Embed Size (px)
Citation preview

This article was downloaded by: [University of Cambridge]On: 08 October 2014, At: 02:19Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Sponges from bathyal depths(1000–1750 m) in the WesternMediterranean SeaMaria-Jesus Uriz a & Dolors Rosell aa Centre d'Estudis Avançats de Blanes , C.S.I.C. Camí de Sta.Bàrbara , 17300, Blanes (Girona), SpainPublished online: 17 Feb 2007.
To cite this article: Maria-Jesus Uriz & Dolors Rosell (1990) Sponges from bathyal depths(1000–1750 m) in the Western Mediterranean Sea, Journal of Natural History, 24:2, 373-391, DOI:10.1080/00222939000770281
To link to this article: http://dx.doi.org/10.1080/00222939000770281
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms& Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

JOURNAL OF NATURAL HISTORY, 1990, 24, 373-391
Sponges from bathyal depths (1000-1750 m) in the Western Mediterranean Sea
MARIA-JESUS URIZ and DOLORS ROSELL
Centre d'Estudis Avanfats de Blanes, C.S.I.C. Cami de Sta. Bdlrbara s/n 17300 Blanes (Girona), Spain
(Accepted 16 October 1989)
This note deals with a study of sponge fauna at seven deep stations (1000-1750 m) in the Western Mediterranean Sea. The species found exhibit a broad geographical distribution, mainly centred in the Northern Atlantic Ocean. Trichostemma sarsi and Hyalonema thomsoni are recorded for the first time in the Mediterranean Sea, and the known bathymetric distribution of Polymastia tissieri has been considerably enlarged. Some taxonomic problems involving the genera Trichostemma, Radiella and Polymastia and the species Hyalonema thomsoni and H. infundibulum are discussed, and arguments supporting the reclassification of the species Radiella tissieri under the genus Polymastia are put forward.
KEYWORDS: Systematics, Demospongiae, Hexactinellida, Bathyal depths, Western Mediterranean.
Introduction Recent research carried out in the deepest regions of the Mediterranean Sea have
considerably improved our general knowledge of the fauna inhabiting such depth (Fredj and Laubier, 1985). However, little progress has been made in our knowledge of certain taxonomic groups, such as Porifera.
The most complete recent work on relatively deep-water Mediterranean sponges was by Vacelet (1969), who mainly studied the sponge fauna from the biocoenoses of rocky bottoms and deep corals on the continental shelf. However, few samples were taken from the bathyal muds, and the maximum depth explored was 800 m. For this reason, the deep-water samples made during the 'Batimar' and 'Bathos-2' cruises, though few in number, acquire interest. Because of the relatively high number of specimens and the characteristic low diversity of these bottoms, the few sponge species found can be considered representative.
Material and methods The material on which this study is based has been deposited in the 'Centre
d'Estudis Avanfats de Blanes (C.S.I.C.)', Spain.
Sampling stations B A T I M A R Cruise. Station 1. 40°57"54'N, 2°04"44'E, 1020-1040m, bottom of
argillaceous mud. Most abundant fauna (in terms of biomass): Trachyrhynchus trachyrhynchus (Risso), Lepidion aques (Risso), M ora mora (Risso) and Phycis blenioides (Briinn) (Rucabado, Del Cerro and Merrett, personal communication) were the most
0022-2933/90 $5-00 © 1990 Taylor & Francis Ltd.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

374 Mafia-Jesus Uriz and Dolors Rosell
abundant fish species; Mesothuria intestinalis (Ascanius) (AlvA, personal communication); Parahomola cuvieri (Risso) and Crustacea in general were the most abundant macroinvertebrate species.
Station 2. 40°54'N, 2°0.8'E, 1220-1277m, bottom of argillaceous mud. Most abundant fauna (in terms of biomass): Lepidion aques (Risso), Polyacanthonotus rissoanus (De Filippi-Vtrany), N othacanthus bonapartei Risso, Alepocephalus rostratus Risso, and Trachyrhychus trachyrhychus (Risso) (Rucabado et al., personal communication) were the most abundant fish species; Mesothuria intestinalis (Ascanius) (AlvA, personal communication), Aristeus antenatus (Risso) and other Crustacea. IsideUa elongata (Esper) and some Hexacorallia, were the most abundant macroinvertebrate species.
Station 3. 40°47-7'N, 1°50.4'E, 1286-1322m, bottom of argillaceous mud. Most abundant fauna (in terms of biomass): Lepidion lepidion (Risso), Polyacanthonotus rissoanus (De Filippi-Vtrany), Alepocephalus rostratus Risso, Bathypterois mediterraneus Bauchot, Coelorhynchus occa (Goode and Beam) (Rucabado et al., personal communication) were the most abundant fish species; Mesothuria intestinalis (Ascanius) (AlvA, personal communication), lsidella elongata (Esper) and Hexacorallia on plastic materials, Brachiopoda and Decapoda were the most abundant macroinvertebrate species.
Station 4. 40 ° 31-4'N, 1 °46"5'E, 1545-1580 m, bottom of argillaceous mud and rocks. Most abundant fauna (in terms of biomass): Alepocephalus rostratus Risso, Bathypterois mediterraneus Bauchot, and Polyacanthonotus rissoanus (De Filippi- Vtrany) (Rucabado et al., personal communication) were the most abundant fish species; Mesothuria intestinalis (Ascanius) (AlvA, personal communication), Isidella elongata (Esper), some Hexacorallia and Decapoda were the most abundant macroinvertebrate species.
BATTHOS-2 Cruise. Station PM-5. 40°20.9'N, l°53-YE, 1716--1750m, bottom of argillaceous mud with some isolated hard substrata.
Station PM-6. 40°53.1'N, 2°2'E, 1267-1329m, argillaceous mud. Station PM-8. 41°7.YN, 2°35-2'E, 1277-1356m, argillaceous mud. The bottom temperature was constant about 13°C. The samples were collected with a special bottom trawl designed to catch
deep-water vagile fauna (Merrett and Marshall, 1981).
