Embed Size (px)
Citation preview

J . Zool., Lond. (1993) 229, 153-161
Spring and summer food habits and habitat use of the European pine marten (Martes martes) on the island of Minorca, Spain
ANTHONY P. CLEVENGER*
Department of Forestry, Wildlife and Fisheries, The University of Tennessee, Knoxville TN 37901, USA
(Accepted 22 October 1991)
(With 1 figure in the text)
The food habits and habitat use of pine marten (Martes marre.? L.) on the Balearic Island of Minorca were studied from March-August 1990. In a 28.2 km2 area, a series ofhiking trails/l'orest roads were used to collect pine marten faeces hi-monthly and study the species' differential use of habitats. A total of28 different food items were identified in 723 faeces. Small mammals were the most important food overall, constituting 34% of the volume. During March-April, Small mammals were the principal food consumed (63% of volume), followed by birds (19"h). In May- June, birds were the main food (40%), then small mammals. Plant material and insects were the most important foods in July-August, both made up 68% ofthe pine marten diet. The abundance of certain foods and the degree of difficulty in obtaining them appear to be important factors that govern pine marten food selection in Minorca. Habitat use was studied along 12 routes totalling 32.6 km. Pine marten showed preference in March-April for Coastal Shrubland habitat, while during May-June they were found most in the Upland Pine type. During July-August, the Upland Pine and Open Pine habitats were most frequented by pine marten. According to bi- monthly food habits data, habitat use seems largely determined by the abundance and availability of their prey items in the available habitat types. Martens may concentrate hunting efforts in areas different from when they are not hunting. The Minorcan pine marten differs from continental populations in its wide use of shrublands, in addition to forests.
Contents Page
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153 Studyarea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154 Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154 Results and discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155
Food habitats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155 Habitatuse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16 1
Introduction
In the southern part of its range, the European pine marten (Martes rnartes L.) is found on six Mediterranean islands, including the Balearic Islands of Minorca and Majorca. These two Spanish islands are geographically isolated and have few species of mammals compared to continental Europe. The only other carnivores on Majorca are the genet (Genetta genettu) and
* Present address: Departamento de Biologia Animal, Universidad de Leon, 24071 Leon, Spain
153 1993 The Zoological Society of London

154 A. P . CLEVENGER
weasel (Mustela niualis), whereas in Minorca just the weasel is present. Small mamma1 species diversity is low on both islands, and microtine rodents, an important prey item throughout the species range, are absent (Alcover, 1988). Because of the lack of competitors and limited prey base, the island pine marten would be expected to exhibit differences in food habits and habitat selection, when compared to the continental pine marten populations.
A recent study of the species’ geographic variation on these two islands and the Iberian Peninsula, showed that cranial measurements of the Minorcan race were significantly larger than those of the continent, greater than any previously known population in its range, and is also characterized by a more robust dentition (Alcover et al., 1986). Virtually nothing is known about the species distribution or ecology on both of these islands other than some preliminary food habits data (Cheylan, 1984; Moreno, Rodriguez & Delibes, 1988). In this paper, there is information on the species’ spring and summer food habits and habitat use on the island of Minorca.
Study area
The island of Minorca is part of the Balearic Island group located in the western Mediterranean approximately 250 km from the Iberian Peninsula. The archipelago consists of 3 main islands, Majorca, Minorca and Ibiza, and 2 lesser islands, Formentera and Cabrera. Minorca is the northernmost of all, and the second largest measuring 69,000 ha. It is approximately 45 km long and 15 km wide.
The climate is humid mediterranean with moderate temperatures whose mean varies slightly during the year. The average temperature for January is 11 ”C and August, 28°C; average yearly rainfall is 220 mm. The island topography is gentle except for several deep river gorges in the south and some mountainous areas in the northern half. The highest point on the island rises 358 m above sea level.
The island vegetation is also mediterranean, consisting of Aleppo pine (Pinus halepensis) and holm oak (Quercus ilex) forests, coastal shrublands and pastureland. The pine and oak forests are fragmented throughout the island and make up 28% ofits area. Typical shrub species include wild olive (Olea europaea), lentisc (Pistacia lentiscus) and tree heather (Erica arborea), and form approximately 25% of the island area. The remaining half of the island consists of many subdivided grazing pastures used by dairy cows. Up until 30 years ago the pine forests were heavily exploited for the production of charcoal and wood fuel for lime extraction. Since these activities have ceased, the forests have recovered, and many are now difficult to enter due to the high and extremely dense understorey vegetation.
