Embed Size (px)
Citation preview

JOURNAL OF TUEWORLD AQUACULTURE SOCmTYTHE WORLD AQUACULTURE SOCIETY
The World Aquaculturc Society is an international organization witb over 2500 members in 80 countries. TheSociety promotes tbe educational, scienûfic and technological dcvelopment of aquacuJturc throughout tbe world,gatbers and disseminates aquaculture information and holds meetings to foster aquaculturc development andinformation exchange. Membership information is available fiom the World Aquaculture Society, MembershipDepartment, 143 1. M. Parker Coliseum, Louisiana State University, Baton Rouge, LA 70803, United States ofAmerica.
Vol. 35, No. 2June, 2004
Spawning Characteristics of the South African MudcrabScylla serrata (Forskai) in Captivity
JEROME A. DAVISSOCIETY OFFICERS-2003-2004President: Kcvin Fitzsimmons Vice-President: John Hargreaves
Thcson, Arizona Baton Rouge, LouisianaUSA USA
President Elect: Daniel Fegan Secretary: Julie DelabbioBangkok Bluefield, West VuginiaThailand USA
Past-President: Geoff Allan Treasurer: Jay ParsonsTaylor's Beach, NSW St. John's, NewfoundlandAustralia Canada
Alejandro Flores-TomLa paz, B.C.S.
Mexico
Michael MasserBryan, Texas
USA
DIRECTORSYnguar OIsen
TrondheimNorway
Gary JensenWashington, DC
USA
lAborarory of Aquaculture anti Artemia Reference Center, Ghent University,Roz/er 44, Ghent, 9000, Belgium anti
Department of Ichthyology anti Fisheries Science, Rhodes University, Box 94,Grahamstown, 6140, South Africa
Gn.ES J. GlillRCHILLANI>THOMASHEcHT
Department of Ichthyology anti Fisheries Science, Rhodes University, Box 94,Grahomstown, 6140. South Africa
PATRICK SOROELOOSSungchul Bai
Nam-GuKorea
Lorenzo JuarezLaredo, Texas
USA
lAboratory of Aquaculture anti Artemia Reference Center, Ghent University,Roz/er 44, Ghent, 9000. Belgium
Abstract
Scylla serrata Is a. potentIal aquaculture specles In Soutbern Africa. Wonnation aOOut ltsreproductive blology Is requlred as a prerequIsIte to establIshing hatchery technology, Adultremale S. serraJ4 were caupt In tbe Umlalazl estuary on tbe subtroplcal east coast or SouthAfrlca and kept In captivity to observe and record spawnlng characterlstics. Data collectedInduded crab size and mass, time In captivity prior to spawnlng, recondlty per batch, relativerecundity, IndividuaI egg mass and slze, size or zoea 1 larvae, Incubation time, and hatchsuccess rale. or tbe 119 crabs kept In captivity 83% spawned In tbe maturation system-most wltbln 40 d or capture. Tbe crabs were hlghly recond (mean relative recondlty per batch= 10,655:: 4,069 eggslg remale) and tbe majorlty or tbe batches hatched witbln 288 h (12 d)at 27 C. Spawnlng In captivity occurred throughout tbe year, wltb a peak in late wlnter/earlyspring. Tbls dlfCers silghtly Crom records or ovarlan maturlty stages or the crabs In tbe wild.A pattern or syncbrooous spawnlng was recorded where tbe remales were observed to extrudetbelr eggs In groups, commonly wlthin 3 d or one anotber, separated by long perlods orlnactivity, suggesting an exogenous spawnlnll cue. A totaI or 1,374,>188zoea larvae were ob-taIned per kg or remale per montb. Thls means that ICsuOident mature remales can be caughtCrom tbe wild, tbese could be used ror stocklng hatchery operadons. Tbe crabs wen easy tomalntaIn, mature, and spawn In captlvity. ThIs wIIl racllltate ruture domestication wblch wllIeventually reduce tbe Deed ror wild caught broodstock. Tbe spawnlng characterlstics or Soutb~can S. serrata fit 10 weil wltb tbose observed ror tbe genus throughout lts dIstrIbution
"ïmplylng that ecologlcal and flsherles management could be simIlar.
CHAPTER REPRESENTATIVESLatin Amerlca Soutbeast AslaOsircs De Melo Michael Rimmer
Reeife Caims, QueenslandBrazil Australia
Unlted States Aquaculture SocietyLaDoo Swann
Ocean Springs, MississippiUSA
JapanShuosuke Koshio
KagoshimaJapan
EDITORIAL BOARD
HarT)' Daniels, Managing EditorPat Edwards, Copy Editor
Kayde Brownlee, Editorial AssistantPoem Gardiner, Editorial Assistant
Sungchul BaiBruce BartonPaul BowserDominique BureauBarT)' Costa-PiercePeter CrocosL. Elizabctb Cruz-SuárczBil! DanielsD. Allen Davis
Associate EditorsAriel Diamant Izhar KhanCarole Engle Shuosuke KoshioDelbcrt Gatlin Don LightnerMayra González-Félix Chhorn LimXirning Guo Junda LinJohn Hargreaves C. Greg LutzReginal Harrell Susan McBrideThomas Hecht Nigel PrestonKatbleen Hartman Steven Rawles
Wendy SealeyMuki ShpigelSandra ShumwayJames TidwellCharles MischkeWagner ValentiKatsuhiko WadaWade WatanabcL. Curry Woods, III
Scylla serrata is a iarge, commercialiyimportant, brackish water, carnivorous crab,distributed throughout the Indo-Pacific re-gion (Brock 1960; Tamura 1970; HilI 1975;Heasman and Fielder 1977). Tbe species isa candidate for aquaculture in Southem Af-rica, but a major bottleneck to the estab-lishment of mudcrab farming worldwide isthe dependence on wild caught juveniles(Williams and Primavera 2001). Hatcherytêéhniques thus need to be developed forthe species in the region. An essential com-
The Journalof the WorldAquaculture Society (ISSN 0893-8849) is published quarterly by tbe World Aqua-culture Society, 143 J. M. Parker Coliseum, Louisiana State University, Baton Rouge, LA 70803. Librarysubscriptions are SI05 (U.S.) per volume in tbe U.S., SliS in Canada, South America and Mexico, SI35 in allotber countries. Individual subscriptions are a benefit of membcrship in tbe World Aquaculture Society. Annualmembership dues are S40 for students, S60 for individuaIs, $250 for corporations and $100 for sustainingindividuals or companies. 1Wenty-five percent of dues is designated for a subscription te tbe Journal ofthe WorldAquaculture Society. Periodicals pastage paid at Baton Rouge, LA and additional mailing offices. POST-MASTER: Send addrcss changes te tbe World Aquaculturc Society, 143 J. M. Parker Coliseum, Louisiana StateUniversity, Baton Rouge, LA 70803.
