Embed Size (px)
Citation preview
letters
nature structural biology • volume 5 number 7 • july 1998 543
A zinc finger library with degenerate α-helices was displayedon the surface of bacteriophage and proteins that bindhuman immunodeficiency virus type-1 (HIV-1) Rev responseelement stem loop IIB (RRE-IIB) RNA or 5S rRNA were iso-lated. DNA encoding affinity selected zinc fingers was shuf-fled by recombination in vitro to isolate proteins with higherRNA binding affinity. Proteins constructed in this way bindRNA specifically both in vitro and in vivo. These resultsdemonstrate that RNA substrate specificity of zinc fingerscan be changed through mutation of α-helices to constructnovel RNA binding proteins.
Zinc fingers constitute the most frequently recognized classof nucleic acid binding motif in the human genome1. C2H2 zincfingers (Fig 1a), though commonly considered a DNA bindingmotif, bind specifically and avidly to both DNA and RNA2–5.Structural conservation and modular interaction of this class ofzinc finger has been exploited to design sequence specific tran-scriptional regulators for potential applications in gene thera-py6–9. A relatively unexplored avenue with potential formolecular intervention is disruption of essential RNA–proteininteractions with sequence-specific RNA binding proteins. Werecently determined that zinc finger interaction with RNA issimilar to DNA in that binding requires a few critical aminoacids in the finger α-helix10, suggesting zinc fingers could pro-vide a stable framework for designing RNA binding proteins.To test this hypothesis we displayed a degenerate zinc fingerlibrary on bacteriophage and selected for zinc finger peptideswith affinity for a specific RNA.
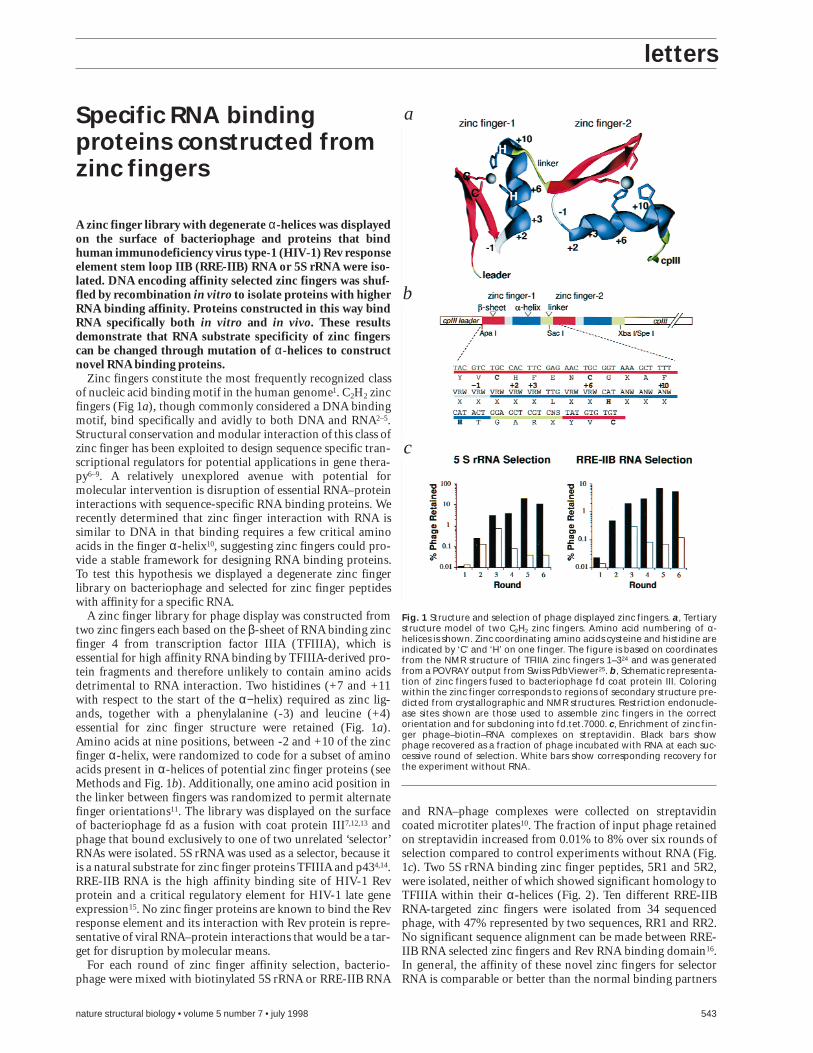
A zinc finger library for phage display was constructed fromtwo zinc fingers each based on the β-sheet of RNA binding zincfinger 4 from transcription factor IIIA (TFIIIA), which isessential for high affinity RNA binding by TFIIIA-derived pro-tein fragments and therefore unlikely to contain amino acidsdetrimental to RNA interaction. Two histidines (+7 and +11with respect to the start of the α−helix) required as zinc lig-ands, together with a phenylalanine (-3) and leucine (+4)essential for zinc finger structure were retained (Fig. 1a).Amino acids at nine positions, between -2 and +10 of the zincfinger α-helix, were randomized to code for a subset of aminoacids present in α-helices of potential zinc finger proteins (seeMethods and Fig. 1b). Additionally, one amino acid position inthe linker between fingers was randomized to permit alternatefinger orientations11. The library was displayed on the surfaceof bacteriophage fd as a fusion with coat protein III7,12,13 andphage that bound exclusively to one of two unrelated ‘selector’RNAs were isolated. 