Embed Size (px)
Citation preview

Roux's Arch Dev Biol (1986) 195:290-295 Roux's Archives of Developmental Biology �9 Springer-Verlag 1986
Stage- and tissue-specific expression of the genes encoding callipllorin, the major larval serum protein of CMliphora vicina Heide Schenkel und Klaus Scheller Zoologisches Institut der Universitfit Wfirzburg, R6ntgenring 10, D-8700 Wfirzburg, Federal Republic of Germany
Summary. The stage- and tissue-specific biosynthesis of cal- liphorin was analysed during the development of the blow- fly, Calliphora vicina. Western blot analyses show that the protein is not present in eggs, whereas it can be detected in fat body, brain, imaginal disk, salivary gland and epider- mis throughout all postembryonic stages, including the adult one. By Northern analysis a unique 2.6 kb mol.wt. mRNA coding for calliphorin is identified exclusively in the fat body tissue of larvae, pupae and adults. Hybridiza- tion experiments of in vivo labelled poly(A) + RNA with filter-bound calliphorin genes indicate that the genes are transcribed until pupariation. However, the translation of the calliphorin mRNA stops at the end of the feeding stage, as shown by [35S]-methionine incorporation.
Key words: Calliphora - Fat body Calliphorin biosynthe- sis - Larval serum proteins - Gene polymorphism - Tran- scription-translation uncoupling
Introduction
Calliphorin belongs to the group of larval serum proteins (also named LSPs, arylphorins or "storage proteins") which could be identified in numerous holometabolous and heterometabolous insects. Among dipterans these proteins show similarities in their size, subunit and amino acid com- position as well as in their developmental profile (for re- view, see Levenbook 1985). Calliphorin serves the following functions: it acts as a storage protein, serves as a carrier protein for ecdysteroid hormones in the haemolymph and is involved in the sclerotization process of cuticle (for review see Scheller 1983). Calliphorin is composed of a series of closely related but heterogeneous hexamers of ~ 500 kilo- daltons (kd) molecular weight. The different polypeptide subunits of 80 kd are encoded by a multigene family of at least 20 genes which are clustered in region 4 a of chromo- some 2 of Calliphora vicina (Schenkel et al. 1985). As we have shown previously the biosynthesis of calliphorin is developmentally regulated. The protein is synthesized in young larvae up to the end of the feeding period, whereas the calliphorin mRNA is present in an untranslatable state until pupariation (Schenkel et al. 1983). In the present arti- cle we extend our description of the calliphorin biosynthesis and show that the expression of the gene family encoding
Offprint requests to: K. Scheller at the above address
calliphorin is stringently regulated so that the transcription occurs only in the fat body cells for a limited period of the development of the blowfly. Furthermore, we describe the translation of the calliphorin mRNA in the fat body and the accumulation of the protein in different tissues.
Materials and methods
Materials. Restriction endonucleases, E. coli DNA poly- merase I and poly(U)-Sepharose were purchased from Pharmacia (Freiburg, FRG), pancreatic DNase I and pro- teinase K from Boehringer (Mannheim, FRG), nitrocellu- lose filters BA 85 from Schleicher and Schiill (Dassel, FRG). Immunostaining reagents were obtained from Sigma and Dakopatts (Hamburg, FRG), c~-[32p]-dATP (> 1000 Ci/mmol), 5,6-[3H] uridine (50 Ci/mmol) and [L-]35 S-methionine (> 800/retool) from Amersham- Buchler (Braunschweig, FRG). Dupont Cronex 4 x-ray films were used for autoradiography.
Animals. Calliphora vicina R.-D. were reared as described in an accompanying paper (K6nig et al. 1986). The animals used in this study derived from the F6-generation of an inbred line, established in our laboratory as the progeny of a single pair of flies.
Immunoprecipitation. Immunoprecipitation of in vivo-la- belled fat body proteins was performed as described else- where (Schenkel and Scheller 1983). Anti-rabbit calliphorin immunoglobulin (IgG) was prepared as described by K6nig et al. (1986).
Dissection of tissues. Tissues from larvae, puparia and adults were dissected under a binocular microscope in ster- ile insect Ringer's solution. Tissues for protein and RNA preparation were collected on dry ice and kept at - 7 5 ~ C until extraction.
