 3minutes at 38” and pH 6.0. The test mixture consisted of 5 to 10 units of enzyme, 0.5 cc. of 0.5 M citrate buffer of pH 6.0, 0.2 cc. of 5 per cent soIubIe starch, and 1 cc. of 0.1 M glucose-l-phosphate, the final volume being 3.5 cc. The glucose-l-phosphate was added to the rest of the mixture after temperature equilibration. The reaction was stopped at some convenient time (5 to 10 minutes) by addition of 5 cc. of 5 per cent trichloroacetic acid and 2 cc. of 2.5 per cent ammonium molybdate’
* Beit Memorial Research Fellow. 1 When ammonium molybdate reagent was added according to the Fiske
and Subbarow phosphate procedure, our test solutions aIways became turbid and made accurate calorimetric readings impossible. Addition of
a55
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
356 Potato Starch Phosphorylase
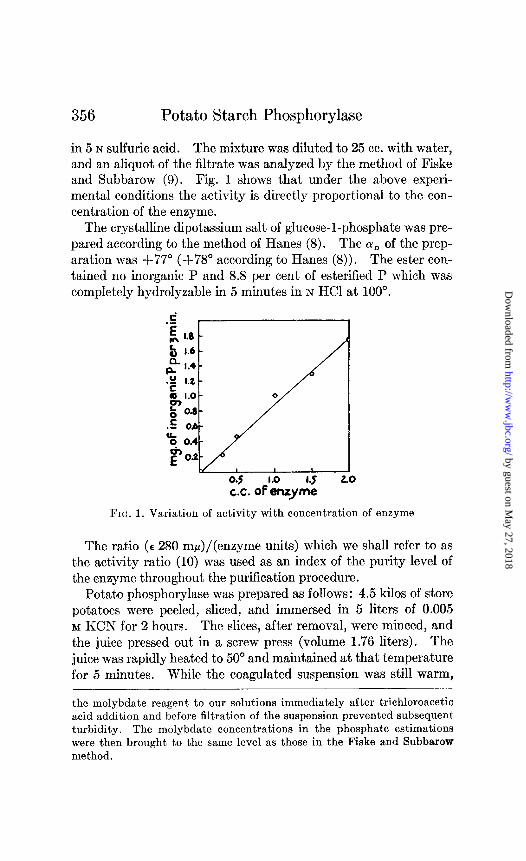
in 5 N sulfuric acid. The mixture was diluted to 25 cc. with water, and an aliquot of the filtrate was analyzed by the method of Fiske and Subbarow (9). Fig. 1 shows that under the above experi- mental conditions the activity is directly proportional to the con- centration of the enzyme.
The crystalline dipotassium salt of glucose-l-phosphate was pre- pared according to the method of Hanes (8). The (Ye of the prep- aration was +77” (+78” according to Hanes (8)). The ester con- tained no inorganic P and 8.8 per cent of esterified P which was completely hydrolyzable in 5 minutes in N HCl at 100’.
0.5 I.0 I.5 2.0 cc. of enzyme
Fro. 1. Variation of activity with concentration of enzyme
The ratio (E 280 mp)/(enzyme units) which we shall refer to as the activity ratio (10) was used as an index of the purity level of the enzyme throughout the purification procedure.
Potato phosphorylase was prepared as follows: 4.5 kilos of store potatoes were peeled, sliced, and immersed in 5 liters of 0.005 M KCN for 2 hours. The slices, after removal, were minced, and the juice pressed out in a screw press (volume 1.76 liters). The juice was rapidly heated to 50” and maintained at that temperature for 5 minutes. While the coagulated suspension was still warm,
the molybdate reagent to our solutions immediately after trichloroacetic
acid addition and before filtration of the suspension prevented subsequent turbidity. The molybdate concentrations in the phosphate estimations were then brought to the same level as those in the Fiske and Subbarow
method.
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
D. E. Green and P. K. Stumpf 357
20 gm. of ammonium sulfate were added for each 100 cc. of fluid. The precipitate was filtered off and discarded. To the clear filtrate were added 15 gm. of ammonium sulfate per 100 cc. of fluid. The precipitate was centrifuged hard and dissolved in 100 cc. of 0.25 M citrate buffer of pH 6.0. The yields of the enzyme are shown in Table I.