Methods
Between March and August, 1990, pine marten scats wcre collected once every 2 months along a series of 12 designated hiking trails and forest roads in a 28.2 km2 area known as ‘La Vall’. The area was selected because of its abundant pine marten population and its excellent representation of the island’s native vegetation and habitat types. The routes were designed to sample all of the habitats in proportion to their representation in the study area. Faeces that might have been confused with those of weasel or by remote chance, feral cats (Felis Iybica), were discarded. Procedures for faeces analysis followed the standard techniques of washing, separating and identifying of different food items (Korschgen, 1980). Mammalian and avian prey remains were identified by microscopic and macroscopic analysis of hair, teeth, bones and feathers (Day, 1966) and compared to a reference collection at the University of Leon. The identified food items were placed in 1 of 6

FOOD AND HABITAT USE OF EUROPEAN PINE MARTEN I55
food categories: Mammals, Birds, Reptiles, Insects, Other Invertebrates and Plant Material. An ocular estimate of percentage volume of each food item was made, and pooling data from all samples, the mean percentage volume and frequency of occurrence of each category of food was calculated.
Pine marten habitat use was determined by indirect methods. Along the 12 routes, stations were marked at 50-m intervals and the respective habitat type and understorey vegetation were classified. Faeces collected along the routes were referenced to the nearest station. Habitat types were categorized as Lowland Pine, Upland Pine, Open Pine, Holm Oak, Coastal Shrubland and Clearing/Forest Ecotone. Understorey vegetation density was subjectively determined for the forested habitats as being, High, Low or Moderate, and Height was categorized as either less than 2 m or greater than 2 m.
To quantify the availability of habitat types, the total number of stations in each were summed and the proportion of each was calculated. Usage was determined by the observed frequency of faeces located in each habitat type. In this paper, habitat preference was determined using the Bonferroni approach (Byers, Steinhorst & Krausman, 1984); selection for and against refers to significant differences at the 0.05 level, between use of a habitat type and its expected use based upon the area sampled.
Results and discussion
Food habits
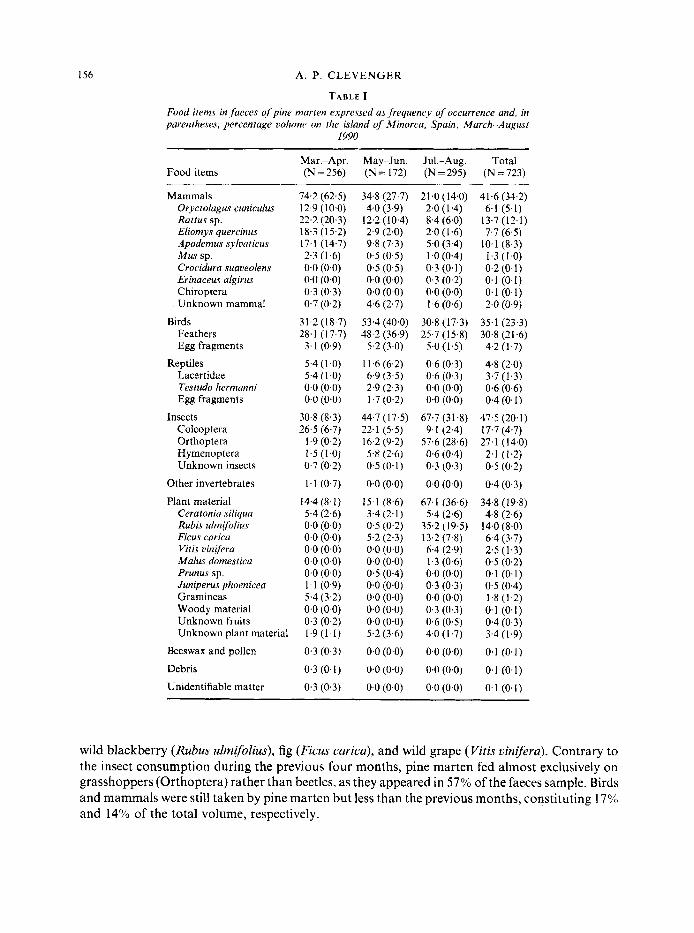
Pine marten in Minorca used a wide variety of foods during spring and summer as 28 different food items were identified in 723 faeces (Table I). Prey items ranged in size from beetles (Coleoptera) to rabbits (Oryetolagus cuniculus). Of noteworthy interest were the remains of a bat (unident. Chiroptera) in one spring faeces and mediterranean tortoise (Testudo hermanni) in five summer faeces; the latter were juveniles in four of the five cases. Mammals, birds, insects and plant material were the four main components of the pine marten diet during the period reported.
