Embed Size (px)
Citation preview

Plant Physiol. (1967) 42, 900-906
The Specific Nature of Plant Cell Wall Polysaccharides'Donald J. Nevins2, Patricia D. English3, and Peter AlbersheimDepartment of Chemistry, University of Colorado, Boulder, Colorado 80302
Received February 13, 1967.
Summa111I(-r1Y. Polysaccharide compositions of cell walls were assessed by quantitativeanialyses of the component sugars. Cell walls were hydrolyzed in 2 N trifluoroaceticacid and the liberated stugars reduced to their respective aldiltols. The alditols wereacetylated and the resulting alditol acetates separated by gas chromatography. Quan-ti,tative assay o,f the alditol acetates was accomplished by electronically integratingthe detector outputt of the gas chromatograph. Iyo-inositol, introduced into thesample pri,or to hydrolysis, served as anl internal standard.
The cell wall polysaccharide compositions of plant varieties within a given speciesare essenti,ally identical. However, differences in the stugar composition were ob-served in cell walls prepared from different species of the same as well as o,f differentgenera. The fact that the wNal,l com,positi,ons of different varieties of the same speciesare the same indicates that the biosynthesis of cell wall polysaccharides is geneticallyregullated. The cell wal,l,s of various morphological parts (roots, hypocotyls, firstinternodes and primary le,aves) of bean plants were each found to have a characteristicsugar composition.
It was found that the cel,l wall stugar composition of suspension-cultured sycamorecells could be altered by growing the cells on different carbon sources. This demon-strates that the biosynthesis of cell wall polysaccharides can be manipulated withoutfatal consequences.
The cell walls of higher plants are complexorgainelles which are composed primarily of poly-saccharide aInd protein (14). In addition, thepresence of smaller amoounts of lipid (18) aindribonutcleic acid (12, 17) constituents has been re-ported, andl a intumber of enzymies have beeni foulndto be localize,d within the cell wall (16). Thestructulral complexity anid anato,mical distinctivenessof dlifferent cell walls are well known from lightanid, more recently, electroni microscope stuidies(1,5,9,22). The structtural complexity of the cellwall anid the pre,sence there of various metabolicallyimportaint components suiggest that the wall imiayhave other functions in addition to its establishedrole of providing mechanical support for the cell.
Polysacchariles are the major constituents ofcell walls. T,he macromole,cules involved are com-posed of both ineuitral sugars and uironiic acids (2).The compositions of cell wall polysaccharides differsomewhat in walls prepare,d from plaints belonigiingto (lifferent families (6, 20). Furthermore, the
1 Supported in part by a grant from the United StatesAtomic Energy Commission No. AT(11-1)-1426.
2 Postdoctoral Fellow of the National Institute ofGeneral Medical Sciences (National Institutes of Health,No. 5-F2-GM-29, 861-02).
3 Predoctoral trainee under National Institutes ofHealth Training Grant HD-172.
compo-sition of wvalls aippears to have a role incelltular growth, for it has been demonstrated thatthe polysaccharide composition chainges duiriing de-velopment (20). Changes in the cell wall poly-saccharide oomposition have also been correlatedwith indoleacetic acid-induced eloingationi of oatcoleoptiles (19). However, the degree to whichthe cell has control over cell wall polysaccharidesynithesis aind the degree of absolute consitancy ofthe polysaccharide comipositi,on of higher plant cel'walls remain to be established. If the cell wallswere randomly synthesized, small chaniges in poly-saccharide composition might have no significance,as these changes c0ould not be correlated with suchwall rellated phentometna as growth. I,f the com-position1 of the wall were precise, however, smallchan1ges in compositilon coutld be of great importance.
One aspect of otur work has beeni concerned withhost-pathogen initerrelationships. In geineral, plantpathogens are able to prodtuce enzymies capable ofbreakinIg down polysacchari,des. 1)ifferences be-tween resistant and susceptilble plaints may involve,at least in part, the ability of the wall polysac-charides to resist enzymatic degradation. In orderto learn whether wall polysaceharides have an im-portant function in disease resi,stance, it must bedemonstrated that cell walls have specific poly-saccharide compositions, composi,tions that are exactand under genetic control. The results reported in
900
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

NEVINS ET AL.-SPECIFIC WALL POLYSACCHARIDES
this paper demonstrate that the composition of cellwall polysaccharides is very precisely determine-dand that the biosynthesis of these molecules is sub-ject to genetic control.
Materials and Methods
Plant .1Materials and Wall Preparation. Seedsof comimon bean (Phaseolus vulgaris L. var. RedKidney, Pinto, and Small W-hiite), mung bean(Phiseoluis aulreus Roxb.), soybean (Glycine maxMerr. var. Harosoy and Harosoy 63), barley(IHordeumt vulgare L. var. Atlas and Atlas 46),and corn (Zea Hnays L. var. Golden r,ocket) weresuirface-sterilized by soaking for 15 minutes in an0.5 % sodium hypoch'lorite (10 % Chlorox) solution.The seeds were planted in trays containing ver-miculite and were watered daily with a one-halfstrength Hoagland's nutrient sol-ution (10). Ironwas suipplied as Fe++-EDTA at a concentration of42 jum. The seedlings were illuminated by a com-mercial reflector fixtuire containing 2 4OW Sylvania"Grow Lux" fluorescent lights apprroximately 15cm above the leaves. A photoperiod regime of 14hours of light followed by 10 hours of darktnesswas employed, and the room temperature was main-tainedl at 25 -+ 20. All planits were harvested 18days after planting with the exception that thebarley was harvested after 12 days. The be-anplants were subdivided into roots, hypocotyls, firstinternodes (between cotyledonary node and thejulnctutre of the first leaves) and primary leafblades, the barley seedlings into leaf blades androots, and the corn seedlings into roots and intoindividual leaves. In an effort to reduice theamouint of starch present in the cells, all plants wereharvested at the end of the dark period. Samplesof plant material were frozen and stored in poly-ethylene bags at -300.
