Embed Size (px)
Citation preview

Specific Potentialities of EmbryonicRat Serotonergic Neurons to Innervate
Different Periventricular Targetsin the Adult Brain
M. DIDIER-BAZES,1* B. VOUTSINOS,1 M. AGUERA,1 C. PEYRON,2
H. AKAOKA,1 AND M.F. BELIN1
1INSERM U 433, Faculte Laennec, 69372 Lyon Cedex 08, France2INSERM U 52, Faculte Rockefeller, 69373 Lyon Cedex 08, France
ABSTRACTDuring the development of the central nervous system, neurons are directed by both
genetic and environmental factors to differentiate and form connections with their targets. Wetook advantage of the abundant homogeneous serotonergic innervations of the ependymaforming the supra- and subependymal plexuses to investigate possible commitment ofembryonic neurons to innervate specific targets during axogenesis in the rat. The origin of thesupraependymal innervation was determined by retrograde transport of cholera toxin (CT)from the ventricles. The supraependymal plexuses of the fourth ventricle mainly originatedfrom neurons in the dorsocaudal region of the raphe dorsalis (DRN), while the rostral DRNand raphe centralis (CRN) contained perikarya projecting into the third ventricle. Thissuggested the existence, along the rostrocaudal axis of the raphe, of different neuronal subsetsable to form distinct supraependymal plexuses in the third or fourth ventricle. To determinewhether serotonergic neurons were committed to innervate specific areas of the ependyma,different embryonic metencephalic segments (rostral, median, or caudal) from 14-day-old ratembryos were independently grafted into the third or fourth ventricle of an adult brain inwhich the serotonergic neurons had been previously destroyed. The distinctive patterns ofre-innervation specific to each of grafted segments indicate that subsets of embryonicserotonergic neurons are indeed committed to innervate certain restricted ependymal areas ofthe adult brain, presumably in response to different neurotropic and/or neurotrophic cues. J.Comp. Neurol. 382:29–45, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: serotonin; ependyma; raphe; development; transplantation; predetermination
For the successful formation of the central nervoussystem (CNS), parallel yet interdependentmaturation andorganization of brain cells is essential. This mainly con-sists of the differentiation of neural cells and the formationof intricate connective patterns between different neuro-nal classes. To determine the mechanisms underlyingthese phenomena in the complex vertebrate CNS, it is firstnecessary to study the individual systems separately.Lineage analysis of and clonal experiments on the cortexsuggest that the initial series of events in neuronal differ-entiation is influenced by local environmental interactionswithin the neuroepithelium, during, or following, final celldivision (Grove et al., 1993; Krushel et al., 1993; McConneland Kaznowski, 1991). As development proceeds, theneuronal growth cones move toward their targets, follow-ing a series of attractive/repulsive molecular cues, specificto classes or subclasses of neurons (Letourneau, 1975;
Jessell, 1988; Patterson, 1988; Goodman and Shatz, 1993).It has also been shown, especially in the peripheralnervous system, that target cells can secrete signalingfactors and promote a second round of late neuronaldifferentiation and/or maturation (Landis, 1990). Al-though themonoaminergic systems of the brainstem (Dahl-strom and Fuxe, 1964) play an important modulatory rolein many brain functions due to their widespread projec-tions throughout the entire neuraxis, little information isavailable regarding themechanisms involved in the forma-
Contract grant sponsor: INSERM; Contract grant number: U433.*Correspondence to: M. Didier-Bazes, INSERM 433, Faculte Laennec,
69372 Lyon Cedex 08, France. E-mail: [email protected] 29 August 1995; Revised 25 November 1996; Accepted 5
January 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 382:29–45 (1997)
r 1997 WILEY-LISS, INC.

tion of the appropriate connections. Concerning the seroton-ergic system, this phenotype first appears in the rat brainbetween days 12–14 of gestation; the serotonergic neuronsthen migrate from the ventral zone toward their finaldestinations (Lauder and Bloom, 1974; Lauder et al., 1982;Lidov and Molliver, 1982a; Wallace and Lauder 1983) andform outgrowing neurites along pre-existing fiber path-ways (Azmitia, 1978; Lidov and Molliver, 1982b; Wallaceand Lauder, 1983). Such guidance of serotonergic fiberoutgrowth has been suggested as one possible cellularbasis for the many widespread connections formed by theserotonergic system (Azmitia, 1978). However, it is notknown whether, at the time of axonogenesis, serotonergicneurons form a homogeneous neuronal population, poten-tially capable of innervating any part of the brain, orwhether distinct groups of serotonergic neurons exist,committed to the innervation of specific targets.Those serotonergic neurons innervating the ventricular
zone are particularly suited to the study of the develop-ment of both junctional and non-junctional serotonergicprojections. Their terminals form dense, homogeneoussupra- or subependymal plexuses, the morphological andneurochemical characteristics of which have been exten-sively studied. In fact, classic ependymocytes, which sepa-rate the cerebrospinal fluid (CSF) from the brain paren-chyma, are innervated by both supra- and sub-ependymalplexuses formed by non-junctional fibers containing respec-tively, either 5-hydroxytryptamine (5-HT) plus g-aminobu-tyric acid (GABA) or 5-HT alone (Lorez and Richard, 1975;Chan, 1976; Belin et al., 1980; Harandi et al., 1986). Incertain regions, the ependyma differentiate into circumven-tricular organs, innervated by serotonergic terminals withother morphological and biochemical characteristics. Thesubcommissural organ (SCO), situated at the junction ofthe third ventricle and the aqueduct under the posteriorcommissura, is one such organ. It is formed by specializedependymocytes, which are richly innervated by an homoge-neous plexus of synaptic terminals containing only 5-HT(Bouchaud andArluison, 1977; Voutsinos et al., 1994). In aprevious study, we showed that the specialized ependymo-cytes of the SCO and the classical ependymocytes of theependyma exert selective tropic and/or trophic effects onembryonic serotonergic neurons grafted into adult rats inwhich the endogenous serotonergic innervation had beenpreviously destroyed (Voutsinos et al., 1994). In fact, theseSCO ependymocytes selectively attract and/or maintainserotonergic synaptic terminals, whereas the classic epen-dymocytes of the third ventricle attract and/or maintainnon-junctional terminals, containing both 5-HT andGABA,as found in the normal rat brain. Such observationsindicate either that targets exert an effect on the matura-tion of serotonergic neurons or that neurons are predeter-mined to innervate defined targets. Our preliminary obser-
vation suggested that embryonic serotonergic neuronsmay be committed to innervate specific targets dependingon the anatomical localization of their perikarya.The aim of the present work was to determine whether
subsets of embryonic serotonergic neurons possess distinctpotentialities to innervate specific targets, i.e. differentareas of the ependyma of the adult rat, at the time ofaxonogenesis. We first determined the location of thoseserotonergic neuronal perikarya projecting to the thirdand fourth ventricles, using retrograde transport of chol-era toxin (CT) from these ventricles. We then studied theability of embryonic serotonergic neurons of differentmetencephalic segments (presumably corresponding todistinct subpopulations of serotonergic neurons defined byretrograde labeling), to innervate specific ventricular zonesby transplanting them into the third or fourth ventricle ofadult rats in which the serotonergic terminals had beenpreviously destroyed.
MATERIALS AND METHODS
Animals
Twenty-eight male Sprague-Dawley rats, 250–300 g,(IFFACREDO, France) were divided into seven groups: (i)three rats injected with CT in the habenular recess of thethird ventricle, (ii) two rats injected with CT in the fourthventricle, (iii) two rats injected with CT in the rostraldorsal raphe nucleus (DRN), (iv) three sham5,7-dihydroxy-tryptamine (5,7-DHT) denervated rats, (v) ten 5,7-DHTdenervated rats implanted with solid transplants in thethird ventricle (4 implanted with embryonic rostral meten-cephalon, 3 with median metencephalon and 3 with caudalmetencephalon), (vi) six 5,7-DHT denervated rats im-planted with solid transplants in the fourth ventricle (twoeach implanted with rostral, median, or caudal embryonicmetencephalon), and (vii) two control rats.All transplanted rats included in the present study
showed successful solid transplants in the habenularrecess of the third ventricle against the posterior commis-sure or in the fourth ventricle, attached both to the roofand the floor of the ventricle, approximately at the rostrallevel of the prepositus hypoglossal nucleus. All experi-ments complied with the guidelines of the Ethics Commit-tee of the French Ministry of Agriculture (Decree 87-848).