Mammals were the most important food during the six-month period, appearing in 41 % of the faeces and constituting 34% of the volume analysed (Fig. 1). Among them, rodents were the dominant food item. All six species of rodents found on the island appeared in the diet, the most frequently taken were the common and Norway rat (Rattus and R. norvegicus, respectively), followed by wood mice (Apodemus sylvaticus), and garden dormice (Eliomys quercinus). Birds, insects and plant material were commonly consumed food items also, occupying 2396, and the latter two categories, 20% of the analysed faeces volume, respectively. Reptiles and Other Invertebrates appeared in the diet in very low quantities throughout the six-month period.
During March and April, 256 faeces were analysed and mammals were by far the most preferred food, constituting 63% of the volume and appearing in 74% of the faeces. Of secondary importance were birds, appearing in 31% of the faeces and forming 19% of its volume. Plant material and insects both totalled 8% of the volume; in the former, grasses, carob fruit (Ceralonia siliqua) and juniper berries (Juniperus phoenicea) were most common, while beetles were the dominant insect eaten.
A change in food preference occurred during May and June when birds were the main food item, constituting 40% of the faeces volume and appearing in 53%) of the 172 analysed. Mammals and insects represented 28% and 18% of the volume, respectively, while plant material remained unchanged from the previous two months, constituting 9%. Reptiles increased from 1% to 6% of the faeces volume between the two bi-monthly periods. Lizards were preyed on most, followed by mediterranean tortoise. Lastly, during the months of July and August, plant material and insects were the main food items. Both appeared in 67% of the 295 faeces analysed and together constituted 68% of the total volume. The predominant plant foods in order of occurrence were,
156 A. P. CLEVENGER
TAHLE I Food items in faeces of pine marten expressed as ,frequency of occurrence and, in purentheses, percentage volume on the island of Minorca, Spain, March-August
I990
Mar.-Apr. May--Jun. JuL-Aug. Total Food items (N = 256) (N = 172) (N = 295) (N = 723)
Mammals Oryctolagus cuniculus Rattus sp. Eliomys quercinus Apodemus sylvuticus Mus sp. Crocidura suaveolens Erinaceus algirus Chiroptera Unknown mammal
Feathers Egg fragments
Lacertidae Testudo hermanni Egg fragments
Coleoptera Orthoptera Hymenoptera Unknown insects
Other invertebrates
Plant material Ceratonia siliqua Rubis ulrnifoliuu Ficus curica Vitis viniferu Malus dornestica Prunus sp. Juniperus phoenicea Gramineas Woody material Unknown fruits Unknown plant material