Cell walls were prepared by chopping the frozentissule into small pieces (about 1 mm) with a razorblade. The chopped material was ground in a\Varing Blendor with approximately 10 volumes(v/w) of cold 100 mzis potassiulm phosphate buffer(pH 7.0). The insoluble material was collected bycentrifugation at 2000 X g for 15 minutes. Thesupernatant fluid was decanted; the precipitate wasresuspended in fresh phosphate buffer, and againcentriifulged. This washing procedure was repeated3 times. The residue was then suspended in acetoneand ground with 10 volumes (v/w) of cold acetonein the Waring Blendor. The residule (i.e. cellwalls) was collected by filtering the acetone sus-pension through 'Whatman No. 1 paper in a Buchnerfunnel. The cell walls were washed 3 times withcold acetone and dried at room temperatuire. Allextraction operations were conducted at 2°. Wehave found that no monomeric sugars are presentin the cell wall material following this extractionprocedure.
Sycamore (Acer pseuidoplatanus L.) cells weregrown in suspension culture in a modified M-6mediuim (21) containing 1 of the following carbonsources: sucrose, maltose, gluco-se, galactose, man-nose or glycerol. Details concerning the growth ofsycamore cells in suspension culture have beenpublished (3). Cells were removed from the cul-ture medium by filtration through a coarse sinteredglass funnel and washed with 300 iml of 100 mMpotassitum phosphate btuffer. The cells were re-suispended in an equal volume (w/v) of the phos-phate bufifer aind broken in a French pressture cellunder a pressure of 1000 pounds per square inch.The cell walls were collected onI a coarse siinteredglass funlnel and washed with several volumes ofbuffer. The walls were resuspended in buffer(1:1 w/v) and again passed through the Frenchpressture cell to ensure that all cells were broken.The suspension was again filtered and the cellwalls washed several more times with buffer. Thiswas followed by extraction with chloroform:methanol (1:1 v/v) and finally with acetone. Thecell wvalls were then air dried at room temperatutre.
Analysis of Cell Walls for Constituent Sugars.The air-dried cell wall samples were further driedovernight in a vacuum dessicator in the presenceof phosphorus pentoxide. Twenty mg samples ofthe dried wall material were weighed into 13 X 100m-m test tubes. Exactly 2 ml of a 2 N trifluoro-acetic acid solution containing 0.5 mg myo-inositolper ml (internal standard) was introduced into thetest tube, and the tube sealed. The sample washeated at 121° for 1 hour. Following hydrolysis,the tube was opened and the residtue (unhydrolyzedportion of the wall) separated from the supernatantfluiid by centrifugation in a clinical centrifuge(1500 X g, 3 min). The supernatant fluid wastransferred to another test tube, and the residuewashed 3 times by resuspension in 0.5 ml portionsof 70 % (v/v) ethanol followed by recentriffugation.The combined supernatant fluid and /70 % ethanolwashings (containing the solt'ble suigars) wereevaporated to dryness at 50° with a stream offiltered air directed on to the fluid surface.
The monosaccharides in the samples were con-verted to their respective alditol acetates, and thesederivatives were quantitatively assayed by gaschromatography. This was accomplished by reduc-tion with 'sodiuim borohvdride, followed bv 5 meth-anol evaporations to remove the borate. The driedsamiple was acetylated in either 1.0 or 0.5 ml ofacetic anhydride bv heating at 1210 for 3 hours.After acetylationi the sa,mples were transferred to3 ml serum v,als, stoppered and kept at -300. One,11 samples of the acetylation mixtures were injectedinto the column of either of 2 Model 810 F & Mduial column gas chromatographs. Separation wasachieved oIn 3 mm (OD) X 120 cm copper columnscontaining approximately 1.5 g of Gas-Chrom Pcoated with a mixture of 0.2 % ethylene glycol adi-pate, 0.2 % ethylene glycol succinate and 0.4 %
901
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
XF1155O (Applied Science Laboratories Inc., StateCollege, Penna.). Thealditol acetatesuweredetectedby flame ionization. The conditions employed forgas chromatography were as follows: injection porttemperatulre 180 to 1900; initial coltumn temperature1200 held constant for 7 mintutes after injection,followed by a programmed temperatuire increase of10 per minutte to 1800; detector temperature 2700;heliuim carrier gas flow 90 ml min-m; oxygen an(dhy-drogen flow rates 100 ancl 45 miii- respectively;electrometer settings range 10, attenutation 1.