Cholera toxin injection and detection
Under chloral hydrate anesthesia, CT b subunit (2 µl ofa 1% solution in 0.9% NaCl) (List Biological Laboratories)was injected stereotaxically, using a Hamilton syringe,into the habenular recess of the third ventricle (injectioncoordinates with reference to the ear bars, incisor bar setat 23 mm and the needle set at an anterior angle of 42degrees: AP 21 mm; DV 15.5 mm; L 10.2) or into thefourth ventricle, via the cisterna magna (injection coordi-nates with reference to the ear bars, the skull position setwith a posterior angle of 22 degrees and the needle set atthe same angle: AP 12.5 mm; DV 14.5 mm; 1 10 mm). CTwas iontophoretically injected into the DRN through aglass micropipette, as previously described (Luppi et al.,1990; Peyron et al., 1996; injection coordinates from theAtlas of Paxinos and Watson (1982) with reference to theear bars, AP12.2 or 11.2 mm; DV 14 mm; L 0).After 48 hours (ventricular injection) or 4 days (DRN
injection), the rats were killed under chloral hydrateanesthesia and intracardiacally perfused with 300 ml offixative (0.1 M phosphate-buffered saline [PBS], pH 7.4,
Abbreviations
BSA bovine serum albuminCSF cerebrospinal fluidCRN central raphe nucleusCT cholera toxin5,7-DHT 5,7-dihydroxytryptamineDRN dorsal raphe nucleusGFAP glial fibrillary acidic protein5-HT 5-hydroxytryptamine (serotonin)PBS phosphate-buffered salineSCO subcommissural organTPOH tryptophan hydroxylase
30 M. DIDIER-BAZES ET AL.
containing 4% paraformaldehyde and 0.2% picric acid).The brains were post-fixed for 12 hours in the samefixative, then immersed in PBS, containing 20% sucrose,
frozen at 260°C and sectioned on a cryostat (20 µm). Thesections were then immunostained, using the biotin-streptavidin system (Hsu et al., 1981). In brief, sections
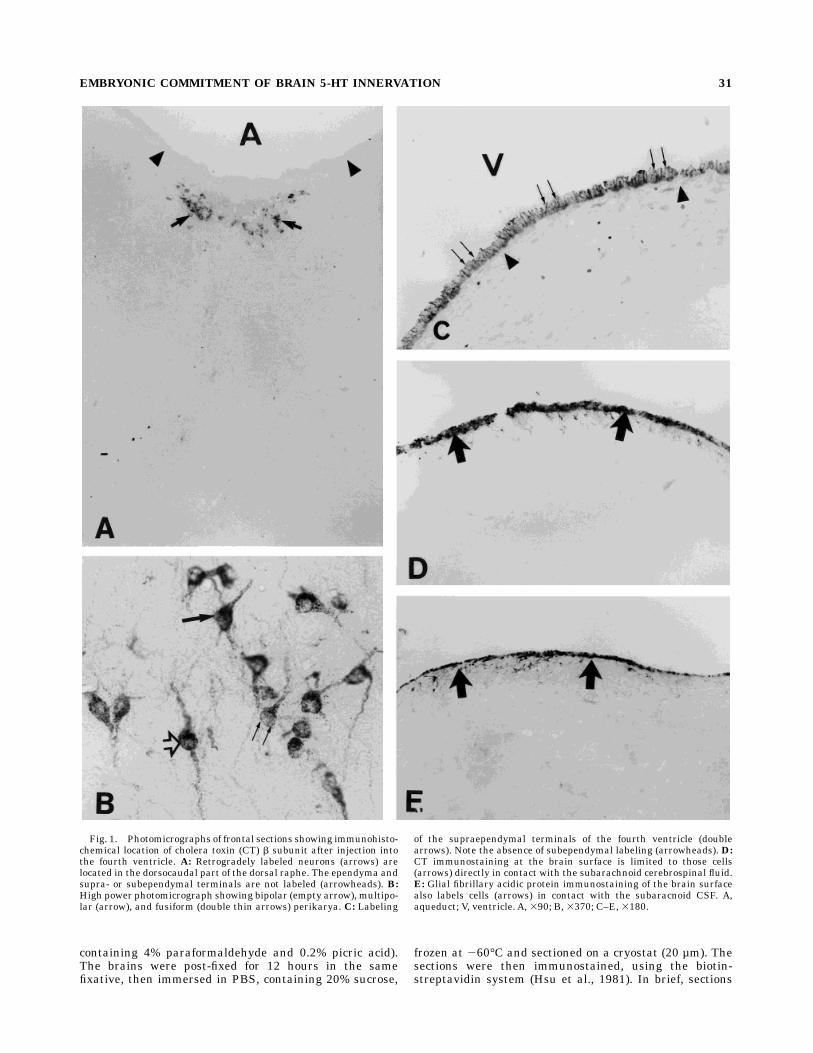
Fig. 1. Photomicrographs of frontal sections showing immunohisto-chemical location of cholera toxin (CT) b subunit after injection intothe fourth ventricle. A: Retrogradely labeled neurons (arrows) arelocated in the dorsocaudal part of the dorsal raphe. The ependyma andsupra- or subependymal terminals are not labeled (arrowheads). B:High power photomicrograph showing bipolar (empty arrow), multipo-lar (arrow), and fusiform (double thin arrows) perikarya. C: Labeling
of the supraependymal terminals of the fourth ventricle (doublearrows). Note the absence of subependymal labeling (arrowheads). D:CT immunostaining at the brain surface is limited to those cells(arrows) directly in contact with the subarachnoid cerebrospinal fluid.E: Glial fibrillary acidic protein immunostaining of the brain surfacealso labels cells (arrows) in contact with the subaracnoid CSF. A,aqueduct; V, ventricle. A, 390; B, 3370; C–E, 3180.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 31
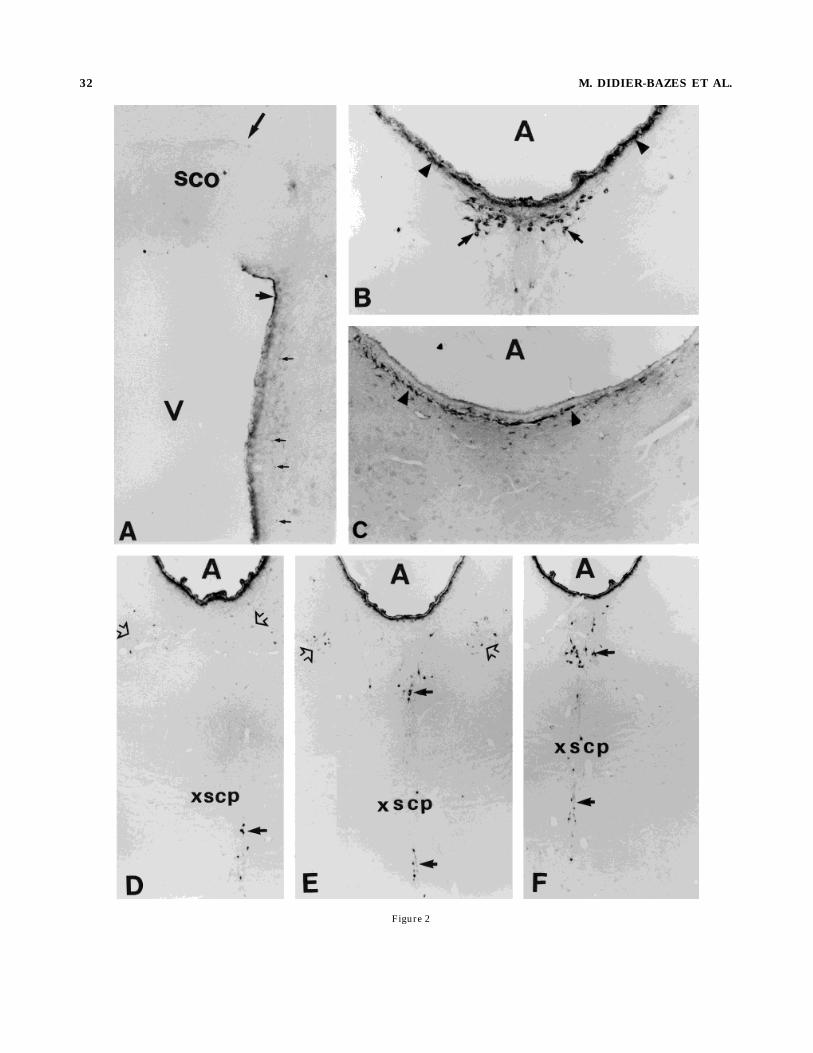
Figure 2
32 M. DIDIER-BAZES ET AL.

were incubated with goat antibody directed against CT bsubunit (List Biological Laboratories; diluted 1/20,000 inPBS containing 0.1% BSA) for 24 hours at 4°C, followed bybiotinylated donkey anti-goat Ig antibodies (Jackson Labo-ratories; diluted 1/1,000 in PBS containing 0.1% BSA),then streptavidin-peroxidase complex (Jackson Laborato-ries; diluted 1/2,000 in PBS containing 0.1% BSA), both for2 hours at room temperature. Peroxidase activity wasvisualized by incubation with 0.03% 3-38-diaminobenzi-dine and 0.003% hydrogen peroxide in 50 mM Tris-HCl,pH 7.5.
Destruction of endogenousserotonin neurons
Serotonergic terminals and neurons were destroyed bystereotaxic injection of 5,7-dihydroxytryptamine creati-nine sulfate (5,7-DHT; Sigma D-0136) 30 minutes afterpretreatment with desipramine (Sigma D-3900; 20 mg/kgi.p.), which protects catecholaminergic neurons from thetoxic effect of 5,7-DHT. The 5,7-DHT was injected underchloral hydrate anesthesia (400 mg/kg i.p.) into the lateralventricle (330 µg in 20 µl of 0.9% NaCl) (injection coordi-nates with reference to the bregma, incisor bar set at 15mm: AP 0 mm; DV 1 3.6 mm; L 11.4 mm) and the medianraphe (66 µg in 4 µl of 0.9% NaCl) (injection coordinateswith reference to the ear bars, incisor bar set at 22.5 mmand the needle set with a posterior angle of 45 degrees: AP13.7 mm; DV 11.9 mm; L 0mm) 10 days before transplan-tation of the embryonic metencephalon.
Tissue dissection and transplantationof embryonic raphe
Fourteen-day-old rat fetuses (day of mating taken as day0) were obtained by cesarean section under ether anesthe-sia and their brains dissected in Hanks’ buffered solution(pH 7.4). The metencephalic brainstem, containing theembryonic serotonergic neurons, which form the dorsaland central raphe nuclei, was excised caudally to themesencephalic flexure, at the level of the isthmic fovea androstrally to the pontine flexures (Fig. 7A). The dissectedbasal plate of the metencephalon was subdivided intothree segments: the rostral, median, and caudal meten-
cephalon, each segmentmeasuring approximately 0.4mm.Under chloral hydrate anesthesia, the different segmentswere injected separately into the habenular recess of thethird ventricle (same coordinates as for injection of CT) orinto the fourth ventricle, via the cisterna magna (injectioncoordinates with reference to the ear bars, the skullposition set with a posterior angle of 23 degrees and theneedle set at the same angle: AP 11.2 mm; DV 11.7 mm; l10 mm). Each group implanted in the third or fourthventricle was subdivided into three subgroups, receivingeither rostral, median, or caudal metencephalic segments.