Beeswax and pollen
Debris
Unidentifiable matter
Birds
Reptiles
Insects
74.2 (62.5) 12.9 (10.0) 22.2 (20.3) 18.3 (1 5.2) 17.1 (14.7) 2.3 (1.6) 0.0 (0.0) 0.0 (0.0) 0.3 (0.3) 0.7 (0.2)
31.2 (18.7) 28.1 (17.7)
3.1 (0.9)
5.4 (1.0) 5.4 ( 1 .O) 0.0 (0.0) 0.0 (0.0)
30.8 (8.3) 26.5 (6.7)
1.9 (0.2) 1-5 (1.0) 0.7 (0.2)
1.1 (0.7)
14.4 (8.1) 5.4 (2.6) 0.0 (0.0) 0.0 (0.0) 0.0 (0.0) 0.0 (0.0) 0.0 (0.0) 1.1 (0.9) 5.4 (3.2) 0.0 (0-0) 0.3 (0.2) 1.9 (1-1)
0.3 (0.3)
0.3 (0. I ) 0.3 (0.3)
34.8 (27.7) 21.0 (14.0) 4.0 (3.9) 2.0 (1.4)
12.2 (10.4) 8.4 (6.0) 2.9 (2.0) 2.0 (1.6) 9.8 (7.3) 5.0 (3.4) 0.5 (0.5) 1.0 (0.4) 0.5 ( 0 3 ) 0.3 (0.1) 0.0 (0.0) 0.3 (0.2)
4.6 (2.7) 1.6 (0.6)
53.4 (40.0) 30.8 (17.3) 48.2 (36.9) 25.7 (15.8)
5.2 (3.0) 5.0 (1.5)
11.6 (6.2) 0.6 (0.3) 6.9 (3.5) 0.6 (0.3) 2.9 (2 3) 0.0 (0.0) 1.7 (0.2) 0.0 (0.0)
44.7 (17.5) 67.7 (31-8) 22.1 (5 .5 ) 9.1 (2.4) 16.2 (9.2) 57.6 (28.6) 5.8 (2.6) 0.6 (0.4) 0.5 (0.1) 0.3 (0.3)
0.0 (0.0) 0.0 (0.0)
0.0 (0.0) 0.0 (0.0)
15.1 (8.6) 67.1 (36.6) 3.4 (2.1) 5.4 (2.6) 0.5 (0.2) 35.2 (19.5) 5.2 (2.3) 13.2 (7.8) 0.0 (0.0) 6.4 (2.9) 0.0 (0.0) 1.3 (0.6) 0.5 (0.4) 0.0 (0.0) 0.0 (0.0) 0.3 (0.3)
0.0 (0.0) 0.3 (0.3) 0.0 (0.0) 0.6 (0.5) 5.2 (3.6) 4.0 (1.7)
0.0 (0.0) 0.0 (0.0)
0.0 (0.0) 0.0 (0.0)
0.0 (0.0) 0.0 (0.0)
0.0 (0.0) 0.0 (0.0)
41.6 (34.2) 6.1 (5.1)
13.7 (12.1) 7.7 (6.5)
10.1 (8.3) 1.3 (1.0) 0.2 (0.1) 0.1 (0.1) 0.1 (0.1) 2.0 (0.9)
35.1 (23.3) 30.8 (21.6) 4.2 (1.7)
4.8 (2.0) 3.7 (1.3) 0.6 (0.6) 0.4 (0.1)
47.5 (20.1) 17.7 (4.7) 27.1 (14.0)
2.1 (1.2) 0.5 (0.2)
0.4 (0.3)
34.8 (19.8) 4.8 (2.6)
14.0 (8.0) 6.4 (3.7) 2.5 (1.3) 0.5 (0.2)
0.5 (0.4) 0.1 (0.1)
1.8 (1.2) 0.1 (0.1) 0.4 (0.3) 3.4 (1.9)
0.1 (0.1)
0.1 (0.1)
0.1 (0.1)
wild blackberry (Rubus ulmifolius), fig (Ficus curica), and wild grape (Vitis vinifera). Contrary to the insect consumption during the previous four months, pine marten fed almost exclusively on grasshoppers (Orthoptera) rather than beetles, as they appeared in 57% of the faeces sample. Birds and mammals were still taken by pine marten but less than the previous months, constituting 17% and 14% of the total volume, respectively.
FOOD AND HABITAT USE OF EUROPEAN PINE MARTEN 157
EX@ Birds Reptiles Mammals EZZ4 Insects Plant mat. 0 Other
' O 0 1 80
60 a - 5 p 40
20
Total 0 Mar.-Apr. May-Jun.
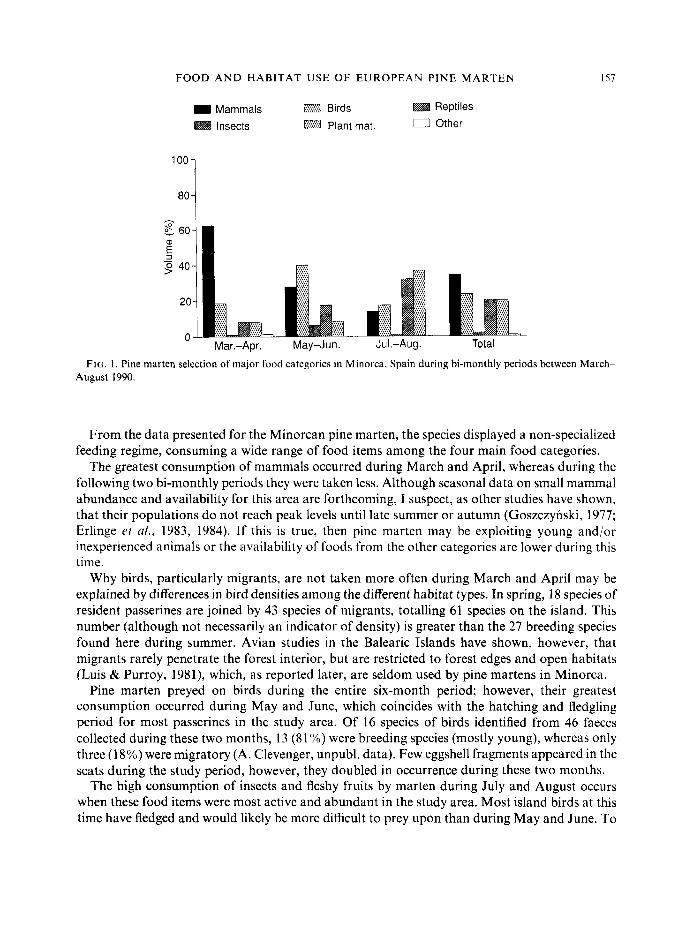
FIG. 1. Pine marten selection of major food categories in Minorca, Spain during bi-monthly periods between March- August 1990.
From the data presented for the Minorcan pine marten, the species displayed a non-specialized feeding regime, consuming a wide range of food items among the four main food categories.