Articles in tbe Journalof the WorldAquaculture Society are indexed or abstracted in: Aquatic Sciences andFisheries Abstracts; ISI SciSearch; ISI Research Alert; ISI Currcnt ContentstAgriculture, Biology & Environ-mental Sciences; Fisheries Review; and Wildlife Review.
ponent of hatchery production is the estab-lishment of a broodstock that can producea predictabie supply of fertilized eggs(Wickins and Lee 2002). A thorough un-derstanding of the reproductive biology ofthe species is thus required. Despite a re-newed research effort worldwide (Wickinsand Lee 2002), driven in part by dwindlingnatura! fisheries for this species (Le Vay2001), little has been published in the pri-mary literature about the spawning charac-teristics of S. serrata. In addition, much of
C Copyriaht by me Wood AquacultuR: Society 2004

122
1
!
1
tbe work tbat was published before Keenanet al.'s (1998) revision of tbe genus leavessome doubt as to tbe precise species fromwhich data were collected.
Soutb African mudcrabs are known tomate and undergo ovarian maturation in es-tuaries (Hill 1975, 1994; Robertson andKruger 1994). Tbey then migrate offshoreto spawn on tbe continental shelf. Fertilizedeggs attach to the pleopods where tbey areincubated. Hatched larvae are released intotbe ocean (HilI 1975; Heasman and Fielder1983; Hill1994; Le Vay 2001). Berried fe-males are caught sporadically in offshoretrawl nets and are extremely rare in estu-aries. Observations on spawning behaviorand data of egg production and hatching aretberefore scant. Tbe only information on re-production of S. serrata in South Africa areseasonal ovarian maturation indices ofcrabs in tbe wild and some anecdotal dataon spawning in captivity (Robertson andKruger 1994).
Tbe aim of tbe research was to gatberinformation about tbe spawning character-istics of wild-caught S. serrata in captivity.This information will be used as a basis forhusbandry practice and to establish hatch-ery techniques tbat will contribute to tbeeventual domestication of tbe species. Tbeinformation can also be used as a compo-nent for conserving tbe wild population ormanaging tbe fishery (Caddy 1989; Koslow1992). In order to achieve this, mature fe-males were caught from tbe wild and main-tained in our research facility on tbe sub-tropical east coast of Soutb Africa (RhodesUniversity Mudcrab Hatchery at MtunziniPrawn Farm).
Materials aod Methods
Broodstock Capture
Mature female crabs were caught fromFebruary 2000 to August 2001 using hoop-and-bag traps in the Umlalazi estuary(31°47'E, 28°57'S) on tbe subtropical eastcoast of Soutb Africa. Tbe traps were notsize selective, and mature males and im-
DA VIS ET AL.SPAWNINO OP MUD CRAB 123
mature females were retumed to tbe waterimmediately. As tbe Umlalazi estuary is aproelaimed estuarine sanctuary we werealso required to return tbe mature femalesunharmed to this estuary on completion ofour observations. Water temperatures in tbeestuaries ranged from 17 to 22 C in winterand from 22 to 30 C in summer. Femaleswere identified as being sexually mature bytheir wide, dark, U-shaped abdomensfringed witb setae (Wamer 1977; Heasman1980; Bames 1987; Robertson and Kruger1994). Fishing was conducted only duringdaylight hours. Traps were checked at 15-min intervals and tbe crabs were removedand transported to our facility in a 200-Lplastic bin covered with a wet hessian sack.Catch per unit effort (CPUE) was calculat-ed for mature females as: (nc/t)/nt where:nc = number of crabs caught; t = soak timein hours; nt = number of traps, and ex-pressed as crabs per trap hour (Robertson1989).
Tbe animals were disinfected againstbacteria and fungi by bathing tbem over-night in an aerated seawater batb containingl00-ppm malachite green and 100-ppm for-malin (Castille and Lawrence 1989; Ha-masaki and Haitai 1993). Size was recorded(carapace widtb in mm and weight ingrams), and each crab was numbered by en-graving tbe carapace using a jewelIer's en-graver.
roof tiles raised on plastic pipes wereplaced on tbe bottom of tbe tank to reducecannibalism and stress. Water was recircu-lated through a 600-L biological trielde fil-ter at a rate of 1Usec. Prior to reaching tbetrickling filter tbe water was passed througha nylon stocking to collect particu1atewaste. The stocking was cleaned daily andreplaced once a montb. Approximately 1 m3of tbe water was replaced daily witb freshseawater at a salinity of 30-32 ppt: pumpedfrom a beach weIl point and filtered to 1fJom.Light was subdued (1-3 Ix, LutronLX-105light meter, Taiwan) by painting tbefluorescent lamp covers green and photo-period was maintained at a constant 14 L:10 d cycle. Tbe system was indoors buttemperature was not controlled, and variedfrom 19 C in winter to 25 C in summer.Crabs were fed to satiation on a feedingtray, twice'per day with squid (Loligo sp.),shrimp (Fenneropenaeus indicus), musseis(Perna perna), or fish roe (assorted species)on successive days. Uneaten food was re-moved dai1y and tbe tank bottom was vac-uumed once a week.
Broodstock Conditioning
Tbe crabs were maintained in a recircu-lating system at a density not exceeding 1.5animals/m2. No male crabs were introducedinto the spawning system, as the femaleswere presumed to have mated in tbe wildand tbus were carrying spermatophores(Robertson and Kruger 1994). Tbe 12-m3maturation tank (4 X 3 m) had a concretebottom that was painted black, and waterdeptb was maintained at 1 m. Nine roundplastic bins (40 cm in diameter, 15 cm inheight) filled witb beach sand were placedon tbe bottom of the tank to facilitatespawning. Twe1ve shelters in tbe form of
Spawning, Incubatidn, and Hatching
Crabs tbat spawned (extruded eggs) wereeasily detected by shining a torch into tbetank and turning over tbe artificial shelters.Berried females were netted out of tbe mat-uration tank as soon as tbey were observedand placed into 60-L glass observationaquaria provided witb an extemal 15-L air-lift biological sand filter. Tbe aquaria werefilled witb seawater from tbe main recircu-lating system, and once tbe crabs had beenintroduced, tbe water was heated with a150-W submerged aquarium heater to 27 Cover 24 h. Tbe crabs were not fed after ex-truding tbeir eggs, but tbe aquaria ~ere si-phoned daily to remove feces and droppedeggs. Dropped eggs were examined undera micr6scope and were commonly found tobe infected witb fungi, polychaet, or nem-atode worms. Females were then treated
)
With 1-ppm and 10-ppm formalin for 24 h.If after treatment tbe female continued to
drop large quantities of eggs, she was con-sidered unsuitable for further work and im-mediate1y returned to tbe estuary of origin.
Once a berried crab was judged to be asuitable hatchery candidate, it was movedto a covered 600-L fiberglass, cylindro-con-ical incubation tank. A window was provid-ed in tbe side of tbe tank for observing tbefemale. Tbe tank was filled with filtered (1fJom),UV sterilized seawater. Tbe water wastecirculated through an 80-L biological air-lift filter and tben passed through a UV ster-ilizer at a rate of 1.7 Umin. Water wasdrained from tbe tank into tbe filter througha 0.3-mm outlet screen to prevent loss ofhatching zoea. A sheet of perforated PVCp1aced at tbe bottom of the cylindrical por-tion of tbe tank pro'vided tbe erabs witb aflat substraturn tbat allowed dropped eggsand fèces to sink into tbe conieal tank bot-tom from where tbey could be removed byopening a valve.
