Embed Size (px)
Citation preview

In the format provided by the authors and unedited.
Supplementary figures sparse panicle1 is required for inflorescence development in Setaria viridis and maize Author affiliations: Pu Huanga,+, Hui Jianga,+, Chuanmei Zhua, Kerrie Barryb, Jerry Jenkinsc, Laura Sandorb, Jeremy Schmutzb,c, Mathew S. Boxa, Elizabeth A. Kellogga, Thomas P. Brutnella,* a Donald Danforth Plant Science Center, 975 N Warson Rd, St. Louis, MO 63132, USA b Department of Energy Joint Genome Institute, Walnut Creek, California, USA c HudsonAlpha Institute for Biotechnology, Huntsville, Alabama, USA + These authors contributed equally to this work
Sparse panicle1 is required for inflorescencedevelopment in Setaria viridis and maize
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
SUPPLEMENTARY INFORMATIONVOLUME: 3 | ARTICLE NUMBER: 17054
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 1
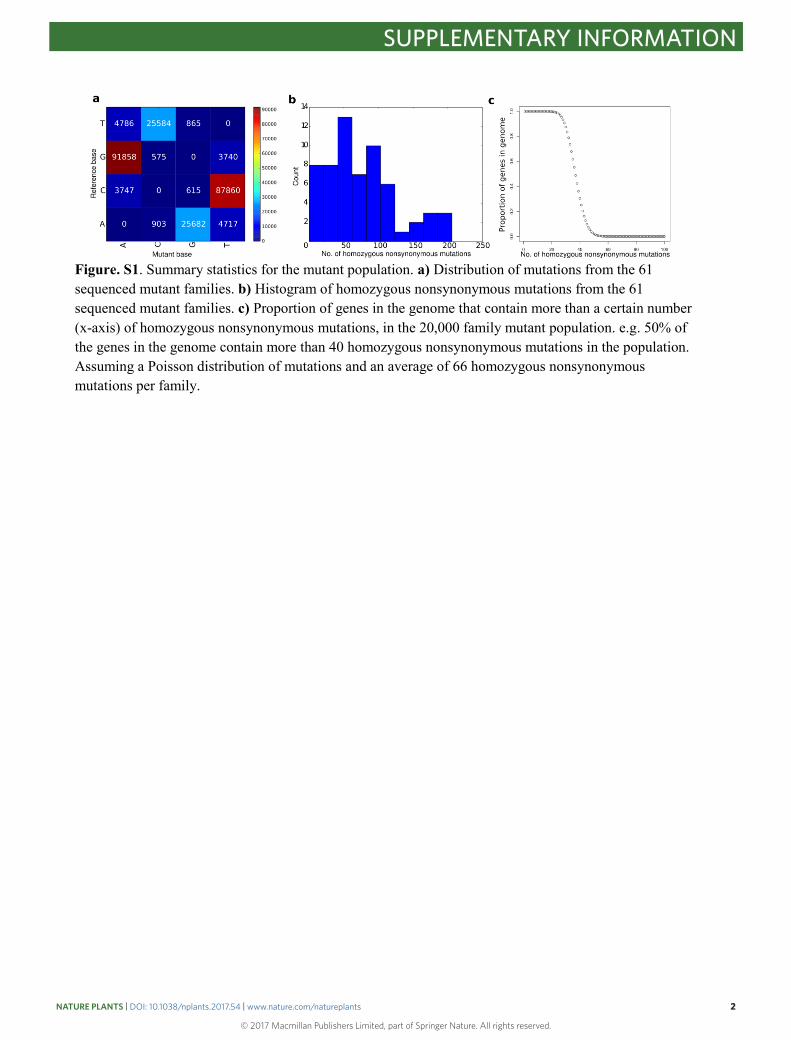
Figure. S1. Summary statistics for the mutant population. a) Distribution of mutations from the 61 sequenced mutant families. b) Histogram of homozygous nonsynonymous mutations from the 61 sequenced mutant families. c) Proportion of genes in the genome that contain more than a certain number (x-axis) of homozygous nonsynonymous mutations, in the 20,000 family mutant population. e.g. 50% of the genes in the genome contain more than 40 homozygous nonsynonymous mutations in the population. Assuming a Poisson distribution of mutations and an average of 66 homozygous nonsynonymous mutations per family.
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 2
SUPPLEMENTARY INFORMATION
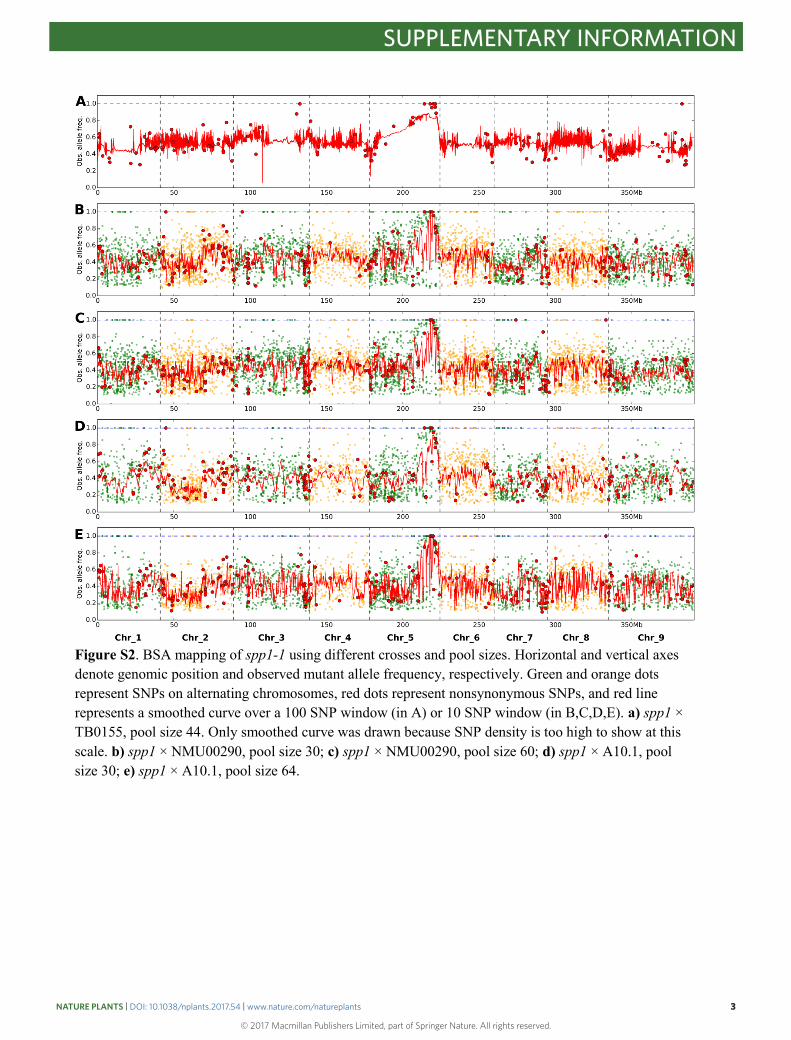
Figure S2. BSA mapping of spp1-1 using different crosses and pool sizes. Horizontal and vertical axes denote genomic position and observed mutant allele frequency, respectively. Green and orange dots represent SNPs on alternating chromosomes, red dots represent nonsynonymous SNPs, and red line represents a smoothed curve over a 100 SNP window (in A) or 10 SNP window (in B,C,D,E). a) spp1 × TB0155, pool size 44. Only smoothed curve was drawn because SNP density is too high to show at this scale. b) spp1 × NMU00290, pool size 30; c) spp1 × NMU00290, pool size 60; d) spp1 × A10.1, pool size 30; e) spp1 × A10.1, pool size 64.
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 3
SUPPLEMENTARY INFORMATION

