Embed Size (px)
Citation preview

Chapter 26 Maturation K. NAKAMURA Faculty of Fisheries, Universiiy of Kagoshima, 4-50-20 Shimoaruta,
Kagoshima-shi 890-0056, Japan
26.1 Introduction
Maturation and reproductive mechanisms in spiny lobsters have only been studied in a few species in detail. Several descriptions of their reproduction biology, basic information on the sexual maturity, annual cycles of reproduction, spawning time and fecundity have come mainly from data on the fisheries’ resources. The available knowledge related to brood stock control comes from rearing experiments accompanied by physiological studies designed to clarify the reproductive processes and the factors controlling them.
26.2 Reproductive system
The reproductive system of the Japanese spiny lobster Panulirus japonicus possesses similar structures to those of other Palinuridae and Nephropidae species. The external and internal reproductive organs as well as the arrangement of the genital apertures were described for P . penicillutus (Matthews, 195 l), P . japonicus (Okamura, 1956) and Jusus lalandii (Paterson, 1968).
26.2.1 Male
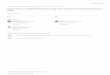
The internal reproductive system of male P. japonicus is shown in Figure 26.1. The matured testis is a semitransparent or slightly yellowish organ, forming elongated lobes which are much smaller than the ovary of females. It is located on the median dorsal of the midgut gland, beneath the heart. It is H-shaped, similar to that in other lobsters (Fielder, 1964; Farmer, 1974; Hunter et al., 1996), with two anterior lobes extending to the dorsolateral stomach, and two posterior lobes lengthening backwards to the first abdominal segment. The posterior lobes lie between the dorso-abdominal muscle situated superficially and main segmental masses of the ventral abdominal muscles in parallel with the midgut. The paired vasa deferentia arise from each posterior lobe and pass downwards to an opening pore at the coxopodite of the fifth pereiopod.
The opening site is provided with a special apparatus which is an ammonitoid protuberance. The tip portion is slightly raised as its axis is not parallel to the basal
474
SPINY L0BSTERS:FISHERIES AND CULTURE B.F. PHILLIPS&J. KITTAKA
Cowriaht 0 200 bv Fishina News Books
SPINY L0BSTERS:FISHERIES AND CULTURE B.F. PHILLIPS&J. KITTAKA
CoDvriaht 0 2000 bv Fishina News Books

Maturation 475
OD
Fig. 26.1 Male and matured female reproductive systems in the Japanese spiny lobster Punulirus juponicus. Upper: dorsal view (male body weight, 132 g; carapace length, 52 mm). Middle: ventral view (male body weight, 186 g; carapace length, 57 mm). Lower: dorsal view (female body weight, 200 g; ovary weight, 19.7 g; carapace length, 61 mm). The testis (T) sends out the vas deferens (VD) which connects to the ammonitoid apparatus (VA) at the coxopodite of the fifth pereiopod (CP). The ovary (OV) shows an H-shape, sending laterally the oviduct (OD) from the median level of the ovarian length.
plane of the proximal whirlpool. The degree of its sclerotization seems to decrease according to ageing of the male; in the intermoult male of 188 g body weight [carapace length (CL) 63 mm] it is completely sclerotized and pigmented like the hard exoskeleton of the thoracic sternum and pereiopod. However, in more aged and larger males, the apparatus is not as hard and pigmented as the neighbouring cuticle. Its arthrodial membrane is slightly thicker than that of the ventral abdomen. This organ was described as an epicuticular rim in the male J. lalandii (Paterson, 1968; Silberbauer, 1971b). However, in P . juponicus the male gonopore is still obscure, although it has been indicated to be at the distal of the apparatus (Okamura, 1956). The author’s histological observation in P. japonicus (Nakamura, 1993) has revealed that it opens at the bottom of the peripheral invagination inside the apparatus, similarly to the case of the epicuticular rim in J. lalundii. The above-mentioned characteristic of the apparatus observed in the young male seems to be unsuitable for copulation if it functions as a genital apparatus. However, it is possible that its hardness and shape change during the moult, which precedes copulation, and this post-moult male with a soft cuticle would copulate. In the case of aged and larger

476 Spiny Lobsters: Fisheries and Culture
males there is a possibility that its distal portion of the apparatus is broken at copulation because two males of 400 g and 750 g (CL 80 mm and 90 mm) had the partially repaired apparatus of its medial portion near the tip.