Systematic descriptions Class tlexactinellida
Order Amplud~scopho~da Family Hyalonematidae Schulze
Hyalonema thomsoni (Marshall, 1875)
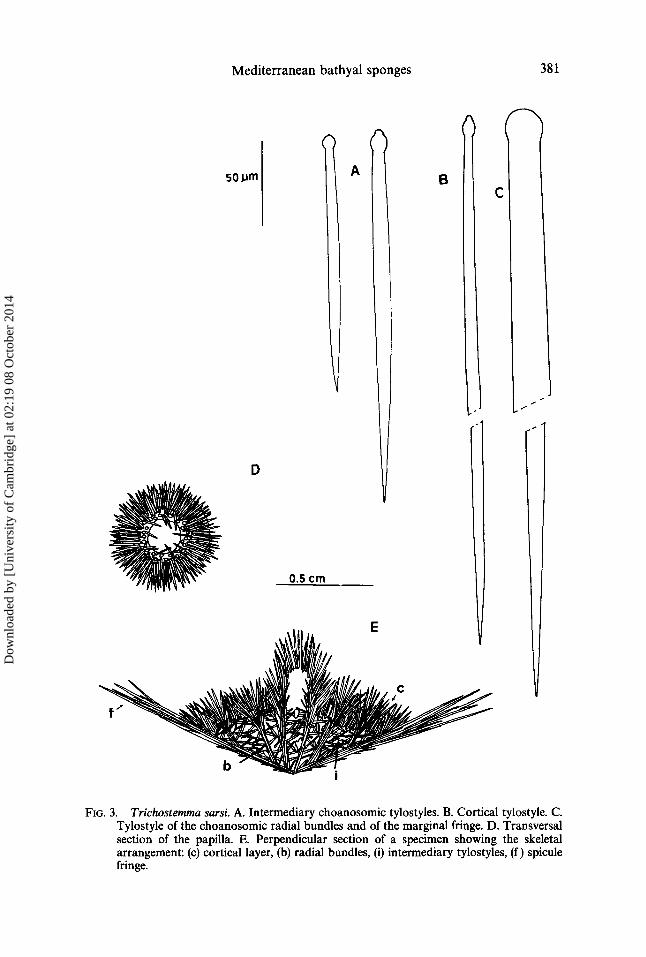
(Figs 1, 5A-D)
Occurrence. Station 4, 1 specimen (Uriz and Rosell, 1986). Station PM-5, 44 specimens.
Morphology Well preserved specimens, 15 cm maximum total length, 4-5 cm length of sponge
body and the rest corresponding to the spicular basal tufts, which probably were longer because they appeared to be broken.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 375
50jJm
"/
+o
Fro. 1. Spicules of Hyalonema thomsoni. A. Smooth diact. B. Smooth pentact. C. Microhexact. D. Authodermal pinnule. E. Medium sized oxyhexacts. F. Spined diact and pentact of the basal pad. G. Amphidiscs I with short umbels. H. Amphidiscs II with long umbels. I. Developing amphidiscs. J. Tetracts of the basal pad. K. Hexacts and pentact of the basal pad.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

376 Maria-Jesus Uriz and Dolors Rosell
Cylindroconical and compressed in shape: the distal part is concave with a very pronounced coniform central process. This cone divides the gastral cavity into four sections and it is supported by a bundle of long spicules traversing the sponge longitudinally and also making up the basal tuft. Uneven surface perforated by numerous orifices of varying size up 2 mm in diameter (parietal oscula?), with the ectosome difficult to remove from the choanosome. Uniform, clean, and evenly hispid gastral cavity. Sponge soft, fragile and easy to tear, greyish brown in colour, masked by mud.
Skeleton Diacts. 700-1530/~m x 8-10 #m, probably larger (most viewed on the slides were
broken), straight or slightly bent with an enlargement off-centre. Generally smooth, except those from the basal pad which have spined ends.
Pentacts and hexacts. Smooth except those from the base which have prickly rays with actines quite variable in size, measuring 180-310/tm x 6-9 #m.
Microhexacts. With rough, straight and somewhat uneven rays, 70-90/~m in diameter.
Medium sized oxyhexacts. Rough with straight or curved actines, measuring 160-195 #m at the point of greatest diameter.
Autodermal pinnules. Paired rays rough, ending abruptly in a point, measuring 22-25 #m x 2.5-4#m; odd ray longer, 100-200#m x 2-5-4 #m in size, with a bushy appearance.
Amphidiscs I. (Macramphidiscs) 120-225/~m x 6-10#m, with a short nearly hemispherical umbel 21-30 #m in length, bearing 4-7 teeth, making up ~ of the total spicule length. Echinated shaft, particularly at the centre where it often has verticillated spines. Small spicules of similar morphology, probably corresponding to developing forms, are also found.
Amphidiscs II. (Mesamphidiscs): 32-108 #m x 2-3/~m, with a bell-shaped umbel making up ½ of the total length, bearing 5-8 teeth. Developing spicules can also be found.
Basal hexacts. Completely echinated, with one axis longer than the other two, 95-148#m x 8-9#m and 68-75/tm x 8-9#m, respectively (thickness not including spines).
Basal tetracts. Very strong, roughly echinated, with one longer axis, 106-120 ~m x 10-11 #m (not including spines), and the other axis practically reduced to a profuse
bundle of large spines, 40-55 #mx 11-16 #m. The ends of the long spicules of the basal tuft were always broken off, so that no
anchors, for attaching the sponge to the substrate, were visible.
Distribution Northern Atlantic Ocean: Azores (Schulze, 1887; Topsent, 1892, 1904), north of the
Shetland Islands (Marshall, 1875), Western Hebrides (Schulze, 1887), Bay of Biscay, and Spanish Bay (35°32'N, 7°7'W) (Schulze, 1887). Species typically dwelling on muddy bottoms with broken shells and Globigerina ooze, from 975m to 1732m in depth.
New record of this genus in the Mediterranean Sea. Taking into account the previous compilation of records by Zibrowius (1985) the number of known species of Hexactinellida in this Sea is at present eight.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 377
Discussion The spicules of this species differ hardly at all from the spicules of the species
H. infundibulum Topsent, 1896. The shape of the upper part of the sponge (with or without a coniform process, respectively) is the sole conspicuous feature allowing separation of the two species. The authors agree with Topsent (1904) and Arnesen (1920) regarding the possibility that the two species are synonymous. The known records of H. infundibulum seem to indicate for this species a geographical and bathymetric distribution (North Atlantic 46°28'N, 7°W (Topsent, 1896) Bay of Biscay, Azores (Topsent, 1904) and Spanish Bay (Arnesen, 1920) (1215-1710 m)) very close to that of H. thomsoni.
Efforts to divide the genus Hyalonema have been made by different authors (Lendenfeld, 1915; Ijima, 1927; Lrvi, 1964) because of the great number of known species (95) currently allocated to it. Following these authors, H. thomsoni and H. infundibulum would be transferred to the genus (or subgenus) Cyliconema. As the differences presently used to separate the species of Hyalonema are inconsistent (variations in the pinnule and amphidiscs form), we prefer presently to maintain the old genus until new information indicates how it should besplit.
Class Demospongiae Order Astrophorida
Family Theneidae SoUas, 1888 Thenea muricata (Bowerbank, 1858)
Synonymy. Thenea sehmidti Sollas, 1888.
