Embed Size (px)
Citation preview

Spermiogenesis in the Imbricate Alligator Lizard,Barisia imbricata (Reptilia, Squamata, Anguidae)
Kevin M. Gribbins,1* Justin L. Rheubert,2 Katherine Touzinsky,1 Jessica Hanover,1
Caroline L. Matchett,1 Gisela Granados-Gonzalez,3 and Oswaldo Hernandez-Gallegos3
1Department of Biology, Wittenberg University, Springfield, Ohio 455012Department of Biological Sciences, Saint Louis University, St. Louis, Missouri 631033Facultad de Ciencias, UAEMex, Instituto Literario # 100 Centro, Toluca, Estado de Mexico C. P. 50000, Mexico
ABSTRACT Although the events of spermiogenesis arecommonly studied in amniotes, the amount of researchavailable for Squamata is lacking. Many studies havedescribed the morphological characteristics of maturespermatozoa in squamates, but few detail the ultrastruc-tural changes that occur during spermiogenesis. Thisstudy’s purpose is to gain a better understanding of thesubcellular events of spermatid development within theImbricate Alligator Lizard, Barisia imbricata. The mor-phological data presented here represent the first com-plete ultrastructural study of spermiogenesis within thefamily Anguidae. Samples of testes from four specimenscollected on the northwest side of the Nevado de Toluca,Mexico, were prepared using standard techniques fortransmission electron microscopy. Many of the ultrastruc-tural changes occurring during spermiogenesis withinB. imbricata are similar to that of other squamates (i.e.,early acrosome formation, chromatin condensation,flagella formation, annulus present, and a prominentmanchette). However, there are a few unique characteris-tics within B. imbricata spermatids that to date have notbeen described during spermiogenesis in other squa-mates. For example, penetration of the acrosomal granuleinto the subacrosomal space to form the basal plate ofthe perforatorium during round spermatid development,the clover-shaped morphology of the developingnuclear fossa of the flagellum, and the bulbous shape tothe perforatorium are all unique to the ImbricateAlligator Lizard. These anatomical character differencesmay be valuable nontraditional data that along withmore traditional matrices (such as DNA sequences andgross morphological data) may help elucidate phyloge-netic relationships, which are historically consideredcontroversial within Squamata. J. Morphol. 274:603–614,2013. � 2013 Wiley Periodicals, Inc.
KEY WORDS: lizard; spermatids; ultrastructure; Ang-uidae; spermiogenesis
INTRODUCTION
The Barisia genus is underrepresented as far asreproductive data and life history characteristics.Much of what is known about the male reproductivebiology in Barisia is more than 20 years old (Vial andStewart, 1985; Guillette and Casas-Andreu, 1987).The most recent information on the reproductivetracts in this genus concentrates on females with mor-
phological descriptions of the corpus luteum, folliculardevelopment, and the histology of the vagina-cloacalregion (Martınez-Torres et al., 2003, 2006; Sanchez-Martınez et al., 2007; Martınez-Torres et al., 2010).Little is known about the testis, the specific histologi-cal aspects of spermatogenesis, and the germ cell de-velopment strategy within male Barisia imbricata.
Reproductive characters such as morphology ofgerm cell types and understanding of the architec-ture of the testis not only provide important repro-ductive indicators as suggested by Guillette andCasas-Andreu (1987), but also have implications onour understanding of the evolution of reproductivemorphology in amniotes. Spermatozoa have longbeen regarded as one of the most differentiated celltypes within vertebrates. The ultrastructural mor-phology of the mature spermatozoon provides acharacter matrix that shows both variability (syna-pomorphies) and conservative features (often sym-plesiomorphies) across chordate taxa (Jamiesonet al., 1995; Jamieson, 1999). Thus, there have beena multitude of recent studies that focus on the mor-phology of the spermatozoa in reptilian sauropsidsand its relevance as a phylogenetic tool for amnioticsystematics and evolution (Scheltinga et al., 2000;Vieira et al., 2005; Tavares-Bastos et al., 2008;
This article was published online on 8 February 2013. An errorwas subsequently identified. This notice is included in the onlineand print versions to indicate that both have been corrected
Contract grant sponsor: Competitive research grants throughWittenberg University; Contract grant sponsor: PIFI (lizardcollections and research, to G. G.-G. and O. H.-G.); Contract grantsponsor: Projects UAMex 3042/2011FS, 9003/2013CAFF andPROMEP FE022/2012.
*Correspondence to: Kevin Gribbins, Department of Biology,Wittenberg University, PO Box 720, Springfield, OH 45501.E-mail: [email protected]
Received 10 October 2012; Revised 21 November 2012;Accepted 4 December 2012
Published online 8 February 2013 inWiley Online Library (wileyonlinelibrary.com)DOI: 10.1002/jmor.20117
JOURNAL OF MORPHOLOGY 274:603–614 (2013)
� 2013 WILEY PERIODICALS, INC.

Tourmente et al., 2008; Rheubert et al., 2010a;Gribbins and Rheubert, 2011).
Rheubert et al. (2011) proposed that structuralcomponents seen in epididymal spermatozoa (wherea majority of the studies are focused) can be charac-terized in an ontogenic fashion during spermiogene-sis and can be seen in the stages just prior to sper-miation. Previous studies have shown that verte-brates follow the same general three-step process ofacrosome development, nuclear elongation/condensa-tion, and flagellar development during spermiogene-sis (see Russell et al., 1990). A thorough investigationat these various stages reveals minute differencesthat developmentally may lead to more prominentdifferences in the mature spermatozoa. For example,some species do not display the microtubules of themanchette (Rheubert et al., 2011), some species showrough endoplasmic reticulum involvement in acro-some formation (Ferreira and Dolder, 2002), andsome species show variation in nuclear condensation(Gribbins and Rheubert, 2011; Rheubert et al., 2012).
Many morphologists have suggested that thereptilian, specifically in Squamata, spermatozoashow more variations across species (apomorphies)than that of other vertebrate taxa (Jamieson et al.,1995, 1996). Their hypotheses are largely based onthe level of compartmentalization and proteinstratification within the acrosome (Healy andJamieson, 1992; Jamieson, 1999, 2007). The eventsleading to this compartmentalization have notbeen well documented and much of the variabilityis not described within a large number of taxawithin the Reptilia. Thus, each study that providesultrastructural details of either the spermatozoa orspermatids supplements our understanding of thedevelopmental processes involved in producingthese highly differentiated male gametes.
Although some studies provide ultrastructuraldescriptions of various aspects of spermiogenesis(acrosome formation, elongation/condensation, andflagellar development) within the 9,0001 species(Reptile-Database, 2012) of lepidosaurians (Clark,1967; Butler and Gabri, 1984; Hondo et al., 1994; Al-Doki, 2004; Mubarak, 2006), only a few species havecomplete ultrastructural descriptions of spermatiddevelopment across the entire process of spermiogen-esis. These comprehensive studies encompass ninefamilies and include 10 species: Sphenodontidae(Sphenodon punctatus; Healy and Jamieson, 1994),Iguanidae (Iguana iguana; Ferreira and Dolder,2002), Dactyloididae (Norops [Anolis] lineatopus;Rheubert et al., 2011), Phrynosomatidae (Sceloporusbicanthalis; Rheubert et al., 2012), Tropiduridae (Tro-pidurus torquatus; Vieira et al., 2001), Lacertidae(Lacerta vivipara; Courtens and Depieges, 1985),Gekkonidae (Hemidactylus turcicus; Rheubert et al.,2011), Scincidae (Chalcides ocellatus; Carcupinoet al., 1989; Scincella lateralis; Gribbins et al., 2007),and Viperidae (Agkistrodon piscivorus; Gribbinset al., 2009). The anguimorpha clade (Shinisauridae,
Lanthanotidae, Varanidae, Helodermatidae, Xeno-sauridae, Anguidae; sensu Wiens et al., 2012), towhich B. imbricata belongs, has no studies encom-passing spermiogenesis and only a single representa-tive with a mature spermatozoon morphologicaldescription (Varanus gouldii; Oliver et al., 1996).Therefore, the purpose of this study is to provide acomplete ultrastructural description of spermiogene-sis in an anguid lizard in order to supply comparativedata from a family of squamate that has little ultra-structural information about the male gamete. Fur-thermore, the recently hypothesized phylogeneticpositions of anguimorphs (sister to Platynota as pro-posed by Conrad, 2008 and Eckstut et al., 2009 ornested within Toxicofera; proposed by Wiens et al.,2012) makes them an interesting clade comparativelyas they will provide spermatid characters from amore derived squamate taxon that is thus far lackingin spermatozoal developmental characters.