The greatest consumption of mammals occurred during March and April, whereas during the following two bi-monthly periods they were taken less. Although seasonal data on small mammal abundance and availability for this area are forthcoming, I suspect, as other studies have shown, that their populations do not reach peak levels until late summer or autumn (Goszczynski, 1977; Erlinge et ul., 1983, 1984). If this is true, then pine marten may be exploiting young and/or inexperienced animals or the availability of foods from the other categories are lower during this time.
Why birds, particularly migrants, are not taken more often during March and April may be explained by differences in bird densities among the different habitat types. In spring, I8 species of resident passerines are joined by 43 species of migrants, totalling 61 species on the island. This number (although not necessarily an indicator of density) is greater than the 27 breeding species found here during summer. Avian studies in the Balearic Islands have shown, however, that migrants rarely penetrate the forest interior, but are restricted to forest edges and open habitats (Luis & Purroy, 1981), which, as reported later, are seldom used by pine martens in Minorca.
Pine marten preyed on birds during the entire six-month period; however, their greatest consumption occurred during May and June, which coincides with the hatching and fledgling period for most passerines in the study area. Of 16 species of birds identified from 46 faeces collected during these two months, 13 (8 1 %) were breeding species (mostly young), whereas only three (18%) were migratory (A. Clevenger, unpubl. data). Few eggshell fragments appeared in the scats during the study period, however, they doubled in occurrence during these two months.
The high consumption of insects and fleshy fruits by marten during July and August occurs when these food items were most active and abundant in the study area. Most island birds at this time have fledged and would likely be more difficult to prey upon than during May and June. To

I58 A . P. CLEVENGER
maximize foraging efficiency, pine marten appear to select the most abundant and easily acquired foods these months.
Pine marten concentrated on food items that were seasonally abundant, as noted by the high consumption of birds during May and June, and insects and fruit during July and August. Consistent with other European pine marten studies (Lockie, 1961 ; Nyholm, 1970; Pulliainen, 1981~; Marchesi & Mermod, 1989), the Minorcan species is a generalistic feeder, however, there was a more balanced use of the available foods in Minorca than has been reported for other populations.
The Minorcan race has been strongly differentiated from the Iberian Peninsula pine marten population in cranial morphology, displaying a broader skull, larger jaw and heavier dentition (Alcover et al., 1986). A similar situation exists in Canada for the North American pine marten ( M . americana) of the Queen Charlotte Islands when compared to the coastal populations (Foster, 1965; Giannico & Nagorsen, 1989). Researchers there were not able to detect anything in the winter diet that would account for the differences in cranial morphology (Nagorsen, Campbell & Giannico, 199 l), however, they indicated that their results are speculative until year-round food habits data are collected.
If the morphological traits reflect selection pressures associated with the size or hardness of prey, as has been hypothesized by Foster (1965) for the Queen Charlotte population, there appears to be nothing in the present diet of the Minorcan pine marten that would account for the observed morphological differences. Nevertheless, historically two prey species, the rabbit and mediterra- nean tortoise, were more abundant on Minorca than at present. Both species were once abundant but have declined in numbers due to disease and human removal. Whether these were important prey items for marten in the past, and resulted in selection for a large size, cannot be proven, but they certainly could have played an important role.
Habitat use
To study habitat use, 652 reference stations were established among the 12 routes, totalling 32.6 km. The average route distance was 2.7 km and ranged from 0.9 to 4.3 km. Between 1 I April and 12 August 1990, a total of 726 faeces were collected along the routes and were used to analyse the differential use of habitats in the study area.