It is possible to divide the vas deferens into three parts: (1) a proximal portion of the highly coiled tube; (2) an intermediate portion of the thick tube of which the initial part is helical; and (3) a distal ejaculatory portion of the slender tube. The spermatophores seem to be produced in the secretory area of these proximal and intermediate vasa deferentia and stored there until copulation. In the case of P . elephas, the distal ejaculatory portion greatly enlarges and the spermatophores are stored therein (Hunter et al., 1996). The testis yields not only spermatids but also nutrient cells observable in the spermatophores. The swollen tube of the intermediate vas deferens changes its diameter according to the maturation cycle (Minagawa & Higuchi, 1997), owing to the quantity of internal matrix embedding the sperms.
26.2.2 Female
The internal reproductive system of female P. japonicus is shown in Figure 26.1. The ovary is located dorsally on the midgut gland, beneath the heart. It is H-shaped, similar to that in other lobsters (Fielder, 1964; Farmer, 1974; Talbot, 1981; Juinio, 1987; Hunter et al., 1996). Its anterior lobes extend to the cephalic area with their tips turning upwards. Its posterior lobes, in the matured condition, elongate to the fourth abdominal segment, parallel to the midgut. The posterior lobes differ in length from each other, the left one commonly being longer. An oviduct derives from the middle level of the ovary length on each side, posterior to the lobe connecting the two halves. It is a semitransparent and slender tube, and opens to the coxopodite of the third pereiopod. There is no genital apparatus at that opening site, differing from the case of the vas deferens. The thelycum is absent, as in J. lalundii (Paterson, 1968) but unlike Nephropidae species. There are a few reports on ovarian development during maturation in spiny lobsters. The ovary is externally white or weakly yellow in the immature condition, and changes to reddish orange or dark orange (Minagawa & Sano, 1997) in the completely mature condition. According to the stage of ovarian development, its shape and size will also change. However, detailed studies related to these processes are extremely limited (Minagawa & Sano, 1997). The author previously investigated this during the breeding season to reveal the relationship between the ovary’s weight and its maturation condition. The value for the gonadosomatic index was 9.5% in a fully matured female (200 g body weight, 61 mm CL).
26.3 Histology of the ovary and developmental oocytes
In P. japonicus, the histological structure of the ovary is as follows. The ovarian wall is composed of two layers, one a superficial connective tissue supplied with blood

Maturation 477
Fig. 26.2 Developmental process of ova in the Japanese spiny lobster Punulirus japonicus. Follicular cells are omitted in these figures. Scale bar = 125 pm; em, egg membrane; vo, vacuole; yg, yolk granules. Each combination of roman numeral and digit corresponds to the stage referred to in the text.
vessels and the other an inner germinal epithelium. Layers of the outer epithelium and smooth muscle are not prepared in this species, in contrast to other spiny or clawed lobsters (Bumpus, 1891; Herrick, 1909; von Bonde, 1936). The connective tissue varies periodically in thickness according to ovarian maturation. For example, its thickness decreases from 0.13 to 0.04 mm in individuals of 150-200 g body weight. The germinal epithelium forms inward folds running through the ovarian length, from which ova grow. They are enveloped in a single layer of flat cells, i.e. follicular cells. However, these follicular cells are difficult to be seen histologically until a certain developmental stage. Matured ova reach around 0.5 mm in diameter.
Developmental stages of the ovary have been assigned by different researchers on the basis of oocyte size, ovary size and ovary colour. There are seven macroscopic stages for J . edwardsii and J. lalandii (= J. edwardsii) (Fielder, 1964), five for P . penicillutus (Juinio, 1987), and six for P . homarus rubellus (Berry, 1971) and P. elephas (Hunter et al., 1996).

478 Spiny Lobsters: Fisheries and Culture
Development of the oocytes in P . japonicus may be classified morphologically into three phases or six stages in haematoxylin-eosin histological preparations (Fig. 26.2). Z Non-vitellogenesis phase: this is an initial and immature stage of development.