Protein extraction and sodium dodecyl-polyacrylamide gel electrophoresis (SDS-PAGE). Tissues were homogenized in sample buffer (80 mM tris-HC1, pH 6.8, 2% SDS, 10% gly- cerol, 0.3 M fl-mercapto-ethanol). After 15 min extraction at room temperature the insoluble material was sedimented by centrifugation (5 rain, 10000g) and the supernatants submitted to PAGE according to Laemmli and Favre (1973) with the modification described by K6nig and Scheller (1986). Immunoblotting of the proteins is described in the accompanying article (K6nig et al. 1986).

Nucleic acid preparation and transfer to nitrocellulose filters. DNA was prepared from single flies by proteinase digestion followed by phenol extraction as described by Schenkel et al. (1985). Total RNA was isolated from different tissues by means of hot phenol (Lepesant et al. 1978). Poly(A) + RNA was prepared from total fat body RNA by poly(U)- Sepharose chromatography according to Scheller and Karl- son (1977). Northern and Southern transfer and hybridiza- tion of the nitrocellulose filters are described in detail else- where (Schenkel et al. 1985).
In vitro labelling of calliphorin genes and marker DNA. The calliphorin gene used as a hybridization probe was sub- cloned in the plasmid pC 223 (see Schenkel et al. 1985). The coding sequences for calliphorin were isolated from pC 223 by digestion with the restriction enzymes Barn HI and Bgl II followed by electro-elution. The purified DNA fragment was labelled by nick-translation according to Rigby et al. (1977) to a specific activity of 3 x 107 cmp/gg using ~_[32p] dATP. Radioactive molecular weight markers for RNA gels were obtained by 3'-end labelling with the Klenow fragment of DNA polymerase I of 2 DNA digested with Hind III.
Filter hybridization of in vivo labelled poly(A) + RNA. Five micrograms of linearized plasmid pC 223 and of )vDNA (controls), were each dissolved in H z O , denatured by boil- ing for 2 min and applied to a nitrocellulose disk of 8 mm in diameter. The DNA was bound to nitrocellulose by bak- ing for 2 h at 80 ~ C. Hybridization of complementary-RNA was performed in HS-buffer - 0.1 M pipes [piperazine-N- N'-bis(2-ethane sulphonic acid)], pH 7.2, 0.5 M NaC1, 10 mM EDTA [ethylendiaminotetraacetic acid], 65% form- amide - containing 5 I~g of in vivo labelled poly(A) + RNA. Each RNA probe to be tested was hybridized together with both the filter-bound pC 223 and ~,DNA. After 24 h of incubation at 58 ~ C the filters were intensively washed: ten times with 1 x SSC (0.15 M NaC1, 15 mM citric acid tri- sodium salt), containing 0.5% SDS, at 65 ~ C and five times with 10 mM tris-HC1, pH 8.0, containing 2 mM EDTA. The last washing solution did not show any radioactivity. The radioactivity retained on the filters was measured in a Packard Prias scintillation counter. Each value for pC 223 was corrected by subtraction of the radioactivity unspecifi- cally bound to the corresponding ~,DNA filter.
Results
Demonstration of calliphorin in different tissues during development
Proteins from fat bodies, salivary glands, epidermis, ima- ginal disks and brains of Calliphora at different develop- mental stages were extracted and separated by PAGE. After transfer of the proteins to nitrocellulose filters, calliphorin was detected by anti-rabbit calliphorin immunoglobulins on Western blots containing the above tissues of 1st, 2nd and 3rd instars and white puparia. Furthermore, calliphorin could be found in whole pupae, pharate adults, adult blow- flies, and, to our surprise, in testes and ovaries. By contrast, in freshly deposited eggs as well as in late embryos calli- phorin was left undetected (Fig. 1). This indicates that calli- phorin appears to be ubiquitous in Calliphora from the beginning of the 1st larval instar and, therefore, was asked
291
Fig. l. Identification of calliphorin in different tissues of Calliphora by immunoblot. Equal amounts (0.05 OD28o) of total protein per lane were resolved by polyacrylamide gel electrophoresis (PAGE), transferred to nitrocellulose filter and visualized by an antibody staining method as described in Materials and methods. The lanes show: a brain, b fat body, c epidermis of 1-day-old larvae, d ima- ginal disks, e fat body-free salivary glands of 2-day-old larvae, ftestis, g fat body, h ovarium of adults, i buffer (negative control), k newly deposited eggs, 1 late embryos, m purified calliphorin (posi- tive control)
whether the protein was synthesized then in different tissues or derives from a common source. To answer this question we examined the appearance of the mRNA coding for calli- phorin by a series of Northern blot analyses.