Occurrence. Station 1, 1 specimen; Station 2, 1 specimen; Station 3, 1 specimen.
Morphology Small, ovoid specimens 6-7 mm in length, with the lateral porous area and the
apical oscula typical of the species deafly visible. The main differences with regard to Mediterranean specimens from shallower depths (Uriz, 1981) were the long, straight and slender anchoring spiculate rootlets and the absence of an equatorial recess.
Skeleton Dichotriaenes. With a straight or curved rhabdome (1150-1500 #m x 25-38/~m in
size) and deuterocladi 380-650/~mx 14-31 #m, longer and more slender than the protodadi (70-110 gm × 15-38 gm in size).
Protriaenes. Very thin, with a long straight rhabdome 360-400 #m x 12-28/~m and cladi 80-95 #mx 10-11 gm; not abundant.
Oxeas. Long and relatively thin, straight, curved, or even flexuous, some are transformed into styles.
Anatriaenes. Present in the anchoring rootlets, with a sinuous rhabdome of up to 1800 pm x 11/~m in size, broad at the base and tapering to a very fine point at the end and with cladi measuring up to 45 gm x 10 pm.
Metasters. With 3-5 straight actines of 50-80/~m x 2-4 #m. Plesiasters. With actines measuring 22-25 #m x 2 gm. Spirasters. Very abundant, measuring 19-23 #m at the point of great diameter.
Distribution Arctic and Atlantic Oceans and Mediterranean Sea. Typically dwelling on muddy
and sandy bottoms, mainly from 100 to 4000 m deep. Also cited from littoral caves (Russ and Ruetzler, 1959; Sara, 1964 a; Ruetzler, 1966).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

378 Maria-Jesus Uriz and Dolors Rosell
Order Hadromerida Family Polymastiidae Gray, 1867 Polymastia tissieri (Vacelet, 1961)
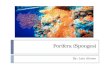
(Fig. 2)
Synonymy. Radiella tissieri Vacelet, 1961.
Occurrence. Station 1, 3 specimens; Station 2, 17 specimens; Station 3, 17 specimens; Station 4, 14 specimens. Station PM-5, 44 specimens; Station PM-6, 7 specimens; Station PM-8, 30 specimens.
Morphology Specimens 0.4-2 cm in diameter and 0-2q3.9 cm thick, hemispherical to flattened in
shape; in general with a convex and minutely hispid upper zone bearing a single small central papilla (usually smaller than 1 mm high) and a lower zone varying in width, exhibiting long hispidity. In some of the specimens this area of a long hispidity was entirely absent, while in others it reached the papillae: this variability was not mentioned by Vacelet (1961). The colour is yellowish white, masked at the bottom by sediment.
These sponges are incrusted not only with mud at the base, as Vacelet (1961) described, but also with small pieces of shells and pebbles, not always completely enclosed by tissue.
Skeleton Cortical layer made up of an outer palisade of small tylostyles and an inner
tangential layer of intermediary tylostyles; these last spicules are also present in the choanosome, exhibiting a rather disarranged disposition.
Radial choanosomic bundles of long tylostyles only pass through the cortical layer at the lower zone of the sponge and cause its long hispidity.
Papilla typical of the genus Polymastia, exhibiting both inhalant and exhalant functions. The inhalant canals are peripheral and the exhalant canal is central, echinated by intermediate tylostyles. Its skeleton shows from outside to inside the same structure of the rest of the sponge: palisade layer, thin tangential layer and longitudinal bundles of long tylostyles densely packed around the exhalant canal.
As a whole, the skeletal arrangement of this species is reminiscent ofP. infrapilosa as redescribed by Boury-Esnault (1987).
Cortical tylostyles. With a pronounced, sometimes trilobate tyle, 95-227 #m x 1- 5 #m in size. Arranged in palisade, they make up the cortex.
Tylostyles of the subcortical tangential layer. With a straight or slightly bent shaft and a well-formed tyle, 260-740 #m x 6-10 #m in size. They are also scattered in the choanosome (intermediary tylostyles).
Tylostyles of the choanosomic radial bundles. 800-2030 #m x 12-22/~m.
Distribution Abundant in the Mediterranean Sea, typically dwelling on epibathyal muds
between 427 and 640m (Vacelet, 1961) (N.B.: the geographical distribution of this species is surely wider than known records indicate. Some unclassified specimens from the northern Atlantic Ocean (exact locality unknown) have been located by one of the authors in the Smithsonian Institution's sponge collection).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014
Mediterranean bathyal sponges 379
A
\
B
1 cm
D E
50 pm
0.5 cm
} G
s
FIG. 2. Polymastia tissieri. A. Tylostyles of the choanosomic bundles. B. Tylostyles of the subcortical tangential layer. C. Cortical tylostyles. D. Schematic section of one specimen showing the substrate (mud and pebbles) completely enclosed. E. Specimens enclosing partially the substrate. F. Transversal section of the papilla. G. Perpendicular section of a specimen showing the skeletal arrangement: (c) cortical layer, (t) tangential layer, (b) radial bundles, (i) intermediary tylostyles, (s) substratum.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

380 Maria-Jesus Uriz and Dolors Rosell
Discussion The genus Polymastia Bowerbank, as redescribed by Ridley and Dendy (1887) and
by Topsent (1900) differs from the genus Trichostemma (=Radiella) mainly in its asymmetrical shape, its lack of a spicule fringe, the aquiferous and skeletal organization of the papillae and in its growth habits (initially fixed to the substratum). However, this last characteristic is not always visible, as it is not uncommon to find species of Polymastia that completely occlude the initial substratum and finish by living freely on the bottom (e.g.P. mamillaris). The constant presence of a tangential or perpendicular subcortical spicule layer in the genus Polymastia (Boury-Esnault, 1987) is another characteristic, not easily visible in Trichostema--at least not in the Mediterranean specimens of T. sarsi--which allows the separation of the two genera.
The substratum included by P. tissieri is often mud but may also comprise small shells or pebbles (the only hard substratum available on the bathyal bottoms inhabited by the sponge). On occasions the substratum is not fully included.
Although after superficial examination it could be considered comparable to the spicule fringe of Trichostemma, the basal hispidity of this species has a different structure. The width of the hispid area is not constant but varies in the different specimens studied (all of which exhibited similar skeletal characteristics) and may even be completely absent. Moreover, basal hispidity is found in some other species of the genus Polymastia, such as P. infrapilosa (Topsent, 1927). The papilla (Figs 2 F and G) also displays both skeletal and aquiferous organization typical of the genus Polymastia, very close to P. infrapilosa (Boury-Esnault, 1987). Lastly, according to our observations, the growth of this species is not always symmetrical. Consequently, the authors consider that, although P. tissieri is clearly an intermediate form between the genera Polymastia and Trichostemma, as Vacelet (1961) pointed out, and it represents a step towards the adaptation of these group of species of bathyal muds, on the basis of its general characteristics, it is closer to the former genus.