Immunocytochemistry and autoradiography
Five months after surgery, control rats, 5,7-DHT treatedrats and grafted rats were anesthetized with chloralhydrate and received an intraventricular injection of 740KBq of [3H]5-HT (0.8 3 1029 moles, NewEnglandNuclear;876.9 GBq/mmol) in 20 µl of 0.9% NaCl. Thirty minuteslater, the animals were killed by intracardiac perfusionwith 2% paraformaldehyde and 2% glutaraldehyde, andthe brains removed and post-fixed for 12 hours in the samefixative. The brains of the 14-day-old rat fetuses, used fordetection of tryptophan hydroxylase (TPOH, the specificrate-limiting enzyme of 5-HT synthesis)-containing cellbodies in the metencephalon (Cash et al., 1985; Weissmanet al., 1987), were fixed by immersion in 4% paraformalde-hyde for 24 hours. Tissue blocks were embedded in polyeth-ylene glycol or paraffin and cut into 7-µm sections, using amicrotome.Sections used for the localization of serotonergic cell
bodies by immunocytochemical detection of TPOH wererinsed for 3 hours in 0.1 M phosphate buffer, containing0.1% BSA and 0.1% Triton X-100, then processed forbiotin-streptavidin immunostaining, as described above.Anti-TPOH antibodies (diluted 1/2,000 in PBS containing0.1% BSA and 1% normal pig serum) were applied for 18hours at 4°C, followed by donkey anti-goat Ig antibodies(Jackson; diluted 1/1,000) for 2 hours. The final step wasidentical to that for the CT b subunit.[3H]5-HT-labeled sections used for the autoradiographic
detection of serotonergic terminals were processed usingIlford K5 emulsion; after 20 days of exposure, the autora-diographs were developed using D19 (Kodak).
RESULTS
Labeling of the cell bodies formingthe ependymal plexuses by retrogradetransport of cholera toxin b subunit
Material injected into the ventricles is transported byCSF flow from the injected ventricles through the aque-duct and into the space surrounding the brain (Wright,1982), thus CT injected into the fourth ventricle diffusesinto this ventricle and the subarachnoid space. Forty-eighthours after injection, the supraependymal terminals of thefourth ventricle were intensely labeled, while the subepen-dymal terminals were unlabeled (Fig. 1C). CT-immunore-active (Fig. 1D) and GFAP-immunolabeled cells (Fig. 1E)were also present at the brain surface; these cells are theastrocytes forming the glia limitans. Numerous neuronalsoma, localized in the dorsocaudal part of the DRN, wereretrogradely labeled by the toxin (30 to 40 cells per slice)(Fig. 1A) (schematically represented in Fig. 4E); labelingwas granular and localized in the soma, dendrites, and
Fig. 2. Photomicrographs of frontal sections illustrating immuno-histochemical location of CT b subunit after injection into the habenul-lar recess of the third ventricle. A: Numerous immunopositive supra-ependymal fibers in the third ventricle are labeled (short arrow). A fewlabeled subependymal terminals are scattered below the ependyma inthe parenchyma (thin arrows). Note the absence of labeling of theterminals innervating the basal region of the subcommissural organ(SCO) (long arrow). B: Neurons that were retrogradely labeled afterCT injection into the fourth ventricle were also labeled by CT injectioninto the third ventricle (arrows). Supra- and subependymal CTlabeling can be seen (arrowheads). C: GFAP immunolabeling showingsubependymal labeling comparable to that seenwith CT immunolabel-ing (arrowheads). D–F: Three different rostrocaudal levels of therostral DRN and the central raphe nucleus (CRN) in which retro-gradely labeled neurons can be seen (arrows) [rostrocaudal coordi-nates with reference to ear bars D 5 12, E 5 11.7, F 5 11.2 from theAtlas of Paxinos andWatson (1982)]. The retrogradely labeled soma inthe rostral DRN form two populations, distinguished by intensity ofthe labeling. One population of weakly labeled neurons is symmetri-cally situated in the wings of the raphe (empty arrows), whereas aheavily labeled cluster is located medially (dark arrow). V, ventricle;SCO, subcommissural organ; xscp, decussation of the superior cerebel-lar peduncles. A and C, 3180; B, 390; D–F, 345.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 33
axons (Fig. 1B). The CT-immunopositive neurons werebipolar, multipolar, or fusiform. Very few CT labeled cellswere seen in the rostral DRN (one or two cells in someslices) or the raphe centralis (CRN).CT injected into the habenular recess of the third
ventricle diffused, via CSF flux, into the third ventricle,aqueduct, fourth ventricle, and subarachnoid space. Forty-eight hours after injection, CT immunoreactivity was seenon the supraependymal fibers of the third ventricle (Fig.2A), aqueduct (Fig. 2B,D–F), and fourth ventricle, as well
as on the glia limitans, surrounding the brain. CT immuno-labeling was also noted just below the ependyma (Fig. 2B),corresponding to GFAP-positive subependymal astrocytes,which border the ventricles and aqueduct and form abarrier between the ventricular CSF and the neuropil (Fig.2C). Scattered subependymal terminals under the epen-dyma of the third ventricle and aqueduct were also immu-noreactive (Fig. 2A,B). The plexuses innervating the basalpart of the SCO (Fig. 2A) and the subependymal region ofthe fourth ventricle were not labeled. As expected, those
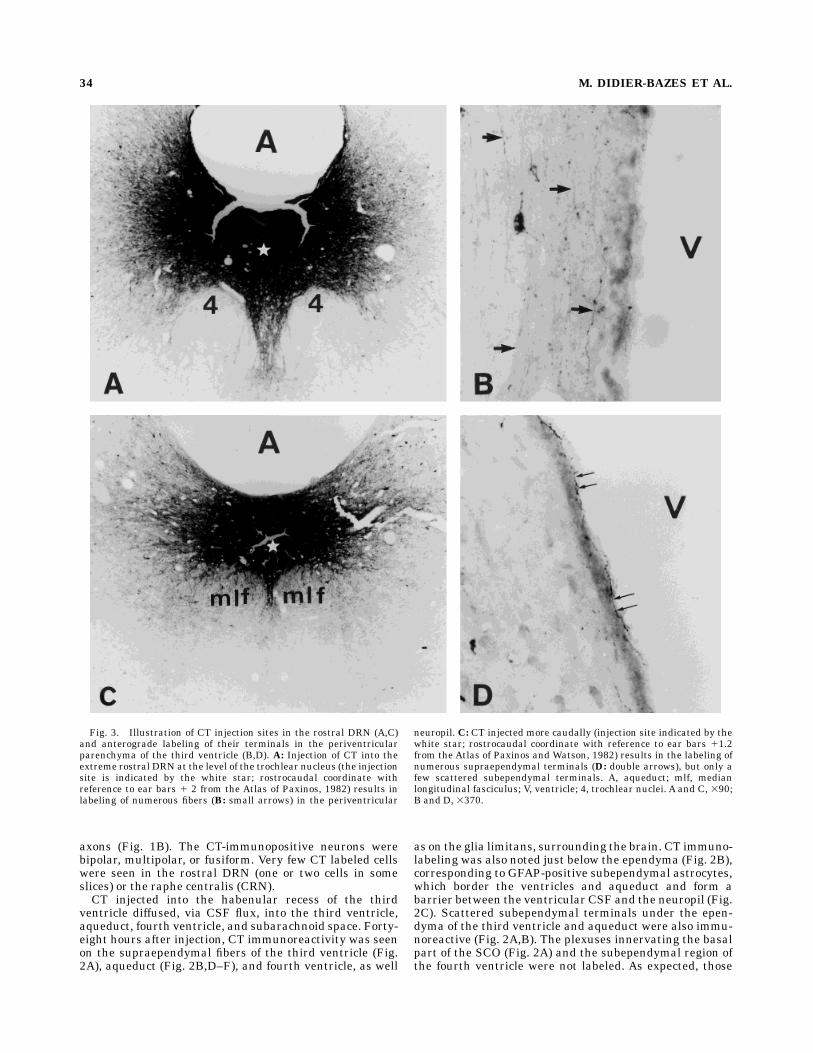
Fig. 3. Illustration of CT injection sites in the rostral DRN (A,C)and anterograde labeling of their terminals in the periventricularparenchyma of the third ventricle (B,D). A: Injection of CT into theextreme rostral DRN at the level of the trochlear nucleus (the injectionsite is indicated by the white star; rostrocaudal coordinate withreference to ear bars 1 2 from the Atlas of Paxinos, 1982) results inlabeling of numerous fibers (B: small arrows) in the periventricular
neuropil. C: CT injected more caudally (injection site indicated by thewhite star; rostrocaudal coordinate with reference to ear bars 11.2from the Atlas of Paxinos and Watson, 1982) results in the labeling ofnumerous supraependymal terminals (D: double arrows), but only afew scattered subependymal terminals. A, aqueduct; mlf, medianlongitudinal fasciculus; V, ventricle; 4, trochlear nuclei. A and C, 390;B and D, 3370.