5S rRNA was used as a selector, because itis a natural substrate for zinc finger proteins TFIIIA and p434,14.RRE-IIB RNA is the high affinity binding site of HIV-1 Revprotein and a critical regulatory element for HIV-1 late geneexpression15. No zinc finger proteins are known to bind the Revresponse element and its interaction with Rev protein is repre-sentative of viral RNA–protein interactions that would be a tar-get for disruption by molecular means.
For each round of zinc finger affinity selection, bacterio-phage were mixed with biotinylated 5S rRNA or RRE-IIB RNA
and RNA–phage complexes were collected on streptavidincoated microtiter plates10. The fraction of input phage retainedon streptavidin increased from 0.01% to 8% over six rounds ofselection compared to control experiments without RNA (Fig.1c). Two 5S rRNA binding zinc finger peptides, 5R1 and 5R2,were isolated, neither of which showed significant homology toTFIIIA within their α-helices (Fig. 2). Ten different RRE-IIBRNA-targeted zinc fingers were isolated from 34 sequencedphage, with 47% represented by two sequences, RR1 and RR2.No significant sequence alignment can be made between RRE-IIB RNA selected zinc fingers and Rev RNA binding domain16.In general, the affinity of these novel zinc fingers for selectorRNA is comparable or better than the normal binding partners
Specific RNA bindingproteins constructed fromzinc fingers
Fig. 1 Structure and selection of phage displayed zinc fingers. a, Tertiarystructure model of two C2H2 zinc fingers. Amino acid numbering of α-helices is shown. Zinc coordinating amino acids cysteine and histidine areindicated by ‘C’ and ‘H’ on one finger. The figure is based on coordinatesfrom the NMR structure of TFIIIA zinc fingers 1–324 and was generatedfrom a POVRAY output from Swiss PdbViewer25. b, Schematic representa-tion of zinc fingers fused to bacteriophage fd coat protein III. Coloringwithin the zinc finger corresponds to regions of secondary structure pre-dicted from crystallographic and NMR structures. Restriction endonucle-ase sites shown are those used to assemble zinc fingers in the correctorientation and for subcloning into fd.tet.7000. c, Enrichment of zinc fin-ger phage–biotin–RNA complexes on streptavidin. Black bars showphage recovered as a fraction of phage incubated with RNA at each suc-cessive round of selection. White bars show corresponding recovery forthe experiment without RNA.