Stage and tissue specificity of calliphorin mRNA
Total RNA was extracted from different tissues. After sepa- ration by electrophoresis on formamide-agarose gels RNA was blotted to nitrocellulose. These Northern blots were hybridized to nick-translated 2.3 kb (2.3 x 103 bases) Barn HI/Bgl II restriction fragments of the plasmid pC 223, con- taining the coding region for calliphorin (see Materials and methods). RNA prepared from brains, imaginal disks, epi- dermis, muscles, salivary glands, eggs, late embryos, testes and ovaries did not cause any hybridization signal, whereas specific hybridization could be detected with RNA of fat bodies from the beginning of the 3rd instar up to adult stage. This suggests that the transcription of the calliphorin gene family is restricted to the fat body and, therefore, tissue-specific and regulated during the development of Cal- liphora. For that reason we further analysed the biosynthe- sis of calliphorin in the fat body with particular interest in the stage-specific appearance, the transcription and the translation of calliphorin mRNA.
The Northern blot analysis depicted in Fig. 2 represents the developmental profiles of the calliphorin gene tran- scripts in fat body cells. No hybridization signal could be observed with RNA from eggs and late embryos even when gels were both overloaded and overexposed. RNA from 1st and 2nd instar fat bodies gave only a very weak signal. This suggests that the calliphorin mRNA may be rapidly utilized and degraded. In contrast, calliphorin gene prod- ucts accumulated to high amounts in the fat body immedi- ately after the second larval moult, reached a maximum in 4-day-old larvae (4d-L) and then continuously decreased,
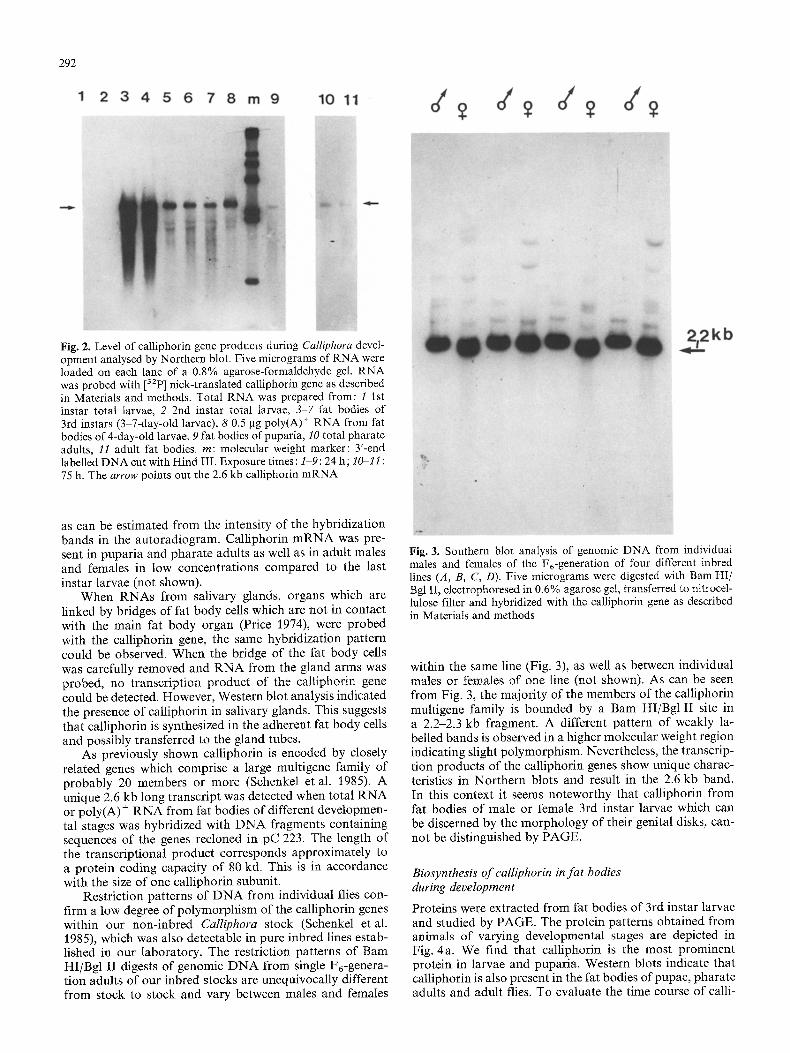
292
Fig. 2. Level of calliphorin gene products during Calliphora devel- opment analysed by Northern blot. Five micrograms of RNA were loaded on each lane of a 0.8% agarose-formaldehyde gel. RNA was probed with [32p] nick-translated calliphorin gene as described in Materials and methods. Total RNA was prepared from: l 1st instar total larvae, 2 2nd instar total larvae, 3-7 fat bodies of 3rd instars (3-7-day-old larvae), 8 0.5 gg poly(A) + RNA from fat bodies of 4-day-old larvae, 9 fat bodies of puparia, 10 total pharate adults, 11 adult fat bodies, m: molecular weight marker: 3'-end labelled DNA cut with Hind III. Exposure times : 1-9:24 h; 1~11 : 75 h. The arrow points out the 2.6 kb calliphorin mRNA
as can be estimated from the intensity of the hybridization bands in the autoradiogram. Calliphorin mRNA was pre- sent in puparia and pharate adults as well as in adult males and females in low concentrations compared to the last instar larvae (not shown).