The detector output of the gas chromatographwas recorded on magnetic tape and the responsepeaks integrated at a later time by means of anIn fotronics playback an(d electronic integrator sys-tem. The integration data were converted to thealbsoluite amotlunts of the vrarious suigars by dividingby appropriate stan(lard valtues. The standlar(dvaluies represent the integrated detector responsefor 1 pkg of suigar divided by the dletector responseof 1 ,lg of myo-inositol. The following valtueswere used: rhamnose 0.44: fucose 0.35;arabinose - 0.42; xylose - 0.54; mannose 0.65;galactose - 0.60; gltucose = n.8s.
All restults presented in this paper represenit themean of at least 2 analyses. We have found thatthis is suifficient to yield data with an accturacyof + 5 %.
(AX detailed description of the proceduire forsuigar analysis has been accepted for ptublication inCarbohydrate Research.)
ResultsIt ux-as not ouir inteintioin in this sttudy- to tunder-
take a comprehensive analysis of the cell walls ofthe taxoniomic hierarchy of plants, for stuch a taskwotuld be overwvhelming evTen if und(lertakeni on alimited scale. Rather, we have attempted to ascer-taini the (legree of specificity inherenit in cell wallpolysaccharides b) stuidying in (letail the polysac-charide composition of cell walls from a selectednumber of agricuiltulral planit varieties andl relatedlspecies and genera. By working with agrictultuiralxvarieties it was possiblle to obtain planits withsimilar genotypes. The selection of the differentvarieties, howe'ver, was arbitrary, as we have littlebasis for knoxving what, if any, phenotypic char-acters might iindicate differences in composition otthe cell wall. .Nevertheless, if the biosynthesis ofthe cell wall is itinder rigid genetic control, thenisimilarities inl wall composition between varietiesof a given species shouil(d be apparent. In order toassess the e,ffect of the genotype oni the compositionof cell wx all polysaccharides, comparisons betweenrelated species aind genera were made. Ouir atteni-tioni was focused -within the Legiumlinaos(ue andGrainijcae families, representatives of the dicoty-ledonis and monocotVyedons respectiv elv.
PolYsa ccha ride Co( 'l;psitio n of Cell IWValls ofPhaseoluis Species. The wall p lxysaccblaride com-
It VI fIa a II.I a .IOYI
o0*11.1.
II-- ,.-
II-.
R AXM.G AX-P C-r- R . -- - . II -I.^,I . . - *-^ XI AGemr Ci R F A X M C_ G' R F A X M C- CB
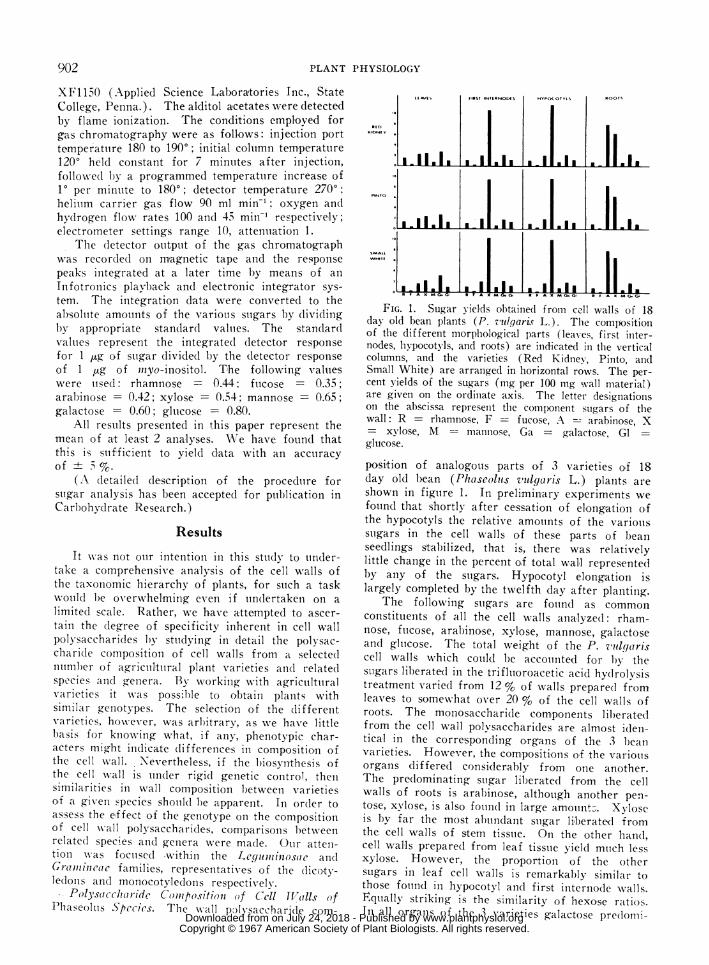
FIG. 1. Sugar vields obtained from cell walls of 18day old bean plants (P. zulgaris L.). Tile compositionof the different morplhological parts (leaves, first inter-nodes, hypocotvls, and roots) are indicated in the verticalcolumns, and the varieties (Red Kidney, Pinto, andSmall White) are arranged in horizontal rows. The per-cent yields of the sugars (mg per 100 mg wall material)are given on the ordiniate axis. The letter designationsoni the abscissa represent the component sugars of thewall: R = rhamnose, F - fucose, A - arabinose, X- xvlose, M m-nanniose, Ga = galactose, Glglucose.
position of analogous parts of 3 varieties of 18day old bean (Phascoluts vldgaris L.) plants areshown in figture 1. In preliminary experiments wefound that shortly after cessation of elongation ofthe hypocotyls the relative amouints of the variouisstigars in the cell walls of these parts of l)eanseedlings sltabilized, that is, there was relativelylittle chainge in the percent of total wall representedby any of the stugars. Hypocotyl elongation islargely completed by the twelfth day after plantiing.