a
b
c
letters
544 nature structural biology • volume 5 number 7 • july 1998
and is highly specific. For example, 5R2 binds 5S rRNA with a Kd
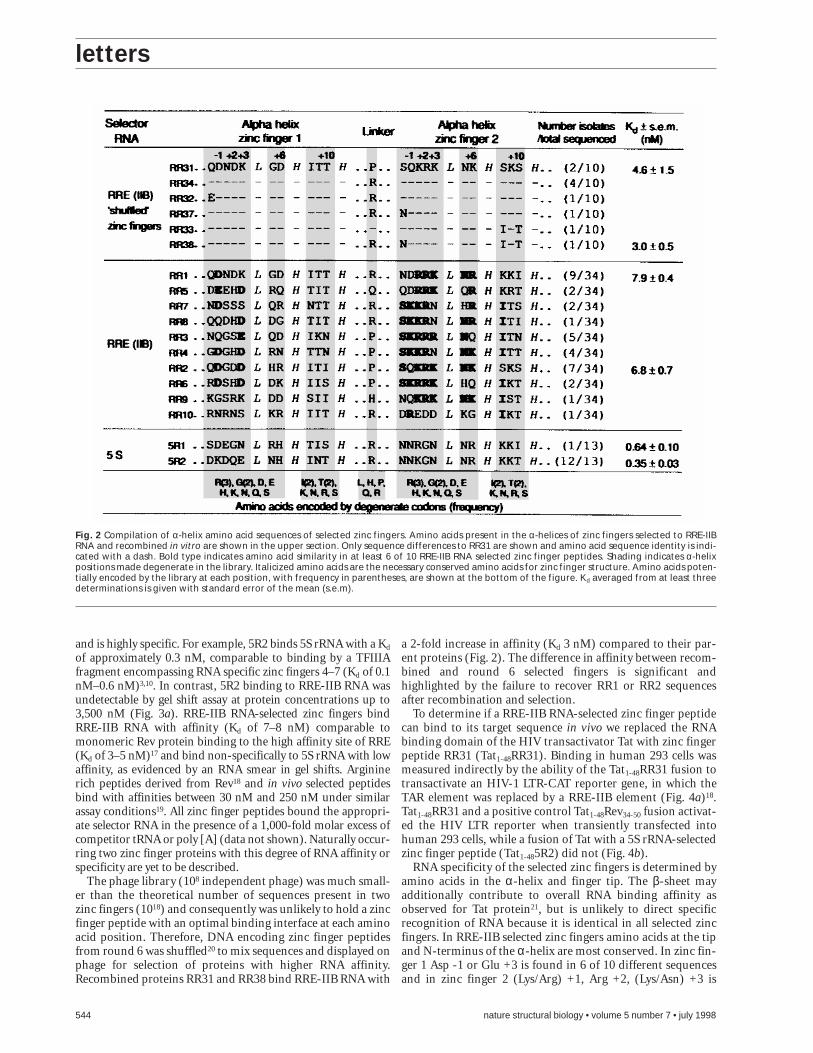
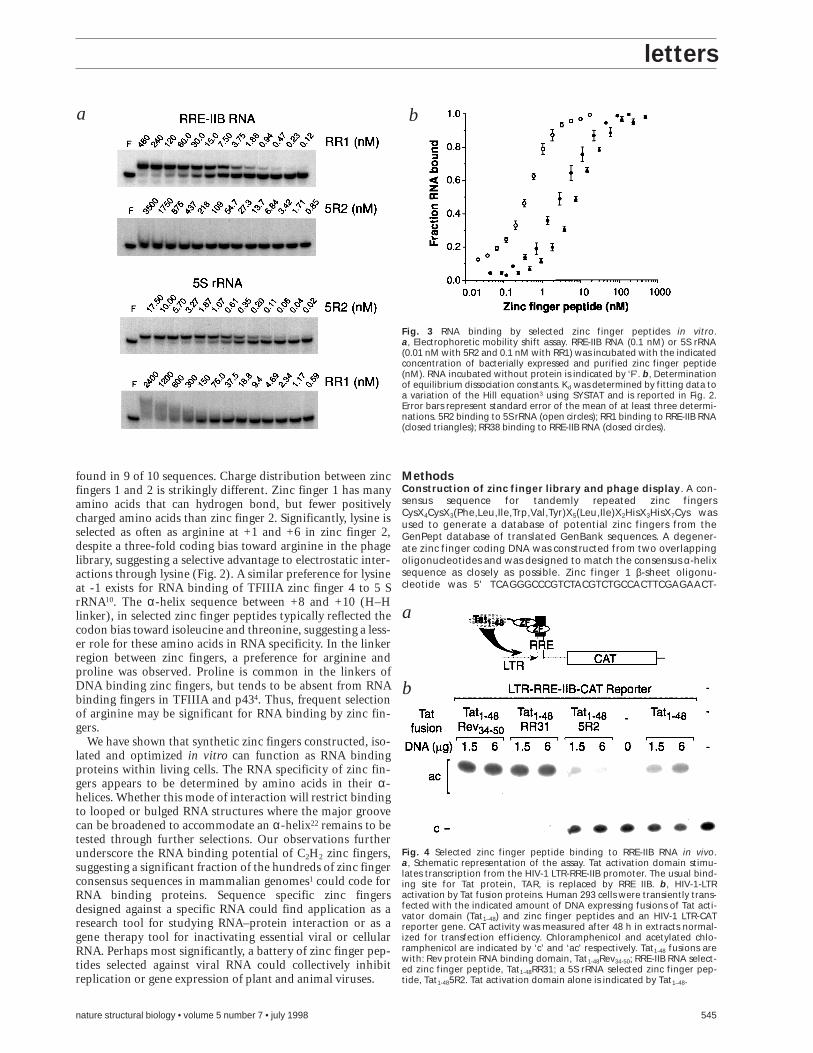
of approximately 0.3 nM, comparable to binding by a TFIIIAfragment encompassing RNA specific zinc fingers 4–7 (Kd of 0.1nM–0.6 nM)3,10. In contrast, 5R2 binding to RRE-IIB RNA wasundetectable by gel shift assay at protein concentrations up to3,500 nM (Fig. 3a). RRE-IIB RNA-selected zinc fingers bindRRE-IIB RNA with affinity (Kd of 7–8 nM) comparable tomonomeric Rev protein binding to the high affinity site of RRE(Kd of 3–5 nM)17 and bind non-specifically to 5S rRNA with lowaffinity, as evidenced by an RNA smear in gel shifts. Argininerich peptides derived from Rev18 and in vivo selected peptidesbind with affinities between 30 nM and 250 nM under similarassay conditions19. All zinc finger peptides bound the appropri-ate selector RNA in the presence of a 1,000-fold molar excess ofcompetitor tRNA or poly [A] (data not shown). Naturally occur-ring two zinc finger proteins with this degree of RNA affinity orspecificity are yet to be described.
The phage library (108 independent phage) was much small-er than the theoretical number of sequences present in twozinc fingers (1018) and consequently was unlikely to hold a zincfinger peptide with an optimal binding interface at each aminoacid position. Therefore, DNA encoding zinc finger peptidesfrom round 6 was shuffled20 to mix sequences and displayed onphage for selection of proteins with higher RNA affinity.Recombined proteins RR31 and RR38 bind RRE-IIB RNA with
a 2-fold increase in affinity (Kd 3 nM) compared to their par-ent proteins (Fig. 2). The difference in affinity between recom-bined and round 6 selected fingers is significant andhighlighted by the failure to recover RR1 or RR2 sequencesafter recombination and selection.