When RNAs from salivary glands, organs which are linked by bridges of fat body cells which are not in contact with the main fat body organ (Price 1974), were probed with the calliphorin gene, the same hybridization pattern could be observed. When the bridge of the fat body cells was carefully removed and RNA from the gland arms was probed, no transcription product of the caltiphorin gene could be detected. However, Western blot analysis indicated the presence of calliphorin in salivary glands. This suggests that calliphorin is synthesized in the adherent fat body cells and possibly transferred to the gland tubes.
As previously shown calliphorin is encoded by closely related genes which comprise a large multigene family of probably 20 members or more (Schenkel et al. 1985). A unique 2.6 kb long transcript was detected when total RNA or poly(A) + RNA from fat bodies of different developmen- tal stages was hybridized with DNA fragments containing sequences of the genes recloned in pC 223. The length of the transcriptional product corresponds approximately to a protein coding capacity of 80 kd. This is in accordance with the size of one calliphorin subunit.
Restriction patterns of DNA from individual flies con- firm a low degree of polymorphism of the calliphorin genes within our non-inbred Calliphora stock (Schenkel et al. 1985), which was also detectable in pure inbred lines estab- lished in our laboratory. The restriction patterns of Barn HI/Bgl II digests of genomic DNA from single F6-genera- tion adults of our inbred stocks are unequivocally different from stock to stock and vary between males and females
Fig. 3. Southern blot anaIysis of genomic DNA from individual males and females of the F6-generation of four different inbred lines (A, B, C, D). Five micrograms were digested with Barn HI/ Bgl II, electrophoresed in 0.6% agarose gel, transferred to nitrocel- lulose filter and hybridized with the calliphorin gene as described in Materials and methods
within the same line (Fig. 3), as well as between individual males or females of one line (not shown). As can be seen from Fig. 3, the majority of the members of the calliphorin multigene family is bounded by a Bam HI/Bgl II site in a 2.2-2.3 kb fragment. A different pattern of weakly la- belled bands is observed in a higher molecular weight region indicating slight polymorphism. Nevertheless, the transcrip- tion products of the calliphorin genes show unique charac- teristics in Northern blots and result in the 2.6 kb band. In this context it seems noteworthy that calliphorin from fat bodies of male or female 3rd instar larvae which can be discerned by the morphology of their genital disks, can- not be distinguished by PAGE.