Figure S3. Phenotypic variation of quantitative traits in SvAUX1 and ZmAUX1 mutants. a) Plant height in S. viridis b) Panicle length in S. viridis. c) Spikelet number in S. viridis (partial), measured from bottom 0.5 to 1.5 cm. d) Root bending angle in S. viridis, measured 24h after plant rotation. e) Plant height in maize. f) Tassel length in maize. g) Spikelet count per row of maize ear. h) Root bending angle in maize, measured 24h after plant rotation. i) Spikelet count of tassel central spike in maize, measured from bottom 0 to 5cm. j) Tassel central spike length in maize. k) Spikelet count of tassel first primary branch, measured from bottom 0 to 5cm. l) Tassel first primary branch length. All bar heights show mean values, error bars show standard deviations, p-values were calculated from two-tailed t-tests and n= number of individuals examined.
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 4
SUPPLEMENTARY INFORMATION

Figure S4. Phylogenetic and expression analysis of AUX1 homologs. a) Phylogeny of AUX1 homologs from A. thaliana and grasses. The tree topology is same as shown in Figure 1F. b) Local synteny between SvAUX1 and ZmAUX1. Plot is based on CoGe (https://genomevolution.org/coge/GEvo.pl). Red and green boxes indicate SvAUX1 and ZmAUX1, respectively. c) Expression profile of SvAUX1 and its homologs in representative tissue/organs in S. viridis. d) Expression profiles of ZmAUX1 and its homologs in representative tissue/organs in maize. All expression data were obtained from phytozome 11 (http://phytozome.jgi.doe.gov/).
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 5
SUPPLEMENTARY INFORMATION

Figure S5. Phenotypic variations of Zmaux1-0. a) WT W22 maize tassel. b) Phenotypic variation of of Zmaux1-0 tassel. Red arrows show nodes without primary branch, white arrow shows branch with very few spikelets. c) Phenotypic variations in maize ear morphology, showing bald top and reduced number of spikelets in Zmaux1-0. Harvested on day 61 after planting.
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
NATURE PLANTS | DOI: 10.1038/nplants.2017.54 | www.nature.com/natureplants 6
SUPPLEMENTARY INFORMATION





