Trichostemma sarsi (Ridley and Denby, 1886)
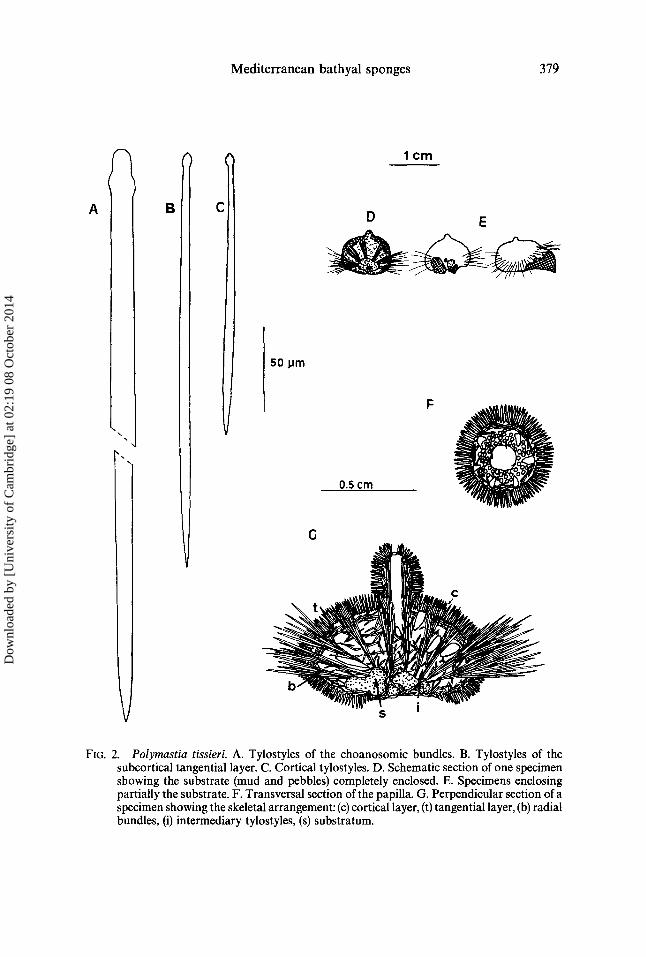
(Figs 3, 5 E and 6)
Occurrence. Station 1, 1 specimen; Station 2, 8 specimens; Station 3, 2 specimens; Station 4, 4 specimens; Station PM-6, 2 specimens.
Morphology Symmetrical sponges conical in shape, 0.5-1 cm in diameter (not including the
spicule fringe) and 0.1~)-6cm thick at the centre. Upper side discoidal, fiat, and somewhat hispid, with a short, central papilla. Lower side, more or less convex, depending on the specimen, glabrous with an outer fringe of long spicules.
There is a well-developed exhalant aquiferous system with a central cavity flowing into the papilla. Yellowish white in colour, masked by an accumulation of sediment between the spicules.
Skeleton Cortical layer, not well defined, of medium-sized tylostyles in a more or less palisade
arrangement. This cortical layer does not extend to the basal zone of the sponge which dearly displays a layer of radial, long choanosomic tylostyles just covered by scattered small-medium tylostyles hardly diverging from the ectosome. The prolongation of the layer of long choanosomic tylostyles makes up the peripheral spicules fringe.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014
Mediterranean bathyal sponges 381
50jJm A B C
' v
FIG. 3. Trichostemma sarsi. A. Intermediary choanosomic tylostyles. B. Cortical tylostyle. C. Tylostyle of the choanosomic radial bundles and of the marginal fringe. D. Transversal section of the papilla. E. Perpendicular section of a specimen showing the skeletal arrangement: (c) cortical layer, (b) radial bundles, (i) intermediary tylostyles, (f) spicule fringe.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

382 Maria-Jesus Uriz and Dolors Rosell
A great number of radial bundles of long tylostyles cross the choanosome from the basal zone passing through the upper cortical layer.
Small-medium sized tylostyles in a some irregular reticulate fashion are also distributed densely through all the choanosome.
Both the cortical spicule layer and the size-categories of tylostyles are more poorly differentiated in this species than in species of the genus Polymastia.
The papilla has a unique central exhalant canal pierced by small-medium spicules. Its skeleton is made up of longitudinal bundles of long tylostyles in the inner region, echinated by medium-sized spicules outside.
Cortical tylostyles. Straight or slightly curved with a well-defined head and a somewhat fusiform shaft, measuring 450-820 #m x 8-11/zm.
Subtylostyles of the radial choanosomic bundles. 950-2980#m x 10-27 #m in size, also covering the lower surface of the sponge and forming the outer fringe.
Intermediary choanosomic tylostyles. 140-300 #m x 6-9 #in in size, similar in shape to the cortical tylostyles.
In addition to the typical spicules, spicule variations can be observed occasionally, including a few subtylostrongyles and longer subtylostyles 3750-4100 pm x 7-8/~m in size, with a blunt point.
Distribution Northern Atlantic Ocean: Madeira, Cape Mogador (Topsent, 1928), Azores (Ridley
and Dendy, 1887; Topsent, 1892; Burton, 1959), Morocco and Madeira (Burton, 1956); 46°4'40"N, 49°2'30"W (Topsent, 1892). Indian Ocean: Saya de Malha (Dendy, 1921), Zanzibar and Maldive region (Burton, 1959), Southern Pacific Ocean: Australia (Ridley and Dendy, 1887; Burton, 1959).
Typically dwelling on muddy bottoms with Globigerina, Pteropoda or broken shells and pebbles, from 225 m to 2630 m.
Discussion The genus Trichostemma was defined by Sars (1872) in order to contain the species
T. hemisphaericum characterized by a hemispherical shape, a peripheral fringe of long spicules, numerous exhalant processes (papillae) and chiefly tylostyles as megascleres.
Previously, Schmidt (1870) had described a Mediterranean species (Radiella sol) which, from the illustration drawn by him, seemed to be similar to T. sarsi (Ridley and Dendy, 1887) but which Hansen (1885), after examination of the holotype, identified with T. hemisphaericum. Topsent (1913) later concurred with Hansen's opinion. Consequently, there was considerable confusion between species of these two genera (Trichostemma and Radiella) which are surely synonymous.