The maximum diameter of oocytes is 120-130 pm. Each nucleus is around 30 pm, and contains commonly a nucleolus as well as dispersed chromatin. The cytoplasm is stained with haematoxylin. ZZ Primary vitellogenesis phase: this phase is divided into three stages. (1) The
oocyte and nucleus diameters are 13&140 pm and 40-50 pm, respectively. The cytoplasm is haematoxylin positive, possessing a few vacuoles peripherally. (2) The diameters of the oocyte and nucleus are around 150 pm and 40-50 pm, respectively. The cytoplasm reduces its haematoxylin-positive character and becomes eosinophi- lic. The peripheral vacuoles increase distinctly in number and are distributed around the nucleus. (3) The maximum diameter of the oocytes is around 250 pm, with the nucleus of 40-50 pm diameter. Eosinophilic granules appear at the peripheral cytoplasm among the distributed vacuoles. The egg membrane appears between the oocyte and follicular cells.
IZZ Secondary vitellogenesis phase: this phase corresponds to a period of yolk accumulation. It may be divided into two stages. (1) The oocytes have a diameter range of 260-480 pm and a nucleus of about 20 pm. In the cytoplasm, yolk appears abundantly as eosinophilic granules. (2) The maximum diameter of the oocytes reaches about 500 pm. The nuclei decrease in size and finally become difficult to see. The thickness of the egg membrane is measured as 4-8 pm. This stage corresponds to the completely mature period of the oocyte.
The above-mentioned classification in P. juponicus needs to be examined in detail before its application to other species. Recently, light- and electron-microscopic observations detailed oogenesis and ovarian development in P . japonicus (Minagawa & Sana, 1997), and eight substages were distinguished.
26.4 Size at maturity
For P. japonicus, only female minimum size at maturity has been reported, with a CL of 38 mm (Inoko et al., 1979). In other cases, it corresponded to 42 mm CL and a body weight of 80 g, at which individuals were assumed to be 1.5-2 years old after the puerulus larvae (Ino, 1947; Kanamori, 1988) or 41.8 mm CL as the estimated size at 50% maturity (Minagawa, 1997). In P . homarus homarus, a value of 3847 mm was reported and the size at onset of oviposition was estimated as 59.5 mm, both as rostra1 CL (Jayakody, 1989).
For other female Punulirus, the CL at first maturity was 86 or 69 mm in P. argus (Evans et al., 1995; Pollock, 1997) and 59 mm in P . guttatus (Evans et al., 1995). In P. longipes longipes, the smallest females with eggs and mature ovaries were 41.8 and 41.4 mm CL, respectively (Gomez et al., 1994).

Maturation 479
For female P. gilchristi, the minimum size at maturity was 55 mm CL (Pollock & Augustyn, 1982; Pollock, 1997).
For female J . lalandii on the west coast of South Africa, the minimum size at maturity was reported as 45 mm or 55-59 mm CL (Matthews, 1962; Heydorn, 1965; Pollock, 1997). For J . edwardsii, the value was between 70 and 97 mm (Bradstock, 1950). This size is known to vary according to a distribution difference of habitat, even when comparing the same species (Street, 1969). For example, female J. lalandii at Queap Bay in South Africa were significantly smaller (53.0 mm) at 50% maturity than those at the higher latitudes of Dassen Island (59.5 mm), Elands Bay (59.9 mm) and Olifantsbos (59.5 mm) (Cockcroft & Goosen, 1995). In J . edwardsii from east and west localities off North Island, New Zealand, the above values for females were 75-79 and 65-69 mm, respectively (Annala et al., 1980).
Further, in P. penicillatus the values were 43 mm in the Saudi Red Sea (Hogarth & Barratt, 1996), 40.8 mm in the Philippines (Juino, 1987), 50 mm in the Gulf of Aqaba (Plaut, 1993) and 62 mm in Enewetak, Marshall Islands (Ebert & Ford, 1986).
Practically, the body weight, total length, CL and rostra1 CL can be used as the maturity index. However, these indices are available only during the breeding season. An ovigerous setae method seems valid, although the estimated size at 50% maturity would be a little smaller than that obtained by an ovigerous female method (Groeneveld & Melville-Smith, 1994).
As for internal indices of the maturity, the increased weight of the testis including accessory genital organs such as the vas deferens is useful in some species (Lindberg, 1955; Heydorn, 1969). In females, the weight, size and colour of the ovary indicate the maturation condition; the ovary changes from the immature colour of creamy white to light orange, then darker orange, and finally brick red in both Jusus and Panulirus species (Fielder, 1964; Berry, 1971).