To determine if a RRE-IIB RNA-selected zinc finger peptidecan bind to its target sequence in vivo we replaced the RNAbinding domain of the HIV transactivator Tat with zinc fingerpeptide RR31 (Tat1-48RR31). Binding in human 293 cells wasmeasured indirectly by the ability of the Tat1-48RR31 fusion totransactivate an HIV-1 LTR-CAT reporter gene, in which theTAR element was replaced by a RRE-IIB element (Fig. 4a)18.Tat1-48RR31 and a positive control Tat1-48Rev34-50 fusion activat-ed the HIV LTR reporter when transiently transfected intohuman 293 cells, while a fusion of Tat with a 5S rRNA-selectedzinc finger peptide (Tat1-485R2) did not (Fig. 4b).
RNA specificity of the selected zinc fingers is determined byamino acids in the α-helix and finger tip. The β-sheet mayadditionally contribute to overall RNA binding affinity asobserved for Tat protein21, but is unlikely to direct specificrecognition of RNA because it is identical in all selected zincfingers. In RRE-IIB selected zinc fingers amino acids at the tipand N-terminus of the α-helix are most conserved. In zinc fin-ger 1 Asp -1 or Glu +3 is found in 6 of 10 different sequencesand in zinc finger 2 (Lys/Arg) +1, Arg +2, (Lys/Asn) +3 is
Fig. 2 Compilation of α-helix amino acid sequences of selected zinc fingers. Amino acids present in the α-helices of zinc fingers selected to RRE-IIBRNA and recombined in vitro are shown in the upper section. Only sequence differences to RR31 are shown and amino acid sequence identity is indi-cated with a dash. Bold type indicates amino acid similarity in at least 6 of 10 RRE-IIB RNA selected zinc finger peptides. Shading indicates α-helixpositions made degenerate in the library. Italicized amino acids are the necessary conserved amino acids for zinc finger structure. Amino acids poten-tially encoded by the library at each position, with frequency in parentheses, are shown at the bottom of the figure. Kd averaged from at least threedeterminations is given with standard error of the mean (s.e.m).