34 M. DIDIER-BAZES ET AL.

neurons labeled after injection into the fourth ventriclewere retrogradely labeled after third ventricle injection(Fig. 2B). In addition, numerous neurons in the rostralpart of the DRN as well as in the CRN and raphe caudallinearis (Fig. 2D–F) were labeled. Neurons from the rostralDRN could be divided into two populations, depending onthe intensity of the labeling. One, in the median axes (Fig.2E,F), was labeled as intensely as the cells in the caudalDRN (Fig. 2A), whereas the other, more laterally situatedin the extreme rostral wings of the DRN (Fig. 2D,E), wasless intensely labeled, suggesting that these neuronsproject axons to regions, such as the subependymal area,in which CT is present at a low concentration. The distinct
projection areas of these two neuronal populations wascorroborated by anterograde labeling with CT injected intodifferent parts of the rostral DRN. When CT was injectediontophoretically into the rostral extremity of the DRNcontaining mainly neurons showing only weak retrogradelabeling (Fig. 3A) (anteroposterior localization correspond-ing to Fig. 2D),marked anterograde labeling of the subepen-dymal fibers of the third ventricle was seen (Fig. 3B),whereas injection into the rostral DRN, at a level contain-ing neurons showing intense retrograde labeling (Fig. 3C)(anteroposterior localization corresponding to Figs. 2F,4C), resulted in anterograde labeling of the supraependy-mal fibers of the third ventricle (Fig. 3D). Localization of
Fig. 4. Schematic representation of transversal sections from theAtlas of Paxinos and Watson (1982), of CT retrogradely labeledneurons. Strongly labeled neurons are represented by dark stars andweakly labeled neurons by empty stars: small stars 5 1 to 10 cells;middle-size stars 5 10 to 20 cells; big stars 5 20 to 30 cells.Rostrocaudal coordinates with reference to ear bars: A, 12.2; B, 11.7;C, 11.2; D, 10.7; E, 10.2; F, 20.3. A, aqueduct; CI, inferior colliculus;CGM, central gray matter; CLi, caudal linear raphe; Cnf, cuneiformnu; DTg, dorsal tegmental nucleus; dtgx, dorsal tegmental decussa-tion; DR, raphe dorsalis; IPA, interpeduncular nucleus, apical; LC,
locus coeruleus; LDTg, laterodorsal nucleus; Me5, nu mesencephalictrigeminal nerve; me5, mesencephalic trigeminal nerve; ml, medianlemniscus; mlf, median longitudinal fasiculus; MnR, median raphe;Pn, pontis; ReIC, recess inf colliculus; RPn, raphe pontis; RRF,retrorubral field; rs, rubrospinal tr; scp, superior cerebellar peduncle;smv, superior medullary velum; SubC, subcoeruleus nu; VTg, ventraltegmental nucleus; xscp, decussation superior cerebellar peduncle; 3,third nuclei; 4, trochlear nuclei; 4n, trochlear nerve; 4V, fourthventricle.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 35

Fig. 5. Photomicrographs of frontal sections processed for trypto-phan hydroxylase (TPOH) immunolabeling at the levels of the raphe(A,B) or third ventricle (C,D).A: TPOH-immunoreactive perikarya areseen in the DRN and CRN of a control rat. B:After 5,7-dihydroxytryp-tamine (5,7-DHT) treatment, the TPOH-positive perikarya are lost.C:Rats grafted with median metencephalon have a SCO reinnervated bya reach plexus of TPOH positive fibers (large arrows), whereas no
supraependymal terminals are detected (fine double arrows). D: Ratsgrafted with rostral metencephalon have a reach plexus of TPOH-positive supraependymal terminals (fine double arrows), whereas onlya few fibers innervate the SCO (large arrows). Rd, raphe dorsalis; Rc,raphe centralis; V, ventricle; SCO, subcommissural organ. A and B,380; C and D, 3370.

the neurons retrogradely labeled by CT is schematicallyrepresented in Figure 4.Given these results indicating a rostrocaudal subdivi-
sion of raphe neurons projecting to the third and fourthventricles, we then dissected the embryonic metencepha-lon (from which the serotonergic neurons of the raphedorsalis and centralis originate) into three adjacent rostro-caudal segments (Fig. 4G), which were then grafted indi-vidually into the third or fourth ventricle of rats in whichthe serotonergic neurons had been previously destroyed by5,7-DHT.
Rats lesioned by 5,7-DHT
Five months after 5,7-DHT injection into the lateralventricle and raphe centralis, apart for a few soma scat-tered rostrocaudally between the medial longitudinal fas-ciculus, no TPOH-immunoreactive neurons were seen inthe raphe dorsalis and centralis (Fig. 5A,B). Tritiated5-HT autoradiography showed that the supra- and sub-ependymal serotonergic plexuses of fibers, which delineatethe ependyma of the third and fourth ventricles (Fig.6A,C–E) in control rats, were not present in 5,7-DHTtreated rats (Fig. 6B,F–H); a few isolated serotonergicfibers were occasionally detected in the parenchyma (Fig.6B).
Rats grafted with embryonicmetencephalic segments
Histology of the embryonic metencephalon before
transplantation. TPOH-immunopositive cells werefound in the region of the metencephalic brainstem be-tween the isthmic fovea and the pontine flexure (Fig. 7A).These cells are situated in the ventral zone; their cellbodies are small (around 10 µm) and bipolar, and theirprocesses primarily elongated in a ventrodorsal direction,consistent with their migration away from the ventricularsurface, as previously described for 14-day-old embryos(Lidov and Molliver, 1982a; Wallace and Lauder, 1983).Localization and histology of the grafts. Trans-
plants injected into the habenular recess of the thirdventricle, rostral to the SCO, were carried away by theCSF and were generally found attached to the habenularrecess, against the posterior commissure (Fig. 7B). Whensuccessfull, transplants injected into the fourth ventricle,via the cisterna magna, were generally found attachedboth to the cerebellum and the floor of the ventricle atabout 2 mm from the interaural line at the rostral level ofthe prepositus hypoglossal nucleus (likely because theywere retained by the fourth ventricular choroid plexuses).The transplants were intensely vascularized, as previ-ously described (Voutsinos et al., 1994), probably by thechoroid plexuses, which makes the ventricle a particularlywell-adapted ‘‘culture bed’’ for the survival of transplantedneurons. Thus, regardless of the origin of the graftedmetencephalic segments (rostral, median or caudal), thetransplants always contained numerous TPOH-immunore-active cells (Fig. 7B,C), with dense ingrowing plexuses offibers. The grafted neurons had themorphological featuresof differentiated 5-HT neurons, being ovoid, fusiform ormultipolar, and similar to their normal counterparts in theadult DRN.Ependymal innervation produced by embryonic met-
encephalic segments grafted into the third ventricle.
The pattern and density of the ependymal innervationproduced by the grafted neurons were dependent on the
rostrocaudal level of the metencephalon from which theembryonic neurons originated. Grafted neurons from therostral part of the embryonic metencephalon projectednumerous outgrowing fibers, whichwere detected by [3H]5-HT binding (Fig. 8A,B) and TPOH immunoreactivity (Fig.5D); these fibers always formed a dense supraependymalplexus covering the apical border of the classic ependymo-cytes on the lateral wall of the third ventricle (Figs. 5D,8B). Irrespective of the rostral graft transplanted, the SCOwere poorly reinnervated (Figs. 5D, 8A). In contrast, graftsfrom the median part of the metencephalon produced farfewer supraependymal terminals (Figs. 5C, 8D), whichnever formed a plexus covering the ependyma, althoughthey did produce dense plexuses of fibers, which wereTPOH-immunopositive (Fig. 5C) or radiolabeled by [3H]5-HT (Fig. 8C), at the basal pole of the SCO specializedependymocytes. The terminals of these plexuses pen-etrated between the ependymocytes in a similar way tothat seen in sham animals (Fig. 6A). Grafts from thecaudal metencephalon produced few serotonergic fibers(which never formed plexuses) innervating the classicependymocytes or the SCO (Fig. 8E,F). Whatever therostrocaudal origin of the grafts, subependymal plexuseswere observed lining the basal border of the ependymo-cytes (Fig. 8B,D,F). As in control animals, the tanycytes,which border the ventral third ventricle, especially at thelevel of the median eminence, were never innervated byserotonergic terminals (data not shown).Ependymal innervation produced by embryonic met-
encephalic segments grafted into the fourth ventricle.
The density of ependymal innervation by metencephalicgrafts was dependent on their rostrocaudal position.Grafted neurons from the caudal and median parts of
the metencephalon (Fig. 9D–I) sent out numerous fibers,covering the velum, located beneath the cerebellum (Fig.9F,H), and the lateral part of the ependyma (Fig. 9D,E); incontrast, the ventromedian ependyma received little or noinnervation (Fig. 9D,E,G,I). Figure 7C shows part of amedianmetencephalic graft, attached to the ventromedianependyma, which contains numerous TPOH-immunoreac-tive neurons, but lacks any supraependymal fiber innerva-tion of the ependyma. Tritiated 5-HT autoradiography alsofailed to detect the presence of supraependymal terminals(data not shown). These data emphasize that embryonicserotonergic neurons are committed to innervate specifictargets.Grafted neurons from the rostral metencephalon pro-
jected only a few very short fibers on to the lateralependyma and velum (Fig. 9A,B), but sent more extensiveoutgrowing fibers to the ventromedian ependyma (Fig.9A,C). The border between the ventromedian and thelateral innervation of the ependyma appeared to be situ-ated below the facial nerve and may correspond to theborder between the prepositus hypoglossal nucleus andthe median vestibular nucleus (Fig. 9E).