MATERIAL AND METHODSTissue Preparation
Barisia imbricata is an anguid lizard found in central Mexico.Four specimens were collected (scientific collector permit: FAUT0186, SEMARNAT) from montane pine forest, consisting of anunderstory of bunch-grass species, just northwest of the Nevadode Toluca at an elevation of 2,590 m (198 090 03@ N, 998 380 00@W) in August 2010 and 2011. Lizards were sacrificed using a0.1% injection of sodium pentobarbitol, testes were thenremoved, cut into small pieces (2–3 mm blocks), submerged inTrumps fixative, and stored under refrigeration. The specimenswere deposited in the Centro de Investigacion en Recursos Bio-ticos, Facultad de Ciencias, UAEMex.
After refrigeration for over 48 h, five small blocks of testicular tis-sues from each testis were washed twice with cacodylate buffer (pH7.4) for 20 min. Tissues were then post-fixed in 2% osmium tetrox-ide for 2 h, washed again in cacodylate buffer (pH 7.4) three timesfor 20 min, dehydrated in increasing concentrations of ethanol(70%, 85%, 95% X2, and 100% X2), and finally cleared twice with10 min. treatments of propylene oxide. The testicular pieces thenwere slowly introduced to a graded series of epoxy resin (Embed812, EMS, Hatfield, PA) (2:1 and 1:1 solutions of propylene oxide:epoxy resin). The final step in the epoxy series was a 24-hour incu-bation in pure Embed 812. Fresh resin was prepared and testiculartissues were embedded in small beam capsules and cured for 48 hat 708C in a Fisher isotemperature vacuum oven (Fisher Scientific,Pittsburg, PA). Once hardened, sections (90 nm) were cut from theblocks via a diamond knife (DDK, Wilmington, DE) on a Leica UC7ultramicrotome (Leica Microsystems, Buffalo Grove, IL). Sectionswere then placed on copper grids and stained for 18 min with ura-nyl acetate and then for 5 min with lead citrate.
Ultrastructural Analysis
The samples were viewed using a JEOL JEM-1200EX IItransmission electron microscope. Spermatids and organellesassociated with spermiogenesis were identified and micrographswere taken with a Gatan 785 Erlangshen digital camera(Gatan, Warrendale, PA). The subsequent micrographs were an-alyzed and composite plates were constructed using Adobe Pho-toshop CS software (Adobe Systems, San Jose, CA).
RESULTS
The beginning stage of spermiogenesis inB. imbricata is marked by the development of an
604 K.M. GRIBBINS ET AL.
Journal of Morphology
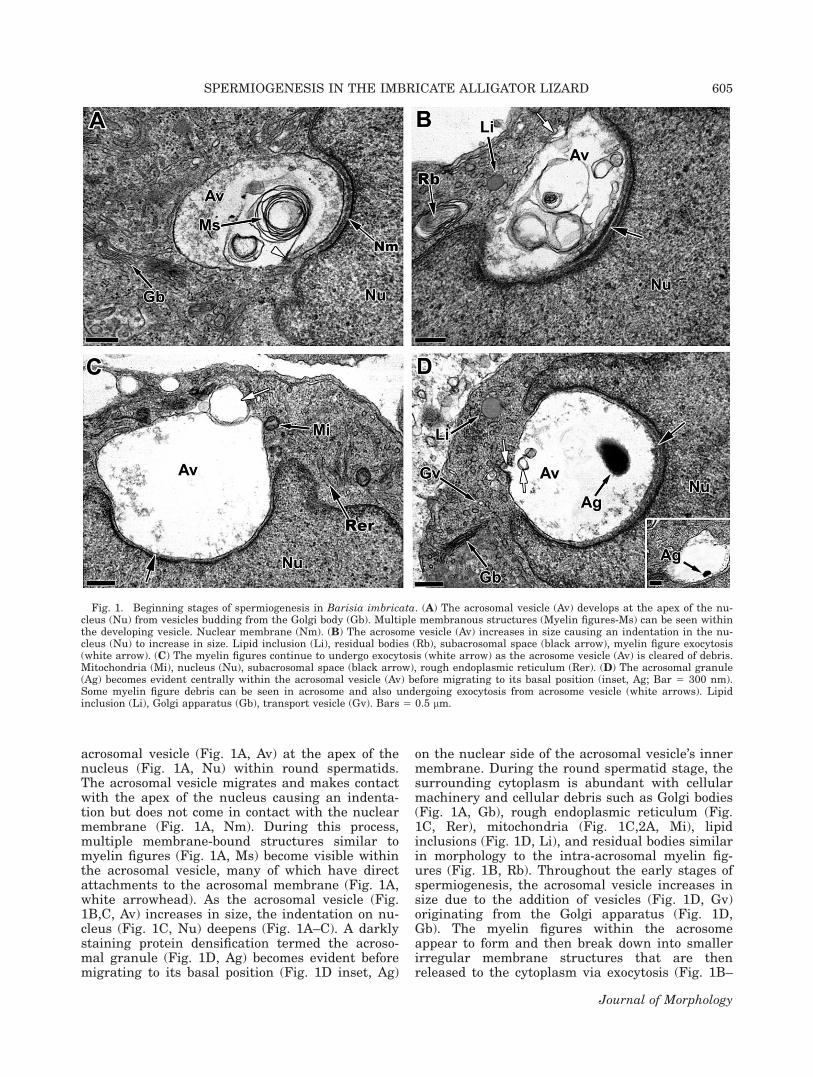
acrosomal vesicle (Fig. 1A, Av) at the apex of thenucleus (Fig. 1A, Nu) within round spermatids.The acrosomal vesicle migrates and makes contactwith the apex of the nucleus causing an indenta-tion but does not come in contact with the nuclearmembrane (Fig. 1A, Nm). During this process,multiple membrane-bound structures similar tomyelin figures (Fig. 1A, Ms) become visible withinthe acrosomal vesicle, many of which have directattachments to the acrosomal membrane (Fig. 1A,white arrowhead). As the acrosomal vesicle (Fig.1B,C, Av) increases in size, the indentation on nu-cleus (Fig. 1C, Nu) deepens (Fig. 1A–C). A darklystaining protein densification termed the acroso-mal granule (Fig. 1D, Ag) becomes evident beforemigrating to its basal position (Fig. 1D inset, Ag)
on the nuclear side of the acrosomal vesicle’s innermembrane. During the round spermatid stage, thesurrounding cytoplasm is abundant with cellularmachinery and cellular debris such as Golgi bodies(Fig. 1A, Gb), rough endoplasmic reticulum (Fig.1C, Rer), mitochondria (Fig. 1C,2A, Mi), lipidinclusions (Fig. 1D, Li), and residual bodies similarin morphology to the intra-acrosomal myelin fig-ures (Fig. 1B, Rb). Throughout the early stages ofspermiogenesis, the acrosomal vesicle increases insize due to the addition of vesicles (Fig. 1D, Gv)originating from the Golgi apparatus (Fig. 1D,Gb). The myelin figures within the acrosomeappear to form and then break down into smallerirregular membrane structures that are thenreleased to the cytoplasm via exocytosis (Fig. 1B–
Fig. 1. Beginning stages of spermiogenesis in Barisia imbricata. (A) The acrosomal vesicle (Av) develops at the apex of the nu-cleus (Nu) from vesicles budding from the Golgi body (Gb). Multiple membranous structures (Myelin figures-Ms) can be seen withinthe developing vesicle. Nuclear membrane (Nm). (B) The acrosome vesicle (Av) increases in size causing an indentation in the nu-cleus (Nu) to increase in size. Lipid inclusion (Li), residual bodies (Rb), subacrosomal space (black arrow), myelin figure exocytosis(white arrow). (C) The myelin figures continue to undergo exocytosis (white arrow) as the acrosome vesicle (Av) is cleared of debris.Mitochondria (Mi), nucleus (Nu), subacrosomal space (black arrow), rough endoplasmic reticulum (Rer). (D) The acrosomal granule(Ag) becomes evident centrally within the acrosomal vesicle (Av) before migrating to its basal position (inset, Ag; Bar 5 300 nm).Some myelin figure debris can be seen in acrosome and also undergoing exocytosis from acrosome vesicle (white arrows). Lipidinclusion (Li), Golgi apparatus (Gb), transport vesicle (Gv). Bars 5 0.5 lm.
SPERMIOGENESIS IN THE IMBRICATE ALLIGATOR LIZARD 605
Journal of Morphology
D, white arrows). Most of these myelin figures andexocytotic events occur during the first two sper-matid steps of early acrosome formation.