During the six-month period, 37% of the faeces were located in Lowland Pine, 22% in Open Pine, 15% in Upland Pine, and 12% in Holm Oak habitat types (Table IIa). Only 5%) and 6% of the
TABLE I Ia Pina marlen hahitar U S E in Minarca during March--Augusi, 195’0
Understorey
Type Density Height
Lowland Pine Moderate 1 2 m Upland Pine Moderate < 2 m Open Pine High > 2 m Holm Oak Low < 2 in Coastal Shrubland ~ - Clearing/Forest Ecotone - -
Pct avail. (N) Pct. use ( N ) Preference*
0450 (297) 0.377 (274) 0.060 (36) 0.151 (110) + 0,210 (137) 0.224 (163) 0 0.140 (94) 0.122 (89) 0 0,040 (23) 0,056 (41) 0 0.100 (65) 0,067 (49) -
652 726
FOOD AND HABITAT USE OF EUROPEAN PINE MARTEN 159
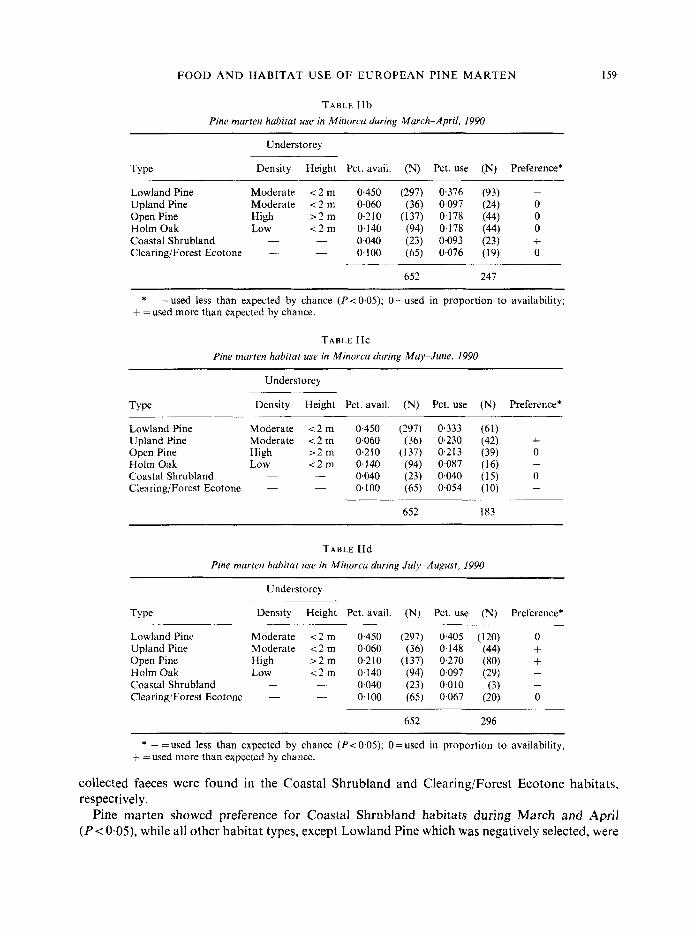
TABLE I lb Pine marten habitat use in Minorca during March-April. 1990
Understorey
TY Pe Density Height Pct. avail. (N) Pct. use (N) Preference*
Lowland Pine Moderate 1 2 m 0,450 (297) 0.376 (93) - Upland Pine Moderate 1 2 m 0.060 (36) 0,097 (24) 0 Open Pine High r 2 m 0.210 (137) 0.178 (44) 0 Holm Oak Low < 2 m 0,140 (94) 0.178 (44) 0 Coastal Shrubland - - 0.040 (23) 0.093 (23) + Clearing/Forest Ecotone - - 0.100 (65) 0.076 (19) 0
652 247 ~~ ~ ~ ~~ ~
* - =used less than expected by chance (P<O.O5), O=used in proportion to availability; + =used more than expected by chance.
TABLE IIc Pine marten habirut use in Minorca during May-June, 1990
Understorey
Type Density Height Pct. avail. (N) Pct. use (N) Preference* ~
Lowland Pine Moderate 1 2 m 0.450 (297) 0.333 (61) - Upland Pine Moderate t 2 m 0,060 (36) 0,230 (42) + Open Pine High > 2 m 0.210 (137) 0.213 (39) 0 Holm Oak Low < 2 m 0.140 (94) 0.087 (16) - Coastal Shrubland __ - 0,040 (23) 0,040 (15) 0 Clearing/Forest Ecotone - - 0.100 (65) 0.054 (10) -
652 183
TABLE IId Pine marten habitat use in Minorca during July-August, 1990
Understorey
Type Density Height Pct. avail. (N) Pct. use (N) Preference*
Lowland Pine Moderate < 2 m 0.450 (297) 0.405 (120) 0 Upland Pine Moderate 1 2 m 0,060 (36) 0,148 (44) + Open Pine High 2 2 m 0.210 (137) 0,270 (80) + Holm Oak Low < 2 m 0.140 (94) 0.097 (29) - Coastal Shrubland - - 0,040 (23) 0.010 (3) - Clearing/Forest Ecotone - - 0.100 (65) 0.067 (20) 0
652 296
* - =used less than expected by chance (P< 0.05); O=used in proportion to availability; + =used more than expected by chance.
collected faeces were found in the Coastal Shrubland and Clearing/Forest Ecotone habitats, respectively.