The subsequent purification involved four successive series of fractionation with ammonium sulfate, the details of which are summarized in Table I. The fractions retained were Fractions II and III in the first fractionation, Fractions B and C in the second,
TABLE I
Results of Fractionations with Ammonium Sulfate
Fraction No. Degree of saturation
I II
III
IV A B
C D
b” c
Y
0.35 470 0.39 2260 0.44 1520
0.51 135 0.38 680 0.41 1470
0.45 1220 0.52 100
0.37 690
0.42 2440 0.47 520
0.42 2425
-
-
- Total enzyme units Activity ratio
1.41 0.84
0.86 4.00
0.94 0.70 0.71
0.46 0.36
0.30
and Fraction b in the third. Careful fractionation of Fraction b yielded our preparation of highest purity.
From a total of 4400 enzyme units in 1.76 liters of potato juice, the final yield was 2425 enzyme units of activity ratio 0.30 in 20 cc. Since 1 unit in the original potato was equivalent to 120 mg. of dry weight, whereas 1 unit of the final preparation was equivalent to 0.324 mg. of dry weight, the degree of concentration was 120/0.324 or 370. The ratio of protein to carbohydrate in the potato is about 1: 8. The degree of concentration in terms of protein was there- fore about 46.
Other methods of purification were explored without success. Alumina absorption was not found effective in the purification of potato phosphorylase. Only about 35 to 40 per cent of the total
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

358 Potato Starch Phosphorylase
enzyme adsorbed could be recovered with various eluting agents. Dialysis for 12 hours against running tap water led to small losses. The enzyme was inactivated after prolonged dialysis against dis- tilled water but not after prolonged dialysis against 0.2 M KCI. Precipitation in the cold with alcohol inactivated the enzyme com- pletely. The enzyme was stable at 0” in dilute ammonium sulfate solution (about pH 6 to 7) for some days; but steady though slow deterioration of activity was observed over a period of weeks. Activity rapidly declined when the enzyme was exposed to solu- tions the pH of which was less than 5 or greater than 9. Stability and activity both were maximum at about pH 6.
The final preparation was always brown in color. Ultracentri- fuge results showed that the substance responsible for the color was not associated with the protein components of the preparation and was apparently of low molecular weight. The colored component could be reduced with NazSz04 to a light yellow substance (non- oxidizable) without affecting the activity of the enzyme. There was no evidence of any functional relationship between the amount of color and the enzymic activity of different preparations.
Samples of our best preparations (activity ratio 0.3) were ex- amined by Dr. Oncley in the ultracentrifuge. Two components were found present in the ratio of 7:3 with sedimentation con- stants @Lo X 1013) of 5.5 to 5.7 and 2.3 to 2.5 respectively. Until the purification is advanced a stage further, it cannot be decided which of the two components is identical with the enzyme.
At the 0.3 activity ratio level of purity, 1 mg. of protein is equivalent to 3.08 enzyme units. Thus, 1 mg. of protein under the conditions of the test catalyzes the formation of 6.16 mg. of inorganic P per hour, which corresponds to the formation of 32 mg. of starch in 1 hour at 38”.
Components of Catalytic System-There are three components necessary for the catalytic condensation of glucose-l-phosphate to starch: (1) enzyme, (2) glucose-l-phosphate, and (3) catalytic amounts of a suitable polysaccharide, e.g. starch, dextrin, or glycogen. The animal enzyme has been shown by Cori and Cori (4) to be a highly dissociated complex of specific protein and adenylic acid. However, the activity of the potato enzyme is neither dependent upon nor influenced by the presence of adenylic acid. The possibility was examined that the potato enzyme was a
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. E. Green and P. K. Stumpf 359
firmly conjugated compound of specific protein and adenylic acid which was not resolved in the course of the purification procedure. This was tested with the animal enzyme by using boiled or acid- hydrolyzed preparations of potato enzyme in concentrated form as a source of adenylic acid. The results of these experiments showed no evidence from this direction that adenylic acid is present in potato phosphorylase.