Biosynthesis o f calliphorin in fa t bodies during development
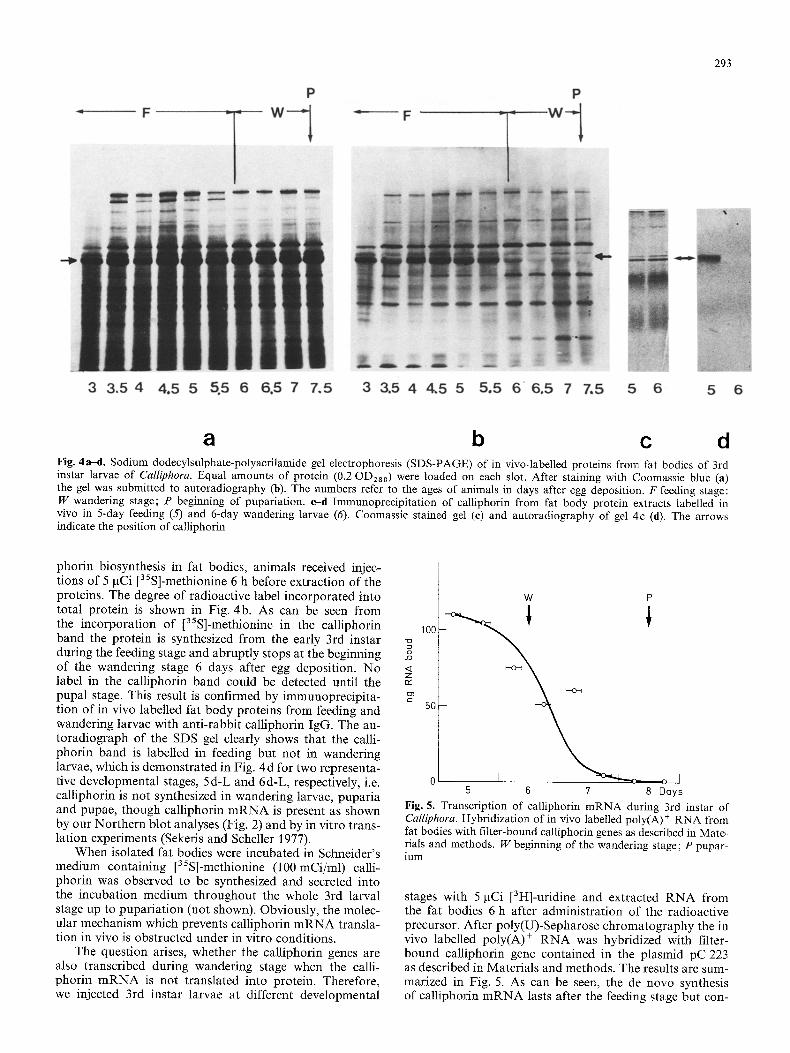
Proteins were extracted from fat bodies of 3rd instar larvae and studied by PAGE. The protein patterns obtained from animals of varying developmental stages are depicted in Fig. 4a. We find that calliphorin is the most prominent protein in larvae and puparia. Western blots indicate that calliphorin is also present in the fat bodies of pupae, pharate adults and adult flies. To evaluate the time course of calli-
293
a b c d Fig. 4a-d. Sodium dodecylsulphate-polyacrilamide gel electrophoresis (SDS-PAGE) of in vivo-labelled proteins from fat bodies of 3rd instar larvae of Calliphora. Equal amounts of protein (0.2 OD280) were loaded on each slot. After staining with Coomassie blue (a) the gel was submitted to autoradiography (b). The numbers refer to the ages of animals in days after egg deposition. F feeding stage: W wandering stage; P beginning of pupariation, e-d Immunoprecipitation of calliphorin from fat body protein extracts labelled in vivo in 5-day feeding (5) and 6-day wandering larvae (6). Coomassie stained gel (e) and autoradiography of gel 4c (d). The arrows indicate the position of calliphorin
phorin biosynthesis in fat bodies, animals received injec- tions of 5 gCi [35S]-methionine 6 h before extraction of the proteins. The degree of radioactive label incorporated into total protein is shown in Fig. 4b. As can be seen from the incorporation of [35S]-methionine in the calliphorin band the protein is synthesized from the early 3rd instar during the feeding stage and abruptly stops at the beginning of the wandering stage 6 days after egg deposition. No label in the calliphorin band could be detected until the pupal stage. This result is confirmed by immunoprecipita- tion of in vivo labelled fat body proteins from feeding and wandering larvae with anti-rabbit calliphorin IgG. The au- toradiograph of the SDS gel clearly shows that the calli- phorin band is labelled in feeding but not in wandering larvae, which is demonstrated in Fig. 4 d for two representa- tive developmental stages, 5d-L and 6d-L, respectively, i.e. calliphorin is not synthesized in wandering larvae, puparia and pupae, though calliphorin m R N A is present as shown by our Northern blot analyses (Fig. 2) and by in vitro trans- lation experiments (Sekeris and Scheller 1977).