Wild-caught crabs were added to tbemain maturation tank to maintain tbe stock-ing density after crabs tbat had spawnedhad been removed. Once hatching had tak-en place, tbe crabs were returned to tbeirestuary of origin.
Measure'!'ent of Parameters
Fecundity in tbe text refers to tbe num-bers of eggs extruded per female per batchand was estimated as follows: berried crabswere weighed shortly after tbe extrusion ofeggs and reweighed after the larvae hadhatched. This gave tbe total mass of tbeeggs. Four crabs starved for the 12-d in-cubation lost 1.14 :!: 0.80% of tbeir bodyweight. This was considered negligible andwas tberefore not included in further cal-culations. Total egg mass was calculated as:
Total egg mass = mass of crab at extrusion- mass.of crab after hatch.
Subsamples of at least 300 eggs were re-moved from tbe egg bolus as soon as pos-sib1eafter.extrusion using sterilized forceps,weighed, and counted in three replicates un-der a dissecting microscope. This provided
124
an .estimate of individual egg mass, where-upon fecundity was estimated by dividingtotal egg mass by individual egg mass. Tovalidate this method, the total number ofeggs produced by tWo females was countedand compared to these estimates.
Egg diameter and larval size (distancebetWeen the lateral spine tips) were mea-sured under a compound microscope usinga graduated eyepiece at 40 times magnifi-cation.
Hatching success was calculated as fol-lows: after hatching in the 600-L tank, thelarvae were mixed weIl by vigorous aera-tion. Nine 50-roL samples were scooped outof the tank at different depths. The larvaein each sample were counted, and an aver-age was calculated as the density of larvaeper 50 roL, allowing for the estimation ofthe number of larvae in the 600-L tank.Hatching success (%) was calculated by:
(Number of larvae hatchedINumber of eggsextruded) X 100.
Statistics
A regression model was fitted to the datato determine the relationship betWeen vari-ables using SPSS 11.0.1 (2001, SPSS, Bel-gium) software. Data was then checked fornormality, linearity, and homo~.cedacity,and a residual plot was done. Pearson's co-efficient was used to exarnine correlationbetWeen variates. Linear, quadratic cubicand power curve estimates were used to de-termine best fit (Sokal and Rohlf 1995).Data are presented as mean :t standard de-viation.
Results
Female Broodstock Capture
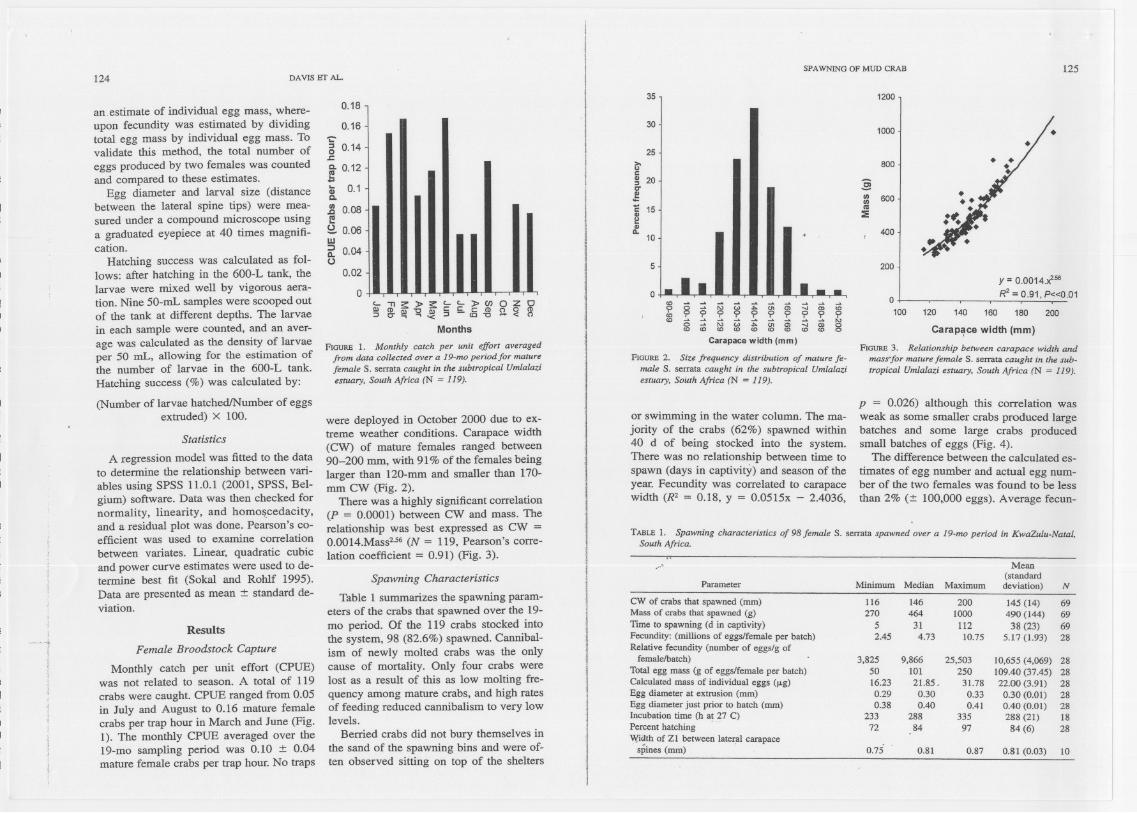
Monthly catch per unit effort (CPUE)was not related to season. A total of 119crabs were caught. CPUE ranged from 0.05in July and August to 0.16 mature femalecrabs per trap hour in March and June (Fig.1). The monthly CPUE averaged over the19-mo sampling period was 0.10 :t 0.04mature female crabs per trap hour. No traps
125
35
SPAWNING OF MUD CRAB
1200
DA VIS ET AL.
0.18
0.16 30
0;:-S 0.14~
g0.12:p 0.1Cl.
1.1 0.08I!2. 0.06w~ 0.04(J
25
~c~ 20CT
!i 15I!:.
10
50.02
o oc..."'I1s:~s:c...c...~cnozo!!! ~~-g~ § 6..E~ g.51 ~
Months
FIGURE 1. Monthly catch per unit effort averagedfrorn data collected over a 19-mo periodfor rnaturefemale S.serrata caught in the subtropical UrnlalaZ!esruary. South Africa (N = 119).
FtGURE2. Sile frequency distribution of rnature fe-male S. serrata caught in the subrropical Urnlalaliesruary. South Africa (N =119).
were deployed in October 2000 due to ex-treme weather conditions. Carapace width(CW) of mature females ranged between90-200 mm, with 91% of the females beinglarger than 120-mm and smaller than 170-mm CW (Fig. 2).