Pine marten showed preference for Coastal Shrubland habitats during March and April ( P < O*OS), while all other habitat types, except Lowland Pine which was negatively selected, were

160 A . P. CLEVENGER
used without any detectable preference (Table IIb). Food habits data during this period showed a high consumption of mammals; however, preliminary small mammal trapping data has indicated highest abundance in the Open Pine habitat type (A. Clevenger, unpubl. data). Pine marten may be found using the Coastal habitat owing to the presence of breeding herring gulls (Larus argentatus). They nest earlier than the forest species in Minorcd, with hatching taking place the second week of April (Muntaner & Congost, 1984). Rat populations are generally high around nesting gull colonies also and may explain the greater use of this habitat. Of six species of birds identified in 11 March-April faeces, all live in the Coastal Shrubland and, to a lesser extent, Open Pine habitats (A. Clevenger, unpubl. data).
During May and June, pine marten showed a strong preference for the Upland Pine habitat (P<O*O5), whereas the Open Pine and Coastal Shrubland were used in proportion to their availability (Table TIC). The other three habitats were used less than expected (P > 0.05). The high incidence of birds in the faeces remains during this period, together with the increased pine marten activity in the Upland Pine forest, suggests that they may be resting in this forest type but preying on breeding bird populations in the Coastal Shrubland and Open Pine habitat types. Of 16 species of birds identified from 46 May-June faeces, only one (Luscinia megarhynchos) can be considered a forest species, while the rest are shrubland inhabitants (A. Clevenger, unpubl. data).
Two habitats were preferred by martens during July and August; the Upland Pine and Open Pine types (P < 0.05), while the Lowland Pine and Clearing/Forest Ecotone were used less than expected (P>O.O5) (Table TId). The main foods during this period were grasshoppers and fleshy fruits, primarily blackberries and figs. These food items are all typical of shrublands, open habitats and forest ecotones, which could suggest that marten may be obtaining food in these habitats but when not hunting return to the pine forest.
In this analysis, usage was determined by the observed frequency of faeces located in each habitat type. This method assumes that the number of faeces in a given habitat reflects its proportional use by pine marten. It does not account for errant faeces marking behaviour in some individuals, but assumes that the rate of marking is constant among all individuals, which has not been proven. Studies have shown that European pine marten are intrasexually territorial (Storch, 1988; Marchesi, 1989; Kruger, 1990), and recently in Minorca this has been verified by radiotelemetry (A. Clevenger, unpubl. data). Therefore, the number of pine marten in a given habitat should be determined by the quality of that habitat (Buskirk & MacDonald, 1989).
The results indicate that pine marten habitat use in Minorca differs from continental Europe, where the species has been reported as being largely restricted to the Eurosiberian forests (Corbet & Southern, 1977; Pulliainen, 19816; Fedyk et al., 1984; Ruiz-Olmo, Parellada & Porta, 1988; Marchesi, 1989). In Minorca, coastal shrublands and open areas form an integral part of the species habitat in addition to forested areas, which is not common among continental populations. The fact that no fossil record exists for the stone marten (M.foina) or pine marten in the Balearic Islands, and it is believed that they were introduced during recent times (Alcover, 1982), the species appears to occupy the typical habitat and food niche of the stone marten. The lack of competition and relatively unrestricted use of food resources on the island may be some of the factors that have resulted in the Minorcan pine marten’s large size.
The majority of the funding for this research came from grants from the Institut Menorqui d’Estudis (IME) and the Gobierno Balearxonselleria d’Agricultura i Pesca. I wish to thank C. Campos, M. A. Campos, A. Onrubia and R. Triay for their enthusiastic help in the field and faeces analysis, J . May01 for logistical support, and R. Squella for permission to conduct this study in La Vall. J. and A. Teinturier kindly allowed me to use their house while in Minorca.

FOOD A N D HABITAT USE O F EUROPEAN PINE MARTEN 161
REFERENCES
Alcover, J. A . (1982). Note on the origin of the present mammalian fauna from the Balearic and Pityusics Islands, Miscnea.
Alcover, J. A. (1988). Els mamifers de les Balears. (2nd edn). Palma de Mallorca: Editorial Moll. Alcover, J. A,, Delibes, M., Gosalbez, M. & Nadal, J. (1986). Martes martes Linnaeus, 1758 ales Balears. Miscnea. zool.
Buskirk, S. W. & MacDonald, L. L. (1989). Analysis of varbdbilily in home-range size of the American marten. J. Wildl.
Byers, C. R., Steinhorst, R. K. & Krausman. P. R. (1984). Clarification ofa technique for analysis ofutilization-availability
Cheylan, G. (1984). Note sur I'alimentation de Martes murtes a Menorca (Balears). Dogma Acta vert. 11: 344-346. Corbet, G. B. & Southern, H. N. (1977). The handbook qfBritish mammals. (2nd edn). Oxford: Blackwell Scientific Publs. Day, M. G. (1966). Identification ofhair and feather remains in the gut and faeces of stoats and weasels. J . Zool., Lond. 148
201 -21 7. Erlinge, S., Goransson, G., Hansson, L., Hogstedt, G., Liberg, 0.. Nilsson, 1. N., Nilsson, T., von Schantz,T. &Sylven, M.