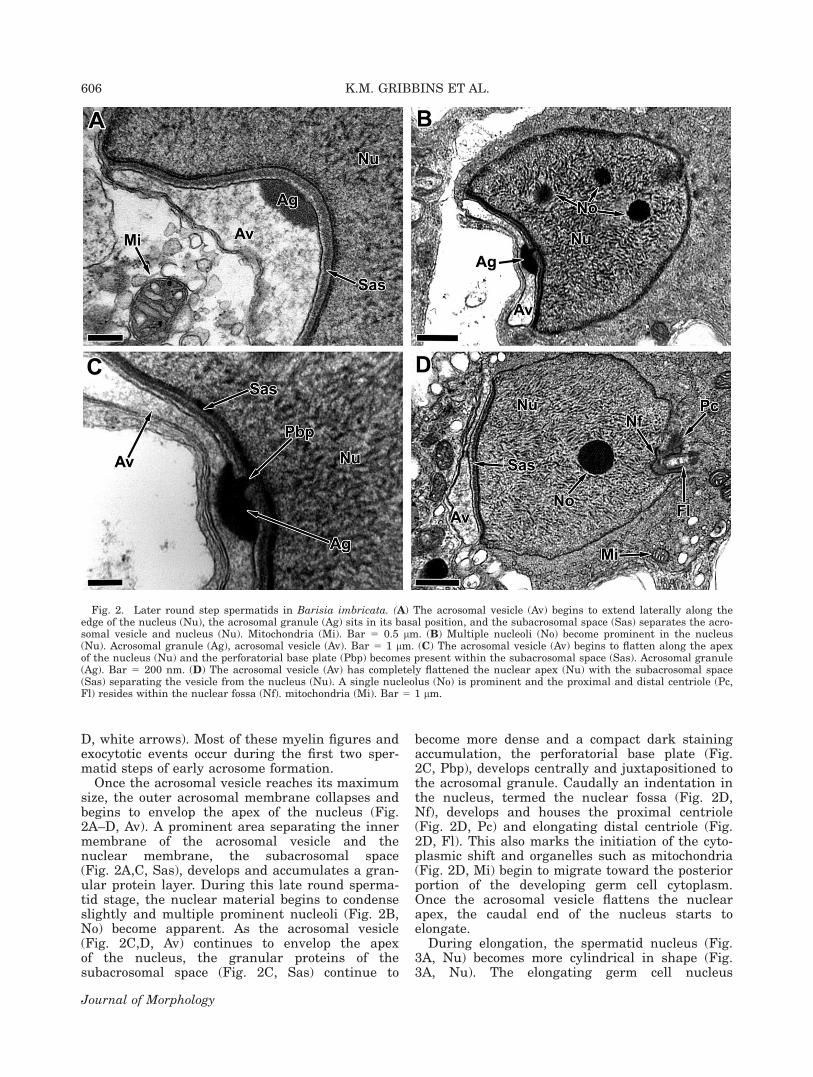
Once the acrosomal vesicle reaches its maximumsize, the outer acrosomal membrane collapses andbegins to envelop the apex of the nucleus (Fig.2A–D, Av). A prominent area separating the innermembrane of the acrosomal vesicle and thenuclear membrane, the subacrosomal space(Fig. 2A,C, Sas), develops and accumulates a gran-ular protein layer. During this late round sperma-tid stage, the nuclear material begins to condenseslightly and multiple prominent nucleoli (Fig. 2B,No) become apparent. As the acrosomal vesicle(Fig. 2C,D, Av) continues to envelop the apexof the nucleus, the granular proteins of thesubacrosomal space (Fig. 2C, Sas) continue to
become more dense and a compact dark stainingaccumulation, the perforatorial base plate (Fig.2C, Pbp), develops centrally and juxtapositioned tothe acrosomal granule. Caudally an indentation inthe nucleus, termed the nuclear fossa (Fig. 2D,Nf), develops and houses the proximal centriole(Fig. 2D, Pc) and elongating distal centriole (Fig.2D, Fl). This also marks the initiation of the cyto-plasmic shift and organelles such as mitochondria(Fig. 2D, Mi) begin to migrate toward the posteriorportion of the developing germ cell cytoplasm.Once the acrosomal vesicle flattens the nuclearapex, the caudal end of the nucleus starts toelongate.
During elongation, the spermatid nucleus (Fig.3A, Nu) becomes more cylindrical in shape (Fig.3A, Nu). The elongating germ cell nucleus
Fig. 2. Later round step spermatids in Barisia imbricata. (A) The acrosomal vesicle (Av) begins to extend laterally along theedge of the nucleus (Nu), the acrosomal granule (Ag) sits in its basal position, and the subacrosomal space (Sas) separates the acro-somal vesicle and nucleus (Nu). Mitochondria (Mi). Bar 5 0.5 lm. (B) Multiple nucleoli (No) become prominent in the nucleus(Nu). Acrosomal granule (Ag), acrosomal vesicle (Av). Bar 5 1 lm. (C) The acrosomal vesicle (Av) begins to flatten along the apexof the nucleus (Nu) and the perforatorial base plate (Pbp) becomes present within the subacrosomal space (Sas). Acrosomal granule(Ag). Bar 5 200 nm. (D) The acrosomal vesicle (Av) has completely flattened the nuclear apex (Nu) with the subacrosomal space(Sas) separating the vesicle from the nucleus (Nu). A single nucleolus (No) is prominent and the proximal and distal centriole (Pc,Fl) resides within the nuclear fossa (Nf). mitochondria (Mi). Bar 5 1 lm.
606 K.M. GRIBBINS ET AL.
Journal of Morphology
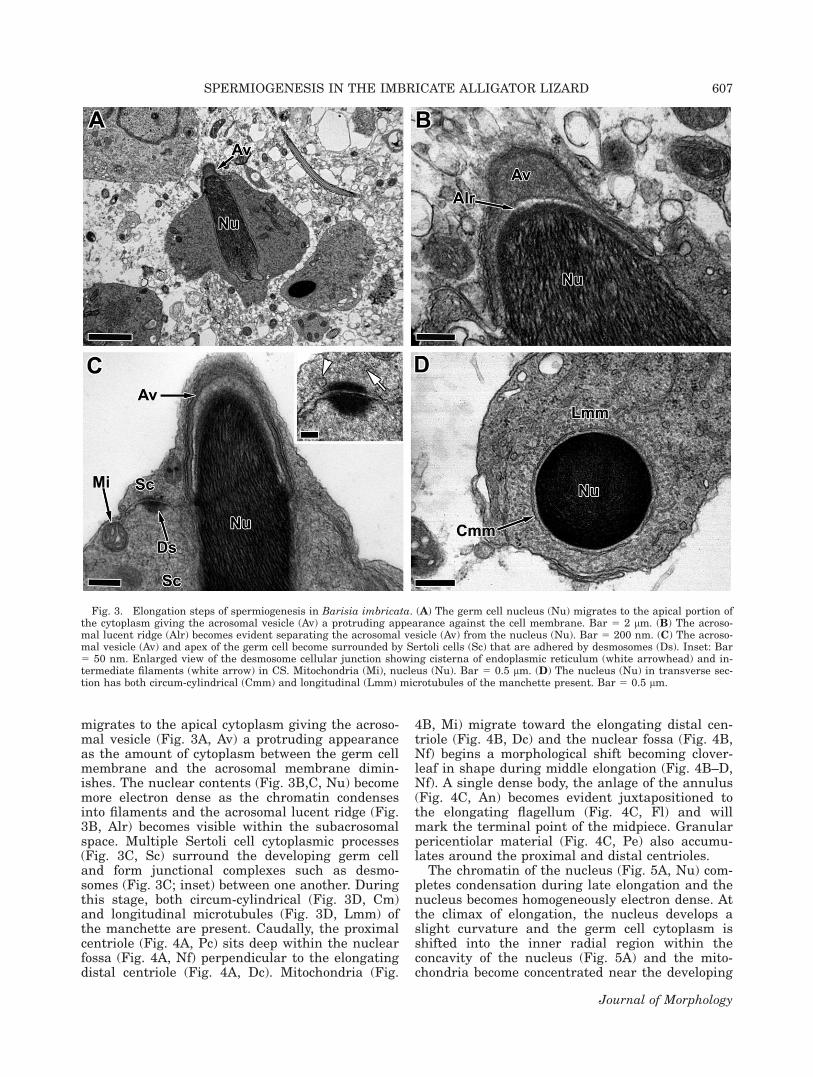
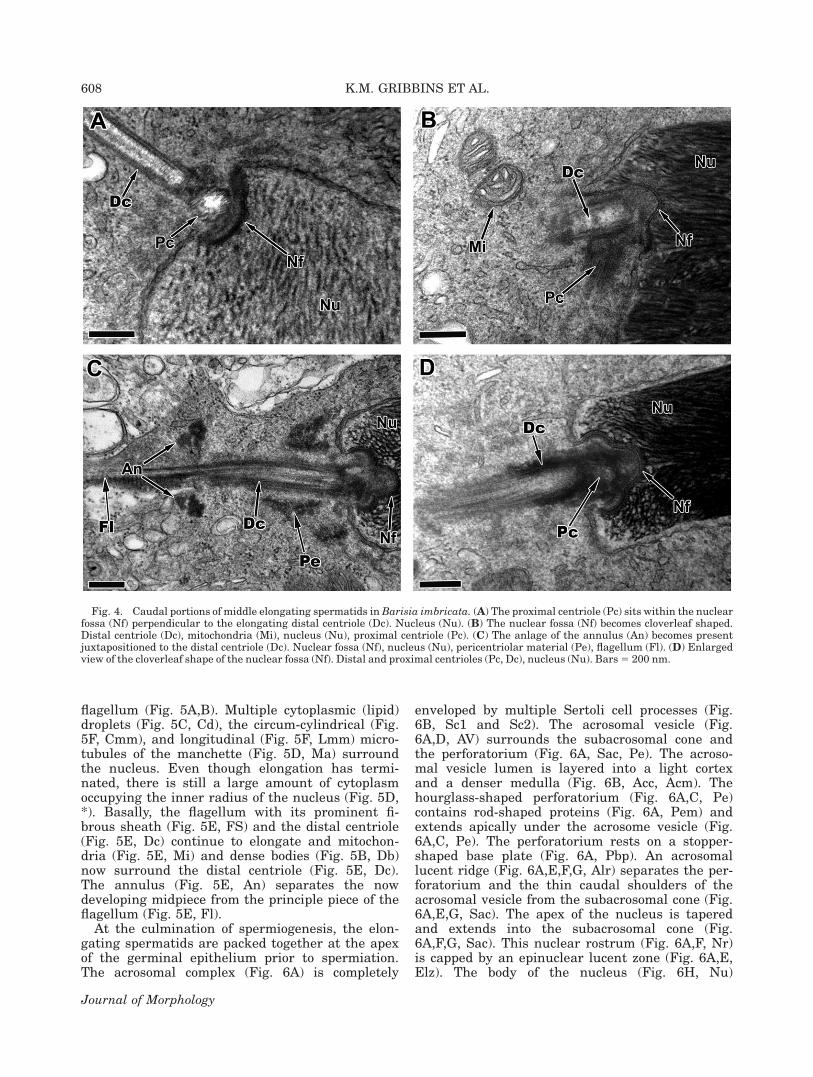
migrates to the apical cytoplasm giving the acroso-mal vesicle (Fig. 3A, Av) a protruding appearanceas the amount of cytoplasm between the germ cellmembrane and the acrosomal membrane dimin-ishes. The nuclear contents (Fig. 3B,C, Nu) becomemore electron dense as the chromatin condensesinto filaments and the acrosomal lucent ridge (Fig.3B, Alr) becomes visible within the subacrosomalspace. Multiple Sertoli cell cytoplasmic processes(Fig. 3C, Sc) surround the developing germ celland form junctional complexes such as desmo-somes (Fig. 3C; inset) between one another. Duringthis stage, both circum-cylindrical (Fig. 3D, Cm)and longitudinal microtubules (Fig. 3D, Lmm) ofthe manchette are present. Caudally, the proximalcentriole (Fig. 4A, Pc) sits deep within the nuclearfossa (Fig. 4A, Nf) perpendicular to the elongatingdistal centriole (Fig. 4A, Dc). Mitochondria (Fig.
4B, Mi) migrate toward the elongating distal cen-triole (Fig. 4B, Dc) and the nuclear fossa (Fig. 4B,Nf) begins a morphological shift becoming clover-leaf in shape during middle elongation (Fig. 4B–D,Nf). A single dense body, the anlage of the annulus(Fig. 4C, An) becomes evident juxtapositioned tothe elongating flagellum (Fig. 4C, Fl) and willmark the terminal point of the midpiece. Granularpericentiolar material (Fig. 4C, Pe) also accumu-lates around the proximal and distal centrioles.