There was a highly significant correlation(P = 0.0001) between CW and mass. Therelationship was best expressed as CW =0.0014.Mass2.56(N = 119, Pearson's corre-lation coefficient = 0.91) (Fig. 3).
or swimming in the water column. The ma-jority of the crabs (62%) spawned within40 d of being stocked into the system.There was no relationship betWeen time tospawn (days in captivitY) and season of theyear. Fecundity was correlated to carapacewidth (R2 = 0.18,Y= 0.0515x- 2.4036,
1000
800
400
200
o100 120
y = 0.0014.x2-56
W = 0.91. P«0.01. . .140 160 160 200
Carap~ce wldth (mm)
FIGURE3. Relationship between carapace width andmass'for rnaturefemale S. serrata caught in the sub-tropical Urnlalazi estuary. South Africa (N = 119).
p = 0.026) although this correlation wasweak as some smaller crabs produced largebatches and some large crabs producedsmall batches of eggs (Fig. 4).
The difference betWeen the calculated es-timates of egg number and actual egg num-ber of the two females was found to be lessthan 2% (:t 100,000 eggs). Average fecun-
Spawning Characteristics
Table 1 summarizes the spawning param-eters of the crabs that spawned over the 19-mo period. Of the 119 crabs stocked intothe system, 98 (82.6%) spawned. Cannibal-ism of newly molted crabs was the onlycause of mortality. Only four crabs werelost as a result of this as 10w molting fre-quency among mature crabs, and high ratesof feeding reduced cannibalism to very lowlevels.
Berried crabs did not bury themselves inthe sand of the spawning bins and were of-ten observed sitting on top of the shelters
N C;; Ä en m'" ....'? '? '? '? '? '? '?N C;; Ä en m '"'" '" '" '" '" '" '" 8
Cara pace wldth (mml
TABLE1. Spawning characteristics of 98 fernale S. serrata spawned over a 19-mo period in KwaZulu-Natal.South Africa.
..'" Mean(standard
Parameter Minimum Median Maximum deviaûon) N
CW of crabs that spawned (mm) 116 146 200 145 (14) 69Mass of crabs that spawned (g) 270 464 1000 490 (144) 69Tlffie to spawning (d in captivity) 5 31 112 38 (23) 69Fecundity: (millions of eggslfemale per batch) 2.45 4.73 10.75 5.17 (1.93) 28Relative fecundity (number of eggslg of
femaleJbatch) 3,825 9,866 25,503 10.655 (4,069) 28Tota! egg mass (g of eggslfemale per batch) 50 101 250 109.40 (37.45) 28Calculated mass of individual eggs (",g) 16.23 21.85. 31.78 22.00 (3.91) 28Egg diameter at extrusion (mm) 0.29 0.30 0.33 0.30 (O.O\) 28Egg diameter just prior to hatch (mm) 0.38 0.40 0.41 0.40 (0.01) 28Incubaûon ûme (h a! 27 C) 233 288 335 288 (2\) 18Percent hatching 72 84 97 84 (6) 28W,ii;\thof ZI between lateraI carapace
spines (mm) . 0.75 0.81 0.87 0.81 (0.03) 10
SPAWNINO OF MUD CRAB
o110 1~ 1ro 1ro 100
Carapace wldth (mm)
FIGURE4. Relationsh/p between carapace wldth
(mm) and lecundity (number ol eggs per lemale perbatch) lor S. serrata (N = 60).
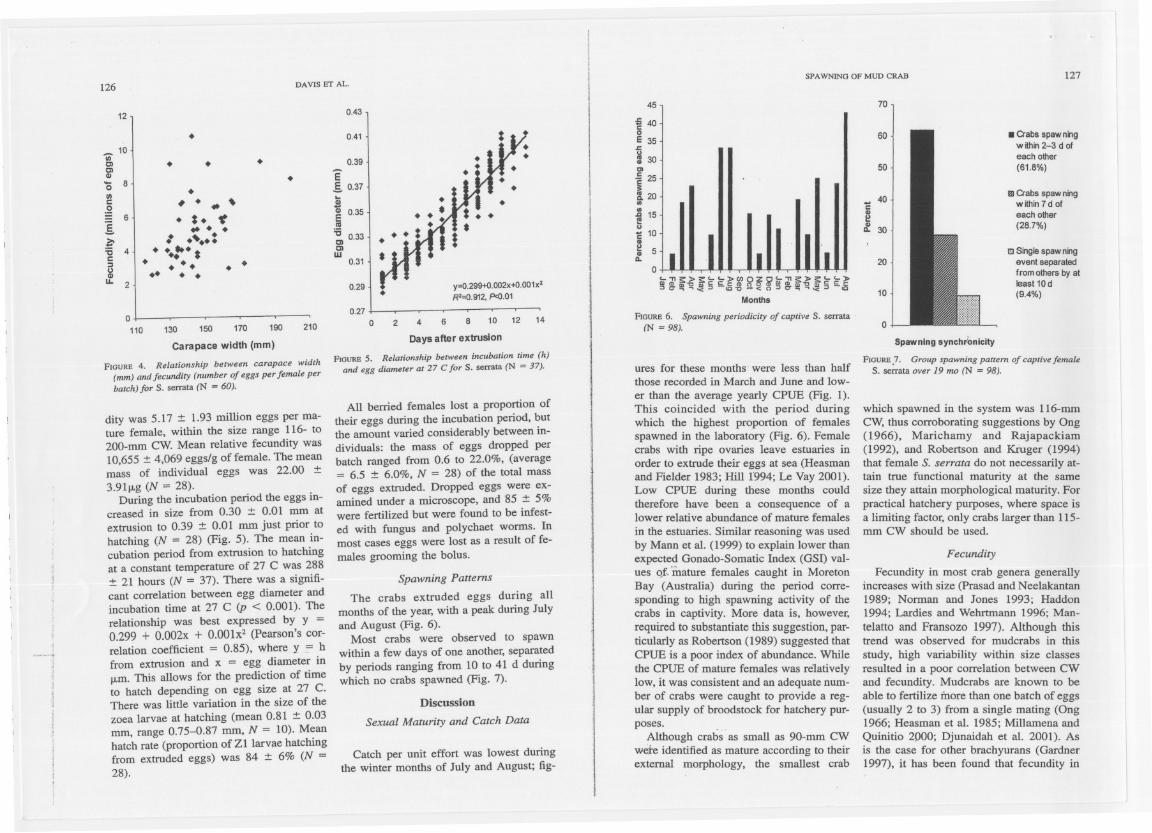
dity was5.17 :t 1.93million eggsper ma-ture female, within the size range 116- to2oo-mm CW. Mean relative fecundity was10,655 :t 4,069 eggs/g of female. The meanmass of individual eggs was 22.00 :t3.91jJ.g (N = 28).
During the incubation period the eggs in-creased in size from 0.30 :t 0.01 mm atextrusion to 0.39 :t 0.01 mm just prior tohatching (N = 28) (Fig. 5). The mean in-cubation period from extrusion to hatchingat a constant temperature of 27 C was 288:t 21 hours (N = 37). There was a signifi-cant cOITelationbetween egg diameter andincubation time at 27 C (p < 0.001). Therelationship was best expressed by y =0.299 + 0.002x + 0.001x2 (Pearson's cor-relation coefficient = 0.85), where y = hfrom extrusion and x = egg diameter in
jJ.m.This allows for the prediction of timeto hatch depending on egg size at 27 C.There was little variation in the size of thezoea larvae at hatching (mean 0.81 :t 0.03mm, range 0.75-0.87 mm, N = 10). Meanhatch rate (proportion of ZI larvae hatchingfrom extrudedeggs) was 84 :t 6% (N =28).