The arguments put forward by Ridley and Dendy (1887) regarding the priority of the genus Trichostemma over Radiella seem to us to be valid (Schmidt's generic diagnosis of this last genus is not recognizable); and if we accept the identity between T. hemisphaericum and R. sol following Hansen (1885) and Topsent (1913), the present specimens should be regarded as belonging not to T. hemisphaericum, but to T. sarsi, as they match perfectly the description of this species made by Ridley and Dendy (1887). Nevertheless, it is not so unlikely that T. hemisphaericum, R. sol and T. sarsi could all be varieties of a single polymorphic species.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 383
Order Poecilosclerida Family Biemnidae Hentschel, 1923
Sigmatoxella annexa (Schmidt, 1870)
Occurrence. Station 2:1 fragmented specimen.
Morphology Four fragments 0.7-1.5 cm x 1.5-2 cm in size which appeared to belong to a single,
soft, fragile, massive sponge, light brown in colour. Ectosomal layer very conspicuous and unevenly hispid. Well developed aquiferous system sometimes sheltering foreign materials.
Skeleton Tylostyles. 330--960/~m x 4-15/tm in size. Toxas. 100-120 #m in length. Sigmata. 1543 #m in diameter. The spicules are slightly larger than those of other Mediterranean specimens taken
at shallower depths (Uriz, 1983).
Distribution Cosmopolitan species with a wide bathymetric (185-1200m) and geographical
distribution. Typically dwelling in detrital, rocky and muddy bottoms.
Family Phorbasidae De Laubenfels, 1936; emended Van Soest, 1984 Phorbas fictitius (Bowerbank, 1866)
(Figs 4 D-F)
Occurrence. Station PM-5, 1 specimen on Caryophyllia sp.
Morphology Encrusting specimen, 1.5-2 mm thick occupying an area of 1 cm x 0.5 cm. Glabrous
surface with conspicuous rounded inhalant areas. Ectosome differentiated, easy to remove from the choanosome. The colour is whitish after preservation in alcohol.
Skeleton Tornotes. 275-350#m, fusiform, straight with short points. They echinate the
choanosomic bundles of acanthostyles, laying tangentially to the ectosoma as well. Acanthostyles. 80-230#m x 6-9 #m, straight, fully spiny with a fine point. They
make up short bundles in the choanosome despite the general thinness of the sponge. Isochelae. 22-28 #m, arcuate, with short teeth.
Distribution Mediterranean Sea (Topsent, 1925; SarA, 1961, 1964a, 1964b; Sara and Siribelli,
1960, 1962; Rubi6, 1971; Bibiloni, 1981; Pulitzer-Finali, 1983). Atlantic Ocean (Bowerbank, 1866; Borojevic et al., 1968; Descatoire, 1969; Rodriguez-Babio and Lorenzo-Gondar, 1978).
Bathymetric distribution: littoral to 2165 m.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

384 Maria-Jesus Uriz and Dolors Rosell
B
A
C
I I
)
G 50~m
lOjJrn C,F,K
I I
I
i I
D ' , J l
; . 4
N
t.I r
q \
r'l
'" \\
\
H ~ t
p
, 4
° ,
. °
s ' ,
r ::l
I
J ~'~
FIG. 4. A-C. Hymedesmia peachi. A. Tomote. B. Acanthostyles. C. Isochaela. D-F. Phorbas fictitius. D. Tornote. E. Acanthostyles. F. Isochaela. G-K. Plocamionida ambi#ua. G. Tornostrongyle. H. Acanthostyle I. I. Acanthostyle II. J. Acanthostrongyles. K. Palmate isochaela.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014
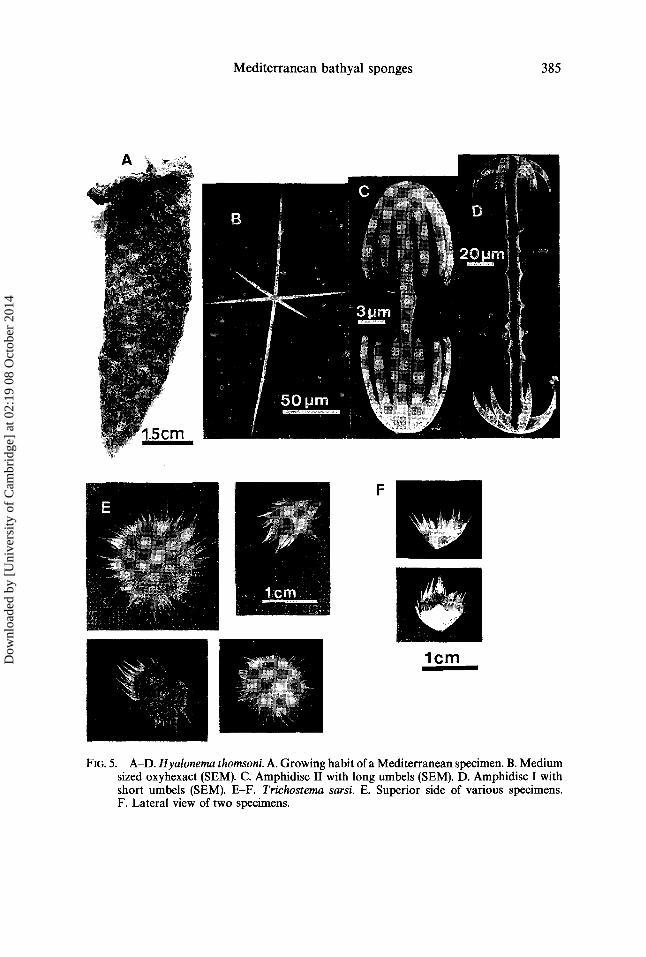
Mediterranean bathyal sponges 385
F
l c m I
FIG. 5. A-D. Hyalonema thomsoni. A. Growing habit of a Mediterranean specimen. B. Medium sized oxyhexact (SEM). C. Amphidisc II with long umbds (SEM). D. Amphidisc I with short umbels (SEM). E-F. Trichostema sarsi. E. Superior side of various specimens. F. Lateral view of two specimens.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014
386 Maria-Jesus Uriz and Dolors Rosell
~-:~<!:!:
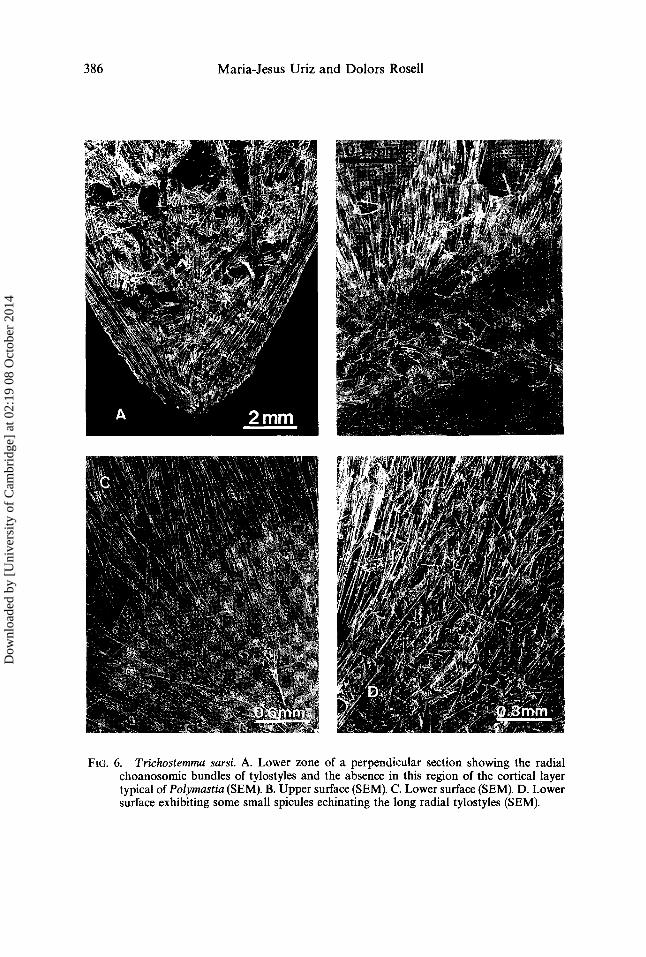
FIG. 6. Trichostemma sarsi. A. Lower zone of a perpendicular section showing the radial choanosomic bundles of tylostyles and the absence in this region of the cortical layer typical ofPolymastia (SEM). B. Upper surface (SEM). C. Lower surface (SEM). D. Lower surface exhibiting some small spicules echinating the long radial tylostyles (SEM).