The chromatin of the nucleus (Fig. 5A, Nu) com-pletes condensation during late elongation and thenucleus becomes homogeneously electron dense. Atthe climax of elongation, the nucleus develops aslight curvature and the germ cell cytoplasm isshifted into the inner radial region within theconcavity of the nucleus (Fig. 5A) and the mito-chondria become concentrated near the developing
Fig. 3. Elongation steps of spermiogenesis in Barisia imbricata. (A) The germ cell nucleus (Nu) migrates to the apical portion ofthe cytoplasm giving the acrosomal vesicle (Av) a protruding appearance against the cell membrane. Bar 5 2 lm. (B) The acroso-mal lucent ridge (Alr) becomes evident separating the acrosomal vesicle (Av) from the nucleus (Nu). Bar 5 200 nm. (C) The acroso-mal vesicle (Av) and apex of the germ cell become surrounded by Sertoli cells (Sc) that are adhered by desmosomes (Ds). Inset: Bar5 50 nm. Enlarged view of the desmosome cellular junction showing cisterna of endoplasmic reticulum (white arrowhead) and in-termediate filaments (white arrow) in CS. Mitochondria (Mi), nucleus (Nu). Bar 5 0.5 lm. (D) The nucleus (Nu) in transverse sec-tion has both circum-cylindrical (Cmm) and longitudinal (Lmm) microtubules of the manchette present. Bar 5 0.5 lm.
SPERMIOGENESIS IN THE IMBRICATE ALLIGATOR LIZARD 607
Journal of Morphology
flagellum (Fig. 5A,B). Multiple cytoplasmic (lipid)droplets (Fig. 5C, Cd), the circum-cylindrical (Fig.5F, Cmm), and longitudinal (Fig. 5F, Lmm) micro-tubules of the manchette (Fig. 5D, Ma) surroundthe nucleus. Even though elongation has termi-nated, there is still a large amount of cytoplasmoccupying the inner radius of the nucleus (Fig. 5D,*). Basally, the flagellum with its prominent fi-brous sheath (Fig. 5E, FS) and the distal centriole(Fig. 5E, Dc) continue to elongate and mitochon-dria (Fig. 5E, Mi) and dense bodies (Fig. 5B, Db)now surround the distal centriole (Fig. 5E, Dc).The annulus (Fig. 5E, An) separates the nowdeveloping midpiece from the principle piece of theflagellum (Fig. 5E, Fl).
At the culmination of spermiogenesis, the elon-gating spermatids are packed together at the apexof the germinal epithelium prior to spermiation.The acrosomal complex (Fig. 6A) is completely
enveloped by multiple Sertoli cell processes (Fig.6B, Sc1 and Sc2). The acrosomal vesicle (Fig.6A,D, AV) surrounds the subacrosomal cone andthe perforatorium (Fig. 6A, Sac, Pe). The acroso-mal vesicle lumen is layered into a light cortexand a denser medulla (Fig. 6B, Acc, Acm). Thehourglass-shaped perforatorium (Fig. 6A,C, Pe)contains rod-shaped proteins (Fig. 6A, Pem) andextends apically under the acrosome vesicle (Fig.6A,C, Pe). The perforatorium rests on a stopper-shaped base plate (Fig. 6A, Pbp). An acrosomallucent ridge (Fig. 6A,E,F,G, Alr) separates the per-foratorium and the thin caudal shoulders of theacrosomal vesicle from the subacrosomal cone (Fig.6A,E,G, Sac). The apex of the nucleus is taperedand extends into the subacrosomal cone (Fig.6A,F,G, Sac). This nuclear rostrum (Fig. 6A,F, Nr)is capped by an epinuclear lucent zone (Fig. 6A,E,Elz). The body of the nucleus (Fig. 6H, Nu)
Fig. 4. Caudal portions of middle elongating spermatids in Barisia imbricata. (A) The proximal centriole (Pc) sits within the nuclearfossa (Nf) perpendicular to the elongating distal centriole (Dc). Nucleus (Nu). (B) The nuclear fossa (Nf) becomes cloverleaf shaped.Distal centriole (Dc), mitochondria (Mi), nucleus (Nu), proximal centriole (Pc). (C) The anlage of the annulus (An) becomes presentjuxtapositioned to the distal centriole (Dc). Nuclear fossa (Nf), nucleus (Nu), pericentriolar material (Pe), flagellum (Fl). (D) Enlargedview of the cloverleaf shape of the nuclear fossa (Nf). Distal and proximal centrioles (Pc, Dc), nucleus (Nu). Bars 5 200 nm.