The following suigars are foulnd as commonconlstitueints of all the cell walls ana,lyzed: rham-nose, ftucose, arabinoise, xylose, mannose, galactoseand glulcose. The total weight of the P. vulgariscell walls which couild be accoutnlted for by thesuigars liberated in the trifluioroacetic acid hy(drolysistreatment varied from 12 % of walls prepared fromleaves to somewhat over 20 % of the cell walls ofroots. The monosaccharide components liberatedfrom the cell wall polysaccharides are almost ideni-tical in the corre;sponding organs of the 3 beanvarieties. However, the compositioons of the variouisorgans differed considerably from one another.The predominating suigar liberated from the cellwalls of roots is arabinose, althotugh another pen-tose, xvlose, is also fouind in large amount:;. Xyloseis by far the most abundant suigar li,berated fromthe cell walls of stem tisstue. On the other hand,cell walls prepared from leaf tissuie yield muich lessxylose. However, the proportion of the othersugars ill leaf cell wvalls is remarkably similar tothose fouind in hypocotyl and first internode w\alls.Equally striking is the similaritv of hexose ratios.In all orgains of the 3 varieties galactose pre(lomiii-
I
I
902
s mis
m a
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
NEVINS ET AL.-SPECIFIC WALL POLYSACCHARIDES
LEAVIS FIRST WTERPOODS HYPOCOTYLS ROOTS
AA X M G. RF!^XM! R F A X MXL F LtX
FIG. 2. Sugar y-ields obtained from cell walls of 4morphological parts of 18 day old mung bean plants (P.aurcus Roxb.). The percent yields are indicated onthe ordinate (mg per 100 mg wall material) and thevarious sugars are designated on the abscissa (see fig 1).
nates, followed by glucose and then by mannosewhich is found in rather minor amounts (less than1 mg/100 mg of wall). Rhamnose is found athigher concentrations in leaves than in the othertissuies analyzed. Fucose is detected in amounts of0.2 to 0.3 % in hydrolysates of all the P. zxulgaristissues studied.
The cell wall compositions of the variotus mor-phological parts of mung bean (Phaseolis aureous)(fig 2) differ significantly from P. vutlgaris.Marked differences between these 2 species areparticularly apparent in the arabinose:xylose ratiosof the r-oot cell walls. The absolute amount ofxylose found in the hypocotyl and first internodetissue is 40 to 60 % less in the mung bean speciesthan in the respective tissues of the common bean.On the other hand, the amount of arabinose isgreater in mting bean leaves than in the leaves ofthe common bean.
Polvsaicch (ri(de Comizpositiont of Cell Walls ofGlycine max. The polysaccharide composition ofthe cell walls prepared from the various morpho-logical parts of 2 varieties of soybean (G. wax)is shown in figture 3. The suigar compositions ofthe cell walls of the 2 varieties are virtually iden-tical. The relative proportions of the variousstigars in the soybean cell walls differ from thoseof the Phaseoluis species. In soybeans, the cellwalls of the 2 stem tissues, the hypccotyl and thefirst internode, differ markedly from each other.In the walls prepared from the hypocotyl there isa significantly larger yield of arabinose than in thewalls prepared from the first internode. Further-more, trifltuoroacetic acid hydrolysis treatmentliberated as monosaccharides a larger proportion of
FIG. 3. Sugar yields obtained from cell walls of 4morphological parts of 18 day old soybean plants (G. max
Merr.). The Harosoy variety is represented by openbars, and the Harosoy 63 by solid bars. The percentyields are indicated on the ordinate (mg per 100 mgwall material) and the various sugars are designatedon the abscissa (see fig 1).
903
the cell walls of soybean leaves (18 % of wallweight) than of the cell walls of Phaseolus leaves(12 % of wall weight). It is noteworthy that theleaf cell wall polysaccharides in soybean containrela,tively large amounts of rhamnose and fucose,almost 2 % of each. Actually, the compositions ofthe soybean walls are more similar to those of truebeans than to those of mung beans. This is par-ticularly evident in the arabinose to xylose ratios ofthe roots and first internodes.
Polysaccharide Comiiposition of Cell Walls ofBarley and Cornt. Evidence for the specificity ofcell wall polysaccharides is further supported bythe analysis of the cell walls of 2 barley varieties.