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 387
Family Hymedesmiidae Topsent, 1928 Hymedesmia peachi Bowerbank, 1882
(Figs 4 A~C)
Occurrence. Station PM-5, 1 specimen on Caryophyllia sp.
Morphology Encrusting specimen 1 mm thick and 1 cm in diameter with minutely hispid surface
and inconspicuous oscules. Ectosome difficult to remove from the choanosome. Whitish cream in colour in alcohol.
Skeleton Tornotes. 290-310 #m x 3-4 #m, asymmetrical points. They are placed tangentially
in the ectosomal layer. Acanthostyles. 85-200pm x 4-10 #m placed perpendicularly to the substratum. Isochelae. Separable into two size categories 16-20#m and 29-35/~m, strongly
arcuate.
Distribution Mediterranean Sea (Topsent, 1936; Boury-Esnault, 1971; Pulitzer-Finali, 1983).
Atlantic Ocean (Topsent, 1891, 1894, 1895; Ferrer-Hernandez, 1914; Burton, 1930; Arndt, 1935; Koltun, 1959; Borojevic et al., 1968).
Bathymetric distribution: littoral to 1750m.
Family Clathriidae Hentschel, 1923 Plocamionida ambigua (Bowerbank, 1866)
(Figs 4 G-K)
Occurrence. Station PM-5, 3 specimens on Caryophyllia sp.
Morphology Soft thin encrusting strongly hispid specimens some miUimetres in diameter.
Ectosome and oscules inconspicuous, Whitish in colour after preservation in alcohol.
Skeleton Acanthostyles 1. Straight or slightly curved, strongly spiny at the basal zone and
with scattered spines along the shaft, 950-1125 #m x 11-12 #m. Acanthostyles II. 135-180#mx 5-7#m, fully spiny. Shaft somewhat fusiform,
either straight or curved at ~ from the base. Acanthostrongyles. 90--100#m x 5-8 btm, curved, with strong spines. Tornostrongyles. 245-280/~m x 3-4#m, straight, with asymmetrical somewhat
swelled ends. Isochelae. Arcuate, wide shaft, three teeth, 28-35/~m in length. The skeletal arrangement is typical of the genus: basal layer of acanthostrongyles
densely arranged; acanthostyles I rising vertically from this layer surrounded by acanthostyles II and by bundles of divergent tornostrongyles.
Distribution North Atlantic Ocean: Sweden (Friestedt, 1885; Alander, 1942), Shetland Islands
(Bowerbank, 1866), Norway (Arnesen, 1903; Burton, 1930), Asturias (Ferrer-
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

388 Maria-Jesus Uriz and Dolors Rosell
Hernandez, 1918), Detroit of Dover (Topsent, 1894), Portugal, Azores, West of Flores (Topsent, 1928). Mediterranean Sea: Monaco (Topsent, 1936), Port Cros (Vacelet, 1976). Arctic Ocean (Arndt, 1935).
Displaying a wide bathymetric distribution (20-2460 m) mainly found dwelling on bottoms of argillaceous or volcanic muds.
Discussion and conclusions According to our observations, the macrofauna inhabiting the argillaceous mud
bottoms at 1000-1750m between the Balearic Islands and continental Spain is composed of few invertebrate filter-feeders (except for Brachiopoda), with detritus and sediment feeders (mainly holothurians), predators and necrophagous species (fishes and Crustacea) predominating. The fauna encountered belongs to the compact muds 'facies', with an extremely reduced community of Isidella elongata, compared with that described by Prr~s and Picard (1964).
Human waste materials (plastics, tins, rags, etc.) accumulate on these deep bottoms in such quantities that their weight sometimes exceeds the animal biomass. Some benthic organisms (e.g., sponges) can turn these foreign materials to their own advantage using them as settling surfaces on bottoms where the absence of hard substrata is a limiting factor.
This study of the sponge fauna collected at the seven bathyal sampling stations in the western Mediterranean has confirmed the presence of two species new to the Mediterranean Sea, Trichostemma sarsi and Hyalonema thomsoni, and expanded the known bathymetric distribution of Polymastia tissieri.
The sponges found enjoy a broad geographical distribution, mainly concentrated in the north Atlantic, as Fredj and Laubier (1985) pointed out for the deep Mediterranean fauna in general. Thenea muricata and Sigmatoxella annexa are frequently found together on muddy and sandy bottoms in the Atlantic (Topsent, 1892, 1904); H. thomsoni, P. ambigua, H. peachi and Ph. fictitius are common Atlantic species: Tricostemma sarsi is widely distributed in the Atlantic, Indian and southern Pacific Oceans; even P. tissieri, until now considered as a Mediterranean endemic, seems also to have an amphi-Atlantic distribution (personal observation).