608 K.M. GRIBBINS ET AL.
Journal of Morphology
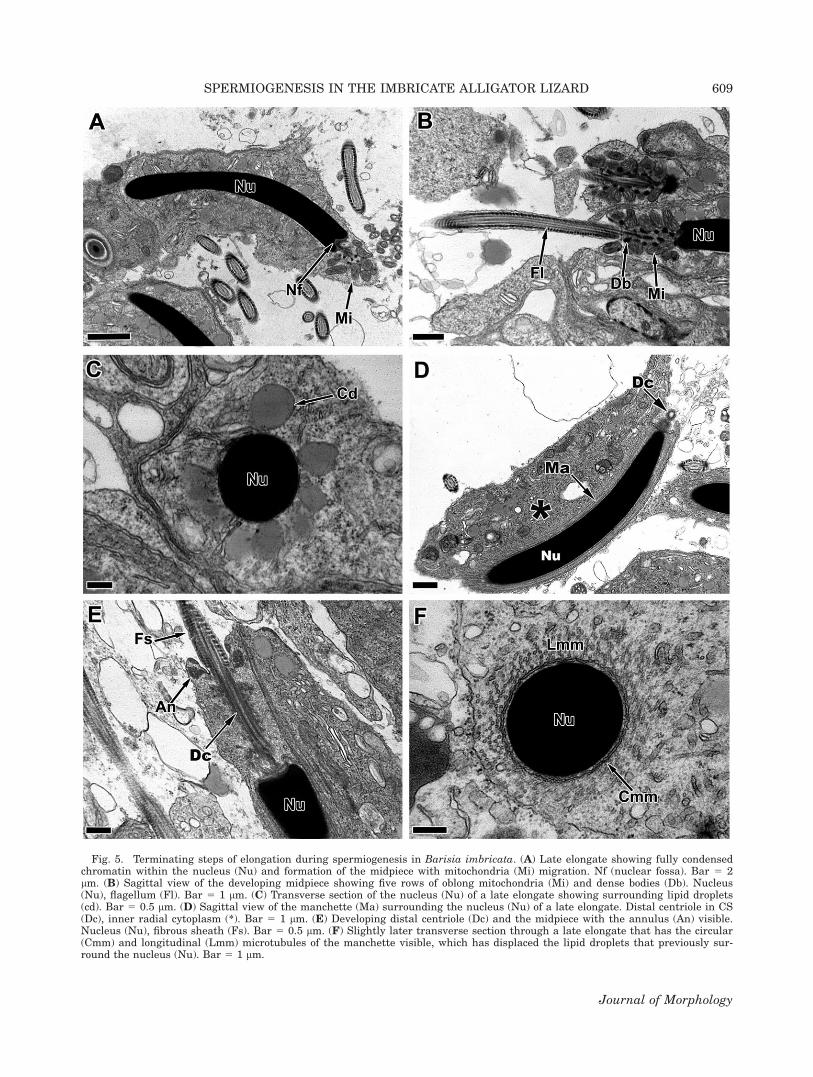
Fig. 5. Terminating steps of elongation during spermiogenesis in Barisia imbricata. (A) Late elongate showing fully condensedchromatin within the nucleus (Nu) and formation of the midpiece with mitochondria (Mi) migration. Nf (nuclear fossa). Bar 5 2lm. (B) Sagittal view of the developing midpiece showing five rows of oblong mitochondria (Mi) and dense bodies (Db). Nucleus(Nu), flagellum (Fl). Bar 5 1 lm. (C) Transverse section of the nucleus (Nu) of a late elongate showing surrounding lipid droplets(cd). Bar 5 0.5 lm. (D) Sagittal view of the manchette (Ma) surrounding the nucleus (Nu) of a late elongate. Distal centriole in CS(Dc), inner radial cytoplasm (*). Bar 5 1 lm. (E) Developing distal centriole (Dc) and the midpiece with the annulus (An) visible.Nucleus (Nu), fibrous sheath (Fs). Bar 5 0.5 lm. (F) Slightly later transverse section through a late elongate that has the circular(Cmm) and longitudinal (Lmm) microtubules of the manchette visible, which has displaced the lipid droplets that previously sur-round the nucleus (Nu). Bar 5 1 lm.
SPERMIOGENESIS IN THE IMBRICATE ALLIGATOR LIZARD 609
Journal of Morphology
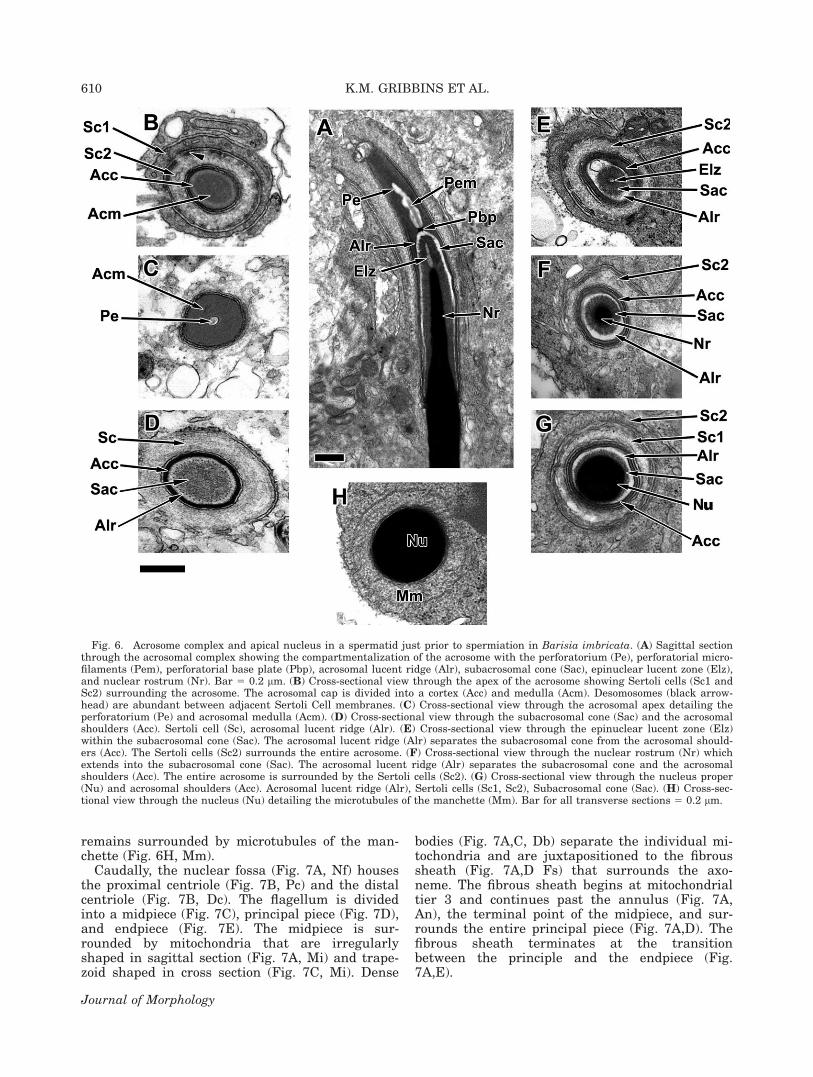
remains surrounded by microtubules of the man-chette (Fig. 6H, Mm).
Caudally, the nuclear fossa (Fig. 7A, Nf) housesthe proximal centriole (Fig. 7B, Pc) and the distalcentriole (Fig. 7B, Dc). The flagellum is dividedinto a midpiece (Fig. 7C), principal piece (Fig. 7D),and endpiece (Fig. 7E). The midpiece is sur-rounded by mitochondria that are irregularlyshaped in sagittal section (Fig. 7A, Mi) and trape-zoid shaped in cross section (Fig. 7C, Mi). Dense
bodies (Fig. 7A,C, Db) separate the individual mi-tochondria and are juxtapositioned to the fibroussheath (Fig. 7A,D Fs) that surrounds the axo-neme. The fibrous sheath begins at mitochondrialtier 3 and continues past the annulus (Fig. 7A,An), the terminal point of the midpiece, and sur-rounds the entire principal piece (Fig. 7A,D). Thefibrous sheath terminates at the transitionbetween the principle and the endpiece (Fig.7A,E).