45
t40oE35ij=~al.5 25~[20
J ..· 11 15
B'!: 10
~ 5Cl11.
o~~~>~~~>moz~~~~>~~~>~~~~~§~~~~~~~~~~~§~á
Monllls
FIOURE 6. Spawn/ng period/city ol capt/ve S.SCl11Ita
(N = 98).
y=o.29~.OO2x+o.OO1x2R2=O.912, P<O.01
210
0.27o 2 4 6 8 10 12 14
Daysafterextruslon
FIOURE S. Reu.rionship between /ncubation time (h)and egg diameter at 27 C lor S. serrata (N =37). ures for these months were less than half
those recorded in March and June and low-er than the average yearly CPUE (Fig. 1).This coincided with the period duringwhich the highest proportion of fe.malesspawned in the laboratory (Fig. 6). Femalecrabs with ripe ovaries leave estuaries inorder to extrude their eggs at sea (Heasmanand Fielder 1983; Hill 1'994;Le Vay 2001).Lew CPUE during these months couldtherefore have been a consequence of alower relative abundance of mature femalesin the estuaries. Similar reasoning was usedby Mann et al. (1999) to explain lower thanexpected Gonado-Somatic Index (GSI) val-ues Qf.Ïnature females caught in MoretonBay (Australia) during the period COITe-sponding to high spawning activity of thecrabs in captivity. More data is, however,required to substantiate this suggestion, par-ticularly as Robertson (1989) suggested thatCPUE is a poor index of abundance. Whilethe CPUE of mature females was relativelylow, it was consistent and an adequate num-ber of crabs were caught to provide a reg-ular supply of broodstock for hatchery pur-poses. .
Although crabs as small as 90-mm CWwete identified as mature according to theirextemal morphology, the smallest crab
AU berried females lost a proportion oftheir eggs during the incubation period, butthe amount varied considerably between in-dividuals: the mass of eggs dropped perbatch ranged from 0.6 to 22.0%, (average= 6.5 :t 6.0%, N = 28) of the tota! massof eggs extruded. Dropped eggs were ex-amined under a microscope, and 85 :t 5%were fertilized but were found to be infest-ed with fungus and polychaet WOITDS.Inmost cases eggs were lost as a result of fe-males grooming the bolus.
Spawning Patterns
The crabs extruded eggs during a11months of the year, with a peak durlng Julyand August (Fig. 6).
Most crabs were observed to spawnwithin a few days of one another, separatedby periods ranging from 10 to 41 d duringwhich no crabs spawned (Fig. 7).
Discussion
Sexual Maturity and Catch Data
Catch per unit effort was lowest duringthe winter months of July and August; fig-
70
60
ro
o
127
. Crabsspawni1gw ithln 2-3 d ofeach other
(61.8%)
el Crabs spaw ningwithln 7 doleach other
(28.7%)
EI Single spaw ningevent separatedIromothers by atleast 10 d
(9.4%)
Spawnlng synchl'llnlclty
FIOURE.7. Group spawning pattem ol captive lemaleS. 5elTata over 19 mo (N =98).
which spawned in the system was 116-mmcw. thus cOIToboratingsuggestions by Ong(1966), Marichamy and Rajapackiam(1992), and Robertson and Kruger (1994)that female S. serrata do not necessarily at-tain true functional maturity at the samesize they attain morphological maturity. Forpractical hatchery purposes, where space isa limiting factor, only crabs larger than 115-mm CW should be used.
Fecundity
Fecundity in most crab genera generallyincreases with size (prasad and Neelakantan1989; NOITDanand Jones 1993; Haddon1994; Lardies and Wehrtmann 1996; Man-telatto and Fransozo 1997). Although thistrend was observed for mudcrabs in thisstudy, high variability within size classesresulted in a poor cOITelationbetween CWand fecundity. Mudcrabs are known to beable to fertilize more than one batch of eggs(usually 2 to 3) from a single mating (Ong1966; Heasman et al. 1985; Millamena andQuinitio 2pQO;Djunaidah et al. 2001). Asis thc case for other brachyurans (Gardner1997), it has been found that fecundity in
126 DAVISEr AL.
121
0.43
. 0.41
_ 10enCl . . . 0.39Cl11I Ê... 8 .0 ..s 0.37en .C . . . ..
i6 . *. ., 0.351 .
... . '6t ...: Cl 0.33
'6 4 . ...... ClC . . W:I . .. . . 0.31.. ..u.2 0.29
..40..ClI!l.
20
10

128 DA VIS ET AL.SPAWNlNG OP MUD CRAB 129
2001; Millamena and Bangcaya 2001;Quinitio et al. 2001).
in February, May, and October found byRobertson and Kruger (1994) for the samepopulation, or their suggestion that spawn-ing probably occurred from late spring toearly autumn. It should, however, be notedthat Robertson and Kruger (1994) recordedhigh variability in OSI values. This givessubstance to the suggestion by Heasman etal. (1985) that OSI values were not a goodindicator of spawning periodicity for mud-crab. Tbe true situation could, however,have been confused by the fact that the an-imals in this study were brought into arti-ficial conditions that may have providedcues different to those that the crabs mighthave experienced in the wild. Mudcrabs areknown to be able to maintain ovarian ma-turity for extended periods before spawning(Heasman et al. 1985). Tbe introduction ofthe females to captiye conditions may haveinduced them to spawn earlier than in thewild. In contrast to the studies by Mann etal. (1999) and Nghia et al. (2001), whofound a strong correlation between the timetaken to spawn in captivity and season, wemade no such observations. Only long-termstudies of spawning and recruitment willclear up the discrepancy of whether mud-crabs in South Africa have a distinct spawn-ing season.
TABU!2. Fecundity anti reproductive output of female Scylla species from different locations.
S. serrata usually decreases with each sub-sequent batch of eggs (Ong 1966). Sincefemales were used for only a single spawn-ing in this study, the smaller egg mass pro-duced by some of the larger females mayhave been the second or third batch from asingle molt and mating, while large batchesproduced by some of the smaller femalesmay have been a fust spawning. This mayexplain the poor correlation between crabsize and batch size. Table 2 lists observedfecundity data of Scylla species. Tbe num-ber of eggs per batch and relative fecundityof South African crabs was high comparedto the range recorded in Australasia (BinJamari 1992; Marichamy and Rajapackiam1992; Jyamanna and Jinadasa 1993; Dat1999; Mann et al. 1999; Srinivasagam et al.2000; Djunaidah et al. 2001; Hai et al.