Acknowledgements We are very grateful to Mr J. Rucabado, 'Batimar' cruise leader, who kindly
facilitated the participation of one of the authors, and to Mr L. del Cerro, Mr V. Alvfi, Dr N. Merrett and Dr L. Saldanha for identifying the non-sponge fauna. Dr N. Boury- Esnault and Dr K. Ruetzler kindly allowed us to examine some slides of the holotype of T. sarsi and some unclassified specimens ofP. tissieri, respectively. Mr F. Pagrs, kindly collected the sponges from the 'Bathos-2' cruise. Special thanks are given to Dr J. Vacelet, Dr N. Boury-Esnault and to Professor C. Lrvi who added information on species bathymetric distribution and whose general comments improved the original manuscript and to Mr J. M. Fortufio and Mr J. Biosca for taking the scanning microphotographs.
References ALANDER, H., 1942. Sponges from the Swedish west-coast and adjacent waters. Goteborg:
H. Struves, Goteborg, 95 pp. ARI~W~SEN, E., 1903. Spongien yon der norwegischen kiiste, Monaxonida: Halichondrina. Bergens
Museums Arbog 1, 1-30.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 389
ARNESEN, E., 1920. Spongia from the 'Michael Sara' North Atlantic deep-sea expedition 1910. Rep. of the 'Michael Sars' North Atlant. Deep Sea Exped. 1910, 3 (2), 1-29.
ARNDT, W., 1935. Sponges. In Gdmp¢ and Wagler (Eds), Die Tierwelt der Nord- und Ostsee. Berlin 3a, 1-140.
BmmONl, M. A., 1981. Estudio sistemfitico de1 orden Poecilosclerida (Demospongia) de la costa Brava (Gerona). Boletin del lnstituto espa~ol de Oceanografia 6, 104-154.
BOROJEVIC, R., CABIOCH, L., and L~w, C., 1968. Inventaire de la Faune Marine de Roscoff. Spongiaires. Editions de la Station Biologique de Roscoff, 1-41.
BOWERBANK, S. J., 1866. II. Synopsis of Genera. In Ray Society London (Eds), A monograph of the British Spongiadae, London 2, 1-388.
BOURY-ESNAULT, N., 1971. Spongiaires de la zone rocheuse de Banyuls-sur Mer. II. Systrmatique. Vie Milieu, ser. B 22 (2), 28%350.
BOURY-ESNAULT, N., 1987. The Polymastia species (Demosponges, Hadromerida) of the Atlantic area. In, J. iCacelet and N. Boury-Esnault (Eds), Taxonomy of Porifera from the N.E. Atlantic and Mediterranean Sea. NATO ASI Series. Berlin: Springer-Verlag G13, pp. 29- 72.
BURTON, M., 1930. Additions to the Sponge Fauna at Plymouth. Journal of the Marine Biological Association of the United Kingdom 16, 489-507.
BURTON, M., 1956. The sponges of the West Africa. Atlantide Report 4, 111-147. BURTON, M., 1959. Sponges. Scientific Reports of the John Murray Expedition 1933-1934.
London: British Natural History, Museum 10 (5), 151-285. DENDY, A., 1921. Report on the Sigmatotetraxonida collected by H.M.S. 'Sealark' in the Indian
Ocean. Transactions of the Linnaean Society of London, ser. 2, Zoology 18 (1), 1-164. DESCATOIRE, A., 1969. Peuplements sessiles de rarchipel de Gl6nan. I. Inventaire: Spongiaires.
Vie Milieu 20 (lb), 177-209. FERREg-HERNANDEZ, F., 1914. Esponjas del Cantfibrico. Parte 2. III Myxospongida. IV
Tetraxonia. V. Triaxonida. Trabajos del Museo Nacional de Ciencias Naturales, Madrid. Ser. Zoologia 17, 3-47.
FERRER-HERNANDEZ, F., 1918. Esponjas del litoral de Asturias. Trabajos del Museo Nacional de Cieneias Naturales, Madrid. Ser. Zoologia 36, 1-39.
FREDJ, G. and LAUBIER, L., 1985. The Deep Mediterranean Benthos. In Moraitov- Apostolopoulou and V. Kiortsis (Eds) Mediterranean marine ecosystems. New York, London: Plenum Press, pp. 109-145.
FRISTEDT, K., 1885. Bidrag till 1/innedomen om de vid Sveriges vestra Kust lefvande Spongiae. Svenska Vetenskaps Akademisk Handlingar 21, 1-56.
HANSEN, G. A., 1885. XIII. Zoologi-Spongiadae. In, Den Norske Nordhaus-Expedition 1876- 1878. Cristiania. pp. 1-25.
IJIMA, I., 1927. The Hexactinellida of the Siboga Expedition. Siboga--Expeditie 4 (Monograph VI), 1-383.
KOLTUN, W. M., 1959. Siliceous Sponges of the North and Far Eastern Seas of the U.S.S.R. Opredelitei po faune S.S.S.R., Izdaterstvo ZIN AN SSSR 67, 1-235 (in Russian).
LENDENFELD, R. YON, 1915. Reports on the scientific results of the expedition to the eastern tropical Pacific etc. XXIX. The Sponges. 3. Hexactinellida. Memoirs of the Museum of Comparative Zoflogy at Harvard College 42, 1-397.
L~vI, C., 1964. Spongiaires des zones bathyale, abyssale et hadale. Galathea Report 7, 63-112. MARSHALL, W., 1875. Untersuchungen fiber Hexactinelliden. Zeitschriftffir Wissenschaftliche
Zoologie, Leipzig 25 (suppl.), 1-142. MEgRETT, N. R. and MAgSrt~LL, N. B., 1981. Observations on the ecology of deep-sea bottom-
living fishes collected off north-west Africa (0-8-27 N). Progress in Oceanography 9, 185- 244.
PfiRLs, J. M. and PIc~o~I), J., 1964. Nouveau manuel de bionomie eenthique de la Mer Mrditerranre. Recueil des Travaux de la Station Marine crEndoume 47 (31), 1-37.
PULITZER-FINALI, G., 1983. A collection of Mediterranean Demospongiae (Porifera) with, in appendix, a list of the Demospongiae hitherto recorded from the Mediterranean Sea. Annali del Museo Civico di Storia Naturale di Genova 84, 445-621.
RIDLEY, S. O. and DENDY, A., 1887. Report on Monaxonida collected by H.M.S. Challenger during the years 1873-1876. Report of the Scientific Results of the Voyage of H.M.S. Challenger, Zoology 20, 1-275.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

390 Maria-Jesus Uriz and Dolors Rosell
RODRIGUEZ-BAmo, C. and LOP~Nzo-GorCOAR, J. E., 1978. Fauna marina de Galicia. II. Contribuci6n al conocimiento de los poriferos del litoral gallego. Monografias de la Universidad de Santiago de Compostela 42, 1-68.