26.5 Spawning time
The typical annual reproductive cycles of female spiny lobster are different among species. From the results of the ovary condition, two spawnings per year are indicated in P. japonicus (Ino, 1950). Of large females with over 47 mm CL, 93% were estimated to spawn twice during the spawning season (Minagawa, 1997). The natural spawning season of this lobster in Japan shows a tendency towards earlier spawning at lower latitudes with higher water temperature in winter, and its range shortens according to the increase in latitude (Inoue, 1981). This season in Japan ranges from early April to early October except in the southern area, Okinawa (Deguchi, 1988). At Okinawa, the spawning season ranges from early January to late October, with its zenith from early May to mid-June.
For P. longipes Zongipes in the Philippines, at least two broods of individuals in rapid succession were reported, particularly during the warmer months of March to

480 Spiny Lobsters: Fisheries and Culture
May (Gomez et al., 1994).For P. penicillatus in the Gulf of Aqaba, Red Sea, females spawned two to four times during the reproductive season from February to October (Plaut, 1993).
For P . homarus rubellus in South Africa and P . polyphagus in the Indian Ocean, three to four broods were yielded annually (Berry, 1971; Kagwade, 1988). However, J . edwardsii in New Zealand and P. delagoae and P. gilchristi in South Africa produced only one brood (Sorensen, 1969; Berry, 1973; Groeneveld & Rossouw, 1995). From these references, it is considered that maturation of all spiny lobsters as well as their oviposition occur at least once in early summer, although there may be a few exceptions, then a second spawning follows, with intervals of 2.5-7 months according to the reproductive cycle of the lobster.
It may be possible to regulate the spawning time by controlling the water temperature of the brood stock. This idea is supported by data on the effect of temperature on the moulting and spawning of the Japanese spiny lobster (Table 26.1) (Deguchi, 1988).
26.6 Copulation and spawning behaviour
For the Japanese spiny lobster, copulation behaviour has been reported in the laboratory (Nagai, 1956) and the procedure was as follows. In mid-May two males and three females were reared in a 16-21°C aquarium with dimensions of 78 x 52 x 70 cm. These sizes were not mentioned. All females were considered to have attained a mature condition, because examination of a dead specimen showed a completely ripe ovary. On 29 May, one female was found to be carrying eggs in the morning. A mucous substance of milky white spermatophore adhered to the ventral surface of its thorax. In the case of P. elephas, two spermatophores were normally deposited on either side of the distal portion of the female’s sternum, below the genital openings (Hunter et al., 1997). From the early morning of the previous day, the female had been observed combing the long and simple setae on each endopodite of the swimmeret with the fifth chelipeds. On the afternoon of 2 June, frequent combing occurred, and a male pursued the female at 21.00 h. The female escaped on several occasions but they finally clung together and copulation took place at 22.50 h. They were in an erect posture, being in contact along the ventral surface of the thorax. It took about 25 s from start until body separation after completion. Such copulation by the same pair occurred four times with 20 or 35 min intervals. The time required for copulation was always within 30 s.
At 3.37 h on 3 June, the female spawned. The female supported her body axis perpendicularly, using the aquarium wall as a prop. Such a posture seems to be general among the spiny lobsters (Berry, 1970). The fourth to sixth segments of the abdomen were flexed and the expanded tail fan covered the gonopore. All swimmerets formed bilaterally temporary side-screens, as if to prevent the loss of extruded eggs. It took about 20 min to carry out the spawning. However, the zenith

Maturation 48 1
Table 26.1 Comparison of time schedules of moulting and spawning in the Japanese spiny lobster reared at water temperatures of 20°C and 25°C
Year and conditionsa Critical dates
1986
WT 25°C CL6.8 cm
BW 295 g 1986 WT 25°C
CL 6.7 cm
BW 270 g 1987
WT 20°C CL 4.2 cm
BW 163 g 1987 WT 20°C
CL 5.9 cm BW 314 g
1987
WT 20°C CL 4.6 cm
BW 174 g
30 April 31 May 6 July 3 August
Moulting Spawning Hatching Moulting
7 May 8 June 12 July
Moulting Spawning Hatching
8 March 7 May 9 May 29 May 12 July
Moulting Spawning Eggs dislodged Eggs dislodged Moulting 6 May 26 June 25 July
Moulting Spawning Eggs dislodged
20 May 24 June 4 July
Moulting Spawning Died while carrying eggs
From Deguchi (1988). WT, water temperature; CL, carapace length; BW, body weight.
of oviposition seemed to have occurred within 10 min. This egg extrusion also took less than 1 h in P . homarus (Berry, 1970); however, it took 2-5 h in J . lalundii and J . edwardsii (von Bonde, 1936; Sorensen, 1969; Silberbauer, 1971a).