Seasonality of Spawning
Pattems of seasonal reproductive activityof mudcrabs worldwide were reviewed byLe Vay (2001). Maturation and spawning ofScylla both in the wild and in captivity hasbeen observed year-round. Tbere are someseasonal peaks, though these differ accord-ing to climate and latitude. In this study thepeak spawning period in captivity over the19-mo period was in late winter (July andAugust). This coincided with the seasonalrise in sea surface temperature and mirroredthe peak in spawning for Sesarma meinerti,another large estuarine crab in the region(Enunerson 1994). This did not, however,fit weIl with the incidence of mature ovaries
Synchronicity of Spawning
Spawning synchronicity is commonamongs.~crustaceans (Olive 1992) and is of-ten associated with food abundance, result-ing in optimum egg quality. It also ensuresthat larvae encounter optimal conditions forgrowth (photoperiod and temperature) andfavorable dispersion of the larvae (currents)(Olive 1992; Emmerson 1994; Oreco andRodriguez 1999).
Although spawning synchronicity of S.serrata in captivity (Fig. 7) has not beenpreviously documented, it has been ob-served in Australia. Pheromones have beenshown to affect sexual behavior in crabs(Oleeson 1991). Evidence also exists thathafching eggs release hormones, which in-ftuence female crab behavior (Oleeson and
Smith 1984; Kamio et al. 2002; Tankersleyet al. 2002). Despite a paucity of evidencein the literature, it is possible that phere-mones could be released during spawningthat could induce other females to spawn.Researchers in the Philippines have, how-ever, not recorded synchronous spawning inmudcrabs kept in the same system. Also,attempts to induce maturation or spawningwith exogenous hormones in decapods suchis shrimp, for example, have been largelyunsuccessful (Chamberlain 1985). Tbereare two other possible explanations for thisphenomenon. FirstIy, that changes in con-ditions in the broodstock maturation systemacted as cues to spawning, but were so sub-tIe as to go undetected, and secondly, thatmudcrabs caught in 'tbe wild have molted,mated, and matured their ovaries with somedegreê of synchrony. This synchrony wasthen carried through the captive maturationprocess, resulting in synchronous spawning.Tbere is also some evidence that the lunarcycle coordinates egg laying and hatchingin the lobster Homarus gammarus, whichhas benefits for the larvae (Ferrero et al.2002). Although Hai et al. (2001) foundthat spawning in Scylla did not followmoon phases, no such correlation was re-corded in this study.
Tbere were no obvious correlations be-tween spawning patterns and parametersmeasured in the broodstock system duringthis study. li, however, spawning is syn-chronized by a naturally occurring cue, itcould be a useful tool for broodstock man-agement and would be a worthwhile area ofresearch. Whatever the mechanisms behindthis phenomenon, the results implied thatwhen using wild-caught females, more thanone batch of eggs was usually available forexperimental or rearing purposes.
Other Aspects of Spawning
As most of the crabs spawned soon afterintroduction to the maturation system, itwas unnecessary to ablate the eyestalk toinduce maturation and spawning, a relative-ly common practice for mudcrabs (Mann et
Fccundity(average Reproducûve
number of outpulCrab size: eggslfemale (number ofCW (rom) per batch X eggslbatch Geographical
Species mass (g) 1()6) per g female) locaûon Reference
Scylla sp. (described 129-175 2.1-4.0 153-433 India Srinivasagam el al.
as tranquebarica) 300-690 2000
Scylla sp. (described as 85-137 0.1-2.3 168-265 India Srinivasagam el al.
serrata) 98-340 2000
Scylla sp. 117-140 1.5-2 N/A India Marichamy and Raja-
N/A pacldam 1992
S. serrata N/A 1.5-3 N/A Sri Lanka Jyamanna and Jinadasa
NIA 1993
S. serrata 164-170 0.4-8.4 543-11.531 Australia Mann el al. 1999
744-820
Scylla (described as ser- N/A 0.8-2 N/A Malaysia Bin Jamari 1992
rata) 350-520
S. paramamosaln NIA 0.6-3.0 NIA Vietnam Hai el al. 200 I
200-300
S. paramamosaln 69-169 0.05-1.8 N/A Vietnam Dat 1999
170-790 (ZI larvae)
S. serrata NIA 1.2-1.6 2.319-5.262 Philippines Millamena and Bang-
350-400 caya 200 1
S. serrata 130-174 0.4-5.2 N/A Philippines Quiniûo el al. 2001
313-950 (ZI larvae)
Scylla sp. NIA 2.7-3.3 N/A lndonesia Djunaidah el al. 200 1
200-300
S. serrata 120-170 2.5-10.8 3.825-25.503 South Africa This study
270-1.000

11~
1.j
i'4
i~
j'~1
1~~'
I
J
1
" 1
. I
130
al. 1999; Millamena and Quinitio 2000;Millamena and Bangcaya 2001). The mat-uration tank size and stocking densitiesused in this study aUowed for an adequatenumber of crabs to be held in captivity toensure a relatively constant supply of viabieeggs.
We observed that when berried femalescould not access the spawning bins due tolarger territorial females occupying them,the eggs did not attach weU to the pleopodsand resulted in a complete loss of the batch.When enough bins were provided, however,the beach sand they contained proved anideal substrate, negating the need for the(more difficult to maintain) mud-based me-dium used by Djunaidah et al. (2001).
Hatching in the incubator tank occurredin the moming between 0800 and 1200 hfor 94% of the crabs. Incubation time at 27C corresponded with that observed by Haiet al. (2001) (9-12 d at 25-31 C) and forthe majority of batches did not vary bymore than 24 h (Tabie I). Moreover, thesmall variance in the increase in egg sizefrom extrusion to hatching between differ-ent crabs (Tabie I) means that, along withontogeny of the embryo, time after extru-sion and egg diameter could be used as atooI to predict the hatch date. This has im-portant implications for the preparation ofthe hatchery and enrichment of live food.
Conclusion
The relatively high fecundity and limitedseasonality of spawning of South AfricanS. serrata may be an adaptation to the var-iable nature of South African estuaries, ofwhich approximately 70% are closed bybarrier sandbars that open interrnittently(BeU et al. 2001; Viljoen and Cyrus 2002).The spawning characteristics of South M-rican S. serrata are nevertheless comparabieto those observed for the genus throughoutits distribution. This should be consideredwhen formulating future fisheries manage-ment or conservation policies if these are to
DAVIS ET AL. SPAWNING OF MUD CRAB 131
be adapted from those established else-where.
Using the fecundity data, the averagepercent hatching success and the percent fe-males that spawn on a monthly basis, a totalof 1,374,488 fust zoea (ZI) larvae could beexpected per kg of female per mo. For prac-tical aquaculture purposes, maintaining arelatively small number of female brood-stock (approximately 20 animals in thisstudy) under the correct artificial conditionscould ensure aregular, year-round supply ofeggs for supplying a hatchery. The com-paratively high fecundity of the crabsmeans that relatively few individuals needto be caught from the wild for stockingcommercial hatcheries, as opposed to pen-aeid shrimp, for example (Wickins and Lee2002). The crabs spawned easily with littlerequirement for special diets or environ-mental manipulation. The life cyc1ehas alsobeen c10sed in captivity (Quinitio et al.2001). This implies that domestication ofthe broodstock will not be difficuit, whichcould reduce the dependence on wild-caught broodstock in the future.
and assistance with the manuscript; Dr. Pe-ter Bossier for reviewing the manuscript.