ROOTS
T
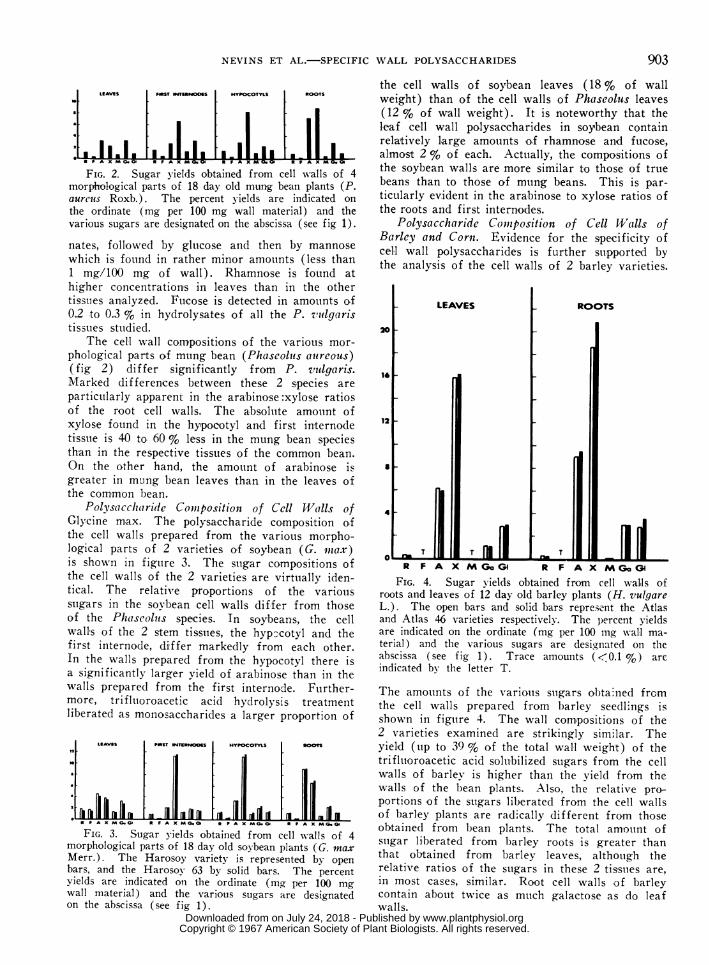
R F A X M GoGl R F A X MGoGFIG. 4. Sugar yields obtained from cell walls of
roots and leaves of 12 day old barley plants (H. vulgareL.). The open bars and solid bars represent the Atlasand Atlas 46 varieties respectively. The percent yieldsare indicated on the ordinate (mg per 100 mg wall ma-terial) and the various sugars are designiated on theabscissa (see fig 1). Trace amounts (<0.1 %) areindicated bv the letter T.
The amoutnts of the variotus suigars obtained frcmthe cell walls prepared from barley seedlings isshown in figuire 4. The wall compositions of the2 varieties examined are strikingly similar. Theyield (tip to 39 % of the total wall weighlt) of thetrifluoroacetic acid soluibilized sugars from the cellwalls of barley is higher than the yield from thewalls of the bean plants. Also, the relative pro-portions of the suigars liberated from the cell wallsof harley plants are radically different from thoseobtainedl from bean plants. The total amouint ofsuigar liberated from barley roots is greater thanthat obtained from barley leaves, althotigh therelative ratios of the suigars in these 2 tissuies are,in most cases, similar. Root cell walls of barleycontain about twice as mtich galactose as do leafwalls.
L.__wl
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
904
I II .I *1 I ElI IaL ...... U A...I.ME..BA . I.~ B -I--
PLANT PHYSIOLOGY
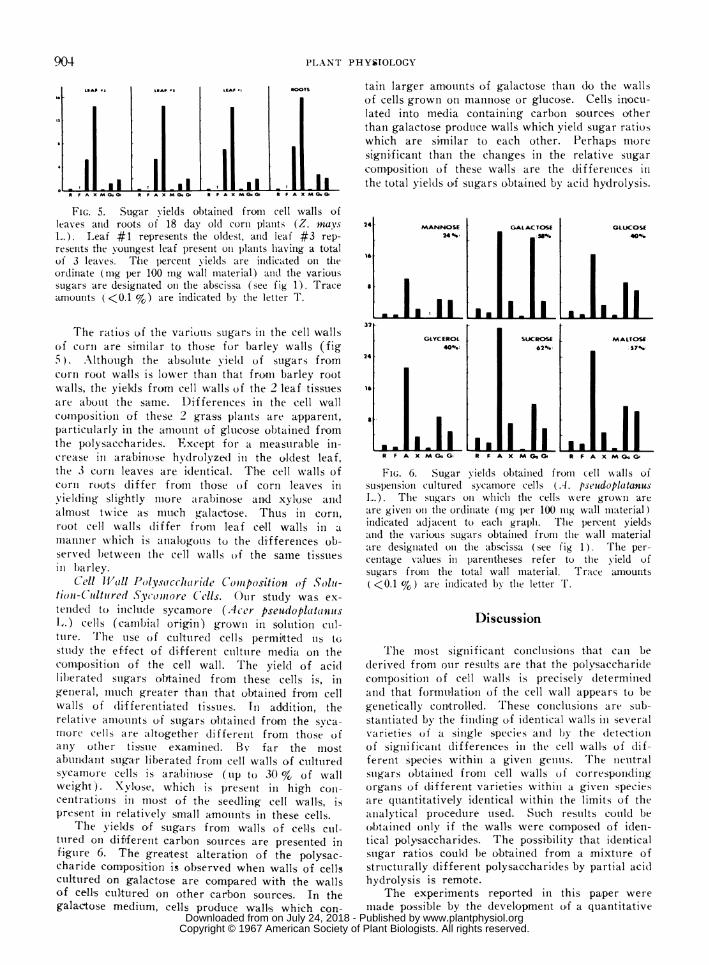
oTS tain larger amoumnts o.f galactose than do the wallsof cells grown on mannose or glucose. Cells inocu-lated into media containing carbon sources otherthan galactose produtce walls which yield sugar ratioswhich are similar to each other. Perhaps mi-oresigni,ficant than the changes in the relative sugarcompositi,on of these walls are the differetnces inthe total yields oIf sugars o,btained by acid hydrolysis.