Fig. 6. Acrosome complex and apical nucleus in a spermatid just prior to spermiation in Barisia imbricata. (A) Sagittal sectionthrough the acrosomal complex showing the compartmentalization of the acrosome with the perforatorium (Pe), perforatorial micro-filaments (Pem), perforatorial base plate (Pbp), acrosomal lucent ridge (Alr), subacrosomal cone (Sac), epinuclear lucent zone (Elz),and nuclear rostrum (Nr). Bar 5 0.2 lm. (B) Cross-sectional view through the apex of the acrosome showing Sertoli cells (Sc1 andSc2) surrounding the acrosome. The acrosomal cap is divided into a cortex (Acc) and medulla (Acm). Desomosomes (black arrow-head) are abundant between adjacent Sertoli Cell membranes. (C) Cross-sectional view through the acrosomal apex detailing theperforatorium (Pe) and acrosomal medulla (Acm). (D) Cross-sectional view through the subacrosomal cone (Sac) and the acrosomalshoulders (Acc). Sertoli cell (Sc), acrosomal lucent ridge (Alr). (E) Cross-sectional view through the epinuclear lucent zone (Elz)within the subacrosomal cone (Sac). The acrosomal lucent ridge (Alr) separates the subacrosomal cone from the acrosomal should-ers (Acc). The Sertoli cells (Sc2) surrounds the entire acrosome. (F) Cross-sectional view through the nuclear rostrum (Nr) whichextends into the subacrosomal cone (Sac). The acrosomal lucent ridge (Alr) separates the subacrosomal cone and the acrosomalshoulders (Acc). The entire acrosome is surrounded by the Sertoli cells (Sc2). (G) Cross-sectional view through the nucleus proper(Nu) and acrosomal shoulders (Acc). Acrosomal lucent ridge (Alr), Sertoli cells (Sc1, Sc2), Subacrosomal cone (Sac). (H) Cross-sec-tional view through the nucleus (Nu) detailing the microtubules of the manchette (Mm). Bar for all transverse sections 5 0.2 lm.
610 K.M. GRIBBINS ET AL.
Journal of Morphology
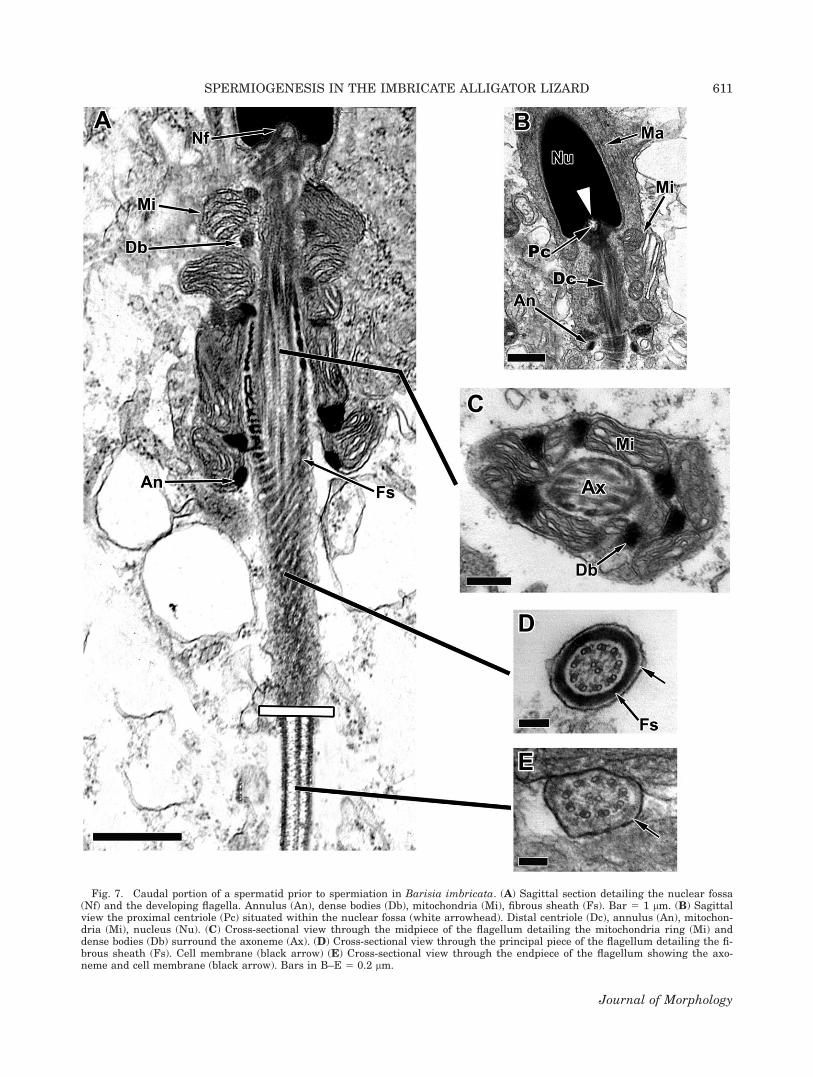
Fig. 7. Caudal portion of a spermatid prior to spermiation in Barisia imbricata. (A) Sagittal section detailing the nuclear fossa(Nf) and the developing flagella. Annulus (An), dense bodies (Db), mitochondria (Mi), fibrous sheath (Fs). Bar 5 1 lm. (B) Sagittalview the proximal centriole (Pc) situated within the nuclear fossa (white arrowhead). Distal centriole (Dc), annulus (An), mitochon-dria (Mi), nucleus (Nu). (C) Cross-sectional view through the midpiece of the flagellum detailing the mitochondria ring (Mi) anddense bodies (Db) surround the axoneme (Ax). (D) Cross-sectional view through the principal piece of the flagellum detailing the fi-brous sheath (Fs). Cell membrane (black arrow) (E) Cross-sectional view through the endpiece of the flagellum showing the axo-neme and cell membrane (black arrow). Bars in B–E 5 0.2 lm.
SPERMIOGENESIS IN THE IMBRICATE ALLIGATOR LIZARD 611
Journal of Morphology

DISCUSSION
The ontogeny of spermatid development withinB. imbricata is similar to that described in othersquamates. Many of the cellular components de-velop in a similar way within the spermatids dur-ing spermiogenesis and most morphological com-parisons suggest that at least some of the sper-matozoal anatomy is highly conserved inSquamata (see Gribbins and Rheubert, 2011;Rheubert et al., 2011, 2012). However, B. imbri-cata has unique features that have not beenobserved before in the squamates studied to dateduring spermatid development. The importance ofsuch apomorphic or possibly autapomorphic char-acters is difficult to evaluate since this is only the11th species, which represents merely 10 families(when B. imbricata is included) within the Squa-mata and Lepidosauria that have comprehensiveultrastructural coverage of spermatid morphologyduring spermiogenesis. Since previous authorshave also stated that spermatid and/or spermato-zoal ultrastructure may be species-specific (Rheu-bert et al., 2012), these data may help furthersupport the taxonomic treatment of the species ofthe B. imbricata complex (B. ciliaris, B. imbri-cata, B. jonesi, and B. planiforms), which are allconsidered sibling species presently (Smith et al.,2002). Though our limited spermiogenic data alsosuggest conservation, like the spermatozoa, inmany aspects of spermatid ultrastructure the dif-ferences seen could infer evolutionary trends orrelationships within squamates and/or other lepi-dosaurians. For example, Jamieson et al. (1995,1996), Vieira et al. (2005), and Rheubert et al.(2010a,b) all suggest that a single extranuclearperforatorium is a synapomorphy for the Squa-mata (which our present study supports). Never-theless, with so few families of lepidosaurianspresently represented, these data remain arbi-trary and preliminary in nature. More taxa suchas B. imbricata representing families that havenot been studied need to be added to this datamatrix to increase the robustness of such hypoth-eses. Also, species within a genus could beexplored using the ultrastructure of sperm devel-opment to see if differences can be seen specifi-cally between species within genera.
Interestingly, the acrosomal complex has alsobeen suggested by other studies to be highly con-served in squamates and probably lepidosaurianspermatozoa (Jamieson, 1999; Vieira et al., 2005;Rheubert et al., 2010a,b, 2011). However, in B.imbricata the most novel character differences inits spermatid’s ultrastructure occur in the acroso-mal complex. The most conspicuous aberrationwithin the acrosome vesicle of B. imbricata is thenumber of myelin figures and membranous debriswithin the vesicle. Though myelin figures and lossof granular material from the acrosome vesicle
during early and middle spermatid developmenthas been suggested previously in Sphenodon punc-tatus (Healy and Jamieson, 1994) and Sceloporusbicanthalis (Rheubert et al., 2012), neither studyshowed the excessive amount of membrane debristhat B. imbricata has early in spermiogenesis. Themembranous myelin figures in some cases occupiedmost of the lumen within the developing acrosomalvesicle and in rare instances even displaced theacrosomal granule peripherally within the vesicle.The high frequency of observation of the myelinfigures provided evidence that they have commonorigins and attachments to the acrosomal mem-brane. These membrane structures appear to windup into layers from the developing acrosomalvesicle’s cell membrane, which has its largestgrowth phase at this time. We hypothesize thatthis is an efficient mechanism to rid the acrosomalvesicle of excess membrane that is delivered bythe multitude of transport vesicles supplied to theacrosomal vesicle via the Golgi apparatus. As theacrosome continues to develop during the roundstage of spermiogenesis, visual evidence of the exo-cytosis of these degrading myelin figures is appa-rent and the newly formed cytoplasmic residualbodies accumulate near the apex of the spermatidnucleus (see Fig. 1, white arrows).