Another male and the females in the same aquarium did not show any positive response to the activities during the above-mentioned behaviour of the couple.
26.7 Effect of eyestalk ablation
Regarding the endocrine control of spiny lobster maturation, a few studies have reported on eyestalk ablation experiments in P . argus and P . hornarus (Quackenbush & Herrnkind, 198 1; Radhakrishnan & Vijayakumaran, 1984). The results indicated the presence of the moult and ovary-inhibiting factors in the eyestalk.

482 Spiny Lobsters: Fisheries and Culture
The author investigated the physiological relation between the eyestalk and ovarian maturation in P . japonicus. Within 2 weeks after eyestalk ablation in early summer (water temperatures of 24-30"C), some of the females of 150-200 g body weight spawned. The number of eggs ranged from 90 000 to 120 000, which is less than the value in other Palinuridae species. For example, 300 000 eggs were reported in P . argus of 75 mm CL (Squires & Riveros, 1978). However, the number is reported to vary according to female body size. In P.japonicus, it ranges from 29 000 to 554 000 for female body lengths of 12.0-25.4 cm (Ino, 1947). The other females did not spawn or in several cases died. The oocytes in the ovary of the non-spawned individuals were at the secondary vitellogenesis phase, accompanying yolk reabsorption. Another non-spawned female of 244 g body weight had entered a pre-moult stage. Its gonadosomatic index was 0.61% and the ova were at the primary vitellogenesis phase. From these results, it can be deduced that the eyestalk not only relates to the ovary-inhibiting mechanism but also is involved in the physiological integration of the moulting metabolism and ovarian development.
26.8 Discussion
For crustacean rearing, similarly to other aquaculture, it is very important to have a basic knowledge of the biology of the species, especially information on growth and maturation. However, most attempts at mariculture of spiny lobsters have been made with insufficient information. This is due to a deficiency of basic data relating to the breeding physiology, because of the slow growth rate of spiny lobsters.
Physiological data are as yet too fragmentary to reveal fully the maturation process and the mechanisms in spiny lobsters. The species-specific and geographical differences observed in the annual reproduction cycle of spiny lobsters suggest the necessity for a new approach from a comparative standpoint.
References
Annala, J.H., McKoy, J.L., Booth, J.D. & Pike, R.B. (1980) Size at the onset of sexual maturity in female Jasus edwardsii (Decapoda: Palinuridae) in New Zealand. N.Z. J . Mar. Freshwat. Res.,
Berry, P.F. (1970) Mating behaviour, oviposition and fertilization in the spiny lobster Panulirus homarus (Linnaeus). Oceanogr. Res. Inst. (Durban) Invest. Rep., 24, 1-16.
Berry, P.F. (1971) The biology of the spiny lobster Panulirus homarus (Linnaeus) off the east coast of southern Africa. Oceanogr. Res. Inst. (Durban) Invest. Rep., 28, 1-75.
Berry, P.F. (1973) The biology of the spiny lobster Palinurus delagoae Barnard, off the coast of Natal, South Africa. Oceanogr. Res. Inst. (Durban) Invest. Rep., 31, 1-27.
Bonde, C. von (1936) The reproduction, embryology and metamorphosis of the Cape crayfish (Jusus lakundii) (Milne-Edwards) Ortman. Invest. Rep. Fish. Mar. Biol, Surv. Div. S. Afr., 6,
14, 217-27.
1-25.

Maturation 483
Bradstock, C.A. (1950) A study of the marine spiny crayfish Jasus lalandii (Milne-Edwards),
Bumpus, H.C. (1891) The embryology of the American lobster. J. Morphol., 5, 215-62. Cockcroft, A.C. & Goosen, P.C. (1995) Shrinkage at moulting in the rock lobster Jasus lalandii and
associated changes in reproductive parameters. S. Afr. J. Mar. Sci., 16, 195-203. Deguchi, Y. (1988) Copulation and spawning; Japanese spiny lobster. In Seed Production of
Decapod Crustaceans (Ed. by R. Hirano), pp. 64-75. Koseisya-Koseikaku, Tokyo (in Japanese). Ebert, T.A. & Ford, R.F. (1986) Population ecology and fishery potential of the spiny lobster
Panulirus penicillatus at Enewetak Atoll, Marshall Islands. Bull. Mar. Sci., 38, 5 M 7 . Evans, C.R., Lockwood, A.P.M., Evans, A.J. & Free, E. (1995) Field studies of the reporductive
biology of the spiny lobsters Panulirus argus (Latreille) and P. gutratus (Latreille) at Bermuda. J. Shellfish Res., 14, 371-81.