FIG. 5. Sugar yields obtained froim cell walls ofleaves aIId roots of 18 day old corn plants (Z. nwNvsL.). ILeaf #1 represents thle oldest, anid leaf #3 rep-resenits the youngest leaf present oni plants having a totalof 3 leaves. The percent yields are indicated oIn theordinate (mg per 100 mg wall material) andt the varioussugars are designated oIn the abscissa (see fig 1). Traceamounts (<0.1 %) are indicated by the letter T.
The ratios of the various sugars in the cell wallsof corin are sinilar to those for barley walls (fig5). Although the absolute yield of sugars fromcorin root walls is lower than that from barley rootwvalls, the yields from cell walls of the 2 leaf tissuesare about the sa,me. Differences in the cell wallcomn,position of these 2 grass plants are apparent,particularly in the a,mount of glu,cose obtained fromthe poly,saccharides. Except for a measurable in-crease in arabinose hydrolyzed in the oldest leaf,the 3 corn< leaves are identical. The cell wvalls ofcorim roo,ts differ from those of corn leaves invieldiing slightly more arabinose and xylose and
almost twice as much galaclt,ose. Thus in corn,
root cell wialls (liffer from leaf cell walls in a
maiinner which is analogotus to the differences ob-served between the cell walls of the same tissuesin blarley.
C'ell latll Polyso cc/h(rri(le Coilipositioit of Solit-tiomi-Cuiltiired Sycomiore Cells. Ouir study was ex-
tended to include sycamore (Acer pseudopltittnutsL.) cells (camrbial origin) grown in solution cuil-ture. The use of cultured cells permi'tted uis tGstuidy the effect of different culltuire media on thec0oml.positionl of the cell wall. The yield of acidlib)erat,ed suigars obtainied from thelse cells is, ingenieral, munch greater thani that obtained from cellwalls of (lifferentiated tissues. In adidition, therelative anmounts of sugar-s obtained from the syca-more cells are altogether differenlt from those of
aniy other tissuie examined. By far the most
abunidaint sugar libterated froni cell walls of cultuiredsyvamore cells is arabinose (up to 30 % of wallweight). Xylose, which is preseint in high con-
ceIntrationls in most of the seedling cell walls, ispresenit in relatively small amoulni'ts in these cells.
The yields of sugars fro-m walls of cellis cul-tuired on dififerenit carbon sources are presented infiguire 6. The greatest alteration of the polysac-charide composition is observed when walls of cell'scultutred on galactose are compared with the wallsojf cells cultured on other carobon source,s. In thegalactose medium, celfls produce walls which con-
GLUCOSE40s
IGLYCEROL SUCROSE MALTOSE40%. 62% 57%
24
10
R F A X MG G, FAX M Ge R F A X MG. G,
FIG. 6. Sugar yields obtained from cell Nvalls ofsuspeensioni culture(d sycamore cells (A. pseudopla(tanusL.). The suigars oii whlichl the cells were grosswIi areare giveII on1 the ordiniate (tilg per 100 nug wall material)indicated adjacenit to eachi graph. The percenit yieldsatnd the various sugars obtainied from the wall materialare designiate(d oni thle abscissa (see fig 1). lThe per-centage values in l)arenltheses refer to the yield ofsugars from the total wall material. Tra--ce amounts<0.1 %) are inidicated by the letter T.
Discussion
The mIost sigiiificanlt conclus-ions that caIn bederived from our restults are that the polysaccharidecomposition of cell walls is precisely determinledandl that formtu-lation of the cell wall appears to begeinetically con,trolled. These conclusionis are sub-stanitiated by the finiding of identical w%,all,s in sev,eralvarieties of a sinigle species aind b)y the detectionof significant differen,ces in the cell wall,s of dif-ferent species within a giveni genuts. The ineutralsuigars obtained fromn. cell walls of correspondciingorganis of different varieties withini a given speciesare quantitatively identical within the limits of theanialytical procesdure used. Such results coutld beobtained only if the walls were com,posed of iden-tical polysaccharides. The possibility that ideniticalsuigar ratios could be obtained from a mixture ofstructurally different polysaccharides by partial aci(dhydrolysis is remote.
The experiments reported in this paiper wereiade possible by the development of a quantitative
c1* X C R F A xIX
IX
IX
LEAP '-a LEAF ' X LEAF *,
R F A [M G. Gd R F A : M 43i Cl R F A
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

905NEVINS ET AL.-SPECIFIC WALL POLYSACCHARIDES
gas chromatographic method for the analysis ofplant cell wall polysaccharides. A controlled tri-fluoroacetic acid hydrolysi,s of the cell walls wasemployed which liberates, as neutral sugars, 11 to60 % of the total cell wall weight. The amountliberated and the relative proportions of the 7neutral suigars found in all plant cell walls areconstant and reproducible for walls prepared froma given sour;ce. Plants grown in different experi-ments for the same length of tiime have the samewall composition. The yields of the various sugarsobtained from cell walls uising thiis procedure arein good agreement with the yields of sugars fronmcell wall preparations reported by other workers(18).