DISCUSSION
In this study, we investigated the potential commitment,at the time of axogenesis, of serotonergic embryonic neu-rons to innervate specific mature targets. The study fo-cused on the projections of serotonergic neurons to theependyma, since they form morphologically and biochemi-cally well-defined homogeneous plexuses of fibers, cover-ing the ependyma. In a first step, using retrograde trans-
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 37

Figure 6
38 M. DIDIER-BAZES ET AL.

port of CT, we determined the location, in the midbrainraphe, of those neuronal perikarya that project to theependyma of the third and fourth ventricles in the adultrat. Using heterotopic transplant experiments, we thendemonstrated the ability of the various rostrocaudal seg-ments of the embryonic metencephalon to innervate spe-cific areas of the third and fourth ventricular ependyma inadult rats.CT b subunit, injected into the parenchyma, is a sensi-
tive retrograde and anterograde tracer, which is taken upby cells from the extracellular space (Luppi et al., 1987;Erickson and Blomqvist, 1988; Luppi et al., 1990). It bindsselectively to themonosialoganglioside (GM1) of the plasmamembrane (Van Heyningen, 1974) and is taken up byinternalization of the toxin-bound ganglioside. When in-jected into the ventricles, it is transported in the CSFcirculatory flow through the ventricles, the aqueduct andthe spaces surrounding the CNS (Wright, 1982). Ourresults show that CT was specifically taken up by thesupraependymal terminals, but not by the ependymocytes,indicating the absence of monosialoganglioside on thesecells. The GFAP-positive astrocytes of the subependymalarea or those forming the glia limitans were also capable oftaking up CT. However, CT-labeled subependymal termi-nals were only seen behind the classic ependyma of thethird ventricle and the aqueduct, but not behind the SCOor the ependyma of the fourth ventricle. This pattern ofterminal labeling probably reflects a difference in toxindiffusion, especially at the level of the SCO, where tightjunctions occur between SCO ependymocytes (Gotow andHashimoto, 1982). Subventricular astrocytes, which takeup CT, also form a barrier that limits the passage ofmolecules from the ventricle to the neuropil. Altogether,this could result in a lower uptake of CT by parenchymalterminals than that observed for supraependymal termi-nals. This was corroborated by the fact that the terminalsof the periventricular parenchyma of the third ventricle,which were anterogradely labeled when CTwas injected inthe rostral DRN, took up only a small amount of CTinjected into the third ventricle, resulting in only weaklabeling of the neurons compared with that seen in themore caudal DRN neurons that project directly into theventricles and were highly retrogradely labeled. The ab-sence of labeled neurons in the periaqueductal graymatter
outside the raphe also emphasizes the weak diffusion ofthe toxin from the ventricles to the periaqueductal paren-chyma. Hence, the highly retrogradely labeled neuronsprobably correspond to neurons projecting into the ven-tricles.Consistent with the serotonergic phenotype of the supra-
ependymal plexuses (Chan Palay, 1976), the highly retro-gradely labeled neurons were located in the serotonergicnuclei of the raphe. Considering the large number ofneurons labeled by the toxin and that serotonergic neuronsare highly collaterized (Van der Kooy and Hattori, 1980;Andersen et al., 1983), it is probable that these neuronsalso project to other brain structures, and therefore areinvolved in coordination and/or integration of informationby interconnecting various brain structures. The existenceof two groups of neurons, one, situated in the caudal rapheand labeled when CT is injected into the fourth ventricle,and another, situated in the rostral raphe and labeled onlywhen CT is injected into the third ventricle, suggest arostrocaudal subdivision of those neuronal subsets thatproject to the third and fourth ventricles. Such a subdivi-sion of the DRN into neuronal subsets, projecting intospecific regions of the CNS, has been previously reported(Steinbusch et al., 1981; O’Hearn and Molliver, 1984).However, it is not known whether the organization of theDRN into neuronal subsets innervating different parts ofthe brain is caused by anatomical constraints, whichcompel the growing embryonic axons to take specific fiberpathways, or whether it results from endogenous neuronalpredetermination. By grafting the serotonergic neurons ofrat embryos into adult hosts in an ectopic position, it ispossible to determine whether such neurons have a re-stricted potential to innervate specific targets. The pres-ence of several clusters, located along the rostrocaudal axisof the DRN and innervating the ventricles, makes itpossible to test the hypothesis of serotonergic embryonicneurons being committed to innervate specific areas, de-pending on their rostrocaudal position.The DRN and CRN originate from bilateral cell columns
situated along the entire length of the embryonic meten-cephalon in the basal plate (Lidov and Molliver, 1982a;Wallace and Lauder, 1983), permitting their subdivisioninto several embryonic rostrocaudal segments. We havepreviously shown that embryonic metencephalons graftedinto the third ventricle of host rats are richly vascularizedand contain neurons that survive and differentiate in thesame way as, and have morphological features comparableto, their normal counterparts (Voutsinos et al., 1994).Thus, serotonergic grafted neurons are able to send out-growing terminals toward classic ependyma and SCOwithdestroyed endogenous serotonergic terminals. In the pre-sent study, we have shown that not all serotonergicembryonic neurons are able to innervate the ependymaand that the pattern of innervation of host ependyma byembryonic grafts depends on the rostrocaudal localizationof the serotonergic neurons in the metencephalon. Thus,neurons from rostral grafts mainly form supraependymalplexuses covering the classic ependymocytes of the lateralwall of the third ventricle and the medioventral wall of thefourth ventricle, at the level of the nucleus prepositushypoglossal. In contrast, neurons from the median andcaudal part of themetencephalon send numerous supraepe-ndymal terminals to the lateral wall of the fourth ventricleand the velum, but do not form supraependymal plexusesin the third ventricle. Finally, SCO ependymocytes are
Fig. 6. Frontal sections at the level of the third (A,B) and fourth(C–H) ventricles, showing serotonergic terminals labeled by [3H]5-HTin control rats (A,C–E) and 5,7-DHT-treated rats (B,F–H). [3H]5-HTuptake by serotonergic terminals permits the highly sensitive visual-ization of 5-HT fibers as clusters of silver grains, forming darkfilaments, whereas scattered silver grains correspond to a diffusereaction (Harandi et al., 1986; Voutsinos et al., 1994). A: In the thirdventricle of a control animal, serotonergic terminals form a plexus atthe basal pole of the SCO ependymocytes (curved arrows) and denseplexuses of supra- (arrowheads) and subependymal (large arrows)fibers. B: In a 5,7-DHT-treated rat, the plexuses innervating the SCOand the classic ependyma are absent. A few scattered serotonergicterminals are seen in the neuropil (arrows). C–E: In the fourthventricle of a control animal, supraependymal terminals, detected by[3H]5-HT uptake, are seen in the dorsal wall corresponding to thevelum (C,D) (arrows), onto the lateral ependyma (C) (arrowheads),and the ventromedian ependyma (C,E) (small arrows). F–H: In a5,7-DHT-treated rat, serotonergic terminals are not present. Cb,cerebellum; mlf, median longitudinal fasciculus; V, ventricle; SCO,subcommissural organ; 7, seventh facial nerve. A and B, 3200; C andF, 380; D,E,G,H, 3320.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 39
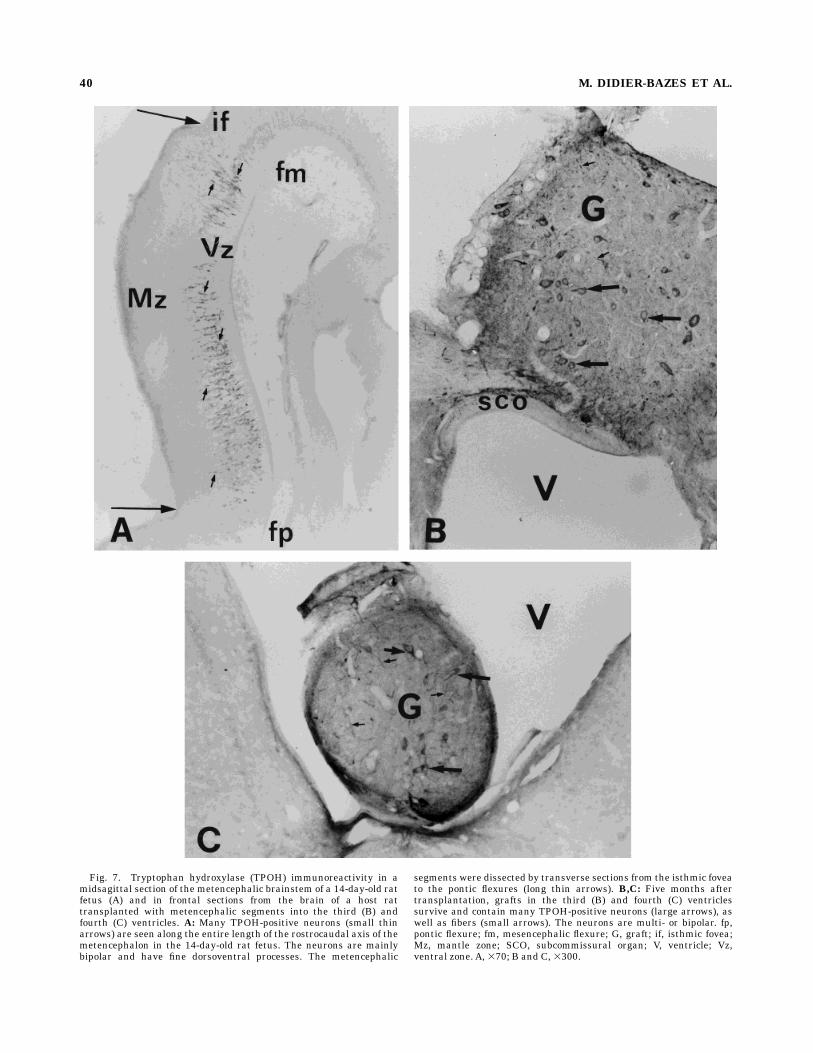
Fig. 7. Tryptophan hydroxylase (TPOH) immunoreactivity in amidsagittal section of the metencephalic brainstem of a 14-day-old ratfetus (A) and in frontal sections from the brain of a host rattransplanted with metencephalic segments into the third (B) andfourth (C) ventricles. A: Many TPOH-positive neurons (small thinarrows) are seen along the entire length of the rostrocaudal axis of themetencephalon in the 14-day-old rat fetus. The neurons are mainlybipolar and have fine dorsoventral processes. The metencephalic
segments were dissected by transverse sections from the isthmic foveato the pontic flexures (long thin arrows). B,C: Five months aftertransplantation, grafts in the third (B) and fourth (C) ventriclessurvive and contain many TPOH-positive neurons (large arrows), aswell as fibers (small arrows). The neurons are multi- or bipolar. fp,pontic flexure; fm, mesencephalic flexure; G, graft; if, isthmic fovea;Mz, mantle zone; SCO, subcommissural organ; V, ventricle; Vz,ventral zone. A, 370; B and C, 3300.