(1983). Predation as a regulating factor on small rodent populations in southern Sweden. Oikos 4 0 36-52. Erlinge, S., Goransson, G., Hogstedt, G., Jansson. G., Liberg, O., Loman, J., Nilsson, I. N., von Schantz, T. & Sylven, M.
(1984). Can vertebrate predators regulate their prey? Am. Nut. 123 125-133. Fedyk, S., Gebczynska, Z., Pucek, M., Raczynski, J. & Sikorski, M. D. (1984). Winter penetration by mammals ofdifferent
habitats in the Biebrza valley. Acta theriol. 2 9 317-336. Foster. 3. B. (1965). The evolulion of the mammals of the Queen Charlotte Islands, British Columbia. Om. Pap. Br.
Columh. prou. Mus. No. 14: 1-130. Giannico, G . R. & Nagorsen, D. W. (1989). Geographic and sexual variation in the skull of Pacific coast marten (Martes
americana). Can. J . Zool. 67: 1386- 1393. Goszczyhki, J. (1977). Connections between predatory birds and mammals and their prey. Acta theriol. 2 2 399--430. Korschgen, L. J. (1980). Procedures for food habits analyses. In Wildlife management techniques manual 113-127.
Kruger, H:H. (1990). Home ranges and patterns of distribution of stone and pine martens. Trans. int. Un. Game Bid.
Lockie, J. D. (1961). The food of the pine marten (Martes martes) in west Ross-shire Scotland. Proc. zool. Soc. Lond. 136
Luis, E. & Purroy, F. J. (1981). Evolucibn estacional de las comunidades de aves en la Isla de Cabrera (Baleares). Studia
Marchesi, P. (1989). Ecology of the pine marten (Martes martes L.) in the Swiss Jura Mountains. PhD diss., University of
Miirchcsi, P. & Mermod, C. (1989). Regime alimentaire de la martre (Marles murtes L.) dans le Jura suisse (Mammalia:
Moreno. S., Rodriguez, A. & Delibes, M. (1988). Summer foods of the pine marten (Martes martes) in Majorcd and
Muntancr. J. & Congost, J. (1984). Avifauna de Menorca. Treb. Mus. Zoo/. Barcelona No. 1: 1-205. (English summary.) Nagorsen, D. W., Campbell, R. W. & Giannico, G. R. (1991). Winter food habits of marten (Martes americana) on the
Nyholm, E. (1970). On the ecology of the pine marten (Martesmartes) in Easterh and Northern Finland. Suomen Riista 2 2
Pulliainen, E. (1981~). Food and feeding habits of the pine marten in Finnish Forest Lapland in winter. Proc. Wldwide
Pulliainen, E. (1981b). Winter habitat selection, home range and movements of the pine marten (Martes martes) in a
Ruiz-Olmo, J., Parelbada, X. & Porta, J. (1988). Sobre la distribucih y el habitat de la marta (Martes martes, L., 1758) en
Storch, 1. ( 1 988). [Home range utilization by pine martens.]. Z . Jagdwiss. 34 115-1 19. [In German with English summary.]
zoo/. Barcelona 6 141 .149.
Barcelona 10 323-333.
Mgmt 53 997-1004.
data. J . Wildl. Mgmt 48: 1050-1053.
Schemnitz, D. (Ed.). Washington, D.C.: The Wildlife Society.
Congress 19 348--349.
187-195.
Oecolhgia 1: 181-223.
Neuchatel.
Mustelidae). Reu. suisse Zool. 9 6 127-146. (English summary.)
Minorca, Balearic Islands. Mammalia 5 2 289-291.
Queen Charlotte Islands. Can. Fld Nut. 105: 55-59.
105 118.
Furhearer Conf. 1: 580-598.
Finnish Lapland forest. Pror. Wldwide Furhearer Conf. 1: 1068-1087.
Cataluria. Pirineos No. 131: 85-94. (English summary.)



















![The mammalian fauna from the Central Himalaya, Nepal3023...Beer Ursus thibetanus G.[Baron] Cuvier, 1823 MCA, SNP, KCA Forest 2200- 3200 Mustelidae Yellow- throated Marten Martes flavigula](https://img.pdfslide.us/doc/110x75/5ff21df818b8563d93274b23/the-mammalian-fauna-from-the-central-himalaya-nepal-3023-beer-ursus-thibetanus.jpg)