Another unique feature to early spermiogenesisin B. imbricata is the development of a proteinrich accumulation centrally and juxtapositioned tothe basal acrosome granule within the subacroso-mal space. We hypothesize this to be the earlystage of perforatorial basal plate formation thatwill provide the basal support to the perforatoriumin later staged spermatids and within the sperma-tozoa. Typically, in lepidosaurians studied to date(see Rheubert et al., 2011) the perforatorial basalplate is not seen until late elongation. Also, endo-plasmic reticulum (Fig. 1C, Rer) is seen close tothe developing acrosome complex; however, no vis-ual observations from this study suggest it playsany role in acrosome formation as proposed byFerreira and Dolder (2002) for Iguana iguana. Themajority of morphological observations of theacrosome complex during spermiogenesis in B.imbricata are similar to what has been describedin other squamates (Clark, 1967; Furieri, 1970;Healy and Jamieson, 1992; Jamieson, 1999; Grib-bins et al., 2007, 2009; Rheubert et al., 2011,2012). These common characteristics included: anelongated nuclear rostrum into the acrosome com-plex, a highly compartmentalized acrosome com-plex with an epinuclear lucent zone, subacrosomalspace, subacrosomal lucent ridge, basal plate, andperforatorium. Lastly, the perforatorium is bulbousat its base (vase-like) within the B. imbricata,which is unique to the more cylinder-like shape ofthe perforatorium in most squamates (Gribbinsand Rheubert, 2011). There also appears to bemultiple arrays of microfilaments within the perfo-
612 K.M. GRIBBINS ET AL.
Journal of Morphology

ratorium in sagittal view, which could be actin fila-ments as hypothesized by Ferreira et al. (2006).
Chromatin condensation begins early in B. imbri-cata during the mid-round spermatid stage and thenucleus at this time has very distinct round granu-lar nucleoli. Condensation of the chromatin is fila-mentous even at this early stage, which is differentthan some squamates and allows chromatin conden-sation to occur in a granular fashion with later chro-matin twisting leading to filamentous condensationsuch as seen in Iguana iguana (Ferreira and Dolder,2002), Hemidactylus turcicus (Rheubert et al.,2011), Agkistrodon piscivorus (Gribbins et al., 2009),and Scincella lateralis (Gribbins et al., 2007). Themanchette of B. imbricata is similar to thatdescribed in other lepidosaurians, in that there iscircular and longitudinal microtubules that makeup its scaffolding (Gribbins et al., 2007). Two ormore Sertoli cells typically arrange themselvesaround developing elongates within B. imbricatasimilar to that described in other squamates (Rheu-bert et al., 2011, 2012). This often leads to desmo-some junctions between Sertoli cells, which mostlikely anchor or secure the germ cells during thefinal stages of maturation within the apex of the ger-minal epithelium.
During elongation, the acrosome complexmatures and chromatin condenses fully within theelongate’s nucleus, all of which are similar to thatof other squamates studied to date (Gribbins andRheubert, 2011; Rheubert et al., 2012). One excep-tion to normal elongation is the development of aclover-shaped nuclear fossa during middle elonga-tion, which has not been reported previously. Theproximal and distal centrioles reside within thefossa comparable to all other squamates and verte-brates studied to date (Gribbins and Rheubert,2011). As the distal centriole elongates to form theflagellum, a distinct annular ring forms markingthe termination point of the midpiece. The elongat-ing flagellum extending from the distal centrioleretains the 9 1 2 microtubule arrangement that istypical of amniotes (Fawcett, 1970). Mitochondriamigrate to the developing midpiece during lateelongation and four to five tiers of mitochondriaare observed within the maturing midpiece sagit-tally within B. imbricata. As the principle pieceforms during elongation, a fibrous sheath is laiddown around the developing axoneme, as is thecase in every species of the Amniota previouslystudied (Healy and Jamieson, 1992). Also, the fi-brous sheath, as in most squamates, penetratesthe length of most of the midpiece, which is a sug-gested synapomorphy for Squamata (Healy andJamieson, 1992; Jamieson, 1999). Within B. imbri-cata, the fibrous sheath begins at mitochondriatier 3 similar to Hemidactylus turcicus (Rheubertet al., 2011) and some teiids (Tupinambis meria-nae, Tavares-Bastos et al., 2002; Crocodilurusamazonicus, Colli et al., 2007; and Dracaena guia-
nensis, Colli et al., 2007). The midpiece in trans-verse section within B. imbricata is an incompletering of mitochondria that are broken up by densebodies, which again is common in Squamata(Jamieson et al., 1996; Ferreria and Dolder, 2002;Gribbins et al., 2007; Rheubert et al., 2011).
The axoneme with its surrounding fibroussheath continues past the midpiece in the Imbri-cate Alligator Lizard as the principle piece. Theperipheral fibers 3 and 8 do continue into the prin-ciple piece but are not as conspicuous as somesquamates (Jamieson et al., 1996; Oliver et al.,1996; Teixeira et al., 1999). The termination of thedeveloping elongating flagellum is the endpiece,which has been found in all lepidosaurians andamniotes studied to date (Jamieson et al., 1996;Jamieson, 1999). The endpiece has no associatedfibrous sheath but retains the 9 1 2 microtubulearrangement of the flagellar axoneme.
Overall, spermiogenesis in B. imbricata is simi-lar to that of other lepidosaurians and all otheramniotes. However, there are a few unique ultra-structural features to the spermatids of B. imbri-cata that may either be autapomorphies for thisspecies or possibly synapomorphies for the familyAnguidae to which the Imbricate Alligator Lizardbelongs. They include their clover-shaped nuclearfossa during elongation, basal plate formation dur-ing early round spermatid development, and thebulbous perforatorium. However, caution must bemaintained until the spermatozoon is examined inthis lizard ultrastructurally and more specieswithin the Anguimorpha clade and other squamatetaxa are studied for both spermatid and spermato-zoal ultrastructure.
LITERATURE CITED
Al-Doki OA. 2004. Electron microscopic study of sperm tail dif-ferentiation in the lizard Bunopus tuberculatus (Squamata,Reptilia). Saudian J Bio Sci 11:111–117.
Butler RD, Gabri MS. 1984. Structure and development of thesperm head in the lizard Podarcis (Lacerta) taurica. J UltraRes 88:261–274.
Carcupino M, Corso G, Pala M. 1989. Spermiogenesis in Chal-cides ocellatus tiligugu (Gmelin) (Squamata, Scincidae): Anelectron microscope study. Boll Zool 56:119–124.
Clark AW. 1967. Some aspects of spermiogenesis in a lizard. AmJ Anat 121:369–400.
Colli GR, Teixeira RD, Schelting DM, Mesquita DO, Wieder-hecker HC, Bao SN. 2007. Comparative study of sperm ultra-structure of five species of teiid lizards (Teiidae, Squamata),and Cercosaura ocellata (Gymnophthalmidae, Squamata). Tis-sue Cell 39:59–78.
Conrad Jl. 2008. Phylogeny and systematics of Squamata (Rep-tilia) based on morphology. Bull Am Mus Nat Hist 310:1–182.
Courtens JL, Depieges A. 1985. Spermiogenesis of Lacertavivipara. J Ultra Res 90:203–220.
Eckstut ME, Sever DM, White ME, Crother BI. 2009. Phyloge-netic analysis of sperm storage in female squamates. In: Ani-mal Reproduction: New Research Developments, LT Dahnof(ed). Nova Science Publishers, Inc., Hauppauge, NY. pp 185–218.
SPERMIOGENESIS IN THE IMBRICATE ALLIGATOR LIZARD 613
Journal of Morphology

Fawcett DW. 1970. A comparative view of sperm ultrastructure.Biol Reprod 2:90–127.
Ferreira A, Dolder H. 2002. Ultrastructural analysis of spermio-genesis in Iguana iguana (Reptilia: Sauria: Iguanidae). EuroJ Morph 40:89–99.
Ferreira A, Mehanna M, Dolder H. 2006. Ultrastructural immuno-cytochemical evidence for actin in the acrosomal complex duringspermiogenesis of the lizard Tropidurus itambere (Rodrigues,1987) (Reptilia: Tropiduridae). Caryologia 59:213–219.