letters
nature structural biology • volume 5 number 7 • july 1998 545
found in 9 of 10 sequences. Charge distribution between zincfingers 1 and 2 is strikingly different. Zinc finger 1 has manyamino acids that can hydrogen bond, but fewer positivelycharged amino acids than zinc finger 2. Significantly, lysine isselected as often as arginine at +1 and +6 in zinc finger 2,despite a three-fold coding bias toward arginine in the phagelibrary, suggesting a selective advantage to electrostatic inter-actions through lysine (Fig. 2). A similar preference for lysineat -1 exists for RNA binding of TFIIIA zinc finger 4 to 5 SrRNA10. The α-helix sequence between +8 and +10 (H–Hlinker), in selected zinc finger peptides typically reflected thecodon bias toward isoleucine and threonine, suggesting a less-er role for these amino acids in RNA specificity. In the linkerregion between zinc fingers, a preference for arginine andproline was observed. Proline is common in the linkers ofDNA binding zinc fingers, but tends to be absent from RNAbinding fingers in TFIIIA and p434. Thus, frequent selectionof arginine may be significant for RNA binding by zinc fin-gers.
We have shown that synthetic zinc fingers constructed, iso-lated and optimized in vitro can function as RNA bindingproteins within living cells. The RNA specificity of zinc fin-gers appears to be determined by amino acids in their α-helices. Whether this mode of interaction will restrict bindingto looped or bulged RNA structures where the major groovecan be broadened to accommodate an α-helix22 remains to betested through further selections. Our observations furtherunderscore the RNA binding potential of C2H2 zinc fingers,suggesting a significant fraction of the hundreds of zinc fingerconsensus sequences in mammalian genomes1 could code forRNA binding proteins. Sequence specific zinc fingersdesigned against a specific RNA could find application as aresearch tool for studying RNA–protein interaction or as agene therapy tool for inactivating essential viral or cellularRNA. Perhaps most significantly, a battery of zinc finger pep-tides selected against viral RNA could collectively inhibitreplication or gene expression of plant and animal viruses.
MethodsConstruction of zinc finger library and phage display. A con-sensus sequence for tandemly repeated zinc fingersCysX4CysX3(Phe,Leu,Ile,Trp,Val,Tyr)X5(Leu,Ile)X2HisX3HisX7Cys wasused to generate a database of potential zinc fingers from theGenPept database of translated GenBank sequences. A degener-ate zinc finger coding DNA was constructed from two overlappingoligonucleotides and was designed to match the consensus α-helixsequence as closely as possible. Zinc finger 1 β-sheet oligonu-cleotide was 5' TCAGGGCCCGTCTACGTCTGCCACTTCGAGAACT-
a
Fig. 3 RNA binding by selected zinc finger peptides in vitro.a, Electrophoretic mobility shift assay. RRE-IIB RNA (0.1 nM) or 5S rRNA(0.01 nM with 5R2 and 0.1 nM with RR1) was incubated with the indicatedconcentration of bacterially expressed and purified zinc finger peptide(nM). RNA incubated without protein is indicated by ‘F’. b, Determinationof equilibrium dissociation constants. Kd was determined by fitting data toa variation of the Hill equation3 using SYSTAT and is reported in Fig. 2.Error bars represent standard error of the mean of at least three determi-nations. 5R2 binding to 5S rRNA (open circles); RR1 binding to RRE-IIB RNA(closed triangles); RR38 binding to RRE-IIB RNA (closed circles).