40 M. DIDIER-BAZES ET AL.
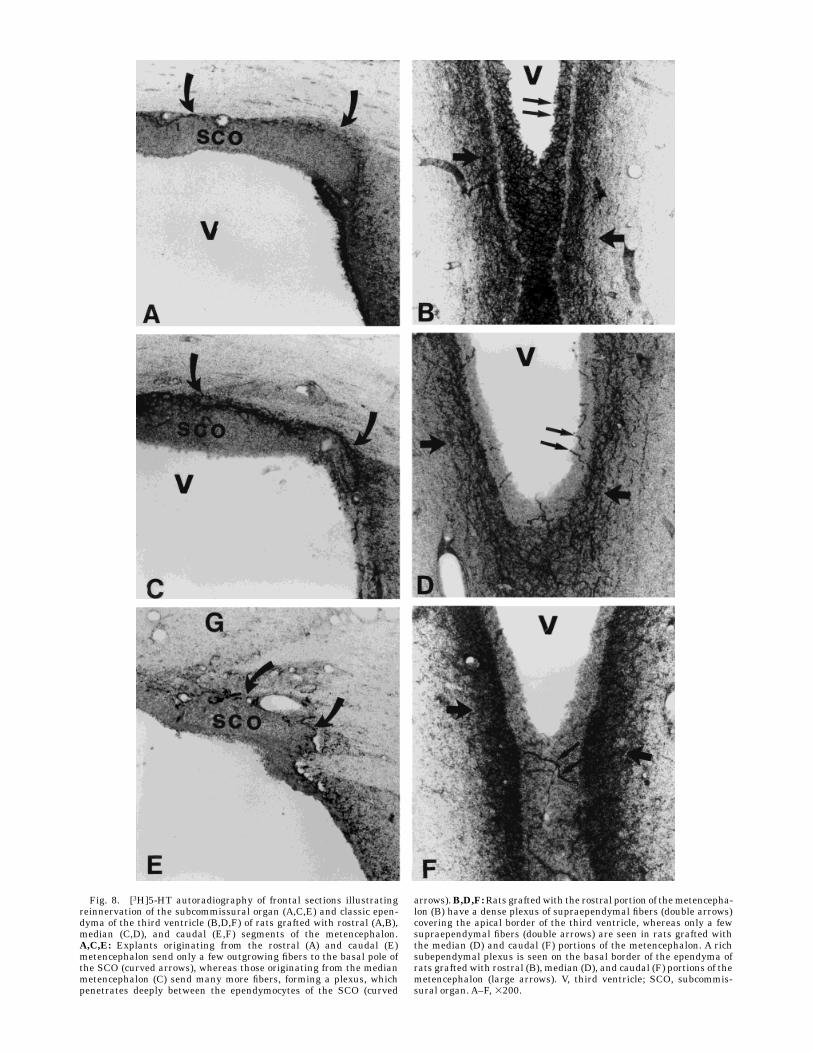
Fig. 8. [3H]5-HT autoradiography of frontal sections illustratingreinnervation of the subcommissural organ (A,C,E) and classic epen-dyma of the third ventricle (B,D,F) of rats grafted with rostral (A,B),median (C,D), and caudal (E,F) segments of the metencephalon.A,C,E: Explants originating from the rostral (A) and caudal (E)metencephalon send only a few outgrowing fibers to the basal pole ofthe SCO (curved arrows), whereas those originating from the medianmetencephalon (C) send many more fibers, forming a plexus, whichpenetrates deeply between the ependymocytes of the SCO (curved
arrows).B,D,F:Rats graftedwith the rostral portion of themetencepha-lon (B) have a dense plexus of supraependymal fibers (double arrows)covering the apical border of the third ventricle, whereas only a fewsupraependymal fibers (double arrows) are seen in rats grafted withthe median (D) and caudal (F) portions of the metencephalon. A richsubependymal plexus is seen on the basal border of the ependyma ofrats grafted with rostral (B), median (D), and caudal (F) portions of themetencephalon (large arrows). V, third ventricle; SCO, subcommis-sural organ. A–F, 3200.

principally innervated by neurons located in the mediansegment. This early commitment of serotonergic embry-onic neurons to innervate different areas of the ventriclesof adult hosts indicates that serotonergic neurons from14-day-old rat embryos form heterogeneous populationswith distinct potentials for innervating mature targets. Italso suggests that these neurons express distinct receptorsfor extracellular neurotropic cues or neurotrophic factors(Letourneau, 1975; Tessier-Lavigne et al., 1988; Thoenen,1991; Goodman and Shatz, 1993). Thus, during embryogen-esis, the growth cones of the various subclasses of seroton-ergic neurons may be sensitive to neurotrophic cues spe-cific for each type of growth cone.Alternatively, serotonergicgrowth cones may respond to a common set of attraction/repulsion cues along pre-existing fiber pathways andselectivity may depend on specific target-released neuro-trophic factors, which permit selective axon elimination orstabilization, as reported for layer 5 cortical neurons(O’Leary and Stanfield, 1989; O’Leary and Koester, 1993).Such a sequential mechanism of target specification isconsistent with the delay period of serotonergic growthcones, seen during development before the establishmentof precise innervation of the target fields (Lidov andMolliver, 1982b). The existence of a delay period alsosuggests a requirement for the targets to be mature tosend signals to the growth cones. The presence of imma-ture cells in the subependymal region (Levison and Gold-man, 1993; Lois and Alvarez-Buylla, 1993) might thusexplain the rich subependymal plexus of non-junctionalserotonergic terminals found in the third ventricle, irrespec-tive of the anteroposterior origin of the embryonicmetence-phalic graft. Indeed, these immature cells may contributeto a persistent embryonic environment favorable to thedevelopment of serotonergic growth cones from all seg-ments, but not to their selection. Thus, serotonergic neu-rons, which have a high potential for regeneration bycollateral sprouting after chemical lesion (Azmitia et al.,1978; Zhou et al., 1995) and to innervate the host brainafter transplantation of fetal neurons (Daszuta et al.,1991; Rajaofetra et al., 1992; Van Luijtellaar et al., 1991)might be committed, at a very early stage during develop-ment, to innervate well-defined targets. This selectivity ofserotonergic neurons for specific targets may also persistinto adulthood, because we have previously shown thatSCO ependymocytes transplanted into the fourth ventricleof adult rats are not innervated by the local supraependy-mal terminals (Didier-Bazes et al., 1991).The formation of subsets of serotonergic neurons inner-
vating the ependyma could be related to the expression ofdifferent co-transmitter phenotypes (Johanson et al., 1981;Gamrani et al., 1984; Hokfelt et al., 1987). However, thisseems unlikely, because the various regions of the supra-ependymal plexuses innervated by the different metence-
phalic segments are homogeneously formed of non-junctional terminals containing both 5-HT and GABA(Belin et al., 1980; Hanrandi et al., 1986). Furthermore,the morphological and biochemical characteristics of theterminals that emerge from the grafted neurons andinnervate their ependymal targets are identical to those oftheir normal counterparts (Voutsinos et al., 1994). Theabsence of serotonergic neurons taking up GABA in disso-ciated cultures of embryonic day 14 rat brain stem (Yama-moto et al., 1981) suggests that the commitment of seroton-ergic neurons toward the GABAergic phenotype occurslater and is dependent on the targets reached by theneurons. In fact, it has been shown, especially in the caseof sympathetic neurons, that targets can cause a switch inneuronal transmitter phenotype (Philippe et al., 1988;Landis, 1990). A similar target influence might explain themorphological and neurochemical homogeneity of the su-praependymal non-junctional 5-HT/GABAterminals, whichcontrasts with the segmental rostrocaudal commitment ofserotonergic neurons innervating the ependyma. The factthat serotonergic neurons that are able to take up GABAcan differentiate and/or survive in vitro only in closecontact with glial cells, while neurons containing only5-HT do not (Belin et al., 1991), supports the hypothesis ofan interaction between glial cells and 5-HT/GABA-containing neurons in the establishment of the neuronalphenotype.The specificity of serotonergic neurons to innervate
selective areas of the ependyma might be related to theearly segmentation of the metencephalon into rhombo-meres, a series of transient transverse domains, distrib-uted along the entire length of the metencephalon andmyelencephalon (Kallen, 1965; Gaunt et al., 1988). Eachrhombomere corresponds to a restricted compartment, theboundary of which cannot be crossed by cells (Fraser et al.,1990). This rhombomerization is dependent on the specificspatial and temporal pattern of expression of homeoboxgenes during early development (reviewed in McGinnisand Krumlauf, 1992). Changes in homeotic genes expres-sion is reported to modify the cellular phenotype andorganization of the developing brain (Chisaka et al., 1992;Kessel, 1993; Zhang et al., 1994). According to the sche-matic maps of McGinnis and Krumlauf (1992) and ofBulfone et al. (1993), the rostral metencephalic segment,which may contain rhombomere R1, and the median andcaudal segments, possibly containing rhombomeres R2and R3, express different sets of homeobox genes, consis-tent with the differential maturation of these regions.Furthermore, the intrinsic positional identity of the rhom-bomeres from an early stage in development is consistentwith a role for this positional identity in neuronal commit-ment, persisting after ectopic transplantation of the rhom-bomeres (Guthrie et al., 1992). Experimental serotonergicinnervation of the ependyma by ectopic transplants of thedifferent rhombomeres could represent a useful model forthe study of a putative role for brain metamerization onthe spatial specification and segregation of neuronal peri-karya and their projections.
CONCLUSION
Our results show that embryonic rat serotonergic neu-rons can be divided into subpopulations that possessdistinct potentialities to innervate mature periventriculartargets as early as embryonic day 14. This finding suggests
TABLE 1. Pattern and Density of Reinnevation of the Ependyma by theRostral, Median, or Caudal Metencephalic Grafts1
Rostralmetencephalon
Medianmetencephalon
Caudalmetencephalon
3VSCO 1 111 1lateral ependyma 111 6 1
4VVelum 6 11 11Lateral ependyma 6 11 11Ventromedian ependyma 11 6 6
1SCO, subcommissural organ; 3V, third ventricle; 4V, fourth ventricle.