Furieri P. 1970. Sperm morphology in some reptiles: Squamataand Chelonia. In: Baccetti B, editor. Comparative Spermatol-ogy. New York: Academic Press. pp 115–132.
Gribbins KM, Rheubert JL. 2011. The ophidian testis, sperma-togenesis and mature spermatozoa. In: Aldridge RD, SeverDM, editors. Reproductive Biology and Phylogeny of Snakes.Boca Raton, FL: CRC Press. pp 183–264.
Gribbins KM, Mills EM, Sever DM. 2007. Ultrastructural exam-ination of spermiogenesis within the testis of the GroundSkink, Scincella lateralis (Squamata, Sauria, Scincidae).J Morph 268:181–192.
Gribbins KM, Rheubert JL, Anzalone ML, Siegel DS, Sever DM.2009. Ultrastructure of spermiogenesis in the Cottonmouth,Agkistrodon piscivorus (Squamata: Viperidae: Crotalinae). JMorphol 271:293–304.
Guillette LR Jr., Casas-Andreu G. 1987. The reproductive biol-ogy of the high elevation Mexican lizard Barisia imbricata.Herpetologica 43:29–36.
Healy JM, Jamieson BGM. 1992. The phylogenetic position ofthe Tuatara, Sphenodon (Spenodontida, Amniota), as indi-cated by cladistics analysis of the ultrastructure of spermato-zoa. Philos Trans Biol Sci 335:207–219.
Healy JM, Jamieson BGM. 1994. The ultrastructure of sperma-togenesis and epididymal spermatozoa of the Tuatara Spheno-don punctatus (Sphenodontida, Amniota). Philos Trans BiolSci 344:187–199.
Hondo E, Kurohmaru M, Toriba M, Hayashi Y. 1994. Seasonalchanges in spermatogenesis and ultrastructure of developingspermatids in the Japanese rat snake, Elaphe climacophora.J Vet Med Sci 56:836–840.
Jamieson BGM. 1999. Spermatozoal phylogeny of the Verte-brata. In: Gagnon C, editor. The Male Gamete: From BasicScience to Clinical Applications. Vienna, VA: Cache RiverPress. pp 303–331.
Jamieson BGM. 2007. Avian spermatozoa: Structure and phy-logeny. In: Jamieson BGM, editor. Reproductive Biology andPhylogeny of Birds, Vol. 6. Enfield, NH: Science Publishers.pp 349–512.
Jamieson BGM, Oliver SC, Scheltinga DM. 1995. The ultra-structure of the spermatozoa of squamata-I. Scincidae, Gek-konidae, and Pygopodidae (Reptilia). Acta Zool 77:85–100.
Jamieson BGM, Oliver SC, Scheltinga DM. 1996. The Ultra-structure of the Spermatozoa of Squamata—I. Scincidae, Gek-konidae and Pygopodidae (Reptilia). Acta Zool 77:85–100.
Martınez-Torres M, Hernandez-Caballero ME, Alvarez-Rodrı-guez C, Luis-Dıaz JA, Ortiz-Lopez G. 2003. Luteal develop-ment and progesterone levels during pregnancy of the vivipa-rous temperate lizard Barisia imbricata imbricata. Gen CompEndo 132:55–65.
Martınez-Torres M, Guzman-Rodrıguez R, Cardenas-Leon M,Brunner-Reynaldo. 2006. Follicular development and ovula-tion determined by ultrasound imaging in the viviparous liz-ard Barisia imbricata (Reptila:Anguidae). Southwest Nat51:401–405.
Martınez-Torres M, Hernandez-Caballero ME, Luis-Dıaz JA,Ortiz-Lopez G, Cardenas-Leon M, Moreno-Fierros L. 2010.Effects of luteectomy in early pregnancy on the maintenanceof gestation and plasma progesterone concentrations in theviviparous temperate lizard Barisia imbricata imbricata.Reprod Biol Endocrinol 8:19.
Mubarak M. 2006. Ultrastructure of sperm tail differentiationof the lizard Stenodactylus dorie (Squamata, Reptilia). J BioSci 6:187–192.
Oliver SC, Jamieson BGM, Scheltinga DM. 1996. The ultra-structure of spermatozoa of Squamata. II. Agamidae, Varani-dae, Colubridae, Elapidae, and Boidae (Reptilia). Herpetolog-ica 52:216–241.
Reptile-Database. 2012. Species numbers. Available at:http://www.reptile-database.org (accessed on February 1, 2012)
Rheubert JL, McMahan CD, Sever DM, Bundy MR, Siegel DS,Gribbins KM. 2010a. Ultrastructure of the reproductivesystem of the black swamp snake (Seminatrix pygaea). VII.spermatozoon morphology and evolutionary trends of spermcharacters in snakes. J Zool Syst Evol Res 48:366–75.
Rheubert JL, Wilson BS, Wolf KW, Gribbins KM. 2010b. Ultra-structural study of spermiogenesis in the Jamaican GrayAnole, Anolis lineatopus (Reptilia: Polychrotidae). Acta Zool91:484–94.
Rheubert JL, Siegel DS, Venable KJ, Sever DM, Gribbins KM.2011. Ultrastructural description of spermiogenesis withinthe Mediterranean Gecko, Hemidactylus turcicus (Squamata:Gekkonidae). Micron 42:680–90.
Rheubert J, Touzinsky K, Hernandez-Gallegos O, Granados-Gonzalez G, Gribbins KM. 2012. Ontogenic development ofspermatids during spermiogenesis in the high altitude bunch-grass lizard (Sceloporus bicanthalis). Spermatogenesis 2:94–103.
Russell LD, Hikim SAP, Ettlin RA, Legg ED. 1990. Histologicaland Histopathological Evaluation of the Testis. Clearwater,FL: Cache River Press.
Sanchez-Martınez PM, Ramırez-Pinilla MR, Miranda-Esquivel.2007. Comparative histology of the vaginal-cloacal region inSquamata and its phylogenetic implications. 2007. Acta Zool88:289–307.
Scheltinga DM, Jamieson BG, Trauth SE, McAllister CT. 2000.Morphology of the spermatozoa of the iguanian lizards Utastansburiana and Urosaurus ornatus (Squamata, Phrynosto-matidae). J Submicrosc Cytol Pathol 32:261–271.
Smith HM, Burg TM, Chiszar D. 2002. Evolutionary speciationin the alligator lizard of the genus Barisia. Bull MarylandHerpet Soc 38:23–26.
Tavares-Bastos L, Teixeira L, Colli GR, Bao SN. 2002. Polymor-phism in the sperm ultrastructure among four species of liz-ards in the genus Tupinambis (Squamata: Teiidae). Acta Zool83:297–307.
Tavares-Bastos L, Colli GR, Bao SN. 2008. The evolution ofsperm ultrastructure among Boidae (Serpentes). Zoomorph127:189–202.
Teixeira RD, Colli GR, Bao SN. 1999. The ultrastucture of thespermatozoa of the lizard Micrablepharus maximiliani (Squa-mata, Gymnophthalmidae), with consideration of the use ofsperm ultrastructure characters in phylogenetic reconstruc-tions. Acta Zool 80:47–59.
Tourmente M, Giojalas L, Chiaraviglio M. 2008. Sperm ultra-structure of Bothrops alternatus and Bothrops diporus (Viper-idae, Serpentes), and its possible relation to the reproductivefeatures of the species. Zoomorph 127:241–248.
Vial JL, Stewart JR. 1985. The reproductive cycle of Barisiamonticola: A unique variation among viviparous lizards.Herpetologica 41:55–57.
Vieira GHC, Wiederhecker HC, Colli GR, Bao SN. 2001. Sper-miogenesis and testicular cycle of the lizard Tropidurus tor-quatus (Squamata, Tropiduridae) in the Cerrado of centralBrazil. Amphibia-Reptilia 22:217–233.
Vieira GHC, Colli GR, Bao SN. 2005. Phylogenetic relationshipsof corytophanid lizards (Iguania, Squamata, Reptilia) basedon partitioned and total evidence analyses of sperm morphol-ogy, gross morphology, and DNA data. Zool Scripta. 34:605–625.
Wiens JJ, Hutter CR, Mulcahy DG, Noonan BP, Townsend TM,Sites JW Jr, Reeder TW. 2012. Resolving the phylogeny of liz-ards and snakes (Squamata) with extensive sampling of genesand species. Biol Lett (in press).
614 K.M. GRIBBINS ET AL.
Journal of Morphology





