A mixture of glucose-l-phosphate (dextrin-free) and the enzyme does not yield starch. On addition of catalytic amounts of starch, dextrin2 or glycogen, synthesis of starch with simultaneous libera- tion of inorganic P ensues. The necessity for added polysaccharide is obscured when a sample of naturally prepared glucose-l-phos- phate is tested. Traces of catalytically active dextrins are in- variably present and these are not removed completely by suc- cessive crystallizations. Thus we have never been able to demon- strate a perfect blank in absence of added polysaccharide, using naturally prepared glucose-l-phosphate. Through the kindness of Dr. C. Cori, we were able to test a sample of synthetic glucose- l-phosphate prepared by the method of Cori et al. (11). The evi- dence was clear (cf. Table II) that in complete absence of poly- saccharide, glucose-l-phosphate was not condensed to starch by the potato phosphorylase.
Specificity of Components-Our experiments confirm the ob- servations of Hanes (8) that glucose-6-phosphate, glucose, fructose, fructose-l-phosphate, and fructose-l : 6-phosphate cannot replace glucose-l-phosphate as substrate.
Starch was the most active polysaccharide catalyst for potato phosphorylase. Dextrin was slightly less active, glycogen showed 77 per cent of the activity of starch, while sucrose, glucose, the Schardinger dextrins, and maltose were inactive.
The form or the origin of the starch employed appeared to be immaterial. Corn, rice, arrow-root, and wheat starches were as effective as potato starch. Similarly, soluble as well as insoluble amyloses were interchangeable. Samples of highly purified corn- starch and glycogen showed no decrease in catalytic activity com- pared to the corresponding crude products. Furthermore, when starch was treated with diastase for a period sufficient to eliminate
* Merck’s dextrin reagent.
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

360 Potato Starch Phosphorylase
staining power with iodine, the catalytic power of the hydrolysate also disappeared. Catalytic action cannot, therefore, be ascribed to a non-polyhexose impurity.
Hanes (8) has reported that maltose in high concentration shows some activity as a catalyst. While our experiments with crude
TABLE II
Effect oj Polysaccharide Catalyst on Starch Formation
Each test mixture consisted of 1 cc. of enzyme, 0.5 cc. of 0.5 M citrate buffer of pH 6, and 1 cc. of 0.1 M glucose-l-phosphate.
Additions
Synthetic glucose-l-phosphate.. . “ I‘ + 12.5 mg. soluble starch.
Glucose-l-phosphate isolated from potato juice.. . ‘I ‘I ‘I ‘I “ + 12.5 mg.
w.
0.00 1.14 0.08
soluble starch............................................ 1.15
Inorganic P per 3 min.
TABLE III
Decrease in Catalytic Effect with Purijication of Maltose
Each test mixture consisted of 1 cc. of enzyme (8 units), 1 cc. of 0.1 M dextrin-free glucose-l-phosphate, and 0.5 cc. of 0.5 M citrate buffer of pH
6.0.
Polysaocharide oat&lyst Catalytic effect, mg. inorganic P
per 3 min.
None.................................................... 5mg.starch............................................. 50 “ commercialmaltose................................
50 “ once recrystallized maltose.. _. 50 “ twice “ “ ..,,.....................
0.00
0.80 0.67 0.35 0.18
maltose confirmed this observation, recrystallization of maltose preparations whittled the catalytic effect down to the vanishing point (cf. Table III). Here again, the presence of catalytic amounts of dextrin impurity had to be reckoned with. This im- purity was not completely eliminated after two successive re- crystallizations of maltose.