In the present studies we have made no attemptto quantitatively determine the uronic acid con-stituents of the cell wall. However, the mode oflinkage of these constituents in the cell walls thatwe have examined will have an effect on the yieldsof stugars, since the acid hydrolysis procedure em-ployed will not hydrolyze aldobiouronic acids. Thatsuch linkages exist is supported by the observationthat treatment of the cell walls with polysaccharidedegrading enzymes frequiently results, upon acidhydrolysis, in increased yields of some of the neu-tral sugars (15). One explanation for thi,s is thatthese enzymes are cleaving aldobiouronic acidlinkages, thereby permitting the detection of theneutral stugar portions of such linkages utpon sub-sequent acid hydrolysis. The acid hydrolysis pro-cedure employed also fails to hydrolyze the glvco-sidic linkages of cellulose and, probably, those ofsuch other acid resistant polymers as 8 1,4-glu-comannan s.
O0ur studies were facilitated by the use of fuillyexpanded morphological parts which were obtainedtfrom young plants and which had no secondarygrowth. \Ve avoid.etd using the distal 5 mm ofroot tips anid took considerable care to harvest theplants at a time when the cell wall composition wasrelatively stable. Prior to this time, the composi-tion of the wall changes radically and rapidly.
The various morphological parts used in thisstudy are made up of all the principal tissue tvpes,i.e., dermal, ground and vrascular, each one ofwhich is, in turn, composed of many specializedcell types. Hence, the wall composition obtainedrepresents an average of the composition contrib-uted by each of these cell types. In addiltion, thesevarious morphological parts contain both primaryand secoondary walls. In order to explaini the com-positional uniformity observed in our stuldies, eitherthe ratio of the variouis cell types within all plantparts mu,st be exceedingly pre-cise and each cellwithin this tissue must have a specific wall com-poisition, or alternatively, all cells within the mor-phological part must have the same wall composition.There is some evidence to support the formeralternative, as it has been shown that the sugarcomposition of cell walls of undifferentiated callus
explants changes upon differentiation into special-ized cell types (11). When the tissue explant isnot induced to differentiate, the composition of thecell wall does not change even upon aging. Wehave observed that walls of sycamore cells grownin ctulture which do not differentiate maintain aconstant composition throughout a 16 day growthcycle.
The nature of the medium in which suspension-cultured sycamore cells are grown markedly effectsthe wall polysaccharide composition. Except foran increase in the yield of galactose from walls ofcells grown on a galactose medium, the major effectresuiltiing from growing cells on di,fferent carbonsources is the variation in the quantity of the wallmaterial hydrolyzed to monosaccharides understandard conditions. The total yield ranges from24 % of walls produced in a mannose medium to62 % of walls prodcuced in a sucrose medium. Itappears that cells control the ratios of these poly-saccharides far more closely than they do theabsolute amounts of these polysaccharides whichare incorporated into the wall. Since the composi-tion of walls is essentially invari,ant when they areprepared from a particular intact plant, it is reason-able to assume that in vivo cells synthesize wallsfrom a rather specific mixture of metabolites. Themedium could effect the composition of the cellwalls by such mechanisms as catabolite repression.Metabolic oontrol of this sort would overrule pri-mary genetic control.
Cox and Gesner (4) have demonstrated thatthe presence of various sugars in the media ofseveral mammalian cell lines in tissue culture canalter the pattern of growth and cell morphology.In particular, these workers have found that thepresence of specific sugars in the medium inter-feres with normal clumping of the cells, and havesuiggested "that the specificity of the effects is dtueto binding of the stugars with complementary siteson the cell suirface leadiilg to suibsequent morpho-logic and metabolic changes." Cox and Gesnerconsider uinlikely the possibility that the moleculesof which the complementary sites are com-posed(heteropolysaccharides) have been modified. Theresults reported here suggest that the sugars in themedium can alter the polysaccharides of the cellsurface. If this were the case, it could directlyeffect a cell's ability to participate in surface inter-actions.
The Phascoluts vitlgaris varieties, the Glvcinemar varieties, and the Hordeum vulgtare varietiesare all differentially resistant to attack by certainpathogens. And yet, the cell wall polysaccharidecompositions within each of these varietal grouipsare essentially identical. From this it would ap-pear that the composition of the cell wall has arather minor role in the expression of differentialdisease resistance within these varieties. Diseaseresistance in plants, however, is often an age re-lated phenomenon. Differential resistance of the
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
Pliaseolus varieties to attack by Colletotrichitumilindemuiiithianumzl (Sacc. and Magn.) Brl. and Cav.is expressed dturing the first 2 weeks of the beanplant's growth (8). Thtis, if resistance is a ftinc-tion of the polysaccharide compositi,on of the cellwalls, then this muist be studied dutring that periodof development when major alterations in the com-position of the cell walls are taking place. Indeed,a resistant plant may be one wvhich reaches thestage of matuire cell walls more quickly than doesthe suscepti,ble plant.
Undoubtedly, allosteric controls and the likehelp to reguilate the synthesis of polysaccharides.This is already known to be the case in the syn-thesis of stich polysaccharidles as starch in leafchloroplasts (7) and glycoproteins in liver (13).Buit it is diffictult to see how stuch control mecha-nisms can regtllate the amotunt of polysaccharidesf,ound in plant cell walls. How, do the elnzymes inthe cytoplasm recognize when a stufficient amountof a particuilar polysaccharide has been synthesize(din the wall? This represents a complex problemwhich, when solved, may well aidd to ouir basicunderstanding of metabolic control an(d lifferen-tiatioll.