42 M. DIDIER-BAZES ET AL.
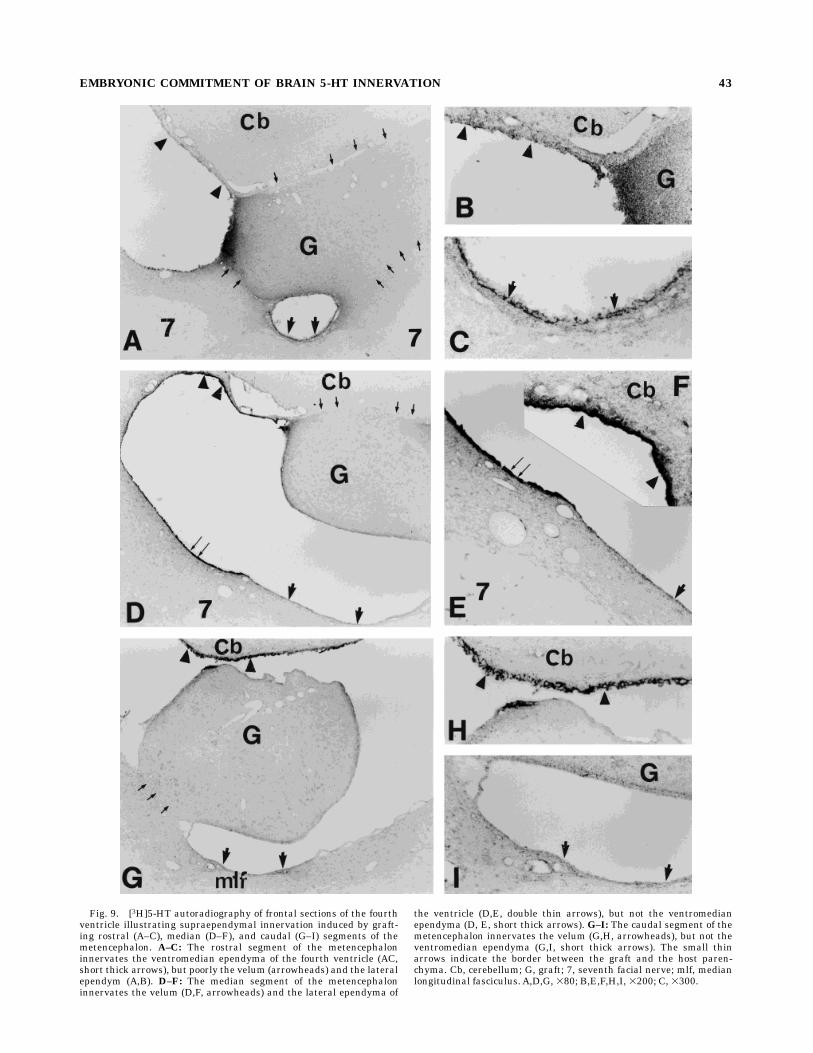
Fig. 9. [3H]5-HT autoradiography of frontal sections of the fourthventricle illustrating supraependymal innervation induced by graft-ing rostral (A–C), median (D–F), and caudal (G–I) segments of themetencephalon. A–C: The rostral segment of the metencephaloninnervates the ventromedian ependyma of the fourth ventricle (AC,short thick arrows), but poorly the velum (arrowheads) and the lateralependym (A,B). D–F: The median segment of the metencephaloninnervates the velum (D,F, arrowheads) and the lateral ependyma of
the ventricle (D,E, double thin arrows), but not the ventromedianependyma (D, E, short thick arrows). G–I: The caudal segment of themetencephalon innervates the velum (G,H, arrowheads), but not theventromedian ependyma (G,I, short thick arrows). The small thinarrows indicate the border between the graft and the host paren-chyma. Cb, cerebellum; G, graft; 7, seventh facial nerve; mlf, medianlongitudinal fasciculus. A,D,G, 380; B,E,F,H,I, 3200; C, 3300.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 43

that serotonergic modulatory neurons, which have numer-ous widespread non-synaptic terminals, are already deter-mined at the time of axogenesis to send out topographi-cally ordered sets of projections as already shown forcortical neurons (reviewed in Franck and Wenner, 1993).This might reflects a more general process available toneuronal systems with widespread projections, especiallythe catecholaminergic system. In addition, the contrastinginnervation of various specific areas of the ependyma bysubsets of serotonergic neurons raises new questions onthe role of this puzzling ventricular innervation. Consider-ing the large number of neurons projecting to the epen-dyma, it is probable that each neuron also projects to otherbrain structures. Thus, the regional selectivity of ependy-mal innervation by subsets of serotonergic neurons mightreflect the involvement of the different ventricular regionsin distinct functional neuronal circuits.
ACKNOWLEDGMENTS
The authors thank V. Thivolle for typing the manuscriptand T. Barkas for revision of the English text. This workwas supported by INSERMU433.
LITERATURE CITED
Andersen, E., B. Rigor, and N. Dafny (1983) Electrophysiological evidenceof concurrent dorsal raphe input to caudate, septum, habenula, thala-mus, hippocampus, cerebellum and olfactory bulb. Int. J. Neurosci.18:107–116.
Azmitia, E.C. (1978) The serotonin-producing neurons of the midbrainmedian and dorsal raphe nuclei. In L.L. Iversen, S.D. Iversen, and S.H.Snyder, (eds): Handbook of Psychopharmacology, Vol. 9. New York:Plenum, pp. 233–314.
Azmitia, E.C., A.M. Buchan, and J.H. Williams (1978) Structural andfonctional restoration by collateral sprouting of hippocampal 5-HTaxons. Nature 274:374–377.
Belin, M.F., H. Gamrani, M. Aguera, A. Calas, and J.F. Pujol (1980)Selective uptake of (3H) gamma-aminobutyrate by rat supra- andsubependymal nerve fibers, histological and high resolution radioauto-graphic study. Neuroscience 5:241–254.
Belin, M.F., M. Fevre-Montange, A. Reboul, M. Didier-Bazes, M. Ehret, M.Maitre, and M. Tardy (1991) Primary dissociated cell culture ofembryonic rat metencephalon: presence of GABA in serotonergic neu-rons. Neurosci. Lett. 125:101–106.
Bouchaud, C., andM.Arluison (1977) Serotoninergic innervation of ependy-mal cells in the rat subcommissural organ. A fluorescence electronmicroscopic and radioautographic study. Biol. Cell 30:61–64.
Bulfone, A., L. Puelles, M.H. Porteus, M.A. Frohman, G.R. Martin, andJ.L.R. Rubenstein (1993) Spatially restricted expression of Dlx-1, Dlx-2(Tes-1), Gbx-2, and Wnt-3 in the embryonic day 12.5 mouse forebraindefines potential transverse and longitudinal segmental boundaries. J.Neurosci. 13:3155–3172.
Cash, C.D., P. Vayer, P. Mandel, and M. Maitre (1985) Tryptophan5-hydroxylase rapid purification from whole rat brain and production ofspecific antiserum. Eur. J. Biochem. 149:239–245.
Chan Palay, V. (1976) Serotonin axons in the supra- and subependymalplexus and leptomeninges: Their roles in local alteration of cerebrospi-nal fluid and vasomotor activity. Brain Res. 102:103–130.
Chisaka, O., T.S. Musci, andM.R. Capecchi (1992) Developmental defects ofthe ear, cranial nerves and hindbrain resulting from targeted disrup-tion of the mouse homeobox gene Hox-I,6. Nature 355:516–520.
Dahlstrom,A., andK. Fuxe (1964) Evidence for the existence of monoamine-containing neurons in the central nervous system. 1. Demonstration ofmonoamines in the cell bodies of brainstem neurons. Acta Physiol.Scand. 62(Suppl. 232):1–55.
Daszuta, A., G. Chazal, S. Garcia, S. Oleskevich, and L. Descarries (1991)Ultrastructural features of serotonin neurons grafted to adult rathippocampus: An immunocytochemical analysis of their cell bodies andaxon terminals. Neuroscience 42:793–811.
Didier-Bazes, M., L. Chouaf, H. Hardin, M. Aguera, M. Fevre-Montange,and M.F. Belin (1991) Developmental neuron-glia interaction: Role of
the serotonin innervation upon the onset of GABA uptake into theependymocytes of the rat subcommissural organ. Dev. Brain Res.63:135–139.
Erickson, H., andA. Blomqvist (1988) Tracing of neuronal connections withcholera toxin subunit B: Light and electron microscopic immunohisto-chemistry using monoclonal antibodies. J. Neurosci. Methods 24:225–235.
Frank, E., and P. Wenner (1993) Environmental specification of neuronalconnectivity. Neuron 10:779–785.
Fraser, S., R. Keynes, and A. Lumsden (1990) Segmentation in the chickembryo hindbrain is defined by cell lineage restrictions. Nature 344:431–435.
Gamrani, H., M. Harandi, M.F. Belin, M.P. Dubois, and A. Calas (1984)Direct electron microscopic evidence for the coexistence of GABAuptake and endogenous serotonin in the same rat central neurons bycoupled radioautographic and immunohistochemical procedures. Neu-rosci. Lett. 48:25–30.
Gaunt, S.J., P.T. Sharpe, and D. Duboule (1988) Spatially restricteddomains of homeo-gene transcripts in mouse embryos: Relation to asegmented body plan. Development 104(suppl.):169–179.
Goodman, C.S., and C.J. Shatz (1993) Developmental mechanisms thatgenerate precise patterns of neuronal connectivity. Cell 72/Neuron 10:(Suppl.) 77–98.
Gotow, T., and P.H. Hashimoto (1982) Intercellular junctions betweenspecialized ependymal cells in the subcommissural organ of the rat. J.Neurocytol. 11:363–379.