Factors Afleeting Activity of Enzyme-The effects of variation of
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
D. E. Green and P. K. Stumpf 361
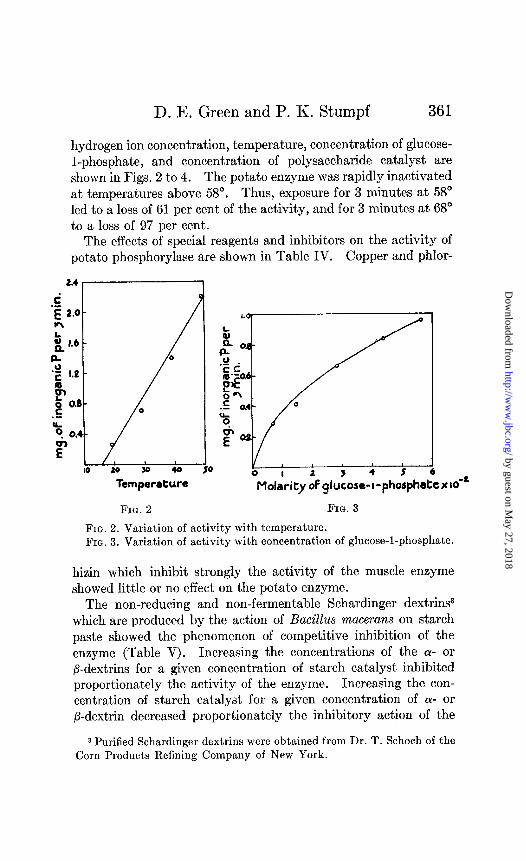
hydrogen ion concentration, temperature, concentration of glucose- l-phosphate, and concentration of polysaccharide catalyst are shown in Figs. 2 to 4. The potato enzyme was rapidly inactivated at temperatures above 58”. Thus, exposure for 3 minutes at 58” led to a loss of 61 per cent of the activity, and for 3 minutes at 68” to a loss of 97 per cent.
The effects of special reagents and inhibitors on the activity of potato phosphorylase are shown in Table IV. Copper and phlor-
2.4
*z 2.0- n
5 I.6 -
P-
*f I.0 - ID 0, “, OB-
.- IL 9 0.4-
?
IO 20
Temperature Molarity of glucose+phosphatexIdt
FIG. 2 FIG. 3
FIG. 2. Variation of activity with temperature. FIG. 3. Variation of activity with concentration of glucose-l-phosphate.
hizin which inhibit strongly the activity of the muscle enzyme showed little or no effect on the potato enzyme.
The non-reducing and non-fermentable Schardinger dextrins3 which are produced by the action of Bacillus macerans on starch paste showed the phenomenon of competitive inhibition of the enzyme (Table V). Increasing the concentrations of the cr- or P-dextrins for a given concentration of starch catalyst inhibited proportionately the activity of the enzyme. Increasing the con- centration of starch catalyst for a given concentration of LY- or fl-dextrin decreased proportionately the inhibitory action of the
3 Purified Schardinger dextrins were obtained from Dr. T. Schoch of the Corn Products Refining Company of New York.
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
362 Potato Starch Phosphorylase
Schardinger dextrins. It would appear that the Schardinger dextrins and starch were competing for the same active group in the
TABLE IV
Effect of Various Reagents on Activity of Enzyme
Reagent Final concentration
-@NOa I‘
M/50,000
M/10,000
CuSOa “
‘I
znsoa
HgClz PbAcz Iodoacetic acid Sodium fluoride Phlorhizin
I‘
Glucose Potassium cyanide Capryl alcohol Sulfanilamide Hydrogen peroxide Nad3zOn
M/50,000 0 M/10,0@ 0 M/l, 000 0 M/25,000 0 M/50,000 0 ~,‘25,000 0 M/27 0 M/27 24 0.7 saturated 18 0.3 “ 0 M/27 0 M/27 0 Saturated solution 0 5.3 mg. y. 0
1% 0 53 mg. % 0
-
TABLE V
Inhibition
per Cent 8
80
Inhibition by Schardinger Dextrins
Each reaction mixture consisted of 1 cc. of enzyme (12 units), 1 cc. of 0.1 M glucose-l-phosphate, and 0.5 cc. of 0.5 M citrate buffer of pH 6.0.
Dextrin Soluble starch Inhibition
mo. ml. per cent
15 12.5 23 25 12.5 52 50 12.5 73 50 12.5 73 50 25.0 50 50 50.0 21
enzyme. This inhibitory action was not shown by inulin, sucrose, or maltose in a similar range of concentration.