Acknowledgments
Thie authors are grateful to Dr. Jack Paxton of theDepartment of Plant Pathology at the University ofIllinois for supplying the soybean seeds, to Dr. CharlesSchaller of the Department of Agronoomy, Universityof California at Davis for supplying the barlev seedsand to Thomas AI. Jones for assistance in the prepara-tion of this manuscript.
Literature Cited
1. ALBERSHEIM, P. 1965. The substruicture anidfunlction of the cell AN-all. In: Plant Biochemistry.J. Boinner alnd J. E. Varner, eds. Academic Press,Inic. Nexv York. 151-6.
2. ALTIERSHEI-M, P. 1965. Bio,genesis of the cell wall.In: Planit Biochemistry. J. Boniner anid J. E. Var-ner, eds. Acadeemic Press, Inc. New York. 298-321.
3. BFECKER, G. E., P. A. Hu-I, AND P. ALBERSHEIM.1964. S! ntthesis of extracelluilar pol]saccharide bysuspenisions of Acer- pscudoplat[() nui; cells. PlaiItPhvsiol. 39: 913-20.
4. Cox, R. P. AND B. NI. GESNER. 1965. Effect ofsiiiple sutgars on the morphology aAcl growth pat-terni of miammalian cell cuiltuires. Proc. NatI. Acad.Sci. U. S. 54: 1571-79.
5. ESAU, K., V. I. CHEAMIL, AND R. H1. GILL. 1966.C\ to'ogN- of differentiating trachearv elements. I.Organelles anid memiibranie systems. Anm. J. Bot-an'V 53: 756-64.
6. GAILLARD, B. I). E. 1965. Colmpar-isoin of theheiicellulloses from plants belongilln to txx-o di f-ferent plant families. Phy-tochenm. 4: 631-34.
7. GHOSH, H. P. AND J. PREISS. 1966. Adenosinediphosphate glucose pyrophosphorylase. A regula-tory enzyme in the biosynthesis of starch in spinachleaf chloroplasts. J. Biol. Chem. 241: 4491-504.
8. GRIFFEY, R. T. AND J. G. LEACH. 1965. The in-fluence of age of tissue on the development ofbean anthracnose lesions. Phytopathology 55:915-18.
9. HEPLER, P. K. AND E. H. NEWN'COMB. 1963. Thefine structure of young tracheary xylem elementsarising by redifferentiation of parenchyma inwounded Coleus stem. J. Exptl. Botany 14: 496-503.
10. HOAGLAND, D. R. AND D. I. ARNON (revised byD. I. Arnon). 1950. The water-culture methodfor groxN-ing plants wxithout soil. College of Ag-riculture, University of California, Berkeley, Cali-fornia. Cir. 347. 32 p.
11. JEFFS, R. A. AND D. H. NORTHCOTE. 1966. Ex-perimental induction of vascular tissue in an un-differentiated plant callus. Biochem. J. 101: 146-52.
12. KIVILAAN, A, T. C. BEAMAN, AND R. S. BAN-DURSKI. 1959. A partial chemical characteriza-tion of Maize coleoptile cell Nx-alls prepared withthe aid of a continually reinewable filter. Nature184: B.A. 81-82.
13. KORNFELD, S., R. KORNFELD, E. F. NEUFELD, ANDP. J. O'BRIEN. 1964. The feedback control ofsugar nucleotide biosynthesis in liver. Proc. Natl.Acad. Sci. U. S. 52: 371-79.
14. LAMPORT, D. T. A. 1960. Hydroxyproline in pri-mary cell x-xalls of higher plants. Nature 188:665-66.
15. McNAB, J. M., D. J. NEVINS, AND P. ALBERSHEIM.1967. Differential resistance of cell xvalls of .Acerpseudoplatanus, 7Triticum vulgare, Hordcumn vul-guarc, and Zca wnays to polysaccharide degradingenzymes. Accepted for publicationi in Phvtopath-ology.
16. NEWCOMB, E. H. 1963. Cytoplasm-cell wall rela-tionships. Ann. Rev. Plant Physiol. 14: 43-64.
17. PHETHEAN, P. D. AND AI. HALLA\VAY. 1965. Thledetection of ribonucleic acid in ptlant cell w-alls.Biochem. J. 96: 9P
18. RAY, P. AI. 1963. Sugar compositioni of oat-coleoptile cell w alls. BiochemI. J. 89: 144-50
19. RAY, P. MI. AND D. B. BAKER. 1965. The effectof auxin Onl synthesis of oat coleoptile cell wallconstituenits. Plant Phy siol. 40: 353-60.
20. THORNBER, J. P. AND D. H. NORIHCOTE. 1961.Clhanges in the chemical composition of a camiibialcell during its differentiationi into x lenm anidphloem tissue in trees. Biocheimi. j. 81: 455-64.
21. TORREY, J. G. AND Y. SHIGEIMURA. 1957. Growtlhand controlled morphogenesis in pea root callustisstie gro\N-ii in liqluid media. Am T.. Botany 44:334-44.
22. WOODING, F. B. P. AND D. H. NoRTrICOTE. 1965.Tlhe fine structure and developmient of the com-panion cell of the phloenx of Accr peidoplautunus.J. Cell Biol. 24: 117-28.
906
www.plantphysiol.orgon July 24, 2018 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.





