Farmer, AS. (1974) Reproduction in Nephrops norvegicus (Decapoda: Nephropidae). J. Zool., 174,
Fielder, D.R. (1964) The spiny lobster, Jaws lalandii (H. Milne-Edwards), in South Australia. 11. Reproduction. Aust. J. Mar. Freshwat. Res., 15, 1 3 3 4 .
Gomez, E.D., Juinio, M.A.R. & Bermas, N.A. (1994) Reproduction of Panulirus longipes longipes in Calatagan, Batangas, Philippines. Crustaceana, 67, 1 10-20.
Groeneveld, J.C. & Melville-Smith, R. (1994) Size at onset of sexual maturity in the South Coast rock lobster Palinurus gilchristi (Decapoda: Palinuridae). S. Afr. J. Mar. Sci., 14, 219-23.
Groeneveld, J.C. & Rossouw, G.J. (1995) Breeding period and size in the South Coast rock lobster, Palinurus gilchristi (Decapoda: Palinuridae). S. Afr. J. Mar. Sci., 15, 17-23.
Herrick, F.H. (1909) Natural history of the American lobster. Bull. U S . Bur. Fish., 29, 149408. Heydorn, A.E.F. (1965) The rock lobster of the South Africa west coast, Jasus lalandii (H. Milne-
Edwards). I. Notes on the reproductive biology and the determination of minimum size limits for commercial catches. S. Afr. Div. Sea Fish. Invest. Rep., 53, 1-32.
Heydorn, A.E.F. (1969) Notes on the biology of Panulirus homarus and on length/weight relationship of Jasus lalandii. S. Afr. Div. Sea Fish. Invest. Rep., 69, 1-26.
Hogarth, P.J. & Barratt, L.A. (1996) Size distribution, maturity and fecundity of the spiny lobster Panulirus penicillatus (Oliver 1791) in the Red Sea. Trop. Zool., 9, 399408.
Hunter, E., Shackley, S.E. & Bennett, D.B. (1996) Recent studies on the crawfish Palinurus elephas in South Wales and Cornwall. J. Mar. Biol. Ass. U.K., 76, 963-83.
Ino, S . (1947) Spawning fecundity and spawning time in Panulirus japonicus De Haan. Nippon Suisan Gakkaishi, 13, 32-3 (in Japanese).
Ino, S. (1950) Observation on the spawning cycle of Ise-ebi Panulirus japonicus v. Siebold. Nippon Suisan Gakkaishi, 15, 725-7 (in Japanese).
Inoko, Y., Kawanishi, M., Hirata, S . & Takaba, M. (1979) Release and research of Gazami larvae- IX. Fisheries and resources analysis concerning the 50’ release group at Edajima Bay. Rep. Hiroshima Pref. Fish. Inst., 10, 51-5 (in Japanese).
Inoue, M. (1981) Studies on breeding of phyllosoma larvae of the Japanese spiny lobster. Rep. Fish. Res. Inst. Kanagawa Pref., 1, 1-91 (in Japanese).
Jayakody, D.S. (1989) Size at onset of sexual maturity and onset of spawning in female Panulirus homarus Crustacea Decapoda Palinuridae in Sri Lanka. Mar. Ecol. Prog. Ser., 57, 83-8.
Juinio, M.A.R. (1987) Some aspects of the reproduction of Panulirus penicillatus (Decapoda: Palinuridae). Bull. Mar. Sci., 41, 242-52.
Kagwade, P.V. (1988) Reproduction in the spiny lobster Panulirus polyphagus (Herbst). J. Mar. Biol. Assoc. India, 30, 3746.
Kanamori, K. (1988) Studies on fisheries managements and resource ecology of Ise-ebi in the Kinann area at Wakayama Pref. Rep. Wakayama Pref. Fish. Inst., 109-209 (in Japanese).
Lindberg, R.G. (1955) Growth, population dynamics, and field behavior in the spiny lobster, Panulirus interruptus (Randall). Univ. Ca lg Publ. Zool., 59, 157-248.
including accounts of autotomy and autopsy. Zool. Publ. Victoria Univ. CON., 7, 1-38.