Properties of Starch Formed in Reaction-A mixture of glucose-
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
D. E. Green and P. K. Stumpf 363
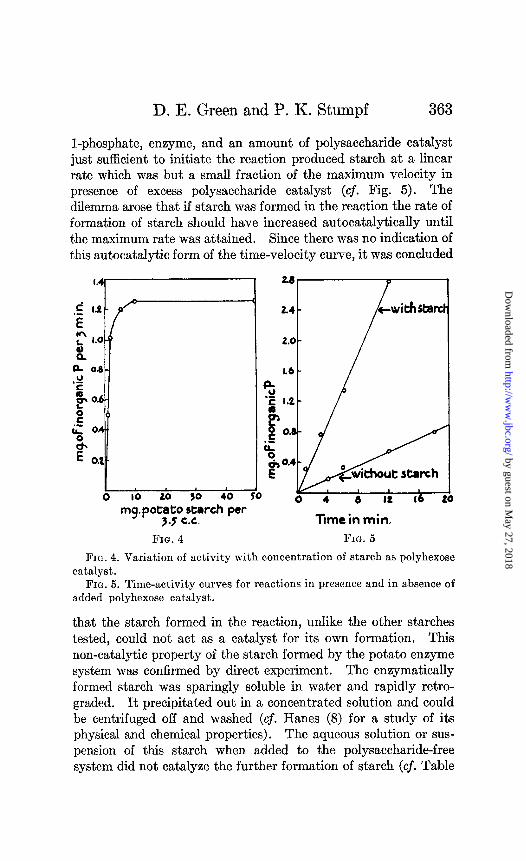
l-phosphate, enzyme, and an amount of polysaccharide catalyst just sufficient to initiate the reaction produced starch at a linear rate which was but a small fraction of the maximum velocity in presence of excess polysaccharide catalyst (cf. Fig. 5). The dilemma arose that if starch was formed in the reaction the rate of formation of starch should have increased autocatalytically until the maximum rate was attained. Since there was no indication of this autocatalytic form of the time-velocity curve, it was concluded
mg.potato starch per 3.5 C.C.
FIG. 4
2.0
Lb t /
Time in min.
FIG. 5
FIG. 4. Variation of activity with concentration of starch as polyhexose
catalyst. FIG. 5. Time-activity curves for reactions in presence and in absence of
added polyhexose catalyst.
that the starch formed in the reaction, unlike the other starches tested, could not act as a catalyst for its own formation. This non-catalytic property of the starch formed by the potato enzyme system was confirmed by direct experiment. The enzymatically formed starch was sparingly soluble in water and rapidly retro- graded. It precipitated out in a concentrated solution and could be centrifuged off and washed (cf. Hanes (8) for a study of its physical and chemical properties). The aqueous solution or sus- pension of this starch when added to the polysaccharide-free system did not catalyze the further formation of starch (cf. Table
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

364 Potato Starch Phosphorylase
VI). It would be of considerable interest to know the chemical basis for this difference in catalytic properties between reagent starch and the starch formed as the reaction product of phosphory- lase action. Comparison of iodine staining power (8), x-ray diagrams (6, 12), and elementary analysis (8) have failed thus far to pick out any significant differences.
TABLE VI
Synthesized Starch As Catalyst
Each reaction mixture consisted of 1 cc. of enzyme, 1 cc. of 0.1 M glucose-
l-phosphate, and 0.5 cc. of 0.5 M citrate buffer of pH 6.0.
Catalytic effect, mg. inorganic P
per 3 min.
Starch synthesized by potato phosphorylase (5 mg.). 0.00 Soluble potato starch (5 mg.). 0.64 Insoluble potato starch (5 mg.). 0.64
TABLE VII
Components of Lima Bean Phosphorylase System
Each reaction mixture consisted of 0.5 cc. of enzyme and 0.5 cc. of 0.5 M
citrate buffer at pH 6.0.
system Ingrggic
3 min.