When isolated fat bodies were incubated in Schneider's medium containing [35S]-methionine (100 mCi/ml) calli- phorin was observed to be synthesized and secreted into the incubation medium throughout the whole 3rd larval stage up to pupariation (not shown). Obviously, the molec- ular mechanism which prevents calliphorin mRNA transla- tion in vivo is obstructed under in vitro conditions.
The question arises, whether the calliphorin genes are also transcribed during wandering stage when the calli- phorin m R N A is not translated into protein. Therefore, we injected 3rd instar larvae at different developmental
W P
100
o
<[ Z 132
50
0 I , o--J 5 6 7 8 Days
Fig. 5. Transcription of calliphorin mRNA during 3rd instar of Calliphora. Hybridization of in vivo labelled poly(A) + RNA from fat bodies with filter-bound calliphorin genes as described in Mate- rials and methods. W beginning of the wandering stage; P pupar- ium
stages with 5 gCi [3H]-uridine and extracted RNA from the fat bodies 6 h after administration of the radioactive precursor. After poly(U)-Sepharose chromatography the in vivo labelled poly(A) § RNA was hybridized with filter- bound calliphorin gene contained in the plasmid pC 223 as described in Materials and methods. The results are sum- marized in Fig. 5. As can be seen, the de novo synthesis of calliphorin mRNA lasts after the feeding stage but con-

294
tinuously decreases during the wandering stage and stops at the beginning of pupariation. This suggests that the bio- synthesis of calliphorin is not regulated on the level of tran- scription in the late 3rd instar Calliphora larvae.
Discussion
Although a wealth of data mostly derived from in vitro studies on larval serum proteins (LSPs) has been accumu- lated over recent years, it could be merely stated that the fat body is the source of LSP-synthesis in dipteran larvae (reviewed by Levenbook 1985). Our Northern analyses now clearly demonstrate that the genes encoding calliphorin, the LSP of Caltiphora, are solely transcribed in the fat body cells, although calliphorin was detected in other larval and pupal tissues. The most direct evidence for this interpreta- tion comes from our experiments with the salivary glands. Calliphorin m R N A detectable in the glands derives from the bridge of fat body cells which connects the gland arms. When RNA from epidermal cells, wing disks and brains were translated in an in vitro system and the newly synthe- sized polypeptides were separated by PAGE, a 80 kd band was labelled but could not be identified by immunological techniques (Schenkel et al. 1983). Obviously, this protein comigrates with calliphorin on the gel but it does not repre- sent calliphorin since the mRNA is lacking in the tissues analysed, as demonstrated by our Northern blots.
Calliphorin shows age-dependent variations in its con- tent. After synthesis in the fat body the protein is secreted into the haemolymph where it accumulates to concentra- tions up to 0.2 mM (Enderle et al. 1983). At the peak of its accumulation calliphorin accounts for 60% of total lar- val protein soluble in diluted salt solution. As shown by our Northern analyses the degree of calliphorin synthesis is correlated with the concentration of calliphorin mRNA. In 1st and 2nd instar larvae the calliphorin m R N A is at a very low level and reaches a maximum in early 3rd instar feeding larvae (3-4 days old), then it slowly declines during the wandering stage and is rapidly reduced after beginning of pupariation. Although calliphorin mRNA is present dur- ing the wandering stage (Fig. 2) it is not translated into protein during this period in fat body tissue (Fig. 4). Our previous observations (Sekeris and Scheller 1977), obtained by analysis of whole animals, are in total agreement with these results, since whole larvae reflect the situation in fat bodies which we have shown to be the only site of calli- phorin biosynthesis. The accumulation of untranslated cal- liphorin m R N A can be explained by the observation that the m R N A is stored in the fat body of wandering larvae in the shape of ribonucleoprotein particles (RNPs). These RNPs appear at the end of the feeding stage and increase continuously until pupariation and are then rapidly de- graded (Schenkel et al. 1983). Further evidence that calli- phorin mRNA is present in fat bodies of wandering larvae is twofold. First, when poly(A) + RNA was translated in a wheat germ system, among the translational products cal- liphorin could be identified (Sekeris and Scheller 1977). Sec- ondly, in vitro cultured fat bodies incorporate [3ss]-methio- nine into caUiphorin (not shown). This suggests that calli- phorin m R N A is artificially reactivated for translation ei- ther by the culture conditions or by the RNA extraction procedure. The main conclusion which can be drawn from our studies is that the tissue-specific biosynthesis of calli- phorin is developmentally regulated. While transcription of
the calliphorin genes is paralleled by translation of the mRNA in young, rapidly growing larvae, translation stops in late larvae which are ready for metamorphosis, although calliphorin mRNA is still present, and transcription of the calliphorin genes continues. Obviously, the biosynthesis of calliphorin is regulated not only at the transcriptional level. Calliphorin mRNA translation is uncoupled from the tran- scription of the calliphorin genes in late larvae.