Literature Cited
Barnes, R. D. 1987. Invertebrate zoology, fifth edition.Saunders College Pubijshing, New York, USA,
Bell, K. N. L, P. D. Cowley, and A. K. Whitfield.200 1. Seasonality in frequency of marine accessto an intermittently open esruary: implications forrecruitment strategies. Esruarine and CoastaI Shelf
Science 52(3):327-337. . ,Bin Jamarl, Z. 1992. Preliminary studies on rearing
tbe larvae of tbe mudcrab (Scylla serrata) in Ma-laysia. Pages 143-147 in C. A. AngelI, editor. Re-port of tbe seminar on mudcrab culrure and tradeheld at Surat Thani, Thailand, 5-8 November1991. Bay of Bengal Programme, Madras, India.
Brock, V. 1960. The introduction of aquatic animalsin Hawaiian waters. Hydrobiology 45:463-480.
Caddy, J. F. 1989. Marine invertebrate fisheries. Theirassessment and management. John WiJey andSons, New York, USA. .
Castille, F. L. and A. L. Lawrence. 1989. Relation-ship betWeen maturation and biochemical com-position of tbe gonads and digestive glands of tbesbrimps Penaeus-Azrecusives and Penaeus setifer-us (L). Journal of Crustacean Biology 9:202-211.
Chamberlaln, G. W. 1985. Biology and coótrol ofsbrimp reproduction. Pages ml-m42 in TexasSbrimp Farming Manual. Texas Agriculrura! Ex-tension Service Publication of Invited Papers pre-sented at tbe Texas Sbrimp Farming Workshop on19-20 November 1985. Corpus Cbristi, Texas,USA.
Dat, H. D. 1999. Preliminary srudies on rearing thelarvae of tbe mudcrab (Scylla paramamosain) inSoutb Vietnam. Pages 147-152 in C. P. Keenanand A. Blackshaw, editors. Mudcrab aquaculrureand biology. Proceedings of an international sci-en9fiè forum held in Darwin, AustraIia, 21-24April 1997. ACIAR proceedings no. 78. WalsonFerguson and Company, Brisbane, Australia.
Djunaldah, L S., D. Suwoyo, M. Wille, E. K. Kon-tara, and P. Sorgeloos. 2001. Investigations ontbe reproductive perfonnance of mudcrab Scyllaspp. broodstock: a research review (abstract). Pag-es 6-7 in D. A. Jones, L. Le Vay, M. Wille, T. T.Nghia, and V. N. Ut, editors. Workshop. on mud-crab rearing, ecology and fisheries, 8-10 January2001. Can Tho University, Vietnam.
Emmerson, W. D. 1994. Seasonal beeding cycles andsex ratios of 8 species of crabs from Mgazana, amangrove esruary in Transkei, Soutbern Africa.Journal of Crustacean Biology 14(3):568-578.
Fet;rero, E. A., N. PrIvIleggi, T. Scovacricchl, and.' G. Van Der Meeren. 2002. Doeslunar cyc1eaf-fect clawed lobster egg hatching and moulting fre-
Acknowledgments
This work was funded by research grantsfrom the Flernish Inter-university Council(VLIR.ZEIN 1998PR21O-913), the LibertyLife Education Foundation, the NRFthrough the South African Network forCoastal and Oceanic Research, and theRhodes University Research Committee.We would like to thank the following peo-ple without whose help this research wouldnot have been possible: Derek Dlamini andPat Khubisa for their help in catching andcaring for the crabs; the management ofAmatikulu Prawn and Robert Landman fortheir professional hospitality; CuIlam Bea-tie of Kwazulu Wildlife for his cooperation;Wendy Davidson late of the OceanographicResearch Institute for sharing her knowl-edge of mudcrabs; Mathieu Wille of theLaboratory of Aquaculture and ArterniaReference Center for his scientific guidance
quency of hatchery-reared juveoiles? OpheUa56(1):13-22.
Gardner, C. 1997. Effect of size on reproductive out-put of giant crabs Pseudocarcinus gigas (La-marck): Oziidae. Marine and Freshwater Research48(7):581-587.
Gleeson, R. A. 1991. Intrinsic factors mediating pher-emone communication in tbe blue crab Callinec-tes sapidus. Pages 17-32 in R. T. Bauer and J. W.Martin, editors. Crustacean sexual biology. Co-lumbia University Press, New YOrk. USA.
Gleeson, R. M. and A. Sm/th. 1984. Characterizationf of a sex pheremone in tbe blue crab Cal/inectes
sapidus: crustacdysone studies. Journal of Chem-ical Ecology 10:913-922.
Greco, L. S. L. and E. M. Rodriguez. 1999. Annualreproduction and growtb of adult crabs Chasmag-nathus granulata (Crustacea, Brachyura. Grapsi-dae). Cahiers De Biologie Marine 40(2):155-164.
Haddon, M. 1994. Size fecundity relationships, mat-ing behaviour and laljval release in tbe New Zea-land paddie crab, Ovalipes catharus (White 1843)(B~achyura, Ponunidae). New Zealand Journal ofMarine and Freshwater Research 28(4):329-334.
Hai, T. N., A. Hassan, A. T. Law and vNo A. Shazili.200 1. Some aspecls on maturation and spawningperfonnance of mudcrabs (Scylla sp.) in captiveconditions (abstract). Pages 8-9 in D. A. Jones,L. Le Vay, M. Wille, T. T. Nghia, and V. N. Ut,editors. Workshop on mudcrab rearing, ecologyand fisheries, 8-10 January 2001. Can Tho Uni-versity, Vietnam.
Hamasaki, K. aod K. Haitai. 1993. Prevention offungal infection in tbe eggs and larvae of tbeswimming crab POrlunis triturbicularus and tbemudcrab Scylla' serrata by batb treatment withfonnalin. Nippon Suisan Gakkaishi 59(6):1067-1072.
Heasman, M. 1980. Aspecls of tbe genera! biologyand fishery of tbe mudcrab Scylla serrata (For-skAI) in Moreton Bay, Queensland. Doctora! tbe-sis. University of Queensland, Queensland, Aus-tralia.
Heasman, M. P. and D. R. Fielder. 1977. The man-agement and exploitation of tbe Queensland mud-crab fishery. Australian Fishery 36:4-8.
Heasman, M. P. aod D. R. Fielder. 1983. Laboratoryspawning and mass rearing of tbe mangrove crab.Scylla serrata (ForskäI). from first zoea to firstcrab stage. Aquaculrure 34:303-316.
Heasmao, M. P., D. R. Fielder, and R. K. Shepard.1985. Mating and spawning in tbe mudcrab, Scyl-la serrata (ForskäI) (Decapoda: Portunidae) inMoreton Bay, Queensland. Australian Journal ofMarine and Freshwater Research 36:773-783.
Hili, B. J. 1975. Abundance, breeding and growtb oftbe crab Scylla serrata (ForskAI) in two Soutb Af-rican esruaries. Marine Biology 32:21-24.