Enzyme......................................................... 0.00 I‘ “ I‘
I‘
+ glucose-l-phosphate.. 0.62
+ “ + soluble starch. 0.67
+ “ + dextrin.. 0.63
+ “ + soluble starch + adenylic acid.. 0.65
Rijle of Polysaccharide Catalyst-The polyhexoses that are catalytically active in starch formation have two characteristics in common: (a) they are all composed of chains of cr-glucose units in 1: 4 glucosidic linkage, and (b) they can all be phosphorylyzed to glucose-l-phosphate in presence of inorganic phosphate and the enzyme. The chain length appears to be of importance. Thus LY and /3 Schardinger dextrins with probable chain lengths of four ar,d six respectively are inactive, whereas the violet-staining dextrins of between six and twelve are fully active. The fact that
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. E. Green and P. K. Stumpf 365
a polyhexose catalyst must be capable of being phosphorylated may bear some relation to its catalytic role in starch formation. Pos- sibly the initial step in starch formation is the transfer of phos- phate from glucose-l-phosphate to the catalyst. Inorganic phosphate would then arise by the spontaneous decomposition of the catalyst-phosphate compound.
The starch phosphorylase enzyme of Lima bean has been pre- pared by the same method as that employed in the purification of the potato enzyme, and has been brought to a comparable degree of purity. Only two components were necessary for starch form- ation, (a) the enzyme and (b) glucose-l-phosphate (cf. Table VII). Addition of starch, dextrin, or glycogen did not increase the rate of condensation of glucose-l-phosphate to starch. This result can- not be explained in terms of some polysaccharide contaminating the enzyme, as is shown by the fact that the preparation of the Lima bean enzyme did not act as a source of polyhexose catalyst for the potato enzyme. There are two possible explanations: either the polyhexose catalyst is an integral part of the Lima bean enzyme molecule or the mechanism of the reaction is different, and does not depend upon the collaboration of a polyhexose.
SUMMARY
The purification and some of the properties of potato phosphory- lase are described. The enzyme was concentrated some 370 times by a series of successive ammonium sulfate fractionations. Ade- nylic acid is not a component of the system, although catalytic amounts of starch, dextrin, or glycogen are required for starch formation from glucose-l-phosphate. Some of the factors affect- ing starch formation by the enzyme are analyzed.
We are grateful to Professor A. B. Hastings for his interest and encouragement. This investigation was assisted by grants from the Ella Sachs Plotz Foundation and the American Association for the Advancement of Science.
BIBLIOGRAPHY
I.. Cori, G. T., and Cori, C. F., Proc. Sot. Ezp. Biol. and Med., 33, 1075 (1935).
2. Cori, G. T., Colowick, S. P., and Cori, C. F., J. Biol. Chem., 133, 381
(1938); 127, 771 (1939).
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Potato Starch Phosphorylase
3. Cori, G. T., and Cori, C. F., J. Bid. Chem., 131,397 (1939). 4. Cori, G. T., and Cori, C. F., J. Biol. Chem., 136, 733 (1940). 5. Cori, G. T., Cori, C. F., and Schmidt, G., J. Biol. Chem., 129,629 (1939). 6. Bear, R. T., and Cori, C. F., J. Biol. Chem., 140, 111 (1941). 7. Hanes, C. S., Proc. Roy. Sot. London, Series B, 128,421 (1940). 8. Hanes, C. S., Proc. Roy. Sot. London, Series B, 129, 174 (1940). 9. Fiske, C. H., and Subbarow, Y., J. Biol. Chem., 66, 375 (1925).
10. Green, D. E., Herbert, D., and Subrahmanyan, V., J. BioZ. Chem., 138, 327 (1941).
11. Cori, C. F., Colowick, S. P., and Cori, G. T., J. BioZ. Chem.; 121, 465 (1937).
12. Astbury, W. T., and Hanes, C. S., Nature, 146, 558 (1940).
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. E. Green and P. K. StumpfPOTATO
STARCH PHOSPHORYLASE OF
1942, 142:355-366.J. Biol. Chem.
http://www.jbc.org/content/142/1/355.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/142/1/355.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from















![Production of tapioca starch [tm] sweet potato.pdf · By applying different reaction conditions - temperature, ... Sweet potato starch is used in the manufacture ... in the popular](https://img.pdfslide.us/doc/110x75/5b612e817f8b9a4f488c1032/production-of-tapioca-starch-tm-sweet-potatopdf-by-applying-different-reaction.jpg)