a
b
Fig. 4 Selected zinc finger peptide binding to RRE-IIB RNA in vivo.a, Schematic representation of the assay. Tat activation domain stimu-lates transcription from the HIV-1 LTR-RRE-IIB promoter. The usual bind-ing site for Tat protein, TAR, is replaced by RRE IIB. b, HIV-1-LTRactivation by Tat fusion proteins. Human 293 cells were transiently trans-fected with the indicated amount of DNA expressing fusions of Tat acti-vator domain (Tat1–48) and zinc finger peptides and an HIV-1 LTR-CATreporter gene. CAT activity was measured after 48 h in extracts normal-ized for transfection efficiency. Chloramphenicol and acetylated chlo-ramphenicol are indicated by ‘c’ and ‘ac’ respectively. Tat1-48 fusions arewith: Rev protein RNA binding domain, Tat1-48Rev34-50; RRE-IIB RNA select-ed zinc finger peptide, Tat1-48RR31; a 5S rRNA selected zinc finger pep-tide, Tat1-485R2. Tat activation domain alone is indicated by Tat1–48.
b

letters
546 nature structural biology • volume 5 number 7 • july 1998
GCGGTAAAGCTTTT 3' and the randomized finger tip and α-helixoligonucleotide was 5' GACGAGCTCCAGTATGWNTWNTWNTATG-WYBWYBCAAWYBWYBWYBWYBWYBAAAAGCTTTACCGCA 3'(where W is A or T; N is A, C, G or T; Y is C or T; B is C, G or T). Thecorresponding oligonucleotides for zinc finger 2 were 5' CATACTGGAGCTCGTCNSTATGTGTGTCACTTTGAAAATTGCGGC-CGTAGTTTC 3' and 5'TGCTCTAGATGCACCGGTGTGWNTWNTWNT-GTGWYBWYBCAAWYBWYBWYBWYBWYBGAAACTACGGCCGCA 3'respectively. Each oligonucleotide pair was annealed and extend-ed with Klenow fragment and the resultant DNA fragments wereligated in tandem into fd.tet.7000 using appropriate restrictionsites to ensure correct orientation (Fig. 1b). Ligation productswere electroporated into DH10B cells and tetracycline resistanttransformants (5.1 × 107) were amplified by growth in 0.27%agarose (SeaPrep, (FMC)) for 30 h at 31 °C . 24 randomly selectedclones had inserts of the correct size and another 25 of 38 clones(65%) produced phage. We estimate 3.4 × 107 viable clones withinserts were screened. Phage isolated from the soft agar (109)were amplified in E. coli K-91 cells and used for selection10. Phage(108–109 colony forming units) were selected following incubationwith 100 nM RNA as previously described7,10. The final two roundsof selection included 10-fold excess of tRNA over biotinylatedRNA.
RRE-IIB (nucleotides 39–81 of HIV RRE with 5' GGG and 3' CCCextensions for stability) and Xenopus laevis oocyte-type 5 S rRNA(120 nucleotides plus one additional 3' U) were synthesized byrun-off transcription with T7 RNA polymerase. Biotin was incorpo-rated into the RNA at approximately 2 mol mol-1 RNA by inclusionof biotin-16-UTP during transcription.
Protein expression, purification, and gel shift analysis.Selected zinc finger DNA was amplified by PCR and subcloned intopET28b(+) for expression as (His)6 fusions. All constructs were con-firmed by automated dideoxy sequencing. Expression in E. coliBL21 cells, purification by nickel column chromatography, proteinfolding, removal of histidine tags, and gel shift conditions were aspreviously reported10.
DNA shuffling and selection. PCR amplified round 6 zinc fingerDNA was shuffled using the method for recombination based onshort sequence homology20. Reassembled DNA was ligated intofd.tet.7000 and electroporated into MC1061 yielding 1.4 × 108 tetra-cycline resistant transformants. Shuffled zinc fingers were selectedfor three rounds with a 1000-fold excess of tRNA competitor. Inrounds 2 and 3, wells were washed with 250 mM NaCl phage elutionbuffer before eluting with 4 M NaCl phage elution buffer10.
Cell transfection assay. Tat-zinc finger fusion constructs weremade in pSV2, and express Tat1-48-AAAAASPPKKKRKVAGGR-zincfinger (nuclear localization signal underlined). Human 293 cellswere transiently transfected by CaPO4 precipitation with 2 µg ofpHIV LTR-IIB CAT, 0.75 µg pCH110, a β-galactosidase expressionplasmid, varying amounts of Tat-fusion plasmids, and carrier DNAup to 10 µg. After 48 h CAT activity was measured in cellularextracts normalized for β-galactosidase activity23.