..~
~
I'I
j
!I~
~
~
~~...;;
132
Hili, B. J. 1994. Offshore spawning by tbe Portunidcrab Scylla serrata (Crustacea. Decapoda). MarineBiology 120(3):379-384.
Jyamanna, S. C. and J. Jinadasa. 1993. Size at ma-turity and spawning periodicity of tbe mudcrabScylla serrata (Forskäl) in tbe Negombo estuary.Joumal of tbe Natura! Scientific Council of SriLanIca 21(10):141-152.
Kamio, M., S. Matsunaga, and N. Fusetani. 2002.Copulation pheremone in the crab Telnessuscheiragonus (Brachyura: Decapoda). MarineEcology Progress Series 234:183-190.
Keenan, C. P., P. J. F. Davie, and D. L. Mann. 1998.A revision of tbe genus Scylla (DeHaan, 1833)(Crustacea: Deeapoda: Brachyura: Portunidae).Rafftes Bulletin of Zoology 46(1):217-245.
Koslow, J. A. 1992. Fecundity and tbe stock recruit-ment relationship. Canadian Joumal of Fisheriesand Aquatic Science 49:210-217.
Lardies, M. A. and J. S. Wehrtmann. 1996. Aspectsof tbe reproductive biology of Petro/isthes laevi-gatus (Guerin 1835) (Decapoda. Anomura, Por-cellanidae), part I. Reproductive output and chem-ical composition of eggs during embryonic devel-oproent. Archive of Fishery and Marine Research43(2):121-135.
Le Vay, L. 2001. Ecology and management of mud-crab Scylla spp. Asian Fisheries Science 14(2):101-112.
Mann, D., T, Asakawa, and A. Blackshaw. 1999.Performance of mudcrab Scylla serrata brood-stock held at Bribie Island Aquaculture ResearchCentre. Pages 10I-I 06 in C. P. Keenan and A.Blackshaw, editors. Mudcrab aquaculture and bi-ology. Proceedings of an international scientificforum held in Darwin, Australia. 21-24 April1997. ACIAR proceedings no. 78. Watson Fer-guson and Company, Brisbane, Australia.
Mantelatto, F. L. M. and A. Fransozo. 1997. Feeun-dity of tbe crab Callinectes ornatus Ordway, 1863(Deeapoda, Brachyura, Portunidae) from tbe Uta-ba region Sao Paulo, BraziI. Crustaceana 70(2):214-226.
Marichamy, R. and S. Rajapacldam. 1992. Experi-ments on larval rearing and seed production of tbemudcrab Scylla serrata (Forskä1). Pages 135-142in C. A. Angell, editor. Report of tbe seminar ontbe mudcrab culture and trade held at Surat Thani,Thailand, 5-8 November 1991. Bay of BengalProgramme, Madras, India.
MllIamena, O. M. and J. P. Bangcaya. 2001. Repro-ductive performance and larval quality of pondraised Scylla serrata females fed various brood-stock diets. Asian Fisheries Science 14:153-159.
Mlllamena, O. M. and E. T. Quinitio. 2000. The ef-fects of diets on tbe reproducti ve performance ofeyestalk ablated and intact mudcrab Scylla serra-ta. Aquaculture 181:81-90.
DA VIS ET AL.
Nghia, T. T., M. WiUe, and P. Sorgeioos. 2001. Ef-fects of light, eyestalk ablation and seasonal cycleon tbe reproductive performance of captive mud-crab (Scylla paramamosain) broodstock in tbeMekong delta (abstract). Pages 4-5 in D. A.Jones, L. Le Vay, M. Wille, T. T. Nghia, and V.N. Ut, editors. Workshop on mudcrab rearing,ecology and fisheries, 8-10 January 2001. CanTho University, Vietnam.
Nonnan, C. P. and M. B. Jones. 1993. Reproductiveecology of tbe velvet swimming crab, Necora-puber (Brachyura, Portunidae) at Plymoutb. Jour-nal of tbe Marine Biological Association of tbeUnited Kingdom 73(2):379-389.
Ollve, P. J. W. 1992. The adaptive significance ofseasonal reproduction in marine invertebrates-tbe importance of distinguishing between modeis.Invertebrate Reproduction and Development22(1-3):165-174.
Ong, K. S. 1966. Observations on the post larvallifehistory of Scylla serrata Forskä1, reared in tbe lab-oratory. The Malaysian Agricultural Joumal45(4):429-443.
Prasad, P. N. and B. Neeiakantan. 1989. Maturityand breeding of tbe mudcrab Scylla serrata (For-skäl) (Decapoda, Brachyura, Portunidae). Pro-ceedings of tbe Indian Academy of Sciences-Anima! Science 98(5):341-349.
QuJnitio, E. T., F. D. Parado-Estepa, O. M. Milla.mena, E. Rodriguez, and E. Borlongan. 2001.Seed production of mudcrab Scylla serrata juve-niles. Asian Fisheries Science 14:161-174.
Robertson, W. D. 1989. Factors affecting catches oftbe crab Scylla serrata. (Forskä1) (Decapoda: Por-tunidae) in baited traps: soak time, time of dayand accessibility of tbe bail Estuarine, Coastaland Shelf Science 29: 161-170.
Robertson, W. D. and A. Kruger. 1994. Size at ma-turity, maling and spawning in tbe Portunid crabScylla serrata (Forskä1) in Natal, Soutb Africa.Estuarine, Coastal and Shelf Science 39:185-200.
Srinivasagam, S., M. Kathirvel, and S. Kulasekara-pandian. 2000. Caplive broodstock development,induced breeding and larval stages of mudcrabs(Scylla spp.). Bullelin no. 12. March 2000. CentralInstitute of Brackishwater Aquaculture (lndianCouncil of Agricultura1 Research), Chennai, India.
Sokal, R. R. and F. J. Rohlf. 1995. Biometry, tbeprinciples and practice of statistics in biologicalresearch, third edition. W. H. Freeman and Com-pany, New York, USA.
Tamura, T. 1970. Propagation of Scylla serrata. Ma-rine Aquaculture 15: 1-3.
Tankersley, R. A., T. M. Bullock, R. B. Forward,and D. Rittschof. 2002. Larval release behavioursin tbe blue crab Callinectes sapidus: role of chem-ical cues. Joumal of Experimental Marine Biologyand Ecology 273(1):1-14.
"
SPAWNING OF MUD CRAB
VlIjoen, A. and D. P. Cyrus. 2002. Temporal varia-tion in tbe recruitment of larval fish into tbe Mhla-tuze estuary in northem KwaZulu-Natal, SoutbAfrica. Marine and Freshwater Research 53(2):439-445.
Warner, G. F. 1977. The biology of crabs..Paul Elek(Scientific Books) Ltd.,.London, England.
....
133
Wlcklns, J. F. and D. O'C. Lee. 2002. Crustaceanfarming. ranching and culture, second edition.Blackwell Science, Oxford, England.
Williams, M. J. and J. H. Primavera. 2001. Choos-ing trapical portunid species for culture, domes-tication and stock enhancement in tbe Indo-Pacif-ic. Asian Fisheries Science 14(2001):121-142.





