Although the LSPs in dipteran insects are immunologi- cally related and show similar physicochemical properties, the regulation of their biosynthesis is different in related species. In Ceratitis the LSPs are synthesized in the late larval and white pupal stage as shown by in vivo labelling of salt-extractable fat body proteins (Chrysanthis et al. 1981, Kaliafas et al. 1984). In vitro translation of poly(A) + RNA suggest the presence of m R N A for LSPs in last larval stages of Sarcophaga (Tahara et al. 1982) and Dacus (Pa- trinou-Georgoula et al. 1983) and Ceratitis (Mintzas et al. 1983). In Drosophila the mRNA coding for LSP 1, which is the protein homologous to calliphorin, is first detectable 1 h after the second moult and remains at a high level until wandering (= climbing) stage and then decreases and is not present in prepupae as shown by in vitro translation experi- ments (Lepesant et al. 1982). Recent studies have shown that the synthesis of LSP 1 also ceases when there is abun- dant translatable LSP 1 mRNA (Powell et al. 1984).
There is evidence from the tests involving the 20-OH- ecdysone-deficient Drosophila mutant ecd 1 that supplemen- tation with exogenous 20-OH-ecdysone enhances in vivo expression of the genes encoding two LSP-1 subunits sug- gesting that the hormone might be involved in the regula- tion of these genes (Lepesant et al. 1982). From experiments of our laboratory (injections of larvae and exposure of iso- lated fat bodies to different 20-OH-ecdysone concentra- tions) we cannot state a relation between the ecdysteroid hormone and the initiation of transcription of calliphorin genes in Calliphora larvae. We note, however, a clear corre- lation between the ecdysteroid titre in Calliphora larvae (Walter and Koolman 1985) and the cessation of calliphorin synthesis and degradation of the calliphorin mRNA. A short rise of the ecdysteroid titre in 5-day-old larvae coin- cides with the shut off of calliphorin mRNA translation and the increase of the ecdysteroid concentration in white puparia parallels the degradation of calliphorin mRNA.
Inspections of Southern blots with genomic DNA from individual males and females from inbred lines revealed a surprising result. The restriction patterns of the calli- phorin genes appear differently in different stocks as well as in males and females of the inbred line. Since the restric- tion patterns also vary between individual males or females, we cannot claim a sex specificity of the calliphorin genes. In spite of the polymorphism of the calliphorin genes a unique 2.6 kb mRNA encodes a uniform protein in all tis- sues and developmental stages tested. It is possible though that different members of the calliphorin gene family are expressed, and/or that the primary gene products are pro- cessed in a variable way. Detailed analysis using cDNA clones are currently underway to resolve this question.
Acknowledgements. We are grateful to Renate Bader and Dr. Erich Bnchner for providing immunochemicals and the equipment for the Western analysis. We thank Andreas Keil, Hans-Georg Spies and Claus Steinlein for excellent technical assistance. This work was supported by grants of the Deutsche Forschungsgemeinschaft (Sche 195/4) and the Universit/itsbund Wiirzburg.