AcknowledgmentsWe thank E. Rebar and C. Pabo for fd.tet.7000; A. Frankel for pSV2TatRev andpHIV LTR-IIB CAT; T. Parslow for pCH110; and E W. Mercer for cell culturefacilities.
Westley J. Friesen and Martyn K. Darby
Department of Microbiology and Immunology, Kimmel Cancer Institute,Thomas Jefferson University, 233 S. 10th St., Philadelphia, Pennsylvania 19107,USA.
Correspondence should be addressed to M.K.D. email: [email protected]
Received 25 March, 1998; accepted 19 May, 1998.
1. Henikoff, S., et al. Science 278, 609–614 (1997).2. Berg, J.M. et al. Science 271, 1081–1085 (1996).3. Clemens, K.R. et al. Science 260, 530–533 (1993).4. Joho, K.E. et al. Cell 61, 293–300 (1990).5. Theunissen, O. et al. Cell 71, 679–690 (1992).6. Choo, Y. et al. Nature 372, 642–645 (1994).7. Rebar, E.J. et al. Science 263, 671–673 (1994).8. Choo, Y. et al. Curr. Opin. Struc. Biol. 7, 117–125 (1997).9. Pomerantz, J.L. et al. Science 267, 93–96 (1995).
10. Friesen, W.J. et al. J. Biol. Chem. 272, 10994–10997 (1997).11. Nakaseko, Y. et al. J. Mol. Biol. 228, 619–636 (1992).12. Rebar, E.J. et al. Meth. Enz. 267, 129–149 (1996).13. Smith, G.P. et al. Meth. Enz. 217, 228–257 (1993).14. Pelham, H. et al. Proc. Natl. Acad. Sci. USA 77, 4170–4174 (1980).15. Hope, T. et al. Curr. Topics Micro. Immun. 193, 91–105 (1995).16. Kjems, J. et al. EMBO J. 11, 1119–1129 (1992).17. Mann, D.A. et al. J. Mol. Biol 241, 193–207 (1994).18. Tan, R. et al. Cell 73, 1031–1040 (1993).19. Harada, K. et al. Nature 380, 175–179 (1996).20. Stemmer, W.P. Proc. Natl. Acad. Sci. USA 91, 10747–10751 (1994).21. Puglisi, J. et al. Science 270, 1200–1203 (1995).22. Battiste, J.L. et al. Science 273, 1547–1551 (1996).23. Sambrook, J. et al. Molecular cloning. A laboratory manual. (Cold Spring Harbor
Laboratory Press; 1989).24. Wuttke, D. et al. J. Mol. Biol. 273, 183–206 (1997).25. Guex, N. et al. Protein Data Bank Quaterly Newsletter 77, 1996).
Site-specific DNA bindingusing a variation of thedouble stranded RNAbinding motif
The integrase family of site-specific recombinases catalyze adiverse array of DNA rearrangements in archaebacteria, eubac-teria and yeast. The solution structure of the DNA bindingdomain of the integrase protein from the conjugative transpo-son Tn916 has been determined using NMR spectroscopy. Thestructure provides the first insights into distal site DNA bind-ing by a site-specific integrase and reveals that the N-terminaldomain is structurally similar to the double stranded RNAbinding domain (dsRBD). The results of chemical shift map-ping experiments suggest that the integrase protein interacts
with DNA using residues located on the face of its three strand-ed β-sheet. This surface differs from the proposed RNA bindingsurface in dsRBDs, suggesting that different surfaces on thesame protein fold can be used to bind DNA and RNA.
Conjugative transposons spread antibiotic resistance amongpathogenic bacteria and are broadly distributed in nature. Theyare extremely promiscuous and can move between unrelatedbacterial cells (strains from different species or genera) duringtransposition (reviewed in refs 1–3). One of the most thorough-ly studied conjugative transposons is the Tn916 element, whichis 18 kilobases in length and carries resistance to tetracycline.Tn916 transposition is performed by two transposon encoded
Fig. 1. The Tn916 transposon. Binding sites for the N- and C-terminaldomains of integrase are shown as triangles and diamonds respectively.Binding sites for the accessory factor xis are shown as hexagons.





