Rum6, M., 1971. Contribuci6n al estudio de la fauna bent6nica del litoral de Blanes. Resumen Tesis Universidad de Barcelona, 21 pp.
Russ, K. and RtrETZLER, K., 1959. Zur Kenntnis der Swammfauna unterseeischer Hfhlen. Publicazioni della Stazione ZooIogica di Napoli 30, 756-787.
RUETZLER, K., 1966. Die Poriferen einer sorrentiner H6hle. Zoologischer Anzeiger 176 (5), 303- 319.
SARA, M., 1961. La fauna di Poriferi delle grotte delle isole Tremiti. Studio ecologico e sistematico. Archivio Zoologico ltaliano 46, 1-61.
SARA, M., 1964 a. Porifera di acque superficiali (0-3 m) del litorale italiano. Annali del Pontificio Istituto Superiore di Scienze e Lettere 'S. Chiara' 14, 299-317.
SARA, M., 1964 b. Distribuzione ed ecologia dei Poriferi in acque superficiali della Riviera ligure di Levante. Archivio Zoologico ltaliano 49, 181-248.
SARA, M. and SISZBELLI, L., 1960. La fauna di Poriferi delle 'seche' del Golfo di Napoli. I. La 'secca' della Gaiola. Annuario delrlstituto e Museo di Zoologia della Universitd di Napoli 12 (3), 1-93.
SARA, M. and SmmELLr, L., 1962. La fauna di Poriferi delle 'seche' del Goifo di Napoli. I. La'secca di Benda Palumno'. Annuario deWIstituto e Museo di Zoologia della Universit& di Napoli 14 (2), 1-62.
SARS, M., 1872. Remarkable forms of animal life from the great deeps off the Norweoian coast. Cristiania, 62 pp.
SCHMIDT, O., 1870. Grundzuge einer Sponoien-fauna des Atlantischen Gebietes. Leipzig: Wilhelm Engelmann, 88 pp.
SCHULZE, E., 1887. Report on the Hexactinellida collected by H.M.S. Challenger during the years 1873-1876. Report of the Scientific Results of the Voyage of H.M.S. Challenger, Zoology 21, 1-513.
TOPSENT, E., 1891. Essai sur la faune des Spongiaires de Roscoff. Archives de Zoologie Exp~rimentale et G~n~ral, 2 s6r 9, 523-554.
TO~ENT, E., 1892. Contribution ~i l'6tude des Spongiaires de l'Atlantique Nord. Resultats des Campaones Scientifiques accomplies par le Prince Albert I. Monaco 2, 1-165.
TOPSENT, E., 1894. Etude sur la Faune des Spongiaires du Pas-de Calais suivie d'une application de la Nomenclature actuelle ~ la Monographie de Bowerbank. Revue de Biolooie du Nort de France 7 (1), 6-28.
TOPSENT, E., 1895. Campagnes du Jacht 'Princesse Alice'. Notice sur les spongiaires recueillies en 1894 et 1895. Bulletin de la Societ~ Zoolooique de France 20, 213-216.
TOPSENT, E., 1896. Eponges. Resultats scientifiques de la Campagne du 'Caudan' dans le Golfe de Gascogne: aout-septembre 1985. Annuaire de rUniverist~ de Lyon 2, 273-295.
TOPSV, NT, E., 1900. Etude monographique des Spongiares de France, III Monaxonida (Hadromerina). Archives de Zoolo#ie Exp~rimentale et G~n~ral, 3 s6r. 8, 1-331.
TOPSENT, E., 1904. Spongiaires des A~ores. Resultats des Campaones Scientifiques accomplies par le Prince Albert I. Monaco 25, 1-279.
TOPSENT, E., 1913. Spongiaires provenant des campagnes scientifiques de la 'Princesse Alice' dans les Mers du Nord (1898-1899). Resultats des Campagnes Scientifiques accomplies par le Prince Albert I. Monaco 45, 3-67.
TOPSENT, E., 1925. Etude de Spongiaires du Golfe de Naples. Archives de Zoologie Exp~rimentale et G~n~ral 63 (5), 623-725.
TO~ENT, E., 1927. Diagnoses d'Eponges nouvelles recueillies par le Prince Albert de Monaco. Bull. Inst. Oc~anogr. 502, 1-19.
TOPSENT, E., 1928. Spongiaires de l'Atlantique et de la M6diterran6e. Resultats des Campaones Scientifiques accomplies par le Prince Albert I. Monaco 74, 1-376.
TOPSENT, E., 1936. Eponges observ6es dans les parages de Monaco. Bulletin de rlnstitut Oc~anographique de Monaco 689, 1-70.
URIZ, M. J., 1981. Estudio sistem~itico de las esponjas Astrophorida (Demospongia) de los fondos de pesca de arrastre entre Tossa y Calella (Catalufia). Bolet'm del Instituto espa~ol de Oceanografia 6, "]-58.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014

Mediterranean bathyal sponges 391
URIZ, M. J., 1983. Contribuei6n a la fauna de esponjas (Demospongia) de Catalufia. Anales de la secci6n de Ciencias del Coleoio Universitario de Gerona. Monografia I, 1-220.
URIZ, M. J. and ROSELL, M. D., 1986. Hyalonema thomsoni Marshall, une nouvelle Hexactinellide en M6diterran6e et ses affinit6s avee H. infundibulum Topsent. Rapports et Proeds Verbaux des R~unions de la Commission Internationale pour rExploration Scientifique de la Met Mdditerrande 30 (2), 11.
VACELET, J., 1961. Quelques 6ponges remarcables de M6diterran6e. Revue des Travaux Institute P~ches maritimes 25 (3), 351-354.
VACELET, J., 1969. Eponges de la Roche du Large et de l'6tage bathyal de M6diterran6e (R6coltes de la soueoupe polongeante Cousteau et dragages). Memoires Museum national d'Histoire Naturelle, ser A 59 (2 et dernier), 145-219.
VACELET, J., 1976. Inventaire des Spongiaires du Pare National de Port-Cros (Var.). Travaux Scientifiques du Parc National de Port-Cros 2, 167-186.
ZmROVaUS, H., 1985. Spongiaires Hexactinellides vivant en mer Ionienne par 2000m de profondeur. Rapports et Proc~s Verbaux des R~unions de la Commission Internationale pour FExploration Scientifique de la Mer M~diterranke 29 (5), 351-354.
Dow
nloa
ded
by [
Uni
vers
ity o
f C
ambr
idge
] at
02:
19 0
8 O
ctob
er 2
014