Grove, E.A., B.P. Williams, D.Q. Li, M. Hajihosseini, A. Friedrich, and J.Price (1993) Multiple restricted lineages in the embryonic rat cerabralcortex. Development 117:553–561.
Guthrie, S., I. Muchamore, A. Kuroiwa, H. Marshall, R. Krumlauf, and A.Lumsden (1992) Neuroectodermal autonomy of Hox-2.9 expressionrevealed by rhombomere transpositions. Nature 356:157–159.
Harandi, M., M. Didier, M. Aguera, A. Calas, and M.F. Belin (1986) GABAand serotonin (5-HT) pattern in the supraependymal fibers of the ratepithalamus: Combined radioautographic and immunocytochemicalstudies. Effect of 5-HT content on 3H GABA accumulation. Brain Res.370:241–249.
Hokfelt, T., D. Millhorn, K. Seroogy, Y. Tsuruo, S. Ceccatelli, B. Lindh, B.Meister, T. Melander, M. Schalling, T. Bartfai, and L. Terenius (1987)Coexistence of peptides with classical neurotransmitters. Experientia43:768–780.
Hsu, S., L. Raine, and H. Fanger (1981) A comparative study of the GFAPantiperoxidase method and an avidin biotin complex method forstudying polypeptides hormones with radioimmunoassay antibodies. J.Clin. Pathol. 75:734–738.
Jessell, T.M. (1988) Adhesion molecules and the hierarchy of neuraldevelopment. Neuron 1:3–13.
Johansson, O., T. Hokfelt, B. Pernow, S.L. Jeffcoate, N. White, H.W.M.Steinbusch, A.A.J. Verhofstad, P.C. Emson, and E. Spindel (1981)Immunohistochemical support for three putative transmitters in oneneuron: Coexistence of 5-hydroxytryptamine, substance P- and thyrotro-pin releasing hormone-like immunoreactivity in medullary neuronsprojecting to the spinal cord. Neuroscience 6:1857–1881.
Kallen, B. (1965) Early morphogenesis and pattern formation in the centralnervous system. In R.L. De Haan, and H. Ursprung (eds): Organogen-esis. NewYork: Holt, Rhinehart and Wilson, pp. 107–128.
Kessel, M. (1993) Reversal of axonal pathways from rhombomere 3correlates with extra Hox expression domains. Neuron 10:379–393.
Krushel, L.A., J.G. Johnston, G. Fishell, R. Tibshirani, and D. VanDer Kooy(1993) Spatially localized neuronal cell lineages in the developingmammalian forebrain. Neuroscience 53:1035–1047.
Landis, S.C. (1990) Target regulation of neurotransmitter phenotype.Trends Neurosci. 8:344–350.
Lauder, J.M., and F.E. Bloom (1974) Ontogeny of monoamine neurons inthe locus coeruleus, raphe nuclei and substantia nigra of the rat. J.Comp. Neurol. 155:469–482.
Lauder, J.M., J.A. Wallace, H. Krebs, P. Petruz, and K. McCarthy (1982) Invivo and in vitro development of serotonergic neurons. Brain Res. Bull.9:605–625.
Letourneau, P.C. (1975) Cell-to-substratum adhesion and guidance ofaxonal elongation. Dev. Biol. 44:92–101.
Levison, S.W., and J.E. Goldman (1993) Both oligodendrocytes and astro-cytes develop from progenitors in the subventricular zone of postnatalrat forebrain. Neuron 10:201–212.
44 M. DIDIER-BAZES ET AL.

Lidov, H.G.W., and M.E. Molliver (1982a) Immunohistochemical study ofthe development of serotonergic neurons in the rat CNS. Brain Res.Bull. 9:559–604.
Lidov, H.G.W., andM.E.Molliver (1982b)An immunohistochemical study ofserotonin neuron development in the rat: Ascending pathways andterminal fields. Brain Res. Bull. 8:389–430.
Lois, C., and A. Alvarez-Buylla (1993) Proliferating subventricular zonecells in the adult mammalian forebrain can differentiate into neuronsand glia. Proc. Natl. Acad. Sci. USA 90:2074–2077.
Lorez, H.P., and J.G. Richard (1975) Supra-ependymal 5-HT axons in therat forebrain after electrolytic and 5–6 DHT induced lesions of themedial forebrain bundle. Brain Res. 100:1–12.
Luppi, P.H., K. Sakai, D. Salvert, P. Fort, and M. Jouvet (1987) Peptidergichypothalamic afferents to the cat raphe pallidus as revealed by a doubleimmunostaining technique using unconjugated cholera toxin as retro-grade tracer. Brain Res. 402:339–345.
Luppi, P.H., P. Fort, and M. Jouvet (1990) Iontophoretic application ofunconjugated cholera toxin B subunit (CTb) combined with immuno-histochemistry of neurochemical substances: A method for transmitteridentification of retrogradely labeled neurons. Brain Res. 534:209–224.
McConnel, S.K., and C.E. Kaznowski (1991) Cell cycle dependence oflaminar determination in developing neocortex. Science 254:282–285.
McGinnis, W.R., and R. Krumlauf (1992) Homeobox genes and axialpatterning. Cell 68:283–302.
O’Hearn, E., and M.E. Molliver (1984) Organization of raphe corticalprojections in rat: A quantative retrograde study. Brain Res. Bull.13:709–726.
O’Leary, D.D., and B.B. Stanfield (1989) Selective elimination of axonsextended by developing cortical neurons is dependent on regionallocale: Experiments utilizing fetal cortical transplants. J. Neurosci.9:2230–2246.
O’Leary, D.D., and S.E. Koester (1993) Development of projection neurontypes, axon patways, and patterned connections of the mammaliancortex. Neuron 10:991–1006.
Patterson, P.H. (1988) On the importance of being inhibited, or say no togrowth cones. Neuron 1:263–267.
Paxinos, G., and C. Watson (1982) The Rat Brain in Stereotaxic Coordi-nates. Orlando, FL: Academic Press.
Peyron, C., P.H. Luppi, P. Fort, C. Rampon, and M. Jouvet (1996) Lowerbrainstem catecholamine afferents to the rat dorsal raphe nucleus. J.Comp. Neurol. 364:402–413.
Philippe, E., M. Garosiand, and B. Droz (1988) Influence of peripheral andcentral targets on subpopulations of sensory neurons expressing calbin-
din immunoreactivity in the dorsal root ganglion of the chick embryo.Neuroscience 26:225–232.
Rajaofetra, N., N. Konig, P. Poulat, L. Marlier, F. Sandillon, M.J. Drian, M.Geffard, and A. Privat (1992) Fate of B1–B2 and B3 rhombencephaliccells transplanted into the transected spinal cord of adult rats: Lightand electron microscopic studies. Exp. Neurol. 117:59–70.
Steinbusch, H.W.M., R. Nieuwenhuys, A.A.J. Verhofstad, and D. Van DerKooy (1981) The nucleus dorsalis raphe of the rat and its projectionupon the caudatoputamen.Acombined cytoarchitectonic, immunohisto-chemical and retrograde transport study. J. Physiol. Paris 77:157–174.
Tessier-Lavigne, M., M. Placzek, A.G.S. Lumsden, J. Dodd, and T.M. Jessel(1988) Chemotropic guidance of developing axons in the mammaliancentral nervous system. Nature 336:775–778.
Thoenen H. (1991) The changing scene of neurotrophic factors. TrendsNeurosci. 14:165–170.
Van der Kooy, D., and T. Hattori (1980) Dorsal raphe cells with collateralprojections to the caudate putamen and substantia nigra: A fluorescentretrograde double labeling study in the rat. Brain Res. 186:1–7.
Van Luijtellaar, M.G.P.A., A.D.M. Tonnaer, A.L. Frankhuijzen, H. Dijkstra,J.J. Hagan, andW.M. Steinbusch (1991) Morphological, neurochemical,and behavioural studies on serotonergic denervation and graft-inducedreinnervation of the rat hippocampus. Neuroscience 42:365–377.
Van Heyningen, W. (1974) Gangliosides as membrane receptors for tetanustoxin, cholera toxin and serotonin. Nature 249:415–417.
Voutsinos, B., L. Chouaf, P. Mertens, P. Ruiz-Flandes, Y. Joubert, M.F.Belin, and M. Didier-Bazes (1994) Tropism of serotonergic neuronstowards glial targets in the rat ependyma. Neuroscience 59:663–672.
Wallace, J., and J.M. Lauder (1983) Development of the serotonergicsystem in the rat embryo: An immunocytochemical study. Brain Res.Bull. 10:459–479.
Weissman, D., M.F. Belin, M.Aguera, C. Meunier, M. Maitre, C.D. Cash, M.Ehret, P. Mandel, and J.F. Pujol (1987) Immunohistochemistry oftryptophan hydroxylase in the rat brain. Neuroscience 23:291–304.
Wright, E.M. (1982) Secretion and circulation of the cerebrospinal fluid.Front. Horm. Res. 9:4–14.
Yamamoto, M., H.W.M. Steinbusch, and T.M. Jessel (1981) Differentialproperties of identified serotonin neurons in dissociated cultures ofembryonic rat brain stem. J. Cell Biol. 91:142–152.
Zhang, M., H.J. Kim, H. Marshall, M. Gendron-Maguire, D.A. Lucas, A.Baron, L.J. Gudas, T. Gridley, R. Krumlauf, and J.F. Grippo (1994)Ectopic Hoxa i induces rhombomere tansformation in mouse hindbrain.Development 120:2431–2442.
Zhou, F.C., E.C. Azmitia, and S. Bledsoe (1995) Rapid serotonergic fibersprouting in response to ibotenic acid lesion in the striatum andhippocampus. Dev. Brain Res. 84:89–98.
EMBRYONIC COMMITMENT OF BRAIN 5-HT INNERVATION 45





