295
R e f e r e n c e s
Chrysanthis G, Marmaras V J, Christodoulou (1981) Major haemo- lymph proteins in Ceratitis capitata: biosynthesis and secretion during development. Wilhelm Roux's Archives 190:33-39
Enderle U, Kfiuser L, Reum L, Scheller K, Koolman J (1983) Ecdysteroids in the haemolymph of the blowfly larvae are bound to calliphorin. In: Scheller K (ed) The larval serum pro- tein of insects: function-biosynthesis-genetic. Thieme, Stuttgart, New York, pp 40-49
Kaliafas AD, Marmaras VJ, Christodoulou C (1984) Immuno- chemical and electrophoretic studies on the localization of the major haemolymph proteins (ceratitins) in Ceratitis capitata during development. Wilhelm Roux's Arch Dev Biol 194:3743
K6nig M, Scheller K (1986) Influence of 20-OH-ecdysone on the incorporation of arylphorin in larval tracheae of Calliphora vicina. Zool Jb Physiol 90:65-76
K6nig M, Agrawal OP, Schenkel H, Scheller K (1986) Incorpora- tion of calliphorin into the cuticle of the developing blowfly, Calliphora vicina. Roux's Arch Dev Biol 195:296-301
Laemmli UK, Favre R (1973) Maturation of the head of bacterio- phage T 4. I. DNA packaging events. J Mol Biol 80 : 575-599
Lepesant JA, Kejzlarovfi-Lepesant J, Garen A (1978) Ecdysone- inducible functions of larval fat bodies in Drosophila. Proc Natl Acad Sci (USA) 75:5570-5574
Lepesant JA, Levine M, Garen A, LepesanVKejzlarov~, Rat L, Somme-Martin G (1982) Developmentally regulated gene ex- pression in Drosophila larval fat bodies. J Molec Appl Gen 1 : 371-383
Levenbook L (1985) Insect storage proteins. In: Kerkut GA, Gil- bert LI (eds) Comprehensive insect physiology, biochemistry and pharmacology, vol 4. Pergamon Press Oxford, pp 307-346
Mintzas AC, Chrysanthis G, Christodoulou C, Marmaras VJ (1983) Translation of the mRNAs coding for the major haemo- lymph proteins of Ceratitis capitata in cell-free system. Compar- ison of the translatable mRNA levels to the respective biosyn- thetic levels of the proteins in the fat body during development. Dev Biol 95:492-496
Patrinou-Georgoula M, Dimitriadis GJ, Sekeris CE (1983) The storage proteins of Dacus oleae: Characterization, correlation between developmental stage appearance and mRNA titer and approach to clone the genes. In: Scheller K (ed) The larval
serum proteins of insects : function-biosynthesis-genetic. Thieme, Stuttgart, New York, pp 61 74
Powell D, Sato JD, Brock HW, Roberts DB (1984) Regulation of synthesis of the larval serum proteins of Drosophila melano- gaster. Dev Biol 102:206-215
Price GM (1974) Protein metabolism by the salivary glands and other organs of the larva of the blowfly, Calliphora erythroce- phala. J Insect Physiol 20:329-347
Rigby PWJ, Dieckmann M, Rhodes C, Berg P (1977) Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J Mol Biol 113:237-251
Scheller K (ed) (1983) The larval serum protein of insects: function - biosynthesis - genetic. Thieme, Stuttgart, New York
Scheller K, Karlson P (1977) Synthesis of poly(A) containing RNA induced by ecdysterone in fat body cells of Calliphora vicina. J Insect Physiol 23:435440
Schenkel H, Scheller K (1983) Ecdeysteroids regulate the synthesis of RNA polymerase molecules in fat body cells of Calliphora larvae. Wilhelm Roux's Arch Dev Biot 192:152-158
Schenkel H, Myllek C, K6nig M, Hausberg P, Scheller K (1983) Calliphorin: Studies on its biosynthesis and function. In: Scheller K (ed) The larval serum proteins of insects: function- biosynthesis-genetic. Thieme, Stuttgart, New York, pp 18-39
Schenkel H, Kejzlarov~-Lepesant J, Berreur P, Moreau J, Scheller K, Br6g6g+re F, Lepesant JA (1985) Identification and molecu- lar analysis of a multigene family encoding calliphorin, the ma- jor larval serum protein of Calliphora vicina. The EMBO Jour- nal 4:2983 2990
Sekeris CE, Scheller K (1977) Calliphorin, a major protein of the blowfly. Correlation between the amount of protein, its biosyn- thesis, and the titre of translatable calliphorin-mRNA during development. Dev Biol 59:12-23
Tahara T, Maeda Y, Kuroiwa A, Ueno K, Obinata M, Natori S (1982) Identification of storage-protein messenger RNA of the fleshfly Sarcophaga peregrina. Biochem J 203:571-575
Walter J, Koolman J (1985) In: Balls M, Bownes M (eds) Ecdyster- oids in insects: how are their concentrations regulated? Claren- don Press, Oxford, pp 198-220
Received December 16, 1985 Accepted March 21, 1986