161-83.

484 Spiny Lobsters: Fisheries and Culture
Matthews, D.C. (1951) The origin, development and nature of the spermatophoric mass of the spiny lobster, Panulirus penicillatus (Oliver). Pac. Sci., 5, 359-71.
Matthews, D.C. (1962) The rock lobster of South West Africa, Jams lalandii (Milne-Edwards). Size frequency, reproduction, distribution, and availability. S. W. Afr. Mar. Res. Lab., Invest. Rep.,
Minagawa, M. (1997) Reproductive cycle and size-dependent spawning of female spiny lobsters (Panulirus japonicus) off Oshima Island, Tokyo, Japan. Mar. Freshwat. Res., 48, 869-74.
Minagawa, M. & Higuchi, S. (1997) Analysis of size, gonadal maturation, and functional maturity in the spiny lobster Panulirus japonicus (Decapoda: Palinuridae). J. Crust. Biol., 17, 70-80.
Minagawa, M. & Sano, M. (1997) Oogenesis and ovarian development cycle of the spiny lobster Panulirus japonicus (Decapoda: Palinuridae). Mar. Freshwat. Res., 48, 875-87.
Nagai, H. (1956) Copulation and spawning behaviours observed in Panulirus japonicus. Suisan- Zosyoku, 4, 9-1 I (in Japanese).
Nakamura, K. (1993) Structure of male accessory apparatus related to the gonopore in the Japanese spiny lobster. Bull. Jpn SOC. Sci. Fish., 59, 1489-93.
Okamura, S. (1956) Anatomy of the Experimental Animals, pp. 141-71. Kazama-Syobou, Tokyo, Japan (in Japanese).
Paterson, N.F. (1968) The anatomy of the cape rock lobster, Jasus Ialandii (H. Milne-Edwards). Ann. S . Afr. Mus., 51, 1-232.
Plaut, I. (1993) Sexual maturity, reproductive season and fecundity of the spiny lobster Panulirus penicillatus from the Gulf of Eilat (Aqaba), Red Sea. Aust. J. Mar. Freshwat. Res., 44, 527-35.
Pollock, D.E. (1997) Egg production and life-history strategies in some clawed and spiny lobster populations. Bull. Mar. Sci., 61, 97-109.
Pollock, D.E. & Augustyn, C. (1982) Biology of the rock lobster Palinurus gilchristi with notes on the South African fishery. Fish. Bull. S. Afr., 16, 57-73.
Quackenbush, L.S. & Herrnkind, W.F. (1981) Regulation of molt and gonadal development in the spiny lobster, Panulirus argus (Crustacea: Palinuridae): effect of eyestalk ablation. Comp. Biochem. Physiol., 69, 523-7.
Radhakrishnan, E.V. & Vijayakumaran, M. (1984) Effect of eyestalk ablation in the spiny lobster Panulirus homarus 3. On gonad maturity. Indian J . Fish., 31, 209-16.
Silberbauer, B.I. (1971a) The biology of the South African rock lobster Jasus lalandii (H. Milne- Edwards). I. Development. S. Afr. Div. Sea Fish. Invest. Rep., 92, 1-70.
Silberbauer, B.I. (1971 b) The biology of the South African rock lobster Jasus lalandii (H. Milne- Edwards). 2. The reproductive organs, mating and fertilisation. S. Afr. Div. Sea Fish. Invest. Rep., 93, 1-46.
Sorensen, J.H. (1969) The New Zealand rock lobster or marine spiny crayfish Jaws edwardsii (Hutton): distribution, growth, embryology and development. N.Z. Mar. Dep. Fish. Tech. Rep., 29, 1 4 6 .
Squires, H.J. & Riveros, G. (1978) Fishery biology of spiny lobsters Panulirus argus of the Guajira Peninsula of Colombian South America 1969-1970. Proc. Natl. SheNfish. Assoc., 68, 63-74.
Street, R.J. (1969) The New Zealand crayfish Jasus edwardsii (Hutton, 1875): an account of growth, moulting and movements in southern waters, with notes on reproduction and predators. N.Z. Mar. Dep. Fish. Tech. Rep., 30, 1-54.
Talbot, P. (1981) The ovary of the lobster, Homarus americanus. I. Architecture of the mature ovary. J . Ultrastruct. Res., 76, 23543.
7, 1-66.